interaction of root-knot nematode, meloidogyne incognita ... · interaction of root-knot nematode,...
TRANSCRIPT

Indian J. Nematol. 4 (1974) : 69-80
INTERACTION OF ROOT-KNOT NEMATODE, MELOIDOGYNE INCOGNITAAND TOBACCO MOSAIC VIRUS IN TOMATO
BY
B. K. GOSWAMI and V. V. CHENULU
Division of Mycology and Plant Pathology, Indian Agricultural Research InstituteNew Delhi, India
A survey of fields around Delhi showed that TMV and root-knot nematode were
commonly associated with tomato plants. Investigations were, therefore, taken up to study thenature of their association as also the effect of virus-nematode complex on plant growth characters.
The plant growth was significantly reduced in simultaneous inoculations with both the pathogens
(N+ V) or when nematode preceded the virus in the inoculations (N +v) in comparison to inoculations with virus (V) or nematode (N) alone or where virus preceded the nematode inoculation
(V +n). Further, there was a significant increase in the nematode population in N + V treatmentsbut virus multiplication was inhibited. As against this, in the treatment where virus preceded
inoculation, the plant growth and virus multiplication was not affected but the nematode population
was suppressed. No significant difference in the trend of these results was recorded between the
observations taken after 20, 30, 40, 50 and 60 days of inoculations and also between the differentinoculation intervals of virus and nematode.
Histopathological studies showed large number of giant cells with necrotic cortical andvascular tissues in the simultaneous inoculations and also in the inoculation where nematodes
were allowed to establish prior to virus. In these treatments nematodes were found to be present
in group of two or more as against single nematode in N or V +n treatments.
A survey of tomato fields in and around Delhi indicated that plantsaffected with virus were also in many cases infected with root-knot nematode (predominantly M. incognita). Investigations were, therefore, undertaken to find outinteraction, if any, of TMV and root-knot nematode and effect on the growth oftomato seedlings. The results of these studies and histopathology of the affectedplants are presentel in this paper.
MATERIALS AND METHODS
Seeds of tomato (Lycopersicon esculentum Mill) var. Pusa ruby (highlysusceptible to both TMV and root-knot nematode) were surface sterilized with

70 B. K. GOSWAMI AND V. V. CHENULU
I : 1000 mercuric chloride solution and sown in pans containing autoclaved sandyloam soil. Twenty days old seedlings were transplanted singly in 6 cm plastic potscontaining sterilized sand-soil mixture. The virus (CPO-TMV of Mathur et al.,1966) was sap inoculated on the upper surface of tomato leaves with a small pieceof sterilized absorbent cotton wool previously soaked in the inoculum containingfinely powdered carborundum (about 600 mesh) to serve as abrassive. Preliminarystudies indicated that virus dilution of 1 : 2000 was convenient for assaying on thelocal lesion host, Chenopodium amaranticolor Costa and Reyn. For estimating theviral concentration of various inocula, local lesion bioassay technique (Holmes,1929) was employed, using half leaf method so that a particular treatment could becompared with the control inoculations of the same leaf.
For inoculations with nematode, infested roots were taken from nema
tode culture pots, egg masses were picked up and allowed to hatch at 25-28°C.Hatched larvae were collected every 24 hours and stored at 4°C in a refrigerator.Before inoculations, the number of larvae per ml of suspension was determined' withthe help of Peter's 1 ml.eelworm counting slide. Approximately equal amount oflarval suspension (about 1000 per pot) was poured in holes (5 to 6 holes made witha glass rod around base of stem) which were closed immediately with top soil.The inoculated pots were kept on a cemented floor to avoid contamination. ABexperiments were conducted under insect proof glass house conditions.
A t the end of the experiment each plant was cut at the base of stem andshoot height and weight were recorded. The roots of these plants were carefullywashed to remove adhering soil particles, gently blotted between folds of blottingpaper and then measured and weighed. One g of nematode affected root fromeach of the nematode treated sets was made into homogeneous root mixture andnumber of galls were counted under a stereoscopic microscope. In case of composite galls, the number of primary galls was estimated according to the methoddescribed by Jones and Nirula(1963). The root mixture used in counting rootgalls were then stained in 0.05 per cent lactophenol-acid fuchsin (Franklin, 1949).The stainecl root pieces were then macerated with 200 ml of water with a few dropsof glacial acetic acid in a waring blendor for 30 ,seconds at high speed and thenumber of larvae along with all the different nematode stages were determined.Population in 5 ml suspension was multiplied with the volume factor to give thetotal population.
Root materials from virus and nematode treatments as also in different
combinations along with healthy ones were transferred to Bouin's fixative and left
in it for 48 hours. The material was then dehydrated in alcohol seri,esand kept in

M. INCOGNITA AND TOBACCO MOSAIC VIRUS 71
methyl benzoate. To obtain a normal appearance of the tissue, the material wasstored in 2% celloidin solution in methyl benzoate for three days and then washedin benzene (three changes of ten minutes each). Paraffin (M. P. 56°C) was used forembedding the roots. Sections were cut at 12ft thick. For staining Johanson's(1940) Safranin-fast green process was used.
RESULTS
Effect of standard extract (SE) of TMV and different nematode levelssingly and in combinations on plant growth, virus concentration and nematodepopulation.
The observations were recorded after two months and the results are
presented in Table 1. A significant reduction in shoot length and weight of plantswas recorded ,at all nematode inoculum levels when inoculated simultaneously withTMV. With an increase in the initial nematode level there was a correspondingreduction in shoot and root length and weight, the maximum reduction being atN 1000+SE. The nematode population increasectwith increasing inoculum levels.In the simultaneous inoculations there were significantly more galls on roots(Plate I-A) as also free soil population of larvae in comparison to nematode aloneseries. Apparently there was greater multiplication of nematode in the presence ofvirus. The virus concentration, on the other hand, was significantly reduced in thepresence of nematode infestation, the maximum reduction being at N1000 level.
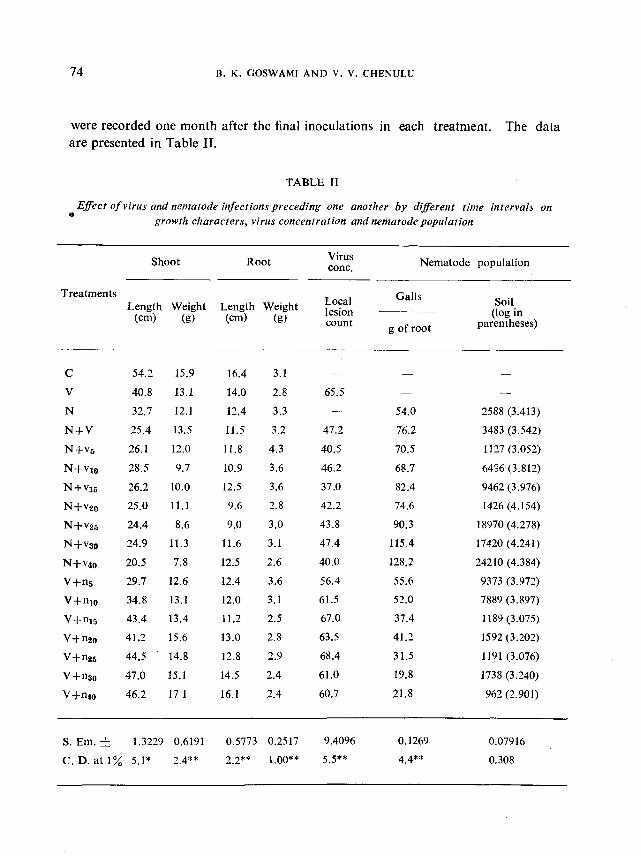
Effect of virus and nematode infections preceding one another by differenttime intervals on.growth characters, virus concentration and nematode population
In the previous experiment a population level of 1000 nematodes per500 g of soil reduced plant growth significantly. Further, significant reduction inplant growth was recorded also in the combined inoculations of nematodes plusSE of virus. These levels were, therefore, used in the present experiment with theobjective of studying the effect of time intervals in the inoculations by the twoorganisms on plant growth as also on virus and nematode multiplication as againstthe single inoculations or simultaneous infections oUhe two organisms.
A total of 18 treatments was set up. In seven of these, nematodeinoculations preceded virus inoculations by time intervals ranging from 5 to 40 days(N + Vsto N + v40). In the next seven treatments, virus preceded nematode inoculations by similar time intervals (V+ns to V+n,o). Single inoculations withnematode (N) and virus (V), simultaneous inoculations by the two (N + V) anduninocnlated cohtrol (C) formed the rest of the four treatments. The observations

TABLE I
Effect of standard extract (SE) of TMV and different inoculum levels of root-knot nematode singly as well as in combinationson plant characters, virus concentration and nematode population
.....•N
Treatments
C
VSB
NI0
Nl00
NIOO
NI000
NloSB
Nl00SE
N500sE
Nl000SE
S.Em.±
C. D. at 1%
Shoot RootVirus cone.Nematode population?'
Galls
~
Length
WeightLength(Weight)Local lesion Soil0(cm)
(g)(cm)(g)countg ofroot(Log in parentheses)0VJ~>65.9
26.818.24.0 -- -i5
60.4
24.315.03.2 57.4>- -Z
t:160.519.217.32.5 -14.8
5000 (3.699):<55.0
18.815.43.1 -57.54875 (3.688):<
50.1
17.612.62.8 85.114350 (4.157)(j- :I:
m43.018.010.83.1 -115.8 37840 (4.578)Z
~56.919.012.02.8 50.235.5 13300 (4.124)
t'"
~51.6
18.215.03.2 48.572.514000 (4.146)
46.0
17.113.63.1 43.6105.018920 (4.277)
29.4
10.512.82.6 30.6165.152000 (4.716)
0.8660
0.75280.83260.3214 1.55662.0912 0.02834
3.5**
3.0**3.3**N.S 7.0**9.0** 0.11**
.* Highly significant; N. S. Not significant

M. INCOGNITA AND TOBACCO MOSAIC VIRUS
PLATE 1
73
A
c
n
D
A. Root systems N (1000 larvae) and N +V (1000 larvae + SE) treatments.B. An adult female nematode feeding on the host cells in N+ V treatment.
C. Multinucleate giant cells ,in N +V treatment.D. Group of giant cells in vascular region in N + V treatment.
74 B. K. GOSWAMI AND V. V. CHENULU
were recorded one month after the final inoculations in each treatment. The dataare presented in Table n.
TABLE IT
Effect of virus and nematode infections preceding one another by different time intervals on•• growth characters, virus concentration qnd nematode population
Shoot
RootVirusNematode populationcone.
Treatments
GallsLength Weight
Length WeightLocal Soil
(cm)(g)(cm)(g)lesion
(log incount g of root
parentheses)
-----
------~-
C
54.215.916.43.1
V
40.813.114.02.865.5
N
32.712.112.43.3-54.0 2588 (3.413)
N+V
25.413.511.53.247.2 76.23483 (3.542)
N+vs
26.112.011.84.340.5 70.51127 (3.052)
N+VIO
28.59.710.93.646.2 68.76436 (3.812)
N+VlS
26.210.012.53.637.0 82.49462 (3.976)
N+V20
25.011.19.62.842.2 74.61426 (4.154)
N+V2S
24.48.69.03.043.8 90.318970 (4.278)
N+vao
24.911.311.63.147.4115.417420 (4.241)
N+V40
20.57.812.52.640.0128.224210 (4.384)
V+ns
29.712.612.43.656.4 55.69373 (3.972)
V+nlO
34.813.112.03.161.5 52.07889 (3.897)
V+n15
43.413.411.22.567.0 37.41189 (3.075)
V+n20
41.215.613.02.863.5 41.21592 (3.202)
V+n2S
44.514.812.82.968.4 31.51191 (3.076)
V+nao
47.015.114.52.461.0 19.81738 (3.240)
V+n40
46.217116.12.460.7 21.8962 (2.901)
S.Em.±
1.32290.61910.57730.25179.40960.12690.07916
C. D. at 1% 5.1*
2.4**2.2**1.00**5.5**4.4**0.308
M. INCOGNlTA AND TOBACCO MOSAIC VIRUS 75
10
10
v~
20 10 40 50 N
'"i• 40
t:~ 1 '0
..
f .0
"
••••m ••
V+n
70 ~-
10 = i'" ~ I40P30 !to 1I10 s-o•·••10",.11,. I20
.. j;24 ::•• I!lJ•• I!.SQ !;••
0:
•• ••••
MYI·. ID ell_'0
OATS
10-;0-;0
N+v
"'~'liij,fll It:i.htlm;
FJ SHOOT ••lENGTH· 0 a ••• I ••
1;1 _ooT LENGTH_O_le. IC•
m SH001 W('GH'. 0 I~ I,•• OOT wlEllHT • 02: c•• ,
~ VI"US CONCENT'lliln<*. o. c•• , toe.' I•••• A
[;J NIEIlATOOf ~no~ • I c-.• 1000 •• /ft.'Of ••
~......N+V-50
, i 40•.I ~i50
"'~ '0z~J
'0 I! r,~I
T - 0g •
'00: • t
III• I5~
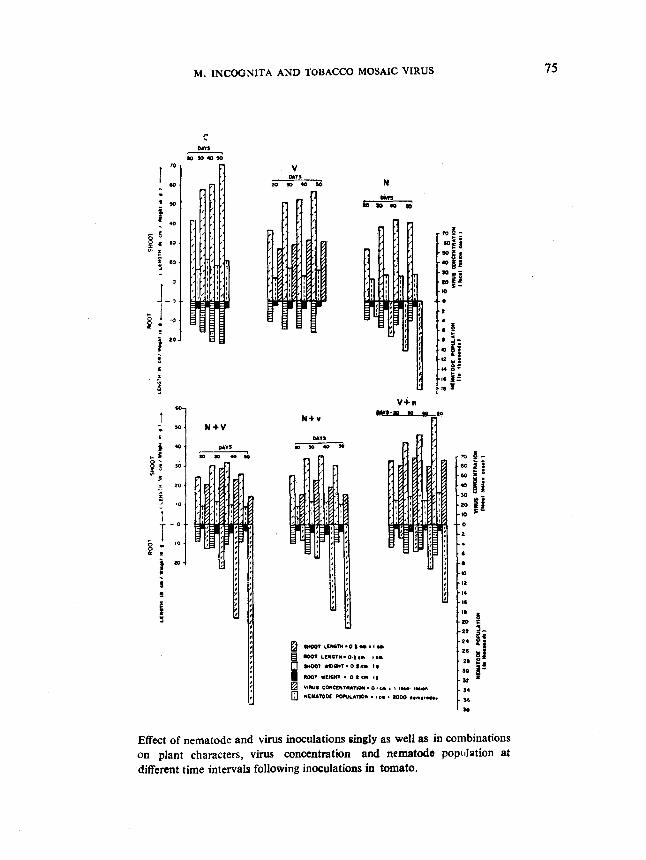
Effect of nematode and virus inoculations singly as well as in combinationson plant characters, virus concentration and nematode population atdifferent time intervals following inoculations in tomato.

76 B. K. GOSWAMI AND V. V. CHENULU
Apart from the fact that significant reductions in plant growth were
obtained with virus as well as nematodes, singly or in simultaneous inoculations in
comparison to the uninoculated control, it is interesting to observe that the time
interval in the establishment of two organisms was an important factor at least in
the multiplication of the two organisms. It may be seen that in comparison to
simultaneous inoculations there was no significant difference, generally speaking, in
shoot length, root length or weight when nematodes preceded virus. But signi
ficantly better growth was obtained by a ten day delay in the nematode inoculations
(V +nl0 and onwards), more strikingly in shoot length than other characters. The
shoot weight however, in some cases showed significantly depressed growth at all
time intervals except where nematode preceded virus by 30 days (N +V30).
Compared to virus alone (V) treatment, virus multiplication was signi
ficantly reduced in treatments where nematodes preceded virus inoculations and also
in the treatments where virus preceded nematode inoculations hy 5 days (V +n5)'
However, except for N +V5, N+V15 and N+V40 virus multiplication was comparableto that obtained in N + V treatment.
The nematode gall formation on root was significantly more in all treat
ments where nematode preceded virus and also where virus preceded nematode
inoculations upto 10 days. M;ore than 10 days interval in nematode inoculations
after virus, n~duced multiplication of the nematode. There was significantly better
multiplication where nematode was established 30 or 40 days before introduction
of virus in plants.
Effect of nematode and virus inoculations singly as well as in combinationson plant characters, virus concentration and nematode population at different timeintervals following inoculations
Generally, experimental observations are recorded a month after inocula
tions where root-knot nematodes are involved. This experiment was initiated with
a view to find out the effect of different time intervals on plant growth, virus con
centration and nematode multiplication in single and combined inoculations.
Six treatments namely, J) virus alone (V), 2) nematode alone (N), 3)nematode and virus si multaneously (N +V), 4) virus inoculations ten days prior tonematode inoculation (V +n), 5) virus inoculation ten days after nematode inocula
tion (N +v) and 6) un inoculated control Cc) were maintained. Each treatment
with four sets was replicated five times and the observations were recorded at inter
vals of ten days starting from 20 days after inoculations and continued upto 50days. The data are presented in Fig. 1.

M. INCOGNITA AND TOBACCO MOSAIC VIRUS 77
. It may be seen that even at 20 days after inoculations significant differences in plant growth, virus concentration and nematode multiplication were noticeable and the same trend continued for 50 days.
A comparison of nematode developmental stages and multiplication innematode alone and nematode plus virus inoculations
Based on the results of the previous experiments it was found that thenematode multiplication increased significantly when the plant was also infectedwith the virus. The present experiment was, therefore, planned to corroboratethis observation and to compare the nematode multiplication and developmentalstages in nematode alone versus nematode plus virus inoculations. The treatmentswere nematode alone (N), simultaneous inoculations with nematode and virus(N+V), nematode preceding virus inoculation by ten days (N+v) and viruspreceding nematode inoculation by ten days (V+n). The data are presented inTable HI.
TABLE III
Nematode multiplication and the developmental stages in nematode alone and nematode
plus virus inoculations
It will be seen that compared to the nematode alone treatment therewas significant increase in the number of galls/ g of root weight as also in the soilpopulation in the simultaneous inoculations (N + V) as well as in the treatmentwhere nematode inoculations preceded virus inoculations (N + V). A similar trend

B. K.. G()SW AMI AND V. V. CHfNULU
was also recorded with regard to developmental stages of nematode with feastnumber of females and males formed in V+n treatment. Apparently inclusion ofvirus as a partner along with nematode or afterwards (10 days) stimulated nematode multiplication.
Histopathologfcal studies
The virus affected roots showed changes in shape and size of thinwalled parenchymatous cells in thl.' cortex as also in the pith region. The cells wereenlarged or narrow and tangentially elongated. Generally the vascular bundles didnot show any effect but in some cases of severe virus infections the phloem parenchyma cells were deformed and elongated abnormally.and the parenchymatous cells.in the cortical region appeared to be multiplying like cambial cells.
With nematode alone (N) or where nematodes were partners with virus.in the inoculations (N+V, N+v and V+n) hypertrophy and hyperplasia of thecells, formation of tyloses and also abnormal cell divisions in different tissues of thehost were prominent. The cortical cells in case of severely affected plants showedsuberization of walls. Generally, in N+V and N+o inoculations the nematodeswere found to be feeding mostly in the xylem or the phloem tissues (Plates I-C andD). The number of adult famales,egg sacs and giant cells with prominent swellingof nuclei were more in N + V and N + v treatments as compared to N or V+n(Plate I-B, C and D). In these treatments, the xylem vessels appeared broader andthinner than the normal and the part near the cambium was largely parenchymatous.The fibres in the initial stages appeared septate and reduced in number. Finallythese disappeared and even the xylem vessels and tracheids ruptured. The phloemcells were almost absent with the disturbance of the entire tissue. Cortical cells in
many places started dividing like cambial cells as a protective measure and hypertrophy and hyperplasia was much more prominent in these two treatments.Generally extensive damage was found in the roots of N+Vand N+v treatmentscompared to other treatments.
DISCUSSION
The data presented above show that a competition exists between thepathogens (TMV and root-knot nematode) when they infect simultaneously acommon host like tomato. The interrelationship between the two pathogens of thedisease complex was basically of antagonistic type where growth and reproductionof one pathogen took place at the cost of the other. The results indicate that thedominance of one pathogen over the other was dependent on the time of estahlish-

M. INCOGNITA AND TOBACCO MOSAIC VIRUS 79
ment of the pathogen. Thus, when virus established first, as in V +n treatment,
the host became unfavourable for the other pathogen (M. incognita) of the disease
complex. Subsequently this resulted in the suppression of soil nematode popula
tion, number of galls produced in the roots as well as the developmental stages of
the nematode within the roots. On the contrary, nematode population and galling
increased in treatments where nematode had precedence over the virus (Table 11).
Generally, nematode established itself in plant roots within forty-eight hours
whereas measurable quantities of virus in the host could be detected only afterninety-six hours.
The growth and development of the host appears to be related to the
type of inoculation. Though virus alone caused a significant reduction in plant
growth characters over control, the results show that the nematode is more dama
ging to tomato than virus singly or in different combinations. The synergistic
effect of the two pathogens on the plant growth characters is apparent in N+V and
N+v inoculations but when virus was allowed to establish first (V +n) the nematode
apparently did not contribute to loss in plant growth vigour.
Swarup & Goswami (1969) earlier recorded significant reductions in
plant growth and increased nematode population in simultaneous infections by root
knot nematode and leaf curl virus on tomato and our results are in conformity with
this finding. Similar observations have also been made by Goffart (1956) for
sugarbeet infected with Heterodera schachtii and yellow disease; Ryder and
Crittenden (1962) for soybean infected with M. incognita acrita and tobacco ring
spot virus, and Khurana et al. (1970) for maize infected with M. incognita and
maize mosaic virus. However, Bird (1969) did not find any synergism between
the tobacco ringspot virus and root-knot nematode in bean plants.
The suppression of nematode population as a result of prior· establish
ment of TMV (V + n treatment) is in agreement with the work of Schlosser (1962)
who demonstrated inhibition of Heterodera schachtii by beet yellows virus. A
similar type of nematode suppression due to virus was reported by Weischer (1970)
who observed 80% and 89% less populations respectively of Aphelenchoider ritzemahosi and Ditylenchus dipsaci in tobacco plants infected with TMV than in virus
free plants.
Histopathological studies also showed that there was increased activity
of the nematode in N +V or N +v treatments than where virus had precedence over
nematode (V+n). Ryder & Crittenden (1962) similarly recorded greater clumpingof nuclei in the vicinity of nematode's head when soy bean plants were inoculated

80 B. K. GOSWAMI AND v. v. CHENULU
with both tobacco ringspot virus and M. incognita acrita. Swarup & Goswami(1969) also recorded more number of giant cells with numerous nuclei in root oftomato plants affected with leaf curl virus and M. incognita than nematode aloneinoculations.
ACKNOWLEDGEMENT
We record our grateful thanks to Dr. S. P. Raychaudhuri, Head,Division of Mycology and Plant Pathology, Indian Agricultural Research Institute,New Delhi, for constant encouragement and providing necessary facilities for work.Thanks are also due to Dr. Gopal Swarup, Plant Pathologist (Nematode) for goingthrough the manuscript and giving valuable suggestions.
REFERENCES
BIRD, A. F. (1969). The influence of tobacco ringspot virus and tobacco mosaic virus on thegrowth of Moloidogyne javanica. Nematologica 15 : 201-209.
FRANKLlN, M. T. (1949). A quick method of demonstrating nematodes of the genus Aphelenchoides in leaves. J. Helm. 23 : 91-93.
GOFFART, H. (1956). Uber das zusammenwirken von rubennemakoden (Heterodera schachtii)und vergilbungskrankheiten (Beta virus 4). Mede. de Statte Gent. 21(1) : 351-360.
HOLMBS, F. O. (1929). Local lesion in tobacco mosaic. Bot. Gaz. 87 : 39.
JONBS, F. G. W. & NIRULA, K. K. (1963). Hatching tests and counts of primary lIalls in assessments of nematicides against Meloidogyne spp. Plant Pathology 12 ; 148-154.
KHURANA, S. M. P., GOSWAMI, B. K. & RAYCHAUDHURI, S. P. (1970). Interaction of maizemosaic with root-knot nematode Meloidogyne incognita (Kofoid and White) Chitwood in maize (Zea mays L.). PhYlOpath. Z. 69 : 267-272.
MATHUR, S. B., MISHRA, S. tb. & TIWARI, V. P. (1966). A new strain of tobacco mosaic virusaffecting chilli pepper variety Puri Orange. PI. Dis. Reptr. 50 : 619-622.
RYDBR, H. W. &CRITTBNDBN, H. W. (1962). Interrelationship of tobacco ringspot virus andMeloidogyne incognita acrita in roots of soybean. Phytopathology 52 : 165-166.
SCHLOSSBR, E. (1962). Unter Suchungen Zum Wirt-Parasit-Verhaltnis der Beta Arten und desRubennematoden (Heterodera schachtii). Zeitschrift fur Pflanzenschutz 69 : 18-20.
SWARUP, G. & GOSWAMI, B. K. (1969). Interrelationship of root-knot nematode and leaf curlvirus in tomato. Indian J. Exptl. Bioi. 7(1) : 64-65.
WBISCHBR, B. (1969). Multiplication and pathogenicity of Aphelenchoides ritzemabosi and Di/y-lenchus dipsaci in healthy and in TMV infested tobacco. Nematologica 15:334-336.
Acceptedfor publication: 20 June 1974