investigation of koi herpesvirus latency in koi - journal of virology
TRANSCRIPT

JOURNAL OF VIROLOGY, May 2011, p. 4954–4962 Vol. 85, No. 100022-538X/11/$12.00 doi:10.1128/JVI.01384-10Copyright © 2011, American Society for Microbiology. All Rights Reserved.
Investigation of Koi Herpesvirus Latency in Koi�
Kathleen E. Eide,1 Tim Miller-Morgan,1,3 Jerry R. Heidel,1 Michael L. Kent,1,2 Rob J. Bildfell,1Scott LaPatra,4 Gregory Watson,1 and Ling Jin1,2*
Department of Biomedical Sciences, College of Veterinary Medicine, Oregon State University, Corvallis, Oregon 973311; Department ofMicrobiology, College of Science, Oregon State University, Corvallis, Oregon 973312; Oregon Sea Grant Extension,
Oregon State University, Corvallis, Oregon 973313; and Research Division,Clear Springs Foods, Inc., Buhl, Idaho 833164
Received 2 July 2010/Accepted 23 February 2011
Koi herpesvirus (KHV) has recently been classified as a member of the family of Alloherpesviridae withinthe order of Herpesvirales. One of the unique features of Herpesviridae is latent infection following aprimary infection. However, KHV latency has not been recognized. To determine if latency occurs inclinically normal fish from facilities with a history of KHV infection or exposure, the presence of the KHVgenome was investigated in healthy koi by PCR and Southern blotting. KHV DNA, but not infectious virusor mRNAs from lytic infection, was detected in white blood cells from investigated koi. Virus shedding wasexamined via tissue culture and reverse transcription-PCR (RT-PCR) testing of gill mucus and feces fromsix koi every other day for 1 month. No infectious virus or KHV DNA was detected in fecal secretion or gillswabs, suggesting that neither acute nor persistent infection was present. To determine if KHV latentinfections can be reactivated, six koi were subjected to a temperature stress regime. KHV DNA andinfectious virus were detected in both gill and fecal swabs by day 8 following temperature stress. KHV DNAwas also detectable in brain, spleen, gills, heart, eye, intestine, kidney, liver, and pancreas in euthanizedkoi 1 month post-temperature stress. Our study suggests that KHV may become latent in leukocytes andother tissues, that it can be reactivated from latency by temperature stress, and that it may be morewidespread in the koi population than previously suspected.
Koi herpesvirus (KHV), a newly identified virus, is highlycontagious to fish and may cause high mortality (80 to 100%)in common carp and koi (Cyprinus carpio) (2, 10, 11). The firstoutbreak of KHV was reported in 1998 in Israel (7). Sincethen, KHV infections have been reported in the United States,Europe, and Asia (2, 10–12). This viral disease affects fish ofvarious ages; however, it causes higher mortality in fry than inolder fish (5, 22). The clinical signs of KHV infection includered and white mottling of the gills, gill hemorrhage, sunkeneyes, and pale patches or blisters on the skin (7). The virus canbe found in the kidney, gill, spleen, fin, intestine, and brain (8).In experimental studies, 82% of fish exposed to the virus at awater temperature of 22°C (which is 7 to 9°C above theirnormal environmental temperature of 13 to 15°C) died within15 days (21).
The complete genome has been sequenced from three KHVstrains from Japan, the United States, and Israel (1). The KHVgenome is about 295 kbp, and contains a 22-kbp terminal directrepeat. KHV thus has the largest genome reported to date forthis family (1). KHV is also known as cyprinid herpesvirus 3(CyHV3) and has been proposed to be a member of Alloher-pesviridae family in the order Herpesvirales (31). Alloherpesvi-ruses are distinct and highly diverged from both the Herpes-viridae and Malacoherpesviridae. KHV is closely related to
CyHV1 (carp pox herpesvirus) and CyHV2 (goldfish hemato-poietic necrosis virus).
One of the unique features of Herpesviridae is latency. La-tency is the most remarkable property of herpesviruses, ensur-ing the maintenance of their genetic information in their hostsfor an extended period in the absence of productive infection(20, 25). There are three subfamilies within Herpesviridae: Al-phaherpesvirinae, Betaherpesvirinae, and Gammaherpesvirinae(25). Members of the three herpesvirus subfamilies infect awide variety of target cells and are able to establish latentinfection, which is associated with a restricted gene expressionof the viral genome with no production of infectious virus (20).Most alphaherpesviruses become latent in the peripheral ner-vous system, such as sensory ganglia and dorsal root ganglia(15, 33). The betaherpesviruses become latent in bone marrow,lymphoid tissue, and kidney (3, 20). Some gammaherpesvirusbecome latent in splenic B cells (20, 23).
Another member of the Alloherpesviridae family, the channelcatfish virus (CCV), has been suggested to become latent fol-lowing a primary infection, based on detection of CCV DNA inperipheral blood leukocytes (9, 29). However, the question ofwhether KHV becomes latent or persists following a primaryinfection remains unresolved (4, 24, 27, 28). There is evidenceshowing that asymptomatic fish are KHV carriers (30). It isimportant to know whether KHV can become latent and re-activate from latency as this characteristic would not only con-stitute a cornerstone for developing detection and controlstrategies for the virus but also shed light on our understandingof herpesvirus evolution. Currently, the only method to screenKHV infection is via detection of KHV antibody by standardserum neutralization assay or by KHV antigen-specific en-
* Corresponding author. Mailing address: Department of Biomedi-cal Sciences, College of Veterinary Medicine, Oregon State University,Corvallis, OR 97331. Phone: (541) 737-9893. Fax: (541) 737-2730.E-mail: [email protected].
� Published ahead of print on 9 March 2011.
4954
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/j
vi o
n 26
Dec
embe
r 20
21 b
y 59
.3.7
5.44
.

zyme-linked immunosorbent assay (ELISA) (27), which maybe limited by test sensitivity. In this study, we examined thestatus of KHV latency in clinically normal koi with a history ofprobable exposure to KHV.
MATERIALS AND METHODS
Source of koi and sampling. Six koi (designated K1 to K6) between 2 and 15years old were from premises that had previous KHV infections or exposure.Three (K2, K4, and K5) were recently imported koi that had been housed in thesame quarantine facility. Prior to donation, K4 and K6 tested positive for KHVby serum antibody ELISAs carried out at the Immunology and Virology Labo-ratory, Veterinary Medicine Teaching Hospital, University of California, Davis(29). The remaining three koi (K1, K2, and K5) were survivors from a pondassociated with a suspected KHV outbreak in 1998 and a confirmed KHVoutbreak in 2003, based on a positive PCR test at the University of GeorgiaInfectious Disease Laboratory. To investigate whether KHV becomes latent inthe peripheral leukocytes, 0.5- to 2.0-ml blood samples from fish K1 to K6 werecollected and stored in EDTA tubes at 2 weeks, 1 month, and 2 months followingarrival of the fish at the Oregon State University, Salmon Disease Research Lab(OSU-SDL). Three sets of blood samples from these six koi were collected toensure the consistency of KHV genome detection as persistence of the genomeover time is a characteristic of latency. The OSU-SDL is specifically designed forconducting in vivo experiments with infectious diseases. The incoming water isfrom a deep well and is pretreated with UV irradiation and is thus not a sourcefor KHV.
Five 2-year-old koi were obtained from facilities with no known history ofKHV problems, and these were designated KI to KVI. An additional four 2-yearold koi were obtained from a local pet store, and these were designated KVI toKIX. These four koi were home bred by a local pet owner, and there are norecords of KHV infection in the breeding facility. Both groups of koi were keptsegregated and maintained at 12°C in 4-ft-diameter tanks at OSU-SDL in ac-cordance with the Animal Care and Use Committee regulations. All bloodsamples were collected via caudal vein puncture after the koi were anesthetizedwith MS-222 (100 ppm). To investigate whether KHV becomes latent in certaintissues, all koi in this study were euthanized via MS-222 (500 ppm) overdose.Tissues, including the brain, spleen, gills, heart, eye, intestine, kidney, liver, andpancreas, were collected at necropsy.
CCB and KF-1 cell lines. Both a common carp brain (CCB) cell line and koifin cell line (KF-1) (gift of Ronald Hedrick, University of California, Davis) werecultured in Dulbecco’s modified Eagle’s medium (DMEM) (Invitrogen, Carls-bad, CA) supplemented with 10% fetal bovine serum (Gemini Bio-Products,West Sacramento, CA), penicillin (100 U/ml), and streptomycin (100 �g/ml)(Sigma-Aldrich, Inc., St. Louis, MO) and incubated at 22°C. The strains of KHVfrom the United States and Israel (KHV-U and KHV-I, respectively) were giftsof Ronald Hedrick.
Detection of persistent infection by examining KHV in body secretions. Thepresence of KHV DNA or infectious virus in body secretions is indicative ofpersistent infection. To determine whether persistent infection was present, weswabbed fish K1 to K6 every other day for 1 month to see if we could detect thepresence of KHV or KHV DNA on gill surfaces and within the anal vent. TheseDacron swabs were placed in 0.5 ml of sterile DMEM (Invitrogen, Carlsbad, CA)containing penicillin (200 U/ml) and streptomycin (200 �g/ml) (Sigma-Aldrich,Inc., St. Louis, MO). Five-microliter aliquots of swab solution were tested byreal-time PCR for KHV DNA, and 0.2 ml of swab solution was inoculated toCCB cells seeded in 12-well plates for KHV virus detection.
Separation of peripheral WBC and total DNA extraction from WBC andplasma. Blood collected from the caudal vein into a syringe previously coatedwith heparin (Sigma-Aldrich, Inc., St. Louis, MO) at 1,000 U/ml in phosphate-buffered saline (PBS) was transferred to an EDTA tube. Blood was centrifugedat 650 � g at 4°C for 10 min; the buffy coat was collected and exposed to 3 to 4volumes of red blood cell lysis buffer (Tris-NH4Cl). The remaining white bloodcells (WBC) were washed twice in sterile DMEM (Invitrogen) by centrifugationat 650 � g at 4°C for 10 min (Becman XJ). Then, WBC from each blood samplewere subjected to a total DNA extraction (yielding WBC total DNA) using aHigh Pure PCR Template Preparation Kit (Roche Diagnostics, Indianapolis,IN). Total DNA was extracted similarly from the plasma pellet that was collectedafter ultracentrifugation of 250 to 500 �l of the plasma layer at 25,000 rpm for1 h min at 4°C (Beckman model XL-70) in an SW28 rotor. Approximately, 0.1 to0.5 �g/�l total DNA can be isolated from WBC obtained from each fish. TheDNA concentration was adjusted to 0.1 �g/�l before use in PCR.
Virus culture and virus isolation. KHV was cultured in CCB or KF-1 cell linesthat were maintained in DMEM (Invitrogen, Carlsbad, CA) supplemented with5% fetal bovine serum (Gemini Bio-Products, West Sacramento, CA), penicillin(100 U/ml), and streptomycin (100 �g/ml) (Sigma-Aldrich, Inc., St. Louis, MO)at 22°C. KHV virus isolation from tissue samples was performed by homogeniz-ing frozen tissue samples in DMEM (1:5 ratio [wt/vol]), centrifuging the tissuehomogenate at 2,000 � g for 10 min, filtering the homogenate through a 0.45-�m-pore-size filter (Waterman), and inoculating supernatant of the tissue ho-mogenate into 25-cm2 flasks seeded with CCB cells (0.5 ml of tissue preparationper 25-cm2 flask). To monitor virus shedding in the gill and feces, each gill orfecal swab medium (0.2 ml/swab) was inoculated onto CCB cells seeded in12-well plates. Visible characteristic KHV cytopathogenic effect (CPE) was con-sidered positive isolation. If initial inoculation failed to produce KHV CPE thefirst time, total cell lysate was reinoculated onto CCB cells seeded in 12-wellplates and incubated at 22°C. Swabs or tissue preparations failing to produceKHV CPE in CCB cells the second time were considered KHV negative. StockKHV cultures were inoculated onto one well of each plate as a positive control.
Purification of KHV DNA. Viral DNA for positive controls was extracted fromeither virions or purified intracellular nucleocapsids as described previously (13).Briefly, the purified virions or nucleocapsids were digested in 10 mM Tris-HCl(pH 8.0), 100 mM EDTA, 1% N-lauroyl sarcosine, and 200 �g/ml proteinase Kovernight at 55°C. The viral DNA was extracted twice with an equal volume ofphenol-chloroform (1:1 [vol/vol]) and then precipitated with two volumes ofethanol and 1/10 volume of sodium acetate. The precipitate was washed once in70% ethanol and dissolved in TE buffer (10 mM Tris-HCl [pH 8.0]–1 mMEDTA).
KHV reactivation by temperature stress. To determine whether KHV latencycan be reactivated, the koi tank water temperature was increased from 12°C to23°C at a rate of 1°C per day. The temperature was then held constant for 4 daysat 23°C before being dropped back to 12°C at a rate of 1°C per day (see Fig. 3A).Anal vent and gill swabs were collected from all six koi every other day startingat day 2 post-temperature increase until day 16 post-temperature stress. Swabswere collected and transported in 0.5 ml of sterile PBS containing penicillin (200U/ml) and streptomycin (200 �g/ml) (Sigma-Aldrich, Inc., St. Louis, MO). Five-microliter aliquots of swab solution were tested by real-time PCR for KHVDNA, and 0.2 ml of swab solution was inoculated onto CCB cells seeded in12-well plates for KHV virus detection.
Two koi, designated K5 and K6, died on day 20 post-temperature stress (i.e.,20°C). These fish were necropsied, and tissue samples were collected, includingthe brain, spleen, gills, heart, eye, intestine, and kidney. One month after com-pletion of the temperature stress regime, the remaining four koi were bled asdescribed above and then euthanized to permit collection of tissues, includingthe brain, spleen, gills, heart, eye, intestine, kidney, liver, and pancreas, vianecropsy.
Total DNA extraction from tissue samples. Tissue samples (approximately 100to 200 mg) obtained from freshly euthanized or dead koi were stored at �80°C.Before DNA extraction, the frozen tissues were homogenized in 800 �l of 1�lysis buffer by 2.5-mm silica beads (Biospec Product) and digested overnight at55°C in the presence of 100 �g of proteinase K. Genomic DNA was thenextracted from the tissue lysates with a High Pure PCR Template PreparationKit according to the manufacturer’s instructions (Roche Diagnostics, Indian-apolis, IN). Approximately 0.1 to 1 �g/�l total DNA could be extracted fromeach tissue. All the tissue DNA was adjusted to 0.1 �g/�l before being used inPCRs. For each sample, 5 �l of total DNA (about 0.5 �g) was used in real-timePCR or PCR.
Total RNA extraction from WBC and KHV-infected KF-1 cells. To determineif persistent infection occurs in the WBC, viral mRNA expressed during lyticinfection was examined in total white blood collected from four healthy koiobtained from a local pet store; these fish had tested positive for KHV DNA inWBC by PCR. Total RNA was extracted from the combined WBC from the fourkoi using TriZol (Invitrogen). As a positive control, total cellular RNA fromKHV-infected KF-1 cells was harvested at 8 days postinfection and extracted byusing TriZol in accordance with the manufacturer’s instructions. Total RNAfrom uninfected KF-1 cells served as a negative control. The isolated total RNAwas resuspended in RNase-free H2O and examined for KHV lytic infection byreverse transcription-PCR (RT-PCR). The extracted total RNA from WBC(WBC total RNA) and KF-1-infected or uninfected cells was adjusted to 0.5�g/ml before use in RT-PCRs.
Primers and probes. Selection of primers for KHV sequence amplification wasbased on conserved DNA sequences of KHV (AF411803). Real-time PCR prim-ers were selected as previously described (8): KHV-86f (5�-GACGCCGGAGACCTTGTG-3�), KHV-163r (5�-CGGGTTCTTATTTTTGTCCTTGTT-3�), andTaqMan probe KHV-109p (5�-6FAM-CTTCCTCTGCTCGGCGAGCACG-TA
VOL. 85, 2011 KHV BECOMES LATENT IN LEUKOCYTES 4955
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/j
vi o
n 26
Dec
embe
r 20
21 b
y 59
.3.7
5.44
.

M-3�, where FAM is 6-carboxyfluorescein and TAM is 6-carboxytetramethylrho-damine). Another real-time PCR primer set specific for a host gene encodingglucokinase was also used as an internal control to equalize the amount of inputtotal DNA. All the real-time PCRs for KHV DNA were run with equal amountsof DNA estimated by real-time PCR of the glucokinase gene. The primer se-quence and TaqMan probe were selected as reported previously (8). The primersused for screening for the presence of viral DNA in tissue samples and tissueculture fluid were complementary to the KHV DNA polymerase and openreading frame 26 (ORF26) sequences: KHVDF-242 and KHVDR-242 for theKHV DNA polymerase gene; KHVF-447 and KHVR-447 for KHV ORF26(Table 1). To further probe the amplified DNA sequence specific for KHV,another set of nested primers was selected: KHVNF242 and KHVNR242 as theDNA polymerase gene probe (Table 1) and KHV263F and KHV263R as theORF26 probe (Table 1).
RT-PCR. First-strand cDNA was synthesized from 2.5 �g of total RNA byusing 10 pmol of random primer and Superscript Reverse Transcriptase III(Invitrogen) according to the manufacturer’s recommendations. PCR amplifica-tion with KHV-specific primers for detection of cDNA of KHV genes expressedduring lytic infection was performed using a 25-�l reaction mixture consisting of22.5 �l of amplification buffer (Platinum PCR Supermix; Invitrogen, Carlsbad,CA), a 1.25 �M concentration of each primer, and 2.5 �l of the completed RTreaction mixture. The mixture was subjected to 34 cycles of 94°C for 30 s, 55°Cfor 30 s, and 72°C for 45 s and then incubated at 72°C for 5 min after the finalcycle.
PCR amplification. PCR amplification with KHV-specific primers for detec-tion of viral DNA in tissues or culture fluid was performed using a 25-�l solutionconsisting of 19 �l of amplification buffer (Platinum PCR Supermix; Invitrogen,Carlsbad, CA), a 0.4 �M concentration of each primer, and 5 �l of total DNA(�0.5 �g) or 5 �l of swab medium. The mixture was subjected to 94°C for 2 min,and 35 cycles of 94°C for 30 s, 55°C for 30 s, and 72°C for 1 min, followed by a10-min elongation reaction at 72°C after the final cycle. Real-time PCR wasperformed according to the manufacturer’s instructions for quantitative PCR(qPCR) Supermix (Platinum qPCR Supermix-UDG with ROX; Invitrogen,Carlsbad, CA).
Plasmid preparation for KHV DNA real-time quantitation. The PCR productamplified from KHV-U DNA with real-time PCR primers was cloned by aTOPO 2.1 PCR cloning vector (Invitrogen, Carlsbad, CA). The correct insert wasscreened by restriction digestion and then sequenced by the Center for GenomeResearch and Biocomputing (CGRB) at Oregon State University. This plasmidproduct was employed to set up the standard curve for measuring the viral DNAcopy number in tissue samples or swab fluid.
Southern blotting. PCR products (40% of total PCR with 0.5 �g of DNAtemplate) generated using WBC or tissue DNA were electrophoresed through a1.5% agarose gel, transferred to a nylon membrane (14), and then UV cross-linked to the membrane. The DNA products were then probed with a digoxi-genin (DIG)-labeled DNA probe. The probe was generated with nested PCRprimers that are specific for the target genes. To make digoxigenin-labeled PCRproducts, digoxigenin-labeled deoxynucleoside triphosphates (Roche Diagnos-tics, Indianapolis, IN) were added to the PCR mixtures according to the manu-facturer’s instructions (Roche Diagnostics, Indianapolis, IN). The membranewas prehybridized with prehybridization buffer (Roche Diagnostics, Indian-apolis, IN) at 68°C and then hybridized with the DIG-labeled DNA probesspecific for the gene coding for DNA polymerase or major capsid protein at 68°C.After incubation with the probe, membranes were washed with 0.1% sodiumdodecyl sulfate and 10% 20� SSC (1� SSC is 0.15 M NaCl plus 0.015 M sodium
citrate) before incubation with an antidigoxigenin antibody conjugated withperoxidase. The membrane was then developed by incubation with a chemilu-minescent peroxidase substrate (Roche Diagnostics, Indianapolis, IN). The blotswere exposed to film (Kodak) at room temperature for 30 min to 2 h. Themolecular masses of the resulting bands were estimated by using a 1-kb DNAladder (Invitrogen, Carlsbad, CA).
DNA sequencing and analysis. The PCR products were cleaned with aChargeSwitch PCR Clean-Up Kit (Invitrogen, Carlsbad, CA) before sequencingand were sequenced by the CGRB at Oregon State University. The nucleotidesequences were analyzed with the Geneiou software.
RESULTS
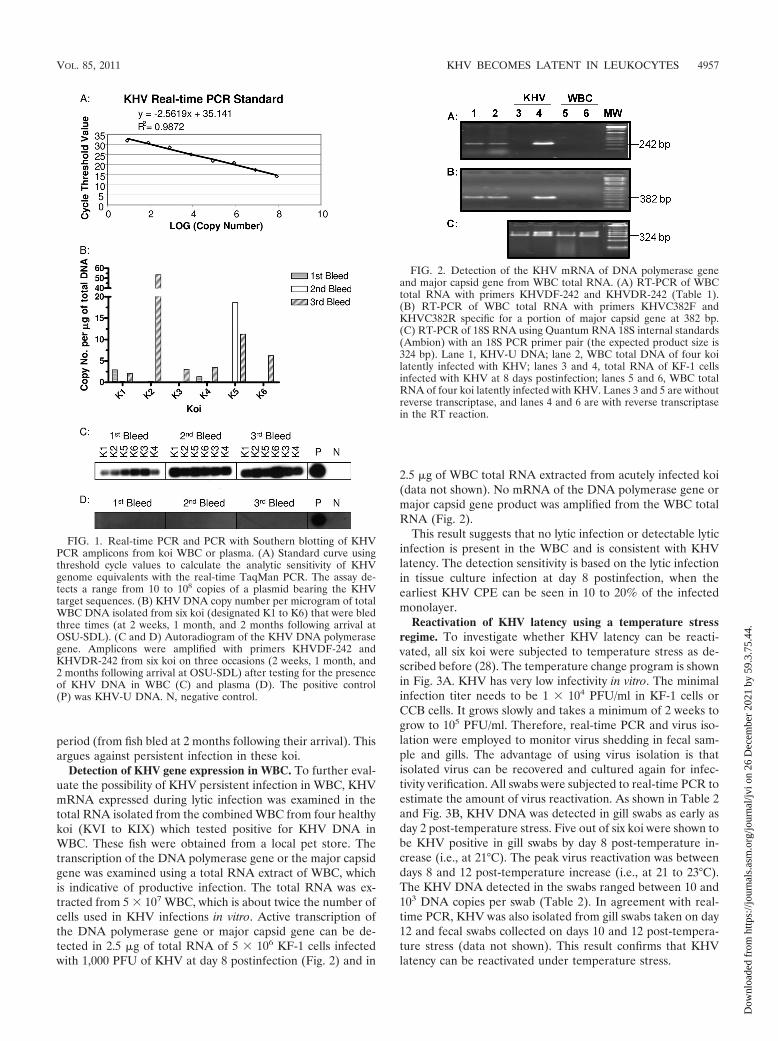
Detection of KHV DNA in leukocytes of previously exposedkoi by real-time PCR and PCR coupled with Southern blot-ting. Herpesvirus latency is characterized by an absence ofinfectious virus and the presence of the viral genome in thelatently infected tissue. To investigate whether KHV becomeslatent in koi peripheral leukocytes, WBC from fish K1 to K6were examined by both real-time PCR (8) and PCR with prim-ers (KHVD242F and KHVD242R) specific for the KHV DNApolymerase gene in blood taken 2, 4, and 8 weeks after arrivalof the fish at OSU-SDL (8). To estimate the amount of DNAin the latently infected tissue, DNA copy number was deter-mined by a real-time PCR standard established by using 10-fold serial dilutions of known KHV DNA from 108 to 0 copiesof the plasmid bearing the KHV DNA insert (Fig. 1A). Al-though KHV DNA was not detected in every blood sample(Fig. 1B), KHV DNA was detected in WBC of all six koi in thethird blood draw, when a minimum of 1-ml of whole blood wasused for WBC isolation. The amount of detectable KHV DNAwas approximately 2 to 60 copies per microgram of the totalWBC DNA, which is very common for herpesvirus latency (26,32). This suggested that KHV may become latent in koi whiteblood cells.
To further prove that the PCR product was specific forKHV DNA, the amplicons with primers KHVD242F andKHVD242R were then hybridized using a DNA probe specificfor the DNA sequence between the region amplified by the twoprimers (Table 1). As shown in Fig. 1C, KHV DNA was de-tected in all six koi and in all three blood draws for each fish.To eliminate the possibility that the virus was present in theplasma, each koi plasma sample was also examined by PCRand Southern blotting. As shown in Fig. 1D, no detectableKHV DNA was present in the plasma, further suggesting thatthe viral DNA resides inside the leukocytes and is not presentas free virus in the plasma.
Detection of KHV in body secretions of previously exposedkoi. The presence of KHV DNA and infectious virus in bodysecretions is indicative of persistent infections. To evaluate thepossibility of KHV persistent infection in these six koi, gillswabs and vent swabs were examined by both real-time PCRand virus isolation. No KHV DNA was detected from any gillswabs or vent swabs taken from all six koi every other day for1 month although positive and negative controls performed asexpected (data not shown). No KHV was isolated from swabswhile positive controls yielded viral growth. In addition, bydirect inoculation of WBC culture supernatant onto CCB cellsseeded in 12-well plates, no KHV infectious virus was detectedin koi WBC taken before this time period (from fish bled at 2weeks and 1 month following their arrival) and after this time
TABLE 1. Primer pairs used to detect KHV DNA and KHVgene transcription
Namea Target Primer sequence
KHVDF-242 DNA polymerase 5�-TGTGCGCCAACTCTCACTAC-3�KHVDR-242 5�-GCCCTTGGTGTAGAGGTTCA-3�KHVNF242 5�-CACGTCCAGAGGGTTCATCT-3�KHVNR242 5�-AGTCCCTCTGCCAGCATCT-3�KHVF-447 ORF26 5�-GCTTACGCCAATTCCACAGT-3�KHVR-447 5�-GCTCGTGGTCGTAGGTATCG-3�KHV263F 5�-CCCAACCACGATAACGAA-3�KHV263R 5�-GGGTGGTAGTGTTCGGGTA-3�KHVC-382F Major capsid 5�-TCTCACCCAGTACACCACCA-3�KHVC-382R 5�-GTTCATGGCGCCAAAGTAGT-3�
a Forward and reverse primers are indicated by F and R, respectively, in theprimer names.
4956 EIDE ET AL. J. VIROL.
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/j
vi o
n 26
Dec
embe
r 20
21 b
y 59
.3.7
5.44
.
period (from fish bled at 2 months following their arrival). Thisargues against persistent infection in these koi.
Detection of KHV gene expression in WBC. To further eval-uate the possibility of KHV persistent infection in WBC, KHVmRNA expressed during lytic infection was examined in thetotal RNA isolated from the combined WBC from four healthykoi (KVI to KIX) which tested positive for KHV DNA inWBC. These fish were obtained from a local pet store. Thetranscription of the DNA polymerase gene or the major capsidgene was examined using a total RNA extract of WBC, whichis indicative of productive infection. The total RNA was ex-tracted from 5 � 107 WBC, which is about twice the number ofcells used in KHV infections in vitro. Active transcription ofthe DNA polymerase gene or major capsid gene can be de-tected in 2.5 �g of total RNA of 5 � 106 KF-1 cells infectedwith 1,000 PFU of KHV at day 8 postinfection (Fig. 2) and in
2.5 �g of WBC total RNA extracted from acutely infected koi(data not shown). No mRNA of the DNA polymerase gene ormajor capsid gene product was amplified from the WBC totalRNA (Fig. 2).
This result suggests that no lytic infection or detectable lyticinfection is present in the WBC and is consistent with KHVlatency. The detection sensitivity is based on the lytic infectionin tissue culture infection at day 8 postinfection, when theearliest KHV CPE can be seen in 10 to 20% of the infectedmonolayer.
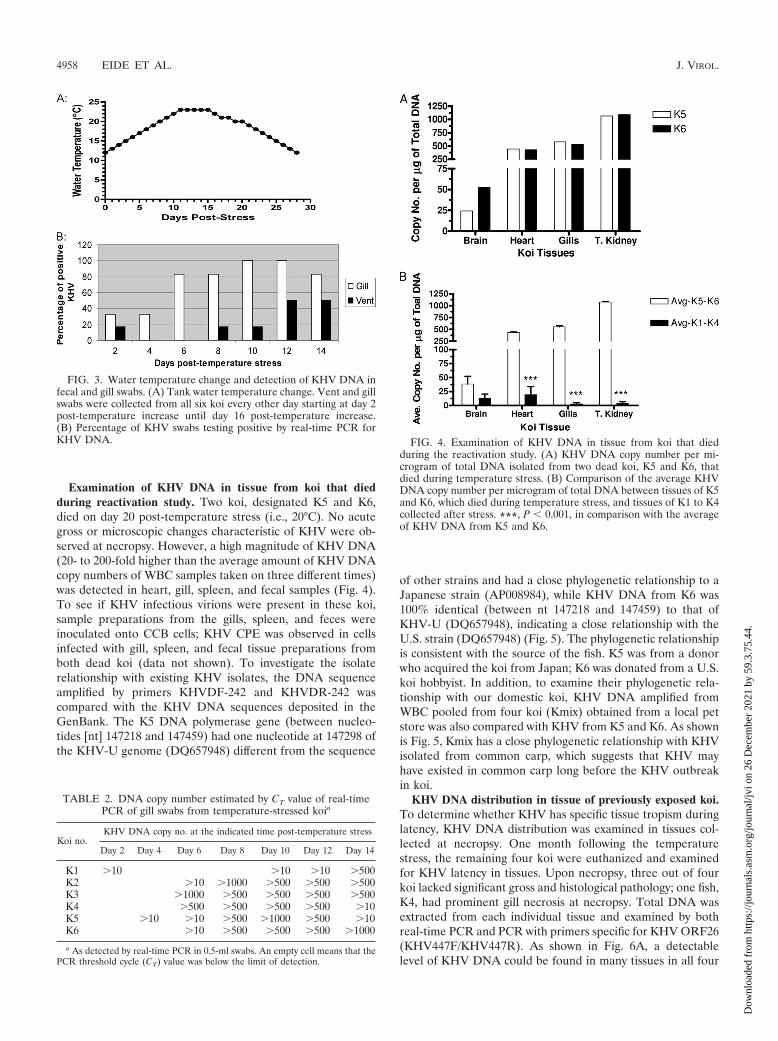
Reactivation of KHV latency using a temperature stressregime. To investigate whether KHV latency can be reacti-vated, all six koi were subjected to temperature stress as de-scribed before (28). The temperature change program is shownin Fig. 3A. KHV has very low infectivity in vitro. The minimalinfection titer needs to be 1 � 104 PFU/ml in KF-1 cells orCCB cells. It grows slowly and takes a minimum of 2 weeks togrow to 105 PFU/ml. Therefore, real-time PCR and virus iso-lation were employed to monitor virus shedding in fecal sam-ple and gills. The advantage of using virus isolation is thatisolated virus can be recovered and cultured again for infec-tivity verification. All swabs were subjected to real-time PCR toestimate the amount of virus reactivation. As shown in Table 2and Fig. 3B, KHV DNA was detected in gill swabs as early asday 2 post-temperature stress. Five out of six koi were shown tobe KHV positive in gill swabs by day 8 post-temperature in-crease (i.e., at 21°C). The peak virus reactivation was betweendays 8 and 12 post-temperature increase (i.e., at 21 to 23°C).The KHV DNA detected in the swabs ranged between 10 and103 DNA copies per swab (Table 2). In agreement with real-time PCR, KHV was also isolated from gill swabs taken on day12 and fecal swabs collected on days 10 and 12 post-tempera-ture stress (data not shown). This result confirms that KHVlatency can be reactivated under temperature stress.
FIG. 1. Real-time PCR and PCR with Southern blotting of KHVPCR amplicons from koi WBC or plasma. (A) Standard curve usingthreshold cycle values to calculate the analytic sensitivity of KHVgenome equivalents with the real-time TaqMan PCR. The assay de-tects a range from 10 to 108 copies of a plasmid bearing the KHVtarget sequences. (B) KHV DNA copy number per microgram of totalWBC DNA isolated from six koi (designated K1 to K6) that were bledthree times (at 2 weeks, 1 month, and 2 months following arrival atOSU-SDL). (C and D) Autoradiogram of the KHV DNA polymerasegene. Amplicons were amplified with primers KHVDF-242 andKHVDR-242 from six koi on three occasions (2 weeks, 1 month, and2 months following arrival at OSU-SDL) after testing for the presenceof KHV DNA in WBC (C) and plasma (D). The positive control(P) was KHV-U DNA. N, negative control.
FIG. 2. Detection of the KHV mRNA of DNA polymerase geneand major capsid gene from WBC total RNA. (A) RT-PCR of WBCtotal RNA with primers KHVDF-242 and KHVDR-242 (Table 1).(B) RT-PCR of WBC total RNA with primers KHVC382F andKHVC382R specific for a portion of major capsid gene at 382 bp.(C) RT-PCR of 18S RNA using Quantum RNA 18S internal standards(Ambion) with an 18S PCR primer pair (the expected product size is324 bp). Lane 1, KHV-U DNA; lane 2, WBC total DNA of four koilatently infected with KHV; lanes 3 and 4, total RNA of KF-1 cellsinfected with KHV at 8 days postinfection; lanes 5 and 6, WBC totalRNA of four koi latently infected with KHV. Lanes 3 and 5 are withoutreverse transcriptase, and lanes 4 and 6 are with reverse transcriptasein the RT reaction.
VOL. 85, 2011 KHV BECOMES LATENT IN LEUKOCYTES 4957
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/j
vi o
n 26
Dec
embe
r 20
21 b
y 59
.3.7
5.44
.
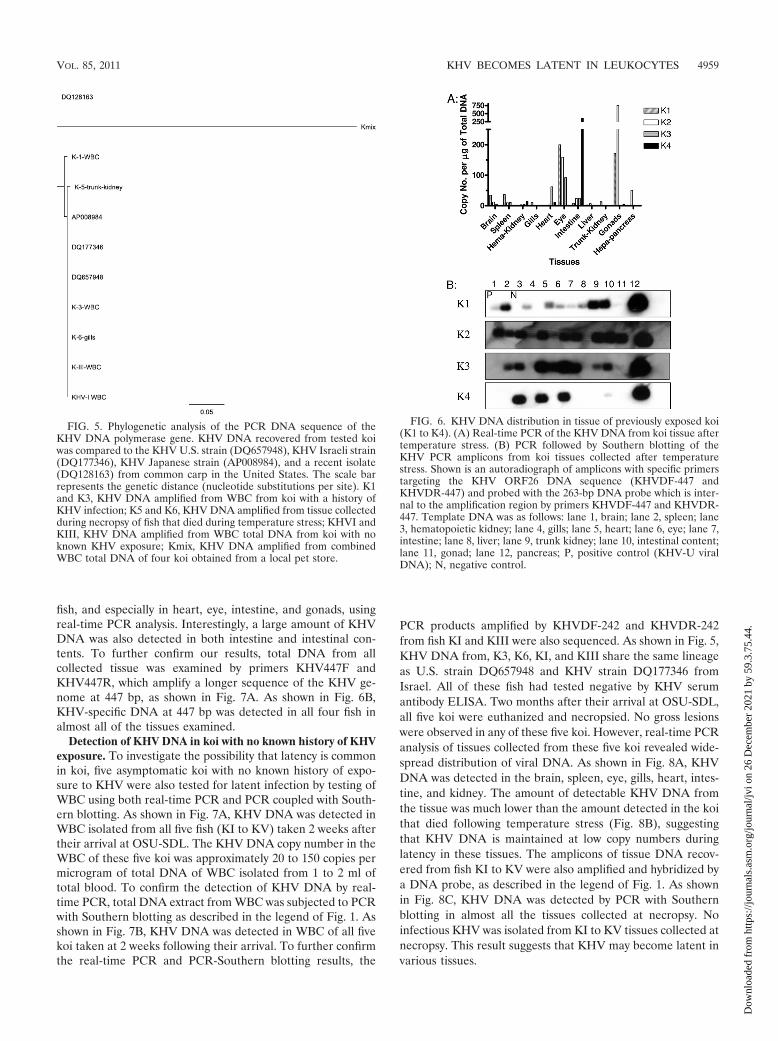
Examination of KHV DNA in tissue from koi that diedduring reactivation study. Two koi, designated K5 and K6,died on day 20 post-temperature stress (i.e., 20°C). No acutegross or microscopic changes characteristic of KHV were ob-served at necropsy. However, a high magnitude of KHV DNA(20- to 200-fold higher than the average amount of KHV DNAcopy numbers of WBC samples taken on three different times)was detected in heart, gill, spleen, and fecal samples (Fig. 4).To see if KHV infectious virions were present in these koi,sample preparations from the gills, spleen, and feces wereinoculated onto CCB cells; KHV CPE was observed in cellsinfected with gill, spleen, and fecal tissue preparations fromboth dead koi (data not shown). To investigate the isolaterelationship with existing KHV isolates, the DNA sequenceamplified by primers KHVDF-242 and KHVDR-242 wascompared with the KHV DNA sequences deposited in theGenBank. The K5 DNA polymerase gene (between nucleo-tides [nt] 147218 and 147459) had one nucleotide at 147298 ofthe KHV-U genome (DQ657948) different from the sequence
of other strains and had a close phylogenetic relationship to aJapanese strain (AP008984), while KHV DNA from K6 was100% identical (between nt 147218 and 147459) to that ofKHV-U (DQ657948), indicating a close relationship with theU.S. strain (DQ657948) (Fig. 5). The phylogenetic relationshipis consistent with the source of the fish. K5 was from a donorwho acquired the koi from Japan; K6 was donated from a U.S.koi hobbyist. In addition, to examine their phylogenetic rela-tionship with our domestic koi, KHV DNA amplified fromWBC pooled from four koi (Kmix) obtained from a local petstore was also compared with KHV from K5 and K6. As shownis Fig. 5, Kmix has a close phylogenetic relationship with KHVisolated from common carp, which suggests that KHV mayhave existed in common carp long before the KHV outbreakin koi.
KHV DNA distribution in tissue of previously exposed koi.To determine whether KHV has specific tissue tropism duringlatency, KHV DNA distribution was examined in tissues col-lected at necropsy. One month following the temperaturestress, the remaining four koi were euthanized and examinedfor KHV latency in tissues. Upon necropsy, three out of fourkoi lacked significant gross and histological pathology; one fish,K4, had prominent gill necrosis at necropsy. Total DNA wasextracted from each individual tissue and examined by bothreal-time PCR and PCR with primers specific for KHV ORF26(KHV447F/KHV447R). As shown in Fig. 6A, a detectablelevel of KHV DNA could be found in many tissues in all four
FIG. 3. Water temperature change and detection of KHV DNA infecal and gill swabs. (A) Tank water temperature change. Vent and gillswabs were collected from all six koi every other day starting at day 2post-temperature increase until day 16 post-temperature increase.(B) Percentage of KHV swabs testing positive by real-time PCR forKHV DNA.
TABLE 2. DNA copy number estimated by CT value of real-timePCR of gill swabs from temperature-stressed koia
Koi no.KHV DNA copy no. at the indicated time post-temperature stress
Day 2 Day 4 Day 6 Day 8 Day 10 Day 12 Day 14
K1 �10 �10 �10 �500K2 �10 �1000 �500 �500 �500K3 �1000 �500 �500 �500 �500K4 �500 �500 �500 �500 �10K5 �10 �10 �500 �1000 �500 �10K6 �10 �500 �500 �500 �1000
a As detected by real-time PCR in 0.5-ml swabs. An empty cell means that thePCR threshold cycle (CT) value was below the limit of detection.
FIG. 4. Examination of KHV DNA in tissue from koi that diedduring the reactivation study. (A) KHV DNA copy number per mi-crogram of total DNA isolated from two dead koi, K5 and K6, thatdied during temperature stress. (B) Comparison of the average KHVDNA copy number per microgram of total DNA between tissues of K5and K6, which died during temperature stress, and tissues of K1 to K4collected after stress. ***, P � 0.001, in comparison with the averageof KHV DNA from K5 and K6.
4958 EIDE ET AL. J. VIROL.
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/j
vi o
n 26
Dec
embe
r 20
21 b
y 59
.3.7
5.44
.
fish, and especially in heart, eye, intestine, and gonads, usingreal-time PCR analysis. Interestingly, a large amount of KHVDNA was also detected in both intestine and intestinal con-tents. To further confirm our results, total DNA from allcollected tissue was examined by primers KHV447F andKHV447R, which amplify a longer sequence of the KHV ge-nome at 447 bp, as shown in Fig. 7A. As shown in Fig. 6B,KHV-specific DNA at 447 bp was detected in all four fish inalmost all of the tissues examined.
Detection of KHV DNA in koi with no known history of KHVexposure. To investigate the possibility that latency is commonin koi, five asymptomatic koi with no known history of expo-sure to KHV were also tested for latent infection by testing ofWBC using both real-time PCR and PCR coupled with South-ern blotting. As shown in Fig. 7A, KHV DNA was detected inWBC isolated from all five fish (KI to KV) taken 2 weeks aftertheir arrival at OSU-SDL. The KHV DNA copy number in theWBC of these five koi was approximately 20 to 150 copies permicrogram of total DNA of WBC isolated from 1 to 2 ml oftotal blood. To confirm the detection of KHV DNA by real-time PCR, total DNA extract from WBC was subjected to PCRwith Southern blotting as described in the legend of Fig. 1. Asshown in Fig. 7B, KHV DNA was detected in WBC of all fivekoi taken at 2 weeks following their arrival. To further confirmthe real-time PCR and PCR-Southern blotting results, the
PCR products amplified by KHVDF-242 and KHVDR-242from fish KI and KIII were also sequenced. As shown in Fig. 5,KHV DNA from, K3, K6, KI, and KIII share the same lineageas U.S. strain DQ657948 and KHV strain DQ177346 fromIsrael. All of these fish had tested negative by KHV serumantibody ELISA. Two months after their arrival at OSU-SDL,all five koi were euthanized and necropsied. No gross lesionswere observed in any of these five koi. However, real-time PCRanalysis of tissues collected from these five koi revealed wide-spread distribution of viral DNA. As shown in Fig. 8A, KHVDNA was detected in the brain, spleen, eye, gills, heart, intes-tine, and kidney. The amount of detectable KHV DNA fromthe tissue was much lower than the amount detected in the koithat died following temperature stress (Fig. 8B), suggestingthat KHV DNA is maintained at low copy numbers duringlatency in these tissues. The amplicons of tissue DNA recov-ered from fish KI to KV were also amplified and hybridized bya DNA probe, as described in the legend of Fig. 1. As shownin Fig. 8C, KHV DNA was detected by PCR with Southernblotting in almost all the tissues collected at necropsy. Noinfectious KHV was isolated from KI to KV tissues collected atnecropsy. This result suggests that KHV may become latent invarious tissues.
FIG. 5. Phylogenetic analysis of the PCR DNA sequence of theKHV DNA polymerase gene. KHV DNA recovered from tested koiwas compared to the KHV U.S. strain (DQ657948), KHV Israeli strain(DQ177346), KHV Japanese strain (AP008984), and a recent isolate(DQ128163) from common carp in the United States. The scale barrepresents the genetic distance (nucleotide substitutions per site). K1and K3, KHV DNA amplified from WBC from koi with a history ofKHV infection; K5 and K6, KHV DNA amplified from tissue collectedduring necropsy of fish that died during temperature stress; KHVI andKIII, KHV DNA amplified from WBC total DNA from koi with noknown KHV exposure; Kmix, KHV DNA amplified from combinedWBC total DNA of four koi obtained from a local pet store.
FIG. 6. KHV DNA distribution in tissue of previously exposed koi(K1 to K4). (A) Real-time PCR of the KHV DNA from koi tissue aftertemperature stress. (B) PCR followed by Southern blotting of theKHV PCR amplicons from koi tissues collected after temperaturestress. Shown is an autoradiograph of amplicons with specific primerstargeting the KHV ORF26 DNA sequence (KHVDF-447 andKHVDR-447) and probed with the 263-bp DNA probe which is inter-nal to the amplification region by primers KHVDF-447 and KHVDR-447. Template DNA was as follows: lane 1, brain; lane 2, spleen; lane3, hematopoietic kidney; lane 4, gills; lane 5, heart; lane 6, eye; lane 7,intestine; lane 8, liver; lane 9, trunk kidney; lane 10, intestinal content;lane 11, gonad; lane 12, pancreas; P, positive control (KHV-U viralDNA); N, negative control.
VOL. 85, 2011 KHV BECOMES LATENT IN LEUKOCYTES 4959
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/j
vi o
n 26
Dec
embe
r 20
21 b
y 59
.3.7
5.44
.

DISCUSSION
Our studies demonstrate that KHV, like mammalian her-pesviruses, can become latent in the peripheral white bloodcells and various tissues. The type of white blood cell latentlyinfected with KHV has yet to be determined. Our preliminarystudies by sorting B cells and T cells demonstrate that B cellsor other non-T cells are the major latency sites of KHV (un-published data). Our results prove that KHV latency can bereactivated by temperature stress between 17°C and 23°C,which may explain the tendency for KHV outbreaks during thesummer. A previous study yielded similar findings, but therelatively brief interval between acute clinical disease and re-activation raised the possibility of persistent infection ratherthan latency (28). In the current study the interval was severalyears, and tests for active infection prior to the temperaturestress treatment were negative, strongly implicating reactiva-tion.
Previous research demonstrated high KHV DNA concen-trations in the gill, kidney, and spleen at the beginning ofinfection and suggested that KHV is lymphoid tropic (8). It ispossible that KHV becomes latent in lymphoid cells because Tcells and B cells are part of the piscine immune system (6,16–18). Since no KHV CPE was observed in CCB cells inoc-ulated with plasma or WBC culture supernatant from KHV-positive fish prior to the temperature stress treatment, KHVDNA, but not infectious virions, was detected, as expected, inperipheral WBC. Our study also suggests that KHV DNAremains at low copy numbers in the latently infected tissues,ranging between 2 to 150 copies per microgram (Fig. 1B, 6A,and 8A.). As shown in Fig. 1, only 30% of the KHV-positivefish were detected by real-time PCR if only 0.5 ml of blood wascollected for WBC isolation. The variation comes from varia-tions in sampling as blood collection from each koi varies
between each time point. However, all fish 50 to 65 cm longtested positive when a minimum of 1 ml of blood was used forWBC isolation (third blood sample from K1 to K6). For fishunder 25 cm long, KHV DNA could be detected in WBC from1 ml of blood (data not shown). However, PCR coupled withSouthern blotting was shown to be a highly sensitive method
FIG. 7. Detection of the KHV PCR amplicons from WBC totalDNA from koi with no known history of KHV exposure. (A) Real-timePCR of KHV DNA from WBC collected at 2 weeks following arrival.The y axis represents the KHV DNA copy number per microgram ofeach koi WBC isolated from 1 ml of blood. (B) Autoradiogram ofKHV PCR amplicons hybridized with KHV DNA probe which isinternal to the PCR primer amplification region shown in Fig. 1. LanesI to V, PCR amplicons of WBC total DNA isolated from KI to KV; P,positive control (KHV-U DNA); N, negative control.
FIG. 8. Detection of the KHV PCR amplicons from tissue totalDNA from koi with no known history of KHV exposure. (A) Real-timePCR of the KHV DNA from koi tissue of KI to KV. x axis, total tissueDNA isolated from 50 to 100 �g of tissue; y axis, KHV DNA copynumber per microgram of total DNA from each tissue. (B) Compar-ison of the average KHV DNA copy number per microgram of totalDNA between tissues of koi K5 and K6, which died during tempera-ture stress, and tissues of KI to KV collected 2 months later. ***, P �0.001, in comparison with the average of KHV DNA from K5 and K6.(C) PCR followed by Southern blotting of the KHV DNA of tissuefrom koi KI to KV. Shown is an autoradiogram of KHV DNA poly-merase gene amplicons, using primers KHVDF-242 and KHVDR-242,hybridized with the KHV DNA probe, which is internal to the PCRprimer amplification region as shown in Fig. 1. The positive control(P) is KHV-U DNA. Template DNA was as follows: lane 1, brain; lane2, spleen; lane 3, hematopoietic kidney; lane 4, gills; lane 5, heart; lane6, eye; lane 7, intestine; lane 8, liver; lane 9, trunk kidney; lane 10,fecal; lane 11, gonad; lane 12, pancreas; P, positive control; N, negativecontrol.
4960 EIDE ET AL. J. VIROL.
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/j
vi o
n 26
Dec
embe
r 20
21 b
y 59
.3.7
5.44
.

for detecting KHV latency in WBC and is more consistent andreliable than the real-time PCR method (Fig. 1C and 7B).
Our studies find that KHV becomes latent in WBC andvarious tissues. Whether the widespread tissue distribution ofKHV DNA reflects circulating latently infected leukocytes oran extremely broad tissue tropism remains uncertain. It ispossible that detection of KHV DNA varies with the numbersof WBC in the tissues during collection at necropsy. It is alsopossible that erythrocytes or other tissue constituents can in-terfere with PCR-based detection of KHV in some tissuessince we noticed that KHV DNA amplification was decreasedin some tissues spiked with positive-control DNA (data notshown). The quality of the viral DNA from each tissue mayalso play an important role in the detection of the virus. If theviral DNA is extensively degraded during necropsy and samplepreparation, it will reduce detection sensitivity. Thus, it is pos-sible that all the tissues contain KHV DNA but that our PCRfailed to detect the viral genome due to degradation of KHVDNA in the degraded tissue.
A large amount of KHV DNA was detected in both intestineand intestinal contents collected at necropsy from some of theclinically normal koi that had recovered from temperaturestress and from the koi with no history of KHV infection (Fig.6A and 8A). This may reflect latency within gastrointestinalepithelial cells that are sloughed into the gut lumen or latencyin lymphocytes undergoing transmigration into the lumen orperhaps simply contamination of the sample with blood due tothe method of “milking” content from the intestinal segmentduring collection.
Koi exposed to KHV do mount an antibody response to thevirus (27). KHV antibody detection by serum neutralization orserum ELISA is the only method used by veterinarians todetect KHV exposure to prevent KHV transmission. However,this assay may miss some fish latently infected with KHV if theantibody titer is below the detection level of the serum neu-tralization or the ELISA method. There are examples of otherherpesvirus infections in which animals have latent infectionsbut do not have detectable antibodies by certain ELISAs (19).Our current study found that the ELISA has very limitedsensitivity in the detection of KHV-positive koi. In addition,although K4 and K6 tested positive prior to our study, theKHV antibody ELISA did not detect antibody in any of the sixkoi sampled at three different times, with the exception of thethird blood sample from K6 (Fig. 9). However, our PCR andSouthern blotting results prove that all six fish were latently in-fected with KHV (Fig. 1C). These latent infections were subse-quently confirmed via reactivation. Furthermore, the detectionof KHV DNA in multiple clinically normal fish with no knownhistory of exposure to a KHV outbreak raises the possibilitythat the distribution of this virus in the koi population is farwider than previously believed (Fig. 2 and 8). Our study sug-gests that the KHV serum ELISA does not always detectKHV-exposed koi and that real-time PCR provides more ac-curacy when WBC are examined directly. Additionally, thePCR coupled with Southern blotting of WBC was shown to bea very sensitive method to detect koi latently infected withKHV. This may allow efficient screening for koi latently in-fected with KHV.
In addition, it is possible that there are many different strainsof KHV present in koi populations. The KHV DNA from fish
at a local pet store evolved from the U.S. strain and is closelyrelated to common carp KHV (Fig. 5). This suggests that KHVmay have originated from a common carp KHV that has yet tobe well characterized.
ACKNOWLEDGMENTS
We thank the American Koi Club Association for funding this study.We thank Ronald Hedrick (University of California, Davis) for
providing the KF-1 cell line and the KHV-I and KHV-U strains usedin this study. We also thank George Rohrmann for helping to edit thelast two versions of the paper.
REFERENCES
1. Aoki, T., et al. 2007. Genome sequences of three koi herpesvirus isolatesrepresenting the expanding distribution of an emerging disease threateningkoi and common carp worldwide. J. Virol. 81:5058–5065.
2. Bondad-Reantaso, M. G., A. Sunarto, and R. P. Subasinghe. 2007. Managingthe koi herpesvirus disease outbreak in Indonesia and the lessons learned.Dev. Biol. (Basel) 129:21–28.
3. Compston, L. I., et al. 2009. Prevalence of persistent and latent viruses inuntreated patients infected with HIV-1 from Ghana, West Africa. J. Med.Virol. 81:1860–1868.
4. Dishon, A., M. Davidovich, M. Ilouze, and M. Kotler. 2007. Persistence ofcyprinid herpesvirus 3 in infected cultured carp cells. J. Virol. 81:4828–4836.
5. Dishon, A., et al. 2005. Detection of carp interstitial nephritis and gillnecrosis virus in fish droppings. Appl. Environ. Microbiol. 71:7285–7291.
6. Flajnik, M. F., and M. Kasahara. 2010. Origin and evolution of the adaptiveimmune system: genetic events and selective pressures. Nat. Rev. Genet.11:47–59.
7. Gilad, O., et al. 2002. Initial characteristics of koi herpesvirus and develop-ment of a polymerase chain reaction assay to detect the virus in koi, Cyprinuscarpio koi. Dis. Aquat. Organ. 48:101–108.
8. Gilad, O., et al. 2004. Concentrations of a koi herpesvirus (KHV) in tissuesof experimentally infected Cyprinus carpio koi as assessed by real-timeTaqMan PCR. Dis. Aquat. Organ. 60:179–187.
9. Gray, W. L., R. J. Williams, R. L. Jordan, and B. R. Griffin. 1999. Detectionof channel catfish virus DNA in latently infected catfish. J. Gen. Virol.80:1817–1822.
10. Grimmett, S. G., J. V. Warg, R. G. Getchell, D. J. Johnson, and P. R. Bowser.2006. An unusual koi herpesvirus associated with a mortality event of com-mon carp Cyprinus carpio in New York State, U.S.A. J. Wildl. Dis. 42:658–662.
11. Iida, T., and M. Sano. 2005. Koi herpesvirus disease. Uirusu 55:145–151. [InJapanese.]
12. Ishioka, T., et al. 2005. Detection and sequence analysis of DNA polymeraseand major envelope protein genes in koi herpesviruses derived from Cyprinuscarpio in Gunma prefecture, Japan. Vet. Microbiol. 110:27–33.
13. Jin, L., et al. 2008. Characterization of a novel alphaherpesvirus associatedwith fatal infections of domestic rabbits. Virology 378:13–20.
14. Jin, L., W. M. Schnitzlein, and G. Scherba. 2000. Identification of thepseudorabies virus promoter required for latency-associated transcript geneexpression in the natural host. J. Virol. 74:6333–6338.
15. Jones, C., and S. Chowdhury. 2007. A review of the biology of bovine
FIG. 9. KHV antibody ELISA. Results of serum collected at threeoccasions (2 weeks, 1 month, and 2 months following arrival at OSU-SDL). An optical density (OD) below 0.24 is considered negative. Thesix koi, K1 to K6, were from populations with a history of KHVinfection. The asterisk indicates that the koi blood is positive for theKHV antibody. �, positive control; �, negative control.
VOL. 85, 2011 KHV BECOMES LATENT IN LEUKOCYTES 4961
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/j
vi o
n 26
Dec
embe
r 20
21 b
y 59
.3.7
5.44
.

herpesvirus type 1 (BHV-1), its role as a cofactor in the bovine respiratorydisease complex and development of improved vaccines. Anim. Health Res.Rev. 8:187–205.
16. Langenau, D. M., and L. I. Zon. 2005. The zebrafish: a new model of T-celland thymic development. Nat. Rev. Immunol. 5:307–317.
17. Lorenzen, N., and S. E. LaPatra. 2005. DNA vaccines for aquacultured fish.Rev. Sci. Tech. 24:201–213.
18. Marchalonis, J. J., S. F. Schluter, R. M. Bernstein, and V. S. Hohman. 1998.Antibodies of sharks: revolution and evolution. Immunol. Rev. 166:103–122.
19. McCaw, M. B., F. A. Osorio, J. Wheeler, J. Xu, and G. A. Erickson. 1997.Effect of maternally acquired Aujeszky’s disease (pseudorabies) virus-spe-cific antibody in pigs on establishment of latency and seroconversion todifferential glycoproteins after low dose challenge. Vet. Microbiol. 55:91–98.
20. Minarovits, J., E. Gonczol, and T. Valyi-Nagy. 2007. Latency strategies ofherpesviruses. Springer, New York, NY.
21. Perelberg, A., A. Ronen, M. Hutoran, Y. Smith, and M. Kotler. 2005. Pro-tection of cultured Cyprinus carpio against a lethal viral disease by an atten-uated virus vaccine. Vaccine 23:3396–3403.
22. Pikarsky, E., et al. 2004. Pathogenesis of acute viral disease induced in fishby carp interstitial nephritis and gill necrosis virus. J. Virol. 78:9544–9551.
23. Pollock, J. L., R. M. Presti, S. Paetzold, and H. W. Virgin IV. 1997. Latentmurine cytomegalovirus infection in macrophages. Virology 227:168–179.
24. Rakus, K. L., et al. 2009. Resistance of common carp (Cyprinus carpio L.) tocyprinid herpesvirus-3 is influenced by major histocompatibility (MH) classII B gene polymorphism. Fish Shellfish Immunol. 26:737–743.
25. Roizman, B., and P. E. Pellett. 2001. The family Herpesviridae: a brief intro-
duction, p. 2381–2397. In D. M. Knipe et al. (ed.), Fields virology, 4th ed.Lippincott Williams & Wilkins, Philadelphia, PA.
26. Sawtell, N. M., D. K. Poon, C. S. Tansky, and R. L. Thompson. 1998. Thelatent herpes simplex virus type 1 genome copy number in individual neuronsis virus strain specific and correlates with reactivation. J. Virol. 72:5343–5350.
27. St-Hilaire, S., N. Beevers, C. Joiner, R. P. Hedrick, and K. Way. 2009.Antibody response of two populations of common carp, Cyprinus carpio L.,exposed to koi herpesvirus. J. Fish Dis. 32:311–320.
28. St-Hilaire, S., et al. 2005. Reactivation of koi herpesvirus infections incommon carp Cyprinus carpio. Dis. Aquat. Organ. 67:15–23.
29. Stingley, R. L., B. R. Griffin, and W. L. Gray. 2003. Channel catfish virusgene expression in experimentally infected channel catfish, Ictalurus punc-tatus (Rafinesque). J. Fish Dis. 26:487–493.
30. Uchii, K., K. Matsui, T. Iida, and Z. Kawabata. 2009. Distribution of theintroduced cyprinid herpesvirus 3 in a wild population of common carp,Cyprinus carpio L. J. Fish Dis. 32:857–864.
31. Waltzek, T. B., et al. 2005. Koi herpesvirus represents a third cyprinidherpesvirus (CyHV-3) in the family Herpesviridae. J. Gen. Virol. 86:1659–1667.
32. Wang, K., T. Y. Lau, M. Morales, E. K. Mont, and S. E. Straus. 2005.Laser-capture microdissection: refining estimates of the quantity and distri-bution of latent herpes simplex virus 1 and varicella-zoster virus DNA inhuman trigeminal ganglia at the single-cell level. J. Virol. 79:14079–14087.
33. Whetstone, C. A., and J. M. Miller. 1989. Two different strains of an alpha-herpesvirus can establish latency in the same tissue of the host animal:evidence from bovine herpesvirus 1. Arch. Virol. 107:27–34.
4962 EIDE ET AL. J. VIROL.
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/j
vi o
n 26
Dec
embe
r 20
21 b
y 59
.3.7
5.44
.