ionic and non-ionic bonds in staining, with special reference to the … · 2006-05-25 ·...
TRANSCRIPT

477
Ionic and non-ionic bonds in staining, with special referenceto the action of urea and sodium chloride on the staining of
elastic fibres and glycogen
By D. J. GOLDSTEIN(From the Medical School, University of the Witwatersrand, Hospital St.,
Johannesburg, S. Africa)
With 2 plates (figs, i and 2)
Summary1. A powerful hydrogen bonding agent such as urea may affect staining in severalways:
(a) by competing for hydrogen bonding sites in the tissue or on the dye particle,it may inhibit staining in which the dye-substrate link is a hydrogen bond;
(6) because it is a dipole, urea may have some affinity for charged sites in the tissue,and thus tend to reduce electrostatic staining;
(c) urea may in some cases decrease staining by increasing the solubility of the dye;(d) by inhibiting association of dye particles in solution, where this is due to hydro-
gen bonding, urea may sometimes facilitate penetration and staining of dense sub-strates.
2. Sodium chloride may inhibit staining due to electrostatic forces by ionic com-petition, but may also decrease staining in some cases by aggregating and precipitat-ing the dye.
3. Prolonged exposure to hot acidic methanol may not only block acidic groups,but also block or destroy non-acidic groups, including the 1,2-glycol group.
4. Urea inhibits the staining of all substrates by Best's carmine. The staining ofglycogen is inhibited also by methylation and by periodic acid, but not by deaminationor sodium chloride. Staining of mucin and nuclei is inhibited by methylation anddeamination, while that of mucin is also inhibited by periodic acid and sodium chloride.It is suggested that the staining of glycogen and mucin by Best's carmine is by hydrogenbonding on to 1,2-glycol groups, the attachment requiring in the case of mucinstabilization by an ionic linkage with some strongly basic group. The staining ofnuclei may also be by hydrogen bonding, on to an unknown group.
5. The staining of elastic fibres by aldehyde-fuchsin, resorcin-fuchsin, orcein,chlorazol black E, and luxol fast blue is inhibited by urea. This is consistent with therole of hydrogen bonding in elastic staining suggested by previous workers.
6. The staining of mast-cell granules and cartilage capsules by aldehyde-fuchsin isinhibited by salt, while that of elastic fibres and interstitial matrix is inhibited by urea.It is suggested that aldehyde-fuchsin contains:
(a) large dye cations which selectively stain negatively charged substrates of highpermeability, such as mast-cell granules and cartilage capsules, and
(6) smaller dye particles which stain elastic fibres and the dense interstitial car-tilage matrix, by hydrogen bonding.
7. Sodium chloride inhibits the staining of mast-cell granules and cartilage matrixby orcein, but not that of nuclei or elastic fibres. In the presence of urea, dilute orceinstains nuclei selectively, apparently by electrostatic attraction reinforced by a linkageof unknown nature.
8. Urea and phthalate both decrease the staining of mucin by a number of basic
[Quart. J. micr. Sci., Vol. 103, pt. 4, pp. 477-92, 1962.]

478 Goldstein—Ionic and non-ionic bonds in staining
dyes, but increase the staining of certain substrates, especially by alcian blue. It issuggested that both substances inhibit staining in which hydrogen bonding is impor-tant, and increase the staining of dense substrates by large-particled dyes by inhibitingaggregation of the dye particles and thus facilitating penetration.
IntroductionSALT (ionic) linkages play an important role in staining (Singer, 1952;Baker, 1958), but are not the only forces involved in the attachment of dye tosubstrate. This is well shown in textile dyeing, where, for example, the woolfibre has a far higher affinity for acid dyes than for inorganic chloride. 'Thisgreater affinity cannot arise in the ionic link itself but must arise from othertypes of binding between the dye molecule and the fibre such as the inter-action of dipoles, hydrogen bonding, and Van der Waals forces, which do notexist in the case of the simple inorganic anions. These forces will in generalincrease with increasing molecular weight and decreasing solubility of thedye . . .' (Vickerstaff, 1954). Similar factors may well apply also in histo-logical staining, and non-ionic bonds may be of primary importance in someprocedures, and contributing factors in others.
Particularly in the staining of elastic fibres, it has been suggested thathydrogen bonds may be of importance. Thus the staining of elastic fibres byorcein and by certain other dyes is relatively insensitive to pH, is enhanced byalcohol, and antagonized by polar groups (Dempsey and others, 1952; Engleand Dempsey, 1954; Fullmer and Lillie, 1956, 1957). These properties sug-gest a non-ionic linkage, and the heat of reaction is consistent with hydrogenbonding (Weiss, 1954). The staining of elastic fibres by Weigert's resorcin-fuchsin method also appears to involve a non-ionic link, possibly a hydrogenbond (Puchtler and others, 1961), although the dye is able to attach to acidicsubstrates by a polar linkage.
In view of these facts, it seemed of interest to investigate the role of hydro-gen bonding in histological procedures, including the staining of elastic fibres.
The addition of urea to a stain solution should inhibit any staining that isdependent on hydrogen bonding, since urea is a powerful hydrogen bondingagent (Haurowitz, 1950), and would presumably compete for hydrogen bond-ing sites on both the dye molecule and the tissue. Similarly, high concentra-tions of sodium chloride might be expected to inhibit staining due to elec-trostatic linkages (Landsmeer, 1951; Singer, 1952; Vickerstaff, 1954; Wolman,1961; Radden, 1961). The present paper reports the effect of urea and sodiumchloride on the action of several elastic stains, Best's carmine (an empiricalmethod for glycogen, of unknown mechanism), and a number of basic dyes.Some of the chief results are illustrated in figs. 1 and 2.
The tables accompanying this paper are printed as an Appendix (p. 490).
Material and methodsThe stomach, duodenum, colon, trachea, and pancreas, of the rat, and the
oesophagus, stomach, small intestine, large intestine, kidney, and skin of

Goldstein—Ionic and non-ionic bonds in staining 479
Xenopus were fixed in 10% formalin and cut in paraffin at 6 ju,. Sections werestained by the following methods:
1. 1 h in aldehyde-fuchsin (Gomori, 1950), ripened for 2 to 5 days.2. 24 h in aldehyde-fuchsin diluted 1:100 with 1% cone. HC1 in 70%
alcohol.3. 30 min in Weigert's resorcin-fuchsin (Gatenby and Painter, 1937).4. 30 min in 1% orcein (Griibler) in 1% cone. HC1 in 70% alcohol.5. 24 hr in o-i% orcein in acid alcohol.6. 15 min in 1% chlorazol black E (G. T. Gurr) in water (Cannon, 1937;
Levine and Morrill, 1941; Salthouse, 1944).7. 30 min in o-i% luxol fast blue (G. T. Gurr) in 70% alcohol (Harms,
IQ57);8. 30 min in Best's carmine (Gatenby and Painter, 1937). Sections were
dehydrated 10 sec in 95% alcohol and 15 sec in absolute alcohol, andmounted in DPX (British Drug Houses).
9. 1 hr in o-i% alcian blue 8GN150 (I.C.I.) in 3% acetic acid (Steedman,1950; Mowry, 1956).
10. 1 hr in o-i% azure A (Nat. Aniline) in 30% alcohol (Kramer andWindrum, 1955).
11. 1 hr in o-oi M (approx. 0-5%) or in o-i% Victoria blue 4R, Bismarckbrown R or Janus green B (all Nat. Aniline dyes) in veronal-acetatebuffer (Hale, 1958) at pH 4-0 to 4-2.
Sections were also stained in the above dye solutions saturated with ureaor containing 10% of NaCl (alcoholic solutions were saturated with NaCl).In all cases stain solutions were filtered before use. Unless otherwise stated,after staining sections were dehydrated in 95% and absolute alcohol (30 secin each), and mounted in DPX.
Additional procedures sometimes performed on sections before stainingincluded:
1. Methylation. 24 h in 1% cone. HC1 in methanol at 56° C (Wiggles-worth, 1952).
2. Deamination. 24 h at room temperature in a solution consisting of 6 gNaNO2) 5 ml glacial acetic acid, and 35 ml water (Monne and Slautter-back, 1950, quoted by Casselman, 1959).
3. Ribonuclease digestion. 1 h at 370 C in 1 mg crystalline ribonuclease(L. Light & Co.) in 1 ml distilled water.
4. Saliva digestion. 30 min at room temperature in two changes of saliva.5. Oxidation in 1% aq. periodic acid for 5 min or 1 h.Phase-contrast microscopy of elastic fibres was carried out by methods
previously described (Goldstein, 1962).
ResultsThe effects of urea and of sodium chloride on staining are summarized in
tables 1 and 3, and illustrated in figs. 1 and 2. In some cases (e.g. orcein and

480 Goldstein—Ionic and non-ionic bonds in staining
aldehyde-fuchsin) a marked effect was only obtained by decreasing the con-centration of the stain, and thus increasing the proportion of urea or salt. Therelatively firm attachment of orcein and aldehyde-fuchsin may explain whyFullmer and Lillie (1956, 1958) were unable to remove these dyes fromsections by differentiation in 6 M urea.
Sodium chloride caused precipitation of some dyes (table 4); staining of allsubstrates was greatly diminished, or abolished, by this salt in these cases.
The results of other procedures are given under the heading of the par-ticular stain tested.
Best's carmine
Methylation abolished subsequent staining of glycogen, and greatlydiminished that of mucin, pancreatic cytoplasmic chromidial substance, andnuclei, but markedly increased the staining of collagen. Deamination markedlyreduced the staining of mucin, nuclei, and chromidial substance, but did notaffect that of glycogen.
Ribonuclease slightly reduced the staining of pancreatic cytoplasmicchromidial substance by Best's carmine, but completely abolished staining byazure A in a control section.
Treatment with periodic acid had no effect on subsequent staining of nucleior cytoplasmic chromidial substance, but reduced or abolished the stainingof glycogen and mucin by Best's carmine (table 2). This effect was not due toremoval of the substrate from the section, since even after an hour in periodicacid the staining of glycogen and mucin by Schiff's reagent was not diminished(table 2).
Orcein
As noted also by Dempsey and others (1952), orcein staining is extra-ordinarily resistant to acid or alkaline differentiation. Six hours' treatment ineither 1 N HC1 or in an alcoholic solution of lithium carbonate had practicallyno effect on the staining of nuclei, cartilage matrix, or elastic fibres.
Dilute (o-1 %) orcein in acid alcohol saturated with urea gave after 24 h anextremely sharp and selective staining of nuclei, with negligible staining ofother tissue components (figs. 1, K; 2,1). This nuclear staining was diminishedor abolished by prior methylation. Deamination increased nuclear staining,and caused very slight staining of cartilage, pancreatic cytoplasmic chromidial
FIG. I (plate). All the photomicrographs represent the trachea of the rat, fixed with forma-lin. Each division of the scale in A represents io>i. This scale applies to all the photomicro-graphs.
A, B, c, stained with Weigert's resorcin-fuchsin, photographed with a yellow filter.D, E, F, stained with Gomori's aldehyde-fuchsin, photographed with a green filter.G, H, 1, stained for 30 min in 1 % orcein, photographed with a green filter.J, K, L, stained for 24 h in o-i% orcein, photographed with a green filter.B, E, H, and K are photographs of slides stained in solutions saturated with urea, while C, F, I,
and L are of slides in solutions saturated with NaCl. Note the effect of sodium chloride on thestaining of cartilage matrix, and the effect of urea on the staining of elastic fibres.
The photographs in each row were exposed and printed under identical conditions.

FIG. r
D. J. GOLDSTEIN
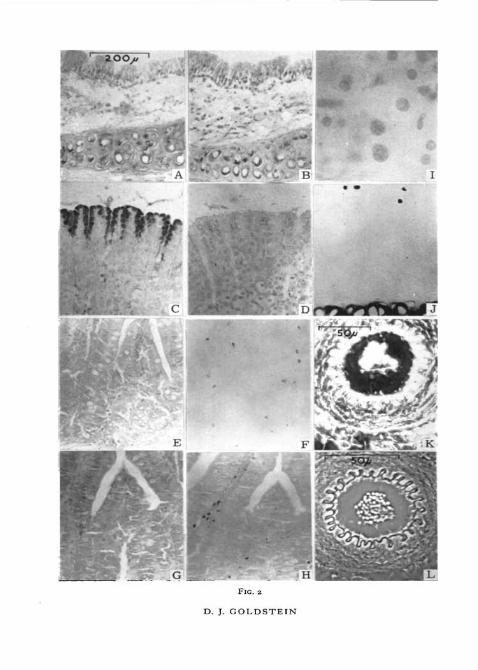
FIG. 2
D. J. GOLDSTEIN

Goldstein—Ionic and non-ionic bonds in staining 481
substance, and the cytoplasm of various epithelia, but had no effect on thestaining of elastic fibres. Oxidation for 5 min in 1% aq. periodic acid had nodetectable effect on subsequent staining in orcein-urea.
Very dilute (o-oi%) orcein in acid alcohol saturated with urea stainednuclei selectively but very weakly after 24 h. In the absence of urea, o-oi%orcein did not stain nuclei as strongly as did o-i% orcein in the presence ofurea, but stained cartilage matrix and elastic fibres slightly while the latterdid not. This indicates that the inhibitory effect of urea on orcein staining ismore specific than that seen with the use of very dilute solutions of the dyewithout urea.
Chlorazol black E
Solutions of the dye made up in water and in 70% alcohol behaved similarly.In addition to elastic fibres, chlorazol black E stained nuclei, some mast-cellgranules, the mucin of goblet cells in the small intestine of the rat, and thesurface epithelial mucin of rat stomach. The granules of eosinophil leucocytestended to stain reddish, apparently staining both with chlorazol black E itselfand with a red acid dye present as a contaminant. In the presence of urea,eosinophil granules stained black, and appeared in improved contrast owingto the decreased staining of other tissue components (fig. 2, G, H). The stainingof eosinophil granules and other structures was decreased or abolished indeaminated sections. Methylation caused a great increase in the intensity ofstaining; in such sections elastic fibres were no longer distinguishable from thesurrounding tissues.
Chlorazol black E appeared to be scarcely soluble in 1 % HC1, only the redcontaminant dissolving. This dye, in acid solution, behaved rather like eosin,staining red blood-corpuscles, cytoplasm, and collagen fibres pink. Dissolved
FIG. 2 (plate). The scale on A applies to all the photomicrographs except K and L. An orangefilter was used for E and F, a green filter for the remainder.
A, rat trachea, stained with Best's carmine. Note the staining of glycogen in the lacunaeof the cartilage.
B, rat trachea, stained with Best's carmine containing 10% NaCl. Glycogen is still stained.c, rat stomach, stained with Best's carmine. Mucin of the superficial epithelium is well
stained.D, rat stomach, stained with Best's carmine containing 10% NaCl. Staining of mucin is
abolished.E, rat small intestine, stained with o-i% luxol fast blue in 70% alcohol. Control for F.F, rat small intestine, stained with 0 1 % luxol fast blue in 70% alcohol saturated with urea.
All staining is abolished, except that of eosinophil granules.C, rat small intestine, stained with 0 1 % aq. chlorazol black E. Control for H.H, rat small intestine, stained with 0 1 % chlorazol black E, saturated with urea. Staining of
all substrates except eosinophil granules is greatly decreased.1, nuclei of rat pancreas stained with 0 1 % orcein in acid alcohol saturated with urea.
Nothing else is stained.J, cartilage and mast-cell granules stained with dilute aldehyde-fuchsin saturated with
reaurea.rea.K, unstained rat artery, mounted in glycerol jelly and examined with negative phase contrast.
Elastic fibres appear bright, indicating that they are of higher R.I. than the medium.L, rat artery in DPX, negative phase contrast. Elastic fibres appear dark, indicating that
they are of lower R.I. than the medium.

482 Goldstein—Ionic and non-ionic bonds in staining
in 0-5% NaOH, chlorazol black E stained cytoplasm and elastic fibres withincreased intensity, but the staining of mucin was somewhat reduced.
Cannon (1937) reported that chlorazol blackE is an excellent stain for chitin'. . . and probably for glycogen which stains pink or red'. In the present studysaliva-treated and control sections stained similarly, and staining of glycogenhas not been observed.
DiscussionAction of sodium chloride on staining
Competition of cations for anionic sites in the tissues probably explainsthe inhibitory action of sodium chloride on the staining by orcein, resorcin-fuchsin, and aldehyde-fuchsin of basiphil substrates such as cartilage matrixand mast-cell granules. Similarly, the inhibition by sodium chloride of thestaining of mucin by Best's carmine probably indicates that this staining in-volves a salt linkage, in this case probably that of a negatively charged dye toan acidophil (basic) substrate (Goldstein, 1962).
Sodium chloride causes many dyes to aggregate (Ostwald, 1933; Valko,1946). This may radically lower the concentration of effective dye particles,even if it does not cause macroscopic precipitation (table 4). Thus the in-hibition by sodium chloride of staining by chlorazol black E, luxol fast blue,alcian blue, Victoria blue 4R, Bismarck brown R, Janus green B, and azureA does not prove conclusively that salt linkages play any role in staining withthese dyes, even though this seems probable on other grounds.
Action of urea on staining
Owing to its hydrogen bonding tendencies, urea could inhibit staining dueto hydrogen bonding by competing with the dye for hydrogen bonding sitesin the tissue, and/or by competing with the tissue for hydrogen bonding groupson the dye molecule. Urea is strongly dipolar, and might also compete withdye ions for charged sites in the substrates. Ion-dipole links are, however, ingeneral weaker than ion-ion links, so that the inhibition by urea of salt linkagesis likely to be less effective than that of hydrogen bonding.
Even where the dye-substrate link is neither a hydrogen bond nor a saltlinkage, the affinity of urea for a dye molecule possessing hydrogen bondinggroups might decrease staining by increasing the solubility of the dye (as itdoes that of proteins: Haurowitz, 1950). Inhibition of staining by urea maytherefore suggest that hydrogen bonding is involved, but cannot, in theabsence of supporting evidence, be regarded as conclusive proof.
In addition to the above actions, urea may affect staining by inhibitingassociation or aggregation of dye molecules in solution, where this is due tohydrogen bonding. This point is discussed more fully below.
Mechanism of Best's carmine
The action of urea in inhibiting staining by Best's carmine suggests thathydrogen bonds may play a role in this method, while the inhibition of mucin

Goldstein—Ionic and non-ionic bonds in staining 483
staining by sodium chloride (table 1; fig. 2, c, D) indicates that in this site,salt linkages are probably involved as well. The solubility of the dye is onlymoderately reduced by the addition of sodium chloride (table 4), and thestaining of glycogen and nuclei is unaffected, so that this salt probably com-petes with dye ions for charged sites in mucin, and does not reduce stainingsimply by causing aggregation and precipitation of the dye. The decrease inthe staining of mucin caused by deamination is consistent with the attachmentof an acid dye to a basic (acidophil) substrate, although it is not conclusiveproof of this.
From the fact that the staining of glycogen by Best's carmine is unaffectedby deamination or the presence of sodium chloride, it appears that saltlinkages are unimportant in the staining of this substrate, and hydrogen bond-ing may here be the main or only factor.
Since periodic acid abolishes the affinity of Best's carmine for glycogen andmucin (table 2), attachment of the dye to these sites is probably dependent on1,2-glycol groups. This is supported by the fact that chromic acid oxidationinduces metachromasia in mucin at the expense of affinity for Best's carmine(Burkl, 1951), since this procedure may form acidic groups from glycols(Lillie, 1951). The postulated dye-substrate hydrogen bond is presumablybetween the —OH of the glycol and a strongly electro-negative group, suchas a carbonyl, on the dye.
The decrease caused by methylation in the staining of both glycogen andmucin by Best's carmine is probably not due to the blocking of acidic groups,since there is no evidence for the presence of positively charged dye particlesin the dye, or for the presence of acidic groups in glycogen. Prolonged methy-lation reduces or abolishes the staining of mucin also by the periodic acid /Schiff (PAS) method (Fisher and Lillie, 1954); so it seems likely that theprocedure can destroy or block glycol groups, or perhaps even remove mucinfrom the section completely. Fairly mild acid hydrolysis removes glycogenfrom sections (Goldstein, 1959), and prolonged methylation may perhapsalso do so.
The increase in the staining of collagen fibres after methylation is prob-ably due to increased acidophilia, resulting from the blocking of acidic groups.
The failure of ribonuclease to abolish the staining of pancreatic cytoplasmicchromidial substance by Best's carmine indicates that the dye probablyattaches to protein in this site rather than to ribonucleic acid (RNA). Themoderate reduction in staining after treatment with the enzyme may be dueto removal of some protein together with the RNA, or to blockage of reactivesites in the tissue by the protein enzyme (French and Benditt, 1953; Morrisand Krikos, 1958; Fullmer 1959; Stolk, 1961).
The staining of nuclei and of cytoplasmic chromidial substance by Best'scarmine appears not to involve a salt linkage, since it is unaffected by sodiumchloride. The decrease in staining after deamination may be due to repulsionof the negatively charged dye by the increased negative charge on the tissue,and does not prove that the dye-substrate link itself is ionic. It is possible that

484 Goldstein—Ionic and non-ionic bonds in staining
the dye attaches by a hydrogen bond (or bonds) to some group destroyed byprolonged methylation. This group is unlikely to be the 1,2-glycol group,since this is not known to occur in nuclei or in cytoplasmic chromidial sub-stance, and the affinity of these sites for Best's carmine is unaffected byperiodic acid (table 2).
To recapitulate, Best's carmine probably stains both glycogen and mucinby hydrogen bonding with 1,2-glycol groups, the attachment requiring inthe latter stabilization by an ionic linkage with some strongly basic group.This basic group may be arginine or lysine (Goldstein, 1962). The stainingby Best's carmine of nuclei and of the protein component of cytoplasmicchromidial substance may be by hydrogen bonding, on to an unknown groupor groups.
The mechanism of elastic staining
Except where it causes precipitation of the dye, as in the case of chlorazolblack E and luxol fast blue, sodium chloride has little or no effect on thestaining of elastic fibres. Urea, on the other hand, markedly decreases orabolishes elastic staining by orcein, resorcin-fuchsin, aldehyde-fuchsin,chlorazol black E, and luxol fast blue. This evidence is consistent with pre-vious suggestions (Dempsey and others, 1952; Engle and Dempsey, 1954;Weiss, 1954; Puchtler and others, 1961) that elastic staining depends on anon-ionic link, probably a hydrogen bond. The less-marked inhibition of thestaining of certain other substrates caused by urea might be due to an increasein the solubility of the dye, or to the dipolar urea competing with dye ionsfor charged sites in the tissue (discussed above).
Moderate methylation (4 h at 6o° C) blocks the staining of mucin by alde-hyde-fuchsin, with little or no effect on the staining of elastic fibres (Spicerand Meyer, 1960), but more prolonged methylation inhibits the staining ofelastic fibres by resorcin-fuchsin, aldehyde-fuchsin, orcein, and other stains(Fullmer and Lillie, 1957). Fullmer and Lillie concluded that the stainsstudied attach to acidic groups in elastic fibres, but it seems possible that here,as with glycogen and mucin, prolonged methylation blocks some hydrogenbonding group. The weak staining of elastic fibres by PAS indicates a relativescarcity of 1,2-glycol groups, but does not exclude the presence of othergroups which might be capable of hydrogen bonding.
The nature of the reactive groups on the dye molecule and in the elasticfibre is quite uncertain, but it is interesting to note that the introduction ofaldehyde groups into basement membranes enhances staining with resorcin-fuchsin (Puchtler and others, 1961), while treatment of elastic fibres witha-naphthol blocks staining with orcein (Dempsey and others, 1952). Further,treatment with nitrous acid, or acetylation, destroys the affinity of orcein forelastic fibres, and this suggests that hydroxyl or amine groups on the dytmolecule are necessary for staining (Fullmer and Lillie, 1956). If hydrogenbonding is indeed involved in elastic staining, it would appear that thehydrogen donor is the molecule of elastic stain, while the electro-negative

Goldstein—Ionic and non-ionic bonds in staining 485
hydrogen acceptor is in the substrate. This is the reverse of the probablesituation with Best's carmine (see discussion above), and may explain whyelastic stains do not stain glycogen, and why Best's carmine does not stainelastic fibres.
Seki (1933, I933#, 1934) attributed certain of the staining properties ofelastic fibres to their supposedly extremely high density. Seki's belief in thehigh density of elastic fibres was probably based on their refractility in sections(compare Fischer's (1899) correlation of the density and refractility of proteingranules), but in fact the R.I. of elastic fibres in sections lies between about1-47 and about 1-54 (fig. 2, K, L, and Goldstein, 1962), so that the refractilityof elastic fibres mounted in DPX or balsam is due to their being of lower R.I.than the medium. Further, elastic fibres have a low electron density, and theR.I. of elastic fibres is only about 1-534 e v e n when dried (Dempsey andLansing, 1954). One must conclude that elastic fibres are of only moderatedensity, and can probably be penetrated and stained by dyes of fairly largeparticle size (Goldstein, 1962). It is conceivable, however, that some colloidaldyes might be unable to penetrate elastic fibres, and this might be a contribu-tory factor in the failure of Best's carmine to stain them.
The R.I., and therefore probably also the density of collagen fibres, ishigher than that of elastic fibres. This correlates well with the fact that collagenfibres may be converted into an elastin-like material by the removal of certainamino-acids, particularly hydroxyproline (Hall, 1959). It is conceivable thatcertain anomalous staining reactions of elastic fibres, 'elastotically degenerated'collagen fibres, and other connective-tissue fibres (Gillman and others, 1954;Fullmer and Lillie, 1958; Hall, 1959) may be due to the inability of dyes oflarge particle size to penetrate and stain fibres of unusually high density.
Complex action of certain elastin stains
It seems likely that some dyes are capable of attaching to substrates bymore than one type of chemical bond.
The insensitivity to pH of the staining of elastic fibres by chlorazol black Eis consistent with a non-ionic linkage, but the somewhat reduced staining ofmucin at high pH suggests that here a salt linkage may also play a part. Thestaining of eosinophil granules by chlorazol black E and luxol fast blue in thepresence of urea (table 1) may also be due to electrostatic forces.
In the case of aldehyde-fuchsin, it is noteworthy that the substrates whichare still stained in the presence of saturated urea are those possessing strongacidic groups, particularly cartilage matrix and mast-cell granules. Aldehyde-fuchsin is probably markedly heterogeneous (Bangle, 1954, 1956), and maycontain positively charged particles carrying no hydrogen bonding groups.Staining by such particles would presumably be unaffected by urea, and thismight account for the staining of strongly acidic substrates in the presenceof urea. In contrast with the perilacunar cartilage capsules, the dense inter-stitial cartilage matrix tends not to stain with aldehyde-fuchsin in the presenceof urea, so that here hydrogen bonds may conceivably play a part in staining.

486 Goldstein—Ionic and non-ionic bonds in staining
It will be remembered that in the staining of acidic substrates aldehyde-fuchsin acts as a large-particled dye, and is apparently unable to penetrateinto and stain dense substances (Goldstein, 1962). The anomalous staining ofthe dense interstitial cartilage matrix would be explicable if the dye could beshown to contain two types of active particle: a fairly small particle capableof staining elastic fibres and interstitial cartilage matrix by hydrogen bonding,and a larger, positively charged particle capable of selectively staining basi-phil substrates of low density (high permeability).
The presence of dye cations in aldehyde-fuchsin and in orcein also explainswhy the affinity of collagen for these dyes is increased by nitrous acid de-amination (Fullmer and Lillie, 1957). This procedure increases basiphilia,but would have little effect on the affinity for such dyes as Weigert's resorcin-fuchsin, which is only slightly ionic in nature.
The selective staining of nuclei by dilute orcein in the presence of urea isunlikely to be simply an ionic staining of an acidic substrate by a basic dye,since staining of the more strongly acidic cartilage matrix and mast-cellgranules is completely inhibited under these conditions. The increase instaining with orcein/urea after deamination suggests that the affinity of thedye for the tissue is increased by a negative charge on the substrate, and thattherefore the effective dye particle probably carries a positive charge. It doesnot prove, however, that the dye-substrate bond is purely ionic. Similarly,the decrease in staining after methylation is consistent with the attachmentof orcein to acidic groups in the tissue, but it must be remembered that inaddition to blocking acidic groups, prolonged methylation may be capable ofdestroying other groups, or even of hydrolysing and removing substancesfrom the section.
The selective staining of nuclei by orcein/urea so closely resembles theresults of the Feulgen method that it seemed possible that prolonged ex-posure to acid alcohol might have liberated aldehyde groups from nuclearDNA, these groups being then demonstrated by a Schiff-like component ofthe orcein. The failure of mucin to stain with orcein/urea even after treatmentwith periodic acid appears, however, to have excluded this possibility.
Mayer's haemalum and similar stains colour nuclei well, but cartilagematrix and mast-cell granules less intensely than Heidenhain's iron-haema-toxylin and basic aniline dyes. It seemed possible that a similar mechanismmight underlie both this relative specificity of Mayer's haemalum and that oforcein/urea, but the following facts do not support this notion. When asection is placed in haemalum the haematein present acts as an acid dye, whilethe alum attaches to acidic (basiphil) substrates. Combination of haemateinwith alum to form a lake is inhibited in Mayer's haemalum solution by thepresence of acetic or citric acid, and only occurs when the section is 'blued' inalkaline tap-water (Baker, 1958). If a section is stained in haemalum and bluedin the usual way, and then placed for a few minutes in 1% aq. haematein,cartilage matrix and mast-cell granules stain far more intensely, indicatingthe presence of bound alum in these sites. The relative failure of cartilage

Goldstein—Ionic and non-ionic bonds in staining 487
matrix and mast-cell granules to stain with Mayer's haemalum is thereforepresumably due not to an absence of substrates capable of binding alum, butto a relative lack of basic (acidophil) substances able to bind the acidic haema-tein. Thus to stain with Mayer's haemalum a tissue structure requires bothbasiphil and acidophil components, and at least some of the selectivity of themethod is due to this. There seems no reason to suppose that a similar factorgoverns staining by orcein/urea, so the reason for the selectivity of this methodmust be sought elsewhere.
One may provisionally conclude that in the presence of urea, positivelycharged orcein particles attach to negatively charged sites in nuclei, but thatthis electrostatic affinity appears to be stabilized by unknown forces, whichare probably not hydrogen bonds. The selectivity and simplicity of the methodmay prove valuable histochemically, if the mechanism and chemical specificitycan be established.
The effect of urea on staining with basic dyes
The decrease caused by urea in the staining of mucin by some basic dyes(table 3) may indicate that the ionic linkage of basic dye to acidic substrate isstabilized in this site by hydrogen bonds. The greater tendency to hydrogenbonding shown by large molecules (Vickerstaff, 1954) could then explainwhy mucin tends to be stained better by basic dyes of large rather than smallparticle size (Goldstein, 1962).
Basic dyes sometimes attach by an alcohol-labile linkage, possibly a hydro-gen bond, to substrates not known to contain strongly acidic groups. Anexample of this is the staining of smooth muscle-cell cytoplasm, red blood-corpuscles, and other structures by saturated azure A in o-i N NaOH (pHabout 13*0) (Goldstein, unpublished). The staining of acidic substrates byazure A is greatly diminished at this pH, and this suggests that only a lowconcentration of the dye cation is present. This in turn suggests that the in-creased non-specific, labile staining seen under these conditions is due tothe unionized (hydroxide) form of the dye. In the case of a weakly basicdye, a significant proportion of the dye would be present in the unionizedform even at fairly low pH. The pK figures for most basic dyes are not avail-able, but contrary to Singer (1952) at least some basic dyes are weak bases(Goldacre and Phillips, 1949; Taylor, 1961). That Victoria blue 4R is a weakbase is suggested by the fact that it is not as soluble in water as in the lessstrongly polar alcohol (Gurr, i960). Alcohol-labile staining of elastic fibresby Victoria blue 4R in veronal buffer at pH 4-2 (Goldstein, unpublished) maythen be due to the unionized form of the dye; it is noteworthy that this stain-ing is inhibited by the presence of urea.
The increase in the staining of some substrates caused by urea also requiresdiscussion.
The selectivity of alcian blue and other basic dyes of large particle sizeprobably depends to a large extent on their inability to penetrate dense sub-strates (Goldstein, 1962). Now it is known that most basic dyes are aggregated
2421.4 L 1

488 Goldstein—Ionic and non-ionic bonds in staining
(associated or 'polymerized') in solution (Valko, 1935, 1946; Robinson, 1935;Rabinowitch and Epstein, 1941; Vickerstaff, 1954). It has further beenpointed out (Valko, 1946) that the same forces are probably responsible forthe intermolecular attraction of dye molecules in solution, and for the affinityof the dye for its substrate. We have seen that the latter affinity may dependpartly on hydrogen bonds. Urea may then be able to inhibit dye association insolution; this would tend to facilitate penetration and staining of dense sub-strates, but would not have the same effect on the staining of readily permeablesubstances such as mucin (Goldstein, 1962).
The suggestion that urea inhibits association of basic dyes in solution issupported by the fact that a solution of a basic dye, saturated with urea,tends to be less reddish in colour than a control solution, when the twosolutions are viewed with a comparator. This effect closely resembles thatseen when one compares a concentrated and a dilute solution of the samedye; the reddish colour of the stronger solution is generally attributed to-association of the dye molecules (Michaelis, 1947; Schubert and Hamerman,1956).
The effect of urea on staining by basic dyes may be simulated by othercompounds. For example, it was reported by Belanger and Hartnett (i960)that toluidine blue in a phthalate / tartaric acid buffer at pH 4-4 stained car-tilage matrix, mast-cell granules, and certain other structures metachroma-tically, but failed to stain mucins of the respiratory and gastro-intestinaltracts. Unpublished work by the author has shown that the staining of mucinby toluidine blue, azure A, and alcian blue is less in a phthalate buffer thanin a veronal buffer at the same low pH, while the staining of nuclei and car-tilage tends to be increased. This effect was particularly noticeable in the caseof alcian blue. The action of phthalate thus closely resembles that of urea,and since the carboxyl groups of the phthalate molecule are capable ofhydrogen bonding (Fieser and Fieser, 1957) it seems possible that the twocompounds act in the same way.
I wish to thank Mr. D. S. Dry and Mr. R. Herman for technical assistance,and Miss A. Andrew and Dr. A. G. Oettle for criticism of the manuscript.The work was performed in the Department of Anatomy under ProfessorP. V. Tobias, whom I wish to thank for his encouragement.
ReferencesBAKER, J. R., 1958. Principles of biological microtechnique. London (Methuen).BANGLE, R., 1954. J. Histochem. Cytochem., 2, 291.
1956. Am. J. Path., 32, 349.BALANCER, L. F., and HARTNETT, A., i960. J. Histochem. Cytochem., 8, 75.BURKL, W., 1951. Anat. Anz., 98, 116.CANNON, H. G., 1937. Nature, 139, 549.CASSELMAN, W. G. B., 1959. Histochemical technique. London (Methuen).DEMPSEY, E. W., and LANSING, A. I., 1954. Int. Rev. Cytol., 3, 437.
VIAL, J. D., LUCAS, R. V., and LANSING, A. I., 1952. Anat. Rec, 113, 197.ENGLE, R. L., and DEMPSEY, E. W., 1954. J. Histochem. Cytochem., 2, 9.

Goldstein—Ionic and non-ionic bonds in staining 489
FIESER, L. F., and FIESER, M., 1957. Introduction to organic chemistry. Boston (Heath).FISCHER, A., 1899. Fixirung, Farbung und Bau des Protoplasvias. Jena (Fischer).FISHER, E. R., and LILLIE, R. D., 1954. J. Histochem. Cytochem., 2, 81.FRENCH, J. E., and BENDITT, E. P., 1953. Ibid., 1, 321.FULLMER, H. M., 1959. Nature, 183, 1274.
and LILLIE, R. D., 1956. J. Histochem. Cytochem., 4, 64.1957. Ibid., 5, 11.
—— 1958. Ibid., 6, 425.GATENBY, J. B., and PAINTER, T., 1937. The microtomists' vademecum, 10th ed. London
(Churchill).GILLMAN, T., PENN, J., BRONKS, D., and Roux, M., 1954. Nature, 174, 789.GOLDACRE, R. J., and PHILLIPS, J. N., 1949. J. chem. Soc... part 3, 1724.GOLDSTEIN, D. J., 1959. Anat. Rec, 134, 217.
1962. Stain Tech., in press.GOMORI, G., 1950. Am. J. clin. Path., 20, 665.HALE, L. J., 1958. 'Biological laboratory data.' London (Methuen).HALL, D. A., 1959. Int. Rev. Cytol., 8, 211.HARMS, H., 1957. Acta Histochem., 4, 314.HAUROWITZ, F., 1950. Chemistry and biology of proteins. New York (Academic Press).KRAMER, H., and WINDRUM, G. M., 1955. J. Histochem. Cytochem., 3, 227.LANDSMEER, J. M. F., 1951. Acta Physiol. Pharmacol. Neerl., 2, 112.LEVINE, N. D., and MORRILL, C. C, 1941. Stain Tech., 16, 121.LILLIE, R. D., 1951. Ibid., z6, 123.MICHAELIS, L., 1947. Symp. Quant. Biol., 12, 131.MORRIS, A. L., and KRIKOS, G. A., 1958. Proc. Soc. exp. Biol. Med., 97, 527.MOWRY, R. W., 1956. J. Histochem. Cytochem., 4, 407.OSTWALD, W., 1933. Trans. Faraday Soc, 29, 347.PUCHTLER, H., SWEAT, F., BATES, R., and BROWN, J. H., 1961. J. Histochem. Cytochem., 9,
553-RABINOWITCH, E., and EPSTEIN, L. F., 1941. J. Am. chem. Soc, 63, 69-78.RADDEN, B. G., 1961. J. Histochem. Cytochem., 9, 165.ROBINSON, C, 1935. Trans. Faraday Soc, 31, 245.SALTHOUSE, T. N., 1944. Stain Tech., 19, 91.SCHUBERT, M., and HAMERMAN, D., 1956. J. Histochem. Cytochem., 4, 159.SEKI, M., 1933. Fol. Anat. Jap., 11, 377.
1933a. Ibid., 391.1934. Ibid., 12, 11.
SINGER, M., 1952. Int. Rev. Cytol., 1, 211.SPICER, S. S., and MEYER, D. B., i960. Am. J. clin. Path., 33, 453.STEEDMAN, H. F., 1950. Quart. J. micr. Sci., 91, 477.STOLK, A., 1961. Nature, 190, 360.TAYLOR, K. B., 1961. Stain Tech., 36, 73.VALK6, E., 1935. Trans. Faraday Soc, 31, 230.
1946. 'Physical chemistry of dyeing.' In Colloid Chemistry, 6, 594 (ed. J. Alexander).New York (Reinhold).
VICKERSTAFF, T., 1954. The physical chemistry of dyeing. 2nd ed. London (Oliver & Boyd).WEISS, J., 1954. J. Histochem. Cytochem., 2, 21.WIGGLESWORTH, V. B., 1952. Quart. J. micr. Sci., 93, 105.WOLMAN, M., 1961. Stain Tech., 36, 21.
Appendix
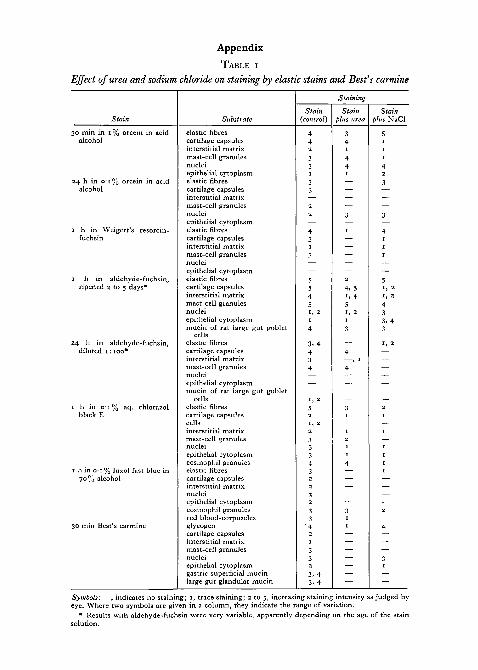
TABLE I
Effect of urea and sodium chloride on staining by elastic stains and Best's carmine
Stain
30 min in 1% orcein in acidalcohol
24 h in O'i% orcein in acidalcohol
1 h in Weigert's resorcin-fuchsin
1 h in aldehyde-fuchsin,ripened 2 to 5 days*
24 h in aldehyde-fuchsin,diluted 1:100*
1 h in 0 1 % aq. chlorazolblack E
1 h in o-i % luxol fast blue in70% alcohol
30 min Best's carmine
Substrate
elastic fibrescartilage capsulesinterstitial matrixmast-cell granulesnucleiepithelial cytoplasmelastic fibrescartilage capsulesinterstitial matrixmast-cell granulesnucleiepithelial cytoplasmelastic fibrescartilage capsulesinterstitial matrixmast-cell granulesnucleiepithelial cytoplasmelastic fibrescartilage capsulesinterstitial matrixmast-cell granulesnucleiepithelial cytoplasmmucin of rat large gut goblet
cellselastic fibrescartilage capsulesinterstitial matrixmast-cell granulesnucleiepithelial cytoplasmmucin of rat large gut goblet
cellselastic fibrescartilage capsulescellsinterstitial matrixmast-cell granulesnucleiepithelial cytoplasmeosinophil granuleselastic fibrescartilage capsulesinterstitial matrixnucleiepithelial cytoplasmeosinophil granulesred blood-corpusclesglycogencartilage capsulesinterstitial matrixmast-cell granulesnucleiepithelial cytoplasmgastric superficial mucinlarge gut glandular mucin
Staining
Stain(control)
442
331
33—2
2
431
3
—
55451, 21
4
3, 4434——
'> 2
52
i , 2
2
333432
2
32
33
• 42
1
332
3. 43. 4
Stainplus urea
341
441
—
—
—
—
3
1
—
—
—
—
—
2
4, 51, 4SI) 2I
3
—
4—, 14——
—
31—
1
21
1
4—————
3
1
—
—
—
—
—
—
—
Stainplus NaCl
5
1
1
42
3
——
3
41
1
1—
—
S1, 21, 2433. 43
1, 2—
—
—
—
—
2
1—
1
—1
1
1
1
—
—
—
—
2
—
4
——
31
—
—
Symbols: —, indicates no staining; 1, trace staining; 2 to 5, increasing staining intensity as judged byeye. Where two symbols are given in a column, they indicate the range of variation.
• Results with aldehyde-fuchsin were vsolution.
hey indicate the range of variation.able, apparently depending on the age of the stain

Goldstein—Ionic and non-ionic bonds in staining 491
TABLE 2
Effect of oxidation by 1% periodic acid on staining with Best's carmine andSchiff's reagent
Stain
Best's carmine (control)S min periodic acid, Best's
carmine60 min periodic acid, Best's
carmine5 min periodic acid, Schiff's
reagent60 min periodic acid, Schiff's
reagent
Staining of substrate
Glycogen
4
2, 3
4
5
Mucin ofXenopus
mucousneck cells
3
2
4
5
Mucin ofratgastric
surfaceepithelium
3. 4
2, 3
4
5
Mucin ofrat smallgut goblet
cells
2, 3
2
5
5
Mucin ofrat large
gut gobletcells
3
2
5
5
Chromidialsubstance
3
3
3
Nuclei
3
3
3
Symbols • As in table
TABLE 3
Effect of urea on staining with basic dyes, correlated with the molecular weight{MW) of dye and R.I. of substrate
Stain
o-i% azure A (MW292) in 30% alcohol
Ditto, saturated withurea
O'Oi M Bismarckbrown R (MW 461)
Ditto, saturated withurea
c o i M Janus green B(MW 483)
Ditto, saturated withurea
001 M Victoria blue4 R (MW 520)
Ditto, saturated withurea
o- i% alcian blue(MW c. 1341), 3%acetic
Ditto, saturated withurea
Mucin ofrat smallgut gobletcells (L)
—, i
—, 1
2
I
4
3
2, 3
—
3
3
Mucin ofrat largegut gobletcells (L)
3
I, 2
4
3 , 4
4. 5
3. 4
3
—
4
4
Sta
Cartilagecapsules (L)
4, 5
4. 5
4
4
5
5
3
2
3
4
ning of substrate
Interstitialcartilagematrix(/, H)
2
2
2
2
3
5
2
—
1
2
Mast-cellgranules
(I,H)
5
S
3
4
4
5
3
4
2> 3
4
Nuclei(I,H)
3
3
2
3
3
3
3
2
1
3
PancreaticUNA (H)
3, 4
4
2
3
3
4, 5
2
2
—
2
Symbols: As in table 1.All slides were stained 15 min and dehydrated in alcohol. Results were somewhat different in air-
dried slides, especially with Victoria blue 4R. Bismarck brown R, Janus green B, and Victoria blue 4Rwere made up in veronal-acetate buffer at pH 40. Molecular weights are from Gurr (i960). H indicatesR.I. above about 1-54; I, about 1-47-1-54; L, 1-47 or lower.

492 Goldstein—Ionic and non-ionic bonds in staining
TABLE 4
Precipitation of dyes by sodium chloride
To an aliquot of stain solution was added 10% NaCl in the case of aqueous solutions, orNaCl to saturation in the case of alcoholic solutions. After standing for about 15 min, thesolution was filtered and matched on a comparator with the original solution. The ratio givenin the table is the optical density of the stain solution to which sodium chloride had been added,
divided by the optical density of the original solution.
Stain solution
0 1 % alcian blue in 3% acetic acid0 1 % Victoria blue 4R in buffer, pH 4-10 1 % azure A in buffer, pH 4-1o-i% luxol fast blue in 70% alcohol0 1 % Bismarck brown R in buffer, pH 4-10 1 % aq. chlorazol black Eo-i% Janus green B in buffer, pH 4-1Best's carmineWeigert's resorcin-fuchsin0 1 % orcein in acid alcoholGomori's aldehyde-fuchsin
Ratio
0 00-13O'is0 1 7
0-180 2 4
O-43C 6 51 0
1 01 0