isoprene emission aids recovery of photosynthetic performance in transgenic nicotiana tabacum...
TRANSCRIPT

Ite
MTa
b
c
d
e
f
a
AA
KAFIMPU
1
i3pvrtpIp
F
h0
Plant Science 226 (2014) 82–91
Contents lists available at ScienceDirect
Plant Science
j ourna l ho me pa g e: www.elsev ier .com/ locate /p lantsc i
soprene emission aids recovery of photosynthetic performance inransgenic Nicotiana tabacum following high intensity acute UV-Bxposure
auro Centrittoa,∗, Matthew Haworthb, Giovanni Marinob,c, Emanuele Pallozzid,e,sonko Tsonevf, Violeta Velikovaf, Isabel Noguesd, Francesco Loretob
Trees and Timber Institute, National Research Council, Via Madonna del Piano 10, 50019 Sesto Fiorentino, FI, ItalyInstitute of Plant Protection, National Research Council, Via Madonna del Piano 10, 50019 Sesto Fiorentino, FI, ItalyDepartment of Biosciences and Territory, University of Molise, Contrada Fonte Lappone, 86090 Pesche, IS, ItalyInstitute of AgroEnvironmental and Forest Biology, National Research Council, Via Salaria km 29.300, 00015 Monterotondo Scalo, RM, ItalyDepartment of Forest Environment and Resources, University of Tuscia, Via San Camillo de Lellis, 01100 Viterbo, ItalyInstitute of Plant Physiology and Genetics, Bulgarian Academy of Sciences, Acad. G. Bonchev Str., Bl. 21, 1113 Sofia, Bulgaria
r t i c l e i n f o
rticle history:vailable online 19 June 2014
eywords:ntioxidant defenseluorescencesoprene
ethanolhotosynthesisV-B irradiance
a b s t r a c t
Isoprene emission by terrestrial plants is believed to play a role in mitigating the effects of abiotic stress onphotosynthesis. Ultraviolet-B light (UV-B) induces damage to the photosynthetic apparatus of plants, butthe role of isoprene in UV-B tolerance is poorly understood. To investigate this putative protective role,we exposed non-emitting (NE) control and transgenic isoprene emitting (IE) Nicotiana tabacum (tobacco)plants to high intensity UV-B exposure. Methanol emissions increased with UV-B intensity, indicatingoxidative damage. However, isoprene emission was unaffected during exposure to UV-B radiation, butdeclined in the 48 h following UV-B treatment at the highest UV-B intensities of 9 and 15 W m−2. Pho-tosynthesis and the performance of photosystem II (PSII) declined to similar extents in IE and NE plantsfollowing UV-B exposure, suggesting that isoprene emission did not ameliorate the immediate impact of
UV-B on photosynthesis. However, after the stress, photosynthesis and PSII recovered in IE plants, whichmaintained isoprene formation, but not in NE plants. Recovery of IE plants was also associated withelevated antioxidant levels and cycling; suggesting that both isoprene formation and antioxidant sys-tems contributed to reinstating the integrity and functionality of cellular membranes and photosynthesisfollowing exposure to excessive levels of UV-B radiation.© 2014 Elsevier Ireland Ltd. All rights reserved.
. Introduction
The level of ultraviolet-B radiation (UV-B, 280–320 nm) reach-ng the Earth’s surface has increased significantly over the past0-years, as anthropogenic emissions of chlorofluorocarbon com-ounds have depleted levels of stratospheric ozone [1]. A largeolume of literature has demonstrated that exposure to UV-Badiation induces a variety of deleterious effects in plants [2], in par-icular to processes such as photosynthesis and the accompanying
rotective physiology that includes UV absorbing compounds [3,4].soprene is the most abundant BVOC (biogenic volatile organic com-ound) emitted by plants, and has been shown to have an important
∗ Corresponding author at: CNR-IVALSA, Via Madonna del Piano 10, 50019 Sestoiorentino, FI, Italy. Tel.: +39 055 52251; fax: +39 055522 5507.
E-mail address: [email protected] (M. Centritto).
ttp://dx.doi.org/10.1016/j.plantsci.2014.06.004168-9452/© 2014 Elsevier Ireland Ltd. All rights reserved.
protective function against abiotic stress [5,6]. Despite their crucialprotective role, there have been relatively few studies on the impactof UV radiation on the emission of BVOCs [7].
The photosynthetic apparatus of plants is sensitive to UV-B radi-ation exposure. The short wavelength and associated energeticsof UV-B light causes damage to the ultrastructure of chloro-plasts, impairs photosystem II (PSII) electron transport, diminishesribulose 1,5-bisphosphate carboxylase/oxygenase activity, andultimately reduces the concentration of chlorophyll. However,plants are able to produce a variety of secondary metabolites thateffectively absorb UV-B, and these metabolites prevent penetrationof harmful UV-B radiation into the mesophyll cells of the leaves[3,8]. For instance, the synthesis and accumulation of UV absorbing
compounds, particularly flavonoids located in the vacuoles of epi-dermal cells, can be elicited in leaves exposed to UV radiation [9,7].Isoprenoid emission can diminish oxidative stress, preven-ting the accumulation of reactive oxygen species (ROS) and

M. Centritto et al. / Plant Science 226 (2014) 82–91 83
F and d) −2
w esultsd
pOfTa[poaah[t[usU[b
ig. 1. Isoprene emission of transgenic and wild type tobacco plants exposed to 3 (a
ere made before, immediately after, 24 h and 48 h after the UV-B treatments. Rifferences at P < 0.05 in response to the UV-B treatments.
rotecting cell membranes against lipid peroxidation [10,11].xidative stress is a consequence of an imbalance between ROS
ormation and the ROS scavenging capacity of the defense system.o scavenge ROS, plants possess enzymatic and non-enzymaticntioxidants, such as ascorbate (ASC) and glutathione (GSH)12,13]. Isoprene is not an effective aqueous antioxidant and isrobably unable to directly scavenge aqueous ROS. However, aque-us and lipid-phase antioxidant capacities are linked (e.g. throughscorbate-mediated regeneration of tocopherol moieties) [14], andny shift in lipid-phase antioxidant capacity may be expected toave an impact upon the capacity of aqueous-phase antioxidants5]. Moreover, isoprene may strengthen membranes thus preven-ing their denaturation when exposed to a variety of harmful agents5]. The general effects of UV radiation on isoprene emission remainnclear [7], and there are presently no studies that clearly demon-
trate a protective role of isoprene against UV-B stress. Exposure toV-B induced no change in isoprenoid emission in Ilex aquifolium15] and Picea abies [16], and increased emission in Quercus gam-elii [17] and in a sub-arctic peatland dominated by Warnstorfia
, 9 (b and e) and 15 (c) W m intensity of UV-B radiation for 90 min. Measurements are means of four replicates ± 1 SEM. Different letters (a–c) indicate significant
exannulata [18], but reduced emission in Pistacia lentiscus [15].These diverse observations may be due to species-specific differ-ences, seasonal effects, and variation in the level and duration ofUV-B exposure among the studies.
To investigate the relationship between UV radiation and BVOCemission, we used transgenic isoprene-emitting (IE) and controlnon-isoprene emitting (NE) Nicotiana tabacum plants. To elicit thepotential mechanism underlying photosynthetic and protectiveresponses of N. tabacum, the plants were exposed to high UV-Bintensities over a short period. We hypothesized that tobacco plantsgenetically manipulated to emit isoprene (IE) are more tolerantthan NE controls when exposed to acute UV-B radiation. The aim ofthe study was: (i) to relate isoprene emission to exposure to highintensities of UV-B radiation; (ii) to assess PSII functionality in IEand NE plants as an indicator of the efficiency of photochemical
reactions at the thylakoid membranes following UV-B exposure;and (iii) to assess the synthesis of protective metabolites followingacute exposure to high intensities of UV-B radiation in IE and NEplants.
84 M. Centritto et al. / Plant Science 226 (2014) 82–91
Fig. 2. Methanol emission of transgenic IE and control NE tobacco plants exposed to 3 (a and d), 9 (b and e) and 15 (c) W m−2 UV-B radiation for 90 min. Measurements werem d in thl e UV-B
2
2
l(u(nppiGusftm
ade before and immediately after UV-B treatment, and then recovery was assesseetters (a–d) indicate significant difference in the mean at P < 0.05 in response to th
. Materials and methods
.1. Plant material
Two isoprene-emitting (IE) transgenic tobacco (N. tabacum)ines (6H and 12H), and their corresponding isoprene non-emittingNE) ‘wild-type’ control equivalents (lines 6A and 12A) weresed for this study. Seeds were provided by Malcolm PossellDepartment of Environmental Sciences, The University of Syd-ey, Australia). The altered N. tabacum does not display alteredlant growth, morphology or photosynthesis as a result of iso-rene biosynthesis [19]. After germination, seedlings were potted
n 4 dm3 pots containing Klasmann compost (Klasmann–DeilmannmbH, Germany) and grown in a greenhouse. Plants were reg-larly watered to pot capacity and fertilized with Hoagland
olution once a week in order to supply mineral nutrients atree access rates [20,21]. Fully expanded leaves (fifth node fromhe apical meristem) of five-week-old plants were used for alleasurements.
e following 24 and 48 h. Results are the means of four replicates ± 1 SEM. Different treatments.
2.2. Light system
A Manhood 31 lamp (Vinci Fine Instruments, Monterotondo,Italy) was used to generate UV-B radiation. The UV-B lamp hasa spectral distribution in the range of 280–345 nm, with a peakwavelength at 310 nm, but also including a portion of UV-A radi-ation (about 15% of total radiation). A DO 9721 photoradiometerequipped with a LP 9021 UV-B probe (Delta Ohm S.r.l., Italy) wasused to measure the level of UV-B radiation reaching the leaves.
2.3. UV-B treatments
Three high-intensity levels of UV-B radiation (3, 9 and15 W m−2) were applied to the tobacco plants to determine anyintensity-response effects. To place these levels of UV-B radiation
into a real world context, the highest recorded global level of UV-B radiation was 3.96 W m−2 in Lhasa, Tibet; this is approximatelythree times greater than standard levels observed in NorthernEurope [22]. Plants were irradiated for 90 min by positioning the
M. Centritto et al. / Plant Science 226 (2014) 82–91 85
F and dw esultsd
UBwaeab(U
2
mS(Uwct
ig. 3. Photosynthesis (A) of transgenic and wild type tobacco plants exposed to 3 (aere made before, immediately after, 24 h and 48 h after the UV-B treatments. Rifferences at P < 0.05 in response to the UV-B treatments.
V-B lamp perpendicular to a fully expanded leaf. Gas exchange,VOC emissions, and images of whole leaf chlorophyll fluorescenceere analysed before and immediately after the UV-B treatments,
nd then 24 and 48 h after the UV-B treatments. Due to thextremely harmful effects of high intensity UV-B radiation on plantsnd the occurrence of an observable effect in isoprene emissioneing apparent only in the Line 6 N. tabacum plants at 9 W m−2
Fig. 1), it was decided to expose only Line 6 tobacco to 15 W m−2
V-B.
.4. Gas exchange measurements
Photosynthesis and BVOC emissions were simultaneouslyeasured in the laboratory under controlled conditions [23].
teady-state net photosynthesis (A) and stomatal conductancegs) were recorded using a LI-6400 IRGA (LI-COR, Lincoln, NE,
SA), by enclosing a portion of the leaf in a 6 cm2 cuvetteith a transparent upper Teflon window. A 300 �mol s−1 flow ofontaminant-free synthetic air, made of N2, O2 and CO2 in concen-rations representative of atmospheric concentrations (80%, 20%
), 9 (b and e) and 15 (c) W m−2 intensity of UV-B radiation for 90 min. Measurements are means of four replicates ± 1 SEM. Different letters (a–d) indicate significant
and 380 �mol mol−1, respectively) was provided to the leaves usinggas tanks and mass flow controllers. Measurements were taken onleaves exposed to a saturating photosynthetic photon flux densityof 1000 �mol m−2 s−1 actinic white light, at a temperature of 30 ◦Cand with the relative humidity of the air within the leaf cuvetteranging between 45 and 55%.
BVOC emission was measured by connecting the exhaust linefrom the leaf cuvette to a proton transfer reaction – mass spec-trometer (PTR-MS, Ionicon Analytik GmbH, Innsbruck, Austria). ThePTR-MS was set in a single ion mode to record traces of protonatedmasses of isoprene (m/z 69) and methanol (m/z 33) (details arereported in Lindinger et al. [24]. The instrument was calibrated dailywith gaseous standards for each compound (Rivoira, Milano, Italy),and measurements were validated with gas-chromatography anal-ysis (Syntech Spectras BTX Analyzer GC 855, Netherlands).
2.5. Chlorophyll fluorescence measurements
A Mini-Imaging-PAM-fluorometer (Heinz Walz GmbH, Effel-trich, Germany) was used to measure chlorophyll a fluorescence
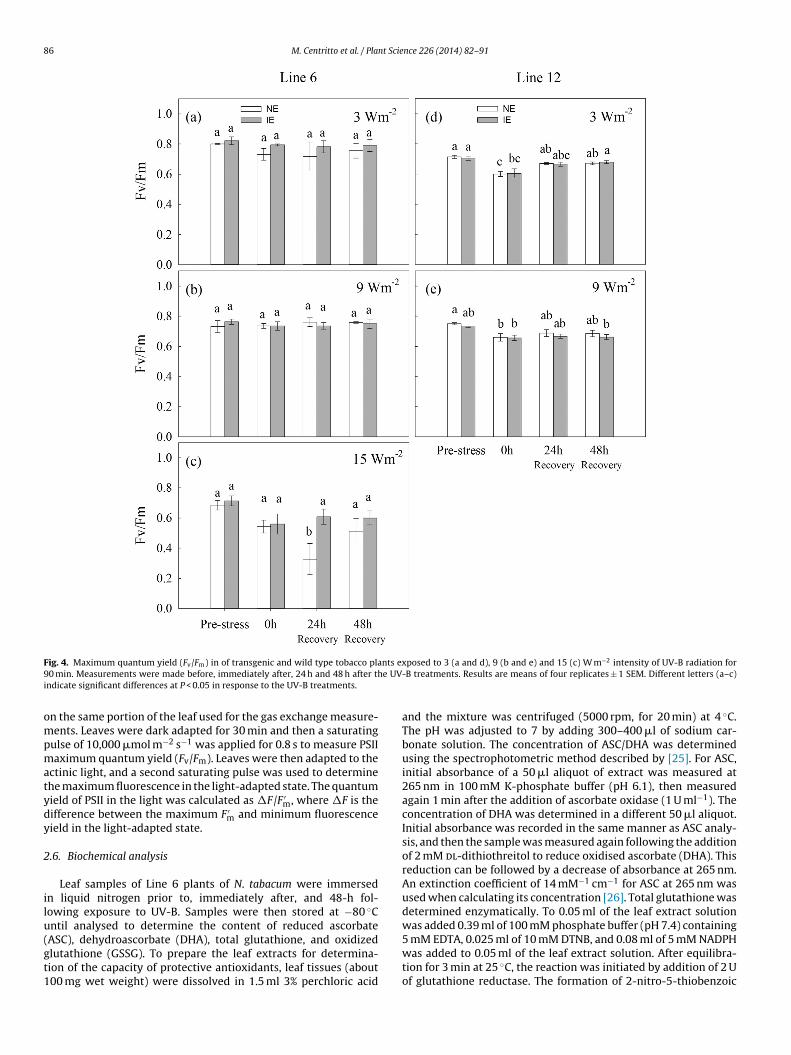
86 M. Centritto et al. / Plant Science 226 (2014) 82–91
F nts ex9 e UVi
ompmatydy
2
ilu(gt1
ig. 4. Maximum quantum yield (Fv/Fm) in of transgenic and wild type tobacco pla0 min. Measurements were made before, immediately after, 24 h and 48 h after th
ndicate significant differences at P < 0.05 in response to the UV-B treatments.
n the same portion of the leaf used for the gas exchange measure-ents. Leaves were dark adapted for 30 min and then a saturating
ulse of 10,000 �mol m−2 s−1 was applied for 0.8 s to measure PSIIaximum quantum yield (Fv/Fm). Leaves were then adapted to the
ctinic light, and a second saturating pulse was used to determinehe maximum fluorescence in the light-adapted state. The quantumield of PSII in the light was calculated as �F/F ′
m, where �F is theifference between the maximum F ′
m and minimum fluorescenceield in the light-adapted state.
.6. Biochemical analysis
Leaf samples of Line 6 plants of N. tabacum were immersedn liquid nitrogen prior to, immediately after, and 48-h fol-owing exposure to UV-B. Samples were then stored at −80 ◦Cntil analysed to determine the content of reduced ascorbate
ASC), dehydroascorbate (DHA), total glutathione, and oxidizedlutathione (GSSG). To prepare the leaf extracts for determina-ion of the capacity of protective antioxidants, leaf tissues (about00 mg wet weight) were dissolved in 1.5 ml 3% perchloric acidposed to 3 (a and d), 9 (b and e) and 15 (c) W m−2 intensity of UV-B radiation for-B treatments. Results are means of four replicates ± 1 SEM. Different letters (a–c)
and the mixture was centrifuged (5000 rpm, for 20 min) at 4 ◦C.The pH was adjusted to 7 by adding 300–400 �l of sodium car-bonate solution. The concentration of ASC/DHA was determinedusing the spectrophotometric method described by [25]. For ASC,initial absorbance of a 50 �l aliquot of extract was measured at265 nm in 100 mM K-phosphate buffer (pH 6.1), then measuredagain 1 min after the addition of ascorbate oxidase (1 U ml−1). Theconcentration of DHA was determined in a different 50 �l aliquot.Initial absorbance was recorded in the same manner as ASC analy-sis, and then the sample was measured again following the additionof 2 mM dl-dithiothreitol to reduce oxidised ascorbate (DHA). Thisreduction can be followed by a decrease of absorbance at 265 nm.An extinction coefficient of 14 mM−1 cm−1 for ASC at 265 nm wasused when calculating its concentration [26]. Total glutathione wasdetermined enzymatically. To 0.05 ml of the leaf extract solutionwas added 0.39 ml of 100 mM phosphate buffer (pH 7.4) containing
5 mM EDTA, 0.025 ml of 10 mM DTNB, and 0.08 ml of 5 mM NADPHwas added to 0.05 ml of the leaf extract solution. After equilibra-tion for 3 min at 25 ◦C, the reaction was initiated by addition of 2 Uof glutathione reductase. The formation of 2-nitro-5-thiobenzoic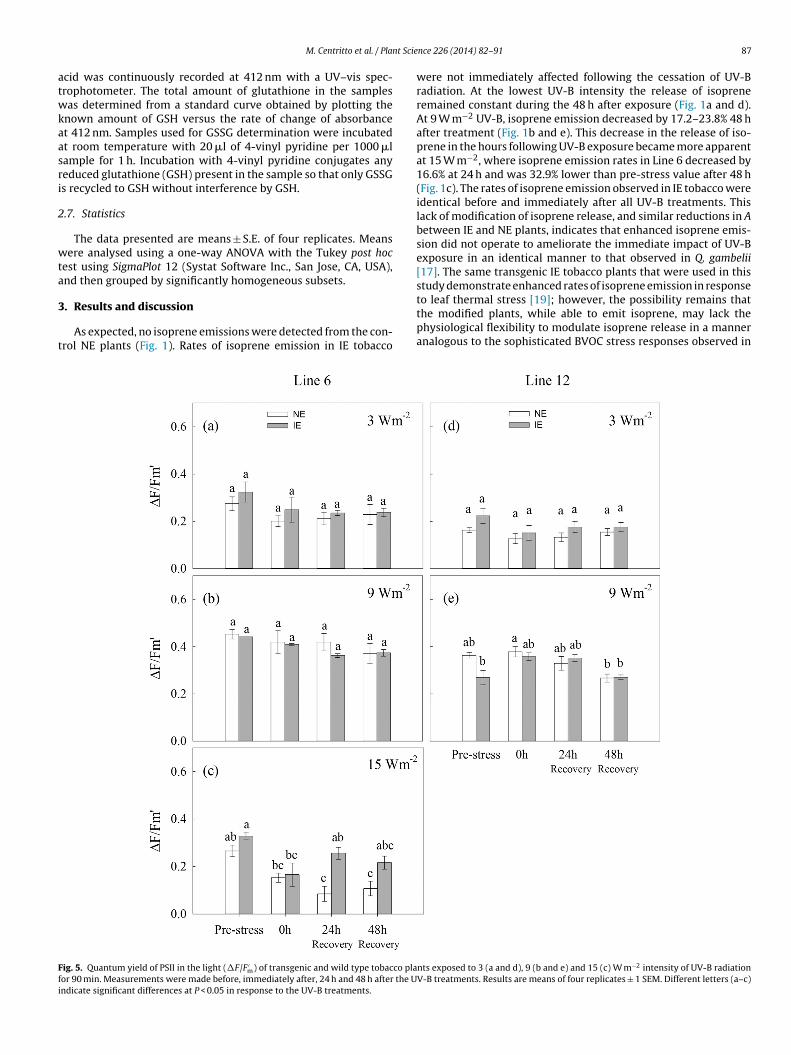
nt Scie
atwkaasri
2
wta
3
t
Ffi
M. Centritto et al. / Pla
cid was continuously recorded at 412 nm with a UV–vis spec-rophotometer. The total amount of glutathione in the samplesas determined from a standard curve obtained by plotting the
nown amount of GSH versus the rate of change of absorbancet 412 nm. Samples used for GSSG determination were incubatedt room temperature with 20 �l of 4-vinyl pyridine per 1000 �lample for 1 h. Incubation with 4-vinyl pyridine conjugates anyeduced glutathione (GSH) present in the sample so that only GSSGs recycled to GSH without interference by GSH.
.7. Statistics
The data presented are means ± S.E. of four replicates. Meansere analysed using a one-way ANOVA with the Tukey post hoc
est using SigmaPlot 12 (Systat Software Inc., San Jose, CA, USA),nd then grouped by significantly homogeneous subsets.
. Results and discussion
As expected, no isoprene emissions were detected from the con-rol NE plants (Fig. 1). Rates of isoprene emission in IE tobacco
ig. 5. Quantum yield of PSII in the light (�F/F ′m) of transgenic and wild type tobacco pla
or 90 min. Measurements were made before, immediately after, 24 h and 48 h after the Undicate significant differences at P < 0.05 in response to the UV-B treatments.
nce 226 (2014) 82–91 87
were not immediately affected following the cessation of UV-Bradiation. At the lowest UV-B intensity the release of isopreneremained constant during the 48 h after exposure (Fig. 1a and d).At 9 W m−2 UV-B, isoprene emission decreased by 17.2–23.8% 48 hafter treatment (Fig. 1b and e). This decrease in the release of iso-prene in the hours following UV-B exposure became more apparentat 15 W m−2, where isoprene emission rates in Line 6 decreased by16.6% at 24 h and was 32.9% lower than pre-stress value after 48 h(Fig. 1c). The rates of isoprene emission observed in IE tobacco wereidentical before and immediately after all UV-B treatments. Thislack of modification of isoprene release, and similar reductions in Abetween IE and NE plants, indicates that enhanced isoprene emis-sion did not operate to ameliorate the immediate impact of UV-Bexposure in an identical manner to that observed in Q. gambelii[17]. The same transgenic IE tobacco plants that were used in thisstudy demonstrate enhanced rates of isoprene emission in response
to leaf thermal stress [19]; however, the possibility remains thatthe modified plants, while able to emit isoprene, may lack thephysiological flexibility to modulate isoprene release in a manneranalogous to the sophisticated BVOC stress responses observed innts exposed to 3 (a and d), 9 (b and e) and 15 (c) W m−2 intensity of UV-B radiationV-B treatments. Results are means of four replicates ± 1 SEM. Different letters (a–c)

88 M. Centritto et al. / Plant Science 226 (2014) 82–91
Fig. 6. Total ascorbate content (ASC + DHA) and redox state ascorbate (ASC/ASC + DHA) in leaves of Line 6 transgenic and wild type tobacco plants exposed to 3 (a and c) and1 de befo < 0.05
Ee
mmaMeUwbdPsargieewt
inaiostt
5 (b and d) W m−2 intensity of UV-B radiation for 90 min. Measurements were maf four replicates ± 1 SEM. Different letters (a–c) indicate significant differences at P
riophorum russeolum, Carex limosa and Populus species followingxposure to UV-B [7,18].
The energetics of UV-B light causes damage to cell walls andembranes through oxidative stress [10,12]. Transient bursts ofethanol are associated with the impairment of cell wall integrity,
s pectins embedded in the cell wall are demethylated [27].ethanol emission was not affected by the lowest level of UV-B
xposure, but became enhanced following exposure to 9 W m−2 ofV-B (Fig. 2b and e). The highest dose of UV-B radiation (15 W m−2)as accompanied by 530–625% increases in methanol release in
oth IE and NE plants (Fig. 2c), indicative of pronounced cell wallegradation. These results are in keeping with those observed byallozzi et al. [7] who showed that UV-A treatments triggeredignificant methanol emissions. Although the results were gener-lly not significant (p < 0.05), the increase in the rate of methanolelease induced at 9 and 15 W m−2 was consistently lower in trans-enic IE than control NE plants, suggesting the possibility thatsoprene emission stabilised cell and thylakoid membranes to somextent [6]. Rates of methanol release observed at the two high-st UV-B intensities subsequently returned to pre-stress levelsithin 24 h indicative of the absence of further cell wall degrada-
ion.Acute exposure to UV-B radiation induced identical reductions
n rates of A in both IE and NE tobacco plants (Fig. 3). This instanta-eous depression in A following UV-B treatment was most apparentt the highest 15 W m−2 intensity of exposure (Fig. 3c), illustrat-ng the deleterious effects of increasing intensities of UV-B light
n photosynthesis. Over the 48 h following 3 W m−2 UV-B expo-ure the plants demonstrated a recovery of photosynthetic rateso pre-stress levels (Fig. 3a and d). However, Lines 6 and 12 N.abacum showed sustained impairment of photosynthetic rates inore, immediately after, 24 h and 48 h after the UV-B treatments. Results are means in response to the UV-B treatments.
the 48 h after UV-B treatment at the higher 9 and 15 W m−2 intensi-ties (Fig. 3b and c). At the highest UV-B intensity IE plants exhibiteda ∼290% increase in A in the 48 h after irradiation relative to valuesobserved immediately after irradiation, whereas photosynthesiscontinued to decline in NE plants exposed to the same treatment(Fig. 3c). The extent of photosynthetic impairment was reliant uponthe intensity of UV-B, with the plants showing relatively few effectsof 3 W m−2 UV-B but sustained damage following radiation levelsof 15 W m−2 (Fig. 3).
Photosystem II is considered the component of the pho-tosynthetic process most sensitive to UV-B [28]. The shortwavelength of UV-B radiation induces oxidative damage tothe thylakoid membranes where the PSII reaction centers arelocated, leading to lower PSII efficiency [29]. Irradiation withUV-B may also induce semiquinone anion formation in the thy-lakoid membranes and lower the rates of PSII electron transport[30]. Exposure to UV-B radiation induced immediate declinesin the efficiency of PSII (Fv/Fm) in Line 12 tobacco, but hadno effect on Line 6 (Fig. 4). The quantum yield of non-cyclicelectron transport (�F/F ′
m) was not affected by 3 and 9 W m−2
UV-B in either tobacco lines (Fig. 5). However, exposure to themost intense UV-B treatment resulted in declines in �F/F ′
m val-ues (Fig. 5c). At 15 W m−2 UV-B, IE plants maintain Fv/Fm values,and �F/F ′
m parameters return to pre-stress levels 24 h after treat-ment, coinciding with the recovery of A (Fig. 3c). In contrast, NEsshowed a reduction in Fv/Fm values 24 h after exposure (Fig. 4c) anddeclines in A and �F/F ′
m were sustained over 48 h (Figs. 3c and 4c).
Increased methanol emissions (Fig. 2c) and corresponding reduc-tions in �F/F ′m (Fig. 5c) exhibited by both IE and NE tobacco plantsused in this study are consistent with UV-B induced cell wall andmembrane damage [7].

M. Centritto et al. / Plant Science 226 (2014) 82–91 89
F G) in l1 de befo P < 0.0
cfdUeeiwtbtef
taciis[iailsern
ig. 7. Total glutathione (GSH + GSSG) and redox state glutathione (GSH/GSH + GSS5 (b and d) W m−2 intensity of UV-B radiation for 90 min. Measurements were maf four replicates ± 1 SEM. Different letters (a–d) indicate significant differences at
The similar reductions in photosynthetic performance and indi-ators of oxidative stress observed in IE and NE plants immediatelyollowing UV-B treatment suggests that the emission of isopreneoes not play a role in the direct stress response of tobacco to acuteV-B radiation. Nonetheless, IE tobacco displayed enhanced recov-ry of A and PSII performance relative to NE plants in the 48 h afterxposure to the highest UV-B level. Photosynthetic rates in IE plantsncreased three-fold in the 48 h after 15 W m−2 UV-B irradiation,
hereas A continued to decline in NE plants exposed to the samereatment (Fig. 3c). This recovery of A in IE plants was accompaniedy sustained emission of isoprene (Figs. 1c and 3c), suggesting thathe release of isoprene, albeit reduced relative to pre-stress lev-ls, may aid in the revitalisation of the photosynthetic apparatusollowing UV-B damage.
Exposure to UV radiation can induce the production of pro-ective compounds within the leaf [4]. Concentrations of thentioxidants ascorbate (ASC) and glutathione (GSH) were signifi-antly increased in both IE and NE plants immediately after UV-Brradiation (Figs. 6 and 7, respectively), suggesting that the antiox-dant capacities of the tobacco plants were rapidly enhanced tocavenge harmful ROS resulting from the interception of UV-B31]. Notably, the stimulation of antioxidant capacities was alreadynduced at 3 W m−2 UV-B irradiation, and was not further enhancedfter exposing plants to higher UV-B levels. Moreover, the antiox-dants were increased to similar levels in IE and NE plants. At theower UV-B of 3 W m−2, ASC levels in IEs and NEs were statistically
−2
imilar in the 48 h following exposure, but at 15 W m IE plantsxhibited higher total ASC than NEs (Fig. 6a and b). Regenerationates of ASC, indicated by the ratio of ASC to total ascorbate, wereot affected by exposure to either UV-B treatment (Fig. 6c and d)eaves of Line 6 transgenic and wild type tobacco plants exposed to 3 (a and c) andore, immediately after, 24 h and 48 h after the UV-B treatments. Results are means5 in response to the UV-B treatments.
[32]. Concentration on a leaf area basis of total GSH in IE and NEplants were identical 48 h after the UV-B treatments (Fig. 7). At3 W m−2 UV-B, total GSH returned to pre-stress levels in the twodays following treatment, but at 15 W m−2 GSH levels increasedby 290% in the hours following treatment; suggesting that the GSHresponse of tobacco is influenced by the intensity of UV-B exposure(Fig. 7a and b). In contrast to the observed stability of ASC contentand regeneration (Fig. 6a and b), the ratio of reduced GSH to totalglutathione declined at 15 W m−2 UV-B in both IE and NE plants inthe 48 h after treatment (Fig. 7d).
Ascorbate has been shown to protect thylakoid membranelipids from oxidative stress through the regeneration of liphophillicantioxidants [33], possibly accounting for the maintenance ofFv/Fm and �F/F ′
m values in IE relative to NE tobacco plants(Figs. 4c and 5c). However, no difference was observed betweenGSH levels following 15 W m−2 UV-B exposure in IE and NE plants(Fig. 7). Peroxidation of lipid membranes is minimised by GSH[34], accounting for both increased total GSH and lower ratios ofreduced GSH to total GSH found in the tobacco lines after UV-Bradiation (Fig. 7). At 15 W m−2 UV-B, the reduced GSH/total GSHratio decreased by ∼20% immediately after the treatment in bothplant types, and there were further ∼23% and ∼16% decreases inthe NE and IE plants respectively over 48 h; but no significant dif-ference was observed in GSH between NE and IE plants in thehours following UV-B treatment. This decrease in the regenera-tion rate of GSH found in both NE and IE plants after the highest
intensity UV-B treatment is an indication of the impairment ofthe anti-oxidant system to prevent long-term damage to photo-synthesis (Fig. 3e). The induction of these antioxidants as well astheir regeneration via the ascorbate–glutathione cycle appears to
9 nt Scie
brmcomtici
iiiprweibppomtaaIwioc
tarseiTUtw[oBrcpFihtlbpttaa
4
gU
[
[
[
[
[
0 M. Centritto et al. / Pla
e independent of the capacity for isoprene emission at low UV-Badiations. At higher UV-B radiations, the capacity of IE leaves toaintain higher ASC levels 48 h after the stress occurrence may
ontribute to more effective regulation of oxidative stress. Thebserved differences in antioxidant capacities between the plantsay suggest that isoprene emission alleviates some of the oxida-
ive stress that diminishes antioxidant content or that emission ofsoprene could be involved in the regulation of ASC but not GSHycles [35] in the response of tobacco to acute irradiation with highntensity UV-B light.
The results of this study suggest that isoprene emission is notnvolved in the immediate plant-stress response to acute high-ntensity UV-B radiation in the transgenic tobacco lines. An increasen the rate of methanol emission and decline in PSII performancearameters does indicate that high intensity UV-B resulted in dis-uption to cell walls and membranes [6], but this was not associatedith the rate of isoprene release from the transgenic tobacco. How-
ver, post UV-B exposure, IE plants did exhibit enhanced recoveryn photosynthetic rates and PSII efficiency. Sustained isopreneiosynthesis was associated with improvements in photosyntheticerformance in the 48 h after irradiation, while the photosynthetichysiology of NE plants exhibited continued deleterious effectsf UV-B radiation (Fig. 3). These findings suggest that isopreneay play a role in the physiological recovery from UV-B radia-
ion, possibly also mediated through enhanced antioxidant contentnd cycling preventing further lipid peroxidation [36], and thusllowing repair of damaged membranes, DNA and proteins [37,38].soprene tends to partition into the lipid phase of membranes
here it may protect membrane integrity and functionality by act-ng as an antioxidant [10,39–41]. This may account for the recoveryf photosynthetic performance observed in IE plants following theessation of UV-B stress.
The physiological response of the control NE and transgenic IEobacco plants appeared to be strongly reliant upon the dose ofcute UV-B radiation received by the leaf. While all UV-B treatmentsesulted in immediate declines in A, this was most apparent andustained at the higher intensities (Fig. 3). The lowest 3 Wm−2 UV-Bxposure did not affect PSII parameters or the emission of methanoln Line 6 tobacco, whereas 15 W m−2 had more pronounced effects.his may suggest that the protective mechanism utilised to absorbV-light [7,42] and disperse excess light energy as heat [43] in the
obacco plants is able to cope with ‘short’ bursts of UV-B radiationithin the ranges of those experienced under natural conditions
23] without experiencing a decline in A [7]. The levels and durationf UV-B radiation used in this study, while not representative of UV-
conditions found under current radiation regimes, do induce plantesponses that contribute to an understanding of the physiologi-al protective processes underpinning photosynthesis and damagerogression when these protective mechanisms are overwhelmed.urthermore, plants may have experienced acute UV-B injury, sim-lar to that induced in this study at mass extinction events in Earthistory when catastrophic losses of stratospheric ozone are thoughto have occurred [44], possibly triggering major events in plant evo-ution through mutation of DNA and metabolic disruption causedy ROS [45,46]. Further studies should be undertaken to exposelants to more realistic UV levels over longer time periods to assesshe possible role of isoprene in ameliorating the effects and con-ributing to enhanced recovery to the effects of long-term exposurend incremental degradation of the photosynthetic and protectivepparatus.
. Conclusions
This study found that the emission of isoprene from trans-enic tobacco did not mitigate immediate plant stress to acuteV-B exposure. This contrasts with observations of enhanced
[
nce 226 (2014) 82–91
isoprene release alleviating the deleterious effects of thermalstress and ozone fumigation in the same transgenic tobacco lines.Irradiation of the leaf with UV-B stimulated the release of methanoland impeded PSII electron transport indicating substantial cell walland membrane damage within plant cells. However, photosyn-thetic rates and PSII performance in IE tobacco recovered in the 48 hfollowing exposure to UV-B, whereas NE plants exhibited contin-ued deleterious effects of UV-B indicative of longer-term damageand photosynthetic impairment. This may suggest that the func-tion of isoprene emission varies depending upon the type of stress,its duration and the extent of any damage to the photosyntheticapparatus. Enhanced levels of ASC and cycling of GSH in IE plantsmay have also contributed to recovery of photosynthetic perfor-mance following UV-B irradiation. This may indicate that both theantioxidant system and isoprene contributed to the recovery of thephotosynthetic process in IE plants. Overall, this study highlights apotential mechanism whereby isoprene emission assists in the sta-bilisation and repair of cell walls and membranes following UV-Binduced damage.
Acknowledgements
The authors would like to extend their sincere thanks to MarcoGiorgetti for his technical assistance in the field. The comments oftwo anonymous reviewers significantly improved this manuscript.This work has been funded by the Italian National Research Coun-cil (project RSTL-DG.RSTL.010.003), the Italian National ResearchCouncil and the Bulgarian Academy of Sciences scientific andtechnological agreement 2013-2014 and Ministero dell’Istruzione,dell’Universita e della Ricerca of Italy (PRIN 2010-2011, PRO-ROOT).
References
[1] J.C. Farman, B.G. Gardiner, J.D. Shanklin, Large losses of total ozone in Antarcticareveal seasonal ClOx/NOx interaction, Nature 315 (1985) 6016–6207.
[2] C.L. Ballaré, M.M. Caldwell, S.D. Flint, A. Robinson, J.F. Bornman, Effects ofsolar ultraviolet radiation on terrestrial ecosystems. Patterns, mechanisms,and interactions with climate change, Photochem. Photobiol. Sci. 10 (2011)226–241.
[3] G. Agati, M. Tattini, Multiple functional roles of flavonoids in photoprotection,New Phytol. 186 (2010) 786–793.
[4] F.R. Li, S.L. Peng, B.M. Chen, Y.P. Hou, A meta-analysis of the responses of woodyand herbaceous plants to elevated ultraviolet-B radiation, Acta Oecol. 36 (2010)1–9.
[5] C.E. Vickers, J. Gershenzon, M.T. Lerdau, F. Loreto, A unified mechanism ofaction for volatile isoprenoids in plant abiotic stress, Nat. Chem. Ecol. 5 (2009)283–291.
[6] F. Loreto, J.P. Schnitzler, Abiotic stresses and induced BVOCs, Trends Plant Sci.15 (2010) 154–166.
[7] E. Pallozzi, A. Fortunati, G. Marino, F. Loreto, G. Agati, M. Centritto, BVOC emis-sion from Populus × canadensis saplings in response to acute UV-A radiation,Physiol. Plant. 148 (2013) 51–61.
[8] L.O. Morales, R. Tegelberg, M. Brosché, M. Keinänen, A. Lindfors, P.J. Aphalo,Effects of solar UV-A and UV-B radiation on gene expression and phenolicaccumulation in Betula pendula leaves, Tree Physiol. 30 (2010) 923–934.
[9] L.O. Morales, R. Tegelberg, M. Brosché, A. Lindfors, S. Siipola, P.J. Aphalo, Tem-poral variation in epidermal flavonoids due to altered solar UV radiation ismoderated by the leaf position in Betula pendula, Physiol. Plant. 143 (2011)261–270.
10] F. Loreto, M. Mannozzi, C. Maris, P. Nascetti, F. Ferranti, S. Pasqualini, Ozonequenching properties of isoprene and its antioxidant role in leaves, PlantPhysiol. 126 (2001) 993–1000.
11] F. Loreto, P. Pinelli, F. Manes, H. Kollist, Impact of ozone on monoterpene emis-sions and evidence for an isoprene-like antioxidant action of monoterpenesemitted by Quercus ilex leaves, Tree Physiol. 24 (2004) 361–367.
12] G. Noctor, C. Foyer, Ascorbate and glutathione: keeping active oxygen undercontrol, Annu. Rev. Plant Physiol. 49 (1998) 249–279.
13] B. Aganchich, S. Wahbi, F. Loreto, M. Centritto, Partial root zone drying: reg-ulation of photosynthetic limitations and antioxidant enzymatic activities inyoung olive (Olea europaea) saplings, Tree Physiol. 29 (2009) 685–696.
14] C. Foyer, A. Trebst, G. Noctor, Signaling and integration of defense functions
of tocopherol, ascorbate and glutathione, in: B. Demmig-Adams, W.W. AdamsIII, A.K. Mattoo (Eds.), Photoprotection, Photoinhibition, Gene Regulation, andEnvironment, Springer, Dordrecht, The Netherlands, 2006, pp. 241–268.15] J. Llusia, L. Llorens, M. Bernal, D. Verdaguer, J. Penuelas, Effects of UV radiationand water limitation on the volatile terpene emission rates, photosynthesis

nt Scie
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[fluxes through time and their influence on plant diversity and speciation, J.
M. Centritto et al. / Pla
rates, and stomatal conductance in four Mediterranean species, Acta Physiol.Plant. 34 (2012) 757–769.
16] J.D. Blande, K. Turunen, J.K. Holopainen, Pine weevil feeding on Norway sprucebark has a stronger impact on needle VOC emissions than enhanced UV-Birradiation, Environ. Pollut. 157 (2009) 174–180.
17] P. Harley, G. Deem, S. Flint, M. Caldwell, Effects of growth under elevated UV-B on photosynthesis and isoprene emission in Quercus gambelii and Mucunapruriens, Global Change Biol. 2 (1996) 149–154.
18] P. Tiiva, R. Rinnan, P. Faubert, J. Räsänen, T. Holopainen, E. Kyrö, J.K. Holopainen,Isoprene emission from a subarctic peatland under enhanced UV-B radiation,New Phytol. 176 (2007) 346–355.
19] C.E. Vickers, M. Possell, C. Cojocariu, V.B. Velikova, J. Laothawornkitkul, A. Ryan,P.M. Mullineaux, C.N. Hewitt, Isoprene synthesis protects transgenic tobaccoplants from oxidative stress, Plant Cell Environ. 32 (2009) 520–531.
20] F. Magnani, M. Centritto, J. Grace, Measurement of apoplasmic and cell-to-cellcomponents of root hydraulic conductance by a pressure-clamp technique,Planta 199 (1996) 296–306.
21] M. Centritto, Photosynthetic limitations and carbon partitioning in cherry inresponse to water deficit and elevated [CO2], Agric. Ecosyst. Environ. 106 (2005)233–242.
22] G. Norsang, L. Kocbach, W. Tsoja, J. Stamnes, A. Dahlback, P. Nema, Ground-based measurements and modeling of solar UV-B radiation in Lhasa, Tibet,Atmos. Environ. 43 (2009) 1498–1502.
23] F. Brilli, T. Tsonev, T. Mahmood, V. Velikova, F. Loreto, M. Centritto, Ultra-dian variation of isoprene emission, photosynthesis, mesophyll conductanceand optimum temperature sensitivity for isoprene emission in water-stressedEucalyptus citriodora saplings, J. Exp. Bot. 64 (2013) 519–528.
24] W. Lindinger, A. Hansel, A. Jordan, Proton-transfer-reaction mass spectrome-try (PTR-MS): on-line monitoring of volatile organic compounds at pptv level,Chem. Soc. Rev. 27 (1998) 347–354.
25] U. Takahama, T. Oniki, Regulation of peroxidase-dependent oxidation of pheno-lics in the apoplast of spinach leaves by ascorbate, Plant Cell Physiol. 33 (1992)379–387.
26] Y. Nakano, K. Asada, Hydrogen peroxide is scavenged by ascorbate-specificperoxidase in spinach chloroplasts, Plant Cell Physiol. 22 (1981) 867–880.
27] I.E. Galbally, W. Kirstine, The production of methanol by flowering plants andthe global cycle of methanol, J. Atmos. Chem. 43 (2002) 195–229.
28] A. Szilárd, L. Sass, Z. Deák, I. Vass, The sensitivity of photosystem II to damage byUV-B radiation depends on the oxidation state of the water-splitting complex,BBA-Bioenergetics 1767 (2007) 876–882.
29] M.A.K. Jansen, V. Gaba, B.M. Greenberg, Higher plants and UV-B radiation:
balancing damage, repair and acclimation, Trends Plant Sci. 3 (1998) 131–135.30] A. Melis, J.A. Nemson, M.A. Harrison, Damage to functional components andpartial degradation of photosystem II reaction center proteins upon chloro-plast exposure to ultraviolet-B radiation, Biochim. Biophys. Acta 1100 (1992)312–320.
[
nce 226 (2014) 82–91 91
31] É. Hideg, C. Barta, T. Kálai, I. Vass, K. Hideg, K. Asada, Detection ofsinglet oxygen and superoxide with fluorescent sensors in leaves understress by photoinhibition or UV radiation, Plant Cell Physiol. 43 (2002)1154–1164.
32] P. Carletti, A. Masi, A. Wonisch, D. Grill, M. Tausz, M. Ferretti, Changes in antioxi-dant and pigment pool dimensions in UV-B irradiated maize seedlings, Environ.Exp. Bot. 50 (2003) 149–157.
33] S. Munné-Bosch, L. Alegre, Interplay between ascorbic acid and lipophilicantioxidant defenses in chloroplasts of water-stressed Arabidopsis plants, FEBSLett. 524 (2002) 145–148.
34] A. Schmidt, K.J. Kunert, Lipid peroxidation in higher plants the role of glu-tathione reductase, Plant Physiol. 82 (1986) 700–702.
35] N.J. Stoke, G.M. Terry, C.N. Hewitt, The impact of ozone, isoprene and propeneon antioxidant levels in two leaf classes of velvet bean (Mucuna pruriens L.), J.Exp. Bot. 49 (1998) 115–123.
36] M.E. Siwko, S.J. Marrink, A.H. de Vries, A. Kozubek, A.J. Schoot Uiterkamp, A.E.Mark, Does isoprene protect plant membranes from thermal shock? A molec-ular dynamics study, BBA-Biomembranes 1768 (2007) 198–206.
37] T.D. Sharkey, E.L. Singsaas, Why plants emit isoprene, Nature 374 (1995)769.
38] A.B. Britt, Repair of DNA damage induced by ultraviolet radiation, Plant Physiol.108 (1995) 891–896.
39] F. Loreto, V. Velikova, Isoprene produced by leaves protects the photosyntheticapparatus against ozone damage, quenches ozone products, and reduces lipidperoxidation of cellular membranes, Plant Physiol. 127 (2001) 1781–1787.
40] T.D. Sharkey, X. Chen, S. Yeh, Isoprene increases thermotolerance offosmidomycin-fed leaves, Plant Physiol. 125 (2001) 2001–2006.
41] F. Loreto, M. Centritto, Leaf carbon assimilation in a water limited world, PlantBiosyst. 142 (2008) 154–161.
42] S. Reuber, J.F. Bornman, G. Weissenböck, A flavonoid mutant of barley (Hordeumvulgare L.) exhibits increased sensitivity to UV-B radiation in the primary leaf,Plant Cell Environ. 19 (1996) 593–601.
43] Y.Y. He, D.P. Häder, UV-B-induced formation of reactive oxygen species andoxidative damage of the cyanobacterium Anabaena sp.: protective effects ofascorbic acid and N-acetyl-l-cysteine, J. Photochem. Photobiol. B: Biol. 66(2002) 115–124.
44] D.J. Beerling, M. Harfoot, B. Lomax, J.A. Pyle, The stability of the stratosphericozone layer during the end-Permian eruption of the Siberian Traps, Philos. T.Roy. Soc. A 365.1856 (2007) 1843–1866.
45] K.J. Willis, K.D. Bennett, H.J.B. Birks, Variability in thermal and UV-B energy
Biogeogr. 36 (2009) 1630–1644.46] J.A. Lake, K.J. Field, M.P. Davey, D.J. Beerling, B.H. Lomax, Metabolomic and phys-
iological responses reveal multi-phasic acclimation of Arabidopsis thaliana tochronic UV radiation, Plant Cell Environ. 32 (2009) 1377–1389.