j.1570-7458.2009.00831.x
TRANSCRIPT

8/8/2019 j.1570-7458.2009.00831.x
http://slidepdf.com/reader/full/j1570-7458200900831x 1/7
© 2009 The Authors Entomologia Experimentalis et Applicata 131: 99–105, 2009
Journal compilation © 2009 The Netherlands Entomological Society 99
DOI: 10.1111/j.1570-7458.2009.00831.x
BlackwellPublishingLtd
TECHNICAL NOTE
Anti-metabolic effects of Galanthus nivalis agglutinin
and wheat germ agglutinin on nymphal stages of
the common brown leafhopper using a novel artificial
diet system
Piotr Tr
é
bicki
1,2
, Rob M. Harding
1
& Kevin S. Powell
2
*
1
Centre for Tropical Crops and Biocommodities, Queensland University of Technology, Queensland, GPO Box 2434,
Brisbane, 4001, Australia, and 2
Department of Primary Industries, Biosciences Research Division, Rutherglen,
Victoria, 3685, Australia
Accepted: 6 January 2009
Key words: lectin, GNA, WGA, cowpea trypsin inhibitor, bioassay, Orosius orientalis
, vector,
oviposition, Homoptera, Cicadellidae
Introduction
The common brown leafhopper, Orosius orientalis
(Matsumura) (Homoptera: Cicadellidae), previously
described as Orosius argentatus
(Evans), is an important
vector of several viruses and phytoplasmas worldwide.
In Australia, phytoplasmas vectored by O. orientalis
cause
a range of economically important diseases, including
legume little leaf (Hutton & Grylls, 1956), tomato big bud
(Osmelak, 1986), lucerne witches broom (Helson, 1951),
potato purple top wilt (Harding & Teakle, 1985), and
Australian lucerne yellows (Pilkington et al., 2004).
Orosius orientalis
also transmits Tobacco yellow dwarf
virus (TYDV; genus Mastrevirus, family Geminiviridae) to
beans, causing bean summer death disease (Ballantyne,
1968), and to tobacco, causing tobacco yellow dwarf
disease (Hill, 1937, 1941). TYDV has only been recorded
in Australia to date. Both diseases result in significant
production and quality losses (Ballantyne, 1968; Thomas,
1979; Moran & Rodoni, 1999). Although direct damage
caused by leafhopper feeding has been observed, it is
relatively minor compared to the losses resulting from
disease (P Tr
E
bicki, unpubl.).
Control strategies for O. orientalis
are primarily based
on the use of chemical insecticides (Paddick et al., 1971;Paddick & French, 1972; Osmelak, 1986). However, due to
the paucity of information available on the life-cycle, popu-
lation dynamics, and disease transmission characteristics
of this leafhopper, the use of insecticides has been largely
ineffective. An alternative approach to controlling sap-
sucking insect pests is the use of anti-metabolites, such as
plant lectins expressed in genetically modified plants
(Gatehouse et al., 1992; Peumans & Van Damme, 1995).
Using artificial diet bioassay systems, a wide range of plant
lectins, including Galanthus nivalis
agglutinin (GNA) and
wheat germ agglutinin (WGA), have been shown to exhibit
anti-metabolic effects against many economically impor-
tant homopteran pests resulting in reduced survival and
delayed development (Habibi et al., 1993; Powell et al.,
1993; Gatehouse et al., 1995; Chen, 2008). Furthermore,transgenic plants expressing lectins have shown enhanced
resistance towards many sap-sucking insects, including the
rice brown planthopper, Nilaparvata lugens
(Stål) and
green leafhopper, Nephotettix
spp. (Peferoen, 1997;
Gatehouse & Gatehouse, 1998; Jouanin et al., 1998). Based
on these studies, the use of anti-metabolites may also
potentially be an effective strategy to control O. orientalis
.
A prerequisite to preliminary screening of potential
anti-metabolites in vitro is the development of a chemically
defined artificial diet for the target pest. The basic
nutritional requirements of O. orientalis
are poorly under-
stood and, although artificial diets for several Cicadellidae
have been developed (Singh, 1977; Cohen, 2004), a diet ordiet feeding system has not been reported for O. orientalis
.
In this study, we describe the development of the first arti-
ficial diet bioassay system for rearing O. orientalis in vitro
and a simple oviposition chamber for collection of newly
emerged first instars. In addition, we describe the use of
this diet bioassay system to assess the anti-metabolic effects
of two plant lectins and a protease inhibitor towards first
instars of the leafhopper.
*
Correspondence: Kevin S. Powell, Department of Primary
Industries, Biosciences Research Division, RMB 1145, Chiltern Valley
Road, Rutherglen, Victoria, 3685, Australia.
E-mail: [email protected]

8/8/2019 j.1570-7458.2009.00831.x
http://slidepdf.com/reader/full/j1570-7458200900831x 2/7
100
Tr
É
bicki et al
.
Materials and methods
Insect culture
Stock colonies of O. orientalis
were obtained from Charles
Darwin University, Darwin, Australia, and from herbaceous
vegetation surrounding commercial tobacco farms in
the Ovens Valley, north-east Victoria, Australia (36
°
37
′
S,
146
°
48
′
E). Leafhoppers were identified to species level
using morphological characteristics of the male genitalia
(Evans, 1966; Ghauri, 1966; Fletcher, 2000). Cultures were
maintained, on 30–40-day-old celery [
Apium graveolens
L. (Apiaceae)] or bean [
Phaseolus vulgaris
L. (Fabaceae)]
plants grown under glasshouse conditions, for eight
generations before fecund female adults were transferred
to oviposition chambers and reared under controlled
environment conditions (25 ± 2 °
C, L14:D10). The
oviposition chambers (Figure 1) were made by modifying
standard plastic Petri dishes (9 ×
2.5 cm). Three access holes
on opposite sides of the dish base were made. A 3–4-cmdiameter opening was made in the lid and sealed with
glued mesh to prevent first-instar escape, facilitate air
circulation, and reduce condensation. One attached leaf of
the host plant was placed in the dish base with the stem
inserted into the access hole, covered with the lid and
sealed with an elastic band. The two remaining access holes
allowed introduction of fecund adult females and removal
of newly emerged first instars. To prevent insect escape
during oviposition or post-hatching, foam was wrapped
around the stem of the leaf and two foam stoppers were
placed into each access hole. Orosius orientalis
oviposits on
the leaf blade, petiole, or stem (Helson, 1942), and using
this chamber design fecund females could freely choose
oviposition sites.
Chemicals and materials
Galanthus nivalis
agglutinin and WGA were obtained from
Abacus Australian Laboratory Supplies, East Brisbane,
Australia. DL-homoserine was purchased from MP
Biomedicals Australasia, Seven Hills, Australia. Cowpea
trypsin inhibitor (CpTi) and all remaining dietary
components were obtained from Sigma-Aldrich, Castle
Hill, Australia. All chemicals utilized in artificial diet
preparation had the highest purity commercially available.
Artificial diet preparation
Two artificial diets were prepared for rearing nymphal
stages of O. orientalis
, MED-1 as previously utilized for thesmaller brown leafhopper, Laodelphax striatellus
Fallen
(Mitsuhashi, 1974), and a modification of MED-1
designated PT-07. Dietary modifications were made to
the amino acid and vitamin profile and concentration, and
by the addition of cholesteryl benzoate (Table 1). All
components were dissolved in sterile ultra pure water
using gentle heat (25 °
C), and the pH was adjusted to 6.5
with 1m
potassium hydroxide. After filtration through a
0.2-
μ
m Millipore disposable filter, diet solutions were
dispensed into 50-ml plastic containers as stock solutions,
and further dispensed into 1.5-ml Eppendorf tubes as
working solutions and stored at –20°
C prior to use.
Feeding trials
The effect of artificial diets, MED-1, and PT-07 on the
development and survival of O. orientalis
was examined
using feeding chambers essentially as described by Powell
et al. (1993), but with a minor modification. The feeding
chamber was placed in a second, larger plastic Petri dish
(9 ×
2.5 cm) containing wet filter paper to increase and
maintain constant humidity (Figure 2). Five newly
emerged first instars of O. orientalis
were removed from
oviposition chambers with a fine wet paint brush and
placed in the feeding chamber (plastic Petri dish,
1 ×
3.5 cm). A single layer of stretched Parafilm M™ wasplaced over the chamber and 100 μ
l of diet was deposited
on the membrane. A second layer of Parafilm M™ was then
stretched over the artificial diet to form a feeding sachet.
Two controls (no diet and water only) were included in
all experiments and 10 replicates were used for each
treatment and control.
To examine the effect of anti-metabolites on the survival
of O. orientalis
, GNA, WGA, and CpTi were separately
Figure 1 Oviposition chamber for Orosius orientalis on (A) host
plant with (B) rubber band closure, (C) fabric mesh ventilation
point, (D) Petri dish, (E) access points for nymph removal and
adult addition with foam plugs to prevent insect escape, and
(F) leaf attached to whole bean or celery host plant.

8/8/2019 j.1570-7458.2009.00831.x
http://slidepdf.com/reader/full/j1570-7458200900831x 3/7
Screening anti-metabolites for leafhopper management
101
incorporated into the PT-07 artificial diet at a concentra-
tion of 0.1% (wt/vol). Five newly emerged first instars
of O. orientalis
were used in each feeding chamber as
described above. ‘No diet’ and PT-07 diet without theinclusion of a plant protein were included as controls. Ten
replicates were used for each treatment and the control.
In all feeding trials, leafhopper survival data were
recorded daily and diets were replaced on alternative days.
To avoid insect escape during the diet changing procedure,
insects were temporarily immobilized by placing the
feeding chamber at –20 °
C for 90 s. To avoid fungal growth
on excreted honeydew, feeding chambers were replaced
weekly under aseptic conditions. Parafilm M™ and feeding
chambers were exposed to ultraviolet light for 20 min
prior to insect introduction. All trials were conducted in
controlled growth rooms (25 ± 2 °
C, L14:D10).
Statistical analysis
Statistical analysis was performed using GenStat software
(10th Edition
©
2007, Lawes Agricultural Trust). A Kaplan–
Meier estimate and log-rank test were used to determinesurvivor distribution and to compare differences between
diet formulations and diets with anti-metabolic compounds.
Corrected mortality calculations (Abbott, 1925) were used
to compare the relative efficacy of treatments.
Results and discussion
Modifications were made to the MED-1 (Mitsuhashi,
1974) diet formulation resulting in the development of a
novel artificial diet PT-07. These modifications included
changing the proportions of amino acids (L-arginine
hydrochloride, L-asparagine, L-glutamic acid, glycine,
Table 1 Composition (mg l−1) of artificial diets, PT-07 and MED-11, used for rearing Orosius orientalis; pH of both diets was adjusted with
KOH to 6.5
Ingredient PT-07 MED-1 Ingredient PT-07 MED-1
L-alanine 1 000 1 000 MgCl2·6H2O 2 000 2 000
γ -amino butyric acid 200 200 KH2PO4 5 000 5 000
L-arginine hydrochloride 3 000 4 000 CaCl2·2H2O 32 31.15
L-asparagine 4 000 3 000 CuCl2·2H2O 3 2.68
L-aspartic acid 1 000 1 000 FeCl3·6H2O 23 22.28
L-cysteine 500 500 MnCl2·4H2O 8 7.93
L-cystine hydrochloride 50 ZnCl2 5 3.96
L-glutamic acid 1 500 2 000
L-glutamine 6 000 6 000 Biotin 1 1
Glycine 400 200 Calcium pantothenate 50 50
L-histidine 1 500 2 000 Choline chloride 500 500
DL-homoserine 8 000 Folic acid 10 10
L-isoleucine 1 500 2 000 Inositol 500 500
L-leucine 1 500 2 000 Nicotinic acid 100 100
L-lysine hydrochloride 1 800 2 000 Pyridoxine hydrochloride 25 25
L-methionine 1 500 1 000 Riboflavin 25 50L-phenylalanine 1 000 1 000 Thiamine hydrochloride 25 25
L-proline 1 000 1 000 Ascorbic acid 1 000
L-serine 1 000 1 000 Sodium ascorbate 1 000
L-threonine 1 500 2 000 Cholesteryl benzoate 25
L-tryptophane 1 000 1 000
L-tyrosine 200 200 Sucrose 50 000 50 000
L-valine 1 500 2 000
1Source: Mitsuhashi, 1974.
Figure 2 Modified feeding chamber for
rearing Orosius orientalis on liquid diet
through a double layer of Parafilm M™.

8/8/2019 j.1570-7458.2009.00831.x
http://slidepdf.com/reader/full/j1570-7458200900831x 4/7
102
Tr
É
bicki et al
.
L-isoleucine, L-leucine, L-lysine hydrochloride, L-
methionine, L-threonine, and L-valine), inorganic salts,
and including cholesteryl benzoate and ascorbic acid,
which resulted in enhanced survival and development of
O. orientalis.
PT-07 artificial diet was significantly more effective for
rearing O. orientalis
than the MED-1 diet (Figure 3),
increasing development and survival. In the absence of
diet, using either no-diet or water-only controls, insects
survived for a maximum of 5 days. In contrast, the survival
of O. orientalis
on the PT-07 diet was significantly
enhanced compared to the MED-1 diet and the controls
(log-rank test: P<0.001). Insects reared on the MED-1 diet
survived for a maximum of 14 days whereas the PT-07 diet
sustained insects for up to 46 days reaching adulthood at
day 42–45. When 100% mortality of leafhoppers was
reached on MED-1 diet, more than 80% of insects
remained alive on the PT-07 diet. Insects reared on MED-1did not develop beyond the third instar (with a large
proportion not surviving ecdysis (data not presented),
whereas PT-07 supported leafhopper development to the
adult stage. On host plants leafhopper survival and develop-
ment varies depending on plant species as this insect is
polyphagous with up to 20 confirmed host plant species
(P Tr
E
bicki, unpubl.). Under laboratory conditions, on
average it takes 25 days till nymphs reach adulthood when
reared on Malva parviflora
L. (Helson, 1942) and up to
35 days on celery (P Tr
E
bicki, unpubl.).
Liquid artificial diets for rearing homopteran pests
have been used for decades (Vanderzant, 1974) along with
a feeding sachet system for delivery which was first
developed for leafhoppers (Carter, 1927). Artificial diets,
which enable the rearing of Cicadellidae and Delphacidae
through successive generations (Mitsuhashi & Koyama,
1972; Mitsuhashi, 1974), have been developed but none
prior to this study was available for O. orientalis
. Optimi-
zation of the concentration and proportion of amino acids
in a diet is an important factor for leafhopper development
and survival as they are known to have phagostimulatory
and phagoinhibitory properties (Sogawa, 1977). Some
leafhoppers also require a source of cholesterol for optimal
development (Lin & Hou, 1981). A number of amino
acids, including L-asparagine, were proven to act as a
sucking stimulant for N. lugens
(Sogawa, 1972) and the
increased concentration of this amino acid may haveimproved survival of O. orientalis
on PT-07 diet.
First instars of O. orientalis
were also exposed to the
PT-07 diet containing 0.1% (wt/vol) concentrations of either
GNA, WGA, or CpTi, with PT-07 diet only and no diet
used as a control. Using the PT-07 control only diet, some
insects were still alive after 43 days. Cowpea trypsin inhibitor
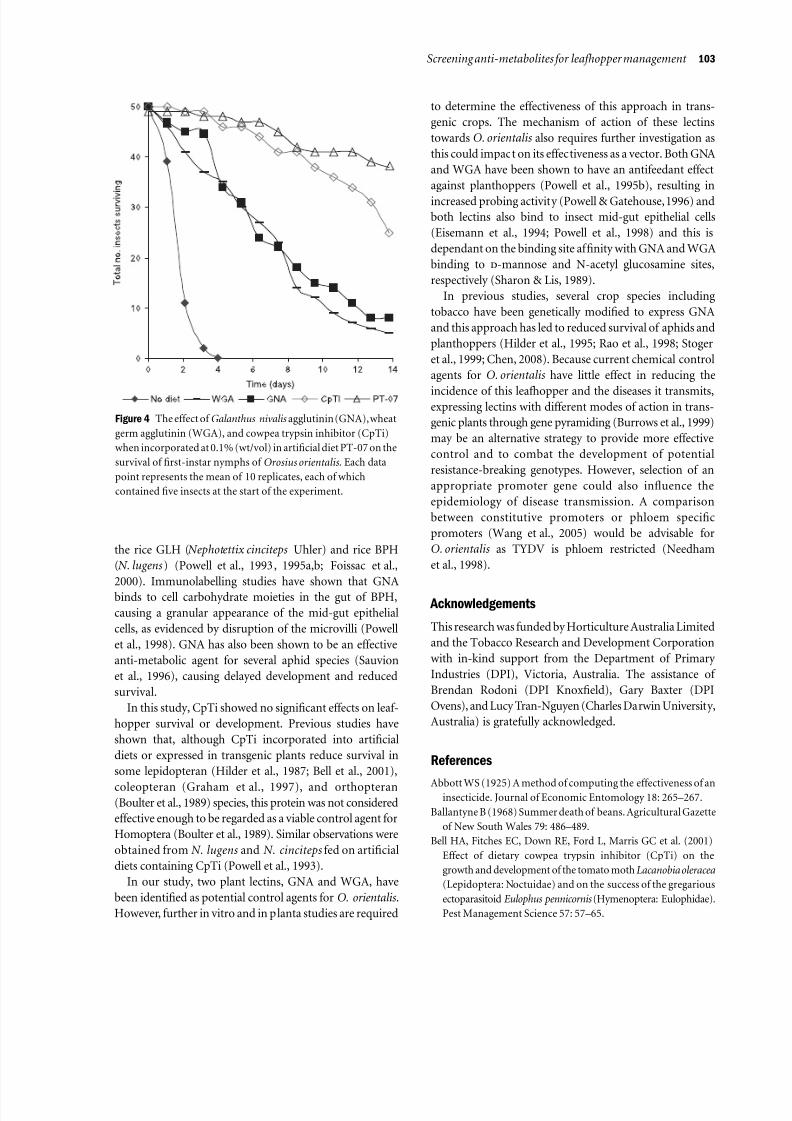
showed no significant effects on leafhopper survival or
development (Figure 4) with a corrected mortality of only
4%. In contrast, GNA and WGA both showed significant
anti-metabolic effects (log-rank test: P<0.001), with
nymphal survival reduced to 22 and 15 days, respectively.
Although the corrected mortality values using WGA andGNA were relatively high at 37 and 35%, respectively and
significantly different to CpTi treatment (log-rank test:
P<0.001), they were not statistically different from one
another (P>0.1). These mortality levels, although lower
than those reported using brown planthopper (BPH)
N. lugens
(up to 76%) (Powell et al., 1993), were nonethe-
less significant. The comparatively broad host plant range
of O. orientalis
(Helson, 1942) may partially explain the
difference in mortality compared to N. lugens
, which is
monophagus on rice. Other studies have shown that
more GNA binds to N. lugens
gut tissue compared to
corresponding tissue of the rice green leafhopper (GLH),
Nephotettix
spp., despite the fact that GLH ingested moreplant sap from GNA-transformed rice plants (Foissac
et al., 2000).
Previous studies, using either an in vitro artificial diet
bioassay system for screening anti-metabolites or by
expressing proteins in transgenic plants, have shown that a
number of plant-derived compounds affect the development
of a range of Homoptera. GNA and WGA, for example,
have shown significant anti-metabolic effects towards
Figure 3 The effect of two artificial diet formulations, MED-1
and PT-07, on the survival and development of first instars of
Orosius orientalis. Each data point represents the mean of 10
replicates, each of which contained five insects at the start of the
experiment.
8/8/2019 j.1570-7458.2009.00831.x
http://slidepdf.com/reader/full/j1570-7458200900831x 5/7
Screening anti-metabolites for leafhopper management
103
the rice GLH (
Nephotettix cinciteps
Uhler) and rice BPH
(
N. lugens
) (Powell et al., 1993, 1995a,b; Foissac et al.,
2000). Immunolabelling studies have shown that GNAbinds to cell carbohydrate moieties in the gut of BPH,
causing a granular appearance of the mid-gut epithelial
cells, as evidenced by disruption of the microvilli (Powell
et al., 1998). GNA has also been shown to be an effective
anti-metabolic agent for several aphid species (Sauvion
et al., 1996), causing delayed development and reduced
survival.
In this study, CpTi showed no significant effects on leaf-
hopper survival or development. Previous studies have
shown that, although CpTi incorporated into artificial
diets or expressed in transgenic plants reduce survival in
some lepidopteran (Hilder et al., 1987; Bell et al., 2001),
coleopteran (Graham et al., 1997), and orthopteran(Boulter et al., 1989) species, this protein was not considered
effective enough to be regarded as a viable control agent for
Homoptera (Boulter et al., 1989). Similar observations were
obtained from N. lugens
and N. cinciteps
fed on artificial
diets containing CpTi (Powell et al., 1993).
In our study, two plant lectins, GNA and WGA, have
been identified as potential control agents for O. orientalis
.
However, further in vitro and in planta studies are required
to determine the effectiveness of this approach in trans-
genic crops. The mechanism of action of these lectins
towards O. orientalis
also requires further investigation as
this could impact on its effectiveness as a vector. Both GNA
and WGA have been shown to have an antifeedant effect
against planthoppers (Powell et al., 1995b), resulting in
increased probing activity (Powell & Gatehouse, 1996) and
both lectins also bind to insect mid-gut epithelial cells
(Eisemann et al., 1994; Powell et al., 1998) and this is
dependant on the binding site affinity with GNA and WGA
binding to d
-mannose and N-acetyl glucosamine sites,
respectively (Sharon & Lis, 1989).
In previous studies, several crop species including
tobacco have been genetically modified to express GNA
and this approach has led to reduced survival of aphids and
planthoppers (Hilder et al., 1995; Rao et al., 1998; Stoger
et al., 1999; Chen, 2008). Because current chemical control
agents for O. orientalis
have little effect in reducing the
incidence of this leafhopper and the diseases it transmits,expressing lectins with different modes of action in trans-
genic plants through gene pyramiding (Burrows et al., 1999)
may be an alternative strategy to provide more effective
control and to combat the development of potential
resistance-breaking genotypes. However, selection of an
appropriate promoter gene could also influence the
epidemiology of disease transmission. A comparison
between constitutive promoters or phloem specific
promoters (Wang et al., 2005) would be advisable for
O. orientalis
as TYDV is phloem restricted (Needham
et al., 1998).
Acknowledgements
This research was funded by Horticulture Australia Limited
and the Tobacco Research and Development Corporation
with in-kind support from the Department of Primary
Industries (DPI), Victoria, Australia. The assistance of
Brendan Rodoni (DPI Knoxfield), Gary Baxter (DPI
Ovens), and Lucy Tran-Nguyen (Charles Darwin University,
Australia) is gratefully acknowledged.
References
Abbott WS (1925) A method of computing the effectiveness of aninsecticide. Journal of Economic Entomology 18: 265–267.
Ballantyne B (1968) Summer death of beans. Agricultural Gazette
of New South Wales 79: 486–489.
Bell HA, Fitches EC, Down RE, Ford L, Marris GC et al. (2001)
Effect of dietary cowpea trypsin inhibitor (CpTi) on the
growth and development of the tomato moth Lacanobia oleracea
(Lepidoptera: Noctuidae) and on the success of the gregarious
ectoparasitoid Eulophus pennicornis (Hymenoptera: Eulophidae).
Pest Management Science 57: 57–65.
Figure 4 The effect of Galanthus nivalisagglutinin (GNA), wheat
germ agglutinin (WGA), and cowpea trypsin inhibitor (CpTi)
when incorporated at 0.1% (wt/vol) in artificial diet PT-07 on the
survival of first-instar nymphs of Orosius orientalis. Each data
point represents the mean of 10 replicates, each of which
contained five insects at the start of the experiment.

8/8/2019 j.1570-7458.2009.00831.x
http://slidepdf.com/reader/full/j1570-7458200900831x 6/7
104 Tr É bicki et al.
Boulter D, Gatehouse AMR & Hilder VA (1989) Use of cowpea
trypsin inhibitor (CpTi) to protect plants against insect
predation. Biotechnology Advances 7: 489–497.
Burrows PR, Barker ADP, Newell CA & Hamilton WDO (1999)
Plant-derived enzyme inhibitors and lectins for resistance
against plant-parasitic nematodes in transgenic crops. Pesticide
Science 52: 176–183.Carter W (1927) A technique for use with homopterous vectors
of plant diseases, with special reference to the sugar beet leaf-
hopper, Eutettix tenallus (Baker). Journal of Agricultural
Research 34: 449–451.
Chen M-S (2008) Inducible direct plant defense against insect
herbivores: a review. Insect Science 15: 101–114.
Cohen AC (2004) Insect Diets: Science and Technology. CRC
Press, Boca Raton, FL, USA.
Eisemann CH, Donaldson RA, Pearson RD, Cadagon LC,
Vuocolo T & Tellam RL (1994) Larvicidal activity of lectins
on Lucilia cuprina: mechanism of action. Entomologia
Experimentalis et Applicata 72: 1–11.
Evans JW (1966) The leafhoppers and froghoppers of Australia
and New Zealand (Homoptera: Cicadelloidea and Cercopoidea).
The Australian Museum Memoir XII: 1–348.
Fletcher MJ (2000) Identification keys and checklists for the
leafhoppers, planthoppers and their relatives occurring in
Australia and New Zealand (Hemiptera: Auchenorrhyncha).
NSW DPI, Orange, New South Wales, Australia. Available at
http://www.agric.nsw.gov.au/Hort/ascu/start.htmOrange-
AgriculturalInstitute.
Foissac X, Thi Loc N, Christou P, Gatehouse AMR & Gatehouse JA
(2000) Resistance to green leafhopper ( Nephotettix virescens)
and brown planthopper ( Nilaparvata lugens ) in transgenic rice
expressing snowdrop lectin (Galanthus nivalis agglutinin;
GNA). Journal of Insect Physiology 46: 573–583.
Gatehouse AMR & Gatehouse JA (1998) Identifying proteinswith insecticidal activity: use of encoding genes to produce
insect-resistant transgenic crops. Pesticide Science 52: 165–
175.
Gatehouse AMR, Boulter D & Hilder VA (1992) Potential of
plant-derived genes in the genetic manipulation of crops for
insect resistance. Plant Genetic Manipulation for Crop Protection
(ed. by AMR Gatehouse, D Boulter & VA Hilder), pp. 135–153.
CAB International, Wallingford, UK.
Gatehouse AMR, Powell KS, Peumans WJ, Van Damme EJM &
Gatehouse JA (1995) Insecticidal properties of plant lectins:
their potential in plant protection. Lectins: Biomedical Per-
spectives (ed. by AJ Pusztai & S Bardocz), pp. 35–57. Taylor &
Francis, London, UK.
Ghauri MSK (1966) Revision of the genus Orosius Distant
(Homoptera: Cicadelloidea). Bulletin of the British Museum
(Natural History), Entomology 18: 231–252.
Graham J, Gordon SC & McNicol RJ (1997) The effect of the
CpTi gene in strawberry against attack by vine weevil (Otio-
rhynchus sulcatus F. Coleoptera: Curculionidae). Annals of
Applied Biology 131: 133–139.
Habibi J, Backus EA & Czapla TH (1993) Plant lectins affect
survival of the potato leafhopper (Homoptera: Cicadellidae).
Journal of Economic Entomology 86: 945–951.
Harding RM & Teakle DS (1985) Mycoplasma-like organisms as
causal agents of potato purple top wilt in Queensland. Australian
Journal of Agricultural Research 36: 443–449.
Helson GAH (1942) The leafhopper Thamnotettix argentata
Evans, a vector of tobacco yellow dwarf. Journal of Council of
Scientific and Industrial Research 15: 175–184.
Helson GAH (1951) The transmission of witches broom virusdisease of lucerne by the common brown leafhopper, Orosius
argentatus (Evans). Biological Science 4: 115–124.
Hilder VA, Gatehouse AMR, Sheerman SE, Barker RF & Boulter
D (1987) A novel mechanism of insect resistance engineered
into tobacco. Nature 330: 160–163.
Hilder VA, Powell KS, Gatehouse AMR, Gatehouse JA, Gatehouse
LN et al. (1995) Expression of snowdrop lectin in transgenic
tobacco plants results in added protection against aphids.
Transgenic Research 4: 18–25.
Hill AV (1937) Yellow dwarf of tobacco in Australia, I. Symptoms.
Journal of Council of Scientific and Industrial Research 10:
228–230.
Hill AV (1941) Yellow dwarf of tobacco in Australia, II. Transmis-
sion by the jassid Thamnotettix argentata (Evans). Journal of
Council of Scientific and Industrial Research 14: 181–186.
Hutton EM & Grylls NE (1956) Legume ‘little leaf’, a virus disease
of subtropical pasture species. Australian Journal of Agri-
cultural Research 7: 85–97.
Jouanin L, Bonade-Bottino M, Girard C, Morrot G & Giband M
(1998) Transgenic plants for insect resistance. Plant Science
131: 1–11.
Lin LC & Hou RF (1981) Dietary requirements of Nephotettix
cinciteps for amino acids and cholesterol. Chinese Journal of
Entomology 1: 41–53.
Mitsuhashi J (1974) Methods for rearing leafhoppers and plant-
hoppers on artificial diets. Review of Plant Protection Research
7: 57–67.Mitsuhashi J & Koyama K (1972) Artificial rearing of the smaller
brown planthopper, Laodelphax striatellus Fallén, with special
reference to rearing conditions for the first instar nymphs.
Japanese Journal of Applied Entomology and Zoology 16: 8–17.
Moran JR & Rodoni B (1999) Strategies for the Control of Tobacco
Yellow Dwarf Virus. Report on a consultancy conducted on
behalf of the Tobacco Corporation of Victoria, Australia.
Needham PD, Atkinson RG, Morris BAM, Gardner RC & Gleave
AP (1998) GUS expression patterns from a tobacco yellow dwarf
virus-based episomal vector. Plant Cell Reports 17: 631–639.
Osmelak JA (1986) Assessment of Various Insecticides for the
Control of the Vector Orosius argentatus (Evans) (Homoptera:
Cicadellidae) and Tomato Big Bud Diseases; Research Report.
Plant Research Institute, Department of Agricultural and
Rural Affairs, Burnley, Victoria, Australia.
Paddick RG & French FL (1972) Suppression of tobacco yellow
dwarf with systematic organophosphorus insecticide. Australian
Journal of Experimental Agriculture and Animal Husbandry
12: 331–334.
Paddick RG, French FL & Turner PL (1971) Control of leafhopper-
borne plant disease possibly due to direct action of systemic
biocides on mycoplasmas. Plant Disease Reporter 55: 291–
293.

8/8/2019 j.1570-7458.2009.00831.x
http://slidepdf.com/reader/full/j1570-7458200900831x 7/7
Screening anti-metabolites for leafhopper management 105
Peferoen M (1997) Insect control with transgenic plants expressing
Bacillus thuringiensis crystal proteins. Advances in Insect
Control: The Role of Transgenic Plants (ed. by N Carozzi & M
Koziel), pp. 21–48. Taylor & Francis, London, UK.
Peumans WJ & Van Damme EJM (1995) Lectins as plant defence
proteins. Plant Physiology 109: 347–352.
Pilkington LJ, Gurr GM, Fletcher MJ, Nikandrow A & Elliott E(2004) Vector status of three leafhopper species for Australian
lucerne yellows phytoplasma. Australian Journal of Entomology
42: 366–373.
Powell KS & Gatehouse JA (1996) Mechanism of mannose-
binding snowdrop lectin for use against brown planthopper in
rice. Rice Genetics III: Proceedings of the Third International
Rice Genetics Symposium (ed. by GS Khush), pp. 753–758.
International Rice Research Institute, Manila, Philippines.
Powell KS, Gatehouse AMR, Hilder VA & Gatehouse JA (1993)
Antimetabolic effects of plant lectins and plant and fungal
enzymes on the nymphal stages of two important rice pests,
Nilaparvata lugens and Nephotettix cinciteps. Entomologia
Experimentalis et Applicata 66: 119–126.
Powell KS, Gatehouse AMR, Hilder VA & Gatehouse JA (1995a)
Antifeedant effects of plant lectins and an enzyme on the adult
stage of the rice brown planthopper, Nilaparvata lugens. Ento-
mologia Experimentalis et Applicata 75: 51–59.
Powell KS, Gatehouse AMR, Hilder VA, Van Damme EJM,
Peumans WJ et al. (1995b) Different antimetabolic effects of
related lectins towards nymphal stages of Nilaparvata lugens.
Entomologia Experimentalis et Applicata 75: 61–65.
Powell KS, Spence J, Bharathi M, Gatehouse JA & Gatehouse
AMR (1998) Immunohistochemical and developmental studies
to elucidate the mechanism of action of the snowdrop lectin on
the rice brown planthopper, Nilaparvata lugens (Stål). Journal
of Insect Physiology 44: 529–539.
Rao KV, Rathore KS, Hodges TK, Fu X, Stoger E et al. (1998)
Expression of snowdrop lectin (GNA) in transgenic rice plants
confers resistance to rice brown planthopper. The Plant Journal
15: 469–477.
Sauvion N, Rahbé Y, Peumans WJ, Van Damme EJM, Gatehouse
JA & Gatehouse AMR (1996) Effects of GNA and other
mannose binding lectins on development and fecundity of the peach potato aphid Myzus persicae. Entomologia Experi-
mentalis et Applicata 79: 285–293.
Sharon N & Lis H (1989) Lectins. Chapman & Hall, London, UK.
Singh P (1977) Artificial Diets for Insects, Mites, and Spiders.
IFI / Plenum, New York, NY, USA.
Sogawa K (1972) Studies on the feeding habits of the brown plant-
hopper. III. Effects of amino acids and other compounds on
the sucking response. Japanese Journal of Applied Entomology
and Zoology 16: 1–7.
Sogawa K (1977) Feeding physiology of the brown planthopper.
The Rice Brown Planthopper, pp. 95–116. Food and Fertilizer
Technology Center for the Asia and Pacific Region, ASPAC,
Taipei, Taiwan.
Stoger E, Williams S, Christou P, Down RE & Gatehouse JA
(1999) Expression of the insecticidal lectin from snowdrop
(Galanthus nivalis agglutinin; GNA) in transgenic wheat
plants: effects on predation by the grain aphid Sitobion avenae.
Molecular Breeding 5: 65–73.
Thomas JE (1979) Tobacco Yellow Dwarf and Bean Summer Death
Viruses. Phd Dissertation. University of Sydney, Australia.
Vanderzant ES (1974) Development, significance and application
of artificial diets for insects. Annual Review of Entomology 19:
139–160.
Wang Z, Zhang K, Sun X, Tang K & Zhang J (2005) Enhancement
of resistance to aphids by introducing the snowdrop lectin gene
gna into maize plants. Journal of Biosciences 30: 627–638.