)joebxj1vcmjtijoh$psqpsbujpo …downloads.hindawi.com/journals/sci/2016/9475981.pdf · mal axonal...
TRANSCRIPT

Research ArticleHDAC6 Inhibitors Rescued the Defective AxonalMitochondrial Movement in Motor Neurons Derived fromthe Induced Pluripotent Stem Cells of Peripheral NeuropathyPatients with HSPB1 Mutation
Ji-Yon Kim1 So-Youn Woo1 Young Bin Hong2 Heesun Choi3 Jisoo Kim3
Hyunjung Choi3 Inhee Mook-Jung3 Nina Ha4 Jangbeen Kyung4 Soo Kyung Koo5
Sung-Chul Jung6 and Byung-Ok Choi7
1Department of Microbiology School of Medicine Ewha Womans University Seoul Republic of Korea2Stem Cell amp Regenerative Medicine Institute Samsung Medical Center Seoul Republic of Korea3Department of Biochemistry and Biomedical Sciences College of Medicine Seoul National University Seoul Republic of Korea4Chong Kun Dang Research Institute Yongin-si Gyeonggi-do Republic of Korea5Division of Intractable Diseases Center for Biomedical Sciences National Institute of Health Chungcheongbuk-do Republic of Korea6Department of Biochemistry School of Medicine Ewha Womans University Seoul Republic of Korea7Departments of Neurology Samsung Medical Center Sungkyunkwan University School of Medicine Seoul Republic of Korea
Correspondence should be addressed to Sung-Chul Jung jungscewhaackr and Byung-Ok Choi bochoi77hanmailnet
Received 18 July 2016 Revised 11 November 2016 Accepted 22 November 2016
Academic Editor Yang D Teng
Copyright copy 2016 Ji-Yon Kim et al This is an open access article distributed under the Creative Commons Attribution Licensewhich permits unrestricted use distribution and reproduction in any medium provided the original work is properly cited
The Charcot-Marie-Tooth disease 2F (CMT2F) and distal hereditary motor neuropathy 2B (dHMN2B) are caused by autosomaldominantly inherited mutations of the heat shock 27 kDa protein 1 (HSPB1) gene and there are no specific therapies available yetHere we assessed the potential therapeutic effect of HDAC6 inhibitors on peripheral neuropathy with HSPB1 mutation using invitro model of motor neurons derived from induced pluripotent stem cells (iPSCs) of CMT2F and dHMN2B patientsThe absolutevelocity of mitochondrial movements and the percentage of moving mitochondria in axons were lower both in CMT2F-motorneurons and in dHMN2B-motor neurons than those in controls and the severity of the defective mitochondrial movement wasdifferent between the two disease models CMT2F-motor neurons and dHMN2B-motor neurons also showed reduced 120572-tubulinacetylation compared with controls The newly developed HDAC6 inhibitors CHEMICAL X4 and CHEMICAL X9 increasedacetylation of 120572-tubulin and reversed axonal movement defects of mitochondria in CMT2F-motor neurons and dHMN2B-motorneurons Our results suggest that the neurons derived from patient-specific iPSCs can be used in drug screening including HDAC6inhibitors targeting peripheral neuropathy
1 Introduction
Charcot-Marie-Tooth disease (CMT) and related neuro-pathies are a heterogeneous group of neurodegenerativedisorders that share a similar phenotype So far almost 40genes have been identified as being responsible for thesedisorders [1] CMT ismainly divided into demyelinating neu-ropathy (CMT1) and axonal neuropathy (CMT2) according
to electrophysiological andhistopathological featuresWithinthese types CMT type 2F (CMT2F) and distal heredi-tary motor neuropathy 2B (dHMN2B) are caused by heatshock 27 kDa protein 1 (HSPB1 also known as HSP27)mutation in chromosome 7q1123 [2] The most frequentpathologies observed in CMT2F and dHMN2B are abnor-mal axonal transport and cytoskeleton organization [3ndash5] However our knowledge of the underlying molecular
Hindawi Publishing CorporationStem Cells InternationalVolume 2016 Article ID 9475981 14 pageshttpdxdoiorg10115520169475981
2 Stem Cells International
mechanisms is still limited and specific therapies are not yetavailable
HSPB1 belongs to a family of small heat shock proteins(sHSPs) that formdynamic quaternary structures in responseto cellular stress via its conserved 120572-crystallin domain Inaddition to its role in protein refolding HSPB1 is involvedin protein translation intracellular reductionoxidation statecytoskeletal structure cell differentiation and apoptosis [6]Although HSPB1 is ubiquitously expressed in various tissuesmutations in HSPB1 appear to cause neuronal degenerationonly in the peripheral nervous system primarily via alter-ation of cytoskeletal components Clinically HSPB1P182L iscausative of dHMN2B whereas HSPB1S135F is causative ofboth CMT2F and dHMN2B [2] Previous studies of trans-fected cell lines indicate that HSPB1S135F expression disruptsthe neurofilament (NF) network and increases toxic aggrega-tion of NFs [3] whereas both HSPB1S135F and HSPB1P182Lexpressions disturb the anterograde transportation of NFsby reducing the binding of kinesin to NFs and inducingcyclin-dependent kinase 5-mediated hyperphosphorylationof NFs [5] In addition mutations in HSPB1 also appearto affect axonal microtubule tracks In stabilized cell linesand presymptomatic transgenic mice HSPB1S135F expressionleads to aberrant stabilization of microtubulin tracks result-ing from hyperactive interaction between HSPB1S135F and 120572-tubulin [4] Moreover symptomatic transgenic HSPB1S135F
and HSPB1P182L mice show reduced acetylation of 120572-tubulinin sciatic nerves and HSPB1S135F expressing mice showfewer moving mitochondria in the dorsal root ganglion[7] However more physiologically relevant and specific cellmodels for CMT2F are needed to identify precise diseasemechanisms and assess the efficacy of candidate drug screen-ing
Histone deacetylase 6 (HDAC6) is a class IIb HDACthat regulates acetylation states of 120572-tubulin [8 9] MoreoverHDAC6 inhibitors reverse axonal transport deficits in mousemodels of CMT2F [7] and cellular models of Parkinsonrsquosdisease [10] and Huntingtonrsquos disease [11] by increasingacetylated 120572-tubulin Therefore we assessed the therapeuticeffect of HDAC6 inhibitors including tubastatin A and newlydeveloped CHEMICAL X4 and CHEMICAL X9 (ChongKun Dang Pharmaceutical Cooperation Seoul Korea) inCMT2F- and dHMN2B-specific motor neurons (MNs) dif-ferentiated from induced pluripotent stem cells (iPSCs) frompatients carrying HSPB1S135F and HSPB1P182L mutations byanalyzing mitochondrial movements and acetylation of 120572-tubulin in vitro
2 Materials and Methods
21 Ethics Statement and Human Samples Skin fibroblastswere obtained from skin biopsy (4mm punch) of twopatients carrying mutant forms of HSPB1 (S135F and P182L)with informed consent after Institutional Review Boardapproval (ECT 11-58-37 Ewha Womans University Mok-Dong Hospital Seoul Korea) All experiments were per-formed in accordance with the approved guidelines andregulations Fresh skin samples were minced and digested
in Dulbeccorsquos Modified Eaglersquos Medium (DMEM) containing10mgmL collagenase type IV 50UmL dispase and 005trypsinEDTA for 40min at 37∘C After filtration througha cell strainer (70120583m pore size) cells were washed twiceand harvested in DMEM containing 20 fetal bovine serum(FBS) and 100 120583gmL penicillinstreptomycin
22 Generation of iPSCs Four types of Sendai virus-containing vectors each expressing different transcriptionfactors (Sox2 Oct4 Klf4 and c-Myc Invitrogen CarlsbadCA USA) were introduced into fibroblasts derived frompatients with a multiplicity of infection of 3 by followingmanufacturerrsquos protocol On day 7 cells were trypsinizedand transferred onto mitomycin C (Sigma St Louis MOUSA)-treated mouse embryonic fibroblasts SNL feeder cells(Cell Biolabs Inc San Diego CA USA) and harvestedwith embryonic stem cell (ESC)iPSC medium (KnockOutGibco Grand Island NY USA) containing 4 ngmL basicfibroblast growth factor (bFGF) The medium was changeddaily On day 30 iPSC colonies were selected based ontheir morphological characteristics Other stem cells usedas controls included human ESCs (WA09 WiCell MadisonWI USA) and human iPSCs (hFSiPS1 National Stem CellBank of Korea Cheongju Korea)
23 Karyotype Analysis CMT2F-iPSCs and dHMN2B-iPSCswere cultured without feeder cells at cellular passages of 10and 20 cells from each type of iPSCs were analyzed for thekaryoptype by Seegene Medical Foundation (Seoul Korea)
24 Teratoma Assay Equal volumes of mixtures of Matrigel(Corning Corning NY USA) and 10 times 106 humanESCs (WA09) human iPSCs (hFSiPS1) CMT2F-iPSCs ordHMN2B-iPSCs were subcutaneously injected into the backsof 5-week-old female NODSCID mice (Laboratory AnimalResource Center Korean Research Institute of Bioscienceand Biotechnology Daejeon Korea) The xenografts wereallowed to grow for 8 weeks and were then explanted bysurgical procedures Teratoma tissues were fixed in 10formaldehyde and embedded in paraffin For histologicalanalysis hematoxylin and eosin staining was performed
25 Embryoid Body- (EB-) Mediated In Vitro DifferentiationAssay ESCs and iPSCs were plated onto ultralow bindingplates (ie uncoated Petri dishes) and cultured in suspensionwith ESCiPSC medium (KnockOut) which was replacedevery other day After 8 days in floating culture EBs weretransferred onto a gelatin-coated (Sigma) chamber slide(NalgeneNunc Rochester NY USA) and cultured in 10FBSDMEM(WelgeneDaegu Korea) to allowdifferentiationrandomly into three germ-layered cells for an additional 8days
26 Reverse Transcription Polymerase Chain Reaction (RT-PCR) For the detection of total and endogenous expres-sion of KLF4 OCT4 SOX2 and c-MYC and HSPB1 RNAextraction from ESCs and iPSCs was performed with TRIzolReverse transcription was performed using AMV reversetranscriptase (Promega Madison WI USA) and PCR was
Stem Cells International 3
performed using Ex Taq polymerase (Takara Bio OtsuJapan) Primer sequences are KLF4 CDR (108 bp) 51015840-CTGCGG CAA AAC CTA CAC AAA-31015840 (forward) and 51015840-GCGAAT TTC CAT CCA CAG CC -31015840 (reverse) KLF4 UTR(96 bp) 51015840-CAT GGT CAA GTT CCC AAC TGA G-31015840(forward) and 51015840-CAC AGA CCC CAT CTG TTC TTT G-31015840 (reverse) OCT34 CDR (161 bp) 51015840-CAG TGC CCG AAACCCACAC-31015840 (forward) and 51015840-GGAGACCCAGCAGCCTCA AA-31015840 (reverse) OCT34 UTR (120 bp) 51015840-GAA AACCTG GAG TTT GTG CCA-31015840 (forward) and 51015840-TCA CCTTCC CTC CAA CCA GTT-31015840 (reverse) SOX2 CDR (131 bp)51015840-TAC CTC TTC CTC CCA CTC C-31015840 (forward) and 51015840-GGTAGTGCTGGGACATGTGA-31015840 (reverse) SOX2UTR(105 bp) 51015840-CCCGGTACGCTCAAAAAGAA-31015840 (forward)and 51015840-GGT TTT TGC GTG AGT GTG GAT-31015840 (reverse)c-MYC CDR (380 bp) 51015840-CGT CCT CGG ATT CTC TGCTC-31015840 (forward) and 51015840-GCTGGTGCATTTTCGGTTGT-31015840 (reverse) c-MYC UTR (328 bp) 51015840-GCG TCC TGG GAAGGG AGA TCC GGA GC-31015840 (forward) and 51015840-TTG AGGGGC ATC GTC GCG GGA GGC TG-31015840 (reverse)
27 Sanger Sequencing Pathogenic mutations (404CgtT and545CgtT) in HSPB1 gene from patients iPSCs were confirmedby Sanger sequencing using a 3730xl DNA Analyzer (Macro-gen Inc Seoul Korea) and analyzed using Sequencherv523 (GeneCodes Corporation Ann Arbor MI USA) Theprimers used for amplifying and sequencing are as follows51015840-TTT CTG AGC AGA CGT CCA GA-31015840 (forward) and 51015840-CTT TAC TTG GCG GCA GTC TC-31015840 (reverse)
28 Directed Differentiation of iPSCs into MNs To gen-erate EBs colonies of ESCs and iPSCs were enzymati-cally dissociated into small clumps and cultured in sus-pension for 2 days in a Petri dish supplemented withESCiPSC medium (KnockOut) containing 10 120583M Rho-associated kinase inhibitor Y27632 (Tocris Bioscience Bris-tol UK) 20 ngmL bFGF (Invitrogen) 10120583M SB435142(Stemgent Cambridge MA) and 02 120583M LDN193189 (Stem-gent) and penicillinstreptomycin On day 5 for caudaliza-tion retinoic acid (1 120583M Sigma) ascorbic acid (04 120583gmLSigma) brain-derived neurotrophic factor (10 ngmL RampDMinneapolis MN USA) and N2 supplement (1 Gibco)were added [12] On day 7 for ventralization the sonic hedge-hog agonist purmorphamine (1 120583M Stemgent) was addedand the dual SMAD inhibitors (SB435142 and LDN193189)were discontinued On day 17 basal medium was changedto neurobasal medium (Invitrogen) containing all previouslyindicated factors with the addition of insulin-like growthfactor-1 (10 ngmL) glial cell line-derived neurotrophic factor(10 ngmL) and B27 (2 Gibco) On day 21 neurosphereswere dissociated with accutase (Gibco) and plated onto poly-L-lysinelaminin-coated culture plates or slide chambers(Nalgene) and supplemented with neurobasal medium con-taining all previously indicated factors with the addition of120573-mercaptoethanol (25120583M Gibco) and glutamic acid (25 120583MSigma)
29 Immunoblot Assay Proteins were collected from dif-ferentiated MNs by conventional methods using RIPA lysis
buffer (150mM NaCl 10 NP-40 05 sodium deoxy-cholate 01 sodium dodecyl sulfate 50mM Tris pH 80)SDS-PAGE gel was transferred to polyvinylidene difluoridemembranes For blotting anti-acetylated 120572-tubulin (1 1000mouse IgG
2b 6-11B-1 Abcam Cambridge UK) and anti-120572-tubulin (1 1000 mouse IgG
1 DM1A Sigma) antibodies were
used Band densities were analyzed with UN-SCAN-IT gelsoftware (Silk Scientific Inc Orem UT USA)
210 Immunocytochemistry Cells were fixed with 4paraformaldehyde and blocked with 10 normal goat serum(Gibco) and 02 Triton X-100 Primary antibodies wereanti-SSEA4 (1 100 mouse IgG
3 MC-813-70 Developmental
Studies Hybridoma Bank (DSHB) Iowa City IA USA)anti-NANOG (1 500 mouse IgG
1 NNG-811 Abcam)
anti-120572-fetoprotein (AFP 1 100 mouse IgG2b 2A9 Abcam)
anti-120572-smooth muscle actin (SMA 1 100 mouse IgG2a 1A4
Abcam) anti-nestin (1 1000 mouse IgG1 10C2 Abcam)anti-HB9 (1 100 mouse IgG1 815C10 DSHB) anti-islet-12 (ISL12 1 50 mouse IgG
2b 394DS DSHB) anti-neurofilament H nonphosphorylated (SMI32 1 500 mouseIgG1 Covance Princeton NJ USA) anti-neuron-specific
beta III tubulin (Tuj1 1 1000 rabbit polyclonal Abcam)anti-microtubule-associated protein 2 (MAP2 1 200rabbit polyclonal Millipore Billerica MA USA) anti-choline acetyltransferase (ChAT 1 1000 rabbit polyclonalAbcam) anti-synapsin 1 (1 1000 rabbit polyclonal Abcam)anti-120572-tubulin (1 500 rabbit polyclonal Abcam) andanti-acetylated 120572-tubulin (1 200 mouse IgG
2b 6-11B-1Abcam) antibodies Secondary antibodies were AlexaFluor 488-conjugated goat anti-rabbit IgG (preadsorbedAbcam) Cy3-conjugated goat anti-rabbit IgG (Abcam)FITC-conjugated goat anti-mouse IgG (Abcam) andCy3-conjugated goat anti-mouse IgG (Abcam) antibodies
211 Microfluidic Culture for Analysis of AxonalMitochondrialMovements Neurospheres derived from ESCs and iPSCswere dissociated with accutase into single cells and seededontomicrochannel plates at a density of 1 times 105 cellsplate andculturedwith neurobasal (Invitrogen) and B27medium for 10days After axons had completely stretched through the 120583m-sized grooves (total length = 8334 120583m) and reached the oppo-site compartment MNs were transfected with mito-dsRED2(Clonetech IncTakara Bio) delivered by lipofectamine 2000(Invitrogen) Within 3 days of transfection mitochondrialimages were captured by a fluorescent microscope at arate of 121 snaps2min and stacked into one file to createkymographs Mitochondrial moving velocity was calculatedby measured angle and distance in kymograph using ImageJsoftware Axonal lengths were also measured using ImageJAxonal lengths were calculated by summation of measuredlength of axon stretched out from a single 120583m-sized grooveand known length of the groove (8334 120583m)
212 Drug Treatment For analyzing protein acetylation levelcells were treated with the HDAC6 inhibitors tubastatinA (5 120583M Selleckchem Houston TX USA) CHEMICALX4 (05 120583M and 5 120583M) and X9 (05 120583M and 5 120583M) for16 hrs For analyzing axonal transport HDAC6 inhibitors
4 Stem Cells International
mentioned above were treated for 3 hrs CHEMICAL X4 andX9 are newly established HDAC6 inhibitors from ChongKun Dang Pharmaceutical Cooperation (Seoul Korea) Thecommon structures of X4 and X9 are composed of threecomponents (the Zn-binder the linker and the cap part) andare very potent hydroxamate-based HDAC6 inhibitors [IC
50
(HDAC6) = 53 and 37 nM resp] Similar to pan-HDACor other HDAC subtype specific inhibitors the hydroxamicacid moiety as a Zn-binding group is very important forblocking the catalytic activity of the HDAC6 Even thoughthese compounds have structurally different cap portions(arylurea and indole) they share the benzyl group as alinker moiety resulting in the sufficient HDAC6-specificity[fold (HDAC1HDAC6) = 400 and gt270 resp] to afford thedesirable efficacies in cells and animals without the toxicitiesassociated with the inhibitions of class I HDAC subtypes
213 Statistical Analysis Values are expressed as mean plusmnstandard error of the mean Groups were compared usingtwo-way analysis of variance using GraphPad Prism (version5 GraphPad Software Inc La Jolla CA USA) Statisticalsignificance was set at 119875 lt 005
3 Results
31 Generation of CMT2F-iPSCs and dHMN2B-iPSCsPatient-specific iPSCs were generated from one CMT2Fpatient (female52-year-old Korean) with 404CgtT (S135F)mutation and one dHMN2B patient (female8-year-oldKorean) with 545CgtT (P182L) mutation of the HSPB1 geneBoth of CMT2F and dHMN2B patients showed predominantdistal leg muscle weakness and toe gait abnormalities Theages at onset were 20 years and 7 years respectively Sensoryneve conduction velocities and action potentials of suralnerves were decreased in CMT2F patient but within normalranges in dHMN2B patient
Skin fibroblasts were reprogrammed into iPSCs by Sendaiviral transduction of four episomal vectors carrying KLF4OCT34 SOX2 and c-MYC (Figures 1(a) and 1(b)) Themorphology of CMT2F-iPSC and dHMN2B-iPSC colonieswhich resembled that of human ESCs (WA09) consisted ofcells having a high nuclear-to-cytoplasm ratio compactedin a flat cobblestone-like appearance with sharp edges (Fig-ure 1(c)) The genetic background of CMT2F-iPSCs anddHMN2B-iPSCs was not changed during the reprogram-ming processes especially at the mutation site of HSPB1(Figure 1(d)) CMT2F-iPSCs and dHMN2B-iPSCs preservedtheir normal karyotype (Figure 1(e)) The expression ofendogenous KLF4 OCT34 SOX2 and c-MYC genes wasdetected by RT-PCR after a few subpassages using primerswith complementary sequences to the intron area of thetarget mRNA (Figure 1(f)) Sendai viral genome contentswere not detected in iPSCs after cellular passage of 10 (seeSupplement Figure S1 in Supplementary Material availableonline at httpdxdoiorg10115520169475981) CMT2F-iPSCs and dHMN2B-iPSCs expressed stem cell markerssuch as NANOG in the nucleus and SSEA in the cyto-plasm (Figure 1(g)) The pluripotency of CMT2F-iPSCs anddHMN2B-iPSCs was verified by the presence of randomly
differentiated AFP-positive endodermal cells SMA-positivemesodermal cells and nestin-positive ectodermal cells via EBformation in vitro (Figure 1(h)) and teratoma formation invivo (Figure 1(i))
32 Derivation of In Vitro Models by MN Differentiation ofCMT2F-iPSCs and dHMN2B-iPSCs To recapitulate periph-eral neuropathy MNs were differentiated from iPSCs byproviding dual SMAD inhibitors (SB435142 and LDN193189)for neuralization retinoic acid for caudalization and pur-morphamine for ventralization according to the methoddescribed by Amoroso et al [12] (Figure 2(a)) Fully differen-tiatedMNs (Figure 2(b)) expressed transcription factors suchas HB9 and ISL12 cytoskeletal markers such as Tuj1 MAP2and SMI32 and synapsin and ChAT (Figure 2(c)) S135F-MNs and P182L-MNs showed no developmental defectsevidenced by no differences between S135F-MNs and P182L-MNs and control WA09-MNs and hFSiPS1-MNs in theproportion of marker-positive cells (Figure 2(d)) WA09-MNs (1236plusmn 23 120583m) and S135F-MNs (1287plusmn 20 120583m) showedno differences in axonal length (Figure 2(e)) Neuromuscularjunctions visualized by 120572-bungarotoxin staining (see Supple-ment Figure S2) formed when MNs were cocultured withmyotube cells differentiated from C2C12 cells
33 Axonal Mitochondrial Transport Defects in S135F-MNsAlthough there is heterogeneity in causative genes for differ-ent CMT2 subtypes many disease subtypes involve abnor-malities in the cellular trafficking system [13] As MNs canhave long axons up to one meter in length defects in axonaltransportation may increase vulnerability to axonopathyIn particular mitochondrial transport is extremely impor-tant for maintaining axonal and synaptic stability in neu-rons During bidirectional trafficking of mitochondria alongmicrotubules quality control is accomplished by dynamicfusion and fission processes that enable mitochondria togenerate ATP to support vital cellular functions and bufferintracellular calcium [14] Therefore we tested whetherS135F-MNs and P182L-MNs have defects in mitochondrialaxonal transport by culturing cells in microchannel plates[15] which compartmentalize axons from soma and den-drites (Figure 3(a)) transfecting cells with mito-dsRED2and analyzing kymograph images (Figure 3(b)) We observedthat the absolute velocity of mitochondrial movements wassignificantly lower in S135F-MNs (019 plusmn 001 120583msec) andslightly lower in P182L-MNs (022 plusmn 001 120583msec) comparedwith control WA09-MNs (024 plusmn 001 120583msec) and hFSiPS1-MNs (025 plusmn 001 120583msec) (Figure 3(c) Supplement FigureS3 and Supplement Table S1) Also the proportion of mov-ing mitochondria was significantly decreased in S135F-MNs(2637 plusmn 506) and P182L-MNs (1419 plusmn 214) comparedto those in control WA09-MNs (3139 plusmn 374) and hFSiPS-MNs (Figure 3(d) and Supplement Table S2)
34 Decreased Acetylation of 120572-Tubulin in S135F-MNs Ax-onal transport is regulated by various posttranslational mod-ifications (eg detyrosylation acetylation and glutamyla-tion) of microtubules through the recruitment of molecular
Stem Cells International 5
Platecells
Addvirus
Plate cells onMEF dishes
Emergingcolonies
Colony selection
20 FBSDMEM 20 KOSRDMEMF12 + bFGF
D-2 D0 D7 D8 D12 3-4 weeks
(a)
Normal HSPB1S135F HSPB1P182L
(b)
S135F-iPSCs P182L-iPSCsSNL WA09-hESCs hFSiPS1
(c)
P182L-iPSCsWA09-hESCs hFSiPS1
AG A T C A C C A T C C C A G T C A C C T T C GA G A T C AC C A T C T C A G T A C C T T C GA G A T C A C C A T C C C A G T C A C C T T C550560550560 540550
545C 545CgtT545C
WA09-hESCs S135F-iPSCshFSiPS1
T G G C T A C A T C T C C C G G T G C T T C A CT G G C T A C A T C T C C C G G T G C T T C A T G G C T A C A T C T T C C G G T G C T T C A410 420 410 420400410 420
404C 404C 404CgtT
(d)
S135F-iPSCs
1 2 3 4 5
6 7 8 9 10 11 12
13 14 15 16 17 18
19 20 21 22 X Y
P182L-iPSCs
1 2 3 4 5
6 7 8 9 10 11 12
13 14 15 16 17 18
19 20 21 22 X Y
(e)
Figure 1 Continued
6 Stem Cells International
Total-KLF4
Total-SOX2
GAPDH
Endo-KLF4Endo-OCT34
Endo-SOX2Endo-c-MYC
Total-OCT34
Total-c-MYC
WA
09-h
ESCs
S135
F-iP
SCs_
clone
1S1
35F-
iPSC
s_clo
ne 2
P182
L-iP
SCs_
clone
1P1
82L-
iPSC
s_clo
ne 2
hFSi
PS1
(f)NANOG DNA NANOGDNA SSEA4 DNA SSEA4DNA
WA
09-h
ESCs
S135
F-iP
SCs
P182
L-iP
SCs
hFSi
PS1
(g)
S135
F-iP
SCs
P182
L-iP
SCs
WA
09-h
ESCs
AFPDNA SMADNA NESTINDNA
hFSi
PS1
(h)
Figure 1 Continued
Stem Cells International 7
Mesoderm EctodermEndoderm
S135
F-iP
SCs
P182
L-iP
SCs
WA
09-h
ESCs
hFSi
PS1
Gl Ca NE
Gl CaM N
Gl Ca NE
NCa
Gl
(i)
Figure 1 Generation of CMT2F patient and dHMN2B patient-derived iPSCs (a) Experimental timeline for iPSC generation KOSRKnockOut serum replacement bFGF basic fibroblast growth factor (b) Morphology of fibroblasts from normal individual and patients(original magnification 50x) Scale bars 200 120583m (c) iPSC colonies showed ESC-like morphology such as a flat cobblestone-like appearancewith individual cells having a high nucleus-to-cytoplasm ratio (original magnification 50x) Scale bars 200120583m (d) CMT2F-iPSCs anddHMN2B-iPSCs had preserved point mutation sites in the HSPB1 gene verified by sequencing of RT-PCR products (e) CMT2F-iPSCs anddHMN2B-iPSCs maintained normal karyotype (f) Expression of total and endogenous Klf4 Oct34 Sox2 and c-Myc in CMT2F-iPSCs anddHMN2B-iPSCs was verified by RT-PCR Two clones from each of the patients-derived iPSCs were tested (clone 1 and clone 2) (g) ESCsand iPSCs expressed stem cell markers such as NANOG (in the nucleus original magnification 200x) and SSEA4 (in the cytoplasm originalmagnification 100x) Scale bars 200 120583m (h) EB-mediated in vitro spontaneous differentiation of ESCs and iPSCs resulted in the expressionof three-germ-layer markers such as AFP (endoderm) SMA (mesoderm) and nestin (ectoderm original magnification 200x) Scale bars100120583m (i) ESCs and iPSCs showed in vivo pluripotency by forming teratomas 8 weeks after subcutaneous injection into NODSCID miceTeratomas consisted of various three-germ-layer tissues including columnar gland epithelial cells with secretions [Gl] for endodermal tissuemuscle [M] and cartilage with calcification [Ca] for mesodermal tissue and neuroectodermal tissue [NE] and nerve axonal bundle [N] forectodermal tissue (original magnification 400x) Scale bars 100120583m
motor proteins [16] Among these modifications acety-lation of 120572-tubulin at the protein site of K40 regulatesbinding of dyneindynactin complexes and kinesin-1 tomicrotubules [11 17] In addition treatment with HDAC6inhibitors which increase acetylation of 120572-tubulin reversestransport defects in cellular models of Huntingtonrsquos disease[11] and LRRK2 mutation-induced Parkinsonrsquos disease [9]Therefore acetylation levels of 120572-tubulin were examined asan indicator of mitochondrial transport defects in S135F-MNs and P182L-MNs We found that S135F-MNs andP182L-MNs showed a significant reduction in acetylated 120572-tubulin levels compared with WA09-MNs or hFSiPS1-MNs(Figure 4)
35 CMT2F- and dHMN2B-Related Disease Phenotypes WereReversed by HDAC6 Inhibition By the treatment withHDAC6 inhibitors including tubastatin A (5 120583M) and newlydeveloped CHEMICAL X4 (05 120583M and 5 120583M) and X9(05 120583M and 5 120583M) acetylation of 120572-tubulin in S135F-MNs
and P182L-MNs was increased (Supplement Figure S4 Fig-ures 5(a) and 5(b)) Moreover the absolute velocity ofmitochondrial movements and the proportion of movingmitochondria in axons were increased significantly by thetreatment of these HDAC6 inhibitors (Figures 5(c) and 5(d)and Supplement Tables S1 and S2)
4 Discussion
Since iPSCs serve as an unlimited source of cells that can giverise to neuronal lineages and help to overcome the obstacleof unavailability of affected neuronal tissues from patientsdue to neuronal postmitotic property in vitro modeling anddrug screening with iPSCs can advance basic research onneurodegenerative diseases Therefore one of the strongestpoints of this study is that CMT2F- and dHMN2B-specificin vitro cellular models can be applied for the screening ofputative therapeuticmolecules such asHDAC6 inhibitors onperipheral neuropathy
8 Stem Cells International
ldquoSelf-renewalrdquo
iPSCs EBs Neurospheres MNs
3-4 wksldquoNeuralizationrdquoBMP inhibitors
ldquoCaudalizationrdquoRA
ldquoVentralizationrdquoShh agonist
(a)
WA09-MNs hFSiPS1-MNs
S135F-MNs P182L-MNs
(b)
WA09-MNs S135F-MNs P182L-MNs
ISL1
2T
uJ1
DN
AH
B9T
uJ1
DN
ASM
I32
MA
P2D
NA
SMI3
2Ch
ATD
NA
Syna
psin
DN
A
hFSiPS1-MNs
(c)
Figure 2 Continued
Stem Cells International 9
SMI32DAPI
0
50
100
150Re
lativ
e exp
ress
ion
()
WA09-MNs hFSiPS1-MNs S135F-MNs P182L-MNs
Relat
ive e
xpre
ssio
n (
)
MAP2DAPI
0
50
100
150
WA09-MNs hFSiPS1-MNs S135F-MNs P182L-MNs(d)
Axonal length
0
500
1000
1500
2000
WA09-MNs S135F-MNs
(120583m
)
(e)
Figure 2 Generation of CMT2F-specific in vitro model by differentiation of patient-derived iPSCs into MNs (a) Schematic presentation ofMN differentiation (b) Differentiated MNs showed typical cellular morphology (original magnification 50x) Scale bars 200 120583m (c) S135F-MNs and P182L-MNs expressedMN-specific transcription factors such asHB9 and ISL12 neuronal cytoskeletalmarkers such as TuJ1 SMI32andMAP2 ChAT and the synaptic vesicularmarker synapsin (originalmagnification 400x and 600x) Scale bars 100 120583m (d) Differentiationefficiency of S135F-MNs and P182L-MNs was comparable to control MNs in terms of neuronal marker expression such as SMI32DAPI andMAP2DAPI (WA09-MNs119873 = 30 hFSiPS1-MNs119873 = 329 S135F-MNs119873 = 1730 and P182L-MNs119873 = 1090) (e) Axonal length of S135F-MNs was comparable to that of control MNs Axonal length was measured by culturing fully differentiated MNs in microchannel plates foran additional 2 weeks (WA09-MNs119873 = 70 and S135F-MNs119873 = 121)
HDAC6 is a type IIb HDAC that modulates deacetylationof cytoplasmic target proteins Because HDAC6 is involvedin cytoskeletal formation modulation of cellular oxida-tionreduction state and the ubiquitin-proteasome systeminhibition of HDAC6 improves pathological conditions inneurons such as impairments in axonal transport oxidativestress and protein aggresome formation in vitro [18 19]In terms of axonal transport HDAC6 inhibitors reverseCMT2F and dHMN2B-related axonal transport by increasingacetylated 120572-tubulin In this study newly developed HDAC6inhibitors having 35-fold lower half-maximal inhibitoryconcentration (IC
50) values than that of tubastatin A showed
better performance on improving mitochondrial axonaltransport at 10 times lower concentration than tubastatin A
CMT2F-iPSCs and dHMN2B-iPSCswere generated frompatient-derived dermal fibroblasts carrying 404CgtT (S135F)and 545CgtT (P182L) mutations of HSPB1 by forced expres-sion of KLF4 OCT34 SOX2 and c-MYC using Sendai virus
vectors After establishing several stem cell colonies twocolony clones from each patient were used for further analysisto avoid differences due to clonal variance and selectediPSCs were differentiated into motor neurons with definedcytokines To recapitulate axonal pathology in vitro patient-specific motor neurons were cultured in microchanneledplates which are designed to separate the microenvironmentsof neuronal compartments In this system specific cellularmodels of CMT2F and dHMN2B showed defects in mito-chondrial transport as the absolute velocity of moving mito-chondria and the percentage of moving mitochondria werelower than control MNs Phenotypic variance between iPSCclones was observed in P182L-MNs in terms ofmitochondrialvelocity (see Figure S3) Defects in the axonal traffic systemare linked to many neurodegenerative diseases Due to thedistinct anatomy of peripheral neurons which have axons upto one meter in length the interchange of genetic materialmetabolites proteins and organelles between soma and distal
10 Stem Cells International
Original magnification 50xBar = 200120583m
(a)
WA09
P182LhFSiPS1
(b)
Absolute velocity (120583msec)
00
01
02
03
WA09-MNs hFSiPS1-MNs S135F-MNs P182L-MNs
(120583m
sec
)
lowast
lowastlowastlowast
(c)
Moving mitochondria ()
0
20
40
60
80
()
WA09-MNs hFSiPS1-MNs S135F-MNs P182L-MNs
lowastlowastlowast
lowastlowastlowast
lowastlowastlowastlowast
(d)
Figure 3 Axonal transport of mitochondria in control MNs S135F-MNs and P182L-MNs (a) Microfluidic culturing system Microchannelplates were made from transparent polymer in a specialized mold to create multicompartments connected by 120583m-sized grooves (8334 120583mlong in length) that only allow growing axons to pass through thereby separating the microenvironments of neuronal components (b)Visualization of mitochondria by Mito-dsRED2 transfection (upper image) and kymograph images for mitochondria movement (middleand lower four images) Mito-dsRED2 vector was introduced into fully differentiated MNs cultured on microchannel plates for an additional1 weekMitochondrial images were taken at a ratio of 121 snaps2min and stacked into kymograph for analysis (originalmagnification 1000x)(c) S135F-MNs show decreased velocity of moving mitochondria compared to control MNs Unpaired t-testslowast119875 lt 005 lowastlowastlowast119875 lt 0001 andlowastlowastlowastlowast119875 lt 00001 Absolute velocity of moving mitochondria was calculated from measured angle and length in kymograph images using
ImageJ (WA09-MNs 119873 = 203 hFSiPS1-MNs 119873 = 512 S135F-MNs 119873 = 428 and P182L-MNs 119873 = 496) (d) S135F-MNs and P182L-MNs show reduced moving proportion of mitochondria in an axon (WA09-MNs 119873 = 9 hFSiPS1-MNs 119873 = 6 S135F-MNs 119873 = 20 andP182L-MNs119873 = 42)
subcellular sites is critical for maintaining neuronal home-ostasis and survival Of the various trafficking cargos thetransport of mitochondria is particularly important becausemitochondria support several cellular processes by supplyingATP buffering intracellular Ca2+ and modulating apoptosis[13] In addition to delivering mitochondria to appropriatesubcellular sites the trafficking of mitochondria is vital forthe recycling of their components through dynamic fusionand fission mechanism [14] Defects in axonal mitochondrialtransport are associated with Alzheimerrsquos disease [20 21]amyotrophic lateral sclerosis [22] and Huntingtonrsquos disease[23 24]
Mitochondria are transported bidirectionally by interac-tions with molecular motor proteins such as kinesin-1 foranterogrademovement and dynein for retrogrademovementand those attach via adaptor proteins such as the mitochon-drial Rho-Milton complex and dynactin respectively [14]The recruitment of molecularmotors tomicrotubule tracks isregulated by posttranslational modification of 120572-tubulin [16]especially acetylation [17] In this study the level of acetylated120572-tubulin was reduced in both S135F-MNs and P182L-MNsBy increasing the acetylation of 120572-tubulin via treatment withHDAC6 inhibitors mitochondrial transport was improvedin terms of both velocity and the percentage of moving
Stem Cells International 11
WA09-MNs hFSiPS1 S135F-MNs P182L-MNs120572
-tubu
linAc
etyl
120572-tu
bulin
Acet
yl120572
-tubu
lin
120572-tu
bulin
DN
A
(a)
00
02
04
06
08
10
WA09-MNs hFSiPS1-MNs S135F-MNs P182L-MNs
Acet
yl120572
-tubu
lin120572
-tubu
lin
lowastlowast
lowast
lowastlowast
lowastlowast
(b)
WA09-MNs hFSiPS1-MNs S135F-MNs P182L-MNs
120572-Tubulin
Acetyl 120572-tubulin
(c)
00
02
04
06
08
10
WA09-MNs hFSiPS1-MNs S135F-MNs P182L-MNs
Acet
yl120572
-tubu
lin120572
-tubu
linlowast
lowast
lowast
lowastlowast
(d)
Figure 4 Reduced acetylation of120572-tubulin in S135F-MNs andP182-MNs (a) Acetylation of120572-tubulinwas visualized by immunostainingwithanti-acetylated 120572-tubulin Abs (red) Total 120572-tubulin was also detected by staining with anti-120572-tubulin Abs (green) (original magnification400x) Scale bars 100120583m (b) The extent of acetylated 120572-tubulin was normalized with corresponding total 120572-tubulin using ImageJ software(WA09-MNs119873 = 3 S135F-MNs119873 = 4 and P182L-MNs119873 = 5) Unpaired 119905-tests lowast119875 lt 005 and lowastlowast119875 lt 001 (c and d) Western blot assayshowed decreased acetylated 120572-tubulin levels in S135F-MNs and P182-MNs (paired 119905-tests lowast119875 lt 005 and lowastlowast119875 lt 001)
mitochondria Hence defects in the axonal trafficking ofmitochondria are associated with decreased acetylation of120572-tubulin which may be the main causative factor foraxonopathy in CMT2F and dHMN2B
In summary iPSCs can be used for valuable resourcesof in vitro disease modeling as well as drug screening Weanticipate that this in vitro system would be a useful modelfor the clinical translational research and drug development
5 Conclusions
This study demonstrates in vitro drug screening mod-els of inherited neuropathy resulting from mutations in
the HSPB1 gene can be developed from patient-specificiPSCs Axonal pathologies were mimicked in microflu-idic culturing system as both S135F-MNs and P182L-MNsshowed a marked decline in the absolute velocity andthe percentage of moving mitochondria These axonal de-fects were associated with decreased acetylation of 120572-tu-bulin and were reversed by treatment of HDAC6 in-hibitors
Competing Interests
The authors declare that they have no conflict of interests
12 Stem Cells International
HDAC6 inhibitors
S135
F-M
Ns
P182
L-M
Ns
Acetyl 120572-tubulin
Acetyl 120572-tubulin
120572-Tubulin
120572-Tubulin
DM
SO
X4_0
5120583
M
X4_5
120583M
X9_0
5120583
M
X9_5
120583M
Tuba
statin
120583M
A_5
(a)
00
05
10
15
20
25
DM
SO
X4_0
5120583
M
X4_5
120583M
X9_0
5120583
M
X9_5
120583M
lowast
lowastlowast
lowastlowast
Acet
yl120572
-tubu
lin120572
-tubu
lin
Acetyl 120572-tubulin in S135F-MNs
00
05
10
15
20
25
DM
SO
X4_0
5120583
M
X4_5
120583M
X9_0
5120583
M
X9_5
120583M
lowast
lowastlowast
lowastlowastAc
etyl
120572-tu
bulin
120572-tu
bulin
Acetyl 120572-tubulin in P182L-MNs
Tuba
statin
120583M
A_5
Tuba
statin
120583M
A_5
(b)
00
01
02
03
04
DM
SO
X4_0
5120583
M
X4_5
120583M
X9_0
5120583
M
X9_5
120583M
lowastlowastlowastlowast
lowastlowastlowastlowast
Abso
lute
velo
city
(120583m
sec
)
00
02
04
06
DM
SO
X4_0
5120583
M
X4_5
120583M
X9_0
5120583
M
X9_5
120583M
lowast
lowastlowastlowastlowast
lowastlowastlowastlowast
lowastlowastlowastlowast
lowastlowast
Abso
lute
velo
city
(120583m
sec
)
S135F-MNs
Absolute velocity
P182L-MNs
Tuba
statin
120583M
A_5
Tuba
statin
120583M
A_5
(c)
Figure 5 Continued
Stem Cells International 13
0
20
40
60
80
Mov
ing
mito
chon
dria
()
DM
SO
X4_0
5120583
M
X4_5
120583M
X4_5
120583M
X9_0
5120583
M
X9_5
120583M
lowast
lowastlowast
lowastlowast
lowastlowastlowastlowast
0
20
40
60
80
Mov
ing
mito
chon
dria
()
DM
SO
X4_0
5120583
M
X9_0
5120583
M
X9_5
120583M
lowastlowastlowastlowast
lowastlowastlowastlowast
lowastlowastlowastlowast
lowastlowastlowastlowast
S135F-MNs
Moving mitochondria
P182L-MNs
Tuba
statin
120583M
A_5
Tuba
statin
120583M
A_5
lowastlowast
(d)
Figure 5 HDAC6 inhibitors reversed axonal defect of mitochondrial transport by increasing acetylation of 120572-tubulin (a and b) Westernblot assay showed that treatment of HDAC6 inhibitors increased acetylated 120572-tubulin levels in S135F-MNs and P182L-MNs (paired 119905-testslowast119875 lt 005 and lowastlowast119875 lt 001) (c) HDAC6 inhibitors increased the absolute velocity of mitochondrial movements in S135F-MNs and P182L-MNs (WA09-MNs119873 = 617 S135F-MNs119873 = 101 and P182L-MNs119873 = 394) (d) HDAC6 inhibitors increased the percentage of movingmitochondria in S135F-MNs and P182L-MNs (WA09-MNs 119873 = 15 S135F-MNs 119873 = 7 and P182L-MNs 119873 = 15) Unpaired 119905-testslowast119875 lt 005 lowastlowast119875 lt 001 and lowastlowastlowastlowast119875 lt 00001
Authorsrsquo Contributions
Ji-Yon Kim was responsible for the iPSC experiments collec-tion and interpretation of the data and preparation of thepaper So-Youn Woo was interpreting the data and editingthe manuscript Inhee Mook-Jung provided microchannelplates and designed the experiment Heesun Choi JisooKim and Hyunjung Choi carried out microfluidic cultureand kymograph analysis Nina Ha Jangbeen Kyung andSoo Kyung Koo designed the experiments and preparationof the manuscript Young-Bin Hong and Sung-Chul Jungwere responsible for conception design of experiments andediting the manuscript Byung-Ok Choi was responsible forthe collection of patient samples and interpretation of thedata
Acknowledgments
This study was supported by the Korean Health TechnologyRampD Project Ministry of Health amp Welfare (HI12C0135HI14C3484 and HI15C1560) and a National Research Foun-dation of Korea grant funded by the Ministry of Science ICTandFuture Planning (NRF-2014R1A2A2A01004240)Humanstem cell line hFSiPS1 was provided by National Stem CellBank of Korea (Korea National Institute of Health Grant no4861-307-210-13)
References
[1] Inherited Peripheral Neuropathies Mutation Database httpwwwmolgenuaacbecmtmutationsmutationsMutByGenecfm
[2] O V Evgrafov IMersiyanova J Irobi et al ldquoMutant small heat-shock protein 27 causes axonal Charcot-Marie-Tooth diseaseand distal hereditary motor neuropathyrdquo Nature Genetics vol36 no 6 pp 602ndash606 2004
[3] J Zhai H Lin J-P Julien andWW Schlaepfer ldquoDisruption ofneurofilament network with aggregation of light neurofilamentprotein a common pathway leading to motor neuron degener-ation due to Charcot-Marie-Tooth disease-linked mutations inNFL andHSPB1rdquoHumanMolecular Genetics vol 16 no 24 pp3103ndash3116 2007
[4] L Almeida-Souza B Asselbergh C drsquoYdewalle et al ldquoSmallheat-shock protein HSPB1 mutants stabilize microtubules inCharcot-Marie-Tooth neuropathyrdquoThe Journal of Neurosciencevol 31 no 43 pp 15320ndash15328 2011
[5] A Holmgren D Bouhy V De Winter et al ldquoCharcot-Marie-Tooth causing HSPB1mutations increase Cdk5-mediated phos-phorylation of neurofilamentsrdquoActa Neuropathologica vol 126no 1 pp 93ndash108 2013
[6] A-P Arrigo ldquoThe cellular ldquonetworkingrdquo of mammalian Hsp27and its functions in the control of protein folding redox stateand apoptosisrdquoAdvances in eXperimental Medicine and Biologyvol 594 pp 14ndash26 2007
14 Stem Cells International
[7] C DrsquoYdewalle J Krishnan D M Chiheb et al ldquoHDAC6inhibitors reverse axonal loss in a mouse model of mutantHSPB1-induced Charcot-Marie-Tooth diseaserdquo NatureMedicine vol 17 no 8 pp 968ndash974 2011
[8] Y Zhang N Li C Caron et al ldquoHDAC-6 interacts with anddeacetylates tubulin and microtubules in vivordquo EMBO Journalvol 22 no 5 pp 1168ndash1179 2003
[9] A Valenzuela-Fernandez J R Cabrero J M Serrador and FSanchez-Madrid ldquoHDAC6 a key regulator of cytoskeleton cellmigration and cell-cell interactionsrdquo Trends in Cell Biology vol18 no 6 pp 291ndash297 2008
[10] V K Godena N Brookes-Hocking AMoller et al ldquoIncreasingmicrotubule acetylation rescues axonal transport and locomo-tor deficits caused by LRRK2 Roc-COR domain mutationsrdquoNature Communications vol 5 article 5245 2014
[11] J P Dompierre J D Godin B C Charrin et al ldquoHistonedeacetylase 6 inhibition compensates for the transport deficitin Huntingtonrsquos disease by increasing tubulin acetylationrdquo TheJournal of Neuroscience vol 27 no 13 pp 3571ndash3583 2007
[12] M W Amoroso G F Croft D J Williams et al ldquoAcceleratedhigh-yield generation of limb-innervating motor neurons fromhuman stem cellsrdquo Journal of Neuroscience vol 33 no 2 pp574ndash586 2013
[13] B J Gentil and L Cooper ldquoMolecular basis of axonal dysfunc-tion and traffic impairments in CMTrdquo Brain Research Bulletinvol 88 no 5 pp 444ndash453 2012
[14] Z-H Sheng and Q Cai ldquoMitochondrial transport in neu-rons impact on synaptic homeostasis and neurodegenerationrdquoNature Reviews Neuroscience vol 13 no 2 pp 77ndash93 2012
[15] J W Park B Vahidi A M Taylor S W Rhee and N LJeon ldquoMicrofluidic culture platform for neuroscience researchrdquoNature Protocols vol 1 no 4 pp 2128ndash2136 2006
[16] SWestermann and KWeber ldquoPost-translational modificationsregulate microtubule functionrdquo Nature Reviews Molecular CellBiology vol 4 no 12 pp 938ndash947 2003
[17] N A Reed D Cai T L Blasius et al ldquoMicrotubule acetylationpromotes kinesin-1 binding and transportrdquoCurrent Biology vol16 no 21 pp 2166ndash2172 2006
[18] C drsquoYdewalle E Bogaert and L Van Den Bosch ldquoHDAC6at the intersection of neuroprotection and neurodegenerationrdquoTraffic vol 13 no 6 pp 771ndash779 2012
[19] C Simoes-Pires V Zwick A Nurisso E Schenker P-ACarrupt and M Cuendet ldquoHDAC6 as a target for neurode-generative diseases what makes it different from the otherHDACsrdquoMolecular Neurodegeneration vol 8 article 7 2013
[20] Y Rui P Tiwari Z Xie and J Q Zheng ldquoAcute impairment ofmitochondrial trafficking by 120573-amyloid peptides in hippocam-pal neuronsrdquo Journal of Neuroscience vol 26 no 41 pp 10480ndash10487 2006
[21] K A Vossel K Zhang J Brodbeck et al ldquoTau reductionprevents A120573-induced defects in axonal transportrdquo Science vol330 no 6001 p 198 2010
[22] K J De vos A L Chapman M E Tennant et al ldquoFamilialamyotrophic lateral sclerosis-linked SOD1 mutants perturbfast axonal transport to reduce axonal mitochondria contentrdquoHumanMolecular Genetics vol 16 no 22 pp 2720ndash2728 2007
[23] E Trushina R B Dyer J D Badger II et al ldquoMutant huntingtinimpairs axonal trafficking inmammalian neurons in vivo and invitrordquo Molecular and Cellular Biology vol 24 no 18 pp 8195ndash8209 2004
[24] A L Orr S Li C-E Wang et al ldquoN-terminal mutant hunt-ingtin associates with mitochondria and impairs mitochondrialtraffickingrdquo Journal of Neuroscience vol 28 no 11 pp 2783ndash2792 2008
Submit your manuscripts athttpwwwhindawicom
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Anatomy Research International
PeptidesInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporation httpwwwhindawicom
International Journal of
Volume 2014
Zoology
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Molecular Biology International
GenomicsInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioinformaticsAdvances in
Marine BiologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Signal TransductionJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
Evolutionary BiologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Biochemistry Research International
ArchaeaHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Genetics Research International
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Advances in
Virolog y
Hindawi Publishing Corporationhttpwwwhindawicom
Nucleic AcidsJournal of
Volume 2014
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Enzyme Research
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
International Journal of
Microbiology

2 Stem Cells International
mechanisms is still limited and specific therapies are not yetavailable
HSPB1 belongs to a family of small heat shock proteins(sHSPs) that formdynamic quaternary structures in responseto cellular stress via its conserved 120572-crystallin domain Inaddition to its role in protein refolding HSPB1 is involvedin protein translation intracellular reductionoxidation statecytoskeletal structure cell differentiation and apoptosis [6]Although HSPB1 is ubiquitously expressed in various tissuesmutations in HSPB1 appear to cause neuronal degenerationonly in the peripheral nervous system primarily via alter-ation of cytoskeletal components Clinically HSPB1P182L iscausative of dHMN2B whereas HSPB1S135F is causative ofboth CMT2F and dHMN2B [2] Previous studies of trans-fected cell lines indicate that HSPB1S135F expression disruptsthe neurofilament (NF) network and increases toxic aggrega-tion of NFs [3] whereas both HSPB1S135F and HSPB1P182Lexpressions disturb the anterograde transportation of NFsby reducing the binding of kinesin to NFs and inducingcyclin-dependent kinase 5-mediated hyperphosphorylationof NFs [5] In addition mutations in HSPB1 also appearto affect axonal microtubule tracks In stabilized cell linesand presymptomatic transgenic mice HSPB1S135F expressionleads to aberrant stabilization of microtubulin tracks result-ing from hyperactive interaction between HSPB1S135F and 120572-tubulin [4] Moreover symptomatic transgenic HSPB1S135F
and HSPB1P182L mice show reduced acetylation of 120572-tubulinin sciatic nerves and HSPB1S135F expressing mice showfewer moving mitochondria in the dorsal root ganglion[7] However more physiologically relevant and specific cellmodels for CMT2F are needed to identify precise diseasemechanisms and assess the efficacy of candidate drug screen-ing
Histone deacetylase 6 (HDAC6) is a class IIb HDACthat regulates acetylation states of 120572-tubulin [8 9] MoreoverHDAC6 inhibitors reverse axonal transport deficits in mousemodels of CMT2F [7] and cellular models of Parkinsonrsquosdisease [10] and Huntingtonrsquos disease [11] by increasingacetylated 120572-tubulin Therefore we assessed the therapeuticeffect of HDAC6 inhibitors including tubastatin A and newlydeveloped CHEMICAL X4 and CHEMICAL X9 (ChongKun Dang Pharmaceutical Cooperation Seoul Korea) inCMT2F- and dHMN2B-specific motor neurons (MNs) dif-ferentiated from induced pluripotent stem cells (iPSCs) frompatients carrying HSPB1S135F and HSPB1P182L mutations byanalyzing mitochondrial movements and acetylation of 120572-tubulin in vitro
2 Materials and Methods
21 Ethics Statement and Human Samples Skin fibroblastswere obtained from skin biopsy (4mm punch) of twopatients carrying mutant forms of HSPB1 (S135F and P182L)with informed consent after Institutional Review Boardapproval (ECT 11-58-37 Ewha Womans University Mok-Dong Hospital Seoul Korea) All experiments were per-formed in accordance with the approved guidelines andregulations Fresh skin samples were minced and digested
in Dulbeccorsquos Modified Eaglersquos Medium (DMEM) containing10mgmL collagenase type IV 50UmL dispase and 005trypsinEDTA for 40min at 37∘C After filtration througha cell strainer (70120583m pore size) cells were washed twiceand harvested in DMEM containing 20 fetal bovine serum(FBS) and 100 120583gmL penicillinstreptomycin
22 Generation of iPSCs Four types of Sendai virus-containing vectors each expressing different transcriptionfactors (Sox2 Oct4 Klf4 and c-Myc Invitrogen CarlsbadCA USA) were introduced into fibroblasts derived frompatients with a multiplicity of infection of 3 by followingmanufacturerrsquos protocol On day 7 cells were trypsinizedand transferred onto mitomycin C (Sigma St Louis MOUSA)-treated mouse embryonic fibroblasts SNL feeder cells(Cell Biolabs Inc San Diego CA USA) and harvestedwith embryonic stem cell (ESC)iPSC medium (KnockOutGibco Grand Island NY USA) containing 4 ngmL basicfibroblast growth factor (bFGF) The medium was changeddaily On day 30 iPSC colonies were selected based ontheir morphological characteristics Other stem cells usedas controls included human ESCs (WA09 WiCell MadisonWI USA) and human iPSCs (hFSiPS1 National Stem CellBank of Korea Cheongju Korea)
23 Karyotype Analysis CMT2F-iPSCs and dHMN2B-iPSCswere cultured without feeder cells at cellular passages of 10and 20 cells from each type of iPSCs were analyzed for thekaryoptype by Seegene Medical Foundation (Seoul Korea)
24 Teratoma Assay Equal volumes of mixtures of Matrigel(Corning Corning NY USA) and 10 times 106 humanESCs (WA09) human iPSCs (hFSiPS1) CMT2F-iPSCs ordHMN2B-iPSCs were subcutaneously injected into the backsof 5-week-old female NODSCID mice (Laboratory AnimalResource Center Korean Research Institute of Bioscienceand Biotechnology Daejeon Korea) The xenografts wereallowed to grow for 8 weeks and were then explanted bysurgical procedures Teratoma tissues were fixed in 10formaldehyde and embedded in paraffin For histologicalanalysis hematoxylin and eosin staining was performed
25 Embryoid Body- (EB-) Mediated In Vitro DifferentiationAssay ESCs and iPSCs were plated onto ultralow bindingplates (ie uncoated Petri dishes) and cultured in suspensionwith ESCiPSC medium (KnockOut) which was replacedevery other day After 8 days in floating culture EBs weretransferred onto a gelatin-coated (Sigma) chamber slide(NalgeneNunc Rochester NY USA) and cultured in 10FBSDMEM(WelgeneDaegu Korea) to allowdifferentiationrandomly into three germ-layered cells for an additional 8days
26 Reverse Transcription Polymerase Chain Reaction (RT-PCR) For the detection of total and endogenous expres-sion of KLF4 OCT4 SOX2 and c-MYC and HSPB1 RNAextraction from ESCs and iPSCs was performed with TRIzolReverse transcription was performed using AMV reversetranscriptase (Promega Madison WI USA) and PCR was
Stem Cells International 3
performed using Ex Taq polymerase (Takara Bio OtsuJapan) Primer sequences are KLF4 CDR (108 bp) 51015840-CTGCGG CAA AAC CTA CAC AAA-31015840 (forward) and 51015840-GCGAAT TTC CAT CCA CAG CC -31015840 (reverse) KLF4 UTR(96 bp) 51015840-CAT GGT CAA GTT CCC AAC TGA G-31015840(forward) and 51015840-CAC AGA CCC CAT CTG TTC TTT G-31015840 (reverse) OCT34 CDR (161 bp) 51015840-CAG TGC CCG AAACCCACAC-31015840 (forward) and 51015840-GGAGACCCAGCAGCCTCA AA-31015840 (reverse) OCT34 UTR (120 bp) 51015840-GAA AACCTG GAG TTT GTG CCA-31015840 (forward) and 51015840-TCA CCTTCC CTC CAA CCA GTT-31015840 (reverse) SOX2 CDR (131 bp)51015840-TAC CTC TTC CTC CCA CTC C-31015840 (forward) and 51015840-GGTAGTGCTGGGACATGTGA-31015840 (reverse) SOX2UTR(105 bp) 51015840-CCCGGTACGCTCAAAAAGAA-31015840 (forward)and 51015840-GGT TTT TGC GTG AGT GTG GAT-31015840 (reverse)c-MYC CDR (380 bp) 51015840-CGT CCT CGG ATT CTC TGCTC-31015840 (forward) and 51015840-GCTGGTGCATTTTCGGTTGT-31015840 (reverse) c-MYC UTR (328 bp) 51015840-GCG TCC TGG GAAGGG AGA TCC GGA GC-31015840 (forward) and 51015840-TTG AGGGGC ATC GTC GCG GGA GGC TG-31015840 (reverse)
27 Sanger Sequencing Pathogenic mutations (404CgtT and545CgtT) in HSPB1 gene from patients iPSCs were confirmedby Sanger sequencing using a 3730xl DNA Analyzer (Macro-gen Inc Seoul Korea) and analyzed using Sequencherv523 (GeneCodes Corporation Ann Arbor MI USA) Theprimers used for amplifying and sequencing are as follows51015840-TTT CTG AGC AGA CGT CCA GA-31015840 (forward) and 51015840-CTT TAC TTG GCG GCA GTC TC-31015840 (reverse)
28 Directed Differentiation of iPSCs into MNs To gen-erate EBs colonies of ESCs and iPSCs were enzymati-cally dissociated into small clumps and cultured in sus-pension for 2 days in a Petri dish supplemented withESCiPSC medium (KnockOut) containing 10 120583M Rho-associated kinase inhibitor Y27632 (Tocris Bioscience Bris-tol UK) 20 ngmL bFGF (Invitrogen) 10120583M SB435142(Stemgent Cambridge MA) and 02 120583M LDN193189 (Stem-gent) and penicillinstreptomycin On day 5 for caudaliza-tion retinoic acid (1 120583M Sigma) ascorbic acid (04 120583gmLSigma) brain-derived neurotrophic factor (10 ngmL RampDMinneapolis MN USA) and N2 supplement (1 Gibco)were added [12] On day 7 for ventralization the sonic hedge-hog agonist purmorphamine (1 120583M Stemgent) was addedand the dual SMAD inhibitors (SB435142 and LDN193189)were discontinued On day 17 basal medium was changedto neurobasal medium (Invitrogen) containing all previouslyindicated factors with the addition of insulin-like growthfactor-1 (10 ngmL) glial cell line-derived neurotrophic factor(10 ngmL) and B27 (2 Gibco) On day 21 neurosphereswere dissociated with accutase (Gibco) and plated onto poly-L-lysinelaminin-coated culture plates or slide chambers(Nalgene) and supplemented with neurobasal medium con-taining all previously indicated factors with the addition of120573-mercaptoethanol (25120583M Gibco) and glutamic acid (25 120583MSigma)
29 Immunoblot Assay Proteins were collected from dif-ferentiated MNs by conventional methods using RIPA lysis
buffer (150mM NaCl 10 NP-40 05 sodium deoxy-cholate 01 sodium dodecyl sulfate 50mM Tris pH 80)SDS-PAGE gel was transferred to polyvinylidene difluoridemembranes For blotting anti-acetylated 120572-tubulin (1 1000mouse IgG
2b 6-11B-1 Abcam Cambridge UK) and anti-120572-tubulin (1 1000 mouse IgG
1 DM1A Sigma) antibodies were
used Band densities were analyzed with UN-SCAN-IT gelsoftware (Silk Scientific Inc Orem UT USA)
210 Immunocytochemistry Cells were fixed with 4paraformaldehyde and blocked with 10 normal goat serum(Gibco) and 02 Triton X-100 Primary antibodies wereanti-SSEA4 (1 100 mouse IgG
3 MC-813-70 Developmental
Studies Hybridoma Bank (DSHB) Iowa City IA USA)anti-NANOG (1 500 mouse IgG
1 NNG-811 Abcam)
anti-120572-fetoprotein (AFP 1 100 mouse IgG2b 2A9 Abcam)
anti-120572-smooth muscle actin (SMA 1 100 mouse IgG2a 1A4
Abcam) anti-nestin (1 1000 mouse IgG1 10C2 Abcam)anti-HB9 (1 100 mouse IgG1 815C10 DSHB) anti-islet-12 (ISL12 1 50 mouse IgG
2b 394DS DSHB) anti-neurofilament H nonphosphorylated (SMI32 1 500 mouseIgG1 Covance Princeton NJ USA) anti-neuron-specific
beta III tubulin (Tuj1 1 1000 rabbit polyclonal Abcam)anti-microtubule-associated protein 2 (MAP2 1 200rabbit polyclonal Millipore Billerica MA USA) anti-choline acetyltransferase (ChAT 1 1000 rabbit polyclonalAbcam) anti-synapsin 1 (1 1000 rabbit polyclonal Abcam)anti-120572-tubulin (1 500 rabbit polyclonal Abcam) andanti-acetylated 120572-tubulin (1 200 mouse IgG
2b 6-11B-1Abcam) antibodies Secondary antibodies were AlexaFluor 488-conjugated goat anti-rabbit IgG (preadsorbedAbcam) Cy3-conjugated goat anti-rabbit IgG (Abcam)FITC-conjugated goat anti-mouse IgG (Abcam) andCy3-conjugated goat anti-mouse IgG (Abcam) antibodies
211 Microfluidic Culture for Analysis of AxonalMitochondrialMovements Neurospheres derived from ESCs and iPSCswere dissociated with accutase into single cells and seededontomicrochannel plates at a density of 1 times 105 cellsplate andculturedwith neurobasal (Invitrogen) and B27medium for 10days After axons had completely stretched through the 120583m-sized grooves (total length = 8334 120583m) and reached the oppo-site compartment MNs were transfected with mito-dsRED2(Clonetech IncTakara Bio) delivered by lipofectamine 2000(Invitrogen) Within 3 days of transfection mitochondrialimages were captured by a fluorescent microscope at arate of 121 snaps2min and stacked into one file to createkymographs Mitochondrial moving velocity was calculatedby measured angle and distance in kymograph using ImageJsoftware Axonal lengths were also measured using ImageJAxonal lengths were calculated by summation of measuredlength of axon stretched out from a single 120583m-sized grooveand known length of the groove (8334 120583m)
212 Drug Treatment For analyzing protein acetylation levelcells were treated with the HDAC6 inhibitors tubastatinA (5 120583M Selleckchem Houston TX USA) CHEMICALX4 (05 120583M and 5 120583M) and X9 (05 120583M and 5 120583M) for16 hrs For analyzing axonal transport HDAC6 inhibitors
4 Stem Cells International
mentioned above were treated for 3 hrs CHEMICAL X4 andX9 are newly established HDAC6 inhibitors from ChongKun Dang Pharmaceutical Cooperation (Seoul Korea) Thecommon structures of X4 and X9 are composed of threecomponents (the Zn-binder the linker and the cap part) andare very potent hydroxamate-based HDAC6 inhibitors [IC
50
(HDAC6) = 53 and 37 nM resp] Similar to pan-HDACor other HDAC subtype specific inhibitors the hydroxamicacid moiety as a Zn-binding group is very important forblocking the catalytic activity of the HDAC6 Even thoughthese compounds have structurally different cap portions(arylurea and indole) they share the benzyl group as alinker moiety resulting in the sufficient HDAC6-specificity[fold (HDAC1HDAC6) = 400 and gt270 resp] to afford thedesirable efficacies in cells and animals without the toxicitiesassociated with the inhibitions of class I HDAC subtypes
213 Statistical Analysis Values are expressed as mean plusmnstandard error of the mean Groups were compared usingtwo-way analysis of variance using GraphPad Prism (version5 GraphPad Software Inc La Jolla CA USA) Statisticalsignificance was set at 119875 lt 005
3 Results
31 Generation of CMT2F-iPSCs and dHMN2B-iPSCsPatient-specific iPSCs were generated from one CMT2Fpatient (female52-year-old Korean) with 404CgtT (S135F)mutation and one dHMN2B patient (female8-year-oldKorean) with 545CgtT (P182L) mutation of the HSPB1 geneBoth of CMT2F and dHMN2B patients showed predominantdistal leg muscle weakness and toe gait abnormalities Theages at onset were 20 years and 7 years respectively Sensoryneve conduction velocities and action potentials of suralnerves were decreased in CMT2F patient but within normalranges in dHMN2B patient
Skin fibroblasts were reprogrammed into iPSCs by Sendaiviral transduction of four episomal vectors carrying KLF4OCT34 SOX2 and c-MYC (Figures 1(a) and 1(b)) Themorphology of CMT2F-iPSC and dHMN2B-iPSC colonieswhich resembled that of human ESCs (WA09) consisted ofcells having a high nuclear-to-cytoplasm ratio compactedin a flat cobblestone-like appearance with sharp edges (Fig-ure 1(c)) The genetic background of CMT2F-iPSCs anddHMN2B-iPSCs was not changed during the reprogram-ming processes especially at the mutation site of HSPB1(Figure 1(d)) CMT2F-iPSCs and dHMN2B-iPSCs preservedtheir normal karyotype (Figure 1(e)) The expression ofendogenous KLF4 OCT34 SOX2 and c-MYC genes wasdetected by RT-PCR after a few subpassages using primerswith complementary sequences to the intron area of thetarget mRNA (Figure 1(f)) Sendai viral genome contentswere not detected in iPSCs after cellular passage of 10 (seeSupplement Figure S1 in Supplementary Material availableonline at httpdxdoiorg10115520169475981) CMT2F-iPSCs and dHMN2B-iPSCs expressed stem cell markerssuch as NANOG in the nucleus and SSEA in the cyto-plasm (Figure 1(g)) The pluripotency of CMT2F-iPSCs anddHMN2B-iPSCs was verified by the presence of randomly
differentiated AFP-positive endodermal cells SMA-positivemesodermal cells and nestin-positive ectodermal cells via EBformation in vitro (Figure 1(h)) and teratoma formation invivo (Figure 1(i))
32 Derivation of In Vitro Models by MN Differentiation ofCMT2F-iPSCs and dHMN2B-iPSCs To recapitulate periph-eral neuropathy MNs were differentiated from iPSCs byproviding dual SMAD inhibitors (SB435142 and LDN193189)for neuralization retinoic acid for caudalization and pur-morphamine for ventralization according to the methoddescribed by Amoroso et al [12] (Figure 2(a)) Fully differen-tiatedMNs (Figure 2(b)) expressed transcription factors suchas HB9 and ISL12 cytoskeletal markers such as Tuj1 MAP2and SMI32 and synapsin and ChAT (Figure 2(c)) S135F-MNs and P182L-MNs showed no developmental defectsevidenced by no differences between S135F-MNs and P182L-MNs and control WA09-MNs and hFSiPS1-MNs in theproportion of marker-positive cells (Figure 2(d)) WA09-MNs (1236plusmn 23 120583m) and S135F-MNs (1287plusmn 20 120583m) showedno differences in axonal length (Figure 2(e)) Neuromuscularjunctions visualized by 120572-bungarotoxin staining (see Supple-ment Figure S2) formed when MNs were cocultured withmyotube cells differentiated from C2C12 cells
33 Axonal Mitochondrial Transport Defects in S135F-MNsAlthough there is heterogeneity in causative genes for differ-ent CMT2 subtypes many disease subtypes involve abnor-malities in the cellular trafficking system [13] As MNs canhave long axons up to one meter in length defects in axonaltransportation may increase vulnerability to axonopathyIn particular mitochondrial transport is extremely impor-tant for maintaining axonal and synaptic stability in neu-rons During bidirectional trafficking of mitochondria alongmicrotubules quality control is accomplished by dynamicfusion and fission processes that enable mitochondria togenerate ATP to support vital cellular functions and bufferintracellular calcium [14] Therefore we tested whetherS135F-MNs and P182L-MNs have defects in mitochondrialaxonal transport by culturing cells in microchannel plates[15] which compartmentalize axons from soma and den-drites (Figure 3(a)) transfecting cells with mito-dsRED2and analyzing kymograph images (Figure 3(b)) We observedthat the absolute velocity of mitochondrial movements wassignificantly lower in S135F-MNs (019 plusmn 001 120583msec) andslightly lower in P182L-MNs (022 plusmn 001 120583msec) comparedwith control WA09-MNs (024 plusmn 001 120583msec) and hFSiPS1-MNs (025 plusmn 001 120583msec) (Figure 3(c) Supplement FigureS3 and Supplement Table S1) Also the proportion of mov-ing mitochondria was significantly decreased in S135F-MNs(2637 plusmn 506) and P182L-MNs (1419 plusmn 214) comparedto those in control WA09-MNs (3139 plusmn 374) and hFSiPS-MNs (Figure 3(d) and Supplement Table S2)
34 Decreased Acetylation of 120572-Tubulin in S135F-MNs Ax-onal transport is regulated by various posttranslational mod-ifications (eg detyrosylation acetylation and glutamyla-tion) of microtubules through the recruitment of molecular
Stem Cells International 5
Platecells
Addvirus
Plate cells onMEF dishes
Emergingcolonies
Colony selection
20 FBSDMEM 20 KOSRDMEMF12 + bFGF
D-2 D0 D7 D8 D12 3-4 weeks
(a)
Normal HSPB1S135F HSPB1P182L
(b)
S135F-iPSCs P182L-iPSCsSNL WA09-hESCs hFSiPS1
(c)
P182L-iPSCsWA09-hESCs hFSiPS1
AG A T C A C C A T C C C A G T C A C C T T C GA G A T C AC C A T C T C A G T A C C T T C GA G A T C A C C A T C C C A G T C A C C T T C550560550560 540550
545C 545CgtT545C
WA09-hESCs S135F-iPSCshFSiPS1
T G G C T A C A T C T C C C G G T G C T T C A CT G G C T A C A T C T C C C G G T G C T T C A T G G C T A C A T C T T C C G G T G C T T C A410 420 410 420400410 420
404C 404C 404CgtT
(d)
S135F-iPSCs
1 2 3 4 5
6 7 8 9 10 11 12
13 14 15 16 17 18
19 20 21 22 X Y
P182L-iPSCs
1 2 3 4 5
6 7 8 9 10 11 12
13 14 15 16 17 18
19 20 21 22 X Y
(e)
Figure 1 Continued
6 Stem Cells International
Total-KLF4
Total-SOX2
GAPDH
Endo-KLF4Endo-OCT34
Endo-SOX2Endo-c-MYC
Total-OCT34
Total-c-MYC
WA
09-h
ESCs
S135
F-iP
SCs_
clone
1S1
35F-
iPSC
s_clo
ne 2
P182
L-iP
SCs_
clone
1P1
82L-
iPSC
s_clo
ne 2
hFSi
PS1
(f)NANOG DNA NANOGDNA SSEA4 DNA SSEA4DNA
WA
09-h
ESCs
S135
F-iP
SCs
P182
L-iP
SCs
hFSi
PS1
(g)
S135
F-iP
SCs
P182
L-iP
SCs
WA
09-h
ESCs
AFPDNA SMADNA NESTINDNA
hFSi
PS1
(h)
Figure 1 Continued
Stem Cells International 7
Mesoderm EctodermEndoderm
S135
F-iP
SCs
P182
L-iP
SCs
WA
09-h
ESCs
hFSi
PS1
Gl Ca NE
Gl CaM N
Gl Ca NE
NCa
Gl
(i)
Figure 1 Generation of CMT2F patient and dHMN2B patient-derived iPSCs (a) Experimental timeline for iPSC generation KOSRKnockOut serum replacement bFGF basic fibroblast growth factor (b) Morphology of fibroblasts from normal individual and patients(original magnification 50x) Scale bars 200 120583m (c) iPSC colonies showed ESC-like morphology such as a flat cobblestone-like appearancewith individual cells having a high nucleus-to-cytoplasm ratio (original magnification 50x) Scale bars 200120583m (d) CMT2F-iPSCs anddHMN2B-iPSCs had preserved point mutation sites in the HSPB1 gene verified by sequencing of RT-PCR products (e) CMT2F-iPSCs anddHMN2B-iPSCs maintained normal karyotype (f) Expression of total and endogenous Klf4 Oct34 Sox2 and c-Myc in CMT2F-iPSCs anddHMN2B-iPSCs was verified by RT-PCR Two clones from each of the patients-derived iPSCs were tested (clone 1 and clone 2) (g) ESCsand iPSCs expressed stem cell markers such as NANOG (in the nucleus original magnification 200x) and SSEA4 (in the cytoplasm originalmagnification 100x) Scale bars 200 120583m (h) EB-mediated in vitro spontaneous differentiation of ESCs and iPSCs resulted in the expressionof three-germ-layer markers such as AFP (endoderm) SMA (mesoderm) and nestin (ectoderm original magnification 200x) Scale bars100120583m (i) ESCs and iPSCs showed in vivo pluripotency by forming teratomas 8 weeks after subcutaneous injection into NODSCID miceTeratomas consisted of various three-germ-layer tissues including columnar gland epithelial cells with secretions [Gl] for endodermal tissuemuscle [M] and cartilage with calcification [Ca] for mesodermal tissue and neuroectodermal tissue [NE] and nerve axonal bundle [N] forectodermal tissue (original magnification 400x) Scale bars 100120583m
motor proteins [16] Among these modifications acety-lation of 120572-tubulin at the protein site of K40 regulatesbinding of dyneindynactin complexes and kinesin-1 tomicrotubules [11 17] In addition treatment with HDAC6inhibitors which increase acetylation of 120572-tubulin reversestransport defects in cellular models of Huntingtonrsquos disease[11] and LRRK2 mutation-induced Parkinsonrsquos disease [9]Therefore acetylation levels of 120572-tubulin were examined asan indicator of mitochondrial transport defects in S135F-MNs and P182L-MNs We found that S135F-MNs andP182L-MNs showed a significant reduction in acetylated 120572-tubulin levels compared with WA09-MNs or hFSiPS1-MNs(Figure 4)
35 CMT2F- and dHMN2B-Related Disease Phenotypes WereReversed by HDAC6 Inhibition By the treatment withHDAC6 inhibitors including tubastatin A (5 120583M) and newlydeveloped CHEMICAL X4 (05 120583M and 5 120583M) and X9(05 120583M and 5 120583M) acetylation of 120572-tubulin in S135F-MNs
and P182L-MNs was increased (Supplement Figure S4 Fig-ures 5(a) and 5(b)) Moreover the absolute velocity ofmitochondrial movements and the proportion of movingmitochondria in axons were increased significantly by thetreatment of these HDAC6 inhibitors (Figures 5(c) and 5(d)and Supplement Tables S1 and S2)
4 Discussion
Since iPSCs serve as an unlimited source of cells that can giverise to neuronal lineages and help to overcome the obstacleof unavailability of affected neuronal tissues from patientsdue to neuronal postmitotic property in vitro modeling anddrug screening with iPSCs can advance basic research onneurodegenerative diseases Therefore one of the strongestpoints of this study is that CMT2F- and dHMN2B-specificin vitro cellular models can be applied for the screening ofputative therapeuticmolecules such asHDAC6 inhibitors onperipheral neuropathy
8 Stem Cells International
ldquoSelf-renewalrdquo
iPSCs EBs Neurospheres MNs
3-4 wksldquoNeuralizationrdquoBMP inhibitors
ldquoCaudalizationrdquoRA
ldquoVentralizationrdquoShh agonist
(a)
WA09-MNs hFSiPS1-MNs
S135F-MNs P182L-MNs
(b)
WA09-MNs S135F-MNs P182L-MNs
ISL1
2T
uJ1
DN
AH
B9T
uJ1
DN
ASM
I32
MA
P2D
NA
SMI3
2Ch
ATD
NA
Syna
psin
DN
A
hFSiPS1-MNs
(c)
Figure 2 Continued
Stem Cells International 9
SMI32DAPI
0
50
100
150Re
lativ
e exp
ress
ion
()
WA09-MNs hFSiPS1-MNs S135F-MNs P182L-MNs
Relat
ive e
xpre
ssio
n (
)
MAP2DAPI
0
50
100
150
WA09-MNs hFSiPS1-MNs S135F-MNs P182L-MNs(d)
Axonal length
0
500
1000
1500
2000
WA09-MNs S135F-MNs
(120583m
)
(e)
Figure 2 Generation of CMT2F-specific in vitro model by differentiation of patient-derived iPSCs into MNs (a) Schematic presentation ofMN differentiation (b) Differentiated MNs showed typical cellular morphology (original magnification 50x) Scale bars 200 120583m (c) S135F-MNs and P182L-MNs expressedMN-specific transcription factors such asHB9 and ISL12 neuronal cytoskeletalmarkers such as TuJ1 SMI32andMAP2 ChAT and the synaptic vesicularmarker synapsin (originalmagnification 400x and 600x) Scale bars 100 120583m (d) Differentiationefficiency of S135F-MNs and P182L-MNs was comparable to control MNs in terms of neuronal marker expression such as SMI32DAPI andMAP2DAPI (WA09-MNs119873 = 30 hFSiPS1-MNs119873 = 329 S135F-MNs119873 = 1730 and P182L-MNs119873 = 1090) (e) Axonal length of S135F-MNs was comparable to that of control MNs Axonal length was measured by culturing fully differentiated MNs in microchannel plates foran additional 2 weeks (WA09-MNs119873 = 70 and S135F-MNs119873 = 121)
HDAC6 is a type IIb HDAC that modulates deacetylationof cytoplasmic target proteins Because HDAC6 is involvedin cytoskeletal formation modulation of cellular oxida-tionreduction state and the ubiquitin-proteasome systeminhibition of HDAC6 improves pathological conditions inneurons such as impairments in axonal transport oxidativestress and protein aggresome formation in vitro [18 19]In terms of axonal transport HDAC6 inhibitors reverseCMT2F and dHMN2B-related axonal transport by increasingacetylated 120572-tubulin In this study newly developed HDAC6inhibitors having 35-fold lower half-maximal inhibitoryconcentration (IC
50) values than that of tubastatin A showed
better performance on improving mitochondrial axonaltransport at 10 times lower concentration than tubastatin A
CMT2F-iPSCs and dHMN2B-iPSCswere generated frompatient-derived dermal fibroblasts carrying 404CgtT (S135F)and 545CgtT (P182L) mutations of HSPB1 by forced expres-sion of KLF4 OCT34 SOX2 and c-MYC using Sendai virus
vectors After establishing several stem cell colonies twocolony clones from each patient were used for further analysisto avoid differences due to clonal variance and selectediPSCs were differentiated into motor neurons with definedcytokines To recapitulate axonal pathology in vitro patient-specific motor neurons were cultured in microchanneledplates which are designed to separate the microenvironmentsof neuronal compartments In this system specific cellularmodels of CMT2F and dHMN2B showed defects in mito-chondrial transport as the absolute velocity of moving mito-chondria and the percentage of moving mitochondria werelower than control MNs Phenotypic variance between iPSCclones was observed in P182L-MNs in terms ofmitochondrialvelocity (see Figure S3) Defects in the axonal traffic systemare linked to many neurodegenerative diseases Due to thedistinct anatomy of peripheral neurons which have axons upto one meter in length the interchange of genetic materialmetabolites proteins and organelles between soma and distal
10 Stem Cells International
Original magnification 50xBar = 200120583m
(a)
WA09
P182LhFSiPS1
(b)
Absolute velocity (120583msec)
00
01
02
03
WA09-MNs hFSiPS1-MNs S135F-MNs P182L-MNs
(120583m
sec
)
lowast
lowastlowastlowast
(c)
Moving mitochondria ()
0
20
40
60
80
()
WA09-MNs hFSiPS1-MNs S135F-MNs P182L-MNs
lowastlowastlowast
lowastlowastlowast
lowastlowastlowastlowast
(d)
Figure 3 Axonal transport of mitochondria in control MNs S135F-MNs and P182L-MNs (a) Microfluidic culturing system Microchannelplates were made from transparent polymer in a specialized mold to create multicompartments connected by 120583m-sized grooves (8334 120583mlong in length) that only allow growing axons to pass through thereby separating the microenvironments of neuronal components (b)Visualization of mitochondria by Mito-dsRED2 transfection (upper image) and kymograph images for mitochondria movement (middleand lower four images) Mito-dsRED2 vector was introduced into fully differentiated MNs cultured on microchannel plates for an additional1 weekMitochondrial images were taken at a ratio of 121 snaps2min and stacked into kymograph for analysis (originalmagnification 1000x)(c) S135F-MNs show decreased velocity of moving mitochondria compared to control MNs Unpaired t-testslowast119875 lt 005 lowastlowastlowast119875 lt 0001 andlowastlowastlowastlowast119875 lt 00001 Absolute velocity of moving mitochondria was calculated from measured angle and length in kymograph images using
ImageJ (WA09-MNs 119873 = 203 hFSiPS1-MNs 119873 = 512 S135F-MNs 119873 = 428 and P182L-MNs 119873 = 496) (d) S135F-MNs and P182L-MNs show reduced moving proportion of mitochondria in an axon (WA09-MNs 119873 = 9 hFSiPS1-MNs 119873 = 6 S135F-MNs 119873 = 20 andP182L-MNs119873 = 42)
subcellular sites is critical for maintaining neuronal home-ostasis and survival Of the various trafficking cargos thetransport of mitochondria is particularly important becausemitochondria support several cellular processes by supplyingATP buffering intracellular Ca2+ and modulating apoptosis[13] In addition to delivering mitochondria to appropriatesubcellular sites the trafficking of mitochondria is vital forthe recycling of their components through dynamic fusionand fission mechanism [14] Defects in axonal mitochondrialtransport are associated with Alzheimerrsquos disease [20 21]amyotrophic lateral sclerosis [22] and Huntingtonrsquos disease[23 24]
Mitochondria are transported bidirectionally by interac-tions with molecular motor proteins such as kinesin-1 foranterogrademovement and dynein for retrogrademovementand those attach via adaptor proteins such as the mitochon-drial Rho-Milton complex and dynactin respectively [14]The recruitment of molecularmotors tomicrotubule tracks isregulated by posttranslational modification of 120572-tubulin [16]especially acetylation [17] In this study the level of acetylated120572-tubulin was reduced in both S135F-MNs and P182L-MNsBy increasing the acetylation of 120572-tubulin via treatment withHDAC6 inhibitors mitochondrial transport was improvedin terms of both velocity and the percentage of moving
Stem Cells International 11
WA09-MNs hFSiPS1 S135F-MNs P182L-MNs120572
-tubu
linAc
etyl
120572-tu
bulin
Acet
yl120572
-tubu
lin
120572-tu
bulin
DN
A
(a)
00
02
04
06
08
10
WA09-MNs hFSiPS1-MNs S135F-MNs P182L-MNs
Acet
yl120572
-tubu
lin120572
-tubu
lin
lowastlowast
lowast
lowastlowast
lowastlowast
(b)
WA09-MNs hFSiPS1-MNs S135F-MNs P182L-MNs
120572-Tubulin
Acetyl 120572-tubulin
(c)
00
02
04
06
08
10
WA09-MNs hFSiPS1-MNs S135F-MNs P182L-MNs
Acet
yl120572
-tubu
lin120572
-tubu
linlowast
lowast
lowast
lowastlowast
(d)
Figure 4 Reduced acetylation of120572-tubulin in S135F-MNs andP182-MNs (a) Acetylation of120572-tubulinwas visualized by immunostainingwithanti-acetylated 120572-tubulin Abs (red) Total 120572-tubulin was also detected by staining with anti-120572-tubulin Abs (green) (original magnification400x) Scale bars 100120583m (b) The extent of acetylated 120572-tubulin was normalized with corresponding total 120572-tubulin using ImageJ software(WA09-MNs119873 = 3 S135F-MNs119873 = 4 and P182L-MNs119873 = 5) Unpaired 119905-tests lowast119875 lt 005 and lowastlowast119875 lt 001 (c and d) Western blot assayshowed decreased acetylated 120572-tubulin levels in S135F-MNs and P182-MNs (paired 119905-tests lowast119875 lt 005 and lowastlowast119875 lt 001)
mitochondria Hence defects in the axonal trafficking ofmitochondria are associated with decreased acetylation of120572-tubulin which may be the main causative factor foraxonopathy in CMT2F and dHMN2B
In summary iPSCs can be used for valuable resourcesof in vitro disease modeling as well as drug screening Weanticipate that this in vitro system would be a useful modelfor the clinical translational research and drug development
5 Conclusions
This study demonstrates in vitro drug screening mod-els of inherited neuropathy resulting from mutations in
the HSPB1 gene can be developed from patient-specificiPSCs Axonal pathologies were mimicked in microflu-idic culturing system as both S135F-MNs and P182L-MNsshowed a marked decline in the absolute velocity andthe percentage of moving mitochondria These axonal de-fects were associated with decreased acetylation of 120572-tu-bulin and were reversed by treatment of HDAC6 in-hibitors
Competing Interests
The authors declare that they have no conflict of interests
12 Stem Cells International
HDAC6 inhibitors
S135
F-M
Ns
P182
L-M
Ns
Acetyl 120572-tubulin
Acetyl 120572-tubulin
120572-Tubulin
120572-Tubulin
DM
SO
X4_0
5120583
M
X4_5
120583M
X9_0
5120583
M
X9_5
120583M
Tuba
statin
120583M
A_5
(a)
00
05
10
15
20
25
DM
SO
X4_0
5120583
M
X4_5
120583M
X9_0
5120583
M
X9_5
120583M
lowast
lowastlowast
lowastlowast
Acet
yl120572
-tubu
lin120572
-tubu
lin
Acetyl 120572-tubulin in S135F-MNs
00
05
10
15
20
25
DM
SO
X4_0
5120583
M
X4_5
120583M
X9_0
5120583
M
X9_5
120583M
lowast
lowastlowast
lowastlowastAc
etyl
120572-tu
bulin
120572-tu
bulin
Acetyl 120572-tubulin in P182L-MNs
Tuba
statin
120583M
A_5
Tuba
statin
120583M
A_5
(b)
00
01
02
03
04
DM
SO
X4_0
5120583
M
X4_5
120583M
X9_0
5120583
M
X9_5
120583M
lowastlowastlowastlowast
lowastlowastlowastlowast
Abso
lute
velo
city
(120583m
sec
)
00
02
04
06
DM
SO
X4_0
5120583
M
X4_5
120583M
X9_0
5120583
M
X9_5
120583M
lowast
lowastlowastlowastlowast
lowastlowastlowastlowast
lowastlowastlowastlowast
lowastlowast
Abso
lute
velo
city
(120583m
sec
)
S135F-MNs
Absolute velocity
P182L-MNs
Tuba
statin
120583M
A_5
Tuba
statin
120583M
A_5
(c)
Figure 5 Continued
Stem Cells International 13
0
20
40
60
80
Mov
ing
mito
chon
dria
()
DM
SO
X4_0
5120583
M
X4_5
120583M
X4_5
120583M
X9_0
5120583
M
X9_5
120583M
lowast
lowastlowast
lowastlowast
lowastlowastlowastlowast
0
20
40
60
80
Mov
ing
mito
chon
dria
()
DM
SO
X4_0
5120583
M
X9_0
5120583
M
X9_5
120583M
lowastlowastlowastlowast
lowastlowastlowastlowast
lowastlowastlowastlowast
lowastlowastlowastlowast
S135F-MNs
Moving mitochondria
P182L-MNs
Tuba
statin
120583M
A_5
Tuba
statin
120583M
A_5
lowastlowast
(d)
Figure 5 HDAC6 inhibitors reversed axonal defect of mitochondrial transport by increasing acetylation of 120572-tubulin (a and b) Westernblot assay showed that treatment of HDAC6 inhibitors increased acetylated 120572-tubulin levels in S135F-MNs and P182L-MNs (paired 119905-testslowast119875 lt 005 and lowastlowast119875 lt 001) (c) HDAC6 inhibitors increased the absolute velocity of mitochondrial movements in S135F-MNs and P182L-MNs (WA09-MNs119873 = 617 S135F-MNs119873 = 101 and P182L-MNs119873 = 394) (d) HDAC6 inhibitors increased the percentage of movingmitochondria in S135F-MNs and P182L-MNs (WA09-MNs 119873 = 15 S135F-MNs 119873 = 7 and P182L-MNs 119873 = 15) Unpaired 119905-testslowast119875 lt 005 lowastlowast119875 lt 001 and lowastlowastlowastlowast119875 lt 00001
Authorsrsquo Contributions
Ji-Yon Kim was responsible for the iPSC experiments collec-tion and interpretation of the data and preparation of thepaper So-Youn Woo was interpreting the data and editingthe manuscript Inhee Mook-Jung provided microchannelplates and designed the experiment Heesun Choi JisooKim and Hyunjung Choi carried out microfluidic cultureand kymograph analysis Nina Ha Jangbeen Kyung andSoo Kyung Koo designed the experiments and preparationof the manuscript Young-Bin Hong and Sung-Chul Jungwere responsible for conception design of experiments andediting the manuscript Byung-Ok Choi was responsible forthe collection of patient samples and interpretation of thedata
Acknowledgments
This study was supported by the Korean Health TechnologyRampD Project Ministry of Health amp Welfare (HI12C0135HI14C3484 and HI15C1560) and a National Research Foun-dation of Korea grant funded by the Ministry of Science ICTandFuture Planning (NRF-2014R1A2A2A01004240)Humanstem cell line hFSiPS1 was provided by National Stem CellBank of Korea (Korea National Institute of Health Grant no4861-307-210-13)
References
[1] Inherited Peripheral Neuropathies Mutation Database httpwwwmolgenuaacbecmtmutationsmutationsMutByGenecfm
[2] O V Evgrafov IMersiyanova J Irobi et al ldquoMutant small heat-shock protein 27 causes axonal Charcot-Marie-Tooth diseaseand distal hereditary motor neuropathyrdquo Nature Genetics vol36 no 6 pp 602ndash606 2004
[3] J Zhai H Lin J-P Julien andWW Schlaepfer ldquoDisruption ofneurofilament network with aggregation of light neurofilamentprotein a common pathway leading to motor neuron degener-ation due to Charcot-Marie-Tooth disease-linked mutations inNFL andHSPB1rdquoHumanMolecular Genetics vol 16 no 24 pp3103ndash3116 2007
[4] L Almeida-Souza B Asselbergh C drsquoYdewalle et al ldquoSmallheat-shock protein HSPB1 mutants stabilize microtubules inCharcot-Marie-Tooth neuropathyrdquoThe Journal of Neurosciencevol 31 no 43 pp 15320ndash15328 2011
[5] A Holmgren D Bouhy V De Winter et al ldquoCharcot-Marie-Tooth causing HSPB1mutations increase Cdk5-mediated phos-phorylation of neurofilamentsrdquoActa Neuropathologica vol 126no 1 pp 93ndash108 2013
[6] A-P Arrigo ldquoThe cellular ldquonetworkingrdquo of mammalian Hsp27and its functions in the control of protein folding redox stateand apoptosisrdquoAdvances in eXperimental Medicine and Biologyvol 594 pp 14ndash26 2007
14 Stem Cells International
[7] C DrsquoYdewalle J Krishnan D M Chiheb et al ldquoHDAC6inhibitors reverse axonal loss in a mouse model of mutantHSPB1-induced Charcot-Marie-Tooth diseaserdquo NatureMedicine vol 17 no 8 pp 968ndash974 2011
[8] Y Zhang N Li C Caron et al ldquoHDAC-6 interacts with anddeacetylates tubulin and microtubules in vivordquo EMBO Journalvol 22 no 5 pp 1168ndash1179 2003
[9] A Valenzuela-Fernandez J R Cabrero J M Serrador and FSanchez-Madrid ldquoHDAC6 a key regulator of cytoskeleton cellmigration and cell-cell interactionsrdquo Trends in Cell Biology vol18 no 6 pp 291ndash297 2008
[10] V K Godena N Brookes-Hocking AMoller et al ldquoIncreasingmicrotubule acetylation rescues axonal transport and locomo-tor deficits caused by LRRK2 Roc-COR domain mutationsrdquoNature Communications vol 5 article 5245 2014
[11] J P Dompierre J D Godin B C Charrin et al ldquoHistonedeacetylase 6 inhibition compensates for the transport deficitin Huntingtonrsquos disease by increasing tubulin acetylationrdquo TheJournal of Neuroscience vol 27 no 13 pp 3571ndash3583 2007
[12] M W Amoroso G F Croft D J Williams et al ldquoAcceleratedhigh-yield generation of limb-innervating motor neurons fromhuman stem cellsrdquo Journal of Neuroscience vol 33 no 2 pp574ndash586 2013
[13] B J Gentil and L Cooper ldquoMolecular basis of axonal dysfunc-tion and traffic impairments in CMTrdquo Brain Research Bulletinvol 88 no 5 pp 444ndash453 2012
[14] Z-H Sheng and Q Cai ldquoMitochondrial transport in neu-rons impact on synaptic homeostasis and neurodegenerationrdquoNature Reviews Neuroscience vol 13 no 2 pp 77ndash93 2012
[15] J W Park B Vahidi A M Taylor S W Rhee and N LJeon ldquoMicrofluidic culture platform for neuroscience researchrdquoNature Protocols vol 1 no 4 pp 2128ndash2136 2006
[16] SWestermann and KWeber ldquoPost-translational modificationsregulate microtubule functionrdquo Nature Reviews Molecular CellBiology vol 4 no 12 pp 938ndash947 2003
[17] N A Reed D Cai T L Blasius et al ldquoMicrotubule acetylationpromotes kinesin-1 binding and transportrdquoCurrent Biology vol16 no 21 pp 2166ndash2172 2006
[18] C drsquoYdewalle E Bogaert and L Van Den Bosch ldquoHDAC6at the intersection of neuroprotection and neurodegenerationrdquoTraffic vol 13 no 6 pp 771ndash779 2012
[19] C Simoes-Pires V Zwick A Nurisso E Schenker P-ACarrupt and M Cuendet ldquoHDAC6 as a target for neurode-generative diseases what makes it different from the otherHDACsrdquoMolecular Neurodegeneration vol 8 article 7 2013
[20] Y Rui P Tiwari Z Xie and J Q Zheng ldquoAcute impairment ofmitochondrial trafficking by 120573-amyloid peptides in hippocam-pal neuronsrdquo Journal of Neuroscience vol 26 no 41 pp 10480ndash10487 2006
[21] K A Vossel K Zhang J Brodbeck et al ldquoTau reductionprevents A120573-induced defects in axonal transportrdquo Science vol330 no 6001 p 198 2010
[22] K J De vos A L Chapman M E Tennant et al ldquoFamilialamyotrophic lateral sclerosis-linked SOD1 mutants perturbfast axonal transport to reduce axonal mitochondria contentrdquoHumanMolecular Genetics vol 16 no 22 pp 2720ndash2728 2007
[23] E Trushina R B Dyer J D Badger II et al ldquoMutant huntingtinimpairs axonal trafficking inmammalian neurons in vivo and invitrordquo Molecular and Cellular Biology vol 24 no 18 pp 8195ndash8209 2004
[24] A L Orr S Li C-E Wang et al ldquoN-terminal mutant hunt-ingtin associates with mitochondria and impairs mitochondrialtraffickingrdquo Journal of Neuroscience vol 28 no 11 pp 2783ndash2792 2008
Submit your manuscripts athttpwwwhindawicom
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Anatomy Research International
PeptidesInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporation httpwwwhindawicom
International Journal of
Volume 2014
Zoology
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Molecular Biology International
GenomicsInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioinformaticsAdvances in
Marine BiologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Signal TransductionJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
Evolutionary BiologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Biochemistry Research International
ArchaeaHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Genetics Research International
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Advances in
Virolog y
Hindawi Publishing Corporationhttpwwwhindawicom
Nucleic AcidsJournal of
Volume 2014
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Enzyme Research
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
International Journal of
Microbiology

Stem Cells International 3
performed using Ex Taq polymerase (Takara Bio OtsuJapan) Primer sequences are KLF4 CDR (108 bp) 51015840-CTGCGG CAA AAC CTA CAC AAA-31015840 (forward) and 51015840-GCGAAT TTC CAT CCA CAG CC -31015840 (reverse) KLF4 UTR(96 bp) 51015840-CAT GGT CAA GTT CCC AAC TGA G-31015840(forward) and 51015840-CAC AGA CCC CAT CTG TTC TTT G-31015840 (reverse) OCT34 CDR (161 bp) 51015840-CAG TGC CCG AAACCCACAC-31015840 (forward) and 51015840-GGAGACCCAGCAGCCTCA AA-31015840 (reverse) OCT34 UTR (120 bp) 51015840-GAA AACCTG GAG TTT GTG CCA-31015840 (forward) and 51015840-TCA CCTTCC CTC CAA CCA GTT-31015840 (reverse) SOX2 CDR (131 bp)51015840-TAC CTC TTC CTC CCA CTC C-31015840 (forward) and 51015840-GGTAGTGCTGGGACATGTGA-31015840 (reverse) SOX2UTR(105 bp) 51015840-CCCGGTACGCTCAAAAAGAA-31015840 (forward)and 51015840-GGT TTT TGC GTG AGT GTG GAT-31015840 (reverse)c-MYC CDR (380 bp) 51015840-CGT CCT CGG ATT CTC TGCTC-31015840 (forward) and 51015840-GCTGGTGCATTTTCGGTTGT-31015840 (reverse) c-MYC UTR (328 bp) 51015840-GCG TCC TGG GAAGGG AGA TCC GGA GC-31015840 (forward) and 51015840-TTG AGGGGC ATC GTC GCG GGA GGC TG-31015840 (reverse)
27 Sanger Sequencing Pathogenic mutations (404CgtT and545CgtT) in HSPB1 gene from patients iPSCs were confirmedby Sanger sequencing using a 3730xl DNA Analyzer (Macro-gen Inc Seoul Korea) and analyzed using Sequencherv523 (GeneCodes Corporation Ann Arbor MI USA) Theprimers used for amplifying and sequencing are as follows51015840-TTT CTG AGC AGA CGT CCA GA-31015840 (forward) and 51015840-CTT TAC TTG GCG GCA GTC TC-31015840 (reverse)
28 Directed Differentiation of iPSCs into MNs To gen-erate EBs colonies of ESCs and iPSCs were enzymati-cally dissociated into small clumps and cultured in sus-pension for 2 days in a Petri dish supplemented withESCiPSC medium (KnockOut) containing 10 120583M Rho-associated kinase inhibitor Y27632 (Tocris Bioscience Bris-tol UK) 20 ngmL bFGF (Invitrogen) 10120583M SB435142(Stemgent Cambridge MA) and 02 120583M LDN193189 (Stem-gent) and penicillinstreptomycin On day 5 for caudaliza-tion retinoic acid (1 120583M Sigma) ascorbic acid (04 120583gmLSigma) brain-derived neurotrophic factor (10 ngmL RampDMinneapolis MN USA) and N2 supplement (1 Gibco)were added [12] On day 7 for ventralization the sonic hedge-hog agonist purmorphamine (1 120583M Stemgent) was addedand the dual SMAD inhibitors (SB435142 and LDN193189)were discontinued On day 17 basal medium was changedto neurobasal medium (Invitrogen) containing all previouslyindicated factors with the addition of insulin-like growthfactor-1 (10 ngmL) glial cell line-derived neurotrophic factor(10 ngmL) and B27 (2 Gibco) On day 21 neurosphereswere dissociated with accutase (Gibco) and plated onto poly-L-lysinelaminin-coated culture plates or slide chambers(Nalgene) and supplemented with neurobasal medium con-taining all previously indicated factors with the addition of120573-mercaptoethanol (25120583M Gibco) and glutamic acid (25 120583MSigma)
29 Immunoblot Assay Proteins were collected from dif-ferentiated MNs by conventional methods using RIPA lysis
buffer (150mM NaCl 10 NP-40 05 sodium deoxy-cholate 01 sodium dodecyl sulfate 50mM Tris pH 80)SDS-PAGE gel was transferred to polyvinylidene difluoridemembranes For blotting anti-acetylated 120572-tubulin (1 1000mouse IgG
2b 6-11B-1 Abcam Cambridge UK) and anti-120572-tubulin (1 1000 mouse IgG
1 DM1A Sigma) antibodies were
used Band densities were analyzed with UN-SCAN-IT gelsoftware (Silk Scientific Inc Orem UT USA)
210 Immunocytochemistry Cells were fixed with 4paraformaldehyde and blocked with 10 normal goat serum(Gibco) and 02 Triton X-100 Primary antibodies wereanti-SSEA4 (1 100 mouse IgG
3 MC-813-70 Developmental
Studies Hybridoma Bank (DSHB) Iowa City IA USA)anti-NANOG (1 500 mouse IgG
1 NNG-811 Abcam)
anti-120572-fetoprotein (AFP 1 100 mouse IgG2b 2A9 Abcam)
anti-120572-smooth muscle actin (SMA 1 100 mouse IgG2a 1A4
Abcam) anti-nestin (1 1000 mouse IgG1 10C2 Abcam)anti-HB9 (1 100 mouse IgG1 815C10 DSHB) anti-islet-12 (ISL12 1 50 mouse IgG
2b 394DS DSHB) anti-neurofilament H nonphosphorylated (SMI32 1 500 mouseIgG1 Covance Princeton NJ USA) anti-neuron-specific
beta III tubulin (Tuj1 1 1000 rabbit polyclonal Abcam)anti-microtubule-associated protein 2 (MAP2 1 200rabbit polyclonal Millipore Billerica MA USA) anti-choline acetyltransferase (ChAT 1 1000 rabbit polyclonalAbcam) anti-synapsin 1 (1 1000 rabbit polyclonal Abcam)anti-120572-tubulin (1 500 rabbit polyclonal Abcam) andanti-acetylated 120572-tubulin (1 200 mouse IgG
2b 6-11B-1Abcam) antibodies Secondary antibodies were AlexaFluor 488-conjugated goat anti-rabbit IgG (preadsorbedAbcam) Cy3-conjugated goat anti-rabbit IgG (Abcam)FITC-conjugated goat anti-mouse IgG (Abcam) andCy3-conjugated goat anti-mouse IgG (Abcam) antibodies
211 Microfluidic Culture for Analysis of AxonalMitochondrialMovements Neurospheres derived from ESCs and iPSCswere dissociated with accutase into single cells and seededontomicrochannel plates at a density of 1 times 105 cellsplate andculturedwith neurobasal (Invitrogen) and B27medium for 10days After axons had completely stretched through the 120583m-sized grooves (total length = 8334 120583m) and reached the oppo-site compartment MNs were transfected with mito-dsRED2(Clonetech IncTakara Bio) delivered by lipofectamine 2000(Invitrogen) Within 3 days of transfection mitochondrialimages were captured by a fluorescent microscope at arate of 121 snaps2min and stacked into one file to createkymographs Mitochondrial moving velocity was calculatedby measured angle and distance in kymograph using ImageJsoftware Axonal lengths were also measured using ImageJAxonal lengths were calculated by summation of measuredlength of axon stretched out from a single 120583m-sized grooveand known length of the groove (8334 120583m)
212 Drug Treatment For analyzing protein acetylation levelcells were treated with the HDAC6 inhibitors tubastatinA (5 120583M Selleckchem Houston TX USA) CHEMICALX4 (05 120583M and 5 120583M) and X9 (05 120583M and 5 120583M) for16 hrs For analyzing axonal transport HDAC6 inhibitors
4 Stem Cells International
mentioned above were treated for 3 hrs CHEMICAL X4 andX9 are newly established HDAC6 inhibitors from ChongKun Dang Pharmaceutical Cooperation (Seoul Korea) Thecommon structures of X4 and X9 are composed of threecomponents (the Zn-binder the linker and the cap part) andare very potent hydroxamate-based HDAC6 inhibitors [IC
50
(HDAC6) = 53 and 37 nM resp] Similar to pan-HDACor other HDAC subtype specific inhibitors the hydroxamicacid moiety as a Zn-binding group is very important forblocking the catalytic activity of the HDAC6 Even thoughthese compounds have structurally different cap portions(arylurea and indole) they share the benzyl group as alinker moiety resulting in the sufficient HDAC6-specificity[fold (HDAC1HDAC6) = 400 and gt270 resp] to afford thedesirable efficacies in cells and animals without the toxicitiesassociated with the inhibitions of class I HDAC subtypes
213 Statistical Analysis Values are expressed as mean plusmnstandard error of the mean Groups were compared usingtwo-way analysis of variance using GraphPad Prism (version5 GraphPad Software Inc La Jolla CA USA) Statisticalsignificance was set at 119875 lt 005
3 Results
31 Generation of CMT2F-iPSCs and dHMN2B-iPSCsPatient-specific iPSCs were generated from one CMT2Fpatient (female52-year-old Korean) with 404CgtT (S135F)mutation and one dHMN2B patient (female8-year-oldKorean) with 545CgtT (P182L) mutation of the HSPB1 geneBoth of CMT2F and dHMN2B patients showed predominantdistal leg muscle weakness and toe gait abnormalities Theages at onset were 20 years and 7 years respectively Sensoryneve conduction velocities and action potentials of suralnerves were decreased in CMT2F patient but within normalranges in dHMN2B patient
Skin fibroblasts were reprogrammed into iPSCs by Sendaiviral transduction of four episomal vectors carrying KLF4OCT34 SOX2 and c-MYC (Figures 1(a) and 1(b)) Themorphology of CMT2F-iPSC and dHMN2B-iPSC colonieswhich resembled that of human ESCs (WA09) consisted ofcells having a high nuclear-to-cytoplasm ratio compactedin a flat cobblestone-like appearance with sharp edges (Fig-ure 1(c)) The genetic background of CMT2F-iPSCs anddHMN2B-iPSCs was not changed during the reprogram-ming processes especially at the mutation site of HSPB1(Figure 1(d)) CMT2F-iPSCs and dHMN2B-iPSCs preservedtheir normal karyotype (Figure 1(e)) The expression ofendogenous KLF4 OCT34 SOX2 and c-MYC genes wasdetected by RT-PCR after a few subpassages using primerswith complementary sequences to the intron area of thetarget mRNA (Figure 1(f)) Sendai viral genome contentswere not detected in iPSCs after cellular passage of 10 (seeSupplement Figure S1 in Supplementary Material availableonline at httpdxdoiorg10115520169475981) CMT2F-iPSCs and dHMN2B-iPSCs expressed stem cell markerssuch as NANOG in the nucleus and SSEA in the cyto-plasm (Figure 1(g)) The pluripotency of CMT2F-iPSCs anddHMN2B-iPSCs was verified by the presence of randomly
differentiated AFP-positive endodermal cells SMA-positivemesodermal cells and nestin-positive ectodermal cells via EBformation in vitro (Figure 1(h)) and teratoma formation invivo (Figure 1(i))
32 Derivation of In Vitro Models by MN Differentiation ofCMT2F-iPSCs and dHMN2B-iPSCs To recapitulate periph-eral neuropathy MNs were differentiated from iPSCs byproviding dual SMAD inhibitors (SB435142 and LDN193189)for neuralization retinoic acid for caudalization and pur-morphamine for ventralization according to the methoddescribed by Amoroso et al [12] (Figure 2(a)) Fully differen-tiatedMNs (Figure 2(b)) expressed transcription factors suchas HB9 and ISL12 cytoskeletal markers such as Tuj1 MAP2and SMI32 and synapsin and ChAT (Figure 2(c)) S135F-MNs and P182L-MNs showed no developmental defectsevidenced by no differences between S135F-MNs and P182L-MNs and control WA09-MNs and hFSiPS1-MNs in theproportion of marker-positive cells (Figure 2(d)) WA09-MNs (1236plusmn 23 120583m) and S135F-MNs (1287plusmn 20 120583m) showedno differences in axonal length (Figure 2(e)) Neuromuscularjunctions visualized by 120572-bungarotoxin staining (see Supple-ment Figure S2) formed when MNs were cocultured withmyotube cells differentiated from C2C12 cells
33 Axonal Mitochondrial Transport Defects in S135F-MNsAlthough there is heterogeneity in causative genes for differ-ent CMT2 subtypes many disease subtypes involve abnor-malities in the cellular trafficking system [13] As MNs canhave long axons up to one meter in length defects in axonaltransportation may increase vulnerability to axonopathyIn particular mitochondrial transport is extremely impor-tant for maintaining axonal and synaptic stability in neu-rons During bidirectional trafficking of mitochondria alongmicrotubules quality control is accomplished by dynamicfusion and fission processes that enable mitochondria togenerate ATP to support vital cellular functions and bufferintracellular calcium [14] Therefore we tested whetherS135F-MNs and P182L-MNs have defects in mitochondrialaxonal transport by culturing cells in microchannel plates[15] which compartmentalize axons from soma and den-drites (Figure 3(a)) transfecting cells with mito-dsRED2and analyzing kymograph images (Figure 3(b)) We observedthat the absolute velocity of mitochondrial movements wassignificantly lower in S135F-MNs (019 plusmn 001 120583msec) andslightly lower in P182L-MNs (022 plusmn 001 120583msec) comparedwith control WA09-MNs (024 plusmn 001 120583msec) and hFSiPS1-MNs (025 plusmn 001 120583msec) (Figure 3(c) Supplement FigureS3 and Supplement Table S1) Also the proportion of mov-ing mitochondria was significantly decreased in S135F-MNs(2637 plusmn 506) and P182L-MNs (1419 plusmn 214) comparedto those in control WA09-MNs (3139 plusmn 374) and hFSiPS-MNs (Figure 3(d) and Supplement Table S2)
34 Decreased Acetylation of 120572-Tubulin in S135F-MNs Ax-onal transport is regulated by various posttranslational mod-ifications (eg detyrosylation acetylation and glutamyla-tion) of microtubules through the recruitment of molecular
Stem Cells International 5
Platecells
Addvirus
Plate cells onMEF dishes
Emergingcolonies
Colony selection
20 FBSDMEM 20 KOSRDMEMF12 + bFGF
D-2 D0 D7 D8 D12 3-4 weeks
(a)
Normal HSPB1S135F HSPB1P182L
(b)
S135F-iPSCs P182L-iPSCsSNL WA09-hESCs hFSiPS1
(c)
P182L-iPSCsWA09-hESCs hFSiPS1
AG A T C A C C A T C C C A G T C A C C T T C GA G A T C AC C A T C T C A G T A C C T T C GA G A T C A C C A T C C C A G T C A C C T T C550560550560 540550
545C 545CgtT545C
WA09-hESCs S135F-iPSCshFSiPS1
T G G C T A C A T C T C C C G G T G C T T C A CT G G C T A C A T C T C C C G G T G C T T C A T G G C T A C A T C T T C C G G T G C T T C A410 420 410 420400410 420
404C 404C 404CgtT
(d)
S135F-iPSCs
1 2 3 4 5
6 7 8 9 10 11 12
13 14 15 16 17 18
19 20 21 22 X Y
P182L-iPSCs
1 2 3 4 5
6 7 8 9 10 11 12
13 14 15 16 17 18
19 20 21 22 X Y
(e)
Figure 1 Continued
6 Stem Cells International
Total-KLF4
Total-SOX2
GAPDH
Endo-KLF4Endo-OCT34
Endo-SOX2Endo-c-MYC
Total-OCT34
Total-c-MYC
WA
09-h
ESCs
S135
F-iP
SCs_
clone
1S1
35F-
iPSC
s_clo
ne 2
P182
L-iP
SCs_
clone
1P1
82L-
iPSC
s_clo
ne 2
hFSi
PS1
(f)NANOG DNA NANOGDNA SSEA4 DNA SSEA4DNA
WA
09-h
ESCs
S135
F-iP
SCs
P182
L-iP
SCs
hFSi
PS1
(g)
S135
F-iP
SCs
P182
L-iP
SCs
WA
09-h
ESCs
AFPDNA SMADNA NESTINDNA
hFSi
PS1
(h)
Figure 1 Continued
Stem Cells International 7
Mesoderm EctodermEndoderm
S135
F-iP
SCs
P182
L-iP
SCs
WA
09-h
ESCs
hFSi
PS1
Gl Ca NE
Gl CaM N
Gl Ca NE
NCa
Gl
(i)
Figure 1 Generation of CMT2F patient and dHMN2B patient-derived iPSCs (a) Experimental timeline for iPSC generation KOSRKnockOut serum replacement bFGF basic fibroblast growth factor (b) Morphology of fibroblasts from normal individual and patients(original magnification 50x) Scale bars 200 120583m (c) iPSC colonies showed ESC-like morphology such as a flat cobblestone-like appearancewith individual cells having a high nucleus-to-cytoplasm ratio (original magnification 50x) Scale bars 200120583m (d) CMT2F-iPSCs anddHMN2B-iPSCs had preserved point mutation sites in the HSPB1 gene verified by sequencing of RT-PCR products (e) CMT2F-iPSCs anddHMN2B-iPSCs maintained normal karyotype (f) Expression of total and endogenous Klf4 Oct34 Sox2 and c-Myc in CMT2F-iPSCs anddHMN2B-iPSCs was verified by RT-PCR Two clones from each of the patients-derived iPSCs were tested (clone 1 and clone 2) (g) ESCsand iPSCs expressed stem cell markers such as NANOG (in the nucleus original magnification 200x) and SSEA4 (in the cytoplasm originalmagnification 100x) Scale bars 200 120583m (h) EB-mediated in vitro spontaneous differentiation of ESCs and iPSCs resulted in the expressionof three-germ-layer markers such as AFP (endoderm) SMA (mesoderm) and nestin (ectoderm original magnification 200x) Scale bars100120583m (i) ESCs and iPSCs showed in vivo pluripotency by forming teratomas 8 weeks after subcutaneous injection into NODSCID miceTeratomas consisted of various three-germ-layer tissues including columnar gland epithelial cells with secretions [Gl] for endodermal tissuemuscle [M] and cartilage with calcification [Ca] for mesodermal tissue and neuroectodermal tissue [NE] and nerve axonal bundle [N] forectodermal tissue (original magnification 400x) Scale bars 100120583m
motor proteins [16] Among these modifications acety-lation of 120572-tubulin at the protein site of K40 regulatesbinding of dyneindynactin complexes and kinesin-1 tomicrotubules [11 17] In addition treatment with HDAC6inhibitors which increase acetylation of 120572-tubulin reversestransport defects in cellular models of Huntingtonrsquos disease[11] and LRRK2 mutation-induced Parkinsonrsquos disease [9]Therefore acetylation levels of 120572-tubulin were examined asan indicator of mitochondrial transport defects in S135F-MNs and P182L-MNs We found that S135F-MNs andP182L-MNs showed a significant reduction in acetylated 120572-tubulin levels compared with WA09-MNs or hFSiPS1-MNs(Figure 4)
35 CMT2F- and dHMN2B-Related Disease Phenotypes WereReversed by HDAC6 Inhibition By the treatment withHDAC6 inhibitors including tubastatin A (5 120583M) and newlydeveloped CHEMICAL X4 (05 120583M and 5 120583M) and X9(05 120583M and 5 120583M) acetylation of 120572-tubulin in S135F-MNs
and P182L-MNs was increased (Supplement Figure S4 Fig-ures 5(a) and 5(b)) Moreover the absolute velocity ofmitochondrial movements and the proportion of movingmitochondria in axons were increased significantly by thetreatment of these HDAC6 inhibitors (Figures 5(c) and 5(d)and Supplement Tables S1 and S2)
4 Discussion
Since iPSCs serve as an unlimited source of cells that can giverise to neuronal lineages and help to overcome the obstacleof unavailability of affected neuronal tissues from patientsdue to neuronal postmitotic property in vitro modeling anddrug screening with iPSCs can advance basic research onneurodegenerative diseases Therefore one of the strongestpoints of this study is that CMT2F- and dHMN2B-specificin vitro cellular models can be applied for the screening ofputative therapeuticmolecules such asHDAC6 inhibitors onperipheral neuropathy
8 Stem Cells International
ldquoSelf-renewalrdquo
iPSCs EBs Neurospheres MNs
3-4 wksldquoNeuralizationrdquoBMP inhibitors
ldquoCaudalizationrdquoRA
ldquoVentralizationrdquoShh agonist
(a)
WA09-MNs hFSiPS1-MNs
S135F-MNs P182L-MNs
(b)
WA09-MNs S135F-MNs P182L-MNs
ISL1
2T
uJ1
DN
AH
B9T
uJ1
DN
ASM
I32
MA
P2D
NA
SMI3
2Ch
ATD
NA
Syna
psin
DN
A
hFSiPS1-MNs
(c)
Figure 2 Continued
Stem Cells International 9
SMI32DAPI
0
50
100
150Re
lativ
e exp
ress
ion
()
WA09-MNs hFSiPS1-MNs S135F-MNs P182L-MNs
Relat
ive e
xpre
ssio
n (
)
MAP2DAPI
0
50
100
150
WA09-MNs hFSiPS1-MNs S135F-MNs P182L-MNs(d)
Axonal length
0
500
1000
1500
2000
WA09-MNs S135F-MNs
(120583m
)
(e)
Figure 2 Generation of CMT2F-specific in vitro model by differentiation of patient-derived iPSCs into MNs (a) Schematic presentation ofMN differentiation (b) Differentiated MNs showed typical cellular morphology (original magnification 50x) Scale bars 200 120583m (c) S135F-MNs and P182L-MNs expressedMN-specific transcription factors such asHB9 and ISL12 neuronal cytoskeletalmarkers such as TuJ1 SMI32andMAP2 ChAT and the synaptic vesicularmarker synapsin (originalmagnification 400x and 600x) Scale bars 100 120583m (d) Differentiationefficiency of S135F-MNs and P182L-MNs was comparable to control MNs in terms of neuronal marker expression such as SMI32DAPI andMAP2DAPI (WA09-MNs119873 = 30 hFSiPS1-MNs119873 = 329 S135F-MNs119873 = 1730 and P182L-MNs119873 = 1090) (e) Axonal length of S135F-MNs was comparable to that of control MNs Axonal length was measured by culturing fully differentiated MNs in microchannel plates foran additional 2 weeks (WA09-MNs119873 = 70 and S135F-MNs119873 = 121)
HDAC6 is a type IIb HDAC that modulates deacetylationof cytoplasmic target proteins Because HDAC6 is involvedin cytoskeletal formation modulation of cellular oxida-tionreduction state and the ubiquitin-proteasome systeminhibition of HDAC6 improves pathological conditions inneurons such as impairments in axonal transport oxidativestress and protein aggresome formation in vitro [18 19]In terms of axonal transport HDAC6 inhibitors reverseCMT2F and dHMN2B-related axonal transport by increasingacetylated 120572-tubulin In this study newly developed HDAC6inhibitors having 35-fold lower half-maximal inhibitoryconcentration (IC
50) values than that of tubastatin A showed
better performance on improving mitochondrial axonaltransport at 10 times lower concentration than tubastatin A
CMT2F-iPSCs and dHMN2B-iPSCswere generated frompatient-derived dermal fibroblasts carrying 404CgtT (S135F)and 545CgtT (P182L) mutations of HSPB1 by forced expres-sion of KLF4 OCT34 SOX2 and c-MYC using Sendai virus
vectors After establishing several stem cell colonies twocolony clones from each patient were used for further analysisto avoid differences due to clonal variance and selectediPSCs were differentiated into motor neurons with definedcytokines To recapitulate axonal pathology in vitro patient-specific motor neurons were cultured in microchanneledplates which are designed to separate the microenvironmentsof neuronal compartments In this system specific cellularmodels of CMT2F and dHMN2B showed defects in mito-chondrial transport as the absolute velocity of moving mito-chondria and the percentage of moving mitochondria werelower than control MNs Phenotypic variance between iPSCclones was observed in P182L-MNs in terms ofmitochondrialvelocity (see Figure S3) Defects in the axonal traffic systemare linked to many neurodegenerative diseases Due to thedistinct anatomy of peripheral neurons which have axons upto one meter in length the interchange of genetic materialmetabolites proteins and organelles between soma and distal
10 Stem Cells International
Original magnification 50xBar = 200120583m
(a)
WA09
P182LhFSiPS1
(b)
Absolute velocity (120583msec)
00
01
02
03
WA09-MNs hFSiPS1-MNs S135F-MNs P182L-MNs
(120583m
sec
)
lowast
lowastlowastlowast
(c)
Moving mitochondria ()
0
20
40
60
80
()
WA09-MNs hFSiPS1-MNs S135F-MNs P182L-MNs
lowastlowastlowast
lowastlowastlowast
lowastlowastlowastlowast
(d)
Figure 3 Axonal transport of mitochondria in control MNs S135F-MNs and P182L-MNs (a) Microfluidic culturing system Microchannelplates were made from transparent polymer in a specialized mold to create multicompartments connected by 120583m-sized grooves (8334 120583mlong in length) that only allow growing axons to pass through thereby separating the microenvironments of neuronal components (b)Visualization of mitochondria by Mito-dsRED2 transfection (upper image) and kymograph images for mitochondria movement (middleand lower four images) Mito-dsRED2 vector was introduced into fully differentiated MNs cultured on microchannel plates for an additional1 weekMitochondrial images were taken at a ratio of 121 snaps2min and stacked into kymograph for analysis (originalmagnification 1000x)(c) S135F-MNs show decreased velocity of moving mitochondria compared to control MNs Unpaired t-testslowast119875 lt 005 lowastlowastlowast119875 lt 0001 andlowastlowastlowastlowast119875 lt 00001 Absolute velocity of moving mitochondria was calculated from measured angle and length in kymograph images using
ImageJ (WA09-MNs 119873 = 203 hFSiPS1-MNs 119873 = 512 S135F-MNs 119873 = 428 and P182L-MNs 119873 = 496) (d) S135F-MNs and P182L-MNs show reduced moving proportion of mitochondria in an axon (WA09-MNs 119873 = 9 hFSiPS1-MNs 119873 = 6 S135F-MNs 119873 = 20 andP182L-MNs119873 = 42)
subcellular sites is critical for maintaining neuronal home-ostasis and survival Of the various trafficking cargos thetransport of mitochondria is particularly important becausemitochondria support several cellular processes by supplyingATP buffering intracellular Ca2+ and modulating apoptosis[13] In addition to delivering mitochondria to appropriatesubcellular sites the trafficking of mitochondria is vital forthe recycling of their components through dynamic fusionand fission mechanism [14] Defects in axonal mitochondrialtransport are associated with Alzheimerrsquos disease [20 21]amyotrophic lateral sclerosis [22] and Huntingtonrsquos disease[23 24]
Mitochondria are transported bidirectionally by interac-tions with molecular motor proteins such as kinesin-1 foranterogrademovement and dynein for retrogrademovementand those attach via adaptor proteins such as the mitochon-drial Rho-Milton complex and dynactin respectively [14]The recruitment of molecularmotors tomicrotubule tracks isregulated by posttranslational modification of 120572-tubulin [16]especially acetylation [17] In this study the level of acetylated120572-tubulin was reduced in both S135F-MNs and P182L-MNsBy increasing the acetylation of 120572-tubulin via treatment withHDAC6 inhibitors mitochondrial transport was improvedin terms of both velocity and the percentage of moving
Stem Cells International 11
WA09-MNs hFSiPS1 S135F-MNs P182L-MNs120572
-tubu
linAc
etyl
120572-tu
bulin
Acet
yl120572
-tubu
lin
120572-tu
bulin
DN
A
(a)
00
02
04
06
08
10
WA09-MNs hFSiPS1-MNs S135F-MNs P182L-MNs
Acet
yl120572
-tubu
lin120572
-tubu
lin
lowastlowast
lowast
lowastlowast
lowastlowast
(b)
WA09-MNs hFSiPS1-MNs S135F-MNs P182L-MNs
120572-Tubulin
Acetyl 120572-tubulin
(c)
00
02
04
06
08
10
WA09-MNs hFSiPS1-MNs S135F-MNs P182L-MNs
Acet
yl120572
-tubu
lin120572
-tubu
linlowast
lowast
lowast
lowastlowast
(d)
Figure 4 Reduced acetylation of120572-tubulin in S135F-MNs andP182-MNs (a) Acetylation of120572-tubulinwas visualized by immunostainingwithanti-acetylated 120572-tubulin Abs (red) Total 120572-tubulin was also detected by staining with anti-120572-tubulin Abs (green) (original magnification400x) Scale bars 100120583m (b) The extent of acetylated 120572-tubulin was normalized with corresponding total 120572-tubulin using ImageJ software(WA09-MNs119873 = 3 S135F-MNs119873 = 4 and P182L-MNs119873 = 5) Unpaired 119905-tests lowast119875 lt 005 and lowastlowast119875 lt 001 (c and d) Western blot assayshowed decreased acetylated 120572-tubulin levels in S135F-MNs and P182-MNs (paired 119905-tests lowast119875 lt 005 and lowastlowast119875 lt 001)
mitochondria Hence defects in the axonal trafficking ofmitochondria are associated with decreased acetylation of120572-tubulin which may be the main causative factor foraxonopathy in CMT2F and dHMN2B
In summary iPSCs can be used for valuable resourcesof in vitro disease modeling as well as drug screening Weanticipate that this in vitro system would be a useful modelfor the clinical translational research and drug development
5 Conclusions
This study demonstrates in vitro drug screening mod-els of inherited neuropathy resulting from mutations in
the HSPB1 gene can be developed from patient-specificiPSCs Axonal pathologies were mimicked in microflu-idic culturing system as both S135F-MNs and P182L-MNsshowed a marked decline in the absolute velocity andthe percentage of moving mitochondria These axonal de-fects were associated with decreased acetylation of 120572-tu-bulin and were reversed by treatment of HDAC6 in-hibitors
Competing Interests
The authors declare that they have no conflict of interests
12 Stem Cells International
HDAC6 inhibitors
S135
F-M
Ns
P182
L-M
Ns
Acetyl 120572-tubulin
Acetyl 120572-tubulin
120572-Tubulin
120572-Tubulin
DM
SO
X4_0
5120583
M
X4_5
120583M
X9_0
5120583
M
X9_5
120583M
Tuba
statin
120583M
A_5
(a)
00
05
10
15
20
25
DM
SO
X4_0
5120583
M
X4_5
120583M
X9_0
5120583
M
X9_5
120583M
lowast
lowastlowast
lowastlowast
Acet
yl120572
-tubu
lin120572
-tubu
lin
Acetyl 120572-tubulin in S135F-MNs
00
05
10
15
20
25
DM
SO
X4_0
5120583
M
X4_5
120583M
X9_0
5120583
M
X9_5
120583M
lowast
lowastlowast
lowastlowastAc
etyl
120572-tu
bulin
120572-tu
bulin
Acetyl 120572-tubulin in P182L-MNs
Tuba
statin
120583M
A_5
Tuba
statin
120583M
A_5
(b)
00
01
02
03
04
DM
SO
X4_0
5120583
M
X4_5
120583M
X9_0
5120583
M
X9_5
120583M
lowastlowastlowastlowast
lowastlowastlowastlowast
Abso
lute
velo
city
(120583m
sec
)
00
02
04
06
DM
SO
X4_0
5120583
M
X4_5
120583M
X9_0
5120583
M
X9_5
120583M
lowast
lowastlowastlowastlowast
lowastlowastlowastlowast
lowastlowastlowastlowast
lowastlowast
Abso
lute
velo
city
(120583m
sec
)
S135F-MNs
Absolute velocity
P182L-MNs
Tuba
statin
120583M
A_5
Tuba
statin
120583M
A_5
(c)
Figure 5 Continued
Stem Cells International 13
0
20
40
60
80
Mov
ing
mito
chon
dria
()
DM
SO
X4_0
5120583
M
X4_5
120583M
X4_5
120583M
X9_0
5120583
M
X9_5
120583M
lowast
lowastlowast
lowastlowast
lowastlowastlowastlowast
0
20
40
60
80
Mov
ing
mito
chon
dria
()
DM
SO
X4_0
5120583
M
X9_0
5120583
M
X9_5
120583M
lowastlowastlowastlowast
lowastlowastlowastlowast
lowastlowastlowastlowast
lowastlowastlowastlowast
S135F-MNs
Moving mitochondria
P182L-MNs
Tuba
statin
120583M
A_5
Tuba
statin
120583M
A_5
lowastlowast
(d)
Figure 5 HDAC6 inhibitors reversed axonal defect of mitochondrial transport by increasing acetylation of 120572-tubulin (a and b) Westernblot assay showed that treatment of HDAC6 inhibitors increased acetylated 120572-tubulin levels in S135F-MNs and P182L-MNs (paired 119905-testslowast119875 lt 005 and lowastlowast119875 lt 001) (c) HDAC6 inhibitors increased the absolute velocity of mitochondrial movements in S135F-MNs and P182L-MNs (WA09-MNs119873 = 617 S135F-MNs119873 = 101 and P182L-MNs119873 = 394) (d) HDAC6 inhibitors increased the percentage of movingmitochondria in S135F-MNs and P182L-MNs (WA09-MNs 119873 = 15 S135F-MNs 119873 = 7 and P182L-MNs 119873 = 15) Unpaired 119905-testslowast119875 lt 005 lowastlowast119875 lt 001 and lowastlowastlowastlowast119875 lt 00001
Authorsrsquo Contributions
Ji-Yon Kim was responsible for the iPSC experiments collec-tion and interpretation of the data and preparation of thepaper So-Youn Woo was interpreting the data and editingthe manuscript Inhee Mook-Jung provided microchannelplates and designed the experiment Heesun Choi JisooKim and Hyunjung Choi carried out microfluidic cultureand kymograph analysis Nina Ha Jangbeen Kyung andSoo Kyung Koo designed the experiments and preparationof the manuscript Young-Bin Hong and Sung-Chul Jungwere responsible for conception design of experiments andediting the manuscript Byung-Ok Choi was responsible forthe collection of patient samples and interpretation of thedata
Acknowledgments
This study was supported by the Korean Health TechnologyRampD Project Ministry of Health amp Welfare (HI12C0135HI14C3484 and HI15C1560) and a National Research Foun-dation of Korea grant funded by the Ministry of Science ICTandFuture Planning (NRF-2014R1A2A2A01004240)Humanstem cell line hFSiPS1 was provided by National Stem CellBank of Korea (Korea National Institute of Health Grant no4861-307-210-13)
References
[1] Inherited Peripheral Neuropathies Mutation Database httpwwwmolgenuaacbecmtmutationsmutationsMutByGenecfm
[2] O V Evgrafov IMersiyanova J Irobi et al ldquoMutant small heat-shock protein 27 causes axonal Charcot-Marie-Tooth diseaseand distal hereditary motor neuropathyrdquo Nature Genetics vol36 no 6 pp 602ndash606 2004
[3] J Zhai H Lin J-P Julien andWW Schlaepfer ldquoDisruption ofneurofilament network with aggregation of light neurofilamentprotein a common pathway leading to motor neuron degener-ation due to Charcot-Marie-Tooth disease-linked mutations inNFL andHSPB1rdquoHumanMolecular Genetics vol 16 no 24 pp3103ndash3116 2007
[4] L Almeida-Souza B Asselbergh C drsquoYdewalle et al ldquoSmallheat-shock protein HSPB1 mutants stabilize microtubules inCharcot-Marie-Tooth neuropathyrdquoThe Journal of Neurosciencevol 31 no 43 pp 15320ndash15328 2011
[5] A Holmgren D Bouhy V De Winter et al ldquoCharcot-Marie-Tooth causing HSPB1mutations increase Cdk5-mediated phos-phorylation of neurofilamentsrdquoActa Neuropathologica vol 126no 1 pp 93ndash108 2013
[6] A-P Arrigo ldquoThe cellular ldquonetworkingrdquo of mammalian Hsp27and its functions in the control of protein folding redox stateand apoptosisrdquoAdvances in eXperimental Medicine and Biologyvol 594 pp 14ndash26 2007
14 Stem Cells International
[7] C DrsquoYdewalle J Krishnan D M Chiheb et al ldquoHDAC6inhibitors reverse axonal loss in a mouse model of mutantHSPB1-induced Charcot-Marie-Tooth diseaserdquo NatureMedicine vol 17 no 8 pp 968ndash974 2011
[8] Y Zhang N Li C Caron et al ldquoHDAC-6 interacts with anddeacetylates tubulin and microtubules in vivordquo EMBO Journalvol 22 no 5 pp 1168ndash1179 2003
[9] A Valenzuela-Fernandez J R Cabrero J M Serrador and FSanchez-Madrid ldquoHDAC6 a key regulator of cytoskeleton cellmigration and cell-cell interactionsrdquo Trends in Cell Biology vol18 no 6 pp 291ndash297 2008
[10] V K Godena N Brookes-Hocking AMoller et al ldquoIncreasingmicrotubule acetylation rescues axonal transport and locomo-tor deficits caused by LRRK2 Roc-COR domain mutationsrdquoNature Communications vol 5 article 5245 2014
[11] J P Dompierre J D Godin B C Charrin et al ldquoHistonedeacetylase 6 inhibition compensates for the transport deficitin Huntingtonrsquos disease by increasing tubulin acetylationrdquo TheJournal of Neuroscience vol 27 no 13 pp 3571ndash3583 2007
[12] M W Amoroso G F Croft D J Williams et al ldquoAcceleratedhigh-yield generation of limb-innervating motor neurons fromhuman stem cellsrdquo Journal of Neuroscience vol 33 no 2 pp574ndash586 2013
[13] B J Gentil and L Cooper ldquoMolecular basis of axonal dysfunc-tion and traffic impairments in CMTrdquo Brain Research Bulletinvol 88 no 5 pp 444ndash453 2012
[14] Z-H Sheng and Q Cai ldquoMitochondrial transport in neu-rons impact on synaptic homeostasis and neurodegenerationrdquoNature Reviews Neuroscience vol 13 no 2 pp 77ndash93 2012
[15] J W Park B Vahidi A M Taylor S W Rhee and N LJeon ldquoMicrofluidic culture platform for neuroscience researchrdquoNature Protocols vol 1 no 4 pp 2128ndash2136 2006
[16] SWestermann and KWeber ldquoPost-translational modificationsregulate microtubule functionrdquo Nature Reviews Molecular CellBiology vol 4 no 12 pp 938ndash947 2003
[17] N A Reed D Cai T L Blasius et al ldquoMicrotubule acetylationpromotes kinesin-1 binding and transportrdquoCurrent Biology vol16 no 21 pp 2166ndash2172 2006
[18] C drsquoYdewalle E Bogaert and L Van Den Bosch ldquoHDAC6at the intersection of neuroprotection and neurodegenerationrdquoTraffic vol 13 no 6 pp 771ndash779 2012
[19] C Simoes-Pires V Zwick A Nurisso E Schenker P-ACarrupt and M Cuendet ldquoHDAC6 as a target for neurode-generative diseases what makes it different from the otherHDACsrdquoMolecular Neurodegeneration vol 8 article 7 2013
[20] Y Rui P Tiwari Z Xie and J Q Zheng ldquoAcute impairment ofmitochondrial trafficking by 120573-amyloid peptides in hippocam-pal neuronsrdquo Journal of Neuroscience vol 26 no 41 pp 10480ndash10487 2006
[21] K A Vossel K Zhang J Brodbeck et al ldquoTau reductionprevents A120573-induced defects in axonal transportrdquo Science vol330 no 6001 p 198 2010
[22] K J De vos A L Chapman M E Tennant et al ldquoFamilialamyotrophic lateral sclerosis-linked SOD1 mutants perturbfast axonal transport to reduce axonal mitochondria contentrdquoHumanMolecular Genetics vol 16 no 22 pp 2720ndash2728 2007
[23] E Trushina R B Dyer J D Badger II et al ldquoMutant huntingtinimpairs axonal trafficking inmammalian neurons in vivo and invitrordquo Molecular and Cellular Biology vol 24 no 18 pp 8195ndash8209 2004
[24] A L Orr S Li C-E Wang et al ldquoN-terminal mutant hunt-ingtin associates with mitochondria and impairs mitochondrialtraffickingrdquo Journal of Neuroscience vol 28 no 11 pp 2783ndash2792 2008
Submit your manuscripts athttpwwwhindawicom
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Anatomy Research International
PeptidesInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporation httpwwwhindawicom
International Journal of
Volume 2014
Zoology
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Molecular Biology International
GenomicsInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioinformaticsAdvances in
Marine BiologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Signal TransductionJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
Evolutionary BiologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Biochemistry Research International
ArchaeaHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Genetics Research International
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Advances in
Virolog y
Hindawi Publishing Corporationhttpwwwhindawicom
Nucleic AcidsJournal of
Volume 2014
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Enzyme Research
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
International Journal of
Microbiology

4 Stem Cells International
mentioned above were treated for 3 hrs CHEMICAL X4 andX9 are newly established HDAC6 inhibitors from ChongKun Dang Pharmaceutical Cooperation (Seoul Korea) Thecommon structures of X4 and X9 are composed of threecomponents (the Zn-binder the linker and the cap part) andare very potent hydroxamate-based HDAC6 inhibitors [IC
50
(HDAC6) = 53 and 37 nM resp] Similar to pan-HDACor other HDAC subtype specific inhibitors the hydroxamicacid moiety as a Zn-binding group is very important forblocking the catalytic activity of the HDAC6 Even thoughthese compounds have structurally different cap portions(arylurea and indole) they share the benzyl group as alinker moiety resulting in the sufficient HDAC6-specificity[fold (HDAC1HDAC6) = 400 and gt270 resp] to afford thedesirable efficacies in cells and animals without the toxicitiesassociated with the inhibitions of class I HDAC subtypes
213 Statistical Analysis Values are expressed as mean plusmnstandard error of the mean Groups were compared usingtwo-way analysis of variance using GraphPad Prism (version5 GraphPad Software Inc La Jolla CA USA) Statisticalsignificance was set at 119875 lt 005
3 Results
31 Generation of CMT2F-iPSCs and dHMN2B-iPSCsPatient-specific iPSCs were generated from one CMT2Fpatient (female52-year-old Korean) with 404CgtT (S135F)mutation and one dHMN2B patient (female8-year-oldKorean) with 545CgtT (P182L) mutation of the HSPB1 geneBoth of CMT2F and dHMN2B patients showed predominantdistal leg muscle weakness and toe gait abnormalities Theages at onset were 20 years and 7 years respectively Sensoryneve conduction velocities and action potentials of suralnerves were decreased in CMT2F patient but within normalranges in dHMN2B patient
Skin fibroblasts were reprogrammed into iPSCs by Sendaiviral transduction of four episomal vectors carrying KLF4OCT34 SOX2 and c-MYC (Figures 1(a) and 1(b)) Themorphology of CMT2F-iPSC and dHMN2B-iPSC colonieswhich resembled that of human ESCs (WA09) consisted ofcells having a high nuclear-to-cytoplasm ratio compactedin a flat cobblestone-like appearance with sharp edges (Fig-ure 1(c)) The genetic background of CMT2F-iPSCs anddHMN2B-iPSCs was not changed during the reprogram-ming processes especially at the mutation site of HSPB1(Figure 1(d)) CMT2F-iPSCs and dHMN2B-iPSCs preservedtheir normal karyotype (Figure 1(e)) The expression ofendogenous KLF4 OCT34 SOX2 and c-MYC genes wasdetected by RT-PCR after a few subpassages using primerswith complementary sequences to the intron area of thetarget mRNA (Figure 1(f)) Sendai viral genome contentswere not detected in iPSCs after cellular passage of 10 (seeSupplement Figure S1 in Supplementary Material availableonline at httpdxdoiorg10115520169475981) CMT2F-iPSCs and dHMN2B-iPSCs expressed stem cell markerssuch as NANOG in the nucleus and SSEA in the cyto-plasm (Figure 1(g)) The pluripotency of CMT2F-iPSCs anddHMN2B-iPSCs was verified by the presence of randomly
differentiated AFP-positive endodermal cells SMA-positivemesodermal cells and nestin-positive ectodermal cells via EBformation in vitro (Figure 1(h)) and teratoma formation invivo (Figure 1(i))
32 Derivation of In Vitro Models by MN Differentiation ofCMT2F-iPSCs and dHMN2B-iPSCs To recapitulate periph-eral neuropathy MNs were differentiated from iPSCs byproviding dual SMAD inhibitors (SB435142 and LDN193189)for neuralization retinoic acid for caudalization and pur-morphamine for ventralization according to the methoddescribed by Amoroso et al [12] (Figure 2(a)) Fully differen-tiatedMNs (Figure 2(b)) expressed transcription factors suchas HB9 and ISL12 cytoskeletal markers such as Tuj1 MAP2and SMI32 and synapsin and ChAT (Figure 2(c)) S135F-MNs and P182L-MNs showed no developmental defectsevidenced by no differences between S135F-MNs and P182L-MNs and control WA09-MNs and hFSiPS1-MNs in theproportion of marker-positive cells (Figure 2(d)) WA09-MNs (1236plusmn 23 120583m) and S135F-MNs (1287plusmn 20 120583m) showedno differences in axonal length (Figure 2(e)) Neuromuscularjunctions visualized by 120572-bungarotoxin staining (see Supple-ment Figure S2) formed when MNs were cocultured withmyotube cells differentiated from C2C12 cells
33 Axonal Mitochondrial Transport Defects in S135F-MNsAlthough there is heterogeneity in causative genes for differ-ent CMT2 subtypes many disease subtypes involve abnor-malities in the cellular trafficking system [13] As MNs canhave long axons up to one meter in length defects in axonaltransportation may increase vulnerability to axonopathyIn particular mitochondrial transport is extremely impor-tant for maintaining axonal and synaptic stability in neu-rons During bidirectional trafficking of mitochondria alongmicrotubules quality control is accomplished by dynamicfusion and fission processes that enable mitochondria togenerate ATP to support vital cellular functions and bufferintracellular calcium [14] Therefore we tested whetherS135F-MNs and P182L-MNs have defects in mitochondrialaxonal transport by culturing cells in microchannel plates[15] which compartmentalize axons from soma and den-drites (Figure 3(a)) transfecting cells with mito-dsRED2and analyzing kymograph images (Figure 3(b)) We observedthat the absolute velocity of mitochondrial movements wassignificantly lower in S135F-MNs (019 plusmn 001 120583msec) andslightly lower in P182L-MNs (022 plusmn 001 120583msec) comparedwith control WA09-MNs (024 plusmn 001 120583msec) and hFSiPS1-MNs (025 plusmn 001 120583msec) (Figure 3(c) Supplement FigureS3 and Supplement Table S1) Also the proportion of mov-ing mitochondria was significantly decreased in S135F-MNs(2637 plusmn 506) and P182L-MNs (1419 plusmn 214) comparedto those in control WA09-MNs (3139 plusmn 374) and hFSiPS-MNs (Figure 3(d) and Supplement Table S2)
34 Decreased Acetylation of 120572-Tubulin in S135F-MNs Ax-onal transport is regulated by various posttranslational mod-ifications (eg detyrosylation acetylation and glutamyla-tion) of microtubules through the recruitment of molecular
Stem Cells International 5
Platecells
Addvirus
Plate cells onMEF dishes
Emergingcolonies
Colony selection
20 FBSDMEM 20 KOSRDMEMF12 + bFGF
D-2 D0 D7 D8 D12 3-4 weeks
(a)
Normal HSPB1S135F HSPB1P182L
(b)
S135F-iPSCs P182L-iPSCsSNL WA09-hESCs hFSiPS1
(c)
P182L-iPSCsWA09-hESCs hFSiPS1
AG A T C A C C A T C C C A G T C A C C T T C GA G A T C AC C A T C T C A G T A C C T T C GA G A T C A C C A T C C C A G T C A C C T T C550560550560 540550
545C 545CgtT545C
WA09-hESCs S135F-iPSCshFSiPS1
T G G C T A C A T C T C C C G G T G C T T C A CT G G C T A C A T C T C C C G G T G C T T C A T G G C T A C A T C T T C C G G T G C T T C A410 420 410 420400410 420
404C 404C 404CgtT
(d)
S135F-iPSCs
1 2 3 4 5
6 7 8 9 10 11 12
13 14 15 16 17 18
19 20 21 22 X Y
P182L-iPSCs
1 2 3 4 5
6 7 8 9 10 11 12
13 14 15 16 17 18
19 20 21 22 X Y
(e)
Figure 1 Continued
6 Stem Cells International
Total-KLF4
Total-SOX2
GAPDH
Endo-KLF4Endo-OCT34
Endo-SOX2Endo-c-MYC
Total-OCT34
Total-c-MYC
WA
09-h
ESCs
S135
F-iP
SCs_
clone
1S1
35F-
iPSC
s_clo
ne 2
P182
L-iP
SCs_
clone
1P1
82L-
iPSC
s_clo
ne 2
hFSi
PS1
(f)NANOG DNA NANOGDNA SSEA4 DNA SSEA4DNA
WA
09-h
ESCs
S135
F-iP
SCs
P182
L-iP
SCs
hFSi
PS1
(g)
S135
F-iP
SCs
P182
L-iP
SCs
WA
09-h
ESCs
AFPDNA SMADNA NESTINDNA
hFSi
PS1
(h)
Figure 1 Continued
Stem Cells International 7
Mesoderm EctodermEndoderm
S135
F-iP
SCs
P182
L-iP
SCs
WA
09-h
ESCs
hFSi
PS1
Gl Ca NE
Gl CaM N
Gl Ca NE
NCa
Gl
(i)
Figure 1 Generation of CMT2F patient and dHMN2B patient-derived iPSCs (a) Experimental timeline for iPSC generation KOSRKnockOut serum replacement bFGF basic fibroblast growth factor (b) Morphology of fibroblasts from normal individual and patients(original magnification 50x) Scale bars 200 120583m (c) iPSC colonies showed ESC-like morphology such as a flat cobblestone-like appearancewith individual cells having a high nucleus-to-cytoplasm ratio (original magnification 50x) Scale bars 200120583m (d) CMT2F-iPSCs anddHMN2B-iPSCs had preserved point mutation sites in the HSPB1 gene verified by sequencing of RT-PCR products (e) CMT2F-iPSCs anddHMN2B-iPSCs maintained normal karyotype (f) Expression of total and endogenous Klf4 Oct34 Sox2 and c-Myc in CMT2F-iPSCs anddHMN2B-iPSCs was verified by RT-PCR Two clones from each of the patients-derived iPSCs were tested (clone 1 and clone 2) (g) ESCsand iPSCs expressed stem cell markers such as NANOG (in the nucleus original magnification 200x) and SSEA4 (in the cytoplasm originalmagnification 100x) Scale bars 200 120583m (h) EB-mediated in vitro spontaneous differentiation of ESCs and iPSCs resulted in the expressionof three-germ-layer markers such as AFP (endoderm) SMA (mesoderm) and nestin (ectoderm original magnification 200x) Scale bars100120583m (i) ESCs and iPSCs showed in vivo pluripotency by forming teratomas 8 weeks after subcutaneous injection into NODSCID miceTeratomas consisted of various three-germ-layer tissues including columnar gland epithelial cells with secretions [Gl] for endodermal tissuemuscle [M] and cartilage with calcification [Ca] for mesodermal tissue and neuroectodermal tissue [NE] and nerve axonal bundle [N] forectodermal tissue (original magnification 400x) Scale bars 100120583m
motor proteins [16] Among these modifications acety-lation of 120572-tubulin at the protein site of K40 regulatesbinding of dyneindynactin complexes and kinesin-1 tomicrotubules [11 17] In addition treatment with HDAC6inhibitors which increase acetylation of 120572-tubulin reversestransport defects in cellular models of Huntingtonrsquos disease[11] and LRRK2 mutation-induced Parkinsonrsquos disease [9]Therefore acetylation levels of 120572-tubulin were examined asan indicator of mitochondrial transport defects in S135F-MNs and P182L-MNs We found that S135F-MNs andP182L-MNs showed a significant reduction in acetylated 120572-tubulin levels compared with WA09-MNs or hFSiPS1-MNs(Figure 4)
35 CMT2F- and dHMN2B-Related Disease Phenotypes WereReversed by HDAC6 Inhibition By the treatment withHDAC6 inhibitors including tubastatin A (5 120583M) and newlydeveloped CHEMICAL X4 (05 120583M and 5 120583M) and X9(05 120583M and 5 120583M) acetylation of 120572-tubulin in S135F-MNs
and P182L-MNs was increased (Supplement Figure S4 Fig-ures 5(a) and 5(b)) Moreover the absolute velocity ofmitochondrial movements and the proportion of movingmitochondria in axons were increased significantly by thetreatment of these HDAC6 inhibitors (Figures 5(c) and 5(d)and Supplement Tables S1 and S2)
4 Discussion
Since iPSCs serve as an unlimited source of cells that can giverise to neuronal lineages and help to overcome the obstacleof unavailability of affected neuronal tissues from patientsdue to neuronal postmitotic property in vitro modeling anddrug screening with iPSCs can advance basic research onneurodegenerative diseases Therefore one of the strongestpoints of this study is that CMT2F- and dHMN2B-specificin vitro cellular models can be applied for the screening ofputative therapeuticmolecules such asHDAC6 inhibitors onperipheral neuropathy
8 Stem Cells International
ldquoSelf-renewalrdquo
iPSCs EBs Neurospheres MNs
3-4 wksldquoNeuralizationrdquoBMP inhibitors
ldquoCaudalizationrdquoRA
ldquoVentralizationrdquoShh agonist
(a)
WA09-MNs hFSiPS1-MNs
S135F-MNs P182L-MNs
(b)
WA09-MNs S135F-MNs P182L-MNs
ISL1
2T
uJ1
DN
AH
B9T
uJ1
DN
ASM
I32
MA
P2D
NA
SMI3
2Ch
ATD
NA
Syna
psin
DN
A
hFSiPS1-MNs
(c)
Figure 2 Continued
Stem Cells International 9
SMI32DAPI
0
50
100
150Re
lativ
e exp
ress
ion
()
WA09-MNs hFSiPS1-MNs S135F-MNs P182L-MNs
Relat
ive e
xpre
ssio
n (
)
MAP2DAPI
0
50
100
150
WA09-MNs hFSiPS1-MNs S135F-MNs P182L-MNs(d)
Axonal length
0
500
1000
1500
2000
WA09-MNs S135F-MNs
(120583m
)
(e)
Figure 2 Generation of CMT2F-specific in vitro model by differentiation of patient-derived iPSCs into MNs (a) Schematic presentation ofMN differentiation (b) Differentiated MNs showed typical cellular morphology (original magnification 50x) Scale bars 200 120583m (c) S135F-MNs and P182L-MNs expressedMN-specific transcription factors such asHB9 and ISL12 neuronal cytoskeletalmarkers such as TuJ1 SMI32andMAP2 ChAT and the synaptic vesicularmarker synapsin (originalmagnification 400x and 600x) Scale bars 100 120583m (d) Differentiationefficiency of S135F-MNs and P182L-MNs was comparable to control MNs in terms of neuronal marker expression such as SMI32DAPI andMAP2DAPI (WA09-MNs119873 = 30 hFSiPS1-MNs119873 = 329 S135F-MNs119873 = 1730 and P182L-MNs119873 = 1090) (e) Axonal length of S135F-MNs was comparable to that of control MNs Axonal length was measured by culturing fully differentiated MNs in microchannel plates foran additional 2 weeks (WA09-MNs119873 = 70 and S135F-MNs119873 = 121)
HDAC6 is a type IIb HDAC that modulates deacetylationof cytoplasmic target proteins Because HDAC6 is involvedin cytoskeletal formation modulation of cellular oxida-tionreduction state and the ubiquitin-proteasome systeminhibition of HDAC6 improves pathological conditions inneurons such as impairments in axonal transport oxidativestress and protein aggresome formation in vitro [18 19]In terms of axonal transport HDAC6 inhibitors reverseCMT2F and dHMN2B-related axonal transport by increasingacetylated 120572-tubulin In this study newly developed HDAC6inhibitors having 35-fold lower half-maximal inhibitoryconcentration (IC
50) values than that of tubastatin A showed
better performance on improving mitochondrial axonaltransport at 10 times lower concentration than tubastatin A
CMT2F-iPSCs and dHMN2B-iPSCswere generated frompatient-derived dermal fibroblasts carrying 404CgtT (S135F)and 545CgtT (P182L) mutations of HSPB1 by forced expres-sion of KLF4 OCT34 SOX2 and c-MYC using Sendai virus
vectors After establishing several stem cell colonies twocolony clones from each patient were used for further analysisto avoid differences due to clonal variance and selectediPSCs were differentiated into motor neurons with definedcytokines To recapitulate axonal pathology in vitro patient-specific motor neurons were cultured in microchanneledplates which are designed to separate the microenvironmentsof neuronal compartments In this system specific cellularmodels of CMT2F and dHMN2B showed defects in mito-chondrial transport as the absolute velocity of moving mito-chondria and the percentage of moving mitochondria werelower than control MNs Phenotypic variance between iPSCclones was observed in P182L-MNs in terms ofmitochondrialvelocity (see Figure S3) Defects in the axonal traffic systemare linked to many neurodegenerative diseases Due to thedistinct anatomy of peripheral neurons which have axons upto one meter in length the interchange of genetic materialmetabolites proteins and organelles between soma and distal
10 Stem Cells International
Original magnification 50xBar = 200120583m
(a)
WA09
P182LhFSiPS1
(b)
Absolute velocity (120583msec)
00
01
02
03
WA09-MNs hFSiPS1-MNs S135F-MNs P182L-MNs
(120583m
sec
)
lowast
lowastlowastlowast
(c)
Moving mitochondria ()
0
20
40
60
80
()
WA09-MNs hFSiPS1-MNs S135F-MNs P182L-MNs
lowastlowastlowast
lowastlowastlowast
lowastlowastlowastlowast
(d)
Figure 3 Axonal transport of mitochondria in control MNs S135F-MNs and P182L-MNs (a) Microfluidic culturing system Microchannelplates were made from transparent polymer in a specialized mold to create multicompartments connected by 120583m-sized grooves (8334 120583mlong in length) that only allow growing axons to pass through thereby separating the microenvironments of neuronal components (b)Visualization of mitochondria by Mito-dsRED2 transfection (upper image) and kymograph images for mitochondria movement (middleand lower four images) Mito-dsRED2 vector was introduced into fully differentiated MNs cultured on microchannel plates for an additional1 weekMitochondrial images were taken at a ratio of 121 snaps2min and stacked into kymograph for analysis (originalmagnification 1000x)(c) S135F-MNs show decreased velocity of moving mitochondria compared to control MNs Unpaired t-testslowast119875 lt 005 lowastlowastlowast119875 lt 0001 andlowastlowastlowastlowast119875 lt 00001 Absolute velocity of moving mitochondria was calculated from measured angle and length in kymograph images using
ImageJ (WA09-MNs 119873 = 203 hFSiPS1-MNs 119873 = 512 S135F-MNs 119873 = 428 and P182L-MNs 119873 = 496) (d) S135F-MNs and P182L-MNs show reduced moving proportion of mitochondria in an axon (WA09-MNs 119873 = 9 hFSiPS1-MNs 119873 = 6 S135F-MNs 119873 = 20 andP182L-MNs119873 = 42)
subcellular sites is critical for maintaining neuronal home-ostasis and survival Of the various trafficking cargos thetransport of mitochondria is particularly important becausemitochondria support several cellular processes by supplyingATP buffering intracellular Ca2+ and modulating apoptosis[13] In addition to delivering mitochondria to appropriatesubcellular sites the trafficking of mitochondria is vital forthe recycling of their components through dynamic fusionand fission mechanism [14] Defects in axonal mitochondrialtransport are associated with Alzheimerrsquos disease [20 21]amyotrophic lateral sclerosis [22] and Huntingtonrsquos disease[23 24]
Mitochondria are transported bidirectionally by interac-tions with molecular motor proteins such as kinesin-1 foranterogrademovement and dynein for retrogrademovementand those attach via adaptor proteins such as the mitochon-drial Rho-Milton complex and dynactin respectively [14]The recruitment of molecularmotors tomicrotubule tracks isregulated by posttranslational modification of 120572-tubulin [16]especially acetylation [17] In this study the level of acetylated120572-tubulin was reduced in both S135F-MNs and P182L-MNsBy increasing the acetylation of 120572-tubulin via treatment withHDAC6 inhibitors mitochondrial transport was improvedin terms of both velocity and the percentage of moving
Stem Cells International 11
WA09-MNs hFSiPS1 S135F-MNs P182L-MNs120572
-tubu
linAc
etyl
120572-tu
bulin
Acet
yl120572
-tubu
lin
120572-tu
bulin
DN
A
(a)
00
02
04
06
08
10
WA09-MNs hFSiPS1-MNs S135F-MNs P182L-MNs
Acet
yl120572
-tubu
lin120572
-tubu
lin
lowastlowast
lowast
lowastlowast
lowastlowast
(b)
WA09-MNs hFSiPS1-MNs S135F-MNs P182L-MNs
120572-Tubulin
Acetyl 120572-tubulin
(c)
00
02
04
06
08
10
WA09-MNs hFSiPS1-MNs S135F-MNs P182L-MNs
Acet
yl120572
-tubu
lin120572
-tubu
linlowast
lowast
lowast
lowastlowast
(d)
Figure 4 Reduced acetylation of120572-tubulin in S135F-MNs andP182-MNs (a) Acetylation of120572-tubulinwas visualized by immunostainingwithanti-acetylated 120572-tubulin Abs (red) Total 120572-tubulin was also detected by staining with anti-120572-tubulin Abs (green) (original magnification400x) Scale bars 100120583m (b) The extent of acetylated 120572-tubulin was normalized with corresponding total 120572-tubulin using ImageJ software(WA09-MNs119873 = 3 S135F-MNs119873 = 4 and P182L-MNs119873 = 5) Unpaired 119905-tests lowast119875 lt 005 and lowastlowast119875 lt 001 (c and d) Western blot assayshowed decreased acetylated 120572-tubulin levels in S135F-MNs and P182-MNs (paired 119905-tests lowast119875 lt 005 and lowastlowast119875 lt 001)
mitochondria Hence defects in the axonal trafficking ofmitochondria are associated with decreased acetylation of120572-tubulin which may be the main causative factor foraxonopathy in CMT2F and dHMN2B
In summary iPSCs can be used for valuable resourcesof in vitro disease modeling as well as drug screening Weanticipate that this in vitro system would be a useful modelfor the clinical translational research and drug development
5 Conclusions
This study demonstrates in vitro drug screening mod-els of inherited neuropathy resulting from mutations in
the HSPB1 gene can be developed from patient-specificiPSCs Axonal pathologies were mimicked in microflu-idic culturing system as both S135F-MNs and P182L-MNsshowed a marked decline in the absolute velocity andthe percentage of moving mitochondria These axonal de-fects were associated with decreased acetylation of 120572-tu-bulin and were reversed by treatment of HDAC6 in-hibitors
Competing Interests
The authors declare that they have no conflict of interests
12 Stem Cells International
HDAC6 inhibitors
S135
F-M
Ns
P182
L-M
Ns
Acetyl 120572-tubulin
Acetyl 120572-tubulin
120572-Tubulin
120572-Tubulin
DM
SO
X4_0
5120583
M
X4_5
120583M
X9_0
5120583
M
X9_5
120583M
Tuba
statin
120583M
A_5
(a)
00
05
10
15
20
25
DM
SO
X4_0
5120583
M
X4_5
120583M
X9_0
5120583
M
X9_5
120583M
lowast
lowastlowast
lowastlowast
Acet
yl120572
-tubu
lin120572
-tubu
lin
Acetyl 120572-tubulin in S135F-MNs
00
05
10
15
20
25
DM
SO
X4_0
5120583
M
X4_5
120583M
X9_0
5120583
M
X9_5
120583M
lowast
lowastlowast
lowastlowastAc
etyl
120572-tu
bulin
120572-tu
bulin
Acetyl 120572-tubulin in P182L-MNs
Tuba
statin
120583M
A_5
Tuba
statin
120583M
A_5
(b)
00
01
02
03
04
DM
SO
X4_0
5120583
M
X4_5
120583M
X9_0
5120583
M
X9_5
120583M
lowastlowastlowastlowast
lowastlowastlowastlowast
Abso
lute
velo
city
(120583m
sec
)
00
02
04
06
DM
SO
X4_0
5120583
M
X4_5
120583M
X9_0
5120583
M
X9_5
120583M
lowast
lowastlowastlowastlowast
lowastlowastlowastlowast
lowastlowastlowastlowast
lowastlowast
Abso
lute
velo
city
(120583m
sec
)
S135F-MNs
Absolute velocity
P182L-MNs
Tuba
statin
120583M
A_5
Tuba
statin
120583M
A_5
(c)
Figure 5 Continued
Stem Cells International 13
0
20
40
60
80
Mov
ing
mito
chon
dria
()
DM
SO
X4_0
5120583
M
X4_5
120583M
X4_5
120583M
X9_0
5120583
M
X9_5
120583M
lowast
lowastlowast
lowastlowast
lowastlowastlowastlowast
0
20
40
60
80
Mov
ing
mito
chon
dria
()
DM
SO
X4_0
5120583
M
X9_0
5120583
M
X9_5
120583M
lowastlowastlowastlowast
lowastlowastlowastlowast
lowastlowastlowastlowast
lowastlowastlowastlowast
S135F-MNs
Moving mitochondria
P182L-MNs
Tuba
statin
120583M
A_5
Tuba
statin
120583M
A_5
lowastlowast
(d)
Figure 5 HDAC6 inhibitors reversed axonal defect of mitochondrial transport by increasing acetylation of 120572-tubulin (a and b) Westernblot assay showed that treatment of HDAC6 inhibitors increased acetylated 120572-tubulin levels in S135F-MNs and P182L-MNs (paired 119905-testslowast119875 lt 005 and lowastlowast119875 lt 001) (c) HDAC6 inhibitors increased the absolute velocity of mitochondrial movements in S135F-MNs and P182L-MNs (WA09-MNs119873 = 617 S135F-MNs119873 = 101 and P182L-MNs119873 = 394) (d) HDAC6 inhibitors increased the percentage of movingmitochondria in S135F-MNs and P182L-MNs (WA09-MNs 119873 = 15 S135F-MNs 119873 = 7 and P182L-MNs 119873 = 15) Unpaired 119905-testslowast119875 lt 005 lowastlowast119875 lt 001 and lowastlowastlowastlowast119875 lt 00001
Authorsrsquo Contributions
Ji-Yon Kim was responsible for the iPSC experiments collec-tion and interpretation of the data and preparation of thepaper So-Youn Woo was interpreting the data and editingthe manuscript Inhee Mook-Jung provided microchannelplates and designed the experiment Heesun Choi JisooKim and Hyunjung Choi carried out microfluidic cultureand kymograph analysis Nina Ha Jangbeen Kyung andSoo Kyung Koo designed the experiments and preparationof the manuscript Young-Bin Hong and Sung-Chul Jungwere responsible for conception design of experiments andediting the manuscript Byung-Ok Choi was responsible forthe collection of patient samples and interpretation of thedata
Acknowledgments
This study was supported by the Korean Health TechnologyRampD Project Ministry of Health amp Welfare (HI12C0135HI14C3484 and HI15C1560) and a National Research Foun-dation of Korea grant funded by the Ministry of Science ICTandFuture Planning (NRF-2014R1A2A2A01004240)Humanstem cell line hFSiPS1 was provided by National Stem CellBank of Korea (Korea National Institute of Health Grant no4861-307-210-13)
References
[1] Inherited Peripheral Neuropathies Mutation Database httpwwwmolgenuaacbecmtmutationsmutationsMutByGenecfm
[2] O V Evgrafov IMersiyanova J Irobi et al ldquoMutant small heat-shock protein 27 causes axonal Charcot-Marie-Tooth diseaseand distal hereditary motor neuropathyrdquo Nature Genetics vol36 no 6 pp 602ndash606 2004
[3] J Zhai H Lin J-P Julien andWW Schlaepfer ldquoDisruption ofneurofilament network with aggregation of light neurofilamentprotein a common pathway leading to motor neuron degener-ation due to Charcot-Marie-Tooth disease-linked mutations inNFL andHSPB1rdquoHumanMolecular Genetics vol 16 no 24 pp3103ndash3116 2007
[4] L Almeida-Souza B Asselbergh C drsquoYdewalle et al ldquoSmallheat-shock protein HSPB1 mutants stabilize microtubules inCharcot-Marie-Tooth neuropathyrdquoThe Journal of Neurosciencevol 31 no 43 pp 15320ndash15328 2011
[5] A Holmgren D Bouhy V De Winter et al ldquoCharcot-Marie-Tooth causing HSPB1mutations increase Cdk5-mediated phos-phorylation of neurofilamentsrdquoActa Neuropathologica vol 126no 1 pp 93ndash108 2013
[6] A-P Arrigo ldquoThe cellular ldquonetworkingrdquo of mammalian Hsp27and its functions in the control of protein folding redox stateand apoptosisrdquoAdvances in eXperimental Medicine and Biologyvol 594 pp 14ndash26 2007
14 Stem Cells International
[7] C DrsquoYdewalle J Krishnan D M Chiheb et al ldquoHDAC6inhibitors reverse axonal loss in a mouse model of mutantHSPB1-induced Charcot-Marie-Tooth diseaserdquo NatureMedicine vol 17 no 8 pp 968ndash974 2011
[8] Y Zhang N Li C Caron et al ldquoHDAC-6 interacts with anddeacetylates tubulin and microtubules in vivordquo EMBO Journalvol 22 no 5 pp 1168ndash1179 2003
[9] A Valenzuela-Fernandez J R Cabrero J M Serrador and FSanchez-Madrid ldquoHDAC6 a key regulator of cytoskeleton cellmigration and cell-cell interactionsrdquo Trends in Cell Biology vol18 no 6 pp 291ndash297 2008
[10] V K Godena N Brookes-Hocking AMoller et al ldquoIncreasingmicrotubule acetylation rescues axonal transport and locomo-tor deficits caused by LRRK2 Roc-COR domain mutationsrdquoNature Communications vol 5 article 5245 2014
[11] J P Dompierre J D Godin B C Charrin et al ldquoHistonedeacetylase 6 inhibition compensates for the transport deficitin Huntingtonrsquos disease by increasing tubulin acetylationrdquo TheJournal of Neuroscience vol 27 no 13 pp 3571ndash3583 2007
[12] M W Amoroso G F Croft D J Williams et al ldquoAcceleratedhigh-yield generation of limb-innervating motor neurons fromhuman stem cellsrdquo Journal of Neuroscience vol 33 no 2 pp574ndash586 2013
[13] B J Gentil and L Cooper ldquoMolecular basis of axonal dysfunc-tion and traffic impairments in CMTrdquo Brain Research Bulletinvol 88 no 5 pp 444ndash453 2012
[14] Z-H Sheng and Q Cai ldquoMitochondrial transport in neu-rons impact on synaptic homeostasis and neurodegenerationrdquoNature Reviews Neuroscience vol 13 no 2 pp 77ndash93 2012
[15] J W Park B Vahidi A M Taylor S W Rhee and N LJeon ldquoMicrofluidic culture platform for neuroscience researchrdquoNature Protocols vol 1 no 4 pp 2128ndash2136 2006
[16] SWestermann and KWeber ldquoPost-translational modificationsregulate microtubule functionrdquo Nature Reviews Molecular CellBiology vol 4 no 12 pp 938ndash947 2003
[17] N A Reed D Cai T L Blasius et al ldquoMicrotubule acetylationpromotes kinesin-1 binding and transportrdquoCurrent Biology vol16 no 21 pp 2166ndash2172 2006
[18] C drsquoYdewalle E Bogaert and L Van Den Bosch ldquoHDAC6at the intersection of neuroprotection and neurodegenerationrdquoTraffic vol 13 no 6 pp 771ndash779 2012
[19] C Simoes-Pires V Zwick A Nurisso E Schenker P-ACarrupt and M Cuendet ldquoHDAC6 as a target for neurode-generative diseases what makes it different from the otherHDACsrdquoMolecular Neurodegeneration vol 8 article 7 2013
[20] Y Rui P Tiwari Z Xie and J Q Zheng ldquoAcute impairment ofmitochondrial trafficking by 120573-amyloid peptides in hippocam-pal neuronsrdquo Journal of Neuroscience vol 26 no 41 pp 10480ndash10487 2006
[21] K A Vossel K Zhang J Brodbeck et al ldquoTau reductionprevents A120573-induced defects in axonal transportrdquo Science vol330 no 6001 p 198 2010
[22] K J De vos A L Chapman M E Tennant et al ldquoFamilialamyotrophic lateral sclerosis-linked SOD1 mutants perturbfast axonal transport to reduce axonal mitochondria contentrdquoHumanMolecular Genetics vol 16 no 22 pp 2720ndash2728 2007
[23] E Trushina R B Dyer J D Badger II et al ldquoMutant huntingtinimpairs axonal trafficking inmammalian neurons in vivo and invitrordquo Molecular and Cellular Biology vol 24 no 18 pp 8195ndash8209 2004
[24] A L Orr S Li C-E Wang et al ldquoN-terminal mutant hunt-ingtin associates with mitochondria and impairs mitochondrialtraffickingrdquo Journal of Neuroscience vol 28 no 11 pp 2783ndash2792 2008
Submit your manuscripts athttpwwwhindawicom
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Anatomy Research International
PeptidesInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporation httpwwwhindawicom
International Journal of
Volume 2014
Zoology
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Molecular Biology International
GenomicsInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioinformaticsAdvances in
Marine BiologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Signal TransductionJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
Evolutionary BiologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Biochemistry Research International
ArchaeaHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Genetics Research International
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Advances in
Virolog y
Hindawi Publishing Corporationhttpwwwhindawicom
Nucleic AcidsJournal of
Volume 2014
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Enzyme Research
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
International Journal of
Microbiology

Stem Cells International 5
Platecells
Addvirus
Plate cells onMEF dishes
Emergingcolonies
Colony selection
20 FBSDMEM 20 KOSRDMEMF12 + bFGF
D-2 D0 D7 D8 D12 3-4 weeks
(a)
Normal HSPB1S135F HSPB1P182L
(b)
S135F-iPSCs P182L-iPSCsSNL WA09-hESCs hFSiPS1
(c)
P182L-iPSCsWA09-hESCs hFSiPS1
AG A T C A C C A T C C C A G T C A C C T T C GA G A T C AC C A T C T C A G T A C C T T C GA G A T C A C C A T C C C A G T C A C C T T C550560550560 540550
545C 545CgtT545C
WA09-hESCs S135F-iPSCshFSiPS1
T G G C T A C A T C T C C C G G T G C T T C A CT G G C T A C A T C T C C C G G T G C T T C A T G G C T A C A T C T T C C G G T G C T T C A410 420 410 420400410 420
404C 404C 404CgtT
(d)
S135F-iPSCs
1 2 3 4 5
6 7 8 9 10 11 12
13 14 15 16 17 18
19 20 21 22 X Y
P182L-iPSCs
1 2 3 4 5
6 7 8 9 10 11 12
13 14 15 16 17 18
19 20 21 22 X Y
(e)
Figure 1 Continued
6 Stem Cells International
Total-KLF4
Total-SOX2
GAPDH
Endo-KLF4Endo-OCT34
Endo-SOX2Endo-c-MYC
Total-OCT34
Total-c-MYC
WA
09-h
ESCs
S135
F-iP
SCs_
clone
1S1
35F-
iPSC
s_clo
ne 2
P182
L-iP
SCs_
clone
1P1
82L-
iPSC
s_clo
ne 2
hFSi
PS1
(f)NANOG DNA NANOGDNA SSEA4 DNA SSEA4DNA
WA
09-h
ESCs
S135
F-iP
SCs
P182
L-iP
SCs
hFSi
PS1
(g)
S135
F-iP
SCs
P182
L-iP
SCs
WA
09-h
ESCs
AFPDNA SMADNA NESTINDNA
hFSi
PS1
(h)
Figure 1 Continued
Stem Cells International 7
Mesoderm EctodermEndoderm
S135
F-iP
SCs
P182
L-iP
SCs
WA
09-h
ESCs
hFSi
PS1
Gl Ca NE
Gl CaM N
Gl Ca NE
NCa
Gl
(i)
Figure 1 Generation of CMT2F patient and dHMN2B patient-derived iPSCs (a) Experimental timeline for iPSC generation KOSRKnockOut serum replacement bFGF basic fibroblast growth factor (b) Morphology of fibroblasts from normal individual and patients(original magnification 50x) Scale bars 200 120583m (c) iPSC colonies showed ESC-like morphology such as a flat cobblestone-like appearancewith individual cells having a high nucleus-to-cytoplasm ratio (original magnification 50x) Scale bars 200120583m (d) CMT2F-iPSCs anddHMN2B-iPSCs had preserved point mutation sites in the HSPB1 gene verified by sequencing of RT-PCR products (e) CMT2F-iPSCs anddHMN2B-iPSCs maintained normal karyotype (f) Expression of total and endogenous Klf4 Oct34 Sox2 and c-Myc in CMT2F-iPSCs anddHMN2B-iPSCs was verified by RT-PCR Two clones from each of the patients-derived iPSCs were tested (clone 1 and clone 2) (g) ESCsand iPSCs expressed stem cell markers such as NANOG (in the nucleus original magnification 200x) and SSEA4 (in the cytoplasm originalmagnification 100x) Scale bars 200 120583m (h) EB-mediated in vitro spontaneous differentiation of ESCs and iPSCs resulted in the expressionof three-germ-layer markers such as AFP (endoderm) SMA (mesoderm) and nestin (ectoderm original magnification 200x) Scale bars100120583m (i) ESCs and iPSCs showed in vivo pluripotency by forming teratomas 8 weeks after subcutaneous injection into NODSCID miceTeratomas consisted of various three-germ-layer tissues including columnar gland epithelial cells with secretions [Gl] for endodermal tissuemuscle [M] and cartilage with calcification [Ca] for mesodermal tissue and neuroectodermal tissue [NE] and nerve axonal bundle [N] forectodermal tissue (original magnification 400x) Scale bars 100120583m
motor proteins [16] Among these modifications acety-lation of 120572-tubulin at the protein site of K40 regulatesbinding of dyneindynactin complexes and kinesin-1 tomicrotubules [11 17] In addition treatment with HDAC6inhibitors which increase acetylation of 120572-tubulin reversestransport defects in cellular models of Huntingtonrsquos disease[11] and LRRK2 mutation-induced Parkinsonrsquos disease [9]Therefore acetylation levels of 120572-tubulin were examined asan indicator of mitochondrial transport defects in S135F-MNs and P182L-MNs We found that S135F-MNs andP182L-MNs showed a significant reduction in acetylated 120572-tubulin levels compared with WA09-MNs or hFSiPS1-MNs(Figure 4)
35 CMT2F- and dHMN2B-Related Disease Phenotypes WereReversed by HDAC6 Inhibition By the treatment withHDAC6 inhibitors including tubastatin A (5 120583M) and newlydeveloped CHEMICAL X4 (05 120583M and 5 120583M) and X9(05 120583M and 5 120583M) acetylation of 120572-tubulin in S135F-MNs
and P182L-MNs was increased (Supplement Figure S4 Fig-ures 5(a) and 5(b)) Moreover the absolute velocity ofmitochondrial movements and the proportion of movingmitochondria in axons were increased significantly by thetreatment of these HDAC6 inhibitors (Figures 5(c) and 5(d)and Supplement Tables S1 and S2)
4 Discussion
Since iPSCs serve as an unlimited source of cells that can giverise to neuronal lineages and help to overcome the obstacleof unavailability of affected neuronal tissues from patientsdue to neuronal postmitotic property in vitro modeling anddrug screening with iPSCs can advance basic research onneurodegenerative diseases Therefore one of the strongestpoints of this study is that CMT2F- and dHMN2B-specificin vitro cellular models can be applied for the screening ofputative therapeuticmolecules such asHDAC6 inhibitors onperipheral neuropathy
8 Stem Cells International
ldquoSelf-renewalrdquo
iPSCs EBs Neurospheres MNs
3-4 wksldquoNeuralizationrdquoBMP inhibitors
ldquoCaudalizationrdquoRA
ldquoVentralizationrdquoShh agonist
(a)
WA09-MNs hFSiPS1-MNs
S135F-MNs P182L-MNs
(b)
WA09-MNs S135F-MNs P182L-MNs
ISL1
2T
uJ1
DN
AH
B9T
uJ1
DN
ASM
I32
MA
P2D
NA
SMI3
2Ch
ATD
NA
Syna
psin
DN
A
hFSiPS1-MNs
(c)
Figure 2 Continued
Stem Cells International 9
SMI32DAPI
0
50
100
150Re
lativ
e exp
ress
ion
()
WA09-MNs hFSiPS1-MNs S135F-MNs P182L-MNs
Relat
ive e
xpre
ssio
n (
)
MAP2DAPI
0
50
100
150
WA09-MNs hFSiPS1-MNs S135F-MNs P182L-MNs(d)
Axonal length
0
500
1000
1500
2000
WA09-MNs S135F-MNs
(120583m
)
(e)
Figure 2 Generation of CMT2F-specific in vitro model by differentiation of patient-derived iPSCs into MNs (a) Schematic presentation ofMN differentiation (b) Differentiated MNs showed typical cellular morphology (original magnification 50x) Scale bars 200 120583m (c) S135F-MNs and P182L-MNs expressedMN-specific transcription factors such asHB9 and ISL12 neuronal cytoskeletalmarkers such as TuJ1 SMI32andMAP2 ChAT and the synaptic vesicularmarker synapsin (originalmagnification 400x and 600x) Scale bars 100 120583m (d) Differentiationefficiency of S135F-MNs and P182L-MNs was comparable to control MNs in terms of neuronal marker expression such as SMI32DAPI andMAP2DAPI (WA09-MNs119873 = 30 hFSiPS1-MNs119873 = 329 S135F-MNs119873 = 1730 and P182L-MNs119873 = 1090) (e) Axonal length of S135F-MNs was comparable to that of control MNs Axonal length was measured by culturing fully differentiated MNs in microchannel plates foran additional 2 weeks (WA09-MNs119873 = 70 and S135F-MNs119873 = 121)
HDAC6 is a type IIb HDAC that modulates deacetylationof cytoplasmic target proteins Because HDAC6 is involvedin cytoskeletal formation modulation of cellular oxida-tionreduction state and the ubiquitin-proteasome systeminhibition of HDAC6 improves pathological conditions inneurons such as impairments in axonal transport oxidativestress and protein aggresome formation in vitro [18 19]In terms of axonal transport HDAC6 inhibitors reverseCMT2F and dHMN2B-related axonal transport by increasingacetylated 120572-tubulin In this study newly developed HDAC6inhibitors having 35-fold lower half-maximal inhibitoryconcentration (IC
50) values than that of tubastatin A showed
better performance on improving mitochondrial axonaltransport at 10 times lower concentration than tubastatin A
CMT2F-iPSCs and dHMN2B-iPSCswere generated frompatient-derived dermal fibroblasts carrying 404CgtT (S135F)and 545CgtT (P182L) mutations of HSPB1 by forced expres-sion of KLF4 OCT34 SOX2 and c-MYC using Sendai virus
vectors After establishing several stem cell colonies twocolony clones from each patient were used for further analysisto avoid differences due to clonal variance and selectediPSCs were differentiated into motor neurons with definedcytokines To recapitulate axonal pathology in vitro patient-specific motor neurons were cultured in microchanneledplates which are designed to separate the microenvironmentsof neuronal compartments In this system specific cellularmodels of CMT2F and dHMN2B showed defects in mito-chondrial transport as the absolute velocity of moving mito-chondria and the percentage of moving mitochondria werelower than control MNs Phenotypic variance between iPSCclones was observed in P182L-MNs in terms ofmitochondrialvelocity (see Figure S3) Defects in the axonal traffic systemare linked to many neurodegenerative diseases Due to thedistinct anatomy of peripheral neurons which have axons upto one meter in length the interchange of genetic materialmetabolites proteins and organelles between soma and distal
10 Stem Cells International
Original magnification 50xBar = 200120583m
(a)
WA09
P182LhFSiPS1
(b)
Absolute velocity (120583msec)
00
01
02
03
WA09-MNs hFSiPS1-MNs S135F-MNs P182L-MNs
(120583m
sec
)
lowast
lowastlowastlowast
(c)
Moving mitochondria ()
0
20
40
60
80
()
WA09-MNs hFSiPS1-MNs S135F-MNs P182L-MNs
lowastlowastlowast
lowastlowastlowast
lowastlowastlowastlowast
(d)
Figure 3 Axonal transport of mitochondria in control MNs S135F-MNs and P182L-MNs (a) Microfluidic culturing system Microchannelplates were made from transparent polymer in a specialized mold to create multicompartments connected by 120583m-sized grooves (8334 120583mlong in length) that only allow growing axons to pass through thereby separating the microenvironments of neuronal components (b)Visualization of mitochondria by Mito-dsRED2 transfection (upper image) and kymograph images for mitochondria movement (middleand lower four images) Mito-dsRED2 vector was introduced into fully differentiated MNs cultured on microchannel plates for an additional1 weekMitochondrial images were taken at a ratio of 121 snaps2min and stacked into kymograph for analysis (originalmagnification 1000x)(c) S135F-MNs show decreased velocity of moving mitochondria compared to control MNs Unpaired t-testslowast119875 lt 005 lowastlowastlowast119875 lt 0001 andlowastlowastlowastlowast119875 lt 00001 Absolute velocity of moving mitochondria was calculated from measured angle and length in kymograph images using
ImageJ (WA09-MNs 119873 = 203 hFSiPS1-MNs 119873 = 512 S135F-MNs 119873 = 428 and P182L-MNs 119873 = 496) (d) S135F-MNs and P182L-MNs show reduced moving proportion of mitochondria in an axon (WA09-MNs 119873 = 9 hFSiPS1-MNs 119873 = 6 S135F-MNs 119873 = 20 andP182L-MNs119873 = 42)
subcellular sites is critical for maintaining neuronal home-ostasis and survival Of the various trafficking cargos thetransport of mitochondria is particularly important becausemitochondria support several cellular processes by supplyingATP buffering intracellular Ca2+ and modulating apoptosis[13] In addition to delivering mitochondria to appropriatesubcellular sites the trafficking of mitochondria is vital forthe recycling of their components through dynamic fusionand fission mechanism [14] Defects in axonal mitochondrialtransport are associated with Alzheimerrsquos disease [20 21]amyotrophic lateral sclerosis [22] and Huntingtonrsquos disease[23 24]
Mitochondria are transported bidirectionally by interac-tions with molecular motor proteins such as kinesin-1 foranterogrademovement and dynein for retrogrademovementand those attach via adaptor proteins such as the mitochon-drial Rho-Milton complex and dynactin respectively [14]The recruitment of molecularmotors tomicrotubule tracks isregulated by posttranslational modification of 120572-tubulin [16]especially acetylation [17] In this study the level of acetylated120572-tubulin was reduced in both S135F-MNs and P182L-MNsBy increasing the acetylation of 120572-tubulin via treatment withHDAC6 inhibitors mitochondrial transport was improvedin terms of both velocity and the percentage of moving
Stem Cells International 11
WA09-MNs hFSiPS1 S135F-MNs P182L-MNs120572
-tubu
linAc
etyl
120572-tu
bulin
Acet
yl120572
-tubu
lin
120572-tu
bulin
DN
A
(a)
00
02
04
06
08
10
WA09-MNs hFSiPS1-MNs S135F-MNs P182L-MNs
Acet
yl120572
-tubu
lin120572
-tubu
lin
lowastlowast
lowast
lowastlowast
lowastlowast
(b)
WA09-MNs hFSiPS1-MNs S135F-MNs P182L-MNs
120572-Tubulin
Acetyl 120572-tubulin
(c)
00
02
04
06
08
10
WA09-MNs hFSiPS1-MNs S135F-MNs P182L-MNs
Acet
yl120572
-tubu
lin120572
-tubu
linlowast
lowast
lowast
lowastlowast
(d)
Figure 4 Reduced acetylation of120572-tubulin in S135F-MNs andP182-MNs (a) Acetylation of120572-tubulinwas visualized by immunostainingwithanti-acetylated 120572-tubulin Abs (red) Total 120572-tubulin was also detected by staining with anti-120572-tubulin Abs (green) (original magnification400x) Scale bars 100120583m (b) The extent of acetylated 120572-tubulin was normalized with corresponding total 120572-tubulin using ImageJ software(WA09-MNs119873 = 3 S135F-MNs119873 = 4 and P182L-MNs119873 = 5) Unpaired 119905-tests lowast119875 lt 005 and lowastlowast119875 lt 001 (c and d) Western blot assayshowed decreased acetylated 120572-tubulin levels in S135F-MNs and P182-MNs (paired 119905-tests lowast119875 lt 005 and lowastlowast119875 lt 001)
mitochondria Hence defects in the axonal trafficking ofmitochondria are associated with decreased acetylation of120572-tubulin which may be the main causative factor foraxonopathy in CMT2F and dHMN2B
In summary iPSCs can be used for valuable resourcesof in vitro disease modeling as well as drug screening Weanticipate that this in vitro system would be a useful modelfor the clinical translational research and drug development
5 Conclusions
This study demonstrates in vitro drug screening mod-els of inherited neuropathy resulting from mutations in
the HSPB1 gene can be developed from patient-specificiPSCs Axonal pathologies were mimicked in microflu-idic culturing system as both S135F-MNs and P182L-MNsshowed a marked decline in the absolute velocity andthe percentage of moving mitochondria These axonal de-fects were associated with decreased acetylation of 120572-tu-bulin and were reversed by treatment of HDAC6 in-hibitors
Competing Interests
The authors declare that they have no conflict of interests
12 Stem Cells International
HDAC6 inhibitors
S135
F-M
Ns
P182
L-M
Ns
Acetyl 120572-tubulin
Acetyl 120572-tubulin
120572-Tubulin
120572-Tubulin
DM
SO
X4_0
5120583
M
X4_5
120583M
X9_0
5120583
M
X9_5
120583M
Tuba
statin
120583M
A_5
(a)
00
05
10
15
20
25
DM
SO
X4_0
5120583
M
X4_5
120583M
X9_0
5120583
M
X9_5
120583M
lowast
lowastlowast
lowastlowast
Acet
yl120572
-tubu
lin120572
-tubu
lin
Acetyl 120572-tubulin in S135F-MNs
00
05
10
15
20
25
DM
SO
X4_0
5120583
M
X4_5
120583M
X9_0
5120583
M
X9_5
120583M
lowast
lowastlowast
lowastlowastAc
etyl
120572-tu
bulin
120572-tu
bulin
Acetyl 120572-tubulin in P182L-MNs
Tuba
statin
120583M
A_5
Tuba
statin
120583M
A_5
(b)
00
01
02
03
04
DM
SO
X4_0
5120583
M
X4_5
120583M
X9_0
5120583
M
X9_5
120583M
lowastlowastlowastlowast
lowastlowastlowastlowast
Abso
lute
velo
city
(120583m
sec
)
00
02
04
06
DM
SO
X4_0
5120583
M
X4_5
120583M
X9_0
5120583
M
X9_5
120583M
lowast
lowastlowastlowastlowast
lowastlowastlowastlowast
lowastlowastlowastlowast
lowastlowast
Abso
lute
velo
city
(120583m
sec
)
S135F-MNs
Absolute velocity
P182L-MNs
Tuba
statin
120583M
A_5
Tuba
statin
120583M
A_5
(c)
Figure 5 Continued
Stem Cells International 13
0
20
40
60
80
Mov
ing
mito
chon
dria
()
DM
SO
X4_0
5120583
M
X4_5
120583M
X4_5
120583M
X9_0
5120583
M
X9_5
120583M
lowast
lowastlowast
lowastlowast
lowastlowastlowastlowast
0
20
40
60
80
Mov
ing
mito
chon
dria
()
DM
SO
X4_0
5120583
M
X9_0
5120583
M
X9_5
120583M
lowastlowastlowastlowast
lowastlowastlowastlowast
lowastlowastlowastlowast
lowastlowastlowastlowast
S135F-MNs
Moving mitochondria
P182L-MNs
Tuba
statin
120583M
A_5
Tuba
statin
120583M
A_5
lowastlowast
(d)
Figure 5 HDAC6 inhibitors reversed axonal defect of mitochondrial transport by increasing acetylation of 120572-tubulin (a and b) Westernblot assay showed that treatment of HDAC6 inhibitors increased acetylated 120572-tubulin levels in S135F-MNs and P182L-MNs (paired 119905-testslowast119875 lt 005 and lowastlowast119875 lt 001) (c) HDAC6 inhibitors increased the absolute velocity of mitochondrial movements in S135F-MNs and P182L-MNs (WA09-MNs119873 = 617 S135F-MNs119873 = 101 and P182L-MNs119873 = 394) (d) HDAC6 inhibitors increased the percentage of movingmitochondria in S135F-MNs and P182L-MNs (WA09-MNs 119873 = 15 S135F-MNs 119873 = 7 and P182L-MNs 119873 = 15) Unpaired 119905-testslowast119875 lt 005 lowastlowast119875 lt 001 and lowastlowastlowastlowast119875 lt 00001
Authorsrsquo Contributions
Ji-Yon Kim was responsible for the iPSC experiments collec-tion and interpretation of the data and preparation of thepaper So-Youn Woo was interpreting the data and editingthe manuscript Inhee Mook-Jung provided microchannelplates and designed the experiment Heesun Choi JisooKim and Hyunjung Choi carried out microfluidic cultureand kymograph analysis Nina Ha Jangbeen Kyung andSoo Kyung Koo designed the experiments and preparationof the manuscript Young-Bin Hong and Sung-Chul Jungwere responsible for conception design of experiments andediting the manuscript Byung-Ok Choi was responsible forthe collection of patient samples and interpretation of thedata
Acknowledgments
This study was supported by the Korean Health TechnologyRampD Project Ministry of Health amp Welfare (HI12C0135HI14C3484 and HI15C1560) and a National Research Foun-dation of Korea grant funded by the Ministry of Science ICTandFuture Planning (NRF-2014R1A2A2A01004240)Humanstem cell line hFSiPS1 was provided by National Stem CellBank of Korea (Korea National Institute of Health Grant no4861-307-210-13)
References
[1] Inherited Peripheral Neuropathies Mutation Database httpwwwmolgenuaacbecmtmutationsmutationsMutByGenecfm
[2] O V Evgrafov IMersiyanova J Irobi et al ldquoMutant small heat-shock protein 27 causes axonal Charcot-Marie-Tooth diseaseand distal hereditary motor neuropathyrdquo Nature Genetics vol36 no 6 pp 602ndash606 2004
[3] J Zhai H Lin J-P Julien andWW Schlaepfer ldquoDisruption ofneurofilament network with aggregation of light neurofilamentprotein a common pathway leading to motor neuron degener-ation due to Charcot-Marie-Tooth disease-linked mutations inNFL andHSPB1rdquoHumanMolecular Genetics vol 16 no 24 pp3103ndash3116 2007
[4] L Almeida-Souza B Asselbergh C drsquoYdewalle et al ldquoSmallheat-shock protein HSPB1 mutants stabilize microtubules inCharcot-Marie-Tooth neuropathyrdquoThe Journal of Neurosciencevol 31 no 43 pp 15320ndash15328 2011
[5] A Holmgren D Bouhy V De Winter et al ldquoCharcot-Marie-Tooth causing HSPB1mutations increase Cdk5-mediated phos-phorylation of neurofilamentsrdquoActa Neuropathologica vol 126no 1 pp 93ndash108 2013
[6] A-P Arrigo ldquoThe cellular ldquonetworkingrdquo of mammalian Hsp27and its functions in the control of protein folding redox stateand apoptosisrdquoAdvances in eXperimental Medicine and Biologyvol 594 pp 14ndash26 2007
14 Stem Cells International
[7] C DrsquoYdewalle J Krishnan D M Chiheb et al ldquoHDAC6inhibitors reverse axonal loss in a mouse model of mutantHSPB1-induced Charcot-Marie-Tooth diseaserdquo NatureMedicine vol 17 no 8 pp 968ndash974 2011
[8] Y Zhang N Li C Caron et al ldquoHDAC-6 interacts with anddeacetylates tubulin and microtubules in vivordquo EMBO Journalvol 22 no 5 pp 1168ndash1179 2003
[9] A Valenzuela-Fernandez J R Cabrero J M Serrador and FSanchez-Madrid ldquoHDAC6 a key regulator of cytoskeleton cellmigration and cell-cell interactionsrdquo Trends in Cell Biology vol18 no 6 pp 291ndash297 2008
[10] V K Godena N Brookes-Hocking AMoller et al ldquoIncreasingmicrotubule acetylation rescues axonal transport and locomo-tor deficits caused by LRRK2 Roc-COR domain mutationsrdquoNature Communications vol 5 article 5245 2014
[11] J P Dompierre J D Godin B C Charrin et al ldquoHistonedeacetylase 6 inhibition compensates for the transport deficitin Huntingtonrsquos disease by increasing tubulin acetylationrdquo TheJournal of Neuroscience vol 27 no 13 pp 3571ndash3583 2007
[12] M W Amoroso G F Croft D J Williams et al ldquoAcceleratedhigh-yield generation of limb-innervating motor neurons fromhuman stem cellsrdquo Journal of Neuroscience vol 33 no 2 pp574ndash586 2013
[13] B J Gentil and L Cooper ldquoMolecular basis of axonal dysfunc-tion and traffic impairments in CMTrdquo Brain Research Bulletinvol 88 no 5 pp 444ndash453 2012
[14] Z-H Sheng and Q Cai ldquoMitochondrial transport in neu-rons impact on synaptic homeostasis and neurodegenerationrdquoNature Reviews Neuroscience vol 13 no 2 pp 77ndash93 2012
[15] J W Park B Vahidi A M Taylor S W Rhee and N LJeon ldquoMicrofluidic culture platform for neuroscience researchrdquoNature Protocols vol 1 no 4 pp 2128ndash2136 2006
[16] SWestermann and KWeber ldquoPost-translational modificationsregulate microtubule functionrdquo Nature Reviews Molecular CellBiology vol 4 no 12 pp 938ndash947 2003
[17] N A Reed D Cai T L Blasius et al ldquoMicrotubule acetylationpromotes kinesin-1 binding and transportrdquoCurrent Biology vol16 no 21 pp 2166ndash2172 2006
[18] C drsquoYdewalle E Bogaert and L Van Den Bosch ldquoHDAC6at the intersection of neuroprotection and neurodegenerationrdquoTraffic vol 13 no 6 pp 771ndash779 2012
[19] C Simoes-Pires V Zwick A Nurisso E Schenker P-ACarrupt and M Cuendet ldquoHDAC6 as a target for neurode-generative diseases what makes it different from the otherHDACsrdquoMolecular Neurodegeneration vol 8 article 7 2013
[20] Y Rui P Tiwari Z Xie and J Q Zheng ldquoAcute impairment ofmitochondrial trafficking by 120573-amyloid peptides in hippocam-pal neuronsrdquo Journal of Neuroscience vol 26 no 41 pp 10480ndash10487 2006
[21] K A Vossel K Zhang J Brodbeck et al ldquoTau reductionprevents A120573-induced defects in axonal transportrdquo Science vol330 no 6001 p 198 2010
[22] K J De vos A L Chapman M E Tennant et al ldquoFamilialamyotrophic lateral sclerosis-linked SOD1 mutants perturbfast axonal transport to reduce axonal mitochondria contentrdquoHumanMolecular Genetics vol 16 no 22 pp 2720ndash2728 2007
[23] E Trushina R B Dyer J D Badger II et al ldquoMutant huntingtinimpairs axonal trafficking inmammalian neurons in vivo and invitrordquo Molecular and Cellular Biology vol 24 no 18 pp 8195ndash8209 2004
[24] A L Orr S Li C-E Wang et al ldquoN-terminal mutant hunt-ingtin associates with mitochondria and impairs mitochondrialtraffickingrdquo Journal of Neuroscience vol 28 no 11 pp 2783ndash2792 2008
Submit your manuscripts athttpwwwhindawicom
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Anatomy Research International
PeptidesInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporation httpwwwhindawicom
International Journal of
Volume 2014
Zoology
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Molecular Biology International
GenomicsInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioinformaticsAdvances in
Marine BiologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Signal TransductionJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
Evolutionary BiologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Biochemistry Research International
ArchaeaHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Genetics Research International
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Advances in
Virolog y
Hindawi Publishing Corporationhttpwwwhindawicom
Nucleic AcidsJournal of
Volume 2014
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Enzyme Research
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
International Journal of
Microbiology

6 Stem Cells International
Total-KLF4
Total-SOX2
GAPDH
Endo-KLF4Endo-OCT34
Endo-SOX2Endo-c-MYC
Total-OCT34
Total-c-MYC
WA
09-h
ESCs
S135
F-iP
SCs_
clone
1S1
35F-
iPSC
s_clo
ne 2
P182
L-iP
SCs_
clone
1P1
82L-
iPSC
s_clo
ne 2
hFSi
PS1
(f)NANOG DNA NANOGDNA SSEA4 DNA SSEA4DNA
WA
09-h
ESCs
S135
F-iP
SCs
P182
L-iP
SCs
hFSi
PS1
(g)
S135
F-iP
SCs
P182
L-iP
SCs
WA
09-h
ESCs
AFPDNA SMADNA NESTINDNA
hFSi
PS1
(h)
Figure 1 Continued
Stem Cells International 7
Mesoderm EctodermEndoderm
S135
F-iP
SCs
P182
L-iP
SCs
WA
09-h
ESCs
hFSi
PS1
Gl Ca NE
Gl CaM N
Gl Ca NE
NCa
Gl
(i)
Figure 1 Generation of CMT2F patient and dHMN2B patient-derived iPSCs (a) Experimental timeline for iPSC generation KOSRKnockOut serum replacement bFGF basic fibroblast growth factor (b) Morphology of fibroblasts from normal individual and patients(original magnification 50x) Scale bars 200 120583m (c) iPSC colonies showed ESC-like morphology such as a flat cobblestone-like appearancewith individual cells having a high nucleus-to-cytoplasm ratio (original magnification 50x) Scale bars 200120583m (d) CMT2F-iPSCs anddHMN2B-iPSCs had preserved point mutation sites in the HSPB1 gene verified by sequencing of RT-PCR products (e) CMT2F-iPSCs anddHMN2B-iPSCs maintained normal karyotype (f) Expression of total and endogenous Klf4 Oct34 Sox2 and c-Myc in CMT2F-iPSCs anddHMN2B-iPSCs was verified by RT-PCR Two clones from each of the patients-derived iPSCs were tested (clone 1 and clone 2) (g) ESCsand iPSCs expressed stem cell markers such as NANOG (in the nucleus original magnification 200x) and SSEA4 (in the cytoplasm originalmagnification 100x) Scale bars 200 120583m (h) EB-mediated in vitro spontaneous differentiation of ESCs and iPSCs resulted in the expressionof three-germ-layer markers such as AFP (endoderm) SMA (mesoderm) and nestin (ectoderm original magnification 200x) Scale bars100120583m (i) ESCs and iPSCs showed in vivo pluripotency by forming teratomas 8 weeks after subcutaneous injection into NODSCID miceTeratomas consisted of various three-germ-layer tissues including columnar gland epithelial cells with secretions [Gl] for endodermal tissuemuscle [M] and cartilage with calcification [Ca] for mesodermal tissue and neuroectodermal tissue [NE] and nerve axonal bundle [N] forectodermal tissue (original magnification 400x) Scale bars 100120583m
motor proteins [16] Among these modifications acety-lation of 120572-tubulin at the protein site of K40 regulatesbinding of dyneindynactin complexes and kinesin-1 tomicrotubules [11 17] In addition treatment with HDAC6inhibitors which increase acetylation of 120572-tubulin reversestransport defects in cellular models of Huntingtonrsquos disease[11] and LRRK2 mutation-induced Parkinsonrsquos disease [9]Therefore acetylation levels of 120572-tubulin were examined asan indicator of mitochondrial transport defects in S135F-MNs and P182L-MNs We found that S135F-MNs andP182L-MNs showed a significant reduction in acetylated 120572-tubulin levels compared with WA09-MNs or hFSiPS1-MNs(Figure 4)
35 CMT2F- and dHMN2B-Related Disease Phenotypes WereReversed by HDAC6 Inhibition By the treatment withHDAC6 inhibitors including tubastatin A (5 120583M) and newlydeveloped CHEMICAL X4 (05 120583M and 5 120583M) and X9(05 120583M and 5 120583M) acetylation of 120572-tubulin in S135F-MNs
and P182L-MNs was increased (Supplement Figure S4 Fig-ures 5(a) and 5(b)) Moreover the absolute velocity ofmitochondrial movements and the proportion of movingmitochondria in axons were increased significantly by thetreatment of these HDAC6 inhibitors (Figures 5(c) and 5(d)and Supplement Tables S1 and S2)
4 Discussion
Since iPSCs serve as an unlimited source of cells that can giverise to neuronal lineages and help to overcome the obstacleof unavailability of affected neuronal tissues from patientsdue to neuronal postmitotic property in vitro modeling anddrug screening with iPSCs can advance basic research onneurodegenerative diseases Therefore one of the strongestpoints of this study is that CMT2F- and dHMN2B-specificin vitro cellular models can be applied for the screening ofputative therapeuticmolecules such asHDAC6 inhibitors onperipheral neuropathy
8 Stem Cells International
ldquoSelf-renewalrdquo
iPSCs EBs Neurospheres MNs
3-4 wksldquoNeuralizationrdquoBMP inhibitors
ldquoCaudalizationrdquoRA
ldquoVentralizationrdquoShh agonist
(a)
WA09-MNs hFSiPS1-MNs
S135F-MNs P182L-MNs
(b)
WA09-MNs S135F-MNs P182L-MNs
ISL1
2T
uJ1
DN
AH
B9T
uJ1
DN
ASM
I32
MA
P2D
NA
SMI3
2Ch
ATD
NA
Syna
psin
DN
A
hFSiPS1-MNs
(c)
Figure 2 Continued
Stem Cells International 9
SMI32DAPI
0
50
100
150Re
lativ
e exp
ress
ion
()
WA09-MNs hFSiPS1-MNs S135F-MNs P182L-MNs
Relat
ive e
xpre
ssio
n (
)
MAP2DAPI
0
50
100
150
WA09-MNs hFSiPS1-MNs S135F-MNs P182L-MNs(d)
Axonal length
0
500
1000
1500
2000
WA09-MNs S135F-MNs
(120583m
)
(e)
Figure 2 Generation of CMT2F-specific in vitro model by differentiation of patient-derived iPSCs into MNs (a) Schematic presentation ofMN differentiation (b) Differentiated MNs showed typical cellular morphology (original magnification 50x) Scale bars 200 120583m (c) S135F-MNs and P182L-MNs expressedMN-specific transcription factors such asHB9 and ISL12 neuronal cytoskeletalmarkers such as TuJ1 SMI32andMAP2 ChAT and the synaptic vesicularmarker synapsin (originalmagnification 400x and 600x) Scale bars 100 120583m (d) Differentiationefficiency of S135F-MNs and P182L-MNs was comparable to control MNs in terms of neuronal marker expression such as SMI32DAPI andMAP2DAPI (WA09-MNs119873 = 30 hFSiPS1-MNs119873 = 329 S135F-MNs119873 = 1730 and P182L-MNs119873 = 1090) (e) Axonal length of S135F-MNs was comparable to that of control MNs Axonal length was measured by culturing fully differentiated MNs in microchannel plates foran additional 2 weeks (WA09-MNs119873 = 70 and S135F-MNs119873 = 121)
HDAC6 is a type IIb HDAC that modulates deacetylationof cytoplasmic target proteins Because HDAC6 is involvedin cytoskeletal formation modulation of cellular oxida-tionreduction state and the ubiquitin-proteasome systeminhibition of HDAC6 improves pathological conditions inneurons such as impairments in axonal transport oxidativestress and protein aggresome formation in vitro [18 19]In terms of axonal transport HDAC6 inhibitors reverseCMT2F and dHMN2B-related axonal transport by increasingacetylated 120572-tubulin In this study newly developed HDAC6inhibitors having 35-fold lower half-maximal inhibitoryconcentration (IC
50) values than that of tubastatin A showed
better performance on improving mitochondrial axonaltransport at 10 times lower concentration than tubastatin A
CMT2F-iPSCs and dHMN2B-iPSCswere generated frompatient-derived dermal fibroblasts carrying 404CgtT (S135F)and 545CgtT (P182L) mutations of HSPB1 by forced expres-sion of KLF4 OCT34 SOX2 and c-MYC using Sendai virus
vectors After establishing several stem cell colonies twocolony clones from each patient were used for further analysisto avoid differences due to clonal variance and selectediPSCs were differentiated into motor neurons with definedcytokines To recapitulate axonal pathology in vitro patient-specific motor neurons were cultured in microchanneledplates which are designed to separate the microenvironmentsof neuronal compartments In this system specific cellularmodels of CMT2F and dHMN2B showed defects in mito-chondrial transport as the absolute velocity of moving mito-chondria and the percentage of moving mitochondria werelower than control MNs Phenotypic variance between iPSCclones was observed in P182L-MNs in terms ofmitochondrialvelocity (see Figure S3) Defects in the axonal traffic systemare linked to many neurodegenerative diseases Due to thedistinct anatomy of peripheral neurons which have axons upto one meter in length the interchange of genetic materialmetabolites proteins and organelles between soma and distal
10 Stem Cells International
Original magnification 50xBar = 200120583m
(a)
WA09
P182LhFSiPS1
(b)
Absolute velocity (120583msec)
00
01
02
03
WA09-MNs hFSiPS1-MNs S135F-MNs P182L-MNs
(120583m
sec
)
lowast
lowastlowastlowast
(c)
Moving mitochondria ()
0
20
40
60
80
()
WA09-MNs hFSiPS1-MNs S135F-MNs P182L-MNs
lowastlowastlowast
lowastlowastlowast
lowastlowastlowastlowast
(d)
Figure 3 Axonal transport of mitochondria in control MNs S135F-MNs and P182L-MNs (a) Microfluidic culturing system Microchannelplates were made from transparent polymer in a specialized mold to create multicompartments connected by 120583m-sized grooves (8334 120583mlong in length) that only allow growing axons to pass through thereby separating the microenvironments of neuronal components (b)Visualization of mitochondria by Mito-dsRED2 transfection (upper image) and kymograph images for mitochondria movement (middleand lower four images) Mito-dsRED2 vector was introduced into fully differentiated MNs cultured on microchannel plates for an additional1 weekMitochondrial images were taken at a ratio of 121 snaps2min and stacked into kymograph for analysis (originalmagnification 1000x)(c) S135F-MNs show decreased velocity of moving mitochondria compared to control MNs Unpaired t-testslowast119875 lt 005 lowastlowastlowast119875 lt 0001 andlowastlowastlowastlowast119875 lt 00001 Absolute velocity of moving mitochondria was calculated from measured angle and length in kymograph images using
ImageJ (WA09-MNs 119873 = 203 hFSiPS1-MNs 119873 = 512 S135F-MNs 119873 = 428 and P182L-MNs 119873 = 496) (d) S135F-MNs and P182L-MNs show reduced moving proportion of mitochondria in an axon (WA09-MNs 119873 = 9 hFSiPS1-MNs 119873 = 6 S135F-MNs 119873 = 20 andP182L-MNs119873 = 42)
subcellular sites is critical for maintaining neuronal home-ostasis and survival Of the various trafficking cargos thetransport of mitochondria is particularly important becausemitochondria support several cellular processes by supplyingATP buffering intracellular Ca2+ and modulating apoptosis[13] In addition to delivering mitochondria to appropriatesubcellular sites the trafficking of mitochondria is vital forthe recycling of their components through dynamic fusionand fission mechanism [14] Defects in axonal mitochondrialtransport are associated with Alzheimerrsquos disease [20 21]amyotrophic lateral sclerosis [22] and Huntingtonrsquos disease[23 24]
Mitochondria are transported bidirectionally by interac-tions with molecular motor proteins such as kinesin-1 foranterogrademovement and dynein for retrogrademovementand those attach via adaptor proteins such as the mitochon-drial Rho-Milton complex and dynactin respectively [14]The recruitment of molecularmotors tomicrotubule tracks isregulated by posttranslational modification of 120572-tubulin [16]especially acetylation [17] In this study the level of acetylated120572-tubulin was reduced in both S135F-MNs and P182L-MNsBy increasing the acetylation of 120572-tubulin via treatment withHDAC6 inhibitors mitochondrial transport was improvedin terms of both velocity and the percentage of moving
Stem Cells International 11
WA09-MNs hFSiPS1 S135F-MNs P182L-MNs120572
-tubu
linAc
etyl
120572-tu
bulin
Acet
yl120572
-tubu
lin
120572-tu
bulin
DN
A
(a)
00
02
04
06
08
10
WA09-MNs hFSiPS1-MNs S135F-MNs P182L-MNs
Acet
yl120572
-tubu
lin120572
-tubu
lin
lowastlowast
lowast
lowastlowast
lowastlowast
(b)
WA09-MNs hFSiPS1-MNs S135F-MNs P182L-MNs
120572-Tubulin
Acetyl 120572-tubulin
(c)
00
02
04
06
08
10
WA09-MNs hFSiPS1-MNs S135F-MNs P182L-MNs
Acet
yl120572
-tubu
lin120572
-tubu
linlowast
lowast
lowast
lowastlowast
(d)
Figure 4 Reduced acetylation of120572-tubulin in S135F-MNs andP182-MNs (a) Acetylation of120572-tubulinwas visualized by immunostainingwithanti-acetylated 120572-tubulin Abs (red) Total 120572-tubulin was also detected by staining with anti-120572-tubulin Abs (green) (original magnification400x) Scale bars 100120583m (b) The extent of acetylated 120572-tubulin was normalized with corresponding total 120572-tubulin using ImageJ software(WA09-MNs119873 = 3 S135F-MNs119873 = 4 and P182L-MNs119873 = 5) Unpaired 119905-tests lowast119875 lt 005 and lowastlowast119875 lt 001 (c and d) Western blot assayshowed decreased acetylated 120572-tubulin levels in S135F-MNs and P182-MNs (paired 119905-tests lowast119875 lt 005 and lowastlowast119875 lt 001)
mitochondria Hence defects in the axonal trafficking ofmitochondria are associated with decreased acetylation of120572-tubulin which may be the main causative factor foraxonopathy in CMT2F and dHMN2B
In summary iPSCs can be used for valuable resourcesof in vitro disease modeling as well as drug screening Weanticipate that this in vitro system would be a useful modelfor the clinical translational research and drug development
5 Conclusions
This study demonstrates in vitro drug screening mod-els of inherited neuropathy resulting from mutations in
the HSPB1 gene can be developed from patient-specificiPSCs Axonal pathologies were mimicked in microflu-idic culturing system as both S135F-MNs and P182L-MNsshowed a marked decline in the absolute velocity andthe percentage of moving mitochondria These axonal de-fects were associated with decreased acetylation of 120572-tu-bulin and were reversed by treatment of HDAC6 in-hibitors
Competing Interests
The authors declare that they have no conflict of interests
12 Stem Cells International
HDAC6 inhibitors
S135
F-M
Ns
P182
L-M
Ns
Acetyl 120572-tubulin
Acetyl 120572-tubulin
120572-Tubulin
120572-Tubulin
DM
SO
X4_0
5120583
M
X4_5
120583M
X9_0
5120583
M
X9_5
120583M
Tuba
statin
120583M
A_5
(a)
00
05
10
15
20
25
DM
SO
X4_0
5120583
M
X4_5
120583M
X9_0
5120583
M
X9_5
120583M
lowast
lowastlowast
lowastlowast
Acet
yl120572
-tubu
lin120572
-tubu
lin
Acetyl 120572-tubulin in S135F-MNs
00
05
10
15
20
25
DM
SO
X4_0
5120583
M
X4_5
120583M
X9_0
5120583
M
X9_5
120583M
lowast
lowastlowast
lowastlowastAc
etyl
120572-tu
bulin
120572-tu
bulin
Acetyl 120572-tubulin in P182L-MNs
Tuba
statin
120583M
A_5
Tuba
statin
120583M
A_5
(b)
00
01
02
03
04
DM
SO
X4_0
5120583
M
X4_5
120583M
X9_0
5120583
M
X9_5
120583M
lowastlowastlowastlowast
lowastlowastlowastlowast
Abso
lute
velo
city
(120583m
sec
)
00
02
04
06
DM
SO
X4_0
5120583
M
X4_5
120583M
X9_0
5120583
M
X9_5
120583M
lowast
lowastlowastlowastlowast
lowastlowastlowastlowast
lowastlowastlowastlowast
lowastlowast
Abso
lute
velo
city
(120583m
sec
)
S135F-MNs
Absolute velocity
P182L-MNs
Tuba
statin
120583M
A_5
Tuba
statin
120583M
A_5
(c)
Figure 5 Continued
Stem Cells International 13
0
20
40
60
80
Mov
ing
mito
chon
dria
()
DM
SO
X4_0
5120583
M
X4_5
120583M
X4_5
120583M
X9_0
5120583
M
X9_5
120583M
lowast
lowastlowast
lowastlowast
lowastlowastlowastlowast
0
20
40
60
80
Mov
ing
mito
chon
dria
()
DM
SO
X4_0
5120583
M
X9_0
5120583
M
X9_5
120583M
lowastlowastlowastlowast
lowastlowastlowastlowast
lowastlowastlowastlowast
lowastlowastlowastlowast
S135F-MNs
Moving mitochondria
P182L-MNs
Tuba
statin
120583M
A_5
Tuba
statin
120583M
A_5
lowastlowast
(d)
Figure 5 HDAC6 inhibitors reversed axonal defect of mitochondrial transport by increasing acetylation of 120572-tubulin (a and b) Westernblot assay showed that treatment of HDAC6 inhibitors increased acetylated 120572-tubulin levels in S135F-MNs and P182L-MNs (paired 119905-testslowast119875 lt 005 and lowastlowast119875 lt 001) (c) HDAC6 inhibitors increased the absolute velocity of mitochondrial movements in S135F-MNs and P182L-MNs (WA09-MNs119873 = 617 S135F-MNs119873 = 101 and P182L-MNs119873 = 394) (d) HDAC6 inhibitors increased the percentage of movingmitochondria in S135F-MNs and P182L-MNs (WA09-MNs 119873 = 15 S135F-MNs 119873 = 7 and P182L-MNs 119873 = 15) Unpaired 119905-testslowast119875 lt 005 lowastlowast119875 lt 001 and lowastlowastlowastlowast119875 lt 00001
Authorsrsquo Contributions
Ji-Yon Kim was responsible for the iPSC experiments collec-tion and interpretation of the data and preparation of thepaper So-Youn Woo was interpreting the data and editingthe manuscript Inhee Mook-Jung provided microchannelplates and designed the experiment Heesun Choi JisooKim and Hyunjung Choi carried out microfluidic cultureand kymograph analysis Nina Ha Jangbeen Kyung andSoo Kyung Koo designed the experiments and preparationof the manuscript Young-Bin Hong and Sung-Chul Jungwere responsible for conception design of experiments andediting the manuscript Byung-Ok Choi was responsible forthe collection of patient samples and interpretation of thedata
Acknowledgments
This study was supported by the Korean Health TechnologyRampD Project Ministry of Health amp Welfare (HI12C0135HI14C3484 and HI15C1560) and a National Research Foun-dation of Korea grant funded by the Ministry of Science ICTandFuture Planning (NRF-2014R1A2A2A01004240)Humanstem cell line hFSiPS1 was provided by National Stem CellBank of Korea (Korea National Institute of Health Grant no4861-307-210-13)
References
[1] Inherited Peripheral Neuropathies Mutation Database httpwwwmolgenuaacbecmtmutationsmutationsMutByGenecfm
[2] O V Evgrafov IMersiyanova J Irobi et al ldquoMutant small heat-shock protein 27 causes axonal Charcot-Marie-Tooth diseaseand distal hereditary motor neuropathyrdquo Nature Genetics vol36 no 6 pp 602ndash606 2004
[3] J Zhai H Lin J-P Julien andWW Schlaepfer ldquoDisruption ofneurofilament network with aggregation of light neurofilamentprotein a common pathway leading to motor neuron degener-ation due to Charcot-Marie-Tooth disease-linked mutations inNFL andHSPB1rdquoHumanMolecular Genetics vol 16 no 24 pp3103ndash3116 2007
[4] L Almeida-Souza B Asselbergh C drsquoYdewalle et al ldquoSmallheat-shock protein HSPB1 mutants stabilize microtubules inCharcot-Marie-Tooth neuropathyrdquoThe Journal of Neurosciencevol 31 no 43 pp 15320ndash15328 2011
[5] A Holmgren D Bouhy V De Winter et al ldquoCharcot-Marie-Tooth causing HSPB1mutations increase Cdk5-mediated phos-phorylation of neurofilamentsrdquoActa Neuropathologica vol 126no 1 pp 93ndash108 2013
[6] A-P Arrigo ldquoThe cellular ldquonetworkingrdquo of mammalian Hsp27and its functions in the control of protein folding redox stateand apoptosisrdquoAdvances in eXperimental Medicine and Biologyvol 594 pp 14ndash26 2007
14 Stem Cells International
[7] C DrsquoYdewalle J Krishnan D M Chiheb et al ldquoHDAC6inhibitors reverse axonal loss in a mouse model of mutantHSPB1-induced Charcot-Marie-Tooth diseaserdquo NatureMedicine vol 17 no 8 pp 968ndash974 2011
[8] Y Zhang N Li C Caron et al ldquoHDAC-6 interacts with anddeacetylates tubulin and microtubules in vivordquo EMBO Journalvol 22 no 5 pp 1168ndash1179 2003
[9] A Valenzuela-Fernandez J R Cabrero J M Serrador and FSanchez-Madrid ldquoHDAC6 a key regulator of cytoskeleton cellmigration and cell-cell interactionsrdquo Trends in Cell Biology vol18 no 6 pp 291ndash297 2008
[10] V K Godena N Brookes-Hocking AMoller et al ldquoIncreasingmicrotubule acetylation rescues axonal transport and locomo-tor deficits caused by LRRK2 Roc-COR domain mutationsrdquoNature Communications vol 5 article 5245 2014
[11] J P Dompierre J D Godin B C Charrin et al ldquoHistonedeacetylase 6 inhibition compensates for the transport deficitin Huntingtonrsquos disease by increasing tubulin acetylationrdquo TheJournal of Neuroscience vol 27 no 13 pp 3571ndash3583 2007
[12] M W Amoroso G F Croft D J Williams et al ldquoAcceleratedhigh-yield generation of limb-innervating motor neurons fromhuman stem cellsrdquo Journal of Neuroscience vol 33 no 2 pp574ndash586 2013
[13] B J Gentil and L Cooper ldquoMolecular basis of axonal dysfunc-tion and traffic impairments in CMTrdquo Brain Research Bulletinvol 88 no 5 pp 444ndash453 2012
[14] Z-H Sheng and Q Cai ldquoMitochondrial transport in neu-rons impact on synaptic homeostasis and neurodegenerationrdquoNature Reviews Neuroscience vol 13 no 2 pp 77ndash93 2012
[15] J W Park B Vahidi A M Taylor S W Rhee and N LJeon ldquoMicrofluidic culture platform for neuroscience researchrdquoNature Protocols vol 1 no 4 pp 2128ndash2136 2006
[16] SWestermann and KWeber ldquoPost-translational modificationsregulate microtubule functionrdquo Nature Reviews Molecular CellBiology vol 4 no 12 pp 938ndash947 2003
[17] N A Reed D Cai T L Blasius et al ldquoMicrotubule acetylationpromotes kinesin-1 binding and transportrdquoCurrent Biology vol16 no 21 pp 2166ndash2172 2006
[18] C drsquoYdewalle E Bogaert and L Van Den Bosch ldquoHDAC6at the intersection of neuroprotection and neurodegenerationrdquoTraffic vol 13 no 6 pp 771ndash779 2012
[19] C Simoes-Pires V Zwick A Nurisso E Schenker P-ACarrupt and M Cuendet ldquoHDAC6 as a target for neurode-generative diseases what makes it different from the otherHDACsrdquoMolecular Neurodegeneration vol 8 article 7 2013
[20] Y Rui P Tiwari Z Xie and J Q Zheng ldquoAcute impairment ofmitochondrial trafficking by 120573-amyloid peptides in hippocam-pal neuronsrdquo Journal of Neuroscience vol 26 no 41 pp 10480ndash10487 2006
[21] K A Vossel K Zhang J Brodbeck et al ldquoTau reductionprevents A120573-induced defects in axonal transportrdquo Science vol330 no 6001 p 198 2010
[22] K J De vos A L Chapman M E Tennant et al ldquoFamilialamyotrophic lateral sclerosis-linked SOD1 mutants perturbfast axonal transport to reduce axonal mitochondria contentrdquoHumanMolecular Genetics vol 16 no 22 pp 2720ndash2728 2007
[23] E Trushina R B Dyer J D Badger II et al ldquoMutant huntingtinimpairs axonal trafficking inmammalian neurons in vivo and invitrordquo Molecular and Cellular Biology vol 24 no 18 pp 8195ndash8209 2004
[24] A L Orr S Li C-E Wang et al ldquoN-terminal mutant hunt-ingtin associates with mitochondria and impairs mitochondrialtraffickingrdquo Journal of Neuroscience vol 28 no 11 pp 2783ndash2792 2008
Submit your manuscripts athttpwwwhindawicom
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Anatomy Research International
PeptidesInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporation httpwwwhindawicom
International Journal of
Volume 2014
Zoology
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Molecular Biology International
GenomicsInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioinformaticsAdvances in
Marine BiologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Signal TransductionJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
Evolutionary BiologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Biochemistry Research International
ArchaeaHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Genetics Research International
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Advances in
Virolog y
Hindawi Publishing Corporationhttpwwwhindawicom
Nucleic AcidsJournal of
Volume 2014
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Enzyme Research
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
International Journal of
Microbiology

Stem Cells International 7
Mesoderm EctodermEndoderm
S135
F-iP
SCs
P182
L-iP
SCs
WA
09-h
ESCs
hFSi
PS1
Gl Ca NE
Gl CaM N
Gl Ca NE
NCa
Gl
(i)
Figure 1 Generation of CMT2F patient and dHMN2B patient-derived iPSCs (a) Experimental timeline for iPSC generation KOSRKnockOut serum replacement bFGF basic fibroblast growth factor (b) Morphology of fibroblasts from normal individual and patients(original magnification 50x) Scale bars 200 120583m (c) iPSC colonies showed ESC-like morphology such as a flat cobblestone-like appearancewith individual cells having a high nucleus-to-cytoplasm ratio (original magnification 50x) Scale bars 200120583m (d) CMT2F-iPSCs anddHMN2B-iPSCs had preserved point mutation sites in the HSPB1 gene verified by sequencing of RT-PCR products (e) CMT2F-iPSCs anddHMN2B-iPSCs maintained normal karyotype (f) Expression of total and endogenous Klf4 Oct34 Sox2 and c-Myc in CMT2F-iPSCs anddHMN2B-iPSCs was verified by RT-PCR Two clones from each of the patients-derived iPSCs were tested (clone 1 and clone 2) (g) ESCsand iPSCs expressed stem cell markers such as NANOG (in the nucleus original magnification 200x) and SSEA4 (in the cytoplasm originalmagnification 100x) Scale bars 200 120583m (h) EB-mediated in vitro spontaneous differentiation of ESCs and iPSCs resulted in the expressionof three-germ-layer markers such as AFP (endoderm) SMA (mesoderm) and nestin (ectoderm original magnification 200x) Scale bars100120583m (i) ESCs and iPSCs showed in vivo pluripotency by forming teratomas 8 weeks after subcutaneous injection into NODSCID miceTeratomas consisted of various three-germ-layer tissues including columnar gland epithelial cells with secretions [Gl] for endodermal tissuemuscle [M] and cartilage with calcification [Ca] for mesodermal tissue and neuroectodermal tissue [NE] and nerve axonal bundle [N] forectodermal tissue (original magnification 400x) Scale bars 100120583m
motor proteins [16] Among these modifications acety-lation of 120572-tubulin at the protein site of K40 regulatesbinding of dyneindynactin complexes and kinesin-1 tomicrotubules [11 17] In addition treatment with HDAC6inhibitors which increase acetylation of 120572-tubulin reversestransport defects in cellular models of Huntingtonrsquos disease[11] and LRRK2 mutation-induced Parkinsonrsquos disease [9]Therefore acetylation levels of 120572-tubulin were examined asan indicator of mitochondrial transport defects in S135F-MNs and P182L-MNs We found that S135F-MNs andP182L-MNs showed a significant reduction in acetylated 120572-tubulin levels compared with WA09-MNs or hFSiPS1-MNs(Figure 4)
35 CMT2F- and dHMN2B-Related Disease Phenotypes WereReversed by HDAC6 Inhibition By the treatment withHDAC6 inhibitors including tubastatin A (5 120583M) and newlydeveloped CHEMICAL X4 (05 120583M and 5 120583M) and X9(05 120583M and 5 120583M) acetylation of 120572-tubulin in S135F-MNs
and P182L-MNs was increased (Supplement Figure S4 Fig-ures 5(a) and 5(b)) Moreover the absolute velocity ofmitochondrial movements and the proportion of movingmitochondria in axons were increased significantly by thetreatment of these HDAC6 inhibitors (Figures 5(c) and 5(d)and Supplement Tables S1 and S2)
4 Discussion
Since iPSCs serve as an unlimited source of cells that can giverise to neuronal lineages and help to overcome the obstacleof unavailability of affected neuronal tissues from patientsdue to neuronal postmitotic property in vitro modeling anddrug screening with iPSCs can advance basic research onneurodegenerative diseases Therefore one of the strongestpoints of this study is that CMT2F- and dHMN2B-specificin vitro cellular models can be applied for the screening ofputative therapeuticmolecules such asHDAC6 inhibitors onperipheral neuropathy
8 Stem Cells International
ldquoSelf-renewalrdquo
iPSCs EBs Neurospheres MNs
3-4 wksldquoNeuralizationrdquoBMP inhibitors
ldquoCaudalizationrdquoRA
ldquoVentralizationrdquoShh agonist
(a)
WA09-MNs hFSiPS1-MNs
S135F-MNs P182L-MNs
(b)
WA09-MNs S135F-MNs P182L-MNs
ISL1
2T
uJ1
DN
AH
B9T
uJ1
DN
ASM
I32
MA
P2D
NA
SMI3
2Ch
ATD
NA
Syna
psin
DN
A
hFSiPS1-MNs
(c)
Figure 2 Continued
Stem Cells International 9
SMI32DAPI
0
50
100
150Re
lativ
e exp
ress
ion
()
WA09-MNs hFSiPS1-MNs S135F-MNs P182L-MNs
Relat
ive e
xpre
ssio
n (
)
MAP2DAPI
0
50
100
150
WA09-MNs hFSiPS1-MNs S135F-MNs P182L-MNs(d)
Axonal length
0
500
1000
1500
2000
WA09-MNs S135F-MNs
(120583m
)
(e)
Figure 2 Generation of CMT2F-specific in vitro model by differentiation of patient-derived iPSCs into MNs (a) Schematic presentation ofMN differentiation (b) Differentiated MNs showed typical cellular morphology (original magnification 50x) Scale bars 200 120583m (c) S135F-MNs and P182L-MNs expressedMN-specific transcription factors such asHB9 and ISL12 neuronal cytoskeletalmarkers such as TuJ1 SMI32andMAP2 ChAT and the synaptic vesicularmarker synapsin (originalmagnification 400x and 600x) Scale bars 100 120583m (d) Differentiationefficiency of S135F-MNs and P182L-MNs was comparable to control MNs in terms of neuronal marker expression such as SMI32DAPI andMAP2DAPI (WA09-MNs119873 = 30 hFSiPS1-MNs119873 = 329 S135F-MNs119873 = 1730 and P182L-MNs119873 = 1090) (e) Axonal length of S135F-MNs was comparable to that of control MNs Axonal length was measured by culturing fully differentiated MNs in microchannel plates foran additional 2 weeks (WA09-MNs119873 = 70 and S135F-MNs119873 = 121)
HDAC6 is a type IIb HDAC that modulates deacetylationof cytoplasmic target proteins Because HDAC6 is involvedin cytoskeletal formation modulation of cellular oxida-tionreduction state and the ubiquitin-proteasome systeminhibition of HDAC6 improves pathological conditions inneurons such as impairments in axonal transport oxidativestress and protein aggresome formation in vitro [18 19]In terms of axonal transport HDAC6 inhibitors reverseCMT2F and dHMN2B-related axonal transport by increasingacetylated 120572-tubulin In this study newly developed HDAC6inhibitors having 35-fold lower half-maximal inhibitoryconcentration (IC
50) values than that of tubastatin A showed
better performance on improving mitochondrial axonaltransport at 10 times lower concentration than tubastatin A
CMT2F-iPSCs and dHMN2B-iPSCswere generated frompatient-derived dermal fibroblasts carrying 404CgtT (S135F)and 545CgtT (P182L) mutations of HSPB1 by forced expres-sion of KLF4 OCT34 SOX2 and c-MYC using Sendai virus
vectors After establishing several stem cell colonies twocolony clones from each patient were used for further analysisto avoid differences due to clonal variance and selectediPSCs were differentiated into motor neurons with definedcytokines To recapitulate axonal pathology in vitro patient-specific motor neurons were cultured in microchanneledplates which are designed to separate the microenvironmentsof neuronal compartments In this system specific cellularmodels of CMT2F and dHMN2B showed defects in mito-chondrial transport as the absolute velocity of moving mito-chondria and the percentage of moving mitochondria werelower than control MNs Phenotypic variance between iPSCclones was observed in P182L-MNs in terms ofmitochondrialvelocity (see Figure S3) Defects in the axonal traffic systemare linked to many neurodegenerative diseases Due to thedistinct anatomy of peripheral neurons which have axons upto one meter in length the interchange of genetic materialmetabolites proteins and organelles between soma and distal
10 Stem Cells International
Original magnification 50xBar = 200120583m
(a)
WA09
P182LhFSiPS1
(b)
Absolute velocity (120583msec)
00
01
02
03
WA09-MNs hFSiPS1-MNs S135F-MNs P182L-MNs
(120583m
sec
)
lowast
lowastlowastlowast
(c)
Moving mitochondria ()
0
20
40
60
80
()
WA09-MNs hFSiPS1-MNs S135F-MNs P182L-MNs
lowastlowastlowast
lowastlowastlowast
lowastlowastlowastlowast
(d)
Figure 3 Axonal transport of mitochondria in control MNs S135F-MNs and P182L-MNs (a) Microfluidic culturing system Microchannelplates were made from transparent polymer in a specialized mold to create multicompartments connected by 120583m-sized grooves (8334 120583mlong in length) that only allow growing axons to pass through thereby separating the microenvironments of neuronal components (b)Visualization of mitochondria by Mito-dsRED2 transfection (upper image) and kymograph images for mitochondria movement (middleand lower four images) Mito-dsRED2 vector was introduced into fully differentiated MNs cultured on microchannel plates for an additional1 weekMitochondrial images were taken at a ratio of 121 snaps2min and stacked into kymograph for analysis (originalmagnification 1000x)(c) S135F-MNs show decreased velocity of moving mitochondria compared to control MNs Unpaired t-testslowast119875 lt 005 lowastlowastlowast119875 lt 0001 andlowastlowastlowastlowast119875 lt 00001 Absolute velocity of moving mitochondria was calculated from measured angle and length in kymograph images using
ImageJ (WA09-MNs 119873 = 203 hFSiPS1-MNs 119873 = 512 S135F-MNs 119873 = 428 and P182L-MNs 119873 = 496) (d) S135F-MNs and P182L-MNs show reduced moving proportion of mitochondria in an axon (WA09-MNs 119873 = 9 hFSiPS1-MNs 119873 = 6 S135F-MNs 119873 = 20 andP182L-MNs119873 = 42)
subcellular sites is critical for maintaining neuronal home-ostasis and survival Of the various trafficking cargos thetransport of mitochondria is particularly important becausemitochondria support several cellular processes by supplyingATP buffering intracellular Ca2+ and modulating apoptosis[13] In addition to delivering mitochondria to appropriatesubcellular sites the trafficking of mitochondria is vital forthe recycling of their components through dynamic fusionand fission mechanism [14] Defects in axonal mitochondrialtransport are associated with Alzheimerrsquos disease [20 21]amyotrophic lateral sclerosis [22] and Huntingtonrsquos disease[23 24]
Mitochondria are transported bidirectionally by interac-tions with molecular motor proteins such as kinesin-1 foranterogrademovement and dynein for retrogrademovementand those attach via adaptor proteins such as the mitochon-drial Rho-Milton complex and dynactin respectively [14]The recruitment of molecularmotors tomicrotubule tracks isregulated by posttranslational modification of 120572-tubulin [16]especially acetylation [17] In this study the level of acetylated120572-tubulin was reduced in both S135F-MNs and P182L-MNsBy increasing the acetylation of 120572-tubulin via treatment withHDAC6 inhibitors mitochondrial transport was improvedin terms of both velocity and the percentage of moving
Stem Cells International 11
WA09-MNs hFSiPS1 S135F-MNs P182L-MNs120572
-tubu
linAc
etyl
120572-tu
bulin
Acet
yl120572
-tubu
lin
120572-tu
bulin
DN
A
(a)
00
02
04
06
08
10
WA09-MNs hFSiPS1-MNs S135F-MNs P182L-MNs
Acet
yl120572
-tubu
lin120572
-tubu
lin
lowastlowast
lowast
lowastlowast
lowastlowast
(b)
WA09-MNs hFSiPS1-MNs S135F-MNs P182L-MNs
120572-Tubulin
Acetyl 120572-tubulin
(c)
00
02
04
06
08
10
WA09-MNs hFSiPS1-MNs S135F-MNs P182L-MNs
Acet
yl120572
-tubu
lin120572
-tubu
linlowast
lowast
lowast
lowastlowast
(d)
Figure 4 Reduced acetylation of120572-tubulin in S135F-MNs andP182-MNs (a) Acetylation of120572-tubulinwas visualized by immunostainingwithanti-acetylated 120572-tubulin Abs (red) Total 120572-tubulin was also detected by staining with anti-120572-tubulin Abs (green) (original magnification400x) Scale bars 100120583m (b) The extent of acetylated 120572-tubulin was normalized with corresponding total 120572-tubulin using ImageJ software(WA09-MNs119873 = 3 S135F-MNs119873 = 4 and P182L-MNs119873 = 5) Unpaired 119905-tests lowast119875 lt 005 and lowastlowast119875 lt 001 (c and d) Western blot assayshowed decreased acetylated 120572-tubulin levels in S135F-MNs and P182-MNs (paired 119905-tests lowast119875 lt 005 and lowastlowast119875 lt 001)
mitochondria Hence defects in the axonal trafficking ofmitochondria are associated with decreased acetylation of120572-tubulin which may be the main causative factor foraxonopathy in CMT2F and dHMN2B
In summary iPSCs can be used for valuable resourcesof in vitro disease modeling as well as drug screening Weanticipate that this in vitro system would be a useful modelfor the clinical translational research and drug development
5 Conclusions
This study demonstrates in vitro drug screening mod-els of inherited neuropathy resulting from mutations in
the HSPB1 gene can be developed from patient-specificiPSCs Axonal pathologies were mimicked in microflu-idic culturing system as both S135F-MNs and P182L-MNsshowed a marked decline in the absolute velocity andthe percentage of moving mitochondria These axonal de-fects were associated with decreased acetylation of 120572-tu-bulin and were reversed by treatment of HDAC6 in-hibitors
Competing Interests
The authors declare that they have no conflict of interests
12 Stem Cells International
HDAC6 inhibitors
S135
F-M
Ns
P182
L-M
Ns
Acetyl 120572-tubulin
Acetyl 120572-tubulin
120572-Tubulin
120572-Tubulin
DM
SO
X4_0
5120583
M
X4_5
120583M
X9_0
5120583
M
X9_5
120583M
Tuba
statin
120583M
A_5
(a)
00
05
10
15
20
25
DM
SO
X4_0
5120583
M
X4_5
120583M
X9_0
5120583
M
X9_5
120583M
lowast
lowastlowast
lowastlowast
Acet
yl120572
-tubu
lin120572
-tubu
lin
Acetyl 120572-tubulin in S135F-MNs
00
05
10
15
20
25
DM
SO
X4_0
5120583
M
X4_5
120583M
X9_0
5120583
M
X9_5
120583M
lowast
lowastlowast
lowastlowastAc
etyl
120572-tu
bulin
120572-tu
bulin
Acetyl 120572-tubulin in P182L-MNs
Tuba
statin
120583M
A_5
Tuba
statin
120583M
A_5
(b)
00
01
02
03
04
DM
SO
X4_0
5120583
M
X4_5
120583M
X9_0
5120583
M
X9_5
120583M
lowastlowastlowastlowast
lowastlowastlowastlowast
Abso
lute
velo
city
(120583m
sec
)
00
02
04
06
DM
SO
X4_0
5120583
M
X4_5
120583M
X9_0
5120583
M
X9_5
120583M
lowast
lowastlowastlowastlowast
lowastlowastlowastlowast
lowastlowastlowastlowast
lowastlowast
Abso
lute
velo
city
(120583m
sec
)
S135F-MNs
Absolute velocity
P182L-MNs
Tuba
statin
120583M
A_5
Tuba
statin
120583M
A_5
(c)
Figure 5 Continued
Stem Cells International 13
0
20
40
60
80
Mov
ing
mito
chon
dria
()
DM
SO
X4_0
5120583
M
X4_5
120583M
X4_5
120583M
X9_0
5120583
M
X9_5
120583M
lowast
lowastlowast
lowastlowast
lowastlowastlowastlowast
0
20
40
60
80
Mov
ing
mito
chon
dria
()
DM
SO
X4_0
5120583
M
X9_0
5120583
M
X9_5
120583M
lowastlowastlowastlowast
lowastlowastlowastlowast
lowastlowastlowastlowast
lowastlowastlowastlowast
S135F-MNs
Moving mitochondria
P182L-MNs
Tuba
statin
120583M
A_5
Tuba
statin
120583M
A_5
lowastlowast
(d)
Figure 5 HDAC6 inhibitors reversed axonal defect of mitochondrial transport by increasing acetylation of 120572-tubulin (a and b) Westernblot assay showed that treatment of HDAC6 inhibitors increased acetylated 120572-tubulin levels in S135F-MNs and P182L-MNs (paired 119905-testslowast119875 lt 005 and lowastlowast119875 lt 001) (c) HDAC6 inhibitors increased the absolute velocity of mitochondrial movements in S135F-MNs and P182L-MNs (WA09-MNs119873 = 617 S135F-MNs119873 = 101 and P182L-MNs119873 = 394) (d) HDAC6 inhibitors increased the percentage of movingmitochondria in S135F-MNs and P182L-MNs (WA09-MNs 119873 = 15 S135F-MNs 119873 = 7 and P182L-MNs 119873 = 15) Unpaired 119905-testslowast119875 lt 005 lowastlowast119875 lt 001 and lowastlowastlowastlowast119875 lt 00001
Authorsrsquo Contributions
Ji-Yon Kim was responsible for the iPSC experiments collec-tion and interpretation of the data and preparation of thepaper So-Youn Woo was interpreting the data and editingthe manuscript Inhee Mook-Jung provided microchannelplates and designed the experiment Heesun Choi JisooKim and Hyunjung Choi carried out microfluidic cultureand kymograph analysis Nina Ha Jangbeen Kyung andSoo Kyung Koo designed the experiments and preparationof the manuscript Young-Bin Hong and Sung-Chul Jungwere responsible for conception design of experiments andediting the manuscript Byung-Ok Choi was responsible forthe collection of patient samples and interpretation of thedata
Acknowledgments
This study was supported by the Korean Health TechnologyRampD Project Ministry of Health amp Welfare (HI12C0135HI14C3484 and HI15C1560) and a National Research Foun-dation of Korea grant funded by the Ministry of Science ICTandFuture Planning (NRF-2014R1A2A2A01004240)Humanstem cell line hFSiPS1 was provided by National Stem CellBank of Korea (Korea National Institute of Health Grant no4861-307-210-13)
References
[1] Inherited Peripheral Neuropathies Mutation Database httpwwwmolgenuaacbecmtmutationsmutationsMutByGenecfm
[2] O V Evgrafov IMersiyanova J Irobi et al ldquoMutant small heat-shock protein 27 causes axonal Charcot-Marie-Tooth diseaseand distal hereditary motor neuropathyrdquo Nature Genetics vol36 no 6 pp 602ndash606 2004
[3] J Zhai H Lin J-P Julien andWW Schlaepfer ldquoDisruption ofneurofilament network with aggregation of light neurofilamentprotein a common pathway leading to motor neuron degener-ation due to Charcot-Marie-Tooth disease-linked mutations inNFL andHSPB1rdquoHumanMolecular Genetics vol 16 no 24 pp3103ndash3116 2007
[4] L Almeida-Souza B Asselbergh C drsquoYdewalle et al ldquoSmallheat-shock protein HSPB1 mutants stabilize microtubules inCharcot-Marie-Tooth neuropathyrdquoThe Journal of Neurosciencevol 31 no 43 pp 15320ndash15328 2011
[5] A Holmgren D Bouhy V De Winter et al ldquoCharcot-Marie-Tooth causing HSPB1mutations increase Cdk5-mediated phos-phorylation of neurofilamentsrdquoActa Neuropathologica vol 126no 1 pp 93ndash108 2013
[6] A-P Arrigo ldquoThe cellular ldquonetworkingrdquo of mammalian Hsp27and its functions in the control of protein folding redox stateand apoptosisrdquoAdvances in eXperimental Medicine and Biologyvol 594 pp 14ndash26 2007
14 Stem Cells International
[7] C DrsquoYdewalle J Krishnan D M Chiheb et al ldquoHDAC6inhibitors reverse axonal loss in a mouse model of mutantHSPB1-induced Charcot-Marie-Tooth diseaserdquo NatureMedicine vol 17 no 8 pp 968ndash974 2011
[8] Y Zhang N Li C Caron et al ldquoHDAC-6 interacts with anddeacetylates tubulin and microtubules in vivordquo EMBO Journalvol 22 no 5 pp 1168ndash1179 2003
[9] A Valenzuela-Fernandez J R Cabrero J M Serrador and FSanchez-Madrid ldquoHDAC6 a key regulator of cytoskeleton cellmigration and cell-cell interactionsrdquo Trends in Cell Biology vol18 no 6 pp 291ndash297 2008
[10] V K Godena N Brookes-Hocking AMoller et al ldquoIncreasingmicrotubule acetylation rescues axonal transport and locomo-tor deficits caused by LRRK2 Roc-COR domain mutationsrdquoNature Communications vol 5 article 5245 2014
[11] J P Dompierre J D Godin B C Charrin et al ldquoHistonedeacetylase 6 inhibition compensates for the transport deficitin Huntingtonrsquos disease by increasing tubulin acetylationrdquo TheJournal of Neuroscience vol 27 no 13 pp 3571ndash3583 2007
[12] M W Amoroso G F Croft D J Williams et al ldquoAcceleratedhigh-yield generation of limb-innervating motor neurons fromhuman stem cellsrdquo Journal of Neuroscience vol 33 no 2 pp574ndash586 2013
[13] B J Gentil and L Cooper ldquoMolecular basis of axonal dysfunc-tion and traffic impairments in CMTrdquo Brain Research Bulletinvol 88 no 5 pp 444ndash453 2012
[14] Z-H Sheng and Q Cai ldquoMitochondrial transport in neu-rons impact on synaptic homeostasis and neurodegenerationrdquoNature Reviews Neuroscience vol 13 no 2 pp 77ndash93 2012
[15] J W Park B Vahidi A M Taylor S W Rhee and N LJeon ldquoMicrofluidic culture platform for neuroscience researchrdquoNature Protocols vol 1 no 4 pp 2128ndash2136 2006
[16] SWestermann and KWeber ldquoPost-translational modificationsregulate microtubule functionrdquo Nature Reviews Molecular CellBiology vol 4 no 12 pp 938ndash947 2003
[17] N A Reed D Cai T L Blasius et al ldquoMicrotubule acetylationpromotes kinesin-1 binding and transportrdquoCurrent Biology vol16 no 21 pp 2166ndash2172 2006
[18] C drsquoYdewalle E Bogaert and L Van Den Bosch ldquoHDAC6at the intersection of neuroprotection and neurodegenerationrdquoTraffic vol 13 no 6 pp 771ndash779 2012
[19] C Simoes-Pires V Zwick A Nurisso E Schenker P-ACarrupt and M Cuendet ldquoHDAC6 as a target for neurode-generative diseases what makes it different from the otherHDACsrdquoMolecular Neurodegeneration vol 8 article 7 2013
[20] Y Rui P Tiwari Z Xie and J Q Zheng ldquoAcute impairment ofmitochondrial trafficking by 120573-amyloid peptides in hippocam-pal neuronsrdquo Journal of Neuroscience vol 26 no 41 pp 10480ndash10487 2006
[21] K A Vossel K Zhang J Brodbeck et al ldquoTau reductionprevents A120573-induced defects in axonal transportrdquo Science vol330 no 6001 p 198 2010
[22] K J De vos A L Chapman M E Tennant et al ldquoFamilialamyotrophic lateral sclerosis-linked SOD1 mutants perturbfast axonal transport to reduce axonal mitochondria contentrdquoHumanMolecular Genetics vol 16 no 22 pp 2720ndash2728 2007
[23] E Trushina R B Dyer J D Badger II et al ldquoMutant huntingtinimpairs axonal trafficking inmammalian neurons in vivo and invitrordquo Molecular and Cellular Biology vol 24 no 18 pp 8195ndash8209 2004
[24] A L Orr S Li C-E Wang et al ldquoN-terminal mutant hunt-ingtin associates with mitochondria and impairs mitochondrialtraffickingrdquo Journal of Neuroscience vol 28 no 11 pp 2783ndash2792 2008
Submit your manuscripts athttpwwwhindawicom
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Anatomy Research International
PeptidesInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporation httpwwwhindawicom
International Journal of
Volume 2014
Zoology
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Molecular Biology International
GenomicsInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioinformaticsAdvances in
Marine BiologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Signal TransductionJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
Evolutionary BiologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Biochemistry Research International
ArchaeaHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Genetics Research International
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Advances in
Virolog y
Hindawi Publishing Corporationhttpwwwhindawicom
Nucleic AcidsJournal of
Volume 2014
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Enzyme Research
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
International Journal of
Microbiology
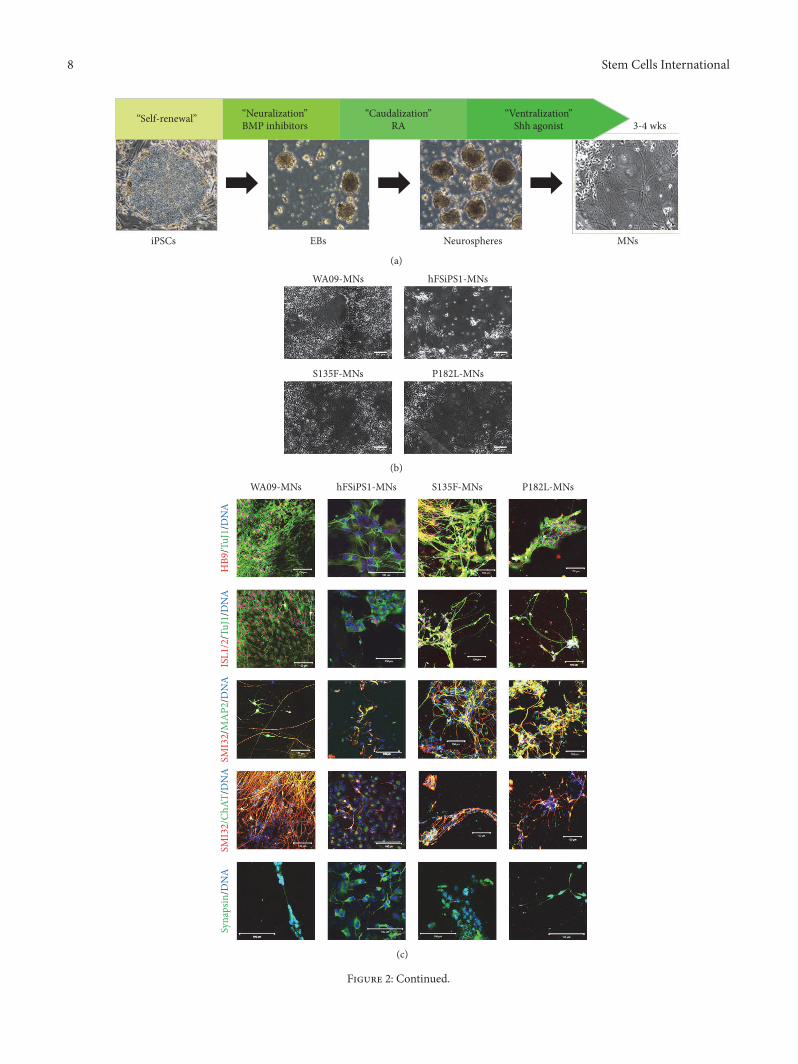
8 Stem Cells International
ldquoSelf-renewalrdquo
iPSCs EBs Neurospheres MNs
3-4 wksldquoNeuralizationrdquoBMP inhibitors
ldquoCaudalizationrdquoRA
ldquoVentralizationrdquoShh agonist
(a)
WA09-MNs hFSiPS1-MNs
S135F-MNs P182L-MNs
(b)
WA09-MNs S135F-MNs P182L-MNs
ISL1
2T
uJ1
DN
AH
B9T
uJ1
DN
ASM
I32
MA
P2D
NA
SMI3
2Ch
ATD
NA
Syna
psin
DN
A
hFSiPS1-MNs
(c)
Figure 2 Continued
Stem Cells International 9
SMI32DAPI
0
50
100
150Re
lativ
e exp
ress
ion
()
WA09-MNs hFSiPS1-MNs S135F-MNs P182L-MNs
Relat
ive e
xpre
ssio
n (
)
MAP2DAPI
0
50
100
150
WA09-MNs hFSiPS1-MNs S135F-MNs P182L-MNs(d)
Axonal length
0
500
1000
1500
2000
WA09-MNs S135F-MNs
(120583m
)
(e)
Figure 2 Generation of CMT2F-specific in vitro model by differentiation of patient-derived iPSCs into MNs (a) Schematic presentation ofMN differentiation (b) Differentiated MNs showed typical cellular morphology (original magnification 50x) Scale bars 200 120583m (c) S135F-MNs and P182L-MNs expressedMN-specific transcription factors such asHB9 and ISL12 neuronal cytoskeletalmarkers such as TuJ1 SMI32andMAP2 ChAT and the synaptic vesicularmarker synapsin (originalmagnification 400x and 600x) Scale bars 100 120583m (d) Differentiationefficiency of S135F-MNs and P182L-MNs was comparable to control MNs in terms of neuronal marker expression such as SMI32DAPI andMAP2DAPI (WA09-MNs119873 = 30 hFSiPS1-MNs119873 = 329 S135F-MNs119873 = 1730 and P182L-MNs119873 = 1090) (e) Axonal length of S135F-MNs was comparable to that of control MNs Axonal length was measured by culturing fully differentiated MNs in microchannel plates foran additional 2 weeks (WA09-MNs119873 = 70 and S135F-MNs119873 = 121)
HDAC6 is a type IIb HDAC that modulates deacetylationof cytoplasmic target proteins Because HDAC6 is involvedin cytoskeletal formation modulation of cellular oxida-tionreduction state and the ubiquitin-proteasome systeminhibition of HDAC6 improves pathological conditions inneurons such as impairments in axonal transport oxidativestress and protein aggresome formation in vitro [18 19]In terms of axonal transport HDAC6 inhibitors reverseCMT2F and dHMN2B-related axonal transport by increasingacetylated 120572-tubulin In this study newly developed HDAC6inhibitors having 35-fold lower half-maximal inhibitoryconcentration (IC
50) values than that of tubastatin A showed
better performance on improving mitochondrial axonaltransport at 10 times lower concentration than tubastatin A
CMT2F-iPSCs and dHMN2B-iPSCswere generated frompatient-derived dermal fibroblasts carrying 404CgtT (S135F)and 545CgtT (P182L) mutations of HSPB1 by forced expres-sion of KLF4 OCT34 SOX2 and c-MYC using Sendai virus
vectors After establishing several stem cell colonies twocolony clones from each patient were used for further analysisto avoid differences due to clonal variance and selectediPSCs were differentiated into motor neurons with definedcytokines To recapitulate axonal pathology in vitro patient-specific motor neurons were cultured in microchanneledplates which are designed to separate the microenvironmentsof neuronal compartments In this system specific cellularmodels of CMT2F and dHMN2B showed defects in mito-chondrial transport as the absolute velocity of moving mito-chondria and the percentage of moving mitochondria werelower than control MNs Phenotypic variance between iPSCclones was observed in P182L-MNs in terms ofmitochondrialvelocity (see Figure S3) Defects in the axonal traffic systemare linked to many neurodegenerative diseases Due to thedistinct anatomy of peripheral neurons which have axons upto one meter in length the interchange of genetic materialmetabolites proteins and organelles between soma and distal
10 Stem Cells International
Original magnification 50xBar = 200120583m
(a)
WA09
P182LhFSiPS1
(b)
Absolute velocity (120583msec)
00
01
02
03
WA09-MNs hFSiPS1-MNs S135F-MNs P182L-MNs
(120583m
sec
)
lowast
lowastlowastlowast
(c)
Moving mitochondria ()
0
20
40
60
80
()
WA09-MNs hFSiPS1-MNs S135F-MNs P182L-MNs
lowastlowastlowast
lowastlowastlowast
lowastlowastlowastlowast
(d)
Figure 3 Axonal transport of mitochondria in control MNs S135F-MNs and P182L-MNs (a) Microfluidic culturing system Microchannelplates were made from transparent polymer in a specialized mold to create multicompartments connected by 120583m-sized grooves (8334 120583mlong in length) that only allow growing axons to pass through thereby separating the microenvironments of neuronal components (b)Visualization of mitochondria by Mito-dsRED2 transfection (upper image) and kymograph images for mitochondria movement (middleand lower four images) Mito-dsRED2 vector was introduced into fully differentiated MNs cultured on microchannel plates for an additional1 weekMitochondrial images were taken at a ratio of 121 snaps2min and stacked into kymograph for analysis (originalmagnification 1000x)(c) S135F-MNs show decreased velocity of moving mitochondria compared to control MNs Unpaired t-testslowast119875 lt 005 lowastlowastlowast119875 lt 0001 andlowastlowastlowastlowast119875 lt 00001 Absolute velocity of moving mitochondria was calculated from measured angle and length in kymograph images using
ImageJ (WA09-MNs 119873 = 203 hFSiPS1-MNs 119873 = 512 S135F-MNs 119873 = 428 and P182L-MNs 119873 = 496) (d) S135F-MNs and P182L-MNs show reduced moving proportion of mitochondria in an axon (WA09-MNs 119873 = 9 hFSiPS1-MNs 119873 = 6 S135F-MNs 119873 = 20 andP182L-MNs119873 = 42)
subcellular sites is critical for maintaining neuronal home-ostasis and survival Of the various trafficking cargos thetransport of mitochondria is particularly important becausemitochondria support several cellular processes by supplyingATP buffering intracellular Ca2+ and modulating apoptosis[13] In addition to delivering mitochondria to appropriatesubcellular sites the trafficking of mitochondria is vital forthe recycling of their components through dynamic fusionand fission mechanism [14] Defects in axonal mitochondrialtransport are associated with Alzheimerrsquos disease [20 21]amyotrophic lateral sclerosis [22] and Huntingtonrsquos disease[23 24]
Mitochondria are transported bidirectionally by interac-tions with molecular motor proteins such as kinesin-1 foranterogrademovement and dynein for retrogrademovementand those attach via adaptor proteins such as the mitochon-drial Rho-Milton complex and dynactin respectively [14]The recruitment of molecularmotors tomicrotubule tracks isregulated by posttranslational modification of 120572-tubulin [16]especially acetylation [17] In this study the level of acetylated120572-tubulin was reduced in both S135F-MNs and P182L-MNsBy increasing the acetylation of 120572-tubulin via treatment withHDAC6 inhibitors mitochondrial transport was improvedin terms of both velocity and the percentage of moving
Stem Cells International 11
WA09-MNs hFSiPS1 S135F-MNs P182L-MNs120572
-tubu
linAc
etyl
120572-tu
bulin
Acet
yl120572
-tubu
lin
120572-tu
bulin
DN
A
(a)
00
02
04
06
08
10
WA09-MNs hFSiPS1-MNs S135F-MNs P182L-MNs
Acet
yl120572
-tubu
lin120572
-tubu
lin
lowastlowast
lowast
lowastlowast
lowastlowast
(b)
WA09-MNs hFSiPS1-MNs S135F-MNs P182L-MNs
120572-Tubulin
Acetyl 120572-tubulin
(c)
00
02
04
06
08
10
WA09-MNs hFSiPS1-MNs S135F-MNs P182L-MNs
Acet
yl120572
-tubu
lin120572
-tubu
linlowast
lowast
lowast
lowastlowast
(d)
Figure 4 Reduced acetylation of120572-tubulin in S135F-MNs andP182-MNs (a) Acetylation of120572-tubulinwas visualized by immunostainingwithanti-acetylated 120572-tubulin Abs (red) Total 120572-tubulin was also detected by staining with anti-120572-tubulin Abs (green) (original magnification400x) Scale bars 100120583m (b) The extent of acetylated 120572-tubulin was normalized with corresponding total 120572-tubulin using ImageJ software(WA09-MNs119873 = 3 S135F-MNs119873 = 4 and P182L-MNs119873 = 5) Unpaired 119905-tests lowast119875 lt 005 and lowastlowast119875 lt 001 (c and d) Western blot assayshowed decreased acetylated 120572-tubulin levels in S135F-MNs and P182-MNs (paired 119905-tests lowast119875 lt 005 and lowastlowast119875 lt 001)
mitochondria Hence defects in the axonal trafficking ofmitochondria are associated with decreased acetylation of120572-tubulin which may be the main causative factor foraxonopathy in CMT2F and dHMN2B
In summary iPSCs can be used for valuable resourcesof in vitro disease modeling as well as drug screening Weanticipate that this in vitro system would be a useful modelfor the clinical translational research and drug development
5 Conclusions
This study demonstrates in vitro drug screening mod-els of inherited neuropathy resulting from mutations in
the HSPB1 gene can be developed from patient-specificiPSCs Axonal pathologies were mimicked in microflu-idic culturing system as both S135F-MNs and P182L-MNsshowed a marked decline in the absolute velocity andthe percentage of moving mitochondria These axonal de-fects were associated with decreased acetylation of 120572-tu-bulin and were reversed by treatment of HDAC6 in-hibitors
Competing Interests
The authors declare that they have no conflict of interests
12 Stem Cells International
HDAC6 inhibitors
S135
F-M
Ns
P182
L-M
Ns
Acetyl 120572-tubulin
Acetyl 120572-tubulin
120572-Tubulin
120572-Tubulin
DM
SO
X4_0
5120583
M
X4_5
120583M
X9_0
5120583
M
X9_5
120583M
Tuba
statin
120583M
A_5
(a)
00
05
10
15
20
25
DM
SO
X4_0
5120583
M
X4_5
120583M
X9_0
5120583
M
X9_5
120583M
lowast
lowastlowast
lowastlowast
Acet
yl120572
-tubu
lin120572
-tubu
lin
Acetyl 120572-tubulin in S135F-MNs
00
05
10
15
20
25
DM
SO
X4_0
5120583
M
X4_5
120583M
X9_0
5120583
M
X9_5
120583M
lowast
lowastlowast
lowastlowastAc
etyl
120572-tu
bulin
120572-tu
bulin
Acetyl 120572-tubulin in P182L-MNs
Tuba
statin
120583M
A_5
Tuba
statin
120583M
A_5
(b)
00
01
02
03
04
DM
SO
X4_0
5120583
M
X4_5
120583M
X9_0
5120583
M
X9_5
120583M
lowastlowastlowastlowast
lowastlowastlowastlowast
Abso
lute
velo
city
(120583m
sec
)
00
02
04
06
DM
SO
X4_0
5120583
M
X4_5
120583M
X9_0
5120583
M
X9_5
120583M
lowast
lowastlowastlowastlowast
lowastlowastlowastlowast
lowastlowastlowastlowast
lowastlowast
Abso
lute
velo
city
(120583m
sec
)
S135F-MNs
Absolute velocity
P182L-MNs
Tuba
statin
120583M
A_5
Tuba
statin
120583M
A_5
(c)
Figure 5 Continued
Stem Cells International 13
0
20
40
60
80
Mov
ing
mito
chon
dria
()
DM
SO
X4_0
5120583
M
X4_5
120583M
X4_5
120583M
X9_0
5120583
M
X9_5
120583M
lowast
lowastlowast
lowastlowast
lowastlowastlowastlowast
0
20
40
60
80
Mov
ing
mito
chon
dria
()
DM
SO
X4_0
5120583
M
X9_0
5120583
M
X9_5
120583M
lowastlowastlowastlowast
lowastlowastlowastlowast
lowastlowastlowastlowast
lowastlowastlowastlowast
S135F-MNs
Moving mitochondria
P182L-MNs
Tuba
statin
120583M
A_5
Tuba
statin
120583M
A_5
lowastlowast
(d)
Figure 5 HDAC6 inhibitors reversed axonal defect of mitochondrial transport by increasing acetylation of 120572-tubulin (a and b) Westernblot assay showed that treatment of HDAC6 inhibitors increased acetylated 120572-tubulin levels in S135F-MNs and P182L-MNs (paired 119905-testslowast119875 lt 005 and lowastlowast119875 lt 001) (c) HDAC6 inhibitors increased the absolute velocity of mitochondrial movements in S135F-MNs and P182L-MNs (WA09-MNs119873 = 617 S135F-MNs119873 = 101 and P182L-MNs119873 = 394) (d) HDAC6 inhibitors increased the percentage of movingmitochondria in S135F-MNs and P182L-MNs (WA09-MNs 119873 = 15 S135F-MNs 119873 = 7 and P182L-MNs 119873 = 15) Unpaired 119905-testslowast119875 lt 005 lowastlowast119875 lt 001 and lowastlowastlowastlowast119875 lt 00001
Authorsrsquo Contributions
Ji-Yon Kim was responsible for the iPSC experiments collec-tion and interpretation of the data and preparation of thepaper So-Youn Woo was interpreting the data and editingthe manuscript Inhee Mook-Jung provided microchannelplates and designed the experiment Heesun Choi JisooKim and Hyunjung Choi carried out microfluidic cultureand kymograph analysis Nina Ha Jangbeen Kyung andSoo Kyung Koo designed the experiments and preparationof the manuscript Young-Bin Hong and Sung-Chul Jungwere responsible for conception design of experiments andediting the manuscript Byung-Ok Choi was responsible forthe collection of patient samples and interpretation of thedata
Acknowledgments
This study was supported by the Korean Health TechnologyRampD Project Ministry of Health amp Welfare (HI12C0135HI14C3484 and HI15C1560) and a National Research Foun-dation of Korea grant funded by the Ministry of Science ICTandFuture Planning (NRF-2014R1A2A2A01004240)Humanstem cell line hFSiPS1 was provided by National Stem CellBank of Korea (Korea National Institute of Health Grant no4861-307-210-13)
References
[1] Inherited Peripheral Neuropathies Mutation Database httpwwwmolgenuaacbecmtmutationsmutationsMutByGenecfm
[2] O V Evgrafov IMersiyanova J Irobi et al ldquoMutant small heat-shock protein 27 causes axonal Charcot-Marie-Tooth diseaseand distal hereditary motor neuropathyrdquo Nature Genetics vol36 no 6 pp 602ndash606 2004
[3] J Zhai H Lin J-P Julien andWW Schlaepfer ldquoDisruption ofneurofilament network with aggregation of light neurofilamentprotein a common pathway leading to motor neuron degener-ation due to Charcot-Marie-Tooth disease-linked mutations inNFL andHSPB1rdquoHumanMolecular Genetics vol 16 no 24 pp3103ndash3116 2007
[4] L Almeida-Souza B Asselbergh C drsquoYdewalle et al ldquoSmallheat-shock protein HSPB1 mutants stabilize microtubules inCharcot-Marie-Tooth neuropathyrdquoThe Journal of Neurosciencevol 31 no 43 pp 15320ndash15328 2011
[5] A Holmgren D Bouhy V De Winter et al ldquoCharcot-Marie-Tooth causing HSPB1mutations increase Cdk5-mediated phos-phorylation of neurofilamentsrdquoActa Neuropathologica vol 126no 1 pp 93ndash108 2013
[6] A-P Arrigo ldquoThe cellular ldquonetworkingrdquo of mammalian Hsp27and its functions in the control of protein folding redox stateand apoptosisrdquoAdvances in eXperimental Medicine and Biologyvol 594 pp 14ndash26 2007
14 Stem Cells International
[7] C DrsquoYdewalle J Krishnan D M Chiheb et al ldquoHDAC6inhibitors reverse axonal loss in a mouse model of mutantHSPB1-induced Charcot-Marie-Tooth diseaserdquo NatureMedicine vol 17 no 8 pp 968ndash974 2011
[8] Y Zhang N Li C Caron et al ldquoHDAC-6 interacts with anddeacetylates tubulin and microtubules in vivordquo EMBO Journalvol 22 no 5 pp 1168ndash1179 2003
[9] A Valenzuela-Fernandez J R Cabrero J M Serrador and FSanchez-Madrid ldquoHDAC6 a key regulator of cytoskeleton cellmigration and cell-cell interactionsrdquo Trends in Cell Biology vol18 no 6 pp 291ndash297 2008
[10] V K Godena N Brookes-Hocking AMoller et al ldquoIncreasingmicrotubule acetylation rescues axonal transport and locomo-tor deficits caused by LRRK2 Roc-COR domain mutationsrdquoNature Communications vol 5 article 5245 2014
[11] J P Dompierre J D Godin B C Charrin et al ldquoHistonedeacetylase 6 inhibition compensates for the transport deficitin Huntingtonrsquos disease by increasing tubulin acetylationrdquo TheJournal of Neuroscience vol 27 no 13 pp 3571ndash3583 2007
[12] M W Amoroso G F Croft D J Williams et al ldquoAcceleratedhigh-yield generation of limb-innervating motor neurons fromhuman stem cellsrdquo Journal of Neuroscience vol 33 no 2 pp574ndash586 2013
[13] B J Gentil and L Cooper ldquoMolecular basis of axonal dysfunc-tion and traffic impairments in CMTrdquo Brain Research Bulletinvol 88 no 5 pp 444ndash453 2012
[14] Z-H Sheng and Q Cai ldquoMitochondrial transport in neu-rons impact on synaptic homeostasis and neurodegenerationrdquoNature Reviews Neuroscience vol 13 no 2 pp 77ndash93 2012
[15] J W Park B Vahidi A M Taylor S W Rhee and N LJeon ldquoMicrofluidic culture platform for neuroscience researchrdquoNature Protocols vol 1 no 4 pp 2128ndash2136 2006
[16] SWestermann and KWeber ldquoPost-translational modificationsregulate microtubule functionrdquo Nature Reviews Molecular CellBiology vol 4 no 12 pp 938ndash947 2003
[17] N A Reed D Cai T L Blasius et al ldquoMicrotubule acetylationpromotes kinesin-1 binding and transportrdquoCurrent Biology vol16 no 21 pp 2166ndash2172 2006
[18] C drsquoYdewalle E Bogaert and L Van Den Bosch ldquoHDAC6at the intersection of neuroprotection and neurodegenerationrdquoTraffic vol 13 no 6 pp 771ndash779 2012
[19] C Simoes-Pires V Zwick A Nurisso E Schenker P-ACarrupt and M Cuendet ldquoHDAC6 as a target for neurode-generative diseases what makes it different from the otherHDACsrdquoMolecular Neurodegeneration vol 8 article 7 2013
[20] Y Rui P Tiwari Z Xie and J Q Zheng ldquoAcute impairment ofmitochondrial trafficking by 120573-amyloid peptides in hippocam-pal neuronsrdquo Journal of Neuroscience vol 26 no 41 pp 10480ndash10487 2006
[21] K A Vossel K Zhang J Brodbeck et al ldquoTau reductionprevents A120573-induced defects in axonal transportrdquo Science vol330 no 6001 p 198 2010
[22] K J De vos A L Chapman M E Tennant et al ldquoFamilialamyotrophic lateral sclerosis-linked SOD1 mutants perturbfast axonal transport to reduce axonal mitochondria contentrdquoHumanMolecular Genetics vol 16 no 22 pp 2720ndash2728 2007
[23] E Trushina R B Dyer J D Badger II et al ldquoMutant huntingtinimpairs axonal trafficking inmammalian neurons in vivo and invitrordquo Molecular and Cellular Biology vol 24 no 18 pp 8195ndash8209 2004
[24] A L Orr S Li C-E Wang et al ldquoN-terminal mutant hunt-ingtin associates with mitochondria and impairs mitochondrialtraffickingrdquo Journal of Neuroscience vol 28 no 11 pp 2783ndash2792 2008
Submit your manuscripts athttpwwwhindawicom
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Anatomy Research International
PeptidesInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporation httpwwwhindawicom
International Journal of
Volume 2014
Zoology
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Molecular Biology International
GenomicsInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioinformaticsAdvances in
Marine BiologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Signal TransductionJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
Evolutionary BiologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Biochemistry Research International
ArchaeaHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Genetics Research International
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Advances in
Virolog y
Hindawi Publishing Corporationhttpwwwhindawicom
Nucleic AcidsJournal of
Volume 2014
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Enzyme Research
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
International Journal of
Microbiology
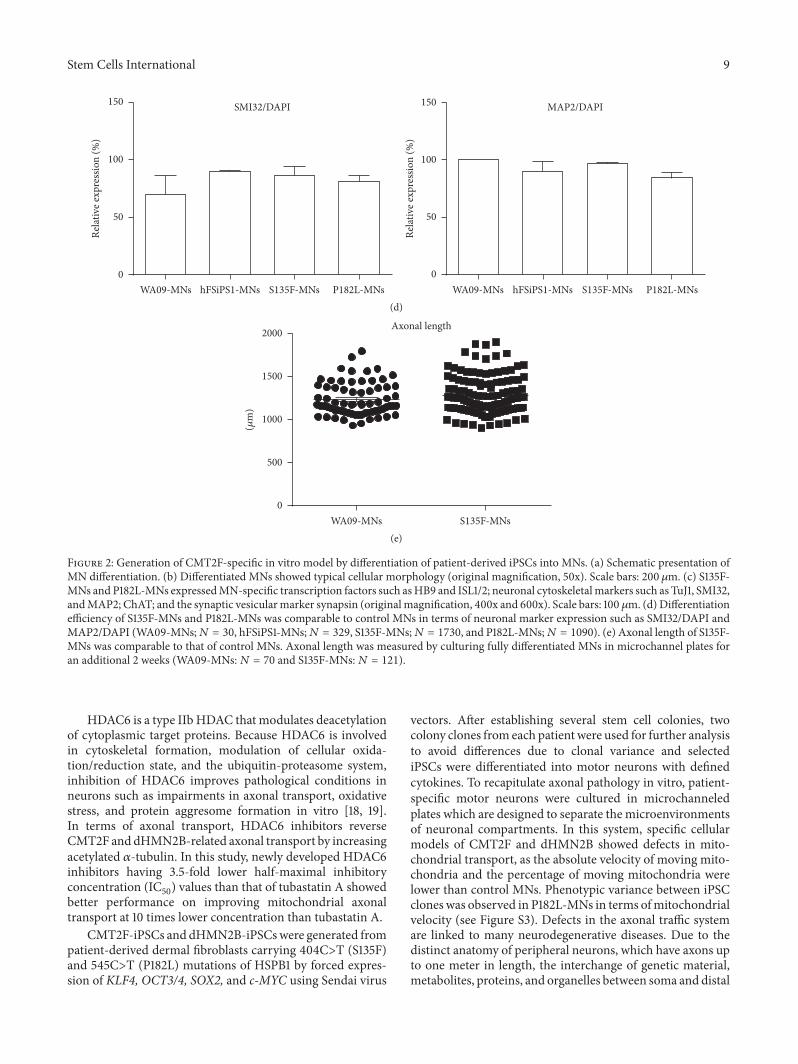
Stem Cells International 9
SMI32DAPI
0
50
100
150Re
lativ
e exp
ress
ion
()
WA09-MNs hFSiPS1-MNs S135F-MNs P182L-MNs
Relat
ive e
xpre
ssio
n (
)
MAP2DAPI
0
50
100
150
WA09-MNs hFSiPS1-MNs S135F-MNs P182L-MNs(d)
Axonal length
0
500
1000
1500
2000
WA09-MNs S135F-MNs
(120583m
)
(e)
Figure 2 Generation of CMT2F-specific in vitro model by differentiation of patient-derived iPSCs into MNs (a) Schematic presentation ofMN differentiation (b) Differentiated MNs showed typical cellular morphology (original magnification 50x) Scale bars 200 120583m (c) S135F-MNs and P182L-MNs expressedMN-specific transcription factors such asHB9 and ISL12 neuronal cytoskeletalmarkers such as TuJ1 SMI32andMAP2 ChAT and the synaptic vesicularmarker synapsin (originalmagnification 400x and 600x) Scale bars 100 120583m (d) Differentiationefficiency of S135F-MNs and P182L-MNs was comparable to control MNs in terms of neuronal marker expression such as SMI32DAPI andMAP2DAPI (WA09-MNs119873 = 30 hFSiPS1-MNs119873 = 329 S135F-MNs119873 = 1730 and P182L-MNs119873 = 1090) (e) Axonal length of S135F-MNs was comparable to that of control MNs Axonal length was measured by culturing fully differentiated MNs in microchannel plates foran additional 2 weeks (WA09-MNs119873 = 70 and S135F-MNs119873 = 121)
HDAC6 is a type IIb HDAC that modulates deacetylationof cytoplasmic target proteins Because HDAC6 is involvedin cytoskeletal formation modulation of cellular oxida-tionreduction state and the ubiquitin-proteasome systeminhibition of HDAC6 improves pathological conditions inneurons such as impairments in axonal transport oxidativestress and protein aggresome formation in vitro [18 19]In terms of axonal transport HDAC6 inhibitors reverseCMT2F and dHMN2B-related axonal transport by increasingacetylated 120572-tubulin In this study newly developed HDAC6inhibitors having 35-fold lower half-maximal inhibitoryconcentration (IC
50) values than that of tubastatin A showed
better performance on improving mitochondrial axonaltransport at 10 times lower concentration than tubastatin A
CMT2F-iPSCs and dHMN2B-iPSCswere generated frompatient-derived dermal fibroblasts carrying 404CgtT (S135F)and 545CgtT (P182L) mutations of HSPB1 by forced expres-sion of KLF4 OCT34 SOX2 and c-MYC using Sendai virus
vectors After establishing several stem cell colonies twocolony clones from each patient were used for further analysisto avoid differences due to clonal variance and selectediPSCs were differentiated into motor neurons with definedcytokines To recapitulate axonal pathology in vitro patient-specific motor neurons were cultured in microchanneledplates which are designed to separate the microenvironmentsof neuronal compartments In this system specific cellularmodels of CMT2F and dHMN2B showed defects in mito-chondrial transport as the absolute velocity of moving mito-chondria and the percentage of moving mitochondria werelower than control MNs Phenotypic variance between iPSCclones was observed in P182L-MNs in terms ofmitochondrialvelocity (see Figure S3) Defects in the axonal traffic systemare linked to many neurodegenerative diseases Due to thedistinct anatomy of peripheral neurons which have axons upto one meter in length the interchange of genetic materialmetabolites proteins and organelles between soma and distal
10 Stem Cells International
Original magnification 50xBar = 200120583m
(a)
WA09
P182LhFSiPS1
(b)
Absolute velocity (120583msec)
00
01
02
03
WA09-MNs hFSiPS1-MNs S135F-MNs P182L-MNs
(120583m
sec
)
lowast
lowastlowastlowast
(c)
Moving mitochondria ()
0
20
40
60
80
()
WA09-MNs hFSiPS1-MNs S135F-MNs P182L-MNs
lowastlowastlowast
lowastlowastlowast
lowastlowastlowastlowast
(d)
Figure 3 Axonal transport of mitochondria in control MNs S135F-MNs and P182L-MNs (a) Microfluidic culturing system Microchannelplates were made from transparent polymer in a specialized mold to create multicompartments connected by 120583m-sized grooves (8334 120583mlong in length) that only allow growing axons to pass through thereby separating the microenvironments of neuronal components (b)Visualization of mitochondria by Mito-dsRED2 transfection (upper image) and kymograph images for mitochondria movement (middleand lower four images) Mito-dsRED2 vector was introduced into fully differentiated MNs cultured on microchannel plates for an additional1 weekMitochondrial images were taken at a ratio of 121 snaps2min and stacked into kymograph for analysis (originalmagnification 1000x)(c) S135F-MNs show decreased velocity of moving mitochondria compared to control MNs Unpaired t-testslowast119875 lt 005 lowastlowastlowast119875 lt 0001 andlowastlowastlowastlowast119875 lt 00001 Absolute velocity of moving mitochondria was calculated from measured angle and length in kymograph images using
ImageJ (WA09-MNs 119873 = 203 hFSiPS1-MNs 119873 = 512 S135F-MNs 119873 = 428 and P182L-MNs 119873 = 496) (d) S135F-MNs and P182L-MNs show reduced moving proportion of mitochondria in an axon (WA09-MNs 119873 = 9 hFSiPS1-MNs 119873 = 6 S135F-MNs 119873 = 20 andP182L-MNs119873 = 42)
subcellular sites is critical for maintaining neuronal home-ostasis and survival Of the various trafficking cargos thetransport of mitochondria is particularly important becausemitochondria support several cellular processes by supplyingATP buffering intracellular Ca2+ and modulating apoptosis[13] In addition to delivering mitochondria to appropriatesubcellular sites the trafficking of mitochondria is vital forthe recycling of their components through dynamic fusionand fission mechanism [14] Defects in axonal mitochondrialtransport are associated with Alzheimerrsquos disease [20 21]amyotrophic lateral sclerosis [22] and Huntingtonrsquos disease[23 24]
Mitochondria are transported bidirectionally by interac-tions with molecular motor proteins such as kinesin-1 foranterogrademovement and dynein for retrogrademovementand those attach via adaptor proteins such as the mitochon-drial Rho-Milton complex and dynactin respectively [14]The recruitment of molecularmotors tomicrotubule tracks isregulated by posttranslational modification of 120572-tubulin [16]especially acetylation [17] In this study the level of acetylated120572-tubulin was reduced in both S135F-MNs and P182L-MNsBy increasing the acetylation of 120572-tubulin via treatment withHDAC6 inhibitors mitochondrial transport was improvedin terms of both velocity and the percentage of moving
Stem Cells International 11
WA09-MNs hFSiPS1 S135F-MNs P182L-MNs120572
-tubu
linAc
etyl
120572-tu
bulin
Acet
yl120572
-tubu
lin
120572-tu
bulin
DN
A
(a)
00
02
04
06
08
10
WA09-MNs hFSiPS1-MNs S135F-MNs P182L-MNs
Acet
yl120572
-tubu
lin120572
-tubu
lin
lowastlowast
lowast
lowastlowast
lowastlowast
(b)
WA09-MNs hFSiPS1-MNs S135F-MNs P182L-MNs
120572-Tubulin
Acetyl 120572-tubulin
(c)
00
02
04
06
08
10
WA09-MNs hFSiPS1-MNs S135F-MNs P182L-MNs
Acet
yl120572
-tubu
lin120572
-tubu
linlowast
lowast
lowast
lowastlowast
(d)
Figure 4 Reduced acetylation of120572-tubulin in S135F-MNs andP182-MNs (a) Acetylation of120572-tubulinwas visualized by immunostainingwithanti-acetylated 120572-tubulin Abs (red) Total 120572-tubulin was also detected by staining with anti-120572-tubulin Abs (green) (original magnification400x) Scale bars 100120583m (b) The extent of acetylated 120572-tubulin was normalized with corresponding total 120572-tubulin using ImageJ software(WA09-MNs119873 = 3 S135F-MNs119873 = 4 and P182L-MNs119873 = 5) Unpaired 119905-tests lowast119875 lt 005 and lowastlowast119875 lt 001 (c and d) Western blot assayshowed decreased acetylated 120572-tubulin levels in S135F-MNs and P182-MNs (paired 119905-tests lowast119875 lt 005 and lowastlowast119875 lt 001)
mitochondria Hence defects in the axonal trafficking ofmitochondria are associated with decreased acetylation of120572-tubulin which may be the main causative factor foraxonopathy in CMT2F and dHMN2B
In summary iPSCs can be used for valuable resourcesof in vitro disease modeling as well as drug screening Weanticipate that this in vitro system would be a useful modelfor the clinical translational research and drug development
5 Conclusions
This study demonstrates in vitro drug screening mod-els of inherited neuropathy resulting from mutations in
the HSPB1 gene can be developed from patient-specificiPSCs Axonal pathologies were mimicked in microflu-idic culturing system as both S135F-MNs and P182L-MNsshowed a marked decline in the absolute velocity andthe percentage of moving mitochondria These axonal de-fects were associated with decreased acetylation of 120572-tu-bulin and were reversed by treatment of HDAC6 in-hibitors
Competing Interests
The authors declare that they have no conflict of interests
12 Stem Cells International
HDAC6 inhibitors
S135
F-M
Ns
P182
L-M
Ns
Acetyl 120572-tubulin
Acetyl 120572-tubulin
120572-Tubulin
120572-Tubulin
DM
SO
X4_0
5120583
M
X4_5
120583M
X9_0
5120583
M
X9_5
120583M
Tuba
statin
120583M
A_5
(a)
00
05
10
15
20
25
DM
SO
X4_0
5120583
M
X4_5
120583M
X9_0
5120583
M
X9_5
120583M
lowast
lowastlowast
lowastlowast
Acet
yl120572
-tubu
lin120572
-tubu
lin
Acetyl 120572-tubulin in S135F-MNs
00
05
10
15
20
25
DM
SO
X4_0
5120583
M
X4_5
120583M
X9_0
5120583
M
X9_5
120583M
lowast
lowastlowast
lowastlowastAc
etyl
120572-tu
bulin
120572-tu
bulin
Acetyl 120572-tubulin in P182L-MNs
Tuba
statin
120583M
A_5
Tuba
statin
120583M
A_5
(b)
00
01
02
03
04
DM
SO
X4_0
5120583
M
X4_5
120583M
X9_0
5120583
M
X9_5
120583M
lowastlowastlowastlowast
lowastlowastlowastlowast
Abso
lute
velo
city
(120583m
sec
)
00
02
04
06
DM
SO
X4_0
5120583
M
X4_5
120583M
X9_0
5120583
M
X9_5
120583M
lowast
lowastlowastlowastlowast
lowastlowastlowastlowast
lowastlowastlowastlowast
lowastlowast
Abso
lute
velo
city
(120583m
sec
)
S135F-MNs
Absolute velocity
P182L-MNs
Tuba
statin
120583M
A_5
Tuba
statin
120583M
A_5
(c)
Figure 5 Continued
Stem Cells International 13
0
20
40
60
80
Mov
ing
mito
chon
dria
()
DM
SO
X4_0
5120583
M
X4_5
120583M
X4_5
120583M
X9_0
5120583
M
X9_5
120583M
lowast
lowastlowast
lowastlowast
lowastlowastlowastlowast
0
20
40
60
80
Mov
ing
mito
chon
dria
()
DM
SO
X4_0
5120583
M
X9_0
5120583
M
X9_5
120583M
lowastlowastlowastlowast
lowastlowastlowastlowast
lowastlowastlowastlowast
lowastlowastlowastlowast
S135F-MNs
Moving mitochondria
P182L-MNs
Tuba
statin
120583M
A_5
Tuba
statin
120583M
A_5
lowastlowast
(d)
Figure 5 HDAC6 inhibitors reversed axonal defect of mitochondrial transport by increasing acetylation of 120572-tubulin (a and b) Westernblot assay showed that treatment of HDAC6 inhibitors increased acetylated 120572-tubulin levels in S135F-MNs and P182L-MNs (paired 119905-testslowast119875 lt 005 and lowastlowast119875 lt 001) (c) HDAC6 inhibitors increased the absolute velocity of mitochondrial movements in S135F-MNs and P182L-MNs (WA09-MNs119873 = 617 S135F-MNs119873 = 101 and P182L-MNs119873 = 394) (d) HDAC6 inhibitors increased the percentage of movingmitochondria in S135F-MNs and P182L-MNs (WA09-MNs 119873 = 15 S135F-MNs 119873 = 7 and P182L-MNs 119873 = 15) Unpaired 119905-testslowast119875 lt 005 lowastlowast119875 lt 001 and lowastlowastlowastlowast119875 lt 00001
Authorsrsquo Contributions
Ji-Yon Kim was responsible for the iPSC experiments collec-tion and interpretation of the data and preparation of thepaper So-Youn Woo was interpreting the data and editingthe manuscript Inhee Mook-Jung provided microchannelplates and designed the experiment Heesun Choi JisooKim and Hyunjung Choi carried out microfluidic cultureand kymograph analysis Nina Ha Jangbeen Kyung andSoo Kyung Koo designed the experiments and preparationof the manuscript Young-Bin Hong and Sung-Chul Jungwere responsible for conception design of experiments andediting the manuscript Byung-Ok Choi was responsible forthe collection of patient samples and interpretation of thedata
Acknowledgments
This study was supported by the Korean Health TechnologyRampD Project Ministry of Health amp Welfare (HI12C0135HI14C3484 and HI15C1560) and a National Research Foun-dation of Korea grant funded by the Ministry of Science ICTandFuture Planning (NRF-2014R1A2A2A01004240)Humanstem cell line hFSiPS1 was provided by National Stem CellBank of Korea (Korea National Institute of Health Grant no4861-307-210-13)
References
[1] Inherited Peripheral Neuropathies Mutation Database httpwwwmolgenuaacbecmtmutationsmutationsMutByGenecfm
[2] O V Evgrafov IMersiyanova J Irobi et al ldquoMutant small heat-shock protein 27 causes axonal Charcot-Marie-Tooth diseaseand distal hereditary motor neuropathyrdquo Nature Genetics vol36 no 6 pp 602ndash606 2004
[3] J Zhai H Lin J-P Julien andWW Schlaepfer ldquoDisruption ofneurofilament network with aggregation of light neurofilamentprotein a common pathway leading to motor neuron degener-ation due to Charcot-Marie-Tooth disease-linked mutations inNFL andHSPB1rdquoHumanMolecular Genetics vol 16 no 24 pp3103ndash3116 2007
[4] L Almeida-Souza B Asselbergh C drsquoYdewalle et al ldquoSmallheat-shock protein HSPB1 mutants stabilize microtubules inCharcot-Marie-Tooth neuropathyrdquoThe Journal of Neurosciencevol 31 no 43 pp 15320ndash15328 2011
[5] A Holmgren D Bouhy V De Winter et al ldquoCharcot-Marie-Tooth causing HSPB1mutations increase Cdk5-mediated phos-phorylation of neurofilamentsrdquoActa Neuropathologica vol 126no 1 pp 93ndash108 2013
[6] A-P Arrigo ldquoThe cellular ldquonetworkingrdquo of mammalian Hsp27and its functions in the control of protein folding redox stateand apoptosisrdquoAdvances in eXperimental Medicine and Biologyvol 594 pp 14ndash26 2007
14 Stem Cells International
[7] C DrsquoYdewalle J Krishnan D M Chiheb et al ldquoHDAC6inhibitors reverse axonal loss in a mouse model of mutantHSPB1-induced Charcot-Marie-Tooth diseaserdquo NatureMedicine vol 17 no 8 pp 968ndash974 2011
[8] Y Zhang N Li C Caron et al ldquoHDAC-6 interacts with anddeacetylates tubulin and microtubules in vivordquo EMBO Journalvol 22 no 5 pp 1168ndash1179 2003
[9] A Valenzuela-Fernandez J R Cabrero J M Serrador and FSanchez-Madrid ldquoHDAC6 a key regulator of cytoskeleton cellmigration and cell-cell interactionsrdquo Trends in Cell Biology vol18 no 6 pp 291ndash297 2008
[10] V K Godena N Brookes-Hocking AMoller et al ldquoIncreasingmicrotubule acetylation rescues axonal transport and locomo-tor deficits caused by LRRK2 Roc-COR domain mutationsrdquoNature Communications vol 5 article 5245 2014
[11] J P Dompierre J D Godin B C Charrin et al ldquoHistonedeacetylase 6 inhibition compensates for the transport deficitin Huntingtonrsquos disease by increasing tubulin acetylationrdquo TheJournal of Neuroscience vol 27 no 13 pp 3571ndash3583 2007
[12] M W Amoroso G F Croft D J Williams et al ldquoAcceleratedhigh-yield generation of limb-innervating motor neurons fromhuman stem cellsrdquo Journal of Neuroscience vol 33 no 2 pp574ndash586 2013
[13] B J Gentil and L Cooper ldquoMolecular basis of axonal dysfunc-tion and traffic impairments in CMTrdquo Brain Research Bulletinvol 88 no 5 pp 444ndash453 2012
[14] Z-H Sheng and Q Cai ldquoMitochondrial transport in neu-rons impact on synaptic homeostasis and neurodegenerationrdquoNature Reviews Neuroscience vol 13 no 2 pp 77ndash93 2012
[15] J W Park B Vahidi A M Taylor S W Rhee and N LJeon ldquoMicrofluidic culture platform for neuroscience researchrdquoNature Protocols vol 1 no 4 pp 2128ndash2136 2006
[16] SWestermann and KWeber ldquoPost-translational modificationsregulate microtubule functionrdquo Nature Reviews Molecular CellBiology vol 4 no 12 pp 938ndash947 2003
[17] N A Reed D Cai T L Blasius et al ldquoMicrotubule acetylationpromotes kinesin-1 binding and transportrdquoCurrent Biology vol16 no 21 pp 2166ndash2172 2006
[18] C drsquoYdewalle E Bogaert and L Van Den Bosch ldquoHDAC6at the intersection of neuroprotection and neurodegenerationrdquoTraffic vol 13 no 6 pp 771ndash779 2012
[19] C Simoes-Pires V Zwick A Nurisso E Schenker P-ACarrupt and M Cuendet ldquoHDAC6 as a target for neurode-generative diseases what makes it different from the otherHDACsrdquoMolecular Neurodegeneration vol 8 article 7 2013
[20] Y Rui P Tiwari Z Xie and J Q Zheng ldquoAcute impairment ofmitochondrial trafficking by 120573-amyloid peptides in hippocam-pal neuronsrdquo Journal of Neuroscience vol 26 no 41 pp 10480ndash10487 2006
[21] K A Vossel K Zhang J Brodbeck et al ldquoTau reductionprevents A120573-induced defects in axonal transportrdquo Science vol330 no 6001 p 198 2010
[22] K J De vos A L Chapman M E Tennant et al ldquoFamilialamyotrophic lateral sclerosis-linked SOD1 mutants perturbfast axonal transport to reduce axonal mitochondria contentrdquoHumanMolecular Genetics vol 16 no 22 pp 2720ndash2728 2007
[23] E Trushina R B Dyer J D Badger II et al ldquoMutant huntingtinimpairs axonal trafficking inmammalian neurons in vivo and invitrordquo Molecular and Cellular Biology vol 24 no 18 pp 8195ndash8209 2004
[24] A L Orr S Li C-E Wang et al ldquoN-terminal mutant hunt-ingtin associates with mitochondria and impairs mitochondrialtraffickingrdquo Journal of Neuroscience vol 28 no 11 pp 2783ndash2792 2008
Submit your manuscripts athttpwwwhindawicom
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Anatomy Research International
PeptidesInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporation httpwwwhindawicom
International Journal of
Volume 2014
Zoology
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Molecular Biology International
GenomicsInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioinformaticsAdvances in
Marine BiologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Signal TransductionJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
Evolutionary BiologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Biochemistry Research International
ArchaeaHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Genetics Research International
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Advances in
Virolog y
Hindawi Publishing Corporationhttpwwwhindawicom
Nucleic AcidsJournal of
Volume 2014
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Enzyme Research
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
International Journal of
Microbiology
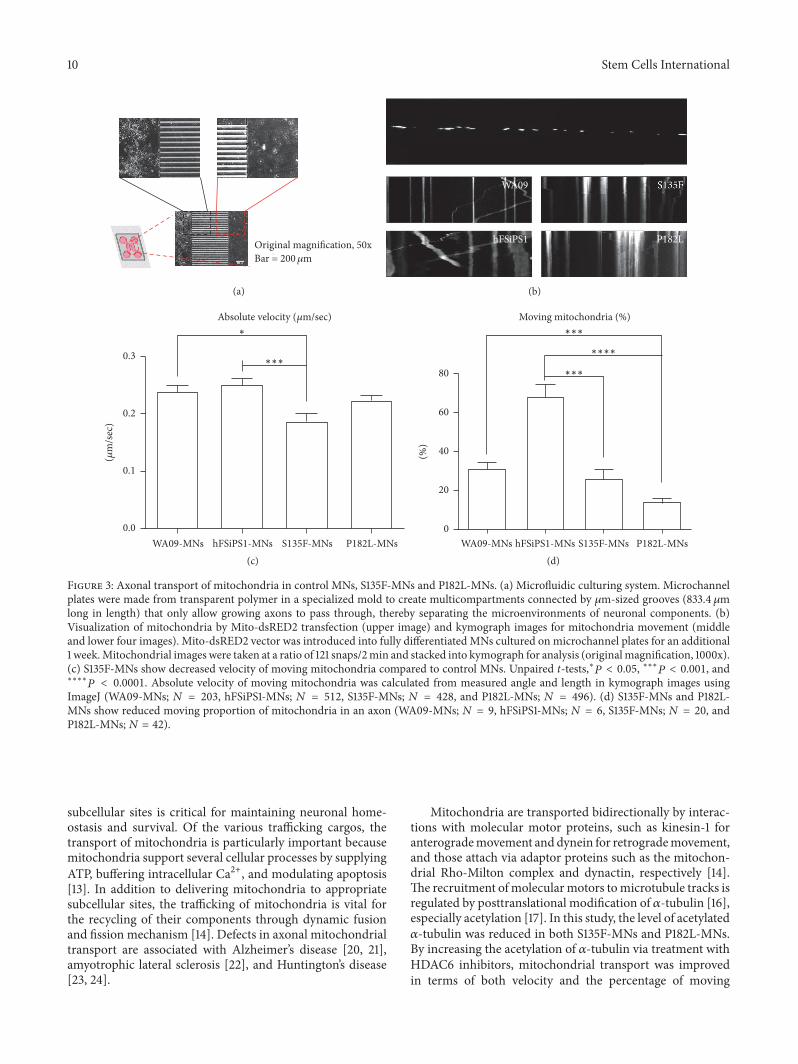
10 Stem Cells International
Original magnification 50xBar = 200120583m
(a)
WA09
P182LhFSiPS1
(b)
Absolute velocity (120583msec)
00
01
02
03
WA09-MNs hFSiPS1-MNs S135F-MNs P182L-MNs
(120583m
sec
)
lowast
lowastlowastlowast
(c)
Moving mitochondria ()
0
20
40
60
80
()
WA09-MNs hFSiPS1-MNs S135F-MNs P182L-MNs
lowastlowastlowast
lowastlowastlowast
lowastlowastlowastlowast
(d)
Figure 3 Axonal transport of mitochondria in control MNs S135F-MNs and P182L-MNs (a) Microfluidic culturing system Microchannelplates were made from transparent polymer in a specialized mold to create multicompartments connected by 120583m-sized grooves (8334 120583mlong in length) that only allow growing axons to pass through thereby separating the microenvironments of neuronal components (b)Visualization of mitochondria by Mito-dsRED2 transfection (upper image) and kymograph images for mitochondria movement (middleand lower four images) Mito-dsRED2 vector was introduced into fully differentiated MNs cultured on microchannel plates for an additional1 weekMitochondrial images were taken at a ratio of 121 snaps2min and stacked into kymograph for analysis (originalmagnification 1000x)(c) S135F-MNs show decreased velocity of moving mitochondria compared to control MNs Unpaired t-testslowast119875 lt 005 lowastlowastlowast119875 lt 0001 andlowastlowastlowastlowast119875 lt 00001 Absolute velocity of moving mitochondria was calculated from measured angle and length in kymograph images using
ImageJ (WA09-MNs 119873 = 203 hFSiPS1-MNs 119873 = 512 S135F-MNs 119873 = 428 and P182L-MNs 119873 = 496) (d) S135F-MNs and P182L-MNs show reduced moving proportion of mitochondria in an axon (WA09-MNs 119873 = 9 hFSiPS1-MNs 119873 = 6 S135F-MNs 119873 = 20 andP182L-MNs119873 = 42)
subcellular sites is critical for maintaining neuronal home-ostasis and survival Of the various trafficking cargos thetransport of mitochondria is particularly important becausemitochondria support several cellular processes by supplyingATP buffering intracellular Ca2+ and modulating apoptosis[13] In addition to delivering mitochondria to appropriatesubcellular sites the trafficking of mitochondria is vital forthe recycling of their components through dynamic fusionand fission mechanism [14] Defects in axonal mitochondrialtransport are associated with Alzheimerrsquos disease [20 21]amyotrophic lateral sclerosis [22] and Huntingtonrsquos disease[23 24]
Mitochondria are transported bidirectionally by interac-tions with molecular motor proteins such as kinesin-1 foranterogrademovement and dynein for retrogrademovementand those attach via adaptor proteins such as the mitochon-drial Rho-Milton complex and dynactin respectively [14]The recruitment of molecularmotors tomicrotubule tracks isregulated by posttranslational modification of 120572-tubulin [16]especially acetylation [17] In this study the level of acetylated120572-tubulin was reduced in both S135F-MNs and P182L-MNsBy increasing the acetylation of 120572-tubulin via treatment withHDAC6 inhibitors mitochondrial transport was improvedin terms of both velocity and the percentage of moving
Stem Cells International 11
WA09-MNs hFSiPS1 S135F-MNs P182L-MNs120572
-tubu
linAc
etyl
120572-tu
bulin
Acet
yl120572
-tubu
lin
120572-tu
bulin
DN
A
(a)
00
02
04
06
08
10
WA09-MNs hFSiPS1-MNs S135F-MNs P182L-MNs
Acet
yl120572
-tubu
lin120572
-tubu
lin
lowastlowast
lowast
lowastlowast
lowastlowast
(b)
WA09-MNs hFSiPS1-MNs S135F-MNs P182L-MNs
120572-Tubulin
Acetyl 120572-tubulin
(c)
00
02
04
06
08
10
WA09-MNs hFSiPS1-MNs S135F-MNs P182L-MNs
Acet
yl120572
-tubu
lin120572
-tubu
linlowast
lowast
lowast
lowastlowast
(d)
Figure 4 Reduced acetylation of120572-tubulin in S135F-MNs andP182-MNs (a) Acetylation of120572-tubulinwas visualized by immunostainingwithanti-acetylated 120572-tubulin Abs (red) Total 120572-tubulin was also detected by staining with anti-120572-tubulin Abs (green) (original magnification400x) Scale bars 100120583m (b) The extent of acetylated 120572-tubulin was normalized with corresponding total 120572-tubulin using ImageJ software(WA09-MNs119873 = 3 S135F-MNs119873 = 4 and P182L-MNs119873 = 5) Unpaired 119905-tests lowast119875 lt 005 and lowastlowast119875 lt 001 (c and d) Western blot assayshowed decreased acetylated 120572-tubulin levels in S135F-MNs and P182-MNs (paired 119905-tests lowast119875 lt 005 and lowastlowast119875 lt 001)
mitochondria Hence defects in the axonal trafficking ofmitochondria are associated with decreased acetylation of120572-tubulin which may be the main causative factor foraxonopathy in CMT2F and dHMN2B
In summary iPSCs can be used for valuable resourcesof in vitro disease modeling as well as drug screening Weanticipate that this in vitro system would be a useful modelfor the clinical translational research and drug development
5 Conclusions
This study demonstrates in vitro drug screening mod-els of inherited neuropathy resulting from mutations in
the HSPB1 gene can be developed from patient-specificiPSCs Axonal pathologies were mimicked in microflu-idic culturing system as both S135F-MNs and P182L-MNsshowed a marked decline in the absolute velocity andthe percentage of moving mitochondria These axonal de-fects were associated with decreased acetylation of 120572-tu-bulin and were reversed by treatment of HDAC6 in-hibitors
Competing Interests
The authors declare that they have no conflict of interests
12 Stem Cells International
HDAC6 inhibitors
S135
F-M
Ns
P182
L-M
Ns
Acetyl 120572-tubulin
Acetyl 120572-tubulin
120572-Tubulin
120572-Tubulin
DM
SO
X4_0
5120583
M
X4_5
120583M
X9_0
5120583
M
X9_5
120583M
Tuba
statin
120583M
A_5
(a)
00
05
10
15
20
25
DM
SO
X4_0
5120583
M
X4_5
120583M
X9_0
5120583
M
X9_5
120583M
lowast
lowastlowast
lowastlowast
Acet
yl120572
-tubu
lin120572
-tubu
lin
Acetyl 120572-tubulin in S135F-MNs
00
05
10
15
20
25
DM
SO
X4_0
5120583
M
X4_5
120583M
X9_0
5120583
M
X9_5
120583M
lowast
lowastlowast
lowastlowastAc
etyl
120572-tu
bulin
120572-tu
bulin
Acetyl 120572-tubulin in P182L-MNs
Tuba
statin
120583M
A_5
Tuba
statin
120583M
A_5
(b)
00
01
02
03
04
DM
SO
X4_0
5120583
M
X4_5
120583M
X9_0
5120583
M
X9_5
120583M
lowastlowastlowastlowast
lowastlowastlowastlowast
Abso
lute
velo
city
(120583m
sec
)
00
02
04
06
DM
SO
X4_0
5120583
M
X4_5
120583M
X9_0
5120583
M
X9_5
120583M
lowast
lowastlowastlowastlowast
lowastlowastlowastlowast
lowastlowastlowastlowast
lowastlowast
Abso
lute
velo
city
(120583m
sec
)
S135F-MNs
Absolute velocity
P182L-MNs
Tuba
statin
120583M
A_5
Tuba
statin
120583M
A_5
(c)
Figure 5 Continued
Stem Cells International 13
0
20
40
60
80
Mov
ing
mito
chon
dria
()
DM
SO
X4_0
5120583
M
X4_5
120583M
X4_5
120583M
X9_0
5120583
M
X9_5
120583M
lowast
lowastlowast
lowastlowast
lowastlowastlowastlowast
0
20
40
60
80
Mov
ing
mito
chon
dria
()
DM
SO
X4_0
5120583
M
X9_0
5120583
M
X9_5
120583M
lowastlowastlowastlowast
lowastlowastlowastlowast
lowastlowastlowastlowast
lowastlowastlowastlowast
S135F-MNs
Moving mitochondria
P182L-MNs
Tuba
statin
120583M
A_5
Tuba
statin
120583M
A_5
lowastlowast
(d)
Figure 5 HDAC6 inhibitors reversed axonal defect of mitochondrial transport by increasing acetylation of 120572-tubulin (a and b) Westernblot assay showed that treatment of HDAC6 inhibitors increased acetylated 120572-tubulin levels in S135F-MNs and P182L-MNs (paired 119905-testslowast119875 lt 005 and lowastlowast119875 lt 001) (c) HDAC6 inhibitors increased the absolute velocity of mitochondrial movements in S135F-MNs and P182L-MNs (WA09-MNs119873 = 617 S135F-MNs119873 = 101 and P182L-MNs119873 = 394) (d) HDAC6 inhibitors increased the percentage of movingmitochondria in S135F-MNs and P182L-MNs (WA09-MNs 119873 = 15 S135F-MNs 119873 = 7 and P182L-MNs 119873 = 15) Unpaired 119905-testslowast119875 lt 005 lowastlowast119875 lt 001 and lowastlowastlowastlowast119875 lt 00001
Authorsrsquo Contributions
Ji-Yon Kim was responsible for the iPSC experiments collec-tion and interpretation of the data and preparation of thepaper So-Youn Woo was interpreting the data and editingthe manuscript Inhee Mook-Jung provided microchannelplates and designed the experiment Heesun Choi JisooKim and Hyunjung Choi carried out microfluidic cultureand kymograph analysis Nina Ha Jangbeen Kyung andSoo Kyung Koo designed the experiments and preparationof the manuscript Young-Bin Hong and Sung-Chul Jungwere responsible for conception design of experiments andediting the manuscript Byung-Ok Choi was responsible forthe collection of patient samples and interpretation of thedata
Acknowledgments
This study was supported by the Korean Health TechnologyRampD Project Ministry of Health amp Welfare (HI12C0135HI14C3484 and HI15C1560) and a National Research Foun-dation of Korea grant funded by the Ministry of Science ICTandFuture Planning (NRF-2014R1A2A2A01004240)Humanstem cell line hFSiPS1 was provided by National Stem CellBank of Korea (Korea National Institute of Health Grant no4861-307-210-13)
References
[1] Inherited Peripheral Neuropathies Mutation Database httpwwwmolgenuaacbecmtmutationsmutationsMutByGenecfm
[2] O V Evgrafov IMersiyanova J Irobi et al ldquoMutant small heat-shock protein 27 causes axonal Charcot-Marie-Tooth diseaseand distal hereditary motor neuropathyrdquo Nature Genetics vol36 no 6 pp 602ndash606 2004
[3] J Zhai H Lin J-P Julien andWW Schlaepfer ldquoDisruption ofneurofilament network with aggregation of light neurofilamentprotein a common pathway leading to motor neuron degener-ation due to Charcot-Marie-Tooth disease-linked mutations inNFL andHSPB1rdquoHumanMolecular Genetics vol 16 no 24 pp3103ndash3116 2007
[4] L Almeida-Souza B Asselbergh C drsquoYdewalle et al ldquoSmallheat-shock protein HSPB1 mutants stabilize microtubules inCharcot-Marie-Tooth neuropathyrdquoThe Journal of Neurosciencevol 31 no 43 pp 15320ndash15328 2011
[5] A Holmgren D Bouhy V De Winter et al ldquoCharcot-Marie-Tooth causing HSPB1mutations increase Cdk5-mediated phos-phorylation of neurofilamentsrdquoActa Neuropathologica vol 126no 1 pp 93ndash108 2013
[6] A-P Arrigo ldquoThe cellular ldquonetworkingrdquo of mammalian Hsp27and its functions in the control of protein folding redox stateand apoptosisrdquoAdvances in eXperimental Medicine and Biologyvol 594 pp 14ndash26 2007
14 Stem Cells International
[7] C DrsquoYdewalle J Krishnan D M Chiheb et al ldquoHDAC6inhibitors reverse axonal loss in a mouse model of mutantHSPB1-induced Charcot-Marie-Tooth diseaserdquo NatureMedicine vol 17 no 8 pp 968ndash974 2011
[8] Y Zhang N Li C Caron et al ldquoHDAC-6 interacts with anddeacetylates tubulin and microtubules in vivordquo EMBO Journalvol 22 no 5 pp 1168ndash1179 2003
[9] A Valenzuela-Fernandez J R Cabrero J M Serrador and FSanchez-Madrid ldquoHDAC6 a key regulator of cytoskeleton cellmigration and cell-cell interactionsrdquo Trends in Cell Biology vol18 no 6 pp 291ndash297 2008
[10] V K Godena N Brookes-Hocking AMoller et al ldquoIncreasingmicrotubule acetylation rescues axonal transport and locomo-tor deficits caused by LRRK2 Roc-COR domain mutationsrdquoNature Communications vol 5 article 5245 2014
[11] J P Dompierre J D Godin B C Charrin et al ldquoHistonedeacetylase 6 inhibition compensates for the transport deficitin Huntingtonrsquos disease by increasing tubulin acetylationrdquo TheJournal of Neuroscience vol 27 no 13 pp 3571ndash3583 2007
[12] M W Amoroso G F Croft D J Williams et al ldquoAcceleratedhigh-yield generation of limb-innervating motor neurons fromhuman stem cellsrdquo Journal of Neuroscience vol 33 no 2 pp574ndash586 2013
[13] B J Gentil and L Cooper ldquoMolecular basis of axonal dysfunc-tion and traffic impairments in CMTrdquo Brain Research Bulletinvol 88 no 5 pp 444ndash453 2012
[14] Z-H Sheng and Q Cai ldquoMitochondrial transport in neu-rons impact on synaptic homeostasis and neurodegenerationrdquoNature Reviews Neuroscience vol 13 no 2 pp 77ndash93 2012
[15] J W Park B Vahidi A M Taylor S W Rhee and N LJeon ldquoMicrofluidic culture platform for neuroscience researchrdquoNature Protocols vol 1 no 4 pp 2128ndash2136 2006
[16] SWestermann and KWeber ldquoPost-translational modificationsregulate microtubule functionrdquo Nature Reviews Molecular CellBiology vol 4 no 12 pp 938ndash947 2003
[17] N A Reed D Cai T L Blasius et al ldquoMicrotubule acetylationpromotes kinesin-1 binding and transportrdquoCurrent Biology vol16 no 21 pp 2166ndash2172 2006
[18] C drsquoYdewalle E Bogaert and L Van Den Bosch ldquoHDAC6at the intersection of neuroprotection and neurodegenerationrdquoTraffic vol 13 no 6 pp 771ndash779 2012
[19] C Simoes-Pires V Zwick A Nurisso E Schenker P-ACarrupt and M Cuendet ldquoHDAC6 as a target for neurode-generative diseases what makes it different from the otherHDACsrdquoMolecular Neurodegeneration vol 8 article 7 2013
[20] Y Rui P Tiwari Z Xie and J Q Zheng ldquoAcute impairment ofmitochondrial trafficking by 120573-amyloid peptides in hippocam-pal neuronsrdquo Journal of Neuroscience vol 26 no 41 pp 10480ndash10487 2006
[21] K A Vossel K Zhang J Brodbeck et al ldquoTau reductionprevents A120573-induced defects in axonal transportrdquo Science vol330 no 6001 p 198 2010
[22] K J De vos A L Chapman M E Tennant et al ldquoFamilialamyotrophic lateral sclerosis-linked SOD1 mutants perturbfast axonal transport to reduce axonal mitochondria contentrdquoHumanMolecular Genetics vol 16 no 22 pp 2720ndash2728 2007
[23] E Trushina R B Dyer J D Badger II et al ldquoMutant huntingtinimpairs axonal trafficking inmammalian neurons in vivo and invitrordquo Molecular and Cellular Biology vol 24 no 18 pp 8195ndash8209 2004
[24] A L Orr S Li C-E Wang et al ldquoN-terminal mutant hunt-ingtin associates with mitochondria and impairs mitochondrialtraffickingrdquo Journal of Neuroscience vol 28 no 11 pp 2783ndash2792 2008
Submit your manuscripts athttpwwwhindawicom
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Anatomy Research International
PeptidesInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporation httpwwwhindawicom
International Journal of
Volume 2014
Zoology
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Molecular Biology International
GenomicsInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioinformaticsAdvances in
Marine BiologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Signal TransductionJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
Evolutionary BiologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Biochemistry Research International
ArchaeaHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Genetics Research International
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Advances in
Virolog y
Hindawi Publishing Corporationhttpwwwhindawicom
Nucleic AcidsJournal of
Volume 2014
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Enzyme Research
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
International Journal of
Microbiology
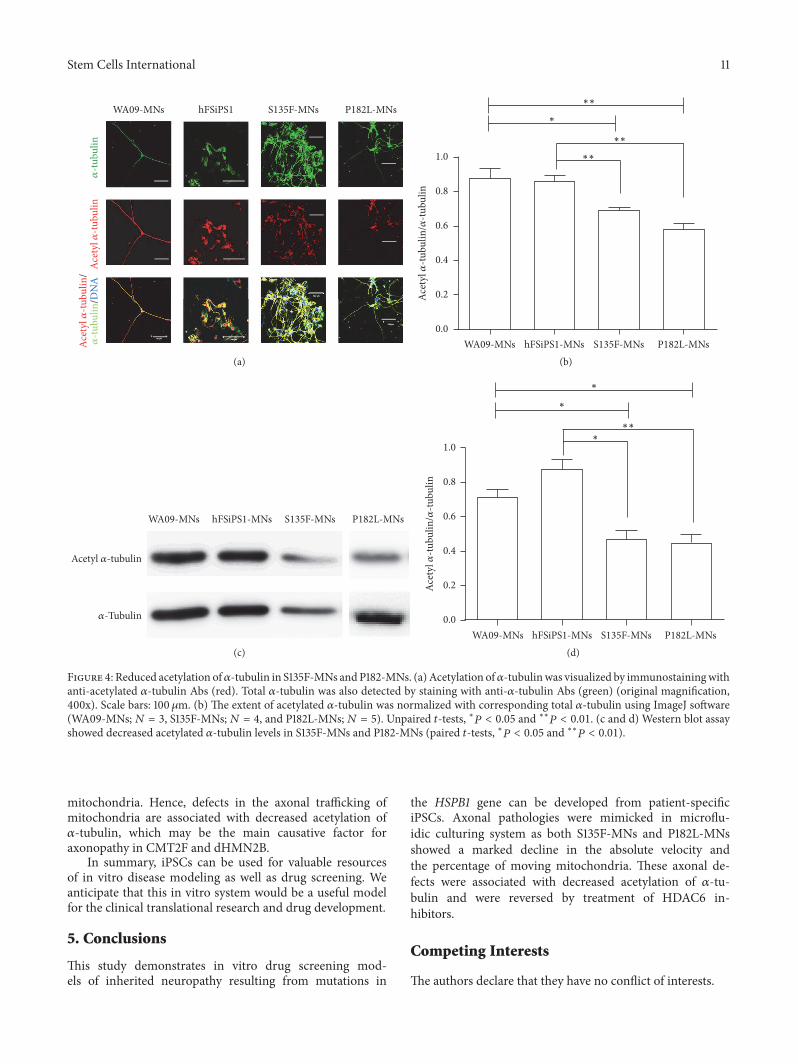
Stem Cells International 11
WA09-MNs hFSiPS1 S135F-MNs P182L-MNs120572
-tubu
linAc
etyl
120572-tu
bulin
Acet
yl120572
-tubu
lin
120572-tu
bulin
DN
A
(a)
00
02
04
06
08
10
WA09-MNs hFSiPS1-MNs S135F-MNs P182L-MNs
Acet
yl120572
-tubu
lin120572
-tubu
lin
lowastlowast
lowast
lowastlowast
lowastlowast
(b)
WA09-MNs hFSiPS1-MNs S135F-MNs P182L-MNs
120572-Tubulin
Acetyl 120572-tubulin
(c)
00
02
04
06
08
10
WA09-MNs hFSiPS1-MNs S135F-MNs P182L-MNs
Acet
yl120572
-tubu
lin120572
-tubu
linlowast
lowast
lowast
lowastlowast
(d)
Figure 4 Reduced acetylation of120572-tubulin in S135F-MNs andP182-MNs (a) Acetylation of120572-tubulinwas visualized by immunostainingwithanti-acetylated 120572-tubulin Abs (red) Total 120572-tubulin was also detected by staining with anti-120572-tubulin Abs (green) (original magnification400x) Scale bars 100120583m (b) The extent of acetylated 120572-tubulin was normalized with corresponding total 120572-tubulin using ImageJ software(WA09-MNs119873 = 3 S135F-MNs119873 = 4 and P182L-MNs119873 = 5) Unpaired 119905-tests lowast119875 lt 005 and lowastlowast119875 lt 001 (c and d) Western blot assayshowed decreased acetylated 120572-tubulin levels in S135F-MNs and P182-MNs (paired 119905-tests lowast119875 lt 005 and lowastlowast119875 lt 001)
mitochondria Hence defects in the axonal trafficking ofmitochondria are associated with decreased acetylation of120572-tubulin which may be the main causative factor foraxonopathy in CMT2F and dHMN2B
In summary iPSCs can be used for valuable resourcesof in vitro disease modeling as well as drug screening Weanticipate that this in vitro system would be a useful modelfor the clinical translational research and drug development
5 Conclusions
This study demonstrates in vitro drug screening mod-els of inherited neuropathy resulting from mutations in
the HSPB1 gene can be developed from patient-specificiPSCs Axonal pathologies were mimicked in microflu-idic culturing system as both S135F-MNs and P182L-MNsshowed a marked decline in the absolute velocity andthe percentage of moving mitochondria These axonal de-fects were associated with decreased acetylation of 120572-tu-bulin and were reversed by treatment of HDAC6 in-hibitors
Competing Interests
The authors declare that they have no conflict of interests
12 Stem Cells International
HDAC6 inhibitors
S135
F-M
Ns
P182
L-M
Ns
Acetyl 120572-tubulin
Acetyl 120572-tubulin
120572-Tubulin
120572-Tubulin
DM
SO
X4_0
5120583
M
X4_5
120583M
X9_0
5120583
M
X9_5
120583M
Tuba
statin
120583M
A_5
(a)
00
05
10
15
20
25
DM
SO
X4_0
5120583
M
X4_5
120583M
X9_0
5120583
M
X9_5
120583M
lowast
lowastlowast
lowastlowast
Acet
yl120572
-tubu
lin120572
-tubu
lin
Acetyl 120572-tubulin in S135F-MNs
00
05
10
15
20
25
DM
SO
X4_0
5120583
M
X4_5
120583M
X9_0
5120583
M
X9_5
120583M
lowast
lowastlowast
lowastlowastAc
etyl
120572-tu
bulin
120572-tu
bulin
Acetyl 120572-tubulin in P182L-MNs
Tuba
statin
120583M
A_5
Tuba
statin
120583M
A_5
(b)
00
01
02
03
04
DM
SO
X4_0
5120583
M
X4_5
120583M
X9_0
5120583
M
X9_5
120583M
lowastlowastlowastlowast
lowastlowastlowastlowast
Abso
lute
velo
city
(120583m
sec
)
00
02
04
06
DM
SO
X4_0
5120583
M
X4_5
120583M
X9_0
5120583
M
X9_5
120583M
lowast
lowastlowastlowastlowast
lowastlowastlowastlowast
lowastlowastlowastlowast
lowastlowast
Abso
lute
velo
city
(120583m
sec
)
S135F-MNs
Absolute velocity
P182L-MNs
Tuba
statin
120583M
A_5
Tuba
statin
120583M
A_5
(c)
Figure 5 Continued
Stem Cells International 13
0
20
40
60
80
Mov
ing
mito
chon
dria
()
DM
SO
X4_0
5120583
M
X4_5
120583M
X4_5
120583M
X9_0
5120583
M
X9_5
120583M
lowast
lowastlowast
lowastlowast
lowastlowastlowastlowast
0
20
40
60
80
Mov
ing
mito
chon
dria
()
DM
SO
X4_0
5120583
M
X9_0
5120583
M
X9_5
120583M
lowastlowastlowastlowast
lowastlowastlowastlowast
lowastlowastlowastlowast
lowastlowastlowastlowast
S135F-MNs
Moving mitochondria
P182L-MNs
Tuba
statin
120583M
A_5
Tuba
statin
120583M
A_5
lowastlowast
(d)
Figure 5 HDAC6 inhibitors reversed axonal defect of mitochondrial transport by increasing acetylation of 120572-tubulin (a and b) Westernblot assay showed that treatment of HDAC6 inhibitors increased acetylated 120572-tubulin levels in S135F-MNs and P182L-MNs (paired 119905-testslowast119875 lt 005 and lowastlowast119875 lt 001) (c) HDAC6 inhibitors increased the absolute velocity of mitochondrial movements in S135F-MNs and P182L-MNs (WA09-MNs119873 = 617 S135F-MNs119873 = 101 and P182L-MNs119873 = 394) (d) HDAC6 inhibitors increased the percentage of movingmitochondria in S135F-MNs and P182L-MNs (WA09-MNs 119873 = 15 S135F-MNs 119873 = 7 and P182L-MNs 119873 = 15) Unpaired 119905-testslowast119875 lt 005 lowastlowast119875 lt 001 and lowastlowastlowastlowast119875 lt 00001
Authorsrsquo Contributions
Ji-Yon Kim was responsible for the iPSC experiments collec-tion and interpretation of the data and preparation of thepaper So-Youn Woo was interpreting the data and editingthe manuscript Inhee Mook-Jung provided microchannelplates and designed the experiment Heesun Choi JisooKim and Hyunjung Choi carried out microfluidic cultureand kymograph analysis Nina Ha Jangbeen Kyung andSoo Kyung Koo designed the experiments and preparationof the manuscript Young-Bin Hong and Sung-Chul Jungwere responsible for conception design of experiments andediting the manuscript Byung-Ok Choi was responsible forthe collection of patient samples and interpretation of thedata
Acknowledgments
This study was supported by the Korean Health TechnologyRampD Project Ministry of Health amp Welfare (HI12C0135HI14C3484 and HI15C1560) and a National Research Foun-dation of Korea grant funded by the Ministry of Science ICTandFuture Planning (NRF-2014R1A2A2A01004240)Humanstem cell line hFSiPS1 was provided by National Stem CellBank of Korea (Korea National Institute of Health Grant no4861-307-210-13)
References
[1] Inherited Peripheral Neuropathies Mutation Database httpwwwmolgenuaacbecmtmutationsmutationsMutByGenecfm
[2] O V Evgrafov IMersiyanova J Irobi et al ldquoMutant small heat-shock protein 27 causes axonal Charcot-Marie-Tooth diseaseand distal hereditary motor neuropathyrdquo Nature Genetics vol36 no 6 pp 602ndash606 2004
[3] J Zhai H Lin J-P Julien andWW Schlaepfer ldquoDisruption ofneurofilament network with aggregation of light neurofilamentprotein a common pathway leading to motor neuron degener-ation due to Charcot-Marie-Tooth disease-linked mutations inNFL andHSPB1rdquoHumanMolecular Genetics vol 16 no 24 pp3103ndash3116 2007
[4] L Almeida-Souza B Asselbergh C drsquoYdewalle et al ldquoSmallheat-shock protein HSPB1 mutants stabilize microtubules inCharcot-Marie-Tooth neuropathyrdquoThe Journal of Neurosciencevol 31 no 43 pp 15320ndash15328 2011
[5] A Holmgren D Bouhy V De Winter et al ldquoCharcot-Marie-Tooth causing HSPB1mutations increase Cdk5-mediated phos-phorylation of neurofilamentsrdquoActa Neuropathologica vol 126no 1 pp 93ndash108 2013
[6] A-P Arrigo ldquoThe cellular ldquonetworkingrdquo of mammalian Hsp27and its functions in the control of protein folding redox stateand apoptosisrdquoAdvances in eXperimental Medicine and Biologyvol 594 pp 14ndash26 2007
14 Stem Cells International
[7] C DrsquoYdewalle J Krishnan D M Chiheb et al ldquoHDAC6inhibitors reverse axonal loss in a mouse model of mutantHSPB1-induced Charcot-Marie-Tooth diseaserdquo NatureMedicine vol 17 no 8 pp 968ndash974 2011
[8] Y Zhang N Li C Caron et al ldquoHDAC-6 interacts with anddeacetylates tubulin and microtubules in vivordquo EMBO Journalvol 22 no 5 pp 1168ndash1179 2003
[9] A Valenzuela-Fernandez J R Cabrero J M Serrador and FSanchez-Madrid ldquoHDAC6 a key regulator of cytoskeleton cellmigration and cell-cell interactionsrdquo Trends in Cell Biology vol18 no 6 pp 291ndash297 2008
[10] V K Godena N Brookes-Hocking AMoller et al ldquoIncreasingmicrotubule acetylation rescues axonal transport and locomo-tor deficits caused by LRRK2 Roc-COR domain mutationsrdquoNature Communications vol 5 article 5245 2014
[11] J P Dompierre J D Godin B C Charrin et al ldquoHistonedeacetylase 6 inhibition compensates for the transport deficitin Huntingtonrsquos disease by increasing tubulin acetylationrdquo TheJournal of Neuroscience vol 27 no 13 pp 3571ndash3583 2007
[12] M W Amoroso G F Croft D J Williams et al ldquoAcceleratedhigh-yield generation of limb-innervating motor neurons fromhuman stem cellsrdquo Journal of Neuroscience vol 33 no 2 pp574ndash586 2013
[13] B J Gentil and L Cooper ldquoMolecular basis of axonal dysfunc-tion and traffic impairments in CMTrdquo Brain Research Bulletinvol 88 no 5 pp 444ndash453 2012
[14] Z-H Sheng and Q Cai ldquoMitochondrial transport in neu-rons impact on synaptic homeostasis and neurodegenerationrdquoNature Reviews Neuroscience vol 13 no 2 pp 77ndash93 2012
[15] J W Park B Vahidi A M Taylor S W Rhee and N LJeon ldquoMicrofluidic culture platform for neuroscience researchrdquoNature Protocols vol 1 no 4 pp 2128ndash2136 2006
[16] SWestermann and KWeber ldquoPost-translational modificationsregulate microtubule functionrdquo Nature Reviews Molecular CellBiology vol 4 no 12 pp 938ndash947 2003
[17] N A Reed D Cai T L Blasius et al ldquoMicrotubule acetylationpromotes kinesin-1 binding and transportrdquoCurrent Biology vol16 no 21 pp 2166ndash2172 2006
[18] C drsquoYdewalle E Bogaert and L Van Den Bosch ldquoHDAC6at the intersection of neuroprotection and neurodegenerationrdquoTraffic vol 13 no 6 pp 771ndash779 2012
[19] C Simoes-Pires V Zwick A Nurisso E Schenker P-ACarrupt and M Cuendet ldquoHDAC6 as a target for neurode-generative diseases what makes it different from the otherHDACsrdquoMolecular Neurodegeneration vol 8 article 7 2013
[20] Y Rui P Tiwari Z Xie and J Q Zheng ldquoAcute impairment ofmitochondrial trafficking by 120573-amyloid peptides in hippocam-pal neuronsrdquo Journal of Neuroscience vol 26 no 41 pp 10480ndash10487 2006
[21] K A Vossel K Zhang J Brodbeck et al ldquoTau reductionprevents A120573-induced defects in axonal transportrdquo Science vol330 no 6001 p 198 2010
[22] K J De vos A L Chapman M E Tennant et al ldquoFamilialamyotrophic lateral sclerosis-linked SOD1 mutants perturbfast axonal transport to reduce axonal mitochondria contentrdquoHumanMolecular Genetics vol 16 no 22 pp 2720ndash2728 2007
[23] E Trushina R B Dyer J D Badger II et al ldquoMutant huntingtinimpairs axonal trafficking inmammalian neurons in vivo and invitrordquo Molecular and Cellular Biology vol 24 no 18 pp 8195ndash8209 2004
[24] A L Orr S Li C-E Wang et al ldquoN-terminal mutant hunt-ingtin associates with mitochondria and impairs mitochondrialtraffickingrdquo Journal of Neuroscience vol 28 no 11 pp 2783ndash2792 2008
Submit your manuscripts athttpwwwhindawicom
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Anatomy Research International
PeptidesInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporation httpwwwhindawicom
International Journal of
Volume 2014
Zoology
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Molecular Biology International
GenomicsInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioinformaticsAdvances in
Marine BiologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Signal TransductionJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
Evolutionary BiologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Biochemistry Research International
ArchaeaHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Genetics Research International
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Advances in
Virolog y
Hindawi Publishing Corporationhttpwwwhindawicom
Nucleic AcidsJournal of
Volume 2014
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Enzyme Research
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
International Journal of
Microbiology

12 Stem Cells International
HDAC6 inhibitors
S135
F-M
Ns
P182
L-M
Ns
Acetyl 120572-tubulin
Acetyl 120572-tubulin
120572-Tubulin
120572-Tubulin
DM
SO
X4_0
5120583
M
X4_5
120583M
X9_0
5120583
M
X9_5
120583M
Tuba
statin
120583M
A_5
(a)
00
05
10
15
20
25
DM
SO
X4_0
5120583
M
X4_5
120583M
X9_0
5120583
M
X9_5
120583M
lowast
lowastlowast
lowastlowast
Acet
yl120572
-tubu
lin120572
-tubu
lin
Acetyl 120572-tubulin in S135F-MNs
00
05
10
15
20
25
DM
SO
X4_0
5120583
M
X4_5
120583M
X9_0
5120583
M
X9_5
120583M
lowast
lowastlowast
lowastlowastAc
etyl
120572-tu
bulin
120572-tu
bulin
Acetyl 120572-tubulin in P182L-MNs
Tuba
statin
120583M
A_5
Tuba
statin
120583M
A_5
(b)
00
01
02
03
04
DM
SO
X4_0
5120583
M
X4_5
120583M
X9_0
5120583
M
X9_5
120583M
lowastlowastlowastlowast
lowastlowastlowastlowast
Abso
lute
velo
city
(120583m
sec
)
00
02
04
06
DM
SO
X4_0
5120583
M
X4_5
120583M
X9_0
5120583
M
X9_5
120583M
lowast
lowastlowastlowastlowast
lowastlowastlowastlowast
lowastlowastlowastlowast
lowastlowast
Abso
lute
velo
city
(120583m
sec
)
S135F-MNs
Absolute velocity
P182L-MNs
Tuba
statin
120583M
A_5
Tuba
statin
120583M
A_5
(c)
Figure 5 Continued
Stem Cells International 13
0
20
40
60
80
Mov
ing
mito
chon
dria
()
DM
SO
X4_0
5120583
M
X4_5
120583M
X4_5
120583M
X9_0
5120583
M
X9_5
120583M
lowast
lowastlowast
lowastlowast
lowastlowastlowastlowast
0
20
40
60
80
Mov
ing
mito
chon
dria
()
DM
SO
X4_0
5120583
M
X9_0
5120583
M
X9_5
120583M
lowastlowastlowastlowast
lowastlowastlowastlowast
lowastlowastlowastlowast
lowastlowastlowastlowast
S135F-MNs
Moving mitochondria
P182L-MNs
Tuba
statin
120583M
A_5
Tuba
statin
120583M
A_5
lowastlowast
(d)
Figure 5 HDAC6 inhibitors reversed axonal defect of mitochondrial transport by increasing acetylation of 120572-tubulin (a and b) Westernblot assay showed that treatment of HDAC6 inhibitors increased acetylated 120572-tubulin levels in S135F-MNs and P182L-MNs (paired 119905-testslowast119875 lt 005 and lowastlowast119875 lt 001) (c) HDAC6 inhibitors increased the absolute velocity of mitochondrial movements in S135F-MNs and P182L-MNs (WA09-MNs119873 = 617 S135F-MNs119873 = 101 and P182L-MNs119873 = 394) (d) HDAC6 inhibitors increased the percentage of movingmitochondria in S135F-MNs and P182L-MNs (WA09-MNs 119873 = 15 S135F-MNs 119873 = 7 and P182L-MNs 119873 = 15) Unpaired 119905-testslowast119875 lt 005 lowastlowast119875 lt 001 and lowastlowastlowastlowast119875 lt 00001
Authorsrsquo Contributions
Ji-Yon Kim was responsible for the iPSC experiments collec-tion and interpretation of the data and preparation of thepaper So-Youn Woo was interpreting the data and editingthe manuscript Inhee Mook-Jung provided microchannelplates and designed the experiment Heesun Choi JisooKim and Hyunjung Choi carried out microfluidic cultureand kymograph analysis Nina Ha Jangbeen Kyung andSoo Kyung Koo designed the experiments and preparationof the manuscript Young-Bin Hong and Sung-Chul Jungwere responsible for conception design of experiments andediting the manuscript Byung-Ok Choi was responsible forthe collection of patient samples and interpretation of thedata
Acknowledgments
This study was supported by the Korean Health TechnologyRampD Project Ministry of Health amp Welfare (HI12C0135HI14C3484 and HI15C1560) and a National Research Foun-dation of Korea grant funded by the Ministry of Science ICTandFuture Planning (NRF-2014R1A2A2A01004240)Humanstem cell line hFSiPS1 was provided by National Stem CellBank of Korea (Korea National Institute of Health Grant no4861-307-210-13)
References
[1] Inherited Peripheral Neuropathies Mutation Database httpwwwmolgenuaacbecmtmutationsmutationsMutByGenecfm
[2] O V Evgrafov IMersiyanova J Irobi et al ldquoMutant small heat-shock protein 27 causes axonal Charcot-Marie-Tooth diseaseand distal hereditary motor neuropathyrdquo Nature Genetics vol36 no 6 pp 602ndash606 2004
[3] J Zhai H Lin J-P Julien andWW Schlaepfer ldquoDisruption ofneurofilament network with aggregation of light neurofilamentprotein a common pathway leading to motor neuron degener-ation due to Charcot-Marie-Tooth disease-linked mutations inNFL andHSPB1rdquoHumanMolecular Genetics vol 16 no 24 pp3103ndash3116 2007
[4] L Almeida-Souza B Asselbergh C drsquoYdewalle et al ldquoSmallheat-shock protein HSPB1 mutants stabilize microtubules inCharcot-Marie-Tooth neuropathyrdquoThe Journal of Neurosciencevol 31 no 43 pp 15320ndash15328 2011
[5] A Holmgren D Bouhy V De Winter et al ldquoCharcot-Marie-Tooth causing HSPB1mutations increase Cdk5-mediated phos-phorylation of neurofilamentsrdquoActa Neuropathologica vol 126no 1 pp 93ndash108 2013
[6] A-P Arrigo ldquoThe cellular ldquonetworkingrdquo of mammalian Hsp27and its functions in the control of protein folding redox stateand apoptosisrdquoAdvances in eXperimental Medicine and Biologyvol 594 pp 14ndash26 2007
14 Stem Cells International
[7] C DrsquoYdewalle J Krishnan D M Chiheb et al ldquoHDAC6inhibitors reverse axonal loss in a mouse model of mutantHSPB1-induced Charcot-Marie-Tooth diseaserdquo NatureMedicine vol 17 no 8 pp 968ndash974 2011
[8] Y Zhang N Li C Caron et al ldquoHDAC-6 interacts with anddeacetylates tubulin and microtubules in vivordquo EMBO Journalvol 22 no 5 pp 1168ndash1179 2003
[9] A Valenzuela-Fernandez J R Cabrero J M Serrador and FSanchez-Madrid ldquoHDAC6 a key regulator of cytoskeleton cellmigration and cell-cell interactionsrdquo Trends in Cell Biology vol18 no 6 pp 291ndash297 2008
[10] V K Godena N Brookes-Hocking AMoller et al ldquoIncreasingmicrotubule acetylation rescues axonal transport and locomo-tor deficits caused by LRRK2 Roc-COR domain mutationsrdquoNature Communications vol 5 article 5245 2014
[11] J P Dompierre J D Godin B C Charrin et al ldquoHistonedeacetylase 6 inhibition compensates for the transport deficitin Huntingtonrsquos disease by increasing tubulin acetylationrdquo TheJournal of Neuroscience vol 27 no 13 pp 3571ndash3583 2007
[12] M W Amoroso G F Croft D J Williams et al ldquoAcceleratedhigh-yield generation of limb-innervating motor neurons fromhuman stem cellsrdquo Journal of Neuroscience vol 33 no 2 pp574ndash586 2013
[13] B J Gentil and L Cooper ldquoMolecular basis of axonal dysfunc-tion and traffic impairments in CMTrdquo Brain Research Bulletinvol 88 no 5 pp 444ndash453 2012
[14] Z-H Sheng and Q Cai ldquoMitochondrial transport in neu-rons impact on synaptic homeostasis and neurodegenerationrdquoNature Reviews Neuroscience vol 13 no 2 pp 77ndash93 2012
[15] J W Park B Vahidi A M Taylor S W Rhee and N LJeon ldquoMicrofluidic culture platform for neuroscience researchrdquoNature Protocols vol 1 no 4 pp 2128ndash2136 2006
[16] SWestermann and KWeber ldquoPost-translational modificationsregulate microtubule functionrdquo Nature Reviews Molecular CellBiology vol 4 no 12 pp 938ndash947 2003
[17] N A Reed D Cai T L Blasius et al ldquoMicrotubule acetylationpromotes kinesin-1 binding and transportrdquoCurrent Biology vol16 no 21 pp 2166ndash2172 2006
[18] C drsquoYdewalle E Bogaert and L Van Den Bosch ldquoHDAC6at the intersection of neuroprotection and neurodegenerationrdquoTraffic vol 13 no 6 pp 771ndash779 2012
[19] C Simoes-Pires V Zwick A Nurisso E Schenker P-ACarrupt and M Cuendet ldquoHDAC6 as a target for neurode-generative diseases what makes it different from the otherHDACsrdquoMolecular Neurodegeneration vol 8 article 7 2013
[20] Y Rui P Tiwari Z Xie and J Q Zheng ldquoAcute impairment ofmitochondrial trafficking by 120573-amyloid peptides in hippocam-pal neuronsrdquo Journal of Neuroscience vol 26 no 41 pp 10480ndash10487 2006
[21] K A Vossel K Zhang J Brodbeck et al ldquoTau reductionprevents A120573-induced defects in axonal transportrdquo Science vol330 no 6001 p 198 2010
[22] K J De vos A L Chapman M E Tennant et al ldquoFamilialamyotrophic lateral sclerosis-linked SOD1 mutants perturbfast axonal transport to reduce axonal mitochondria contentrdquoHumanMolecular Genetics vol 16 no 22 pp 2720ndash2728 2007
[23] E Trushina R B Dyer J D Badger II et al ldquoMutant huntingtinimpairs axonal trafficking inmammalian neurons in vivo and invitrordquo Molecular and Cellular Biology vol 24 no 18 pp 8195ndash8209 2004
[24] A L Orr S Li C-E Wang et al ldquoN-terminal mutant hunt-ingtin associates with mitochondria and impairs mitochondrialtraffickingrdquo Journal of Neuroscience vol 28 no 11 pp 2783ndash2792 2008
Submit your manuscripts athttpwwwhindawicom
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Anatomy Research International
PeptidesInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporation httpwwwhindawicom
International Journal of
Volume 2014
Zoology
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Molecular Biology International
GenomicsInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioinformaticsAdvances in
Marine BiologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Signal TransductionJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
Evolutionary BiologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Biochemistry Research International
ArchaeaHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Genetics Research International
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Advances in
Virolog y
Hindawi Publishing Corporationhttpwwwhindawicom
Nucleic AcidsJournal of
Volume 2014
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Enzyme Research
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
International Journal of
Microbiology
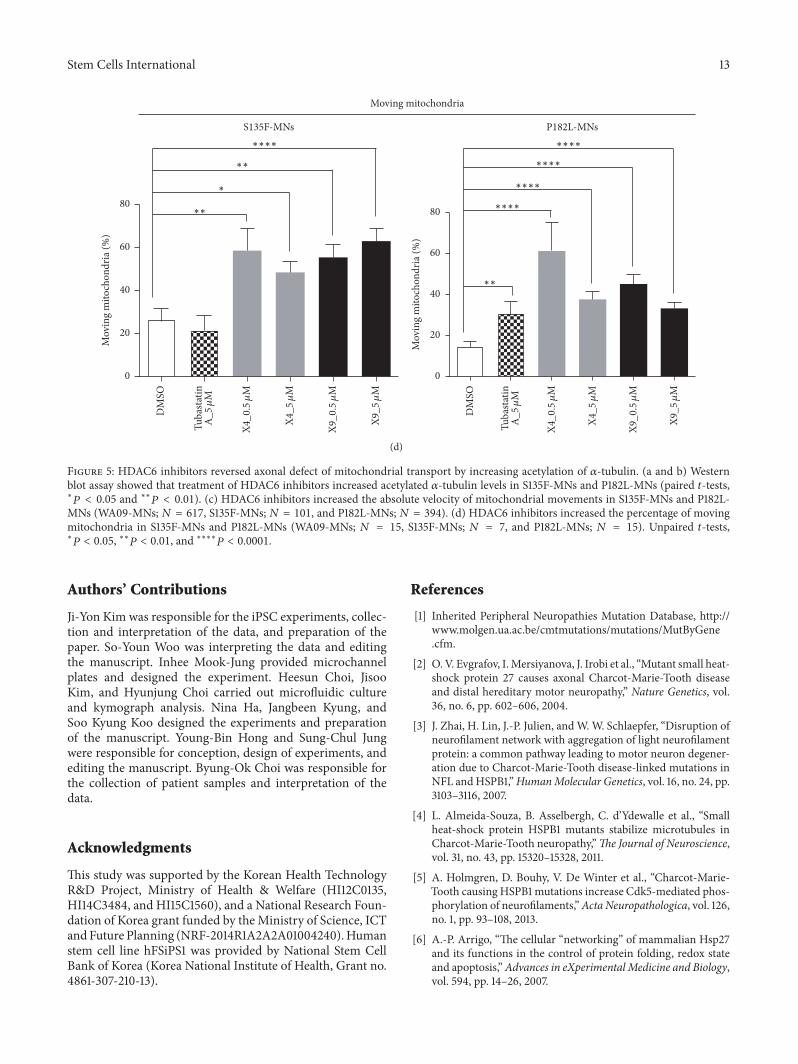
Stem Cells International 13
0
20
40
60
80
Mov
ing
mito
chon
dria
()
DM
SO
X4_0
5120583
M
X4_5
120583M
X4_5
120583M
X9_0
5120583
M
X9_5
120583M
lowast
lowastlowast
lowastlowast
lowastlowastlowastlowast
0
20
40
60
80
Mov
ing
mito
chon
dria
()
DM
SO
X4_0
5120583
M
X9_0
5120583
M
X9_5
120583M
lowastlowastlowastlowast
lowastlowastlowastlowast
lowastlowastlowastlowast
lowastlowastlowastlowast
S135F-MNs
Moving mitochondria
P182L-MNs
Tuba
statin
120583M
A_5
Tuba
statin
120583M
A_5
lowastlowast
(d)
Figure 5 HDAC6 inhibitors reversed axonal defect of mitochondrial transport by increasing acetylation of 120572-tubulin (a and b) Westernblot assay showed that treatment of HDAC6 inhibitors increased acetylated 120572-tubulin levels in S135F-MNs and P182L-MNs (paired 119905-testslowast119875 lt 005 and lowastlowast119875 lt 001) (c) HDAC6 inhibitors increased the absolute velocity of mitochondrial movements in S135F-MNs and P182L-MNs (WA09-MNs119873 = 617 S135F-MNs119873 = 101 and P182L-MNs119873 = 394) (d) HDAC6 inhibitors increased the percentage of movingmitochondria in S135F-MNs and P182L-MNs (WA09-MNs 119873 = 15 S135F-MNs 119873 = 7 and P182L-MNs 119873 = 15) Unpaired 119905-testslowast119875 lt 005 lowastlowast119875 lt 001 and lowastlowastlowastlowast119875 lt 00001
Authorsrsquo Contributions
Ji-Yon Kim was responsible for the iPSC experiments collec-tion and interpretation of the data and preparation of thepaper So-Youn Woo was interpreting the data and editingthe manuscript Inhee Mook-Jung provided microchannelplates and designed the experiment Heesun Choi JisooKim and Hyunjung Choi carried out microfluidic cultureand kymograph analysis Nina Ha Jangbeen Kyung andSoo Kyung Koo designed the experiments and preparationof the manuscript Young-Bin Hong and Sung-Chul Jungwere responsible for conception design of experiments andediting the manuscript Byung-Ok Choi was responsible forthe collection of patient samples and interpretation of thedata
Acknowledgments
This study was supported by the Korean Health TechnologyRampD Project Ministry of Health amp Welfare (HI12C0135HI14C3484 and HI15C1560) and a National Research Foun-dation of Korea grant funded by the Ministry of Science ICTandFuture Planning (NRF-2014R1A2A2A01004240)Humanstem cell line hFSiPS1 was provided by National Stem CellBank of Korea (Korea National Institute of Health Grant no4861-307-210-13)
References
[1] Inherited Peripheral Neuropathies Mutation Database httpwwwmolgenuaacbecmtmutationsmutationsMutByGenecfm
[2] O V Evgrafov IMersiyanova J Irobi et al ldquoMutant small heat-shock protein 27 causes axonal Charcot-Marie-Tooth diseaseand distal hereditary motor neuropathyrdquo Nature Genetics vol36 no 6 pp 602ndash606 2004
[3] J Zhai H Lin J-P Julien andWW Schlaepfer ldquoDisruption ofneurofilament network with aggregation of light neurofilamentprotein a common pathway leading to motor neuron degener-ation due to Charcot-Marie-Tooth disease-linked mutations inNFL andHSPB1rdquoHumanMolecular Genetics vol 16 no 24 pp3103ndash3116 2007
[4] L Almeida-Souza B Asselbergh C drsquoYdewalle et al ldquoSmallheat-shock protein HSPB1 mutants stabilize microtubules inCharcot-Marie-Tooth neuropathyrdquoThe Journal of Neurosciencevol 31 no 43 pp 15320ndash15328 2011
[5] A Holmgren D Bouhy V De Winter et al ldquoCharcot-Marie-Tooth causing HSPB1mutations increase Cdk5-mediated phos-phorylation of neurofilamentsrdquoActa Neuropathologica vol 126no 1 pp 93ndash108 2013
[6] A-P Arrigo ldquoThe cellular ldquonetworkingrdquo of mammalian Hsp27and its functions in the control of protein folding redox stateand apoptosisrdquoAdvances in eXperimental Medicine and Biologyvol 594 pp 14ndash26 2007
14 Stem Cells International
[7] C DrsquoYdewalle J Krishnan D M Chiheb et al ldquoHDAC6inhibitors reverse axonal loss in a mouse model of mutantHSPB1-induced Charcot-Marie-Tooth diseaserdquo NatureMedicine vol 17 no 8 pp 968ndash974 2011
[8] Y Zhang N Li C Caron et al ldquoHDAC-6 interacts with anddeacetylates tubulin and microtubules in vivordquo EMBO Journalvol 22 no 5 pp 1168ndash1179 2003
[9] A Valenzuela-Fernandez J R Cabrero J M Serrador and FSanchez-Madrid ldquoHDAC6 a key regulator of cytoskeleton cellmigration and cell-cell interactionsrdquo Trends in Cell Biology vol18 no 6 pp 291ndash297 2008
[10] V K Godena N Brookes-Hocking AMoller et al ldquoIncreasingmicrotubule acetylation rescues axonal transport and locomo-tor deficits caused by LRRK2 Roc-COR domain mutationsrdquoNature Communications vol 5 article 5245 2014
[11] J P Dompierre J D Godin B C Charrin et al ldquoHistonedeacetylase 6 inhibition compensates for the transport deficitin Huntingtonrsquos disease by increasing tubulin acetylationrdquo TheJournal of Neuroscience vol 27 no 13 pp 3571ndash3583 2007
[12] M W Amoroso G F Croft D J Williams et al ldquoAcceleratedhigh-yield generation of limb-innervating motor neurons fromhuman stem cellsrdquo Journal of Neuroscience vol 33 no 2 pp574ndash586 2013
[13] B J Gentil and L Cooper ldquoMolecular basis of axonal dysfunc-tion and traffic impairments in CMTrdquo Brain Research Bulletinvol 88 no 5 pp 444ndash453 2012
[14] Z-H Sheng and Q Cai ldquoMitochondrial transport in neu-rons impact on synaptic homeostasis and neurodegenerationrdquoNature Reviews Neuroscience vol 13 no 2 pp 77ndash93 2012
[15] J W Park B Vahidi A M Taylor S W Rhee and N LJeon ldquoMicrofluidic culture platform for neuroscience researchrdquoNature Protocols vol 1 no 4 pp 2128ndash2136 2006
[16] SWestermann and KWeber ldquoPost-translational modificationsregulate microtubule functionrdquo Nature Reviews Molecular CellBiology vol 4 no 12 pp 938ndash947 2003
[17] N A Reed D Cai T L Blasius et al ldquoMicrotubule acetylationpromotes kinesin-1 binding and transportrdquoCurrent Biology vol16 no 21 pp 2166ndash2172 2006
[18] C drsquoYdewalle E Bogaert and L Van Den Bosch ldquoHDAC6at the intersection of neuroprotection and neurodegenerationrdquoTraffic vol 13 no 6 pp 771ndash779 2012
[19] C Simoes-Pires V Zwick A Nurisso E Schenker P-ACarrupt and M Cuendet ldquoHDAC6 as a target for neurode-generative diseases what makes it different from the otherHDACsrdquoMolecular Neurodegeneration vol 8 article 7 2013
[20] Y Rui P Tiwari Z Xie and J Q Zheng ldquoAcute impairment ofmitochondrial trafficking by 120573-amyloid peptides in hippocam-pal neuronsrdquo Journal of Neuroscience vol 26 no 41 pp 10480ndash10487 2006
[21] K A Vossel K Zhang J Brodbeck et al ldquoTau reductionprevents A120573-induced defects in axonal transportrdquo Science vol330 no 6001 p 198 2010
[22] K J De vos A L Chapman M E Tennant et al ldquoFamilialamyotrophic lateral sclerosis-linked SOD1 mutants perturbfast axonal transport to reduce axonal mitochondria contentrdquoHumanMolecular Genetics vol 16 no 22 pp 2720ndash2728 2007
[23] E Trushina R B Dyer J D Badger II et al ldquoMutant huntingtinimpairs axonal trafficking inmammalian neurons in vivo and invitrordquo Molecular and Cellular Biology vol 24 no 18 pp 8195ndash8209 2004
[24] A L Orr S Li C-E Wang et al ldquoN-terminal mutant hunt-ingtin associates with mitochondria and impairs mitochondrialtraffickingrdquo Journal of Neuroscience vol 28 no 11 pp 2783ndash2792 2008
Submit your manuscripts athttpwwwhindawicom
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Anatomy Research International
PeptidesInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporation httpwwwhindawicom
International Journal of
Volume 2014
Zoology
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Molecular Biology International
GenomicsInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioinformaticsAdvances in
Marine BiologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Signal TransductionJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
Evolutionary BiologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Biochemistry Research International
ArchaeaHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Genetics Research International
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Advances in
Virolog y
Hindawi Publishing Corporationhttpwwwhindawicom
Nucleic AcidsJournal of
Volume 2014
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Enzyme Research
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
International Journal of
Microbiology

14 Stem Cells International
[7] C DrsquoYdewalle J Krishnan D M Chiheb et al ldquoHDAC6inhibitors reverse axonal loss in a mouse model of mutantHSPB1-induced Charcot-Marie-Tooth diseaserdquo NatureMedicine vol 17 no 8 pp 968ndash974 2011
[8] Y Zhang N Li C Caron et al ldquoHDAC-6 interacts with anddeacetylates tubulin and microtubules in vivordquo EMBO Journalvol 22 no 5 pp 1168ndash1179 2003
[9] A Valenzuela-Fernandez J R Cabrero J M Serrador and FSanchez-Madrid ldquoHDAC6 a key regulator of cytoskeleton cellmigration and cell-cell interactionsrdquo Trends in Cell Biology vol18 no 6 pp 291ndash297 2008
[10] V K Godena N Brookes-Hocking AMoller et al ldquoIncreasingmicrotubule acetylation rescues axonal transport and locomo-tor deficits caused by LRRK2 Roc-COR domain mutationsrdquoNature Communications vol 5 article 5245 2014
[11] J P Dompierre J D Godin B C Charrin et al ldquoHistonedeacetylase 6 inhibition compensates for the transport deficitin Huntingtonrsquos disease by increasing tubulin acetylationrdquo TheJournal of Neuroscience vol 27 no 13 pp 3571ndash3583 2007
[12] M W Amoroso G F Croft D J Williams et al ldquoAcceleratedhigh-yield generation of limb-innervating motor neurons fromhuman stem cellsrdquo Journal of Neuroscience vol 33 no 2 pp574ndash586 2013
[13] B J Gentil and L Cooper ldquoMolecular basis of axonal dysfunc-tion and traffic impairments in CMTrdquo Brain Research Bulletinvol 88 no 5 pp 444ndash453 2012
[14] Z-H Sheng and Q Cai ldquoMitochondrial transport in neu-rons impact on synaptic homeostasis and neurodegenerationrdquoNature Reviews Neuroscience vol 13 no 2 pp 77ndash93 2012
[15] J W Park B Vahidi A M Taylor S W Rhee and N LJeon ldquoMicrofluidic culture platform for neuroscience researchrdquoNature Protocols vol 1 no 4 pp 2128ndash2136 2006
[16] SWestermann and KWeber ldquoPost-translational modificationsregulate microtubule functionrdquo Nature Reviews Molecular CellBiology vol 4 no 12 pp 938ndash947 2003
[17] N A Reed D Cai T L Blasius et al ldquoMicrotubule acetylationpromotes kinesin-1 binding and transportrdquoCurrent Biology vol16 no 21 pp 2166ndash2172 2006
[18] C drsquoYdewalle E Bogaert and L Van Den Bosch ldquoHDAC6at the intersection of neuroprotection and neurodegenerationrdquoTraffic vol 13 no 6 pp 771ndash779 2012
[19] C Simoes-Pires V Zwick A Nurisso E Schenker P-ACarrupt and M Cuendet ldquoHDAC6 as a target for neurode-generative diseases what makes it different from the otherHDACsrdquoMolecular Neurodegeneration vol 8 article 7 2013
[20] Y Rui P Tiwari Z Xie and J Q Zheng ldquoAcute impairment ofmitochondrial trafficking by 120573-amyloid peptides in hippocam-pal neuronsrdquo Journal of Neuroscience vol 26 no 41 pp 10480ndash10487 2006
[21] K A Vossel K Zhang J Brodbeck et al ldquoTau reductionprevents A120573-induced defects in axonal transportrdquo Science vol330 no 6001 p 198 2010
[22] K J De vos A L Chapman M E Tennant et al ldquoFamilialamyotrophic lateral sclerosis-linked SOD1 mutants perturbfast axonal transport to reduce axonal mitochondria contentrdquoHumanMolecular Genetics vol 16 no 22 pp 2720ndash2728 2007
[23] E Trushina R B Dyer J D Badger II et al ldquoMutant huntingtinimpairs axonal trafficking inmammalian neurons in vivo and invitrordquo Molecular and Cellular Biology vol 24 no 18 pp 8195ndash8209 2004
[24] A L Orr S Li C-E Wang et al ldquoN-terminal mutant hunt-ingtin associates with mitochondria and impairs mitochondrialtraffickingrdquo Journal of Neuroscience vol 28 no 11 pp 2783ndash2792 2008
Submit your manuscripts athttpwwwhindawicom
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Anatomy Research International
PeptidesInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporation httpwwwhindawicom
International Journal of
Volume 2014
Zoology
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Molecular Biology International
GenomicsInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioinformaticsAdvances in
Marine BiologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Signal TransductionJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
Evolutionary BiologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Biochemistry Research International
ArchaeaHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Genetics Research International
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Advances in
Virolog y
Hindawi Publishing Corporationhttpwwwhindawicom
Nucleic AcidsJournal of
Volume 2014
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Enzyme Research
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
International Journal of
Microbiology

Submit your manuscripts athttpwwwhindawicom
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Anatomy Research International
PeptidesInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporation httpwwwhindawicom
International Journal of
Volume 2014
Zoology
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Molecular Biology International
GenomicsInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioinformaticsAdvances in
Marine BiologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Signal TransductionJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
Evolutionary BiologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Biochemistry Research International
ArchaeaHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Genetics Research International
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Advances in
Virolog y
Hindawi Publishing Corporationhttpwwwhindawicom
Nucleic AcidsJournal of
Volume 2014
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Enzyme Research
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
International Journal of
Microbiology