juvenile hormone biosynthesis in the cockroach, …...the jh biosynthetic pathway in ca of day 6...
TRANSCRIPT

Juvenile hormone biosynthesis in the cockroach, Diploptera punctata: the characterization of the
biosynthetic pathway and the regulatory roles of allatostatins and NMDA receptor
by
Juan Huang
A thesis submitted in conformity with the requirements for the degree of Doctor of Philosophy
Department of Cell and Systems Biology University of Toronto
© Copyright by Juan Huang (2015)

ii
Juvenile hormone biosynthesis in the cockroach, Diploptera
punctata: the characterization of the biosynthetic pathway and the
regulatory roles of allatostatins and NMDA receptor
Juan Huang
Doctor of Philosophy (2015), Department of Cell and Systems Biology, University of Toronto
Abstract
The juvenile hormones (JH) play essential roles in regulating growth, development,
metamorphosis, ageing, caste differentiation and reproduction in insects. Diploptera punctata,
the only truly viviparous cockroach is a well-known model system in the study of JH
biosynthesis and its regulation. The physiology of this animal is characterized by very stable and
high rates of JH biosynthesis and precise and predictable reproductive events that correlate well
with rates of JH production. Many studies have been performed on D. punctata to determine the
function of JH. However, the pathway of JH biosynthesis has not been identified. In addition,
although many factors are known to regulate JH biosynthesis, the exact mechanisms remain
unclear. The aim of my research was to elucidate the JH biosynthetic pathway in D. punctata and
study the mechanisms by which allatostatin (AST) and N-methyl-D-aspartate (NMDA) receptor
regulate JH production. I have (1) identified genes in the JH biosynthetic pathway, and
determined their roles in JH biosynthesis; (2) investigated the mode of action of AST by
determining the signaling pathway of AstR and the target of AST action; (3) determined the role
of the NMDA receptor in JH biosynthesis using RNA interference and treatment with an NMDA
receptor antagonist. To validate the application value of my research, AST analogs with high JH
inhibitory activity were designed and their activities on JH biosynthesis were measured by in
vitro and in vivo bioassays.

iii
Acknowledgements
It has been four and a half years since I began my studies at the University of Toronto. I still
remember the first day in Toronto. I was excited and nervous. Starting from the first day I went
to the lab, I was surrounded by friendly faces and help. My supervisor Stephen S. Tobe and his
wife Martha Tobe helped me to get used to the culture on another continent. Jinrui Zhang helped
me to find a place to live. And all the documents were handled in one day with the help of
Ekaterina F. Hult and Jane Linley. They successfully took away all my nervousness and made
me feel that my life in Toronto would be exciting.
Before I came to Canada, I barely spoke English. Language has been troublesome for me. I was
worried that no one would like to talk to me because of my poor English. But again, people in
my lab, Ekaterina F. Hult, Jinrui Zhang, Koichiro J. Yagi, Shirley H. Tiu, Elisabeth Marchal and
Ilke van Hazel helped me to get over my trouble. They have been super patient, and always
encouraged me to speak. My English was greatly improved with their help.
In regard to research, there are many people to thank. First, my supervisor Stephen S. Tobe; he
has an interesting way to train his students. To start my project, Steve asked me to read papers
and find a project I am interested in, instead of assigning me one. It was difficult in the
beginning, but when I look back now, I find it was great training. Now I am able to start and
complete a project independently, thanks to him. In addition, Steve always provided great
suggestions for my projects, and he always encouraged me to try new things. He taught me:
never to be afraid of failure, because that is part of PhD training. I would never have finished my
PhD without his support and supervision.

iv
And I would also like to thank Ekaterina F. Hult and Elisabeth Marchal for their great help
during my PhD study. I was a chemist before I came to Toronto. I knew very little biology.
Ekaterina F. Hult not only helped me to start my project, but also taught me many techniques in
biology. Most importantly, she has an ability to make me feel good about myself. She
encouraged me many times when I was frustrated with my failed experiments. Elisabeth Marchal
is the most kind and sweet person I have ever met. She is always very thoughtful and nice, and
she always came up with great ideas. We were a great team and worked on several projects
together. I learned a lot from her, not only her knowledge in biology, but also her attitude to
research and life.
I would also like to acknowledge my other collaborators who contributed to my research
projects: Jinrui Zhang, who taught me Radiochemical assay, and cockroach dissection; Koichiro
J. Yagi, who helped me set up HPLC and gave suggestions for my projects; Prof Barbara Stay,
who taught me cockroach dissection and provided suggestions for my projects; Prof Jozef
Vanden Broeck, who provided the equipment and reagents to run the AstR functional assays;
Sven Zels, who taught me the technique of receptor functional assays; Ilke van Hazel, who
helped me with the cell culture and the expression of NMDA receptors.
I would like to express my gratitude to my committee members: Belinda Chang, Ian Orchard,
William G. Bendena, David Lovejoy and Les Buck. I thank them for their great suggestions for
my projects and my thesis.
I would like to thank all my friends in Toronto and in China. They brought so much happiness
and joy to my life, and made my life in Toronto pleasant and colorful. Lastly, I would like to
thank my parents and my brother for their support in the past four and a half years.

v
Table of Contents
Abstract ……………………………………………………………………………………….ii
Acknowledgement …………………………………………………………………………….iii
Table of Contents……………………………………………………………………………...v
Table of Figures………………………………………………………………………………..vii
Table of Tables…………………………………………………………………………………ix
Abbreviations…………………………………………………………………………………..x
Chapter 1: General Introduction
1.1 Juvenile hormones………………………………………………………………… 1
1.2 JH biosynthetic pathway…………………………………………………………...7
1.3 JH signaling pathway……………………………………………………………... 26
1.4 Diploptera punctata………………………………………………………………. 32
1.5 Regulation of JH titre……………………………………………………………... 38
1.6 Rational and objectives of my study……………………………………………... 49
1.7 References………………………………………………………………………….. 52
Chapter 2: Characterization of the Juvenile Hormone pathway in the viviparous
cockroach, Diploptera punctata
2.1 Summary…………………………………………………………………………... 69
2.2 Introduction……………………………………………………………………….. 69
2.3 Materials and Methods…………………………………………………………… 73
2.4 Results…………………………………………………………………………….... 80
2.5 Discussion………………………………………………………………………….. 90
2.6 Supplementary data………………………………………………………………. 96
2.7 References………………………………………………………………………… 100
Chapter 3: Mode of action of allatostatins in the regulation of juvenile hormone
biosynthesis in the cockroach, Diploptera punctata
3.1 Summary………………………………………………………………………….. 105

vi
3.2 Introduction………………………………………………………………………. 105
3.3 Materials and Methods…………………………………………………………... 108
3.4 Results……………………………………………………………………………...114
3.5 Discussion………………………………………………………………………….126
3.6 Supplementary data……………………………………………………………….131
3.7 References………………………………………………………………………… 134
Chapter 4: Identification and characterization of the NMDA receptor and its role in
regulating reproduction in the cockroach, Diploptera punctata
4.1 Summary………………………………………………………………………….. 138
4.2 Introduction………………………………………………………………………. 138
4.3 Materials and Methods…………………………………………………………... 141
4.4 Results……………………………………………………………………………...144
4.5 Discussion………………………………………………………………………….155
4.6 Supplementary data……………………………………………………………….160
4.7 References………………………………………………………………………… 166
Chapter 5: General discussion
5.1 Function of JH in reproduction...……………………………………………….. 169
5.2 Evolution of the JH biosynthetic pathway...……………………………………. 170
5.3 Regulation of JH biosynthesis….………………………………………………... 172
5.4 The value of my study in insect control…..……………………………………...175
5.5 Future perspective……………………………………………………………….. 176
5.6 References………………………………………………………………………….177
Chapter 6: Appendices……………………………………………………………………… 180

vii
Table of Figures
Figure 1.1 Structures of the JH homologues in insects 2
Figure 1.2 The JH biosynthetic pathway 8
Figure 1.3 Dynamics of JH metabolism throughout the life cycle of D. punctata 35
Figure 2.1 Scheme of JH biosynthetic pathway 71
Figure 2.2 Tissue specific expression of genes encoding JH biosynthetic enzymes 82
Figure 2.3 Developmental expression of genes encoding JH biosynthetic enzymes
during the first gonadotrophic
cycle of D. punctata
84
Figure 2.4 The effect of JH precursors on JH biosynthesis by CA from mated female
D. punctata
86
Figure 2.5 Efficiency of HMGR-JHAMT RNAi-mediated knockdown and the effect
of silencing on the transcription of the other genes encoding enzymes in the
JH biosynthetic pathway in day 4 mated female D. punctata.
86
Figure 2.6 JH regulates ovarian development 88
Figure 2.7 Transverse sections of the basal oocytes from day 4 control and HMGR-
JHAMT
dsRNA-treated animals.
89
Figure 3.1 Relative expression levels of Dippu-AstR and Dippu-AST mRNA in tissues
of day 4 males and mated females.
115
Figure 3.2 Relative expression levels of Dippu-AstR and Dippu-AST mRNA during the
first
gonadotrophic cycle
116
Figure 3.3 The effect of Dippu-AST dsRNA on JH biosynthesis by the CA. 120
Figure 3.4 Dose-response curves for ASTs in CHO-WTA11 cells expressing Dippu-
AstR.
121
Figure 3.5 Dose-response curves for the bioluminescence response induced in
CHOPAM28 and HEK293 cells expressing Dippu-AstR
122
Figure 3.6 The effect of Dippu-AstR dsRNA on JH biosynthesis by the CA and on the
expression of genes encoding enzymes in the JH biosynthetic pathway of
D. punctata
123
Figure 3.7 The effect of AST on the expression levels of genes encoding enzymes in
the JH biosynthetic pathway in CA of day 6 mated female D. punctata
124
Figure 3.8 JH precursors rescue the AST-induced JH inhibition. 124
Figure 4.1 Amino acid sequence alignment of the two Diploptera NR1 subunit
(DpNR1A, DpNR1B), and homologous receptors from D. melanogaster
and T. castaneum
145
Figure 4.2 Phylogram depicting the relationship between the NR1 subunits from
Diploptera and orthologues of this receptor from other insects.
146
Figure 4.3 Molecular characterization of DpNR2. 149

viii
Figure 4.4 Graphic representation of the relative tissue distribution of (A) DpNR1A
transcript levels, (B) DpNR1B transcript levels and (C) DpNR2 transcript
levels in tissues of day 4 adult male and mated female D. punctata.
150
Figure 4.5 Relative transcript levels of DpNR1A, DpNR1B and DpNR2 in brains of
mated female D. punctata from day 0-day 7 after ecdysis.
151
Figure 4.6 Relative transcript levels of DpNR1A, DpNR1B and DpNR2 in CA of of
mated female D. punctata from day 0-day 7 after ecdysis
151
Figure 4.7 Relative transcript levels of DpNR1A, DpNR1B and DpNR2 in testes of
differentages of male D. punctata.
152
Figure 4.8 The effect of DpNR2 dsRNA treatment on JH biosynthesis and basal
oocyte growth, and the interactions among these genes in mated female D.
punctata.
152
Figure 4.9 In vivo effect of MK-801 on JH biosynthesis, basal oocyte growth and
relative Vg mRNA levels.
154
Figure 6.1 The effect of topical application of K15 and W206 on JH biosynthesis and
oocyte growth
183

ix
Table of Tables
Table 2.1 q-RT-PCR primer sequences and reaction efficiencies and correlation
coefficients in the q-RT-PCR assay
77
Table 2.2 Primers for dsRNA construction 79
Table 3.1 Potency of Dippu-ASTs a: activation of AstR in CHO-WTA11 cells
(EC50) or inhibitory effect on JH release (IC50)
125
Table 6.1 Structure of AST analogs 181
Table 6.2 Potency of Dippu-AST analogs a: inhibitory effect on JH release (IC50)
and activation of AstR in CHO-WTA11 cells (EC50)
182

x
Abbreviations
20E 20-hydroxyecdysone
AC adenylate cyclase
AST allatostatin
AstR Allatostatin receptor
AT Allatotropin
CA corpora allata
CHO Chinese hamster ovary
CRE cAMP responsive element
DMMP diphosphomevalonate
DMPP dimethylallyl pyrophosphate
FALD Farnesal dehydrogenase
FOLD Farnesol dehydrogenase
FPP farnesyl diphosphate
FPPP Farnesyl diphosphate pyrophosphatase
FPPS Farnesyl diphosphate synthase
GnRH gonadotropin releasing hormone
GPP Geranyl pyrophosphate
HEK human embryonic kidney
HMGR 3-hydroxy-3-methylglutaryl-CoA reductase
HMGS 3-hydroxy-3-methylglutaryl-CoA synthase
IPP isopentenyl pyrophosphate
IPPI Isopentenyl diphosphate isomerase
JH juvenile hormones
JHMAT Juvenile hormone acid O-methyltransferase
Kr-h1 Krüpel-homologues 1
LH luteinizing hormone
MA mevalonic acid
Met Methoprene-tolerant
MF methyl farnesoate
MK Mevalonate kinase
NMDAR N-methyl-D-aspartate receptor
PMK Phosphomevalonate kinase
PPMD Diphosphomevalonate decarboxylase
PTX pertussis toxin
RXR retinoid X receptor
Thiol Acetoacetyl-CoA thiolase
USP Ultraspiracle
Vg vitellogenin
Vn vitellin

1
Chapter 1
General Introduction
1 Juvenile hormones
The juvenile hormones (JH), a family of acyclic sesquiterpenoids, play essential roles in
regulating growth, development, metamorphosis, aging, caste differentiation and reproduction in
insects. This family of hormones has been extensively studied because of its central role in insect
development and reproduction and their potential value in pest control. This section will review
the current knowledge of JHs, including the JH homologues, enzymes in the JH biosynthetic
pathway and signal pathways of JH in insects.
1.1 JH homologues
JHs are synthesized and secreted by specialized, paired endocrine glands, the corpora allata (CA).
As early as 1934, Wigglesworth pointed out that insect metamorphosis was controlled by a
hormone produced by CA, a gland near the insect brain (Wigglesworth, 1934). In 1956, a highly
active extract, which produced anomalies in metamorphosis, was obtained from Cecropia Moth
Hyalophora cecropia (Williams, 1956). The structure of the first JH homologue was later
elucidated by Röller et al (Röller et al., 1967), as methyl (2E,6E,10-cis)-10,11-epoxy-7-ethyl-3,
11-dimethyl-2,6-tridecadienoate. The structure was further confirmed as the 2E,6E,10-cis isomer
(Dahm et al., 1968), and the absolute configuration of the chiral centers (C10 and C11) was
determined to be 10R,11S (Faulkner and Petersen, 1971; Meyer et al., 1971; Nakanishi et al.,
1971). This JH homologue was known as JH I (Fig. 1.1) (Goodman and Cusson, 2012).

2
COOCH3
O
COOCH3
O
JH I
JH II
COOCH3
O
JH III
COOCH3
O
JH 0
COOCH3
O
iso-JH 0
COOCH3
O O
JHB3
COOCH3
O
JHSB3
O
COOCH3
O
4'-Hydroxy JH III
COOCH3
O
HO
HO
COOCH3
O
8'-Hydroxy JH III
12'-Hydroxy JH III
HO
COOCH3
MF
23
4
5
67
8
9
1011
1
Figure 1.1 Structures of the JH homologues in insects. Figure adapted from Goodman and
Cusson (2012)

3
JH I, which has only been identified in the Lepidoptera, not only plays important roles in
regulating development, morphogenesis and reproduction in the Lepidoptera, but also has an
effect on the development of other insects (Fisher and Mayer, 1982; Granger et al., 1979;
Granger et al., 1982; Shalaby et al., 1990; Steiner et al., 1999). In larval development of Corcyra,
JH I treatment on ligated early-last instar resulted in a stimulation on DNA synthesis with a
consequent increase in DNA content and DNA concentration (Lakshmi and Dutta-Gupta, 1990).
During the last half of the larval molt of the tobacco hornworm, M. sexta, the presence of JH I at
the peak of the ecdysteroid titer is important in inducing dopa decarboxylase (DDC), an enzyme
which converts dopa to dopamine (Hiruma and Riddiford, 1985). In other insects, topical
application of the synthetic JH I to adult Musca domestica vicina Macq resulted in a shortened
gonoadotrophic cycle, decreased number of eggs and reduced hatching rate (Shalaby et al., 1990).
In addition, addition of JH I to the culture medium improved the development of single two-cell-
stage embryos of a polyembryonic wasp Copidosoma floridanum (Iwabuchi, 1995).
A second JH homologue, JH II (methyl (2E, 6E, 10-cis)-10,11-epoxy-3,7,11-trimethyl-2,6-
tridecadienoate) was identified in H. cecropia extracts (Meyer et al., 1970; Meyer et al., 1968).
JH II is the 2E, 6E, 10-cis isomer, as in JH I, but differs from JH I by a methyl group at C7 (Fig.
1.1). The absolute configuration of natural JH II at the C10, C11 positions has not yet been
determined (Goodman and Cusson, 2012). Same as JH I, JH II has only been identified in the
Lepidoptera. Nevertheless, relatively little research has been performed on JH II. In Trichoplusia
ni, the JH (JH I and JH II) and ecdysteroid titres were determined from the egg to the pupal molt
(Grossniklaus-Burgin and Lanzrein, 1990). Very little JH was detected in the freshly laid eggs of
T. ni, while in larval stages, JH II appeared to be the predominant or exclusive juvenile hormone
to interact with ecdysteroids to regulate the larval development. JH II, which is the most

4
abundant JH in Sesamia nonagrioides, is involved both in diapause programming and diapause
manifestation in this animal (Eizaguirre et al., 2005). In addition, JH II appears to be able to
initiate male production followed by sexual reproduction in the water flea Daphnia magna
(Cladocera, Crustacea). Exposure of D. magna to either JH I or JH II reduces the reproduction
rate, and induces parthenogenetically reproducing D. magna to produce male neonates (Oda et
al., 2005).
JH III was first identified from organ cultures of CA of the tobacco hornworm moth, Manduca
sexta (Judy et al., 1973). JH III displays the same E, E configuration at C2, C3 and C6, C7;
however, it differs from other JH homologues, with methyl groups at the C3, C7, and C11
positions. This hormone only contains one chiral carbon (C10), which displays the 10R
configuration in insects (Fig. 1.1). Of the juvenile hormone family, JH III is the most ubiquitous
JH homologue since it is the only JH biosynthesized and released in Orthoptera, Coleoptera,
Diptera, Hymenoptera, Dictyoptera, Lepidoptera, and the primitive ametamorphic Thysanura
(Baker et al., 1984; Tobe and Stay, 1985b). In larvae and adults of many insects, JH III is the
principal or only JH homologue identified, such as in the cockroach, Nauphoeta cinerea,
Diploptera punctata and the firebrat, Thermobia domestica (Baker et al., 1984; Tobe et al., 1985).
JH 0 and its isomer 4-methyl JH I (iso-JH 0) were identified in M. sexta eggs (Bergot et al.,
1981). Differing from JH III, JH 0 contains ethyl groups at the C3, C7, and C11 positions (Fig.
1.1). To date, JH 0 and its isomer (iso-JH 0) have been identified only in the Lepidoptera and
their functions in insects were unclear. JH III bisepoxide (JHB3), which contains a second
epoxide substitution at C6, C7, was first identified in Drosophila melanogaster (Richard et al.,
1989b). JHB3 was determined to be the major in vitro JH product of larval ring glands and of
adult CA-corpus cardiacum (CC) complexes of D. melanogaster. Later study identified JHB3 in

5
various dipteran species, such as Ceratitis capitata, Lucilia cuprina, Phormia regina,
Sarcophaga bullata (Bylemans et al., 1998; Lefevere et al., 1993; Moshitzky and Applebaum,
1995; Moshitzky et al., 2003). It has been demonstrated that the higher cyclorrhaphous Diptera
produce JHB3 predominantly, and JHB3 is believed to be restricted to the higher Diptera
(Richard et al., 1989a; Richard et al., 1989b). In L. cuprina, JHB3 is the only juvenile hormone
biosynthesized in vitro (Lefevere et al., 1993). Although it appeared that JHB3 production was
restricted to higher Diptera, JHB3 was reported to be synthesized by CA and the male accessory
glands of the mosquitoes, A. aegypti in vitro (Borovsky et al., 1994). However, recent work by Li
et al. (Li et al., 2003) was unable to detect any JHB3 synthesized by CA complex of A. aegypti.
The existence of JHB3 in other orders has yet to be confirmed.
Even though JHs were identified and characterized in various species of insects, the structure of
the JH in order Hemiptera has been a matter of controversy (Kotaki, 1993, 1996). Although JH
III and methyl farnesoate (MF) were reported as the products of CA in vitro in Dysdercus
fasciatus (Bowers et al., 1983; Feldlaufer et al., 1982) and the presence of JH I in the
hemolymph of Riptortus clavatus (Numata et al., 1992), there were no significant levels of the
known JH or related compounds in the milkweed bug, Oncopeltus fasciatus (Baker et al., 1988).
The presence of an unknown Heteropteran JH was suggested (Miyawaki et al., 2006). The
mystery regarding the JH in Hemiptera was not resolved until 2009, when Kotaki et al (2009)
identified a new JH homologue, JH III skipped bisepoxide (JHSB3; Fig. 1.1) from Plautia stali, a
member of the family Pentatomidae, suborder Heteroptera, order Hemiptera using a novel
approach. The term “skipped” refers to a second epoxide substitution switching from C6, C7 as
in JHB3 to C2, C3. The absolute chemical structure of the novel skipped bisepoxide JH was
characterized by the screening of a JH molecular library, and the juvenilizing activity of JHSB3

6
with different configurations on C2 and C3 and chirality on C10 was determined. Their result
shows that the (2R, 3S) configuration is more important for biological activity than the chirality
of C10, C11. JHSB3 with the 2R, 3S-configuration was more potent than those with the 2S, 3R-
configuration and 2,3-double bond (Kotaki et al., 2011). The function of JHSB3 was determined
in the last instars and adults of P. stali (Kotaki et al., 2011). Topical application of JHSB3 to last
instar nymphs inhibited their metamorphosis, and JHSB3 application in allatectomized and
diapausing adults stimulated the development of ovaries and ectadenia in females and males,
respectively.
Another family of JH homologues, hydroxylated JHs (HJHs; Fig. 1.1) was identified from the
African locust Locusta migratoria (Mauchamp et al., 1999). JH III was identified as the main
product released by the CA in vitro of L. migratoria (Mauchamp et al., 1985), while later studies
discovered three different hydroxylated forms of JH III (4-OH, 8-OH, and 12-OH JH III)
exhibited JH-like biological effects (Darrouzet et al., 1997; Mauchamp et al., 1999), in which 12-
OH JH III was found to be 100-fold more active than JH III.
Methyl farnesoate (MF; Fig. 1.1), a JH precursor without a C10, C11 epoxidation as in JH III,
was first isolated to function as a JH from the hemolymph of the spider crab Libinia emarginata
(Laufer et al., 1987). Recent studies have shown multifunctional roles of MF in crustaceans,
including reproduction, molting, larval development, morphogenesis, behaviour and general
protein synthesis (Chang et al., 2001; Nagaraju, 2007). Studies in insects suggest that MF may
also serve as a hormone in some insects. In the embryos of the cockroach N. cinerea, the
predominant product released by the embryonic CA is MF until the stage of breaking of the
chorion, and this substance circulates in embryonic haemolymph (Bürgin and Lanzrein, 1988;
Lanzrein et al., 1984). MF was also found to be biosynthesized by ring glands of larval D.

7
melanogaster (Richard et al., 1989a; Richard et al., 1989b), the embryonic CA of D. punctata
(Cusson et al., 1991b), the larval CA of Pseudaletia unipuncta (Cusson et al., 1991b) and the
adult CA of Phormia regina (Yin et al., 1995). In addition, MF was also identified from the
hemolymph of five orders of insects, including D. melanogaster (order Diptera), Schistocerca
americana (order Orthoptera), three species of true bugs (order Hemiptera), worker honeybees,
Apis mellifera (order Hymenoptera), and three species of beetle (order Coleoptera) (Teal et al.,
2014). Based on the study on the activity of MF, Goodman and Cusson (2012) reviewed the in
vivo biological role of MF in larvae and adults of D. melanogaster: During the larval stage, MF
is more active in blocking adult development than JH III or JHB3, whereas JH III or JHB3 is
more active once pupariation has been initiated.
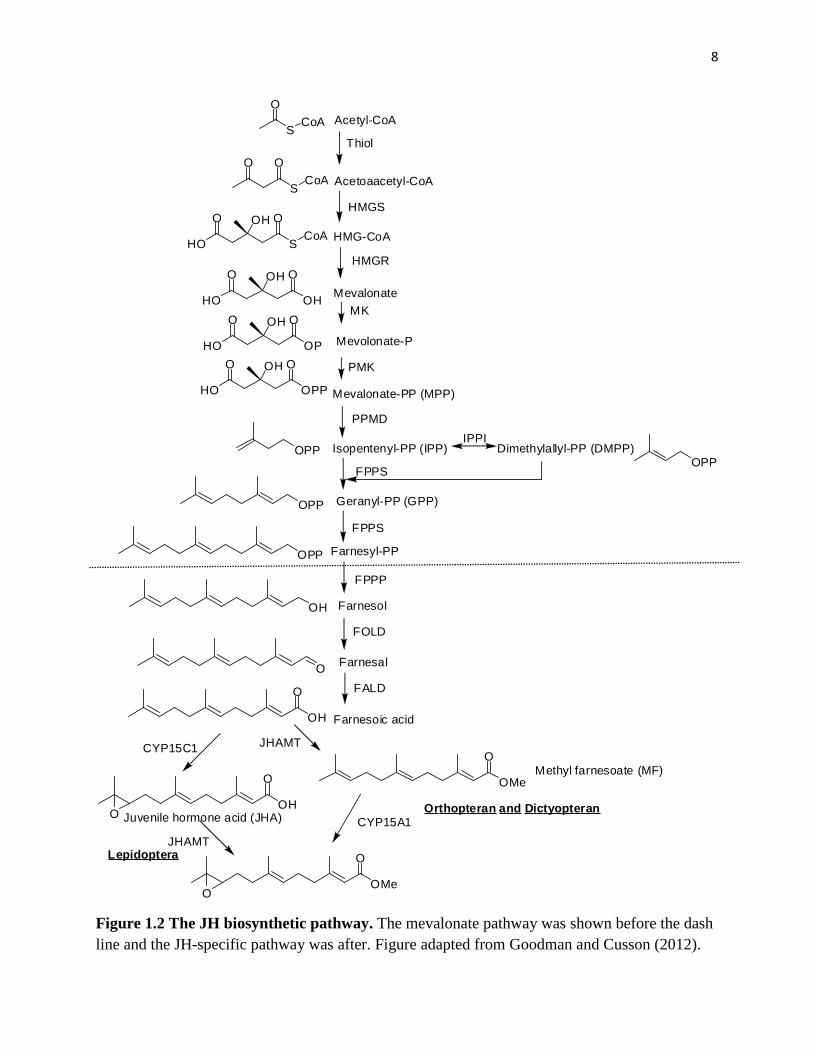
2. JH biosynthetic pathway
The JH biosynthetic pathway, which comprises 13 discrete enzymatic steps, can be divided into
two distinct biosynthetic parts: the mevalonate pathway and the JH-specific pathway (Fig. 1.2).
The mevalonate pathway is an important cellular metabolic pathway present in all higher
eukaryotes and many bacteria. The isoprenoids produced by the mevalonate pathway are vital for
diverse cellular functions, including the synthesis of cholesterol, haem A, ubiquinone, dilochol,
and farnesylated proteins, growth control, and electron transport (Goldstein and Brown, 1990).
The most well-studied product of mevalonate pathway is cholesterol, because of its role in
maintaining cell membranes and its implications for human cardiovascular diseases (Goldstein
and Brown, 1990). The initial stages of biosynthesis of JH to the formation of farnesyl
diphosphate (FPP) proceeds through the mevalonate pathway, which is shared in vertebrates and
invertebrates (Belles et al., 2005; Goodman and Cusson, 2012).
8
S
O
CoA
Thiol
S
O
CoA
O
HMGS
Acetyl-CoA
HMG-CoA
Acetoaacetyl-CoA
S
O
CoA
OH
HO
O
Mevalonate
HMGR
OH
OOH
HO
O
MK
PMK
Mevalonate-PP (MPP)
Isopentenyl-PP (IPP)
PPMD
Dimethylallyl-PP (DMPP)
Farnesyl-PP
Farnesol
Farnesal
Farnesoic acid
FPPS
FPPP
FOLD
FALD
OPP
OOH
HO
O
OPPOPP
IPPI
OPP
OH
O
OH
O
OH
O OMe
O
OMe
O
O
O
JHAMT
JHAMT
CYP15A1
Mevolonate-POP
OOH
HO
O
Geranyl-PP (GPP)OPP
FPPS
Orthopteran and Dictyopteran
Lepidoptera
Methyl farnesoate (MF)
Juvenile hormone acid (JHA)
CYP15C1
Figure 1.2 The JH biosynthetic pathway. The mevalonate pathway was shown before the dash
line and the JH-specific pathway was after. Figure adapted from Goodman and Cusson (2012).

9
Insects and other arthropods, however, do not produce cholesterol as a final product of the
mevalonate pathway, because they lack the enzymes squalene synthetase (farnesyl-diphosphate
farnesyltransferase) and lanosterol synthase, which are required for the production of cholesterol
(Clark and Bloch, 1959). Thus, the second portion of the JH biosynthetic pathway comprises
enzymatic steps unique to JH-producing organisms. Earlier studies on the JH biosynthetic
pathway focused on the activity of enzymes in the mevalonate pathway (Casals et al., 1996;
Couillaud and Feyereisen, 1991; Feyereisen and Farnsworth, 1987a). Thanks to whole genome
sequencing and CA trancriptomics studies, the identification and characterization of JH
biosynthetic enzymes have greatly improved, especially for genes encoding enzymes in the JH-
specific pathway (Consortium, 2006; Group, 2004; Mita et al., 2004; Noriega et al., 2006). Since
JH III is the most common JH homologs in insects, our review focuses on the study of enzymes
directly involved in the biosynthesis of JH III.
2.1 Acetoacetyl-CoA thiolase (ACAT, Thiol)
The synthesis of JH begins with acetyl-CoA, which condenses with another acetyl-CoA through
the catalysis of Thiol to form Acetoacetyl-CoA (Fig. 1.2). The gene encoding Thiol has been
identified in the genome of D. melanogaster, B. mori, Anopheles gambiae and A. aegypti, A.
mellifera and in an EST of the scolytid beetle Ips pini (Bomtorin et al., 2014; Eigenheer et al.,
2003; Keeling et al., 2004; Kinjoh et al., 2007; Nouzova et al., 2011). In vertebrates, the
functional Thiol is comprised by a tetramer of identical subunits and has two cysteine residues at
the active sites (Gehring and Harris, 1970), which are conserved in insects. In B. mori, Thiol was
almost exclusively expressed in the CA-CC complex, wheras the transcription of Thiol in A.
aegypti and A. mellifera was expressed in many tissues, including the ovary and fat body
(Bomtorin et al., 2014; Kinjoh et al., 2007; Nouzova et al., 2011).

10
2.2 3-hydroxy-3-methylglutaryl-CoA synthase (HMGS)
The function of HMGS is to catalyze the condensation of acetyl-CoA and acetoacetyl-CoA to
yield HMG-CoA (Fig. 1.2). The enzymatic activity of HMGS in the CA was determined in the
adult female of D. punctata. The results show that enzymatic activity of HMGS has the same
pattern as JH III biosynthesis, which suggests that HMGS influences the rate of JH biosynthesis.
However, the fact that the enzyme activity declined after the dramatic decrease of JH on day 6
indicates that changes in the activity of HMGS enzymes are apparently not responsible for the
collapse in JH synthetic ability on day 6 (Couillaud and Feyereisen, 1991).
The gene encoding HMGS was first isolated in the cockroach B. germanica (Buesa et al., 1994;
Martinez-Gonzalez et al., 1993), and later in D. melanogaster (Spradling et al., 1999), the
scolytid beetle Dendroctonus jeffreyi (Tittiger et al., 2000), B. mori (Kinjoh et al., 2007), A.
aegypti (Nouzova et al., 2011), and A. mellifera (Bomtorin et al., 2014). HMGS predominantly
expressed in CA-CC complex of B. mori (Kinjoh et al., 2007), A. aegypti (Nouzova et al., 2011),
and A. mellifera (Bomtorin et al., 2014), and the transcript levels correspond to JH biosynthesis
in the CA.
In B. germanica, two HMGS enzymes (HMGS-1 and HMGS-2) with 69% amino acid identity
were demonstrated (Buesa et al., 1994), and the gene encoding HMGS-1 was considered to be a
functional retrogene derived from HMGS-2 by retrotransposition (Buesa et al., 1994; Cabano et
al., 1997; Casals et al., 2001). In mammals, two forms of HMGS (a mitochondrial form and a
cytoplasmic form) have been detected, which are encoded by two different genes (Ayte et al.,
1990). In B. germanica, none of the two HMGS enzymes show any recognizable N-terminal
leader peptide to target the protein to mitochondria, which suggests that the enzyme is cytosolic

11
in insects (Buesa et al., 1994). Both HMGS are highly expressed in the adult ovary, coordinately
regulated in the ovary during the gonadotrophic cycle, but expressed differently throughout
development (Ayte et al., 1990; Martinez-Gonzalez et al., 1993). The expression and enzymatic
activities of both HMGS were also determined in the fat body of B. germanica (Casals et al.,
1996). HMGS-1 did not show any significant mRNA level or detectable protein level in the fat
body, which indicates a limited role for HMGS-1 in the fat body. HMGS-2, on the other hand,
shows a clear pattern in the fat body, which was consistent with that of vitellogenin production.
In D. jeffreyi, HMGS transcript localizes mainly in the metathorax and abdomen (Tittiger et al.,
2000). Topical application of JH III induced a dose- and time-dependent increase in HMGS
transcripts in the male metathoracic-abdominal region, whereas no increase in the transcript
levels was observed in the JH III-treated female. The JH III-mediated regulation of HMGS
suggests that in addition to its function in the JH biosynthetic pathway, HMGS appears to control
the isoprenoid pathway (Tittiger et al., 2000).
2.3 3-hydroxy-3-methylglutaryl-CoA reductase (HMGR)
HMGR catalyzes the first committed step of the isoprenoid biosynthetic pathway, the conversion
of HMG-CoA to mevalonate (Fig. 1.2). It is believed to play an important role in the regulation
of sterol synthesis and is generally referred to as the rate-limiting enzyme in cholesterol synthesis
in vertebrates (Goldstein and Brown, 1990). Two distinct classes of HMGR have been identified
(Bochar et al., 1999): in eukaryotes, HMGR consist of a highly conserved C-terminal catalytic
domain and a poorly conserved N-terminal membrane anchor domain, which contains two to
eight inferred transmembrane helices. The second class of HMGR was discovered in the Archaea
and the true bacterium Pseudomonas mevalonii, which lack the N-terminal membrane anchor
domain. Both HMGR serve the same function in different species.

12
The presence of HMGR in the CA was first reported in M. sexta, which convert HMG-CoA to
mevalonate (Bergot et al., 1979). The activity of HMGR was further studied in D. punctata
(Feyereisen and Farnsworth, 1987a) and the grasshopper, Schistocerca nitens (Baker and
Schooley, 1981). HMGR in the insect undergoes phosphorylation (inactive form) and
dephosphorylation (active form), which could be altered by Mg-ATP or NaF (Monger and Law,
1982). In M. sexta, the activity of HMGR in the CA parallels, in most cases, the ability of the
gland to synthesize JH (Baker and Schooley, 1981). In D. punctata, the activity of HMGR
parallels JH biosynthesis until day 5 during the first gonadotrophic cycle. JH biosynthesis drops
dramatically on day 6, while the activity of HMGR remains high till day 8. In addition, the half-
life of HMGR was not related to the half-life of JH III biosynthesis (Feyereisen and Farnsworth,
1987a). The results suggest that HMGR is not the ‘the rate-limiting enzyme’ in JH biosynthesis
in D. punctata.
To date, the gene encoding HMGR was cloned in many insect species, including D.
melanogaster (Gertler et al., 1988), B. germanica (Martinezgonzalez et al., 1993), the I. pini
(Hall et al., 2002), Ips paraconfusus (Tittiger et al., 1999), D. jeffreyi (Tittiger et al., 2003), the
moth Agrotis ipsilon (Duportets et al., 2000), B. mori (Kinjoh et al., 2007), A. aegypti (Nouzova
et al., 2011), and A. mellifera (Bomtorin et al., 2014). The identification of HMGR genes has
permitted further study of the function and the regulation of HMGR. For instance, a HMGR gene
encoding 916 amino acid was identified in D. melanogaster. The 56% identity in C-terminal
region to the hamster HMGR reflects the essential function of the C-terminal region. On the
other hand, the high similarity in the membrane-spanning regions between the mammalian and
Drosophila HMGR suggests that the transmembrane domains may be essential for recognizing
specific mevalonate derivatives or their binding proteins. In addition, the identification of

13
HMGR has allowed the expression of Drosophila HMGR in Schneider cells. Addition of
mevalonate suppresses the transcript level and the enzymatic activity of Drosophila HMGR,
which indicates a feedback regulation involved in the mevalonate pathway (Gertler et al., 1988).
The expression and the function of HMGR were also determined in many other insects. In B.
germanica (Martinezgonzalez et al., 1993), A. ipsilon (Duportets et al., 2000) and A. mellifera
(Bomtorin et al., 2014), HMGR was found to express in many tissues, including fat body, ovary,
muscle, brain and CA, whereas its expression in B. mori (Kinjoh et al., 2007) and A. aegypti
(Nouzova et al., 2011) was predominantly in the CA. Study of HMGR in Ips paraconfusus and D.
jeffreyi suggests that HMGR is not only involved in the regulation of JH biosynthesis, but also
the production of monoterpenoid pheromones in the males. (Tittiger et al., 2003; Tittiger et al.,
1999).
2.4 Mevalonate kinase (MK)
MK is responsible for the phosphorylation of mevalonate to produce the 5-phosphomevalonate
(mevalonate-P), which is metabolized in fungal, plant, and vertebrate systems to isopentenyl
pyrophosphate (IPPI) (Cornforth et al., 1960) (Fig. 1.2). The isolation, purification, and
characterization of MK was first reported in larval S. bullata by Goodfellow and Barnes (1971).
MK is distributed in the muscle and brain complex cytosol fractions of S. bullata (Cornforth et
al., 1960). The gene encoding MK was identified in D. melanogaster and A. gambiae genomes,
the EST database of D. punctata (Noriega et al., 2006), B. mori (Kinjoh et al., 2007), A. aegypti
(Nouzova et al., 2011), and A. mellifera (Bomtorin et al., 2014). In B. mori and A. aegypti, the
transcript level of MK is highest in the CA, whereas in A. mellifera, the highest expression of
MK is in the brain (Bomtorin et al., 2014; Kinjoh et al., 2007; Nouzova et al., 2011). In addition,

14
the expression profile of MK in the CA of B. mori and A. aegypti corresponded to changes in JH
biosynthesis.
2.5 Phosphomevalonate kinase (PMK)
Phosphomevalonate kinase (PMK) catalyzes the phosphorylation of mevalonate-P into 5-
diphosphomevalonate (MPP), an essential step in isoprenoid biosynthesis (Fig. 1.2). Two non-
orthologous genes encoding PMK have been identified: the Saccharomyces cerevisiae ERG8
gene, which is found in eubacteria, fungi and plants, and the human PMK gene, which is present
only in animals (Houten and Waterham, 2001). In insects, the gene encoding PMK was
identified in D. melanogaster and A. gambiae genomes, B. mori (Kinjoh et al., 2007), A. aegypti
(Nouzova et al., 2011), and A. mellifera (Bomtorin et al., 2014). In B. mori, PMK is expressed in
multiple tissues, and the expression profile in the larval CA shows a similar pattern as that of the
JH biosynthesis (Kinjoh et al., 2007). In A. aegypti, the PMK was highly expressed in the CA,
followed by ovary. The expression of PMK in the CA also coordinated with JH biosynthesis
(Nouzova et al., 2011).
2.6 Diphosphomevalonate decarboxylase (PPMD)
Mevalonate diphosphate decarboxylase (PPMD) is an enzyme in the mevalonate pathway that
catalyzes the decarboxylation of the six-carbon MPP to the five-carbon isopentenyl diphosphate
(IPP). This reaction involves the dehydration of the substrate and the hydrolysis of one molecule
of ATP, and Mg2+ is required (Jabalquinto et al., 1988). In rats, PPMD is a key enzyme in the
MVA pathway that is essential for the biosynthesis of the isoprenoids. However, there are not
many studies on PPMD in insects. The gene encoding PPMD was identified from the genomes
of D. melanogaster and A. gambiae, the EST database of I. pini (Keeling et al., 2004) and later in

15
B. mori (Kinjoh et al., 2007), and A. aegypti (Nouzova et al., 2011). In B. mori, PPMD was
exclusively expressed in CC-CA complex of 4th instar larvae, and its expression paralleled the JH
titre in 4th and 5th instars. In A. aegypti, PPMD was also exclusively expressed in the CA and its
expression coordinated with JH biosynthesis by CA, which indicates that PPMD is involved in
the regulation of JH biosynthesis in insects (Kinjoh et al., 2007; Nouzova et al., 2011).
2.7 Isopentenyl diphosphate isomerase (IPPI)
In the mevalonate pathway, IPP is the sole product of the ATP-dependent decarboxylation of
MPP and must be isomerized to DMPP to form Geranyl pyrophosphate (GPP) (Fig. 1.2). The
enzyme isopentenyl diphosphate isomerase (IPPI), which catalyzes the isomerization of
isopentenyl pyrophosphate (IPP) to dimethylallyl pyrophosphate (DMPP) (Fig. 1.2), plays a
central role in isoprenoid biosynthesis (Ramos-Valdivia et al., 1997). Two isoforms of IPPI have
been identified. Type 1 IPPI (IPPI-1) is a metalloprotein that is found in eukaryotes, and the
optimal functioning of IPPI-1 requires a divalent metal cation (Mg2+ or Mn2+). Zinc has also
been identified as an essential cofactor for the catalysis activity of IPPI-1 from E. coli (Carrigan
and Poulter, 2003). The type 2 isoform (IPPI-2) is a flavoenzyme found in plant chloroplasts and
bacteria; the isomerase activity requires not only a divalent metal cation, but also a reduced
flavin coenzyme (de Ruyck et al., 2014).
In insect species, IPPI was first partially characterized from the extracts of B. mori, in which that
the isomerase activity is dependent on the metal ions. Mn2+ was a better activator than Mg2+,
especially at low concentrations (Koyama et al., 1985). Later study identified the sequence of
IPPI in the genome database of D. melanogaster and A. gambiae, the EST database of I. pini
(Keeling et al., 2004), B. mori (Kinjoh et al., 2007), A. aegypti (Diaz et al., 2012; Nouzova et al.,

16
2011), the spruce budworm, Choristoneura fumiferana (Sen et al., 2012), , M. sexta (Sen et al.,
2012) and A. mellifera (Bomtorin et al., 2014). The IPPI gene identified in A. aegypti (AaIPPI)
encodes a 244 amino acid (aa) protein with high similarity to IPPI-1 in other organisms (Diaz et
al., 2012). Two important motifs which are associated with the catalytic roles of IPPI-1 are well
conserved: a TNACCSHPL motif containing a conserved cysteine residue and a WGEHEIDY
motif that contains a conserved glutamate residue. The function of IPPI-1 in other organisms
requires the binding of a divalent metal cation (Mg2+, Mn2+ or Zn2+) (Carrigan and Poulter, 2003),
whereas the enzymatic assay shows that the full activity of AaIPPI requires Mg2+ or Mn2+ but not
Zn2+, and its activity can be completely inhibited by iodoacetamide (Diaz et al., 2012).
Insects in the order Lepidoptera produce five JH homologues (JH 0, JH I, 4-methyl JH I, JH II,
and JH III) (See section 1.1). For the biosynthesis of different JH homologues, homologs of IPP
and DMPP are involved into the mevalonate pathway, which requires the IPPI in Lepidoptera to
catalyze the isomerization of homoisopentenyl diphosphate (HIPP) to homodimethylallyl
diphosphate (HDMPP). Earlier studies in pig demonstrated that the isomerization of HIPP by
porcine IPPI produced very little HDMPP. However, studies in insects demonstrated that CA
homogenates of adult female M. sexta and purified IPPI from B. mori regiospecificly catalyzed
the isomerization of HIPP to HDMPP, which suggests that the lepidopteran IPPI enzyme is
structurally distinct from other isomerases (Baker et al., 1981; Koyama et al., 1985). Further
study on IPPI in C. fumiferana and M. sexta confirmed the function of IPPI in catalyzing the
isomerization of HIPP, and the homology models of the CfIPPI and HIPP isomerization study
revealed that the lepidopteran IPPI enzyme has a larger active site cavity, to allow binding of
larger substrates and to stabilize the high-energy intermediate formed during substrate
isomerization (Sen et al., 2012).

17
The expression of IPPI mRNA varies in different tissues, but in most insects, the highest
transcript level of IPPI was found in the CA. In B. mori, IPPI mRNA levels are expressed almost
exclusively in the CA of 4th instar larvae, with relative low levels in other tissues (Kinjoh et al.,
2007). In A. mellifera and C. fumiferana, mRNA of IPPI was expressed in multiple tissues, with
the highest transcript level in the CA (Bomtorin et al., 2014; Sen et al., 2012). IPPI mRNA of A.
aegypti expressed in various tissues, including CA-CC, ovary, hindgut, brain, midgut, fat body of
the female, and testis and accessory glands of the male (Diaz et al., 2012). The ubiquitous
expression of IPPI suggests that IPPI might be involved in many metabolic pathways. The
pattern of change of IPPI mRNA in the CA-CC during female pupal and adult development was
consistent with the changes in JH biosynthesis, which suggests that the transcription of IPPI is
partially responsible for JH biosynthesis (Diaz et al., 2012; Nouzova et al., 2011). Similar results
were found in B. mori (Kinjoh et al., 2007). The expression of BmIPPI in the CA of 4th, 5th
larvae, and pupae coordinate with the JH titre.
2.8 Farnesyl diphosphate synthase (FPPS)
The condensation of IPP and DMPP forms an intermediate compound GPP, which then
undergoes a second condensation step to generate farnesyl diphosphate (FPP) (Fig. 1.2). This
process is catalyzed by the enzyme Farnesyl diphosphate synthase (FPPS), a type of
prenyltransferase. FPPS is a homodimeric protein, which is formed by tightly coupled subunits
ranging from 32 to 44 kDa in size (Vandermoten et al., 2009a). The activity of FPPS requires
divalent metal cations (Mg2+ or Mn2+). Sequence analysis of FPPSs revealed seven conserved
regions, including two substrate binding regions, regions II and VI. Both regions contain an
aspartate-rich motif, DDx(xx)xD (x represents any amino acid) (Liang et al., 2002). Region II,
which includes the first aspartate-rich motif (FARM), is responsible for the determination of

18
chain-length, while region VI, which contains the second aspartate-rich motif (SARM), is
considered to be the IPP binding site (Liang et al., 2002). The first FPPS genes in insects were
cloned in A. ipsilon by Castillo-Gracia and Couillaud (1999) with high identity (about 40%) with
other FPPSs and high transcript level in the CA. Additional insect FPPSs were identified in many
insect orders, including Lepidoptera (Sen and Sperry, 2002), Diptera (Nouzova et al., 2011; Sen
et al., 2007), Coleoptera (Taban et al., 2009), Hemiptera (Lewis et al., 2008; Sun and Li, 2012;
Zhang and Li, 2008), Hymenoptera (Bomtorin et al., 2014) and Blattodea (Noriega et al., 2006).
In Lepidoptera, two distinct forms of FPPS were identified, designated type-1 and type-2 FPPS
(FPPS-I and FPPS-II) (Cusson et al., 2006), while two slightly different isoforms of type-2 FPPS
are present in B. mori (FPPS-2 and FPPS-3) (Kinjoh et al., 2007). Like the prenyltransferases in
other organisms, FPPS in M. sexta requires the divalent cation (Mg2+ or Mn2+) for its activity.
The presence of detergent, glycerol, and non-specific protein-protein interactions improves the
stability and catalytic activity of FPPS (Sen and Sperry, 2002). As described in section 1.1, the
Lepidoptera produce five JH homologues (Fig. 1.1), which requires the synthesis of
ethyl/methyl-substituted FPP by FPPS. The preference of prenyltransferase to ethyl/methyl-
substituted DMPP was determined using M. sexta CA homogenates, and the results suggest that
the selectivity of the enzyme incline to the ethyl-substituted substrate (Sen et al., 1996).
Additional studies on the selectivity of this prenyltransferase were performed using different
substrate analogs (Sen et al., 2006). Compared to pig liver FPPS, the lepidopteran enzyme
derived from CA homologues displays greater steric latitude around the C-3 and C-7 alkyl
positions of DMAPP and geranyl diphosphate (GPP). The enzymes generate more ethyl-
branched geranyl/farnesyl diphosphate and the substrate specificity related to the enzyme
localization (Sen et al., 2006). The structure analysis of FPPS-I and FPPS-II in C. fumiferana

19
revealed that FPPS-1 displays several unique active site substitutions, whereas FPPS-II has a
more conventional catalytic cavity which indicates FPPS-I are better suited than FPPS-II for
generating ethyl-substituted products. However, tissue distribution of FPPS mRNA showed that
FPPS-I is ubiquitous whereas FPPS-II is predominately expressed in the CA (Cusson et al.,
2006). The result is consistent with the distribution of FPPS 1-3 mRNA in 4th instar B. mori
(Kinjoh et al., 2007). These results suggest that FPPS-II may play a leading role in lepidopteran
JH biosynthesis despite its apparently more conventional catalytic cavity. In other species, FPPS
forms a homodimeric protein. The recombinant C. fumiferana FPPS-2 was active in producing
FPP; However, expression of FPPS-1 (CfFPPS1, Pseudaletia unipuncta FPPS1, and A. ipsilon
FPPS1) in E. coli failed to display any FPPS activity in vitro. Surprisingly, the combination of
CfFPPS1 and CfFPP2 enhanced the enzyme activity, and an association between CfFPPS1 and
CfFPPS2 was observed, which suggests that FPPS-I and FPPS-II may derive from a heteromer to
play a role in JH biosynthesis in moths. Whether these two enzymes form heterodimers in vivo
has not yet been verified.
Two FPPS genes that encode proteins with about 80% identity were identified in the green peach
aphid, Myzus persicae, and in the bird cherry-oat aphid Rhopalosiphum padi (Sun and Li, 2012;
Zhang and Li, 2008). Enzyme activity of FPPS in R. padi shows both enzymes could catalyze the
formation of FPP from IPP and DMAPP (Sun and Li, 2012). However, the function of FPPS in
M. persicae was not determined. Without the feature of the FPPS in M. persicae, it is premature
to conclude that these genes are involved JH biosynthesis. On the other hand, other
prenyltransferase genes displaying dual geranyl diphosphate (GPP)/farnesyl diphosphate (FPP)
synthase activity in vitro were identified in M. persicae. These two prenyltransferase genes
encode very similar proteins, apart from the presence of a mitochondrial leader sequence (Lewis

20
et al., 2008). It is interesting to note that the prenyltransferase enzyme in aphid is not unique to
the aphid species from which it was cloned. The molecular dynamics of the enzyme are
responsible to maintain the balance between the production of GPP and FPP (Vandermoten et al.,
2009b).
Only single copies of the FPPS gene were identified in insects from other orders, such as Diptera
(Nouzova et al., 2011; Sen et al., 2007) and Coleoptera (Taban et al., 2009). The scanning of the
A. mellifera genome showed the presence of seven copies (Consortium, 2006). The expression of
the first six copies of the FPPS gene revealed that FPPS3 is the bona fide gene involved in JH
biosynthesis in honey bees (Bomtorin et al., 2014).
2.9 Farnesyl diphosphate pyrophosphatase (FPPP)
Farnesyl diphosphate pyrophosphatase (FPPP) catalyzes the hydrolysis of farnesyl diphosphate
(FPP) to farnesol (FOL). Relatively little was known about FPPP in insects. Cao et al. (2009)
screened the D. melanogaster genome and identified the first FPPP genes in the insects. FPPP
belongs to the haloalkanoic acid dehalogenase (HAD) super family that catalyzes phosphoryl
transfer reactions (Allen and Dunaway-Mariano, 2004). Members of the HAD phosphatase
superfamily have four conserved amino acid signature motifs, which are also well conserved in
the DmFPPP. Nyati et al. (2013) identified 3 putative FPPP (AaFPPP-1, -2, and -3) paralogs
through a search for orthologs of the DmFPPP in the CA of A. aegypti. Recombinant AaFPPP-1
and AaFPPP-2 displayed the features of FPPP in their ability to hydrolyze FPP into FOL, and the
FPPP activity of the CA extracts was found to be Mg2+-dependent. The determination of the
function of FPPPs in JH biosynthesis using RNAi reveals that FPPP-1 plays the predominant
function in JH biosynthesis. Unlike mRNA of enzymes in the mevalonate pathway which are

21
predominately expressed in the CA, AaFPPPs mRNA are expressed in various tissues, with
FPPP-1 highly expressed in midgut and Malpighian tubules, FPPP-2 in Malpighian tubules, and
FPPP-3 in brain and ovary. The ubiquitous expression of FPPP may result from the pleiotropic
functions of farnesol and farnesal. In spite of the ubiquitous expression, the expression of FPPP-
1 and -2 in the CA correlated with JH biosynthesis in sugar-fed females, which suggests FPPP
may play a role in the regulation of JH biosynthesis.
2.10 Farnesol dehydrogenase (FOLD)
A Farnesol dehydrogenase (FOLD) is responsible for the catalysis of the conversion of farnesol
(FOL) to farnesal (FAL) (Fig. 1.2). In vertebrates, plants, and fungi, the oxidation of FOL to
FAL is mediated by nicotinamide-dependent dehydrogenases (Chayet et al., 1973; Inoue et al.,
1984; Keung, 1991). Study of FOL oxidation using CA homogenates of the adult female M sexta,
revealed that farnesol and/or farnesal dehydrogenase were NAD+-dependent enzymes (Baker et
al., 1983). However, the conversion of FOL to FAL in larval M. sexta was not affected by
nicotinamide. The enzyme, which oxidizes FOL to FAL in larval M. sexta, appears to be an
oxygen-dependent enzyme, perhaps a flavin and/or iron-dependent oxidase (Sperry and Sen,
2001). A FOLD enzyme was identified and functionally characterized in the CA of adult A.
aegypti (Mayoral et al., 2009a). In CA of adult female M sexta, the FOLD enzyme was
ineffective in the addition of NADP+ (Baker et al., 1983). However, the FOLD in A. aegypti was
characterized to be a NADP+-dependent farnesol-dehydrogenase. It is possible that moths utilize
a different mechanism for FOL oxidation. In A. aegypti, FOLD is expressed in various tissues,
and with a relatively low transcript level in the CA. On the other hand, the transcript levels of
FOLD in the CA coordinate with JH biosynthesis (Mayoral et al., 2009a).

22
2.11 Farnesal dehydrogenase (FALD)
Farnesal dehydrogenase (FALD), which catalyzes the oxidation of farnesal to FA, was one of the
less understood steps in JH synthesis (Fig. 1.2). An early study using the CA homogenates of the
adult female, M sexta predicted that FALD is an NAD+-dependent aldehyde dehydrogenase and
this aldehyde dehydrogenase showed some substrate specificity for the 2E isomer (Baker et al.,
1983). Rivera-Perez et al. (2013) identified and characterized a FALD enzyme in female A.
aegypti. Two FALD genes with 50% amino acid identity were identified, in which FALD-1
produces four different transcripts and FALD-2 produces one. All five FALD variants exhibit the
activity to convert FAL to farnesoic acid (FA), and the oxidation is stimulated in the presence of
NAD+. mRNA of FALD variants have unique tissue distribution profiles, with each
predominantly expressed in one unique tissue: FALD1-A in ovaries, FALD1-B in Malpighian
tubes, FALD1-C in hindgut, FALD1-D in nervous tissue, while FALD2 are relatively low in
transcript level comparing to FALD1. The reduction of FALD activity results in accumulation of
farnesol, which subsequently converts back into farnesol, resulting in farnesol leaking out of the
CA. Oxidation of farnesal may be a rate limiting step in JH synthesis in mosquito after blood
feeding.
2.12 Juvenile hormone acid O-methyltransferase (JHAMT)
In the final two steps of JH biosynthesis, FA is converted to JH III through a methyl transfer and
an epoxidation (Fig. 1.2). The order of the final two steps is insect order dependent. In
Orthoptera, Dictyoptera, Coleoptera and Diptera, FA undergoes the methylation of the carboxyl
group to produce methyl farnesoate (MF), which is epoxidized by a P450 monooxygenase at C10,
C11 position to generate JH III. In Lepidoptera, however, a reverse step order occurs:

23
epoxidation precedes methylation. Other ethyl-branched JH homologues are also synthesized
through the same order, but with different precursors derived from homomevalonate and
mevalonate (Shinoda and Itoyama, 2003). The modeling of A. gambiae, B. mori, D.
melanogaster and T. castaneum JHAMTs and docking simulation shows that all insect JHAMTs
are able to esterify both FA and JHA. The order of the methylation/epoxidation may be
controlled by the specificity of the epoxidase. The epoxidase in Lepidoptera might have higher
affinity than JHAMT for FA, which results in epoxidation precedes methylation. In other insects,
however, the epoxidase can only selectively catalyze MF. Thus, esterification of FA to MF by
JHAMT occurs in earlier step (Defelipe et al., 2011).
The first JHAMT was cloned and functionally characterized in, B. mori (Shinoda and Itoyama,
2003). The sequence analysis reveals that JHAMT belongs to the SAM-dependent
methyltransferases family, with a conserved S-adenosyl-L-methionine (SAM) binding motif. The
protein encoded by the JHAMT gene was able to not only convert JH III acid to JH III, but also
catalyze the conversion of JHA I, II, and FA to their cognate JH methyl esters in the presence of
S-adenosyl-L-methionine (SAM). Northern blot analysis shows that BmJHAMT mRNA is
exclusively expressed in the CA. Expression of JHAMT correlates well with the JH biosynthetic
activity of the CA in 4th and 5th instar larvae, pupae and adults, and especially in 5th instar larvae
in which the shutdown of the expression of JHAMT appears to be the primary reason for the
decline in JH biosynthesis. These results suggest that JHAMT is the rate limiting enzyme in the
JH biosynthesis in B. mori (Kinjoh et al., 2007; Shinoda and Itoyama, 2003). Orthologues of
JHAMT have also been cloned and characterized in other insect species, including T. castaneum
(Minakuchi et al., 2008a), D. melanogaster (Niwa et al., 2008), the Eri silkworm, Samia cynthia
ricini (Sheng et al., 2008), A. aegypti (Mayoral et al., 2009b), the desert locust Schistocerca

24
gregaria (Marchal et al., 2011) and A. mellifera (Bomtorin et al., 2014). In all species, JHAMT
is expressed predominately in the CA and the recombinant JHAMT protein from these insect can
catalyze the methylation of FA into MF, as well as JHA into JH III. In T. castaneum, silencing
the JHAMT gene using RNAi induced precocious metamorphosis, while in D. melanogaster
(Minakuchi et al., 2008a), JHAMT overexpression resulted in a pharate adult lethal phenotype
(Niwa et al., 2008). In S. gregaria, knockdown of JHAMT not only resulted in lower JH release,
but also a suppression in FA-stimulated JH release. A delay in sexual maturation was also
observed in JHAMT-silenced animals (Marchal et al., 2011). In many insects, such as T.
castaneum (Minakuchi et al., 2008a), D. melanogaster (Niwa et al., 2008), S. cynthia ricini
(Sheng et al., 2008), S. gregaria (Marchal et al., 2011) and A. mellifera (Bomtorin et al., 2014),
the expression of JHAMT in the CA correlates well with JH biosynthesis.
Farnesoic acid O-methyltransferase (FAMeT), which was first reported in a crustacean, was
initially considered to be the enzyme converting FA to MF in crustaceans (Gunawardene et al.,
2001). And a FAMeT was cloned in D. melanogaster (Burtenshaw et al., 2008).
Immunohistochemical analysis shows the presence of FAMeT in the CA portion of the ring
gland. However, recombinant FAMeT did not show any enzymatic activity in catalyzing the
conversion of FA or JHA. In S. gregaria, FAMeT mRNA was expressed in several tissues, and
its expression in the CA did not correlate with JH biosynthesis. In addition, silencing FAMeT
has no effect on either JH release or MF content of the CA (Marchal et al., 2011). These results
suggest that FAMeT does not encode a functional methyltransferase.
2.13 Juvenile hormone epoxidase (CYP15A1/CYP14C1)

25
Juvenile hormone epoxidase is involved in the epoxidation of FA in the Lepidotera or the
epoxidation of MF in the Orthoptera, Dictyoptera, Coleoptera, and Diptera (Fig. 1.2). Early
studies demonstrated that this epoxidase is a microsomal cytochrome P450 enzyme (Hammock,
1975). The first JH epoxidase, named CYP15A1, was cloned from D. punctata (Helvig et al.,
2004). This enzyme contains all the features of a typical microsomal P450, and its recombinant
protein catalyzed the epoxidation of MF to JH III in the presence of NADPH. The product of
epoxidation is mostly the (10R)-enantiomer. Studies on JHAMT reveal that the order of the final
two steps is primarily determined by the substrate specificity of epoxidase (Section 1.2.12).
CYP15A1 from D. punctata showed strong substrate specificity to the natural substrate MF.
Substitution of natural MF with geometrical isomers of MF and other terpenoids, such as
farnesol, farnesal, FA, and JH III showed little or no activity. DpCYP15A1 mRNA was
exclusively expressed in the CA and the expression level is higher in CA with high JH
biosynthetic activity. The expression and function of CYP15A1 has also been investigated in
many other insects, including A. aegypti (Nouzova et al., 2011), S. gregaria (Marchal et al., 2011)
and A. mellifera (Bomtorin et al., 2014). In all, CYP15A1 was selectively expressed in the CA.
In A. aegypti and A. mellifera, the expression of CYP15A1 does not correlate with JH
biosynthesis, whereas in S. gregaria, CYP15A1 shows high levels of transcription in active CA.
Silencing CYP15A1 in S. gregaria resulted in a reduction in JH release and an accumulation of
MF within the CA.
CYP15C1, an ortholog of CYP15A1, was recently identified and characterized in the
Lepidoptera, B. mori (Daimon et al., 2012). And CYP15C1 shares high homology with the
CYP15A1 in D. punctata. The dimolting (mod) mutation, which causes precocious larval-pupal
metamorphosis, results in a null mutation in the coding sequence of CYP15C1. CYP15C1 was

26
then expressed in Drosophila S2 cells and enzymological analysis revealed that CYP15C1
converts FA to JHA in a highly stereospecific manner. Further study showed that CYP15C1 is
responsible for the mod mutant of B. mori and its molecular defect results in the absence of JHs
(JH I and JH II) in B. mori, indicating CYP15C1 plays essential roles in JH biosynthesis. In
addition, CYP15C1 is specifically expressed in the CA. The expression of CYP15C1, on the
other hand, did not show any change during development, which suggests that CYP15C1 is not
the rate-limiting enzyme in the JH biosynthetic pathway. The CYP15 gene is not found in higher
dipterans such as D. melanogaster, probably because the production of JHB3 requires a different
epoxidase.
3. JH signaling pathway
3.1 Ultraspiracle (USP) as a JH receptor
Ultraspiracle (USP) is a homologue of the vertebrate retinoid X receptor (RXR), which can form
heteromers with other nuclear receptors to bind with genomic response elements (Henrich et al.,
1994). In Lepidoptera and Diptera, USP displays high identity with RXR only in the DNA-
binding domain, whereas the similarity in the ligand-binding domains is relatively low. In other
insect orders and other arthropods, USP shows higher similarity to vertebrate (Iwema et al.,
2007). USP can interact with several nuclear receptors, such as EcR and DHR38 (Sutherland et
al., 1995; Yao et al., 1992). The USP:EcR complex, which is formed by the heterodimerization
between EcR and USP, is required for the binding of the steroid hormone 20-hydroxyecdysone
(Yao et al., 1992).
Using a fluorescence assay, Jones and Sharp (1997) demonstrated a protein-ligand interaction
between the natural Drosophila JHs (JH III ester monoepoxide and bisepoxide, respectively) and

27
recombinant D. melanogaster USP. This interaction cannot be influenced by the addition of
farnesol or 20E, which indicates the specificity of the binding. On the other hand, JH acid, which
itself did not change the fluorescence of USP, affects the interaction between JH III ester and
USP. The action of JH acid indicates that JH acid may bind to USP in a different manner than JH
III ester. Further studies on D. melanogaster USP showed that it could also specifically bind to
JH III, which changes the conformation of USP and stabilizes the dimeric/oligomeric quaternary
structure of USP. The JH III agonist methoprene shows a competitive inhibition in the the JH III-
USP interaction (Jones et al., 2001). Furthermore, the binding affinities between USP and other
natural farnesoid products of the ring gland of D. melanogaster were determined (Jones et al.,
2006). MF exhibited a nanomolar affinity to USP, and the addition of an epoxide across a double
bond or any substitution on C1 (Fig. 1.1) other than methyl ester, resulted in a decrease in
affinity to USP. Mutational analysis showed that the binding of JH III to USP was strongly
reduced by the mutation C472A/H475L.
Although USP has been demonstrated to interact with JHs, further studies on USP did not show
its effect on JH action. The structure-based analysis of a Heliothis virescens USP protein shows
that JHs could fit into the ligand binding pocket (LBP) of USP. However, the percentage of
occupancy of LBP was relatively low, which raised concerns over the validity of USP as an JH
receptor (Sasorith et al., 2002). In vivo USP activation assays in third instar Drosophila larvae
demonstrated that neither natural JHs (JHI, JHII and JHIII) nor JH analogs (pyriproxifen and
methoprene) were able to activate USP, whereas fenoxycarb, a carbamate insecticide that mimics
the action of JH, induced a weak activation. Beck et al. (2009) determined the activation of USP
by JHs and their analogs in transgenic animals expressing either the GAL4-TcUSP (T.
castaneum) or the GAL4-DmUSP (D. melanogaster). Their results show that JHs and their

28
analogs were not able to activate USP. In particular, MF, which displayed a nanomolar affinity to
DmUSP (Jones et al., 2001), did not show any effect on the activation of USP. On the other hand,
pre-incubation of organs with JH III lead to the repression of GAL4-TcUSP and the GAL4-
DmUSP activated by 20E. Thus, USP may not act as an JH receptor, but may interact with JH in
the EcR/USP complex.
3.2 Methoprene-tolerant (Met) as a JH receptor
Methoprene-tolerant (Met) is a basic-helix-loop-helix (bHLH)/Per-Arnt-Sim (PAS) protein
containing a HLH structure and two PAS domains (A and B). The Met gene was first discovered
by an ethyl methane sulfonate mutagenesis screen (Wilson and Fabian, 1986). The Met mutation
conferred a 100-fold-increased resistance to methoprene and JH III, and was resistant to
methoprene-induced pseudotumor formation in larvae and to JH III- or methoprene-induced
vitellogenic oocyte development in adult females, suggesting Met might be a JH receptor. The
hydroxyapatite (HAP) binding assay of JH III to JH III-binding protein in fat body cells revealed
a 10-fold lower binding affinity in the Drosophila Met strain (Shemshedini and Wilson, 1990).
In vitro synthesized Drosophila Met bound to JH III with high affinity (Kd = 5.3 ± 1.5 nM, mean
± SD). And the effectiveness of JHs in the activation of Met expressed in Drosophila S2 cells is
JH III>JH II>JH I>methoprene (Miura et al., 2005). A similar affinity was also found in the
binding of Tribolium Met to JH III (Kd = 2.94 ± 0.68 nM, mean ± SD) (Charles et al., 2011). The
function of Met and the high affinity of Met to JH suggest that Met may act as a JH receptor.
As a potential JH receptor, the ligand-binding properties of Met were determined (Charles et al.,
2011). To examine which part of the Tribolium Met protein is responsible for binding JH III, the
truncated proteins with part of the conserved domain were synthesized for a ligand-binding assay.

29
The results show that Met specifically binds JH III through its C-terminal PAS domain (PAS-B
domain plus C-terminal region). As a bHLH protein, Met requires either a homo- or heterodimer
partner for its activity (Kewley et al., 2004). In A. aegypti, a Ftz-F1-interacting steroid receptor
coactivator (FISC) was identified as a functional partner of Met in mediating JH-induced gene
expression (Li et al., 2011). Microarray analysis and RNAi studies revealed an ortholog of FISC
in T. castaneum, named steroid receptor co-activator (SRC), which was responsible for the
formation of heterodimer with Met (Zhang et al., 2011). The closest relative of FISC/SRC in
Drosophila is Taiman (Tai) (Charles et al., 2011). The study on the interaction between Tai and
Met suggested a model of JH action on Met in Drosophila. Met forms a homodimer in the
absence of JH, while the binding of JH to the PAS-B domain of Met results in conformational
changes to release Met from the homophilic complex and allows it to bind Tai (Charles et al.,
2011). Another JH-dependent heterodimeric partner of Met, Cycle (CYC) was identified in A.
aegypti (Shin et al., 2012). The binding between Met and CYC only occurs in the presence of JH
III, and is induced by JH III in a dose dependent manner. Both Met and CYC specifically binds
to the E-box-like motif from the Kr-h1 gene promoter. Silencing CYC, Met or SRC/FISC using
RNAi, impaired the circadian activation of Kr-h1 and Hairy genes. Based on the previous studies,
it currently appears that the JH receptor is composed of two DNA-binding bHLH/PAS
transcription factors, in which Met is an obligatory component and the partner of Met varies in
different insects.
In Drosophila, the Met mutant showed a high resistance to the toxic and morphogenetic effects
of JHs and their analogs (Shemshedini et al., 1990; Wilson and Ashok, 1998; Wilson and Fabian,
1986). However, although Met-null mutants show reduced oogenesis, they are viable. The
phenotype of complete absence of Met is too subtle to conclude that Met is a genuine JH

30
receptor (Wilson and Ashok, 1998). The possible reasons for the lack of an expected phenotype
are: (1) JH has a weak effect on preadult Drosophila (Wilson and Ashok, 1998), (2) a paralogous
of Met, germ-cell expressed (gce), exists in Drosophila. gce function as a JH receptor in the
absence of Met (Abdou et al., 2011). To determine the clear role of Met, Met was studied in
another insect model, T. castaneum, which only possess one Drosophila Met/gce gene. In this
insect, silencing Met display a more clear effect. The loss of Met in early-instar larvae resulted in
the production of premature pupae or heterochronic larva-pupa intermediates (Konopova and
Jindra, 2007). In addition, the knockdown of Met in the final larval instars disrupted the larval-
pupal ecdysis and induced precocious development of adult structures (Parthasarathy et al.,
2008). In the true bug, Pyrrhocoris apterus, knockdown of Met also results in the similar
phenotype as JH depletion, which causes precocious development of adult color pattern, wings
and genitalia (Konopova et al., 2011). Met regulates premetamorphosis and metamorphosis in
both holometabolous and hemimetabolous insects as in the JH signaling pathway.
JHs not only play important roles in insect metamorphosis, but also regulate the reproduction of
insects. As a key factor in the JH signaling pathway, the function of Met in reproduction has also
been determined. In the migratory locust, Locusta migratoria, silencing Met or Tai results in an
arrest of ovarian development and a reduction in vitellogenin gene expression in the fat body
(Song et al., 2014). In D. punctata, the silencing of Met blocked basal oocyte development,
suppressed the transcription of vitellogenin in the fat body and the uptake of vitellogenin by
ovary. In addition, the typical profile of JH biosynthesis was disrupted in Met-knockdown
animals, which results in the failure of patency (Marchal et al., 2014).
3.3 Krüpel-homologues 1 (Kr-h1) in downstream of JH signaling pathway

31
Kr-h1, a transcription factor with a DNA-binding motif of eight C2H2 zinc fingers, was shown to
be the JH-inducible target of Met. In Drosophila, the expression of Kr-h1 is in JH-dependent
manner and ectopic expression of Kr-h1 caused a phenotype similar to application of JH
(Minakuchi et al., 2008b). In a B. mori cell line, subnanomolar levels of natural JHs were able to
induce BmKr-h1 rapidly (Kayukawa et al., 2012). This induction involves BmMet2 and BmSRC:
JH ligand to BmMet2 and interact with BmSRC to form a JH/BmMet2/BmSRC complex, which
activates BmKr-h1 by interacting with a JH response element (kJHRE). The transcription of Kr-
h1 demonstrates that the induction of BmKr-h1 by JH occurs only in the epidermis of
penultimate-instars, but not in the prepupal stage (Kayukawa et al., 2014).
The role of Kr-h1 in conveying the JH signal to regulate metamorphosis has been studied in both
hemimetabolous and holometabolous insects, including B. germanica (Lozano and Belles, 2011),
P. apterus (Konopova et al., 2011; Smykal et al., 2014b), R. prolixus (Konopova et al., 2011),
D. melanogaster (Minakuchi et al., 2008b), T. castaneum (Minakuchi et al., 2009)
and B. mori (Kayukawa et al., 2014; Smykal et al., 2014b). Kr-h1 represses the adult
morphogenesis. Knocking down Kr-h1 in the third instar or penultimate-instar larvae resulted in
precocious adult development. The repressing action of Kr-h1 on morphogenesis involves the
expression of the broad gene in holometabolous insects (Minakuchi et al., 2009), but not in
hemimetabolous insects (Konopova et al., 2011; Smykal et al., 2014b). Even though the absence
of Kr-h1 is necessary for adult morphogenesis, it is not required to maintain the larval program
during the first two larval instars. The early stages of insect development appear to be initially
independent of JH (Smykal et al., 2014b).
Aside from the function of Kr-h1 in metamorphosis, its roles in reproduction were also examined.
In the migratory locust, L. migratoria, Kr-h1 was demonstrated to convey the JH signal for the

32
induction of Vg. Depletion of Kr-h1 results in a drastic reduction in Vg expression in the fat
body, and subsequently resulted in unsuccessful egg production (Song et al., 2014). However, in
P. apterus, knockdown of Kr-h1 did not block ovarian development or suppress Vg expression in
the fat body(Smykal et al., 2014a). A similar result was also observed in
T. castaneum (Parthasarathy et al., 2010). Depletion of JHAMT or Met caused a significant
reduction in Vg mRNA level, whereas the knockdown of Kr-h1 only caused a 30% reduction.
The downstream signal of JH appears to vary among stages and species.
4. Diploptera punctata
The Pacific beetle cockroach, D. punctata, has proven to be a valuable model insect in the study
of the dynamics of regulation of juvenile hormone (JH) biosynthesis and metabolism,
particularly during late nymph development and reproduction, as a consequence of several other
unique physiological attributes: 1) strikingly high rates of JH biosynthesis compared to other
insects, 2) maintenance of constant in vitro rates of JH biosynthesis, 3) precise and predictable
reproductive events correlated with rates of JH production, 4) uniformity among colony
members of the same age. In addition, Diploptera are easy to rear and handle in the laboratory,
their CA are easily excised and the animals demonstrate high survival rates following surgical
manipulations (Roth and Stay, 1961; Stay, 1999; Tobe and Stay, 1977). Thus, I chose D.
punctata as my study model. In this section, the life cycle and the endocrine control of
development and reproduction of D. punctata will be reviewed.
4.1 Nymphal development
On average, female D. punctata have four nymphal stages, whereas males have three;, however,
the number of nymphal molts can vary depending on conditions (Holbrook and Schal, 1998,

33
2004). Based on observations of our laboratory colony of D. punctata, stadium durations of
females are 13, 14, 16, and 21 days for the first, second, penultimate and final stages,
respectively. Diploptera exhibit sexual dimorphism, and as adults, males are smaller and more
slender than females. In contrast to 64 days of female nymphal life, males generally undergo
imaginal ecdysis sooner, within approximately 50 days, with the first, penultimate and final
stadia lasting 13, 16, and 22 days, respectively (Yagi et al., 1991).
During the growth and development of Diploptera, JH and ecdysteroids play critical roles in
determining nymphal or imaginal developmental pathways. The circulating titer of JH and
ecdysteroids in each stage are shown in Fig. 1.3A (Kikukawa and Tobe, 1986b). In the early
nymphal stadia, higher rates of JH production are found in the latter half of the stadium. In final
instars, JH release declines to undetectable rates after day 10, as hemolymph ecdysteroid titer
starts to rise (Kikukawa and Tobe, 1986a, b). The decline in JH titer and the increase in
circulating ecdysteroids in the final instars allow imaginal ecdysis to occur. During the first 8
days of the penultimate 3rd stadium, JH is critical for maintaining nymphal characters at the next
molt. At this time, reduction in the JH titer results in precocious adult metamorphosis at the next
ecdysis (Kikukawa and Tobe, 1986a). Furthermore, JH and ecdysteroids are critical to regulate
the normal duration of the stadium (Kikukawa, 1989; Kikukawa and Tobe, 1986b). In final
instars, ecdysteroid titer becomes elevated only after JH release declines. How JH interacts with
ecdysteroids to regulate the development of D. punctata was investigated by the application of
JH analog prior to day 10 of last instar females. This resulted in the desynchronization of
ecdysteroid release (Kikukawa, 1989; Kikukawa and Tobe, 1986b; Tobe et al., 1985), possibly
because high JH titer blocks the release or synthesis of ecdysteroids. However, allatectomy on
the first day of the last stadium resulted in a prolongation of low levels of ecdysteroids and the

34
low titer of JH did not promote the release of ecdysteroids (Kikukawa and Tobe, 1986b). Further
investigation is needed to determine the interaction between JH and ecdysteroids in D. punctata.
4.2 Reproduction
After emergence, adult females mate immediately while still teneral, whereas adult males must
be about a week old to successfully court and mate (Stay and Roth, 1958; Woodhead, 1986).
Mating and spermatophore transfer stimulate CA activity. Subsequently, rates of JH biosynthesis
and oocyte growth increase rapidly (Roth and Stay, 1961; Ruegg et al., 1983; Stay and Tobe,
1977). By day 2 of the gonadotrophic cycle vitellogenesis commences (Fig. 1.3B) as Vg, the
precursor of vitellin (Vn), becomes detectable in the hemolymph and fat body. At this time,
oocytes reach about 0.8mm in length and begin to accumulate Vn (Mundall et al., 1981).
Elevated JH titer stimulates this synthesis and uptake, as the denervation of virgin female CA, or
the implantation of mated female CA with ovaries into males, similarly results in Vg synthesis
and uptake by the oocytes (Mundall et al., 1979; Stay and Tobe, 1977). As rates of JH
biosynthesis rise, CA volume and cell number also increase in parallel, reaching a peak around
day 4 to 5 (Szibbo and Tobe, 1981).
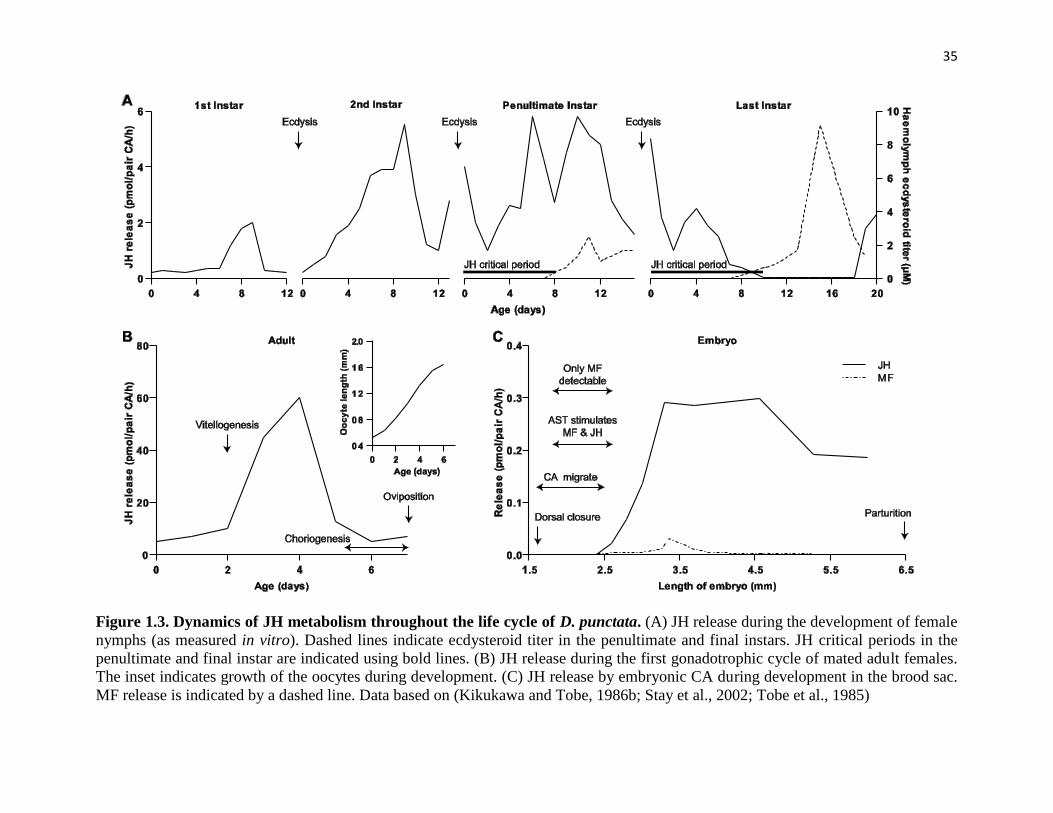
35
Figure 1.3. Dynamics of JH metabolism throughout the life cycle of D. punctata. (A) JH release during the development of female
nymphs (as measured in vitro). Dashed lines indicate ecdysteroid titer in the penultimate and final instars. JH critical periods in the
penultimate and final instar are indicated using bold lines. (B) JH release during the first gonadotrophic cycle of mated adult females.
The inset indicates growth of the oocytes during development. (C) JH release by embryonic CA during development in the brood sac.
MF release is indicated by a dashed line. Data based on (Kikukawa and Tobe, 1986b; Stay et al., 2002; Tobe et al., 1985)

36
As oocyte growth and Vn accumulation continues, rates of JH biosynthesis begin to decline
around day 5. When oocytes reach approximately 1.6mm in length, the accumulation of Vn
slows, the spaces between the follicle cells close and the cells begin to deposit the chorion (Stay
et al., 1984; Woodhead et al., 2003). Hemolymph Vg titer also declines at this time (Mundall et
al., 1981). As the oocytes approach their maximal length, 20-hydroxyecdysone becomes
detectable in the oocytes, and levels rise during chorion formation (Stay et al., 1984). Ecdysone
remains in the ovary where ecdysteroid conjugates are likely stored for use by the embryos
during development (Stay unpublished). By day 7, the rate of JH production has declined to day
1 levels (Fig. 1.3B), choriogenesis is complete, and ovulation occurs. As the basal oocytes leave
the common oviduct they are fertilized, pairs of fertilized eggs are then covered by a reduced
oothecal membrane, and finally retracted into the brood sac where embryogenesis proceeds
(Roth, 1970; Woodhead et al., 2003).
Unlike members of many other insect orders, cockroaches demonstrate a clear correlation
between cycles of JH biosynthetic activity by the CA, and patterns of oocyte growth and
vitellogenesis. Oocyte maturation, fat body vitellogenin (Vg) production and the uptake of Vg by
the ovaries are JH-dependent processes in this group (Engelmann, 1979; Rankin and Stay, 1984;
Roth and Stay, 1961; Stay and Tobe, 1978), whereas in higher orders, such as the Diptera,
endocrine regulation of oocyte development requires both JH and ecdysteroids and synthesis of
Vg occurs in both the fat body and ovary (see (Raikhel et al., 2005; Valle, 1993)).
4.3 Embryo development
Embryos, usually 12 to 14, develop within the brood sac for approximately 63 days, and grow
from 1.5mm at oviposition to 6.5mm by parturition. Dorsal closure of the body wall and

37
completion of the gut occurs relatively early, around 12 days after oviposition, when the embryos
are 1.6mm in length. At this time, the CA begin to migrate to their final dorsal position (Stay and
Coop, 1973). Early embryonic CA produce and release the JH precursor, MF, in the virtual
absence of JH, from day 16 until day 26 when embryos reach 2.6mm (Fig. 1.3C). During this
time, allatostatins (ASTs), a family of regulatory neuropeptides (see below) which inhibit JH
production in all post-embryonic developmental stages, stimulate MF and JH biosynthesis (Stay
et al., 2002). Surprisingly, this is similar to the function of these peptides in some crustaceans
(Kwok et al., 2005). The mechanism by which this occurs merits further study in Diploptera as
these finding may have important implications for the evolutionary history of these peptides
(Hult et al., 2008).
Unlike ovoviviparous blaberids, D. punctata is truly viviparous; females provide nourishment to
the embryos via nutritive milk, rich in carbohydrates and lipocalin-like proteins that is secreted
in large quantities by glandular cells in the brood sac epithelium. Secretion begins shortly after
dorsal closure. Once the gut is completely formed, the embryos start to drink the fluid and begin
to increase dramatically in dry weight (Stay and Coop, 1973; Stay and Coop, 1974; Williford et
al., 2004). Elevated levels of JH can disrupt milk production and trigger a decline in production,
but the humoral factor which promotes competence for milk production remains unidentified
(Evans and Stay, 1989, 1995; Stay and Lin, 1981). Currently, 25 Milk protein cDNAs have been
identified; these genes exist in multiple copies at several loci. Further study of this gene family is
critical, and may have implications for the evolution of viviparity in insects (Williford et al.,
2004).
Several days before parturition, a second cycle of oocyte growth begins, and re-mating is not
required as females store sperm after copulation (Mundall et al., 1981; Rankin and Stay, 1985).

38
As a consequence of viviparity Diploptera oviposit fewer oocytes, less frequently than do
oviparous species. However, nymphs are more developed at birth and reach reproductive
maturity more quickly, with fewer nymphal molts, than other blaberids (Roth, 1970; Stay and
Coop, 1973).
5 Regulation of JH titre
As a hormone which plays essential roles in insects, JH is tightly regulated to maintain the
development, metamorphosis and reproduction of insects. The circulating JH titer in the
hemolymph is determined not only by the activity of the CA, but also by other processes, such as
enzymatic degradation, binding to carrier proteins and uptake by target organs. In this section,
the factors involved in the regulation of JH titre in the CA will be summarized, mainly focusing
on the regulation in D. punctata.
CA are connected with the corpora cardiaca (CC) via nervi corporis allati (NCA) I, and in turn,
through the CC with the brain via the nervi corporis cardiaci (NCC) I and II to medial and lateral
neurosecretory cells of the brain respectively (Tobe and Stay, 1985a). JH, which is produced in
the CA, is regulated by both neural and humoral inputs, including neuropeptides,
neurotransmitters, ecdysteroids and even JH itself.
5.1 Allatostatins
ASTs are neuropeptides originally described to inhibit JH biosynthesis rapidly and reversibly in
insects. Their existence was originally hypothesized by Tobe and Stay (1980) based on the
relationship between JH biosynthesis and oocyte length. Aside from their ability to inhibit JH
biosynthesis, ASTs have also been demonstrated to suppress muscular activity, inhibit
vitellogenesis and modulate the activity of certain midgut digestive enzymes (Stay and Tobe,

39
2007). In insects, ASTs were initially grouped into three families, YXFGL-amide-allatostatins
(FLGa/ASTs), W2W9-allatostatins (MIP/ASTs) and PISCF-allatostatins (PISCF/ASTs) (Coast
and Schooley, 2011; Stay and Tobe, 2007). However, MIP/AST was originally characterized to
be a myoinhibitory peptide family, and therefore, the regulation of JH biosynthesis will be
discussed only in FLGa/ASTs and PISCF/ASTs (Coast and Schooley, 2011).
The FLGa/AST was first discovered from brain extracts of D. punctata (Pratt et al., 1989; Stay
and Tobe, 2007; Woodhead et al., 1989). In D. punctata, thirteen AST peptides are encoded by a
single neuropeptide precursor (Donly et al., 1993). Members of the FGLa family of ASTs share a
core C-terminal pentapeptide region (Tyr/Phe-Xaa-Phe-Gly-Leu/Ile-NH2) which is the main
functional region needed for inhibition of JH biosynthesis (Pratt et al., 1991; Stay et al., 1991).
Studies have demonstrated the pleiotropic nature of these peptides and their wide distribution.
ASTs have been found in many cells of the central nervous system, gut and ovary in D. punctata
(Garside et al., 2002; Stay et al., 1994b; Woodhead et al., 2003). A well-documented role for
FLGa/AST is the inhibitory effect on JH biosynthesis by the CA (Pratt et al., 1989; Stay et al.,
1994a; Woodhead et al., 1994; Woodhead et al., 1989), but they can also function as potent
inhibitors of muscle contraction in the gut (Lange et al., 1995; Stay et al., 1994b), stimulate
carbohydrate-metabolizing enzymes in the midgut and inhibit Vg production and cardiac activity
(Fuse et al., 1999; Garside et al., 2002; Martin et al., 1996; Vilaplana et al., 1999; Yu et al.,
1995b). Physiological and molecular work in D. punctata has confirmed the inhibitory function
of FGLa AST (for review see (Stay and Tobe, 2007)). This effect occurs through paracrine
release of FLGa/AST within the CA from cells in the pars lateralis of the brain sending axons in
the NCC II, to the CA (Stay et al., 1992). The expression of the FLGa/AST precursor in the
whole brain is negatively correlated with JH biosynthesis by the CA (Garside et al., 2003).

40
Furthermore, a negative correlation was also observed between sensitivity of CA to ASTs and JH
biosynthesis during the first gonadotrophic cycle of mated adult females. The sensitivity of the
CA to ASTs declines during vitellogenesis and increases again before choriogenesis occurs (Pratt
et al., 1990; Stay et al., 1991; Unnithan and Feyereisen, 1995).
Putative FLGa/AST receptors were characterised using affinity labeling and radio-ligand binding
assays in the CA and the brain (Cusson et al., 1991a, 1992a; Yu et al., 1995a). More recently, a
putative AST receptor was cloned belonging to the G-protein coupled receptor family of
mammalian galanin receptors. Its relative expression level shows a steep rise on day 6 in the CA
and the brain of the adult female, after which the level declines again. This downregulation is
consistent with the loss of sensitivity of CA to AST following ovulation (Pratt et al., 1990; Stay
et al., 1991; Unnithan and Feyereisen, 1995). The peak on day 6 suggests that AST and its
receptor may be responsible for the decline of JH biosynthesis at this time. Initial RNAi
experiments showed that silencing of the putative AST receptor results in an increase in JH
production (Lungchukiet et al., 2008a; Lungchukiet et al., 2008b). The signal transduction
pathway downstream of the FLGa/AST receptor likely involves the 1,4,5-inositol
triphosphate(IP3)/Diacylglycerol (DAG) pathway, as well as protein kinase C (PKC) (Rachinsky
et al., 1994). The IP3/DAG pathway is initiated upon binding of a neuropeptide to a G protein
bound to the membrane. Increased IP3 induces release of intracellular Ca2+, whereas DAG
activates PKC. Activators of PKC (phorbol esters) are potent inhibitors of JH III biosynthesis in
D. punctata (Feyereisen and Farnsworth, 1987b). Steps early in the JH pathway are likely the
target of AST, more specifically the export of citrate from the mitochondria or the cytosolic
conversion to acetyl-CoA, which will then enter the mevalonate pathway (Sutherland and
Feyereisen, 1996; Wang et al., 1994). However, more work is needed to further elucidate the

41
signal transduction pathway and to clearly identify the link between the AST receptor and the
exact target of this neuropeptide in JH biosynthesis.
The insect PISCF/AST was first identified in the tobacco hornworm, M. sexta (Kramer et al.,
1991). PISCF/AST is a 15 amino acid peptide with the sequence qIRYRQCYFNPISCF, with a
blocked N-terminus (pGlu), a free C-terminus, and a disulphide bridge linking two Cys residues
at positions 7 and 14. Using HPLC- MALDI-TOF, PISCF/AST was isolated and characterized in
the female mosquito A. aegypti (Li et al., 2006). AePISCF/AST inhibits JH biosynthesis in dose-
dependent manner, with maximal inhibition in the nanomolar range. This inhibitory effect can be
rescued by farnesoic acid, indicating that the target of AePISCF/AST is located before the
formation of farnesoic acid. PISCF/AST plays important roles in regulating JH biosynthesis in A.
aegypti. The levels of AePISCF/AST in the brain correlated with the sensitivity of CA to AST
and the activity of CA in JH biosynthesis (Li et al., 2006). However, in other insects, such as D.
melanogaster, PISCF/ASTs did not appear to regulate JH biosynthesis, but rather acts as a
myotropin (Price et al., 2002). In A. pisum and M. persicae, PISCF/AST shows a significant
dose-dependent feeding suppression effect, resulting in mortality, reduced growth and fecundity
(Down et al., 2010; Matthews et al., 2010).
Two PISCF/AST receptor (PISCF/AstR) genes were identified in D. melanogaster using a
reverse pharmacological approach. They are G-protein-coupled receptors, which are the insect
homologs of the mammalian opioid/somatostatin receptors. Site-directed mutagenesis studies
demonstrated that a residue in transmembrane region 3 and the loop between transmembrane
regions 6 and 7 affect ligand binding (Kreienkamp et al., 2002). Two PISCF/AstR paralogs were
also isolated in A. aegypti (Mayoral et al., 2010). The tissue distribution of the two PISCF/AstR
shows that both genes are expressed highest in the abdominal ganglia, whereas the expression

42
differs in other tissues, such as Malpighian tubules. In Malpighian tubules, PISCF/AstRB
displays high expression, whereas the transcript level of PISCF/AstRA is relatively low.
Developmental expression of PISCF/AstR mRNA also displays different patterns, which
suggests that the pleiotropic effects of PISCF/AST in mosquitoes might be mediated by the
different receptor paralogs (Mayoral et al., 2010). The PISCF/AstR identified in T. castaneum
could be activated by TcAST and T. castaneum allatostatin double C (Trica-ASTCC) as well as
M. sexta PISCF/AST in a dose-dependent manner (Audsley et al., 2013). TcPISCF/AstR is
widely distributed, with the highest transcript level in the head and the gut. Whole mount
immunocytochemistry localised TcPISCF/AstR in the median and lateral neurosecretory cells of
the brain, in the CC, throughout the ventral nerve cord and in midgut neurosecretory cells, but
not in Malpighian tubules, indicating that TcPISCF/AstR is close to PISCF/AstRA in A. aegypti.
5.2 Allatotropin
Allatotropin (AT) was first purified from the extracts of heads of pharate adult M. sexta (Kataoka
et al., 1989). This 13-residue peptide (GFKNVEMMTARGF-NH2) was shown to stimulate JH
biosynthesis in the CA of adult female M. sexta, but not in the CA of larval M. sexta, the
mealworm beetle Tenebrio molitor, the grasshopper S. nitens and the cockroach Periplaneta
americana (Kataoka et al., 1989). Further studies demonstrated that Manse-AT could also be
active on the CA in other adult insects, including the Lepidoptera Heliothis virescens (Teal,
2002), Spodoptera frugiperda (Oeh et al., 2000), and Lacanobia oleracea (Audsley et al., 1999).
In addition, Manse-AT was also found to stimulate JH biosynthesis in the larvae of L. oleracea
(Audsley et al., 2000), in Diptera Phormia regina (Tu et al., 2001) and in Orthoptera Romalea
microptera (Li et al., 2005).

43
Much work has attempted to identify other ATs. A putative AT has been extracted from the
subesophageal ganglion of male cricket G. bimaculatus. The extract was able to stimulate JH
biosynthesis in CA from G. bimaculatus and another cricket, Acheta domesticus (Lorenz and
Hoffmann, 1995). Extracts of the brain-subesophageal ganglion-CC-CA from P. apterus, also
display activity in the stimulation of JH biosynthesis (Hodkova et al., 1996). A methanolic
extract of the suboesophageal ganglia (SOG)-CC of the Mythimna loreyi virgin males stimulates
the synthesis of JH III acid and iso-JH II, whereas synthetic Manse-AT had no significant effect
(Kou and Chen, 2000). Unfortunately, ATs in G. bimaculatus or P. apterus have not been
isolated and identified. Another AT was isolated and identified from the abdominal ganglia of
the mosquito A. aegypti (Veenstra and Costes, 1999). The sequence of this peptide was
determined to be APFRNSEMMTARGF-NH2. The cDNA clone encoding this novel
neuropeptide was shown to encode a single copy of this peptide. AT in A. aegypti (refer as AaAT)
has a stimulatory effect on the JH biosynthesis of adult female CA, and it appears that the
stimulation results from the increased ability of CA to convert FA to JH III induced by AaAT (Li
et al., 2003). AaAT was also shown to induce JH biosynthesis in CA from newly emerged
females (Li et al., 2003).
In M. sexta, Manse-AT was shown to stimulate JH biosynthesis in the CA of adult females, but
not in larval or pupal CA of M. sexta. However, Manse-AT mRNA and immunoreactivity
showed the expression of Manse-AT not only in the nervous system of adults but also in that of
larvae, indicating that Manse-AT may play multiple roles in insects (Bhatt and Horodyski, 1999).
Later studies showed that in addition to regulating JH biosynthesis by the CA, AT are
multifunction in different insects, including the stimulation of myoactivity (Bhatt and Horodyski,
1999; Paemen et al., 1991; Rudwall et al., 2000), inhibition of midgut ion transport (Lee et al.,

44
1998), stimulation of foregut movement (Duve et al., 2000; Duve et al., 1999), and
cardioacceleratory effects (Koladich et al., 2002).
5.3 Neurotransmitters regulating JH biosynthesis
Several neurotransmitters are involved in the regulation of JH biosynthesis in insects.
Octopamine inhibits JH biosynthesis in the CA of D. punctata in vitro (Thompson et al., 1990).
In B. germanica, dopamine can have a stimulatory or inhibitory effect on the CA depending on
the stage of the ovarian cycle (Pastor et al., 1991). In M. sexta, dopamine stimulates JH
biosynthesis in CA from the first 2 days of the last larval stadium, but inhibits in CA from larvae
in the beginning of the prepupal period. The stimulatory or inhibitory effect of dopamine is
related to the the adenylyl cyclase system of CA (Granger et al., 1996). Ontogenetic differences
in the control of JH biosynthesis by dopamine have also been demonstrated in Drosophila
females in inhibiting or stimulating JH degradation (Gruntenko et al., 2005). The excitatory
neurotransmitter L-glutamate, which acts through ionotropic receptors to raise intracellular Ca2+
concentration, stimulates JH biosynthesis in the CA of D. punctata through the action of NMDA-,
kainate- and/or quisqualate-sensitive subtypes of ionotropic L-glutamate receptors (Chiang et al.,
2002b; Pszczolkowski et al., 1999).
5.4 Second messengers
The signal transduction pathways for neuropeptides and neurotransmitters usually involve
intracellular reaction cascades regulating levels of second messengers. The effect on JH
biosynthesis has been measured for several of these messengers.
Calcium is considered unique as a second messenger in the regulation of JH biosynthesis in D.
punctata, since intracellular Ca2+ can function as a second messenger on its own, and, at the same

45
time, Ca2+ is essential for the function of cyclic nucleotides and the IP3/DAG second messenger
system. Incubation of CA in medium lacking Ca2+, and blockage of non-specific Ca2+ channels,
inhibits JH release. Because no buildup of JH or MF occurs in the CA as a consequence of such
blockage, it seems likely that Ca2+ affects overall JH biosynthesis, instead of release (Kikukawa
et al., 1987). JH biosynthesis rises as Ca2+ concentrations are increased in vitro. Moreover,
elevated levels of extracellular Ca2+ can counteract the effect of brain extracts (containing AST)
on JH biosynthesis (Aucoin et al., 1987). Two types of Ca2+ channels have been proposed for this
action: 1) Ca2+ enters the CA cells through voltage-gated Ca2+ channels ((McQuiston et al., 1990;
Thompson and Tobe, 1986), as reviewed by (Rachinsky and Tobe, 1996)) or 2) there is a
glutamate-induced Ca2+ influx via an ionotropic L-glutamate receptor, later reported to be the
NMDA receptor (Chiang et al., 2002a; Chiang et al., 2002b; Pszczolkowski et al., 1999). Release
of Ca2+ from intracellular stores also appears to stimulate JH biosynthesis in vitro. The Ca2+-
ATPase inhibitor thapsigargin was shown to increase intracellular Ca2+ and induce JH
biosynthesis (Rachinsky and Tobe, 1996; Rachinsky et al., 1994).
Compounds that increase intracellular cAMP concentrations were found to inhibit JH
biosynthesis in an in vitro assay (Meller et al., 1985). The addition of brain extracts showed a
dose-dependent elevation of cAMP levels (Aucoin et al., 1987). Together, these results suggest
that cAMP could act as a second messenger for AST from the brain. A later study by Cusson et
al. (Cusson et al., 1992b) measured levels of cGMP and cAMP in virgin and mated females
following addition of AST. The mechanism of AST inhibition does not involve either cGMP or
cAMP, suggesting that cyclic nucleotides are likely second messengers of another inhibitory
signal affecting JH biosynthesis. A possible candidate is the neurotransmitter octopamine, which

46
in high concentrations can induce a rise in the level of cAMP in vitro (see above and (Thompson
et al., 1990)).
5.5 Ovarian factor
The normal pattern of JH biosynthesis in the CA of D. punctata during the first gonadotrophic
cycle of mated females requires the presence of the ovary (Rankin and Stay, 1984; Stay and
Tobe, 1978; Stay et al., 1983). Ovariectomy of females soon after the adult molt resulted in a low
level of JH biosynthesis throughout the first gonadotrophic cycle (Stay and Tobe, 1978). The
cyclic pattern in JH biosynthesis could be rescued following implantation into females of at least
one-half an ovary from a day 0 mated female (Stay et al., 1983). Similarly, implantation of
ovarioles with vitellogenic basal oocytes into male animals with denervated CA also resulted in
an increase in JH biosynthesis (Hass et al., 2003; Rankin and Stay, 1984).
The effect of the ovary on JH biosynthesis appears to be stage-dependent. In a study by
Sutherland et al. (2000), young ovaries were found to have a dual role: stimulation of JH
biosynthesis and repression of the mRNA levels of CYP4C7, a cytochrome P450 enzyme coding
a gene involved in JH catabolism. These roles were reversed in post-vitellogenic ovaries. Rankin
and Stay (1984) have shown that the size of the basal oocyte is a good indicator of the degree
with which the ovary can stimulate JH biosynthesis. The ovary acquires this ability at the start of
vitellogenesis but loses it post-vitellogenesis.
Currently, the exact nature of the ovarian factors involved in this modulation of JH production
remains unknown. A study by Unnithan et al. (1998) suggested that a factor produced by ovaries
in all stages can induce stable stimulation of JH synthesis. The effect of this ovarian factor is
antagonized by a factor from the brain, distinct from ASTs (Unnithan et al., 1998). In contrast, a

47
study by Elliott et al. (Elliott et al., 2006) provided evidence for a different stage-specific
peptidergic factor from the ovary that stimulates JH production. This ovarian factor is produced
when basal oocyte lengths range from 0.76 to 1.15 mm (day 2 and 3 adult females). Ovaries with
oocytes that are larger were no longer stimulatory in in vitro assays. This factor is considered the
same as that described earlier by Rankin and Stay (1984), but to date, the exact peptidergic
structure remains unknown.
An inhibitory peptidergic factor was reported from post-vitellogenic ovaries, which induced a
rise in the cGMP content of CA in vitro (Chang et al., 2005). Elevated cGMP signaling was
suggested to cause a long-term arrest on JH biosynthesis indirectly, through atrophy of the CA
cells, reduction in protein content and depletion of the cellular machinery. This factor is thought
to be the same post-vitellogenic ovary factor described by Sutherland et al. (2000) controlling the
expression of CYP4C7. However, this peptide is not likely to be AST since the quantities of AST
detected in the hemolymph are too low to exert a physiological effect on the CA (Stay et al.,
1994a). ASTs in the ovary probably act locally by facilitating ovulation or by preventing
glycosylation of Vg and its release from the fat body (Garside et al., 2002; Rankin and Stay,
1984; Woodhead et al., 2003).
5.6 JH feedback
Negative feedback regulation of an endocrine gland, induced by its own product is a well-known
endocrine control mechanism. A direct or indirect suppression of JH biosynthesis by the CA was
first shown in a study assessing the effect of a JH analog. Topical application of this analog
suppressed JH biosynthesis in a dose dependent fashion as did topical treatment with JH itself
(Tobe and Stay, 1979). A subsequent study suggested an indirect feedback of JH on the CA

48
through the brain, by comparing the effect of JH analog on males with CA connected or
disconnected with the brain (Stay et al., 1994a). In both cases, the rates of JH biosynthesis were
reduced compared to controls, intact CA more so than denervated ones. However, in animals in
which the CA were disconnected from the brain, AST levels in the hemolymph were greatly
elevated. These results indicate that the exogenous JH analog acts on the brain to inhibit JH
biosynthesis, by paracrine release of AST in the intact CA and through the hemolymph in the
disconnected CA (Ruegg et al., 1983; Stay et al., 1994a).
In vertebrates, the bulk end-product of mevalonate pathway, cholesterol, has a feedback
regulation on the transcription and post-transcription of enzymes in the mevalonate pathway,
which in turn mediates the production of cholesterol (Chang and Limanek, 1980; Clarke et al.,
1987; Goldstein and Brown, 1990). The addition of farnesol to cultured cells accelerated the
degradation of HMGR, while the addition of mevalonate raised the activity of FPPP expressed in
CHO cells (Meigs et al., 1996; Meigs and Simoni, 1997). In insects, the accumulation of JH
precursor or depletion of one enzyme in the pathway also results in a change in the activity of
other enzymes. In CA with low FALD activity, farnesal accumulated and was converted back to
farnesol that leaks from the CA, to balance the enzyme activity and the JH precursor pool
(Rivera-Perez et al., 2013). The expression and activity of enzymes in the JH biosynthetic
pathway appears to be involved in JH feedback regulation of JH titre, since the rate of JH
synthesis is regulated by the rate of flux of isoprenoids and the expression of genes in the
pathway (Details seen Section 1.2). However, further investigation is required to prove this
hypothesis.
5.7 Ecdysteroids

49
Injection of 20-hydroxyecdysone (20E) reduced oocyte growth and Vn content of basal oocytes
(Friedel et al., 1980) and inhibited JH biosynthesis in D. punctata (Stay et al., 1980). However,
no inhibitory effect was observed following treatment of CA with 20E in vitro (Paulson and
Stay, 1987). In M. sexta, incubation of brain-CC-CA complexes with 20E resulted in a reduction
in the production of JH acid (Granger and Janzen, 1987). The inhibitory effect of 20E in JH
biosynthesis may be related to dopamine: 20E down regulates dopamine, which is responsible
for the stimulation of JH biosynthesis (Granger et al., 1996). On the other hand, in D. punctata, a
significant rise in DNA synthesis in CA from male adults treated with 20E suggested that
ecdysteroids may affect JH biosynthesis by regulating proliferation of CA cells (Chiang et al.,
1995; Tsai et al., 1995). However, this conflicts with the observed 20E-induced decrease in CA
cell number of ovariectomized D. punctata females (Tobe et al., 1984). The pathway through
which ecdysteroids inhibit JH production by the CA is unclear.
6 Rationale and objectives of my study
The juvenile hormones are essential in regulating insect growth, development, metamorphosis,
aging, caste differentiation and reproduction. The important roles of JH in insect development
and reproduction and its value in pest control have led to multiple studies on the biosynthesis,
regulation and function of JH. Among cockroach families, endocrine regulatory mechanisms are
easier to discern in members of the Blaberidae. In these species, only the basal oocyte of each
ovariole undergoes vitellogenesis during a given gonadotrophic cycle, and a long period of
ovarian quiescence occurs while the oviposited fertilized eggs develop in the brood sac of the
female (Roth, 1970). Among these species, D. punctata (Dictyoptera; Blattaria; Blaberidae), the
only known truly viviparous species in that the brood sac provides nourishment to the embryos

50
(Roth, 1970), has become a model system for studies on reproduction and the regulation of JH
production.
Recently, the JH biosynthetic pathway has been established in insects such as B. mori (Kinjoh et
al., 2007), A. aegypti (Nouzova et al., 2011) and A. mellifera (Bomtorin et al., 2014). However,
there are fewer studies on the JH biosynthetic pathway in D. punctata. Thus, in Chapter 2 of this
thesis, the genes involved in the JH biosynthetic pathway were identified and characterized. This
is the first report of the characterization of the JH biosynthetic pathway in hemimetabolous
insects. The function of JH as a regulator of female cockroach reproduction, and the role of JH
biosynthetic enzymes in regulating JH biosynthesis were established. In particular, our research
revealed an interaction between the transcriptions of genes in the biosynthetic pathway i.e. a
feedback mechanism is involved in the regulation of the expression of enzymes in the JH
biosynthetic pathway.
In terms of the regulation of JH biosynthesis, ASTs have been demonstrated to be one of the
essential inhibitory factors, particularly for D. punctata, in which no AT or other allatoregulatory
neuropeptides have been discovered to date. Although the function of AST in the regulation of
JH has been well-studied, the mode of action of AST remains unclear. In Chapter 3, the mode of
action of AST was studied by determining the signaling pathway in AST action, and the target of
AST action. DpAstR was expressed in multiple vertebrate cell lines, and the activation activities
of the 13 ASTs were determined. Furthermore, my study showed that the activation of AstR
results in the elevation of intracellular calcium and cAMP, which indicates that Ca2+ and cAMP
can server as second messenger in the signaling pathway of AST action. In addition, the target of
AST action was studied using RNAi and the measurement of JH in the presence or absence of JH

51
precursors. Our result indicates that AST probably affects JH biosynthesis prior to the entry of
Acetyl-CoA into the JH biosynthetic pathway.
As a second messenger, Ca2+ has been demonstrated to modulate/regulate JH biosynthesis in D.
punctata. NMDAR is an ionotropic receptor with a high Ca2+ permeability, and its activators
glutamate and NMDA have been shown to stimulate JH biosynthesis, probably by activating the
NMDAR channel. Previous studies also showed that NMDA stimulates JH biosynthesis in vitro
in the D. punctata (Chiang et al., 2002a). Thus, it was of interest to determine the function of
NMDAR in JH biosynthesis and reproduction. In chapter 4, the genes encoding NMDAR were
identified in D. punctata and characterized in this thesis. The function of NMDAR in
reproduction was also examined by knocking down the genes encoding NMDAR subunit 2, and
by the application of NMDAR antagonist MK-801. However, neither JH biosynthesis nor oocyte
growth was affected, suggesting that NMDAR does not play important roles in the regulation of
JH biosynthesis or reproduction in female D. punctata.

52
References
Abdou, M.A., He, Q., Wen, D., Zyaan, O., Wang, J., Xu, J., Baumann, A.A., Joseph, J., Wilson, T.G., Li, S.,
2011. Drosophila Met and Gce are partially redundant in transducing juvenile hormone action. Insect Biochem
Mol Biol 41, 938-945.
Allen, K.N., Dunaway-Mariano, D., 2004. Phosphoryl group transfer: evolution of a catalytic scaffold. Trends
Biochem Sci 29, 495-503.
Aucoin, R.R., Rankin, S.M., Stay, B., Tobe, S.S., 1987. Calcium and cyclic AMP Involvement in the
regulation of juvenile hormone biosynthesis in Diploptera punctata. Insect Biochem 17, 965-969.
Audsley, N., Vandersmissen, H.P., Weaver, R., Dani, P., Matthews, J., Down, R., Vuerinckx, K., Kim, Y.J.,
Vanden Broeck, J., 2013. Characterisation and tissue distribution of the PISCF allatostatin receptor in the red
flour beetle, Tribolium castaneum. Insect Biochem Mol Biol 43, 65-74.
Audsley, N., Weaver, R.J., Edwards, J.P., 1999. Juvenile hormone synthesis by corpora allata of tomato moth
Lacanobia oleracea (Lepidoptera: Noctuidae), and the effects of allatostatins and allatotropin in vitro. . Eur J
Entomol 96, 287–293.
Audsley, N., Weaver, R.J., Edwards, J.P., 2000. Juvenile hormone biosynthesis by corpora allata of larval
tomato moth, Lacanobia oleracea, and regulation by Manduca sexta allatostatin and allatotropin. Insect
Biochem Mol Biol 30, 681-689.
Ayte, J., Gil-Gomez, G., Haro, D., Marrero, P.F., Hegardt, F.G., 1990. Rat mitochondrial and cytosolic 3-
hydroxy-3-methylglutaryl-CoA synthases are encoded by two different genes. Proc Natl Acad Sci U S A 87,
3874-3878.
Baker, F.C., Lanzrein, B., Miller, C.A., Tsai, L.W., Jamieson, G.C., Schooley, D.A., 1984. Detection of only
JH III in several life-stages of Nauphoeta cinerea and Thermobia domestica. Life Sciences 35, 1553-1560.
Baker, F.C., Lee, E., Bergot, B.J., Schooley, D.A., 1981. Isomerization of isopentenyl pyrophosphate and
homoisopentenyl pyrophosphate by Manduca sexta corpora cardiaca-corpora allata homogenates. Dev
Endocrinol 15, 67-80.
Baker, F.C., Mauchamp, B., Tsai, L.W., Schooley, D.A., 1983. Farnesol and farnesal dehydrogenase(s) in
corpora allata of the tobacco hornworm moth, Manduca sexta. J Lipid Res 24, 1586-1594.
Baker, F.C., Schooley, D.A., 1981. Biosynthesis of 3-hydroxy-3-methylglutaryl-CoA, 3-hydroxy-3-
ethylglutaryl-CoA, mevalonate and homomevalonate by insect corpus allatum and mammalian hepatic tissues.
Biochim Biophys Acta 664, 356-372.
Baker, F.C., Tsai, L.W., Reuter, C.C., Schooley, D.A., 1988. The absence of significant levels of the known
juvenile hormones and related compounds in the milkweed bug, Oncopeltus fasciatus. Insect Biochem 18,
453–462.
Beck, Y., Delaporte, C., Moras, D., Richards, G., Billas, I.M., 2009. The ligand-binding domains of the three
RXR-USP nuclear receptor types support distinct tissue and ligand specific hormonal responses in transgenic
Drosophila. Dev Biol 330, 1-11.
Belles, X., Martin, D., Piulachs, M.D., 2005. The mevalonate pathway and the synthesis of juvenile hormone
in insects. Annu Rev Entomol 50, 181-199.
Bergot, B.J., Baker, F.C., Cerf, D.C., Jamieson, G., Schooley, D.A., 1981. Qualitative and quantitative aspects
of juvenile hormone titers in developing embryos of several insect species: discovery of a new JH-like
substance extracted from eggs of Manduca sexta, in: Pratt, G.E., Brooks, G.T. (Eds.), Juvenile Hormone
Biochemistry, pp. 33-45.
Bergot, B.J., Baker, F.C., Lee, E., Schooley, D.A., 1979. Absolute configuration of homomevalonate and 3-
hydroxy-3-ethylglutaryl- and 3-hydroxy-3-methylglutaryl CoA, produced by cell-free extracts of insect

53
corpora allata; cautionary note on prediction of absolute stereochemistry based on liquid chromatographic
elution order of diastereomeric derivatives. J Am Chem Soc 101, 7432–7434.
Bhatt, T.R., Horodyski, F.M., 1999. Expression of the Manduca sexta allatotropin gene in cells of the central
and enteric nervous systems. J Comp Neurol 403, 407-420.
Bochar, D.A., Stauffacher, C.V., Rodwell, V.W., 1999. Sequence comparisons reveal two classes of 3-
hydroxy-3-methylglutaryl coenzyme A reductase. Mol Genet Metab 66, 122-127.
Bomtorin, A.D., Mackert, A., Rosa, G.C., Moda, L.M., Martins, J.R., Bitondi, M.M., Hartfelder, K., Simoes,
Z.L., 2014. Juvenile hormone biosynthesis gene expression in the corpora allata of honey bee (Apis mellifera
L.) female castes. PLoS One 9, e86923.
Borovsky, D., Carlson, D.A., Hancock, R.G., Rembold, H., van Handel, E., 1994. De novo biosynthesis of
juvenile hormone III and I by the accessory glands of the male mosquito. Insect Biochem Mol Biol 24, 437-
444.
Bowers, W.S., Marsella, P.A., Evans, P.H., 1983. Identification of an hemipteran juvenile hormone: in vitro
biosynthesis of JH III by Dysdercus fasciatus. J Exp Zool 228, 555–559.
Buesa, C., Martínez-Gonzalez, J., Casals, N., Haro, D., Piulachs, M.D., Bellés, X., Hegardt, F.G., 1994.
Blattella germanica has two HMG-CoA synthase genes. Both are regulated in the ovary during the
gonadotrophic cycle. J Biol Chem 269, 11707–11713.
Bürgin, C., Lanzrein, B., 1988. Stage dependent biosynthesis of methyl farnesoate and juvenile hormone III
and metabolism of juvenile hormone III in embryos of the cockroach, Nauphoeta cinera. . Insect Biochem 18,
3–9.
Burtenshaw, S.M., Su, P.P., Zhang, J.R., Tobe, S.S., Dayton, L., Bendena, W.G., 2008. A putative farnesoic
acid O-methyltransferase (FAMeT) orthologue in Drosophila melanogaster (CG10527): relationship to
juvenile hormone biosynthesis? Peptides 29, 242-251.
Bylemans, D., Borovsky, D., Ujvary, I., De Loof, A., 1998. Biosynthesis and regulation of Juvenile Hormone
III, Juvenile Hormone III Bisepoxide, and Methyl Farnesoate during the reproductive cycle of the grey fleshfly,
Neobellieria (Sarcophaga) bullata. Arch Insect Biochem Physiol 37, 248-256.
Cabano, J., Buesa, C., Hegardt, F.G., Marrero, P.F., 1997. Catalytic properties of recombinant 3-hydroxy-3-
methylglutaryl coenzyme A synthase-1 from Blattella germanica. Insect Biochem Mol Biol 27, 499-505.
Cao, L., Zhang, P., Grant, D.F., 2009. An insect farnesyl phosphatase homologous to the N-terminal domain of
soluble epoxide hydrolase. Biochem Biophys Res Commun 380, 188-192.
Carrigan, C.N., Poulter, C.D., 2003. Zinc is an essential cofactor for type I isopentenyl
diphosphate:dimethylallyl diphosphate isomerase. J Am Chem Soc 125, 9008-9009.
Casals, N., Buesa, C., Marrero, P.F., Belles, X., Hegardt, F.G., 2001. 3-Hydroxy-3-methylglutaryl coenzyme A
synthase-1 of Blattella germanica has structural and functional features of an active retrogene. Insect Biochem
Mol Biol 31, 425-433.
Casals, N., Buesa, C., Piulachs, M.D., Cabano, J., Marrero, P.F., Belles, X., Hegardt, F.G., 1996. Coordinated
expression and activity of 3-hydroxy-3-methylglutaryl coenzyme A synthase and reductase in the fat body of
Blattella germanica (L.) during vitellogenesis. Insect Biochem Mol Biol 26, 837-843.
Castillo-Gracia, M., Couillaud, F., 1999. Molecular cloning and tissue expression of an insect farnesyl
diphosphate synthase. Eur J Biochem 262, 365-370.
Chang, E.S., Chang, S.A., Mulder, E.P., 2001. Hormones in the lives of crustaceans: an overview. Am Zool 41,
1090–1097.
Chang, L.W., Tsai, C.M., Yang, D.M., Chiang, A.S., 2005. Cell size control by ovarian factors regulates
juvenile hormone synthesis in corpora allata of the cockroach, Diploptera punctata. Insect Biochem Mol Biol
35, 41-50.

54
Chang, T.Y., Limanek, J.S., 1980. Regulation of cytosolic acetoacetyl coenzyme A thiolase, 3-hydroxy-3-
methylglutaryl coenzyme A synthase, 3-hydroxy-3-methylglutaryl coenzyme A reductase, and mevalonate
kinase by low density lipoprotein and by 25-hydroxycholesterol in Chinese hamster ovary cells. J Biol Chem
255, 7787-7795.
Charles, J.P., Iwema, T., Epa, V.C., Takaki, K., Rynes, J., Jindra, M., 2011. Ligand-binding properties of a
juvenile hormone receptor, Methoprene-tolerant. Proc Natl Acad Sci U S A 108, 21128-21133.
Chayet, L., Pont-Lezica, R., George-Nascimento, C., Cori, O., 1973. Biosynthesis of sequiterpene alcohols and
aldehydes by cell free extracts from orange flavedo. . Phytochemistry 12, 95–101.
Chiang, A.S., Lin, W.Y., Liu, H.P., Pszczolkowski, M.A., Fu, T.F., Chiu, S.L., Holbrook, G.L., 2002a. Insect
NMDA receptors mediate juvenile hormone biosynthesis. Proc Natl Acad Sci U S A 99, 37-42.
Chiang, A.S., Pszczolkowski, M.A., Liu, H.P., Lin, S.C., 2002b. Ionotropic glutamate receptors mediate
juvenile hormone synthesis in the cockroach, Diploptera punctata. Insect Biochem Mol Biol 32, 669-678.
Chiang, A.S., Tsai, W.H., Schal, C., 1995. Neural and hormonal regulation of growth of corpora allata in the
cockroach, Diploptera punctata. Mol Cell Endocrinol 115, 51-57.
Clark, A.J., Bloch, K., 1959. The absence of sterol synthesis in insects. J Biol Chem 234, 2578–2582.
Clarke, C.F., Tanaka, R.D., Svenson, K., Wamsley, M., Fogelman, A.M., Edwards, P.A., 1987. Molecular
cloning and sequence of a cholesterol-repressible enzyme related to prenyltransferase in the isoprene
biosynthetic pathway. Mol Cell Biol 7, 3138-3146.
Coast, G.M., Schooley, D.A., 2011. Toward a consensus nomenclature for insect neuropeptides and peptide
hormones. Peptides 32, 620-631.
Consortium, T.H.B.G.S., 2006. Insights into social insects from the genome of the honeybee Apis mellifera. .
Nature 443, 931–949.
Cornforth, R.H., Fletcher, K., Hellig, H., Popjak, G., 1960. Stereospecificity of enzymic reactions involving
mevalonic acid. Nature 185, 923-924.
Couillaud, F., Feyereisen, R., 1991. Assay of HMG-CoA synthase in Diploptera punctata corpora allata. Insect
Biochem 21, 131-135.
Cusson, M., Beliveau, C., Sen, S.E., Vandermoten, S., Rutledge, R.G., Stewart, D., Francis, F., Haubruge, E.,
Rehse, P., Huggins, D.J., Dowling, A.P., Grant, G.H., 2006. Characterization and tissue-specific expression of
two lepidopteran farnesyl diphosphate synthase homologs: implications for the biosynthesis of ethyl-
substituted juvenile hormones. Proteins 65, 742-758.
Cusson, M., Prestwich, G.D., Stay, B., Tobe, S.S., 1991a. Photoaffinity labeling of allatostatin receptor
proteins in the corpora allata of the cockroach, Diploptera punctata. Biochem Biophys Res Commun 181, 736-
742.
Cusson, M., Prestwich, G.D., Stay, B., Tobe, S.S., 1992a. Evidence for allatostatin receptor Proteins in the
corpora allata of the cockroach, Diploptera punctata, as demonstrated by photoaffinity-labeling. Insect
Juvenile Hormone Research, 183-191.
Cusson, M., Yagi, K.J., Ding, Q., Duve, H., Thorpe, A., McNeil, J.N., Tobe, S.S., 1991b. Biosynthesis and
release of juvenile hormone and its precursors in insects and crustaceans: the search for a unifying arthropod
endocrinology. Insect Biochem 21, 1-6.
Cusson, M., Yagi, K.J., Guan, X.C., Tobe, S.S., 1992b. Assessment of the role of cyclic nucleotides in
allatostatin-induced inhibition of juvenile hormone biosynthesis in Diploptera punctata. Mol Cell Endocrinol
89, 121-125.
Dahm, K.H., Roller, H., Trost, B.M., 1968. The juvenile hormone. IV. Stereochemistry of juvenile hormone
and biological activity of some of its isomers and related compounds. Life Sci II 7, 129-137.

55
Daimon, T., Kozaki, T., Niwa, R., Kobayashi, I., Furuta, K., Namiki, T., Uchino, K., Banno, Y., Katsuma, S.,
Tamura, T., Mita, K., Sezutsu, H., Nakayama, M., Itoyama, K., Shimada, T., Shinoda, T., 2012. Precocious
metamorphosis in the juvenile hormone-deficient mutant of the silkworm, Bombyx mori. PLoS Genet 8,
e1002486.
Darrouzet, E., Mauchamp, B., Prestwich, G.D., Kerhoas, L., Ujvary, I., Couillaud, F., 1997. Hydroxy juvenile
hormones: new putative juvenile hormones biosynthesized by locust corpora allata in vitro. Biochem Biophys
Res Commun 240, 752-758.
de Ruyck, J., Janczak, M.W., Neti, S.S., Rothman, S.C., Schubert, H.L., Cornish, R.M., Matagne, A., Wouters,
J., Poulter, C.D., 2014. Determination of kinetics and the crystal structure of a novel type 2 isopentenyl
diphosphate: dimethylallyl diphosphate isomerase from Streptococcus pneumoniae. Chembiochem 15, 1452-
1458.
Defelipe, L.A., Dolghih, E., Roitberg, A.E., Nouzova, M., Mayoral, J.G., Noriega, F.G., Turjanski, A.G., 2011.
Juvenile hormone synthesis: "esterify then epoxidize" or "epoxidize then esterify"? Insights from the structural
characterization of juvenile hormone acid methyltransferase. Insect Biochem Mol Biol 41, 228-235.
Diaz, M.E., Mayoral, J.G., Priestap, H., Nouzova, M., Rivera-Perez, C., Noriega, F.G., 2012. Characterization
of an isopentenyl diphosphate isomerase involved in the juvenile hormone pathway in Aedes aegypti. Insect
Biochem Mol Biol 42, 751-757.
Donly, B.C., Ding, Q., Tobe, S.S., Bendena, W.G., 1993. Molecular cloning of the gene for the allatostatin
family of neuropeptides from the cockroach Diploptera punctata. Proc Natl Acad Sci U S A 90, 8807-8811.
Down, R.E., Matthews, H.J., Audsley, N., 2010. Effects of Manduca sexta allatostatin and an analog on the
pea aphid Acyrthosiphon pisum (Hemiptera: Aphididae) and degradation by enzymes from the aphid gut.
Peptides 31, 489-497.
Duportets, L., Belles, X., Rossignol, F., Couillaud, F., 2000. Molecular cloning and structural analysis of 3-
hydroxy-3-methylglutaryl coenzyme A reductase of the moth Agrotis ipsilon. Insect Mol Biol 9, 385-392.
Duve, H., Audsley, N., Weaver, R.J., Thorpe, A., 2000. Triple co-localisation of two types of allatostatin and
an allatotropin in the frontal ganglion of the lepidopteran Lacanobia oleracea (Noctuidae): innervation and
action on the foregut. Cell Tissue Res 300, 153-163.
Duve, H., East, P.D., Thorpe, A., 1999. Regulation of lepidopteran foregut movement by allatostatins and
allatotropin from the frontal ganglion. J Comp Neurol 413, 405-416.
Eigenheer, A.L., Keeling, C.I., Young, S., Tittiger, C., 2003. Comparison of gene representation in midguts
from two phytophagous insects, Bombyx mori and Ips pini, using expressed sequence tags. Gene 316, 127-136.
Eizaguirre, M., Schafellner, C., Lopez, C., Sehnal, F., 2005. Relationship between an increase of juvenile
hormone titer in early instars and the induction of diapause in fully grown larvae of Sesamia nonagrioides. J
Insect Physiol 51, 1127-1134.
Elliott, K.L., Woodhead, A.P., Stay, B., 2006. A stage-specific ovarian factor with stable stimulation of
juvenile hormone synthesis in corpora allata of the cockroach Diploptera punctata. J Insect Physiol 52, 929-
935.
Engelmann, F., 1979. Insect vitellogenin: identification, biosynthesis, and role in vitellogenesis. Adv in Insect
Phys 14, 59.
Evans, L.D., Stay, B., 1989. Humoral induction of milk synthesis in the viviparous cockroach Diploptera
punctata. Invert Reprod Dev 15, 171-176.
Evans, L.D., Stay, B., 1995. Regulation of competence for milk production in Diploptera punctata: Interaction
between mating, ovaries and the corpus allatum. Invert Reprod Dev 28, 161-170.
Faulkner, D.J., Petersen, M.R., 1971. Synthesis of C-18 Cecropia juvenile hormone to obtain optically active
forms of known absolute configuration. J Am Chem Soc 93, 3766-3767.

56
Feldlaufer, M.F., Bowers, W.S., Soderlund, D.M., Evans, P.H., 1982. Biosynthesis of the sesquiterpenoid
skeleton of juvenile hormone 3 by Dysdercus fasciatus corpora allata in vitro. . J Exp Zool 223, 295–298.
Feyereisen, R., Farnsworth, D.E., 1987a. Characterization and regulation of HMG-CoA Reductase during a
cycle of Juvenile Hormone synthesis. Mol Cell Endocrinol 53, 227-238.
Feyereisen, R., Farnsworth, D.E., 1987b. Inhibition of Insect juvenile hormone synthesis by Phorbol 12-
myristate 13-acetate. FEBS Lett 222, 345-348.
Fisher, C.W., Mayer, R.T., 1982. Characterization of house fly microsomal mixed function oxidases: inhibition
by juvenile hormone i and piperonyl butoxide. Toxicology 24, 15-31.
Friedel, T., Feyereisen, R., Mundall, E.C., Tobe, S.S., 1980. The allatostatic effect of 20-hydroxyecdysone on
the adult viviparous cockroach, Diploptera punctata. J Insect Physiol 26, 665-670.
Fuse, M., Zhang, J.R., Partridge, E., Nachman, R.J., Orchard, I., Bendena, W.G., Tobe, S.S., 1999. Effects of
an allatostatin and a myosuppressin on midgut carbohydrate enzyme activity in the cockroach Diploptera punctata. Peptides 20, 1285-1293.
Garside, C.S., Bendena, W.G., Tobe, S.S., 2003. Quantification and visualization of Dippu-AST mRNA in the
brain of adult Diploptera punctata: mated females vs. virgin females vs. males. J Insect Physiol 49, 285-291.
Garside, C.S., Koladich, P.M., Bendena, W.G., Tobe, S.S., 2002. Expression of allatostatin in the oviducts of
the cockroach Diploptera punctata. Insect Biochem Mol Biol 32, 1089-1099.
Gehring, U., Harris, J.I., 1970. The active site cysteines of thiolase. Eur J Biochem 16, 492-498.
Gertler, F.B., Chiu, C.Y., Richter-Mann, L., Chin, D.J., 1988. Developmental and metabolic regulation of the
Drosophila melanogaster 3-hydroxy-3-methylglutaryl coenzyme A reductase. Mol Cell Biol 8, 2713-2721.
Goldstein, J.L., Brown, M.S., 1990. Regulation of the mevalonate pathway. Nature 343, 425-430.
Goodfellow, R.D., Barnes, F.J., 1971. Mevalonate kinase from the larva of the fleshfly, Sarcophaga bullata.
Insect Biochem 1, 271-275.
Goodman, W.G., Cusson, M., 2012. The juvenile hormones, in: Gilbert, L.I. (Ed.), Insect Endocrinology.
Academic Press, London, p. 55.
Granger, N.A., Bollenbacher, W.E., Vince, R., Gilbert, L.I., Baehr, J.C., Dray, F., 1979. In vitro biosynthesis
of juvenile hormone by the larval corpora allata of Manduca sexta: quantificationby radioimmunoassay. Mol
Cell Endocrinol 16, 1-17.
Granger, N.A., Janzen, W.P., 1987. Inhibition of Manduca sexta corpora allata in vitro by a cerebral
allatostatic neuropeptide. Mol Cell Endocrinol 49, 237-248.
Granger, N.A., Niemiec, S.M., Gilbert, L.I., Bollenbacher, W.E., 1982. Juvenile hormone synthesis in vitro by
larval and pupal corpora allata of Manduca sexta. Mol Cell Endocrinol 28, 587-604.
Granger, N.A., Sturgis, S.L., Ebersohl, R., Geng, C., Sparks, T.C., 1996. Dopaminergic control of corpora
allata activity in the larval tobacco hornworm, Manduca sexta. Arch Insect Biochem Physiol 32, 449-466.
Grossniklaus-Burgin, C., Lanzrein, B., 1990. Qualitative and quantitative analyses of juvenile hormone and
ecdysteroids from the egg to the pupal molt in Trichoplusia ni. Arch Insect Biochem Physiol 14, 13-30.
Group, B.B.A., 2004. A draft sequence for the genome of the domesticated silkworm (Bombyx mori). Science
306, 1937-1940.
Gruntenko, N.E., Karpova, E.K., Alekseev, A.A., Chentsova, N.A., Saprykina, Z.V., Bownes, M.,
Rauschenbach, I.Y., 2005. Effects of dopamine on juvenile hormone metabolism and fitness in Drosophila
virilis. J Insect Physiol 51, 959-968.

57
Gunawardene, Y.I.N.S., Chow, B.K.C., He, J.G., Chan, S.M., 2001. The shrimp FAMeT cDNA is encoded for
a putative enzyme involved in the methylfarnesoate (MF) biosynthetic pathway and is temporally expressed in
the eyestalk of different sexes. Insect Biochem Mol Biol 31, 1115-1124.
Hall, G.M., Tittiger, C., Andrews, G.L., Mastick, G.S., Kuenzli, M., Luo, X., Seybold, S.J., Blomquist, G.J.,
2002. Midgut tissue of male pine engraver, Ips pini, synthesizes monoterpenoid pheromone component
ipsdienol de novo. Naturwissenschaften 89, 79-83.
Hammock, B.D., 1975. NADPH dependent epoxidation of methyl farnesoate to juvenile hormone in the
cockroach Blaberus giganteus L. Life Sciences 17, 323-328.
Hass, J.K., Cassias, K.A., Woodhead, A.P., Stay, B., 2003. The effect of ovary implants on juvenile hormone
production by corpora allata of male Diploptera punctata. J Insect Sci 3, 30.
Helvig, C., Koener, J.F., Unnithan, G.C., Feyereisen, R., 2004. CYP15A1, the cytochrome P450 that catalyzes
epoxidation of methyl farnesoate to juvenile hormone III in cockroach corpora allata. Proc Natl Acad Sci U S
A 101, 4024-4029.
Henrich, V.C., Szekely, A.A., Kim, S.J., Brown, N.E., Antoniewski, C., Hayden, M.A., Lepesant, J.A., Gilbert,
L.I., 1994. Expression and function of the ultraspiracle (usp) gene during development of Drosophila
melanogaster. Dev Biol 165, 38-52.
Hiruma, K., Riddiford, L.M., 1985. Hormonal regulation of dopa decarboxylase during a larval molt. Dev Biol
110, 509-513.
Hodkova, M., Okuda, T., Wagner, R., 1996. Stimulation of corpora allata by extract from neuroendocrine
complex; Comparison of reproducing and diapausing Pyrrhocoris apterus. Eur J Entomol 93, 535–543.
Holbrook, G.L., Schal, C., 1998. Social influences on nymphal development in the cockroach, Diploptera
punctata. Physiological Entomology 23, 121-130.
Holbrook, G.L., Schal, C., 2004. Maternal investment affects offspring phenotypic plasticity in a viviparous
cockroach. Proc Natl Acad Sci U S A 101, 5595-5597.
Houten, S.M., Waterham, H.R., 2001. Nonorthologous gene displacement of phosphomevalonate kinase. Mol
Genet Metab 72, 273-276.
Hult, E.F., Weadick, C.J., Chang, B.S.W., Tobe, S.S., 2008. Reconstruction of ancestral FGLamide-type insect
allatostatins: A novel approach to the study of allatostatin function and evolution. J Insect Physiol 54, 959-968.
Inoue, H., Tsuji, H., Uritani, I., 1984. Characterization and activity change of farnesol dehydrogenase in black
rot fungus-infected sweet potato. . Agric Biol Chem 48, 733–738.
Iwabuchi, K., 1995. Effect of juvenile hormone on the embryogenesis of a polyembryonic wasp, Copidosoma
floridanum, in vitro. In Vitro Cell Dev Biol Anim 31, 803-805.
Iwema, T., Billas, I.M., Beck, Y., Bonneton, F., Nierengarten, H., Chaumot, A., Richards, G., Laudet, V.,
Moras, D., 2007. Structural and functional characterization of a novel type of ligand-independent RXR-USP
receptor. EMBO J 26, 3770-3782.
Jabalquinto, A.M., Alvear, M., Cardemil, E., 1988. Physiological aspects and mechanism of action of
mevalonate 5-diphosphate decarboxylase. Comp Biochem Physiol B 90, 671-677.
Jones, G., Jones, D., Teal, P., Sapa, A., Wozniak, M., 2006. The retinoid-X receptor ortholog, ultraspiracle,
binds with nanomolar affinity to an endogenous morphogenetic ligand. FEBS J 273, 4983-4996.
Jones, G., Sharp, P.A., 1997. Ultraspiracle: An invertebrate nuclear receptor for juvenile hormones. Proc Natl
Acad Sci U S A 94, 13499-13503.
Jones, G., Wozniak, M., Chu, Y., Dhar, S., Jones, D., 2001. Juvenile hormone III-dependent conformational
changes of the nuclear receptor ultraspiracle. Insect Biochem Mol Biol 32, 33-49.

58
Judy, K.J., Schooley, D.A., Dunham, L.L., Hall, M.S., Bergot, B.J., Siddall, J.B., 1973. Isolation, Structure,
and Absolute Configuration of a New Natural Insect Juvenile Hormone from Manduca sexta. Proc Natl Acad
Sci U S A 70, 1509-1513.
Kataoka, H., Toschi, A., Li, J.P., Carney, R.L., Schooley, D.A., Kramer, S.J., 1989. Identification of an
allatotropin from adult Manduca sexta. Science 243, 1481-1483.
Kayukawa, T., Minakuchi, C., Namiki, T., Togawa, T., Yoshiyama, M., Kamimura, M., Mita, K., Imanishi, S.,
Kiuchi, M., Ishikawa, Y., Shinoda, T., 2012. Transcriptional regulation of juvenile hormone-mediated
induction of Kruppel homolog 1, a repressor of insect metamorphosis. Proc Natl Acad Sci U S A 109, 11729-
11734.
Kayukawa, T., Murata, M., Kobayashi, I., Muramatsu, D., Okada, C., Uchino, K., Sezutsu, H., Kiuchi, M.,
Tamura, T., Hiruma, K., Ishikawa, Y., Shinoda, T., 2014. Hormonal regulation and developmental role of
Kruppel homolog 1, a repressor of metamorphosis, in the silkworm Bombyx mori. Dev Biol 388, 48-56.
Keeling, C.I., Blomquist, G.J., Tittiger, C., 2004. Coordinated gene expression for pheromone biosynthesis in
the pine engraver beetle, Ips pini (Coleoptera: Scolytidae). Naturwissenschaften 91, 324-328.
Keung, W.M., 1991. Human liver alcohol dehydrogenases catalyze the oxidation of the intermediary alcohols
of the shunt pathway of mevalonate metabolism. Biochem Biophys Res Commun 174, 701-707.
Kewley, R.J., Whitelaw, M.L., Chapman-Smith, A., 2004. The mammalian basic helix-loop-helix/PAS family
of transcriptional regulators. Int J Biochem Cell Biol 36, 189-204.
Kikukawa, S., Smith, C., Tobe, S.S., 1989. Morphogenetic and gonadotrophic effects of a juvenile hormone
analogue ((7S)-hydroprene) in last instar female larvae of Diploptera punctata. Physiological Entomology 14,
165-172.
Kikukawa, S., Tobe, S.S., 1986a. Critical periods for juvenile hormone sensitivity during larval life of female
Diploptera punctata. J Insect Physiol 32, 1035-1042.
Kikukawa, S., Tobe, S.S., 1986b. Juvenile hormone biosynthesis in female larvae of Diploptera punctata and
the effect of allatectomy on hemolymph ecdysteroid titer. J Insect Physiol 32, 981-986.
Kikukawa, S., Tobe, S.S., Solowiej, S., Rankin, S.M., Stay, B., 1987. Calcium as a regulator of juvenile
hormone biosynthesis and release in the cockroach Diploptera punctata. Insect Biochem 17, 179-187.
Kinjoh, T., Kaneko, Y., Itoyama, K., Mita, K., Hiruma, K., Shinoda, T., 2007. Control of juvenile hormone
biosynthesis in Bombyx mori: Cloning of the enzymes in the mevalonate pathway and assessment of their
developmental expression in the corpora allata. Insect Biochem Mol Biol 37, 808-818.
Koladich, P.M., Cusson, M., Bendena, W.G., Tobe, S.S., McNeil, J.N., 2002. Cardioacceleratory effects of
Manduca sexta allatotropin in the true armyworm moth, Pseudaletia unipuncta. Peptides 23, 645-651.
Konopova, B., Jindra, M., 2007. Juvenile hormone resistance gene Methoprene-tolerant controls entry into
metamorphosis in the beetle Tribolium castaneum. Proc Natl Acad Sci U S A 104, 10488-10493.
Konopova, B., Smykal, V., Jindra, M., 2011. Common and distinct roles of juvenile hormone signaling genes
in metamorphosis of holometabolous and hemimetabolous insects. PLoS One 6, e28728.
Kotaki, T., 1993. Biosynthetic products by heteropteran corpora allata in vitro. Invert Reprod Dev 31, 225–230.
Kotaki, T., 1996. Evidence for a new juvenile hormone in a stink bug, Plautia stali. J Insect Physiol 42, 279–
286.
Kotaki, T., Shinada, T., Kaihara, K., Ohfune, Y., Numata, H., 2009. Structure determination of a new juvenile
hormone from a heteropteran insect. Org Lett 11, 5234-5237.
Kotaki, T., Shinada, T., Kaihara, K., Ohfune, Y., Numata, H., 2011. Biological activities of juvenile hormone
III skipped bisepoxide in last instar nymphs and adults of a stink bug, Plautia stali. J Insect Physiol 57, 147-
152.

59
Kou, R., Chen, S.J., 2000. Allatotropic activity in the suboesophageal ganglia and corpora cardiaca of the adult
male loreyi leafworm, Mythimna loreyi. Arch Insect Biochem Physiol 43, 78-86.
Koyama, T., Matsubara, M., Ogura, K., 1985. Isoprenoid enzyme systems of silkworm. I. Partial purification
of isopentenyl pyrophosphate isomerase, farnesyl pyrophosphate synthetase, and geranylgeranyl
pyrophosphate synthetase. J Biochem 98, 449-456.
Kramer, S.J., Toschi, A., Miller, C.A., Kataoka, H., Quistad, G.B., Li, J.P., Carney, R.L., Schooley, D.A., 1991.
Identification of an allatostatin from the tobacco hornworm Manduca sexta. Proc Natl Acad Sci U S A 88,
9458-9462.
Kreienkamp, H.J., Larusson, H.J., Witte, I., Roeder, T., Birgul, N., Honck, H.H., Harder, S., Ellinghausen, G.,
Buck, F., Richter, D., 2002. Functional annotation of two orphan G-protein-coupled receptors, Drostar1 and -2,
from Drosophila melanogaster and their ligands by reverse pharmacology. J Biol Chem 277, 39937-39943.
Kwok, R., Zhang, J.R., Tobe, S.S., 2005. Regulation of methyl farnesoate production by mandibular organs in
the crayfish, Procambarus clarkii: A possible role for allatostatins. J Insect Physiol 51, 367-378.
Lakshmi, M., Dutta-Gupta, A., 1990. Juvenile hormone mediated DNA synthesis during larval development of
Corcyra cephalonica (Insecta). Biochem Int 22, 269-278.
Lange, A.B., Bendena, W.G., Tobe, S.S., 1995. The effect of the 13 Dip-allatostatins on myogenic and induced
contractions of the cockroach (Diploptera punctata) hindgut. J Insect Physiol 41, 581-588.
Lanzrein, B., Imboden, H., Biirgin, C., Briining, E., H., G., 1984. On titers, origin, and functions of juvenile
hormone III, methyl farnesoate and ecdysteroids in embryonic development of the ovoviviparous cockroach,
Nauphoeta cinerea. , in: Hoffmann, J., Porchet, M. (Eds.), Biosynthesis, Metabolism and Mode of Action of
lnvertebrate Hormones Springer, pp. 454-465.
Laufer, H., Borst, D., Baker, F.C., Reuter, C.C., Tsai, L.W., Schooley, D.A., Carrasco, C., Sinkus, M., 1987.
Identification of a juvenile hormone-like compound in a crustacean. Science 235, 202-205.
Lee, K.Y., Horodyski, F.M., Chamberlin, M.E., 1998. Inhibition of midgut ion transport by allatotropin (Mas-
AT) and Manduca FLRFamides in the tobacco hornworm Manduca sexta. J Exp Biol 201, 3067-3074.
Lefevere, K.S., Lacey, M.J., Smith, P.H., Roberts, B., 1993. Identification and quantification of juvenile
hormone biosynthesized by larval and adult Australian sheep blowfly Lucilia cuprina (Diptera: Calliphoridae).
Insect Biochem Mol Biol 23, 713-720.
Lewis, M.J., Prosser, I.M., Mohib, A., Field, L.M., 2008. Cloning and characterisation of a prenyltransferase
from the aphid Myzus persicae with potential involvement in alarm pheromone biosynthesis. Insect Mol Biol
17, 437-443.
Li, M., Mead, E.A., Zhu, J., 2011. Heterodimer of two bHLH-PAS proteins mediates juvenile hormone-
induced gene expression. Proc Natl Acad Sci U S A 108, 638-643.
Li, S., Ouyang, Y.C., Ostrowski, E., Borst, D.W., 2005. Allatotropin regulation of juvenile hormone synthesis
by the corpora allata from the lubber grasshopper, Romalea microptera. Peptides 26, 63-72.
Li, Y., Hernandez-Martinez, S., Fernandez, F., Mayoral, J.G., Topalis, P., Priestap, H., Perez, M., Navare, A.,
Noriega, F.G., 2006. Biochemical, molecular, and functional characterization of PISCF-allatostatin, a regulator
of juvenile hormone biosynthesis in the mosquito Aedes aegypti. J Biol Chem 281, 34048-34055.
Li, Y., Unnithan, G.C., Veenstra, J.A., Feyereisen, R., Noriega, F.G., 2003. Stimulation of JH biosynthesis by
the corpora allata of adult female Aedes aegypti in vitro: effect of farnesoic acid and Aedes allatotropin. J Exp
Biol 206, 1825-1832.
Liang, P.H., Ko, T.P., Wang, A.H., 2002. Structure, mechanism and function of prenyltransferases. Eur J
Biochem 269, 3339-3354.
Lorenz, M.W., Hoffmann, K.H., 1995. Allatotropic activity in the subesophageal ganglia of crickets, Gryllus bimaculatus and Acheta domesticus. . J Insect Physiol 41, 191–196.

60
Lozano, J., Belles, X., 2011. Conserved repressive function of Kruppel homolog 1 on insect metamorphosis in
hemimetabolous and holometabolous species. Sci Rep 1, 163.
Lungchukiet, P., Donly, B.C., Zhang, J.R., Tobe, S.S., Bendena, W.G., 2008a. Molecular cloning and
characterization of an allatostatin-like receptor in the cockroach Diploptera punctata. Peptides 29, 276-285.
Lungchukiet, P., Zhang, J.R., Tobe, S.S., Bendena, W.G., 2008b. Quantification of allatostatin receptor mRNA
levels in the cockroach, Diploptera punctata, using real-time PCR. J Insect Physiol 54, 981-987.
Marchal, E., Hult, E.F., Huang, J., Pang, Z., Stay, B., Tobe, S.S., 2014. Methoprene-Tolerant (Met)
Knockdown in the Adult Female Cockroach, Diploptera punctata Completely Inhibits Ovarian Development.
PLoS One 9, e106737.
Marchal, E., Zhang, J.R., Badisco, L., Verlinden, H., Hult, E.F., Van Wielendaele, P., Yagi, K.J., Tobe, S.S.,
Vanden Broeck, J., 2011. Final steps in juvenile hormone biosynthesis in the desert locust, Schistocerca
gregaria. Insect Biochem Mol Biol 41, 219-227.
Martin, D., Piulachs, M.D., Belles, X., 1996. Inhibition of vitellogenin production by allatostatin in the
German cockroach. Mol Cell Endocrinol 121, 191-196.
Martinez-Gonzalez, J., Buesa, C., Piulachs, M.D., Belles, X., Hegardt, F.G., 1993. 3-Hydroxy-3-
methylglutaryl-coenzyme-A synthase from Blattella germanica. Cloning, expression, developmental pattern
and tissue expression. Eur J Biochem 217, 691-699.
Martinezgonzalez, J., Buesa, C., Piulachs, M.D., Belles, X., Hegardt, F.G., 1993. Molecular cloning,
developmental pattern and tissue expression of 3-hydroxy-3-methylglutaryl coenzyme a reductase of the
cockroach Blattella germanica. Eur J Biochem 213, 233-241.
Matthews, H.J., Down, R.E., Audsley, N., 2010. Effects of Manduca sexta allatostatin and an analogue on the
peach-potato aphid Myzus persicae (hemiptera: aphididae) and degradation by enzymes in the aphid gut. Arch
Insect Biochem Physiol 75, 139-157.
Mauchamp, B., Couillaud, F., Malosse, C., 1985. Gas chromatography-mass spectroscopy analysis of juvenile
hormone released by insect corpora allata. Anal Biochem 145, 251-256.
Mauchamp, B., Darrouzet, E., Malosse, C., Couillaud, F., 1999. 4´-OH-JH-III: an additional hydroxylated
juvenile hormone produced by locust corpora allata in vitro. . Insect Biochem Mol Biol 29, 475–480.
Mayoral, J.G., Nouzova, M., Brockhoff, A., Goodwin, M., Hernandez-Martinez, S., Richter, D., Meyerhof, W.,
Noriega, F.G., 2010. Allatostatin-C receptors in mosquitoes. Peptides 31, 442-450.
Mayoral, J.G., Nouzova, M., Navare, A., Noriega, F.G., 2009a. NADP(+)-dependent farnesol dehydrogenase,
a corpora allata enzyme involved in juvenile hormone synthesis. Proc Natl Acad Sci U S A 106, 21091-21096.
Mayoral, J.G., Nouzova, M., Yoshiyama, M., Shinod, T., Hernandez-Martinez, S., Dolghih, E., Turjanski,
A.G., Roitberg, A.E., Priestap, H., Perez, M., Mackenzie, L., Li, Y.P., Noriega, F.G., 2009b. Molecular and
functional characterization of a juvenile hormone acid methyltransferase expressed in the corpora allata of
mosquitoes. Insect Biochem Mol Biol 39, 31-37.
McQuiston, A.R., Thompson, C.S., Tobe, S.S., 1990. Voltage-dependent sodium and calcium channels of the
corpora allata of the cockroach Diploptera punctata, in: Borkovec, A.B., Masler, E.P. (Eds.), Insect
Neurochemistry and Neurophysiology. Humana Press, Clifton, pp. 329-332.
Meigs, T.E., Roseman, D.S., Simoni, R.D., 1996. Regulation of 3-hydroxy-3-methylglutaryl-coenzyme A
reductase degradation by the nonsterol mevalonate metabolite farnesol in vivo. J Biol Chem 271, 7916-7922.
Meigs, T.E., Simoni, R.D., 1997. Farnesol as a regulator of HMG-CoA reductase degradation: characterization
and role of farnesyl pyrophosphatase. Arch Biochem Biophys 345, 1-9.
Meller, V.H., Aucoin, R.R., Tobe, S.S., Feyereisen, R., 1985. Evidence for an inhibitory role of Cyclic-AMP
in the control of juvenile hormone biosynthesis by cockroach corpora allata. Mol Cell Endocrinol 43, 155-163.

61
Meyer, A.S., Hanzmann, E., Murphy, R.C., 1971. Absolute configuration of Cecropia juvenile hormone. Proc
Natl Acad Sci U S A 68, 2312-2315.
Meyer, A.S., Hanzmann, E., Schneiderman, H.A., Gilbert, L.I., Boyette, M., 1970. The isolation and
identification of the two juvenile hormones from the Cecropia silk moth. Arch Biochem Biophys 137, 190-213.
Meyer, A.S., Schneiderman, H.A., Hanzmann, E., Ko, J.H., 1968. The two juvenile hormones from the
cecropia silk moth. Proc Natl Acad Sci U S A 60, 853-860.
Minakuchi, C., Namiki, T., Shinoda, T., 2009. Kruppel homolog 1, an early juvenile hormone-response gene
downstream of Methoprene-tolerant, mediates its anti-metamorphic action in the red flour beetle Tribolium
castaneum. Dev Biol 325, 341-350.
Minakuchi, C., Namiki, T., Yoshiyama, M., Shinoda, T., 2008a. RNAi-mediated knockdown of juvenile
hormone acid O-methyltransferase gene causes precocious metamorphosis in the red flour beetle Tribolium
castaneum. Febs Journal 275, 2919-2931.
Minakuchi, C., Zhou, X., Riddiford, L.M., 2008b. Kruppel homolog 1 (Kr-h1) mediates juvenile hormone
action during metamorphosis of Drosophila melanogaster. Mech Dev 125, 91-105.
Mita, K., Kasahara, M., Sasaki, S., Nagayasu, Y., Yamada, T., Kanamori, H., Namiki, N., Kitagawa, M.,
Yamashita, H., Yasukochi, Y., Kadono-Okuda, K., Yamamoto, K., Ajimura, M., Ravikumar, G., Shimomura,
M., Nagamura, Y., Shin, I.T., Abe, H., Shimada, T., Morishita, S., Sasaki, T., 2004. The genome sequence of
silkworm, Bombyx mori. DNA Res 11, 27-35.
Miura, K., Oda, M., Makita, S., Chinzei, Y., 2005. Characterization of the Drosophila Methoprene -tolerant
gene product. Juvenile hormone binding and ligand-dependent gene regulation. FEBS J 272, 1169-1178.
Miyawaki, R., Tanaka, S.I., Numata, H., 2006. Role of juvenile hormone in the control of summer diapause in
adult Poecilocoris lewisi (Heteroptera: Scutelleridae). Formosan Entomol 26, 1–10.
Monger, D.J., Law, J.H., 1982. Control of juvenile hormone biosynthesis. Evidence for phosphorylation of the
3-hydroxy-3-methylglutaryl coenzyme A reductase of insect corpus allatum. J Biol Chem 257, 1921-1923.
Moshitzky, P., Applebaum, S.W., 1995. Pathway and regulation of JHIII-Bisepoxide biosynthesis in adult
Drosophila melanogaster corpus allatum. Arch Insect Biochem Physiol 30, 225-237.
Moshitzky, P., Miloslavski, I., Aizenshtat, Z., Applebaum, S.W., 2003. Methyl palmitate: a novel product of
the Medfly (Ceratitis capitata) corpus allatum. Insect Biochem Mol Biol 33, 1299-1306.
Mundall, E.C., Tobe, S.S., Stay, B., 1979. Induction of vitellogenin and growth of implanted oocytes in male
cockroaches. Nature 282, 97-98.
Mundall, E.C., Tobe, S.S., Stay, B., 1981. Vitellogenin fluctuations in hemolymph and fat body and dynamics
of uptake into oocytes during the reproductive cycle of Diploptera punctata. J Insect Physiol 27, 821-827.
Nagaraju, G.P.C., 2007. Is methyl farnesoate a crustacean hormone? Aquaculture 272, 39-54.
Nakanishi, K., Schooley, D.A., Koreeda, M., Dillon, J., 1971. Absolute configuration of the C18-juvenile
hormone: application of a new circular dichroism method using tris(dipivaloylmethanato) praseodymium.
Chem Commun, 1235–1236.
Niwa, R., Niimi, T., Honda, N., Yoshiyama, M., Itoyama, K., Kataoka, H., Shinoda, T., 2008. Juvenile
hormone acid O-methyltransferase in Drosophila melanogaster. Insect Biochem Mol Biol 38, 714-720.
Noriega, F.G., Ribeiro, J.M.C., Koener, J.F., Valenzuela, J.G., Hernandez-Martinez, S., Pham, V.M.,
Feyereisen, R., 2006. Comparative genomics of insect juvenile hormone biosynthesis. Insect Biochem Mol
Biol 36, 366-374.
Nouzova, M., Edwards, M.J., Mayoral, J.G., Noriega, F.G., 2011. A coordinated expression of biosynthetic
enzymes controls the flux of juvenile hormone precursors in the corpora allata of mosquitoes. Insect Biochem
Mol Biol 41, 660-669.

62
Numata, H., Numata, A., Takahashi, C., Nakagawa, Y., Iwatani, K., Takahashi, S., Miura, K., Chinzei, Y.,
1992. Juvenile hormone I is the principal juvenile hormone in a hemipteran insect, Riptortus clavatus. .
Experientia 48, 606–610.
Nyati, P., Nouzova, M., Rivera-Perez, C., Clifton, M.E., Mayoral, J.G., Noriega, F.G., 2013. Farnesyl
phosphatase, a corpora allata enzyme involved in juvenile hormone biosynthesis in Aedes aegypti. PLoS One 8.
Oda, S., Tatarazako, N., Watanabe, H., Morita, M., Iguchi, T., 2005. Production of male neonates in Daphnia
magna (Cladocera, Crustacea) exposed to juvenile hormones and their analogs. Chemosphere 61, 1168-1174.
Oeh, U., Lorenz, M.W., Dyker, H., Losel, P., Hoffmann, K.H., 2000. Interaction between Manduca sexta
allatotropin and manduca sexta allatostatin in the fall armyworm Spodoptera frugiperda. Insect Biochem Mol
Biol 30, 719-727.
Paemen, L., Tips, A., Schoofs, L., Proost, P., Van Damme, J., De Loof, A., 1991. Lom-AG-myotropin: a novel
myotropic peptide from the male accessory glands of Locusta migratoria. Peptides 12, 7-10.
Parthasarathy, R., Sun, Z., Bai, H., Palli, S.R., 2010. Juvenile hormone regulation of vitellogenin synthesis in
the red flour beetle, Tribolium castaneum. Insect Biochem Mol Biol 40, 405-414.
Parthasarathy, R., Tan, A., Palli, S.R., 2008. bHLH-PAS family transcription factor methoprene-tolerant plays
a key role in JH action in preventing the premature development of adult structures during larval-pupal
metamorphosis. Mech Dev 125, 601-616.
Pastor, D., Piulachs, M.D., Cassier, P., Andre, M., Belles, X., 1991. Invivo and invitro studies of the action of
dopamine on oocyte growth and juvenile hormone production in Blattella germanica (L) (Dictyoptera,
Blattellidae). Comptes Rendus De L Academie Des Sciences Serie Iii-Sciences De La Vie-Life Sciences 313,
207-212.
Paulson, C.R., Stay, B., 1987. Humoral inhibition of the corpora allata in larvae of Diploptera punctata - Role
of the brain and ecdysteroids. J Insect Physiol 33, 613-622.
Pratt, G.E., Farnsworth, D.E., Feyereisen, R., 1990. Changes in the sensitivity of adult cockroach corpora
allata to a brain allatostatin. Mol Cell Endocrinol 70, 185-195.
Pratt, G.E., Farnsworth, D.E., Fok, K.F., Siegel, N.R., Mccormack, A.L., Shabanowitz, J., Hunt, D.F.,
Feyereisen, R., 1991. Identity of a 2nd type of allatostatin from cockroach brains - an octadecapeptide amide
with a tyrosine-rich address sequence. Proc Natl Acad Sci U S A 88, 2412-2416.
Pratt, G.E., Farnsworth, D.E., Siegel, N.R., Fok, K.F., Feyereisen, R., 1989. Identification of an allatostatin
from adult Diploptera punctata. Biochem Biophys Res Commun 163, 1243-1247.
Price, M.D., Merte, J., Nichols, R., Koladich, P.M., Tobe, S.S., Bendena, W.G., 2002. Drosophila
melanogaster flatline encodes a myotropin orthologue to Manduca sexta allatostatin. Peptides 23, 787-794.
Pszczolkowski, M.A., Lee, W.S., Liu, H.P., Chiang, A.S., 1999. Glutamate-induced rise in cytosolic calcium
concentration stimulates in vitro rates of juvenile hormone biosynthesis in corpus allatum of Diploptera
punctata. Mol Cell Endocrinol 158, 163-171.
Rachinsky, A., Tobe, S.S., 1996. Role of second messengers in the regulation of juvenile hormone production
in insects, with particular emphasis on calcium and phosphoinositide signaling. Arch Insect Biochem Physiol
33, 259-282.
Rachinsky, A., Zhang, J., Tobe, S.S., 1994. Signal transduction in the inhibition of juvenile hormone
biosynthesis by allatostatins - roles of diacylglycerol and calcium. Mol Cell Endocrinol 105, 89-96.
Raikhel, A.S., Brown, M.R., Belles, X., 2005. Hormonal control of reproductive processes, in: Gilbert, L.I.,
Iatrou, K., Gill, S.S. (Eds.), Comprehensive molecular insect science. Elsevier, Oxford, pp. 433-491.
Ramos-Valdivia, A.C., van der Heijden, R., Verpoorte, R., 1997. Isopentenyl diphosphate isomerase: a core
enzyme in isoprenoid biosynthesis. A review of its biochemistry and function. Nat Prod Rep 14, 591-603.

63
Rankin, S.M., Stay, B., 1984. The changing effect of the ovary on rates of juvenile hormone synthesis in
Diploptera punctata. Gen Comp Endocrinol 54, 382-388.
Rankin, S.M., Stay, B., 1985. Regulation of juvenile hormone synthesis during pregnancy in the cockroach,
Diploptera punctata. J Insect Physiol 31, 145-157.
Richard, D.S., Applebaum, S.W., Gilbert, L.I., 1989a. Developmental regulation of juvenile hormone
biosynthesis by the ring gland of Drosophila melanogaster. J Comp Physiol B 159, 383-387.
Richard, D.S., Applebaum, S.W., Sliter, T.J., Baker, F.C., Schooley, D.A., Reuter, C.C., Henrich, V.C., Gilbert,
L.I., 1989b. Juvenile hormone bisepoxide biosynthesis in vitro by the ring gland of Drosophila melanogaster: a
putative juvenile hormone in the higher Diptera. Proc Natl Acad Sci U S A 86, 1421-1425.
Rivera-Perez, C., Nouzova, M., Clifton, M.E., Garcia, E.M., LeBlanc, E., Noriega, F.G., 2013. Aldehyde
dehydrogenase 3 converts farnesal into farnesoic acid in the corpora allata of mosquitoes. Insect Biochem Mol
Biol 43, 675-682.
Röller, H., Dahm, D.H., Sweeley, C.C., Trost, B.M., 1967. The structure of the juvenile hormone. Angew
Chem Int Ed 6, 179–180.
Roth, L.M., 1970. Evolution and taxonomic significance of reproduction in Blattaria. Annual Review of
Entomology 15, 75-97.
Roth, L.M., Stay, B., 1961. Oocyte development in Diploptera punctata (Eschscholtz) (Blattaria). J Insect
Physiol 7, 186-202.
Rudwall, A.J., Sliwowska, J., Nassel, D.R., 2000. Allatotropin-like neuropeptide in the cockroach abdominal
nervous system: myotropic actions, sexually dimorphic distribution and colocalization with serotonin. J Comp
Neurol 428, 159-173.
Ruegg, R.P., Lococo, D.J., Tobe, S.S., 1983. Control of corpus allatum activity in Diploptera punctata - Roles
of the pars intercerebralis and pars lateralis. Experientia 39, 1329-1334.
Sasorith, S., Billas, I.M., Iwema, T., Moras, D., Wurtz, J.M., 2002. Structure-based analysis of the ultraspiracle
protein and docking studies of putative ligands. J Insect Sci 2, 25.
Sen, S.E., Ewing, G.J., Thurston, N., 1996. Characterization of lepidopteran prenyltransferase in Manduca
sexta corpora allata. . Arch Insect Biochem Physiol 32, 315–332.
Sen, S.E., Hitchcock, J.R., Jordan, J.L., Richard, T., 2006. Juvenile hormone biosynthesis in M. sexta:
substrate specificity of insect prenyltransferase utilizing homologous diphosphate analogs. Insect Biochem
Mol Biol 36, 827-834.
Sen, S.E., Sperry, A.E., 2002. Partial purification of a farnesyl diphosphate synthase from whole-body
Manduca sexta. Insect Biochem Mol Biol 32, 889-899.
Sen, S.E., Tomasello, A., Grasso, M., Denton, R., Macor, J., Beliveau, C., Cusson, M., Crowell, D.N., 2012.
Cloning, expression and characterization of lepidopteran isopentenyl diphosphate isomerase. Insect Biochem
Mol Biol 42, 739-750.
Sen, S.E., Trobaugh, C., Beliveau, C., Richard, T., Cusson, M., 2007. Cloning, expression and characterization
of a dipteran farnesyl diphosphate synthase. Insect Biochem Mol Biol 37, 1198-1206.
Shalaby, A.M., Abd el-Ghaffar, H., el Zayat, M.A., Youssef, N.S., 1990. Effect of juvenile hormone on Musca
domestica vicina Macq. II. The gonotrophic cycle. Folia Morphol (Praha) 38, 160-168.
Shemshedini, L., Lanoue, M., Wilson, T.G., 1990. Evidence for a juvenile hormone receptor involved in
protein synthesis in Drosophila melanogaster. J Biol Chem 265, 1913-1918.
Shemshedini, L., Wilson, T.G., 1990. Resistance to juvenile hormone and an insect growth regulator in
Drosophila is associated with an altered cytosolic juvenile hormone-binding protein. Proc Natl Acad Sci U S
A 87, 2072-2076.

64
Sheng, Z., Ma, L., Cao, M.X., Jiang, R.J., Li, S., 2008. Juvenile hormone acid methyl transferase is a key
regulatory enzyme for juvenile hormone synthesis in the Eri silkworm, Samia cynthica ricini. Arch Insect
Biochem Physiol 69, 143-154.
Shin, S.W., Zou, Z., Saha, T.T., Raikhel, A.S., 2012. bHLH-PAS heterodimer of methoprene-tolerant and
Cycle mediates circadian expression of juvenile hormone-induced mosquito genes. Proc Natl Acad Sci U S A
109, 16576-16581.
Shinoda, T., Itoyama, K., 2003. Juvenile hormone acid methyltransferase: a key regulatory enzyme for insect
metamorphosis. Proc Natl Acad Sci U S A 100, 11986-11991.
Smykal, V., Bajgar, A., Provaznik, J., Fexova, S., Buricova, M., Takaki, K., Hodkova, M., Jindra, M., Dolezel,
D., 2014a. Juvenile hormone signaling during reproduction and development of the linden bug, Pyrrhocoris apterus. Insect Biochem Mol Biol 45, 69-76.
Smykal, V., Daimon, T., Kayukawa, T., Takaki, K., Shinoda, T., Jindra, M., 2014b. Importance of juvenile
hormone signaling arises with competence of insect larvae to metamorphose. Dev Biol 390, 221-230.
Song, J., Wu, Z., Wang, Z., Deng, S., Zhou, S., 2014. Kruppel-homolog 1 mediates juvenile hormone action to
promote vitellogenesis and oocyte maturation in the migratory locust. Insect Biochem Mol Biol 52, 94-101.
Sperry, A.E., Sen, S.E., 2001. Farnesol oxidation in insects: evidence that the biosynthesis of insect juvenile
hormone is mediated by a specific alcohol oxidase. Insect Biochem Mol Biol 31, 171-178.
Spradling, A.C., Stern, D., Beaton, A., Rhem, E.J., Laverty, T., Mozden, N., Misra, S., Rubin, G., 1999. The
Berkely Drosophila Genome Project gene disruption project: single P-element insertions mutating 25% of vital
Drosophila genes. . Genetics 153, 135–177.
Stay, B., 1999. Diploptera punctata, in: Huber, I., Masler, E.P., Rao, B.R. (Eds.), Cockroaches as models for
neurobiology: Applications in biomedical research CRC press, Boca Raton, FL., pp. 880-889.
Stay, B., Bachmann, J.A.S., Stoltzman, C.A., Fairbairn, S.E., Yu, C.G., Tobe, S.S., 1994a. Factors affecting
allatostatin release in a cockroach (Diploptera punctata) - Nerve section, juvenile hormone analog and ovary. J
Insect Physiol 40, 365-372.
Stay, B., Chan, K.K., Woodhead, A.P., 1992. Allatostatin-immunoreactive neurons projecting to the corpora
allata of adult Diploptera punctata. Cell Tissue Res 270, 15-23.
Stay, B., Coop, A., 1973. Developmental stages and chemical composition in embryos of cockroach,
Diploptera punctata, with observations on effect of diet. J Insect Physiol 19, 147-171.
Stay, B., Coop, A.C., 1974. Milk Secretion for Embryogenesis in a Viviparous Cockroach. Tissue & Cell 6,
669-693.
Stay, B., Friedel, T., Tobe, S.S., Mundall, E.C., 1980. Feedback control of juvenile hormone synthesis in
cockroaches - Possible role for ecdysterone. Science 207, 898-900.
Stay, B., Joshi, S., Woodhead, A.P., 1991. Sensitivity to allatostatins of corpora allata from larval and adult
female Diploptera punctata. Journal of Insect Physiology 37, 63-70.
Stay, B., Lin, H.L., 1981. The inhibition of milk synthesis by juvenile hormone in the viviparous cockroach,
Diploptera punctata. J Insect Physiol 27, 551-&.
Stay, B., Ostedgaard, L.S., Tobe, S.S., Strambi, A., Spaziani, E., 1984. Ovarian and hemolymph titers of
ecdysteroid during the gonadotropic cycle in Diploptera punctata. J Insect Physiol 30, 643-651.
Stay, B., Roth, L.M., 1958. The reproductive behavior of Diploptera punctata (Blattaria: Diplopteridae).
Proceedings 10th international congress of entomology 2, 5.
Stay, B., Tobe, S.S., 1977. Control of juvenile hormone biosynthesis during the reproductive cycle of a
viviparous cockroach 1. Activation and inhibition of corpora allata. Gen Comp Endocrinol 33, 531-540.

65
Stay, B., Tobe, S.S., 1978. Control of juvenile hormone biosynthesis during reproductive cycle of a viviparous
cockroach 2. Effects of unilateral allatectomy, implantation of supernumerary corpora allata, and ovariectomy.
Gen Comp Endocrinol 34, 276-286.
Stay, B., Tobe, S.S., 2007. The role of allatostatins in juvenile hormone synthesis in insects and crustaceans.
Annual Review of Entomology 52, 277-299.
Stay, B., Tobe, S.S., Bendena, W.G., 1994b. Allatostatins - identification, primary structures, functions and
distribution. Advances in Insect Physiology, Vol 25 25, 267-337.
Stay, B., Tobe, S.S., Mundall, E.C., Rankin, S., 1983. Ovarian stimulation of juvenile hormone biosynthesis in
the viviparous cockroach, Diploptera punctata. Gen Comp Endocrinol 52, 341-349.
Stay, B., Zhang, J.R., Tobe, S.S., 2002. Methyl farnesoate and juvenile hormone production in embryos of
Diploptera punctata in relation to innervation of corpora allata and their sensitivity to allatostatin. Peptides 23,
1981-1990.
Steiner, B., Pfister-Wilhelm, R., Grossniklaus-Burgin, C., Rembold, H., Treiblmayr, K., Lanzrein, B., 1999.
Titres of juvenile hormone I, II and III in Spodoptera littoralis (Noctuidae) from the egg to the pupal moult and
their modification by the egg-larval parasitoid Chelonus inanitus (Braconidae). J Insect Physiol 45, 401-413.
Sun, X.F., Li, Z.X., 2012. In silico and in vitro analyses identified three amino acid residues critical to the
catalysis of two aphid farnesyl diphosphate synthase. Protein J 31, 417-424.
Sutherland, J.D., Kozlova, T., Tzertzinis, G., Kafatos, F.C., 1995. Drosophila hormone receptor 38: a second
partner for Drosophila USP suggests an unexpected role for nuclear receptors of the nerve growth factor-
induced protein B type. Proc Natl Acad Sci U S A 92, 7966-7970.
Sutherland, T.D., Feyereisen, R., 1996. Target of cockroach allatostatin in the pathway of juvenile hormone
biosynthesis. Mol Cell Endocrinol 120, 115-123.
Sutherland, T.D., Unnithan, G.C., Feyereisen, R., 2000. Terpenoid omega-hydroxylase (CYP4C7) messenger
RNA levels in the corpora allata: a marker for ovarian control of juvenile hormone synthesis in Diploptera
punctata. J Insect Physiol 46, 1219-1227.
Szibbo, C.M., Tobe, S.S., 1981. Cellular and volumetric changes in relation to the activity cycle in the corpora
allata of Diploptera punctata. J Insect Physiol 27, 655-665.
Taban, A.H., Tittiger, C., Blomquist, G.J., Welch, W.H., 2009. Isolation and characterization of farnesyl
diphosphate synthase from the cotton boll weevil, Anthonomus grandis. Arch Insect Biochem Physiol 71, 88-
104.
Teal, P.E., 2002. Effects of allatotropin and allatostatin on in vitro production of juvenile hormones by the
corpora allata of virgin females of the moths of Heliothis virescens and Manduca sexta. Peptides 23, 663-669.
Teal, P.E., Jones, D., Jones, G., Torto, B., Nyasembe, V., Borgemeister, C., Alborn, H.T., Kaplan, F., Boucias,
D., Lietze, V.U., 2014. Identification of methyl farnesoate from the hemolymph of insects. J Nat Prod 77, 402-
405.
Thompson, C.S., Tobe, S.S., 1986. Electrical properties of membranes and cells of the corpora allata of the
cockroach Diploptera punctata: evidence for the presence of voltage-sensitive calcium channels., in: Borkovec,
A.B., Gelman, D.B. (Eds.), Insect Neurochemistry and Neurophysiology. Humana Press, Totowa, pp. 375-378.
Thompson, C.S., Yagi, K.J., Chen, Z.F., Tobe, S.S., 1990. The effects of octopamine on juvenile hormone
biosynthesis, electrophysiology, and cAMP content of the corpora allata of the cockroach Diploptera punctata.
Journal of Comparative Physiology B-Biochemical Systemic and Environmental Physiology 160, 241-249.
Tittiger, C., Barkawi, L.S., Bengoa, C.S., Blomquist, G.J., Seybold, S.J., 2003. Structure and juvenile
hormone-mediated regulation of the HMG-CoA reductase gene from the Jeffrey pine beetle, Dendroctonus
jeffreyi. Mol Cell Endocrinol 199, 11-21.

66
Tittiger, C., Blomquist, G.J., Ivarsson, P., Borgeson, C.E., Seybold, S.J., 1999. Juvenile hormone regulation of
HMG-R gene expression in the bark beetle Ips paraconfusus (Coleoptera: Scolytidae): implications for male
aggregation pheromone biosynthesis. Cell Mol Life Sci 55, 121-127.
Tittiger, C., O'Keeffe, C., Bengoa, C.S., Barkawi, L.S., Seybold, S.J., Blomquist, G.J., 2000. Isolation and
endocrine regulation of an HMG-CoA synthase cDNA from the male Jeffrey pine beetle, Dendroctonus
jeffreyi (Coleoptera: Scolytidae). Insect Biochem Mol Biol 30, 1203-1211.
Tobe, S.S., Clarke, N., Stay, B., Ruegg, R.P., 1984. Changes in cell number and activity of the corpora allata
of the cockroach Diploptera punctata - a role for mating and the ovary. Canadian Journal of Zoology-Revue
Canadienne De Zoologie 62, 2178-2182.
Tobe, S.S., Ruegg, R.P., Stay, B.A., Baker, F.C., Miller, C.A., Schooley, D.A., 1985. Juvenile hormone titer
and regulation in the cockroach Diploptera punctata. Experientia 41, 1028-1034.
Tobe, S.S., Stay, B., 1977. Corpus allatum activity in vitro during reproductive cycle of viviparous cockroach,
Diploptera punctata (Eschscholtz). Gen Comp Endocrinol 31, 138-147.
Tobe, S.S., Stay, B., 1979. Modulation of juvenile hormone synthesis by an analog in the cockroach. Nature
281, 481-482.
Tobe, S.S., Stay, B., 1980. Control of juvenile hormone biosynthesis during the reproductive cycle of a
viviparous cockroach. III. Effects of denervation and age on compensation with unilateral allatectomy and
supernumerary corpora allata. Gen Comp Endocrinol 40, 89-98.
Tobe, S.S., Stay, B., 1985a. Structure and regulation of the corpus allatum. Advances in Insect Physiology 18,
305-432.
Tobe, S.S., Stay, B., 1985b. Structure and regulation of the corpus allatum. Advances in Insect Physiology 18,
305-432.
Tsai, W.H., Holbrook, G.L., Schal, C., Chiang, A.S., 1995. In vitro growth of corpora allata from Diploptera punctata. In Vitro Cellular & Developmental Biology-Animal 31, 542-546.
Tu, M., Kou, R., Wang, Z., Stoffolano, J.G., Jr., Yin, C., 2001. Immunolocalization and possible effect of a
moth allatotropin-like substance in a fly, Phormia regina (Diptera: Calliphoridae). J Insect Physiol 47, 233-
244.
Unnithan, G.C., Feyereisen, R., 1995. Experimental acquisition and loss of allatostatin sensitivity by corpora
allata of Diploptera punctata. J Insect Physiol 41, 975-980.
Unnithan, G.C., Sutherland, T.D., Cromey, D.W., Feyereisen, R., 1998. A factor causing stable stimulation of
juvenile hormone synthesis by Diploptera punctata corpora allata in vitro. J Insect Physiol 44, 1027-1037.
Valle, D., 1993. Vitellogenesis in insects and other groups--a review. Mem Inst Oswaldo Cruz 88, 1-26.
Vandermoten, S., Haubruge, E., Cusson, M., 2009a. New insights into short-chain prenyltransferases:
structural features, evolutionary history and potential for selective inhibition. Cell Mol Life Sci 66, 3685-3695.
Vandermoten, S., Santini, S., Haubruge, E., Heuze, F., Francis, F., Brasseur, R., Cusson, M., Charloteaux, B.,
2009b. Structural features conferring dual geranyl/farnesyl diphosphate synthase activity to an aphid
prenyltransferase. Insect Biochem Mol Biol 39, 707-716.
Veenstra, J.A., Costes, L., 1999. Isolation and identification of a peptide and its cDNA from the mosquito
Aedes aegypti related to Manduca sexta allatotropin. Peptides 20, 1145-1151.
Vilaplana, L., Maestro, J.L., Piulachs, M.D., Belles, X., 1999. Modulation of cardiac rhythm by allatostatins in
the cockroach Blattella germanica (L.) (Dictyoptera, Blattellidae). J Insect Physiol 45, 1057-1064.
Wang, Z.W., Ding, Q., Yagi, K.J., Tobe, S.S., 1994. Terminal stages in juvenile hormone biosynthesis in
corpora allata of Diploptera punctata - developmental changes in enzyme activity and regulation by
allatostatins. J Insect Physiol 40, 217-223.

67
Wigglesworth, V.B., 1934. The physiology of ecdysis in Rhodnius prolixus (Hemiptera). II. Factors controlling
molting and ‘metamorphosis’. Quart. J Microsc Sci s2–77, 191–222.
Williams, C.M., 1956. The juvenile hormone of insects. Nature 178, 212–213.
Williford, A., Stay, B., Bhattacharya, D., 2004. Evolution of a novel function: nutritive milk in the viviparous
cockroach, Diploptera punctata. Evo Dev 6, 67-77.
Wilson, T.G., Ashok, M., 1998. Insecticide resistance resulting from an absence of target-site gene product.
Proc Natl Acad Sci U S A 95, 14040-14044.
Wilson, T.G., Fabian, J., 1986. A Drosophila melanogaster mutant resistant to a chemical analog of juvenile
hormone. Dev Biol 118, 190-201.
Woodhead, A.P., 1986. Male age - Effect on mating behavior and success in the cockroach Diploptera punctata. Animal Behaviour 34, 1874-1879.
Woodhead, A.P., Khan, M.A., Stay, B., Tobe, S.S., 1994. 2 new allatostatins from the brains of Diploptera
punctata. Insect Biochem Mol Biol 24, 257-263.
Woodhead, A.P., Stay, B., Seidel, S.L., Khan, M.A., Tobe, S.S., 1989. Primary structure of 4 allatostatins -
neuropeptide inhibitors of juvenile hormone synthesis. Proc Natl Acad Sci U S A 86, 5997-6001.
Woodhead, A.P., Thompson, M.E., Chan, K.K., Stay, B., 2003. Allatostatin in ovaries, oviducts, and young
embryos in the cockroach Diploptera punctata. J Insect Physiol 49, 1103-1114.
Yagi, K.J., Konz, K.G., Stay, B., Tobe, S.S., 1991. Production and utilization of farnesoic acid in the juvenile
hormone biosynthetic pathway by corpora allata of larval Diploptera punctata. Gen Comp Endocrinol 81, 284-
294.
Yao, T.P., Segraves, W.A., Oro, A.E., McKeown, M., Evans, R.M., 1992. Drosophila ultraspiracle modulates
ecdysone receptor function via heterodimer formation. Cell 71, 63-72.
Yin, C.M., Zou, B.X., Jiang, M.G., Li, M.F., Qin, W.H., Potter, T.L., Stoffolano, J.G., 1995. Identification of
juvenile hormone III bisepoxide (JHB3), juvenile hormone III and methyl farnesoate secreted by the corpus
allatum of Phormia regina (Meigen), in vitro and function of JHB3. Either applied alone or as a part of a
juvenoid blend. J Insect Physiol 41, 473-479.
Yu, C.G., Hayes, T.K., Strey, A., Bendena, W.G., Tobe, S.S., 1995a. Identification and partial characterization
of receptors for allatostatins in brain and corpora allata of the cockroach Diploptera punctata using a binding
assay and photoaffinity labeling. Regul Pept 57, 347-358.
Yu, C.G., Stay, B., Ding, Q., Bendena, W.G., Tobe, S.S., 1995b. Immunochemical identification and
expression of allatostatins in the gut of Diploptera punctata. J Insect Physiol 41, 1035-1043.
Zhang, Y.L., Li, Z.X., 2008. Two different farnesyl diphosphate synthase genes exist in the genome of the
green peach aphid, Myzus persicae. Genome 51, 501-510.
Zhang, Z., Xu, J., Sheng, Z., Sui, Y., Palli, S.R., 2011. Steroid receptor co-activator is required for juvenile
hormone signal transduction through a bHLH-PAS transcription factor, methoprene tolerant. J Biol Chem 286,
8437-8447.

68
Chapter 2
Characterization of the Juvenile Hormone pathway in the
viviparous cockroach, Diploptera punctata
This chapter is an adapted reprint of my article:
Juan Huang, Elisabeth Marchal, Ekaterina F. Hult, and Stephen S. Tobe, Characterization of the
Juvenile Hormone pathway in the viviparous cockroach, Diploptera punctata. PloS one,
accepted.
Authors’ contribution:
E. Marchal performed part of the sequencing of the JH biosynthetic genes, and the tissue
distribution and expression profiles. I performed the RNAi and RCA. The paper was written by
E. Marchal and me with editing by E.F. Hult and S.S. Tobe

69
Summary
Juvenile hormones (JHs) are key regulators of insect development and reproduction. The JH
biosynthetic pathway is known to involve 13 discrete enzymatic steps. In the present study, we
have characterized the JH biosynthetic pathway in the cockroach Diploptera punctata. The effect
of exogenous JH precursors on JH biosynthesis was also determined. Based on sequence
similarity, orthologs for the genes directly involved in the pathway were cloned, and their spatial
and temporal transcript profiles were determined. The effect of shutting down the JH pathway in
adult female cockroaches was studied by knocking down genes encoding HMG-CoA reductase
(HMGR) and Juvenile hormone acid methyltransferase (JHAMT). As a result, oocyte
development slowed as a consequence of reduction in JH biosynthesis. Oocyte length, fat body
transcription of Vg and ovarian vitellin content significantly decreased. In addition, silencing
HMGR and JHAMT resulted in a decrease in the transcript levels of other genes in the pathway.
Introduction
Juvenile hormones (JHs) play key roles in regulating growth, development, metamorphosis,
aging, caste differentiation and reproduction in insects (as reviewed by Goodman and Cusson,
2012; Hartfelder, 2000). The multiple processes in which JH is involved and the critical role
which JH plays in metamorphosis and reproduction emphasize the importance of elucidating the
JH biosynthetic pathway and the factors that regulate its biosynthesis.
JHs are sesquiterpenoid compounds that are synthesized and secreted by specialized, paired
endocrine glands, the corpora allata (CA). The complete biosynthetic pathway of JH III (the most
widespread and predominant JH homologue in insects) comprises 13 discrete enzymatic steps.
This pathway can be divided into two metabolic parts (Fig. 2.1): the early portion comprises the

70
mevalonate pathway to the formation of farnesyl diphosphate (FPP) and is conserved in both
vertebrates and invertebrates; the later part of the pathway is specific to insects and other
arthropods. In this later part, FPP is cleaved to farnesol, which is then oxidized to the carboxylic
acid (farnesoic acid; FA), followed by methyl esterification, epoxidation and formation of JH
(Belles et al., 2005). The order in which these two final steps in JH biosynthesis occurs, appears
to be insect order dependent. In orthopteran, coleopteran, dipteran and dictyopteran insects, FA
is first methylated to MF, which in turn undergoes a C10, C11 epoxidation to the functional JH.
In Lepidoptera, however, the opposite situation prevails: epoxidation precedes methylation
(Defelipe et al., 2011; Marchal et al., 2011; Shinoda and Itoyama, 2003).
Recent studies have reported on the molecular elucidation of the JH pathway in several
holometabolous insects such as the silkworm Bombyx mori, the mosquito, Aedes aegypti and the
honeybee, Apis mellifera. In B. mori, all genes encoding enzymes involved in the mevalonate
pathway (Kinjoh et al., 2007) and the isoprenoid branch of JH biosynthesis (Cheng et al., 2014;
Shinoda and Itoyama, 2003) have been isolated. Each enzyme in the mevalonate pathway is
encoded by a single gene, except farnesyl diphosphate synthase (FPPS), which comprises three
homologs. The genes encoding enzymes in the isoprenoid branch of JH biosynthesis, however,
underwent gene duplication to create multiple copies. The transcripts for most JH enzymes are
highly enriched or exclusively expressed in the CA (Ueda et al., 2009). The expression pattern of
the genes encoding these enzymes in the CA correlates well with rates of JH biosynthesis
(Kinjoh et al., 2007). In A. aegypti, changes in the transcription of 11 of the enzymes are
responsible in part for the dynamic changes in JH biosynthesis (Nouzova et al., 2011). The
expression of genes in the JH biosynthetic pathway was also determined in female castes of A.
mellifera and was found to correlate with the JH hemolymph titre in adult worker bees, but, not

71
Fig. 2.1. Scheme of JH biosynthetic pathway (Adapted from Belles et al. (2005) and Nouzova
et al. (2011)). The Insect- and Arthropod-specific pathway is represented in the dashed box.
Precursors are in bold and connected by arrows. Enzymes are in italics. Abbreviations for the
enzymes are given in brackets.
Acetyl-CoA
Acetoacetyl-CoA
HMG-CoA
Mevalonic acid
Phosphomevalonate
Diphosphomevalonate
Isopentenyl diphosphate Dimethylallyl diphosphate
Geranyl diphosphate
Farnesyl diphosphate
Farnesol
Farnesal
Farnesoic acid
Methyl farnesoate
Juvenile hormone III
Acetoacetyl-CoA thiolase (1- Thiol)
HMG-CoA synthase (2- HMGS)
HMG-CoA reductase (3- HMGR)
Mevalonate kinase (4- MK)
Phosphomevalonate kinase (5- PMK)
Diphosphomevalonate decarboxylase (6- PPMD)
Isophentenyl diphosphate isomerase (7- IPPI)
Farnesyl diphosphate synthase (8- FPPS)
Farnesyl diphosphate synthase (8- FPPS)
Farnesyl diphosphate pyrophosphatase (9- FPPP)
Farnesol dehydrogenase (10 – FOLD)
Farnesal dehydrogenase (11 – FALD)
Juvenile hormone acid methytransferase (12- JHAMT)
Methyl farnesoate epoxidase (13- CYP15A1)

72
in larvae (Bomtorin et al., 2014). Until recently, no structural or molecular data were available
on FPP pyrophosphatase (FPPP) or farnesal dehydrogenase (FALD). Current studies in the
dipterans, the fly, Drosophila melanogaster and A. aegypti have characterized the gene encoding
an FPP phosphatase (FPPP) (Cao et al., 2009; Nyati et al., 2013). Moreover, genes encoding an
aldehyde dehydrogenase (FALD) have now been functionally characterized in A. aegypti
(Rivera-Perez et al., 2013).
Diploptera punctata, the only truly viviparous cockroach is a well-known model system in the
study of JH biosynthesis and its regulation. The physiology of this animal is characterized by
very stable and high rates of JH biosynthesis and precise and predictable reproductive events that
correlate well with rates of JH production (see review by Marchal et al. (2013a)). In adult
females, JH regulates oocyte maturation, fat body vitellogenin (Vg) production and the uptake of
Vg by the developing oocytes (Rankin and Stay, 1984; Stay and Tobe, 1978). Aside from the
original molecular identification of CYP15A1 (Helvig et al., 2004), the gene encoding the
epoxidase producing the functional JH, JH-related research in D. punctata has mainly focused on
examining JH titre and physiological aspects of JH function (see Marchal et al. (2013a)).
An earlier study predicted 4 genes encoding enzymes in JH biosynthetic pathway in D. punctata
based on sequence similarity with Drosophila and Anopheles gambiae genes (Noriega et al.,
2006). To further characterize the JH biosynthetic pathway in this important model system on a
molecular level, we have confirmed and identified 11 out of 13 genes encoding the JH
biosynthetic enzymes, and have also analyzed the tissue distribution and developmental
transcript profile of these genes during the first gonadotropic cycle of the female cockroach. The
predominant expression of these genes in the CA and the correlation between their transcript
levels and the rates of JH biosynthesis suggests that the 11 genes cloned in our study are

73
encoding functional enzymes. In addition, the transcript levels of the 11 genes indicate that the
expression of these genes partly regulate JH biosynthesis. Addition of exogenous JH precursors
was able to stimulate JH biosynthesis in CA with low activity suggesting that JH production was
also regulated by flux of substrates in the pathway.
We also used RNA interference (RNAi) to study the effect of silencing genes in the pathway on
rates of JH biosynthesis and ovarian development. HMGR and JHAMT were chosen as the target
genes for the RNAi, because both are well-studied genes whose expression patterns clearly
correlate with rates of JH biosynthesis (Goodman and Cusson, 2012). Our results show that JH
biosynthesis decreased and ovarian development slowed in HMGR-JHAMT dsRNA treated
animals. Of particular interest is our discovery that manipulation of individual genes encoding
the JH biosynthetic enzymes using dsRNA technology resulted in a significant decrease in the
transcript levels of other genes, which indicates a feedback mechanism is involved in the
regulation of the expression of enzymes in the JH biosynthetic pathway.
Materials and methods
Animals - The D. punctata colony was maintained at 27 °C in constant darkness and animals
were fed lab chow and water at libitum. To obtain pools of synchronised animals, newly molted
female adult cockroaches were picked from the colony, placed in separate containers and
provided with water and lab chow. Mated status was confirmed by the presence of a
spermatophore.
Tissue collection and RNA extraction - D. punctata were dissected in modified cockroach ringer
solution (150 mM NaCl, 12 mM KCl, 10 mM CaCl2.2H2O, 3 mM MgCl2.6H2O, 10 mM HEPES,
40 mM glucose, pH 7.2) using a dissecting microscope. Samples were flash-frozen in liquid N2

74
to prevent RNA degradation and were stored at -80 ºC until further processing. For each
dissected female, basal oocyte lengths were measured to determine the physiological age. CA
samples were taken from day 0 to 7 of adult females for the developmental profiles of genes
encoding enzymes in the JH biosynthetic pathway. For each time point, three biologically
independent pools of 10 animals each were collected. To determine the tissue distribution of the
genes of interest, the following tissues were dissected from 3 independent pools of 10 animals
each: brain (Br), nerve cord (NC), corpora allata (CA), fat body (FB), midgut (MG), Malpighian
tubules (MT), ovary (Ov) from females, and accessory gland (AG) and testes (Te) from males.
For the RNAi experiments (§2.6), 3 biologically independent pools comprising CA from 7
animals were collected. Pooled samples were homogenised with RNase-free pestles and total
RNA was extracted using the RNeasy Mini Kit (Qiagen) according to the manufacturer’s
instructions. An additional DNase treatment (RNase-free DNase set, Qiagen) was performed to
eliminate potential genomic DNA contamination. Because of the small size of the CA, RNA
from this tissue was extracted using the RNAqueous®-Micro Kit (Ambion), followed by the
recommended DNase step. Quality and concentration of the resulting RNA samples were
measured using a Nanodrop spectrophotometer (Thermo Scientific.).
Sequencing genes involved in the JH biosynthetic pathway - Since no genome or full
transcriptome sequence data are currently available for D. punctata, a first set of degenerate
primers was developed based on a multiple sequence alignment of known orthologous sequences
from different insects. Such an alignment was made for Acetoacetyl-CoA thiolase (Thiol),
HMG-CoA reductase (HMGR), Phosphomevalonate kinase (PK), Diphosphomevalonate
decarboxylase (PPMD), Isopentenyl diphosphate isomerase (IPPI), Juvenile hormone acid
methyltransferase (JHAMT) and the JH target vitellogenin (Vg). Degenerate primers are listed in

75
Supplementary table S1. For Methyl farnesoate epoxidase (CYP15A1) the full sequence was
already characterised (GenBank accession number AY509244) (Helvig et al., 2004). For HMG-
CoA synthase (HMGS), Mevalonate kinase (MK), Farnesyl diphosphate synthase (FPPS) and
Farnesol dehydrogenase (FOLD), a partial sequence was found in the D. punctata CA EST
database (Noriega et al., 2006). Their sequence was partially confirmed using gene-specific
primers (Table S1). A temperature-gradient polymerase chain reaction (PCR) was run using Taq
DNA polymerase (Sigma-Aldrich Co.) with D. punctata day 4 CA cDNA as a template. Bands
of the expected size were cut out and further purified using the GenEluteTM
Gel extraction Kit
(Sigma-Aldrich Co.). The resulting DNA fragments were subcloned into a CloneJET™ cloning
vector using the CloneJET™ PCR Cloning Kit (Fermentas) and sequenced. Sequences for PMK,
PPMD and JHAMT identified using degenerate primers, were too short to submit to NCBI’s
GenBank; therefore, RACE (Rapid Amplification of cDNA Ends) was performed. Their
sequences were confirmed using primers listed in Table S1.
Radiochemical assay (RCA) - The in vitro radiochemical assay (RCA) for JH biosynthesis was
performed (Feyereisen and Tobe, 1981; Tobe and Clarke, 1985). CA were incubated in TC199
medium for 3h, then transferred to new medium containing JH precursors for another 3h
incubation. JH biosynthesis in both incubations was measured, and first incubation
measurements were used as a control. JH precursors acetyl CoA, mevalonic acid (MA),
diphosphomevalonate (DPPM) and farnesol (Sigma-Aldrich, Canada) were dissolved in water
before use.
cDNA synthesis and quantitative real-time PCR (q-RT-PCR) - cDNA was transcribed from an
equal amount of RNA using the SuperScript™ III First-Strand Synthesis SuperMix for q-RT-
PCR in a final volume of 20 µl following the manufacturer’s instructions (Invitrogen Life

76
Technologies). All samples were reverse transcribed together in a single run. The resulting
cDNA samples were diluted 10-fold with PCR grade water. A calibrator sample was prepared by
pooling 5 µl of each cDNA sample. In the same run, negative control reactions were set up
without reverse transcriptase enzyme to test for genomic DNA contamination.
q-RT-PCR primers were designed using IDT’s (Integrated DNA Technologies) PrimeQuest
design tool (http://eu.idtdna.com/PrimerQuest/Home/Index). Primer sets were subsequently
validated by determining relative standard curves for each gene transcript using a five-fold serial
dilution of the calibrator cDNA sample. Efficiency and correlation coefficients (R²) were
determined for each primer pair. Primers used for q-RT-PCR profiling are listed in Table 2.1.
q-RT-PCR reactions were carried out in triplicate in a total volume of 10 µl containing 5 µl of
IQ™ SYBR® Green Supermix (Bio-Rad), 1 µl forward and reverse primer (5 µM), 2 µl of MQ-
water and 1 µl of cDNA. All reactions were performed using Bio-Rad’s CFX384 Touch ™ Real-
Time PCR Detection System using a two-step thermal cycling profile: 95ºC for 3 min, followed
by 40 cycles of 95ºC for 10 s and 59ºC for 30 s. Upon completion of every run, a dissociation
protocol (melt curve analysis) was performed to check for formation of primer dimers. A few
representative PCR products were run on a 1.2% agarose gel containing GelRedTM (Biotium)
and visualised under UV to confirm target specificity. Prior to target gene profiling, previously
described housekeeping genes were tested for their stability in the designed tissue distribution,
temporal profiling and RNAi experiments. The optimal housekeeping genes were selected using
the geNorm and Normfinder software as described previously (Marchal et al., 2013b). For each
tested cDNA sample, the normalization factor for the reference genes relative to the calibrator
samples was calculated and used to determine the normalized expression levels of the target
genes relative to the calibrator (Vandesompele et al., 2002).
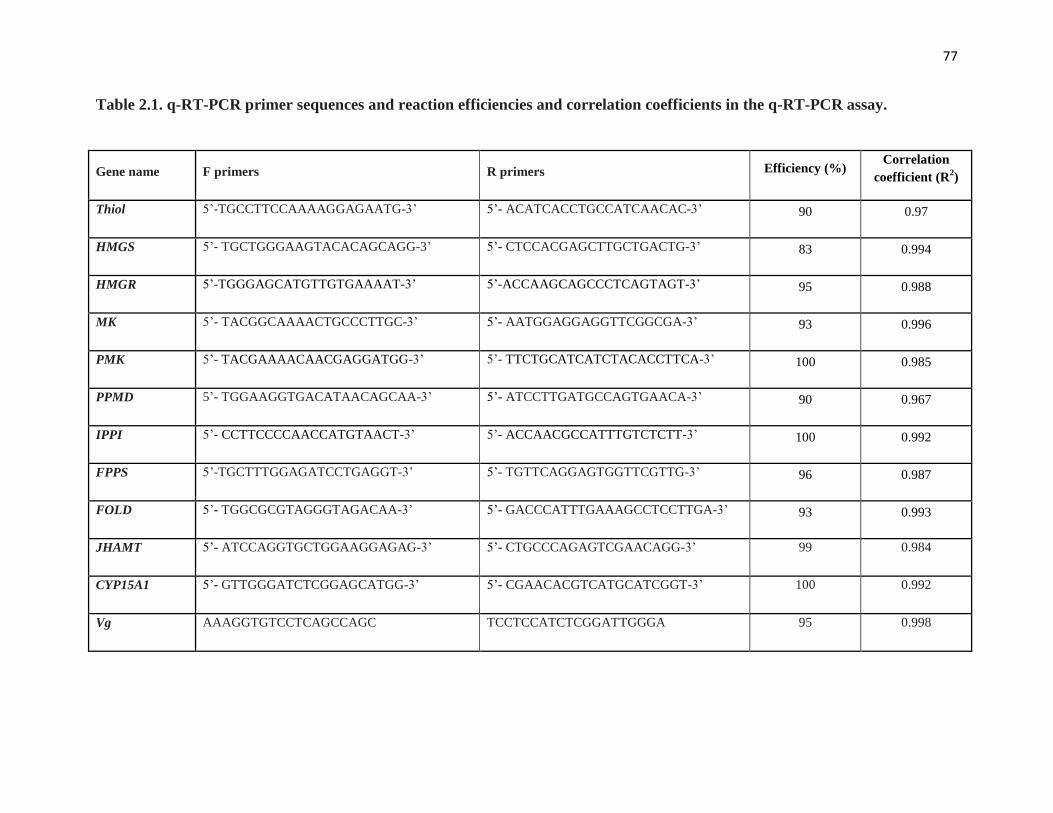
77
Table 2.1. q-RT-PCR primer sequences and reaction efficiencies and correlation coefficients in the q-RT-PCR assay.
Gene name F primers R primers Efficiency (%) Correlation
coefficient (R2)
Thiol 5’-TGCCTTCCAAAAGGAGAATG-3’ 5’- ACATCACCTGCCATCAACAC-3’ 90 0.97
HMGS 5’- TGCTGGGAAGTACACAGCAGG-3’ 5’- CTCCACGAGCTTGCTGACTG-3’ 83 0.994
HMGR 5’-TGGGAGCATGTTGTGAAAAT-3’ 5’-ACCAAGCAGCCCTCAGTAGT-3’ 95 0.988
MK 5’- TACGGCAAAACTGCCCTTGC-3’ 5’- AATGGAGGAGGTTCGGCGA-3’ 93 0.996
PMK 5’- TACGAAAACAACGAGGATGG-3’ 5’- TTCTGCATCATCTACACCTTCA-3’ 100 0.985
PPMD 5’- TGGAAGGTGACATAACAGCAA-3’ 5’- ATCCTTGATGCCAGTGAACA-3’ 90 0.967
IPPI 5’- CCTTCCCCAACCATGTAACT-3’ 5’- ACCAACGCCATTTGTCTCTT-3’ 100 0.992
FPPS 5’-TGCTTTGGAGATCCTGAGGT-3’ 5’- TGTTCAGGAGTGGTTCGTTG-3’ 96 0.987
FOLD 5’- TGGCGCGTAGGGTAGACAA-3’ 5’- GACCCATTTGAAAGCCTCCTTGA-3’ 93 0.993
JHAMT 5’- ATCCAGGTGCTGGAAGGAGAG-3’ 5’- CTGCCCAGAGTCGAACAGG-3’ 99 0.984
CYP15A1 5’- GTTGGGATCTCGGAGCATGG-3’ 5’- CGAACACGTCATGCATCGGT-3’ 100 0.992
Vg AAAGGTGTCCTCAGCCAGC TCCTCCATCTCGGATTGGGA 95 0.998

78
RNA interference (RNAi) - CA cDNA was used as a template to amplify fragments of genes to be
used in dsRNA construction. These fragments were subcloned and sequenced as described above.
Two separate constructs were designed in different regions of the genes to eliminate off-target
effects. Primers used are given in Table 2.2. dsRNA was synthesized using the MEGAscript
RNAi kit (Ambion). The procedure is based on the high-yield transcription reaction of a user-
provided linear transcript with a T7 promoter sequence. Transcription was carried out at 37 °C
overnight. The reaction mixture was treated with DNase I and RNase, and then purified by
phase-solid phase adsorption purification, according to the manufacturer’s instructions (Ambion).
The dsRNA concentration was determined using a Nanodrop spectrophotometer (Thermo
Scientific.). Diluted dsRNA was run on a 1.2% agarose gel to examine integrity of the construct
and efficiency of duplex formation. The negative control construct (-pJET) was designed in a
non-coding region of the pJET 1.2/blunt cloning vector (CloneJet PCR Cloning Kit, Thermo
Scientific).
Several RNAi trials with different injection timings and dsRNA construct concentrations were
investigated to obtain an efficient gene silencing (data not shown). Newly molted mated females
were injected with 3 μg HMGR dsRNA or control dsRNA on day 0 and day 2, and with 3 μg
JHAMT dsRNA or control dsRNA on day 1 and day 3. CA, ovary and fat body samples were
taken on day 4 as described above. Fat body was immediately stored in liquid N2 prior to RNA
extraction. CA were dissected and cleaned in TC199 medium (GIBCO; 1.3 mM Ca2+, 2% Ficoll,
methionine-free) for use in the RCA or flash frozen in liquid N2 to prevent RNA degradation.
Basal oocyte length was measured and the ovaries were collected for vitellogenin measurements
(§ 2.7) and histology (§ 2.8).
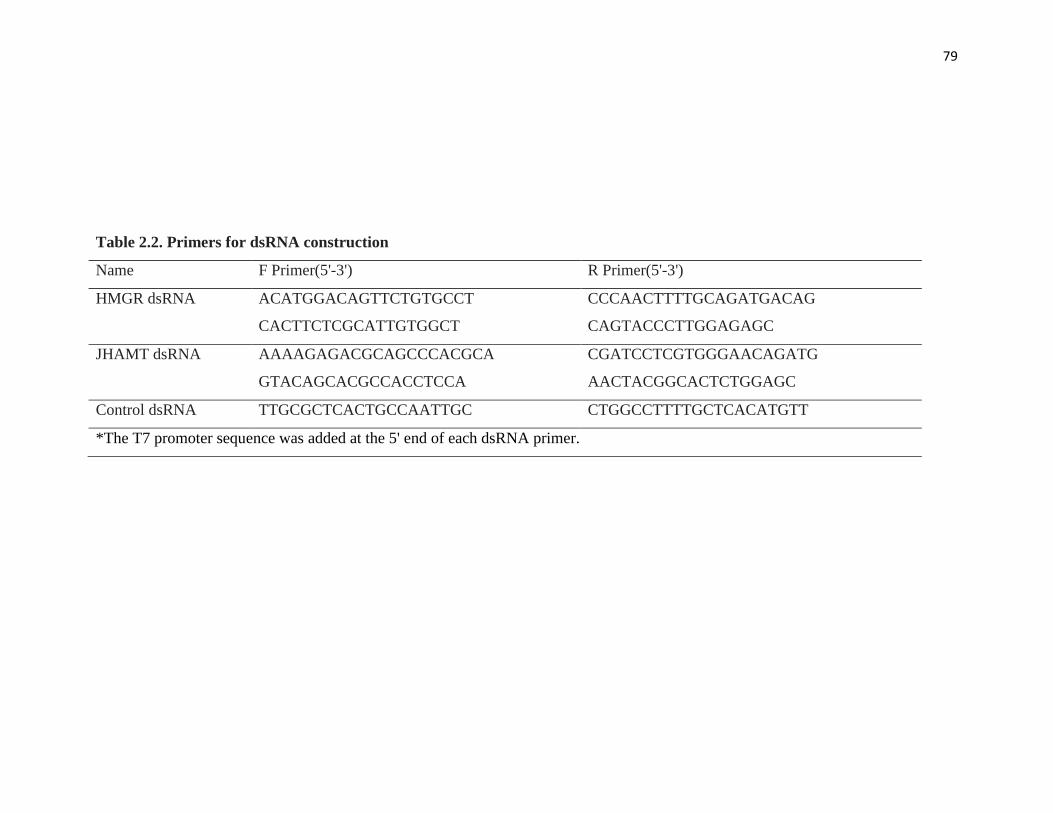
79
Table 2.2. Primers for dsRNA construction
Name F Primer(5'-3') R Primer(5'-3')
HMGR dsRNA ACATGGACAGTTCTGTGCCT CCCAACTTTTGCAGATGACAG
CACTTCTCGCATTGTGGCT CAGTACCCTTGGAGAGC
JHAMT dsRNA AAAAGAGACGCAGCCCACGCA CGATCCTCGTGGGAACAGATG
GTACAGCACGCCACCTCCA AACTACGGCACTCTGGAGC
Control dsRNA TTGCGCTCACTGCCAATTGC CTGGCCTTTTGCTCACATGTT
*The T7 promoter sequence was added at the 5' end of each dsRNA primer.

80
Vitellogenin ELISA - Single dissected ovaries were homogenized and extracted twice in 50 µl
PBS according to Mundall et al. (1981). Total ovary protein content was determined using a
Bradford assay (Bradford, 1976) performed according to the manufacturer’s (Sigma-Aldrich) 96
well plate protocol.
Vitellin was quantified using an indirect enzyme-linked immunosorbent assay (ELISA)
following the protocol described in Marchal et al. (2014). A rabbit polyclonal antibody made
against D. punctata mature egg homogenate was used as the first antibody (Stoltzman and Stay,
1997). Goat anti-rabbit IgG, HRP-linked antibody (1:3000 in 1% BSA) was then added as the
second antibody. Wells were treated with 100ul of TMB for 10 min. Absorbance was measured
at 650 nm with a Molecular Devices SpectraMax Pus 384 microplate reader.
Histology and microscopy - To view overall structure, a subset of oocytes was fixed for 3 days in
3% glutaraldehyde in 0.1 M phosphate buffer, post-fixed for 1 hr with 1% OsO4 in 0.1 M
phosphate buffer, dehydrated in ascending ethanol series, and embedded in Spurr’s resin.
Sections (1 µm) were cut with a Leica EM UC6 ultramicrotome, then mounted and stained with
methylene blue and toluidine blue. All light microscopy was conducted using a Leica DMI3000
inverted microscope.
Results
Identification of genes encoding JH biosynthetic enzymes and Vg in D. punctata.
CYP15A1 was previously identified in D. punctata and the genes encoding HMGS, MK, FPPS
and FOLD were present in a CA EST database of D. punctata (Helvig et al., 2004; Noriega et al.,
2006). We confirmed the sequence of HMGS, MK, FPPS and FOLD. For the remaining enzymes
in the JH biosynthetic pathway, we aligned (predicted) orthologous sequences from different

81
insect orders and designed degenerate primers based on conserved domains. We succeeded in
cloning an additional five (partial) sequences for enzymes in the conserved mevalonate pathway:
Thiol, HMGR, PMK, PPMD and IPPI and one extra sequence encoding an enzyme in the JH-
specific part of the pathway, JHAMT. The complete sequences for HMGR and JHAMT were
obtained. The ORFs of HMGR and JHAMT encode proteins of 825 and 274 amino acids in
length, respectively. Amino acid alignments of these proteins with known insect orthologs are
given in Supplementary Fig. S2.1. Moreover, a partial sequence of Vg was cloned from D.
punctata adult female fat body. The sequences for the genes were deposited in NCBI’s GenBank
with accession numbers shown in Table S2.1.
Tissue specificity and developmental transcript profiles during the first gonadotropic cycle are
consistent with roles of these genes in JH biosynthesis.
Using q-RT-PCR, the relative transcript levels of 11 genes in the JH biosynthetic pathway were
determined in several tissues of day 4 mated females and males. Relative transcript levels for the
Diploptera orthologs of genes in the JH pathway were normalized to transcript levels of the
reference genes EF1a and Tubulin (Marchal et al., 2013b). To represent the tissue distribution
data for the 11 enzyme-encoding genes in one figure, the transcript levels were normalized to the
transcript level of PMK measured in the calibrator sample. The orthologous genes encoding
enzymes in the JH biosynthetic pathway were most highly expressed in the CA, which is
consistent with the functions of the enzymes (Fig. 2.2). Most genes appear to be exclusively
expressed in the day 4 female CA: Thiol, HMGS, HMGR, MK, PPMD, FPPS, JHAMT and
CYP15A1; whereas a few show a broader tissue distribution: PMK, IPPI and FOLD. These latter
3 appear to be expressed not only in nervous tissues but also in peripheral tissues such as the
ovary, fat body and Malpighian tubules. Moreover, there are 1000-fold differences in relative

82
Figure 2.2. Tissue specific expression of genes encoding JH biosynthetic enzymes. All tissues were dissected from day 4 mated
adult females, except accessory glands (AG) and testes (Test) were from day 4 adult male. Abbreviations on the X-axis: Brain (Br),
corpora allata (CA), nerve cord (NC), midgut (Mg), Malpighian tubules (MT), fat body (Fb), and ovary (Ov). Bars represent the mean
of three biologically independent pools of ten animals run in triplicate and normalized to Tubulin and EF1α. Vertical error bars
indicate SEM.

83
mRNA levels of the enzyme-encoding genes, with PMK, PPMD, IPPI and FOLD being present
in low abundance, Thiol, HMGS and HMGR being of intermediate abundance and MK, FPPS,
JHAMT and CYP15A1 being represented in high abundance (Fig. S2.2).
Our next step was to follow the relative transcript levels in the CA of mated females throughout
the first gonadotropic cycle. Target gene expression was normalized to transcript levels of EF1a
and Armadillo according to a previous study (Marchal et al., 2013b). In general, the expression
of the 11 genes is correlated with the in vitro rates of JH biosynthesis in the CA and relative
mRNA levels for DippuVg measured in the female fat body (Fig. 2.3). The relative mRNA levels
in the CA for most of the genes of interest were low at the beginning of the adult female stage
when JH biosynthesis is low; rose during the vitellogenic portion of the first gonadotropic cycle
reaching a peak on day 3-4 and thereafter began to decline on day 5, attaining a low level during
oviposition on day 7. However, there appear to be three exceptions to this pattern: PMK, IPPI
and FPPS. Relative mRNA quantities for PMK and IPPI were very low compared to other genes
and did not display dramatic changes during the first gonadotropic cycle. FPPS transcripts were
high on day 0, remained high until day 4 and then declined on day 5 when vitellogenesis slows.
Addition of JH precursors stimulates JH biosynthesis in CA with low JH biosynthetic activity in
vitro.
Previous studies suggested that JH synthesis is controlled by the rate of flux of isoprenoids in A.
aegypti (Nouzova et al., 2011). To determine the role of other JH precursors in regulating JH
biosynthesis, we tested the rate of JH biosynthesis with the addition of JH precursors in the early
steps of mevalonate pathway or the addition of farnesol. The addition of acetyl CoA, DPPM or
farnesol to the incubation medium had a significant stimulatory effect on JH biosynthesis,

84
Figure 2.3. Developmental expression of genes encoding JH biosynthetic enzymes during the first gonadotrophic cycle of D.
punctata. Measurements were taken every day during the cycle (day 0 to day 7 after the final molt). Bars represent the mean of three
biologically independent pools of ten animals run in triplicate and normalized to Armadillo and EF1α. Vertical error bars indicate
SEM. Inset at the right bottom shows JH biosynthesis per individual CA (n=12) and the transcript level of DpVg in fat body during the
first gonadotrophic cycle (Marchal et al., 2014). Vertical error bars indicate SEM.

85
whereas MA did not (Fig. 2.4A). For the first time, we demonstrated that acetyl CoA, as the first
precursor in the JH biosynthetic pathway, was able to stimulate JH biosynthesis. The rank order
of the stimulatory effects of the different JH precursors on JH biosynthesis is as follows:
farnesol > acetyl CoA > DMMP > MA.
We also evaluated the sensitivity of CA to farnesol during the first gonadotrophic cycle. CA
were dissected from day 0, 3, 4, 5, and 7, and incubated with medium containing 40 µM farnesol.
On days 3 and 4, when the CA show high JH biosynthetic activity, the addition of farnesol had
no effect on in vitro JH biosynthesis (Fig. 2.4B). On the other hand, JH biosynthesis in CA of
day 0, 5 and 7 mated females was significantly increased.
Injection of HMGR-JHAMT dsRNA resulted in a significant downregulation of the target genes
but also of other genes in the JH biosynthetic pathway.
A systemic RNAi response was observed following injection of HMGR- and JHAMT dsRNA
into animals every other day during the first gonadotropic cycle. Relative transcript levels were
measured in the CA of day 4 animals using q-RT-PCR. A significant knockdown of 64% and 94%
was measured for HMGR and JHAMT, respectively. Off-target effects were investigated by
checking the Ct value of housekeeping genes . Moreover, two different dsRNA constructs were
used to eliminate off-target effects (primers listed in Table 2). Both constructs yielded a similar
phenotype.
q-RT-PCR was used to determine the relative mRNA levels of the other genes involved in the JH
biosynthetic pathway using the control and treated CA described above. A significant reduction
in relative expression levels was observed for all genes encoding the enzymes directly involved
in the pathway, with the exception of the genes encoding FOLD and the epoxidase, CYP15A1
(Fig. 2.5).
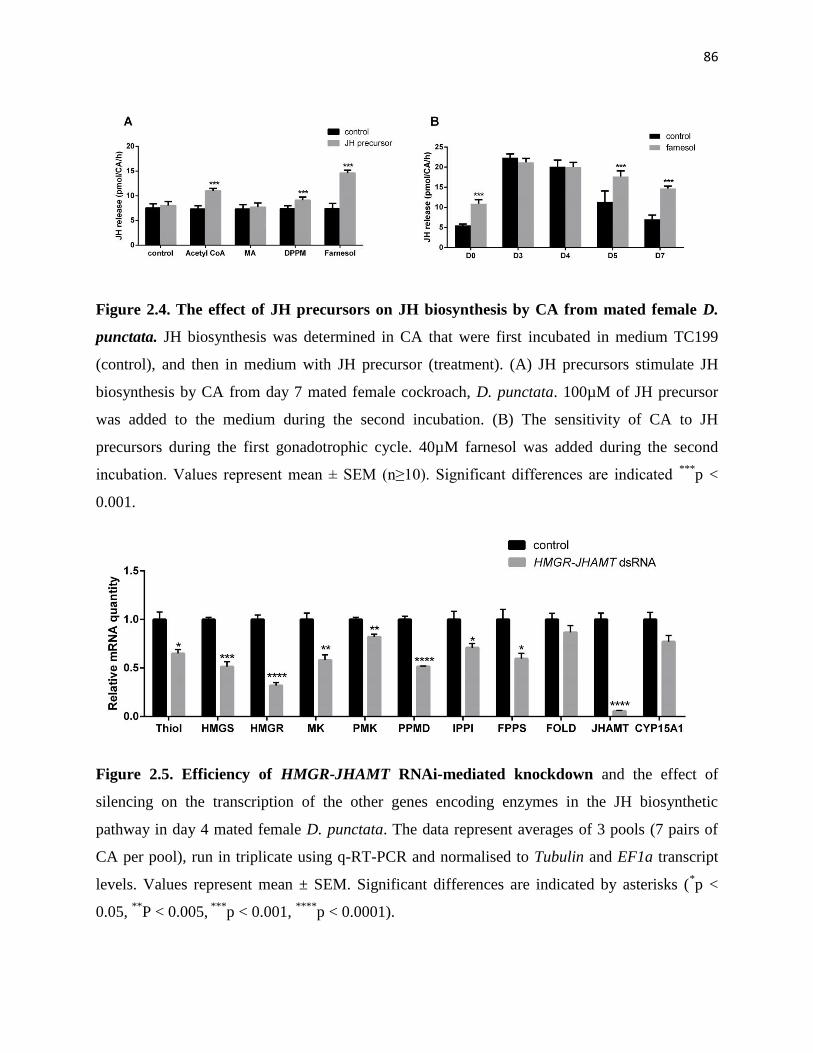
86
Figure 2.4. The effect of JH precursors on JH biosynthesis by CA from mated female D.
punctata. JH biosynthesis was determined in CA that were first incubated in medium TC199
(control), and then in medium with JH precursor (treatment). (A) JH precursors stimulate JH
biosynthesis by CA from day 7 mated female cockroach, D. punctata. 100µM of JH precursor
was added to the medium during the second incubation. (B) The sensitivity of CA to JH
precursors during the first gonadotrophic cycle. 40µM farnesol was added during the second
incubation. Values represent mean ± SEM (n≥10). Significant differences are indicated ***
p <
0.001.
Figure 2.5. Efficiency of HMGR-JHAMT RNAi-mediated knockdown and the effect of
silencing on the transcription of the other genes encoding enzymes in the JH biosynthetic
pathway in day 4 mated female D. punctata. The data represent averages of 3 pools (7 pairs of
CA per pool), run in triplicate using q-RT-PCR and normalised to Tubulin and EF1a transcript
levels. Values represent mean ± SEM. Significant differences are indicated by asterisks (*p <
0.05, **
P < 0.005, ***
p < 0.001, ****
p < 0.0001).

87
Silencing HMGR and JHAMT resulted in a significant reduction in rates of JH biosynthesis and
slows ovarian development.
The downregulation of HMGR and JHAMT resulted in a 73% reduction in the rates of JH
biosynthesis in the CA of day 4 adult females as measured using the in vitro RCA (Fig. 2.6A).
To confirm the role of JH in inducing Vg transcription in the fat body and uptake in the
developing basal oocytes, fat body and ovaries were dissected from four day old control and
treated adult cockroaches. The basal oocyte length was significantly decreased following
silencing of HMGR and JHAMT. The average oocyte length in the HMGR-JHAMT RNAi
animals measured 0.8 mm compared to the control of 1.21 mm (Fig. 2.6B). The transcript level
of the Vg in the fat body of the HMGR-JHAMT dsRNA-treated animals was reduced 80% (Fig.
2.6C). In addition, the knockdown of HMGR-JHAMT mRNA resulted in a significant reduction
in vitellin content compared to the controls (Fig. 2.6D).
We also studied the histology of the developing basal oocytes in control and treated females.
Control oocytes were fully patent on day 4, showing large intercellular spaces between the
follicle cells and the clear presence of yolk spheres. In the HMGR-JHAMT RNAi animals, on the
other hand, patency was not fully induced and as a result, yolk was not deposited in these
oocytes (Fig. 2.7).
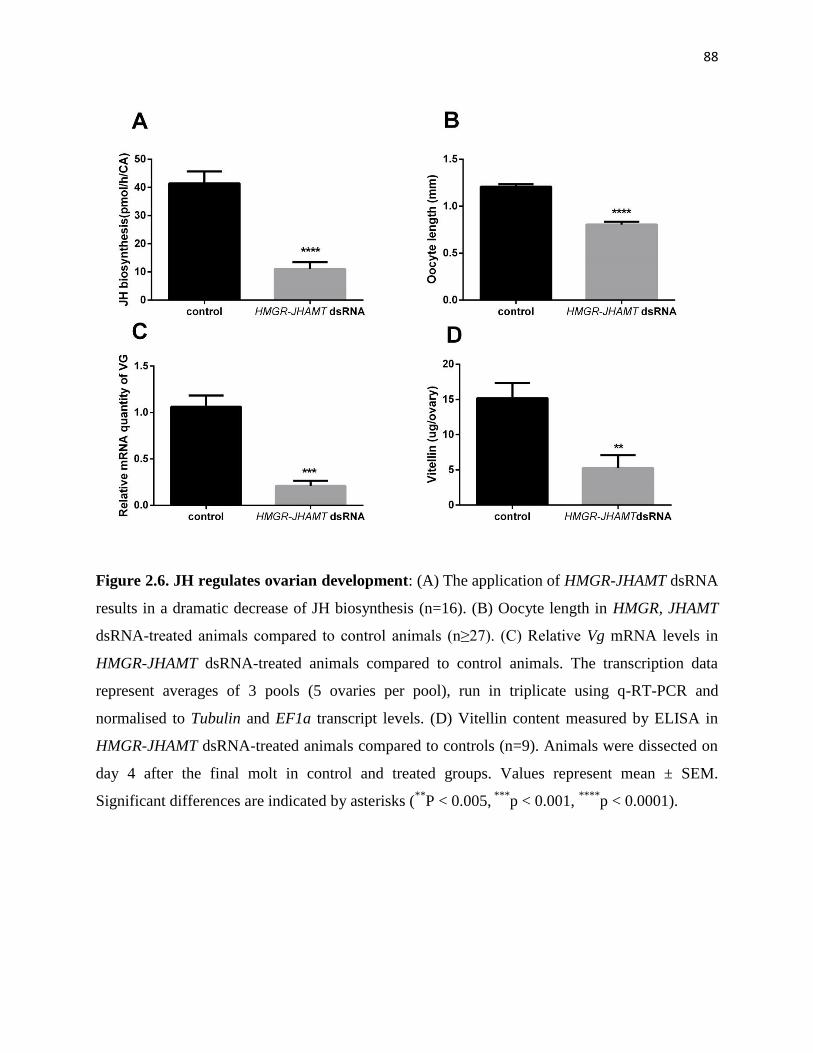
88
Figure 2.6. JH regulates ovarian development: (A) The application of HMGR-JHAMT dsRNA
results in a dramatic decrease of JH biosynthesis (n=16). (B) Oocyte length in HMGR, JHAMT
dsRNA-treated animals compared to control animals (n≥27). (C) Relative Vg mRNA levels in
HMGR-JHAMT dsRNA-treated animals compared to control animals. The transcription data
represent averages of 3 pools (5 ovaries per pool), run in triplicate using q-RT-PCR and
normalised to Tubulin and EF1a transcript levels. (D) Vitellin content measured by ELISA in
HMGR-JHAMT dsRNA-treated animals compared to controls (n=9). Animals were dissected on
day 4 after the final molt in control and treated groups. Values represent mean ± SEM.
Significant differences are indicated by asterisks (**
P < 0.005, ***
p < 0.001, ****
p < 0.0001).

89
Figure 2.7. Transverse sections of the basal oocytes from day 4 control and HMGR-JHAMT dsRNA-treated animals. Follicle
cells (Fc), intercellular gaps (arrows), yolk spheres (arrowheads) and lipid spheres (L) are indicated. Scale bars represent 200 µm.

90
Discussion
Sequence of genes encoding enzymes in the JH biosynthetic pathway - Through in silico data
mining and degenerate primer PCR and RACE, we have successfully characterized 11 of the 13
enzyme-encoding genes in the JH biosynthetic pathway of D. punctata. Cheng et al (2014)
showed that enzymes in the early steps of the mevalonate pathway were generated by single
copy genes in many insects, whereas genes encoding FPPS and enzymes in the JH-specific
pathway probably underwent gene duplication in Lepidoptera. In D. punctata, only a single copy
of these genes was identified. However, at this point, the existence of multiple copies of several
of these genes cannot be ruled out and this issue will have to await the assembly of the
Diploptera genome.
The full-length HMGR and JHAMT were cloned from D. punctata CA. HMGR encodes a protein
of 825 amino acids in length. Following a conserved domain search, HMGR was observed to
contain the typical conserved motifs found in members of the HMG-CoA reductase superfamily
of proteins. JHAMT encodes a protein of 274 amino acids in length and contains the motifs
typically found in AdoMet-dependent methyltransferases. Still missing are sequences for
farnesyl diphosphate pyrophosphatase (FPPP) and farnesal dehydrogenase (FALD). Molecular
analysis of these genes was long hampered as a consequence of the minute size of the CA but
using a newly developed sensitive assay for measuring JH precursor pools employing fluorescent
tags, the genes encoding FPPP and FALD were recently characterized in A. aegypti (Nyati et al.,
2013; Rivera-Perez et al., 2013; Rivera-Perez et al., 2012). These were, however, not included in
our study because the availability of only dipteran sequences made their characterization in the
phylogenetic basal D. punctata problematic and therefore remain uncharacterized in this species
to date.

91
Transcription of enzymes in the JH biosynthetic pathway partly regulates JH biosynthesis - The
11 orthologs of JH biosynthesis genes cloned in our model D. punctata are predominately
expressed in the CA, and their relative transcript levels correlate well with the rates of JH
biosynthesis (Fig. 2.2 and 2.3). Our results suggest that CA are highly specialized tissues for the
biosynthesis of JH, and genes identified in our study are encoding functional enzymes in the JH
pathway. Enzymes in the mevalonate pathway are responsible for not only JH production but
also for the production of other terpenoids such as defensive secretions and pheromones (as
reviewed by (Belles et al., 2005; Goldstein and Brown, 1990)). It is therefore of interest that all
mevalonate enzyme-encoding genes except PMK and IPPI are exclusively expressed in the CA
with trace amounts in other tissues. This can be explained by the strikingly high rates of JH
biosynthesis in the CA of day 4 adult female D. punctata. For the later enzymes involved in the
JH specific steps of the pathway, the transcription of JHAMT and CYP15A1 is CA-specific,
whereas FOLD is expressed in many tissues. Similar results were observed in A. aegypti and A.
mellifera (Bomtorin et al., 2014; Mayoral et al., 2009a; Nouzova et al., 2011). Because of the
multiple functions of farnesol, the oxidation of farnesol to farnesal does not appear to be a JH
biosynthesis-specific reaction (Mayoral et al., 2009a).
Changes in rates of JH biosynthesis during the first gonadotropic cycle of D. punctata are
dynamic, corresponding to specific reproductive events (see below) and are therefore tightly
regulated. The transcript levels of enzyme-encoding genes in JH biosynthesis are highly
coordinated with the rates of JH biosynthesis (Fig. 2.3), suggesting that at least part of the
regulation of JH biosynthesis involves coordinated changes in the transcription of the genes in
the biosynthetic pathway, in agreement with studies performed in A. aegypti (Nouzova et al.,
2011) and B. mori (Kinjoh et al., 2007), although the ability of each gene to regulate JH

92
biosynthesis appears to differ. In addition, the expression patterns of HMGS and HMGR mRNA
were similar to the patterns of their enzyme activities (Couillaud and Feyereisen, 1991;
Feyereisen and Farnsworth, 1987). Although the transcription of most genes in the biosynthetic
pathway showed a correlation with the JH biosynthesis, there are a few exceptions, including
PMK, IPPI and FPPS. PMK and IPPI, which are expressed in multiple tissues (Fig. 2.2), and
may not be specifically regulated in the CA. For FPPS, 3 FPPS homologs were found in B. mori
(Ueda et al., 2009), and 7 in A. mellifera (Bomtorin et al., 2014). Although we currently have
identified only 1 FPPS gene in D. punctata, additional FPPS genes may exist in the CA that
regulate JH biosynthesis.
Effect of exogenous JH precursors on rates of JH biosynthesis - We have found that JH
precursors are able to stimulate JH biosynthesis (Fig. 2.4A), which suggests that the rate of JH
biosynthesis is not only controlled by transcription of the genes in the pathway, but also by the
flux of substrates in the pathway. Farnesol stimulates JH biosynthesis by CA from day 0, 5 and 7
mated females. However, application of farnesol to CA showing high JH biosynthetic activity
(day 3 and 4) does not result in a significant change in JH biosynthesis (see Fig. 2.4B). These
results suggest that during the first gonadotrophic cycle of D. punctata, the supply of this
precursor is rate-limiting in CA with showing low JH biosynthetic activity, whereas there are
other factors, including nutrients and neurotransmitters that control JH production at high rates of
JH biosynthesis. Nevertheless, 14 JH precursors are involved in the JH biosynthetic pathway,
and each JH precursor connects to the ‘upstream’ and ‘downstream’ enzymes. Many factors
could affect the stimulatory effect of JH precursor in JH production, including cell permeability
of the added compounds, the activity of upstream/downstream enzymes, and the size of other JH
precursor pools.

93
A feedback mechanism involved in the regulation of the transcription of JH biosynthetic genes -
HMGR and JHAMT have been studied in the JH pathway since they are representative enzymes
involved in the well-conserved mevalonate pathway and in the JH-specific portion of the
pathway respectively (Debernard et al., 1994; Li et al., 2013; Marchal et al., 2011;
Martinezgonzalez et al., 1993; Mayoral et al., 2009c; Monger et al., 1982; Wang et al., 2013).
We therefore chose to silence HMGR and JHAMT on a posttranscriptional level using RNAi
during the first gonadotropic cycle of D. punctata. RNAi trials have shown that fewer injections
of HMGR or JHAMT dsRNA and with lower concentrations of construct, did not result in a
significant silencing of our target gene mRNA levels (data not shown). We therefore conclude
that the RNAi response in D. punctata appears to be systemic, but is not as effective as in
another hemimetabolous insect model, the desert locust, Schistocerca gregaria in which smaller
quantities of the JHAMT dsRNA construct induced a longer downregulation of the same target
gene during the first gonadotropic cycle (Marchal et al., 2011). A further in-depth study on the
RNAi machinery D. punctata should be performed to explain the differences in sensitivity
compared to other hemimetabolous insect species.
JH biosynthesis is controlled by many factors including neuropeptides (allatostatins, allatotropins,
short neuropeptide F) (Kaneko and Hiruma, 2014; Weaver and Audsley, 2009),
neurotransmitters (octopamine, dopamine and glutamate) (Granger et al., 1996; Pszczolkowski et
al., 1999; Thompson et al., 1990) and JH itself (Goodman and Cusson, 2012; Marchal et al.,
2013a). Topical application of JH or a JH analog resulted in a suppression of JH biosynthesis,
which suggests a negative feedback regulation of CA in D. punctata (Tobe and Stay, 1979). In
higher animals, cholesterol, the bulk end-product of mevalonate pathway, shows a feedback
regulation on the transcription and post-transcription of enzymes in the mevalonate pathway

94
(Chang and Limanek, 1980; Clarke et al., 1987; Goldstein and Brown, 1990). Later studies
showed that farnesol, an intermediate product in the JH biosynthetic pathway, is able to
accelerate the degradation of HMGR, and the increase of mevalonate flux raises the activity of
FPPP expressed in CHO cells (Meigs et al., 1996; Meigs and Simoni, 1997). In our study,
regulating transcription of the JH biosynthetic enzymes seems to affect the entire biosynthetic
pathway, rather than individual steps. The silencing of HMGR and JHAMT resulted in a
significant decrease in the transcript level of several other genes in the pathway (Fig. 2.5). A
possible explanation is that the accumulation of JH precursors as a result of RNAi treatment
resulted in a feedback on the expression of other genes in the JH biosynthetic pathway to balance
the size of JH precursor pools and the enzyme activity.
The essential role of JH in reproduction - Our results clearly show that select silencing of genes
encoding enzymes in the JH biosynthetic pathway effectively reduces JH biosynthesis in vitro
(Fig. 2.6). As a well-known model for the study of the regulation of JH biosynthesis, D. punctata
displays a consistent characteristic profile of JH production during the first gonadotropic
cyclethat is closely correlated with specific reproductive events such as vitellogenesis and
chorionation. Once JH production rises on day 2, vitellogenin synthesis in the fat body
commences, and vitellogenin is taken up from the hemolymph and is incorporated into the
developing basal oocytes. The elevated JH titer results in patency in the follicular epithelium of
the basal oocytes, permitting the uptake of Vg and subsequent yolk formation (Marchal et al.,
2014; Pratt and Davey, 1972). On day 5, the spaces in the follicular epithelium close as rates of
JH biosynthesis decline and Vg transcription in the fat body and circulating Vg levels decrease.
At this point, choriogenesis begins and JH biosynthesis remains low until oviposition on day 7
(as reviewed by Marchal et al., 2013a). Our RNAi study has focused on day 4 of the first

95
gonadotropic cycle, a time when JH titre is high in controls and animals are vitellogenic (Tobe et
al., 1985) as characterized by high DippuVg mRNA transcript levels in the fat body and JH-
dependent induction of patency in the maturing basal oocytes. Following the reduction in JH
production by the silencing of DippuHMGR and DippuJHAMT, mRNA levels of DippuVg in the
fat body are significantly reduced, incorporation of vitellin into the oocytes is impaired and
patency does not occur (Fig. 2.6 and 2.7). Previous reports have shown that in S. gregaria,
oocyte length is significantly affected following treatment of females with JHAMT dsRNA
(Marchal et al., 2011). RNAi was also used in the cotton bollworm Helicoverpa armigera to
effectively silence HMGR, resulting in reduced Vg expression (Wang et al., 2013). Each of these
studies highlights specific aspects of JH-regulated reproductive events. The results described in
the current paper confirm the central role played by JH during the female reproductive cycle of D.
punctata.
JH biosynthesis is a fundamental process in regulating insect development, metamorphosis and
reproduction (Goodman and Cusson, 2012). Although the genes directly involved in JH
biosynthesis have been characterized using molecular techniques in several insect species, there was
no similar study in the hemimetabolous insects. Thus, our study is the first to characterize the
majority of the genes directly involved in JH biosynthesis in a hemimetabolous insect and a well-
known model for studying the physiology of JH. This paper now provides the molecular tools to
study the regulatory mechanisms of JH production in D. punctata (for a first study, see (Huang et
al., 2014)). In addition, the role of JH biosynthetic enzymes and the JH precursor supplies in
regulating JH biosynthesis have been demonstrated, as has the critical function of JH as the
master regulator of cockroach female reproduction.

96
Supplementary data
A.
BlageHMGR 1 M-VGRLFRAHGQFCASHPWEVIVATLTLTVCMLTVDQ-------------RP-LGLPPGWGHNC---------------------------------ITLEEYNAADMIVMTLIRCVAVLYSYYQFCHLQKLGSKYILGI
DippuHMGR 1 M-VGWLFLAHGQFCASHPWEVIVATFTLTACMLTVDP-------------HP-LGLPPGWRKNC---------------------------------ISLEEYNAADVIVMTLIRCVAVLYSYYQFCHLQKLGSKYILGI
ApimeHMGR 1 M-LTRLFEIHGRFCAGHPLEVIVTTFTLTACILNMETG----------NGHPSVPLTAHCGPGR---------------------------------CNSDDLNAADVIVMTIIRCLAILFTYHQFRNLQKLGSKYILGI
TricaHMGR 1 M-TTRLFRAHGEFCATHPWEVIVATLTLAACMLTVDQQ----------HPAPPPKPTLRYCAEC---------------------------------LQEAEYNAADLIVMTLIRCLAVLYCYYQFCNLEKLGSKYILGI
AedaeHMGR 1 MKVGRLFRAHGEFCASHPWEVIVALVTLTACIITFDKG----------SADPFQQSSSRSGSGRPC----------------PSWRFSSVSPDEAFRCEEVEQNGIDVILMTIVRCSAILYCYYHFCNLQKLGSKYILGI
DromeHMGR 1 M-IGRLFRAHGEFCASHPWEVIVALLTITACMLTVDKNNTLDASSGLGTATASAAAAGGSGSGAGSGASGTIPPSSMGGSATSSRHRPCHGWSQSCDGLEAEYNAADVILMTIVRCTAVLYCYYQFCSLHRLGSKYVLGI
BommoHMGR 1 M---KVWGAHGEFCARHQWEVIVATLALLACAASVERHG----------PGNRSEHCAGWARACP--------------------------------GLEAEYQAADAVIMTFVRCAALLYAYYQVLNLHKIASKYLLII
BlageHMGR 93 AGLFTVFSSFVFSSSVINFLGSDVSDLKDALFFFLLLIDLSKATVLAQFALSSRSQDEVKHNIARGIAMLGPTITLDTVVETLVIGVGMLSGVRRLEVLCCFACMSVIVNYVVFMTFYPACLSLILELSRSGESGRPAWH
DippuHMGR 93 AGLFTVFSSFVFSSSVINFLGSDISDLKDALFFFLLLIDLSKATLLAQFALSSCSQDEVKHNIARGIAMLGPTITLDTVVETLVIGVGTLSGVRRLEVLCCFACMSVIVNYIVFMTFYPAFLSLILELARSGEGGRPAWH
ApimeHMGR 97 AGLFTVFSSVVFTSSVVNFGRSDISDLKDALFFFLLIIDLSKAAVLAQLALSSRNKEEVRANIARGMSLLGPTITLDTLVETLLISIGSLSGVRRLEILCSFACLGVVVNYIVFMTFYPACLSLILELSRETNTMKPLSA
TricaHMGR 97 AGLFTVFSSFVFTSTVLNLLWIDVSDLKDALFFFLLLIDLSKAAMLAQSALSASNQEEVKSNIARGMAVLGPTITLDTIVETLVIGVGTLSGVHRLEMLSYFACLSVLVNYIVFMTFYPACLSLILELSRTTN---IYGN
AedaeHMGR 115 AGLFTVFSSFIFTSTVINFLGSEVSDLKDALFFFLLLIDLSKAAILAQLALCGSSQSEVTMNIARGMEILGPAISLDTLVETLVIGVGTLSGVQRLELLSGFAVLSAIVNYIVFMTFYPACLSLILDLSRNAGNLIQKNK
DromeHMGR 140 AGLFTVFSSFIFTTAIIKFLGSDISELKDALFFLLLVIDLSNSGRLAQLALSGSNQAEVTQNIARGLELLGPAISLDTIVEVLLVGVGTLSGVQRLEVLCMFAVLSVLVNYVVFMTFYPACLSLIFDLSRSGVDMSVVRE
BommoHMGR 96 AGVFSTFASFIFTSALASLFWSELASIKDAPFLFLLVADVARGARMAKAGWSAG--EDQGKGVGRALSLLGPTATLDTLLAILLVGVGALSGVPRLEHMCTFACLALLVDYIVFITFYPACLSLVADFATNRK---EIAH
BlageHMGR 233 DKS--LIIKALHEEDQKPNPVVQRVKVIMSAGLMLVHAHRWVRCLSIALWPDLTSLRYFCTHCDTGVSYSRWSFASEGEELPTVKLVTG--DSVVNSNST----DDAQLHYYIMRWLTVSADHIVILILLLALAVKFVFF
DippuHMGR 233 DRS--LIMKALHEEDQKPNPVVQRVKVIMSAGLMLVHAH------------------------------SRWSFASEGEELPIVNLVTG--DSVVITNST----QDAQIHDYIMRWLTVSADHIVILILLLALAVKFVFF
ApimeHMGR 237 DKI--FMMHPLDEEDQKPNPVVERVKLIMIAGLFVVHAN------------------------------SRFKSE-ESEDTVEGKVSST--NSHVIVNSYNETEDSSEVKGYLMNWLSVSADNIVILILLLALAIKFIFF
TricaHMGR 234 KQS--LIARALKEEDHKSNPVVQRVKLIMSAGLMLVHAR------------------------------SRWPFK--EDDVENIRPLVV--EQHMTLNRT----EDTTLHEYIMKWVTVSADHIVILILLLALVVKFVFF
AedaeHMGR 255 KEN--LLARVLTEEDQKPNPVVQRVKLIMSSGLMIVHVL------------------------------SRLAISEKDSDTAEHIIASHSHEHLAAMNKT----EPNEISEFIMRWLSISTEQIVTYILIIALGVKFVFF
DromeHMGR 280 KAKGSLLLKSLTEEEQKANPVLQRVKLIMTTGLMAVHIY------------------------------SRVAFSGSDYDAVDKTLTPT-LSLNVSNNRT----ESGEIADIIIKWLTMSADHIVISIVLIALVVKFICF
BommoHMGR 231 DSP-------FSEEDLKPNPVVQRVKMIMAAGLLCVHLT------------------------------SRWPWS------ANHGIIEG--PIDASIPVP----HDNILLHSYVKWFSVSADYIVIATLLCALIIKFIFF
BlageHMGR 365 ETRDELTTTRGMDGWVEVSSP------VEHKYVQTE----------------------------------QPSCSAP------EQPLEEPPASN------------RSIDECLSVCK-SDVGAQ--ALSDCEVMALVTS-
DippuHMGR 335 EVKDELNTTRGMDGWVEVSSP------VEHKFVQTD----------------------------------QVCWELPSSS---QEPDEQPLPCD------------RPVQECLAICK-SDAGAN--SLSDSEVMSLVSS-
ApimeHMGR 342 EDKEDIAKQLQFKVEDDTEEK------IENEYMKEKKFEIEYVKDKENENEDMNFSFLTTKMPFKLSFAKMQTISIPWIDGKEEQIDEKQCTVNTSNDKNLFSQIPRSVEECLKIYK-SELGAN--GLTDEEVIQLVKN-
TricaHMGR 334 ENKEELAEQLRAHISTESVDPGKKDNRFKMPLIKTQSFFLTNNTKED---------------------SACEDKEVQTDIGRLESEFEKLPAGN-----DVSVKTCRSLEECLKIYNDSNLGGA--ALSDDEVILLVKN-
AedaeHMGR 359 D-RHNLSDQILLSVANEAAAAAAAAAAAAQKQKQLELELERPQPVATFT--------LAETVPVEEKATQSELCLLPGRGQRAKSMGADDELNLDDLEDELVEREPRPLEMCLKILNETDDGAV--GLTDEEIKMIVRAG
DromeHMGR 385 DNRDPLPDQLRQSGPVAIAAKASQTTPIDEEHVEQEKDTENS--------------------------AAVRTLLFTIEDQSSANASTQTDLLPLRHRLVGPIKPPRPVQECLDILNSTEEGSGPAALSDEEIVSIVHAG
BommoHMGR 322 EEQRNWVYDMDDMTVKEVINDTDLS--RKPKFSVGD---------------------------------DSNSEVSTQTDEAGNVEDMEWPTLSPSSSASKLNAKKRPMVECLELYR---SGACT-SLSDEEVIMLVEQ-
BlageHMGR 443 -GHIAGYQLEKVVRNPERGVGIRRQILTKTADL-KDALDNLPYKNYDYLKVMGACCENVIGYMPVPVGVAGPLNLDGRLVHVPLATTEGCLVASTNRGMRALMRCGVTSRIVADGMTRGPVVRFPNIDRASEAMLWMQVP
DippuHMGR 416 -GHVAAYQLEKMVGDPERGVGIRRKILTQKADL-KDALDNLPYKNYDYTKVMGACCENVIGYMPVPVGVAGPLKLDGCLVYVPLATTEGCLVASTNRGSRALMRCGVTSRIVADGMTRGPVVRFPNIERASEAMLWMQAA
ApimeHMGR 472 -NHIAAYQLEKAVGDMERGVEIRRFIIGEAGNF-LDYLSNLPYKDYDYSKVLGACCENVMGYVPVPLGIAGPLLLDGELYYVPMATTEGCLVASTNRGSRALLKCGVTSRVVADGMTRGPVVRFPNIVRASEAMAWMQDP
TricaHMGR 445 -KHIPAYQIEKAVDDPERGVGIRRKILAREGNF-SEALTDLPFRNYDYAKVMGACCENVIGYMPVPVGYAGPLNLDGRHVYVPMATTEGCLVASTNRGCRALLDCGVTSRVVSDGMTRGPVVRFPSITKASEAMSWMKCS
AedaeHMGR 488 NGYCPLYKIETVIGDAERGVKIRRDMIQKEANLPANAFKHLSYKNYDYSKVMNACCENVLGYVQIPVGYAGPLILDGVRYYVPMATTEGALVASTNRGCKAISTRGVTSFVEDIGMTRAPCIKFPNVLRAAQAKRWMETP
DromeHMGR 499 GTHCPLHKIESVLDDPERGVRIRRQIIGSRAKMPVGRLDVLPYEHFDYRKVLNACCENVLGYVPIPVGYAGPLLLDGETYYVPMATTEGALVASTNRGCKALSVRGVRSVVEDVGMTRAPCVRFPSVARAAEAKSWIEND
BommoHMGR 422 -SHIPMHRLEAVLEDPLRGVRLRRRVIASRFNN-ETAIKQLPYLNYDYSKVLNACCENVIGYIGVPVGYAGPLVVDGKPYMIPMATTEGALVASTNRGAKAIGSRGVTSVVEDVGMTRAPAVKLPNVVRAHECRQWIDNK
BlageHMGR 581 YNFEQIKKNFDSTSRFARLSKIHIRVAGRHLFIRFIATTGDAMGMNMLSKGTEVALAYVQQVYPDMEILSLSGNFCTDKKPAAVNWIEGRGKSVVCEAIVPADIIKSVLKTSVQALMDVNITKNLIGSAVAGSIGGFNAH
DippuHMGR 554 QNFEAMKKHFDSTSRYARLSKIHIRVAGRHLFIRFVATTGDAMGMNMLSKGTEVALSFVQQMFPDMEILSLSGNFCTDKKPAAVNWIEGRGKSVVCEAVVPADVVSSVLKTSVQALVDLNITKNFYGSAIAGSVGGNNAH
ApimeHMGR 610 DNFKEMKNSFNLTSRFARLTKINIRIAGRHLFIRFVATTGDAMGMNMLSKGTEKSLNTVKEHFPDMEILSLSGNFCTDKKPAAVNWVCGRGKSVVCEAVVPADIVTNVLKTSVHALVDVNISKNMIGSAIAGSIGGFNAH
TricaHMGR 583 QNFEAMKQQFDSSSRFARLSKLLIKIAGRHLFVRFEAKTGDAMGMNMVSKGTEMSLKYVQKQFPEMEILSLSGNFCTDKKPAAVNWIEGRGKSVVCEAIIPSEIVKKVLKTSTPALVDVNNSKNMIGSAVAGSIGGFNAH
AedaeHMGR 628 ENFAVIKKAFDSTSRFARLQELHIAMDGPILYARFRALTGDAMGMNMVSKGSEMALREVHRSFPDMQIISLSGNFCSDKKPAAINWIKGRGKRVICEAIVPADKLRTILKTNARTLVQCNKLKNMTGSAVAGSIGGNNAH
DromeHMGR 639 ENYRVVKTEFDSTSRFGRLKDCHIAMDGPQLYIRFVAITGDAMGMNMVSKGAEMALRRIQLQFPDMQIISLSGNFCCDKKPAAINWIKGRGKRVVTECTISAATLRSVLKTDAKTLVECNKLKNMGGSAMAGSIGGNNAH
BommoHMGR 560 ENYALLKEAFDSTSRFARLQEIHVGVDGATLYLRFRATTGDAMGMNMVSKGAENALKLLKTFFRDMEVISLSGNYCSDKKAAAINWIKGRGKRVVCETVISSENLRTIFKTDAKTLSRCNKIKNLSGSALAGSIGGNNAH
BlageHMGR 721 AANIVTAIFIATGQDPAQNVGSSNCMTLMEPWGEDGKDLYVSCTMPSIEIGTIGGGTVLPPQAACLDMLGVRGANEMCPGENANTLARIVCGTVLAGELSLMSALAAGHLVKSHMRHNRSSVSTS----------GSEPS
DippuHMGR 694 AANIVTAIFIATGQDPAQNVGSSNCITIMEPWGDDKKDLYVSCSMPSIEIGTVGGGTILPPQAACLDMLGVKGANAVCPGENANMLARIVCGTVLAGELSLMSALAAGHLVKSHLRHNRSSVTTS----------GSEPS
ApimeHMGR 750 AANIVTAIFIATGQDPAQNVGSSNCMTLMEPWGTDGSDLYVSCTMPSIEIGTVGGGTILPAQGACLSILGVKGAHSDEPGENASRLARIVCATVLAGELSLMAALTAGHLVKSHLRHNRSSTTVTNAMSVPQKYTGTKLS
TricaHMGR 723 AANIVTAIYIATGQDPAQNIGSSNCMTLMEPWGETGEDLYVSCTMPSIEIGTVGGGTVLPAQSSCLEMLRVKGSHPDCPGENASQLARIVCGTVLAGELSLMAALTAGHLVRSHLRHNRSTTTIP-----------EEFS
AedaeHMGR 768 AANMVTAIYIATGQDPAQNVTSSNCSTNMEPYGDNGEDLYMTCTMPSLEVGTVGGGTGLPGQGACLDMLGVRGAHPTHPAENSKQLARVICATVMAGELSLMAALVNSDLVKSHMRHNRSSVAVNPG----LPATAQLQS
DromeHMGR 779 AANMVTAVFLATGQDPAQNVTSSNCSTAMECWAENSEDLYMTCTMPSLEVGTVGGGTGLPGQSACLEMLGVRGAHATRPGDNAKKLAQIVCATVMAGELSLMAALVNSDLVKSHMRHNRSSIAVN--------------S
BommoHMGR 700 AANMVTAIFIATGQDPAQNVTSSNCSTNMEAYGENGEDLYVTCTMPSLEVGTVGGGTVLTGQGACLEILGVKGAG-TRPAENSARLASLICATVLAGELSLMAALVNSDLVKSHMRHNRSTLNVQ------------TAN
BlageHMGR 851 TP--------ACKS-------------
DippuHMGR 824 KS-------------------------

97
ApimeHMGR 890 VPNLLQPVQNVCKGLLEKS--------
TricaHMGR 852 QNRYHIP---PCKDI------------
AedaeHMGR 904 APSLLTACNSSSSGSSSSIGTSLSAKQ
DromeHMGR 905 ANNPLNVTVSSCSTIS-----------
BommoHMGR 827 VEPYTVALKVPPS--------------
B.
SchgrJHAMT 1 MDKAELYSRSNGLQRWEASAALEAAWPALRWPAPP-LRVLDVGCGAGDVTVDLLLPRLPP-HTQLVGTDVSAAMVEHAAELYGAAHPGLSFQLLDIADPDIDASPVYQLAPFDKIFSFFCLHWVPEQRQAAENLHRLLKP
DippuJHAMT 1 MHKAELYSSSHGLQKRDAAHALTEYLDHMTWR-PG-DRVLDVGCGPGFVTAQELMPRLPQDFAILVGTDVSHAMVQHATSTY--VQPKLKFAHLDISSTHIDK-ELWEPG-FDKIFSFYCLHWIPDQRTAVNNIYHLLRP
TricaJHAMT 1 MNKASLYSKYSGLQKNDASFVIDNYLRLIKWK-PN-ANILDIGSGDGNVIFELLLPKIPKHFAKFVGTDISEEMVLFAKNQCN--DPKIDFLQMDIS----ATIPPEFHEYFDHIFSFYCLHWVVEQRQAMKNIFDMLKP
DromeJHAMT 1 MNQASLYQHANQVQRHDAKLILDEFASTMQWRSDGEDALLDVGSGSGNVLMDFVKPLLP-IRGQLVGTDISSQMVHYASKHYQR-EERTRFQVLDIGCER-LPEELS--GRFDHVTSFYCLHWVQNLKGALGNIYNLLKP
AedaeJHAMT 1 MNKPNLYHRANGVQRRDAKEILDEHGHLLRWKEENEDSLLDIGCGSGDVLIDFVIPMVPPKRARVLGTDVSEQMVRFARKVHSD-VENLFFETLDIEGD--ISSFLNKWGCFDHITSFYCLHWVRSQRSAFSNIYNLMAP
BommoJHAMT 1 MNNADLYRKSNSLQKRDALRCLEEHANKIKWKKIG-DRVIDLGCADG-SVTDILKVYMPKNYGRLVGCDISEEMVKYANKHHG--FGRTSFRVLDIEGD--LTADLK--QGFDHVFSFYTLHWIRDQERAFRNIFNLLGD
ApimeJHAMT 1 MFLVEEYVKASTIQYRDAADIIGEFAEEMSEMKGK---CLDIGCGPGIVTKELILPNLSP-EAKLVGMDISRPMIEYAKNMYHD-EERLSFQLLDIET---MDLPKDTFDQFNNVLSFYCLHWCQNFRKAFDNIYKLLRP
SchgrJHAMT 139 GG-EVVLSLLAHCPIFSVYEGLAHKPQWKEYMEDARRFISPYHHSEDPAREMNELLCRAGFRVTLCTRQQRSFTFPGHSALIEAVTAVNPFVERLPETLQQEFLEDCMKEVLRQKLVSIEDDADSNNNSNGSNRSGNNAV
DippuJHAMT 135 AG-EALVLLMAKCPVFNVYTAQSNKPKWQQYMKDASRYISPYHQLKDPKSEFINIVEDVGFHVVDCDCRQNKFNYGTLERLKDAIKAVNPFMDRIPEELQEEYLNDCLSEARRIKCT------ESNNN-----------V
TricaJHAMT 133 GG-EMLLTFLASNPIYDIYERMAKSNKWGPYMNNLKKYISPYHHSEDPETELENLLKKEGFITHLCRVENRSYTFPSFSVLSKSVSAVNPFIKKLPENEIDTYIEDYLKEVRKLKTIT----IETCNN------------
DromeJHAMT 136 EGGDCLLAFLASNPVYEVYKILKTNDKWSTFMQDVENFISPLHYSLSPGEEFSQLLNDVGFVQHNVEIRNEVFVYEGVRTLKDNVKAICPFLERMPADLHEQFLDDFIDIVISMNLQQ-------GEN------------
AedaeJHAMT 138 NG-DCLLGFLARNPIFDIYDQLSNSAKWSMYMTDVDKYISPYQYCENPVGEIEEILSSVGFTKYKIHIADKIYVYEGIDSLKKAVQAVNPFSERMPLDLQEDFLNDYIAVVRRMSLSEN-----CCGN------------
BommoJHAMT 133 EG-DCLLLFLGHTPIFDVYRTLSHTEKWHSWLEHVDRFISPYHDNEDPEKEVKKIMERVGFSNIEVQCKTLFYVYDDLDVLKKSVAAINPFN--IPKDILEDFLEDYIDVVREMRLLDR-----CNNN------------
ApimeJHAMT 133 GG-KGLFMLLSWNDGFDVYKKLYANPRYRPYMQEPERFIPIFHECKDRRVNLRKILETTGFEILHCSEREKSYIYKNSEIMKKHIMAINPFISRIPNSLKKEFEDEITREIVNMKIQLLN----KDEN------------
SchgrJHAMT 278 DANSKKLTTRYSVLTVVAAKAAAQNGVRTVR-----------
DippuJHAMT 257 EEVT---TVSYDIIVAHIKKP---------------------
TricaJHAMT 256 NDNEEKIHVPYKLFVTFASKPV--------------------
DromeJHAMT 257 NEDQKFLSP-YKLVVAYARKTPEFVNNVFLEPTHQNLVKGIN
AedaeJHAMT 260 ENDYKFITP-YKLVVVYAVK----------------------
BommoJHAMT 253 VGESVSIKFNYKVISVYARKLCLSLM----------------
ApimeJHAMT 256 GEQEYNILDRYQIFVTYIRKPVC-------------------
Figure S2.1. Amino acid sequence alignment of Diploptera proteins with several functionally characterized ortholog insect
proteins. (A) DippuHMGR with HMGR from the German cockroach Blattella germanica (GenBank accession number: CAA49628.1
(Martinezgonzalez et al., 1993), the honeybee Apis mellifera (GenBank accession number: XP_623118.1), the red flour beetle
Tribolium castaneum (GenBank accession number: XP_973850.1), the mosquito Aedes aegypti (GenBank accession number:
XP_001659923.1), the fly Drosophila melanogaster (GenBank accession number: NP_732900.1) and the silkworm Bombyx mori
(GenBank accession number: BAF62108.1 (Kinjoh et al., 2007)) (B) DippuJHAMT with JHAMT from the desert locust, S. gregaria
(GenBank accession number: ADV17350.1 (Marchal et al., 2011)), the red flour beetle T. castaneum (GenBank accession number:
NP_001120783.1 (Minakuchi et al., 2008)), the fly D. melanogaster (GenBank accession number: AB113579.1 (Niwa et al., 2008)),
the mosquito A. aegypti (GenBank accesson number: ABD65474.1 (Mayoral et al., 2009b)), the silkworm B. mori (GenBank
accession number: NP_001036901.1 (Shinoda and Itoyama, 2003)) and the honeybee A. mellifiera (GenBank accession number:
AGG79412.1 (Li et al., 2013)).

98
PM
K
PP
MD
IPP
I
FO
LD
Th
iol
HM
GS
HM
GR
MK
FP
PS
JH
AM
T
CY
P15A
1
0
1 0
2 0
5 0
1 0 0
1 5 0
2 0 0
5 0 0
1 0 0 0
1 5 0 0
2 0 0 0
2 5 0 0
3 0 0 0
3 5 0 0
Re
lati
ve
mR
NA
qu
an
tity
Figure S2.2. Comparison of transcript levels of genes encoding JH biosynthetic enzymes in
CA of day 4 mated female. Bars represent the mean of three biologically independent pools of
ten animals run in triplicate and normalized to Tubulin and EF1α. Vertical error bars indicate
SEM. Enzyme abbreviations are as in Fig. 2.1.

99
Table S2.1. (Degenerate) primer sequences for cloning of (partial) sequences for orthologous genes encoding JH biosynthesis
enzymes in D. punctata. Gene name abbreviations were shown in Fig. 2.1.
Gene name F Primer(5'-3') R Primer(5'-3') Accession number (NCBI)
Thiol GGDCARAATCCWGCDAGRCARGC TTGHGCWGCRAANGCTTCRTT KJ188021
HMGS AGAGTGAACTGGAAGTGCA TATGAAGCACGCACTCCTC KJ188022
HMGR AATGGTAGGGTGGCTGTTTCTA GTATTTCATGATTTGCTGGGTTC KJ188023
MK CTGCCCCCGGTAAAGTTATC GATGCTTGACTTCCTGCACC KJ188024
PMK ATGGAAGTAAACAAAATTTCAG CACTCTGTTTCTGCATCATCTAC KJ188025
PPMD TGTTCAGAAAAYAATTTYCCNAC TGATTNSWRTCYTTCATTGT KJ188026
IPPI CAYMGDGCVTTYAGTKTDTTY TCDAWYTCRTGTTCKCCCCA KJ188027
FPPS TGGCGCGTAGGGTAGACAA GATCCATAACATTCTGCAAACATTG KJ188028
FOLD ATGCAGCGTTGGACTGGA TCCAGCATTGTTGATGAGAA KJ188029
JHAMT ATGCACAAAGCAGAACTGTATTC TCAAGGCTTTTTAATATGAGCGACAAT KJ188030
CYP15A1 ATGGTCATCGCTCTTATTGTCATC ACCATTCATTTCCTTGGAATCAACT AY509244
Vg TGGAACRCDCTICTCTGYTGYCT TTKAKSAYGTTRAYTTCCCAG KJ188031

100
References
Belles, X., Martin, D., Piulachs, M.D., 2005. The mevalonate pathway and the synthesis of juvenile hormone
in insects. Annu Rev Entomol 50, 181-199.
Bomtorin, A.D., Mackert, A., Rosa, G.C., Moda, L.M., Martins, J.R., Bitondi, M.M., Hartfelder, K., Simoes,
Z.L., 2014. Juvenile hormone biosynthesis gene expression in the corpora allata of honey bee (Apis mellifera
L.) female castes. PLoS One 9, e86923.
Bradford, M.M., 1976. A rapid and sensitive method for the quantitation of microgram quantities of protein
utilizing the principle of protein-dye binding. Anal Biochem 72, 248-254.
Cao, L., Zhang, P., Grant, D.F., 2009. An insect farnesyl phosphatase homologous to the N-terminal domain of
soluble epoxide hydrolase. Biochem Biophys Res Commun 380, 188-192.
Chang, T.Y., Limanek, J.S., 1980. Regulation of cytosolic acetoacetyl coenzyme A thiolase, 3-hydroxy-3-
methylglutaryl coenzyme A synthase, 3-hydroxy-3-methylglutaryl coenzyme A reductase, and mevalonate
kinase by low density lipoprotein and by 25-hydroxycholesterol in Chinese hamster ovary cells. J Biol Chem
255, 7787-7795.
Cheng, D., Meng, M., Peng, J., Qian, W., Kang, L., Xia, Q., 2014. Genome-wide comparison of genes
involved in the biosynthesis, metabolism, and signaling of juvenile hormone between silkworm and other
insects. Genet Mol Biol 37, 444-459.
Clarke, C.F., Tanaka, R.D., Svenson, K., Wamsley, M., Fogelman, A.M., Edwards, P.A., 1987. Molecular
cloning and sequence of a cholesterol-repressible enzyme related to prenyltransferase in the isoprene
biosynthetic pathway. Mol Cell Biol 7, 3138-3146.
Couillaud, F., Feyereisen, R., 1991. Assay of HMG-CoA synthase in Diploptera punctata corpora allata. Insect
Biochem 21, 131-135.
Debernard, S., Rossignol, F., Couillaud, F., 1994. The HMG-CoA reductase inhibitor fluvastatin inhibits insect
juvenile hormone biosynthesis. Gen Comp Endocrinol 95, 92-98.
Defelipe, L.A., Dolghih, E., Roitberg, A.E., Nouzova, M., Mayoral, J.G., Noriega, F.G., Turjanski, A.G., 2011.
Juvenile hormone synthesis: "esterify then epoxidize" or "epoxidize then esterify"? Insights from the structural
characterization of juvenile hormone acid methyltransferase. Insect Biochem Mol Biol 41, 228-235.
Feyereisen, R., Farnsworth, D.E., 1987. Characterization and regulation of HMG-CoA reductase during a cycle
of juvenile hormone synthesis. Mol Cell Endocrinol 53, 227-238.
Feyereisen, R., Tobe, S.S., 1981. A rapid partition assay for routine analysis of juvenile hormone release by
insect corpora allata. Anal Biochem 111, 372-375.
Goldstein, J.L., Brown, M.S., 1990. Regulation of the mevalonate pathway. Nature 343, 425-430.
Goodman, W.G., Cusson, M., 2012. The Juvenile Hormones, in: Gilbert, L.I. (Ed.), Insect Endocrinology, pp.
310-365.
Granger, N.A., Sturgis, S.L., Ebersohl, R., Geng, C., Sparks, T.C., 1996. Dopaminergic control of corpora
allata activity in the larval tobacco hornworm, Manduca sexta. Arch Insect Biochem Physiol 32, 449-466.
Hartfelder, K., 2000. Insect juvenile hormone: from "status quo" to high society. Braz J Med Biol Res 33, 157-
177.
Helvig, C., Koener, J.F., Unnithan, G.C., Feyereisen, R., 2004. CYP15A1, the cytochrome P450 that catalyzes
epoxidation of methyl farnesoate to juvenile hormone III in cockroach corpora allata. Proc Natl Acad Sci USA
101, 4024-4029.

101
Huang, J., Marchal, E., Hult, E.F., Zels, S., Vanden Broeck, J., Tobe, S.S., 2014. Mode of action of
allatostatins in the regulation of juvenile hormone biosynthesis in the cockroach, Diploptera punctata. Insect
Biochem Mol Biol 54C, 61-68.
Kaneko, Y., Hiruma, K., 2014. Short neuropeptide F (sNPF) is a stage-specific suppressor for juvenile
hormone biosynthesis by corpora allata, and a critical factor for the initiation of insect metamorphosis. Dev
Biol 393, 312-319.
Kinjoh, T., Kaneko, Y., Itoyama, K., Mita, K., Hiruma, K., Shinoda, T., 2007. Control of juvenile hormone
biosynthesis in Bombyx mori: cloning of the enzymes in the mevalonate pathway and assessment of their
developmental expression in the corpora allata. Insect Biochem Mol Biol 37, 808-818.
Li, W.F., Huang, Z.Y., Liu, F., Li, Z.G., Yan, L.M., Zhang, S.W., Chen, S.L., Zhong, B.X., Su, S.K., 2013.
Molecular cloning and characterization of juvenile hormone acid methyltransferase in the honey bee, Apis
mellifera, and its differential expression during caste differentiation. PLoS One 8.
Marchal, E., Hult, E.F., Huang, J., Pang, Z., Stay, B., Tobe, S.S., 2014. Methoprene-tolerant (Met) knockdown
in the adult female cockroach, Diploptera punctata completely inhibits ovarian development. PLoS One 9,
e106737.
Marchal, E., Hult, E.F., Huang, J., Stay, B., Tobe, S.S., 2013a. Diploptera punctata as a model for studying the
endocrinology of arthropod reproduction and development. Gen Comp Endocr 188, 85-93.
Marchal, E., Hult, E.F., Huang, J., Tobe, S.S., 2013b. Sequencing and validation of housekeeping genes for
quantitative real-time PCR during the gonadotrophic cycle of Diploptera punctata. BMC Res Notes 6, 237.
Marchal, E., Zhang, J., Badisco, L., Verlinden, H., Hult, E.F., Van Wielendaele, P., Yagi, K.J., Tobe, S.S.,
Vanden Broeck, J., 2011. Final steps in juvenile hormone biosynthesis in the desert locust, Schistocerca
gregaria. Insect Biochem Mol Biol 41, 219-227.
Martinezgonzalez, J., Buesa, C., Piulachs, M.D., Belles, X., Hegardt, F.G., 1993. Molecular cloning,
developmental pattern and tissue expression of 3-hydroxy-3-methylglutaryl coenzyme a reductase of the
cockroach Blattella germanica. European Journal of Biochemistry 213, 233-241.
Mayoral, J.G., Nouzova, M., Navare, A., Noriega, F.G., 2009a. NADP(+)-dependent farnesol dehydrogenase,
a corpora allata enzyme involved in juvenile hormone synthesis. Proc Natl Acad Sci U S A 106, 21091-21096.
Mayoral, J.G., Nouzova, M., Yoshiyama, M., Shinod, T., Hernandez-Martinez, S., Dolghih, E., Turjanski,
A.G., Roitberg, A.E., Priestap, H., Perez, M., Mackenzie, L., Li, Y.P., Noriega, F.G., 2009b. Molecular and
functional characterization of a juvenile hormone acid methyltransferase expressed in the corpora allata of
mosquitoes. Insect Biochem Mol Biol 39, 31-37.
Mayoral, J.G., Nouzova, M., Yoshiyama, M., Shinoda, T., Hernandez-Martinez, S., Dolghih, E., Turjanski,
A.G., Roitberg, A.E., Priestap, H., Perez, M., Mackenzie, L., Li, Y., Noriega, F.G., 2009c. Molecular and
functional characterization of a juvenile hormone acid methyltransferase expressed in the corpora allata of
mosquitoes. Insect Biochem Mol Biol 39, 31-37.
Meigs, T.E., Roseman, D.S., Simoni, R.D., 1996. Regulation of 3-hydroxy-3-methylglutaryl-coenzyme A
reductase degradation by the nonsterol mevalonate metabolite farnesol in vivo. J Biol Chem 271, 7916-7922.
Meigs, T.E., Simoni, R.D., 1997. Farnesol as a regulator of HMG-CoA reductase degradation: characterization
and role of farnesyl pyrophosphatase. Arch Biochem Biophys 345, 1-9.
Minakuchi, C., Namiki, T., Yoshiyama, M., Shinoda, T., 2008. RNAi-mediated knockdown of juvenile
hormone acid O-methyltransferase gene causes precocious metamorphosis in the red flour beetle Tribolium castaneum. Febs Journal 275, 2919-2931.
Monger, D.J., Lim, W.A., Kezdy, F.J., Law, J.H., 1982. Compactin inhibits insect HMG-CoA reductase and
juvenile hormone biosynthesis. Biochem Biophys Res Commun 105, 1374-1380.

102
Mundall, E.C., Tobe, S.S., Stay, B., 1981. Vitellogenin fluctuations in haemolymph and fat body and dynamics
of uptake into oöcytes during the reproductive cycle of Diploptera punctata. J Insect Physiol 27, 821-827.
Niwa, R., Niimi, T., Honda, N., Yoshiyama, M., Itoyama, K., Kataoka, H., Shinoda, T., 2008. Juvenile
hormone acid O-methyltransferase in Drosophila melanogaster. Insect Biochem Mol Biol 38, 714-720.
Noriega, F.G., Ribeiro, J.M., Koener, J.F., Valenzuela, J.G., Hernandez-Martinez, S., Pham, V.M., Feyereisen,
R., 2006. Comparative genomics of insect juvenile hormone biosynthesis. Insect Biochem Mol Biol 36, 366-
374.
Nouzova, M., Edwards, M.J., Mayoral, J.G., Noriega, F.G., 2011. A coordinated expression of biosynthetic
enzymes controls the flux of juvenile hormone precursors in the corpora allata of mosquitoes. Insect Biochem
Mol Biol 41, 660-669.
Nyati, P., Nouzova, M., Rivera-Perez, C., Clifton, M.E., Mayoral, J.G., Noriega, F.G., 2013. Farnesyl
phosphatase, a corpora allata enzyme involved in juvenile hormone biosynthesis in Aedes aegypti. PLoS One 8.
Pratt, G.E., Davey, K.G., 1972. Corpus Allatum and Oogenesis in Rhodnius-Prolixus (Stal) .1. Effects of
Allatectomy. Journal of Experimental Biology 56, 201-214.
Pszczolkowski, M.A., Lee, W.S., Liu, H.P., Chiang, A.S., 1999. Glutamate-induced rise in cytosolic calcium
concentration stimulates in vitro rates of juvenile hormone biosynthesis in corpus allatum of Diploptera
punctata. Mol Cell Endocrinol 158, 163-171.
Rankin, S.M., Stay, B., 1984. The changing effect of the ovary on rates of juvenile hormone synthesis in
Diploptera punctata. Gen Comp Endocr 54, 382-388.
Rivera-Perez, C., Nouzova, M., Clifton, M.E., Garcia, E.M., LeBlanc, E., Noriega, F.G., 2013. Aldehyde
dehydrogenase 3 converts farnesal into farnesoic acid in the corpora allata of mosquitoes. Insect Biochem Mol
Biol 43, 675-682.
Rivera-Perez, C., Nouzova, M., Noriega, F.G., 2012. A quantitative assay for the juvenile hormones and their
precursors using fluorescent tags. PLoS One 7, e43784.
Shinoda, T., Itoyama, K., 2003. Juvenile hormone acid methyltransferase: a key regulatory enzyme for insect
metamorphosis. Proc Natl Acad Sci U S A 100, 11986-11991.
Stay, B., Tobe, S.S., 1978. Control of juvenile hormone biosynthesis during reproductive cycle of a viviparous
cockroach 2. Effects of unilateral allatectomy, implantation of supernumerary corpora allata, and ovariectomy.
Gen Comp Endocr 34, 276-286.
Stoltzman, C.A., Stay, B., 1997. Gonadotrophic and morphogenetic effects of a juvenile hormone analog
treatment and ovary presence on last instar male and female Diploptera punctata (Blattaria: Blaberidae). Eur J
Entomol 94, 335-348.
Thompson, C.S., Yagi, K.J., Chen, Z.F., Tobe, S.S., 1990. The effects of octopamine on juvenile hormone
biosynthesis, electrophysiology, and cAMP content of the corpora allata of the cockroach Diploptera punctata.
J Comp Physiol B 160, 241-249.
Tobe, S.S., Clarke, N., 1985. The effect of L-methionine concentration on juvenile hormone biosynthesis by
corpora allata of the cockroach Diploptera punctata. Insect Biochem 15, 175-179.
Tobe, S.S., Ruegg, R.P., Stay, B.A., Baker, F.C., Miller, C.A., Schooley, D.A., 1985. Juvenile hormone titer
and regulation in the cockroach Diploptera punctata. Experientia 41, 1028-1034.
Tobe, S.S., Stay, B., 1979. Modulation of juvenile hormone synthesis by an analog in the cockroach. Nature
281, 481-482.
Ueda, H., Shinoda, T., Hiruma, K., 2009. Spatial expression of the mevalonate enzymes involved in juvenile
hormone biosynthesis in the corpora allata in Bombyx mori. J Insect Physiol 55, 798-804.

103
Vandesompele, J., De Preter, K., Pattyn, F., Poppe, B., Van Roy, N., De Paepe, A., Speleman, F., 2002.
Accurate normalization of real-time quantitative RT-PCR data by geometric averaging of multiple internal
control genes. Genome Biol 3.
Wang, Z., Dong, Y., Desneux, N., Niu, C., 2013. RNAi silencing of the HaHMG-CoA reductase gene inhibits
oviposition in the Helicoverpa armigera cotton bollworm. PLoS One 8, e67732.
Weaver, R.J., Audsley, N., 2009. Neuropeptide regulators of juvenile hormone synthesis: structures, functions,
distribution, and unanswered questions. Ann N Y Acad Sci 1163, 316-329.

104
Chapter 3
Mode of action of allatostatins in the regulation of juvenile hormone
biosynthesis in the cockroach, Diploptera punctata
This chapter is an adapted reprint of my article:
Juan Huang, Elisabeth Marchal, Ekaterina F. Hult, Sven Zels, Jozef Vanden Broeck, Stephen S.
Tobe, Mode of action of allatostatins in the regulation of juvenile hormone biosynthesis in the
cockroach, Diploptera punctata, Insect Biochem Mol Biol, 2014, 54, 61-68.
Authors’ contribution:
I designed and performed all experiments in this study and wrote the paper with editing by E.
Marchal, E.F. Hult, and S.S. Tobe.

105
Summary
The FGLamide allatostatins (FGL/ASTs) are a family of neuropeptides with pleiotropic
functions, including the inhibition of juvenile hormone (JH) biosynthesis, vitellogenesis and
muscle contraction. In the cockroach, Diploptera punctata, thirteen FGLa/ASTs and one
allatostatin receptor (AstR) have been identified. However, the mode of action of ASTs in
regulation of JH biosynthesis remains unclear. Here, we determined the tissue distribution of
Dippu-AstR and Dippu-AST. The transcript level of Dippu-AstR in the CA corresponds to the JH
biosynthesis, while the expression of Dippu-AST mRNA in the brain did not show any
significant change during the first gonadotrophic cycle. In addition, silencing Dippu-AstR results
in a significantly increase of JH biosynthesis, while the knockdown of Dippu-AST has no effect.
These results suggest that it is the change of AST receptor in the CA, instead of AST levels in
the brain that control the AST-mediated inhibition of JH production.
To determine the signal pathway of Dippu-AstR, we expressed Dippu-AstR in vertebrate cell
lines, and activated the receptor with the Dippu-ASTs. Our results show that all thirteen ASTs
activated Dippu-AstR in a dose dependent manner, albeit with different potencies. Functional
analysis of AstR in multiple cell lines demonstrated that activation of the AstR receptor resulted
in elevated levels of Ca2+ and cAMP, which suggests that Dippu-AstR can act through the Gαq
and Gαs protein pathways. The study on the target of AST action reveals that FGL/AST affects
JH biosynthesis prior to the entry of acetyl-CoA into the JH biosynthetic pathway.
Introduction
Allatostatins (ASTs), a family of pleiotropic neuropeptides, were originally named for their
ability to inhibit juvenile hormone (JH) biosynthesis by corpora allata (CA) rapidly and

106
reversibly (Bendena et al., 1999). Three families of ASTs have been identified in insects and
named as: FGLa/ASTs (A-type), MIP/ASTs (B-type) and the PISCF/ASTs (C-type) (Coast and
Schooley, 2011). The distribution and function of the three families of ASTs has been reviewed
by Stay and Tobe (Stay and Tobe, 2007). The best documented role of FGLa/AST is their ability
to inhibit juvenile hormone (JH) biosynthesis by corpora allata (CA) (Stay and Tobe, 2007).
Later studies demonstrated other functions of FGLa/ASTs, including regulation of myotropic
activity in gut tissues (Duve et al., 1995; Lange et al., 1995) and cardiac rhythm (Vilaplana et al.,
1999), inhibition of vitellogenin synthesis in the fat body (Martin et al., 1996), and stimulation of
enzyme activity in the lumen of the midgut (Fuse et al., 1999).
As neuropeptides, ASTs exert their effects by binding to a G protein-coupled receptor, AstR.
FGLa/AST receptors, which are structurally related to the mammalian galanin receptor, were
first identified in the fly Drosophila melanogaster DAR-1 (Birgul et al., 1999; Lenz et al., 2000)
and DAR-2 (Birgul et al., 1999; Lenz et al., 2000), and later in the silkworm, Bombyx mori
(Secher et al., 2001) and the stick insect, Carausius morosus (Auerswald et al., 2001). In D.
punctata, putative AstRs have previously been partially characterized using photoaffinity
labeling and a radioligand-binding assay in the CA and brain (Cusson et al., 1991, 1992; Yu et
al., 1995). Lungchukiet et al (Lungchukiet et al., 2008b) identified a putative FGLa/AST
receptor gene in D. punctata. Silencing this gene resulted in a significant increase in JH
biosynthesis (Lungchukiet et al., 2008a; Lungchukiet et al., 2008b).
The responses of FGLa/AST receptor to FGLa/ASTs have been assayed in D. melanogaster
(DAR-1 and DAR-2) (Birgul et al., 1999; Larsen et al., 2001), B. mori (Secher et al., 2001) and
the cockroach, Periplaneta americana (Gade et al., 2008). The studies in B. mori and P.
americana focused on the response of AstR to exogenous ASTs. The signaling pathway of AstR

107
was, however, not completely elucidated. Larsen et al (Larsen et al., 2001) expressed Drosophila
FGLa/AST receptors, DAR-1 and DAR-2 in CHO cells and activated them with four putative
FGLa/ASTs from D. melanogaster AST and four FGLa/ASTs from D. punctata in the presence
or absence of pertussis toxin (PTX). They found that PTX caused a complete loss of Ca2+ signal
in cells expressing DAR-1, and a decreased Ca2+ signal in cells expressing DAR-2. Their results
suggested that the activation of DAR-1 and DAR-2 by FGLa/ASTs coupled to multiple signaling
pathways, including Gi/o protein and other, PTX-insensitive G-proteins.
Although the function of AST has been well-studied, little is known about the precise target of
AST action. Previous studies focused on select enzymes in the JH biosynthetic pathway. It is
now known that thirteen enzymes are involved in the JH biosynthetic pathway (Fig. 2.1) (Belles
et al., 2005; Nouzova et al., 2011). The potential targets of action of ASTs were originally
studied by employing different known JH precursors (Pratt et al., 1991; Pratt et al., 1989). The
results suggested that the inhibitory action of AST on JH biosynthesis resides in step(s) prior to
mevalonate. However, neither HMG-CoA synthase nor HMG-CoA reductase activity was
affected by ASTs (Sutherland and Feyereisen, 1996). These authors proposed that the target of
AST action on JH biosynthesis is in step(s) prior to the JH biosynthetic pathway, and may be
related to the transport of citrate from mitochondria to cytosol and/or to the cleavage of citrate to
yield acetyl-CoA.
Our study focuses on the viviparous cockroach, Diploptera punctata, in which ASTs were first
characterized. The first gonadotropic cycle is characterized by a precise regulation of JH
biosynthesis necessary to coordinate a specific series of reproductive events closely correlated
with oocyte growth. This makes D. punctata an ideal model for a more in depth study of the
mode of action of AST and its signal transduction pathway. We have analyzed the spatial

108
expression pattern of the AST precursor and AstR which is consistent with their roles in
regulating JH biosynthesis. Moreover, by using an aequorin-based assay and expressing the
receptor in a mammalian system, we have unambiguously identified that AST is a ligand for the
candidate AstR (Lungchukiet et al., 2008a; Lungchukiet et al., 2008b) and that this receptor can
couple to Ca2+ and cAMP.
Thirteen FGLa/ASTs have been identified in D. punctata. These peptides share a conserved C-
terminal Tyr (Phe)- Xaa-Phe-Gly-Leu-NH2, which is believed to be the main functional region
for the inhibition of JH biosynthesis (Donly et al., 1993; Marchal et al., 2013a). The FGLa/ASTs
inhibit JH biosynthesis by CA at low concentrations in vitro but with different potencies (Tobe et
al., 2000). Our study has also examined the relationship between binding affinity and potency
with a view to understanding the sites of action of the peptides.
The precise target of AST action remains unclear so far. Our study did not find any significant
changes in the transcript level of genes encoding enzymes in JH biosynthetic pathway. The
rescue of AST-induced JH inhibition by JH precursors suggest that the target of AST action is
prior to the entry of Acetyl-CoA into the JH biosynthetic pathway.
Materials and Methods
Insects - D. punctata were reared in cages and fed with lab chow and water at libitum at 27 ºC in
a dark room. Newly molted female adult cockroaches were picked from the colony and raised in
separate containers. Mated status was confirmed by the presence of a spermatophore.
Tissue collection - Cockroach tissues were dissected under a dissecting microscope. Basal oocyte
length was measured to determine the physiological age of cockroaches. Selected tissues were
dissected and cleaned in sterile cockroach ringer solution (150 mM NaCl, 12 mM KCl, 10 mM

109
CaCl2.2H2O, 3 mM MgCl2.6H2O, 10 mM HEPES, 40 mM Glucose, pH 7.2), and stored at -80 ºC
to prevent degradation.
RNA extraction and cDNA synthesis - Pooled samples were homogenized using a plastic pestle
and RNA was extracted using the RNeasy Mini Kit (Qiagen) according to the manufacturer’s
instructions. An additional DNase treatment (RNase-free DNase set, Qiagen) was performed to
eliminate potential genomic DNA contamination. RNA of CA was extracted using the
RNAqueous®-Micro Kit (Ambion). DNase treatment was performed to eliminate genomic DNA
contamination. The quantity and quality of RNA was determined using a Nanodrop
spectrophotometer (Thermo Scientific). An equal amount of RNA was transcribed with
Superscript III reverse transcriptase (Invitrogen Life Technologies) utilizing random hexamers as
described in the protocol. The resulting cDNA was diluted tenfold.
Quantitative Real Time-PCR (q-RT-PCR) - Prior to target gene profiling, Tubulin and EF1a were
chosen as the optimal housekeeping genes according to a previous study (Marchal et al., 2013b).
The q-RT-PCR reactions were performed in triplicate on a CFX384 Touch ™ Real-Time PCR
Detection System (Bio-Rad) in a final volume of 10 µl, containing 1 µl of cDNA, 5 µl IQ™
SYBR® Green Supermix (Bio-Rad), 1 µl forward and reverse primer (5 µM) and 2 µl of MQ-
water. The reaction was incubated for 3 min in 95ºC, followed by 40 cycles with following
thermal profile: 95ºC, 10 s; 59ºC, 30 s. Target specificity was confirmed by performing a
dissociation protocol (melt curve analysis) and running a few representative q-RT-PCR products
on an agarose gel containing GelRed™ (Biotium). Realtime primers used for q-RT-PCR are
listed in Table S3.1. Primer sets were validated by determining relative standard curves for each
gene transcript using a five-fold serial dilution of a calibrator cDNA sample. Efficiency and
correlation coefficient (R²) are shown in Table S1. The primer sets for genes in the JH

110
biosynthetic pathway were chosen as in Chapter 2. The quantity of mRNA for each tested gene
relative to reference genes was determined as described by Vandesompele et al. (Vandesompele
et al., 2002).
Peptides and substrates - Thirteen Dippu-ASTs were custom synthesized by GL Biochemical
Ltd., Shanghai (China). Peptides were purified by high performance liquid chromatography
(HPLC) (purity ≥95%). Peptides were dissolved in water to obtain a concentration of 1mM.
Peptide solutions were stored at -80 ºC prior to further processing and dilution.
Acetyl-CoA, mevalonic acid (MA), diphosphomevalonate (DPPM) and farnesol were purchased
from Sigma-Aldrich Canada. Substrates were dissolved in water before use.
Cell culture and transfection - Chinese hamster ovary (CHO) WTA11 and PAM28 cells stably
expressing apoaequorin (Euroscreen, Belgium) and human embryonic kidney (HEK) 293 cells
were cultured in monolayers in Dulbecco’s Modified Eagles Medium nutrient mixture F12-Ham
(DMEM/F12) (Sigma) supplemented with 10% heat-inactivated fetal calf serum (Invitrogen),
100 IU/ml penicillin and 100 µg/ml streptomycin (Invitrogen). An additional 250 µg/ml Zeocin
(Invitrogen) was added to the medium for CHO-WTA11 cells, and an additional 5 µg/ml
Puromycin (Sigma) was added to the medium for PAM cells. The cells were cultured at 37ºC
with a constant supply of 5% CO2.
Transfections with pcDNA3.1D-Dippu-AstR or empty pcDNA3.1D vector were carried out in
T75 flasks at 60 to 80% confluency. Transfection medium for CHO cells was prepared using the
Lipofectamine LTX kit (Invitrogen) with 3.75ml Opti-MEM, 7.5 µg vector construct and 18.75
µl Plus™ Reagent in a 5 ml polystyrene round-bottom tube. After 5 min incubation at room
temperature, 45 µl LTX (Invitrogen) was added. Transfection medium was added dropwise to
the cells after 30 min incubation at room temperature. The transfection medium of the HEK293

111
cells was similar to that of the CHO cells, except that 9 µg DNA construct (6 µg of vector
construct and 3 µg of reporter gene plasmid) was added to the medium. The luciferase ORF,
downstream of a cAMP responsive element (CRE) served as the reporter gene. After transfection,
cells were incubated overnight and an additional 10 ml of culture medium was added. The cells
were allowed to grow for another night prior to intracellular Ca2+ or cAMP measurements.
Aequorin assay - CHO cells were detached using 1× Phosphate Buffered Saline (PBS)
containing 0.2% EDTA (pH 8.0), collected and pelleted by centrifugation in DMEM/F12
medium. The number of viable and nonviable cells was determined using a NucleoCounter NC-
100™ (Chemometic). The cells were then resuspended to a density of 5×106 cells/ml in sterile
filtered DMEM/bovine serum albumin (BSA) medium (DMEM/F12 with L-glutamine and 15
mM HEPES, without Phenol red, supplemented with 0.1% BSA). A concentration of 5 µM
coelenterazine h (Invitrogen) was added and the cells were incubated for 4 h in the dark at room
temperature with gentle shaking to reconstitute the holo-enzyme aequorin. The cells were then
diluted 10-fold in BSA medium and incubated for another 30 min in the dark with gentle shaking.
Peptides were dissolved in BSA medium and dispensed in 50 µl aliquots into the wells of a white
96-well plate. Fifty µl of the cell suspension was injected into each well and light emission was
recorded using a Mithras LB 940 multimode microplate reader (Berthold Technologies) over 30
sec. The cells were lysed by injection of 50 µl 0.3% Triton X-100 and light emission was
monitored for an additional 8 sec. The total response (ligand + Triton-X100) is the representative
for the quantity of viable cells present in the well. BSA medium was used as a negative control
in each row of the plate, and 1 µM ATP served as a positive control. The negative response was
subtracted from the luminescence measured in wells of the same row. Calculations were made

112
using the output file from Microwin software (Berthold Technologies) in Excel (Microsoft).
Further analysis was done in Excel and GraphPad Prism 6.
cAMP reporter assay - HEK cells were detached using 1× PBS containing 0.2% EDTA (pH 8.0),
collected and pelleted by centrifugation in DMEM/F12 medium. The number of viable and
nonviable cells was determined using a NucleoCounter NC-100™ (Chemometic). The cells were
then resuspended to a concentration of 1×106 cell/ml in DMEM/F12 containing 200 µM 3-
isobutyl-1-methylxanthine (IBMX, Sigma) to prevent cAMP breakdown. Peptides were
dissolved in IBMX medium in the presence or absence of 20 µM forskolin, and were dispensed
in 50 µl aliquots into the wells of a white 96-well plate. Fifty µl of cell suspension was added
into each well and the plate was incubated in a CO2 incubator (5% CO2) at 37ºC for 3.5 h. Fifty
µl of Steadylite Plus substrate (PerkinElmer) was then added to each well and the plate was
incubated in the dark for 15 min while gently shaking. Light emission resulting from the
luciferase enzymatic activity was recorded for 5 s/well using a Mithras LB940. Results were
analyzed using the output of MicroWin and further processed by Excel and GraphPad Prism 6.
Radiochemical assay (RCA) - The in vitro RCA for JH biosynthesis was performed as described
by Feyereisen and Tobe (Feyereisen and Tobe, 1981) and modified by Tobe and Clarke (Tobe
and Clarke, 1985). Two incubations were conducted in the JH biosynthesis rescue experiments.
CA were incubated in TC199 medium with 10-6 M AST7 for 3h, and then transferred to fresh
medium with 10-6 M AST7 and JH precursors for a second 3h incubation. JH biosynthesis was
determined after each incubation. The rate of JH biosynthesis during the first incubation was
used as the control value.
RNA interference (RNAi) - Dippu-AstR and Dippu-AST dsRNA constructs were prepared using
the MEGAscript® RNAi Kit (Ambion). Primers used are given in Table S3.2. The Fragments

113
were amplified by PCR using forward and reverse primers with T7 promoters
(TAATACGACTCACTATAGGGAGA) attached to the 5’ end. Fragments were then subcloned
and sequenced to verify the presence of the T7 promoter. The amplified fragment was used in an
RNA transcription reaction and incubated overnight to yield annealed dsRNA transcripts. A
nuclease digestion was subsequently performed to remove ssRNA and DNA remaining in the
product. The dsRNA was further purified according to the manufacturer’s instructions (Ambion).
Concentration of the dsRNA construct was determined using a nanodrop instrument (Thermo
Fisher Scientific Inc.). Five-fold diluted dsRNA was run on a 1.2% agarose gel to examine the
quality and integrity of the construct. The control dsRNA was prepared using a PCR fragment
amplified from a non-coding region of pJET1.2 cloning vector (Thermo Scientific).
Dippu-AstR dsRNA was diluted in cockroach saline to a concentration of 250ng/µl. Each adult
female was injected with 4 µl of dsRNA solution on day 0, 2 and 4 after the final moult. CA
were dissected on day 6 and stored in liquid nitrogen prior to RNA extraction. A second set of
cockroaches was injected with Dippu-AstR dsRNA in the same scheme as described above. CA
were dissected on day 6 and cleaned in TC199 medium (GIBCO; 1.3 mM Ca2+, 2% Ficoll,
methionine-free) for their use in the radiochemical assay (RCA) determining the effect of AST
on JH biosynthesis. Basal oocyte length was measured during dissection.
Dippu-AST dsRNA was diluted in cockroach saline to a concentration of 250ng/µl. Each adult
female was injected with 4 µl of dsRNA solution on day 0, 2, 4 and 5 after the final moult. Brain
were dissected on day 6 and stored in liquid nitrogen prior to RNA extraction. CA were dissected
on day 6 and cleaned in TC199 medium (GIBCO; 1.3 mM Ca2+, 2% Ficoll, methionine-free) for
their use in the radiochemical assay (RCA) determining the effect of AST on JH biosynthesis.
Basal oocyte length was measured during dissection.

114
Data analysis – Data set were analyzed by T-test to determine the significant difference. All
results were expressed as mean ± SEM, and considered significantly different at P ≤ 0.05.
Results
Tissue distribution of Dippu-AST and Dippu-AstR
The tissue distribution of Dippu-AST and Dippu-AstR was determined in adult male and female
cockroaches using q-RT-PCR (Fig. 3.1). Nine tissues were used to examine the tissue specificity
of Dippu-AST and Dippu-AstR: brain (Br), nerve cord (NC), corpora allata (CA), fat body (Fb),
ovary (Ov), midgut (MG) and Malpighian tubules (MT) from females and accessory gland (AG)
and testes (Te) from males. The Dippu-AST gene is expressed in brain, nerve cord and midgut,
which is consistent with the pleiotropic functions of ASTs (Fig. 3.1). Dippu-AstR shows the
highest transcription levels in the CA, followed by nerve cord, brain and fat body. All the other
tissues tested showed either negligible or undetectable levels of Dippu-AstR mRNA.
Developmental expression during the first gonadotrophic cycle of females
We determined the developmental profile of Dippu-AstR in the CA, brain and fat body of mated
females during their first gonadotrophic cycle (Fig. 3.2). In CA, the Dippu-AstR mRNA declined
on days 2-4 and then increased significantly on day 6. The transcript level of Dippu-AstR in the
CA was inversely correlated with the rate of JH biosynthesis (Fig. 3.2A). In brain, on the other
hand, transcript levels of Dippu-AstR and Dippu-AST showed only minor variations (Fig. 3.2B).
The transcript level of Dippu-AstR in fat body was relatively low, with the highest level of
mRNA expression on day 4, and negligible on all other days (data not shown).
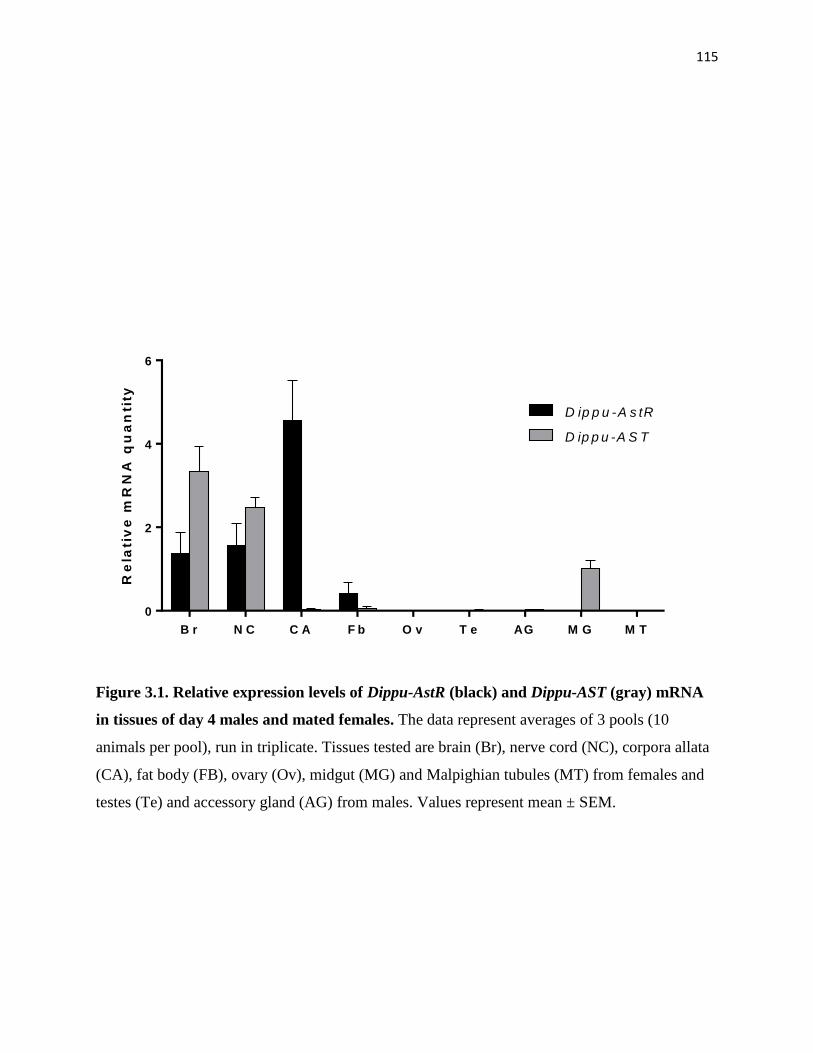
115
B r N C C A F b O v T e AG M G M T
0
2
4
6
Re
lati
ve
mR
NA
qu
an
tity
D ip p u -A s tR
D ip p u -A S T
Figure 3.1. Relative expression levels of Dippu-AstR (black) and Dippu-AST (gray) mRNA
in tissues of day 4 males and mated females. The data represent averages of 3 pools (10
animals per pool), run in triplicate. Tissues tested are brain (Br), nerve cord (NC), corpora allata
(CA), fat body (FB), ovary (Ov), midgut (MG) and Malpighian tubules (MT) from females and
testes (Te) and accessory gland (AG) from males. Values represent mean ± SEM.

116
D 0 D 1 D 2 D 3 D 4 D 5 D 6 D 7
0
5
1 0
1 5
2 0
0
5
1 0
1 5
2 0
2 5
Re
lati
ve
mR
NA
qu
an
tity
of
As
tR JH
bio
sy
nth
es
is(p
mo
l/h/C
A)
D ip p u -A s tR
J H b io s y n th e s is
D 0 D 1 D 2 D 3 D 4 D 5 D 6 D 7
0 .0
0 .5
1 .0
1 .5
2 .0
0
1
2
3
4
Re
lati
ve
mR
NA
qu
an
tity
of
As
tRR
ela
tive
mR
NA
qu
an
tity o
f AS
T
D ip p u -A s tR
D ip p u -A S T
A
B
Figure 3.2. Relative expression levels of Dippu-AstR and Dippu-AST mRNA during the first
gonadotrophic cycle. (A) Dippu-AstR mRNA expression and JH biosynthesis in CA. Dippu-
AstR mRNA levels were quantified by q-RT-PCR and normalized against levels of Armadillo
and EF1a mRNA (Marchal et al., 2013b). The data represent the average of 3 biologically
independent pools (10 animals per pool), run in triplicate. Rate of JH biosynthesis was measured
using the RCA, n≥10. (B) Dippu-AstR and Dippu-AST mRNA expression in brain. Dippu-AstR
and Dippu-AST mRNA levels were normalized against levels of Tubulin and EF1a mRNA
(Marchal et al., 2013b). Values represent mean ± SEM.

117
Effect of Dippu-AST dsRNA on JH biosynthesis
To determine the controlling role of AST in JH biosynthesis, we knocked down Dippu-AST
using RNAi. Injecting Dippu-AST dsRNA results in a 91% decrease of Dippu-AST mRNA in the
brain. However, JH biosynthesis of CA did not show any significant change in the dsRNA
treated animals (Fig. 3.3).
Functional activation of Dippu-AstR with ASTs
We initially expressed Dippu-AstR in CHO-WTA11 cells. This cell line expresses the Ca2+
reporter apoaequorin and Gα16, a promiscuous G protein that couples to most GPCRs with
subsequent mobilization of intracellular Ca2+ (Stables et al., 1997). As shown in Fig. 3.4, all 13
tested ASTs induced clear dose-dependent bioluminescence responses in AstR expressing cells.
Most of the peptides showed similar degrees of biological efficacy (ability to activate AstR) but
differed considerably in potency, with EC50 values ranging from 0.2 nM for AST6 to 30 nM, in
the case of AST13 (Table 3.1). In general, the abilities of ASTs to activate AstR corresponded to
their potencies as inhibitors of JH production by the CA, with the exception of AST1, AST5 and
AST6 (Tobe et al., 2000) (Table 3.1).
To determine the second messenger pathways involved in Dippu-AstR activation, we first
expressed Dippu-AstR in CHO-PAM28 cells (lacking the promiscuous Gα16). The expression of
apoaequorin in the cell allows testing whether the receptor can couple naturally through
intracellular Ca2+. Two ASTs (AST5 and 6) were chosen because of their high potency in
inhibiting JH biosynthesis or in activating Dippu-AstR. As shown in Fig. 3.5A, AST5 and 6
induced dose-dependent intracellular Ca2+ responses in AstR-transfected CHO-PAM28 cells,
with EC50 values of 21.4 nM and 1.1 nM, respectively.

118
We subsequently tested whether the receptor coupled with cAMP in the signal transduction
pathway. We expressed Dippu-AstR in HEK293 cells, which contain the luciferase gene under
the control of a cAMP response element. We assayed different concentrations of AST5 and
AST6 in the presence or absence of forskolin. Forskolin activates adenylate cyclase (AC), which
increases intracellular levels of cAMP. If AstR would couple negatively to AC, we would expect
lower levels of cAMP following application of ASTs. This was not the case. Application of
AST5 or AST6 to transfected HEK cells did not cause any significant change in cAMP level.
However, in the absence of forskolin, AST5 and AST6 treatments resulted in a dose-dependent
increase in cAMP concentration, as measured by assay of luciferase activity (Fig. 3.5B). The
EC50 values were 287.8 nM for AST5 and 24.3 nM for AST6. The EC50 values for the cAMP-
based luciferase assay in HEK293 cells were higher than for the Ca2+ responses detected in either
of the two CHO cell lines, but the relative order of ligand potency was maintained. CHO-
WTA11, CHO-PAM28 and HEK293 cells transfected with pcDNA3.1D (empty vector) did not
show any response to ASTs.
AST does not affect the transcript level of enzymes in the JH biosynthetic pathway
The injection of AstR dsRNA on day 0, 2 and 4 resulted in a 67% knockdown of AstR mRNA
levels in CA of day 6 females (Fig. 3.6A). The JH biosynthetic activity of CA was measured
using the RCA. In AstR knockdown animals, JH production increased by 60% and the response
of CA to AST decreased by 28% (Fig. 3.6B and 3.6C).
Eleven of 13 genes encoding enzymes in the JH biosynthetic pathway have been identified
(Chapter 2). To determine the target of AST action in the JH biosynthetic pathway, we measured
the mRNA levels of the 11 genes encoding enzymes catalyzing different steps in the JH
biosynthetic pathway. Although AstR dsRNA treatment resulted in an increase in the rate of JH

119
biosynthesis, none of the relative transcript levels showed significant changes (Fig. 3.6D). The
high variation of the transcript level of JHAMT on day 6 can be explained by previous results
that show a great degree of fluctuation in JHAMT mRNA levels at times when rates of JH
biosynthesis are highly dynamic (see Chapter 2).
To confirm our results, we incubated CA in medium containing 10-7 M AST7 for 3 hours, which
results in a 60 to 70% decrease in JH biosynthesis (Tobe et al., 2000), and determined the
transcript level of the enzymes in the CA. As in the AstR RNAi experiments, the mRNA levels of
the 11 genes tested show no significant changes (Fig. 3.7).
JH precursors reverse AST-induced inhibition of JH biosynthesis
Earlier studies showed that mevalonate and farnesol were able to partially reverse AST-induced
inhibition of JH biosynthesis (Pratt et al., 1991; Pratt et al., 1989). To determine which enzyme(s)
is/are affected by ASTs in the JH biosynthetic pathway, we treated CA with select JH precursors
in the presence of 10-6 M AST7. If the activity of enzymes was inhibited by AST, the JH
precursors prior to the enzyme will not be able to reverse the inhibition of AST. As shown in Fig.
3.8, all exogenous JH precursors acetyl-CoA, mevalonic acid, diphosphomevalonate and farnesol
significantly stimulated the rate of JH biosynthesis in the presence of AST7.
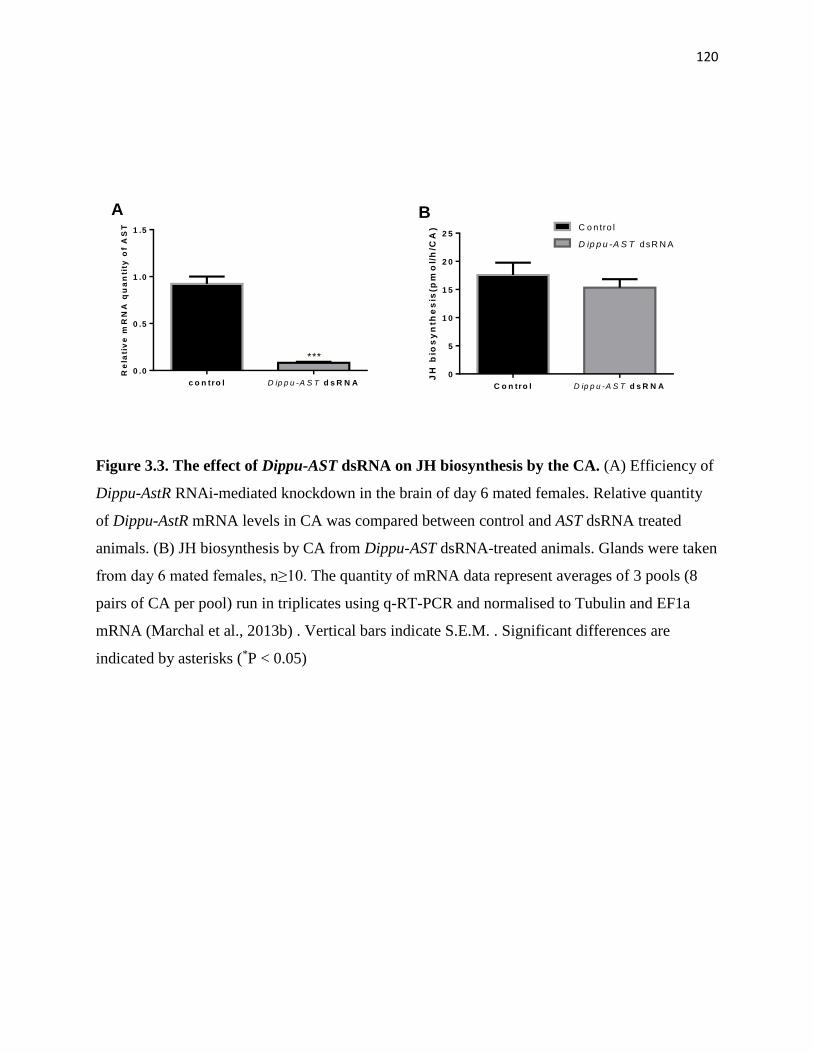
120
c o n tro l D ip p u -A S T d s R N A
0 .0
0 .5
1 .0
1 .5
Re
lati
ve
mR
NA
qu
an
tity
of
AS
T
***
C o n tro l D ip p u -A S T d s R N A
0
5
1 0
1 5
2 0
2 5
JH
bio
sy
nth
es
is(p
mo
l/h
/CA
) C o n tro l
D ip p u -A S T dsR N A
A B
Figure 3.3. The effect of Dippu-AST dsRNA on JH biosynthesis by the CA. (A) Efficiency of
Dippu-AstR RNAi-mediated knockdown in the brain of day 6 mated females. Relative quantity
of Dippu-AstR mRNA levels in CA was compared between control and AST dsRNA treated
animals. (B) JH biosynthesis by CA from Dippu-AST dsRNA-treated animals. Glands were taken
from day 6 mated females, n≥10. The quantity of mRNA data represent averages of 3 pools (8
pairs of CA per pool) run in triplicates using q-RT-PCR and normalised to Tubulin and EF1a
mRNA (Marchal et al., 2013b) . Vertical bars indicate S.E.M. . Significant differences are
indicated by asterisks (*P < 0.05)
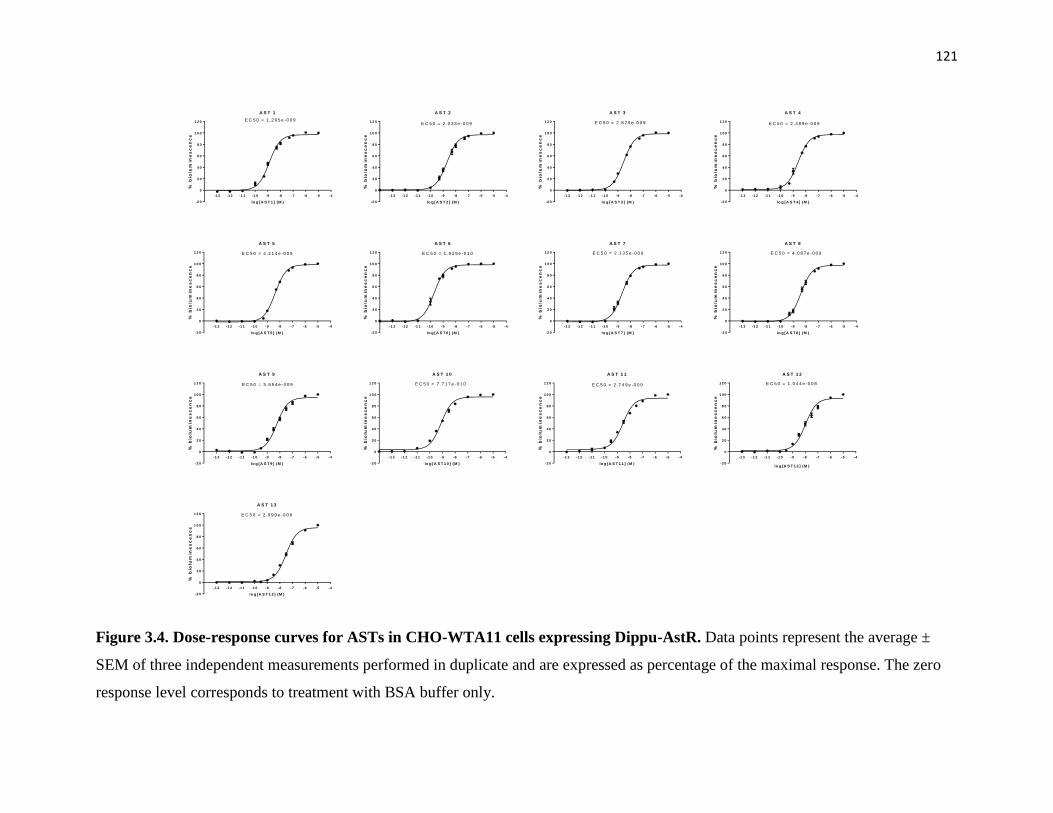
121
-1 3 -1 2 -1 1 -1 0 -9 -8 -7 -6 -5 -4
-2 0
0
2 0
4 0
6 0
8 0
1 0 0
1 2 0
A S T 1
lo g [A S T 1 ] (M )
% b
iolu
min
es
ce
nc
e
E C 5 0 = 1 .2 9 5 e -0 0 9
-1 3 -1 2 -1 1 -1 0 -9 -8 -7 -6 -5 -4
-2 0
0
2 0
4 0
6 0
8 0
1 0 0
1 2 0
A S T 2
lo g [A S T 2 ] (M )
% b
iolu
min
es
ce
nc
e
E C 5 0 = 2 .0 3 3 e -0 0 9
-1 3 -1 2 -1 1 -1 0 -9 -8 -7 -6 -5 -4
-2 0
0
2 0
4 0
6 0
8 0
1 0 0
1 2 0
A S T 3
lo g [A S T 3 ] (M )
% b
iolu
min
es
ce
nc
e
E C 5 0 = 2 .8 2 9 e -0 0 9
-1 3 -1 2 -1 1 -1 0 -9 -8 -7 -6 -5 -4
-2 0
0
2 0
4 0
6 0
8 0
1 0 0
1 2 0
A S T 4
lo g [A S T 4 ] (M )
% b
iolu
min
es
ce
nc
e
E C 5 0 = 2 .4 8 9 e -0 0 9
-1 3 -1 2 -1 1 -1 0 -9 -8 -7 -6 -5 -4
-2 0
0
2 0
4 0
6 0
8 0
1 0 0
1 2 0
A S T 5
lo g [A S T 5 ] (M )
% b
iolu
min
es
ce
nc
e
E C 5 0 = 4 .2 1 4 e -0 0 9
-1 3 -1 2 -1 1 -1 0 -9 -8 -7 -6 -5 -4
-2 0
0
2 0
4 0
6 0
8 0
1 0 0
1 2 0
A S T 6
lo g [A S T 6 ] (M )
% b
iolu
min
es
ce
nc
e
E C 5 0 = 1 .9 2 9 e -0 1 0
-1 3 -1 2 -1 1 -1 0 -9 -8 -7 -6 -5 -4
-2 0
0
2 0
4 0
6 0
8 0
1 0 0
1 2 0
A S T 7
lo g [A S T 7 ] (M )
% b
iolu
min
es
ce
nc
e
E C 5 0 = 2 .1 3 5 e -0 0 9
-1 3 -1 2 -1 1 -1 0 -9 -8 -7 -6 -5 -4
-2 0
0
2 0
4 0
6 0
8 0
1 0 0
1 2 0
A S T 8
lo g [A S T 8 ] (M )
% b
iolu
min
es
ce
nc
e
E C 5 0 = 4 .0 8 7 e -0 0 9
-1 3 -1 2 -1 1 -1 0 -9 -8 -7 -6 -5 -4
-2 0
0
2 0
4 0
6 0
8 0
1 0 0
1 2 0
A S T 9
lo g [A S T 9 ] (M )
% b
iolu
min
es
ce
nc
e
E C 5 0 = 5 .6 5 4 e -0 0 9
-1 3 -1 2 -1 1 -1 0 -9 -8 -7 -6 -5 -4
-2 0
0
2 0
4 0
6 0
8 0
1 0 0
1 2 0
A S T 1 0
lo g [A S T 1 0 ] (M )
% b
iolu
min
es
ce
nc
e
E C 5 0 = 7 .7 1 7 e -0 1 0
-1 3 -1 2 -1 1 -1 0 -9 -8 -7 -6 -5 -4
-2 0
0
2 0
4 0
6 0
8 0
1 0 0
1 2 0
A S T 1 1
lo g [A S T 1 1 ] (M )
% b
iolu
min
es
ce
nc
e
E C 5 0 = 2 .7 4 9 e -0 0 9
-1 3 -1 2 -1 1 -1 0 -9 -8 -7 -6 -5 -4
-2 0
0
2 0
4 0
6 0
8 0
1 0 0
1 2 0
A S T 1 2
% b
iolu
min
es
ce
nc
e
E C 5 0 = 1 .0 4 4 e -0 0 8
lo g [A S T 1 2 ] (M )
-1 3 -1 2 -1 1 -1 0 -9 -8 -7 -6 -5 -4
-2 0
0
2 0
4 0
6 0
8 0
1 0 0
1 2 0
A S T 1 3
lo g [A S T 1 3 ] (M )
% b
iolu
min
es
ce
nc
e
E C 5 0 = 2 .9 9 9 e -0 0 8
Figure 3.4. Dose-response curves for ASTs in CHO-WTA11 cells expressing Dippu-AstR. Data points represent the average ±
SEM of three independent measurements performed in duplicate and are expressed as percentage of the maximal response. The zero
response level corresponds to treatment with BSA buffer only.

122
-1 4 -1 3 -1 2 -1 1 -1 0 -9 -8 -7 -6 -5 -4
-2 0
0
2 0
4 0
6 0
8 0
1 0 0
1 2 0
D ip p u -A s tR in C H O -P A M 2 8
C o n c e n t r a t io n lo g [ M ]
% b
iolu
min
es
ce
nc
e A S T 5
A S T 6
-1 2 -1 1 -1 0 -9 -8 -7 -6 -5 -4
-2 0
0
2 0
4 0
6 0
8 0
1 0 0
1 2 0
D ip p u -A s tR in H E K 2 9 3
C o n c e n t r a t io n lo g [ M ]%
bio
lum
ine
sc
en
ce A S T 5
A S T 6
A B
Figure 3.5. Dose-response curves for the bioluminescence response induced in (A) CHO-
PAM28 and (B) HEK293 cells expressing Dippu-AstR. Data points represent the average ±
SEM of three independent measurements performed in duplicate and are expressed as a
percentage of the maximal response. The zero response level corresponds to treatment with BSA
buffer only. Peptides tested are AST5 (squares) and AST6 (triangles).

123
c o n tro l A s tR d s R N A
0 .0
0 .5
1 .0
1 .5
2 .0
Re
lati
ve
mR
NA
qu
an
tity
of
As
tR
*
c o n tro l A s tR d s R N A
0
2 0
4 0
6 0
8 0
% J
H b
ios
yn
the
sis
in
hib
itio
n
*
c o n tro l A s tR d s R N A
0
5
1 0
1 5
JH
bio
sy
nth
es
is(p
mo
l/h
/CA
)
*
T h io l H M G S H M G R M K P M K P P -M e v D IP P I F P P S F O L D J H A M T C Y P 1 5
0
1
2
3
4
5
Re
lati
ve
mR
NA
qu
an
tity
c o n tro l
A s tR d s R N A
A B C
D
Figure 3.6. The effect of Dippu-AstR dsRNA on JH biosynthesis by the CA and on the
expression of genes encoding enzymes in the JH biosynthetic pathway of D. punctata. (A)
Efficiency of Dippu-AstR RNAi-mediated knockdown in mated females. Relative quantity of
Dippu-AstR mRNA levels in CA was compared between control and AstR dsRNA treated
animals. (B) JH biosynthesis by CA from Dippu-AstR dsRNA-treated animals. Glands were
taken from day 6 mated females, n≥10. (C) Effect of 10-7 M Dippu-AST7 on JH biosynthesis by
CA from control and AstR knockdown animals. CA were incubated first in normal medium for
3h and subsequently incubated in medium with 10-7 M AST7 for another 3h. The percentage of
inhibition was calculated using the rate of JH biosynthesis during the first and second incubation.
(D) The effect of silencing Dippu-AstR on the transcript levels of genes encoding enzymes in the
JH biosynthetic pathway. Enzyme abbreviations are as described in Figure S1. The mRNA
quantity data represent averages of 3 pools (8 pairs of CA per pool) run in triplicates using q-RT-
PCR. Vertical bars indicate S.E.M.. Significant differences are indicated by asterisks (*P < 0.05).

124
AC AT H M G S H M G R M K P M K P P M e v D IP P I F P P S F AL D J H AM T E P O X
0
5
1 0
1 5
2 0R
ela
tiv
e m
RN
A q
ua
nti
ty c o n tro l
tre a tm e n t
Figure 3.7. The effect of AST on the expression levels of genes encoding enzymes in the JH
biosynthetic pathway in CA of day 6 mated female D. punctata. CA were incubated for 3h in
normal (control) medium or medium supplemented with 10-7 M AST7 (treatment) and then
washed with cockroach saline prior to the RNA extraction. RNA was extracted from CA in
control and treatment groups to determine the transcript level of genes encoding enzymes in JH
biosynthetic pathway. Enzyme abbreviations are as described in Figure S1. The data represent
averages of 3 pools (8 pairs of CA per pool) run in triplicate using q-RT-PCR. Vertical bars
indicate S.E.M.
Ac e tyl C o A M A M AP T F a r n e s o l
0
5
1 0
1 5
JH
re
lea
se
(p
mo
l/C
A/h
)
c o n tro l
J H p re c u rs o r
***
******
***
Figure 3.8. JH precursors rescue the AST-induced JH inhibition. JH biosynthesis was
evaluated in CA that were first incubated in medium TC199 with 10-6 M AST7 (control), and
then in medium with 10-6 M AST7 together with the individual precursors: Acetyl-CoA (100
µM), MA (100 µM), DPPM (100 µM) or farnesol (40 µM). CA were dissected from day 7 mated
females. Each data point represents mean ± SEM (n ≥ 10) Significant differences are indicated
by asterisks (***P < 0.001).

125
Table 3.1. Potency of Dippu-ASTs a: activation of AstR in CHO-WTA11 cells (EC50) or
inhibitory effect on JH release (IC50)
Allatostatin EC50
1
/nM
Rank1
order
(EC50)
IC502
/nM
Rank2
order
(IC50)
Sequence
AST1 1.295 3 9.566 11 Leu-Tyr-Asp-Phe-Gly-Leu-NH2
AST2 2.033 4 0.4254 3 Ala-Tyr-Ser-Tyr-Val-Ser-Glu-Tyr-Lys-Arg-Leu-Pro-Val- Tyr-Asn-Phe-Gly-Leu-NH2
AST3 2.829 8 12.54 12 Ser-Lys-Met-Tyr-Gly-Phe-Gly-Leu-NH2
AST4 2.489 6 1.139 5 Asp-Gly-Arg-Met-Tyr-Ser-Phe-Gly-Leu-NH2
AST5 4.214 10 0.1063 1 Asp-Arg-Leu-Tyr-Ser-Phe-Gly-Leu-NH2
AST6 0.1929 1 2.725 8 Ala-Arg-Pro-Tyr-Ser-Phe-Gly-Leu-NH2
AST7 2.135 5 0.4118 2 Ala-Pro-Ser-Gly-Ala-Gln-Arg-Leu-Tyr-Gly-Phe-Gly-Leu-NH2
AST8 4.087 9 3.275 10 Gly-Gly-Ser-Leu-Tyr-Ser-Phe-Gly-Leu-NH2
AST9 5.654 11 2 6 Gly-Asp-Gly-Arg-Leu-Tyr-Ala-Phe-Gly-Leu-NH2
AST10 0.7717 2 0.9225 4 Pro-Val-Asn-Ser-Gly-Arg-Ser-Ser-Gly-Ser-Arg-Phe-Asn-Phe-Gly-Leu-NH2
AST11 2.749 7 2.795 9 Tyr-Pro-Gln-Glu-His-Arg-Phe-Ser-Phe-Gly-Leu-NH2
AST12 10.44 12 2.011 7 Pro-Phe-Asn-Phe-Gly-Leu-NH2
AST13 29.99 13 12.84 13 Ile-Pro-Met-Tyr-Asp-Phe-Gly-Ile-NH2
a Potency is defined as the dose required to achieve a given level of activation of AstR or
inhibition of JH biosynthesis, and are listed in rank order. 1 EC50 was determined by AstR
activation assay in CHO-WTA11 cells (details seen Fig. 1). 2 IC50 values are as reported by
(Tobe et al., 2000) (CA from day 2 virgin female).

126
Discussion
Our observations on the expression of the Dippu-AstR and Dippu-AST provide insights into the
tissue-specific interaction between the ligands and the receptor. In brain and nerve cord, both
Dippu-AstR and Dippu-AST are expressed, whereas in CA, only the receptor is expressed. This is
consistent with observations that AST is delivered to the CA by nerves from neurosecretory cells
in the brain (Stay et al., 1992). In the midgut, however, Dippu-AST is expressed, but its receptor
is not (Fig. 3.1); tissue distribution of Dippu-AstR and Dippu-AST in day 7 animals provided
similar results (Fig. S3.1). Previous studies suggest that ASTs can induce myotropic activity in
gut tissues (Duve et al., 1995; Lange et al., 1995). Furthermore, putative receptors for ASTs in
midgut were partially characterized using a radioligand-binding assay (Bowser and Tobe, 2000).
Therefore, the undetectable transcript level of Dippu-AstR in midgut suggests that one or more
additional AstR in D. punctata may exist, just as two receptors for allatostatins have been
identified in Drosophila melanogaster (Birgul et al., 1999; Lenz et al., 2000).
The sensitivity of CA to ASTs differs between animals of different ages (Pratt et al., 1990; Stay
et al., 1991). CA appear more sensitive to AST treatment when JH biosynthetic activity is low
than at stages of high JH production. In the present study, we have shown that the transcript
levels of Dippu-AstR in the CA correlate with changes in the sensitivity of the CA to ASTs (Fig.
3.2A). It is therefore reasonable to suggest that the changing transcript levels of Dippu-AstR are
partly responsible for the changes in JH biosynthesis during the first gonadotropic cycle. The
ability of ASTs to inhibit JH biosynthesis depends not only on the sensitivity of the CA, but also
on the concentration of the ASTs (Stay et al., 1996). q-RT-PCR of the AST precursor gene
showed that there was no significant change in the transcript level of ASTs (Fig. 3.2B). Lloyd et
al (2000) determined the quantity of AST in CA and brain by ELISA using antibody against

127
Dippu-AST7, and found no significant change in the content of AST7 during the first
gonadotropic cycle. Meanwhile, silencing the AST precursor gene did not cause any significant
increase of JH production (Fig. 3.3). Same result was observed in the German cockroach, B.
germanica (Maestro and Belles, 2006). These results suggest that the AST-inhibition of JH
biosynthesis by AST is controlled by the change of expression of the AST receptor.
To functionally analyze AstR, we expressed the Dippu-AstR in mammalian cells and activated it
with Dippu-AST. We first expressed Dippu-AstR in CHO cells, demonstrating that it is indeed a
Diploptera allatostatin receptor. All thirteen ASTs are very potent in activating AstR expressed
in CHO-WTA11 cells with EC50 values in the low nanomolar range (Fig. 3.4). Despite the
structural similarities, there were some differences in the response to individual peptides. AST13,
which contains isoleucine at the C-terminus, exhibited the highest EC50. This result reinforces
the importance of the C-terminal pentapeptide motif Y/FXFGL-NH2. Moreover, the amino acids
outside of the C-terminal pentapeptide motif appear to determine the affinity of AST for AstR as
well. AST5 and AST6 have the same pentapeptide motif (Table 3.1), whereas their ability to
activate the receptor is significantly different. The change of Pro to Leu, and Ala to Asp
decreases AST affinity of AstR by 20-fold.
A P. americana allatostatin receptor was expressed in Xenopus oocytes, and the potency of
different Periplaneta ASTs in activating Peram-AstR was determined (Gade et al., 2008). Five
Dippu-ASTs (AST 1, 2, 3, 6 and 13) have the same structure as in P. americana (AST 1, 2, 3, 6
and 14). And the rank order of potency of these ASTs in activating Dippu-AstR expressed in
CHO-WTA11 cells is similar to that of P. americana receptor (Gade et al., 2008). However, the
potencies of AST 1, 5 and 6 in activating AstR differ from their capacities to inhibit JH
production in vitro (Table 1). AST 5, which showed the highest inhibitory activity, demonstrated

128
only a moderate potency in activating the expressed Dippu-AstR, ranking 10 out of 13. In
contrast, AST1, which was one of the weakest inhibitors of JH biosynthesis in vitro (ranking 11
out of 13), appears to be one of the most potent peptides in receptor activation. AST6, the
structure of which is conserved in several insect orders, showed the highest potency in activating
expressed Dippu-AstR (Bendena et al., 1999). Its ability to inhibit JH biosynthesis, on the other
hand, was moderate. Different reasons could contribute to the differences between potency in
activating the receptor and inhibiting JH biosynthesis in vitro. The susceptibility of ASTs to
degradation by enzymes in the CA differs between ASTs, which could result in different
potencies in inhibiting JH biosynthesis. AST5, which showed high resistance to the degradation
by hemolymph and membrane preparations, presented the highest activity in inhibiting JH
biosynthesis (Garside et al., 1997a, b). Furthermore, tests with cloned receptors in mammalian
cells might not fully reflect the actual situation in insects. The posttranslational modifications of
AstR or the interacting proteins present in receptor-expressing cells could affect the
conformational and functional properties of this receptor. Nevertheless, we here chose
mammalian cell lines because there may be fewer interacting proteins in this heterologous
system than in insect cell lines.
The inhibition of JH biosynthesis by Dippu-ASTs is a complex process, probably involving more
than one second messenger. Previous studies showed that ASTs regulate JH biosynthesis by
acting through the inositol trisphosphate (IP3)/diglyceride (DAG) second messenger systems, in
which protein kinase C (PKC) and Ca2+ are involved (Feyereisen and Farnsworth, 1987b;
Rachinsky and Tobe, 1996; Rachinsky et al., 1994). However, the role of Ca2+ in regulation of
JH biosynthesis is confounding. JH production shows dose-dependency on extracellular Ca2+ in
the medium, whereas the Ca2+ ionophore A23187 caused a rapid decline in JH release

129
(Kikukawa et al., 1987). Inhibition of JH biosynthesis by brain extracts was antagonized by
inorganic Ca2+channel blockers (Feyereisen and Farnsworth, 1987a). However, elevation of
intracellular Ca2+ levels by treating CA with the Ca2+-mobilizing drug thapsigargin diminished
the inhibitory effect of ASTs (Rachinsky et al., 1994). The activation of Dippu-AstR induced a
dose-dependent intracellular Ca2+ response in CHO-PAM28 cells (Fig. 3.5A), which supports the
involvement of Ca2+ as a second messenger of AST. Aucoin et al (1987) suggested that there
may be both a Ca2+-dependent and a Ca2+-independent pathway (cAMP) involved in the
inhibition of JH biosynthesis. Incubating CA with brain extracts resulted in an increase in the
level of cAMP and a decrease in JH biosynthesis. Forskolin, which causes a dose-dependent
accumulation of cAMP, led to a rapid and dose-dependent inhibition of JH biosynthesis.
Moreover, the sensitivity of CA to forskolin shows the same pattern as its sensitivity to AST
during the first gonadotrophic cycle (Feyereisen and Farnsworth, 1987a; Meller et al., 1985). In
contrast, another study suggest that levels of cAMP do not increase following treatment of CA
with ASTs. Our findings demonstrated that the activation of Dippu-AstR induced a dose-
dependent cAMP response in HEK293 cells, which suggests that cAMP may be involved in the
signaling pathway of AST cells (Fig. 3.5B). Dippu-AstR appears to dually couple through the
Gαs and Gαq pathways and triggers the release of intracellular Ca2+ and the increase in cAMP
levels. Studies on Drosophila AstRs suggested that DAR-1 and DAR-2 couple to Gi/o mediated
signaling and other G-proteins (Birgul et al., 1999; Larsen et al., 2001). However, activation of
AstR in the presence of forskolin did not significantly influence the cAMP signal. This result
suggests that there is no involvement of Gαi in the Dippu-AstR pathway.
AST regulates JH biosynthesis by affecting either enzymes directly involved in the JH
biosynthetic pathway or steps prior to the production of acetyl-CoA. We have studied the effect

130
of AST on the transcript levels of genes in the JH biosynthetic pathway. The inhibitory effect of
AST on JH biosynthesis was rapid and reversible, which suggests that AST may not regulate JH
biosynthesis through transcript levels ((Pratt et al., 1991; Pratt et al., 1989). Our study confirmed
this earlier assumption by revealing that the transcript levels of genes in the JH biosynthetic
pathway were not affected by AstR silencing or AST treatment in vitro (Fig. 3.6 and 3.7).
Previous studies indicate that ASTs regulate JH biosynthesis through steps(s) prior to the
production of mevalonate (Pratt et al., 1991; Pratt et al., 1989). However, the enzyme activities
of HMG-CoA synthase or HMG-CoA reductase were not affected by AST (Sutherland and
Feyereisen, 1996). To determine which enzyme in the JH biosynthetic pathway was influenced
by AST, we used several JH precursors to reverse the AST-induced inhibition of JH biosynthesis.
All the JH precursors (including acetyl-CoA) were able to partially restore JH biosynthesis,
thereby suggesting that the enzyme activities were not affected by AST. The target of AST
action probably lies prior to the start of the JH biosynthetic pathway in regulating the production
of precursors of acetyl-CoA such as the transport of citrate from mitochondria to cytosol and/or
the cleavage of citrate to yield acetyl-CoA (Sutherland and Feyereisen, 1996). Further
investigation is needed to identify the exact target of AST.
In D. punctata, ASTs play an important role in the regulation of JH biosynthesis. The potency of
the 13 ASTs in activating AstR reveals the structure-activity relationship of AST action. The
activation of AstR in CHO and HEK cells suggests that both Ca2+ and cAMP can be involved in
the signal transduction of AST. Our results show that AST does not affect the transcript levels
and that the activities of enzymes in the JH biosynthetic pathway remain normal following
treatment with AST. AST probably affects JH biosynthesis prior to the entry of precursors into
the JH biosynthetic pathway. The exact target of AST action remains an open question.
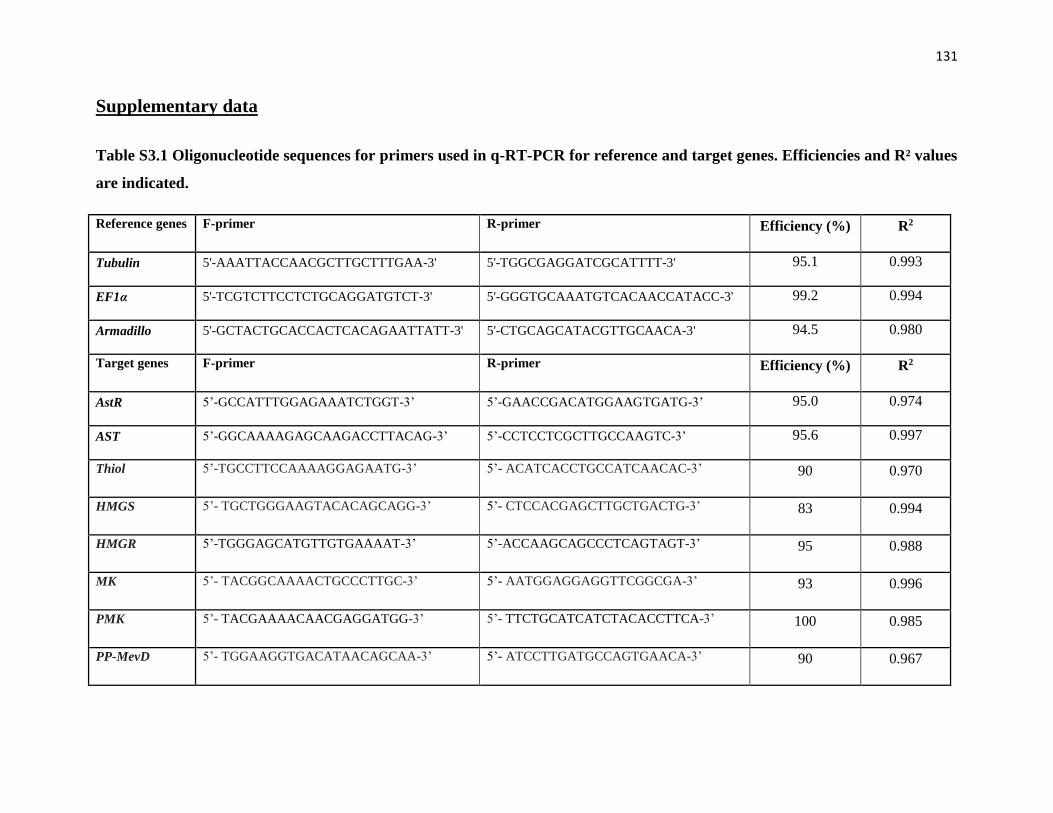
131
Supplementary data
Table S3.1 Oligonucleotide sequences for primers used in q-RT-PCR for reference and target genes. Efficiencies and R² values
are indicated.
Reference genes F-primer R-primer Efficiency (%) R2
Tubulin 5'-AAATTACCAACGCTTGCTTTGAA-3' 5'-TGGCGAGGATCGCATTTT-3' 95.1 0.993
EF1α 5'-TCGTCTTCCTCTGCAGGATGTCT-3' 5'-GGGTGCAAATGTCACAACCATACC-3' 99.2 0.994
Armadillo 5'-GCTACTGCACCACTCACAGAATTATT-3' 5'-CTGCAGCATACGTTGCAACA-3' 94.5 0.980
Target genes F-primer R-primer Efficiency (%) R2
AstR 5’-GCCATTTGGAGAAATCTGGT-3’ 5’-GAACCGACATGGAAGTGATG-3’ 95.0 0.974
AST 5’-GGCAAAAGAGCAAGACCTTACAG-3’ 5’-CCTCCTCGCTTGCCAAGTC-3’ 95.6 0.997
Thiol 5’-TGCCTTCCAAAAGGAGAATG-3’ 5’- ACATCACCTGCCATCAACAC-3’ 90 0.970
HMGS 5’- TGCTGGGAAGTACACAGCAGG-3’ 5’- CTCCACGAGCTTGCTGACTG-3’ 83 0.994
HMGR 5’-TGGGAGCATGTTGTGAAAAT-3’ 5’-ACCAAGCAGCCCTCAGTAGT-3’ 95 0.988
MK 5’- TACGGCAAAACTGCCCTTGC-3’ 5’- AATGGAGGAGGTTCGGCGA-3’ 93 0.996
PMK 5’- TACGAAAACAACGAGGATGG-3’ 5’- TTCTGCATCATCTACACCTTCA-3’ 100 0.985
PP-MevD 5’- TGGAAGGTGACATAACAGCAA-3’ 5’- ATCCTTGATGCCAGTGAACA-3’ 90 0.967

132
Table S3.1 (Continue) Oligonucleotide sequences for primers used in q-RT-PCR for reference and target genes. Efficiencies
and R² values are indicated.
Target genes F-primer R-primer Efficiency (%) R2
IPPI 5’- CCTTCCCCAACCATGTAACT-3’ 5’- ACCAACGCCATTTGTCTCTT-3’ 100 0.992
FPPS 5’-TGCTTTGGAGATCCTGAGGT-3’ 5’- TGTTCAGGAGTGGTTCGTTG-3’ 96 0.987
FOLD 5’- TGGCGCGTAGGGTAGACAA-3’ 5’- GACCCATTTGAAAGCCTCCTTGA-3’ 93 0.993
JHAMT 5’- ATCCAGGTGCTGGAAGGAGAG-3’ 5’- CTGCCCAGAGTCGAACAGG-3’ 99 0.984
CYP15 5’- GTTGGGATCTCGGAGCATGG-3’ 5’- CGAACACGTCATGCATCGGT-3’ 100 0.992
Table S3.2. Primers for dsRNA construction
Name F Primer(5'-3') R Primer(5'-3')
AstR dsRNA TGTAATCATACGGCTAACGGATC
AATGGTAGAACGTAGTCTGTTGC
AST dsRNA GTCTACCGTCGGCTCTTGTA
TGCTTCTCTTGCCGAGACC
Control dsRNA TTGCGCTCACTGCCAATTGC CTGGCCTTTTGCTCACATGTT
*The T7 promoter sequence was added at the 5' end of each dsRNA primer.
133
B r N C C A F b O v T e AG M G M T
0
5
1 0
1 5
Re
lati
ve
mR
NA
qu
an
tity
D ip p u -A s tR
D ip p u -A S T
Figure S3.1. Relative transcript levels of Dippu-AstR (black) and Dippu-AST (gray) mRNA
in tissues of day 7 male and mated female D. punctata. Dippu-AstR/AST mRNA were
quantified by q-RT-PCR. The data represent the average of 3 biologically independent pools (10
animals per pool), run in triplicate. Tissue abbreviations are as described in Figure 2.1. Values
represent mean ± SEM.

134
References
Aucoin, R.R., Rankin, S.M., Stay, B., Tobe, S.S., 1987. Calcium and cyclic-AMP involvement in the
regulation of juvenile hormone biosynthesis in Diploptera Punctata. Insect Biochem 17, 965-969.
Auerswald, L., Birgul, N., Gade, G., Kreienkamp, H.J., Richter, D., 2001. Structural, functional, and
evolutionary characterization of novel members of the allatostatin receptor family from insects. Biochem
Biophys Res Commun 282, 904-909.
Belles, X., Martin, D., Piulachs, M.D., 2005. The mevalonate pathway and the synthesis of juvenile hormone
in insects. Annu Rev Entomol 50, 181-199.
Bendena, W.G., Donly, B.C., Tobe, S.S., 1999. Allatostatins: a growing family of neuropeptides with
structural and functional diversity. Annals of the New York Academy of Sciences 897, 311-329.
Birgul, N., Weise, C., Kreienkamp, H.J., Richter, D., 1999. Reverse physiology in Drosophila: identification
of a novel allatostatin-like neuropeptide and its cognate receptor structurally related to the mammalian
somatostatin/galanin/opioid receptor family. Embo J 18, 5892-5900.
Bowser, P.R.F., Tobe, S.S., 2000. Partial characterization of a putative allatostatin receptor in the midgut of the
cockroach Diploptera punctata. Gen Comp Endocrinol 119, 1-10.
Coast, G.M., Schooley, D.A., 2011. Toward a consensus nomenclature for insect neuropeptides and peptide
hormones. Peptides 32, 620-631.
Cusson, M., Prestwich, G.D., Stay, B., Tobe, S.S., 1991. Photoaffinity-Labeling of Allatostatin Receptor
Proteins in the Corpora Allata of the Cockroach, Diploptera-Punctata. Biochem Biophys Res Commun 181,
736-742.
Cusson, M., Prestwich, G.D., Stay, B., Tobe, S.S., 1992. Evidence for allatostatin receptor proteins in the
corpora allata of the cockroach, Diploptera punctata, as demonstrated by photoaffinity labeling. Insect
Juvenile Hormone Res, 183-191.
Donly, B.C., Ding, Q., Tobe, S.S., Bendena, W.G., 1993. Molecular cloning of the gene for the allatostatin
family of neuropeptides from the cockroach Diploptera punctata. Proc Natl Acad Sci U S A 90, 12055-12055.
Duve, H., Wren, P., Thorpe, A., 1995. Innervation of the foregut of the cockroach Leucophaea maderae and
inhibition of spontaneous contractile activity by callatostatin neuropeptides. Physiol Entomol 20, 33-44.
Feyereisen, R., Farnsworth, D.E., 1987a. Comparison of the Inhibitory Effects of Brain Extract, High K+ and
Forskolin on Juvenile-Hormone Synthesis by Diploptera-Punctata Corpora Allata. Insect Biochem 17, 939-942.
Feyereisen, R., Farnsworth, D.E., 1987b. Inhibition of Insect Juvenile-Hormone Synthesis by Phorbol 12-
Myristate 13-Acetate. Febs Lett 222, 345-348.
Feyereisen, R., Tobe, S.S., 1981. A Rapid Partition Assay for Routine Analysis of Juvenile-Hormone Release
by Insect Corpora Allata. Anal Biochem 111, 372-375.
Fuse, M., Zhang, J.R., Partridge, E., Nachman, R.J., Orchard, I., Bendena, W.G., Tobe, S.S., 1999. Effects of
an allatostatin and a myosuppressin on midgut carbohydrate enzyme activity in the cockroach Diploptera
punctata. Peptides 20, 1285-1293.
Gade, G., Marco, H.G., Richter, D., Weaver, R.J., 2008. Structure-activity studies with endogenous
allatostatins from Periplaneta americana: Expressed receptor compared with functional bioassay. J Insect
Physiol 54, 1557-1557.
Garside, C.S., Hayes, T.K., Tobe, S.S., 1997a. Degradation of Dip-allatostatins by hemolymph from the
cockroach, Diploptera punctata. Peptides 18, 17-25.
Garside, C.S., Hayes, T.K., Tobe, S.S., 1997b. Inactivation of Dip-allatostatin 5 by membrane preparations
from the cockroach Diploptera punctata. Gen Comp Endocrinol 108, 258-270.

135
Kikukawa, S., Tobe, S.S., Solowiej, S., Rankin, S.M., Stay, B., 1987. Calcium as a regulator of juvenile
hormone biosynthesis and release in the cockroach Diploptera punctata. Insect Biochem 17, 179-187.
Lange, A.B., Bendena, W.G., Tobe, S.S., 1995. The effect of the 13 Dip-allatostatins on myogenic and induced
contractions of the cockroach (Diploptera punctata) hindgut. J Insect Physiol 41, 581-588.
Larsen, M.J., Burton, K.J., Zantello, M.R., Smith, V.G., Lowery, D.L., Kubiak, T.M., 2001. Type A
allatostatins from Drosophila melanogaster and Diplotera puncata activate two Drosophila allatostatin
receptors, DAR-1 and DAR-2, expressed in CHO cells. Biochem Biophys Res Commun 286, 895-901.
Lenz, C., Williamson, M., Grimmelikhuijzen, C.J.P., 2000. Molecular cloning and genomic organization of a
second probable allatostatin receptor from Drosophila melanogaster. Biochem Biophys Res Commun 273,
571-577.
Lloyd, G.T., Woodhead, A.P., Stay, B., 2000. Release of neurosecretory granules within the corpus allatum in
relation to the regulation of juvenile hormone synthesis in Diploptera punctata. Insect Biochem Mol Biol 30,
739-746.
Lungchukiet, P., Donly, B.C., Zhang, J.R., Tobe, S.S., Bendena, W.G., 2008a. Molecular cloning and
characterization of an allatostatin-like receptor in the cockroach Diploptera punctata. Peptides 29, 276-285.
Lungchukiet, P., Zhang, J.R., Tobe, S.S., Bendena, W.G., 2008b. Quantification of allatostatin receptor mRNA
levels in the cockroach, Diploptera punctata, using real-time PCR. J Insect Physiol 54, 981-987.
Maestro, J.L., Belles, X., 2006. Silencing allatostatin expression using double-stranded RNA targeted to
preproallatostatin mRNA in the German cockroach. Archives of insect biochemistry and physiology 62, 73-79.
Marchal, E., Hult, E.F., Huang, J., Stay, B., Tobe, S.S., 2013a. Diploptera punctata as a model for studying the
endocrinology of arthropod reproduction and development. Gen Comp Endocrinol 188, 85-93.
Marchal, E., Hult, E.F., Huang, J., Tobe, S.S., 2013b. Sequencing and validation of housekeeping genes for
quantitative real-time PCR during the gonadotrophic cycle of Diploptera punctata. BMC Res Notes 6, 237.
Martin, D., Piulachs, M.D., Belles, X., 1996. Inhibition of vitellogenin production by allatostatin in the
German cockroach. Mol Cell Endocrinol 121, 191-196.
Meller, V.H., Aucoin, R.R., Tobe, S.S., Feyereisen, R., 1985. Evidence for an inhibitory role of cyclic AMP in
the control of juvenile hormone biosynthesis by cockroach corpora allata. Mol Cell Endocrinol 43, 155-163.
Nouzova, M., Edwards, M.J., Mayoral, J.G., Noriega, F.G., 2011. A coordinated expression of biosynthetic
enzymes controls the flux of juvenile hormone precursors in the corpora allata of mosquitoes. Insect Biochem
Mol Biol 41, 660-669.
Pratt, G.E., Farnsworth, D.E., Feyereisen, R., 1990. Changes in the sensitivity of adult cockroach corpora
allata to a brain allatostatin. Mol Cell Endocrinol 70, 185-195.
Pratt, G.E., Farnsworth, D.E., Fok, K.F., Siegel, N.R., McCormack, A.L., Shabanowitz, J., Hunt, D.F.,
Feyereisen, R., 1991. Identity of a second type of allatostatin from cockroach brains: an octadecapeptide amide
with a tyrosine-rich address sequence. Proc Natl Acad Sci U S A 88, 2412-2416.
Pratt, G.E., Farnsworth, D.E., Siegel, N.R., Fok, K.F., Feyereisen, R., 1989. Identification of an Allatostatin
from Adult Diploptera-Punctata. Biochem Biophys Res Commun 163, 1243-1247.
Rachinsky, A., Tobe, S.S., 1996. Role of second messengers in the regulation of juvenile hormone production
in insects, with particular emphasis on calcium and phosphoinositide signaling. Archives of insect
biochemistry and physiology 33, 259-282.
Rachinsky, A., Zhang, J., Tobe, S.S., 1994. Signal transduction in the inhibition of juvenile hormone
biosynthesis by allatostatins: roles of diacylglycerol and calcium. Mol Cell Endocrinol 105, 89-96.

136
Secher, T., Lenz, C., Cazzamali, G., Sorensen, G., Williamson, M., Hansen, G.N., Svane, P.,
Grimmelikhuijzen, C.J.P., 2001. Molecular cloning of a functional allatostatin gut/brain receptor and an
allatostatin preprohormone from the silkworm Bombyx mori. J Biol Chem 276, 47052-47060.
Stables, J., Green, A., Marshall, F., Fraser, N., Knight, E., Sautel, M., Milligan, G., Lee, M., Rees, S., 1997. A
bioluminescent assay for agonist activity at potentially any G-protein-coupled receptor. Anal Biochem 252,
115-126.
Stay, B., Chan, K.K., Woodhead, A.P., 1992. Allatostatin-immunoreactive neurons projecting to the corpora
allata of adult Diploptera punctata. Cell Tissue Res 270, 15-23.
Stay, B., Fairbairn, S., Yu, C.G., 1996. Role of allatostatins in the regulation of juvenile hormone synthesis.
Archives of insect biochemistry and physiology 32, 287-297.
Stay, B., Joshi, S., Woodhead, A.P., 1991. Sensitivity to allatostatins of corpora allata from larval and adult
female Diploptera Punctata. J Insect Physiol 37, 63-70.
Stay, B., Tobe, S.S., 2007. The role of allatostatins in juvenile hormone synthesis in insects and crustaceans.
Annual review of entomology 52, 277-299.
Sutherland, T.D., Feyereisen, R., 1996. Target of cockroach allatostatin in the pathway of juvenile hormone
biosynthesis. Mol Cell Endocrinol 120, 115-123.
Tobe, S.S., Clarke, N., 1985. The Effect of L-Methionine Concentration on Juvenile-Hormone Biosynthesis by
Corpora-Allata of the Cockroach Diploptera-Punctata. Insect Biochem 15, 175-179.
Tobe, S.S., Zhang, J.R., Bowser, P.R., Donly, B.C., Bendena, W.G., 2000. Biological activities of the
allatostatin family of peptides in the cockroach, Diploptera punctata, and potential interactions with receptors.
J Insect Physiol 46, 231-242.
Vandesompele, J., De Preter, K., Pattyn, F., Poppe, B., Van Roy, N., De Paepe, A., Speleman, F., 2002.
Accurate normalization of real-time quantitative RT-PCR data by geometric averaging of multiple internal
control genes. Genome biology 3, RESEARCH0034.
Vilaplana, L., Maestro, J.L., Piulachs, M.D., Belles, X., 1999. Modulation of cardiac rhythm by allatostatins in
the cockroach Blattella germanica (L.) (Dictyoptera, Blattellidae). J Insect Physiol 45, 1057-1064.
Yu, C.G., Hayes, T.K., Strey, A., Bendena, W.G., Tobe, S.S., 1995. Identification and partial characterization
of receptors for allatostatins in brain and corpora allata of the cockroach Diploptera punctata using a binding
assay and photoaffinity labeling. Regul Pept 57, 347-358.

137
Chapter 4
Identification and characterization of the NMDA receptor and its
role in regulating reproduction in the cockroach, Diploptera
punctata
This chapter is an adapted reprint of my article:
Juan Huang, Ekaterina F. Hult, Elisabeth Marchal, Stephen S. Tobe, Identification and
characterization of the NMDA receptor and its role in regulating reproduction in the cockroach,
Diploptera punctata, 2014, paper submitted to ‘Journal of experimental biology’
Authors’ contribution:
I designed and performed all experiments in this study and wrote the paper with editing by E.
Marchal, E.F. Hult, and S.S. Tobe.

138
Summary
The NMDA receptor (NMDAR) plays important roles in excitatory neurotransmission and in the
regulation of reproduction in mammals. NMDAR in insects comprises two subunits, NR1 and
NR2. In this study, we identified two NR1 paralogs and eleven NR2 alternative splicing variants
in D. punctata. This is the first report of NR1 paralogs in insects. The tissue distribution and
expression profile of DpNR1A, DpNR1B and DpNR2 in different tissues were also investigated.
Previous studies have demonstrated NMDA-stimulated JH biosynthesis in the corpora allata (CA)
through the influx of extracellular Ca2+ in Diploptera punctata. However, our data show that the
transcript levels of DpNR1A, DpNR1B and DpNR2 were low in the CA. In addition, neither
partial knockdown of DpNR2 nor in vivo treatment with a physiologically relevant dose of MK-
801 resulted in any significant change in JH biosynthesis by CA or basal oocyte growth.
Injection of animals with a high dose of MK-801 (30 µg/animal/injection), which paralyzed the
animals for 4-5 h, resulted in a significant decrease in JH biosynthesis on days 4 and 5. However,
the reproductive events during the first gonadotrophic cycle in female D. punctata were
unaffected. Thus, NMDAR does not appear to play important roles in the regulation of JH
biosynthesis or mediate reproduction of female D. punctata.
Introduction
L-glutamate (Glu), a major excitatory amino acid transmitter, mediates diverse physiological
functions in the vertebrate nervous system (Mahesh and Brann, 2005). The Glu receptors (GluR)
have been classified into three major subtypes: the α-amino-3-hydroxy-5-methyl-4-
isoxazolepropionic acid (AMPA) receptor, N-methyl-D-aspartate (NMDA) receptor and kainate
receptor (Madden, 2002). The NMDA receptors (NMDAR) are distinguished from other

139
ionotropic receptors by their unique properties, including selective agonists and antagonists, high
Ca2+ permeability, and voltage-dependent Mg2+ blockade (McBain and Mayer, 1994). The
unique properties of NMDAR allow it to play key roles in excitatory neurotransmission and
important neurological processes, including learning, memory and behavior (Mussig et al., 2010;
Newcomer and Krystal, 2001; Xia et al., 2005).
NMDAR in vertebrates is composed of two subunits, NR1 and NR2, and in some cases NR3
subunits (Madden, 2002). The NR1 subunit is essential for the basic channel activity of NMDAR,
whereas the NR2 subunit contributes to enhance and modulate the receptor function (Sydow et
al., 1996). Although much is known about the function of NMDAR in vertebrates, little
information is available on NMDARs in insects. Thus far, two subunits, NR1 and NR2, have
been identified in insects. The NR1 and NR2 subunits were previously reported to be distributed
throughout the brain of Drosophila melanogaster and Apis mellifera (Wu et al., 2007; Xia et al.,
2005; Zannat et al., 2006).
The importance of NMDAR in the regulation of reproduction of mammals is well-known.
NMDAR mediates reproduction through the regulation of pulse and surge gonadotropin-
releasing hormone (GnRH)/luteinizing hormone (LH) secretion (Maffucci et al., 2009; Mahesh
and Brann, 2005). In insects, juvenile hormones (JHs) are key regulators of growth, development,
metamorphosis, aging, caste differentiation and reproduction (Goodman and Granger, 2005;
Hartfelder, 2000). As a result of the importance of JH in physiological processes, its biosynthesis
is tightly regulated by many factors, including neuropeptides (allatostatins, allatotropins) (Stay
and Tobe, 2007) and neurotransmitters (octopamine, dopamine and glutamate) (Granger et al.,
1996; Pszczolkowski et al., 1999; Thompson et al., 1990). Chiang et al (2002) demonstrated that
JH biosynthesis by corpora allata (CA) of the cockroach, Diploptera punctata, is stimulated by

140
an NMDA-induced influx of Ca2+ ions and this elevation was significantly reduced by NMDAR
antagonists including Mg2+, MK-801 or conantokin T. Furthermore, a Drosophila larval mutant
for NMDAR1 showed reduced mRNA levels of the gene encoding JH acid methyltransferase
(JHAMT), a key regulatory enzyme of JH biosynthesis in this species (Huang et al., 2011).
These studies suggest that NMDAR plays a role in the regulation of JH biosynthesis.
As a high-affinity antagonist of NMDARs, MK-801 has been used to study the function of
NMDAR in both vertebrates and invertebrates (Rawls et al., 2009; Sircar et al., 1987; Troncoso
and Maldonado, 2002). In addition to the inhibitory effect of MK-801 in NMDA-stimulated JH
biosynthesis in D. punctata, MK-801 was found to influence ovarian development and
vitellogenesis in the flesh fly Neobellieria bullata and the locust Schistocerca gregaria (Begum
et al., 2004; Chiang et al., 2002). In S. gregaria, the inhibition of vitellogenesis was overcome by
treatment with JH. A later study on the butterfly Bicyclus anynana and the cricket Gryllus
bimaculatus showed that MK-801 affects JH biosynthesis in vitro and JH titres in both species,
and subsequently regulates insect reproduction (Geister et al., 2008).
D. punctata is a well-known model in studying the physiology of JH biosynthesis and regulation;
in this animal, JH biosynthesis is high and stable, and the reproductive events correlate very well
with rates of JH production (see review by Marchal et al. (2013a)). In this study, we chose D.
punctata as our model to determine the role of NMDAR in the regulation of JH biosynthesis and
reproduction. We identified the genes encoding the subunits of NMDAR in D. punctata, and
examined the expression of NMDAR in several tissues. In addition, we investigated the roles of
NMDAR in the regulation of JH biosynthesis, vitellogenesis and oocyte growth in vivo using
RNA interference (RNAi) and MK-801 treatments.

141
Materials and Methods
Insects - D. punctata were reared in cages and fed with lab chow and water at libitum at 27 -28
ºC in a dark room. Newly molted male and female adult cockroaches were picked from the
colony and raised in separate containers. Mated status in the females was confirmed by the
presence of a spermatophore.
Tissue collection - Cockroach tissues were dissected under a dissecting microscope. Basal oocyte
length was measured to determine the physiological age of female cockroaches. Selected tissues
were dissected and cleaned in sterile cockroach ringer solution (150 mM NaCl, 12 mM KCl, 10
mM CaCl2.2H2O, 3 mM MgCl2.6H2O, 10 mM HEPES, 40 mM Glucose, pH 7.2), flash-frozen in
liquid nitrogen to prevent RNA degradation and stored at -80 ºC until further processing.
RNA extraction and cDNA synthesis - Selected tissues were collected from adult females and
male for gene sequence, tissue distribution and developmental profiling. RNA extraction and
cDNA synthesis were performed as described by Marchal et al. (2013b). For the tissue
distribution and developmental profiling, three biologically independent pools of 10 animals
each were collected. For RNAi and MK-801 treatment experiments, 3 biologically independent
pools of brain and fat body were collected, each pool containing tissue from 5 animals.
Sequencing of DpNR1A, DpNR1B and DpNR2 - Degenerate primer sequences were designed for
DpNR1A, DpNR1B and DpNR2 based on conserved amino acid sequences of several insect
orthologs. Primers used for degenerate PCR are listed in Table S4.1. Partial sequences were
obtained using these primers in a standard T-gradient PCR using Taq DNA polymerase (Sigma-
Aldrich) and a D. punctata brain cDNA sample. After purification, the resulting DNA fragments
were subcloned into a pJET 1.2/blunt cloning vector (CloneJet PCR Cloning Kit, Thermo

142
Scientific) and sequenced following the protocol outlined in the ABI PRISM BigDye Terminator
Ready Reaction Cycle Sequencing Kit (Applied Biosystems). The complete sequence of
DpNR1A, DpNR1B and DpNR2 was obtained using 5’-RACE (Rapid Amplification of cDNA
Ends) and 3’-RACE strategies, following the protocol outlined in the Roche 5'/3' RACE Kit.
Primers used for RACE are listed in Table S4.1.
Phylogenetic analysis - For the phylogenetic analysis of DpNR1 genes, the NR1 sequences of 15
insect species (identified or predicted) were used. These sequences were aligned using ClustalW
as implemented within MEGA 6.06 (Tamura et al., 2013). Poorly aligned positions and gaps
were removed, which resulted in 854 amino acid residues. The obtained alignment was used to
construct a phylogenetic tree in PhyML 3.0 (Guindon and Gascuel, 2003) based on the
maximum-likelihood principle, using the WAG substitution model (Whelan and Goldman, 2001).
Four substitution rate categories were used to estimate the gamma parameter shape with 100
bootstrap replicates to assess branch support (Felsenstein, 1985; Yang, 1994). The resulting tree
was then rooted using the sea slug, Aplysia californica as an outgroup.
Quantitative Real Time-PCR (q-RT-PCR) - Primers used for q-RT-PCR are shown in Table S4.2.
Primer sets were validated by determining relative standard curves for each gene transcript using
a five-fold serial dilution of a calibrator cDNA sample. Efficiency and correlation coefficient (R²)
can be found in Table S2. Reactions were performed in triplicate on a CFX384 Touch ™ Real-
Time PCR Detection System (Bio-Rad) as described previously by Marchal et al. (2013b).
Target specificity was confirmed by running a few representative q-RT-PCR products on an
agarose gel containing GelRed™ (Biotium). The optimal housekeeping genes for target gene
profiling and RNAi experiments were chosen according to a previous study (Marchal et al.,

143
2013b). The quantity of mRNA for each tested gene relative to reference genes was determined
as described by Vandesompele et al. (2002).
RNA interference (RNAi) - Primers for DpNR2 and control pJET dsRNA constructs with T7
promoters are shown in Table S4.3. Double-stranded RNA (dsRNA) constructs were prepared
using the MEGAscript® RNAi Kit (Ambion). A PCR using forward and reverse primers with
attached T7 promoters was performed to amplify the fragment, which was subcloned and
sequenced to verify the presence of the T7 promoter. The amplified fragment was used in an
RNA transcription reaction which was incubated overnight to obtain a high yield of annealed
dsRNA construct. A nuclease digestion was subsequently performed to remove ssRNA and DNA
remaining in the product. The dsRNA was further purified according to the manufacturer’s
instructions (Ambion). Concentration of the dsRNA construct was determined using a Nanodrop
instrument (Thermo Fisher Scientific Inc., Canada). Five-fold diluted dsRNA was run on a 1.2%
agarose gel to examine the quality and integrity of the construct.
Newly molted adult female cockroaches (day 0) were injected with 2 µg of either DpNR2
dsRNA or pJET dsRNA diluted in 5 µl of cockroach saline. This treatment was repeated on days
1, 2 and 3, and the effect of DpNR2 dsRNA was determined on day 4. Brains were dissected and
stored in liquid nitrogen prior to RNA extraction. CA were dissected and cleaned in TC199
medium (GIBCO; 1.3 mM Ca2+, 2% Ficoll, methionine-free) for use in the radiochemical assay
(RCA) (see below). Basal oocyte length was measured during dissection.
MK-801 in vivo assay - MK-801 was dissolved in ddH2O to a concentration of 6 µg/µl and
injected into animals using a Hamilton syringe. Newly molted adult females were injected with
30 µg of MK-801 on days 0, 1 and 3, and the effect of MK-801 was determined from day 4 to
day 8. Fat body was dissected from day 4 animals and stored in liquid nitrogen prior to RNA

144
extraction. CA were dissected and cleaned in TC199 medium (GIBCO; 1.3 mM Ca2+, 2% Ficoll,
methionine-free) for use in the RCA (see below). Basal oocyte length was also measured.
Results
Identification of DpNR1 subunits
Two DpNR1 were identified with open reading frames of 2871bp (DpNR1A) and 2703bp
(DpNR1B). This was accomplished employing a degenerate PCR approach with adult brain
cDNA. 5’-RACE and 3’-RACE experiments were performed to complete the sequences. The two
DpNR1 sequences were deposited in the NCBI GenBank and received accession numbers:
KJ747198 and KJ747199. Unlike the alternative splicing variants of NR1 genes in other insects,
the differences between the two variants in D. punctata are distributed throughout the whole
gene (Fig. 4.1) and may be the result of gene duplication. A phylogenetic tree of NR1 was
constructed by maximum likelihood methods (Fig. 4.2). Generally, the sequences of NR1 were
grouped based on insect orders, and the two DpNR1s (Blattodea) cluster together with
confidence. On the other hand, the relationship between orders is not well-resolved as indicated
by low bootstrap values at deeper nodes. The lack of power to resolve these nodes could be
explained by insufficient sampling among hemimetabolous insects.

145
Fig. 4.1. Amino acid sequence alignment of the two Diploptera NR1 subunit (DpNR1A, DpNR1B), and homologous receptors
from D. melanogaster (DNR1, GenBank acc. no. NP_730940.1) and T. castaneum (TNR1, GenBank acc. no. XP_969654.1,
predicted sequence). Conservatively substituted residues are highlighted in yellow, and the different residues between DpNR1A and
DpNR1B in green. Three putative hydrophobic transmembrane regions (TM1, TM3, and TM4) and one hydrophobic pore-forming
segment (TM2) are highlighted in the boxes. Agonist-binding domain (S1 and S2 domains) are underlined.

146
Fig. 4.2. Phylogram depicting the relationship between the NR1 subunits from Diploptera
and orthologues of this receptor from other insects. Phylogenetic analysis was conducted in
PhyML 3.0 using WAG substitution model with 100 bootstrap replicates. Poorly aligned amino
acid were eliminated by eye resulting in 854 positions. The bar represents 0.2 substitutions per
site. The sea slug, Aplysia californica was used as outgroup to root the tree.

147
Identification of DpNR2 subunits
DpNR2 undergoes alternative splicing, mostly at the 3’ end, generating eleven different
transcripts, which may encode nine different proteins. Full-length cDNAs for all eleven variants
have been isolated and their sequences were deposited in the NCBI GenBank with accession
numbers: KJ747200 to KJ747210. The deduced amino acid sequence and nucleotide sequence of
DpNR2 subunits was shown in Supplementary Fig. S4.1. In the eleven transcripts, there are two
5’ untranslated region, two insertions, and four different 3’ ends (Fig. 4.3A). The sequence of
insertions, deletions and different 3’-ends are shown in Fig. 4.3B. All eleven DpNR2 variants
contain the domain structure, four hydrophobic regions (TM1-TM4) and two ligand binding
domains. Amino acid sequence comparisons between DpNR2A-1 and NR2 subunits from D.
melanogaster, A. mellifera, and A. aegypti were determined (Table S4.4). DpNR2A-1 shares the
highest similarity with NR2 subunits from A. mellifera.
Expression of DpNR1A, DpNR1B and DpNR2
Previous studies of NMDAR in insects have focused on its function in brain (Wu et al., 2007;
Xia et al., 2005; Zannat et al., 2006). In the present study, we were interested in assessing the
role of NMDAR in other tissues. We therefore determined the localization and relative
abundance of DpNR1A, DpNR1B and DpNR2 mRNA in day 4 adult male and female
cockroaches using q-RT-PCR (Fig. 4.4). Specific q-RT-PCR primers for each DpNR1 variant
were designed to determine whether transcripts for the different DpNR1 paralogs are present in
the same tissue. Q-RT-PCR primers for DpNR2 are located in the conserved region of DpNR2
common to all putative splice variants. In mated female cockroaches, DpNR1A, DpNR1B and
DpNR2 were highly expressed in brain, followed by nerve cord. Previous study has shown that
NMDA stimulated JH biosynthesis in the CA of Diploptera (Chiang et al., 2002). However, the

148
transcript level of DpNR1A, DpNR1B and DpNR2 in the CA was relatively low compared to
other tissues. In male cockroaches, the highest transcript levels of DpNR1A, DpNR1B and
DpNR2 were measured in the brain (Fig. 4.4). It is interesting to see that the transcript level of
DpNR2 in testes was very low whereas both genes encoding NR1 were high.
To further study the function of NMDAR in D. punctata, we determined the developmental
profile of DpNR1A, DpNR1B and DpNR2 in the brain, CA and testes. DpNR1A and DpNR1B
were stably transcribed in brain of day 0 to day 7 post-emergence mated female D. punctata.
DpNR2 mRNA in the brain showed a slight increase on days 2, 3 and 7, but none of the changes
were significant (Fig. 4.5). In the CA, the transcript level of DpNR1B and DpNR2 remained very
low and did not show any significant change during the first gonadotrophic cycle (Fig. 6).
DpNR1A mRNA levels exhibited a significant increase on day 6. In male cockroaches, we
studied the expression of NMDAR in testes of males of differing ages. The transcript level of
DpNR2 was very low throughout all the ages tested (Fig. 4.7).
Effect of DpNR2 dsRNA on JH biosynthesis and oocyte growth
To investigate the role of NMDAR in reproduction, DpNR2 was silenced using RNAi. The
injection of 2 µg of DpNR2 dsRNA on days 0, 1, 2 and 3 resulted in a 48.7% knockdown of
DpNR2 mRNA levels in brains of day 4 females (Fig. 4.8a). Knockdown of DpNR2 also resulted
in a decrease in the transcript levels of DpNR1A and DpNR1B. However, the JH biosynthetic
activity of CA and basal oocyte lengths in dsRNA-treated animals did not show any significant
difference from control animals (Fig. 4.8b, 4.8c). A similar result was also observed in DpNR1B
dsRNA- treated animals (Fig. S4.2).

149
Fig. 4.3. Molecular characterization of DpNR2. (A) The schematic structures of the 11
variants of the NR2. NR2 variants are generated by alternative splicing, including two 5’
untranslated region, two insertions, and four different 3’ ends. The 5’ end untranslated region is
shown with black or dash line. The position of insertion 1 is shown by the black arrow and
insertion 2 by a gray arrow. The alternated carboxyl-terminal sequences are indicated by colored
boxes: A1 (Green), A2 (Red), B (Gray), and C (Purple). Four putative transmembrane segments
(TM I – IV) are shown by bold black lines. The agonist-binding domains S1 and S2 are
indicated by the hatched boxes. The accession number is displayed under the gene number. (B)
The sequence of insertion, deletion and the alternative created carboxyl-terminal.

150
B r N C C A F b M G M T O v B r T e A G
0
1
2
3
Re
lati
ve
mR
NA
qu
an
tity
of
NR
1A
B r N C C A F b M G M T O v B r T e A G
0 .0
0 .5
1 .0
1 .5
2 .0
2 .5
Re
lati
ve
mR
NA
qu
an
tity
of
NR
1B
B r N C C A F b M G M T O v B r T e A G
0
1
2
3
4
5
Re
lati
ve
mR
NA
qu
an
tity
of
NR
2
A
B
C
F em a le M ale
F em a le M ale
F em a le M ale
Fig. 4.4. Graphic representation of the relative tissue distribution of (A) DpNR1A transcript
levels, (B) DpNR1B transcript levels and (C) DpNR2 transcript levels in tissues of day 4
adult male and mated female D. punctata. Relative mRNA quantity was normalized against
levels of Tubulin and EF1α mRNA (Marchal et al., 2013b). The data represents an average of 3
pools (10 animals per pool), run in triplicate. Abbreviations used: Br brain, NC nerve cord, CA
corpora allata, Fb fat body, Ov ovary, MG midgut, MT Malpighian tubules, AG accessory gland
and Te testes. Values represent mean ± SEM.
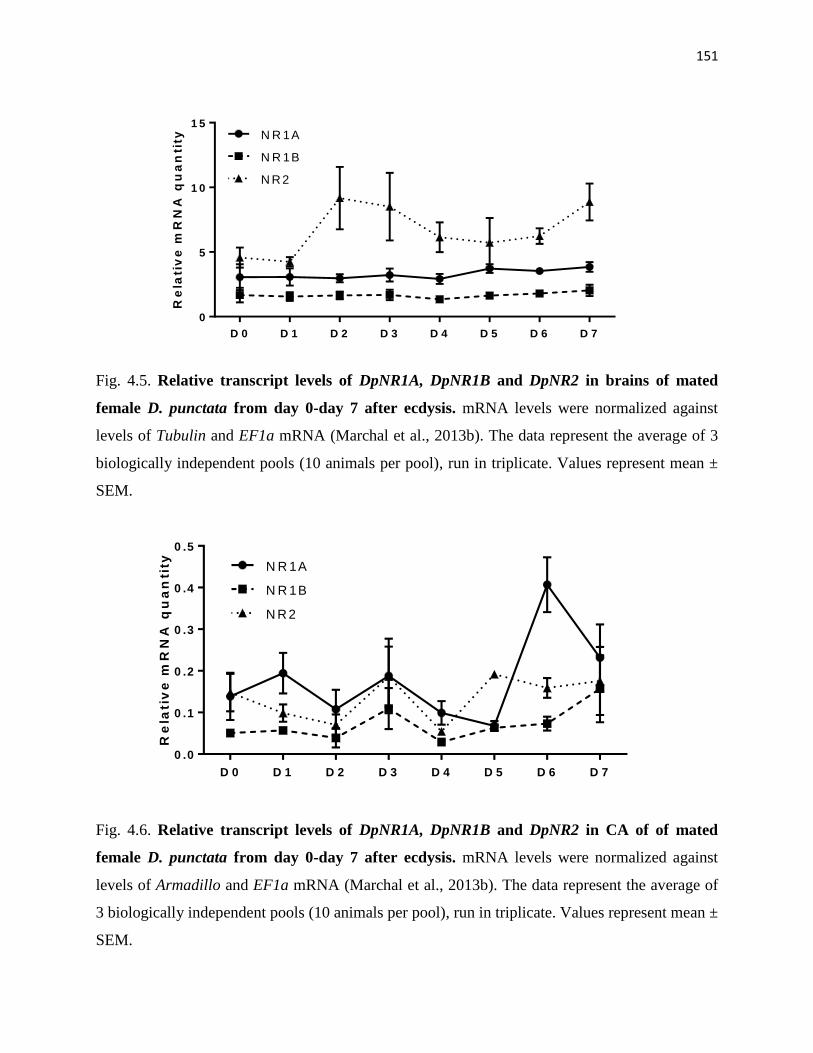
151
D 0 D 1 D 2 D 3 D 4 D 5 D 6 D 7
0
5
1 0
1 5
Re
lati
ve
mR
NA
qu
an
tity N R 1A
N R 1B
N R 2
Fig. 4.5. Relative transcript levels of DpNR1A, DpNR1B and DpNR2 in brains of mated
female D. punctata from day 0-day 7 after ecdysis. mRNA levels were normalized against
levels of Tubulin and EF1a mRNA (Marchal et al., 2013b). The data represent the average of 3
biologically independent pools (10 animals per pool), run in triplicate. Values represent mean ±
SEM.
D 0 D 1 D 2 D 3 D 4 D 5 D 6 D 7
0 .0
0 .1
0 .2
0 .3
0 .4
0 .5
Re
lati
ve
mR
NA
qu
an
tity N R 1A
N R 1B
N R 2
Fig. 4.6. Relative transcript levels of DpNR1A, DpNR1B and DpNR2 in CA of of mated
female D. punctata from day 0-day 7 after ecdysis. mRNA levels were normalized against
levels of Armadillo and EF1a mRNA (Marchal et al., 2013b). The data represent the average of
3 biologically independent pools (10 animals per pool), run in triplicate. Values represent mean ±
SEM.

152
D 0 D 2 D 4 D 6 D 8 D 1 0 D 1 5 D 2 5
0
1
2
3
4
Re
lati
ve
mR
NA
qu
an
tity
N R 1A
N R 1B
N R 2
Fig. 4.7. Relative transcript levels of DpNR1A, DpNR1B and DpNR2 in testes of different
ages of male D. punctata. mRNA levels were normalized against levels of Tubulin and EF1a
mRNA (Marchal et al., 2013b). The data represent the average of 3 biologically independent
pools (10 animals per pool), run in triplicate. Values represent mean ± SEM.
N R 1 A N R 1 B N R 2
0 .0
0 .2
0 .4
0 .6
0 .8
1 .0
Re
lati
ve
mR
NA
qu
an
tity
*
c o n tro l
N R 2 dsR N A
c o n tro l N R 2 d s R N A
0
1 0
2 0
3 0
4 0
JH
bio
sy
nth
es
is(p
mo
l/h
/CA
)
c o n tro l N R 2 d s R N A
0 .0
0 .5
1 .0
1 .5
Oo
cy
te l
en
gth
(m
m)
AB C
Fig. 4.8. The effect of DpNR2 dsRNA treatment on JH biosynthesis and basal oocyte growth,
and the interactions among these genes in mated female D. punctata. (A) Relative quantity of
DippuNR1A, DippuNR1B and DippuNR2 mRNA levels in brain between control and dsRNA
treated animals. mRNA levels were normalized against levels of Tubulin and EF1a mRNA
(Marchal et al., 2013b). The data represent the average of 3 biologically independent pools (5
animals each pool), run in triplicate. (B) JH biosynthesis by CA from control and dsRNA-treated
animals. (C) Basal oocyte length in control and dsRNA-treated animals. Values represent mean ±
SEM. Levels of significance to the control are indicated with the asterisk symbol: *P < 0.05

153
In vivo effect of NMDAR antagonist MK-801 on JH biosynthesis and ovarian development
The effect of NMDAR on JH biosynthesis and ovarian development was also determined by
injecting the NMDAR non-competitive antagonist MK-801 into the animals. To determine the
optimal dose of injection, newly molted adult females were injected with 0 (control), 3, 12 and
30 µg MK-801 on days 0, 1 and 3. JH biosynthesis and basal oocyte length were determined on
day 4. As shown in Fig. S4.3, there was no significant effect on JH biosynthesis in animals
treated with 3 µg or 12 µg MK-801.
To further study the effect of MK-801 on reproduction, mated female cockroaches were injected
with 30 µg MK-801 on days 0, 1, and 3 following ecdysis. Rates of JH biosynthesis and basal
oocyte lengths in control and treated animals were determined from day 4 to day 8. As shown in
Fig. 4.9A, application of MK-801 resulted in a 34% and 26% decrease of JH biosynthesis in day
4, and 5 animals, respectively. On day 6, however, MK-801 treated animals showed higher rates
of JH biosynthesis than control animals. On days 7 and 8, there was no significant difference in
JH biosynthesis between control and MK-801 treated animals. Furthermore, MK-801 did not
have any effect on basal oocyte length (Fig. 4.9B). All animals oviposited on day 8. To study the
effect of MK-801 on the Vg synthesis, we determined the transcript level of DpVg in the fat body
of day 4 animal. No significant change was found in the transcription of DpVg in MK-801-
treated animals (Fig. 4.9C).

154
Figure 4.9. In vivo effect of MK-801 on JH biosynthesis (A), basal oocyte growth (B) and relative Vg mRNA levels (C). Females
were injected with MK-801 on days 0, 1 and 3 following ecdysis (30µg/animal). Control was injected with ddH2O. JH biosynthesis (A)
and oocyte length (B) were determined on day 4 to day 8. All animals oviposited on day 8. The mRNA level of Vg was determined in
the fat body of day 4 animals. mRNA levels were normalized to levels of Tubulin and EF1a mRNA (Marchal et al., 2013b). The data
represent the average of 3 biologically independent pools (5 animals each pool), run in triplicate. Values represent mean ± SEM. (A)
N≥15; (B) N≥10. Levels of significance to the control are indicated with an asterisk: *P < 0.05, **P < 0.01, ***P < 0.001
d a y 4 d a y 5 d a y 6 d a y 7 d a y 8
0
1 0
2 0
3 0
4 0
5 0
6 0
7 0
A G E
JH
bio
sy
nth
es
is(p
mo
l/h
/CA
)
c o n tro l
M K -8 0 1
***
**
*
d a y 4 d a y 5 d a y 6 d a y 7
1 .2
1 .4
1 .6
1 .8
2 .0
2 .2
A G E
Ba
sa
l o
oc
yte
le
ng
th (
mm
)
c o n tro l
M K -8 0 1
c o n tro l M K -8 0 1
0 .0
0 .5
1 .0
1 .5
Re
lati
ve
mR
NA
qu
an
tity
of
VG
A B C

155
Discussion
We have identified two distinct NR1 genes and eleven NR2 variants in D. punctata. The major
functional domains appear to be well conserved in both DpNR1 and DpNR2 amino acid
sequences (Fig. 4.1 and 4.3). The protein contains three hydrophobic transmembrane regions
(TM1, 3-4), a hydrophobic pore-forming segment (TM2), and two ligand binding domains (S1
and S2) (Fig. 4.1 and 4.3) (Dingledine et al., 1999; Kuryatov et al., 1994; Stern-Bach et al.,
1994). The asparagine residue, which was predicted to control Ca2+ permeability and voltage-
dependent Mg2+ blockade, is present in the TM2 domain of DpNR1 (N630 in DpNR1A and
N608 in DpNR1B) (Burnashev et al., 1992). The overall amino acid sequence identity between
DpNR1, DpNR2 and NMDAR subunits in other species is shown in Table S4.4. DpNR1A has
higher sequence identity to NR1 in other species than DpNR1B. Both DpNR1 and DpNR2
subunits show highest homology to NMDAR of A. mellifera.
The two NR1 subunits are 65.6% identical at the amino acid level and the differences are
distributed throughout the entire gene, which suggests that the two NR1 paralogs result from
gene duplication (Fig. 4.1). The other NR1 paralogs were isolated from the Zebrafish (Cox et al.,
2005). They encode the same length of protein with only a few amino acid differences. No NR1
paralogs other than in Zebrafish have been identified in vertebrate or invertebrate species.
Phylogenetic analysis of DpNR1 with NR1s from other insect species shows that the two
DpNR1s are grouped together, which suggests that these are paralogs resulting from a gene
duplication event in D. punctata that did not occur in other insects. However, given that our
dataset was composed primarily of higher insects, and the small number of insect species in
which NR1 has been identified, it is difficult to make definitive conclusions about the origin of
the paralogs. DpNR2 undergoes alternative splicing, generating eleven different transcripts that

156
may encode nine protein isoforms (Fig. 4.3). Unlike NR2 in Drosophila in which alternative
splicing occurs mainly at the 5’ untranslated region, DpNR2 undergoes alternative splicing
principally at the 3’-end (Xia et al., 2005).
Tissue distribution shows that both DpNR1 and DpNR2 mRNA accumulated in brain and nerve
cord, which is consistent with the role of NMDAR in learning and memory (Xia et al., 2005).
The NR1 subunit is essential for the basic channel activity of NMDAR, whereas the NR2 subunit
is regarded as the rate-limiting molecule in controlling the optimal channel properties of
NMDAR (Monyer et al., 1992; Sprengel et al., 1998; Tang et al., 1999). In D. punctata, we
observed that both DpNR1 paralogs are stably expressed in brain, whereas the relative DpNR2
mRNA level underwent a slight change, probably to regulate the activity of NMDAR (Fig. 4.5).
In rats, D-aspartic acid (D-Asp) can induce testosterone synthesis through the activation of
NMDAR in testis (Santillo et al., 2014). A high transcript level of DpNR1 was observed in adult
D. punctata testes, indicating that NMDAR may also play a role in regulation of reproduction in
our model insect (Fig. 4.6). However, the transcript level of DpNR2 in testes was negligible
compared to that of DpNR1, which suggests that NR1 may form a homomeric functional channel,
as was observed in an earlier study in which expression of Drosophila NR1 alone produced a
weak but significant NMDA response (Ultsch et al., 1993; Xia et al., 2005). An alternative
explanation for the low expression of DpNR2 in the testes could be the existence of one or more
additional DpNR2 subunits in D. punctata. Four distinct NR2 subunits (A-D) were identified in
mammalian species. All four NR2 subunits are expressed in brain (Cull-Candy et al., 2001). In
testis, however, the expression of NR2 subunits varies (Hu et al., 2004; Santillo et al., 2014). In
the rat testis, only NR2A and NR2D are strongly expressed, whereas NR2B and NR2C are
undetectable (Santillo et al., 2014). In mouse testis, on the other hand, only NR2B subunit is

157
highly expressed (Hu et al., 2004). Thus, it is possible that an additional NR2 gene is expressed
in the testis of D. punctata.
The function of NMDAR in reproduction is well-studied in mammals (Mahesh and Brann, 2005).
However, clear evidence linking NMDAR to reproduction in insects remains limited. The
possible role of NMDAR in reproduction of insects was described by Chiang et al. (2002). In
that study, NMDA stimulates JH biosynthesis by inducing a Ca2+-influx from the extracellular
environment in the CA of D. punctata (Chiang et al., 2002). However, our study demonstrates
that the transcript levels of DpNR1 and DpNR2 in the CA are very low throughout the first
gonadotrophic cycle.
Treatment with MK-801 in vivo blocked vitellogenesis in S. gregaria and resulted in a reduction
in JH titre in G. bimaculatus (Begum et al., 2004; Geister et al., 2008). For this reason, the role
of NMDAR in JH biosynthesis and reproduction was further examined by the administration of
MK-801 in vivo in D. punctata. Administration of a high dose of MK-801 results in a significant
decrease in JH biosynthesis by day 4 and 5 CA, indicating that MK-801 does have an effect on
JH biosynthesis. However, the effect of MK-801 on JH biosynthesis is not apparent at doses less
than 12 µg/animal/injection (about 60 µg/g body mass) (Fig. S4.3). The result of administration
of MK-801 in S. gregaria suggests that the synthesis of Vg in the fat body was affected by MK-
801 injection (Begum et al., 2004). In D. punctata, the production of Vg in fat body and the
uptake of Vg by the basal oocytes are JH-dependent events (Marchal et al., 2013a; Rankin and
Stay, 1984; Stay and Tobe, 1978). Although rates of JH biosynthesis on days 4 and 5 in MK-
801-treated animals are lower than in controls, the change in transcript level of DpVg was not
significant relative to controls (Fig. 4.9C), and the pattern of JH biosynthesis during the first
gonadotrophic cycle is similar to that in control animals (Fig. 4.9A). Oocyte growth in treated

158
animals is also similar to the controls (Fig. 4.9B). All females oviposited on day 8. Overall,
administration of MK-801 does not have a significant effect on reproduction in female D.
punctata.
Treatment of animals with an extremely high dose of MK-801 in vivo significantly reduced JH
biosynthesis on days 4 and 5, even though MK-801 did not show any significant effect on JH
biosynthesis in vitro. Partial knockdown of DpNR2 did not show any effect on JH biosynthesis or
on basal oocyte growth (Fig. 4.8). To date, there is no clear evidence showing that the effect of
MK-801 on JH biosynthesis is mediated through NMDAR. In rats, injection of 0.2 µg/g MK-801
resulted in a failure to elevate LH, FSH, or progesterone (Luderer et al., 1993). In rats and mice,
a dose of 0.1 µg/g MK-801 was the maximum dose that could be used without causing
sensorimotor impairments and/or signs of intoxication (Van der Staay et al., 2011). Injection of
30 µg/animal of MK-801 paralyzed the cockroaches for 4-5 h. Therefore, it is possible that the
significant decrease in JH biosynthesis on days 4 and 5 in D. punctata was the result of
physiological stress induced by MK-801 treatment, rather than the action of NMDAR.
Accordingly, NMDAR does not appear to play important roles in the regulation of JH
biosynthesis or reproduction in female D. punctata.
In conclusion, two NR1 paralogs and eleven NR2 alternative splicing variants have been
identified in D. punctata. The expression of NMDAR subunits suggests that NMDAR may play
a role in the reproduction of male cockroaches. However, in the female cockroach, although a
previous study suggested that NMDAR mediates JH biosynthesis in D. punctata, our data reveal
a different story. Neither in vivo treatment of MK-801 nor partial knockdown of DpNR2 have
any effect on JH biosynthesis. The decrease in JH biosynthesis at a high dose in MK-801-treated
animals appears to result from physiological stress, rather than the direct action of NMDAR. In

159
addition, no reproductive events were affected following the blocking of NMDAR activity
through RNAi or MK-801 treatment. A reexamination of the function of NMDA and receptors in
the reproduction of insects now appears to be appropriate and timely.

160
Supplementary data
Table S4.1 Degenerate primer and RACE primer sequences for cloning DpNR1A, DpNR1B
and DpNR2 cDNAs.
Name Symbol Degenerate/RACE primer sequences
NMDA receptor
subunit 1 variant
A
NR1A F 5'- CTRTCGCCCGATGGTCARTTYGG -3'
R 5'- CAGTATGAAYACYCCTGCCATRT -3'
Fn 5'- ACGTATTTCAACATYGGYGGHGT-3'
Rn 5'- CCAAATTGGCCATCCGGCGACAA-3'
3’Race 5'- GGATTCGCCATGATCATTGTGGCA -3'
5’Race 5'- GGATCCATTGAAATCGCTGTGGA -3'
NMDA receptor
subunit 1 variant
B
NR1B F 5'- CTRTCGCCCGATGGTCARTTYGG -3'
R 5'- CAGTATGAAYACYCCTGCCATRT -3'
Fn 5'- ATCCACAGCTCCGACACDGAYGG -3'
Rn 5'- GTACCTTCTCCTATGCCACTATT-3'
3’Race 5'- TAAATGATGCGAGACTGCGTA-3'
5’Race 5'- CCACACATCAGCTTGATGGGA -3'
NMDA receptor
subunit 2
NR2 F 5'- ATACCGGTCATCKCVTGGAAYGC -3'
R 5'- AACATCCAAGARGCYGTRTCRAA -3'
Fn 5' - CTCGGAGAGGGAAGCAGTTGTGG -3’
Rn 5' -TCGAGCAGTCGYTTRTTRAACAT - 3'
3’Race 5'- CCAGAACCGATCCACTGTAGCA -3'
5’Race 5'- CTCTCCGCTCAAGGCCGGAATT -3'

161
Table S4.2 Oligonucleotide sequences for primers used in q-RT-PCR for reference and target genes. Efficiencies and R² values are
indicated.
Reference
genes F-primer R-primer
Amplicon
size (bp)
Efficiency
(%) R2
Tubulin 5'-AAATTACCAACGCTTGCTTTGAA-3' 5'-TGGCGAGGATCGCATTTT-3' 58 95.1 0.993
EF1α 5'-TCGTCTTCCTCTGCAGGATGTCT-3' 5'-GGGTGCAAATGTCACAACCATACC-3' 109 99.2 0.994
Armadillo 5'-GCTACTGCACCACTCACAGAATTATT-
3' 5'-CTGCAGCATACGTTGCAACA-3' 64 94.5 0.980
Target genes F-primer R-primer Amplicon
size (bp)
Efficiency
(%) R2
DpNR1A 5’- ATCGAGAAGCGGAAAACACT -3’ 5’- GTTGCTGGATCATTGACACC -3’ 80 90.2 0.984
DpNR1B 5’- GCACACTTTGGGACTCAAGA -3’ 5’- CCTCCAGCAACTAGCATGAA -3’ 54 100 0.983
DpNR2 5’- AAGAACCAGAACCGATCCAC -3’ 5’- GGCCACTAGGAAATCCAAAA -3’ 105 91.7 0.974
DpVg 5’-AAAGGTGTCCTCAGCCAGC-3’ 5’-TCCTCCATCTCGGATTGGGA-3’ 105 95.1 0.998

162
Table S4.3 Nucleotide sequences of primers used in making the dsRNA constructs.
Gene F-primer (5'-3') R-primer (5'-3')
DpNR2 ACAGGATATGGTATCGCCTTTAGC TCTTGAGCTTCGAAAACTGCAC
DpNR12 TAACTGGGGACAGTCCACAC TCAGGTGAGAGTCCAACATGG
pJET TTGCGCTCACTGCCAATTGC CTGGCCTTTTGCTCACATGTT
Table S4.4 Amino acid identities (in %) between NMDAR subunits in D. punctata and
other insects. DpNR1 was compared with NR1 in other species, and DpNR2A-1 was
compared with NR2 in other species.
DpNR1B D. melanogaster A. mellifera A. aegypti
DpNR1A 65.6 65.4 75.4 68.2
DpNR1B --- 55.6 62.7 58.8
DpNR2A-1 --- 67.3% 75.6% 71.0%

163
AGCTCGACCTTTGGGACTATGATTGTTGGGCAAAGTTGCATCGTAATTCTCTTGTTTACGGTTACCATGGTGACTACTTCGAGGTCTCCTTGGTTGGGTGTGACTCCAGCGGTATCCAACCTTTCGTCGAGTTCGCCCAGATTGTCAGCTTGGAGAGAAAATAACTCTGGGCCGCACCATCACCATCATCAAGGGGAGAAGAAA
GGAAGAGAAGGAAACTTGTCCATTGGACTTATTGTGCCGTACACTAACTTTGGCGTAAGGGACTATATTCGGGCAGTGAAGAGTGCTGTGGAGAAATTGGCGAAACCAAGAGGCAGGAGACTCAACTTTTTCAAGAAGTACAATTTCTCTCCCAACGAAGTTCACAGCGTCATGATGTCACTAACTCCAAGCCCCACTGCCATTCTTAACTCACTCTGCAAGGAGTTTCTCTCTGTGAACGTCTCCGCCATATTGTACCTG
1 - ATGAACTATGAGAAATATGGAAGGAGTACGGCGTCTGCGCAATATTTCTTGCAGCTTGCGGGCTACTTGGGAATCCCGGTTATCGCCTGG - 90 1 - M N Y E K Y G R S T A S A Q Y F L Q L A G Y L G I P V I A W - 30
91 - AACGCGGATAATTCCGGCCTTGAGCGGAGAGCGTCTCAATCAAGCCTTCAGCTGCAGTTAGCACCATCTCTGGAGCACCAGACGGCCGCC - 180 31 - N A D N S G L E R R A S Q S S L Q L Q L A P S L E H Q T A A - 60
181 - ATGTTGAGTATATTGGAGAGATACAAGTGGCATCAGTTCTCTGTGGTCACCAGTCAGATAGCGGGTCACGATGACTTCCTACAGGCGGTC - 270 61 - M L S I L E R Y K W H Q F S V V T S Q I A G H D D F L Q A V - 90
271 - AGAGAGAGGATCACCGAAGTGCAAGATAGATTCAAGTTCACGATCCTGAATCAAGTGTTGGTCACAAAGCCGGTAGATTTACTAGACCTG - 360 91 - R E R I T E V Q D R F K F T I L N Q V L V T K P V D L L D L - 120
361 - GTCAACTCTGAGTCTCGAGTGATGTTACTCTACGCCACCAGAGAAGAGGCCATACACATCTTGAAAGCCGCCAGAGATTACCAGATCACT - 450 121 - V N S E S R V M L L Y A T R E E A I H I L K A A R D Y Q I T - 150
451 - GGAGAGAACTACGTATGGGTCGTCACCCAGAGCGTCATGGAGAACCTCCAGACACCTTTCGGTTTCCCTGTCGGCATGCTCGGTGTCCAT - 540 151 - G E N Y V W V V T Q S V M E N L Q T P F G F P V G M L G V H - 180
541 - TTCGATACAAGCAGCACATCCCTTGTGAACGAAATCACCACCGCCATTAGGGTGTACGCATACGGGGTAGAGGATTTTGTGAATGATCCC - 630 181 - F D T S S T S L V N E I T T A I R V Y A Y G V E D F V N D P - 210
631 - AGAAATGTCAATCTCTCTCTCAGCACACAATTATCTTGTGAAGGGATGGGAGACTCCAGATGGAAAACAGGAGACAGATTCTTCAGGTAC - 720 211 - R N V N L S L S T Q L S C E G M G D S R W K T G D R F F R Y - 240
721 - CTCCGGAACGTGAGTGTTGAGGGGGATACAGGAAAGCCACATGTAGAATTCACACCGGAAGGAGTTCTGAAGGCAGCAGAGTTGAAAATA - 810 241 - L R N V S V E G D T G K P H V E F T P E G V L K A A E L K I - 270
811 - ATGAATTTGAGGCCTGGTGTTAGCAAGCAGCTTGTGTGGGAAGAGATCGGAGTGTGGAAATCTTGGGAGAAGGAAGGTCTGGACATCAAA - 900 271 - M N L R P G V S K Q L V W E E I G V W K S W E K E G L D I K - 300
901 - GACATCGTGTGGCCTGGGAACAGTCACACTCCGCCTCAGGGAGTTCCTGAAAAGTTCCACCTGAAGATAACTTTCCTGGAAGAACCTCCA - 990 301 - D I V W P G N S H T P P Q G V P E K F H L K I T F L E E P P - 330
991 - TATATCAACCTTGCGCCCCCGGATCCCGTCACCGGAAAATGCAGCATGAACAGGGGCGTTCTCTGCCGGGTGGCCAAGGAAGAAGAAATG - 1080 331 - Y I N L A P P D P V T G K C S M N R G V L C R V A K E E E M - 360
1081 - GAAAAGGTGGATGTACCGATGGCACATAAAAACGGCAGTTTCTACCAATGCTGCTCTGGGTTCTGCATAGACCTGCTAGAAAAATTCGCA - 1170 361 - E K V D V P M A H K N G S F Y Q C C S G F C I D L L E K F A - 390
S1 1171 - GAAGAACTTGGGTTCACATACGAACTCGTGAGGGTGGAAGATGGAAAATGGGGGACTCTGGAGAACGGGAAGTGGAACGGATTGATAGCG - 1260 391 - E E L G F T Y E L V R V E D G K W G T L E N G K W N G L I A - 420
1261 - GATCTCGTCAACCGGAAGACAGACATGGTGATGACGTCACTGATGATAAACTCGGAGAGGGAAGCAGTTGTGGACTTCACTGTGCCCTTT - 1350 421 - D L V N R K T D M V M T S L M I N S E R E A V V D F T V P F - 450
1351 - ATGGAGACTGGCATTGCCATTTTGGTGGCCAAGAGAACTGGTATCATATCTCCCACAGCTTTTCTTGAACCGTTTGACACGGCCTCCTGG - 1440
451 - M E T G I A I L V A K R T G I I S P T A F L E P F D T A S W - 480
1441 - ATGCTGGTAGGTGTAGTAGCGATCCAGGCAGCCACTTTTACTATTTTCCTTTTCGAATGGCTGAGTCCTTCAGGATTCGACATGAAGAAT - 1530 481 - M L V G V V A I Q A A T F T I F L F E W L S P S G F D M K N - 510
TM 1 1531 - GTGGATGGTTTCCAGCAAGTGTCTCCGTCCCCAAATCACCGGTTTTCATTGTTCAGGACCTATTGGTTAGTGTGGGCTGTCCTCTTCCAA - 1620 511 - V D G F Q Q V S P S P N H R F S L F R T Y W L V W A V L F Q - 540
TM 2 1621 - GCAGCCGTACATGTAGATTCCCCAAGAGGCTTCACTGCCAGGTTCATGACCAATGTGTGGGCCATGTTTGCAGTGGTGTTCCTTGCTATT - 1710 541 - A A V H V D S P R G F T A R F M T N V W A M F A V V F L A I - 570
TM 3 1711 - TACACTGCCAACCTGGCTGCCTTCATGATCACAAGAGAAGAATTTCATGAATTTTCTGGCCTCGACGATTCTAGGCTGTCAAAACCATTC - 1800 571 - Y T A N L A A F M I T R E E F H E F S G L D D S R L S K P F - 600
1801 - AGTCACAAACCCATGTACAGGTTTGGTACTATCCCATGGAGCCACACCGACTCAACTCTTAGTAAATATTTTGCCCCCATGCATGCCTAC - 1890 601 - S H K P M Y R F G T I P W S H T D S T L S K Y F A P M H A Y - 630
S2 1891 - ATGAAGAACCAGAACCGATCCACTGTAGCAGAGGGAATAGAAGCTGTTCTTAGTGGTGAACTGGATGCTTTCATTTATGATGGCACAGTT - 1980 631 - M K N Q N R S T V A E G I E A V L S G E L D A F I Y D G T V - 660
1981 - TTGGATTTCCTAGTGGCCCAGGATGAGGACTGCCGTCTGCTCACTGTGGGATCATGGTATGCCATGACAGGATATGGTATCGCCTTTAGC - 2070 661 - L D F L V A Q D E D C R L L T V G S W Y A M T G Y G I A F S - 690
2071 - CGCAACTCTAAATATGTTCAAATGTTCAACAAGCAAATGCTTGATTTTCGGGAAAATGGGGACTTGGAGAGACTACGGAGGTATTGGATG - 2160 691 - R N S K Y V Q M F N K Q M L D F R E N G D L E R L R R Y W M - 720
2161 - ACCGGGACATGTAAGCCTGGTAAACAAGAACATAAATCCAGTGACCCTCTAGCCCTGGAGCAGTTTTTATCTGCATTTTTGCTGCTCATG - 2250 721 - T G T C K P G K Q E H K S S D P L A L E Q F L S A F L L L M - 750
TM 4 2251 - TCTGGTATCCTACTAGCTGCAGTACTCTTAGCACTAGAACATGTGTATTTCAAATATGTTAGAAAACATTTGGCCAAAACAGACAGAGGA - 2340 751 - S G I L L A A V L L A L E H V Y F K Y V R K H L A K T D R G - 780
2341 - GGTTGCTGTGCTCTAATAAGTTTGAGCATGGGCAAATCATTAACGTTCCGCGGTGCAGTTTTCGAAGCTCAAGATATATTAAGACATCAT - 2430 781 - G C C A L I S L S M G K S L T F R G A V F E A Q D I L R H H - 810
2431 - CGTTGTCGGGATCCTATATGTGATACACATTTATGGAAAGTGAAACACGAGTTGGATCTCGCCCGTATGAAGATTCGACAATTGCAGAAA - 2520 811 - R C R D P I C D T H L W K V K H E L D L A R M K I R Q L Q K - 840
2521 - GAACTTGAAGCTCACGGCATCAAGCCAAGTAGAAGGAAGAAGAAGAAACGCGTCCCGTCCTGCTGGGTCTCCTGCTTCACACGAAACCAG - 2610 841 - E L E A H G I K P S R R K K K K R V P S C W V S C F T R N Q - 870

164
2611 - CCCGCGGGAGAGCAGGCGTCCCAATTGTTGAATGCAGATGACGTCCTCAAACCGCGTGATATGACAAGAAATCACGTGACATCTACAGAA - 2700 871 - P A G E Q A S Q L L N A D D V L K P R D M T R N H V T S T E - 900
2701 - TTCTCCGGCCGTTTTCACAGTGCGGGCCAGTTATACAGGTAA - 2742 901 - F S G R F H S A G Q L Y R * - 930
Fig S4.1. The nucleotide sequence and deduced amino acid sequence of DpNR2B-2. Agonist-
binding domains (S1 and S2 domains) are underlined; three hydrophobic transmembrane regions
(TM1, TM3, and TM4) and one hydrophobic pore-forming segment (TM2) are highlighted in the
boxes. Two insertions, and alternative 3’-end are highlighted in yellow, green and purple,
respectively.
Fig. S4.2. The effect of DpNR1B dsRNA treatment on JH biosynthesis and basal oocyte
growth, and the interactions among these genes in mated female D. punctata. (A) Relative
quantity of DippuNR1A, DippuNR1B and DippuNR2 mRNA levels in brain between control and
dsRNA treated animals. mRNA levels were normalized against levels of Tubulin and EF1a
mRNA. The data represent the average of 3 biologically independent pools (5 animals each pool),
run in triplicate. (B) JH biosynthesis by CA from control and dsRNA-treated animals. (C) Basal
oocyte length in control and dsRNA-treated animals. Values represent mean ± SEM. Levels of
significance to the control are indicated with the asterisk symbol: *P < 0.05, **P < 0.01
N R 1 1 N R 1 2 N R 2
0 .0
0 .5
1 .0
1 .5
2 .0
Re
lati
ve
mR
NA
qu
an
tity
***
N R 12 d sR N A
c o n tro l
C o n tro l N R 1 2 d s R N A
0
2 0
4 0
6 0
8 0
JH
bio
sy
nth
es
is(p
mo
l/h
/CA
)
C o n tro l N R 1 2 d s R N A
0 .0
0 .5
1 .0
1 .5
Ba
sa
l o
oc
yte
le
ng
th (
mm
)
A B C

165
Fig. S4.3. JH biosynthesis of CA in day 4 mated female D. punctata injected with different
doses of MK-801 on days 0, 1 and 3. Control was injected with ddH2O. Values represent mean
± SEM (n ≥17). Levels of significance to the control are indicated with the asterisk symbol: ***P
< 0.001.
3 µ g 1 2 µ g 3 0 µ g
0
2 0
4 0
6 0
JH
bio
sy
nth
es
is (
pm
ol/
h/C
A) c o n tro l
M K -8 0 1
***

166
References
Begum, M., Breuer, M., Kodrik, D., M, M.R., De Loof, A., 2004. The NMDA receptor antagonist MK-801
inhibits vitellogenesis in the flesh fly Neobellieria bullata and in the desert locust Schistocerca gregaria. J.
Insect Physiol. 50, 927-934.
Burnashev, N., Schoepfer, R., Monyer, H., Ruppersberg, J.P., Gunther, W., Seeburg, P.H., Sakmann, B., 1992.
Control by asparagine residues of calcium permeability and magnesium blockade in the NMDA receptor.
Science 257, 1415-1419.
Chiang, A.S., Lin, W.Y., Liu, H.P., Pszczolkowski, M.A., Fu, T.F., Chiu, S.L., Holbrook, G.L., 2002. Insect
NMDA receptors mediate juvenile hormone biosynthesis. P. Natl. Acad. Sci. U.S.A. 99, 37-42.
Cox, J.A., Kucenas, S., Voigt, M.M., 2005. Molecular characterization and embryonic expression of the family
of N-methyl-D-aspartate receptor subunit genes in the zebrafish. Dev. Dynam. 234, 756-766.
Cull-Candy, S., Brickley, S., Farrant, M., 2001. NMDA receptor subunits: diversity, development and disease.
Curr. Opin. Neurobiol. 11, 327-335.
Dingledine, R., Borges, K., Bowie, D., Traynelis, S.F., 1999. The glutamate receptor ion channels. Pharmacol.
Rev. 51, 7-61.
Felsenstein, J., 1985. Confidence limits on phylogenies: An approach using the bootstrap. Evolution 39, 783-
791.
Geister, T.L., Lorenz, M.W., Hoffmann, K.H., Fischer, K., 2008. Effects of the NMDA receptor antagonist
MK-801 on female reproduction and juvenile hormone biosynthesis in the cricket Gryllus bimaculatus and the
butterfly Bicyclus anynana. J. Exp. Biol. 211, 1587-1593.
Goodman, W.G., Granger, N.A., 2005. The juvenile hormones, in: Gilbert, L.I., Iatrou, K., Gill, S.S. (Eds.),
Comprehensive Molecular Insect Science. Elsevier, Oxford, pp. 319-408.
Granger, N.A., Sturgis, S.L., Ebersohl, R., Geng, C., Sparks, T.C., 1996. Dopaminergic control of corpora
allata activity in the larval tobacco hornworm, Manduca sexta. Arch. Insect Biochem. Physiol. 32, 449-466.
Guindon, S., Gascuel, O., 2003. A simple, fast, and accurate algorithm to estimate large phylogenies by
maximum likelihood. Syst. Biol. 52, 696-704.
Hartfelder, K., 2000. Insect juvenile hormone: from "status quo" to high society. Braz. J. Med. Biol. Res. 33,
157-177.
Hu, J.H., Yang, N., Ma, Y.H., Jiang, J., Zhang, J.F., Fei, J., Guo, L.H., 2004. Identification of glutamate
transporters and receptors in mouse testis. Acta Pharmacol. Sin. 25, 366-371.
Huang, J., Tian, L., Peng, C., Abdou, M., Wen, D., Wang, Y., Li, S., Wang, J., 2011. DPP-mediated TGFbeta
signaling regulates juvenile hormone biosynthesis by activating the expression of juvenile hormone acid
methyltransferase. Development 138, 2283-2291.
Kuryatov, A., Laube, B., Betz, H., Kuhse, J., 1994. Mutational analysis of the glycine-binding site of the
NMDA receptor: structural similarity with bacterial amino acid-binding proteins. Neuron 12, 1291-1300.
Luderer, U., Strobl, F.J., Levine, J.E., Schwartz, N.B., 1993. Differential gonadotropin responses to N-methyl-
D,L-aspartate in metestrous, proestrous, and ovariectomized rats. Biol. Reprod. 48, 857-866.
Madden, D.R., 2002. The structure and function of glutamate receptor ion channels. Nat. Rev. Neurosci. 3, 91-
101.
Maffucci, J.A., Noel, M.L., Gillette, R., Wu, D., Gore, A.C., 2009. Age- and hormone-regulation of N-methyl-D-aspartate receptor subunit NR2b in the anteroventral periventricular nucleus of the female rat: implications
for reproductive senescence. J. Neuroendocrinol. 21, 506-517.

167
Mahesh, V.B., Brann, D.W., 2005. Regulatory role of excitatory amino acids in reproduction. Endocrine 28,
271-280.
Marchal, E., Hult, E.F., Huang, J., Stay, B., Tobe, S.S., 2013a. Diploptera punctata as a model for studying the
endocrinology of arthropod reproduction and development. Gen. Comp. Endocr. 188, 85-93.
Marchal, E., Hult, E.F., Huang, J., Tobe, S.S., 2013b. Sequencing and validation of housekeeping genes for
quantitative real-time PCR during the gonadotrophic cycle of Diploptera punctata. BMC Res. Notes. 6, 237.
McBain, C.J., Mayer, M.L., 1994. N-methyl-D-aspartic acid receptor structure and function. Physiol. Rev. 74,
723-760.
Monyer, H., Sprengel, R., Schoepfer, R., Herb, A., Higuchi, M., Lomeli, H., Burnashev, N., Sakmann, B.,
Seeburg, P.H., 1992. Heteromeric NMDA receptors: molecular and functional distinction of subtypes. Science
256, 1217-1221.
Mussig, L., Richlitzki, A., Rossler, R., Eisenhardt, D., Menzel, R., Leboulle, G., 2010. Acute disruption of the
NMDA receptor subunit NR1 in the honeybee brain selectively impairs memory formation. J. Neurosci. 30,
7817-7825.
Newcomer, J.W., Krystal, J.H., 2001. NMDA receptor regulation of memory and behavior in humans.
Hippocampus 11, 529-542.
Pszczolkowski, M.A., Lee, W.S., Liu, H.P., Chiang, A.S., 1999. Glutamate-induced rise in cytosolic calcium
concentration stimulates in vitro rates of juvenile hormone biosynthesis in corpus allatum of Diploptera
punctata. Mol. Cell. Endocrinol. 158, 163-171.
Rankin, S.M., Stay, B., 1984. The changing effect of the ovary on rates of juvenile hormone synthesis in
Diploptera punctata. Gen. Comp. Endocrinol. 54, 382-388.
Rawls, S.M., Thomas, T., Adeola, M., Patil, T., Raymondi, N., Poles, A., Loo, M., Raffa, R.B., 2009.
Topiramate antagonizes NMDA- and AMPA-induced seizure-like activity in planarians. Pharmacol. Biochem.
Be. 93, 363-367.
Santillo, A., Falvo, S., Chieffi, P., Burrone, L., Chieffi Baccari, G., Longobardi, S., Di Fiore, M.M., 2014. D-
aspartate affects NMDA receptor-extracellular signal-regulated kinase pathway and upregulates androgen
receptor expression in the rat testis. Theriogenology 81, 744-751.
Sircar, R., Rappaport, M., Nichtenhauser, R., Zukin, S.R., 1987. The novel anticonvulsant Mk-801 - a potent
and specific ligand of the brain phencyclidine sigma-receptor. Brain Res. 435, 235-240.
Sprengel, R., Suchanek, B., Amico, C., Brusa, R., Burnashev, N., Rozov, A., Hvalby, O., Jensen, V., Paulsen,
O., Andersen, P., Kim, J.J., Thompson, R.F., Sun, W., Webster, L.C., Grant, S.G., Eilers, J., Konnerth, A., Li,
J., McNamara, J.O., Seeburg, P.H., 1998. Importance of the intracellular domain of NR2 subunits for NMDA
receptor function in vivo. Cell 92, 279-289.
Stay, B., Tobe, S.S., 1978. Control of juvenile hormone biosynthesis during the reproductive cycle of a
viviparous cockroach. II. Effects of unilateral allatectomy, implantation of supernumerary corpora allata, and
ovariectomy. Gen. Comp. Endocrinol. 34, 276-286.
Stay, B., Tobe, S.S., 2007. The role of allatostatins in juvenile hormone synthesis in insects and crustaceans.
Annu. Rev. Entomol. 52, 277-299.
Stern-Bach, Y., Bettler, B., Hartley, M., Sheppard, P.O., O'Hara, P.J., Heinemann, S.F., 1994. Agonist
selectivity of glutamate receptors is specified by two domains structurally related to bacterial amino acid-
binding proteins. Neuron 13, 1345-1357.
Sydow, S., Kopke, A.K.E., Blank, T., Spiess, J., 1996. Overexpression of a functional NMDA receptor subunit
(NMDAR1) in baculovirus-infected Trichoplusia ni insect cells. Mol. Brain Res. 41, 228-240.
Tamura, K., Stecher, G., Peterson, D., Filipski, A., Kumar, S., 2013. MEGA6: Molecular Evolutionary
Genetics Analysis version 6.0. Mol. Biol. Evol. 30, 2725-2729.

168
Tang, Y.P., Shimizu, E., Dube, G.R., Rampon, C., Kerchner, G.A., Zhuo, M., Liu, G., Tsien, J.Z., 1999.
Genetic enhancement of learning and memory in mice. Nature 401, 63-69.
Thompson, C.S., Yagi, K.J., Chen, Z.F., Tobe, S.S., 1990. The effects of octopamine on juvenile hormone
biosynthesis, electrophysiology, and cAMP content of the corpora allata of the cockroach Diploptera punctata.
J. Comp. Physiol. B 160, 241-249.
Troncoso, J., Maldonado, H., 2002. Two related forms of memory in the crab Chasmagnathus are differentially
affected by NMDA receptor antagonists. Pharmacol. Biochem. Be. 72, 251-265.
Ultsch, A., Schuster, C.M., Laube, B., Betz, H., Schmitt, B., 1993. Glutamate receptors of Drosophila
melanogaster. Primary structure of a putative NMDA receptor protein expressed in the head of the adult fly.
Febs Lett 324, 171-177.
Van der Staay, F.J., Rutten, K., Erb, C., Blokland, A., 2011. Effects of the cognition impairer MK-801 on
learning and memory in mice and rats. Behav. Brain Res. 220, 215-229.
Vandesompele, J., De Preter, K., Pattyn, F., Poppe, B., Van Roy, N., De Paepe, A., Speleman, F., 2002.
Accurate normalization of real-time quantitative RT-PCR data by geometric averaging of multiple internal
control genes. Genome Biol. 3, 1-12.
Whelan, S., Goldman, N., 2001. A general empirical model of protein evolution derived from multiple protein
families using a maximum-likelihood approach. Mol. Biol. Evol. 18, 691-699.
Wu, C.L., Xia, S., Fu, T.F., Wang, H., Chen, Y.H., Leong, D., Chiang, A.S., Tully, T., 2007. Specific
requirement of NMDA receptors for long-term memory consolidation in Drosophila ellipsoid body. Nat.
Neurosci. 10, 1578-1586.
Xia, S., Miyashita, T., Fu, T.F., Lin, W.Y., Wu, C.L., Pyzocha, L., Lin, I.R., Saitoe, M., Tully, T., Chiang,
A.S., 2005. NMDA receptors mediate olfactory learning and memory in Drosophila. Curr. Biol. 15, 603-615.
Yang, Z., 1994. Maximum likelihood phylogenetic estimation from DNA sequences with variable rates over
sites: approximate methods. J. Mol. Evol. 39, 306-314.
Zannat, M.T., Locatelli, F., Rybak, J., Menzel, R., Leboulle, G., 2006. Identification and localisation of the
NR1 sub-unit homologue of the NMDA glutamate receptor in the honeybee brain. Neurosci. Lett. 398, 274-
279.

169
Chapter 5
General Discussion
1. Function of JH in reproduction
The original function of JH in insects was considered to be a regulator of reproduction (Tobe and
Bendena, 1999). Several different types of hormones are involved in the endocrine control of
female reproduction, of which JHs and ecdysteroids are most studied. In most hemimetabolous
insects, the major events of reproduction are controlled by JH, while in higher insects, both JH
and ecdysteroids were involved (Raikhel et al., 2005). In D. punctata, allatectomy or
implantation of CA with ovaries into male animals showed that JH stimulates Vg synthesis and
Vg uptake (Mundall et al., 1979; Stay and Tobe, 1977). Moreover, in several other cockroaches
species, treatment with JH or JH analogs achieved the same result (Comas et al., 1999; Don-
Wheeler and Engelmann, 1997; Weaver and Edwards, 1990). In this thesis, the function of JH in
regulating female reproduction was further investigated by repressing JH biosynthesis using
RNAi. As expected, the lowered JH biosynthesis on day 4 in the CA resulted in a significant
reduction of Vg mRNA levels, uptake of Vn into the oocytes, and in blocking patency (Chapter
2). The increase of JH biosynthesis on day 3 and 4 is essential for the development of oocytes.
On the other hand, the decrease in JH production starting on day 5 is required to complete the
regular growth of oocytes. An abnormal high titre of JH after day 5 resulted in a failure in
oviposition (Hult et al, data unpublished). It is obvious that JH plays critical functions in
regulating cockroach female reproduction and therefore must be tightly regulated.

170
2. Evolution of the JH biosynthetic pathway
2.1 The loss of sterols biosynthesis in arthropods
In most eukaryotic animal and plant systems, cholesterol and related sterols, which are essential
component of cell membranes, are biosynthesized through the mevalonate pathway. Arthropods,
however, do not appear to synthesize cholesterol, but rather produce sesquiterpenoids. The final
product of the mevalonate pathway is JH, rather than cholesterol, as a consequence of the
absence of genes encoding squalene synthase and other subsequent enzymes to produce sterols
(Belles et al., 2005). The biosynthesis of JH and cholesterol share the mevalonate pathway until
the production of FPP. In Arthropoda, FPP is converted to FOL and thereafter to FA (Figure
1.1), while it is catalyzed to squalene in animals producing cholesterol. In some plants, on the
other hand, cholesterol and JH III and their related compounds are synthesized, suggesting that
both the JH and cholesterol biosynthetic pathway are present in plant system (Yang et al., 2013).
Tobe and Bendena (1999) suggest that the ability to biosynthesize cholesterol was maintained in
the ancestor of the arthropod/annelid lineage, but lost in the arthropod line, probably because of
the abundance of cholesterol in the diet. The biosynthesis of JH in some plants is the result of
convergence, which is unrelated to its occurrence in arthropods. Nevertheless, the reason for
losing the ability to biosynthesize cholesterol de novo in arthropods remains unknown.
2.2 Enzymes in the JH biosynthetic pathway controlling the end product of the JH
biosynthetic pathway
As shown in Figure 1.1, various JHs are produced in insects. These JHs shares the main carbon
skeleton, but with different methyl/ethyl substitutions and carbon-sites for epoxidation. Most
ethyl substituted JH III products occur in the order of the Lepidoptera (Details seen Chapter 1

171
section 1). The reason for the various end products of the JH biosynthetic pathway in
Lepidoptera can be found in the catalytic selectivity of enzymes in the pathway. Unlike in other
insects, both acetyl-CoA and propionyl-CoA are involved in the early steps of JH biosynthesis to
produce ethyl-branched JHs in M. sexta (Brindle et al., 1987; Brindle et al., 1988). The
branched-chain amino acid transaminase, which generates propionyl-CoA by the metabolism of
leucine and valine, is found only in the CA of Lepidoptera (Brindle et al., 1988). In addition,
enzymes in the JH biosynthetic pathway have evolved to utilize different substrates for JH
biosynthesis. For instance, IPPI in M. sexta and C. fumiferana contains an active site cavity,
which allows it to bind to larger substrates (ethyl-substituted IPP) and to stabilize the high-
energy intermediates to form ethyl- substituted DMPP (Sen et al., 2012). FPPS in M. sexta is also
adapted to catalyze both ethyl- and methyl-substituted DMPP and GPP, and the selectivity of the
enzyme inclines to the ethyl-substituted substrate (Sen et al., 1996; Sen et al., 2006).
As noted in Chapter 1, section 1.2.13, CYP15 genes have been characterized to encode
cytochrome P450 (CYP) enzymes that epoxidize MF to JH in Orthoptera, Dictyoptera and
Coleoptera, or FA to JHA in Lepidotera. However, in higher Diptera that produce bisepoxy
forms of JH III, no CYP15 gene has been found (Daimon and Shinoda, 2013). Different CYP
enzymes that could catalyze the bisepoxidization of MF may be present in the higher Diptera to
compensate the loss of CYP15, resulting in the production of JHB3. However, in P. stali that
produces JHSB3, a CYP15 ortholog was identified. The function of this CYP15 ortholog has not
yet been determined. Whether there are other epoxidases involved in the synthesis of JHSB3
remains unclear. In addition, in the crustacean D. pulex, no CYP15 ortholog was found in the
genome. The lack of CYP15 genes may be the reason for crustaceans using MF as their JH-like
bioactive compound.

172
The catalytic selectivity of the enzymes in the JH biosynthetic pathway determines not only the
structure of the JH products, but also the order of steps in the JH biosynthetic pathway. As
shown in Figure 1.2, the last two steps (methylation and epoxidation) of the JH biosynthetic
pathways differ between Lepidoptera and other insects. Two different epoxidase enzymes,
CYP15A1 and CYP15C1 are involved in the last steps of JH biosynthesis in different insect
orders. The order of the methylation and epoxidation steps appears to be controlled by the
selectivity of the epoxidase. CYP15C1, the functional epoxidase in Lepidoptera, specifically
converts FA to JHA (Daimon et al., 2012). Therefore, epoxidation occurs before methylation in
Lepidoptera. However, in other insect orders, CYP15A1 has been characterized as the active
epoxidase. The substrate selectivity of CYP15A1 to MF determines that methylation precedes
epoxidation (Helvig et al., 2004).
D. punctata, as a more basal species in the Insecta, did not appear to undergo any significant
modification of the enzymes in the JH biosynthetic pathway. Thus, only JH III has been
identified as the JH biosynthetic product of CA.
3. Regulation of JH biosynthesis
3.1 Enzymes in the biosynthetic pathway
Regulation of JH biosynthesis is a complicated process involving multiple regulators. The best-
characterized regulators are the neuropeptides, including ATs and ASTs. Enzymes in the JH
biosynthetic pathway were also shown to regulate JH biosynthesis (Couillaud and Feyereisen,
1991; Feyereisen and Farnsworth, 1987; Kinjoh et al., 2007; Nouzova et al., 2011). Some
enzymes have been proposed to catalyze rate-limiting steps in the pathway (e.g. FALD in A.
aegypti (Rivera-Perez et al., 2013); JHAMT in B. mori (Shinoda and Itoyama, 2003)). In this

173
thesis, the transcript levels of genes in the JH biosynthetic pathway were determined in relation
to the JH biosynthesis in D. punctata. Similar to other species, the expression of most genes is
well-correlated with the JH biosynthetic activity, suggesting that JH biosynthesis is in part
controlled by the transcription of enzymes. However, no rate-limiting enzyme could be identified
in D. punctata. My study has also shown that precursor supply mediates JH biosynthesis. In
addition, silencing individual genes encoding the JH biosynthetic enzymes resulted in a
significant decrease in the transcript levels of other genes. Therefore, a feedback among genes in
JH biosynthetic pathway was proposed: the expression of enzymes in the pathway is correlated
with other enzymes to obtain the balance between enzyme activity and JH precursor supply.
Further experiments to determine the amount of JH precursors in the CA after silencing
individual genes would provide evidence to support the hypothesis.
3.2 Neuropeptides
In cockroaches, ASTs compose a family of 13 to 14 neuropeptides with the conserved C-
terminal sequence (Y/FXFGL/I-NH2) and variable N-terminal sequences. All the peptides are
encoded by a single gene. Evolutionary analysis of this gene indicates that the AST sequences
were generated by a gene duplication event before the species diverged (Belles et al., 1999).
Structure-activity studies show that all ASTs are able to inhibit JH biosynthesis effectively, albeit
with different potencies (Tobe et al., 2000). This result suggests that AstRs in cockroaches are
relatively unselective, binding ASTs with different structures. In my thesis, the selectivity of
DpAstR to its ligand was determined by the activation of DpAstR expressed in CHO cells using
13 DpASTs and AST analogs. All ASTs and AST analogs were able to activate DpAstR, and the
selectivity of DpAstR to DpASTs was shown to be structure-dependent (Table 3.1 and Table 6.1).

174
The core sequence (Y/FXFGL/I-NH2) is essential for binding, while the N-terminal region
regulates binding affinity.
In general, the ability of AST to bind AstR correlates well with its potency to inhibit JH
production by the CA (Table 3.1). AST is a pleiotropic neuropeptide, mediating different
physiological effects, such as midgut enzyme activity and hindgut muscle contraction. However,
in these functions, binding of AST to its receptor does not seem to correlate with its activity
(Lange et al., 1995). In addition, the AstR identified in the CA of D. punctata was not found to
be expressed in the midgut, where the precursor of its ligand AST was highly expressed. My
study supports the hypothesis proposed by Tobe and Bendena (1999) that there are different AST
receptor subtypes in different tissues. Unfortunately, no second AstR could be identified in D.
punctata.
As mentioned before (Chapter 1, section 3.1), there are different families of allatostatic
neuropeptides. The FGLa/ASTs which inhibits JH biosynthesis in cockroaches, do not show the
allato-inhibitory effect on the CA of Lepidoptera or Diptera. Instead, the PISCF/AST family is
involved to regulate JH biosynthesis in these insect orders. Unlike in cockroaches where 14
FGLa/ASTs with very high sequence homology are encoded by a single gene, the PISCF/AST
precursor gene only encodes one AST peptide in Lepidoptera and Diptera (Kramer et al., 1991;
Li et al., 2006). In addition to PISCF/AST, another type of neuropeptide, termed allatotropin
(AT), which stimulates JH biosynthesis in the CA, was also identified in Lepidoptera and Diptera.
3.3 Second messenger (Ca2+)
Calcium, an intracellular second messenger, is a critical factor in controlling JH biosynthesis.
However, the role of calcium in the JH biosynthesis seems to vary depending on species and

175
stage (Allen et al., 1992; Dale and Tobe, 1988; Kikukawa et al., 1987). To make it more
complicated, calcium was determined to act as a second messenger in the signal pathway of AST
action to inhibit JH biosynthesis (Chapter 3). Calcium can also act as the second messenger of
Manse-AT to enhance CA activity (Horodyski et al., 2011). In this thesis, the function of
NMDAR as an ionotropic calcium channel was investigated. A previous study demonstrated that
NMDA stimulates JH biosynthesis in the CA through the influx of Ca2+ into the CA cells
(Chiang et al., 2002). However, our study, using RNAi and in vivo treatment with NMDAR
antagonists, revealed that NMDAR does not appear to regulate JH biosynthesis (Chapter 4).
Clearly, a complex mechanism is involved in the regulation of JH biosynthesis by calcium.
4. The value of my study in insect control
As an essential hormone controlling many physiological processes in insects, JH production is
tightly regulated to allow normal development. This characteristic makes JH a potential target for
insect control. Thousands of JH analogs have been synthesized and bioassayed to discover
potential insecticides (Goodman and Cusson, 2012). In addition to JH analogs, ASTs which
inhibit JH biosynthesis at low doses, have been employed in the design of potential insect growth
regulators (IGRs). Previous studies have determined the distribution, activity and structure-
activity relationship of the thirteen ASTs in D. punctata, which has permitted the development of
analogs of the naturally occurring ASTs (Donly et al., 1993; Rankin and Stay, 1987; Tobe et al.,
2000). The development of AST analogs as potential insecticides need to overcome two major
problems: (1) maintaining the high allato-inhibitory activity being a shorter peptide; (2) having a
high resistance to the peptidases in the CA and hemolymph (Marchal et al., 2013). In this thesis,
two series of AST analogs were designed and synthesized based on the previous rules. Some
AST analogs not only exhibited similar potencies in inhibiting JH biosynthesis in vitro to the

176
naturally occurring AST (Table 6.2), but were also able to regulate JH biosynthesis and oocyte
growth in an in vivo bioassay (Fig. 6.1). Preliminary degradation results show that the AST
analogs have higher resistance to peptidases in hemolymph than natural AST. These AST
analogs can be considered as potential IGRs. Further modification of these AST analgos is now
required to obtain higher allato-inhibitory activity and higher resistance to peptidases in the
cockroach.
5. Future perspectives
This thesis aimed to characterize the JH biosynthetic pathway and determine factors regulating
JH biosynthesis in D. punctata. Genes in the JH biosynthetic pathways were identified,
functionally characterized, and regulatory factors including AST and NMDAR were studied.
However, many questions still remain unanswered. The exact target of AST action remains
unknown. The role of AST in regulating JH biosynthesis is well-established, while the roles of
other regulatory factors including second messengers, JH feedback and ecdysteroids remain
unclear. In addition to the regulation of JH biosynthesis, the JH action and cross-talking with
other hormone also require further investigate.
D. punctata is a great model in studying the endocrinology of arthropods. In this animal the JH
titre is relatively high and stable, only JH III is is synthesized, and reproductive events are well-
correlated with JH biosynthesis. The genome of D. punctata is currently being assembled which
will allow high-throughput and more in depth studies aimed at identifying the regulatory
mechanisms targeting JH biosynthesis. A first study is currently ongoing involving differential
transcriptomics of CA with low versus high JH biosynthetic activity.

177
References
Allen, C.U., Janzen, W.P., Granger, N.A., 1992. Manipulation of intracellular calcium affects in vitro juvenile
hormone synthesis by larval corpora allata of Manduca sexta. Mol Cell Endocrinol 84, 227-241.
Belles, X., Graham, L.A., Bendena, W.G., Ding, Q., Edwards, J.P., Weaver, R.J., Tobe, S.S., 1999. The
molecular evolution of the allatostatin precursor in cockroaches. Peptides 20, 11-22.
Belles, X., Martin, D., Piulachs, M.D., 2005. The mevalonate pathway and the synthesis of juvenile hormone
in insects. Annu Rev Entomol 50, 181-199.
Brindle, P.A., Baker, F.C., Tsai, L.W., Reuter, C.C., Schooley, D.A., 1987. Sources of propionate for the
biogenesis of ethyl-branched insect juvenile hormones: Role of isoleucine and valine. Proc Natl Acad Sci U S
A 84, 7906-7910.
Brindle, P.A., Schooley, D.A., Tsai, L.W., Baker, F.C., 1988. Comparative metabolism of branched-chain
amino acids to precursors of juvenile hormone biogenesis in corpora allata of lepidopterous versus
nonlepidopterous insects. J Biol Chem 263, 10653-10657.
Chiang, A.S., Lin, W.Y., Liu, H.P., Pszczolkowski, M.A., Fu, T.F., Chiu, S.L., Holbrook, G.L., 2002. Insect
NMDA receptors mediate juvenile hormone biosynthesis. Proc Natl Acad Sci U S A 99, 37-42.
Comas, D., Piulachs, M.D., Belles, X., 1999. Fast induction of vitellogenin gene expression by juvenile
hormone III in the cockroach Blattella germanica (L.) (Dictyoptera, Blattellidae). Insect Biochem Mol Biol 29,
821-827.
Couillaud, F., Feyereisen, R., 1991. Assay of HMG-CoA synthase in Diploptera punctata corpora allata. Insect
Biochem 21, 131-135.
Daimon, T., Kozaki, T., Niwa, R., Kobayashi, I., Furuta, K., Namiki, T., Uchino, K., Banno, Y., Katsuma, S.,
Tamura, T., Mita, K., Sezutsu, H., Nakayama, M., Itoyama, K., Shimada, T., Shinoda, T., 2012. Precocious
metamorphosis in the juvenile hormone-deficient mutant of the silkworm, Bombyx mori. PLoS Genet 8,
e1002486.
Daimon, T., Shinoda, T., 2013. Function, diversity, and application of insect juvenile hormone epoxidases
(CYP15). Biotechnol Appl Biochem 60, 82-91.
Dale, J.F., Tobe, S.S., 1988. Differences in the stimulation by calcium ionophore of juvenile hormone III
release from corpora allata of solitarious and gregarious Locusta migratoria. Experientia 44, 240-242.
Don-Wheeler, G., Engelmann, F., 1997. The biosynthesis and processing of vitellogenin in the fat bodies of
females and males of the cockroach Leucophaea maderae. Insect Biochem Mol Biol 27, 901-918.
Donly, B.C., Ding, Q., Tobe, S.S., Bendena, W.G., 1993. Molecular cloning of the gene for the allatostatin
family of neuropeptides from the cockroach Diploptera punctata. Proc Natl Acad Sci U S A 90, 8807-8811.
Feyereisen, R., Farnsworth, D.E., 1987. Characterization and regulation of HMG-CoA Reductase during a
cycle of Juvenile Hormone synthesis. Mol Cell Endocrinol 53, 227-238.
Goodman, W.G., Cusson, M., 2012. The juvenile hormones, in: Gilbert, L.I. (Ed.), Insect Endocrinology.
Academic Press, London, p. 55.
Helvig, C., Koener, J.F., Unnithan, G.C., Feyereisen, R., 2004. CYP15A1, the cytochrome P450 that catalyzes
epoxidation of methyl farnesoate to juvenile hormone III in cockroach corpora allata. Proc Natl Acad Sci U S
A 101, 4024-4029.
Horodyski, F.M., Verlinden, H., Filkin, N., Vandersmissen, H.P., Fleury, C., Reynolds, S.E., Kai, Z.P., Broeck,
J.V., 2011. Isolation and functional characterization of an allatotropin receptor from Manduca sexta. Insect Biochem Mol Biol 41, 804-814.

178
Kikukawa, S., Tobe, S.S., Solowiej, S., Rankin, S.M., Stay, B., 1987. Calcium as a regulator of juvenile
hormone biosynthesis and release in the cockroach Diploptera punctata. Insect Biochem 17, 179-187.
Kinjoh, T., Kaneko, Y., Itoyama, K., Mita, K., Hiruma, K., Shinoda, T., 2007. Control of juvenile hormone
biosynthesis in Bombyx mori: Cloning of the enzymes in the mevalonate pathway and assessment of their
developmental expression in the corpora allata. Insect Biochem Mol Biol 37, 808-818.
Kramer, S.J., Toschi, A., Miller, C.A., Kataoka, H., Quistad, G.B., Li, J.P., Carney, R.L., Schooley, D.A., 1991.
Identification of an allatostatin from the tobacco hornworm Manduca sexta. Proc Natl Acad Sci U S A 88,
9458-9462.
Lange, A.B., Bendena, W.G., Tobe, S.S., 1995. The effect of the thirteen Dip-Allatostatins on myogenic and
induced contractions of the cockroach (Diploptera Punctanta) hindgut. J Insect Physiol 41, 581-588.
Li, Y., Hernandez-Martinez, S., Fernandez, F., Mayoral, J.G., Topalis, P., Priestap, H., Perez, M., Navare, A.,
Noriega, F.G., 2006. Biochemical, molecular, and functional characterization of PISCF-allatostatin, a regulator
of juvenile hormone biosynthesis in the mosquito Aedes aegypti. J Biol Chem 281, 34048-34055.
Marchal, E., Hult, E.F., Huang, J., Stay, B., Tobe, S.S., 2013. Diploptera punctata as a model for studying the
endocrinology of arthropod reproduction and development. Gen Comp Endocr 188, 85-93.
Mundall, E.C., Tobe, S.S., Stay, B., 1979. Induction of vitellogenin and growth of implanted oocytes in male
cockroaches. Nature 282, 97-98.
Nouzova, M., Edwards, M.J., Mayoral, J.G., Noriega, F.G., 2011. A coordinated expression of biosynthetic
enzymes controls the flux of juvenile hormone precursors in the corpora allata of mosquitoes. Insect Biochem
Mol Biol 41, 660-669.
Raikhel, A.S., Brown, M.R., Belles, X., 2005. Hormonal control of reproductive processes, in: Gilbert, L.I.,
Iatrou, K., Gill, S.S. (Eds.), Comprehensive molecular insect science. Elsevier, Oxford, pp. 433-491.
Rankin, S.M., Stay, B., 1987. Distribution of allatostatin in the adult cockroach, Diploptera punctata and
effects on corpora allata in vitro. Journal of Insect Physiology 33, 551-558.
Rivera-Perez, C., Nouzova, M., Clifton, M.E., Garcia, E.M., LeBlanc, E., Noriega, F.G., 2013. Aldehyde
dehydrogenase 3 converts farnesal into farnesoic acid in the corpora allata of mosquitoes. Insect Biochem Mol
Biol 43, 675-682.
Sen, S.E., Ewing, G.J., Thurston, N., 1996. Characterization of lepidopteran prenyltransferase in Manduca sexta corpora allata. . Arch Insect Biochem Physiol 32, 315–332.
Sen, S.E., Hitchcock, J.R., Jordan, J.L., Richard, T., 2006. Juvenile hormone biosynthesis in M. sexta:
substrate specificity of insect prenyltransferase utilizing homologous diphosphate analogs. Insect Biochem
Mol Biol 36, 827-834.
Sen, S.E., Tomasello, A., Grasso, M., Denton, R., Macor, J., Beliveau, C., Cusson, M., Crowell, D.N., 2012.
Cloning, expression and characterization of lepidopteran isopentenyl diphosphate isomerase. Insect Biochem
Mol Biol 42, 739-750.
Shinoda, T., Itoyama, K., 2003. Juvenile hormone acid methyltransferase: a key regulatory enzyme for insect
metamorphosis. Proc Natl Acad Sci U S A 100, 11986-11991.
Stay, B., Tobe, S.S., 1977. Control of juvenile hormone biosynthesis during the reproductive cycle of a
viviparous cockroach 1. Activation and inhibition of corpora allata. Gen Comp Endocrinol 33, 531-540.
Tobe, S.S., Bendena, W.G., 1999. The regulation of juvenile hormone production in arthropods. Functional
and evolutionary perspectives. Ann N Y Acad Sci 897, 300-310.
Tobe, S.S., Zhang, J.R., Bowser, P.R.F., Donly, B.C., Bendena, W.G., 2000. Biological activities of the
allatostatin family of peptides in the cockroach, Diploptera punctata, and potential interactions with receptors.
Journal of Insect Physiology 46, 231-242.

179
Weaver, R.J., Edwards, J.P., 1990. The role of the corpora allata and associated nerves in the regulation of
ovarian cycles in the oviparous cockroach Periplaneta americana. J Insect Physiol 36, 51-59.
Yang, H., Kim, H.S., Jeong, E.J., Khiev, P., Chin, Y.W., Sung, S.H., 2013. Plant-derived juvenile hormone III
analogues and other sesquiterpenes from the stem bark of Cananga latifolia. Phytochemistry 94, 277-283.

180
Chapter 6
Appendices
ASTs which inhibit JH biosynthesis at low doses, have been employed in the design of potential
insecticides. Furthermore, molecular cloning, structure determination, distribution and activity of
the thirteen ASTs in D. punctata, have permitted the development of analogs of the naturally
occurring ASTs (Donly et al., 1993; Rankin and Stay, 1987; Tobe et al., 2000). Although ASTs
inhibit JH biosynthesis effectively in vitro, they have some shortcomings as potential pesticides.
Firstly, the longer the peptide, the more unwanted side-products that are produced during
chemical synthesis. For ASTs, even the shortest AST peptide has 6 amino acids (Tobe et al.,
2000). Secondly, ASTs are susceptible to metabolic inactivation by peptidases in hemolymph
and midgut (Garside et al., 1997a, b). For example the half-life of Dippu-AST 5 (AST5: Asp-
Arg-Leu-Tyr-Ser-Phe-Gly-Leu-amide) is less than 1 hour when incubated with membrane
preparations of brain and midgut (Garside et al., 1997b). Therefore, much effort has been
expended to overcome this problem and create better AST pseudopeptides. In this thesis, two
series of AST anaologs were designed and their activities in regulating JH biosynthesis in vitro
and in vivo in D. punctata were shown in this chapter.

181
Table 6.1. Structure of AST analogs
Analogs Structure
W201
Gly
O
Phe Gly Leu NH2
W202 Gly
O
Phe Gly Leu NH2
W203
Gly
O
Phe Gly Leu NH2
W204 Gly
O
Phe Gly Leu NH2
W205
Gly
O
Phe Gly Leu NH2
W206
W70
Gly
O
Phe Gly Leu NH2
Br
W71
Gly
O
Phe Gly Leu NH2
Br
K15
Gly
O
Phe Gly Leu NH2
Br
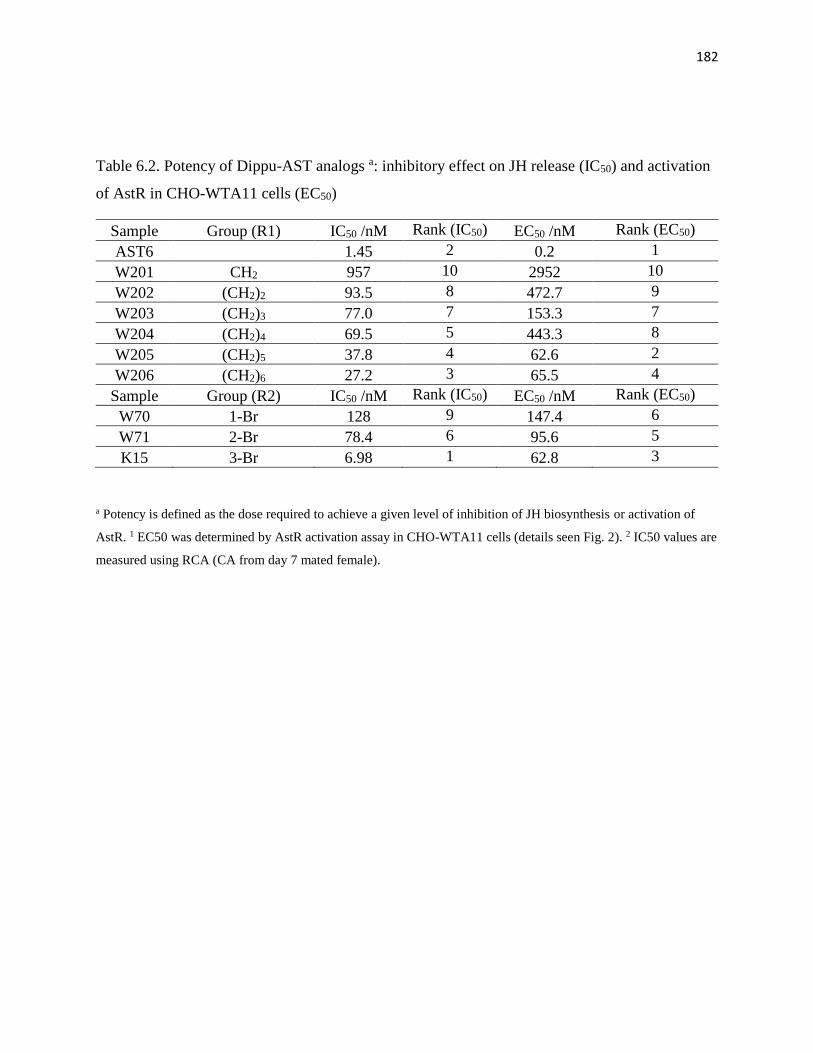
182
Table 6.2. Potency of Dippu-AST analogs a: inhibitory effect on JH release (IC50) and activation
of AstR in CHO-WTA11 cells (EC50)
Sample Group (R1) IC50 /nM Rank (IC50) EC50 /nM Rank (EC50)
AST6
1.45 2 0.2 1
W201 CH2 957 10 2952 10
W202 (CH2)2 93.5 8 472.7 9
W203 (CH2)3 77.0 7 153.3 7
W204 (CH2)4 69.5 5 443.3 8
W205 (CH2)5 37.8 4 62.6 2
W206 (CH2)6 27.2 3 65.5 4
Sample Group (R2) IC50 /nM Rank (IC50) EC50 /nM Rank (EC50)
W70 1-Br 128 9 147.4 6
W71 2-Br 78.4 6 95.6 5
K15 3-Br 6.98 1 62.8 3
a Potency is defined as the dose required to achieve a given level of inhibition of JH biosynthesis or activation of
AstR. 1 EC50 was determined by AstR activation assay in CHO-WTA11 cells (details seen Fig. 2). 2 IC50 values are
measured using RCA (CA from day 7 mated female).

183
Fig. 6.1. The effect of topical application of K15 and W206 on JH biosynthesis (A) and
oocyte growth (B). AST analogs were diluted in 20% DMSO and 80% acetone to a
concentration of 10-3 M, and applied 5 µl to the dorsal abdomen of each D. punctata female on
day 0. The effect of AST analogs on JH biosynthesis and basal oocyte growth was determined
from day 4 to day 8.

184
References
Donly, B.C., Ding, Q., Tobe, S.S., Bendena, W.G., 1993. Molecular cloning of the gene for the allatostatin
family of neuropeptides from the cockroach Diploptera punctata. Proceedings of the National Academy of
Sciences of the United States of America 90, 8807-8811.
Garside, C.S., Hayes, T.K., Tobe, S.S., 1997a. Degradation of dip-allatostatins by hemolymph from the
cockroach, Diploptera punctata. Peptides 18, 17-25.
Garside, C.S., Hayes, T.K., Tobe, S.S., 1997b. Inactivation of dip-allatostatin 5 by membrane preparations
from the cockroach Diploptera punctata. General and comparative endocrinology 108, 258-270.
Rankin, S.M., Stay, B., 1987. Distribution of allatostatin in the adult cockroach, Diploptera punctata and
effects on corpora allata in vitro. Journal of Insect Physiology 33, 551-558.
Tobe, S.S., Zhang, J.R., Bowser, P.R.F., Donly, B.C., Bendena, W.G., 2000. Biological activities of the
allatostatin family of peptides in the cockroach, Diploptera punctata, and potential interactions with receptors.
Journal of Insect Physiology 46, 231-242.

185
Copyright Acknowledgements
The author would like to thank:
Elisabeth Marchal, Ekaterina F. Hult, and Stephen S. Tobe for permitting to use the following
material: (1) Diploptera punctata as a model for studying the endocrinology of arthropod
reproduction and development. Gen Comp Endocr , 2013, 188, 85-93. (2) Characterization of the
Juvenile Hormone pathway in the viviparous cockroach, Diploptera punctata. PloS one,
accepted. (3) Identification and characterization of the NMDA receptor and its role in regulating
reproduction in the cockroach, Diploptera punctata, 2014, paper submitted to ‘Journal of
experimental biology’
Elisabeth Marchal, Ekaterina F. Hult, Sven Zels, Jozef Vanden Broeck, and Stephen S. Tobe for
permit to use the following material: Mode of action of allatostatins in the regulation of juvenile
hormone biosynthesis in the cockroach, Diploptera punctata, Insect Biochem Mol Biol, 2014, 54,
61-68.
And Journal: ‘Insect biochemistry and molecular biology’ for allowing the author to reprint the
article in the institutions open scholarly website (institutional repository).