kiaulių miostatino geno polimorfizmas ir jo ryšys su …gyvŪnŲ veisimo ir genetikos katedra k....
TRANSCRIPT

1
LIETUVOS VETERINARIJOS AKADEMIJA GYVULININKYSTöS TECHNOLOGIJOS
FAKULTETAS GYVŪNŲ VEISIMO IR GENETIKOS KATEDRA
K. Janušausko gyvūnų genetikos laboratorija
Vaida Palačenkait÷
Kiaulių miostatino geno polimorfizmas ir jo ryšys su m÷sos kokybiniais bei kiekybiniais rodikliais
Magistro darbas
Darbo vadovas: e.prof.p. Ilona Miceikien÷
Kaunas, 2008

2
Magistro darbas atliktas 2007 – 2008 metais Lietuvos veterinarijos
akademijoje, Gyvūnų veisimo ir genetikos katedroje, K.Janušausko gyvūnų
genetikos laboratorijoje.
Magistro darbą paruoš÷: Vaida Palačenkait÷ (v., pavard÷) (parašas)
Magistro darbo vadovas: e.prof. p. dr. I.Miceikien÷
(LVA, Gyvūnų veisimo ir genetikos katedra)
(parašas)
Recenzentas:
(parašas)

3
TURINYS
Santrumpų sąrašas.......................................................................................................................4
Įvadas..........................................................................................................................................6
1. LITERATŪROS APŽVALGA...............................................................................................7
1.1. Genai, įtakojantys kiaulienos kiekį ir kokybę......................................................................7
1.2. Miogenino (MYOG) genas..................................................................................................7
1.3. Leptino (LEP) genas............................................................................................................8
1.4. Augimo hormono (GH) genas..............................................................................................8
1.5. Rapole (RN) genas...............................................................................................................9
1.6. Halotano (HAL) genas.........................................................................................................9
1.7. Melanokortino 4 receptoriaus (MC4R) genas....................................................................11
1.8. Kiaulių selekcija genetinių žymenų pagalba......................................................................11
2. TYRIMŲ METODAI IR MEDŽIAGA................................................................................13
2.1. Tyrimų medžiaga ir metodai..............................................................................................13
2.2. Kiaulių fenotipinių požymių duomenų baz÷s....................................................................14
2.3.DNR skyrimas iš plauko svogūn÷lio ląstelių......................................................................14
2.4. DNR švarumo ir koncentracijos nustatymas......................................................................14
2.5. Polimerazin÷ grandinin÷ reakcija.......................................................................................15
2.5.1. Kiaulių miostatino geno MYF5 (PGR-RFIP) tyrimo metodika………………………..15
2.6. Statistin÷ duomenų analiz÷.................................................................................................16
3. TYRIMŲ REZULTATAI IR APTARIMAS........................................................................17
3.1. Kiaulių miostatino geno polimorfizmo ištyrimas..............................................................17
4. IŠVADOS.............................................................................................................................22
5. SUMMARY..........................................................................................................................23
6. LITERATŪROS SĄRAŠAS................................................................................................24

4
SANTRUMPŲ SĄRAŠAS
ATP – adenozintrifosfatas
bp – bazių pora
CRC – halotono genas
DFD – tamsios, kietos, sausos m÷sos sindromas (dark, firm, dry)
DNR – dezoksiribonukleinin÷ rūgštis
DTT - dithiotreitolis
ESR – estrogenų receptorius
GH – augimo hormono genas
GHR – augimo hormono receptorius
GHRH – augimo hormoną atpalaiduojančio receptoriaus genas
GHRHR – augimo hormoną atpalaiduojanč receptorius
H-FABP –riebiąsias rūgštis jungiantis baltymo genas
IGF-I – insulino augimo faktorius
MC1R - melanocitus stimuliuojančio hormono receptoriaus genas
MC4R – melanokortino 4 receptorius
MHS – stresinį sindromą sąlygojantis genas
MYOG – miogeno genas
PGR – polimerazin÷ grandinin÷ reakcija
PIT-1 – hipofiz÷s transkripcijos faktorius
PSE – blyškios, minkštos, vandeningos m÷sos sindromas (pale, soft, exudative)
RFIP – restrikcinių fragmentų ilgio polimorfizmas
RN – Rapole genas
QTL – kokybinių požymių lokusai
OD - optinis tankis
TaqI – restriktaz÷
TAE – TAE buferis

5
Darbo tikslas
Ištirti miostatino geno įvairovę kiaulių tarpe ir nustatyti polimorfizmo įtaką kiaulių
produktyvumo savyb÷ms.
Darbo uždaviniai
• Surinkti ir išanalizuoti mokslinę literatūrą apie miostatino geną.
• Įdiegti kiaulių miostatino geno tyrimo metodiką LVA K.Janušausko Gyvūnų
genetikos laboratorijoje.
• Ištirti miostatino geno įvairovę kiaulių tarpe, naudojant sumodeliuotus pradmenis
pagal geno seką.
• Suformuoti fenotipinių požymių duomenų bazę.
• Ištirti miostatino geno įvairov÷s įtaką kiaulių produktyvumo savyb÷ms.
Darbo naujumas
• LVA Gyvūnų genetikos laboratorijoje įdiegta kiaulių miostatino geno, ištyrimo
metodika.
• Pirmą kartą ištirtas miostatino geno polimorfizmas bei jo variantų įtaka kiaulių
produktyvumo savyb÷ms
Praktinis pritaikymas
Įdiegta metodika leidžia ištirti kiaulių genome miostatino geną, nustatyti alelių
polimorfizmą ir įvertinti jo poveikį kiaulių fenotipin÷ms savyb÷ms.
Siūloma atsižvelgti į kiaulių genome esančio miostatino geno polimorfizmą, atrenkant
ir parenkant gyvulius, turinčius ekonomiškai naudingus alelius, kad būtų galima pagerinti
produktyviąsias savybes.

6
ĮVADAS
Kiaulių produktyviąsias savybes veikia tiek fenotipiniai, tiek genetiniai faktoriai.
Kiaulių autosomose atrasti genai arba genų grup÷s, lemiantys vislumą, pen÷jimosi spartą,
raumeningumą, m÷sos kokybę, rezistentiškumą ligoms. Gyvulių atrinkimas veisimui, pagal
konkrečius genus, leidžia suformuoti bandas su pageidaujamais požymiais.
Pasaulyje, paskutiniu dešimtmečiu kiaulių genetikoje susidom÷ta esamais kiekybinių
požymių lokusais bei jų įtaka skerdenos sud÷ties pokyčiams (t.y. riebalų šalinimui bei liesos
m÷sos didinimui) ir skerdenos kokybei , tai svarbiausios šios charakteristikos apibr÷žiančios
skerdenos kokybę ir vartotojų poreikį. Kiekybinių požymių lokusų įtakos galvijų bei kiaulių
augimo greičiui ir skerdenos sud÷čiai tyrimai buvo aprašyta eil÷je publikacijų (Alexandra et
al., 1997; Switonski, 2002; Di Stasio et al., 2003).
Tačiau, kadangi aukštus kokybinius ir kiekybinius rodiklius įtakoja eil÷ genų bei genų
grupių, kurių ekspresija ne visada būna vienoda, tod÷l, prieš įtraukiant į selekcijos
programas, geno žymenio įtaką būtina įvertinti konkrečiose kiaulių veisl÷se (Chikuni et al.,
1997; Grochowska et al., 1999; Curi et al., 2005).
Lietuvos veterinarijos akademijoje veikianti K.Janušausko gyvūnų genetikos
laboratorija vykdo naminių gyvūnų ir paukščių paveldimų ligų ir ydų tyrimus, užsiima
selekcijos efektyvumo, gyvulių produktyvumo bei pieno ir m÷sos kokyb÷s gerinimu
molekulinių genetinių metodų pagalba, vykdo žem÷s ūkio gyvūnų genetinių išteklių
monitoringą. LVA gyvūnų genetikos laboratorijoje buvo atlikti tyrimai ir nustatyta
melanokortino 4 receptoriaus (MC4R) geno įtaka Lietuvos baltųjų kiaulių augimo greičiui ir
skerdienos kokybei, o taip pat buvo analizuota kiaulių stresinį sindromą sąlygojančio geno
(MHS) įtaka kiaulių sveikatingumui, produktyvumui ir reprodukcin÷ms savyb÷ms (Jokubka
and Miceikien÷, 2001; Malevičiūt÷ et al., 2002; Jokubka and Miceikien÷,2003:).
Yra žinoma visa eil÷ genų įtakojančių kiaulienos produkcijos kiekį bei kokybę. Tai
miostatino, leptino, augimo hormono, augimo hormono receptoriaus, halotano ir kt. genai
(Switonski, 2002). Lietuvoje miostatino geno įtaka kiaulių produktyvumui, produkcijos
kokybei bei reprodukcin÷ms savyb÷ms nebuvo tyrin÷ta.

7
1. LITERATŪROS APŽVALGA
1.1. Genai, įtakojantys kiaulienos kiekį ir kokybę
Kiaulių DNR yra išsid÷sčiusi 19 chromosomų porų ir sudaryta iš daugiau kaip 100
tūkstančių funkcionuojančių genų. Kiaulių atrinkimas pagal genetinius markerius gali tur÷ti
svarbią įtaką kiaulienos kiekiui ir kokybei.
1.2. Miogenino (MYOG) genas
Miogeno (MYOG) geno pagrindinis vaidmuo yra raumenų diferenciacija, kontroliuojant
miofibrilių susidarymą. M÷siniuose gyvuliuose, tokiuose kaip kiaul÷s ir galvijai, miofibrilių
skaičius yra susijęs su augimo galia. Saumillion A. ir kt. (Saumillion, 1997) išanalizavo
kiaulių MYOG geno genetinį kintamumą ir jo ryšį su augimo savyb÷mis. MYOG geno
fragmentai buvo išskirti naudojant PGR ir patikrinti pagal genominę biblioteką su keturių
žmonių MyoD DNR fragmentais. Ir egzonų, ir promotoriaus regionai buvo labai panašūs į
žmogaus ir pel÷s genus. Sautern bloto 105 negiminingų kiaulių analiz÷ parod÷ tris
polimorfines MspI sritis, esančias geno promotoriaus regione, antrame introne ir 3’ krašte.
Buvo sukurti PGR-RFIP testai, randantys keturis MYOG alelius. PGR-RFIP testai ir
mikrosatelitų žymenys suteik÷ galimybę genotipuoti didelį kiaulių skaičių, norint išanalizuoti
m÷sos atsid÷jimo ir augimo savybių genetinį ryšį. MyoD geno šeima sudaryta iš keturių
struktūriškai artimų genų – tai MYOD1, MYOG, MYF5 ir MYF6. Šie genai koduoja
pagrindinius spiral÷-kilpa-spiral÷ (bHLH) baltymus ir susiję su raumeninių ląstelių
diferenciacija tiek in vitro, tiek in vivo. MYOG yra vienintelis MyoD genas, kuris randamas
visų skeleto raumenų ląstelių linijose. Pagrindinis MYOG vaidmuo raumenų diferencijos
procese, manoma, gali būti susijęs su skirtingais mioblastų ir miofibrilių kiekiais. Norint
nustatyti skirtingų MYOG alelių įtaką kiaulių raumenų vystimuisi ir augimui, reikia išskirti
genominius MYOG fragmentus ir apibūdinti keturis haplotipus, esančius MspI srityje. Greitas
PGR-RFIP testas suteikia galimybę nustatyti ryšius susijusius su augimo savyb÷mis
(Saumillion, 1997). Myf-5 ir MyoD1 genai ekspresuojami proliferuojančiose pirmin÷se
ląstel÷se, vadinamomis mioblastais. Myf-6 genas iš esm÷s ekspresuojamas pogimdyminiu
periodu (Pas, 1999).

8
1.3. Leptino (LEP) genas
Kiaulių riebumas yra vienas iš ekonomiškai svarbių požymių, kadangi rinka pageidauja
liesesn÷s kiaulienos, be to, riebalų kiekio padid÷jimas reikalauja didesnių pašarų sąnaudų.
Leptinas yra 16 kDa baltymas, išskiriamas baltųjų riebalų ląstelių (adipocitų), kuris reguliuoja
pašaro įsisavinimą, energijos sunaudojimą bei viso organizmo energijos balansą. Leptino
geno mutacijos nulemia visiškai nutukusio fenotipo ob/ob pasireiškimą pel÷se (Jiang, 1999).
Leptiną išskiria riebalinis audinys ir nulemia svorio praradimą graužikams, sumažinant
pašaro įsisavinimą ir padidinamas energijos suvartojimas. Pastaraisiais metais daug tyrimų
atliekama norint išsiaiškinti leptino įkatą svorio reguliavime, paaiškinant nutukimo fiziologiją.
Leptinas yra svarbus ne tik svorio reguliavimui ir energijos balansui, bet taip pat veikia kaip
metabolinis ir neuroendokrininis hormonas. Leptino receptoriai buvo rasti choroidiniame
rezginyje ir pagumburyje, taip pat hipofizio liaukoje skirtingose rūšyse, tokiose kaip žiurk÷s ir
pel÷s, kiaul÷s, avys ir žmogus.(Barata et.al., 2002; Saleri et al., 2002).
1.4. Augimo hormono (GH) genas
Augimo hormono geno pasireiškimo kelias susideda iš skirtingų tarpusavyje susijusių
genų, tokių kaip GH (augimo hormonas), IGF1 (insulino augimo faktorius), PIT1 (hipofiz÷s
transkripcijos faktorius), GHRH (augimo hormoną atpalaiduojantis hormonas),
somatostatinas, GHRHR (augimo hormoną atpalaiduojančio hormono receptorius), GHR
(augimo hormono receptorius) veiklos. Riebalų storis ir vidutinis dienos prieaugis yra labai
svarbūs požymiai kiaulienos produkcijai, nes jie koreliuoja su augimu ir liesos m÷sos kiekiu
skerdenoje. GH genas yra susijęs su šiais dviem požymiais. Tyrin÷damas ApaI ir HinpI
polimorfizmą, Knorr ir kiti (1997) rado ryšį tarp genų variantų ir riebalų savybių ir padar÷
išvadą, kad GH lokusas turi būti atidžiai ištirtas ir įtrauktas į naudingas selekcija žymenų
pagalba programas. Panašiai, Yu ir kiti (1995, 1999), Franco Mauricio ir kiti (2005) ištyr÷ ryšį
tarp PIT1 polimorfizmo ir skerdenos bei požymių savybių ir pareišk÷, kad šis genas gali būti
genas kandidatas, tiriant kiaulių QTL (kokybinius požymių lokusus) (Pierzchala et al., 2004).
Kiaulių augimo hormono genas yra sudarytas iš 190 amino rūgščių ir jį išskiria
priešakin÷s hipofiz÷s somatotropin÷s ląstel÷s. GH genas yra randamas 12 chromosomoje. GH
genas yra žinomas, kaip turintis didelę įtaką kiaulių skerdenos kokybei (Sun et al., 1997).

9
1.5. Rapole (RN) genas
Jeigu didžioji dalis pjetr÷nų savo genome turi streso (halotano) geną, tai hempšyrų ir su
jais poruotų kiaulių 15-toje chromosomoje yra atrastas RN genas. Pastarasis genas taip pat
blogina m÷sos kokybę, ypač kumpio.
RN geno neigiamų pasekmių priežastis yra glikogeno kiekio padid÷jimas raumenyje,
tose kiaul÷se, kurios turi geną. Šis genas egzistuoja 2 alelinių genų formoje: rn + alelinis
genas (normalus alelinis genas) ir RN-alelinis-genas (alelinis genas su neigiamu efektu N).
Ligi šiol, šis genas buvo surastas Hempšyrų veisl÷je ir Hempšyrų mišrūnų linijose.
Nepalankus alelinis genas (RN-) paveikia technologinę m÷sos kokybę, radikaliai didindamas
glikogeno kiekį. Glikogenas yra susikaupusio cukraus kiekis, kurio atsargos yra kaupiamos
kepenyse ir raumenyse. Po skerdimo, glikogenas raumenyse yra paverčiamas į pieno rūgštį,
kuri sumažina raumens pH. Tod÷l, kuo daugiau glikogeno yra raumenyje, tuo pieno rūgšties
bus daugiau pagaminta ir tuo bus žemesnis galutinis raumens pH. Padidintas pieno rūgšties
kiekis, gali būti priežastis to, kad raumens pH nukrenta žemiau 5,5 per 24 valandas po
skerdimo. "Rūgšti m÷sa" labai panaši savo savyb÷mis į išblyškusią, minkštą ir vandeningą
(PSE) m÷są, sukeltą PSS geno. Iš tikrųjų, kai PSS genas dalyvauja, jis stiprina padarinį RN
geno m÷sos kokybei. Skirtingai nuo Hal geno, RN genas turi poveikį turiniui, bet ne normai,
pH nukritimo (pH 24 val. <5.5), kuris priveda prie “ rūrščios m÷sos”, geriau žinomos kaip
Hampšyro padarinys. RN-alelinio geno pasekm÷ yra vandeninga m÷sa (4% skirtumas) ir
padid÷ję virimo nuostoliai (8 % skirtumas), sąlygojantis didelius finansinius nuostolius.
Neseni atradimai molekulin÷je genetikoje, leido nustatyti RN geną 15 chromosomoje ir kitimą
PRKAG3 gene, susijusiame su glikogeno kiekiu. Jie gali būti identifikuoti nauju molekuliniu
metodu, geresnei šio geno kontrolei (Hamilton et al., 2001)
1.6. Halotano (HAL) genas
Halotano genas turi kelis pavadinimus. Jis dar vadinamas streso (STRES) genu, MH
(hipertermijos) genu, CRC (kalcio pralaidumo kanalo) genu, RYR (riadonino receptoriaus)
genu.
Intensyvinant selekciją m÷singumo gerinimo kryptimi, padid÷jo stresams jautrių kiaulių
skaičius. Šį reiškinį sukelia kiaulių 6-oje chromosomoje sutinkamas recesyvinis halotano
genas (n). Jeigu šis genas yra homozigotin÷s būkl÷s (nn), tokios kiaul÷s yra jautrios stresams.
Literatūros ir atliktų tyrimų duomenimis, stresams jautrios kiaul÷s sunkiau negu jiems

10
atsparios išgyvena stresines situacijas, blogiau prisitaiko prie pakitusių aplinkos sąlygų,
jautrios stresams kiaul÷s blogiau penisi, jų m÷sa šviesi, minkšta, vandeninga (angl., pale, soft,
exudative – sutrumpintai žymima PSE) arba atvirkščiai – tamsi, kieta ir sausa (angl., dark,
firm, dry – sutrumpintai žymima DFD). Jautrios stresams kiaul÷s yra raumeningesn÷s, tačiau
jų m÷sa, kuriai būdingos PSE savyb÷s, yra blogos kokyb÷s, ne taip tinka perdirbti, ypač
rūkytiems gaminiams, prastesn÷ jos prekin÷ išvaizda. Tokią m÷są perdirbant did÷ja gamybos
kaštai, maž÷ja produkcijos išeiga, o galiausiai – blog÷ja ekonominiai kiaulienos gamybos
rodikliai (Leach et al., 1996; Hamilton et al., 2001)
Įvairių veislių kiaul÷s į stresus reaguoja skirtingai. Labai produktyvių veislių gyvuliai
yra jautresni. Dažniausiai tai m÷sinių veislių kiaul÷s, ilgai selekcionuotos m÷singumo
gerinimo kryptimi. Tokių gyvulių organizme vykstantys intensyvūs metabolizmo procesai
skatina mas÷s augimą, o adaptacin÷s savyb÷s prast÷ja. Apibendrintais įvairių užsienio šalių
literatūros duomenimis, jautriausi stresams yra pjetr÷nai ir įvairios selekcijos landrasai. Be to,
stresams jautresni yra kuiliukai negu kiaulait÷s (Kriauzien÷ ir kt., 2005)
Kiaulininkyst÷je daugelį stresinių situacijų būtina pašalinti, tačiau jų visiškai
išgyvendinti neįmanoma. Tod÷l greta m÷singumo gerinimo turi būti vykdoma ir stresams
atsparesnių kiaulių atranka. Kiaulių stresinę reakciją galima nustatyti daugeliu metodų, tačiau
tiksliausią stresams atsparesnių kiaulių atranką galima padaryti, naudojant genotipinio
įvertinimo metodus. Daugelyje pasaulio šalių sparčiai diegiamas genetinių žymenų metodas
(polimerazin÷s grandinin÷s reakcijos metodas, sutrumpintai žymimas PGR). Juo galima ištirti
įvairaus amžiaus kiaules. Tam užtenka iškirpti ausies gabaliuką, galima paimti spermos,
kraujo arba šerių su šaknel÷mis pavyzdžius, kurie laboratorijoje specialiai apdorojami ir iš jų
išskiriama DNR (deoksiribonukleinin÷ rūgštis). Analizuojant DNR grandinę, genetinių
žymenų metodu nustatomos ne tik jautrios stresams kiaul÷s (nn), bet ir tos, kurios yra
heterozigotin÷s (Nn) ir su visiškai dominuojančiais atsparumo stresams genais (NN). Šis
metodas įdiegtas ir Lietuvoje.
1.7. Melanokortino 4 receptoriaus (MC4R) genas
Genas yra lokalizuotas kiaulių 1 chromosomoje q22-27 regione. MC4R genas vaidina
svarbų vaidmenį reguliuojant kiaulių medžiagų apykaitos energijos balansą, riebalų
atsid÷jimą, augimo greitį ir maisto medžiagų pasisavinimą. Kiaulių energijos balansas remiasi
energijos suvartojimu iš maisto medžiagų kūno energijai ir medžiagų apykaitai.

11
Melanokortino 4 receptoriaus lokuso nustatytas pleotropinis poveikis produktyvumo
savyb÷ms: paros priesvoriui (g), nugaros lašinių storiui (mm), raumeningumui (%). Šis
genetinis žymuo gali būti naudojamas veisimo tikslais atrenkant gyvulius pagal tokius
požymius kaip riebalų atsid÷jimas, augimo greitis bei pašarų pasisavinimas. Atlikus tyrimus
Lietuvoje nustatyta, melanokortino 4 receptoriaus (MC4R) geno įtaka Lietuvos baltųjų kiaulių
augimo greičiui ir skerdienos kokybei. (Jokubka ir kt., 2003; Miceikien÷ ir kt., 2007).
1.8. Kiaulių selekcija genetinių žymenų pagalba
Dabartin÷s kiaulininkyst÷s tikslą galima būtų įvardinti kaip selekcijos proceso
spartinimą veisiant aukšto produktyvumo, atsparias ligoms bei aplinkos veiksniams, genetinių
defektų neturinčias kiaules, iš kurių gaunama produkcija būtų saugi žmonių mitybai, o jos
perdirbimas produktų gamybai reikalautų kuo mažiau ekonominių, energetinių bei laiko
sąnaudų. Vienas iš būdų per trumpesnį laiką pasiekti šiuos tikslus yra naujų biotechnologinių
metodų, tokių kaip genetinių žymenų (MAS), diegimas ir taikymas selekcijoje. Genas ar genų
grup÷, lemianti tam tikrą požymį, yra vadinama genetiniu žymeniu. Genetinių žymenų
įjungimas į veisimo programas leidžia padidinti selekcijos efektyvumą ir prapl÷sti požymių,
kuriais galima efektyviai manipuliuoti, spektrą. Panaudojant žymenis, galima vykdyti
selekciją tiek pagal vieną požymį, kurį lemia vienas genas, tiek pagal vieną ar kelis požymius,
kuriuos lemia keletas genų. Kiaulių veislininkyst÷je naudojamus genetinius žymenis galima
suskirstyti į dvi grupes:
Kai požymį lemia vienas genas:
HAL genas - esantis 6 chromosomoje, lemiantis jautrumą stresams, pablog÷jusią m÷sos
kokybę ir tuo pačiu įtakojantis didesnį raumeningumą;
RN genas - esantis 15 chromosomoje, lemiantis pablog÷jusią m÷sos kokybę;
ESR genas - esantis 1 chromosomoje, lemiantis paršelių skaičių lizde.
Kai požymį lemia genų grup÷, vadinama "kiekybinių požymių lokusu":
OPN lokusas - esantis 8 chromosomoje, lemiantis paršelių skaičių lizde;
Lokusas 4 chromosomoje, įtakojantis pen÷jimosi spartą;
K 88 lokusas, esantis 13 chromosomoje, lemiantis atsparumą kai kurioms ligoms.
Taigi, atsiradus naujiems - molekuliniams - genotipo įvertinimo metodams, atsiveria naujos
galimyb÷s kiaulių selekcijoje. Ją galima vykdyti pagal šiuos genetinius žymenis (EPL -
ekonominių požymių lokusus):

12
• m÷sos kokyb÷s rodiklius - m÷sos pH (rūgštingumą), vandens rišlumą,
tarpraumeninių riebalų kiekį sąlygojančius genų žymenis;
• reprodukcines savybes - paršelių skaičių vadoje sąlygojantį estrogeno
receptoriaus žymenį;
• produktyvumo požymius - pen÷jimosi intensyvumo lokuso žymenį, m÷singumo
lokuso žymenį;
• sveikatingumo požymius – organizmo bendro atsparumo ligoms (MHC) lokuso
žymenis, organizmo atsparumo stresams (HAL) geno žymenį.
Išvardinti pagrindiniai rodikliai yra svarbūs kaip objektas, atliekant kiaulių genetinius
tyrimus. Žem÷s ūkio gyvulių genomų tyrimai sudar÷ sąlygas žem÷s ūkio gyvulių genetinių
žem÷lapių sudarymui, o jų palyginimas leido atlikti genų, susijusių su kiekybinių požymių
lokusais, paiešką. Gyvulių genomo genetiniai žem÷lapiai yra pritaikyti atliekant kelių lokusų
sąsajų, lokusų ir genų, susijusių su kiaulių ekonomiškai naudingais požymiais, tyrimus.
Nežiūrint į tai, nustatytų kiekybinių požymių lokusų skaičius vis dar yra nedidelis, o nustatytų
atskirų genų, kurie yra identifikuoti kaip atsakingi už kiekybinius požymius skaičius – dar
mažesnis. Nustatyti pavieniai genai, susiję su kiekybinių požymių lokusais, įtraukiami į
veisimo programas, derinant juos su įprastiniais fenotipiniais selekciniais metodais – atliekant
gyvulių selekciją genetinių žymenų pagalba (MAS). Toks kombinuotas populiacijos
selekcijos būdas daug efektyvesnis ir rezultatai pasiekiami per trumpesnį laiko tarpą.
Genetinių žymenų, kurie gali būti panaudoti selekcijoje, paieška yra pagrįsta dvejomis
strategijomis: kiekybinių požymių lokuso kartografavimu ir potencialių genų įtakos
nustatymu. Kiekybinių požymių lokusų paieška, atliekant pilną genomo skenografiją, yra
apibūdinama kaip chromosomų segmentų, kuriuose yra lokalizuoti kandidatiniai genai,
atsakingi už požymį, tyrimas, nenustatant konkretaus geno ar mutacijos. Kandidatinio geno
tyrimo būdas yra parankus, kadangi yra tiriami vienas ar keletas konkrečių genų, kurie,
manoma, turi įtakos tiriamiems požymiams (Miceikien÷ ir kt., 2007).
13
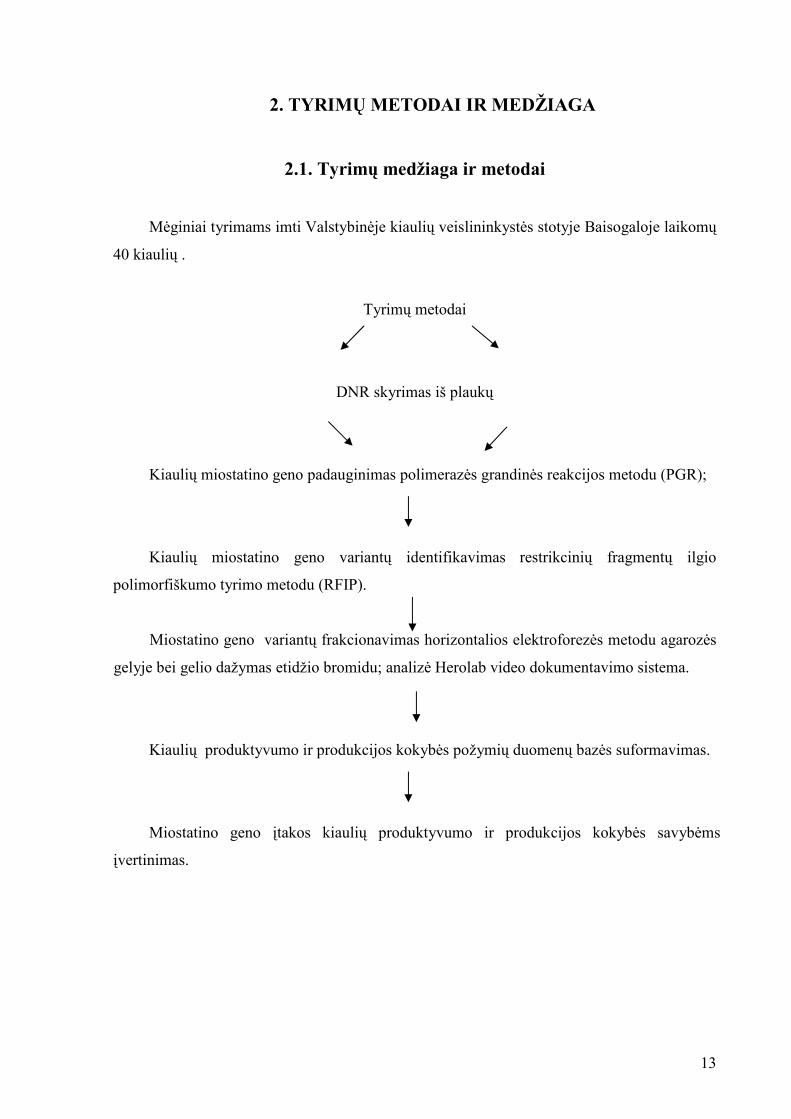
2. TYRIMŲ METODAI IR MEDŽIAGA
2.1. Tyrimų medžiaga ir metodai
M÷giniai tyrimams imti Valstybin÷je kiaulių veislininkyst÷s stotyje Baisogaloje laikomų
40 kiaulių .
Tyrimų metodai
DNR skyrimas iš plaukų
Kiaulių miostatino geno padauginimas polimeraz÷s grandin÷s reakcijos metodu (PGR);
Kiaulių miostatino geno variantų identifikavimas restrikcinių fragmentų ilgio
polimorfiškumo tyrimo metodu (RFIP).
Miostatino geno variantų frakcionavimas horizontalios elektroforez÷s metodu agaroz÷s
gelyje bei gelio dažymas etidžio bromidu; analiz÷ Herolab video dokumentavimo sistema.
Kiaulių produktyvumo ir produkcijos kokyb÷s požymių duomenų baz÷s suformavimas.
Miostatino geno įtakos kiaulių produktyvumo ir produkcijos kokyb÷s savyb÷ms
įvertinimas.

14
2.2. Kiaulių fenotipinių požymių duomenų baz÷s
Buvo surinkti duomenys ir suformuota kiaulių sekančių požymių duomenų baz÷:
Pen÷jimosi savyb÷s –
amžius skerdimo metu, d.; amžius esant 100 kg mas÷s, d.; priesvoris per parą, g.; suvartota
pašarų per pen÷jimosi laikotarpį; pašarų sąnaudos 1 kg priesvorio.
M÷sin÷s savyb÷s –
šiltos skerdienos mas÷, kg; skerdienos pusel÷s ilgis, cm; bekono pusel÷s ilgis, cm; nugaros
raumens plotas, cm²; kumpio mas÷, kg; lašinių storis ties 6-7 šonkauliu, mm; lašinių storis ties
10 šonkauliu, mm; lašinių storis už paskutinio šonkaulio, mm; lašinių storis ties paskutiniu
juosmens slanksteliu; lašinių storis taške Fat1, mm; lašinių storis taške Fat2, mm; raumeninis
storis taške Fat2, mm; raumeningumas, %.
2.3. DNR skyrimas iš plauko svogūn÷lio ląstelių
Nukerpami 4-5 plaukų svogūn÷liai ir patalpinami į m÷gintuv÷lius. Paruošiamas lizavimo
mišinys (DTT -7,5 µl, Chelex- 200µl, Protenaze K (20mg/ml)- 10.7µl). M÷gintuv÷lio turinys
užpilamas lizavimo mišiniu (vienam pavyzdžiui imama 218,2µl paruošto mišinio).
M÷gintuv÷liai 30s. maišomi maišykl÷s „Vortex" pagalba. Centrifuguojami l0 s 13500
aps./min. greičiu. Pavyzdžiai 30 min. inkubuojami 56°C temperatūroje. Po inkubacijos m÷giniai
(po 10µm) pakaitinami 94°C temperatūroje (inaktyvuojami - amplifikatoriuje) bei paliekami
nakčiai.
2.4. DNR švarumo ir koncentracijos nustatymas
Genomin÷s DNR kiekis ir grynumas nustatomas spektrofotometrinio metodo pagalba.
Tam tikslui paruošiamas l00 µl skiestos DNR tirpalas: imama l0 µl koncentruotos DNR ir
skiedžiama 90µl distiliuotu vandeniu. DNR kiekis nustatomas išmatuojant skiesto tirpalo
optinį tankį (OD) prie 260nm bangos ilgio. Kai OD = 1, tai l ml tirpalo yra 50ug dvigrand÷s
DNR.
DNR kokyb÷ įvertinama išmatavus skiesto tirpalo optinius tankius prie 260 ir 280 nm
bangos ilgių. Švarumą rodo santykis OD260/OD280. Švarių DNR tirpalų santykis yra 1.8-2.0.
Jei tirpale yra baltymų ar fenolio priemaišų, šis santykis bus mažesnis nei nurodyta. Baltymų

15
koncentracija neturi viršyti 0.5 mg/ml ribos. Jei tirpale priemaišų yra daugiau, reikia atlikti
pakartotinį genomin÷s DNR valymą.
.
2.5. Polimerazin÷ grandinin÷ reakcija (PGR)
Polimerazin÷ grandinin÷ reakcija yra metodas, pavadintas pagal DNR polimerazę,
fermentą, vykdantį DNR replikaciją ląstel÷je. Tai yra nukleino rūgščių sintez÷s in vitro
metodas, kuriuo laboratoriniame m÷gintuv÷lyje gali būti specifiškai padauginti (amplifikuoti)
atskiri DNR fragmentai. PGR metodas yra labai jautrus, tod÷l padauginti pasirinkta DNR
atkarpa gali sudaryti netgi vieną milijoninę bendro DNR pavyzdžio dalį. Tai reiškia, kad,
naudojant PGR galima amplifikuoti net ir vienintelį pasirinktą geną.
2.5.1. Kiaulių miostatino geno MYF5 (PGR-RFIP) tyrimo metodika
MYF-5 geno pradmenys:
Pirminis (5‘ CTCCGAATTAGTGTGGCTTC-3‘)
Atvirkštinis (5‘ –GTTCTTTCGGGACCAGACAGGCCTC-3‘)
Mišinys PGR reakcijai:
PGR komponentai: Vienai reakcijai atlikti (µl)
Geros kokyb÷s dejonizuotas vanduo ddH2O 2,75
Buferis 10xPCR (be MgCl2) 2,5
MgCl2 (konc. 50mM) 1,5
dNTP miksas (2mM) 2,5
Pirminis pradmuo (darbin÷ konc. 10pmol) 2,5
Atvirkštinis pradmuo (darbin÷ konc. 10pmol) 2,5
BSA 0,25
Taq polymeraz÷ (1vnt./µl) aktyvumo 0,5
PGR atliekama automatiniame amplifikatoriuje GeneAmp ® PGR sitema 2700, AB Applied
Biosystems.

16
PGR režimas:
95 OC, 5 min (pirmin÷ denatūracija)
94 OC, 45s (denatūracija),
60 OC 1 min (oligonukleotidų prisijungimas),
72 OC 1 min (DNR grandin÷l÷s sintez÷).
72 OC, Sintez÷s užbaigimui 10 min. Laikyti: 4 OC
Reakcija atliekama 35 ciklus
MYF-5 geno variantų ištyrimas
PGR produktas – 322 bp
DNR fragmentų dydis po karpymo restrikciniu fermentu HinfI, 370C per naktį.
A (alelis) -322 bp.
B (alelis) - 170 bp, 143 bp.
A B 322bp ��� 170bp ��� 143bp ���
2.6. Statistin÷ duomenų analiz÷
Kiaulių miostatino alelių ir genotipų dažnumai tirtoje grup÷je, faktinis ir
prognozuojamas heterozigotiškumas tirtuose lokusuose buvo apskaičiuoti R – statistiniu
paketu.
Vertinant miostatino geno įtaką kiaulienos kiekybiniams ir kokybiniams rodikliams
buvo atlikta vienfaktorin÷ dispersin÷ analiz÷ Vienfaktorin÷s analiz÷s būdu buvo įvertinta
atskirų genų ir agreguotų genotipų įtaka fenotipiniams požymiams.

17
3. TYRIMŲ REZULTATAI IR APTARIMAS
3.1. Kiaulių miostatino geno polimorfizmo ištyrimas
Viso buvo paimta 40 kiaulių m÷ginių, iš kurių buvo išskirta DNR, nustatyta jos
koncentracija bei švarumas. Tiriamas genas buvo padaugintas polimerazin÷s grandinin÷s
reakcijos metodu, o aleliai identifikuoti skaldant gautus PGR produktus karpymo fermentu ...
Kiaulių miostatino geno rasti 2 skirtingi aleliai (1 lentel÷, 1,paveikslas).
1 lentel÷. Rastas MYF5 geno alelių skaičius tirtoje kiaulių grup÷je
Lokusas Alelių skaičius
MYF5 2
1 paveikslas.Miostatinas (MYF – 5):1-markeris, 2, 4, 6, 8, 10, 12, 14, 16, 18 – PGR ; 3, 9, 11, 19 - AA genotipas;5, 7, 13, 15, 17 – AB genotipas.
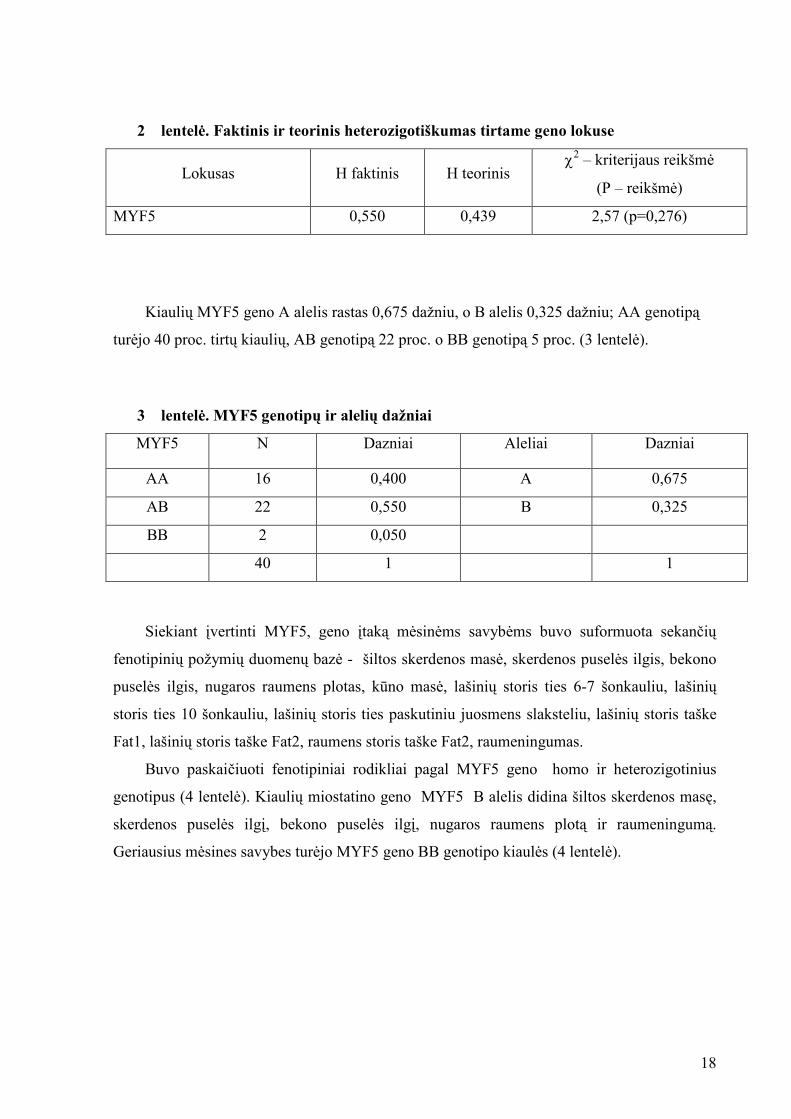
Įvertinus faktinį ir teorinį heterozigotiškumą visoje tirtoje gyvulių grup÷je faktinis
heterozigotiškumas buvo rastas didesnis, nei teorinis, kas rodo pakankamą genetin÷s
įvairov÷s kiekį tirtame lokuse, nors skirtumas n÷ra statistiškai reikšmingas. (2 lentel÷).
18
2 lentel÷. Faktinis ir teorinis heterozigotiškumas tirtame geno lokuse
Lokusas H faktinis H teorinis χ
2 – kriterijaus reikšm÷
(P – reikšm÷)
MYF5 0,550 0,439 2,57 (p=0,276)
Kiaulių MYF5 geno A alelis rastas 0,675 dažniu, o B alelis 0,325 dažniu; AA genotipą
tur÷jo 40 proc. tirtų kiaulių, AB genotipą 22 proc. o BB genotipą 5 proc. (3 lentel÷).
3 lentel÷. MYF5 genotipų ir alelių dažniai
MYF5 N Dazniai Aleliai Dazniai
AA 16 0,400 A 0,675
AB 22 0,550 B 0,325
BB 2 0,050
40 1 1
Siekiant įvertinti MYF5, geno įtaką m÷sin÷ms savyb÷ms buvo suformuota sekančių
fenotipinių požymių duomenų baz÷ - šiltos skerdenos mas÷, skerdenos pusel÷s ilgis, bekono
pusel÷s ilgis, nugaros raumens plotas, kūno mas÷, lašinių storis ties 6-7 šonkauliu, lašinių
storis ties 10 šonkauliu, lašinių storis ties paskutiniu juosmens slaksteliu, lašinių storis taške
Fat1, lašinių storis taške Fat2, raumens storis taške Fat2, raumeningumas.
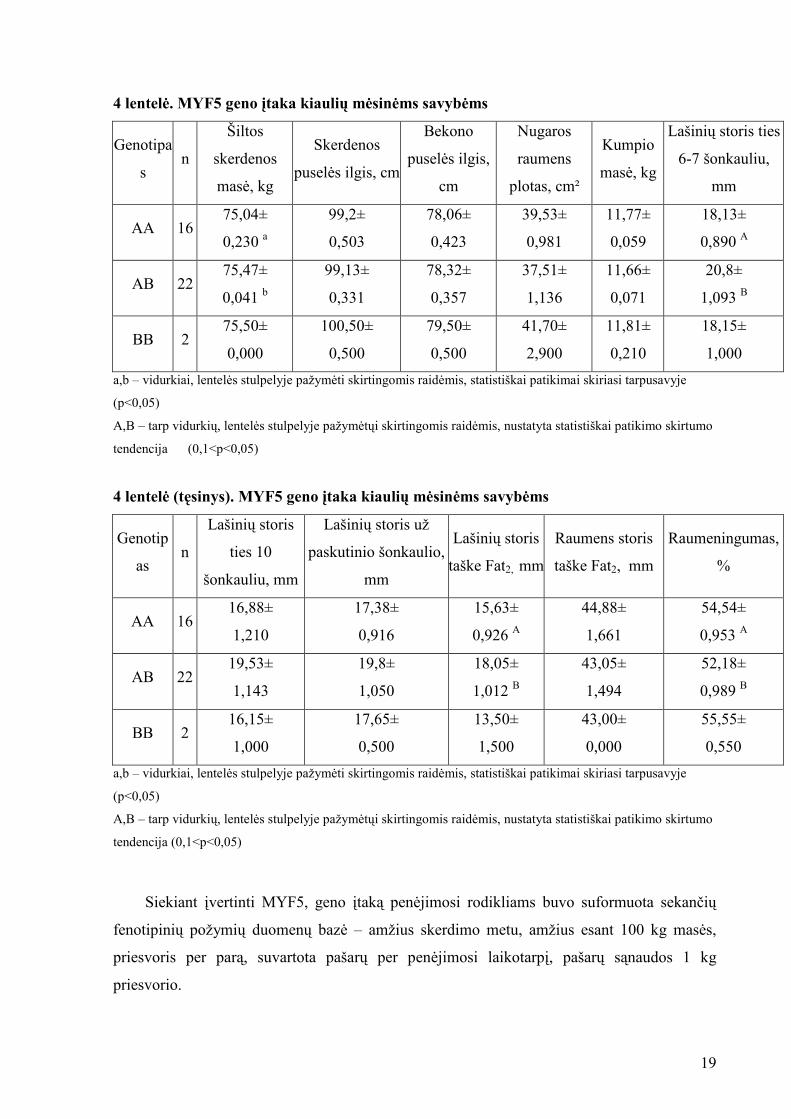
Buvo paskaičiuoti fenotipiniai rodikliai pagal MYF5 geno homo ir heterozigotinius
genotipus (4 lentel÷). Kiaulių miostatino geno MYF5 B alelis didina šiltos skerdenos masę,
skerdenos pusel÷s ilgį, bekono pusel÷s ilgį, nugaros raumens plotą ir raumeningumą.
Geriausius m÷sines savybes tur÷jo MYF5 geno BB genotipo kiaul÷s (4 lentel÷).
19
4 lentel÷. MYF5 geno įtaka kiaulių m÷sin÷ms savyb÷ms
Genotipa
s n
Šiltos
skerdenos
mas÷, kg
Skerdenos
pusel÷s ilgis, cm
Bekono
pusel÷s ilgis,
cm
Nugaros
raumens
plotas, cm²
Kumpio
mas÷, kg
Lašinių storis ties
6-7 šonkauliu,
mm
AA 16 75,04±
0,230 a
99,2±
0,503
78,06±
0,423
39,53±
0,981
11,77±
0,059
18,13±
0,890 A
AB 22 75,47±
0,041 b
99,13±
0,331
78,32±
0,357
37,51±
1,136
11,66±
0,071
20,8±
1,093 B
BB 2 75,50±
0,000
100,50±
0,500
79,50±
0,500
41,70±
2,900
11,81±
0,210
18,15±
1,000
a,b – vidurkiai, lentel÷s stulpelyje pažym÷ti skirtingomis raid÷mis, statistiškai patikimai skiriasi tarpusavyje
(p<0,05)
A,B – tarp vidurkių, lentel÷s stulpelyje pažym÷tųi skirtingomis raid÷mis, nustatyta statistiškai patikimo skirtumo
tendencija (0,1<p<0,05)
4 lentel÷ (tęsinys). MYF5 geno įtaka kiaulių m÷sin÷ms savyb÷ms
Genotip
as n
Lašinių storis
ties 10
šonkauliu, mm
Lašinių storis už
paskutinio šonkaulio,
mm
Lašinių storis
taške Fat2, mm
Raumens storis
taške Fat2, mm
Raumeningumas,
%
AA 16 16,88±
1,210
17,38±
0,916
15,63±
0,926 A
44,88±
1,661
54,54±
0,953 A
AB 22 19,53±
1,143
19,8±
1,050
18,05±
1,012 B
43,05±
1,494
52,18±
0,989 B
BB 2 16,15±
1,000
17,65±
0,500
13,50±
1,500
43,00±
0,000
55,55±
0,550
a,b – vidurkiai, lentel÷s stulpelyje pažym÷ti skirtingomis raid÷mis, statistiškai patikimai skiriasi tarpusavyje
(p<0,05)
A,B – tarp vidurkių, lentel÷s stulpelyje pažym÷tųi skirtingomis raid÷mis, nustatyta statistiškai patikimo skirtumo
tendencija (0,1<p<0,05)
Siekiant įvertinti MYF5, geno įtaką pen÷jimosi rodikliams buvo suformuota sekančių
fenotipinių požymių duomenų baz÷ – amžius skerdimo metu, amžius esant 100 kg mas÷s,
priesvoris per parą, suvartota pašarų per pen÷jimosi laikotarpį, pašarų sąnaudos 1 kg
priesvorio.
20
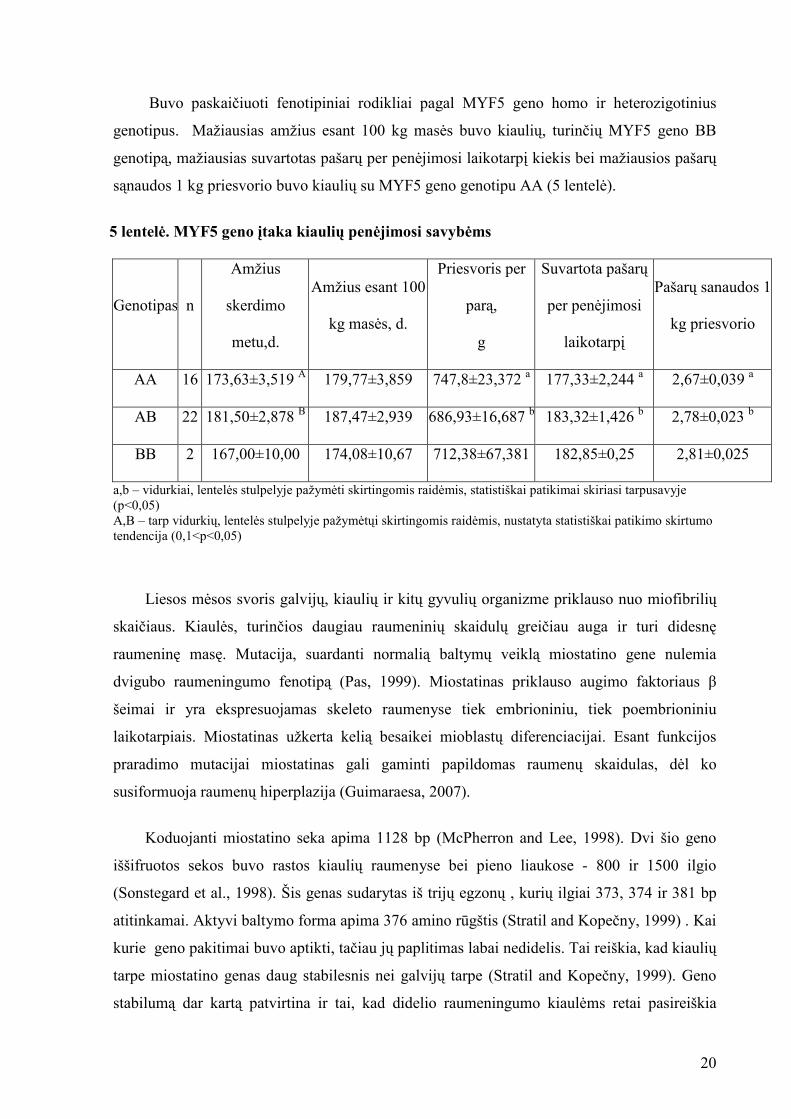
Buvo paskaičiuoti fenotipiniai rodikliai pagal MYF5 geno homo ir heterozigotinius
genotipus. Mažiausias amžius esant 100 kg mas÷s buvo kiaulių, turinčių MYF5 geno BB
genotipą, mažiausias suvartotas pašarų per pen÷jimosi laikotarpį kiekis bei mažiausios pašarų
sąnaudos 1 kg priesvorio buvo kiaulių su MYF5 geno genotipu AA (5 lentel÷).
5 lentel÷. MYF5 geno įtaka kiaulių pen÷jimosi savyb÷ms
Genotipas n
Amžius
skerdimo
metu,d.
Amžius esant 100
kg mas÷s, d.
Priesvoris per
parą,
g
Suvartota pašarų
per pen÷jimosi
laikotarpį
Pašarų sanaudos 1
kg priesvorio
AA 16 173,63±3,519 A 179,77±3,859 747,8±23,372 a 177,33±2,244 a 2,67±0,039 a
AB 22 181,50±2,878 B 187,47±2,939 686,93±16,687 b 183,32±1,426 b 2,78±0,023 b
BB 2 167,00±10,00 174,08±10,67 712,38±67,381 182,85±0,25 2,81±0,025
a,b – vidurkiai, lentel÷s stulpelyje pažym÷ti skirtingomis raid÷mis, statistiškai patikimai skiriasi tarpusavyje (p<0,05) A,B – tarp vidurkių, lentel÷s stulpelyje pažym÷tųi skirtingomis raid÷mis, nustatyta statistiškai patikimo skirtumo tendencija (0,1<p<0,05)
Liesos m÷sos svoris galvijų, kiaulių ir kitų gyvulių organizme priklauso nuo miofibrilių
skaičiaus. Kiaul÷s, turinčios daugiau raumeninių skaidulų greičiau auga ir turi didesnę
raumeninę masę. Mutacija, suardanti normalią baltymų veiklą miostatino gene nulemia
dvigubo raumeningumo fenotipą (Pas, 1999). Miostatinas priklauso augimo faktoriaus β
šeimai ir yra ekspresuojamas skeleto raumenyse tiek embrioniniu, tiek poembrioniniu
laikotarpiais. Miostatinas užkerta kelią besaikei mioblastų diferenciacijai. Esant funkcijos
praradimo mutacijai miostatinas gali gaminti papildomas raumenų skaidulas, d÷l ko
susiformuoja raumenų hiperplazija (Guimaraesa, 2007).
Koduojanti miostatino seka apima 1128 bp (McPherron and Lee, 1998). Dvi šio geno
iššifruotos sekos buvo rastos kiaulių raumenyse bei pieno liaukose - 800 ir 1500 ilgio
(Sonstegard et al., 1998). Šis genas sudarytas iš trijų egzonų , kurių ilgiai 373, 374 ir 381 bp
atitinkamai. Aktyvi baltymo forma apima 376 amino rūgštis (Stratil and Kopečny, 1999) . Kai
kurie geno pakitimai buvo aptikti, tačiau jų paplitimas labai nedidelis. Tai reiškia, kad kiaulių
tarpe miostatino genas daug stabilesnis nei galvijų tarpe (Stratil and Kopečny, 1999). Geno
stabilumą dar kartą patvirtina ir tai, kad didelio raumeningumo kiaul÷ms retai pasireiškia

21
raumenų hiperplazija. Tačiau joms būdinga raumenų hipertrofija, ypač Pjetr÷nų ir Belgijos
landrasų veisl÷ms. Remiantis atliktais tyrimais teigiama, jog Jorkšyrų veisl÷s kiaul÷s geba
geriau sintetinti baltymus raumenyse (Ezewke ir Martin, 1975), tod÷l jos raumeningesn÷s nei
to paties amžiaus ir svorio kitų veislių kiaul÷s.
Specialistų grup÷ ištyr÷ 105 skirtingų veislių kaules ir 3‘ polimorfizmą jie aptiko visose
tirtose kiaulių veisl÷se, išskyrus Meishan populiaciją. Ernst ir kolegos (1993) surado
dominuojantį alelinį MYOD geną skirtingose kinų kiaulių veisl÷se (Fengjing, Meishan,
Minzu), tai paaiškina kod÷l šių veislių kiaulių JAV ir Europoje yra tiek mažai.
Pagal mūsų gautus tyrimų rezultatus kiaulių miostatino genas veik÷ tiek m÷sines tiek
pen÷jimosi savybes. Panašius tyrimų rezulatatus gavo ir kiti mokslininkai (Saumillion et
al.,1997; Cieslak et al., 2003; Fausto et al.,2005).
Intensyvios gyvulininkyst÷s sąlygomis, kai pagrindinius reikalavimus diktuoja rinka,
ypač svarbu yra tinkamai pritaikyti selekcijos ir genetikos pasiekimus. Kiaulininkyst÷je
selekcijos procesas labiau nukreiptas į efektyvų heteroz÷s efekto panaudojimą. Pagrindiniais
kiaulių gerinimo būdais išlieka kryžminimas bei linijų selekcija. Elitiniuose veislynuose yra
išvedamos vertingos linijos, kurios dauginamos reprodukciniuose ūkiuose, o iš jų patenka į
pramoninius, kuriose pl÷tojama komercin÷ kiaulininkyst÷ ir auginami hibridai m÷sai.
Laikantis tokios selekcijos sistemos, kiaulininkyst÷je yra siekiama pagerinti kiaulių
pen÷jimosi, m÷sines ir reprodukcines savybes, sukurti bekoninio ir m÷sinio tipo hibridus.
Tačiau pastaruoju metu kiaulių veislininkyst÷je drauge su ekonominiais, iki šiol laikytais
pagrindiniais, selekcionuojamais požymiais vis didesnis d÷mesys kreipiamas į antraeilius –
kokyb÷s ir sveikatos požymius. Manoma, kad ateityje sudarant kiaulių veislininkyst÷s
programas būtina atkreipti d÷mesį į svarbiausių selekcionuojamų požymių biologinius ir
genetinius ryšius, daugiau d÷mesio skirti gaunamos produkcijos kokybei, gyvulių
sveikatingumui bei gerbūvio sąlygoms, o nustatant kiaulių veislinę vertę, taikyti metodus,
kurie apjungtų fenotipo ir genetinę informaciją (Miceikien÷ ir kt., 2007).

22
4. IŠVADOS
1. Kiaulių MYF5 geno A alelis rastas 0,675 dažniu, o B alelis 0,325 dažniu; AA
genotipą tur÷jo 40 proc. tirtų kiaulių, AB genotipą 22 proc. o BB genotipą 5 proc
2. Įvertinus faktinį ir teorinį heterozigotiškumą faktinis heterozigotiškumas buvo
rastas didesnis, nei teorinis, kas rodo pakankamą genetin÷s įvairov÷s kiekį ir
sudaro prielaidas vykdyti gyvulių selekciją pagal miostatino geną.
3. Kiaulių miostatino geno MYF5 B alelis statistiškai reikšmingai didina šiltos
skerdenos masę ir raumeningumą.
4. Geriausius m÷sines savybes tur÷jo MYF5 geno BB genotipo kiaul÷s.
5. Mažiausias amžius esant 100 kg mas÷s buvo kiaulių, turinčių MYF5 geno BB
genotipą.
6. Mažiausios pašarų sąnaudos 1 kg priesvorio buvo kiaulių su MYF5 geno
genotipu AA.

23
5. SUMMARY
Master's work;
Master: Vaida Palačenkait÷
Topic of Master degree thesis: Polymorphism of pig myostatin gene and it‘s inluence to
meat quantitative and qualitative traits. Tutor: Ilona Miceikien÷
Lithuanian Veterinary Academy, Department of Animal Breeding and Genetics, K.
Janušauskas Laboratory of Animal Genetics.
Master‘s work accomplished in the year 2007 – 2009, volume of Master work 26 pages
original, 5 tables.
Object and tasks of work. Analyse and summarize literature about myostatin gene in pig.
Introduce pig Myf 5 gene research methodology at K. Janušauskas Laboratory of Animal
Genetics, LVA..Investigate Myf 5 gene polymorphism and distribution of different alleles and
genotypes in pigs, create data base of phenotypic traits, investigate myostatin gene
polymorhism influence to pig productive traits.
Research methodology. DNA extraction from hair roots; PGR to amplify MYF5 gene;RFLP
method to identify mutation; Electrophoresis in agarose gel;.Staining with etidium Bromide
Genotyping;.Statistical analysis of data.
Results and conclusions. There were found two alleles for pocine MYF5 gene allele with
frequency 0,675 , B allele with frequency 0,325 ; AA genotype had 40 percent of tested
pigs, AB genotype 22 percent, BB genotype 5 percent. Observed heterozigosity was found
higher than expected what showed enough genetical variation and gave oportunity to use
miostaytin gene as genetic marker in selection programs. Pig myostatin gene MYF5 B allele
statistically significantly increased warm carcas weight and muscularity. The best meat traits
had pigs with genotype BB. Also they had the lowest age while reaching 100 kg. Smallest
feed intake for 1 kg makeweigt had pigs with genotype AA.

24
6. LITERATŪROS SĄRAŠAS
1. Alexandra C. McPherron and Se-Jin Lee. Double muscling in cattle due to mutations in
the myostatin gene. Proc. Natl. Acad. Sci. USA, 1997.Vol. 94, pp. 12457-12461.
2. Barata M., Saleri R., Mainardi G.L., Valle D., Giustina A., Tamanini C. Leptin Regulates
GH Gene Expression and Secretion and Nitric Oxide Production in Pig Pituitary Cells.
Endocrinology.2002. V.143(2), P.551-557.
3. Chikuni K., Tanabe R., Muroya S., Fukumoto Y. & Ozawa S. A simple method for
genotyping thr bovine growth hormone gene. Animal Genetics. 1997. 28, 230
4. Cieslak D., Blicharski T., Kapelanski W., Pierzchala M. Investigation of polymorphisms
in the porcine myostatin (GDF8; MSTN)gene. Czech J Anim Sci. 2003. 48: 69 – 75.
5. Curi R.A., Oliveira H.N., Silveira A.C. and Lopes C.R. Effects of polymorphic
microsatellites in the regulatory region of IGF1 and GHR on growth and carcass traits in
beef cattle. Animal Genetics. 2005. Vol. 36. P. 58-62.
6. Ernst C.W., Vaske D.A., Larson R.G., Rothschild M.F. Rapid communication: MspI
restriction fragment length polymorphism at the swine MYOG locus. Czech J Anim
Sci.1993. 71, 3479.
7. Ezekwe, M.O. and R.J. Martin. Cellular characteristics of skeletal muscle in selected
strains of pigs and mice and the unselected controls. 1975. Growth. 39:95.
8. Franco Mauricio M., Antunes Robson C., Silva Heyder D., Goulart Luiz R. Association of
PIT1, GH and GHRH polymorphisms with performance and carcass traits in Landrace
pigs. J. Appl.Genet. 2005. v. 46(2), P.195-200.
9. Fausto Morena da Silva, Guimaraes Simone Eliza Facioni, Lopes Paulo Savio, Pires
Aldrin Vieira, Guimaraes Marta Fonseca Martins, Barbosa da Silva Marcos Vinicius
Gualberto, Schierholt Alexs Sandro, Silva Kleibe de Moraes and Gomide Lucio Alberto
de Miranda. Association of MYF5 gene allelic variants with production traists in pigs.
Genetic and Molecular Biology. 2005. 28, 3, 363 – 369.
10. Hamilton D.N., Ellis M., Miller K.D., McKeith F.K., Parrett D.F. The effect of the
halothane and rendement napole genes on carcass and meat quality characteristics of
pigs. Journal of Animal Science. 2001. N. 78. P. 2862 – 2867.
11. Grochowska R., Zwierschowski L., Snochowski M. and Reklewski Z. Stimulated growth
hormone (GH) release in Friesian cattle with respect to GH genotypes. Reproduction
Nutrition Development. 1999. Vol. 39. P. 171-180.

25
12. Guimaraes Simone, Stahl Chad, Lonergan Steven, Geiger Becky, Rothschild Max.
Myostatin promoter analysis and expression pattern in pigs. Livestock Science. 2007. V.
112, P. 143-150.
13. Jokubka R., Miceikien÷ I. Polymerase chain reaction and restriction fragment length
polymorphism screening for the HAL gene. Proceedings of the Latvian Academy of
Sciences. ISSN 1407-009X. 2001. N. 5/6. P.237-241.
14. Jokubka R., Miceikien÷ I. A missence variation of melanocortin – 4 receptior gene
(MC4R) in Lithuanian White pig breed. Proceedings of the 9th Baltic Animal Breeding
Conference. Sigulda. 2003. P.65-67.
15. Yu, T-P., Tuggle C.K., Schmitz C.B., Rothschild M.F.. Association of PIT1
polymorphism with growth and carcass traits in pigs. J. Anim. Sci. 1995. V. 73, P. 282-
288.
16. Jiang Zhi-Hua, Gibson John P. Genetic polymorphisms in the leptin gene and their
association with fatness in four pig breeds. 1999. Mammalian Genome. V.10, P.191-193.
17. Knorr C., Moser G., Muller E. and Geldermann H. Association of GH gene variants with
performance traits in F-2 generations of European wild boar. Pietrain and Meishan pigs.
Animal Genetics. 1997. Vol. 28. P. 124-128.
18. Kriauzien÷ Janina, Miceikien÷ Ilona, Masiulien÷ Angel÷, Baltr÷nait÷ Lina. HAL geno
įvairov÷ S linijos kiaulių bandoje. Geno įtaka kiaulių reprodukcin÷ms savyb÷ms 2005.
Veterinarija ir Zootechnika. T.30 (52).
19. Leach L.M., Ellis M., Sutton D.S. et al. The growth performance, carcass
characteristics, and meat quality of halothane carrier and negative pigs. Journal of
Animal Science. 1996. N.74. P. 934 – 943.
20. Malevičiut÷ J., Baltr÷nait÷ L., Pečiulaitien÷ N., Miceikien÷ I. Genetinių žymeklių
panaudojimas gyvulių selekcijoje. Ūkininko patar÷jas. 2002. Nr.131 (1425).
21. McPherron A.C., Lee S. Double muscling in cattle due to mutations in the myostatin
gene. 1998. Proc. Natl. Acad. Sci. USA, 94, 12457-12461.
22. Miceikien÷ I., Baltr÷nait÷ L., Pečiulaitien÷ N., Kriauzien÷ J. Kiaulių veisimas ir
genetika. Kaunas: Terra Publica. 2007. 58 psl.
23. Pas M.F.W., Soumillion A.,. Harders F. L,. Verburg F. J, Bosch, Galesloot P., The
Meuwissen Influences of Myogenin Genotypes on Birth Weight, Growth Rate, Carcass
Weight, Backfat Thickness, and Lean Weight of Pigs. 1999. J. Anim. Sci.. V.77. P.123-
126.

26
24. Pierzchała Mariusz, Blicharski Tadeusz, Kurył Jolanta. Growth rate and carcass quality in
relation to GH/MspI and GH/HaeII PCR-RFLP polymorhism in pigs. Poland, 2004.
Animal Science Papers and Reports V. 22 N. 1, P. 57-64.
25. Saleri R., Mainardi G.L., Valle D., Giustina A., Tamanini C. Leptin Regulates GH Gene
Expression and Secretion and Nitric Oxide Production in Pig Pituitary Cells.
Endocrinology.2002. V.143(2), P.551-557.
26. Soumillion Ann, Erkens Jo H.F., Lenstra Johannes A., Günther Rettenberger, Pas Marinus
F.W. Genetic variation in porcine myogenin gene locus. 1997. Mammalian Genome. V.8,
N.8, P. 564-568.
27. Sonstegard T.S., Rohrer G.A., Smith T.P.L. Myostatin maps to porcine chromosome 15
by linkage and physical analyses. 1998. Anim. Genet., 29, 19-22.
28. Stasio L. Di., Brugiapaglia A., Destefanis G., Albera A., Sartore S. GH1 as candidate
gene for variability of meat production traits. J. Anim. Breed. Genet. 2003. 120, 358 –
361.
29. Stratil A., Kopečny M. Genomic organization, sequence and polymorphism of the
porcine myostatin (GDF8; MSTN) gene. 1999. Anim. Genet., 30, 468-470.
30. Switonski M. Molecular genetics in beef cattle breding a review. Animal Sciense Papers
and Reports. 2002. Vol. 20. Supplement 1, 7-18.
31. Sun H.,.Taylor C, Robic A.,. Wang L, Rothschild M. F., Tuggle C.K. Mapping of growth
hormone receptor of swine chromosome 18. 1997. Animal Genetics. V. 28, P. 351-353.