l22 genome evolution 15 (1)
DESCRIPTION
afdsTRANSCRIPT
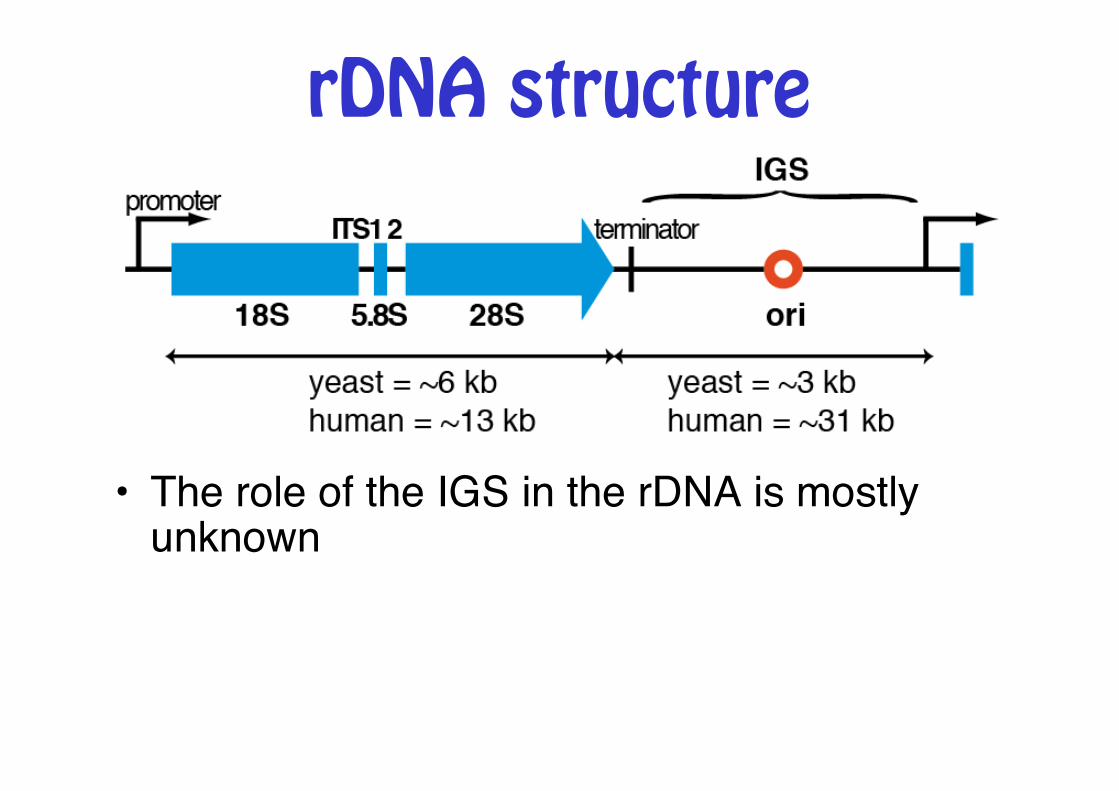
rDNA structure
• The role of the IGS in the rDNA is mostly unknown"

Phylogenetic footprinting identi!ed noncoding promoter

Characterisation of the noncoding promoter
Kobayashi & Ganley, 2005 Science 309: 1581-1584
rDNA copy number usually recovers if some are deleted
The noncoding IGS promoter is required
for this rDNA amplification

Detecting positive selection • We will look at three methods for detecting
positive selection:"• More non-synonymous than synonymous
mutations (Dn/Ds ratio)"• Phylogenetic Dn/Ds ratio"• Selective sweeps"

dN/dS ratio redux • The reverse logic for the dN/dS ratio can be
used to find evidence for positive selection"• Under positive selection, you expect more
non-synonymous mutations than synonymous mutations!
• Therefore a dN/dS ratio of greater than 1 suggests positive selection"

Example: heterochromatin protein 1 variant
• Looking at a protein called heterochromatin protein 1 (HP1), which functions in specifying heterochromatin"
• In Drosophila melanogaster there there are five HP1 paralogs"
• Found that one (HP1-rhino) showed evidence of evolving via positive selective"
• Suggests its involved in some aspect of adaptation"

Example: HP1_rhino protein
Other HP1 paralogs are evolving under negative selection, but the HP1-rhino shows a high dN/dS ratio"
Vermaack et al, 2005. PLoS Genetics 1: e9

Limitations of dN/dS for positive selection
• Similar to those for the dN/dS ratio with negative selection"
• Only works on protein-coding regions of the genome"
• Has limited sensitivity – typically only a few sites are undergoing positive selection"
• Need to take bias in mutations into account"

Phylogenetic dN/dS ratio • The dN/dS ratio averages over all evolution
that has occurred between species"• However, the positive selection may have
only occurred during one part of the evolutionary history – difficult to detect"
• Therefore, phylogenetic approaches are used to estimate the dN/dS ratios on each branch of a phylogenetic tree"

Lineage-speci!c dN/dS Maybe positive selection only occurred in this gene at this point in time?"
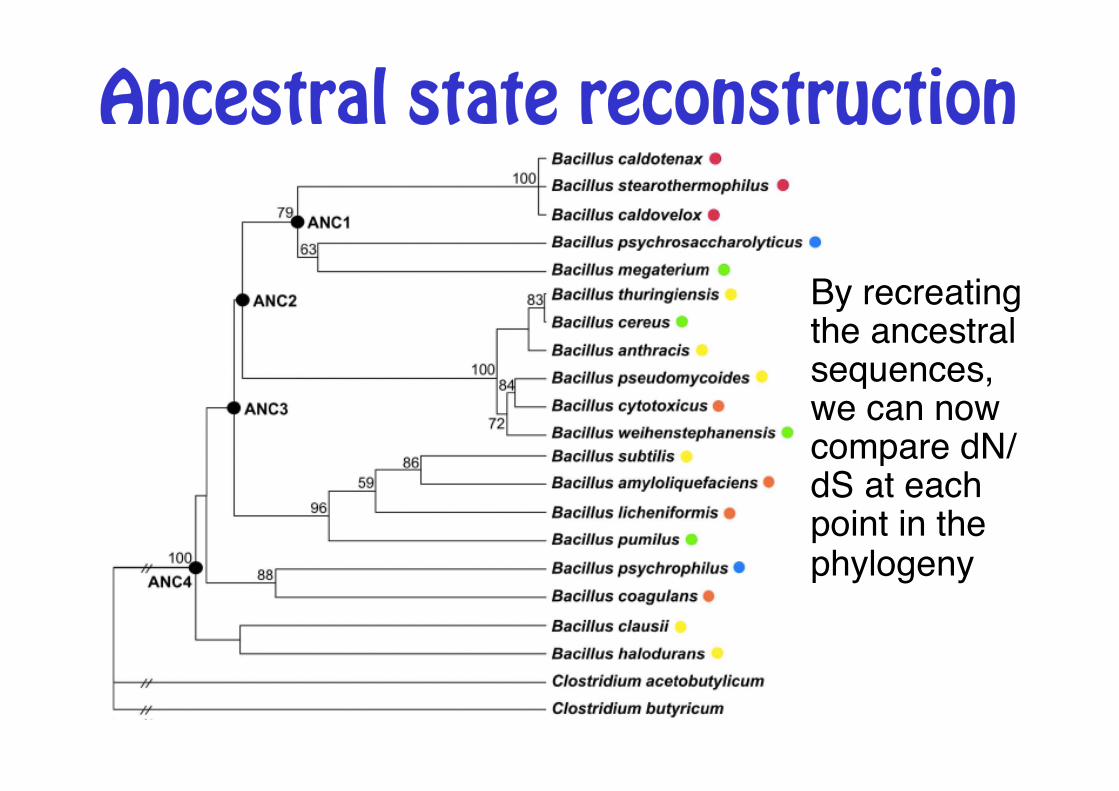
Ancestral state reconstruction
By recreating the ancestral sequences, we can now compare dN/dS at each point in the phylogeny"

Limitations of phylogenetic dN/dS for positive selection
• Similar to those for the simple dN/dS ratio "• In addition, ancestral state reconstruction is
not simple, and the dN/dS relies on it being accurate, which it might not be"

Selective sweeps • When a mutation is selected, it will take
along neighbouring SNPs that are linked"• This is called a “selective sweep”, and the
the neighbouring SNPs are said to “hitchhike” along with the positively selected mutation"
• The extent of DNA that hitchhikes along with the selected mutation is governed by meiotic recombination"
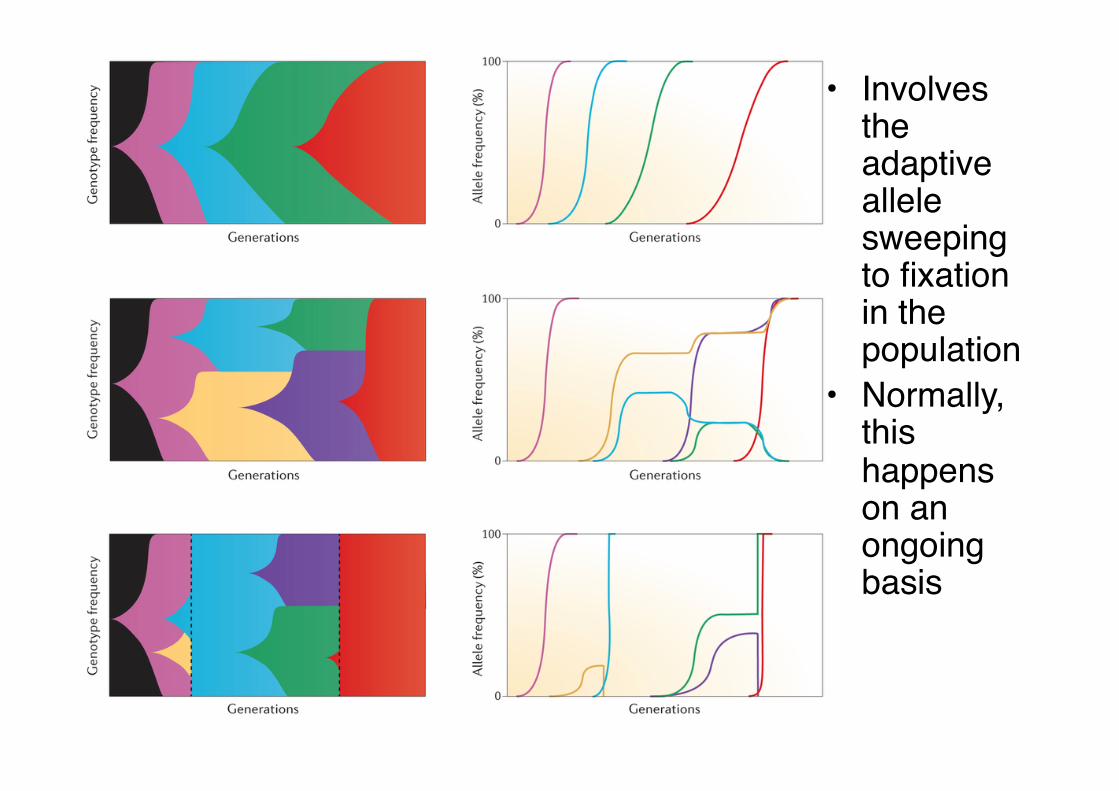
• Involves the adaptive allele sweeping to fixation in the population"
• Normally, this happens on an ongoing basis"

Selective sweeps Positive selection for green mutation
Neutral evolution and selection against red mutations

Selective sweeps Results in a signature of decreased genetic variation around the positively-selected mutation

Detecting selective sweeps • Selective sweeps can be detected by
scanning the genome in a population for regions of low polymorphism/high linkage disequilibrium"
• However, limitations arise because other factors can also result in this pattern"
• In particular, variation in the rate of meiotic recombination across the genome can cause dramatic differences in the level of linkage disequilibrium "

Horizontal gene transfer • One unusual way a genome can evolve is
by picking up bits from other species’ genomes"
• Known as horizontal gene transfer (HGT) – exchange of genetic material between non parent-offspring individuals"
• HGT is very common in bacteria – they seem to swap parts of their genome with abandon – but is much more controversial in eukaryote evolution "

Horizontal gene transfer

Horizontal gene transfer Soucy et al, 2015 Nature Reviews Genetics 16: 472-482

Mitochondria/chloroplasts • Mitochondria and chloroplasts are examples
of HGT"• They are bacterial endosymbionts of
eukaryote cells"• In both cases many genes have been
horizontally transferred from mitochondria and chloroplasts into the nuclear genome"
• Because of the endosymbiotic relationship, these are considered special cases"

Detecting HGT • The gold standard for detecting HGT is
phylogenetic incongruence"• When the phylogenetic tree for a region of
the genome disagrees with the “established” species phylogeny"
• Such incongruence is consistent with that region having been horizontally transferred"
• However, other explanations are possible: e.g. frequent gene gains/losses, or rapid evolution of a gene in one lineage"
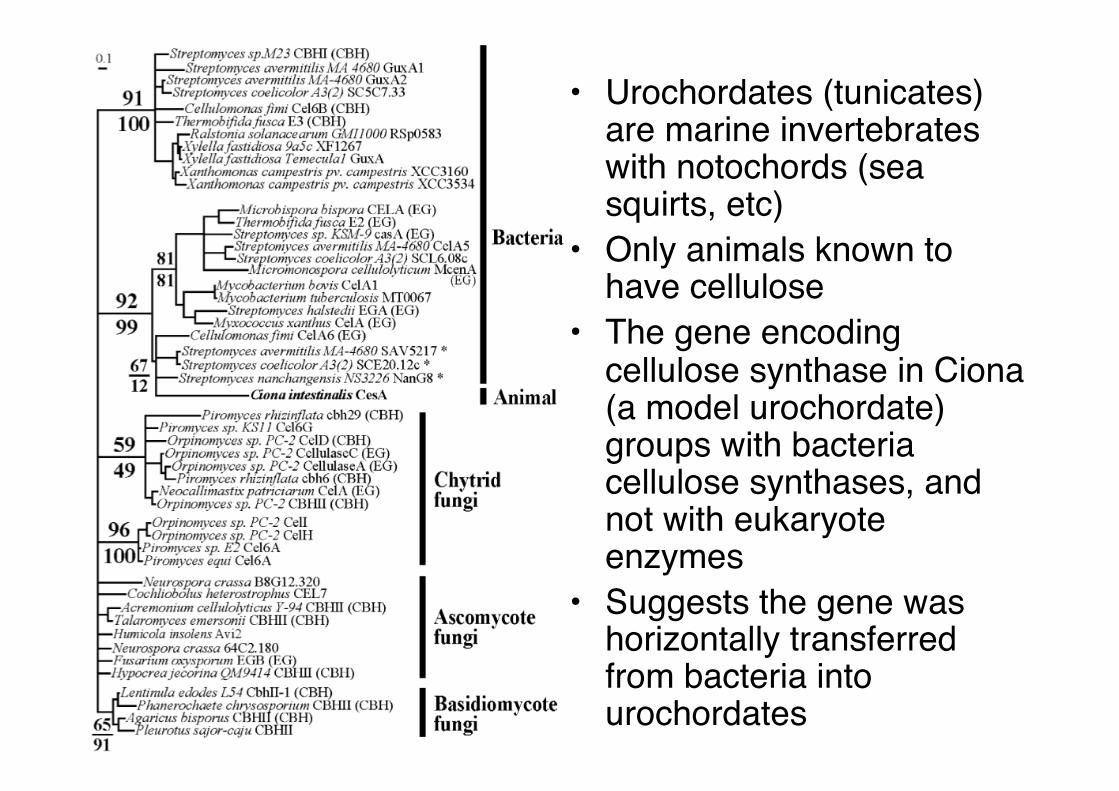
• Urochordates (tunicates) are marine invertebrates with notochords (sea squirts, etc)"
• Only animals known to have cellulose"
• The gene encoding cellulose synthase in Ciona (a model urochordate) groups with bacteria cellulose synthases, and not with eukaryote enzymes"
• Suggests the gene was horizontally transferred from bacteria into urochordates"

Care is needed • The original paper describing the human
genome in 2001 found 113 genes that are likely to have been horizontally transferred to vertebrates from bacteria"
• Based on BLAST searches where the closest match outside of vertebrates was a bacterial gene"
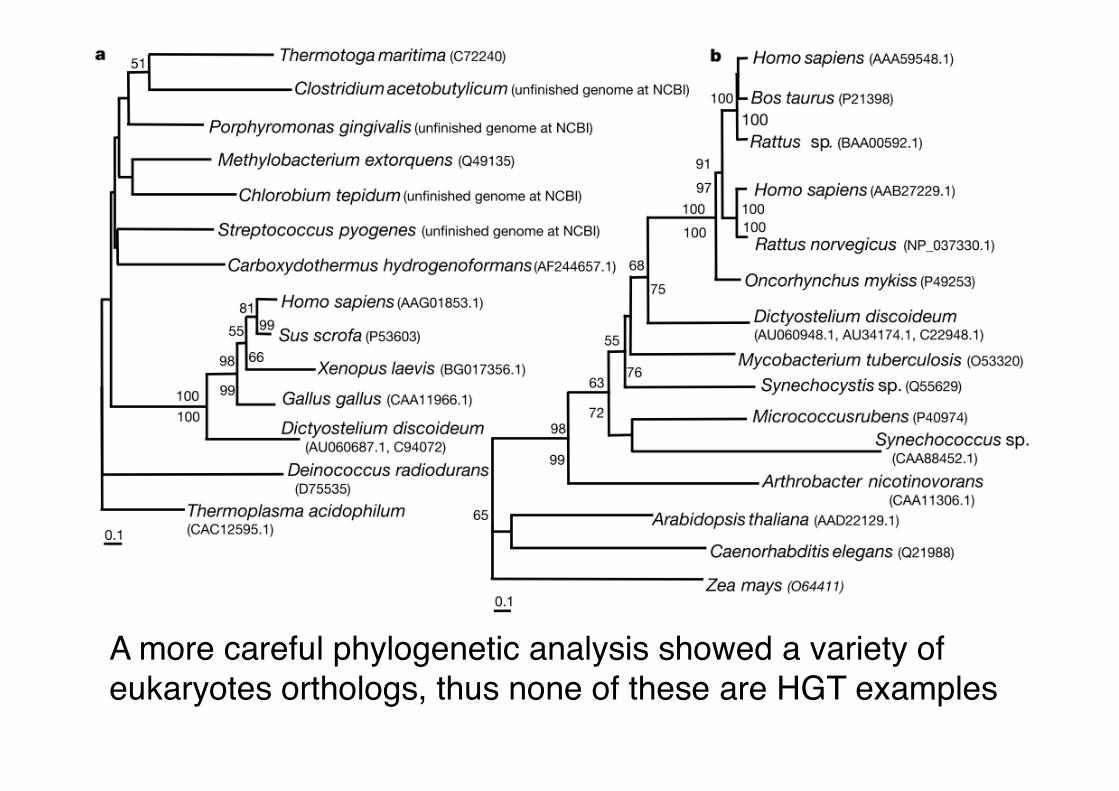
A more careful phylogenetic analysis showed a variety of eukaryotes orthologs, thus none of these are HGT examples"

• A gene that is evolving quickly in one lineage but not others will show up as a long branch in a phylogenetic tree"
Long branch problems
http://mesquiteproject.org/mesquite1.0/Mesquite_Folder/docs/mesquite/studies/study002/index.html

• These often falsely group together in phylogenetic trees, or otherwise end up in weird places"
Long branch problems

• Problems like long branches mean simple phylogenetic incongruence can be misleading"
• Therefore, convincing evidence for HGT: the HGT recipient lineages nest within the donor lineages, to the exclusion of other lineages related to the recipient lineages"
Nested phylogenetic incongruence: gold standard!

• This pattern could result from a variety of causes, including, but not limited to, HGT"
Simple incongruence is not enough
Gene X species phylogeny
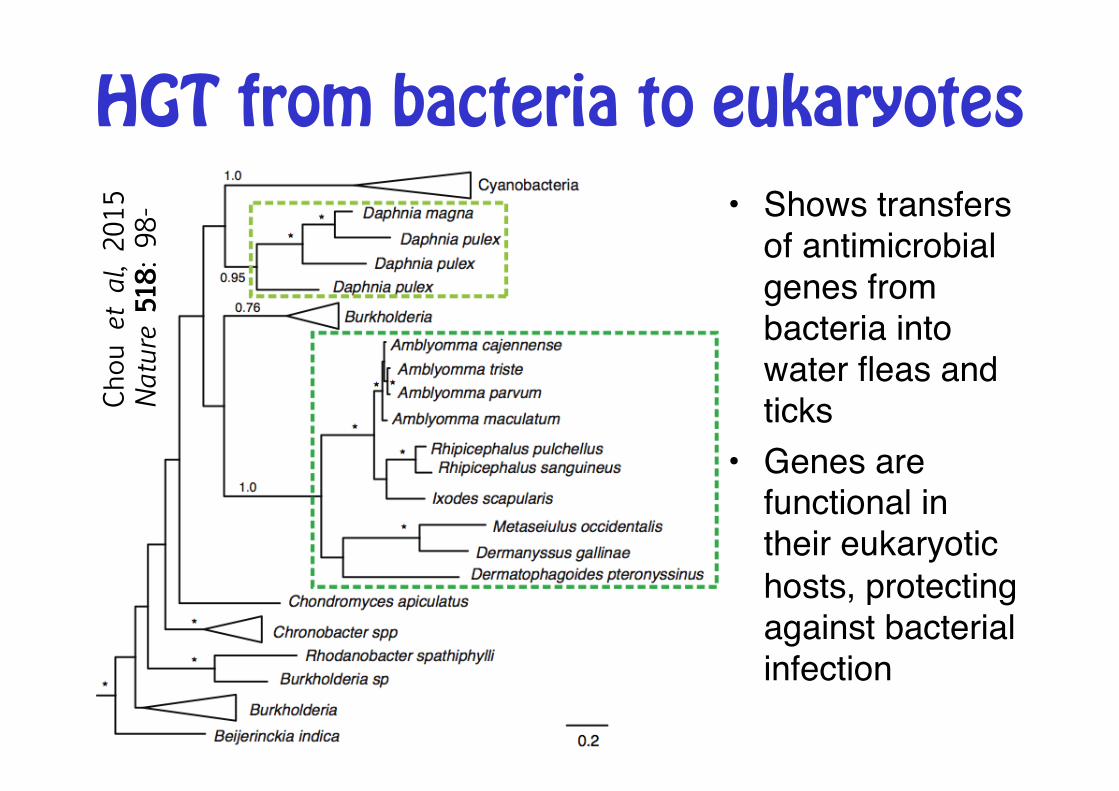
Chou
et al, 20
15
Nature 5
18: 98
- • Shows transfers of antimicrobial genes from bacteria into water fleas and ticks"
• Genes are functional in their eukaryotic hosts, protecting against bacterial infection"
HGT from bacteria to eukaryotes

• HGT of genes involved in breakdown of seaweed cell walls from algal parasites to human gut microbes"
• So far, only found in Japanese people, giving them a unique ability to use seaweed as a food source!"
• Placental syncytin genes came from HGT of viral genes into mammals"
Final cool examples