lake biwa in aquatic - 《湖泊科学》 aquatic systems and environments lake biwa okuda ·...
TRANSCRIPT

123
S P R I N G E R B R I E F S I N B I O LO G Y
Noboru Okuda · Katsutoshi WatanabeKayoko Fukumori · Shin-ichi NakanoTakefumi Nakazawa
Biodiversity in Aquatic Systems and Environments Lake Biwa
Okuda · Watanabe · Fukum
ori

SpringerBriefs in Biology
For further volumes: http://www.springer.com/series/10121


Noboru Okuda • Katsutoshi Watanabe Kayoko Fukumori • Shin-ichi Nakano Takefumi Nakazawa
Biodiversity in Aquatic Systems and Environments
Lake Biwa

ISSN 2192-2179 ISSN 2192-2187 (electronic)ISBN 978-4-431-54149-3 ISBN 978-4-431-54150-9 (eBook) DOI 10.1007/978-4-431-54150-9 Springer Tokyo Heidelberg New York Dordrecht London
Library of Congress Control Number: 2013951142
© The Author(s) 2014 This work is subject to copyright. All rights are reserved by the Publisher, whether the whole or part of the material is concerned, specifi cally the rights of translation, reprinting, reuse of illustrations, recitation, broadcasting, reproduction on microfi lms or in any other physical way, and transmission or information storage and retrieval, electronic adaptation, computer software, or by similar or dissimilar methodology now known or hereafter developed. Exempted from this legal reservation are brief excerpts in connection with reviews or scholarly analysis or material supplied specifi cally for the purpose of being entered and executed on a computer system, for exclusive use by the purchaser of the work. Duplication of this publication or parts thereof is permitted only under the provisions of the Copyright Law of the Publisher’s location, in its current version, and permission for use must always be obtained from Springer. Permissions for use may be obtained through RightsLink at the Copyright Clearance Center. Violations are liable to prosecution under the respective Copyright Law. The use of general descriptive names, registered names, trademarks, service marks, etc. in this publication does not imply, even in the absence of a specifi c statement, that such names are exempt from the relevant protective laws and regulations and therefore free for general use. While the advice and information in this book are believed to be true and accurate at the date of publication, neither the authors nor the editors nor the publisher can accept any legal responsibility for any errors or omissions that may be made. The publisher makes no warranty, express or implied, with respect to the material contained herein.
Printed on acid-free paper
Springer is part of Springer Science+Business Media (www.springer.com)
Noboru Okuda Center for Ecological Research Kyoto University Otsu, Japan
Kayoko Fukumori Section of Integrative Biology The University of Texas at Austin Austin , TX , USA
Takefumi Nakazawa Department of Life Sciences The College of Biosciences
and BiotechnologyNational Cheng Kung University Tainan , Taiwan
Katsutoshi Watanabe Department of Zoology Graduate School of ScienceKyoto University Kyoto , Japan
Shin-ichi NakanoCenter for Ecological ResearchKyoto UniversityOtsu, Japan

v
Foreword
While the twentieth century was the century when researchers tried to discover “the general basic principles of organisms,” the twenty-fi rst century is expected to be the century when researchers try to understand “the evolution and diversity of organ-isms” on the basis of such general principles of organisms by integrating various disciplines such as morphology, physiology, and ecology.
The chief diffi culty in studying “the evolution and diversity of organisms” lies in the fact that we have to consider factors at various levels ranging from the genome to the ecosystem. As taking various factors into account may cause a loss of focus, traditional studies have been restricted to analyzing only one individual level or fac-tor. However, unfortunately, the current research and education system based on such a compartmentalized approach is inadequate for incisively studying “the evo-lution and diversity of organisms.”
In order to solve these problems, we should strongly emphasize the necessity for joint studies and integration of the education programs between micro-level biology (genomic science, evolutionary developmental biology, genetic science, cell biol-ogy, neurobiology, molecular physiology, and molecular evolutionary studies) and macro-level biology (primatology, anthropology, ethology, environmental biology, evolutionary taxonomy, and so on) to young biologists. We launched a new educa-tion program in Kyoto University, called “Global COE program for Evolution and Biodiversity Research” to promote such integrative studies at various levels, and have succeeded in initiating novel currents of study of biodiversity that led rather than followed those in the rest of the world. To this aim, we decided to publish six books in “SpringerBriefs in Biology” which we hope will stimulate interest in such novel approaches on the evolution and diversity of organisms in the world and among young biologists.
This book covers a broad range of aquatic biology from molecular biology, through ecology, to paleobiology in Lake Biwa, a fascinating ancient lake that is also famed as a biodiversity hotspot. It will be a useful resource not only for researchers and technicians but also for graduate students who want to learn aquatic science. Naturalists and nature lovers can enjoy here a virtual tour of the world of biodiversity in this ancient lake. That biodiversity ranges over vast differences of

vi
scale: We describe the latest molecular genetics techniques that have revealed a relationship between the lake’s microbial diversity and its ecosystem functioning. We also describe the evolutionary history of the lake’s biodiversity, especially fi sh diversity, as well as laboratory experiments that revealed that diversity of fi sh feed-ing morphology affects the body size of plankton communities and consequently the overall ecosystem properties.
Unfortunately, Lake Biwa is also known as a tragic lake which has suffered from a variety of human disturbances, such as eutrophication, habitat destruction, exotic species introduction, and global warming, and that is now threatened by biodiver-sity loss and ecosystem deterioration. This book thus will appeal to conservationists and policy makers who are interested in biodiversity conservation and ecosystem management. As well, it will provide an opportunity for all readers to think deeply about the need to conserve the earth’s biodiversity.
Kiyokazu Agata Professor, Department of Biophysics, Kyoto University
Project Leader of Kyoto University Global COE program“Evolution and Biodiversity”
Foreword

vii
Pref ace
Two decades have passed since the Earth Summit, at which the Convention on Biological Diversity (CBD) came into force to encourage the conservation of bio-logical diversity on the earth and the fair and equitable sharing of benefi ts arising from the sustainable use of its components. At present, the loss of biological diver-sity has progressed at an accelerating rate and is particularly severe in freshwater ecosystems, in which environmental alterations have been extreme due to drastic changes in land-use patterns. For most of us, it is intuitively understandable why fi shery resources should be conserved, because they provide humankind with ben-efi ts such as food sources, i.e., a provisioning service from aquatic ecosystems. However, it is not so easy to answer the question of why fi sh diversity is necessary for human well-being. We also have little knowledge on how diversity of other aquatic biota, especially of microbes, affects ecosystem properties such as organic production, water purifi cation, and nutrient cycling. Or rather, we human beings have no inherent sense to discern the invisible nature of diversity in a microscopic world. For aquatic biologists, therefore, it is a great challenge to perceive the entity of biological diversity in aquatic ecosystems, trace its evolutionary history, and dis-cover the ecological consequences of its evolutionary diversifi cation, toward the integration of biodiversity sciences.
Here we introduce the latest topics of ecological and evolutionary research on aquatic biological diversity from bacteria to fi shes, with special reference to the case of Lake Biwa. Lake Biwa, in western Japan, is an ancient lake with a geological history of four million years, making it the third oldest lake in the world. It is called a biodiversity hotspot, where 1,769 aquatic species including 61 endemics are recorded, providing a rare opportunity to study the evolutionary diversifi cation of aquatic biota and its ecological consequences. In this book, we fi rst narrate the evo-lutionary history of biological diversity, especially of fi sh diversity in this lake, focusing on its paleogeography and molecular phylogeny (Chap. 1 by Katsutoshi Watanabe). The reader will learn how physical features of the lake have promoted evolutionary diversifi cation of feeding functions in fi shes.

viii
Functional diversifi cation of fi shes, which play important ecological roles as keystone predators of lake ecosystems, can sometimes be a major driver for altering an entire lake community through their top-down trophic cascading effects (Chap. 2 by Noboru Okuda and Kayoko Fukumori). We demonstrate that predator functional diversity has the potential for altering species composition, size structure, and tro-phic relations of an entire biological community, which in turn affects ecosystem functioning such as energy fl ows and nutrient cycling.
In lakes, the leading actors in ecosystem functioning, such as production and mineralization of organic matter, are microbes, which are often regarded as ecosys-tem engineers altering aquatic environments because of their overwhelming domi-nance in number and mass. A few decades ago, the diversity of aquatic microbes in nature remained unexplored because a large portion of bacteria never grow under cultivation to be isolated for taxonomic identifi cation. However, the recent explo-sion of advances in molecular techniques has vigorously encouraged microbiolo-gists to characterize genetic and functional diversity of in situ microbial communities (Chap. 3 by Shin-ichi Nakano).
While respecting the blessings of invaluable biological diversity in lake ecosys-tems, we have to warn that modern human activities have brought a crisis of biodi-versity loss in lakes worldwide, which will result in deterioration of ecosystem functioning and thus of ecosystem services. In the face of such human disturbances, ecosystem alterations are often catastrophic, a phenomenon known as ecological regime shift. To understand how human-mediated environmental changes can induce regime shifts in lake ecosystems and to evaluate how their biological diver-sity, especially intraspecifi c phenotypic diversity, affects ecosystem resilience, i.e., dampening the incidence and intensity of regime shifts, a promising approach is construction of theoretical models, which is considered a kind of thought experi-ment to test probable future scenarios (Chap. 4 by Takefumi Nakazawa).
Again, we stress that Lake Biwa can be an ideal model system in which to prac-tice biodiversity conservation with the promotion of ecological and evolutionary understanding of biological diversity in lake ecosystems. This opportunity exists because Lake Biwa has experienced a variety of serious environmental problems that are shared by many other lakes worldwide, and also because the lake currently shows symptoms of recovery from the biodiversity loss and ecosystem deteriora-tion, recognized only by our persistent efforts in biodiversity research. Through this book, the reader will learn how the evolutionary diversifi cation of aquatic organ-isms interplays with the ecology of their populations and communities, and vice versa, and why we should conserve biological diversity at different levels of bio-logical organization from genes to ecosystems.
Otsu , Japan Noboru Okuda
Preface

ix
Contents
1 Origin and Diversification of Freshwater Fishes in Lake Biwa .............. 11.1 Introduction: Lake Biwa and Its Fish Fauna ........................................ 11.2 A Brief Review on the Origins and Evolution
of Freshwater Fishes ............................................................................ 81.3 Molecular Phylogenetic Approaches ................................................... 10
1.3.1 Origin of Endemic Lineages .................................................... 101.3.2 Population Establishment by Endemic
and Non-endemic Species ........................................................ 141.4 Conclusions and Perspectives .............................................................. 15References ..................................................................................................... 17
2 Predator Diversity Changes the World: From Gene to Ecosystem ........ 212.1 Why is Biodiversity Important? ........................................................... 212.2 Biodiversity at Three Levels ................................................................ 222.3 What is Phenotypic Polymorphism? .................................................... 222.4 Phenotypic Polymorphism in Aquatic Ecosystems ............................. 23
2.4.1 Trophic Polymorphism and Adaptive Divergence ................... 232.4.2 Ecological Speciation and Species Pair ................................... 252.4.3 The Ancient Lake Biwa as an Evolutionary Hot Spot ............. 262.4.4 Life History Polymorphism ..................................................... 272.4.5 Rapid Evolution of Trophic Polymorphism ............................. 282.4.6 Genetic Mechanisms of Trophic Polymorphism ..................... 30
2.5 Niche Construction .............................................................................. 312.6 Predator Diversity and Ecosystem Functioning ................................... 33
2.6.1 Mesocosm Experiments ........................................................... 352.6.2 Food Web Properties as a Universal Measure ......................... 37
2.7 Eco-Evolutionary Feedbacks ............................................................... 402.7.1 Feedback Loops ....................................................................... 412.7.2 Community Genomic Dynamics ............................................. 43
2.8 Why is Predator Diversity Important? ................................................. 44References ..................................................................................................... 45

x
3 Biodiversity Researches on Microbial Loop in Aquatic Systems ............ 513.1 Introduction .......................................................................................... 513.2 Towards More Complicated Microbial Food Web............................... 523.3 Detection of Microorganisms Using Molecular
Biological Techniques.......................................................................... 543.3.1 Biodiversity of Bacteria ........................................................... 543.3.2 Biodiversity of Protists ............................................................ 56
3.4 Recent Development in Studies on Food Chain Between Bacteria and Protists ............................................................. 57
3.5 Predation on Protists ............................................................................ 593.6 Biodiversity Researches on Microbial Loop in Lake Biwa ................. 593.7 Concluding Remarks ........................................................................... 61References ..................................................................................................... 62
4 A Dynamic Resilience Perspective Toward Integrated Ecosystem Management: Biodiversity, Landscape, and Climate .............................. 694.1 Introduction .......................................................................................... 694.2 Regime Shift and Resilience as Key Concepts
in Ecosystem Management .................................................................. 704.3 Regime Shifts Mediated by Ontogenetic Niche Shifts ........................ 724.4 Effects of Ecological Contexts on the Occurrence
of Regime Shifts Mediated by Ontogenetic Niche Shifts .................... 754.4.1 Food-Web Topology ................................................................ 764.4.2 Spatial Confi guration ............................................................... 784.4.3 Seasonality ............................................................................... 814.4.4 Other Factors............................................................................ 83
4.5 Toward Integrated Ecosystem Management ........................................ 854.5.1 From Static to Dynamic Resilience Perspectives .................... 854.5.2 Implications for Lake Ecosystem Management....................... 86
References ..................................................................................................... 88
Contents

1N. Okuda et al., Biodiversity in Aquatic Systems and Environments: Lake Biwa, SpringerBriefs in Biology, DOI 10.1007/978-4-431-54150-9_1, © The Author(s) 2014
Abstract Lake Biwa is the largest and oldest lake in Japan. It harbors more than a thousand animal and plant species, including 67 indigenous freshwater fi sh species/subspecies with 16 endemic or semi-endemic forms. Paleogeographical and paleonto-logical studies have revealed that the location and environment of the lake have changed during its history of more than four million years. The current Lake Biwa, with its unique environments, was established 0.4 million years ago. Endemic fi shes in Lake Biwa are traditionally divided into old “relict species” and new “species that evolved in the lake,” the latter being assumed to have evolved through ecological adaptations to the present lake environment. However, recent molecular phylogenetic/population genetic analyses have revealed that many of the “species that evolved in the lake” have origins that are older than the present Lake Biwa, while other species likely evolved in the present environment through ecological adaptation. On the other hand, many fi sh populations that inhabit Lake Biwa, irrespective of their origin, show similar patterns involving recent population expansions that occurred several tens of thousands of years ago. Extensive community level analyses of “lacustrine syn-dromes” that are found in Lake Biwa are necessary based on morphology, physiology, phylogenetics, population genetics, and also new “omics” approaches.
Keywords Adaptation • Ancient lake • Divergence time • Ecological speciation • Endemic species • Fossil records • Mitochondrial DNA (mtDNA) • Phylogeny • Phylogeography
1.1 Introduction: Lake Biwa and Its Fish Fauna
Lake Biwa, located in central Honshu Island, Japan (35°20′N, 136°10′E, 84.4 m above sea level), is a representative ancient lake in East Asia. It is the largest lake in Japan in terms of surface area (670.3 km 2 ) and volume (27.5 km 3 ), and is also the oldest lake in Japan, with its origin dating back to four million years ago (Mya)
Chapter 1 Origin and Diversifi cation of Freshwater Fishes in Lake Biwa
2
(Yokoyama 1984 ; Kawabe 1994 ). Like other ancient lakes, Lake Biwa harbors many species (>1,000), including diverse endemic animal and plant taxa (~60 spe-cies/subspecies; Nishino 2003 ; Nishino and Hamabata 2005 ). Thus, it has been rec-ognized as a central research area for studying the biodiversity and speciation of freshwater organisms in Japan (Kawanabe 1996 ; Rossiter 2000 ).
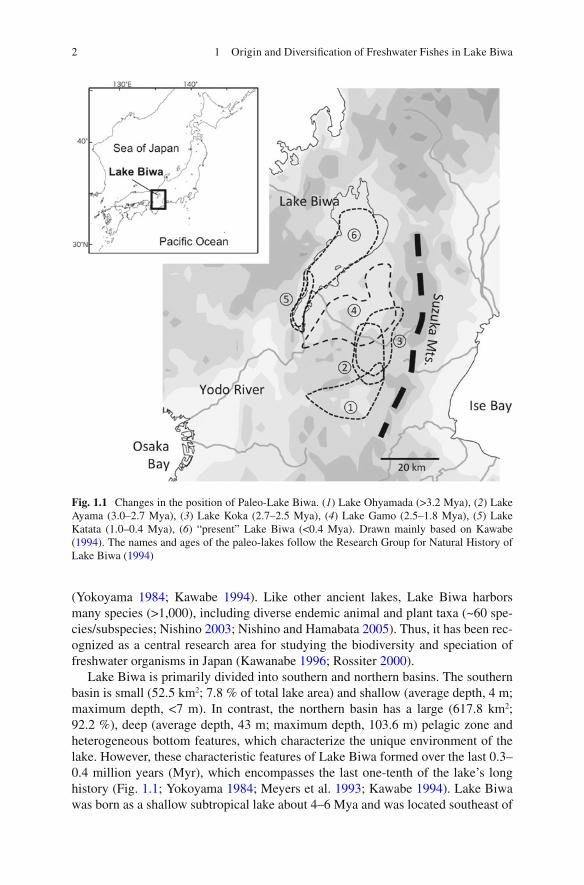
Lake Biwa is primarily divided into southern and northern basins. The southern basin is small (52.5 km 2 ; 7.8 % of total lake area) and shallow (average depth, 4 m; maximum depth, <7 m). In contrast, the northern basin has a large (617.8 km 2 ; 92.2 %), deep (average depth, 43 m; maximum depth, 103.6 m) pelagic zone and heterogeneous bottom features, which characterize the unique environment of the lake. However, these characteristic features of Lake Biwa formed over the last 0.3–0.4 million years (Myr), which encompasses the last one-tenth of the lake’s long history (Fig. 1.1 ; Yokoyama 1984 ; Meyers et al. 1993 ; Kawabe 1994 ). Lake Biwa was born as a shallow subtropical lake about 4–6 Mya and was located southeast of
Fig. 1.1 Changes in the position of Paleo-Lake Biwa. ( 1 ) Lake Ohyamada (>3.2 Mya), ( 2 ) Lake Ayama (3.0–2.7 Mya), ( 3 ) Lake Koka (2.7–2.5 Mya), ( 4 ) Lake Gamo (2.5–1.8 Mya), ( 5 ) Lake Katata (1.0–0.4 Mya), ( 6 ) “present” Lake Biwa (<0.4 Mya). Drawn mainly based on Kawabe ( 1994 ). The names and ages of the paleo-lakes follow the Research Group for Natural History of Lake Biwa ( 1994 )
1 Origin and Diversifi cation of Freshwater Fishes in Lake Biwa
3
its present position (the Ueno Basin, Mie Prefecture) (Kawabe 1994 ; Yokoyama 1984 ). After experiencing drastic geological changes (e.g., Yokoyama 1968 , 1984 ), the lake reformed about 1 Mya in the location of the current southern basin; the transition was followed by the formation of the northern basin (<0.4 Mya).
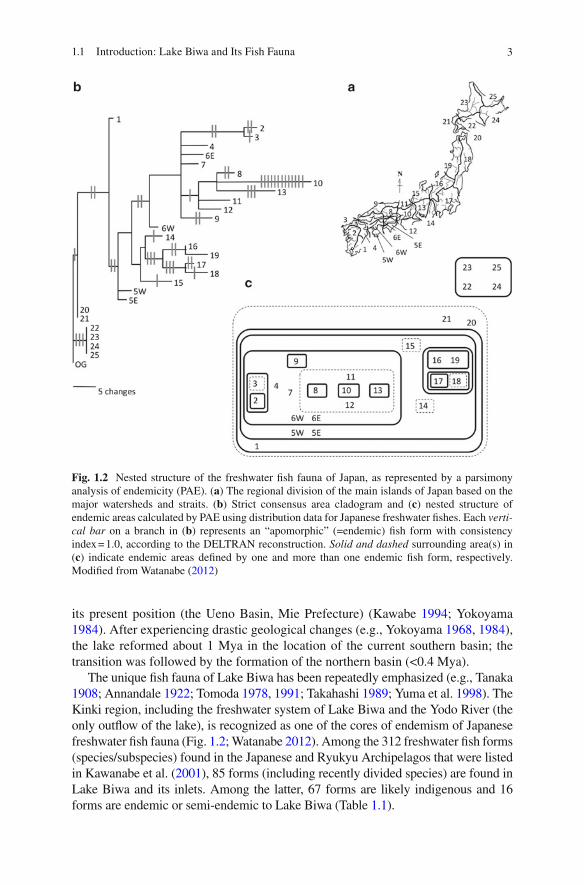
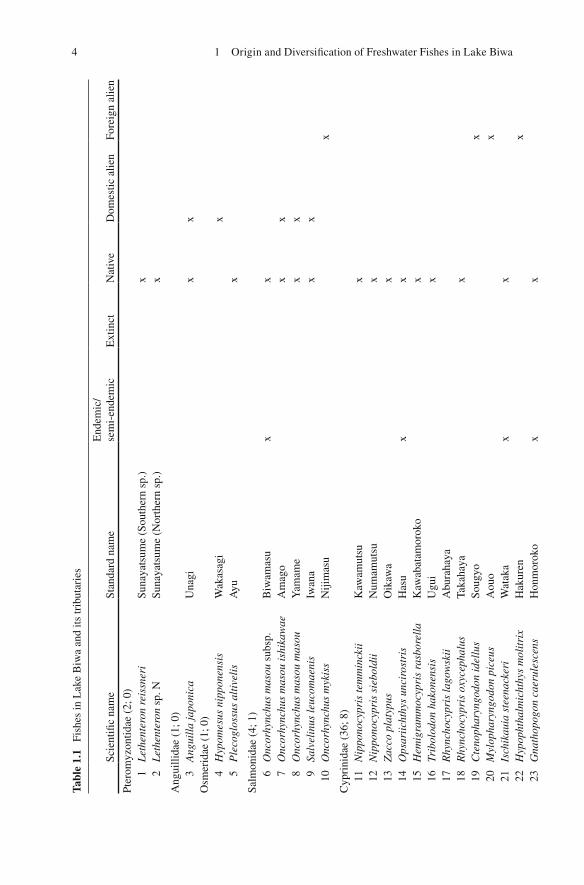
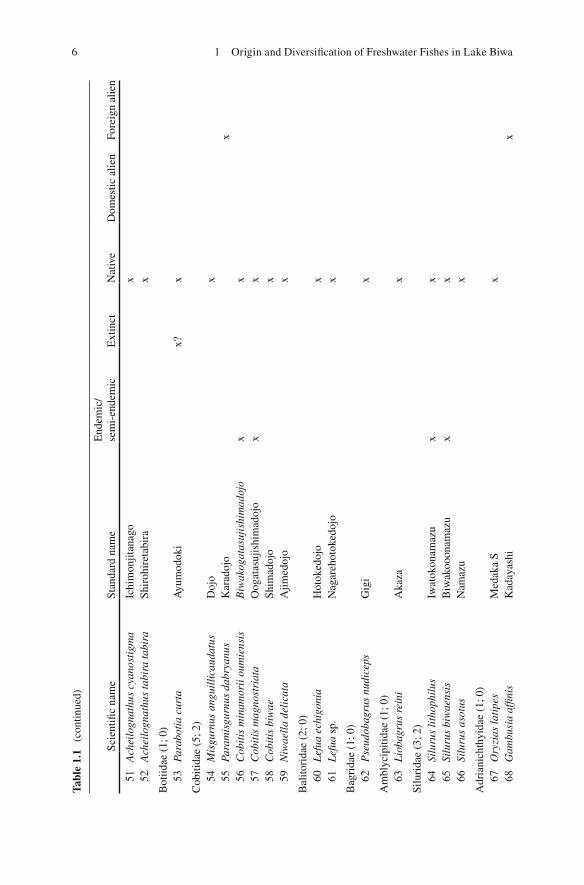
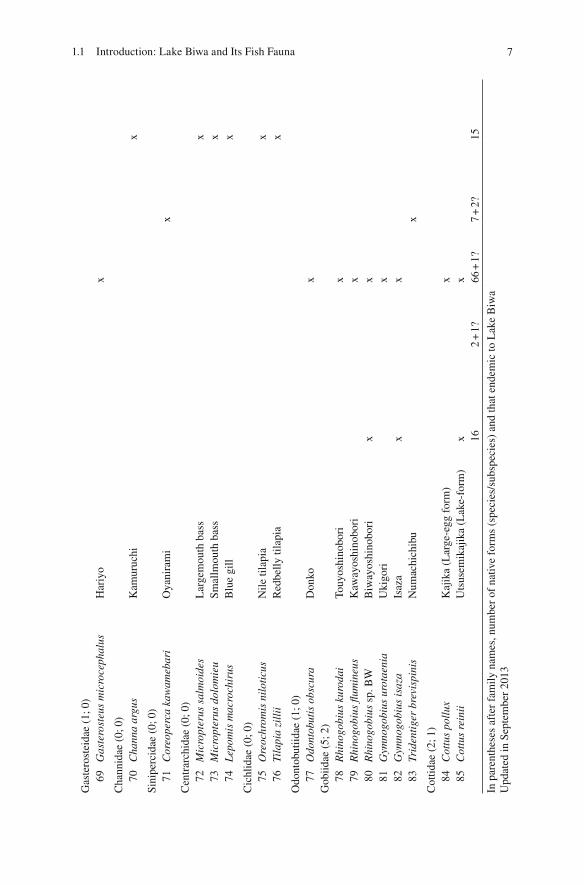
The unique fi sh fauna of Lake Biwa has been repeatedly emphasized (e.g., Tanaka 1908 ; Annandale 1922 ; Tomoda 1978 , 1991 ; Takahashi 1989 ; Yuma et al. 1998 ). The Kinki region, including the freshwater system of Lake Biwa and the Yodo River (the only outfl ow of the lake), is recognized as one of the cores of endemism of Japanese freshwater fi sh fauna (Fig. 1.2 ; Watanabe 2012 ). Among the 312 freshwater fi sh forms (species/subspecies) found in the Japanese and Ryukyu Archipelagos that were listed in Kawanabe et al. ( 2001 ), 85 forms (including recently divided species) are found in Lake Biwa and its inlets. Among the latter, 67 forms are likely indigenous and 16 forms are endemic or semi-endemic to Lake Biwa (Table 1.1 ).
Fig. 1.2 Nested structure of the freshwater fi sh fauna of Japan, as represented by a parsimony analysis of endemicity (PAE). ( a) The regional division of the main islands of Japan based on the major watersheds and straits. ( b) Strict consensus area cladogram and ( c ) nested structure of endemic areas calculated by PAE using distribution data for Japanese freshwater fi shes. Each verti-cal bar on a branch in ( b) represents an “apomorphic” (=endemic) fi sh form with consistency index = 1.0, according to the DELTRAN reconstruction. Solid and dashed surrounding area(s) in ( c) indicate endemic areas defi ned by one and more than one endemic fi sh form, respectively. Modifi ed from Watanabe ( 2012 )
1.1 Introduction: Lake Biwa and Its Fish Fauna
4
Tabl
e 1.
1 Fi
shes
in L
ake
Biw
a an
d its
trib
utar
ies
Scie
ntifi
c na
me
Stan
dard
nam
e E
ndem
ic/
sem
i-en
dem
ic
Ext
inct
N
ativ
e D
omes
tic a
lien
Fore
ign
alie
n
Pter
omyz
ontid
ae (
2; 0
)
1 Le
then
tero
n re
issn
eri
Suna
yats
ume
(Sou
ther
n sp
.)
x
2 Le
then
tero
n sp
. N
Suna
yats
ume
(Nor
ther
n sp
.)
x
Ang
uilli
dae
(1; 0
)
3 A
ngui
lla
japo
nica
U
nagi
x
x O
smer
idae
(1;
0)
4
Hyp
omes
us n
ippo
nens
is
Wak
asag
i x
5
Ple
cogl
ossu
s al
tive
lis
Ayu
x
Salm
onid
ae (
4; 1
)
6 O
ncor
hync
hus
mas
ou s
ubsp
. B
iwam
asu
x x
7
Onc
orhy
nchu
s m
asou
ishi
kaw
ae
Am
ago
x x
8
Onc
orhy
nchu
s m
asou
mas
ou
Yam
ame
x x
9
Salv
elin
us le
ucom
aeni
s Iw
ana
x x
10
O
ncor
hync
hus
myk
iss
Niji
mas
u x
Cyp
rini
dae
(36;
8)
11
N
ippo
nocy
pris
tem
min
ckii
K
awam
utsu
x
12
N
ippo
nocy
pris
sie
bold
ii
Num
amut
su
x
13
Zacc
o pl
atyp
us
Oik
awa
x
14
Ops
arii
chth
ys u
ncir
ostr
is
Has
u x
x
15
Hem
igra
mm
ocyp
ris
rasb
orel
la
Kaw
abat
amor
oko
x
16
Trib
olod
on h
akon
ensi
s U
gui
x
17
Rhy
ncho
cypr
is la
gow
skii
A
bura
haya
18
Rhy
ncho
cypr
is o
xyce
phal
us
Taka
haya
x
19
C
teno
phar
yngo
don
idel
lus
Soug
yo
x
20
Myl
opha
ryng
odon
pic
eus
Aou
o x
21
Is
chik
auia
ste
enac
keri
W
atak
a x
x
22
Hyp
opht
halm
icht
hys
mol
itri
x H
akur
en
x
23
Gna
thop
ogon
cae
rule
scen
s H
onm
orok
o x
x
1 Origin and Diversifi cation of Freshwater Fishes in Lake Biwa

5
24
G
nath
opog
on e
long
atus
Ta
mor
oko
x x?
25
Pun
gtun
gia
herz
i M
ugits
uku
x
26
Pse
udor
asbo
ra p
arva
M
otsu
go
x x
27
Sa
rcoc
heil
icht
hys
biw
aens
is
Abu
rahi
gai
x x
28
Sa
rcoc
heil
icht
hys
vari
egat
us
mic
rooc
ulus
B
iwah
igai
x
x
29
P
seud
ogob
io e
soci
nus
Kam
atsu
ka
x
30
Abb
otti
na r
ivul
aris
T
such
ifuk
i x?
x?
31
Biw
ia z
ezer
a Z
ezer
a x
32
B
iwia
yod
oens
is
Yod
ozez
era
x
33
Squa
lidu
s ch
anka
ensi
s bi
wae
Su
gom
orok
o x
x
34
Squa
lidu
s ch
anka
ensi
s ts
uchi
gae
Kou
raim
orok
o x
35
Sq
uali
dus
japo
nicu
s ja
poni
cus
Dem
emor
oko
x
36
Squa
lidu
s gr
acil
is g
raci
lis
Itom
orok
o x
37
H
emib
arbu
s ba
rbus
N
igoi
x
38
H
emib
arbu
s la
beo
Kor
aini
goi
x
39
Hem
ibar
bus
long
iros
tris
Z
unag
anig
oi
x
40
Cyp
rinu
s ca
rpio
(na
tive
type
) K
oi (
wild
type
) x
41
C
ypri
nus
carp
io (
intr
oduc
ed)
Koi
x
42
C
aras
sius
aur
atus
gra
ndoc
ulis
N
igor
obun
a x
x
43
Car
assi
us c
uvie
ri
Gen
goro
buna
x
x
44
Car
assi
us a
urat
us la
ngsd
orfi i
G
inbu
na
x
45
Tana
kia
lanc
eola
ta
Yar
itana
go
x
46
Tana
kia
lim
bata
A
bura
bote
x
47
R
hode
us o
cell
atus
oce
llat
us
Tair
ikub
arat
anag
o x
48
R
hode
us o
cell
atus
kur
umeu
s N
ippo
nbar
atan
ago
x x
49
A
chei
logn
athu
s rh
ombe
us
Kan
ehir
a x
50
A
chei
logn
athu
s lo
ngip
inni
s It
asen
para
x
x
(con
tinue
d)
1.1 Introduction: Lake Biwa and Its Fish Fauna
6
Scie
ntifi
c na
me
Stan
dard
nam
eE
ndem
ic/
sem
i-en
dem
icE
xtin
ctN
ativ
eD
omes
tic a
lien
Fore
ign
alie
n
51
A
chei
logn
athu
s cy
anos
tigm
a Ic
him
onjit
anag
o x
52
A
chei
logn
athu
s ta
bira
tabi
ra
Shir
ohir
etab
ira
x
Bot
iidae
(1;
0)
53
Pa
rabo
tia
curt
a A
yum
odok
i x?
x
Cob
itida
e (5
; 2)
54
M
isgu
rnus
ang
uill
icau
datu
s D
ojo
x
55
Para
mis
gurn
us d
abry
anus
K
arad
ojo
x
56
Cob
itis
min
amor
ii o
umie
nsis
B
iwak
ogat
asuj
ishi
mad
ojo
x x
57
C
obit
is m
agno
stri
ata
Oog
atas
ujis
him
adoj
o x
x
58
Cob
itis
biw
ae
Shim
adoj
o x
59
N
iwae
lla
deli
cata
A
jimed
ojo
x
Bal
itori
dae
(2; 0
)
60
Lefu
a ec
higo
nia
Hot
oked
ojo
x
61
Lefu
a sp
. N
agar
ehot
oked
ojo
x
Bag
rida
e (1
; 0)
62
P
seud
obag
rus
nudi
ceps
G
igi
x
Am
blyc
ipiti
dae
(1; 0
)
63
Liob
agru
s re
ini
Aka
za
x
Silu
rida
e (3
; 2)
64
Si
luru
s li
thop
hilu
s Iw
atok
onam
azu
x x
65
Si
luru
s bi
wae
nsis
B
iwak
ooon
amaz
u x
x
66
Silu
rus
asot
us
Nam
azu
x
Adr
iani
chth
yida
e (1
; 0)
67
O
ryzi
as la
tipe
s M
edak
a S
x
68
Gam
busi
a af
fi nis
K
aday
ashi
x
Tabl
e 1.
1 (c
ontin
ued)
1 Origin and Diversifi cation of Freshwater Fishes in Lake Biwa
7
Gas
tero
stei
dae
(1; 0
)
69
Gas
tero
steu
s m
icro
ceph
alus
H
ariy
o x
Cha
nnid
ae (
0; 0
)
70
Cha
nna
argu
s K
amur
uchi
x
Sini
perc
idae
(0;
0)
71
C
oreo
perc
a ka
wam
ebar
i O
yani
ram
i x
Cen
trar
chid
ae (
0; 0
)
72
Mic
ropt
erus
sal
moi
des
Lar
gem
outh
bas
s x
73
M
icro
pter
us d
olom
ieu
Smal
lmou
th b
ass
x
74
Lepo
mis
mac
roch
irus
B
lue
gill
x
Cic
hlid
ae (
0; 0
)
75
Ore
ochr
omis
nil
otic
us
Nile
tila
pia
x
76
Tila
pia
zill
ii
Red
belly
tila
pia
x
Odo
ntob
utiid
ae (
1; 0
)
77
Odo
ntob
utis
obs
cura
D
onko
x
Gob
iidae
(5;
2)
78
R
hino
gobi
us k
urod
ai
Touy
oshi
nobo
ri
x
79
Rhi
nogo
bius
fl um
ineu
s K
away
oshi
nobo
ri
x
80
Rhi
nogo
bius
sp.
BW
B
iway
oshi
nobo
ri
x x
81
G
ymno
gobi
us u
rota
enia
U
kigo
ri
x
82
Gym
nogo
bius
isaz
a Is
aza
x x
83
Tr
iden
tige
r br
evis
pini
s N
umac
hich
ibu
x
Cot
tidae
(2;
1)
84
C
ottu
s po
llux
K
ajik
a (L
arge
-egg
for
m)
x
85
Cot
tus
rein
ii
Uts
usem
ikaj
ika
(Lak
e-fo
rm)
x x
16
2 +
1?
66 +
1?
7 +
2?
15
In p
aren
thes
es a
fter
fam
ily n
ames
, num
ber
of n
ativ
e fo
rms
(spe
cies
/sub
spec
ies)
and
that
end
emic
to L
ake
Biw
a U
pdat
ed in
Sep
tem
ber
2013
1.1 Introduction: Lake Biwa and Its Fish Fauna

8
At least for the primary freshwater fi shes, all of the species in Lake Biwa are categorized into the “warm-temperature group,” which is defi ned physiologically and biogeographically. Therefore, they are considered to consist of East Asian tem-perate faunal components. Some landlocked or freshwater forms of diadromous fi shes from the “cool temperature group” also occur in and around Lake Biwa (e.g., a lamprey, Lethenteron sp. “Northern forms,” some salmonids from Salvelinus and Oncorhynchus ).
The fi shes that occur in Lake Biwa use several types of habitats in and around the lake (Miura 1966 ; Tokui and Kawanabe 1984 ; Yuma et al. 1998 ; Fujita et al. 2008 ). Some use different habitats during different life-history stages and seasons. The species of Lake Biwa have been categorized into eight life-history types based on their main habitats and migration patterns (Hosoya 2005 ). Among the available habitats, the rocky bottom area and the pelagic zone of Lake Biwa are unique in Japanese fresh waters. Also, vast areas of sandy bottom provide unique habitats for some benthic/benthopelagic fi shes (e.g., Pseudogobio esocinus and Biwia zezera ). Lagoons, or shallow satellite lakes (called “Naiko”), around Lake Biwa were origi-nally very important habitats for many species, especially for reproduction and as nursery areas (Fujita et al. 2008 ). However, due to recent reclamation, the total area of these satellite lakes has decreased to about 10 % of the original area (32.4 km 2 in ~1925, 4.25 km 2 in 1995; Nishino 2005 ).
The number of endemic species (~60 animals and plants, including a dozen fi sh forms) is not very large when compared to levels of endemism in other ancient lakes throughout the world (e.g., >600 species in Lake Tanganyika; ~1,000 species in Lake Baikal; Martens 1997 ). The ratio of endemic forms in Lake Biwa is <10 % (24 % for fi sh) (Table 1.1 ). The taxonomic composition of the freshwater fi sh assemblage in Lake Biwa does not differ greatly from that of rivers in western Japan. There is no prominent taxonomic group in terms of endemism, as found in the African Great Lakes with cichlids or in Lake Baikal with sculpins. Therefore, endemism in Lake Biwa cannot be characterized as “species fl ocks” (e.g., Greenwood 1984 ). Instead, the assemblage of Lake Biwa consists of several sets of one or a few endemic and non-endemic forms from various taxonomic groups; in a manner of speaking, it is like a “salad bowl.”
1.2 A Brief Review on the Origins and Evolution of Freshwater Fishes
Several authors have discussed the origins of the endemic fi sh fauna of Lake Biwa. Fossil records are the only direct evidence of past faunal characteristics and their historical changes. Since Kodera and Nakamura ( 1976 ) discovered pharyngeal teeth of Cyprinus , fragmented fossil remains such as the pharyngeal teeth of cyprinids, crania and spines of siluriforms, and a jaw of a channid have been reported from Paleo-Lake Biwa (=the “Kobiwako” Group) (see below). Mollusk and diatom
1 Origin and Diversifi cation of Freshwater Fishes in Lake Biwa

9
fossils have also provided important keys for understanding changes (succession/alternation) in the aquatic biota of Paleo-Lake Biwa (Research Group for Natural History of Lake Biwa 1994 ).
Paleogeographical and paleontological data suggest that there was a large fresh-water system that consisted of a series of rivers and lakes in western Japan in the Pliocene (called the “second Setouchi river/lake system”). This system would have harbored a rich basal freshwater fi sh fauna that was closely related to the continental fauna. The initial stages of Paleo-Lake Biwa and its eastern neighbor, Paleo-Lake Tokai, were located at the eastern end of the second Setouchi river/lake system. The initial stage of Paleo-Lake Biwa (>3.2 Mya; “Lake Ohyamada”) was inferred to be a subtropical shallow lake, from which cyprinids, including Cyprininae, Xenocyprininae, Cultrinae, Gobioninae, Leuciscinae, and Acheilognathinae (Nakajima 1987 ), bagrid and silurid catfi shes (Kobayakawa 1994 ; Watanabe and Uyeno 1999 ), and a channid (Tanimoto and Okuyama 2003 ), have been discovered. Cyprinins, especially Cyprinus spp . , were the dominant components of this paleofauna (called the “lake of carp”) and xenocyprinins were also common, although the latter do not occur in present-day Japan (Nakajima 1994 ).
According to a quantitative comparison of cyprinid pharyngeal teeth among the for-mations of Paleo-Lake Biwa (Nakajima 1994 ), the paleo-ichthyofauna after Lake Ohyamada became rather simple. Cyprinin genera, Cyprinus and Carassius, were con-spicuously dominant in “Lake Ayama” (3.0–2.7 Mya), “Lake Koka (or Sayama)” (2.7–2.5 Mya), and “Lake Gamo” (2.5–1.8 Mya) (called the “lake of crusian carp”). Stratigraphical studies have suggested that large-scale standing waters disappeared after Lake Gamo was lost in this region. Then, “Lake Katata” formed around the present southern basin of Lake Biwa (1.0–0.4 Mya) (Fig. 1.1 ). It had a rich ichthyofauna includ-ing cyprinid Cyprininae, Xenocyprininae, Cultrinae, Gobioninae, and Leuciscinae (Nakajima 1994 ), and a bagrid catfi sh (Kobayakawa 1994 ). These fossil remains could include those related to current endemic species in Lake Biwa and they are the fi rst recognized endemics in this paleolake (the cyprinin Carassius cuvieri and the cultrin Ischikauia steenackeri ; Nakajima 1994 ). Fossil pharyngeal teeth similar to those of the specialized phytoplankton feeder C. cuvieri, which is endemic to Lake Biwa, exhibit several levels of specialization (Kodera 1985 ). Nakajima ( 1994 ) suggested that those ancestral forms would produce endemic species through ecological adaptations to the environments of the succeeding “present Lake Biwa.”
As above, the fossil record has contributed to our understanding of major fea-tures and succession in the ichthyofauna of Paleo-Lake Biwa that occurred over a period of 4 Myr (Tomoda 1991 ; Nakajima 1994 ). However, low taxonomic resolu-tion and possible bias in the sampling of fossils have prevented a detailed recon-struction of the origins and formation of the endemic fauna of Lake Biwa.
Another approach to understanding the origins of endemism in Lake Biwa involves comparisons of faunal and species traits (e.g., Ueno 1943 ; Kawanabe 1978 , 1996 ; Tomoda 1978 ). Kawanabe ( 1978 ) divided the endemic forms into two catego-ries: species that differentiated in situ and relic species. This division was probably based on the geographical distributions of putative ancestral forms and their eco-logical features. The former, “species that evolved in the lake,” were represented by
1.2 A Brief Review on the Origins and Evolution of Freshwater Fishes

10
the gudgeon Gnathopogon caerulescens (from G . elongatus ), Carassius cuvieri (from C . auratus ), Gymnogobius isaza (from G . urotaenia ), and so on (Kawanabe 1978 , 1996 ; the scientifi c names have been changed to match the recent literature).
Tomoda ( 1978 ) clearly dealt with the “ecological speciation” scenario (see Rundle and Nosil 2005 ) for some fi sh groups such as Carassius carps and Silurus catfi shes in Lake Biwa, mainly by considering modifi cations in feeding habits/appa-ratus, habitat types, and migration patterns in endemic species. Takahashi ( 1989 ) focused on the Lake Biwa fl oating goby Gymnogobius isaza and examined the eco-logical speciation process that followed the development of the present environment of Lake Biwa. It would be reasonable to consider that ecological adaptation to the present environment (large, deep pelagic zone/heterogeneous bottom features) occurred after the development of these environmental conditions. Therefore, some authors have concluded that the “species that evolved in Lake Biwa” originated dur-ing the last 0.3–0.4 Myr (Nakajima 1987 ; Takahashi 1989 ). However, the derivation of endemic lineages from closest extant lineages may or may not occur at the same time as adaptive evolution (Nakajima 1994 ). A reliable phylogenetic framework is required to test ecological speciation scenarios for the origins of endemic species, as well as to reexamine the categorization of endemic species that was proposed by Kawanabe ( 1978 ).
1.3 Molecular Phylogenetic Approaches
Recent experimental and theoretical developments in molecular genetics can allow us to discuss historical dynamics in the evolutionary diversifi cation of organisms within a phylogenetic framework (see, e.g., Avise 2004 ). The origins of endemic biodiversity in the world’s ancient lakes have been the focus of research using molecular phylogenetic/population genetic approaches (e.g., Salzburger and Meyer 2004 for the African Great Lakes; Kontula et al. 2003 for Lake Baikal; also see Cristescu et al. 2010 ). These studies have revealed evolutionary patterns and the tempo of explosive adaptive radiations in ancient lakes. In contrast, until very recently, only a few molecular studies have examined the biodiversity of Lake Biwa. Here, I illustrate some examples of previous studies as well as ongoing research into the origin and establishment of endemic species in Lake Biwa.
1.3.1 Origin of Endemic Lineages
As mentioned above, the traditional view of the origin of endemic “species that evolved in Lake Biwa” is concerned with ecological adaptation after the develop-ment of the present Lake Biwa environment, i.e., over the last 0.4 Myr. It is diffi cult to determine when adaptive evolution occurred in a genetic sense (Barrett and Schluter 2008 ). However, using molecular phylogenetic approaches, we can test the
1 Origin and Diversifi cation of Freshwater Fishes in Lake Biwa

11
hypothesis that endemic species evolved from putative ancestral species (extant sis-ter lineages) after the development of the present Lake Biwa environment.
The Lake Biwa fl oating goby Gymnogobius isaza is a typical species that adapted to the pelagic environment of Lake Biwa (Takahashi 1989 ). The goby completes its whole life history within the lake; it lives in the pelagic zone and exhibits diel verti-cal migrations, except during the reproductive season when it spawns in shallow areas near the shore. Based on a phylogenetic analysis of Gymnogobius gobies using mitochondrial DNA sequence data, Harada et al. ( 2002 ) rejected two previous hypotheses, that G. isaza was derived from a presumed ancestor, G. urotaenia, that lived around Lake Biwa; and that the former lineage originated after the develop-ment of the present Lake Biwa. Instead, using molecular clocks for fi shes, they found that G. isaza was derived from the common ancestor of G. urotaenia and G. petschiliensis (referred to as Gymnogobius sp. 1) in the Pliocene (>2 Mya). This result was supported by subsequent studies (Sota et al. 2005 ; Tabata and Watanabe 2013 ) that dated the phylogenetic tree using geological calibration data (i.e., the formation of the Tsushima Strait, which presumably divided the freshwater species into continental and Japanese species).
Similarly, our mtDNA analysis, which used a relaxed molecular clock model with geological constraints, provided evidence that the gudgeon Gnathopogon cae-rulescens , a representative species that exhibits limnetic adaptation, was derived from the closest monophyletic group within G. elongatus 1.7 Mya [0.5–3.5 Mya, 95 % highest posterior density region (HPD)] (Kakioka et al. 2013 ). The old (>4 Myr) derivation of the Carassius cuvieri lineage from C. auratus was also inferred based on a molecular clock (1.52 % pairwise distance/Myr) (Takada et al. 2010 ). Furthermore, according to our ongoing exhaustive phylogenetic analyses of endemic fi shes in Lake Biwa (R. Tabata et al., unpublished data), derivation of most of the presumed “species that evolved in Lake Biwa” from their closest lineages is estimated to date back to the early Pleistocene, the Pliocene, or even the Late Miocene. These patterns do not agree with the hypothesis that the divergence of endemic species occurred in the present Lake Biwa (<0.4 Mya) unless we fi rmly assume that all of the direct ancestral forms of these endemic species have already gone extinct during the last <0.4 Myr.
A few endemic species, however, are little differentiated from their extant sister forms. Benthopelagic gudgeons Sarcocheilichthys are good examples of such spe-cies. The gudgeons show conspicuous trophic polymorphism, especially in the head part, and two species/subspecies are described from Lake Biwa; S. variegatus microoculus (endemic subspecies), which has a short to long head, and S. biwaensis (endemic species), which has a long head and a darker body color. No signifi cant differentiation is found between the two species in neutral genetic markers, mtDNA, or microsatellites (Fig. 1.3 ; Komiya et al. 2011 ), and the level of differentiation between Lake Biwa and neighboring populations is not large (Komiya et al. 2013 ). Head shape and body color are likely adaptive traits because they obviously corre-spond to habitat characteristics; S. v. microoculus with a short head and yellowish body inhabits sandy bottom habitats, whereas S. v. microoculus and S. biwaensis with longer heads and darker bodies inhabit rocky bottom habitats that are formed
1.3 Molecular Phylogenetic Approaches

12
in restricted areas in the northern basin of Lake Biwa (Komiya et al. 2011 ). Hence, their trophic traits and body colors probably evolved rapidly following the develop-ment of the rocky shore environment in the present Lake Biwa, although it is pos-sible that variation in those traits can be attributed to phenotypic plasticity. Another endemic gudgeon subspecies Squalidus chankaensis biwae , which shows adapta-tions to the pelagic zone, also seems to be a young endemic form (R. Kakioka et al., unpublished data). Further, Oohara and Okazaki ( 1996 ) suggested that the Biwa salmon, Oncorhynchus masou rhodurus , originated in the present Lake Biwa (~0.5 Mya) based on an mtDNA molecular clock of 2 % pairwise distance/Myr. Our mtDNA analysis results, which were obtained using a relaxed molecular clock model with geological constraints, roughly agrees with this inference (0.4–0.7 Myr 95 % HPD, R. Tabata et al., unpublished data).
Fig. 1.3 mtDNA haplotype group frequencies ( a ), statistical parsimony network of mtDNA ( b ), and the result of a Bayesian genetic assignment analysis ( K = 2) based on 14 microsatellites ( c ) for Sarcocheilichthys fi shes. Sb , S. biwaensis ; others, S. variegatus microoculus . In both ( a ) and ( b ), each clade is shown in the same color. No distinct genetic divergence between the species/local samples was detected in mtDNA ( b ) or microsatellites ( c ). Modifi ed from Komiya et al. ( 2011 )
1 Origin and Diversifi cation of Freshwater Fishes in Lake Biwa
13
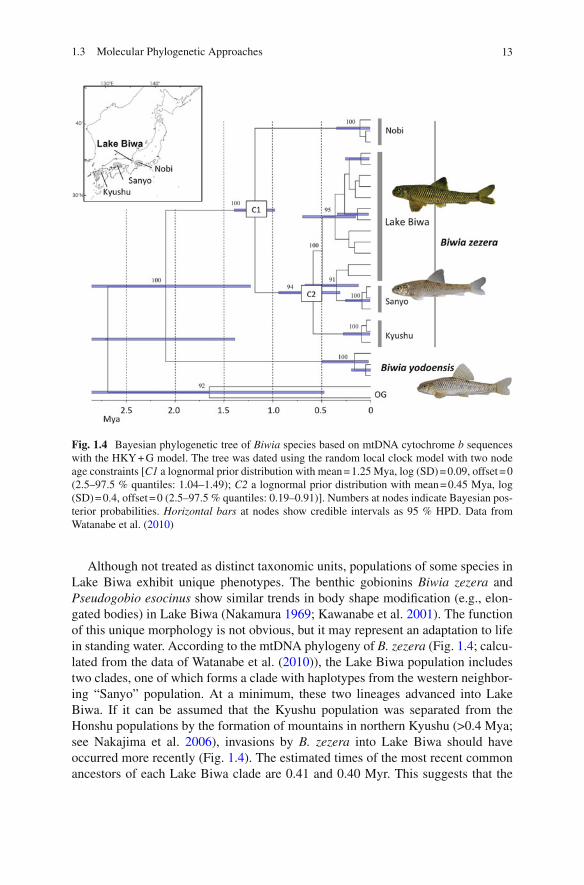
Although not treated as distinct taxonomic units, populations of some species in Lake Biwa exhibit unique phenotypes. The benthic gobionins Biwia zezera and Pseudogobio esocinus show similar trends in body shape modifi cation (e.g., elon-gated bodies) in Lake Biwa (Nakamura 1969 ; Kawanabe et al. 2001 ). The function of this unique morphology is not obvious, but it may represent an adaptation to life in standing water. According to the mtDNA phylogeny of B. zezera (Fig. 1.4 ; calcu-lated from the data of Watanabe et al. ( 2010 )), the Lake Biwa population includes two clades, one of which forms a clade with haplotypes from the western neighbor-ing “Sanyo” population. At a minimum, these two lineages advanced into Lake Biwa. If it can be assumed that the Kyushu population was separated from the Honshu populations by the formation of mountains in northern Kyushu (>0.4 Mya; see Nakajima et al. 2006 ), invasions by B. zezera into Lake Biwa should have occurred more recently (Fig. 1.4 ). The estimated times of the most recent common ancestors of each Lake Biwa clade are 0.41 and 0.40 Myr. This suggests that the
Fig. 1.4 Bayesian phylogenetic tree of Biwia species based on mtDNA cytochrome b sequences with the HKY + G model. The tree was dated using the random local clock model with two node age constraints [ C1 a lognormal prior distribution with mean = 1.25 Mya, log (SD) = 0.09, offset = 0 (2.5–97.5 % quantiles: 1.04–1.49); C2 a lognormal prior distribution with mean = 0.45 Mya, log (SD) = 0.4, offset = 0 (2.5–97.5 % quantiles: 0.19–0.91)]. Numbers at nodes indicate Bayesian pos-terior probabilities. Horizontal bars at nodes show credible intervals as 95 % HPD. Data from Watanabe et al. ( 2010 )
1.3 Molecular Phylogenetic Approaches

14
time scale for adaptation to Lake Biwa was less than ~0.4 Myr, i.e., adaptation occurred after the development of the present Lake Biwa. Pseudogobio esocinus presents similar results (K. Tominaga et al., unpublished data). A unique landlocked form of Ayu, Plecoglossus altivelis , in Lake Biwa is also estimated to have differen-tiated after formation of the present Lake Biwa (0.1 Mya from Nei’s genetic dis-tance for allozyme data; Nishida 1985 ).
In summary, molecular phylogenetic analyses are revealing that most of the endemic lineages in Lake Biwa differentiated from extant related lineages long before the development of the present Lake Biwa (<0.4 Mya). However, irrespective of taxonomical treatment, adaptive phenotypes that are endemic to Lake Biwa can be inferred to have evolved in the present Lake Biwa environment. Exhaustive phy-logenetic analyses of Lake Biwa endemics will further alter our view of the pattern and time scale of the origin of endemic lineages in the lake.
1.3.2 Population Establishment by Endemic and Non-endemic Species
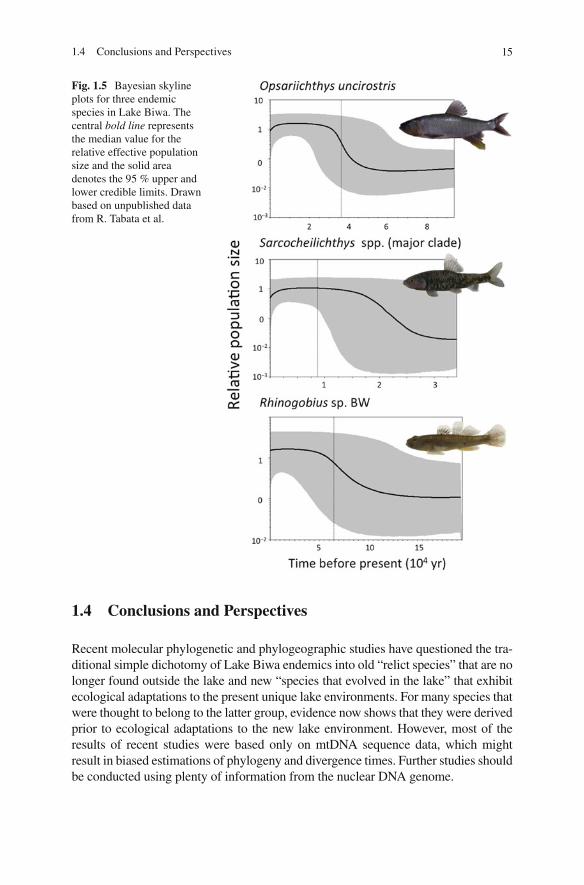
The divergence time for endemic lineages in Lake Biwa varies greatly among spe-cies, as shown above. However, our ongoing comparative demographic analysis of Lake Biwa populations has shown similar patterns among many species. A Bayesian skyline plot analysis (Drummond et al. 2005 ), as well as traditional mismatch dis-tribution analysis, has provided information regarding historical demographic changes using molecular genetic data (Ho and Shapiro 2011 ). Many populations, including both old and young endemic lineages of “species that evolved in Lake Biwa” as well as “relict species,” show a pattern of recent population expansion that occurred several tens of thousands of years ago (Fig. 1.5 ). This pattern has also been observed for many non-endemic species (R. Tabata et al., unpublished data). This means that the present populations in Lake Biwa colonized the lake in the late Pleistocene, after the present lake environment developed, or they experienced severe bottlenecks and subsequently became reestablished during that period.
Drastic climate changes in the Pleistocene are believed to have had large impacts on freshwater fi sh populations. Rising sea levels in the interglacial period would have reduced the habitats available to freshwater fi shes, especially considering island topography, and the cool and dry climate in the glacial period would also have had negative impacts on populations that were adapted to a temperate monsoon climate. Even in Lake Biwa, reductions in population size in the late Pleistocene have been inferred for many species. However, the lake would have also functioned as a “reservoir” for many species, which are represented by the “relict endemic species” as well as other general species.
1 Origin and Diversifi cation of Freshwater Fishes in Lake Biwa
15
Fig. 1.5 Bayesian skyline plots for three endemic species in Lake Biwa. The central bold line represents the median value for the relative effective population size and the solid area denotes the 95 % upper and lower credible limits. Drawn based on unpublished data from R. Tabata et al.
1.4 Conclusions and Perspectives
Recent molecular phylogenetic and phylogeographic studies have questioned the tra-ditional simple dichotomy of Lake Biwa endemics into old “relict species” that are no longer found outside the lake and new “species that evolved in the lake” that exhibit ecological adaptations to the present unique lake environments. For many species that were thought to belong to the latter group, evidence now shows that they were derived prior to ecological adaptations to the new lake environment. However, most of the results of recent studies were based only on mtDNA sequence data, which might result in biased estimations of phylogeny and divergence times. Further studies should be conducted using plenty of information from the nuclear DNA genome.
1.4 Conclusions and Perspectives

16
During drastic climate changes in the late Pleistocene, the large water body of Lake Biwa would have functioned as a reservoir for endemic and non-endemic spe-cies. Under such conditions, both old and new lineages from various taxonomic groups would have been facilitated to adapt ecologically to the unique lake environ-ments, such as the pelagic zone and rocky bottom. They evolved similar sets of phenotypic traits in parallel, which can be treated as “lacustrine syndromes” (e.g., pelagic plankton feeders, sustained swimmers, rocky bottom specialists). Such par-allel adaptive evolution at the community level might have occurred repeatedly in western Japan at several time points since the Pliocene. Unlike other typical cases in the world’s ancient lakes, no remarkable species fl ocks are found in the present Lake Biwa. However, the multiple colonizations and independent radiations inferred in East African cichlids (e.g., Salzburger and Meyer 2004 ) are likely shared with the situation in Lake Biwa. The geographical location and history may be attributable to the less developed species fl ocks in Lake Biwa, which has never been glaciated and has maintained some connections with river systems adjacent to the lake. Considering possible unique aspects of the endemic community in Lake Biwa, par-allel adaptive evolution should be examined throughout the fi sh assemblage based on phylogeny, population genetics, ecology, and traditional morphological/physio-logical comparisons.
Currently, several “omics” studies have also been initiated to examine lacustrine adaptations in Lake Biwa (e.g., proteomics, QTL association mapping with RAD- seq; Kakioka et al. 2013 ). These studies will contribute to identifying the genetic basis of some aspects of adaptation. Furthermore, genome-level studies may pro-vide a clue to answering “when” such adaptive evolution occurred (Barrett and Schluter 2008 ). Were the previous inhabitants of Paleo-Lake Biwa able to adapt to the present Lake Biwa more easily than newcomers due to their standing variation? Interactions between adaptations and community structuring over multiple time scales remain an open question. Such evolutionary interactions would have caused feedback into ecosystem processes, which might have prompted further adaptation and speciation in the lake. To learn more from Lake Biwa, we should concentrate on reducing human impacts on the lake ecosystem, which currently suffers from eutro-phication, destruction of the ecotone area, and destruction of endemic communities through introductions of invasive alien species, such as sunfi shes.
Acknowledgments I express my sincere thanks to Noboru Okuda for providing me the opportunity to write this review. I also thank Takefumi Komiya, Ryoichi Tabata, Ryo Kakioka, Koji Tominaga, and Shigefumi Kanao for providing information on the results of their studies, which will be pub-lished in the near future and will enhance this incomplete review on the origin of Lake Biwa endemics. Kohji Mabuchi provided constructive suggestions for improving the manuscript. This study was supported in part by the Grants-in-Aid from the Ministry of Education, Culture, Sports, Science and Technology, Japan (nos. 18570086, 21370035, and “Formation of a Strategic Base for Biodiversity and Evolutionary Research: from Genome to Ecosystem” of the GCOE) and the WEC Research Fund for Ecology and Civil Engineering (2009-6).
1 Origin and Diversifi cation of Freshwater Fishes in Lake Biwa

17
References
Annandale N (1922) The macroscopic fauna of Lake Biwa. Annot Zool Jpn 10:127–153 Avise JC (2004) Molecular markers, natural history, and evolution, 2nd edn. Sinauer, Sunderland Barrett RDH, Schluter D (2008) Adaptation from standing genetic variation. Trends Ecol Evol
23:38–44 Cristescu ME, Adamowicz SJ, Vaillant JJ, Haffner DG (2010) Ancient lakes revisited: from the
ecology to the genetics of speciation. Mol Ecol 19:4837–4851 Drummond AJ, Rambaut A, Shapiro B, Pybus OG (2005) Bayesian coalescent inference of past
population dynamics from molecular sequences. Mol Biol Evol 22:1185–1192 Fujita T, Nishino M, Hosoya K (2008) The original scenery of the “Naiko” lagoons around Lake
Biwa inferred by fi sh specimen survey. Jpn J Ichthyol 55:77–93 (in Japanese with English abstract)
Greenwood PH (1984) What is a species fl ock? In: Echelle AA, Kornfi eld I (eds) Evolution of fi sh species fl ocks. University of Maine at Orono Press, Orono
Harada S, Jeon SR, Kinoshita I, Tanaka M, Nishida M (2002) Phylogenetic relationships of four species of fl oating gobies ( Gymnogobius ) as inferred from partial mitochondrial cytochrome b gene sequences. Ichthyol Res 49:324–332
Ho SYW, Shapiro B (2011) Skyline-plot methods for estimating demographic history from nucle-otide sequences. Mol Ecol 11:423–434
Hosoya K (2005) Migration patterns of freshwater fi shes in Lake Biwa and roles of “Naiko”. In: Nishino M, Hamabata E (eds) Messages from “Naiko”: conservation of the biodiversity in the wetland around Lake Biwa and its rehabilitation. Sunrise Publishing, Otsu (in Japanese)
Kakioka R, Kokita T, Komada H, Watanabe K, Okuda N (2013) A RAD-based linkage map and comparative genomics in the gudgeons (genus Gnathopogon , Cyprinidae). BMC Genomics 14:32. doi: 10.1186/1471-2164-14-32
Kakioka R, Kokita T, Tabata R, Mori S, Watanabe K (2013) The origins of limnetic forms and cryptic divergence in Gnathopogon fi shes (Cyprinidae) in Japan. Environ Biol Fish 96:631–644. doi: 10.1007/s10641-012-0054-x
Kawabe T (1994) Biwako no Oitachi (formation of Lake Biwa). In: Research Group for Natural History of Lake Biwa (ed) Biwako no shizenshi (The natural history of Lake Biwa). Yasaka Shobo, Tokyo (in Japanese)
Kawanabe H (1978) Some biological problems. Verh Int Ver Limnol 20:2674–2677 Kawanabe H (1996) Asian great lakes, especially Lake Biwa. Environ Biol Fish 47:219–234 Kawanabe H, Mizuno N, Hosoya K (2001) Freshwater fi shes of Japan, 3rd edn. Yama-Kei
Publishers, Tokyo Kobayakawa M (1994) Catfi shes. In: Research Group for Natural History of Lake Biwa (ed)
Biwako no shizenshi (The natural history of Lake Biwa). Yasaka Shobo, Tokyo (in Japanese) Kodera H (1985) Paleontological evidence for appearance of a species, Gengoro-buna Carassius
cuvieri Temminck et Schlegel. A comparative study of the dental tissues between living and fossil species. Earth Sci (Chikyu Kagaku) 39:272–281 (in Japanese with English abstract)
Kodera H, Nakamura T (1976) The fi rst discovery of fossil carp from the Kobiwako Group, Shiga Prefecture, Japan. J Geol Soc Jpn 81:591–594 (in Japanese with English abstract)
Komiya T, Fujita S, Watanabe K (2011) A novel resource polymorphism in fi sh, driven by differential bottom environments: an example from an ancient lake in Japan. PLoS One 6:e17430. doi: 10.1371/journal.pone.0017430
Komiya T, Fujita-Yanahibayashi S, Watanabe K (2013) Multiple colonizations of Lake Biwa by Sarcocheilichthys fi shes and their population history. Environ Biol Fish. doi: 10.1007/s10641-013-0176-9
Kontula T, Kirilchik SV, Väinolä R (2003) Endemic diversifi cation of the monophyletic cottid fi sh species fl ock in Lake Baikal explored with mtDNA sequencing. Mol Phylogenet Evol 27:143–155
Martens K (1997) Speciation in ancient lakes. Trends Ecol Evol 12:177–182
References

18
Meyers PA, Takemura K, Horie S (1993) Reinterpretation of late Quaternary sediment chronology of Lake Biwa, Japan, from correlation with marine glacial–interglacial cycles. Quat Res 39:154–162
Miura T (1966) Ecological notes of the fi shes and interspecifi c relations among them in Lake Biwa. Jpn J Limnol 27:49–72
Nakajima T (1987) Formation of ichthyofauna and speciation in Lake Biwa. In: Mizuno N, Goto A (eds) Freshwater fi shes in Japan: their distribution, variation and speciation. Tokai University Press, Tokyo (in Japanese)
Nakajima T (1994) Cyprinid fi shes. In: Research Group for Natural History of Lake Biwa (ed) Biwako no shizenshi (The natural history of Lake Biwa). Yasaka Shobo, Tokyo (in Japanese)
Nakajima J, Onikura N, Matsui S, Oikawa S (2006) Geographical distribution of genuine freshwa-ter fi shes in Fukuoka Prefecture, northern Kyushu, Japan. Jpn J Ichthyol 53:117–131
Nakamura M (1969) Cyprinid fi shes of Japan. Special Publication, Research Institute of Natural Resources, no 4, Shigen Kagaku Kenkyusyo, Tokyo (in Japanese)
Nishida M (1985) Substantial genetic differentiation in Ayu Plecoglossus altivelis of the Japanese and Ryukyu Islands. Bull Jpn Soc Sci Fish 51:1269–1274
Nishino M (2003) Biwako no Koyushu wo Meguru Mondai. Koyushu list no Ichibu Shusei nit suite (The problem of endemics in Lake Biwa). Oumia 76:3–4 (in Japanese)
Nishino M (2005) Wetland environment of Japan and “Naiko” lagoons around Lake Biwa. In: Nishino M, Hamabata E (eds) Messages from “Naiko”: conservation of the biodiversity in the wetland around Lake Biwa and its rehabilitation. Sunrise Publishing, Otsu (in Japanese)
Nishino M, Hamabata E (2005) Messages from “Naiko”: conservation of the biodiversity in the wetland around Lake Biwa and its rehabilitation. Sunrise Publishing, Otsu (in Japanese)
Oohara I, Okazaki T (1996) Genetic relationship among three subspecies of Oncorhynchus masou determined by mitochondrial DNA sequence analysis. Zool Sci 13:189–198
Research Group for Natural History of lake Biwa (1994) Biwako no shizenshi (The natural history of Lake Biwa). Yasaka Shobo, Tokyo (in Japanese)
Rossiter A (2000) Lake Biwa as a topical ancient Lake. Adv Ecol Res 31:571–598 Rundle HD, Nosil P (2005) Ecological speciation. Ecol Lett 8:336–352 Salzburger W, Meyer A (2004) The species fl ocks of East African cichlid fi shes: recent advances
in molecular phylogenetics and population genetics. Naturwissenschaften 91:277–290 Sota T, Mukai T, Shinozaki T, Sato H, Yodoe K (2005) Genetic differentiation of the gobies
Gymnogobius castaneus and G. taranetzi in the region surrounding the Sea of Japan as inferred from a mitochondrial gene genealogy. Zool Sci 22:87–93
Tabata R, Watanabe K (2013) Hidden mitochondrial DNA divergence in the Lake Biwa endemic goby Gymnogobius isaza : implications for its evolutionary history. Environ Biol Fish 96:701–712. doi: 10.1007/s10641-012-0062-x
Takada M, Tachihara K, Kon K, Yamamoto G, Iguchi K, Miya M, Nishida M (2010) Biogeography and evolution of the Carassius auratus -complex in East Asia. BMC Evol Biol 10:7
Takahashi S (1989) A review of the origins of endemic species in Lake Biwa with special reference to the goby fi sh, Chaenogobius isaza . J Paleolimnol 1:279–292
Tanaka S (1908) On some fi shes from Lake Biwa, with description of one new species and a list of all the fi sh species hitherto known from that locality. Annot Zool Jpn 7:1–15
Tanimoto M, Okuyama S (2003) A fossil channid found from the Lower Pliocene Ueno Formation of the Kobiwako Group at Ohyamada-mura, Ayama-gun, Mie Prefecture. Chigakukenkyu 51:195–199
Tokui T, Kawanabe H (1984) Fishes. In: Horie S (ed) Lake Biwa. Monographiae Biologicae 54. Dr W Junk Publishers, Dordrecht
Tomoda Y (1978) Lake Biwa and catfi sh (Biwako to namazu). Chobunsha, Tokyo (in Japanese) Tomoda Y (1991) Fish of Lake Biwa: endemic fi sh of Lake Biwa and fi sh remains collected from
Paleo Lake Biwa (Kobiwako Group). In: Nature Conservation Foundation of Shiga Prefecture (ed) Landscape and environment of Shiga: scientifi c studies of Shiga Prefecture, Japan (Shigaken shizenshi). Daiichihoki, Tokyo (in Japanese with English summary)
1 Origin and Diversifi cation of Freshwater Fishes in Lake Biwa

19
Ueno M (1943) Freshwater life in some regions of the West Pacifi c. In: Society of the Pacifi c (ed) Pacifi c ocean and inlandwater (Taiheiyo no kaiyo to rikusui). Iwanami, Tokyo (in Japanese)
Watanabe K (2012) Faunal structure of Japanese freshwater fi shes and its artifi cial disturbance. Environ Biol Fish 94:533–547. doi: 10.1007/s10641-010-9601-5
Watanabe K, Uyeno T (1999) Fossil bagrid catfi shes from Japan and their zoogeography, with description of a new species, Pseudobagrus ikiensis . Ichthyol Res 46:397–412
Watanabe K, Kawase S, Mukai T, Kakioka R, Miyazaki J-I, Hosoya K (2010) Population diver-gence of Biwia zezera (Cyprinidae: Gobioninae) and the discovery of a cryptic species, based on mitochondrial and nuclear DNA sequence analyses. Zool Sci 27:647–655
Yokoyama T (1968) Transition of paleo-lake Biwa during the pliocene deduced from paleocurrent directions and other sedimentological features of sandy facies in the Kobiwako group, Central Japan: the research of younger cenozoic strata in Kinki province, part 11. J Geol Soc Jpn 74:623–632
Yokoyama T (1984) Stratigraphy of the Quaternary system around Lake Biwa and geohistory of the ancient Lake Biwa. In: Horie S (ed) Lake Biwa, Monographiae Biologicae 54. Dr W Junk Publishers, Dordrecht
Yuma M, Hosoya K, Nagata Y (1998) Distribution of the freshwater fi shes of Japan: an historical overview. Environ Biol Fish 52:97–124
References

21N. Okuda et al., Biodiversity in Aquatic Systems and Environments: Lake Biwa, SpringerBriefs in Biology, DOI 10.1007/978-4-431-54150-9_2, © The Author(s) 2014
Abstract Trophic polymorphism, defi ned as the state in which two or more clearly different feeding functional traits exist within the same population of a species, is general in fi shes. In aquatic ecosystems, the fi shes often have strong impacts on prey communities as keystone predators, so that phenotypic divergence of their feeding traits can alter biotic and abiotic attributes of environments through changes in the form of trophic interactions. The predator-induced environmental alterations may, in turn, drive evolutionary changes in adaptive traits of themselves as well as of other members in the communities. This process, in which ecology and evolution reciprocally interplay over contemporary time-scales, is termed “eco-evolutionary feedback.” In this chapter, we review how and when trophic polymorphism has been generated in fi sh populations and then discuss what consequences it has in ecologi-cal and evolutionary aspect. Special references are made to the case in the ancient Lake Biwa, which has a geological history long enough for divergent fi sh popula-tions to come to ecological speciation and thus provides a good opportunity to understand how such an evolutionary process diversifi es biological communities and consequently ecosystem properties in lakes.
Keywords Biodiversity • Eco-evolutionary feedback • Ecological speciation • Ecosystem functioning • Keystone predator • Mesocosm • Species pair • Trophic polymorphism
2.1 Why is Biodiversity Important?
What is the ecological signifi cance of conserving biological diversity within a spe-cies? In the context of conservation biology, it could be argued that maintenance of intraspecifi c phenotypic variation has the advantage of increasing population viabil-ity because it provides the population with the adaptability to respond to changing environments. However, such a traditional view has been replaced by recent
Chapter 2 Predator Diversity Changes the World: From Gene to Ecosystem

22
ecological fi ndings that the intraspecifi c diversity of a keystone species impacts substantially on biological communities and their ecosystem functioning. In this chapter, we fi rst focus on phenotypic polymorphism in fi shes as keystone predators of aquatic ecosystems, referring to genetic mechanisms underlying their polymor-phism. We also introduce some of the latest studies, which demonstrate that intra-specifi c functional diversity of fi sh predators alters aquatic community structure and ecosystem processes. Finally, we show that environments modifi ed by a predator’s functional diversity can serve as a driver for the evolutionary diversifi cation of aquatic biota through eco-evolutionary feedbacks.
2.2 Biodiversity at Three Levels
According to the Convention on Biological Diversity (CBD), biological diversity includes diversity within species, between species and of ecosystems, being viewed at different levels of biological organization from genes to ecosystems. In terms of conservation biology, the maintenance of intraspecifi c diversity is of ecological importance because a population with a variety of phenotypes, which have the potential for adaptation against unpredictable environmental changes in the future, such as global warming, habitat alteration and prevalence of emerging pathogens, may be able to prevent extinction. Furthermore, recent developments in community genetics have provided a new insight into the ecological and evo-lutionary consequences of intraspecifi c diversity. In a population of terrestrial plant, for example, cottonwood Populus , individual phenotypes can harbor dif-ferent arthropod communities according to their anti-herbivore defense capacity, which is based on genotypes (Whitham et al. 2006 ). The genotypic difference among individuals further causes intraspecifi c variation in condensed tannin pro-duction in leaves, which in turn affects litter decomposition processes attribut-able to forest fl oor microbes. Since this fi nding, an increasing number of studies have looked at the cascading effects of intraspecifi c biodiversity on community structure and ecosystem processes, attracting much attention from both commu-nity and evolutionary ecologists.
2.3 What is Phenotypic Polymorphism?
Phenotypic polymorphism, defi ned as the state in which two or more clearly differ-ent phenotypic traits exist within the same population of a species, is a common phenomenon found in a variety of taxonomic groups from prokaryotes to higher vertebrates. The polymorphic phenotypes include behavioral, morphological, phys-iological and life history traits (Gross 1996 ). In a narrower sense, phenotypic poly-morphism is confi ned to discontinuous phenotypic variation within a Mendelian population in which interbreeding individuals share a common gene pool. In a broad
2 Predator Diversity Changes the World: From Gene to Ecosystem

23
sense, however, it is applicable to the case in which intraspecifi c phenotypic varia-tion results from secondary contact between multiple populations under different selection regimes and/or undergoing different genetic drifts in different locations. In addition, some studies have recently reported that the polymorphic state is attained by phenotypic plasticity within a genetically monomorphic population (Agrawal 2001 ; Miner et al. 2005 ). To understand the evolution of phenotypic polymorphism, it is essential to elucidate the origins of polymorphic phenotypes and the underlying mechanisms for polymorphic expression. But yet, such a subject is at the state of the art. In this chapter, when treating with cases in which there exists discontinuous phenotypic variation within a given population at a given time, we regard all of them as phenotypic polymorphism without referring to the processes and mechanisms for generating it.
2.4 Phenotypic Polymorphism in Aquatic Ecosystems
In aquatic organisms, phenotypic polymorphism is often observed in traits related to predator–prey interactions. Prey organisms often show a polymorphic pattern of traits in relation to inedibility, termed anti-predator defensive traits. For example, phytoplankton can avoid zooplankton predation by means of colony formation, while zooplankton under high predation pressure express their sharp helmet and extended tail spine, which functions to deter fi sh and invertebrate predation (Lass and Spaak 2003 ). In contrast, fi sh predators show polymorphism in behavioral and morphological traits associated with feeding habits, which is referred to as trophic polymorphism (Smith and Skulason 1996 ). Intraspecifi c variation in feeding traits leads to discrete variation in species trophic niche through functional differences in prey size selectivity or prey species-specifi c foraging effi ciency (Robinson 2000 ).
2.4.1 Trophic Polymorphism and Adaptive Divergence
In the East African Great Lakes, which have a long geological history spanning hundreds of thousands to tens of millions of years, cichlid fi shes are well known to be a group showing adaptive radiation. In this group, it has been believed that a small number of ancestral species evolved to the diversity of existing species through specialization of their habitat and food resource utilization (Seehausen 2006 ). The ancestral species, which was the fi rst to colonize these lakes, would have intensifi ed intraspecifi c competition for food resources and habitats in the process of its population expansion under environments in which there existed few or no predators and competitors. As an effi cient way to reduce intraspecifi c competition, the fi sh might have partitioned their resources to occupy alternative empty niches as the population became saturated. Individual variation in plastic foraging behavior may have been the fi rst step of niche specialization ( Bolnick et al. 2003 ). When the
2.4 Phenotypic Polymorphism in Aquatic Ecosystems

24
foraging behavior is partly inherited in the progeny, behavioral specialization can facilitate the subsequent physiological and morphological adaptations to specifi c resource utilization (Sacotte and Magnan 2006 ). Such a phenotypic adaptation, which enables each individual to utilize different resources more effi ciently, will favor disruptive selection within the population (Knudsen et al. 2010 ). If it is accom-panied by reproductive isolation due to sexual selection, then the adaptive diver-gence can lead to ecological speciation (Seehausen et al. 2008 ). Marked interspecifi c diversity of trophic niches and morphology found in African cichlids may be an evolutionary consequence of resource partitioning among individuals derived from a colonized population (Bootsma et al. 1996 ; Genner et al. 1999 ).
There is excellent evidence supporting the evolutionary scenario of adaptive radiation in cichlids, in which colonized populations of ancestral species partitioned resources among individuals and subsequently diversifi ed their morphology, lead-ing to ecological speciation. In a Nicaraguan crater lake, which was formed by volcanic activity less than 23,000 years ago, two cichlid species coexist and form a monophyletic assemblage (Barluenga et al. 2006 ). One species, which is wide-spread among adjacent lakes, lives in the littoral habitat and another endemic spe-cies lives in the limnetic habitat of the deep crater lake. The two species also differ greatly in their feeding habits and morphology, with the limnetic species possessing a more slender body. Molecular data revealed that these two species were reproduc-tively isolated less than 10,000 years ago, suggesting that morphological divergence through habitat/food preference and subsequent reproductive isolation through assortative mating led to sympatric speciation. This fi nding seems to reconfi rm assumptions of adaptive radiation of cichlids in African Great Lakes in which mor-phologically diverse species have derived from a few ancestors. In the case of Nicaraguan cichlids, colonization to newly created environments, where competi-tors are originally absent, would have facilitated phenotypic divergence of trophic morphology and the subsequent ecological speciation, which provides unique opportunities to understand the evolutionary process of adaptive diversifi cation within a lake environment.
Although trophic polymorphism has been found in phylogenetically different groups of fi shes, some of its features are shared among them. One of the most remarkable, in this respect, is that the evolutionary pattern of morphological diver-gence associated with feeding specialization is similar across species. For example, individuals with a deeper body, a shorter snout and a wider mouth feed on benthic prey preferentially, while those with a slenderer body and an elongated snout fre-quently feed on plankton in pelagic waters. In the former case, the wider mouth has the advantage of readily consuming benthic prey on the lake bottom (Robinson 2000 ) and the deeper body allows the development of larger pharyngeal muscles, which are advantageous in the crushing of the hard shells of benthic invertebrates, such as gastropods and decapods (Wainwright et al. 1991 ; Osenberg et al. 2004 ). By contrast, in the latter case, the slender body is suited to optimize swimming perfor-mance, which would be required to extensively search for planktonic prey (Hendry et al. 2011 ). In addition, these individuals have a greater gill raker number and nar-rower gill raker spacing, which are both effective in the fi ltration of small particles
2 Predator Diversity Changes the World: From Gene to Ecosystem

25
such as plankton (Robinson 2000 ). Such a morphological divergence is closely associated with habitat-specifi c food resource distribution, characterized by typical lake food webs, i.e., plankton prey derived from a pelagic food chain in the limnetic habitat and benthos prey from a benthic food chain in the littoral habitat. For many fi sh species with phenotypic polymorphism, even if there are interspecifi c varia-tions, their morphological divergence is usually associated with two different tro-phic niches, planktivory and benthivory.
2.4.2 Ecological Speciation and Species Pair
Trophic polymorphism has also been frequently observed in freshwater fi sh species inhabiting postglacial lakes in northern temperate environments: e.g., salmonids, sticklebacks and osmerids (Taylor 1999 ). The reason for this is associated with the geological history of these lakes. In high latitudes of the Northern Hemisphere, the land surface was extensively covered by ice during the last glaciation, which extended from approximately 110,000 to 10,000 years ago. According to the most predominant hypothesis, fi sh species that were the fi rst to colonize in postglacial lakes would have undergone morphological divergence in order to utilize unused food resources in the absence of competitors. If that is true, the evolution of trophic polymorphism in high latitudes must have occurred during the last 10,000 years. This time scale is too short for the fi sh population to diverge into different biological species with complete reproductive isolation (i.e., speciation) but it may be long enough for the population to undergo adaptive divergence (Hendry et al. 2009 ).
The above situation reported for freshwater fi shes in high latitudes is often regarded as a species pair. A species pair is defi ned as a pair of populations that are typically distinguished from each other by differences in ecology, behavior and/or morphology, but are taxonomically classifi ed as the same species because of their incomplete reproductive isolation (Taylor 1999 ). However, sexual selection follow-ing divergent natural selection will lead to the evolution of reproductive isolation through assortative mating based on ecologically selected traits, which can be the principal mechanism of ecological speciation (McKinnon and Rundle 2002 ). Recent molecular studies also provide strong support for the hypothesis that adaptive diver-gence contributing to reduction of gene fl ows will cause genetic incompatibility responsible for reproductive isolation and promote ecological speciation between members of a sympatric species pair (Rogers and Bernatchez 2007 ).
In traditional systematics based on morphological taxonomy, it does not matter if the species pair accords with the entity of biological species. For freshwater fi shes whose local populations have experienced isolation and reorganization repeatedly through geological events such as river and lake capture, it is not easy to defi ne a boundary for a biological species. Whether discrete variation in feeding traits found in a focal taxa is regarded as intraspecifi c or interspecifi c variation relies on how a researcher classifi es a biological entity into “single species” or “different species”. However, “species” is just an operational taxonomic unit and its defi nition is not an
2.4 Phenotypic Polymorphism in Aquatic Ecosystems

26
important issue for understanding how a diversity of feeding functions has evolved from an ancestral lineage. It is more important to recognize that polymorphic popu-lations observed in nature can be arrayed somewhere along a continuum from pan-mixia to complete reproductive isolation (Hendry 2009 ).
2.4.3 The Ancient Lake Biwa as an Evolutionary Hot Spot
The ancient Lake Biwa, which has a geological history longer than postglacial lakes (tens of thousands years) and shorter than the oldest African Great Lakes (tens of millions years), provides a good opportunity to observe how the process of ecologi-cal speciation leads to species diversifi cation. This lake was formed south far from its current location ca. four million years ago (see Chap. 1 ). It is a tectonic lake, i.e., the lake formed as a result of tectonic movement, having gradually moved north-wards. The lake deepened through faulting at the current location ca. 400 thousand years ago and then has stood still there. At present, it shows a high biodiversity with 1,769 aquatic species recorded, including 61 endemics (Timoshkin et al. 2011 ). Similar to other ancient lakes created by tectonic movements, Lake Biwa is charac-terized by the development of deep pelagic habitats, which support many pelagic fi shes (Okuda et al. 2012 ). Recent molecular techniques have revealed that some endemic fi shes, which have adapted to pelagic environments, are derived from lit-toral or fl uvial ancestral species (see Chap. 1 ). Many of these pelagic species share adaptive traits, such as a slender body and narrow gill raker spacing, which are advantageous to inhabiting pelagic waters. Because such an evolutionary diver-gence is found among phylogenetically distant groups, it is suggested that conver-gent evolution of phenotypes adaptive to the pelagic habitats has occurred following the appearance of newly created deep water environments in Lake Biwa.
Even if divergent evolution progressed over geological time scales in this lake, reproductive isolation may remain incomplete. This is the case in sister species of a minnow, Gnathopogon , living in Lake Biwa (Fig. 2.1 ). Gnathopogon elongatus elon-gatus is a common freshwater fi sh inhabiting rivers and ponds in western Japan, while G. caerulescens is endemic to Lake Biwa. In Lake Biwa, these two species coexist but segregate their habitats: i.e., G. elongatus elongatus lives in littoral and fl uvial habi-tats, whereas G. caerulescens lives in limnetic habitats. The former is typical of ben-thivores and the latter of planktivores. The planktivorous G. caerulescens has a slender body and narrower gill raker spacing, which is advantageous to feeding on plankton in pelagic waters. An advanced molecular technique, with a next generation sequencer, elucidated the genetic basis of such a morphological adaptation (Kakioka 2013 ; also see Sect. 2.4.6 ). Although there are remarkable morphological differences between these coexisting fi shes, their phylogenetic relationship is closer than that of allopatric populations within the species G. elongatus elongates (Kakioka et al. 2013a ; Fig. 2.1 ). More interestingly, they are reproductively compatible to each other in captive condi-tions and their hybrids sometimes occur in nature (Nakamura 1969 ; T. Kokita, unpub-lished data), suggesting that discrete variation in their feeding morphology stands for
2 Predator Diversity Changes the World: From Gene to Ecosystem

27
trophic polymorphism within a species pair rather than interspecifi c variation between sister species. This phenomena may also be true for sister species of some other fi sh groups in Lake Biwa.
2.4.4 Life History Polymorphism
In freshwater fi shes, another example of phenotypic polymorphism is often found in life history traits. In many species of salmon and sticklebacks in which trophic poly-morphism is well known, two different life history forms coexist within a popula-tion, i.e., a diadromous form, which migrates between freshwater and marine habitats, and a non-diadromous (or landlocked) form, which occurs in freshwater environments throughout its entire life (Taylor 1999 ). In these polymorphic popula-tions the non-diadromous form generally has a smaller body size at maturity and more gill rakers or narrower gill raker spacing than those in the anadromous form (Foote et al. 1999 ; McKinnon and Rundle 2002 ). For migratory threespine stickle-backs with a marine origin, molecular phylogenetic evidence suggests that the migration-related morphological traits (e.g., the pattern, number, and size of the bony lateral plates) have evolved repeatedly in populations from different locations (Colosimo et al. 2004 ).
Planktivorous
Benthivorous
Feeding habits and morphology
Sympatric lacustine population(G. Caerulescens)
Allopatric lacustrine population(G. elongatus elongatus)
Sympatric lacustrine population(G. elongatus elongatus)
Allopatric fluvial population(G. elongatus elongatus)
Molecular phylogeny
Allopatric fluvial population
Sympatric lacustine population
Allopatric lacustrine population
Sympatric lacustrine population
G. Caerulescens
G. elongatus elongatus
Fig. 2.1 Feeding habits, morphology and molecular phylogeny of a genus Gnathopogon in Japan. The phylogenetic tree was modifi ed after Kakioka et al. ( 2013a )
2.4 Phenotypic Polymorphism in Aquatic Ecosystems

28
An alewife Alosa pseudoharengus (Clupeidae) is one example where relationships between life history, trophic morphology and feeding habits have been intensively investigated (Palkovacs and Post 2008 ). In this species, anadromous fi sh migrate from the ocean to freshwater lakes for breeding in spring. Their young-of- the-year (YOY) fi sh spend several months in nursery lakes to preferentially prey upon large bodied zooplankton before migrating to the ocean in autumn. However, if there are any physi-cal barriers to their migration route, the migratory populations become landlocked, completing their entire life in the nursery lakes. While migratory populations are sometimes landlocked by natural barriers such as beaver dams and wind throws for years to decades, some of them have been completely isolated by artifi cial barriers such as man-made dams, resulting in the prevention of gene fl ows.
In alewives, non-migratory landlocked populations have smaller gape width and narrower gill raker spacing than anadromous populations. In lakes with land-locked alewives, zooplankton size is small all year round, as compared to lakes with anadromous alewives, because of constant fi sh predation of large bodied zooplankton. In a situation where intra- and/or inter-specifi c food competition is intense and constant among planktivores coexisting in lakes, evolution might favor the landlocked morph with the smaller gape width and narrower gill raker spacing, which are more adept at foraging on small-bodied prey items. Estimations based on molecular data suggest that such a divergent evolution occurred between 5000 and 270 years ago, during which time dams were built in this region (Palkovacs et al. 2008 ).
For migratory fi shes, geographic barriers temporarily isolate local populations in terms of geological time scales, while their high mobility promotes the reorga-nization of different populations under different selection regimes, creating spatio- temporal heterogeneity in trait distribution within and/or among popula-tions. As a consequence, a multimodal distribution of phenotypes may be observed at a given time and space in relation to trophic morphology as well as life history traits. This may explain why polymorphic phenomena, such as trophic polymor-phism or a species pair, have often been reported for freshwater fi shes with a diadromous origin.
2.4.5 Rapid Evolution of Trophic Polymorphism
How rapidly can trophic polymorphism emerge within a population after coloni-zation of a new environment? Human introductions of non-native species, which have been considered a serious conservation issue worldwide, can provide unex-pected opportunities to study the evolutionary process of trophic polymorphism. The bluegill sunfi sh Lepomis macrochirus was introduced into Japan from the United States in 1960 and, thereafter, rapidly spread throughout the country. Surprisingly, all of the introduced fi sh are derived from only 18 individuals col-lected from the Mississippi River at Guttenberg, Iowa, and genetic diversity is very low in the respective introduced populations, probably due to a founder
2 Predator Diversity Changes the World: From Gene to Ecosystem

29
effect arising from introduction of a small number of individuals with subsequent genetic drift (Kawamura et al. 2006 ). In the original population there are two feeding morphs. The littoral or benthic morph has an enlarged body depth and long pectoral fi ns, indicating a feeding specialization towards benthic prey, while the limnetic or pelagic morph has a slender body and short pectoral fi ns, which give an advantage in searching for zooplankton in pelagic waters (Ehlinger 1990 , 1991 ; Wilson 1998 ).
In Lake Biwa, there are records indicating that introduced bluegills mainly fed on shrimp in the early phase of their colonization during the 1970s (Terashima 1980 ), suggesting that their founders were originally the littoral morph. More than half a century after the colonization, however, Yonekura et al. ( 2002 ) found that the introduced population showed specialization of feeding habits, i.e., individual food preferences for zooplankton, zoobenthos or submerged plants, and that they indi-vidually differed in morphology corresponding to their feeding specialization. Their trophic morph-specifi c feeding habits were also consistent through the time, as suggested by their stable isotopic signatures, which revealed that each of the morphs occupied a different trophic niche (Uchii et al. 2007 ). Such a feeding spe-cialization might have been promoted by food resource partitioning among intro-duced bluegills because their habitats are confi ned to a narrow zone of shallow waters in this deep lake. Yonekura et al. ( 2007b ) demonstrated that the smaller gill raker spacing characteristic of the pelagic morph increases foraging effi ciency for zooplankton prey, whereas the wider mouth characteristic of the benthic morph increases foraging effi ciency for zoobenthos prey. More interestingly, submerged plant feeders have developed an intermediate morph between benthic and limnetic morphs. Uchii ( 2007 ) revealed that bluegills in captive conditions could not com-pensate for their growth by feeding on submerged plants exclusively, suggesting that the submerged plant feeders found naturally adopt “the best of a bad situation” strategy, because their intermediate morph is not specialized for either benthos or plankton feeding.
The example of introduced bluegills implies that newly colonized populations can achieve phenotypic divergence much faster than expected from the evolutionary time required for a species pair, which is estimated as ten thousand years. Unfortunately, it remains untested whether trophic polymorphism of introduced bluegills is attributed to genetic divergence during their colonization, which gives direct evidence for the rapid evolution. However, recent molecular data suggests the possibility that polymorphic variation in feeding traits is under the control of phe-notypic plasticity (see Sect. 2.4.6 ). Another excellent example of the rapid evolu-tion, if not of trophic polymorphism, in fi shes is the case of sockeye salmon, in which introduced populations derived from a common ancestry colonized adjacent river and lake habitats separately, resulting in phenotypic divergence. In these intro-duced populations, Hendry et al. ( 2000 ) demonstrated that reproductive isolation rapidly evolved between two divergent populations within fewer than 13 genera-tions of their colonization. Therefore, it is possible for newly colonized populations to evolve trophic polymorphism followed by assortative mating between the same morphs on an ecological time-scale, sometimes even within decades.
2.4 Phenotypic Polymorphism in Aquatic Ecosystems

30
2.4.6 Genetic Mechanisms of Trophic Polymorphism
Although it remains unknown how phenotypic polymorphism is maintained within a population, there are two mechanisms for generating it. One is genetic variation, with the other being phenotypic plasticity, defi ned as ontogenetic phenotypic changes in response to changes in the environment. Some studies have reported that trophic polymorphism in fi shes has at least partly a genetic basis. For example, Proulx and Magnan ( 2004 ) conducted common garden experiments with a lacus-trine brook charr, in which both pelagic and littoral morphs coexisted. Young-of- the-year (YOY) fi sh of each morph were transplanted to a tank, either simulating a pelagic or a littoral habitat, and fed pelagic or benthic food respectively. Some vari-ation in feeding morphology was accounted for by genetic effects, whereas some was environmental. More interestingly, additional experiments, under which fi sh habitats were alternated between each other during their growth, indicated that some of the morphological traits were plastically changed to adjust to a new habitat, suggesting the ecological signifi cance of phenotypic plasticity during ontogenetic habitat shifts. For many species showing trophic polymorphism, phenotypic plastic-ity plays an important role in intraspecifi c phenotypic variation, as discussed later.
Recent advances in molecular techniques have stimulated studies to elucidate the genetic mechanisms of phenotypic polymorphism in fi shes: the techniques include comparative genomics, genetic mapping, candidate gene approaches, transcrip-tomics and so on (Ellegren and Sheldon 2008 ). Quantitative trait loci (QTL) analy-sis, one of the linkage mapping, is a powerful approach for identifying genome regions that contain genes responsible for variation in quantitative traits. Since the QTL analysis requires intercross and backcross between different populations, which have divergent phenotypic traits but are reproductively compatible to each other, species pairs are considered a promising subject for this analysis. Threespine sticklebacks are among the best understood species in fi shes in relation to the genetic basis on life history and trophic polymorphisms with the application of the QTL (Peichel et al. 2001 ; Colosimo et al. 2004 ; Cresko et al. 2004 ; Shapiro et al. 2004 ; Kimmel et al. 2005 ). Although we have diffi culty in applying this technique to wild populations of non-model organisms because of the labor intensive and time consuming work required to get their genomic resources, the development of high throughput next generation sequencers will enable us to effi ciently perform it with wild fi sh populations (Baird et al. 2008 ; Kakioka et al. 2013b ).
Because the QTL can only detect genome regions containing or linked to target genes, positional cloning and nomination of candidate genes are required to fi nd causative genetic variants. Colosimo et al. ( 2005 ) is the fi rst to identify a particular gene involved in the expression of divergent phenotypes in wild fi sh populations. In threespine sticklebacks, landlocked populations evolved from fully armored anad-romous ancestors and experienced armor reduction repeatedly in different locations (Barrett 2010 ). Colosimo et al. ( 2005 ) demonstrated that the ectodysplasin gene, which is involved in proper development of a number of ectodermal derivatives and dermal bones in mammals, was responsible for the armor reduction in landlocked threespine sticklebacks by means of genetic mapping, sequencing and transgenic
2 Predator Diversity Changes the World: From Gene to Ecosystem

31
techniques. Surprisingly, the causative alleles were shared among widely separated populations, suggesting a molecular genetic basis of the rapid parallel evolution of phenotypes adaptive to freshwater habitats.
Transcriptome profi ling is also a promising tool for unraveling complex molecular genetic basis of phenotypic divergence between closely relatives by examining their differential gene expression on the microarray. Using this technique, for example, Derome and others demonstrated in polymorphic populations of lake whitefi sh Coregonus clupeaformis that two sympatric morphs, a dwarf morph adaptive to lim-netic habitats and a normal morph to benthic habitats, showed differential expression of genes involved in energetic metabolism and regulation of muscle contraction, both of which are associated with swimming activity, and that their differential gene expres-sion was parallel among different lakes (Derome and Bernatchez 2006 ; Derome et al. 2006 ). This is reasonable when considering that the dwarf morphs are selected for high swimming performance in the limnetic habitats. More interestingly, the expres-sion of the same genes involved in modulating swimming activity were upregulated for a sympatric congener Coregonus artedi , a specialist competitor for the limnetic trophic niche, providing the molecular evidence of parallel phenotypic evolution.
While an increasing number of studies have reported evidence for the molecular genetic basis of phenotypic polymorphism in fi shes, they do not usually reject the pos-sibility of its environmental determination, i.e., phenotypic plasticity. Many research-ers have used common garden experiments to demonstrate that phenotypic plasticity of feeding morphology is induced by diet and/or habitat manipulation in fi sh popula-tions with polymorphic phenotypes (Hegrenes 2001 ; Andersson 2003 ; Andersson et al. 2005 ; Olsson and Eklöv 2005 ). In introduced populations, which have been recently colonized by a small number of individuals and, therefore, possess low genetic variation, trophic polymorphism may be substantially controlled by pheno-typic plasticity, as in the case of bluegills introduced into Japan (Yonekura et al. 2007a ; also see Sect. 2.4.5 ). Although it is hypothesized that the phenotypic plasticity can be controlled by epigenetics (i.e., heritable changes in gene expression or cellular phenotype caused by mechanisms other than changes in the underlying DNA sequence), its molecular mechanisms are poorly understood. In particular there is lit-tle published information regarding which key genes are involved in phenotypic plas-ticity of polymorphic traits and how they control developmental plasticity through a gene regulatory network (Sibthorpe et al. 2006 ). Without understanding the molecular mechanisms for phenotypic plasticity, diffi culties remain in discriminating between genetic and environmental effects on trophic polymorphism. Further advances in molecular techniques and their application to wild fi sh populations are needed to elu-cidate the genetic mechanisms underlying life history and trophic polymorphisms.
2.5 Niche Construction
A member of a biological community embedded within an ecosystem has a number of effects on other individuals simply by utilizing the space and energy provided by the ecosystem. These effects can sometimes alter ecosystem structure and
2.5 Niche Construction

32
functioning through ecological interactions between individuals or between biotic and abiotic components. In such an interaction network, species that have strong impacts on community and ecosystem processes are termed “keystone species” or “ecosystem engineers.” When “environment” is defi ned as “physical, chemical and biological properties outside the organism under consideration,” phenotypes of the keystone species can shape their environments. The environments altered by the keystone species may, in turn, act as a driver for the evolution of both itself and other members of the community. The process by which phenotypes expressed by a given species shapes the adaptive landscape of phenotypic evolution for itself or other species through alteration of biotic and abiotic attributes of their environments is termed “niche construction” (Odling-Smee et al. 2003 ). 1
In aquatic ecosystems, although there are a large number of examples of niche construction by fi sh acting as keystone predators, two key mechanisms of trophic cascade and nutrient transportation can be recognized (Fig. 2.2 ). Trophic cascade is defi ned as a food web consequence in which predators have indirect effects on non- prey organisms at trophic levels lower than those of their prey, through trophic inter-actions within a food web (Carpenter et al. 1985 ). For example, in the presence of planktivorous fi sh, large-bodied zooplankton are preferentially eliminated from the plankton community through fi sh predation (Brooks and Dodson 1965 ). This preda-tion effect can alter microbial communities including microalgae, bacteria and pro-tozoa. Since large-bodied zooplankton such as Daphnia have higher grazing rates, they depress growth of their microbial prey to a greater extent. Despite their micro-scopic size, microalgae and bacteria have large impacts on ecosystem processes, including production and decomposition rates, because of their large biomass and high metabolic turnover. When the abundance of planktivorous fi sh increases, microbial communities enhance their production and decomposition through alle-viation of predation by large-bodied zooplankton, resulting in an alteration of aquatic environments.
The mechanism of nutrient transportation is more effective in a situation where predator fi sh move from productive to less productive habitats (Vanni 2002 ). For example, when a large shoal of anadoromous fi sh migrate from the ocean to oligo-trophic streams and lakes, or when benthic fi sh exhibit vertical feeding migration from the lake bottom to surface waters in which nutrients are prone to be depleted, a supply of nutrients is transported to the less productive habitat as their excreta or their body mass themselves. The transported nutrients are quickly consumed by microalgae and bacteria, increasing their production and decomposition rates. This fi sh-driven nutrient transportation can have a substantial effect on material cycling within an ecosystem.
In this way, fi shes that are considered keystone predators of aquatic ecosystems have the potential to drastically alter community structure and ecosystem processes
1 Post and Palkovacs ( 2009 ) recommended restricting the use of the term “niche construction” to the process by which organisms shape environments in order to separate it from the process by which biologically constructed environments shape selection regimes for the evolution of organis-mal traits.
2 Predator Diversity Changes the World: From Gene to Ecosystem
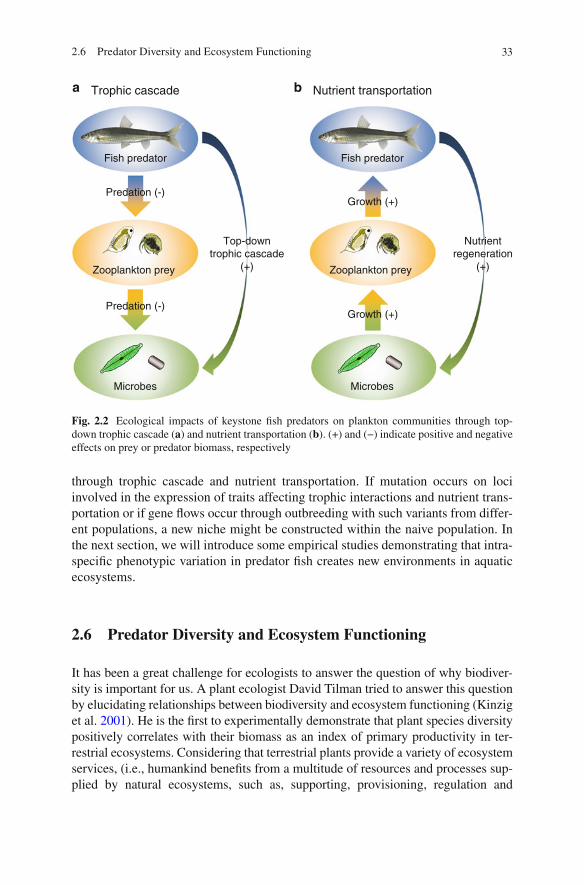
33
through trophic cascade and nutrient transportation. If mutation occurs on loci involved in the expression of traits affecting trophic interactions and nutrient trans-portation or if gene fl ows occur through outbreeding with such variants from differ-ent populations, a new niche might be constructed within the naive population. In the next section, we will introduce some empirical studies demonstrating that intra-specifi c phenotypic variation in predator fi sh creates new environments in aquatic ecosystems.
2.6 Predator Diversity and Ecosystem Functioning
It has been a great challenge for ecologists to answer the question of why biodiver-sity is important for us. A plant ecologist David Tilman tried to answer this question by elucidating relationships between biodiversity and ecosystem functioning (Kinzig et al. 2001 ). He is the fi rst to experimentally demonstrate that plant species diversity positively correlates with their biomass as an index of primary productivity in ter-restrial ecosystems. Considering that terrestrial plants provide a variety of ecosystem services, (i.e., humankind benefi ts from a multitude of resources and processes sup-plied by natural ecosystems, such as, supporting, provisioning, regulation and
Predation (-)
Top-downtrophic cascade
(+)
Fish predator
Zooplankton prey
Microbes
Trophic cascadea b
Predation (-)
Growth (+)
Fish predator
Zooplankton prey
Microbes
Nutrient transportation
Growth (+)
Nutrientregeneration
(+)
Fig. 2.2 Ecological impacts of keystone fi sh predators on plankton communities through top- down trophic cascade ( a ) and nutrient transportation ( b ). (+) and (−) indicate positive and negative effects on prey or predator biomass, respectively
2.6 Predator Diversity and Ecosystem Functioning

34
cultural services; see Millennium Ecosystem Assessment 2005 for defi nition), it is obvious that we profi t from the conservation of plant species diversity, which results in increased primary production. Inspired by his work, several theoretical models have been proposed and subsequently an increasing number of empirical studies have been conducted, for both animal and plant communities, to test the hypothesis that biodiversity enhances ecosystem functioning (Waide et al. 1999 ).
What about the role of predators? Unfortunately, very few studies have examined how the species diversity of predators affects community composition and ecosystem processes (Straub and Snyder 2006 ; Schmitz 2009 ). There are various reasons why studies on predator diversity effects are so limited. First, we have diffi culty in experi-mentally manipulating the species composition of predators, which have an extensive home range associated with their high mobility. Secondly, it is not easy to theoreti-cally predict how a combination of different predator species, which feed on prey at different trophic levels and from different trophic pathways, will impact on an ecosys-tem. Even if predator diversity effects are detected, we cannot easily interpret their mechanisms using theoretical models. Furthermore, even when effects on community composition and ecosystem processes can be quantifi ed for each predator species, it is impossible to conclude what traits actually lead to such community and ecosystem consequences because the species are different from each other in traits other than those related to feeding (Chalcraft and Resetarits Jr. 2003 ; Wesner 2012 ). However, there is a measure to overcome such diffi culties in demonstrating predator functional diversity effects on community structure and ecosystem processes, which is, to manipulate a diversity of trophic morphs or a species pair of fi sh predators.
As mentioned previously, trophic morphs are usually different only in trophic and/or life history traits and, otherwise, share all other ecological characteristics. By comparing different morphs or by manipulating a combination of these morphs within a predator population, we can examine how phenotypic diversity of a focal trait affects community and ecosystem properties. Based on this idea, Post et al. ( 2008 ) tried to demonstrate that intraspecifi c phenotypic variation among predator fi sh populations can alter prey community structure under natural conditions.
Post et al. ( 2008 ) focused on geographic variation in migration behavior and feed-ing morphology among alewife populations from 19 lakes in Connecticut, USA. They compared body size and biomass of zooplankton prey among lakes either with anad-romous or landlocked alewives or entirely without alewives. As stated in Sect. 2.4.4 , since the anadromous alewives have larger gape width and gill raker spacing than the landlocked alewives, the former preferentially feed on large- bodied zooplankton prey. In the anadromous lakes, alewives were absent during the winter season, so that large-bodied zooplankton dominated in the absence of strong zooplankton predators. However, spring breeding migration by the anadromous alewives and the subsequent recruitment of their YOY facilitated selective predation on large-bodied zooplankton, seasonally eliminating larger individuals from the zooplankton communities. By con-trast, in landlocked lakes in which alewives reside through the year, zooplankton com-munities are always exposed to intense and indiscriminate predation by landlocked YOY with smaller gape and gill raker spacing, perpetually skewing the zooplankton community size towards a smaller distribution.
2 Predator Diversity Changes the World: From Gene to Ecosystem

35
Intraspecifi c geographic variation over their life history and trophic polymor-phisms of alewives can also alter lake environments through top-down trophic cas-cades (Post et al. 2008 ). In the anadromous lakes, phytoplankton biomass increases during the seasonal reduction in zooplankton body size and biomass, suggesting that the anadromous YOY have positive, indirect effects on phytoplankton by depressing the grazing intensity of large-bodied zooplankton. The work of Post et al. ( 2008 ) is noteworthy as a milestone study demonstrating that intraspecifi c variation in predator feeding functions affects ecosystem processes such as primary productivity.
2.6.1 Mesocosm Experiments
Even though variations in prey community structure and ecosystem processes were found between lakes with different feeding functions within a predator species, they might be attributed to geographic variation in limno-physical and -chemical charac-teristics or to the density of other planktivorous fi sh. In order to reveal the ecosys-tem consequence of intraspecifi c predator functional diversity, experiments are required in which environmental factors, with the exception of predator feeding functions, are controlled. One promising approach is that of mesocosm experiments.
Since Forbes ( 1887 ) proposed the idea of “The Lake as a Microcosm,” in which lakes are considered as self-contained and circulating systems, the term “micro-cosm” has often been used in microbiology to refer to microbial cultivation systems containing essential components for their sustainable growth. Following experi-mental attempts to simulate lake ecosystems containing macro organisms such as zoobenthos and fi sh, the microcosms have been too small to ensure their sustain-ability. In order to fulfi ll energy and space requirements for the macro organisms, we have to provide them with a larger cultivation system. Such a self- contained experimental system, which is larger than microbial cultivation systems but smaller than natural ecosystems (i.e., on the meso-scale), is termed a “mesocosm.” Mesocosm experiments enable us to manipulate a given factor by controlling all other physical, chemical and biological environments and, thereby, to assess the ecosystem effects of the focal factor (Fig. 2.3 ).
An interesting mesocosm experiment considering trophic polymorphism in ale-wives was undertaken by Palkovacs and Post ( 2009 ) who constructed solid walled polyethylene enclosures with 2 m diameter and 6 m depth in Rogers Lake, Connecticut, USA. Plankton communities were exposed to different predation regimes by introducing 15 YOY fi sh of either anadromous or landlocked alewives into each enclosure in order to examine effects of predator feeding functions on prey communities. Some enclosures were left unstocked (i.e., no fi sh) as a control. Mesocosm experiments were conducted during the summer season when the YOY fi sh cause top-down trophic cascades in natural zooplankton communities. Weekly sampling of zooplankton communities from the enclosures was undertaken.
2.6 Predator Diversity and Ecosystem Functioning

36
Experimental results were consistent with those reported for natural lakes, with summer zooplankton body size and biomass signifi cantly reduced only in systems containing anadromous alewives. In addition, the presence of anadromous alewives led to a marked reduction in species richness and the diversity index of zooplankton communities. Palkovacs and Post ( 2009 ) were successful in demonstrating that intra-specifi c variation in predator feeding functions alone could alter zooplankton size distribution and species composition using these mesocosm experiments. This may also support the previous conclusion that variation between lakes in primary produc-tivity is due to differential trophic cascading effects caused by alewife morphs.
Similar results have been reported for another system exhibiting trophic poly-morphism in fi sh predators. Harmon et al. ( 2009 ) conducted mesocosm experiments with a species pair of threespine sticklebacks that underwent adaptive divergence to benthic (i.e., benthivorous) and limnetic (i.e., planktivorous) morphs from a gener-alist ancestor. To examine the effects of evolutionary diversifi cation of the threes-pine sticklebacks on ecosystem properties, they manipulated a combination of two specialist morphs or one generalist ancestor in large tanks to artifi cially simulate pond ecosystems including the plankton and benthos communities. They found strong effects of predator functional diversifi cation on prey community composition
Fig. 2.3 Twelve 2,000 L mesocosms tanks set up in Center for Ecological Research, Kyoto University. It is possible to automatically control water temperature, light intensity and L:D length. They are useful for experiments to examine effects of fi sh predator manipulation on ecosystem properties (see Sect. 2.6.2 )
2 Predator Diversity Changes the World: From Gene to Ecosystem

37
and ecosystem processes such as primary productivity and water chemistry. One of the more remarkable outcomes was that they could detect synergistic effects of the predators’ adaptive diversifi cation on the pond ecosystems as well as differential effects of each morph, by comparing ecosystem consequences between simple and diverse feeding functions. The results have ecological implications for the evolu-tionary diversifi cation of the sticklebacks because these morphs often coexist in natural ponds and lakes.
Intraspecifi c phenotypic diversity effects of fi sh predators can also appear through nutrient transportation. In guppies inhabiting streams in Trinidad, local populations are often isolated by natural barriers such as waterfalls and are exposed to different predation regimes from piscivorous fi sh. The high predation risk favors the evolution of life history traits in guppies, including a younger age at maturity, a smaller body size at maturity and higher reproductive rates. These life history traits are also linked with metabolic rate, which determines physiological performance such as respiration and excretion rate. The common garden experiments to manipu-late fi sh from different populations showed that phenotypic diversifi cation in gup-pies results in the divergence of community structure (benthic algal and invertebrate biomass) and ecosystem processes (gross primary productivity, leaf decomposition rates and nutrient fl ux), which are altered through nutrient transportation driven by intraspecifi c variation in the excretion rate of fi sh from different populations (Palkovacs et al. 2009 ; Bassar et al. 2010 ).
2.6.2 Food Web Properties as a Universal Measure
In previous sections, we cited some examples in which intraspecifi c phenotypic diversity in fi sh predators alters ecosystem properties. However, some ecosystem properties are system-specifi c (e.g., species composition) and others are just a func-tional trait of a specifi c guild (e.g., primary productivity by algae and decomposi-tion rate by bacteria and fungi) rather than ecosystem functions performed by an entire community. How then can we generalize from a variety of results from differ-ent systems and what is a universal measure for ecosystem properties, which are ubiquitous and intrinsic to all aquatic systems, thus allowing comparisons to be made between systems? Here we focus on structural properties of food web as a universal measure for the entire community.
A food web is a diagram to depict the overall energy fl ows through trophic inter-actions within an entire community (Lindeman 1942 ). Since the Eltonian pyramid (Elton 1927 ), a variety of ideas have been proposed to describe structural properties of intricate food webs in nature. One of promising measures to characterize food web properties is food chain length (FCL), defi ned as the number of trophic trans-fers of energy from primary producers to a top predator, which has been easily measured and comparable across ecosystems, with the recent development of isoto-pic technique (Post 2002 ). In lake ecosystems, the FCL can be the best explained by the lake size in volume (Post et al. 2000 ). However, it also varies temporally even
2.6 Predator Diversity and Ecosystem Functioning
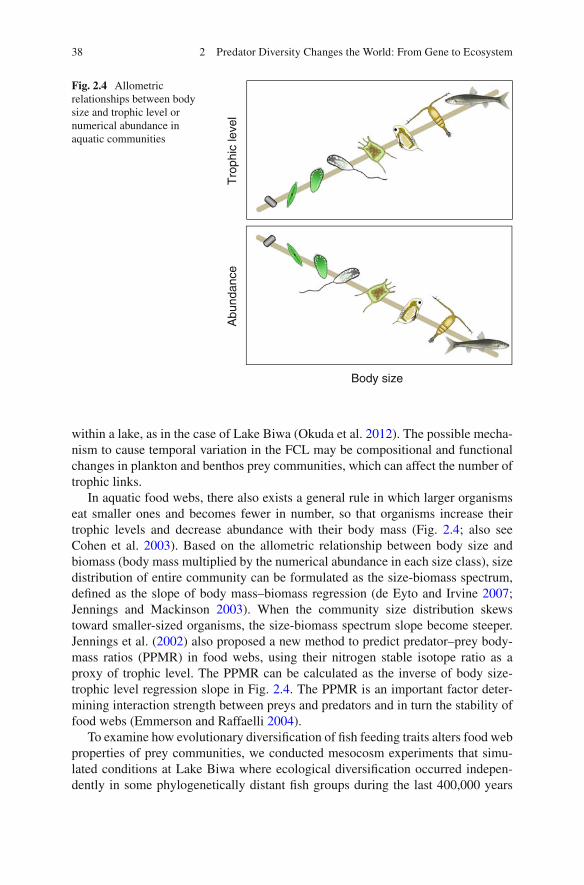
38
within a lake, as in the case of Lake Biwa (Okuda et al. 2012 ). The possible mecha-nism to cause temporal variation in the FCL may be compositional and functional changes in plankton and benthos prey communities, which can affect the number of trophic links.
In aquatic food webs, there also exists a general rule in which larger organisms eat smaller ones and becomes fewer in number, so that organisms increase their trophic levels and decrease abundance with their body mass (Fig. 2.4 ; also see Cohen et al. 2003 ). Based on the allometric relationship between body size and biomass (body mass multiplied by the numerical abundance in each size class), size distribution of entire community can be formulated as the size-biomass spectrum, defi ned as the slope of body mass–biomass regression (de Eyto and Irvine 2007 ; Jennings and Mackinson 2003 ). When the community size distribution skews toward smaller-sized organisms, the size-biomass spectrum slope become steeper. Jennings et al. ( 2002 ) also proposed a new method to predict predator–prey body- mass ratios (PPMR) in food webs, using their nitrogen stable isotope ratio as a proxy of trophic level. The PPMR can be calculated as the inverse of body size- trophic level regression slope in Fig. 2.4 . The PPMR is an important factor deter-mining interaction strength between preys and predators and in turn the stability of food webs (Emmerson and Raffaelli 2004 ).
To examine how evolutionary diversifi cation of fi sh feeding traits alters food web properties of prey communities, we conducted mesocosm experiments that simu-lated conditions at Lake Biwa where ecological diversifi cation occurred indepen-dently in some phylogenetically distant fi sh groups during the last 400,000 years
Tro
phic
leve
l
Body size
Abu
ndan
ce
Fig. 2.4 Allometric relationships between body size and trophic level or numerical abundance in aquatic communities
2 Predator Diversity Changes the World: From Gene to Ecosystem

39
after the development of deep pelagic habitats (see Sect. 2.4.3 ). We set up 12 meso-cosm tanks in which abiotic environments (i.e., light intensity, L:D length, water temperature and nutrient levels) were controlled and then a plankton community and supplementary zoobenthos prey were provided (Fig. 2.3 ). We manipulated the phenotypic diversity of feeding functions in fi sh predators, using a species pair of Gnathopogon that had diverged into planktivorous G. caerulescens and benthivo-rous G . elongatus elongatus but remained reproductively compatible to each other (see Sect. 2.4.3 ). We conducted experiments with a full-factorial design for four treatments: no fi sh (NO), benthivorous G . elongatus elongatus (B), and both plank-tivorous G. caerulescens and benthivorous G . elongatus elongatus (PB), and plank-tivorous G. caerulescens (P), assuming historical change in diversity of fi sh feeding functions in a lake (Fig. 2.5 ).
We measured the size-biomass spectrum, FCL and PPMR of plankton prey com-munities in the mesocosm tanks. It was predicted that planktivorous fi sh would skew the plankton communities toward a smaller size distribution through a top- down trophic cascading effect, under which a large zooplankton biomass would be depressed and a smaller plankton biomass would be indirectly enlarged, resulting in
Planktivores (P)
Benthivores (B)No fish (NO)
Benthivore & planktivore (BP)
G. Caerulescens
G. Caerulescens
G. Caerulescens
G. elongatus elongatus
G. elongatus elongatus
G. elongatus elongatus
a
c
b
d
Fig. 2.5 An experimental design of predator manipulation. Each treatment corresponds to histori-cal change in diversity of fi sh feeding functions in a lake. The lake has no fi sh in original condition ( a ), an ancestral benthivorous fi sh colonizes the lake ( b ), and then the colonized population experi-ences ecological speciation into benthivorous and planktivorous species ( c ). However, fi sh diver-sity decreases if either of these two becomes extinct. The ancestral benthivorous species may go extinct for some reason, e.g., destruction of coastal habitats under human disturbances ( d )
2.6 Predator Diversity and Ecosystem Functioning

40
a steeper slope of size-biomass spectrum in the order of NO < B < PB < P. Contrary to this prediction, the plankton size-biomass spectrum did not signifi cantly differ among these four treatments though the presence of fi sh predator increased the bio-mass of phytoplankton (Fukumori et al. unpublished data). The benthivorous G . elongatus elongatus feeds mainly on benthos but also less effi ciently on plank-ton, as is often the case in other benthivorous fi shes. The mesocosm tanks with a volume of 2,000 L might not be large enough to control predation of ben-thivorous G . elongatus elongates on zooplankton.
However, we found signifi cant differences in the FCL and PPMR among the treatments. Under the presence of benthivorous fi sh, the plankton FCL, defi ned as the highest trophic level observed for all samples of size-fractionated plankton com-munities, increased and the PPMR of plankton communities decreased (Sakai 2013 ). Although underlying mechanisms for these results remain unclear, selective predation of the two fi shes on different species of large-sized zooplankton (e.g., cladocerans versus copepods) is likely to alter trophic relations within plankton communities. More interestingly, we detected synergetic effects of predator feeding functional diversity on both indices. Under the coexistence of fi sh with different feeding functions, fi sh production, particularly for benthivorous fi sh, increased, possibly due to reduced food resource competition between planktivorous and ben-thivorous fi sh: the benthivore can exploit benthic food exclusively in the absence of benthic competitors (Fukumori et al., unpublished data). Assuming that the nutrient excretion rate scales with fi sh body mass (Torres and Vanni 2007 ; McIntyre et al. 2008 ), the benthivorous fi sh coexisting with the planktivorous fi sh might enhance nutrient transportation from the benthic to the pelagic habitats, resulting in syner-getic effects of top-down and bottom-up trophic cascades on the plankton food webs. Our experiments demonstrate that predator phenotypic diversity can alter food web properties of prey communities and thus the overall energy fl ows within an ecosystem.
2.7 Eco-Evolutionary Feedbacks
There is no doubt that intraspecifi c phenotypic diversity of fi sh predators is an important driver for altering prey community composition and thus ecosystem prop-erties. However, we must also consider an overlooked point, which is that ecosys-tem alteration, as a consequence of fi sh phenotypic diversifi cation, creates new aquatic environments, i.e., niche construction, which can shape the adaptive land-scape for the evolution of predators themselves as well as prey species. Ecological interactions between a keystone species and other community members drive evo-lutionary changes in their adaptive traits through environmental alterations and, in turn, the adaptive evolutionary changes shape the form of ecological interactions. This process, in which ecology and evolution reciprocally interplay over contempo-rary time-scales, is termed “eco-evolutionary feedback” (Post and Palkovacs 2009 ;
2 Predator Diversity Changes the World: From Gene to Ecosystem

41
Fig. 2.6 ). Although eco-evolutionary feedbacks are poorly understood, some studies have provided evidence that such feedbacks do operate in natural conditions.
2.7.1 Feedback Loops
A good example of a widely studied system is that of alewife-zooplankton interac-tions in lakes. In landlocked lakes, intense and constant predation by alewives elimi-nates large-bodied zooplankton and skews the prey size toward a smaller distribution throughout the year. This results in individuals with smaller gape width and gill raker spacing being favored when foraging on smaller prey. Such a situation will place the landlocked alewives under strong directional selection for feeding on small-bodied zooplankton, driving the adaptive evolution of the landlocked morph from an anadromous ancestor (Palkovacs and Post 2008 ). In addition, sporadic occurrences of physical barriers to fi sh migration between freshwater and marine habitats promotes the parallel evolution of landlocked morphs independently in dif-ferent locations, resulting in spatio-temporal heterogeneity in the geographic distri-bution of feeding traits within the species (Palkovacs et al. 2008 ).
It has been also suggested for Gnathopogon fi shes that planktivorous morphs evolved from the fl uvial ancestors with benthivorous morph independently in some lakes where
Nicheconstruction
Gene Individual
Ecosystem properties
Keystone predators
Expression
ImpactNatural selection
Environment
Population&
Community
ActionCharacterization
Evolutionary diversification
Fig. 2.6 A conceptual model for eco-evolutionary feedbacks. In aquatic ecosystems, fi sh preda-tors often have evolved phenotypic polymorphism as a consequence of intraspecifi c resource par-titioning in colonized populations or under newly created environments. When they alter ecosystem properties through compositional changes of prey communities, a new niche is constructed, which in turn molds fi tness landscapes for themselves and other species, feeding back to metagenomic changes in the whole community
2.7 Eco-Evolutionary Feedbacks

42
there exist few fi sh competitors for zooplankton prey (Kakioka et al. 2013b ). Among these populations with the great geographic variation in geometric morphology, a popu-lation endemic to Lake Biwa seems to have been most adapted for plankton feeding in pelagic environments, with its longest evolutionary history (Kakioka 2013 ).
Geographic variation in keystone predator phenotypes also has the potential to drive evolutionary divergence in their prey as well as to alter food web properties of prey communities. Walsh and Post ( 2011 ) recently evaluated the link between pheno-typic variation in predators and their prey species by rearing water fl eas, Daphnia , from lakes containing predatory alewives with different phenotypes under different water temperature and resource conditions, and then comparing their life history traits among these lakes. They found that Daphnia grew faster, matured earlier, and pro-duced more offspring in lakes with anadromous alewives than in lakes with land-locked or no alewives. Although life history theory predicts that increased predation on large sized-zooplankton selects for maturity at smaller size, laboratory experiments showed that Daphnia matured at a slightly larger size in anadromous lakes, in which alewives have both a larger gape width and gill raker spacing to preferentially forage larger prey (see Sect. 2.4.4 ). This result seems to be inconsistent with the theory. The evolutionary response of Daphnia to divergent predation regimes can be best explained as an adaptation to a colder environment and shorter growing season known as coun-tergradient variation ( sensu Conover et al. 2009 ). For Daphnia from the anadromous lakes, the colder environment and shorter growing season results from the seasonal migration and predation by anadromous alewives, which substantially extirpated reproductive populations of Daphnia early in the summer. In this prey–predator sys-tem, prey life history evolution is facilitated by seasonal occurrence of increased pre-dation, associated with predator life history traits rather than predation intensity and size-specifi c predation associated with predator feeding traits.
Walsh and Post ( 2012 ) also focused on how predator evolutionary divergence can mold the evolution of phenotypic plasticity in prey life history traits. It is well known that Daphnia alternate between sexual and asexual phases, producing parthenogenetic eggs (i.e., fully developed eggs without fertilization) throughout most of the year and sexual resting eggs during periods when environmental conditions deteriorate. Increased occurrence of predators selects for prey individuals that pass through the period of high predation as resting eggs. In lakes with landlocked alewives, Daphnia are under intense year-round predation, whereas they experience temporal but predict-able predation in the anadromous lakes into which alewives seasonally migrate from marine habitats. According the theory, the temporal variation of alewife occurrence should affect a response of Daphnia engaged in sexual reproduction. To test this pre-diction, Daphnia from lakes with different alewife phenotypes were exposed to ale-wife kairomones as an environmental cue for the presence of predators. They found that, when exposed to the fi sh kairomones, Daphnia from the anadromous lakes invested more in sexual reproduction than those from the landlocked or alewife free lakes, suggesting a fl exible life history trade-off in response to the predation risk. In conclusion, the divergent evolution of predator phenotypes has evolutionary conse-quences for phenotypic plasticity of prey life history traits.
2 Predator Diversity Changes the World: From Gene to Ecosystem

43
2.7.2 Community Genomic Dynamics
In lake ecosystems, eco-evolutionary feedback loops can also cascade from Daphnia to microbes, such as bacteria, microalgae and protozoa, because Daphnia has strong impacts on microbial communities through resource com-petition and nutrient regeneration, due to its high grazing ability and excretion rate (Carpenter et al. 1985 ; Carpenter et al. 1992 ). However, such cascading effects have been hitherto evaluated only as the overall response of each tro-phic guild (e.g., primary producer, primary consumer or decomposer) or func-tional group (i.e., pico-, nano- or micro- plankton) rather than of each microbial taxon. Microbial diversity, particularly of bacteria, is primarily important in determining ecosystem functioning and, thus, ecosystem services for human-kind, including the decomposition of organic matter, nutrient cycling and water purification (Bell et al. 2005 ; Cardinale 2011 ). Prompt responses of bacterial communities to changing environments are achieved by rapid evolution rates as well as by species turnover (Fukami et al. 2007 ; Gravel et al. 2011 ). Decades ago, the largest obstacle to disentangling the intricate relationships between bacterial diversity and ecosystem functioning was the difficulty in character-izing bacterial species composition in situ because most of them are unculti-vable and thus cannot be isolated for species identification. However, recent advances in molecular techniques and bioinformatics have developed metage-nomics or ecogenomics to characterize a profile of community-level genetic diversity in situ or comprehensively search for functional genes involved in phenotypic expression (see Chap. 3 ). The advent of next generation sequencer technologies has also accelerated ecological research as well as advancing our understanding of the molecular phylogenetics of aquatic microbial communi-ties (Kalyuzhnaya et al. 2008 ).
As discussed previously, plankton communities can be affected by pheno-typic polymorphism in fish predators. Recent molecular studies on fish pheno-typic polymorphism suggest that alleles at a small number of loci have large effects on adaptively divergent traits (Colosimo et al. 2005 ), contrary to tradi-tional views of quantitative genetics that consider polygenic variation in a large number of loci to have additive small effects on phenotypes. If it is general in fishes, such a gene with large effects may act as a “keystone gene” to influence metagenomic dynamics of whole plankton communities and, consequently, ecosystem properties. This is known to occur in terrestrial ecosystems where genetic variation within a population of foundation plant species is inherited through its phenotypic expression (e.g., concentration of chemical defense sub-stances) to community and ecosystem phenotypes (e.g., species composition and interactions, nutrient cycling, and decomposition rate) (Whitham et al. 2006 ). It will not be long before we can examine the nature of the eco-evolu-tionary feedbacks as community genomic dynamics under water, through a molecular-scope.
2.7 Eco-Evolutionary Feedbacks

44
2.8 Why is Predator Diversity Important?
Traditionally, evolutionary biologists have searched for an answer to the questions of how and when current biological communities diversifi ed using fossil and molec-ular records, which help to infer the past changes in morphology and DNA sequence. However, ecologists have been more interested in what are the ecological implica-tions for evolutionary diversifi cation of current biological communities, often assuming that species’ traits are static rather than variable. As demonstrated by an increasing number of studies, we now perceive that organismal traits are evolving at a faster rate than was previously assumed (Hairston et al. 2005 ), which leads to a new idea that the evolution of the organismal traits dynamically interplays with the ecology of populations, communities and ecosystems, and vice-versa (Post and Palkovacs 2009 ). Biological diversity observed in nature can therefore be viewed not only as a consequence of past evolutionary events but also as a product of on- going evolution in existing communities. Diversity currently found within/among keystone predator populations can have the potential to infl uence the future direc-tion of diversity at three levels of biological organization, from genes to ecosystems, through eco-evolutionary feedbacks.
At present, species extinction has progressed rapidly, with an accelerating rate, due to anthropogenic disturbances ranging from local human activities to global climate change. Biodiversity loss is particularly severe in freshwater ecosystems (Secretariat of the Convention on Biological Diversity 2010 ). Although much atten-tion has been paid to the conservation of species, specifi cally those that are endan-gered, we also have to conserve the functional diversity of keystone species within/among populations, which may not necessarily be endangered. The effects of preda-tor functional diversity can sometimes extend to ecosystem functioning, such as energy fl ows and nutrient cycling, from which we can enjoy a variety of ecosystem services profi table for human well-being.
In this chapter, the ecological and evolutionary importance of functional diver-sity of fi sh predators in the context of ecosystem management, as well as of tradi-tional conservation biology, has been discussed. In order to deepen our understanding of the processes and mechanisms for generating and maintaining biological diver-sity, we have to further accumulate empirical data and establish a framework to integrate the theory of eco-evolutionary feedbacks. This idea is expected to develop a new paradigm for biological synthesis, linking from molecular biology to macroecology.
Acknowledgements We thank Dr. T. Kokita for his comments on our manuscript. We also thank R. Kakioka and Y. Sakai for providing their unpublished data. This manuscript was under the sup-port of the Global COE Program (06) from the MEXT, Japan, and The Environment Research and Technology Development Fund (D-1102 and S-9) of the Ministry of the Environment, Japan, and the JSPS Grant-in Aid (No.20370009 and No.23657019).
2 Predator Diversity Changes the World: From Gene to Ecosystem

45
References
Agrawal AA (2001) Phenotypic plasticity in the interactions and evolution of species. Science 294:321–326. doi: 10.1126/science.1060701
Andersson J (2003) Effects of diet-induced resource polymorphism on performance in arctic charr ( Salvelinus alpinus ). Evol Ecol Res 5:213–228. http://cat.inist.fr/?aModele=affi cheN&cpsidt=14588743
Andersson J, Byström P, Persson L, De Roos AM (2005) Plastic resource polymorphism: effects of resource availability on Arctic char ( Salvelinus alpinus ) morphology. Biol J Linn Soc 85:341–351. doi: 10.1111/j.1095-8312.2005.00501.x
Baird NA, Etter PD, Atwood TS, Currey MC, Shiver AL, Lewis ZA, Selker EU, Cresko WA, Johnson EA (2008) Rapid SNP discovery and genetic mapping using sequenced RAD markers. PLoS One 3:e3376. doi: 10.1371/journal.pone.0003376
Barluenga M, Stölting KN, Salzburger W, Muschick M, Meyer A (2006) Sympatric speciation in Nicaraguan crater lake cichlid fi sh. Nature 439:719–723
Barrett RDH (2010) Adaptive evolution of lateral plates in three-spined stickleback Gasterosteus aculeatus: a case study in functional analysis of natural variation. J Fish Biol 77:311–328. doi: 10.1111/j.1095-8649.2010.02640.x
Bassar RD, Marshall MC, López-Sepulcre A, Zandonà E, Auer SK, Travis J, Pringle CM, Flecker AS, Thomas SA, Fraser DF, Reznick DN (2010) Local adaptation in Trinidadian guppies alters ecosystem processes. Proc Natl Acad Sci U S A 107:3616–3621. doi: 10.1073/pnas.0908023107
Bell T, Newman JA, Silverman BW, Turner SL, Lilley AK (2005) The contribution of species rich-ness and composition to bacterial services. Nature 436:1157–1160. doi: 10.1038/nature-03891Bolnick DI
Bolnick DI, Svanbäck R, Fordyce JA, Yang LH, Davis JM, Hulsey CD, Forister ML (2003) The ecol-ogy of individuals: incidence and implications of individual specialization. Am Nat 161:1–28. http://www.jstor.org/stable/10.1086/343878
Bootsma HA, Hecky RE, Hesslein RH, Turner GF (1996) Food partitioning among Lake Malawi nearshore fi shes as revealed by stable isotope analyses. Ecology 77:1286–1290. http://www.jstor.org/discover/10.2307/2265598?uid=3739920&uid=2134&uid=2&uid=70&uid=4&uid=3739256&sid=55941630443
Brooks JL, Dodson SI (1965) Predation, body size, and competition of plankton. Science 150:28–35. http://www.jstor.org/discover/10.2307/1717947
Cardinale BJ (2011) Biodiversity improves water quality through niche partitioning. Nature 472:86–89. doi: 10.1038/nature09904
Carpenter SR, Kitchell JF, Hodgson JR (1985) Cascading trophic interactions and lake productiv-ity. BioSci 35:634–639. http://www.jstor.org/discover/10.2307/1309989?uid=3739920&uid=2&uid=4&uid=3739256&sid=55941682303
Carpenter SR, Cottingham KL, Schindler DE (1992) Biotic feedbacks in lake phosphorus cycles. Trends Ecol Evol 7:332–336. doi.org/10.1016/0169-5347(92)90125-U
Chalcraft DR, Resetarits Jr. WJ (2003) Predator identity and ecological impacts: functional redun-dancy or functional diversity? Ecology 84:2407–2418. doi.org/10.1890/02-0550
Cohen JE, Jonsson T, Carpenter SR (2003) Ecological community description using the food web, species abundance, and body size. Proc Natl Acad Sci U S A 100:1781–1786
Colosimo PF, Peichel CL, Nereng K, Blackman BK, Shapiro MD, Schluter D, Kingsley DM (2004) The genetic architecture of parallel armor plate reduction in threespine sticklebacks. PLoS Biol 2:0635–0641. doi: 10.1371/journal.pbio.0020109
Colosimo PF, Hosemann KE, Balabhadra S, Villarreal G Jr, Dickson M, Grimwood J, Schmutz J, Myers RM, Schluter D, Kingsley DM (2005) Widespread parallel evolution in sticklebacks by repeated fi xation of ectodysplasin alleles. Science 307:1928–1933. doi: 10.1126/science.1107239
References

46
Conover DO, Duffy TA, Hice LA (2009) The covariance between genetic and environmental Infl uences across ecological gradients: reassessing the evolutionary signifi cance of countergradient and cogra-dient variation. Ann N Y Acad Sci 1168:100–129. doi: 10.1111/j.1749-6632.2009.04575.x
Cresko WA, Amores A, Wilson C, Murphy J, Currey M, Phillips P, Bell MA, Kimmel CB, Postlethwait JH (2004) Parallel genetic basis for repeated evolution of armor loss in Alaskan threespine stickleback populations. Proc Natl Acad Sci U S A 101:6050–6055. doi: 10.1073/pnas.0308479101
de Eyto E, Irvine K (2007) Assessing the status of shallow lakes using an additive model of bio-mass size spectra. Aquat Conserv Mar Freshw Ecosyst 17:724–736
Derome N, Bernatchez L (2006) The transcriptomics of ecological convergence between 2 lim-netic coregonine fi shes (Salmonidae). Mol Biol Evol 23:2370–2378
Derome N, Duchesne P, Bernatchez L (2006) Parallelism in gene transcription among sympatric lake whitefi sh ( Coregonus clupeaformis Mitchill) ecotypes. Mol Ecol 15:1239–1249
Ehlinger TJ (1990) Habitat choice and phenotype-limited feeding effi ciency in bluegill: individual differences and trophic polymorphism. Ecology 71:886–896. http://www.jstor.org/discover/10.2307/1937360?uid=3739920&uid=2&uid=4&uid=3739256&sid=55941682303
Ehlinger TJ (1991) Allometry and analysis of morphometric variation in the bluegill, Lepomis macrochirus . Copeia 1991:347–357. http://www.jstor.org/discover/10.2307/1446584?uid=3739920&uid=2&uid=4&uid=3739256&sid=55941682303
Ellegren H, Sheldon BC (2008) Genetic basis of fi tness differences in natural populations. Nature 452:169–175
Elton C (1927) Animal ecology. Sidgwick and Jackson, London Emmerson MC, Raffaelli D (2004) Predator–prey body size, interaction strength and the stability
of a real food web. J Anim Ecol 73:399–409 Foote CJ, Moore K, Stenberg K, Craig KJ, Wenburg JK, Wood CC (1999) Genetic differentiation in
gill raker number and length in sympatric anadromous and nonanadromous morphs of sockeye salmon, Oncorhynchus nerka . Environ Biol Fish 54:263–274. doi: 10.1023/A:1007548807233
Forbes SA (1887) The lake as a microcosm. Bulletin of the Peoria Scientifi c Association 77–87 Fukami T, Beaumont HJE, Zhang X-X, Rainey PB (2007) Immigration history controls diversifi -
cation in experimental adaptive radiation. Nature 446:436–439. doi: 10.1038/nature05629 Genner MJ, Turner GF, Barker S, Hawkins SJ (1999) Niche segregation among Lake Malawi
cichlid fi shes? Evidence from stable isotope signatures. Ecol Lett 2:185–190. doi: 10.1046/j.1461-0248.1999.00068.x
Gravel D, Bell T, Barbera C, Bouvier T, Pommier T, Venail P, Mouquet N (2011) Experimental niche evolution alters the strength of the diversity–productivity relationship. Nature 469:89–92. doi: 10.1038/nature09592
Gross MR (1996) Alternative reproductive strategies and tactics: diversity within sexes. Trends Ecol Evol 11:92–98. http://dx.doi.org/10.1016/0169-5347(96)81050-0
Hairston NG Jr, Ellner SP, Geber MA, Yoshida T, Fox JA (2005) Rapid evolution and the convergence of ecological and evolutionary time. Ecol Lett 8:1114–1127. doi: 10.1111/j.1461-0248.2005.00812.x
Harmon LJ, Matthews B, Des Roches S, Chase JM, Shurin JB, Schluter D (2009) Evolutionary diversifi cation in stickleback affects ecosystem functioning. Nature 458:1167–1170. doi: 10.1038/nature07974
Hegrenes S (2001) Diet-induced phenotypic plasticity of feeding morphology in the orangespotted sunfi sh, Lepomis humilis . Ecol Freshw Fish 10:35–42. doi: 10.1034/j.1600-0633.2001.100105.x
Hendry AP (2009) Ecological speciation! Or the lack thereof? Can J Fish Aquat Sci 66:1383–1398. doi: 10.1139/F09-074
Hendry AP, Wenburg JK, Bentzen P, Volk EC, Quinn TP (2000) Rapid evolution of reproductive isolation in the wild: evidence from introduced salmon. Science 290:516–518
Hendry AP, Bolnick DI, Berner D, Peichel CL (2009) Along the speciation continuum in stickle-backs. J Fish Biol 75:2000–2036
Hendry AP, Hudson K, Walker JA, Räsänen K, Chapman LJ (2011) Genetic divergence in mor-phology–performance mapping between Misty Lake and inlet stickleback. J Evol Biol 24:23–35. doi: 10.1111/j.1420-9101.2010.02155.x
2 Predator Diversity Changes the World: From Gene to Ecosystem

47
Jennings S, Mackinson S (2003) Abundance–body mass relationships in size-structured food webs. Ecol Let 6:971–974
Jennings S, Warr KJ, Mackinson S (2002) Use of size-based production and stable isotope analy-ses to predict trophic transfer effi ciencies and predator–prey body mass ratios in food webs. Mar Ecol Prog Ser 240:11–20
Kakioka R (2013) Adaptive divergences of cyprinid fi shes in relation to divergent habitat use. Doctoral Thesis, Kyoto University, Kyoto
Kakioka R, Kokita T, Tabata R, Mori S, Watanabe K (2013a) The origins of limnetic forms and cryptic divergence in Gnathopogon fi shes (Cyprinidae) in Japan. Environ Biol Fish 96:631–644. doi: 10.1007/s10641-012-0054-x
Kakioka R, Kokita T, Kumada H, Watanabe K, Okuda N (2013b) A RAD-based linkage map for QTL analysis and comparative genomics in the gudgeons (genus Gnathopogon , Cyprinidae). BMC Genomics 14:32. http://www.biomedcentral.com/1471-2164/14/32
Kalyuzhnaya MG, Lapidus A, Ivanova N, Copeland AC, McHardy AC, Szeto E, Salamov A, Grigoriev IV, Suciu D, Levine SR, Markowitz VM, Rigoutsos I, Tringe SG, Bruce DC, Richardson PM, Lidstrom ME, Chistoserdova L (2008) High-resolution metagenomics targets specifi c functional types in complex microbial communities. Nat Biotechnol 26:1029–1034. doi: 10.1038/nbt.1488
Kawamura K, Yonekura R, Katano O, Taniguchi Y, Saitoh K (2006) Origin and dispersal of blue-gill sunfi sh, Lepomis macrochirus , in Japan and Korea. Mol Ecol 15:613–621. doi: 10.1111/j.1365-294X.2006.02823.x
Kimmel CB, Ullmann B, Walker C, Wilson C, Currey M, Phillips PC, Bell MA, Postlethwait JH, Cresko WA (2005) Evolution and development of facial bone morphology in threespine stick-lebacks. Proc Natl Acad Sci U S A 102:5791–5796. doi: 10.1073/pnas.0408533102
Kinzig AP, Pacala SW, Tilman D (2001) The functional consequences of biodiversity: empirical progress and theoretical extensions. Princeton University Press, Princeton
Knudsen R, Primicerio R, Amundsen P-A, Klemetsen A (2010) Temporal stability of indi-vidual feeding specialization may promote speciation. J Anim Ecol 79:161–168. doi: 10.1111/j.1365-2656.2009.01625.x
Lass S, Spaak P (2003) Chemically induced anti-predator defences in plankton: a review. Hydrobiologia 491:221–239. doi: 10.1023/A:1024487804497
Lindeman RL (1942) The trophic-dynamic aspect of ecology. Ecology 23:399–417 McIntyre PB, Flecker AS, Vanni MJ, Hood JM, Taylor BW, Thomas SA (2008) Fish distributions
and nutrient cycling in streams: can fi sh create biogeochemical hotspots? Ecology 89:2335–2346. http://dx.doi.org/10.1890/07-1552.1
McKinnon JS, Rundle HD (2002) Speciation in nature: the threespine stickleback model systems. Trends Ecol Evol 17:480–488. http://dx.doi.org/10.1016/S0169-5347(02)02579-X
Millennium Ecosystem Assessment (2005) Ecosystems and human well-being: synthesis. Island, Washington, DC
Miner BG, Sultan SE, Morgan SG, Padilla DK, Relyea RA (2005) Ecological consequences of phe-notypic plasticity. Trends Ecol Evol 20:685–692. http://dx.doi.org/10.1016/j.tree.2005.08.002
Nakamura M (1969) Cyprinid fi shes in Japan. Midori Shobo, Tokyo (in Japanese) Odling-Smee FJ, Laland KN, Feldman MW (2003) Niche construction: the neglected process in
evolution. Princeton University Press, Princeton Okuda N, Takeyama T, Komiya T, Kato Y, Okuzaki Y, Karube J, Sakai Y, Hori M, Tayasu I, Nagata
T (2012) A food web and its long-term dynamics in Lake Biwa: a stable isotope approach. In: Kawanabe H (ed) Lake Biwa: interactions between nature and people, Springer, Amsterdam, pp 205–210
Olsson J, Eklöv P (2005) Habitat structure, feeding mode and morphological reversibility: factors infl uencing phenotypic plasticity in perch. Evol Ecol Res 7:1109–1123
Osenberg CW, Huckins CJF, Kaltenberg A, Martinez A (2004) Resolving within- and between- population variation in feeding ecology with a biomechanical model. Oecologia 141:57–65. doi: 10.1007/s00442-004-1650-z
References

48
Palkovacs EP, Post DM (2008) Eco-evolutionary interactions between predators and prey: can predator-induced changes to prey communities feed back to shape predator foraging traits? Evol Ecol Res 10:699–720. http://www.yale.edu/post_lab/pdfs/Palkovacs%20and%20Post%202008%20%28EER%29.pdf
Palkovacs EP, Post DM (2009) Experimental evidence that phenotypic divergence in predators drives community divergence in prey. Ecology 90:300–305. http://www.esajournals.org/doi/pdf/10.1890/08-1673.1
Palkovacs EP, Dion KB, Post DM, Caccone A (2008) Independent evolutionary origins of land-locked alewife populations and rapid parallel evolution of phenotypic traits. Mol Ecol 17:582–597. doi: 10.1111/j.1365-294X.2007.03593.x
Palkovacs EP, Marshall MC, Lamphere BA, Lynch BR, Weese DJ, Fraser DF, Reznick DN, Pringle CM, Kinnison MT (2009) Experimental evaluation of evolution and coevolution as agents of ecosystem change in Trinidadian streams. Philos Trans R Soc Lond B 364:1617–1628. doi: 10.1098/rstb.2009.0016
Peichel CL, Nereng KS, Ohgi KA, Cole BLE, Colosimo PF, Buerkle CA, Schluter D, Kingsley DM (2001) The genetic architecture of divergence between threespine stickleback species. Nature 414:901–905. doi: 10.1038/414901a
Post DM (2002) The long and short of food-chain length. Trends Ecol Evol 17:269–277 Post DM, Palkovacs EP (2009) Eco-evolutionary feedbacks in community and ecosystem ecology:
interactions between the ecological theatre and the evolutionary play. Philos Trans R Soc Lond B 364:1629–1640. doi: 10.1098/rstb.2009.0012
Post DM, Pace ML, Hairston NG Jr (2000) Ecosystem size determines food-chain length in lakes. Nature 405:1047–1049
Post DM, Palkovacs EP, Schielke EG, Dodson SI (2008) Intraspecifi c variation in a predator affects com-munity structure and cascading trophic interactions. Ecology 89:2019–2032. doi: 10.1890/07-1216.1
Proulx R, Magnan P (2004) Contribution of phenotypic plasticity and heredity to the trophic poly-morphism of lacustrine brook charr ( Salvelinus fontinalis M.). Evol Ecol Res 6:503–522. http://www.bgc-jena.mpg.de/bgc-organisms/uploads/People/Proulx-Magnan-EER-2004.pdf
Robinson BW (2000) Trade offs in habitat-specifi c foraging effi ciency and the nascent adaptive divergence of sticklebacks in lakes. Behaviour 137:865–888. http://www.jstor.org/discover/10.2307/4535745?uid=3739920&uid=2&uid=4&uid=3739256&sid=55942013823
Rogers SM, Bernatchez L (2007) The genetic architecture of ecological speciation and the associa-tion with signatures of selection in natural lake whitefi sh ( Coregonus sp . Salmonidae) species pairs. Mol Biol Evol 24:1423–1438. doi: 10.1093/molbev/msm066
Sacotte S, Magnan P (2006) Inherited differences in foraging behaviour in the offspring of two forms of lacustrine brook charr. Evol Ecol Res 8:843–857. http://www.uqtr.ca/GREA/pdf/publi/sacotte.pdf
Sakai Y (2013) Spatio-temporal dynamics of planktonic food webs in the coastal ecosystem of Lake Biwa. Doctoral Thesis, Kyoto University, Kyoto
Schmitz OJ (2009) Effects of predator functional diversity on grassland ecosystem function. Ecology 90:2339–2345. http://dx.doi.org/10.1890/08-1919.1
Secretariat of the Convention on Biological Diversity (2010) Global Biodiversity Outlook 3. An open access publication ( http://creativecommons.org/licenses/by-nc/3.0/ ), Montréal
Seehausen O (2006) African cichlid fi sh: a model system in adaptive radiation research. Proc R Soc Lond B 273:1987–1998. doi: 10.1098/rspb.2006.3539
Seehausen O, Terai Y, Magalhaes IS, Carleton KL, Mrosso HDJ, Miyagi R, van der Sluijs I, Schneider MV, Maan ME, Tachida H, Imai H, Okada N (2008) Speciation through sensory drive in cichlid fi sh. Nature 455:620–626. doi: 10.1038/nature07285
Shapiro MD, Marks ME, Peichel CL, Blackman BK, Nereng KS, Jónsson B, Schluter D, Kingsley DM (2004) Genetic and developmental basis of evolutionary pelvic reduction in threespine sticklebacks. Nature 428:717–723. doi: 10.1038/nature02415
Sibthorpe D, Sturlaugsdóttir R, Kristjansson BK, Thorarensen H, Skúlason S, Johnston IA (2006) Characterisation and expression of the paired box protein 7 (Pax7) gene in polymorphic Arctic charr ( Salvelinus alpinus ). Comp Biochem Physiol B 145:371–383
Smith TB, Skulason S (1996) Evolutionary signifi cance of resource polymorphisms in fi shes, amphibians, and birds. Ann Rev Ecol Syst 27:111–133. http://www.jstor.org/discover/10.2307/2097231?uid=3739920&uid=2&uid=4&uid=3739256&sid=55943041433
2 Predator Diversity Changes the World: From Gene to Ecosystem

49
Straub CS, Snyder WE (2006) Species identity dominates the relationship between predator biodi-versity and herbivore suppression. Ecology 87:277–282. http://dx.doi.org/10.1890/05-0599
Taylor EB (1999) Species pairs of north temperate freshwater fi shes: Evolution, taxonomy, and conservation. Rev Fish Biol Fish 9:299–324. doi: 10.1023/A:1008955229420
Terashima A (1980) Bluegill: a vacant ecological niche in Lake Biwa. In: Kawai T, Kawanabe H, Mizuno N (eds) Japanese freshwater organisms: ecology of invasion and disturbance. Tokai University Press, Tokyo, pp 63–70
Timoshkin OA, Grygier MJ, Wada E, Nakai K, Nishino M, Genkal SI, Biserov VI, Gagarin VG, Semernoy VP, Jankowski AW, Stepanjants SD, Tsalolikhin SY, Starobogatov YI, Alexeev VR, Tuzovskij PV, Okuneva GL, Sheveleva NG, Pomazkova GI, Arov IV, Mazepova GF, Janz H, Obolkina LA, Chernyshev AV, Morino H, Matsuda M, Ohtsuka T, Kawakatsu M, Maehata M, Masuda Y, Faubel A, Yahiro K, Hirasawa R, Tuji A, Kusuoka Y, Kameda K, Ishida T, Itoh T, Ichise S, Wakabayashi T, Okubo I, Seki S, Nagasawa K, Ogawa K, Masunaga K,Gamo J (2011) Biodiversity of Lake Biwa: new discoveries and future potential. In: Timoshkin OA (ed) Index of animal species inhabiting Lake Baikal and its Catchment Area. Volume II: Basins and Channels in the South of East Siberia and North Mongolia, vol 2. Russian Academy of Sciences, Siberian Division, Limnological Institute, Novosibirsk, pp 1439–1513
Torres LE, Vanni MJ (2007) Stoichiometry of nutrient excretion by fi sh: interspecifi c variation in a hypereutrophic lake. Oikos 116:259–270. doi: 10.1111/j.0030-1299.2007.15268.x
Uchii K (2007) Adaptive signifi cance of a herbivorous morph in blugill sunfi sh introduced into Lake Biwa. Doctoral Thesis, Kyoto University, Kyoto
Uchii K, Okuda N, Yonekura R, Karube Z, Matsui K, Kawabata Z (2007) Trophic polymorphism in bluegill sunfi sh ( Lepomis macrochirus ) introduced into Lake Biwa: evidence from stable isotope analysis. Limnology 8:59–63. doi: 10.1007/s10201-006-0196-7
Vanni MJ (2002) Nutrient cycling by animals in freshwater ecosystems. Ann Rev Ecol Syst 33:341–370. doi: 10.1146/annurev.ecolsys.33.010802.150519
Waide RB, Willig MR, Steiner CF, Mittelbach G, Gough L, Dodson SI, Juday GP, Parmenter R (1999) The relationship between productivity and species richness. Ann Rev Ecol Syst 30:257–300. http://www.jstor.org/stable/221686
Wainwright PC, Osenberg CW, Mittelbach GG (1991) Trophic polymorphism in the pumpkinseed sunfi sh ( Lepomis gibbosus Linnaeus): effects of environment on ontogeny. Funct Ecol 5:40–55. http://www.jstor.org/discover/10.2307/2389554?uid=3739920&uid=2&uid=4&uid=3739256&sid=55943041433
Walsh MR, Post DM (2011) Interpopulation variation in a fi sh predator drives evolutionary diver-gence in prey in lakes. Proc R Soc Lond B 278:2628–2637. doi: 10.1098/rspb.2010.2634
Walsh MR, Post DM (2012) The impact of intraspecifi c variation in a fi sh predator on the evolution of phenotypic plasticity and investment in sex in Daphnia ambigua . J Evol Biol 25:80–89. doi: 10.1111/j.1420-9101.2011.02403.x
Wesner JS (2012) Predator diversity effects cascade across an ecosystem boundary. Oikos 121:53–60. doi: 10.1111/j.1600-0706.2011.19413.x
Whitham TG, Bailey JK, Schweitzer JA, Shuster SM, Bangert RK, LeRoy CJ, Lonsdorf EV, Allan GJ, DiFazio SP, Potts BM, Fischer DG, Gehring CA, Lindroth RL, Marks JC, Hart SC, Wimp GM, Wooley SC (2006) A framework for community and ecosystem genetics: from genes to ecosystems. Nat Rev Genet 7:510–523. doi: 10.1038/nrg1877
Wilson DS (1998) Adaptive individual differences within single populations. Philos Trans R Soc Lond B 353:199–205. doi: 10.1098/rstb.1998.0202
Yonekura R, Nakai K, Yuma M (2002) Trophic polymorphism in introduced bluegill in Japan. Ecol Res 17:49–57. doi: 10.1046/j.1440-1703.2002.00462.x
Yonekura R, Kawamura K, Uchii K (2007a) A peculiar relationship between genetic diversity and adaptability in invasive exotic species: bluegill sunfi sh as a model species. Ecol Res 22:911–919. doi: 10.1007/s11284-007-0357-0
Yonekura R, Kohmatsu Y, Yuma M (2007b) Difference in the predation impact enhanced by mor-phological divergence between introduced fi sh populations. Biol J Linn Soc 91:601–610. doi: 10.1111/j.1095-8312.2007.00821.x
References

51N. Okuda et al., Biodiversity in Aquatic Systems and Environments: Lake Biwa, SpringerBriefs in Biology, DOI 10.1007/978-4-431-54150-9_3, © The Author(s) 2014
Abstract The food linkage between heterotrophic bacteria and protists is so-called “microbial loop,” functioning as important matter cycling in pelagic food webs. Biomass of heterotrophic bacteria sometimes predominates total heterotrophic bio-mass in lakes, and organic matter transfer started from heterotrophic bacteria has been intensively studied by numerous researchers all over the world. The roles of plank-tonic protists, such as heterotrophic nanofl agellates and ciliates, in microbial loop are to consume bacteria that are too small to serve directly as major prey items for most zooplankters, and to be themselves utilized by the zooplankton. There is a consensus that food linkages between bacteria and protists are substantial in many lakes. In addi-tion, bacterial loss due to viral lysis has also been paid great attention by many researchers since the late 1990. The present review provides the overview on the trend and future stage of microbial loop researches in freshwater systems, with special ref-erence to culture-independent molecular techniques such as PCR cloning and sequenc-ing, fl uorescently in situ hybridization or denaturing gradient gel electrophoresis for phylogenetic analyses on microbial communities. A review on ecology and biodiver-sity researches on microbial loop in Lake Biwa is also provided.
Keywords Bacteria • Biodiversity • Ciliates • Culture-independent molecular tech-niques • Lakes • Microbial food web • Microbial loop • Nanofl agellates • Protists
3.1 Introduction
In pelagic areas of lakes or oceans, matter cycling is mainly driven by planktonic food web. It is well known that the herbivorous food web where phytoplankton are preyed on zooplankton is one important matter cycling in pelagic ecosystems. By contrast, the food linkage between heterotrophic bacteria and protists, so-called “microbial loop,” has been known as another important matter cycling in pelagic food webs (Fig. 3.1 ) (Azam et al. 1983 ). Phytoplankton release dissolved organic matter (DOM)
Chapter 3 Biodiversity Researches on Microbial Loop in Aquatic Systems

52
as intermediate products of photosynthesis and/or as autolytic products (Riemann and Søndergaard 1986 ). DOM thus released is utilized for growth of heterotrophic bacte-ria, followed by protistan grazing on bacteria. Protists thereafter are consumed by meso-zooplankton such as cladocerans and/or copepods. Biomass of heterotrophic bacteria sometimes predominates total heterotrophic biomass in lakes (Kawabata and Nakanishi 1996 ), and organic matter transfer started from heterotrophic bacteria has been intensively studied by numerous researchers all over the world.
The roles of planktonic protists, such as heterotrophic nanofl agellates and cili-ates, in microbial loop are to consume bacteria that are too small to serve directly as major prey items for most zooplankters, and to be themselves utilized by the zoo-plankton (Azam et al. 1983 ). There is a consensus that food linkages between bac-teria and protists are substantial in many lakes (Nagata 1988 ; Bloem and Bar-Gilissen 1989 ; Sanders et al. 1989 ; Šimek et al. 1990 , 1995 ; Šimek and Straskrabova 1992 ; Nakano et al. 1998a , b ). In addition, bacterial loss due to viral lysis has also been paid attention by many researchers since the late 1990 (Suttle 2007 ).
The present review provides the overview on the trend and future stage of micro-bial loop researches mainly in freshwater systems, together with some related researches in marine systems, during the last decade. In addition, I review biodiversity researches on microbial loop in Lake Biwa, together with a perspective on them.
3.2 Towards More Complicated Microbial Food Web
Bergh et al. ( 1989 ) reported density of virus in freshwaters (254 × 10 6 particles mL −1 ) is higher than that of marine waters (up to 14.9 × 10 6 particles mL −1 ). Due to stimulation by Bergh et al. ( 1989 ), aquatic microbial ecologists examined abun-dance of bacteria infected by viruses (0.7–9 %, Weinbauer and Höfl e 1998 ). Based on the measurements of primary production, bacterial production, protistan bacte-rivory, bacterial lysis by viruses, and production of viruses, Bratbak et al. ( 1992 ) have concluded importance of DOM release by viral lysis of bacteria in carbon cycle in a microbial loop, and they proposed “viral loop” which included the DOM release in a basic microbial loop (Fig. 3.1 ). Wilhelm and Suttle ( 1999 ) pointed out that viruses were also infectious to other planktonic organisms such as phytoplank-ton, suggesting “viral shunt” which included DOM releases from viral lysis of bac-teria, phytoplankton and their grazers.
Since virus infection is basically species specifi c, viral lysis of bacteria may occur at population level. So, viruses may preferentially reduce the bacterial popu-lation with active growth (“kill the winner” model, Thingstad 2000 ). The model shows coexistence of bacterial populations by host-specifi c viruses that prevent the best bacterial competitors from building up a high biomass (Thingstad 2000 ). Since the late 1990s, some researchers have made comparison in loss of bacterial abun-dance between protistan grazing and viral lysis, and the relative importance between the two may be dependent on oxygen environment in a water column (Weinbauer and Höfl e 1998 ; Pradeep Ram et al. 2010 ). Some researchers have reported differ-ence in the relative importance among bacterial phylogenies. In Rimov Reservoir,
3 Biodiversity Researches on Microbial Loop in Aquatic Systems
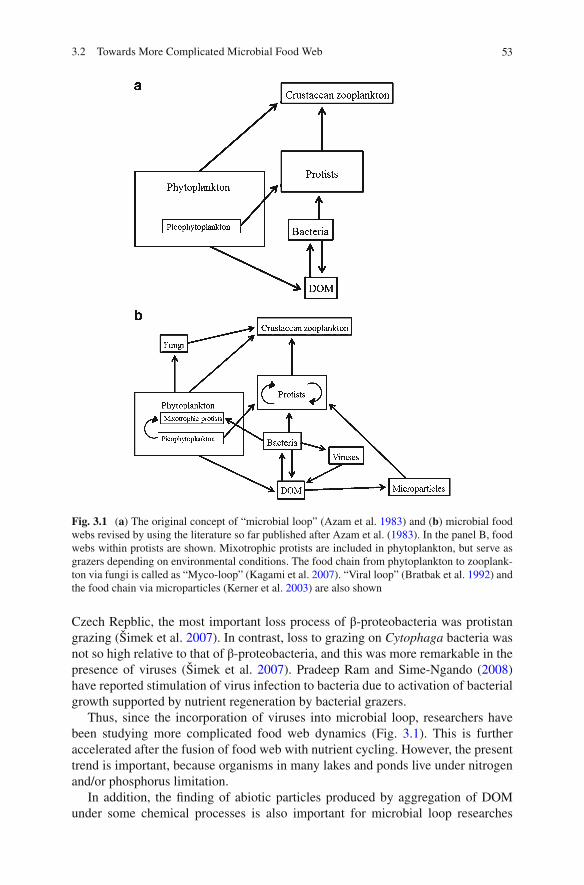
53
Czech Repblic, the most important loss process of β-proteobacteria was protistan grazing (Šimek et al. 2007 ). In contrast, loss to grazing on Cytophaga bacteria was not so high relative to that of β-proteobacteria, and this was more remarkable in the presence of viruses (Šimek et al. 2007 ). Pradeep Ram and Sime-Ngando ( 2008 ) have reported stimulation of virus infection to bacteria due to activation of bacterial growth supported by nutrient regeneration by bacterial grazers.
Thus, since the incorporation of viruses into microbial loop, researchers have been studying more complicated food web dynamics (Fig. 3.1 ). This is further accelerated after the fusion of food web with nutrient cycling. However, the present trend is important, because organisms in many lakes and ponds live under nitrogen and/or phosphorus limitation.
In addition, the fi nding of abiotic particles produced by aggregation of DOM under some chemical processes is also important for microbial loop researches
Fig. 3.1 ( a ) The original concept of “microbial loop” (Azam et al. 1983 ) and ( b ) microbial food webs revised by using the literature so far published after Azam et al. ( 1983 ). In the panel B, food webs within protists are shown. Mixotrophic protists are included in phytoplankton, but serve as grazers depending on environmental conditions. The food chain from phytoplankton to zooplank-ton via fungi is called as “Myco-loop” (Kagami et al. 2007 ). “Viral loop” (Bratbak et al. 1992 ) and the food chain via microparticles (Kerner et al. 2003 ) are also shown
3.2 Towards More Complicated Microbial Food Web

54
(Kerner et al. 2003 ). Some of those abiotic particles have the sizes similar to those of planktonic bacteria, containing DNA, lipids, sugars and/or proteins those of which make the C:N ratio of the particle as 4.1–6.8 (Kerner et al. 2003 ). Those results strongly suggest that the particles are important food items for bacteria graz-ers such as protists. More interestingly, production rate of the particle was similar to that of bacterial production (Kerner et al. 2003 ). In future studies, we may have to include organic matter transfer from the particles to bacterial grazers to fi gure out more reliable food web dynamics and/or matter cycling in planktonic systems (Fig. 3.1 ).
3.3 Detection of Microorganisms Using Molecular Biological Techniques
3.3.1 Biodiversity of Bacteria
Culture-independent molecular techniques such as PCR cloning and sequencing, fl uorescently in situ hybridization (FISH) or Denaturing Gradient Gel Electrophoresis (DGGE) are now commonly used for phylogenetic analyses on microbial communi-ties (DeLong and Karl 2005 ). FISH, which demonstrated the predominant bacterial clade SAR11 (α-proteobacteria) in oceans (Giovannoni and Stingl 2005 ), is one of the most powerful techniques to detect particular microbial populations. In freshwa-ter systems, phylogenetic analyses on bacterial community structure have been con-ducted with several methods including clone library or FISH. There are 34 phylogenetic clusters of closely related sequences that are restricted to freshwater and/or dominated by freshwater sequences (Zwart et al. 2002 ). Some dominant clusters such as the ACK-M1 cluster (acI-A) of Actinobacteria , the LD12 cluster (alfV) of Alphaproteobacteria , the Rhodoferax sp. BAL47 cluster (including the R-BT065 cluster) (beta I), the Polynucleobacter necessarius cluster (beta II) of Betaproteobacteria , and the SOL cluster (including the LD2 cluster) of Bacteroidetes have so far been reported from freshwater systems (Zwart et al. 2003 ; Warnecke et al. 2004 , 2005 ; Schauer and Hahn 2005 ; Šimek et al. 2005 ; Hahn 2006 ; Newton et al. 2011 ; Salcher et al. 2011 ; Hahn et al. 2012 ; Watanabe et al. 2012 ). Members of the R-BT065 subgroup and the newly proposed genus Limnohabitans are included in the Rhodoferax sp. BAL47 cluster (Šimek et al. 2001 ; Hahn et al. 2010a , b ; Kasalický et al. 2010 ).
Ecological roles of some of those dominant bacterial phylogenetic clusters have already been elucidated. Abundance of Polynucleobacter necessarius ssp. asymbi-oticus (PnecC) has positive correlation with concentration of lake humic substances, suggesting importance of the bacteria as decomposers of the organic matter (Jezberová et al. 2010 ). The bacteria in Limnohabitans cluster have high growth rates, serving as important food sources for heterotrophic nanofl agellates (Horňák et al. 2006 ; Šimek et al. 2006 ). For bacterial phylogenetic groups other than
3 Biodiversity Researches on Microbial Loop in Aquatic Systems

55
Limnohabitans cluster bacteria, phosphorus availability may be more important than grazing (Horňák et al. 2006 ).
Recently, pyrosequencing (next-generation sequencing) technique has been remarkably applied to analyze microbial community structure, and those studies have so far demonstrated enormous bacterial and/or archaeal species (Operational Taxonomy Unit, OTU) diversity in marine environments (Sogin et al. 2006 ; Brazelton et al. 2010 ). Those studies have revealed many undescribed taxa and lineages of bacterial and/or archaeal species, and have indicated the presence of large numbers of rare taxa among the relatively few abundant species that dominate these assem-blages in any particular environment, so called “rare biosphere” (Sogin et al. 2006 ; Brazelton et al. 2010 ). Those rare biosphere species have low abundance, suggesting less mortality due to limited encounter with their grazers and/or specifi c viruses (Sogin et al. 2006 ; Caron and Countway 2009 ). In addition, protistan grazers may not consume those rare biosphere species so much because of their small cell size and/or low growth rate (Pedrós-Alió 2006 ). Those rare biosphere species may serve as a potentially inexhaustible genomic reservoir, which could explain how microbial communities respond to subtle environmental changes (Sogin et al. 2006 ).
For bacterial community structure analyses in freshwater systems using pyrose-quencers, not so many studies have so far been conducted. Those studies examined bacterial community structure in biofi lms developed in a drinking water distribution system (Hong et al. 2010 ), bacterial community structure of Amazon River (Ghai et al. 2011 ), difference in community structure of air-borne bacteria inoculated into sterilized Swedish rock pool waters (Langenheder and Székely 2011 ), seasonal changes in bacterial OTU composition in relation to some environmental events such as phytoplankton blooms, wax and wane of zooplankton and nitrifi cation in Lake Erken, Sweden (Eiler et al. 2012 ) and bacterial OTU richness in fourteen Swedish lakes with different lake area, catchment area and trophic status (Logue et al. 2012 ). Eiler et al. ( 2012 ) successfully demonstrated the responses of bacterial community to those biological changes, identifying ecological characteristics of some bacterial phylogenetic groups. Logue et al. ( 2012 ) tried to elucidate the mechanisms that underlie and maintain bacterial biodiversity and patterns in its spatial distribution in fourteen nutrient-poor Swedish lakes. The authors found a positive correlation between bacterial OTU richness and nutrient availability, but the relationships between bacterial OTU richness and lake/catchment area were negative. Those new studies are ecologically interesting, and more collection of bacterial community structure in freshwater systems using next-generation sequencing will be needed.
Besides exploring genetic diversity of bacteria, we need to know ecophysiology of each bacterial species. To do that, isolation and incubation of particular bacterial species are necessary. It is well known that almost all bacterial species are non- culturable, and some isolation and incubation techniques have been developed for higher-throughput of bacterial cultures (Connon and Giovannoni 2002 ). This is also the case for freshwater bacteria. Most of bacteria in 34 freshwater phylogenetic clusters are non-culturable (Zwart et al. 2002 ), but some studies have developed successful culture methods for some dominant freshwater bacterial clusters (Hahn 2003 ; Watanabe et al. 2009 ).
3.3 Detection of Microorganisms Using Molecular Biological Techniques

56
3.3.2 Biodiversity of Protists
Molecular biological techniques have also been largely introduced into protistan biodiversity researches (Massana et al. 2006 ). Moon-van der Staay et al. ( 2001 ) demonstrated high diversity of picoeukaryotes in the sequences of the 18S ribo-somal RNA gene amplifi ed from a picoplanktonic assemblage collected in the equa-torial Pacifi c Ocean. The sequences of those picoeukaryotes were previously unknown which might be assigned to some major marine phyla such as prasino-phytes, haptophytes, dinofl agellates, stramenopiles, choanofl agellates and acanthar-ians. Stramenopiles which include many of so-called fl agellates are one of the most commonly detected marine phyla (Massana et al. 2006 ). Most of fl agellates do not have distinct morphological characteristics which should be used for identifi cation for genus or species, and cells of some fl agellate species are easily deformed by addition of fi xatives. So, some DNA probes to detect certain fl agellate populations by FISH method have been developed (Lim et al. 1999 ). Massana et al. ( 2009 ) used two oligonucleotide probes prepared with 18S rRNA genes derived from chryso-phyceae fl agellates in Blanes Bay, Spain, and respectively detected two populations with average 19 % and 3 % within the total fl agellate community. In addition, some of those fl agellates ingested fl uorescently labeled bacteria (Massana et al. 2009 ).
Countway et al. ( 2005 , 2007 ) investigated protistan diversity by T-RFLP method in the waters collected from the North Atlantic and North Pacifi c oceans, and found that dominant OTUs of the protists from both oceans contributed only to Atlantic or Pacifi c OTUs, and the OTUs detected in both the Atlantic and Pacifi c Ocean sam-ples accounted only for 22 % of the total. Pyrosequencing technique has been also introduced to examine protistan diversity, and Logares et al. ( 2012 ) have demon-strated high diversity of marine stramenopile fl agellates in European coastal waters in relation to their geographical distribution and habitat condition.
The term “rare biosphere” is also valid for protistan diversity: the presence of large numbers of rare taxa among the relatively few abundant species that dominate these assemblages in any particular environment, so called “rare biosphere” (Caron and Countway 2009 ). Countway et al. ( 2005 ) incubated a seawater sample collected from the Western North Atlantic and found changes in overall eukaryotic diversity predominated by protists and species composition expressed as phylotype diversity. The eukaryotic diversity estimated with species richness showed minor changes. However, changes in the dominant eukaryotic phylotypes were substantial, and this was also the case for “rare biosphere” species. Those results again suggest that pro-tistan species in “rare biosphere” serve as a potentially inexhaustible genomic res-ervoir, which could explain how protistan communities respond to subtle environmental changes (Caron and Countway 2009 ).
In freshwater systems, we already have plenty of information about ciliate diver-sity (Beaver and Crisman 1982 , Finlay et al. 1998 and literature therein, Foissner 1999 and literature therein). By contrast, the information about fl agellate diversity is still very limited (Auer and Arndt 2001 ). However, if we consider freshwater protistan species in “rare biosphere,” we do not have any information so far. To
3 Biodiversity Researches on Microbial Loop in Aquatic Systems

57
detect “rare biosphere” microorganisms, we need to use some molecular biological techniques. Studies with some molecular biological approaches are necessary to have the information about freshwater protistan species in “rare biosphere.”
One of fl agellate genera commonly detected in freshwaters is Spumella . Many of previous studies have determined dominant fl agellate genera using cell morphology, and many small colorless chrysophytes have been identifi ed as the genus Spumella or Spumella -like (Sanders et al. 1989 ). Pfandl et al. ( 2009 ) investigated the molecu-lar microdiversity for 13 Spumella -like fl agellate strains isolated from two Austrian freshwaters, followed by examination of their affi liation to different ecotypes with distinct small subunit ribosomal ribonucleic acid (SSU rRNA) genotypes. The eco-physiological tolerances of the fl agellates to temperature or salinity were also exam-ined. The authors have found that those strains consisted of different ecotypes and genotypes, and molecular distance was correlated to temperature and salinity toler-ances. Those results contradict the assumption that variations in SSU rRNA are due to accumulation of neutral mutations. So, the authors have concluded that current taxonomy of species on the Spumella -like fl agellate strains is inappropriate at least with respect to ecological or ecophysiological studies.
3.4 Recent Development in Studies on Food Chain Between Bacteria and Protists
As we discussed earlier, molecular biological techniques have enabled us to have detection of microbial genus/species level and analysis of microbial diversity. So, the researchers of microbial loop have been paying attention to difference in vulner-ability for grazing among bacterial phylogenetic groups. In marine studies so far conducted, we have not yet found any particular bacterial phylogenetic groups vul-nerable to grazing (Yokokawa and Nagata 2005 ; Massana et al. 2009 ). By contrast, some bacterial phylogenetic groups in some freshwaters have been reported as vul-nerable to grazing (Šimek et al. 2007 ).
The commonest method to determine grazing rate on bacteria by protists may be the one which uses surrogate food particles such as fl uorescently labeled bacteria (Sherr et al. 1987 ; Nakano et al. 1998a , b ) or beads (Ichinotuska et al. 2006 ; Okamura et al. 2012 ). There are two great advantages of this method. The fi rst is that incubation times are short. The second is that we can observe protists which ingest surrogate particles under an appropriate microscope, and this enables us to identify which protists (species or genus) are bacterivores, and how much percent-age of protists conduct bacterivory. Fluorescently labeled bacteria (FLB) are most frequently used as surrogate particles (Sherr et al. 1987 ; Nakano et al. 1998a , b ). However, this method has some challenges (Landry 1994 ). The most serious point is that protists select among a variety of particle types. Some protists ingest surro-gate particles added, and the others do not show any response even if they are bac-terivores. No general pattern of preference has been found for either heat-killed
3.4 Recent Development in Studies on Food Chain Between Bacteria and Protists

58
bacteria or artifi cial latex beads (Sanders et al. 1989 ; Montagnes and Lessard 1999 ). Some studies have demonstrated protistan prey preference for living over non- living microorganisms (Landry et al. 1991 ; Montagnes and Lessard 1999 ; Massana et al. 2009 ). A stramenopile fl agellate Spumella egested FLB ingested into its food vacu-ole after a vacuole passage time of about three minutes, while the vacuole passage time of unstained living bacteria exceeded twenty minutes. Massana et al. ( 2009 ) prepared FLB using Brevundimonas diminuta and two FISH probes which respec-tively detected two bacterial strains Nereida sp. and Dokdonia sp. FLB of B. diminuta was heat-killed, while other two bacterial strains were also prepared as alive. The authors used natural fl agellate grazers in a water sample collected in Blanes Bay, Spain, and two stramenopile fl agellate groups, MAST-4 and MAST-1C, were detected using two FISH probes. Using those stuffs, the authors made com-parisons between grazing rates determined with the FLB and with the living bacte-rial strains. For MAST-4 fl agellates, their grazing rates determined with the living bacterial strains were higher than those with the FLB, similar to the results in previ-ous studies (Boenigk et al. 2001 ). By contrast, for MAST-1C fl agellates, the grazing rates determined with the FLB were higher than those with the alive bacteria. In addition, MAST-1C fl agellates did not ingest Dokdonia sp. The authors concluded that small cell size of Dokdonia sp. made the bacteria inedible for MAST-1C fl agel-lates. Researchers of microbial ecology in freshwaters have so far examined bacte-rial response to grazing. Pernthaler ( 2005 ) summarized phenotypic properties of bacteria which may provide protection from protistan grazing: fi lamentation of cell morphology, changes in cell surface potential, cell size reduction, changes in motil-ity patterns, toxin production, cell-to-cell communication, and production of exo-polymer wrapping cells inside. Among those, fi lamentous form of bacteria is mostly known as protection by protistan grazing, and fi lamentous bacteria were often abun-dant at high protozoan grazing pressure to resist grazing (Pernthaler 2005 ; Young 2006 ). Mysteriously, microbial ecology researchers in marine environments have not paid great attention to bacterial response to grazing so far (Sherr and Sherr 2002 ). The information so far collected by freshwater researchers will be useful also for marine researchers, though I do not know the reason for much less studies on bacterial response to grazing in marine environments.
After 1990s, bacterivory by mixotrophic protists has also been paid great atten-tion (Fig. 3.1b ). Studies on mixotrophs’bacterivory have been conducted both in freshwater and marine systems, and relative importance of bacterivory between mixotroph and heterotrophs has been examined. Zubkov and Tarran ( 2008 ) made comparisons of grazing rate on bacteria by plastidic and aplastidic protists in the euphotic layer of the North Atlantic Ocean. The former was considered as mixotro-phic and numerically dominant. The authors especially referred on plastidic protists with small cell size (<2 μm). The latter was heterotrophic and less numerically abundant relative to the former one. Specifi c ingestion rates (bacterial number ingested one cell of protist per unit time) of plastidic protists were lower than those of aplastidic protists. As a result, community ingestion rates (bacterial number ingested by protists in one ml of water per unit time) of small plastidic protists were at levels similar to those of aplastidic protists.
3 Biodiversity Researches on Microbial Loop in Aquatic Systems

59
In freshwater systems, there are good researches on bacterivory by mixotrophic protists (Kimura and Ishida 1985 ; Bird and Kalff 1986 ; Urabe et al . 2000 ; Jones 2000 ).Tittel et al. ( 2003 ) studied on ecology of mixotrophic fl agellate, genus Ochromonas , together with formation mechanism of deep subsurface chlorophyll a maximum in an acidifi ed lake whose food web structure was simple with poor fl ora and fauna. Ochromonas could kept its high abundance and grazing activity by organic matter production of its photosynthesis even after consuming bacteria and small algae to low levels, and the fl agellate had further grazing activity on those food items. Abundance of metazoan predators decreased due to the decrease in abundance of small algae which was food resource also for the predators. So, it is likely that Ochromonas takes advantage for bacterivory over heterotrophic protists under less predation pressure by metazoans. In addition, the formation of deep sub-surface chlorophyll a maximum was elucidated as accumulation of phytoplankton which escaped from Ochromonas grazing conducted in epilimnion.
Studies on relative importance of bacterivory between mixotroph and hetero-trophs after 2000 thus have revealed high grazing pressure on bacteria by mixotro-phic protists. Further, high grazing activity of those mixotrophs occasionally have trophic cascade to phytoplankton and zooplankton. So, the ecological importance of those mixotrophs may be more signifi cant that we evaluated before (Fig. 3.1b ).
3.5 Predation on Protists
In freshwaters, studies about predation on fl agellates by ciliates, rotifers, cladocer-ans and copepods (Burns and Schallenberg 2001 ; Nakano et al. 2001 ; Yoshida et al. 2001a , b ) (Fig. 3.1b ) and about predation on ciliates by rotifers, cladocerans and copepods (Hansen 2000 ; Adrian et al. 2001 ; Burns and Schallenberg 2001 ; Nakano et al. 2001 ) are available. However, we still have only limited information about predation on protists (Sherr and Sherr 2002 ) even at the present, though this may be important transfers of organic matter within a food web (Fig. 3.1 ). This is also the case for marine waters.
The reason for less availability of studies about predation on protists may be due to fragility of protistan cells and/or to researchers’ interest in bacteria which are relatively easy materials for applying molecular biological techniques.
3.6 Biodiversity Researches on Microbial Loop in Lake Biwa
There is a large accumulation of scientifi c information about Lake Biwa (Kawanabe and Maehata 2012 ). In the pelagic area of the lake, planktonic food web is important matter cycling. We already have excellent publications on long term changes in abundance and species composition of phytoplankton ( Hsieh et al. 2010 ; Tsujimura and Ichise 2012 ) and zooplankton (Miura and Cai 1990 ; Tsugeki et al. 2003 ;
3.6 Biodiversity Researches on Microbial Loop in Lake Biwa

60
Kawabata 2012 ). Plenty of ecological researches on phytoplankton (Nakanishi et al. 1992a ; Tezuka and Nakano 1993 ; Miyajima et al. 1994 , 1995 ; Nakano et al. 1996 ) and zooplankton (Kawabata 1987a , b , 1989 , 1991 , 1993 , 1995 ) are also available. Food web dynamics in the herbivorous food chain is also well documented (Nakanishi et al. 1992b ; Kawabata and Nakanishi 1996 ).
In the present review, I would like to introduce research development on micro-bial loop in Lake Biwa.
A driving power of microbial loop is DOM supply from phytoplankton primary production, and there are some studies on bacterial utilization of DOM released by phytoplankton in Lake Biwa (Tanaka et al. 1974 , 1975 ; Ohara et al. 1993 ). Tanaka et al. ( 1974 , 1975 ) found that phytoplankton in Lake Biwa released glycolate as a major component of released photosynthetic products, and that glycolate-utilizing bacteria dominated when the glycolate excretion rate of the phytoplankton was high. Ohara et al. ( 1993 ) reported that dominant phytoplankton species partly deter-mined the dominant bacterial populations in the lake, through release of DOM. From the point of view of biodiversity researches on microbial loop, those three studies may be pioneer studies in Lake Biwa.
Food chain interactions, together with matter cycling within a food chain, have been intensively studied on microbial loop in freshwater and marine systems. A pio-neer and representative work on microbial loop in Lake Biwa is Nagata ( 1988 ), and his work was followed by some functional studies such as nutrient regeneration through microbial loop (Nakano 1994 ) and top-down/bottom-up control on bacterial abundance (Gurung and Urabe 1999 ; Gurung et al. 2000 ). Those three studies paid special attention to phosphorus cycling through microbial loop, because Lake Biwa is a severely P limited lake. For biodiversity in microbial loop in Lake Biwa, there are some studies by Nagata and his colleagues. Nishimura et al. ( 2005 ) examined three types of ecology among bacteria with very high nucleic acid contents, bacteria with high nucleic acid contents, and bacteria with low nucleic acid contents, suggesting that P limitation exerts relatively severe constraints on the growth of bacterial groups with higher nucleic acid contents, which allows bacteria with low nucleic acid con-tents to be competitive and become an important component of the microbial loop. Using FISH, Nishimura and Nagata ( 2007 ) also found dominance by α-proteobacteria in the epilimnion of Lake Biwa. Since many previous studies have reported that a dominant phylogenetic group in lakes is β-proteobacteria (Glöckner et al. 1999 ), Nishimura and Nagata ( 2007 ) provide unique information about bacterial diversity in Lake Biwa. Okazaki et al. ( 2013 ) also used FISH and found that a predominant bacte-rial phylogenetic group in the oxygenated hypolimnion of Lake Biwa affi liated with the CL500-11 cluster (phylum Chlorofl exi ) which may be a potentially important component of the pelagic biogeochemical cycling in the lake.
We still have only limited information about ecological roles of viruses in micro-bial loop in Lake Biwa (Honjo et al. 2007 ; Pradeep Ram et al. 2010 ). Pradeep Ram et al. ( 2010 ) reported high bacterial loss to viral infection in the epilimnion and moderate in the hypolimnion. This result to some extent contradicts to those of other studies (Weinbauer and Höfl e 1998 ; Bettarel et al. 2004 ; Colombet et al. 2006 ), and we have to collect more information about loss of bacteria due to virus infection in Lake Biwa.
3 Biodiversity Researches on Microbial Loop in Aquatic Systems

61
Our information about ecological roles of protists in microbial loop in Lake Biwa is still limited, similar to the case of viruses. The only literature which reports ciliate diversity in the lake is Yoshida et al. ( 2001b ), and no information about fl ag-ellate diversity of the lake is available. The information about grazing on bacteria by heterotrophic/autotrophic fl agellates in Lake Biwa is available (Nakano 1994 ; Nakano et al. 1998b ; Urabe et al. 2000 ), but this is not the case for ciliates. In addi-tion, Yoshida et al. ( 2001a ) is the only publication which reports predation on pro-tists in Lake Biwa.
Lake Biwa may be the lake where microbial loop studies have been most inten-sively conducted in Japan. However, even so, there are still many ecological and/or biodiversity studies which should be conducted in future. Some molecular biologi-cal techniques such as next-generation sequencing may provide us new and massive information about microbial diversity in Lake Biwa. It is essential for us microbial ecologists to use those techniques, because many microorganisms such as bacteria or small protists have limited morphological characteristics for microscopic identi-fi cation. To have more profound understanding on microbial ecology in the lake, we need to relate biodiversity information derived from those molecular biological techniques to ecological functions or processes.
3.7 Concluding Remarks
Some authors have so far revised the basic structure of “microbial loop” (Bratbak et al. 1992 ; Caron and Finlay 1994 ; Kerner et al. 2003 ). Figure 3.1b shows a revised version of microbial loop after inclusion of all the possible components, though I have skipped to fi gure out numerous food linkages within protists. Protists have several feeding modes such as fi lter, raptorial, diffusion and cytopharyngeal basket feedings, and protists probably have web-like food chains. In addition, in freshwater systems, there is another “loop” so-called “myco-loop” (Kagami et al. 2007 ). In myco-loop, large inedible phytoplankton species are infected by fungi, nutrients within these cells are consumed by parasitic fungi, and some of which in turn are grazed by zooplankton (Kagami et al. 2007 ).
There are four advantages when we conduct studies on microbial ecology in freshwater systems. The fi rst, if we consider lakes, a lake environment is a closed system. So, we have relatively clear border between a lake and another system. Also, a lake system is more susceptible to changes in external environments than marine system.
The second, we can conduct in situ experiments in many freshwater systems. In those experiments, we have natural condition on experimental systems, taking appropriate care of them. This is important for ecological researches. Cole et al. ( 2006 ) followed organic matter transfer within a lake food web by daily addition of bicarbonate labeled with stable isotope carbon. The organic matter supply to the lake was dominated by terrestrial origin. However, only 2 % of the organic matter supply reached to zooplankton via microbial loop. Elaborated and precise studies such as Cole et al. ( 2006 ) may be feasible only in lake systems.
3.7 Concluding Remarks

62
The third, lake systems are usually much smaller than marine systems. So, costs needed for scientifi c researches in lakes should be smaller than those in marine systems. In addition, many freshwater systems are easily accessible in terms of geography and/or economy, and this makes frequent visit possible.
The forth, freshwater systems consist of lentic and lotic ones. In rivers or streams, there are biofi lms on solid surfaces. Biofi lms provide unique ecosystems with high microbial diversity and concentrated microbial biomass, consisted of microalgae, fungi, bacteria and protists (Fukuda et al. 2004 ). Only limited studies have so far been conducted for biofi lms in lotic environments, and further studies are needed to elucidate the unique systems.
Freshwater systems are susceptible to human activities, due to high accessibility relative to other ecosystems. So, studies in freshwater systems are important to elu-cidate complex interactions within ecosystems and to determine what can be done for the better management of ecosystems for the sustainable existence of human beings and other living organisms. We need to have up-to-date, modern, sophisti-cated and well-organized research system for future human well-being.
References
Adrian R, Wickham SA, Butler NM (2001) Trophic interactions between zooplankton and the microbial community in contrasting food webs: the epilimnion and deep chlorophyll maximum of a mesotrophic lake. Aquat Microb Ecol 24:83–97
Auer B, Arndt H (2001) Taxonomic composition and biomass of heterotrophic fl agellates in rela-tion to lake trophy and season. Freshw Biol 46:959–972
Azam F, Fenchel T, Field JG, Gray JS, Meyer-Reil LA, Thingstad F (1983) The role of water- column microbes in the sea. Mar Ecol Prog Ser 10:257–263
Beaver JR, Crisman TL (1982) The trophic response of ciliated protozoans in freshwater lakes. Limnol Oceanogr 27:246–253
Bergh O, Borsheim YK, Bratbak G, Heldal M (1989) High abundance of viruses found in aquatic environments. Nature 340:467–468
Bettarel Y, Sime-Ngando T, Amblard C, Dolan J (2004) Viral activity in two contrasting lake eco-systems. Appl Environ Microbiol 70:2941–2951
Bird DF, Kalff J (1986) Bacterial grazing by planktonic lake algae. Science 231:493–495 Bloem J, Bar-Gilissen M-JB (1989) Bacterial activity and protozoan grazing potential in a strati-
fi ed lake. Limnol Oceanogr 34:297–309 Boenigk J, Arndt H, Cleven EJ (2001) The problematic nature of fl uorescently labeled bacteria
(FLB) in Spumella feeding experiments—an explanation by using video microscopy. Arch Hydrobiol 152:329–338
Bratbak G, Heldal M, Thingstad TF, Riemann B, Haslund OH (1992) Incorporation of viruses into the budget of microbial C-transfer. A fi rst approach. Mar Ecol Prog Ser 83:273–280
Brazelton WJ, Ludwig KA, Sogin ML, Andreishcheva EN, Kelley DS, Shen CC, Edwards RL, Baross JA (2010) Archaea and bacteria with surprising microdiversity show shifts in domi-nance over 1,000-year time scales in hydrothermal chimneys. Proc Natl Acad Sci 107:1612–1617
Burns CW, Schallenberg M (2001) Calanoid copepods versus cladocerans: consumer effects on protozoa in lakes of different trophic status. Limnol Oceanogr 46:1558–1565
Caron DA, Countway PD (2009) Hypotheses on the role of the protistan rare biosphere in a chang-ing world. Aquat Microb Ecol 57:227–238
3 Biodiversity Researches on Microbial Loop in Aquatic Systems

63
Caron DA, Finlay BJ (1994) Protozoan links in food webs. In: Hausmann K, Hulsmann N (eds) Progress in protozoology, Gustav Fisher Verlag, Stuttgart, pp 125–130
Cole JJ, Carpenter SR, Pace ML, Van de Bogert MC, Kitchell JL, Hodgson JR (2006) Differential support of lake food webs by three types of terrestrial organic carbon. Ecol Lett 9:558–568
Colombet J, Sime-Ngando T, Cauchie HM, Fonty G, Hoffmann L, Demeure G (2006) Depth- related gradients of viral activity in Lake Pavin. Appl Environ Microbiol 72:4440–4445
Connon SA, Giovannoni SJ (2002) High-throughput methods for culturing microorganisms in very-low-nutrient media yield diverse new marine isolates. Appl Environ Microbiol 68:3878–3885
Countway PD, Gast RJ, Savai P, Caron DA (2005) Protistan diversity estimates based on 18S rDNA from seawater incubations in the Western North Atlantic. J Eukaryot Microbiol 52:95–106
Countway PD, Gast RJ, Dennett MR, Savai P, Rose JM, Caron DA (2007) Distinct protistan assemblages characterize the euphotic zone and deep sea (2500 m) of the western North Atlantic (Sargasso Sea and Gulf Stream). Environ Microbiol 9:1219–1232
DeLong EF, Karl DM (2005) Genomic perspectives in microbial oceanography. Nature 437:336–342
Eiler A, Heinrich F, Bertilsson S (2012) Coherent dynamics and association networks among lake bacterioplankton taxa. ISME J 6:330–342
Finlay BJ, Esteban GF, Fenchel T (1998) Protozoan diversity: converging estimates of the global number of free-living ciliate species. Protist 149:29–37
Foissner W (1999) Protist diversity: estimates of the near-imponderable. Protist 150:363–368 Fukuda M, Ashida A, Tomaru Y, Nakano S (2004) An improved method for collecting heterotro-
phic microorganisms inhabiting on pebbles in streams. Limnology 5:41–46 Ghai R, Rodriguez-Valera F, McMahon KD, Toyama D, Rinke R, de Oliveira TCS, Garcia JW, de
Miranda P, Henrique-Silva F (2011) Metagenomics of the water column in the pristine upper course of the Amazon River. PLoS One 6:e23785
Giovannoni SJ, Stingl U (2005) Molecular diversity and ecology of microbial plankton. Nature 437:343–348
Glöckner FO, Fuchs BM, Amann R (1999) Bacterioplankton compositions of lakes and oceans: a fi rst comparison based on fl uorescence in situ hybridization. Appl Environ Microbiol 65:3721–3726
Gurung TB, Urabe J (1999) Temporal and vertical difference in factors limiting growth rate of heterotrophic bacteria in Lake Biwa. Microb Ecol 38:136–145
Gurung TB, Nakanishi M, Urabe J (2000) Seasonal and vertical difference in negative and positive effects of grazers on heterotrophic bacteria in Lake Biwa. Limnol Oceanogr 45:1689–1696
Hahn MW (2003) Isolation of strains belonging to the cosmopolitan Polynucleobacter necessaries cluster from freshwater habitats located in three climatic zones. Appl Environ Microbiol 69:5248–5254
Hahn MW (2006) The microbial diversity of inland waters. Curr Opin Biotechnol 17:256–261 Hahn MW, Kasalický V, Jezbera J, Brandt U, Jezberová J, Šimek K (2010a) Limnohabitans curvus
gen. nov., sp. nov., a planktonic bacterium isolated from a freshwater lake. Int J Syst Evol Microbiol 60:1358–1365
Hahn MW, Kasalický V, Jazbera J, Brandt U, Šimek K (2010b) Limnohabitans australis sp. nov., isolated from freshwater pond, and emended description of the genus Limnohabitans. Int J Syst Evol Microbiol 60:2946–2950
Hahn MW, Sheuerl T, Jezberová J, Koll U, Jezbera J, Šimek K, Vannini C, Petron G, Wu QL (2012) The passive yet successful way of planktonic life: genomic and experimental analysis of the ecology of a free-living polynucleobacter population. PLoS One 7:e32772
Hansen AM (2000) Response of ciliates and Cryptomonas to the spring cohort of a cyclopoid in a shallow hypereutrophic lake. J Plankton Res 22:185–203
Hong PY, Hwang C, Ling F, Anersen GL, LeChevallier MW, Liu WT (2010) Analysis of bacgterial biofi lm communities in water meters of a drinking water distribution system via pyrosequenc-ing. Appl Environ Microbiol. doi: 10.1128/AEM.00281-10
References

64
Honjo M, Matsui K, Ishii N, Nakanishi M, Kawabata Z (2007) Viral abundance and its related factors in a stratifi ed lake. Fundam Appl Limnol 168:105–112
Horňák K, Jezbera J, Nedoma J, Gasol JM, Šimek K (2006) Effects fo resource availability and bacterivory on leucine incorporation in different groups of freshwater bacterioplankton, assessed using microautoradiography. Aquat Microb Ecol 45:277–289
Hsieh CH, Ishikawa K, Sakai Y, Ishikawa T, Ichise S, Yamamoto Y, Kuo TC, Park H-D, Yamamura N, Kumagai M (2010) Phytoplankton community reorganization driven by eutrophication and warming in Lake Biwa. Aquat Sci 72:467–483
Ichinotuska D, Ueno H, Nakano S (2006) The relative importance of nanofl agellates and ciliates as consumers of bacteria in a coastal sea area (Japan), where the oligotrichous Strombidium spp. and Strobilidium spp. dominate. Aquat Microb Ecol 42:139–147
Jezberová J, Jezbera J, Brandt U, Lindström ES, Langenheder S, Hahn MW (2010) Ubiquity of Polynucleobacter necessarius ssp. asymbioticus in lentic freshwater habitats of a heterogenous 2000 km 2 area. Environ Microbiol 12:658–669
Jones R (2000) Mixotrophy in planktonic protists: an overview. Freshw Biol 45:219–226 Kagami M, de Bruin A, Ibelings BW, Van Donk E (2007) Parasitic chytrids: their effects on phy-
toplankton communities and food-web dynamics. Hydrobiologia 578:113–129 Kasalický V, Jezbera J, Šimek K, Hahn MW (2010) Limnohabitans planktonicus sp. nov., and
Limnohabitans parvus sp. nov., two novel planktonic Betaproteobacteria isolated from a fresh-water reservoir and emended description of the genus Limnohabitans. Int J Syst Evol Microbiol 60:2710–2714
Kawabata K (1987a) Ecology of large phytoplankters in Lake Biwa: population dynamics and food relations with zooplankters. Bull Plankton Soc Jpn 34:165–172
Kawabata K (1987b) Abundance and distribution of Eodiaptomus japonicus (Copepoda: Calanoida) in Lake Biwa. Bull Plankton Soc Jpn 34:173–183
Kawabata K (1989) Seasonal changes in abundance and vertical distribution of Mesocyclops ther-mocyclopoides , Cyclops vicinus and Daphnia longispina in Lake Biwa. Jpn J Limnol 50:9–13
Kawabata K (1991) Ontogenic changes in copepod behavior: an ambush cyclopoid predator and a calanoid prey. J Plankton Res 13:27–34
Kawabata K (1993) Mortality rate of Eodiaptomus japonicas (Copepoda: Calanoida) in Lake Biwa. Jpn J Limnol 54:131–136
Kawabata K (1995) Ongogenic niches of a planktonic copepod in Lake Biwa studied on a fi ne temporal scale. Ecol Res 10:207–215
Kawabata K (2012) Pelagic copepods. In: Kawanabe H, Nishino M, Maehata M (eds) Lake Biwa: interactions between nature and people. Springer, Tokyo, pp 93–96
Kawabata K, Nakanishi M (1996) Food web structure and biodiversity in lake ecosystems. In: Abe T, Levin S, Higashi M (eds) Biodiversity: an ecological perspective. Springer, New York, pp 203–213
Kawanabe NM, Maehata M (2012) Lake Biwa: interactions between nature and people. Springer, Tokyo
Kerner M, Hohenberg H, Ertl S, Reckermann M, Spitzy A (2003) Self-organization of dissolved organic matter to micelle-like microparticles in river water. Nature 422:150–154
Kimura B, Ishida Y (1985) Photophagotrophy in Uroglena Americana, Chrysophyceae. Jpn J Limnol 46:315–318
Landry MR (1994) Methods and controls for measuring the grazing impact of planktonic protists. Mar Microb Food Webs 8:37–57
Landry MR, Lehner-Fournier JM, Sundstrom JA, Fagerness VL, Selph KE (1991) Discrimination between living and heat-killed prey by a marine zoofl agellate, Paraphysomonas vestita (Stokes). J Exp Mar Biol Ecol 146:139–151
Langenheder S, Székely AJ (2011) Species sorting and neutral processes are both important during the initial assembly of bacterial communities. ISME J 5:1086–1094
Lim EL, Dennet MR, Caron DA (1999) The ecology of Paraphysomonas imperforata based on studies employing oligonucleotide probe identifi cation in coastal water samples and enrich-ment cultures. Limnol Oceanogr 44:37–51
3 Biodiversity Researches on Microbial Loop in Aquatic Systems

65
Logares R, Audic S, Santini S, Pernice MC, de Vargas C, Massana R (2012) Diversity patterns and activity of uncultured marine heterotrophic fl agellates unveiled with pyrosequencing. ISME J 6:1823–1833
Logue JB, Langenheder S, Andersson AF, Bertilsson S, Drakare S, Lanzén A, Lindström ES (2012) Freshwater bacterioplankton richness in oligotrophic lakes depends on nutrient avail-ability rather than on species-area relationships. ISME J 6:1127–1136
Massana R, Gillou L, Diez B, Pedrós-Alió C (2002) Unveiling the organisms behind novel eukary-otic ribosomal DNA sequences from the ocean. Appl Environ Microbiol 68:4554–4558
Massana R, Castresana J, Balagué V, Guillou L, Romari K, Groisillier A (2004) Phylogenetic and ecological analysis of novel marine stramenopiles. Appl Environ Microbiol 70:3528–3534
Massana R, Terrado R, Forn I, Lovejoy C, Pedrós-Alió C (2006) Distribution and abundance of uncultured heterotrophic fl agellates in the world oceans. Environ Microbiol 8:1515–1522
Massana R, Unrein F, Rodriguez-Martinez R, Forn I, Lefort T, Pinhassi J, Not F (2009) Grazing rates and functional diversity of uncultured heterotrophic fl agellates. ISME J 3:588–596
Miura T, Cai QH (1990) Annual and seasonal occurrences of the zooplankters observed in the North Basin of Lake Biwa from 1965 to 1979. Contribution from Otsu Hydrobiological Station, Kyoto University 337:1–35
Miyajima T, Nakanishi M, Nakano S, Tezuka Y (1994) An autumnal bloom of a diatom Melosira granulata in a shallow eutrophic lake: physical and chemical constraints on its population dynamics. Arch Hydrobiol 130:143–162
Miyajima T, Nakano S, Nakanishi M (1995) Planktonic diatoms in pelagic silicate cycle in Lake Biwa. Jpn J Limnol 56:211–220
Montagnes DJS, Lessard EJ (1999) Population dynamics of the marine planktonic ciliate Strombidinopsis multiauris : its potential to control phytoplankton blooms. Aquat Microb Ecol 20:167–181
Moon-van der Staay SY, De Wachter R, Vaulot D (2001) Oceanic 18S rDNA sequences from picoplankton reveal unsuspected eukaryotic diversity. Nature 409:607–610
Nagata T (1988) The microfl agellate-picoplankton food linkage in the water column of Lake Biwa. Limnol Oceanogr 33:504–517
Nakanishi M, Miyajima T, Nakano S, Tezuka Y (1992a) Studies on the occurrence of Anabaena and Microcystis blooms in Akanoi Bay of the south basin of Lake Biwa, with special attention to nutrient levels. Ann Rept Interdiscipl Res Inst Environ Sci 11:67–75
Nakanishi M, Tezuka Y, Narita T, Mitamura O, Kawabata K, Nakano S (1992b) Phytoplankton primary production and its fate in a pelagic area of Lake Biwa. Arch Hydrobiol Beih Ergebn Limnol 35:47–67
Nakano S (1994) Estimation of phosphorus release rate by bacterivorous fl agellates in Lake Biwa. Jpn J Limnol 55:201–211
Nakano S, Seike Y, Sekino T, Okumura M, Kawabata K, Fujinaga K, Nakanishi M, Mitamura O, Kumagai M, Hashitani H (1996) A rapid growth of Aulacoseira granulata (Bacillariophyceae) during the typhoon season in the south basin of Lake Biwa. In: Robarts RD, Kumagai M (eds) BITEX special issue, Jpn J Limnol 57:493–500
Nakano S, Ishii N, Manage PM, Kawabata Z (1998a) Trophic roles of heterotrophic nanofl agel-lates and ciliates among planktonic organisms in a hypereutrophic pond. Aquat Microb Ecol 16:153–161
Nakano S, Koitabashi T, Ueda T (1998b) Seasonal changes in abundance of heterotrophic nanofl a-gellates and their consumption of bacteria in Lake Biwa with special reference to trophic inter-actions with Daphnia galeata . Arch Hydrobiol 142:21–34
Nakano S, Manage PM, Nishibe Y, Kawabata Z (2001) Trophic linkage among heterotrophic nanofl agellates, ciliates and metazoan zooplankton in a hypereutrophic pond. Aquat Microb Ecol 25:259–270
Newton RJ, Jones SE, Eiler A, McMahon KD, Bertilsson S (2011) A guide to the natural history of freshwater lake bacteria. Microbiol Mol Biol Rev 75:14–49
Nishimura Y, Nagata T (2007) Alphaproteobacterial dominance in a large mesotrophic lake (Lake Biwa, Japan). Aquat Microb Ecol 48:231–240
References

66
Nishimura Y, Kim C, Nagata T (2005) Vertical and seasonal variations of bacterioplankton sub-groups with different nucleic acid contents: possible regulation by phosphorus. Appl Environ Microbiol 71:5828–5836
Ohara S, Fukami K, Ishida Y (1993) Algal effects on the bacterial assemblage in Lake Biwa. Jpn J Limnol 54:261–268
Okamura T, Mori Y, Nakano S, Kondo R (2012) Abundance and bacterivory of heterotrophic nanofl agellates in the meromictic Lake Suigetsu, Japan. Aquat Microb Ecol 66:149–158
Okazaki Y, Hodoki Y, Nakano S (2013) Seasonal dominance of CL500-11 bacterioplankton (Phylum Chlorofl exi ) in the oxygenated hypolimnion of Lake Biwa, Japan. FEMS Microbiol Ecol 83(1):82–92
Pedrós-Alió C (2006) Marine microbial diversity: can it be determined? Trends Microbiol 14:257–263
Pernthaler J (2005) Predation on prokaryotes in the water column and its ecological implications. Nat Rev Microbiol 3:537–546
Pfandl K, Chatzinotas A, Dyal P, Boenigk J (2009) SSU rRNA gene variation resolves population heterogeneity and ecophysiological differentiation within a morphospecies (Stramenopiles, Chrysophyceae). Limnol Oceanogr 54:171–181
Pradeep Ram AS, Sime-Ngando T (2008) Functional responses of prokaryotes and viruses to grazer effects and nutrient additions in freshwater microcosms. ISME J 2:498–509
Pradeep Ram AS, Nishimura Y, Tomaru Y, Nagasaki K, Nagata T (2010) Seasonal variation in viral-induced mortality of bacterioplankton in the water column of a large mesotrophic lake (Lake Biwa, Japan). Aquat Microb Ecol 58:249–259
Riemann B, Søndergaard M (1986) Carbon dynamics in eutrophic temperate lakes. Elsevier, The Netherlands
Salcher MM, Pernthaler J, Posch T (2011) Seasonal bloom dynamics and ecophysiology of the freshwater sister clade of SAR11 bacteria ‘that rule the waves’ (LD12). ISME J 5:1242–1252
Sanders RW, Porter KG, Bennett SJ, Debiase AE (1989) Seasonal patterns of bacterivory by fl agel-lates, ciliates, rotifers, and cladocerans in a freshwater planktonic community. Limnol Oceanogr 34:673–687
Schauer M, Hahn MW (2005) Diversity and phylogenetic affi liations of morphologically con-spicuous large fi lamentous bacteria occurring in the pelagic zones of broad spectrum of fresh-water habitats. Appl Environ Microbiol 71:1931–1940
Sherr EB, Sherr BF (2002) Signifi cance of predation by protists in aquatic microbial food webs. Antonie van Leeuwenhoek 81:293–308
Sherr BF, Sherr EB, Fallon RD (1987) Use of monodispersed, fl uorescently labeled bacteria to estimate in situ protozoan bacterivory. Appl Environ Microbiol 53:958–965
Šimek K, Bobkova K, Macek M, Nedoma J (1995) Ciliate grazing on picoplankton in a eutrophic reservoir during the summer phytoplankton maximum: a study at the species and community level. Limnol Oceanogr 40:1077–1090
Šimek K, Macek M, Seda J, Vyhnalek V (1990) Possible food chain relationships between bacte-rioplankton, protozoans and cladocerans in a reservoir. Int Rev Gesam Hydrobiol 75:583–596
Šimek K, Pernthaler J, Weinbauer MG, Hornák K, Dolan JR, Nedoma J et al (2001) Changes in bacte-rial community composition and dynamics and viral mortality rates associated with enhanced fl ag-ellate grazing in a mesoeutrophic reservoir. Appl Environ Microbiol 67:2723–2733
Šimek K, Straskrabova V (1992) Bacterioplankton production and protozoan bacterivory in a mesotrophic reservoir. J Plankton Res 14:773–787
Šimek K, Horňák K, Jezbera J, Mašín M, Nedoma J, Gasol JM, Schauer M (2005) Infl uence of top-down and bottom-up manipulations on the R-BT065 subcluster of b-Proteobacteria, an abundant group in bacterioplankton of a freshwater reservoir. Appl Environ Microbiol 71:2381–2390
Šimek K, Hornák K, Jezbera J, Nedoma J, Vrba J, Strakrábová V, Macek M, Dolan JR, Hahn MW (2006) Maximum growth rates and possible life strategies of different bacterioplankton groups in relation to phosphorus availability in a freshwater reservoir. Environ Microbiol 8:1613–1624
Šimek K, Weinbauer MG, Hornak K, Jezbera J, Nedoma J, Dolan JR (2007) Grazer and virus- induced mortality of bacterioplankton accelerates development of Flectobacillus populations in a freshwater community. Environ Microbiol 9:789–800
3 Biodiversity Researches on Microbial Loop in Aquatic Systems

67
Sogin ML, Morrison HG, Huber JA, Welch DM, Huse SM, Neal PR, Arrieta JM, Herndl GJ (2006) Microbial diversity in the deep sea and the underexplored “rare biosphere”. Proc Natl Acad Sci 103:12115–12120
Suttle CA (2007) Marine viruses – major players in the global ecosystem. Nat Rev Microbiol 5:801–812
Tanaka N, Nakanishi M, Kadota H (1974) The excretion of photosynthetic product by natural phytoplankton population in Lake Biwa. Jpn J Limnol 35:91–98
Tanaka N, Nakanishi M, Kadota H (1975) Seasonal variation of glycollate-utilizing bacteria in the water column of Lake Biwa. Bull Jpn Soc Sci Fish 41:1129–1134
Tezuka Y, Nakano S (1993) Induction of Anabaena bloom by nutrient enrichment to the lake water collected from the south basin of Lake Biwa. Jpn J Limnol 54:85–90
Thingstad TF (2000) Elements of a theory for the mechanisms controlling abundance, diversity, and biogeochemical role of lytic viruses in aquatic systems. Limnol Oceanogr 45:1320–1328
Tittel J, Bissinger V, Zippel B, Gaedke U, Bell E, Lorke A, Kamjunke N (2003) Mixotrophs com-bine resource use to outcompete specialists: Implications for aquatic food webs. Proc Natl Acad Sci 100:12776–12781
Tsugeki N, Oda H, Urabe J (2003) Fluctuation of the zooplankton community in Lake Biwa during the 20th century: a paleolimnological analysis. Limnology 4:101–107
Tsujimura S, Ichise S (2012) Characteristics of the phytoplankton fl ora and long term changes in the phytoplankton community of Lake Biwa. In: Kawanabe H, Nishino M, Maehata M (eds) Lake Biwa: Interactions between nature and people. Springer, Tokyo, pp 41–48
Urabe J, Grung TB, Yoshida T, Sekino T, Nakanishi M (2000) Diel changes in phagotrophy by Cryptomonas in Lake Biwa. Limnol Oceanogr 45:1558–1563
Warnecke F, Amman R, Pernthaler J (2004) Actinobacterial 16S rRNA genes from freshwater habitats cluster in four distinct lineages. Environ Microbiol 6:242–253
Warnecke F, Sommaruga R, Sekar R, Hofer JS, Pernthaler J (2005) Abundance, identity, and growth state of Actinobacteria in mountain lakes of different UV transparency. Appl Environ Microbiol 71:5551–5559
Watanabe K, Komatsu N, Ishii Y, Negishi M (2009) Effective isolation of bacterioplankton genus Polynucleobacter from freshwater environments grown on photochemicaly degraded dissolved organic matter. FEMS Microbiol Ecol 67:57–68
Watanabe K, Komatsu N, Kitamura T, Ishii Y, Park H-D, Miyata R, Noda N, Sekiguchi Y, Satou T, Watanabe M, Yamamura S, Imai A, Hayashi S (2012) Ecological niche separation in the Polynucleobacter subclusters linked to quality of dissolved organic matter: a demonstration using a high sensitivity cultivation-based approach. Environ Microbiol. doi: 10.1111/j.1462-2920.2012.02815.x
Weinbauer MG, Höfl e MG (1998) Signifi cance of viral lysis and fl agellate grazing as factors con-trolling bacterioplankton production in a eutrophic lake. Appl Environ Microbiol 64:431–438
Wilhelm SW, Suttle CA (1999) Viruses and nutrient cycles in the sea. Bioscience 49:781–788 Yokokawa T, Nagata T (2005) Growth and grazing mortality rates of phylogenetic groups of bac-
terioplankton in coastal marine environments. Appl Environ Microbiol 71:6799–6807 Yoshida T, Gurung TB, Kagami M, Urabe J (2001a) Contrasting effects of a cladoceran ( Daphnia
galeata )and a calanoid copepod ( Eodiaptomus japonicus ) on algal and microbial plankton in a Japanese lake, Lake Biwa. Oecologia 129:602–610
Yoshida T, Kagami M, Gurung TB, Urabe J (2001b) Seasonal succession of zooplankton in the north basin of Lake Biwa. Aquat Ecol 35:19–29
Young KD (2006) The selective value of bacterial shape. Microbiol Mol Biol Rev 3:660–703 Zubkov MV, Tarran GA (2008) High bacterivory by the smallest phytoplankton in the North
Atlantic Ocean. Nature 455:224–227 Zwart G, Crump BC, Kamst-van Agterveld MP, Hagen F, Han SK (2002) Typical freshwater bac-
teria: an analysis of available 16S rRNA gene sequences from plankton of lakes and rivers. Aquat Microb Ecol 28:141–155
Zwart G, van Hannen EJ, Kamst-van Agterveld MP, Vander Gucht K, Lindström ES, Van Wichelen J, Lauridsen T, Crump BC, Han SK, Declerck S (2003) Rapid screening for freshwater bacte-rial group by using reverse line brot hybridization. Appl Environ Microbiol 69:5875–5883
References

69N. Okuda et al., Biodiversity in Aquatic Systems and Environments: Lake Biwa, SpringerBriefs in Biology, DOI 10.1007/978-4-431-54150-9_4, © The Author(s) 2014
Abstract Ecosystems often show sudden and drastic shifts in their states following relatively small environmental changes, yet the environmental restoration does not necessarily easily recover the original state. The resilience theory has played a piv-otal role in ecosystem management by providing a theoretical basis for such abrupt and irreversible phase transitions (i.e., regime shifts). However, a major concern remains that the existing theory considers ecosystem responses along only a single disturbance axis (e.g., eutrophication), despite the fact that natural ecosystems are subject to multiple anthropogenic disturbances. In this chapter, I introduce ontoge-netic niche shifts (i.e., changes in resource use or predation vulnerability during individual growth) as a possible common mechanism of regime shifts. Based on this framework, I show how additional factors not accounted for the basic resilience theory (e.g., species extinction and invasion, habitat loss and fragmentation, and phenological shifts) may affect whether or where regime shifts occur along environ-mental gradients. I conclude that these results taken together illustrate the impor-tance of interdisciplinary research integrating biodiversity conservation, landscape protection, and climate change adaptation for more effective management of lake ecosystems.
Keywords Alternative stable states • Bistability • Ecosystem disturbance • Ecosystem restoration • Food-web dynamics • Global change • Positive feedback • Warming
4.1 Introduction
We enjoy various benefits from lake ecosystems such as drinking water, fishery resources, and opportunities for leisure, sport, and education (Postel and Carpenter 1997). These ecosystem services are a driving force for the development and main-tenance of human society, as ancient people achieved highly advanced civilization
Chapter 4A Dynamic Resilience Perspective Toward Integrated Ecosystem Management: Biodiversity, Landscape, and Climate

70
around big rivers such as the Nile River and the Tigris–Euphrates basin. We exploit the natural resources of lakes, especially large lakes, in which a large amount of fresh water is available and diverse aquatic species, especially fish, coexist. However, the human activities around these ecosystems inevitably cause various anthropo-genic disturbances. In the last several decades, lake-ecosystem services have been significantly degraded through anthropogenic disturbances such as eutrophication (Smith et al. 2006; Smith and Schindler 2009), biodiversity degradation (Lovel 1997; Chapin et al. 2000; Sala et al. 2000), and landscape modification (Fahrig 2003; Cushman 2006; Collinge 2009). In addition, climate change creates a serious global problem for ecosystem services (Lovejoy and Hannah 2005; Hannah 2010; Kernan et al. 2010). These ecosystem disturbances may reduce water quality, fish-ery catch, and the cultural value of lakes, thereby potentially preventing sustainable development of human society.
This is true and important for Lake Biwa and people living around the lake as well. Lake Biwa has been subject to various ecosystem disturbances during the lat-ter half of the twentieth century such as eutrophication (Hsieh et al. 2010, 2011), biological invasion (Nakai 1999), lakeshore development (Nakanish and Sekino 1996), and climate change (Hsieh et al. 2010, 2011). As a result, the ecosystem services of Lake Biwa (in particular, water supply and fishery) have been seriously degraded, which is essential for our life and culture (Kawanabe et al. 2012). Establishing effective ecosystem management is therefore a pressing concern in ecological research. In this chapter, I will present a novel and widely applicable perspective for ecosystem management based on the resilience (regime shift) the-ory, and consider the possibility that it could help improve the ecosystem manage-ment of lakes including Lake Biwa.
4.2 Regime Shift and Resilience as Key Concepts in Ecosystem Management
Monitoring is a pivotal part of ecosystem management. We monitor ecosystems by assuming that changes in ecosystem states reflect changes in environmental condi-tions. As such, we expect that we can predict future ecosystem states and prevent environmental disturbances from becoming serious. However, the relationships between the ecosystem state and environmental condition are not necessarily linear (see below). As a result, ecosystems often undergo sudden and rapid shifts follow-ing relatively small environmental changes. Such abrupt state transitions are gener-ally called regime shifts (Scheffer et al. 2001; Scheffer and Carpenter 2003; Folke et al. 2004; Schröder et al. 2005). Understanding the dynamic nature of such eco-systems is crucial for establishing more effective ecosystem management.
Ecological regime shift is defined as rapid and drastic reorganization of systems (e.g., ecosystem and community), from one relatively stable state to another, in response to relatively small environmental changes (Scheffer et al. 2001; Schröder
4 A Dynamic Resilience Perspective Toward Integrated Ecosystem Management…
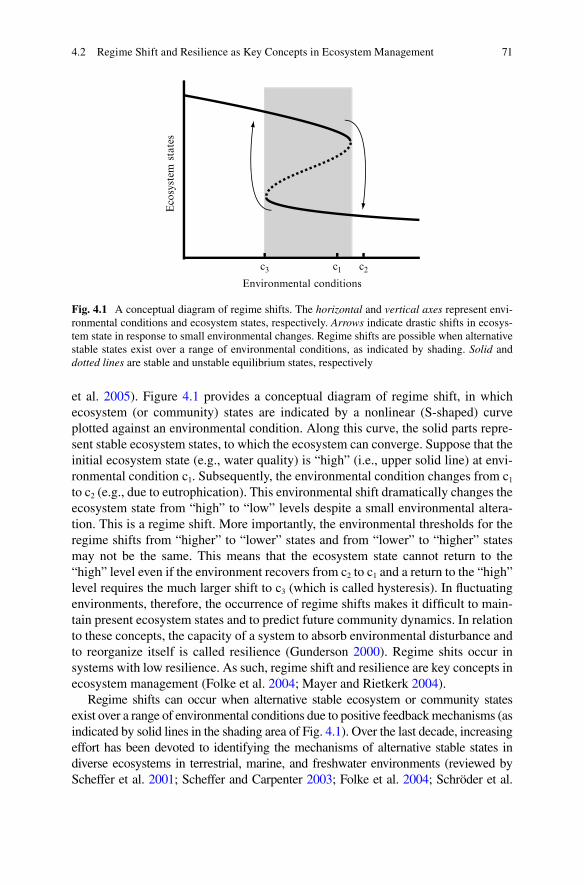
71
et al. 2005). Figure 4.1 provides a conceptual diagram of regime shift, in which ecosystem (or community) states are indicated by a nonlinear (S-shaped) curve plotted against an environmental condition. Along this curve, the solid parts repre-sent stable ecosystem states, to which the ecosystem can converge. Suppose that the initial ecosystem state (e.g., water quality) is “high” (i.e., upper solid line) at envi-ronmental condition c1. Subsequently, the environmental condition changes from c1 to c2 (e.g., due to eutrophication). This environmental shift dramatically changes the ecosystem state from “high” to “low” levels despite a small environmental altera-tion. This is a regime shift. More importantly, the environmental thresholds for the regime shifts from “higher” to “lower” states and from “lower” to “higher” states may not be the same. This means that the ecosystem state cannot return to the “high” level even if the environment recovers from c2 to c1 and a return to the “high” level requires the much larger shift to c3 (which is called hysteresis). In fluctuating environments, therefore, the occurrence of regime shifts makes it difficult to main-tain present ecosystem states and to predict future community dynamics. In relation to these concepts, the capacity of a system to absorb environmental disturbance and to reorganize itself is called resilience (Gunderson 2000). Regime shits occur in systems with low resilience. As such, regime shift and resilience are key concepts in ecosystem management (Folke et al. 2004; Mayer and Rietkerk 2004).
Regime shifts can occur when alternative stable ecosystem or community states exist over a range of environmental conditions due to positive feedback mechanisms (as indicated by solid lines in the shading area of Fig. 4.1). Over the last decade, increasing effort has been devoted to identifying the mechanisms of alternative stable states in diverse ecosystems in terrestrial, marine, and freshwater environments (reviewed by Scheffer et al. 2001; Scheffer and Carpenter 2003; Folke et al. 2004; Schröder et al.
Environmental conditions
Eco
syst
em s
tate
s
c3 c1 c2
Fig. 4.1 A conceptual diagram of regime shifts. The horizontal and vertical axes represent envi-ronmental conditions and ecosystem states, respectively. Arrows indicate drastic shifts in ecosys-tem state in response to small environmental changes. Regime shifts are possible when alternative stable states exist over a range of environmental conditions, as indicated by shading. Solid and dotted lines are stable and unstable equilibrium states, respectively
4.2 Regime Shift and Resilience as Key Concepts in Ecosystem Management

72
2005). In lake ecosystems, for example, researchers have focused on regime shifts between a macrophyte-dominated clear water phase and a phytoplankton- dominated turbid water phase along a gradient nutrient loading (Scheffer 1998; Genkai-Kato and Carpenter 2005). In this case, the positive feedback involves antagonistic interactions between two producers (macrophytes and phytoplankton) as follows. When phyto-plankton are dominant, they suppress submerged macrophytes by reducing the light intensity that is available at the bottom. When macrophytes are dominant, they suppress phytoplankton by inhibiting the resuspension of organic matter from lake sediments into the water. This antagonistic relationship leads to the prediction that lake ecosys-tems will converge to either a macrophyte-dominated or phytoplankton-dominated state, depending on the initial conditions, and the sudden change in ecosystem state from a clear to a turbid water phase can occur following eutrophication. In addition, such regime shifts can result in an alteration of ecosystem functions such as lake pri-mary productivity (Genkai- Kato et al. 2012). Many other types of alternative stable states and regime shifts have also been identified in aquatic and terrestrial ecosystems. Examples of these regimes shifts include shifts between the presence and absence of piscivorous fish (Jansson et al. 2007; Persson et al. 2007), forest and savanna (Peterson 2002), social concern and indifference for ecosystem management (Suzuki and Iwasa 2009), and many others (see reviews by Scheffer et al. 2001; Scheffer and Carpenter 2003; Folke et al. 2004; Schröder et al. 2005).
4.3 Regime Shifts Mediated by Ontogenetic Niche Shifts
Here, I introduce a recently identified, but very important, mechanism of alternative stable states. This mechanism is ontogenetic niche shift, that is, the changes in eco-logical niches, such as resource use and predation vulnerability, during individual growth. Virtually all multi-cellular organisms on the earth exhibit individual growth between birth and death and most species experience ontogenetic niche shifts (Wilbur 1980; Werner and Gilliam 1984). A well-known example of ontogenetic niche shift is the metamorphosis of amphibians and insects whereby they change their morphology drastically. However, ontogenetic niche shifts without clear mor-phological changes are also common in nature (Werner and Gilliam 1984; Nakazawa and Yamamura 2007; also see Sect. 4.4.2).
Importantly, ontogenetic niche shifts divide a species into distinct groups (e.g., stage and size) that allow individuals of the same species to occupy different eco-logical niches within a community (Nakazawa et al. 2011; Miller and Rudolf 2011). As a result, ontogenetic niche shifts play an important role in determining commu-nity structure and dynamics (Nakano and Murakami 2001; Baxter et al. 2005; Knight et al. 2005). In aquatic ecosystems, for example, the ontogenetic niche shifts of fish from planktivory to benthivory connect pelagic and benthic trophic pathways (Persson et al. 1996; Reñones et al. 2002; Xu et al. 2007; Briones et al. 2012), gen-erally called pelagic-benthic coupling (Schindler and Scheuerell 2002). Several theoretical studies have recently suggested that ontogenetic niche shifts may
4 A Dynamic Resilience Perspective Toward Integrated Ecosystem Management…

73
mediate the occurrence of regime shifts by generating alternative stable community states along environmental gradients.
In one such study, Schreiber and Rudolf (2008) developed a simple one con-sumer–two resource model that included consumer ontogenetic diet shift, as follows:
dR dtJ J J J J J J Jr R K R a C R( )/ /= 1 � � (4.1a)
dR dtA A A A A A A Ar R K R a C R( )/ /= 1 � � (4.1b)
dC dtJ A A A A J J J J J Jb a C R b a C R d C/ = � � (4.1c)
dC dtA J J J J A Ab a C R d C/ = � (4.1d)
In the model, Rh (h = J or A) represents the density of a resource used by either juveniles (J) or adults (A), and Ch is the density of the consumer at each stage. The resources exhibit logistic growth and are reduced by either juvenile or adult con-sumers (Eqs. 4.1a and 4.1b). Resource consumption at the adult stage promotes reproduction and supports recruitment into the juvenile stage (Eq. 4.1c). Likewise, resource consumption at the juvenile stage promotes maturation and supports recruitment into the adult stage (Eq. 4.1d). The juveniles are lost by maturation and natural mortality (Eq. 4.1c), whereas the adults are lost only by natural mortality (Eq. 4.1d). The parameters are defined as follows: rh intrinsic growth rate, Kh carry-ing capacity, ah resource consumption rate, bh energy conversion efficiency, and dh death rate. Using this model, Schreiber and Rudolf (2008) illustrated how ontoge-netic shifts can result in alternative stable community states and how the system can undergo regime shifts in response to environmental changes (left panel of Fig. 4.2). Regime shifts in this model result from the positive feedback caused by the apparent competition-like interactions between the juvenile and adult resources. Suppose that the resource that the juveniles use becomes more abundant, promoting matura-tion. In this case, the resource used by the adults is affected negatively through consumption by the increased density of adults. The decrease in the adult resource, in turn, leads to a decrease in reproduction and an increase in the juvenile resource, as the density of juveniles decreases. This results in positive feedback and the sys-tem converges to either juvenile- or adult-dominated states, depending on the initial conditions. In this system, therefore, a sudden and drastic shift in population stage- structure and community structure can occur following changes in environmental conditions such as resource carrying capacity or consumer death rate.
Guill (2009) also explored a one-consumer–two-resource model containing a consumer’s ontogenetic niche diet shift. His model is conceptually similar to that of Schreiber and Rudolf (2008). One major difference of his model is that it is based on biomass dynamics while Schreiber and Rudolf (2008) based their model on the dynamics of population abundance. As such, Guill (2009) assumed that if the resource is not sufficiently available, the body mass of the consumer may decrease or the consumer may cease to reproduce because of the associated metabolic costs.
4.3 Regime Shifts Mediated by Ontogenetic Niche Shifts
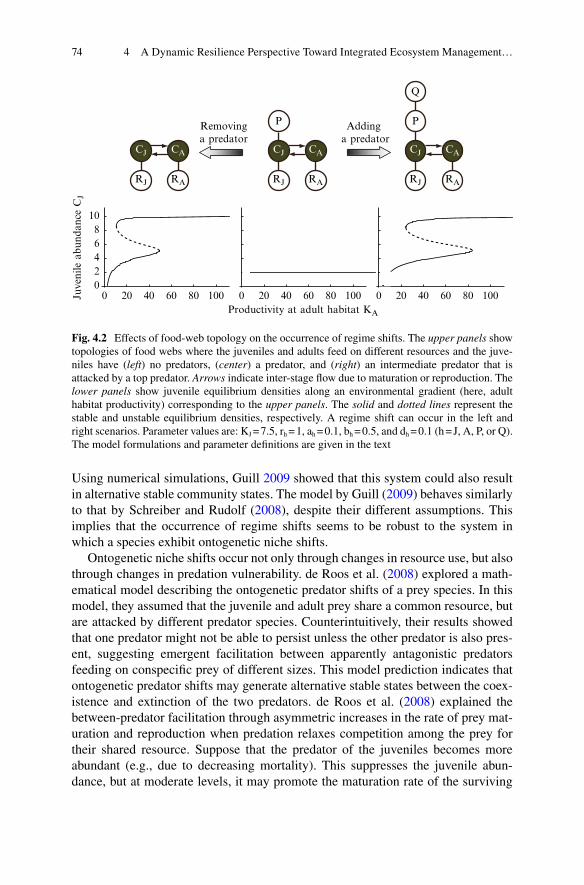
74
Using numerical simulations, Guill 2009 showed that this system could also result in alternative stable community states. The model by Guill (2009) behaves similarly to that by Schreiber and Rudolf (2008), despite their different assumptions. This implies that the occurrence of regime shifts seems to be robust to the system in which a species exhibit ontogenetic niche shifts.
Ontogenetic niche shifts occur not only through changes in resource use, but also through changes in predation vulnerability. de Roos et al. (2008) explored a math-ematical model describing the ontogenetic predator shifts of a prey species. In this model, they assumed that the juvenile and adult prey share a common resource, but are attacked by different predator species. Counterintuitively, their results showed that one predator might not be able to persist unless the other predator is also pres-ent, suggesting emergent facilitation between apparently antagonistic predators feeding on conspecific prey of different sizes. This model prediction indicates that ontogenetic predator shifts may generate alternative stable states between the coex-istence and extinction of the two predators. de Roos et al. (2008) explained the between-predator facilitation through asymmetric increases in the rate of prey mat-uration and reproduction when predation relaxes competition among the prey for their shared resource. Suppose that the predator of the juveniles becomes more abundant (e.g., due to decreasing mortality). This suppresses the juvenile abun-dance, but at moderate levels, it may promote the maturation rate of the surviving
1086420
Productivity at adult habitat KA
Juve
nile
abu
ndan
ce C
J
Q
P
CJ CA
RJ RA
P
CJ CA
RJ RA
CJ CA
RJ RA
Adding a predator
Removinga predator
200 40 60 80 100 0 20 40 60 80 100 0 20 40 60 80 100
Fig. 4.2 Effects of food-web topology on the occurrence of regime shifts. The upper panels show topologies of food webs where the juveniles and adults feed on different resources and the juve-niles have (left) no predators, (center) a predator, and (right) an intermediate predator that is attacked by a top predator. Arrows indicate inter-stage flow due to maturation or reproduction. The lower panels show juvenile equilibrium densities along an environmental gradient (here, adult habitat productivity) corresponding to the upper panels. The solid and dotted lines represent the stable and unstable equilibrium densities, respectively. A regime shift can occur in the left and right scenarios. Parameter values are: KJ = 7.5, rh = 1, ah = 0.1, bh = 0.5, and dh = 0.1 (h = J, A, P, or Q). The model formulations and parameter definitions are given in the text
4 A Dynamic Resilience Perspective Toward Integrated Ecosystem Management…

75
juveniles because of food-dependent individual growth. The increased maturation rate may positively affect the predator of the adults (also see de Roos et al. 2007). Similarly, moderate predation of the adults may promote reproduction among the surviving individuals, thereby positively affecting the predator of the juveniles. de Roos et al. (2008) further showed that the emergent facilitation between stage- specific predators is also possible when a shared prey species has more than two stages.
These theoretical models all assume that environmental conditions (e.g., resource availability and mortality risk) at one life stage strongly affect the performance of other stages via density-dependent effects; for example, higher maturation rates intensify resource competition at the adult stage and reduce the per-capita reproduc-tion rate. This implies that alternative stable states may not exist when maturation or reproduction are density-independent (McCoy et al. 2009) or are governed by other rules (de Roos et al. 2002; Takimoto 2003; Nakazawa and Yamamura 2007). To test the key assumption, empirical studies have recently examined to what extent stage-specific performance is density-dependent (Walter and Kiutchell 2001; Ohlberger et al. 2012; also see reviews by Abrams 2009; Vincenzi et al. 2012). On the other hand, direct tests of model predictions are rather difficult because they require long-term data on community dynamics. Even so, the occurrence of regime shifts mediated by ontogenetic niche shifts is expected to be widespread in nature because ontogenetic niche shifts are ubiquitous among organisms (Wilbur 1980; Werner and Gilliam 1984). Notably, Persson et al. (2007) have recently illustrated, via whole-lake manipulations, that a reduction in the density of large prey individu-als caused an increase in the abundance of small individuals, allowing their predator (piscivores) to recover. They explained that the underlying mechanism would be the prey’s ontogenetic shift in predation vulnerability. The results of Persson et al. (2007) suggest that the presence and absence of the piscivores could be alternative stable states in natural environments.
4.4 Effects of Ecological Contexts on the Occurrence of Regime Shifts Mediated by Ontogenetic Niche Shifts
The occurrence of regime shifts that are mediated by ontogenetic niche shifts seems to be intrinsic to the dynamics of size/stage-structured (i.e. almost all) ecological communities. Although this has significant implications for ecosystem management (see Sect. 4.2), many questions remain. In particular, it is important to determine whether regime shifts always occur in size-structured communities, and if not, under what conditions they occur. Ontogenetic niche shifts may not necessarily generate alternative stable states in the presence of additional factors that diminish or eliminate their effects. I explain below how the occurrence of regime shifts that are mediated by ontogenetic niche shifts are altered in complex ecosystems. I focus on three factors: (i) food-web topology, (ii) spatial configuration, and (iii)
4.4 Effects of Ecological Contexts on the Occurrence of Regime Shifts Mediated…

76
seasonality. The reason that I focused on these factors is not only that real food webs are much more complex than assumed in the simple models described above. At present, both aquatic and terrestrial ecosystems are under various anthropogenic disturbances such as biodiversity degradation due to species extinction and invasion (Lovel 1997; Chapin et al. 2000; Sala et al. 2000), habitat loss and fragmentation due to landscape modification (Fahrig 2003; Cushman 2006; Collinge 2009), and phenological shifts due to climate change (Lovejoy and Hannah 2005; Hannah 2010; Kernan et al. 2010). Understanding the conditions that cause regime shifts in various ecological contexts (i.e., species composition and their relationships in space and time), will lead to a better understanding of how ecosystem resilience varies with anthropogenic disturbance.
4.4.1 Food-Web Topology
In this section, I examine how regime shifts mediated by ontogenetic niche shifts can be altered by changes in food-web topology. First, suppose that juveniles and adults are both primary consumers feeding on different resources and only the juve-niles are attacked by a predator species (Nakazawa 2011a). The dynamics of this system are described as follows:
dR dtJ J J J J J J Jr R K R a C R( )/ /= 1 � � (4.2a)
dR dtA A A A A A A Ar R K R a C R( )/ /= 1 � �
(4.2b)
dC dtJ A A A A J J J J J J P Jb a C R b a C R d C a C P/ = � � � (4.2c)
dC dtA J J J J A Ab a C R d C/ = � (4.2d)
dP dt/ = b a C P d PP P J P� (4.2e)
In the model, the predator P increases by feeding on the juveniles and decreases due to natural mortality (Eq. 4.2e). The parameters aP, bP, and dP are the attack rate, energy conversion efficiency, and death rate. The other model variables and parameters are the same as in the model by Schreiber and Rudolf (2008) (see above). This system has only one stable equilibrium, where CJ* = dP/bPaP and CA* = KJbJaJdP(rJbPaP ‐ aJdP)/rJb-P
2aP2dA (center panel of Fig. 4.2). Given that alternative stable states can exist in the
absence of the predator (left panel of Fig. 4.2; see also Schreiber and Rudolf 2008), this result indicates that stage-specific predation suppresses alternative stable states and prevents regime shifts after environmental changes. This can be explained intui-tively as follows: The introduced predator P relaxes resource competition among juve-niles by controlling juvenile abundance. This breaks down the positive feedback caused by the apparent competition-like interactions between the resources at the juve-nile and adult stages.
4 A Dynamic Resilience Perspective Toward Integrated Ecosystem Management…

77
When another species is added as a new top predator feeding on predator P, the model is modified as follows:
dR dtJ J J J J J J Jr R K R a C R( )/ /= 1- - (4.3a)
dR dtA A A A A A Ar R K a C R( )/ /= −1 - (4.3b)
dC dtJ A A A A J J J J J J P Jb a C R b a C R d C a C P/ = - - - (4.3c)
dC dtA J J J J A Ab a C R d C/ = - (4.3d)
dP dt PQ/ = b a C P d P aP P J P Q- -
(4.3e)
dQ dt PQ/ = −b a d QQ Q Q
(4.3f)
The variable Q represents the abundance of the newly added predator. The parameters aQ, bQ, and dQ are the attack rate, energy conversion efficiency, and death rate, respectively, of predator Q. This model predicts that the system recovers the potential for alternative stable states (right panel of Fig. 4.2; see Nakazawa 2011a for analysis). This is because the new top predator Q controls intermediate predator P abundance and cancels the suppression of alternative stable states caused by pred-ator P. The same qualitative result is obtained when the presence of predators is manipulated at the adult stage or at both juvenile and adult stages (Nakazawa 2011a). This means that regime shifts can occur if either of the following two condi-tions is satisfied for both juveniles and adults: they are top predators (left panel of Fig. 4.2), or they are consumed by an intermediate predator that is attacked by a top predator (right panel of Fig. 4.2). Notably, juvenile and adult trophic levels do not affect the results (Nakazawa 2011a). Therefore, top-down regulation by predators critically affects the potential for regime shifts. On the other hand, the bottom-up effects of resource availability (e.g., stage-specific resource productivity and trophic levels) alters the likelihood that a system will be in a particular alternative stable state (i.e., the parameter space) when regime shifts are possible; however, bottom- up effects do not affect whether regime shifts can occur (Nakazawa 2011a).
The potential for alternative stable states in systems wherein juveniles and adults are generalist consumers can be explored by manipulating the number of alternative resources that are available at each life-stage (Nakazawa 2011b). A one-consumer n-resource system without predators is the simplest case, and can be modeled as:
dR dtJ i J i J i J i J i J i J i J ir R K R a C R( ), , , , , , , ,/ /= 1- -
(4.4a)
dR dtA i A i A i A i A i A i A i A ir R K R a C R( ), , , , , , , ,/ /= 1- -
(4.4b)
dC dtJ A i A i A i A J i J i J i J J i J ib a R C b a R C d C/ , , , , , , , ,= ( ) ( )∑ ∑- -
(4.4c)
4.4 Effects of Ecological Contexts on the Occurrence of Regime Shifts Mediated…

78
dC dtA J i J i J i J A i A ib a R C d C/ , , , , ,= ( )∑ -
(4.4d)
In this model, i (i = 1 to nh) denotes the ith resource of the juveniles or adults and nh (h = J or A) is the number of resources used by the juveniles or adults. Analysis of the model showed that the relative balance of total resource availability is crucial for the development of alternative stable states. This is probably because the relative balance of maturation and reproduction flow allows for the development of the posi-tive feedback that is caused by apparent competition-like interactions between the juvenile and adult resources (see above). The same qualitative results are obtained when the juveniles or adults utilize allochthonous subsidies from outside the system (Nakazawa 2011b).
The results of the models in this section indicate that food-web topology is a key determinant of the regime shift occurrence. Anthropogenic changes in species composition, due to extinction and invasion, may cause or prevent regime shifts by altering the resilience of ecosystems and communities to environmental pertur-bations. For example, regime shifts may become more likely when a native top predator goes extinct (from center to left panels of Fig. 4.2) or when a predator is introduced and feeds on the native top predator (from center to right panels of Fig. 4.2). While this is an oversimplification of real-life situations, it is sufficient to illustrate the point.
4.4.2 Spatial Configuration
In the field of spatial ecology, researchers have increasingly recognized that spa-tially distinct food webs are frequently connected by the movement of organisms and that local food-web dynamics are tightly regulated by biotic and abiotic con-ditions within distinct habitats. This idea has been considered from a variety of perspectives, including meta-community (Holyoak et al. 2005), spatial subsidy and cross-ecosystem linkage (Polis et al. 2004, food web theory (Rooney et al. 2006), and meta-ecosystem perspectives (Loreau et al. 2003). Even though onto-genetic food-web coupling is fairly common in nature, ontogenetic niche shifts have rarely been considered by spatial ecologists as a major factor coupling dis-tinct food webs. As a well-known example, aquatic insect and amphibian meta-morphosis couples aquatic and terrestrial food webs (Nakano and Murakami 2001; Baxter et al. 2005; Knight et al. 2005). In addition, the so-called breeding migration that has been observed in a variety of animal taxa couples the spatially distinct food webs that are in feeding and breeding grounds (see reviews in Nakazawa and Yamamura 2007). Ontogenetic food-web coupling is widely observed on a smaller scale within localized areas. For example, many fish species switch from planktivory to benthivory as they grow, coupling the pelagic and lit-toral food webs of aquatic ecosystems (Persson et al. 1996; Reñones et al. 2002; Xu et al. 2007; Briones et al. 2012).
4 A Dynamic Resilience Perspective Toward Integrated Ecosystem Management…

79
The development of alternative stable states in meta-community dynamics driven by ontogenetic niche shifts can also be explored through modeling (Nakazawa 2011b). Suppose here that both juveniles and adults are primary consumers and they have multiple stage-specific habitats, in which they exploit different resources. For simplicity, assume that the habitat colonization processes are random and the juve-niles and adults do not move among stage-specific habitats. The dynamics of this system are described as follows:
dR dtJ i J i J i J i J i J i J i J ir R K R a C R( ), , , , , , , ,/ /= 1- -
(4.5a)
dR dtA i A i A i A i A i A i A i A ir R K R a C R( ), , , , , , , ,/ /= −1 -
(4.5b)
dC dtJ i J A i A i A i A i J i J i J i J i J i Jn b a C R b a C R d C, , , , , , , , , ,/ /= ( ) ( )∑1 - - ,,i
(4.5c)
dC dtA i A J i J i J i J i A i A in b a C R d C, , , , , , ,/ /= ( ) ( )∑1 �
(4.5d)
In the model, i (i = 1 to nh) denotes the ith habitat of the juveniles or adults and nh (h = J or A) is the number of habitats used by the juveniles or adults. Definitions of the other variables and parameters are the same as in the models presented previ-ously. The juveniles and adults are recruited into each stage-specific habitat at the rate of (1/nh) ∑ (bh,iah,iCh,iRh,i) (i.e., the total reproduction or maturation production divided by the stage-specific habitat number).
First, consider the simple situation in which parameter values are identical in all juvenile or adult habitats (i.e., rh,i = rh, Kh,i = Kh, ah,i = ah, bh,i = bh and dh,i = dh). Under these conditions, Rh,i* = Rh*and Ch,i* = Ch*, which allows rigorous mathematical analysis (see Nakazawa 2011b for details). The model predicts that alternative sta-ble states exist (i.e., regime shifts can occur) when the number of juvenile and adult habitats are comparable. This condition is consistent with the argument outlined above that the relative balance of the total resource availability in the juvenile and adult habitats is essential for the development of alternative stable states. This also suggest that if maturation rate surpasses reproduction rate (e.g., due to higher pro-ductivity in juvenile habitats than in adult habitats), then an alternative stable state could exist when there are fewer juvenile habitats than adult habitats. In reality, of course, it is not easy to rigorously count the number of stage-specific habitats or to fully characterize the habitat properties that determine maturation and reproduction rates. Nevertheless, these results clearly indicate that spatial configuration such as habitat area and connectivity critically affect the occurrence of regime shifts.
On the basis of these predictions, I emphasize that changes in spatial configura-tion due to habitat loss or fragmentation may critically affect the occurrence of regime shifts. Suppose that both juvenile and adult stages have two habitats in the initial spatial configuration (i.e., nh = 2), alternative stable states exist over a certain range of environmental conditions, and regime shifts can occur through changes in adult habitat productivity (center panel of Fig. 4.3). However, regime shifts are pre-vented when a juvenile habitat is lost (left panel of Fig. 4.3). This can be explained
4.4 Effects of Ecological Contexts on the Occurrence of Regime Shifts Mediated…
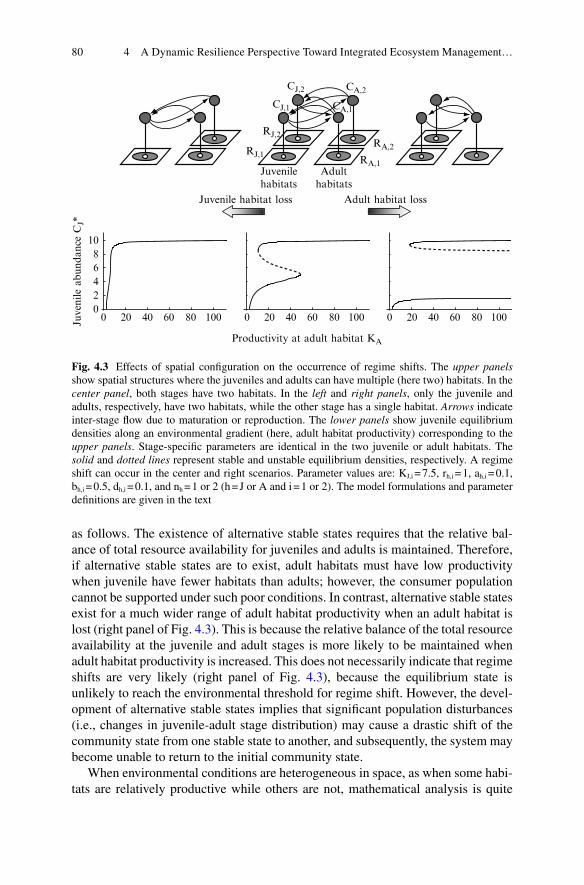
80
as follows. The existence of alternative stable states requires that the relative bal-ance of total resource availability for juveniles and adults is maintained. Therefore, if alternative stable states are to exist, adult habitats must have low productivity when juvenile have fewer habitats than adults; however, the consumer population cannot be supported under such poor conditions. In contrast, alternative stable states exist for a much wider range of adult habitat productivity when an adult habitat is lost (right panel of Fig. 4.3). This is because the relative balance of the total resource availability at the juvenile and adult stages is more likely to be maintained when adult habitat productivity is increased. This does not necessarily indicate that regime shifts are very likely (right panel of Fig. 4.3), because the equilibrium state is unlikely to reach the environmental threshold for regime shift. However, the devel-opment of alternative stable states implies that significant population disturbances (i.e., changes in juvenile-adult stage distribution) may cause a drastic shift of the community state from one stable state to another, and subsequently, the system may become unable to return to the initial community state.
When environmental conditions are heterogeneous in space, as when some habi-tats are relatively productive while others are not, mathematical analysis is quite
Productivity at adult habitat KA
Juve
nile
abu
ndan
ce C
J*
Adult habitat lossJuvenile habitat loss
Juvenilehabitats
Adulthabitats
1086420
CJ,1
CJ,2 CA,2
RJ,2
RJ,1RA,2
RA,1
CA,1
40 60 80 100200 40 60 80 100200 40 60 80 100200
Fig. 4.3 Effects of spatial configuration on the occurrence of regime shifts. The upper panels show spatial structures where the juveniles and adults can have multiple (here two) habitats. In the center panel, both stages have two habitats. In the left and right panels, only the juvenile and adults, respectively, have two habitats, while the other stage has a single habitat. Arrows indicate inter-stage flow due to maturation or reproduction. The lower panels show juvenile equilibrium densities along an environmental gradient (here, adult habitat productivity) corresponding to the upper panels. Stage-specific parameters are identical in the two juvenile or adult habitats. The solid and dotted lines represent stable and unstable equilibrium densities, respectively. A regime shift can occur in the center and right scenarios. Parameter values are: KJ,i = 7.5, rh,i = 1, ah,i = 0.1, bh,i = 0.5, dh,i = 0.1, and nh = 1 or 2 (h = J or A and i = 1 or 2). The model formulations and parameter definitions are given in the text
4 A Dynamic Resilience Perspective Toward Integrated Ecosystem Management…

81
difficult. However, preliminarily numerical simulations show that multiple (i.e., more than two) stable states can exist in spatially heterogeneous environments (Nakazawa 2011b). Assume that both juveniles and adults have two habitats (i.e., nh = 2) and the productivities of the two juvenile habitats (i.e., KJ,1 and KJ,2) are dif-ferent. Under these conditions, at least three alternative stable states are found for some parameter settings (Nakazawa 2011b). One of these stable states is an adult-dominated state, and the other two are juvenile-dominated states, in which juvenile abundance is dominated by either of the two subpopulations. This result suggests that understanding spatial community dynamics is much more difficult than we have supposed and it may be impossible without sufficiently detailed information regarding the initial spatial distribution of species. This idea is apparently consistent with the notion of historical contingency of community assembly, which states that equilibrium community structure is governed by initial conditions and stochastic processes (Law 1999; Fukami 2004; but see Fukami and Nakajima 2011). The model results also indicate that juvenile abundance can be higher in the less produc-tive habitat than in the more productive habitat (Nakazawa 2011b). This pattern is produced by a source-sink relationship among stage-specific habitats; that is, the more productive juvenile habitat can support the juvenile subpopulation in the less productive habitat through facilitation of maturation. The existence of multiple sta-ble states suggests that mismatches between local environmental conditions and local community structure are possible, that the source-sink relationships depend on the initial conditions, and that these relationships may suddenly collapse (i.e., regime shifts occur) following environmental change. These predictions are still preliminary but potentially quite important for ecosystem management in space. Future work is needed to more fully elucidate how the regime shifts are influenced by changes in spatial configuration and local environmental conditions.
4.4.3 Seasonality
Phenology, the timing of life-history events such as breeding and migration, is cru-cial for population dynamics. The importance of phenology in population dynamics is illustrated by the match/mismatch hypothesis. This hypothesis states that if the most energy-expensive period of the consumer’s reproductive phenology is close to the peak period of the resource availability, then recruitment of the consumer will be high (Cushing 1990). This is becoming a key concept in seasonal community dynamics (Nakazawa and Doi 2012). This idea can also be applied to a stage- structured system. Assume that juveniles and adults are primary consumers and they change their maturation or reproduction efficiencies seasonally, as follows:
dR dtJ J J J J J J Jr R K R a C R/ /= ( )1 � �
(4.6a)
dR dtA A A A A A A Ar R K R a C R( )/ /= 1� � (4.6b)
4.4 Effects of Ecological Contexts on the Occurrence of Regime Shifts Mediated…

82
dC dtJ A A A A A J J J J J J Jp t b a C R p t b a C R d C/ = ( ) ( )� �
(4.6c)
dC dtA J J J J J A Ap t b a C R d C/ = ( ) �
(4.6d)
In the model, the time-varying function ph(t) (h = J or A) represents the phenol-ogy of maturation or reproduction. Note that the phenology in the model indicates seasonal variations in behavioral efficiency or activity that are governed by environ-mental cues (e.g., temperature and precipitation), rather than by resource consump-tion. The functional form of ph(t) is determined by empirical data or specific assumptions. For simplicity, and as is common in the literature (Namba 1984; Abrams 2004; Nakazawa and Doi 2012), I opt here for sinusoidal functions, ph(t) = 1 + uhsin{2π(t ‐ vh)/T}, where T, uh, and vh represent seasonal period, amplitude, and peak timing of maturtion or reproduction, respectively.
Using the model, numerical simulations of long-term (i.e., over many seasonal cycles) average community states were performed with initial population abun-dances of Rh = 1 and Ch = 1. When the model is run along a gradient of adult habitat productivity (KA) for differing degrees of phenological mismatch between matura-tion and reproduction (i.e., vJ − vA), the results show that a regime shift-like transi-tion occurs around KA = 15 when vJ − vA = 0 and that the severity or threshold of the shift varied with changes in the maturation-reproduction phenological mismatch (Fig. 4.4a). When the long-term average abundances of both juveniles and adults along a gradient of maturation-reproduction phenological mismatches are evalu-ated, the results show that juvenile and adult abundances are highly variable, depending on maturation and reproduction phenology. Most notably, the stage dis-tribution may suddenly and drastically shift when maturation–reproduction pheno-logical mismatch changes (Fig. 4.4b). Detecting regime shifts in scenarios like this is difficult because the system undergoes seasonal (i.e., ever-fluctuating) changes, making equilibrium states impossible to evaluate. Nevertheless, the above results clearly indicate that long-term community dynamics may change drastically with match/mismatch of maturation and reproduction phenology.
Climate change has altered individual growth rates and, as a result, the timing of ontogenetic niche shifts (Post et al. 2008; Seebens et al. 2009; Ozgul et al. 2010; Yang and Rudolf 2010) and reproduction (Beebee 1995; Crick et al. 1997; Taylor 2007). Therefore, climate change-indued phenological shifts may also alter the resilience of communities. However, it is still not clear how climate change advances or delays species-specific and trait-specific phenology. This is because life-history parameters are bioenergetically related to one another and climate change regulates multiple life-history traits in complex ways (Ozgul et al. 2010). Accumulating phe-nology data is necessary in order to better understand the occurrence of regime shifts in seasonal community dynamics and to more accurately predict their responses to climate change.
4 A Dynamic Resilience Perspective Toward Integrated Ecosystem Management…
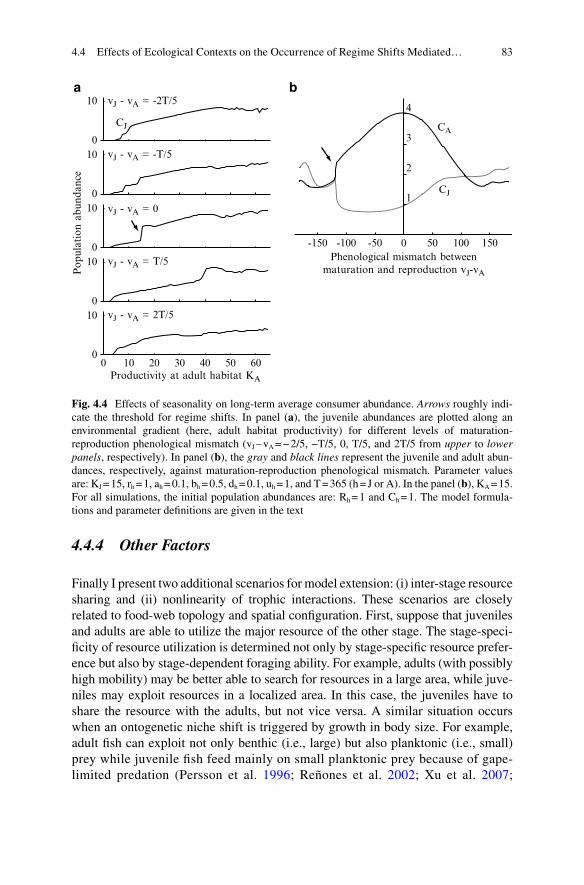
83
4.4.4 Other Factors
Finally I present two additional scenarios for model extension: (i) inter-stage resource sharing and (ii) nonlinearity of trophic interactions. These scenarios are closely related to food-web topology and spatial configuration. First, suppose that juveniles and adults are able to utilize the major resource of the other stage. The stage-speci-ficity of resource utilization is determined not only by stage-specific resource prefer-ence but also by stage-dependent foraging ability. For example, adults (with possibly high mobility) may be better able to search for resources in a large area, while juve-niles may exploit resources in a localized area. In this case, the juveniles have to share the resource with the adults, but not vice versa. A similar situation occurs when an ontogenetic niche shift is triggered by growth in body size. For example, adult fish can exploit not only benthic (i.e., large) but also planktonic (i.e., small) prey while juvenile fish feed mainly on small planktonic prey because of gape-limited predation (Persson et al. 1996; Reñones et al. 2002; Xu et al. 2007;
Phenological mismatch betweenmaturation and reproduction vJ-vAP
opul
atio
n ab
unda
nce
CA
CJ
4
3
2
1
10
010
010
010
010
00
Productivity at adult habitat KA
CJ
vJ - vA = 2T/5
vJ - vA = -2T/5
vJ - vA = T/5
vJ - vA = -T/5
vJ - vA = 0
10 20 30 40 50 60
-150 -100 -50 0 50 100 150
a b
Fig. 4.4 Effects of seasonality on long-term average consumer abundance. Arrows roughly indi-cate the threshold for regime shifts. In panel (a), the juvenile abundances are plotted along an environmental gradient (here, adult habitat productivity) for different levels of maturation- reproduction phenological mismatch (vJ – vA = − 2/5, −T/5, 0, T/5, and 2T/5 from upper to lower panels, respectively). In panel (b), the gray and black lines represent the juvenile and adult abun-dances, respectively, against maturation-reproduction phenological mismatch. Parameter values are: KJ = 15, rh = 1, ah = 0.1, bh = 0.5, dh = 0.1, uh = 1, and T = 365 (h = J or A). In the panel (b), KA = 15. For all simulations, the initial population abundances are: Rh = 1 and Ch = 1. The model formula-tions and parameter definitions are given in the text
4.4 Effects of Ecological Contexts on the Occurrence of Regime Shifts Mediated…

84
Briones et al. 2012). This example may be categorized as ontogenetic niche expan-sion. I developed a one-consumer two-resource model with inter-stage resource shar-ing as follows:
dR dtJ J J J J J J J J A J A Jr R K R a C R a C R( )/ / , ,= 1� � �
(4.7a)
dR dtA A A A A J A J A A A A Ar R K R a C R a C R( )/ / , ,= 1� � �
(4.7b)
dC dtJ A J A J J A A A A A A J J J J J J A J A A Jb a R b a R C b a R b a R C/ , , , , , , , ,= +( ) +( )� ��d CJ i J i, , (4.7c)
dC dtA J J J J J J A J A A J A i A ib a R b a R C d C/ , , , , , ,= +( ) �
(4.7d)
where aJ,A and aA,J determine the rates at which juveniles or adults consume the resource of the other stage. Analysis of the model shows that alternative stable states are generally suppressed as these parameters increase in value (Nakazawa 2011b); that is, inter-stage resource sharing suppresses the occurrence of regime shifts. This result can be explained as follows: If the adults exploit the juvenile resource with high efficiency, thereby regulating the amount of the juvenile resource, then it can interrupt the positive feedback caused by apparent competition-like interactions between juvenile and adult resources.
The nonlinearity of trophic interactions is also crucial for understanding the occurrence of regime shifts. Schreiber and Rudolf (2008) showed through numeri-cal simulations that regime shifts are generally smoothed or prevented when the juveniles or adults have a long handling time (i.e., Holling type-II functional responses) in the simplest one-consumer-two resource system (model 1). Similar patterns are observed when there is strong interference competition within stages (Nakazawa 2011a). This is because low food-dependent maturation or reproduction interrupts the positive feedback that is necessary for the development of alternative stable states. Other species (e.g., predators) may also have nonlinear functional responses that have significant effects on the occurrence of regime shifts. Suppose that juveniles have a top predator with a nonlinear functional response. Nakazawa (2011a) showed that, in contrast to systems with linear functional responses of pred-ators (center panel of Fig. 4.2), alternative stable states can exist when the predator exhibits strong interference competition. This is because interference competition controls prey (i.e., the juveniles) in a density-dependent manner similar to the con-trol exerted when a new top predator attacks the predator (right panel of Fig. 4.2). On the other hand, regime shifts remain impossible when the predator has a Holling type-II functional response (Nakazawa 2011a). These results suggest that the non-linearity of trophic interactions may either cause or prevent the occurrence of regime shifts, depending on the basic food-web configuration.
4 A Dynamic Resilience Perspective Toward Integrated Ecosystem Management…

85
4.5 Toward Integrated Ecosystem Management
4.5.1 From Static to Dynamic Resilience Perspectives
The occurrence of regime shifts that are mediated by ontogenetic niche shifts can be significantly altered by various factors, such as food-web topology, spatial configu-ration, and seasonality. These results shed light on our understanding of regime shifts in complex ecosystems. At the same time, they open new avenues for future research and for establishing more effective ecosystem management based on resil-ience theory. What is the most important is to more accurately evaluate ecological contexts (i.e., species compositions and their relationships in space and time) to understand the occurrence of regime shifts. This information will allow us to under-stand long-term community dynamics and ecosystem responses to disturbances. This idea is straightforward, but has not been established by empirical research on regime shifts and ecosystem management. This is because ecosystem disturbances (e.g., biodiversity degradation, landscape modification, and phenological shift) and regime shift (or resilience) have conventionally been studied independently of one another in ecosystem management research (Peterson et al. 1998; Awiti 2011). In particular, the primary concern in the previous study of regime shifts has been the determination of the point at which regime shifts occur along environmental gradi-ents (e.g., nutrient loading) and how to predict them (Andersen et al. 2009). These attempts implicitly assume that ecological contexts are invariant. The only excep-tions, to my knowledge, are the work of Genkai-Kato and Carpenter (2005) and Genkai-Kato (2007). These researchers studied how regime shifts in shallow eutro-phic lakes would be altered by changes in the relative pelagic-to-littoral habitat area and the associated presence of macrophytes or piscivores, respectively. Genkai- Kato and Carpenter (2005) argued that the resilience is determined by lake size and morphology, but their results can be used to suggest that resilience may be altered by lakeshore development through decreases in macrophyte-dominated littoral area. Genkai-Kato (2007) incorporated the effects of macrophytes and piscivores by manipulating parameter values associated with zooplankton grazing rate or plankti-vore death rate, although he did not explicitly describe their population dynamics. Despite these studies, however, regime shifts in complex ecosystems remain poorly understood. More detailed studies on regime shift and resilience should be taken into account in future, and even past, studies of regime shift and resilience.
The resilience of ecosystems is determined by ecological contexts. This implies that changes in ecological context due to anthropogenic disturbances may affect whether or where regime shifts occur along environmental gradients. Recognizing the importance of the novel concept, here I wish to propose “dynamic resilience perspective,” in contrast to the conventional view that ecological resilience is defined in constant ecological contexts (Gunderson 2000). This concept is quite important in ecosystem management because of current various ecosystem distur-bances. Acceleration of biodiversity loss and biological invasion are serious con-cerns all over the world (Lovel 1997; Chapin et al. 2000; Sala et al. 2000). Apex
4.5 Toward Integrated Ecosystem Management

86
predatory species are especially vulnerable to extinction (Myers and Worm 2003; Prugh et al. 2009), while invasive predators frequently occupy the top-predator niche, exploiting the resident prey community (Eby et al. 2006). In addition, habitat loss and fragmentation due to landscape modification (Fahrig 2003; Cushman 2006; Collinge 2009) and phenological shifts due to climate change (Lovejoy and Hannah 2005; Hannah 2010; Kernan et al. 2010) are increasingly serious worldwide. Theoretical predictions based on the dynamic resilience perspective suggest that these anthropogenic ecosystem disturbances may modify the community dynamics and spatial distribution of species in ways that are not yet fully understood. An emerging concern is that these disturbances may together create a negative spiral, whereby ecosystem disturbances make ecological communities more susceptible to environmental change-induced regime shifts and regime shifts make ecosystems more vulnerable to human disturbance. In order to avoid such a resilience melt-down, we should be more aware of the causal linkages between ecological context and ecosystem resilience, conventionally studied independently. One may think that it is virtually impossible to completely describe the real world, and that such attempts are not promising. Although it is certainly challenging, one possibility may stem from the fact that there are the general rules governing the occurrence of regime shifts that are mediated by ontogenetic niche shifts. These rules describe the ways in which top-down regulation by predators and bottom-up regulation by resources control the positive feedback that is necessary for the development of alternative stable states (Nakazawa 2011a, b). We may be able to predict the occur-rence of regime shifts in complex ecosystems by determining the extent to which basic mechanisms that underlie alternative stable states are density-dependent and how they interact with other factors and processes. In this chapter, I have focused on ontogenetic niche shifts as generators of alternative stable states. However, the same arguments apply to regime shifts caused by other mechanisms and are therefore generally applicable to ecosystem management.
4.5.2 Implications for Lake Ecosystem Management
The occurrence of regime shifts that are mediated by ontogenetic niche shifts would be likely in lake ecosystems (e.g., Persson et al. 2007). This is because aquatic ani-mals grow by several orders of magnitude in size between hatching and death and they typically exhibit clear ontogenetic niche shifts (Hildrew et al. 2007), and also because lakes are basically closed systems to which simple theoretical analysis is relatively applicable. I also emphasize that lake ecosystems are highly susceptible to various anthropogenic disturbances including biodiversity degradation and habi-tat modification resulting from human activity (Myers and Worm 2003; Eby et al. 2006; Collinge 2009). As a consequence, the dynamic resilience perspective may be useful to manage lake ecosystems successfully.
From this viewpoint, I end this chapter by briefly discussing possible implications of the present theoretical predictions for ecosystem management of Lake Biwa.
4 A Dynamic Resilience Perspective Toward Integrated Ecosystem Management…

87
Fortunately, a large amount of biological and environmental data has been accumu-lated by regular monitoring and sampling programs in Lake Biwa. These data may provide valuable opportunities to consider the possibility. First, Lake Biwa was invaded by piscivorous fishes (bluegill and largemouth bass) in the 1960–1970s (Nakai 1999; Nakazawa et al. 2007), as with many temperate lakes in North America and Europe (Rahel 2000; Hickley and Chare 2004). The dynamic resilience perspec-tive suggests that these species invasions might have significantly influenced how the native lake community responds to, and absorb, environmental disturbances by alter-ing cascading top-down effects upon lower trophic levels (see Fig. 4.2). If this mech-anism operates, some species may have become less likely to recover from environmental change-induced population declines in the presence of the introduced piscivores. Secondly, lakes are not completely closed systems but can be link to one another by rivers, canals, and mobile animals. Furthermore, aquatic organisms often form spatial population structure in large lakes. For example, previous studies using stable isotopes and DNA markers have shown that some species would not be eco-logically or genetically uniform in Lake Biwa with a surface area of about 670 km2 (Takai and Sakamoto 1999; Iguchi et al. 2008; but see Takahashi and Ohara 2004). I also note that Lake Biwa is connected with many influent rivers and nearby satellite lakes. Some fish species exhibit ontogenetic or breeding migration between Lake Biwa and these surrounding waters (Shibata et al. 2010). Therefore, lakeshore devel-opment in the past may have affected the ecosystem resilience mediated by spatial structure through decreases in or degradation of littoral habitats (see Fig. 4.3; also see Nakanish and Sekino 1996). Finally, climate change can also impact on lake ecosys-tems. Recent studies showed that warming is an important driver of plankton com-munity dynamics in Lake Biwa (Hsieh et al. 2010, 2011; Tsugeki et al. 2010), although long-term variations in intra-annual population dynamics (i.e., phenologi-cal shifts) remain unclear especially for higher trophic levels. Intriguingly, these eco-system responses to climate change and eutrophication seem interactive. This may imply that climate change also affects the ecosystem resilience to environmental disturbance (see Fig. 4.4). Overall, although these theoretical implications are not yet fully elucidated by empirical evidence, the dynamic resilience perspective suggests that the ecosystem management of Lake Biwa (and other lakes) may benefit by being more interdisciplinary and taking into account multiple ecosystem disturbances associated with biodiversity, landscape, and climate.
In conclusion, we are still in the early stages of understanding how the resilience of complex lake ecosystems is determined and how it is altered by anthropogenic disturbances. Both theoretical and empirical advances are needed to refine the exist-ing conception of ecological resilience and to develop more effective ecosystem management that integrates biodiversity conservation, habitat protection, and cli-mate change adaptation. Models and experiments that explicitly consider additional factors are inevitably more complex than those that ignore them. However, given the importance in ecological dynamics, the potential gains in understanding warrant the additional effort required to include these factors. I conclude that ecosystem man-agement should be made using the dynamic resilience perspective. The scenario- based approach I outlined here will serve as the initial step.
4.5 Toward Integrated Ecosystem Management

88
Acknowledgements I thank anonymous reviewers for valuable comments on the manuscript. This research was supported by the Japan Society for Promotion of Science Research Fellowship for Young Scientists.
References
Abrams P (2004) When does periodic variation in resource growth allow robust coexistence of competing consumer species? Ecology 85:372–382
Abrams P (2009) When does greater mortality increase population size? The long history and diverse mechanisms underlying the hydra effect. Ecol Lett 12:462–474
Andersen T, Carstensen J, Hernández-García E, Duarte CM (2009) Ecological thresholds and regime shifts: approaches to identification. Trends Ecol Evol 24:49–57
Awiti AO (2011) Biological diversity and resilience: lessons from the recovery of cichlid species in Lake Victoria. Ecol Soc 16:9
Baxter CV, Fausch K, Saunders WC (2005) Tangled webs: reciprocal flows of invertebrate prey link streams and riparian zones. Freshw Biol 50:201–220
Beebee TJC (1995) Amphibian breeding and climate. Nature 374:219–220Briones JC, Tsai CH, Nakazawa T, Sakai Y, Papa RD, Hsieh CH, Okuda N (2012) Long-term
changes in the diet of Gymnogobius isaza from Lake Biwa, Japan: effects of body size and environmental prey availability. PLoS One 7:e53167. doi:10.1371/journal.pone.0053167
Chapin FS, Zavaleta ES, Eviner VT, Naylor RL, Vitousek PM, Reynolds HL, Hooper DU, Lavorel S, Sala OE, Hobbie SE, Mack MC, Díaz S (2000) Consequences of changing biodiversity. Nature 405:234–242
Collinge SK (2009) Ecology of fragmented landscapes. Johns Hopkins University Press, BaltimoreCrick HQP, Dudley C, Glue DE, Thomson DL (1997) UK birds are laying eggs earlier. Nature
388:526Cushing DH (1990) Plankton production and year-class strength in fish populations: an update of
the match mismatch hypothesis. Adv Mar Biol 26:249–293Cushman SA (2006) Effects of habitat loss and fragmentation on amphibians: a review and pro-
spectus. Biol Conserv 128:231–240de Roos AM, Leonardsson K, Persson L, Mittelbach GG (2002) Ontogenetic niche shifts and flex-
ible behavior in size-structured populations. Ecol Monogr 72:271–292de Roos AM, Schellekens T, van Kooten T, van de Wolfshaar K, Claessen D, Persson L (2007)
Food-dependent growth leads to overcompensation in stage-specific biomass when mortal-ity increases: he influence of maturation versus reproduction regulation. Am Nat 170:E59–E76
de Roos AM, Schellekens T, Van Kooten T, Persson L (2008) Stage-specific predator species help each other to persist while competing for a single prey. Proc Natl Acad Sci U S A 105:13930–13935
Eby LA, Roach WJ, Crowder LB, Stanford JA (2006) Effects of stocking-up freshwater food webs. Trends Ecol Evol 21:576–584
Fahrig L (2003) Effects of habitat fragmentation on biodiversity. Annu Rev Ecol Evol Syst 34:487–515
Folke C, Carpenter S, Walker B, Scheffer M, Elmqvist T, Gunderson L, Holling CS (2004) Regime shifts, resilience, and biodiversity in ecosystem management. Annu Rev Ecol Evol Syst 35:557–581
Fukami T (2004) Community assembly along a species pool gradient: implications for multiple- scale patterns of species diversity. Popul Ecol 46:137–147
Fukami T, Nakajima M (2011) Community assembly: alternative stable states or alternative tran-sient states? Ecol Lett 14:973–984
Genkai-Kato M (2007) Macrophyte refuges, prey behaviour and trophic interactions: conse-quences for lake water clarity. Ecol Lett 10:105–114
4 A Dynamic Resilience Perspective Toward Integrated Ecosystem Management…

89
Genkai-Kato M, Carpenter SR (2005) Eutrophication due to phosphorus recycling in relation to lake morphometry, temperature, and macrophytes. Ecology 86:210–219
Genkai-Kato M, Vadeboncoeur Y, Liboriussen L, Jeppesen E (2012) Benthic–planktonic coupling, regime shifts, and whole-lake primary production in shallow lakes. Ecology 93:619–631. http://dx.doi.org/10.1890/10-2126.1
Guill C (2009) Alternative dynamical states in stage-structured consumer populations. Theor Popul Biol 76:168–178
Gunderson LH (2000) Ecological resilience–in theory and application. Annu Rev Ecol Evol Syst 31:425–439
Hannah LJ (2010) Climate change biology. Elsevier, BurlingtonHickley P, Chare S (2004) Fisheries for non-native species in England and Wales: angling or the
environment? Fish Manage Ecol 11:203–212Hildrew AG, Raffaelli DR, Edmonds-Brown R (2007) Body size: the structure and function of
aquatic ecosystems. Cambridge University Press, CambridgeHolyoak M, Leibold MA, Holt RD (2005) Metacommunities: spatial dynamics and ecological
communities. University of Chicago Press, ChicagoHsieh CH, Ishikawa K, Sakai Y, Ishikawa T, Ichise S, Yamamoto Y, Kuo TC, Park HD, Yamamura
N, Kumagai M (2010) Phytoplankton community reorganization driven by eutrophication and warming in Lake Biwa. Aquat Sci 72:467–483
Hsieh CH, Sakai Y, Ban S, Ishikawa K, Ishikawa T, Ichise S, Yamamura N, Kumagai M (2011) Eutrophication and warming effects on long-term variation of zooplankton in Lake Biwa. Biogeosciences 8:1383–1399
Iguchi K, Ohkawa T, Nishida M (2008) Genetic structure of land-locked ayu within the Biwa Lake system. Fish Sci 68:138–143
Jansson M, Persson L, de Roos AM, Jones RI, Tranvik LJ (2007) Terrestrial carbon and intraspe-cific size-variation shape lake ecosystems. Trends Ecol Evol 22:316–322
Kawanabe H, Nishino M, Maehata M (2012) Lake Biwa: interactions between nature and people. Springer Academic, Amsterdam
Kernan M, Battarbee RW, Moss BR (2010) Climate change impacts on freshwater ecosystems. Wiley-Blackwell, West Sussex
Knight TM, McCoy MW, Chase JM, McCoy KA, Holt RD (2005) Trophic cascades across ecosystems. Nature 437:880–883
Law R (1999) Theoretical aspects of community assembly. In: McGlade J (ed) Advanced ecologi-cal theory. Blackwell, Oxford
Loreau M, Mouquet N, Holt RD (2003) Meta-ecosystems: a theoretical framework for a spatial ecosystem ecology. Ecol Lett 6:673–679
Lovejoy T, Hannah L (2005) Climate change and biodiversity. Yale University Press, New HavenLovel G (1997) Biodiversity: global change through invasion. Nature 388:627–628Mayer AL, Rietkerk M (2004) The dynamic regime concept for ecosystem management and res-
toration. BioScience 54:1013–1020McCoy MW, Barfield M, Holt RD (2009) Predator shadows: complex life histories as generators
of spatially patterned indirect interactions across ecosystems. Oikos 118:87–100Miller TEX, Rudolf VHW (2011) Thinking inside the box: community-level consequences of
stage structured populations. Trends Ecol Evol 26:457–466Myers RA, Worm B (2003) Rapid worldwide depletion of predatory fish communities. Nature
423:280–283Nakai K (1999) Recent, faunal changes in Lake Biwa, with particular reference to the bass fishing
boom in Japan. In: Kawanabe H, Coulter GW, Roosevelt AC (eds) Ancient lakes: their cultural and biological diversity. Kenobi Productions, Ghent, pp 227–241
Nakanish M, Sekino T (1996) Recent drastic changes in Lake Biwa bio-communities, with special attention to exploitation of the littoral zone. GeoJournal 40:63–67
Nakano S, Murakami M (2001) Reciprocal subsidies: dynamic interdependence between terres-trial and aquatic food webs. Proc Natl Acad Sci U S A 98:166–170
Nakazawa T (2011a) Ontogenetic niche shift, food-web coupling, and alternative stable states. Theor Ecol 4:479–492
References

90
Nakazawa T (2011b) Alternative stable states generated by ontogenetic niche shift in the presence of multiple resource use. PLoS One 6:e14667
Nakazawa T, Doi H (2012) A perspective on match/mismatch of phenology in community con-texts. Oikos 121:489–495
Nakazawa T, Yamamura N (2007) Breeding migration and population stability. Popul Ecol 49:101–113
Nakazawa T, Ishida N, Kato M, Yamamura N (2007) Larger body size with higher predation rate. Ecol Freshw Fish 16:362–372
Nakazawa T, Ushio M, Kondoh M (2011) Scale dependence of predator–prey mass ratio: determi-nants and applications. In: Belgrano A, Reiss J (eds) The role of body size in multispecies systems. Adv Ecol Res 45:269–302
Namba T (1984) Competitive co-existence in a seasonally fluctuating environment. J Theor Biol 111:369–386
Ohlberger J, Langangen Ø, Edeline E, Claessen D, Winfield I, Stenseth N, Vollestad A (2012) Stage-specific biomass overcompensation by juveniles in response to increased adult mortality in a wild fish population. Ecology. doi:10.1890/11-0410.1
Ozgul A, Childs DZ, Oli MK, Armitage KB, Blumstein DT, Olson LE, Tuljapurkar S, Coulson T (2010) Coupled dynamics of body mass and population growth in response to environmental change. Nature 466:482–485
Persson L, Andersson J, Wahlstrom E, Eklov P (1996) Size-specific interactions in lake systems: predator gape limitation and prey growth rate and mortality. Ecology 77:900–911
Persson L, Amundsen PA, de Roos AM, Klementsen A, Knudsen R, Primicerio P (2007) Culling prey promotes predator recovery–alternative stable states in a whole lake experiment. Science 316:1743–1746
Peterson GD (2002) Forest dynamics in the southeastern U S: managing multiple stable states. In: Gunderson LH, Pritchard L Jr (eds) Resilience and the behavior of large-scale ecosystems. Island, Washington, DC, pp 227–246
Peterson G, Allen CR, Holling CS (1998) Ecological resilience, biodiversity, and scale. Ecosystems 1:6–18
Polis GA, Power ME, Huxel GR (2004) Food webs at the landscape level. University of Chicago Press, Chicago
Post E, Pedersen C, Wilmers CC, Forchhammer MC (2008) Phenological sequences reveal aggre-gate life history response to climatic warming. Ecology 89:363–370
Postel S, Carpenter SR (1997) Freshwater ecosystem services. In: Daily G (ed) Nature’s services. Island, Washington, DC
Prugh LR, Stoner CJ, Epps CW, Bean WT, Ripple WJ, Laliberte AS, Brashares JS (2009) The rise of the mesopredator. Bioscience 59:779–791
Rahel FJ (2000) Homogenization of fish faunas across the United States. Science 288:854–856Reñones O, Polunin NVC, Goni R (2002) Size related dietary shifts of Epinephelus marginatus in
a western Mediterranean littoral ecosystem: an isotope and stomach content analysis. J Fish Biol 61:122–137
Rooney N, McCann K, Gellner G, Moore JC (2006) Structural asymmetry and the stability of diverse food webs. Nature 442:265–269
Sala OE, Chapin FS, Armesto JJ, Berlow E, Bloomfield J, Dirzo R, Huber-Sanwald E, Huenneke LF, Jackson RB, Kinzig A, Leemans R, Lodge DM, Mooney HA, Oesterheld M, Poff NL, Sykes MT, Walker BH, Walker M, Wall DH (2000) Global biodiversity scenarios for the year 2100. Science 287:1770–1774
Scheffer M (1998) Ecology of shallow lakes. Chapman and Hall, New YorkScheffer M, Carpenter SR (2003) Catastrophic regime shifts in ecosystems: linking theory to
observation. Trends Ecol Evol 18:648–656Scheffer M, Carpenter S, Foley JA, Folke C, Walker B (2001) Catastrophic shifts in ecosystems.
Nature 413:591–596Schindler DE, Scheuerell MD (2002) Habitat coupling in lake ecosystems. Oikos 98:177–189
4 A Dynamic Resilience Perspective Toward Integrated Ecosystem Management…

91
Schreiber S, Rudolf VHW (2008) Crossing habitat boundaries: coupling dynamics of ecosystems through complex life cycles. Ecol Lett 11:576–587
Schröder A, Persson L, de Roos AM (2005) Direct experimental evidence for alternative stable states: a review. Oikos 110:3–19
Seebens H, Einsle U, Straile D (2009) Copepod life cycle adaptations and success in response to phytoplankton spring bloom phenology. Glob Change Biol 15:1394–1404
Shibata J, Karube Z, Oishi M, Yamaguchi M, Goda Y, Okuda N (2010) Physical structure of habi-tat network differently affects migration patterns of native and invasive fishes in Lake Biwa and its tributary lagoons: stable isotope approach. Popul Ecol 53:143–153
Smith VH, Schindler DW (2009) Eutrophication science: where do we go from here? Trends Ecol Evol 24:201–207
Smith VH, Joye SB, Howarth RW (2006) Eutrophication of freshwater and marine ecosystems. Limnol Oceanogr 51:351–355
Suzuki S, Iwasa Y (2009) The coupled dynamics of human socio-economic choice and lake water system: the interaction of two sources of nonlinearity. Ecol Res 24:479–489
Takahashi D, Ohara K (2004) Genetic variations estimated from PCR-RFLP analysis of two morphs of the freshwater goby Rhinogobius in the Lake Biwa water system. Ichthyol Res 51:99–105
Takai N, Sakamoto W (1999) Identification of local populations of Lake Biwa catfish Silurus biwaensis in Japan on the basis of δ13C and δ15N analyses. Can J Zool 77:258–266
Takimoto G (2003) Adaptive plasticity in ontogenetic niche shifts stabilizes consumer-resource dynamics. Am Nat 162:93–109
Taylor SG (2007) Climate warming causes phenological shift in Pink Salmon, Oncorhynchus gor-buscha, behavior at Auke Creek, Alaska. Glob Change Biol 14:229–235
Tsugeki NK, Urabe J, Hayami Y, Kuwae M, Nakanishi M (2010) Phytoplankton dynamics in Lake Biwa during the 20th century: complex responses to climate variation and changes in nutrient status. J Paleolimnol 44:69–83
Vincenzi S, Satterthwaite WH, Mangel M (2012) Spatial and temporal scale of density-dependent body growth and its implications for recruitment, population dynamics and management of stream-dwelling salmonid populationse. Rev Fish Biol Fish. doi:10.1007/s11160-011-9247-1
Walter C, Kiutchell JF (2001) Cultivation/depensation effects on juvenile survival and recruitment: implications for the theory of fishing. Can J Fish Aquat Sci 58:39–50
Werner EE, Gilliam JF (1984) The ontogenetic niche and species interactions in size-structured populations. Annu Rev Ecol Syst 15:393–425
Wilbur HM (1980) Complex life cycles. Annu Rev Ecol Syst 11:67–93Xu J, Zhang M, Xie P (2007) Size-related shifts in reliance on benthic and pelagic food webs by
lake anchovy. Ecoscience 14:170–177Yang LH, Rudolf VHW (2010) Phenology, ontogeny and the effects of climate change on the tim-
ing of species interactions. Ecol Lett 13:1–10
References