lateral hyporheic exchange throughout the mississippi river network
TRANSCRIPT

LETTERSPUBLISHED ONLINE: 11 MAY 2014 | DOI: 10.1038/NGEO2157
Lateral hyporheic exchange throughout theMississippi River networkBrian A. Kiel1,2 and M. Bayani Cardenas1*River water circulates vertically through the river bed andlaterally through bank sediments1–4. This hyporheic exchangebrings river water into contact with microbes in the adjacentsediments, otherwise known as the hyporheic exchangezone. As such, hyporheic zones act as hotspots for thebiogeochemical cycling of carbon, metals and nutrients inrivers1,2,5–8, and can, for example, influence the export ofnitrogen todownstreamecosystems5,9,10.Here,wecalculate theextent and duration of lateral hyporheic exchange throughouttheMississippiRivernetwork, usingaphysics-basednumericalmodel that takes into account the distribution of groundwaterbaseflow, river discharge, alluvium permeability and rivermorphology3,4.Wefindthat inoursimulations, lateralexchangeoccurs throughout the network: practically all of the riverwater that reaches the mouth of theMississippi River networkhas circulated through the lateral hyporheic zone. River waterresidence time in the hyporheic zone ranges from less thanan hour in headwaters to over a month in larger channels.Comparison of the residence times that we derive to apreviously reportedresidence timethreshold fordenitrificationsuggests that aroundonequarter of the lateral hyporheic zonesin theMississippiRivernetwork favourdenitrification, and thusnitrogen loss. Given that river water can circulate many timesthrough the river bank before reaching the rivermouth, and canundergo vertical exchange with the river bed, we suggest thatour estimates serve as a lower limit for potential nitrogen loss.
River morphology forces water to circulate through river bedand bank alluvium1–4. This process, known as hyporheic exchange,occurs vertically and laterally and affects downstream nitrogenexport5,9,10 as hyporheic zones are reaction hotspots that act asbuffers1,2,5–8. By proportion, larger nitrogen inputs are buffered byhyporheic zones in smaller streams, but larger rivers buffer moremass7,9,10. Here, we calculated lateral hyporheic exchange every tenchannel widths using a physics-based model3,4 that considered dis-tributed groundwater baseflow, river discharge, alluviumpermeabil-ity and river morphology throughout theMississippi River network.The analyses revealed that lateral exchange occurs throughout thenetwork. Practically all of the river discharge reaching the net-work outlet had circulated through the lateral hyporheic zone. Themean length needed for 50% exchange is 1.64 km. Comparison ofhyporheic residence times to previously reported denitrificationtimescales show that ∼1/4 of lateral hyporheic zones favour deni-trification, with the rest favouring nitrification; a small portion oflateral hyporheic zones are net nitrogen sinks. Thus, when con-sidered independently, lateral hyporheic zones may have a lim-ited capacity to buffer nitrogen loading. However, considerationfor vertical hyporheic exchange and that nitrogen routed downthe network is serially exchanged would increase the collectivenetwork-scale buffering of nitrogen.
Rivers and their adjacent alluvial beds and banks are hydrauli-cally interconnected through a mosaic of shallow groundwater flowpaths beginning and ending at rivers. Referred to as hyporheicexchange or flow, these are driven by hydraulic head gradients alongthe river-alluvium boundary. The planform shape of rivers producesgradients that lead to lateral exchange of water between rivers andtheir banks3,4. This lateral exchange, along with vertical exchangewith the river bed,is largely responsible for bringing river water incontact withmicrobes in the sediment, which in turn process water-borne carbon, metals and nutrients1,2,5,7,8. This exchange servesmyriad critical natural functions which collectively affect channel-to network-scale solute dynamics and budgets9–13.
Nutrient export from the Mississippi River network (MRN) isresponsible for the dead zone in the Gulf ofMexico14. Managing thisenvironmental impact necessitates an understanding of network-scale solute transformation processes, which requires integrationof local-scale processes. Previous investigations showed that inproportion to inputs, the processing of nitrogen and carbon arelarger in lower order (first–fourth Strahler order) streams7,9,15–18,but by absolute amounts, higher order channels (>fifth order)account for more mass of solutes transformed9,11,15. Previous modelswere developed from the perspective of the river, with uptake ratesdetermined through regressions with surface residence time ordischarge15,17. The underlying principle for regression-basedmodelsis that the reactions are lumped under some sufficient effectivesurface residence time which allows for reaction completion. Arecent comprehensive study7, the LINXII project, conducted acrossbiomes and scales quantified nitrate uptake rates using instreamisotopic tracer injections and related these to instream nitrateconcentration. The uptake models were then implemented in afifth-order network model which further illustrated and verifiedthe relative roles of lower and higher order channels on nitrogenbuffering. Thus, previous network-scale models represented local-scale hyporheic exchange processes through some abstraction thatdid not explicitly represent the physics of hyporheic exchange.
The goal of this study was to use physically based models3,4to quantify lateral hyporheic zone (LHZ) processes throughoutthe MRN to determine their extent, magnitude and potentialbiogeochemical impacts. We quantified local LHZ fluxes, areasand residence times using measured or estimated channel width,depth, sinuosity and valley slope; estimated permeability of the bankalluvium; and groundwater baseflow. The spatially distributed LHZmetrics were then integrated into network-scale indices. Themodelsassume that lateral exchange occurs because river water infiltratesinto the LHZ at the upstream half of a channel meander andreturns to the river along its downstream half. Thus, the lateralhyporheic exchange that is considered here is naturally forced by thelongitudinal hydraulic head gradient along the river water surfaceimposed on a sinuous channel.
1Geological Sciences, The University of Texas at Austin, Austin, Texas 78712, USA, 2Bureau of Economic Geology, The University of Texas at Austin, Austin,Texas 78712, USA. *e-mail: [email protected]
NATURE GEOSCIENCE | VOL 7 | JUNE 2014 | www.nature.com/naturegeoscience 413
© 2014 Macmillan Publishers Limited. All rights reserved.

LETTERS NATURE GEOSCIENCE DOI: 10.1038/NGEO2157
Channel geometric properties were taken from US Geologi-cal Survey (USGS) National Hydrography Dataset (NHD, http://nhd.usgs.gov/); high-resolution NHD data for channels classifiedas perennially flowing were used. Sinuosity and channel widthwere measured directly from the NHD data set. Channel widthwas calculated from NHD river polygons; width was calculatedevery 1.5 km along the entire network, except within reservoirs,which were filtered out. Sinuosity was calculated for every seg-ment greater than ten bankfull widths, following previous stud-ies19. To assign sinuosity for smaller channels whose widths werenot measurable, sinuosity was interpolated across the watershedbased on channels with calculated sinuosity. Valley slopes werederived through regression using information from the USGSHydrologic Land Regions (HLR) data set (http://water.usgs.gov/lookup/getspatial?hlrus) and the US Environmental ProtectionAgency’s (EPA) field measurements of water surface slope andlong-reach sinuosity20. Sediment permeability was estimated us-ing empirical equations21,22 based on median grain diameter (D50)
compiled from available USEPA data sets23,24. Groundwater base-flow discharge was calculated using the baseflow index from theUSGS (http://water.usgs.gov/lookup/getspatial?bfi48grd) and mul-tiplying this by the mean river discharge taken from the USGSNational Water Information System (NWIS, http://waterdata.usgs.gov/). The analyses were conducted for three sub-watersheds ofthe MRN: the Ohio River, the Upper Mississippi River and theMissouri River. Data were sometimes analysed separately by wa-tershed, whereas other calculations were done collectively for theentire network.
We calculated LHZ metrics for ∼500,000 km of river channels.The calculations for LHZ metrics were for each elementary lengthunit (L) which we defined as half the channel meander arc length,which in turn was calculated from width and sinuosity. For eachL, we calculated the ratio of volumetric lateral hyporheic discharge(QE) to river discharge (QR):
F=QE/QR
The probability of a given water parcel traversing a length unit whilenot being exchanged is:
PNE=(1−F)
This assumes that the river is well-mixed within each segment.The probability of a water molecule not being exchanged starting
from a given segment and until it reaches some downstream point(PN) is thus the product of the PNE values of all the segmentstraversed. From this, the length it takes for PN to be belowsome threshold can be calculated. We calculated the lengths atwhich PN≤0.5 (L50) and PN≤0.1 (L90), which corresponds tothe length it takes for 50% and 90% of the river discharge tohave been recirculated through the LHZ. Finally, the fractionof all water reaching some chosen outlet within the networkthat had been exchanged with the LHZ is the incrementaldischarge-weighted average:
FO=1−(∑
PNQRi
)/(∑QRi
)with PN calculated for each segment and terminating at the outlet,and where QRi is the incremental river discharge for the samesegment such that the total of all QRi is the river discharge at theoutlet QO, that is, QO≈
∑QRi.
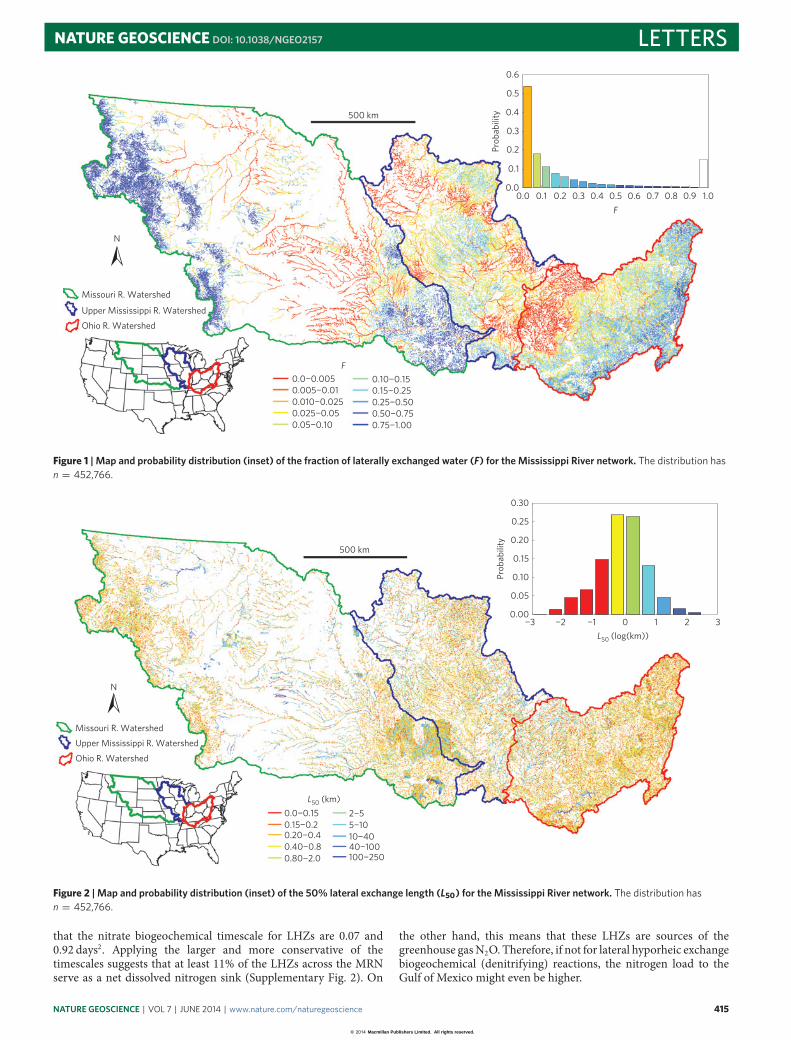
The fraction of river water that is exchanged at each reach (F)generally increases from the headwaters going downstream (Fig. 1and Supplementary Fig. 1) and follows an exponential distributionwith a mean of 0.147 and standard deviation of 0.218. But althoughthe majority of reaches have F < 0.15, many smaller rivers have
F > 0.75. A previous study showed that local exchange turnover(analogous to F) at river segments that are hundreds of metreslong, with discharge ranging from 1–100 l s−1, reaches as high as40% (ref. 13), which is consistent with our results. (Note that Fig. 1includes a relatively large number of values for F in the range 0.9–1.0as some points that had F slightly larger than 1.0 were all lumpedinto the above category). F is related toQR (inm3 s−1) via (R2
= 0.97):
F(QR)=0.0673Q−0.447R
Lateral hyporheic exchange persists across the network; at thescale of our calculations, no area had F=0.0. We found FO=0.996at the most downstream outlet of the portion of the MRN weconsidered—the confluence of the Ohio and Upper MississippiRivers. Practically all of the water that flows from the MRNhas cycled through the LHZ during the course of its journeydownstream. The mean and median L50 are 1.64 and 0.67 km(Fig. 2), respectively, and the mean and median for L90 are 2.77and 1.58 km. The exchange lengths are generally shorter near theheadwaters, because F is generally larger there.
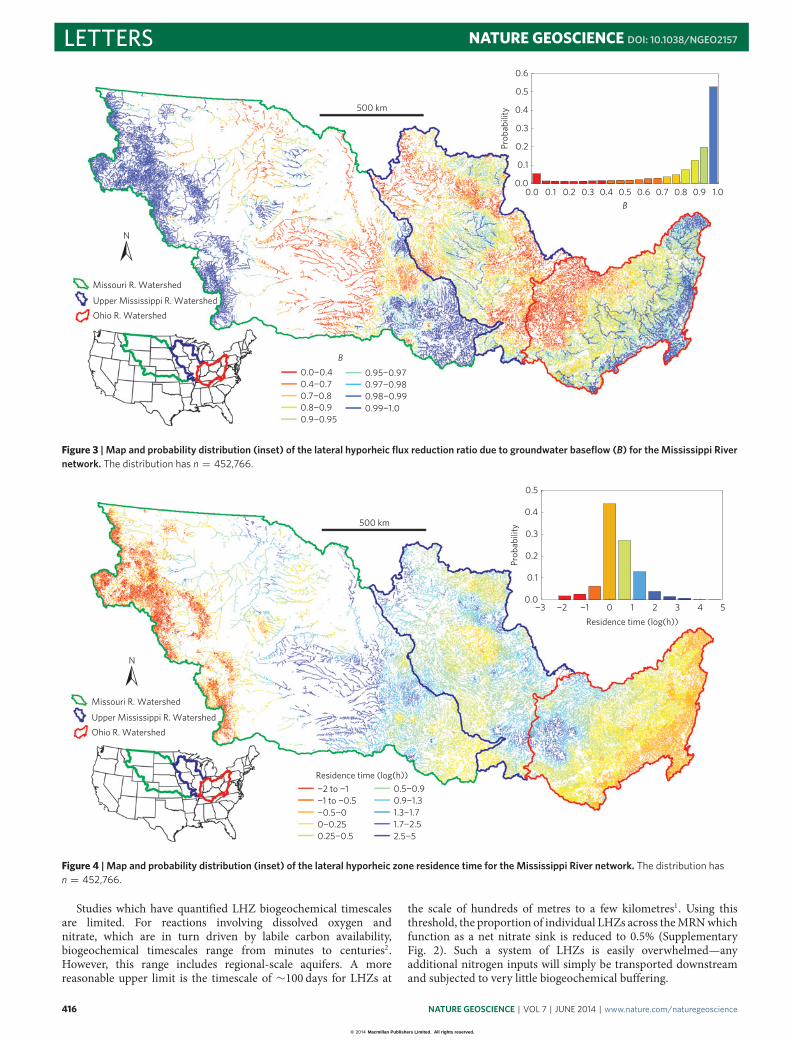
Groundwater baseflow dictates the extent of lateral hyporheicexchange because groundwater flow towards a river counters lateralhyporheic flow. Our calculation of the proportion of the hyporheicflux calculated when baseflow is taken into account to the fluxthat would occur without baseflow (B=QE/QE,Max) shows that thisspans the entire possible range from 0, where lateral exchange iscompletely inhibited by baseflow, to 1, where there is no effect frombaseflow (Fig. 3 and Supplementary Fig. 1). B follows a negativeexponential distribution, with most values closer to 1.0; its meanis 0.88 with a standard deviation of 0.24. Thus, baseflow has ameasurable effect on network-scale exchange. This also indicatesa susceptibility of LHZ processes to any modification of regionalgroundwater flows and budgets, which have been markedly alteredas a result of groundwater extraction25.
Lateral hyporheic exchange does not affect net fluid massbalance, but it has substantial effects on reactive solute budgets.The balance between the rate at which reactants are deliveredto and the amount of time they spend within the reactionzones determine biogeochemical reaction rates26. Thus, the fluidresidence time through the LHZ is a critical parameter1,2,26–28.The LHZ residence times we calculated ranged from less thanan hour, in the headwaters, to more than a month in thelarger meandering channels (Fig. 4). Aerobic conditions generallyprevail when the ratio of residence or transport time to reaction(oxygen consumption) time is small, that is, less than 1, andnitrification becomes dominant. Inversely, when this ratio is large,anaerobic conditions which favour denitrification prevail. Wethus compared the exchange residence times we calculated to apreviously determined threshold time of 6.9 h in the LHZ of athird-order stream27 belowwhich nitrification is favoured and abovewhich denitrification is dominant (see Supplementary Informationfor further discussion of the timescale thresholds). Based on thisthreshold, 24.2% of the LHZs favour denitrification whereas the restfavour nitrification (Fig. 4 and Supplementary Fig. 2). Thus, theindividual LHZs across the MRN independently have limitations intheir capacity to buffer nitrate, and may be producing it instead.
The timescale for reactions can vary over several orders ofmagnitude2. Despite this, the biogeochemical timescale, the time atwhich a reactive solute originating from the stream and enteringthe LHZ has undergone an exponential-fold drop relative to itsoriginal concentration when it entered the LHZ, is a useful metric1.This better characterizes the relative role of the LHZ as a netnitrogen sink, because LHZ flow paths typically transition fromaerobic to anaerobic; the terminal processes along flow paths tendto be reducing, for example, denitrification. Application of thebiogeochemical timescale concept to two field studies26,29 showed
414 NATURE GEOSCIENCE | VOL 7 | JUNE 2014 | www.nature.com/naturegeoscience
© 2014 Macmillan Publishers Limited. All rights reserved.
NATURE GEOSCIENCE DOI: 10.1038/NGEO2157 LETTERS
500 km
Missouri R. Watershed
Upper Mississippi R. Watershed
Ohio R. Watershed
0.0−0.0050.005−0.010.010−0.0250.025−0.050.05−0.10
0.10−0.150.15−0.250.25−0.500.50−0.750.75−1.00
F
N
0.0 0.1 0.2 0.3 0.4 0.5 0.6 0.7 0.8 0.9 1.00.0
0.1
0.2
0.3
0.4
0.5
0.6
F
Prob
abili
ty
Figure 1 | Map and probability distribution (inset) of the fraction of laterally exchanged water (F) for the Mississippi River network. The distribution hasn = 452,766.
0.0−0.150.15−0.20.20−0.40.40−0.80.80−2.0
2−55−1010−4040−100100−250
L50 (km)
Missouri R. Watershed
N
Upper Mississippi R. Watershed
Ohio R. Watershed
500 km
−3 −2 −1 0 1 2 30.00
0.05
0.10
0.15
0.20
0.25
0.30
L50 (log(km))
Prob
abili
ty
Figure 2 | Map and probability distribution (inset) of the 50% lateral exchange length (L50) for the Mississippi River network. The distribution hasn = 452,766.
that the nitrate biogeochemical timescale for LHZs are 0.07 and0.92 days2. Applying the larger and more conservative of thetimescales suggests that at least 11% of the LHZs across the MRNserve as a net dissolved nitrogen sink (Supplementary Fig. 2). On
the other hand, this means that these LHZs are sources of thegreenhouse gasN2O.Therefore, if not for lateral hyporheic exchangebiogeochemical (denitrifying) reactions, the nitrogen load to theGulf of Mexico might even be higher.
NATURE GEOSCIENCE | VOL 7 | JUNE 2014 | www.nature.com/naturegeoscience 415
© 2014 Macmillan Publishers Limited. All rights reserved.
LETTERS NATURE GEOSCIENCE DOI: 10.1038/NGEO2157
0.0−0.40.4−0.70.7−0.80.8−0.90.9−0.95
0.95−0.970.97−0.980.98−0.990.99−1.0
B
Missouri R. Watershed
N
Upper Mississippi R. Watershed Ohio R. Watershed
500 km
0.0 0.1 0.2 0.3 0.4 0.5 0.6 0.7 0.8 0.9 1.00.0
0.1
0.2
0.3
0.4
0.5
0.6
B
Prob
abili
ty
Figure 3 | Map and probability distribution (inset) of the lateral hyporheic flux reduction ratio due to groundwater baseflow (B) for the Mississippi Rivernetwork. The distribution has n = 452,766.
−3 −2 −1 0 1 2 3 4 50.0
0.1
0.2
0.3
0.4
0.5
Prob
abili
ty
−2 to −1−1 to −0.5−0.5−00−0.250.25−0.5
0.5−0.90.9−1.31.3−1.71.7−2.52.5−5
Residence time (log(h))
Missouri R. Watershed
N
Upper Mississippi R. Watershed Ohio R. Watershed
500 km
Residence time (log(h))
Figure 4 | Map and probability distribution (inset) of the lateral hyporheic zone residence time for the Mississippi River network. The distribution hasn = 452,766.
Studies which have quantified LHZ biogeochemical timescalesare limited. For reactions involving dissolved oxygen andnitrate, which are in turn driven by labile carbon availability,biogeochemical timescales range from minutes to centuries2.However, this range includes regional-scale aquifers. A morereasonable upper limit is the timescale of ∼100 days for LHZs at
the scale of hundreds of metres to a few kilometres1. Using thisthreshold, the proportion of individual LHZs across theMRNwhichfunction as a net nitrate sink is reduced to 0.5% (SupplementaryFig. 2). Such a system of LHZs is easily overwhelmed—anyadditional nitrogen inputs will simply be transported downstreamand subjected to very little biogeochemical buffering.
416 NATURE GEOSCIENCE | VOL 7 | JUNE 2014 | www.nature.com/naturegeoscience
© 2014 Macmillan Publishers Limited. All rights reserved.

NATURE GEOSCIENCE DOI: 10.1038/NGEO2157 LETTERSOur analysis and comparison of residence times as a metric for
dissolved nitrogen buffering potential is for individual and indepen-dent LHZs. A nitrogen (and water) molecule would be circulatedmany times through the LHZs as it travels down the network,particularly in light of the relatively small values for L50; it wouldbe serially processed by the LHZs. Moreover, our analysis did notconsider vertical hyporheic exchange which occurs at smaller andshorter timescales and whose reaction thresholds may be different.Vertical exchange provides further opportunities for denitrifica-tion. Future efforts should be directed at integrating these aspectsof exchange—multi-dimensionality, temporal dynamics and serialprocessing. Our current estimates for nitrogen buffering capacitysets the lower limit. As almost all of the water flowing down thenetwork flows through the hyporheic zone at some point, hyporheicexchange influences the biogeochemical cycling of nitrogen andtherefore the amount delivered ultimately to the Gulf of Mexico.
Received 16 September 2013; accepted 4 April 2014;published online 11 May 2014
References1. Boano, F., Demaria, A., Revelli, R. & Ridolfi, L. Biogeochemical zonation due to
intrameander hyporheic flow.Wat. Resour. Res. 46,W02511 (2010).2. Gomez, J. D., Wilson, J. L. & Cardenas, M. B. Residence time distributions in
sinuosity-driven hyporheic zones and their biogeochemical effects.Wat.Resour. Res. 48,W09533 (2012).
3. Cardenas, M. A model for lateral hyporheic flow based on valley slope andchannel sinuosity.Wat. Resour. Res. 45,W01501 (2009).
4. Cardenas, M. Stream-aquifer interactions and hyporheic exchange in gainingand losing sinuous streams.Wat. Resour. Res. 45,W06429 (2009).
5. Battin, T. J. et al. Biophysical controls on organic carbon fluxes in fluvialnetworks. Nature Geosci. 1, 95–100 (2008).
6. Boano, F., Camporeale, C., Revelli, R. & Ridolfi, L. Sinuosity-driven hyporheicexchange in meandering rivers. Geophys. Res. Lett. 33, L18406 (2006).
7. Mulholland, P. J. et al. Stream denitrification across biomes and its response toanthropogenic nitrate loading. Nature 452, 202–246 (2008).
8. Fischer, H., Kloep, F., Wilzcek, S. & Pusch, M. T. A river’s liver—microbialprocesses within the hyporheic zone of a large lowland river. Biogeochemistry76, 349–371 (2005).
9. Stewart, R. J. et al. Separation of river network-scale nitrogen removal amongthe main channel and two transient storage compartments.Wat. Resour. Res.47,W00J10 (2011).
10. Ye, S. et al. Dissolved nutrient retention dynamics in river networks: Amodeling investigation of transient flows and scale effects.Wat. Resour. Res. 48,W00J17 (2012).
11. Ensign, S. H. & Doyle, M. W. Nutrient spiraling in streams and river networks.J. Geophys. Res. 111, G04009 (2006).
12. Fisher, S. G., Grimm, N. B., Marti, E., Holmes, R. M. & Jones, J. B. Materialspiraling in stream corridors: A telescoping ecosystem model. Ecosystems 1,19–34 (1998).
13. Covino, T., McGlynn, B. & Mallard, J. Stream-groundwater exchange andhydrologic turnover at the network scale.Wat. Resour. Res. 47,W12521 (2011).
14. Turner, R. E. & Rabalais, N. N. Coastal eutrophication near the Mississippiriver delta. Nature 368, 619–621 (1994).
15. Seitzinger, S. P. et al. Nitrogen retention in rivers: model development andapplication to watersheds in the northeastern USA. Biogeochemistry 57,199–237 (2002).
16. Wollheim, W. M., Voosmarty, C. J., Peterson, B. J., Seitzinger, S. P. &Hopkinson, C. S. Relationship between river size and nutrient removal.Geophys. Res. Lett. 33, L06410 (2006).
17. Alexander, R. B., Smith, R. A. & Schwarz, G. E. Effect of stream channelsize on the delivery of nitrogen to the Gulf of Mexico. Nature 403,758–761 (2000).
18. Peterson, B. J. et al. Control of nitrogen export from watersheds by headwaterstreams. Science 292, 86–90 (2001).
19. Soar, P. J. & Thorne, C. R. Channel Restoration Design for Meandering Rivers.Report No. ERDC/CHL-CR-01-1, 454 (US Army Corps of Engineers, 2001).
20. Kaufmann, P. R., Levine, P., Robison, E. G., Seeliger, C. & Peck, D. V.Quantifying Physical Habitat in Wadeable Streams. Report No.EPA/620/R-99/003, 149 (US Environmental Protection Agency, 1999).
21. Shepherd, R. G. Correlations of permeability and grain-size. Ground Wat. 27,633–638 (1989).
22. Lu, C. P., Chen, X. H., Cheng, C., Ou, G. X. & Shu, L. C. Horizontal hydraulicconductivity of shallow streambed sediments and comparison with thegrain-size analysis results. Hydrol. Proc. 26, 454–466 (2012).
23. Paulsen, S. G. et al. Condition of stream ecosystems in the US: An overview ofthe first national assessment. J. N. Am. Benthol. Soc. 27, 812–821 (2008).
24. Stoddard, J. L. et al. An Ecological Assessment of Western Streams and Rivers.Report No. 620/R-05/005, (US Environmental Protection Agency, 2005).
25. Wada, Y. et al. Global depletion of groundwater resources. Geophys. Res. Lett.37, L20402 (2010).
26. Zarnetske, J. P., Haggerty, R., Wondzell, S. M., Bokil, V. A. &Gonzalez-Pinzon, R. Coupled transport and reaction kinetics control thenitrate source-sink function of hyporheic zones.Wat. Resour. Res. 48,W11508 (2012).
27. Zarnetske, J. P., Haggerty, R., Wondzell, S. M. & Baker, M. A. Dynamics ofnitrate production and removal as a function of residence time in the hyporheiczone. J. Geophys. Res. 116, G01025 (2011).
28. Gu, C. H., Hornberger, G. M., Mills, A. L., Herman, J. S. & Flewelling, S. A.Nitrate reduction in streambed sediments: Effects of flow and biogeochemicalkinetics.Wat. Resour. Res. 43,W12413 (2007).
29. Pinay, G., O’Keefe, T. C., Edwards, R. T. & Naiman, R. J. Nitrate removalin the hyporheic zone of a salmon river in Alaska. Riv. Res. Appl. 25,367–375 (2009).
AcknowledgementsM.B.C. thanks the NSF (grant EAR-0955750) for support. The Geology Foundation at theUniversity of Texas at Austin provided additional support.
Author contributionsM.B.C. conceived this study with input from B.A.K. The data analysis, compilation andmodelling were performed by B.A.K. The paper was written by M.B.C. with input fromB.A.K., and both authors interpreted the results.
Additional informationSupplementary information is available in the online version of the paper. Reprints andpermissions information is available online at www.nature.com/reprints.Correspondence and requests for materials should be addressed to M.B.C.
Competing financial interestsThe authors declare no competing financial interests.
NATURE GEOSCIENCE | VOL 7 | JUNE 2014 | www.nature.com/naturegeoscience 417
© 2014 Macmillan Publishers Limited. All rights reserved.