lay, rupert - das leben - wesen und werden
TRANSCRIPT

DER CHRIST IN DER WELT
EINE ENZYKLOPÄDIE
In verantwortlicher Zusammenarbeit mit deutschen und französischen Autoren
unter beratender Mitwirkung von P. Johannes Hirschmann S J
II. Reihe
DIE WELT, IN DER WIR LEBEN
Band 4
PAUL PATTLOCH VERLAG • ASCHAFFENBURG

DAS LEBEN
WESEN UND WERDEN
von
RUPERT LAY
PAUL PATTLOCH VERLAG ASCHAFFENBURG

Rupert Lay, geboren am 14. Juni 1929; Mitglied des Jesuitenordens; Studium der theoretischen Physik und Philosophie; Promotion in Bonn; seit 1965 Dozent für Naturphilosophie und Wissenschaftstheorie. Ab 1. 4. 1967 ordentlicher Professor für Philosophie an der Philosophisch-Theologischen Hochschule St. Georgen in Frankfurt/Main. Veröffentlichungen: Arbeiten zur Naturphilosophie und Transzendentalontologie.
1969 © by Paul Pattloch Verlag Aschaffenburg 1969
Umschlagentwurf: Willy Kretzer Gesamtherstellung: Walter Pieper, Würzburg

EINFÜHRUNG
Das Geheimnis des Lebens ist sicher eines der erregendsten Geheimnisse, denen der Menschengeist sich je gegenübersah. «Leben ist für jeden denkenden Menschen ein so verständlicher und zugleich doch rätselhafter Begriff. Sein Sinn, so hätte es den Anschein, müßte für alle Zeiten und Völker klar und eindeutig sein» (A. I. Oparin, Das Leben, Jena 1963, 1). Aber es scheint nur so. An kaum einem anderen Begriff entzweite sich das philosophische Denken der Menschheit so sehr, wie gerade an dem des Lebens. Was ist ‹Leben›? Was bedeutet Leben? Wie ist es geworden? Fragen über Fragen. Dabei scheint eine Antwort gar nicht so schwer: erfahren wir Menschen doch in uns selbst, was Leben ist, was Leben bedeutet. Konnte man von hierher nicht eine Bestimmung dessen, was Leben meint, versuchen? Nun, solche Versuche gibt es, aber sie haben sich als untauglich erwiesen. Der legitime Ansatz für die Beantwortung unserer durchaus philosophischen Fragen wird von der Biologie, der Lehre von den Erscheinungen des Lebendigen, ausgehen müssen. Erst in einem Weiterfragen, erst im Deuten, dem Fragen nach der Bedeutung biologischer Erscheinung, werden wir den Geheimnissen des Lebendigen und damit auch denen des Lebens auf die Spur kommen können. Diese für den modernen Leser fast trivale Feststellung ist jedoch keineswegs so selbstverständlich, wie es auf den ersten Blick scheinen möchte. Am Anfang des abendländischen Mühens um die Probleme des Lebens stehen durchaus dogmatische Theorien über das Leben: Theorien, die nicht ausgehen von der ganz unbefangenen Musterung der Erscheinungen der ‹Natur›. Eine solche Durchmusterung führt zu der zunächst ganz unreflexen Erkenntnis, daß es doch offenbar eine Vielfalt von ‹Ordnungen›, von Strukturen gebe in dieser ‹Natur›: ‹tote› Materie, Pflanzen, Tiere, Menschen, denen spezifische Funktionsweisen zugeordnet sind. Zu den dogmatischen Lebenstheorien möchten wir alle Monismen zählen, Theorien also, die einen dieser Erscheinungskomplexe universalisieren: Alles ist belebt, alles ist durchgeistet, alles ist materiell. Solche Monismen sind keineswegs Geschichte, abwegige Geschichte des menschlichen Denkens. Man kann ihnen auch heute noch begegnen. Sie werden - auch im Zeitalter der modernen Naturwissenschaften — immer dann auftauchen, wenn eine für einen der genannten Erscheinungskomplexe gesicherte wissenschaftliche Methode, auf Grund deren wir zu 5 bestimmten ‹Naturgesetzen› gelangten und gelangen, derart verallgemeinert wird, daß man sie auch auf die jeweils anderen Komplexe anwendet. Ein Physiker wird, wenn er mit seiner Methode an die Erscheinungen des Lebendigen herangeht, nichts anderes entdecken als physikalische Gesetze. Modifizierte physikalische Gesetze vielleicht - aber eben doch Gesetze, die ebenso - wenigstens grundsätzlich - auch im Nichtbelebten gelten können. Das ist sicher eine wissenschaftstheoretisch erlaubte Methode, solange der Physiker nicht behauptet, etwas über das Lebewesen als Lebewesen auszusagen. Er sagt - möglicherweise etwas durchaus Richtiges - über das Lebewesen als auch materielle Erscheinung. Die Tendenz zum wissenschaftstheoretischen äußerst bedenklichen Verabsolutieren einer Methodik — bedenklich, weil die Methode dem Objekt anzupassen ist und nicht das Objekt der Methode - liegt in dem nur allzu menschlichen Suchen nach Einheit begründet. Das ist heute nicht sehr viel anders als vor 2500 Jahren. So darf es uns nicht wundern, daß die ersten Theorien über die Natur des Lebendigen durchaus monistisch waren.
1. Der Hylezoismus (alles ist belebt) ARISTOTELES berichtet: Die meisten unter den älteren Denkern wollten überall nur materielle Urgründe annehmen, denn woraus alle Gegenstände bestehen, woraus sie sich entwickeln und worin sie sich endlich wieder auflösen . . . das ist nach ihrer Auffassung der . . . Urgrund aller Gegenstände (Mph. I 3; 983 b 7ff.). THALES, der Begründer dieser Richtung, erklärte das Wasser zum Urgrund . . . Vielleicht kam er zu dieser Auffassung durch die Beobachtung, daß die Nahrung von allem feucht ist, und daß das Warme aus dem Wasser komme und von ihm lebe (ibd. 2o ff.). «Einige lassen die Seele in das All verwoben sein. So scheint auch Thales zu seinem Ausspruch gekommen sein, es sei alles voller Götter» (de anima I, 5; 411 a 7 f.). ANAXIMENES lehrte nach dem Zeugnis des Aristoteles die Luft als Urgrund, HERAKLIT das Feuer (Mph. I, 3; 9843 5 ff.). Daß die älteren jonischen Naturphilosophen ihren Urgrund nicht materialistisch, sondern durchaus durchlebt verstanden, ist zumindest für Anaximenes und Heraklit nicht zu bestreiten. Erneuert wurde der Hylezoismus in der Philosophie der Renaissance. Parazelsus, Cardanus, später G Bruno, P. Gassendi, B. Spinoza, H. More, Goethe (WW XXV, 132 f.) waren Hylezoisten. KANT versuchte eine apriorische Widerlegung des Hylezoismus: «... die Möglichkeit einer lebenden Materie läßt sich nicht einmal den- 6 ken; die einer belebten Materie . . . kann nur sofern dürftigerweise gebraucht werden, als sie uns an der Organisation derselben im Kleinen in der Erfahrung offenbart wird, keineswegs aber a priori ihrer Möglichkeit nach eingesehen werden» (KU § 73). Dennoch wurden auch nach Kant noch manche hylezoistischen Lebenstheorien vertreten (Schule Schellings, Lotze, Fechner, E. von Hartmann, . . .). Dem Hylezoismus verwandt und nicht immer scharf von ihm abzugrenzen ist der

2. Panpsychismus (alles ist beseelt) Sicherlich könnte man auch die frühen Jonier zu den Panpsychisten zählen. Nach PLATON schuf der Demiourgos die Welt als ein beseeltes und vernunftbegabtes Wesen, als eine selige Gottheit (Timaios, 346). ARISTOTELES schreibt: «Es entstehen in der Erde und im Wasser Tiere und Pflanzen, weil in der Erde Feuchtigkeit, im Wasser Lebensodem und überall seelische Wärme ist. So ist im gewissen Sinn alles voller Seele . . .» (de generat, anim. III, 11; 62 a). Doch auch in der philosophischen Neuzeit wurde der Panpsychismus vertreten: SPINOZA hält die Dinge aller «Grade» für beseelt (animata) (Ethik, II, prop. 13 schol.). Ganz ähnlich LEIBNIZ (Monadol. 68 f.). Auch SCHELLING behauptet: «Alles im Universum ist beseelt» (WW I, 6; 217) und E. HAECKEL erklärt: «Jedes Atom besitzt eine inhärente Summe von Kraft und ist in diesem Sinne ‹beseelt› . . .» Lust und Unlust, Begierde und Abneigung, Anziehung und Abstoßung müssen allen Massen-Atomen gemeinsam sein! (Die Perigenese der Plastidulen, 38). Die Plastidulen (= belebte Atomkomplexe) haben ein unbewußtes Gedächtnis. Masse und Äther besitzen Empfindung und Willen (Welträtsel. 254). Nach A. RIEHL ist der Panpsychismus «eine reine Spekulation, für welche die psychophvsischen Tatsachen keine Handhabe bieten. - Der Dichter mag die Dinge ringsum beseelen; als Denker aber sollten wir aufhören, von einem Lieben und Hassen der Elemente und von Atomempfindungen zu träumen» (Zur Einführung in die Philosophie der Gegenwart, 1903, 161 f.). Monismus ist auch der
3. Materialismus (alles ist Materie) Der Materialismus hat im Laufe der Geschichte sehr verschiedene Formen angenommen. Geeignet unterscheiden wir den mechanistischen Materialismus vom psychologischen Materialismus (nur diese beiden 7 Formen interessieren uns hier zunächst). Der mechanistische Materialismus gründet im Mechanismus, einer Theorie, die alle physikalischen Prozesse auf mechanische zu reduzieren suchte. Eine Extrapolation des Mechanismus auf alle Bereiche führt konsequent zum mechanistischen Materialismus. Mit der Aufgabe des Mechanismus durch die moderne Physik hat auch der mechanistische Materialismus seine Grundlage verloren und wird heute nur noch in popularisierten Formen des Materialismus gefunden. Der psychische Materialismus begegnet uns in sehr verschiedenen Formen: Geist (und Analoges gilt auch für eventuelle Prinzipien des Lebendigen) ist eine bestimmte «Art» der Materie; Geist ist eine «Ausscheidung», ein Produkt der Materie; Geist ist Funktion der Materie; Geist ist einer der Zustände der Materie. Der Hylezoismus der Jonier ist ein «organischer» Materialismus. Der Atomismus der Antike [R. Lay, Welt des Stoffes (abgekürzt: WS) I, Aschaffenburg 1965, 20] ist ein mechanistischer Materialismus. Ähnliches gilt nach dem Zeugnis des DIOGENES LAERTIOS auch für die Epikureer. Von den christlichen Philosophen hielt vor allem TERTULLIAN die Seele für materiell (PL II, 697 B; 186 A; 819 B). Unter dem Einfluß der Philosophie des Descartes blühte der Materialismus aller Formen während des 18. und 19. Jahrhunderts auf. J.O.DE LAMETTRIE (1709-1751) hält den Menschen für eine «Maschine, die sich selbst aufzieht» (L'homme 1875, 25). Die Seele ist nichts als ein Teil des Gehirns (ibd. 66). Denken ist eine Eigenschaft der Materie, deren Wesen aber unbekannt ist (ibd. 74). Für L. FEUERBACH ist alles Wirkliche körperlich (WW II, 321). C. VOGT erklärt, daß «die Gedanken etwa in demselben Verhältnis zum Gehirn stehen, wie die Galle zur Leber oder der Urin zu den Nieren» (Köhlerglaube und Wissenschaft, 1854). Eine besondere Ausprägung fand der Materialismus im DIAMAT. Die für unsere Problematik sauberste Formulierung fand er in der kleinen Schrift J. Stalins, «Über historischen und dialektischen Materialismus», die weitgehend kaum etwas anderes ist als ein kurzes Resume der Hauptthesen von Marx und Engels. Ganz zu Beginn heißt es: «Der dialektische Materialismus ist die Weltanschauung der marxistisch-leninistischen Partei. Diese Weltanschauung heißt darum dialektischer Materialismus, weil ihr Herangehen an die Naturerscheinungen die dialektische ist und weil ihre Deutung der Naturerscheinungen, ihre Theorie, materialistisch ist.» Zur Dialektik schreibt er: Die Natur ist ein Ganzes; die Einzeldinge sind organisch miteinander verbunden und müssen aus diesen Beziehungen heraus verstanden werden. Alles befindet sich in unaufhörlicher Bewegung .. . 8 Diese Bewegung schreitet gradlinig fort und ist ein Aufstieg vom Einfachen zum Komplizierten, vom Niedern zum Höheren; was die Entwicklung vorwärtstreibt, sind die Gegensätze, die inneren Widersprüche der Naturerscheinungen. Zum Sinn des Materialismus meint er: Die Wirklichkeit ist nicht Idee oder Bewußtsein, bedarf auch keines Weltgeistes, sondern ist ausschließlich Materie, die sich nach ihren eigenen, durch die dialektische Methode festzustellenden Gesetzen bewegt; Sein ist gleich Materie; Materie ist darum immer das Primäre, Quelle der Empfindungen, der Vorstellungen, des Bewußtseins. «Denken ist ein Produkt der Materie, die in ihrer Entwicklung einen hohen Grad von Vollkommenheit erreicht hat, und zwar ein Produkt des Gehirns.» -«Denken ist von Materie nicht zu trennen.» Solche monistischen Theorien sind - wie gesagt - samt und sonders dogmatischer Art: d. h. sie gehen mit einem Wissensapriori an die Erscheinung heran, das den eigentlichen (philosophischen) Erscheinungshintergrund grundsätzlich vorwissenschaftlich fixiert. Das ist schlechte Philosophie. Gute Philosophie basiert auf einer möglichst

breiten empirischen Basis und fragt dann den gesamten empirischen Befund nach seinen philosophischen Voraussetzungen, Implikationen und Konsequenzen. Wenn wir also wissen wollen, was denn Leben eigentlich sei, müssen wir zunächst eine möglichst breite empirische Basis absichern, zunächst einmal einen möglichst einwandfreien Katalog von Eigenschaften des Lebendigen erstellen und uns dann nach den Bedingungen der Möglichkeit dieser Eigenschaften fragen. Diese Frage verläßt im Regelfall den einzelwissenschaftlichen Raum und stößt in das Gebiet der Philosophie vor: so ist denn die Antwort auf die Frage, was denn Leben eigentlich sei, durchaus philosophischer Art. Die Biologie sucht und findet als empirische Wissenschaft mehr oder weniger gut gesicherte ‹Gesetze›, ohne aber angeben zu können, warum gerade diese Gesetze und keine anderen für den Bereich ihrer Beobachtungen gelten. 9
ERSTER TEIL
DAS WESEN DES LEBENDIGEN
10 11 12 ERSTER ABSCHNITT
EMPIRISCHE DATEN ZUR ‹NATUR› DES LEBENDIGEN
Mit der Frage nach dem Wesen des Lebendigen haben wir uns ein recht großartiges Thema gestellt. Wir wissen noch gar nicht so recht, ob uns eine überzeugende Antwort gelingen möchte. Einiges wissen wir immerhin: welchen Weg wir einschlagen müssen, um zum Ziel zu kommen: es ist der Weg über eine biologische beschreibende ‹Definition› der Erscheinung des Lebendigen, eine Definition, die Momente zusammenfaßt, die dem unbestreitbar Lebendigen zukommen. Beginnen wir also mit einer solchen Zusammenstellung von Eigenschaften, die allem Lebendigen gemeinsam sind. Alle Lebewesen haben bestimmte chemische und ‹biologische› Eigenschaften gemeinsam: wir wollen hier - ohne Anspruch auf Vollständigkeit - drei vorstellen: a) Stoffwechsel, b) Vermehrung, c) Mutabilität der Formen. Dabei läßt es sich nicht vermeiden, daß wir einige chemische Zusammenhänge aufzeichnen, ist doch gerade die neuere Biologie über die Aufhellung des Mechanismus dieser drei typisch biologischen Funktionen von der reinen Beschreibung des makrokosmischen Phänomens zu einer genaueren Bestimmung dessen gelangt, was diesen Phänomen zugrunde liegt, zu dem, was sie eigentlich ausmachen. 13 ERSTES KAPITEL
STOFFWECHSEL
Unter Stoffwechsel versteht man eine funktionale Einheit, die aus Stoffaufnahme aus der Umwelt in den Organismus, dem Umbau in körpereigene Stoffe (Assimilation) und dem Abbau von solchen Stoffen, sowie deren Ausscheidung besteht. Dient die Stoffaufnahme vorwiegend — wie beim heranwachsenden Organismus — dem Aufbau des Lebewesens, spricht man von Baustoffwechsel, dient sie der Gewinnung von Energie für Lebensleistungen, so spricht man von Betriebsstoffwechsel. Alle diese Stoffwechselabläufe beruhen geeinzelt betrachtet auf chemischen und physikalischen Vorgängen, deren kausalanalytische Aufhellung allerdings in vielen Fällen noch nicht geglückt ist. Der Betriebsstoffwechsel ist in der Regel an einen Gasstoffwechsel gebunden. Molekularer Sauerstoff (O2) wird von jeder Zelle aufgenommen und zur (chemisch komplizierten) Zellatmung benutzt, meist wird dabei CO2 freigesetzt. Manche Organismen (die anaeroben) führen jedoch eine anoxydative (Spaltungs-)Atmung durch, welche meist als Gärung geschieht. Häufig ist diese Art der Spaltungsatmung bei einer Reihe von Bakterien (z. B. Bact. amylobacter und andere die Buttersäuregärung auslosende Bakterien) und einigen Pilzen (manche haben auch fakultativ beide Atmungen, etwa der Hefepilz Saccharomyces cerevisiae). Aber auch Tiere, die in sauerstoffarmer Umgebung leben, wie Schlammbewohner (etwa die Amöbe Pelomyxa) oder Darmparasitcn (Darmamöben, Bandwürmer, Spulwürmer) können Glycogen gämngs-artig ohne Sauerstoff zu flüchtigen Fettsäuren und Kohlensäure abbauen und dadurch Energie gewinnen. Der oxydative Abbau von Kohlehydraten, Fetten, Aminosäuren geschieht als Kettenring von Reaktionen, dem sogenannten Zitronensäure-Zyklus: dabei wird in mehreren Reaktionen Wasserstoff freigesetzt, dessen allzu schnelles Oxydieren durch eine Reihe von Fermenten verzögert wird. Der freie Wasserstoff wird zunächst von energiereichen Pyridinfermenten aufgenommen und von ihnen an das gelbe Atmungs-ferment (Flavinferment) weitergegeben, das seinerseits auf ein Cytochrom (diese Verbindung enthält ein Ringsystem, wie wir es beim Chlorophyll kennen lernen werden, doch befindet sich in der Mitte statt

14 eines Mg ein Fe Atom) einwirkt. Dabei wird das dreiwertige Eisen zum zweiwertigen reduziert, der Wasserstoff ionisiert (er verliert sein Elektron), der sich mit ebenfalls durch Cytochrome ionisiertem Sauerstoff zu Wasser verbindet. Die dabei freiwerdende Energie wird als chemische Energie gebunden und als ATP (s. S. 17) gespeichert. Die biologische Oxydation besteht also eigentlich nicht in Sauerstoffaufnahme, sondern im Abbau von Wasserstoff. Bei der Beschreibung des Energie- bzw. Betriebsstoffwechsels wollen wir uns auf die Darstellung eines recht einfachen Beispiels beschränken: das der Photosynthese. Diese Beschränkung ist wohl gerechtfertigt, da dieser Stoffwechselvorgang sicher eine der Voraussetzungen für alles höhere Leben auf der Erde ist. Es geht um die Entstehung eines einfachen Zuckers (C6H12O6) aus Wasser und Kohlendioxyd: Nach den Untersuchungen von M. CALVIN (geb. 1911) wird das Kohlendioxyd (CO2) in einen Zucker (Ribulose 1,5 phosphat) eingebaut. Es entsteht eine ihrer Struktur nach noch unbekannte, ziemlich instabile Verbindung (X). Diese zerfällt in zwei Verbindungen mit je drei Koh-lenstoffatomen (Phosphoglycerinsäure). Unter dem Einfluß von Adenosintriphosphat (ATP) und Nicotin-amid-adenin-dinukleotid-phosphat (NADP), das zwei Wasserstoffionen gebunden hat (NADPH+H+) und Abspaltung von Wasser bildet sich Triose 3 phosphat. Zwei dieser Molekel bilden nun unseren einfachen Zucker (Hexose) und einen Rest, der sich unter Einfluß von ATP und NADPH+H+ zu Ribulose 5 phosphat und nochmaligem Einwirken von ATP und NADPH+H+ wieder zu unserem eingangs genannten Zucker (Ribulose 1,5 diphosphat) wandelt. Damit ist der Kreislauf geschlossen: Aus Wasser und Kohlendioxyd ist Hexose geworden. Es bleibt uns nun noch zu erklären, wie ATP und NADPH+H+ entstehen. Ausgangssubstanzen sind Adenosindiphosphat (ADP) und NADP. Unter dem Einfluß von Licht und bei Anwesenheit von Chlorophyll dissoziieren sich zwei Wassermolekel in zwei H-Ionen und zwei OH-Ionen. Die beiden H-Ionen verwandeln das NADP in NADPH+H+ und die beiden OH-Ionen des ADP über mehrere noch unbekannte chemische Zwischenstufen unter Abspaltung eines Wassermolekels und eines Sauerstoffatorns in ATP. Beim Aufbau der Hexose werden sie dann wieder zu ADP und NADP. Ausgangsprodukt der Photosynthese sind also: Wasser und Kohlendioxyd. Bei Anwesenheit von Ribolose 1,5 phosphat, ADP, NADP, Chlorophyll und Licht werden sie zur Hexose unter Abgabe von Sauerstoff. Dem chemisch interessierten Leser seien die Strukturen der genannten chemischen Verbindungen vorgestellt: 15 1. Ribulose 1,5 diphosphat: 2. X
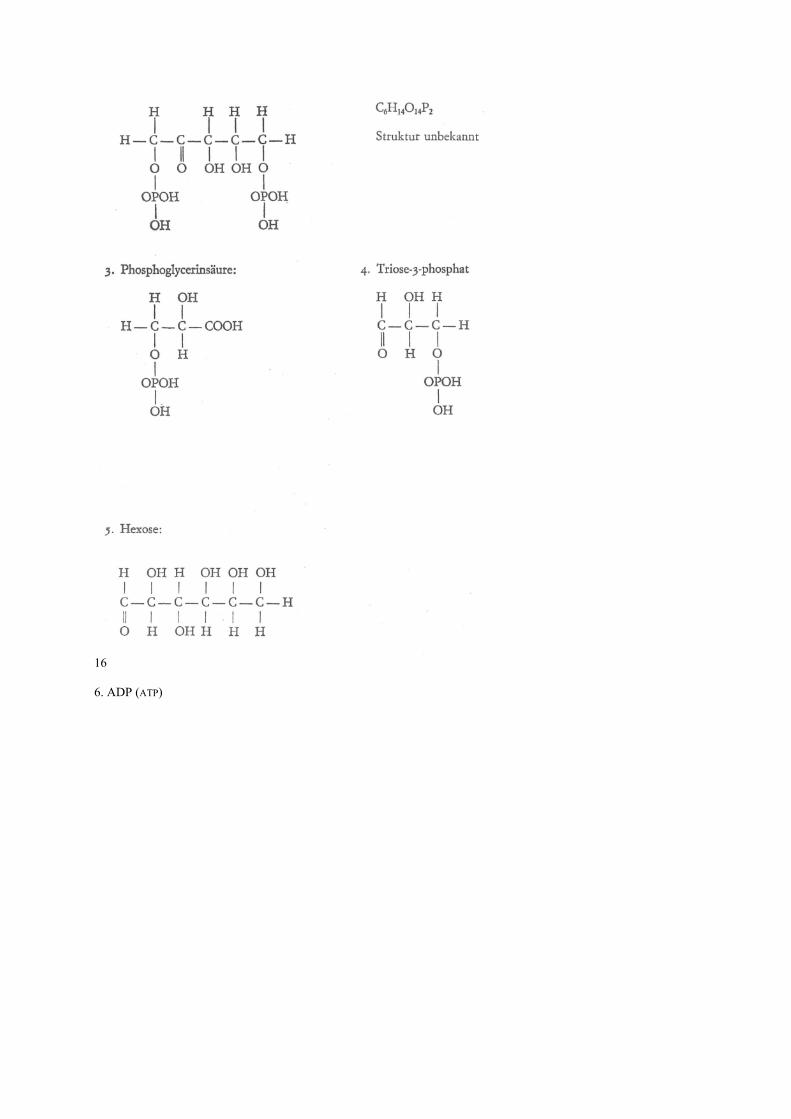
16 6. ADP (ATP)
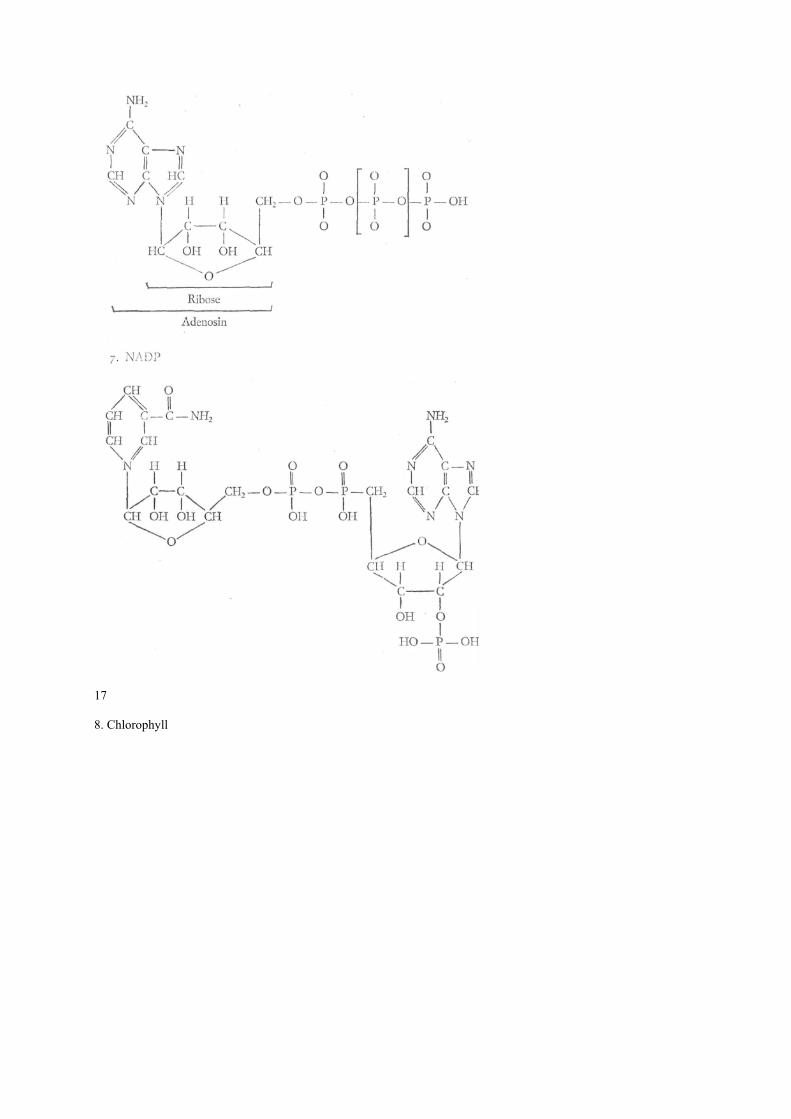
17 8. Chlorophyll

18 ZWEITES KAPITEL
VERMEHRUNG
Neben dem Bau- und Energiestoffwechsel ist die Vermehrung durch quasiidentische Reduplikation das wesentlichste Merkmal einer biologischen Struktur. Eine solche Reduplikation setzt eine Verdoppelung (oder Vervielfältigung) des genetischen Materials voraus. Dieses genetische Material besteht bei allen uns bekannten Lebewesen aus Nukleinsäuren (NS), die oft in Chromosomenfäden eingebaut sind. Meist handelt es sich dabei um Desoxyribonukleinsäuren (DNS), kettenförmige Molekel von gewaltiger Länge (etwa 1 mm und Molekulargewichten bis zu einigen Milliarden). Nukleinsäurefäden als Informationsspeicher welche die Informationen für die gesamte Individualentwicklung eines Lebewesens, insofern sie genetisch gesteuert ist, bestimmen, finden sich jedoch auch in Mikroorganismen, die keine eigentlichen Chromosome besitzen. Es sei hier als Beispiel der Bakteriophage T 5 genannt, eine Virusart, die sich in Escherichia-Coli-Bakterien identisch vervielfacht. Wie manche mikrobiologischen Strukturen [von denen wir philosophisch nicht sicher ausmachen können, ob sie ‹leben› (mehr darüber im Kapitel über Viren)] hat er nur ein DNS-Molekül, während die zellulär strukturierten (sicher lebenden) Organismen mehrere DNS-Moleküle besitzen. Wahrscheinlich ist jedes Chromosom dieser Organismen aus sehr vielen DNS Fäden aufgebaut. Jeder quasiidentischen Reduplikation einer solchen Einheit liegt also eine quasiidentische Reduplikation der für ein Lebewesen spezifischen DNS Molekel zu Grunde. Wir sprechen von ‹quasiidentischer› Reduplikation, weil und insofern die reduplizierte DNS-Apparatur geringfügig chemisch verändert sein kann. Solche Veränderungen (Genmutationen) konnten bei dem Ausgangs-DNS-Bestand ontogenetisch nicht mehr wirksam werden, oder treten erst bei der Reduplikation selbst auf. Das Reduplikat heißt Mutante.
1. Der chemische Aufbau der DNS Jedes DNS-Molekül besteht aus einer langen Kette aneinandergereihter und chemisch gekoppelter Nukleotide. Diese

Nukleotide enthalten als Bausteine eine Pentose (Desoxyribose), eine Phosphatgruppe sowie eine 19 stickstoffhaltige Base. Dabei kann es sich entweder um die Purinbasen Guanin und Adenin oder die Pyrimidinbasen Cytosin und Thymin handeln. Nach den Basen kann man also vier verschiedene Nukleotide, die am Aufbau der DNS beteiligt sind, unterscheiden. Desoxyribose: Die Zählung der Kohlenstoffatome beginnt bei dem unterstrichenen und läuft dann im Uhrzeigersinn weiter.
20 Die Punkte in den Strukturformeln der Basen deuten an, welches Atom an die Pentose gebunden ist. Die Strukturaufklärung der DNS gelang 1953 James Dewey WATSON (geb. 1928) und Francis Harry Compton CRICK (geb. 1916), die 1962 für ihre Arbeiten den Nobelpreis für Medizin erhielten: 1. Die Base des Nukleotids ist an das Kohlenstoffatom 1 der Pentose gebunden, der Phosphatrest hängt am Kohlenstoffatom 5. 2. Bei der Verknüpfung der Nukleotide zu Polynukleotiden, den Nukleinsäuren, erfolgt die Bindung zwischen dem Phosphatrest und dem Kohlenstoffatom 3 der Pentose des benachbarten Nukleotids. Für die genetische Wirkung des DNS-Moleküls ist vor allem die Basenfolge wichtig. Ein Ausschnitt aus einem DNS-Molekül hat also etwa folgende Struktur:

Die Sequenz der Basen wäre Cytosin, Guanin, Thymin (CGT). 21 3. Das Molekül besteht (in der Regel) aus zwei Ketten, die um eine gemeinsame Achse gewunden sind. Diese Doppelhelix hat genau bekannte geometrische Abmessungen. Die Basen weisen auf das Innere der Doppelhelix und sind stets so geordnet, daß dem Thymin des einen Stranges ein Adenin des anderen und einem Cytosin ein Guanin gegenüberliegt. Der Zusammenhalt der beiden Faden geschieht über Wasserstoffbrücken zwischen entsprechenden Basen. Solche Wasserstoffbrückenbindungen sind chemisch gesehen dann möglich, wenn der an ein stark elektronegatives Element gebundene Wasserstoff in der Nähe eines Atoms mit einem ‹einsamen› Elektronenpaar steht. Die Bindungen (sowie die molekularen Distanzen und die Neigungen der Basen zur Kette) lassen sich wie folgt niederschreiben: Thymin-Adenin
Die N—H—O Brücke der Adenin- Thyminbindung mißt 2,80 Å, die N—H—N Brücke 3,00 Å
22

Cytosin-Guanin Die N—H—O Brücken der Cytosin-
Guaninbindung messen 2,90 Å, die N—Η—N Brücke 3,00 Å
Weil sich in der Regel zwei komplementäre Basen gegenüberstehen, besitzt jedes DNS-Molekül die charakteristischen Basenfolgen so, daß nach der Reduplikation die neuen Doppelketten gleich sind.
2. Die Reduplikation der DNS Um die DNS zu verdoppeln, werden die beiden Polynukleotidfäden zunächst reißverschlußartig voneinander getrennt, indem sich die Was-serstoffbrückenbindungen losen. Wenn wir uns das DNS-Molekel als recht verquirlte Strickleiter vorstellen mit den Pentose-Phosphorsäure-rest-Bindungen als Tauen und den durch Wasserstoffbrücken verbundenen Basen als Sprossen, erhalten wir also zunächst zwei Halbleitern. Für das Lösen der Basenpaare ist ein Enzym notwendig. Die Halbleiter wirkt nun als Matrix, d. h. sie nimmt sofort (über Wasserstoff-brückenbindungen der Basen) freie Nukleotide, die im Zellraum meist reichlich vorhanden sind, auf. Darauf werden die einzelnen Nukleotide mittels eines zweiten Enzyms aneinandergekettet. So entstehen zwei untereinander und mit der Ausgangs-DNS per se identische DNS-Moleküle. S. M. FRIEDEMANN und J. B. WEINSTEIN haben 1965 diesen «semi-konservativen Mechanismus» (je eine Helix wird von den beiden neuen DNS-Molekeln von der der Ausgangs-DNS übernommen) durch Mär-kierungsversuche mit 15N in Bakterien-DNS aufgeklärt. 23 Viele Fragen der ‹Technik› der DNS-Reduplikation sind aber noch nicht gelöst. So ist noch nicht sicher bekannt, wie sich die DNS, trotz der spiraligen Verdrillung der beiden Fäden, verdoppeln kann. Es gibt jedoch Hinweise, die vermuten lassen, daß viele DNS-Molekel ringförmig konstruiert sind oder sich doch wenigstens unmittelbar vor der Reduplikation ringförmig schließen. John CAIRNS konnte für die DNS des Escherichia coli nach vorhergehender Markierung mit 3H diese Anordnung nachweisen. Cytosin-Guanin . Die N—H—O Brücken der Cytosin- Guaninbindung messen 2,90 Å, die N—Η—N Brücke 3,00 Å Weil sich in der Regel zwei komplementäre Basen gegenüberstehen, besitzt jedes DNS-Molekül die charakteristischen Basenfolgen so, daß nach der Reduplikation die neuen Doppelketten gleich sind. 2. Die Reduplikation der DNS Um die DNS zu verdoppeln, werden die beiden Polynukleotidfäden zunächst reißverschlußartig voneinander getrennt, indem sich die Was-serstoffbrückenbindungen losen. Wenn wir uns das DNS-Molekel als recht verquirlte Strickleiter vorstellen mit den Pentose-Phosphorsäure-rest-Bindungen als Tauen und den durch Wasserstoffbrücken verbundenen Basen als Sprossen, erhalten wir also zunächst zwei Halbleitern. Für das Lösen der Basenpaare ist ein Enzym notwendig. Die Halbleiter wirkt nun als Matrix, d. h. sie nimmt sofort (über Wasserstoff-brückenbindungen der Basen) freie Nukleotide, die im Zellraum meist reichlich vorhanden sind, auf. Darauf werden die einzelnen Nukleotide mittels eines zweiten Enzyms aneinandergekettet. So entstehen zwei untereinander und mit der Ausgangs-DNS per se identische DNS-Moleküle. S. M. FRIEDEMANN und J. B. WEINSTEIN haben 1965 diesen «semi-konservativen Mechanismus» (je eine Helix wird von den beiden neuen DNS-Molekeln von der der Ausgangs-DNS übernommen) durch Mär-kierungsversuche mit 15N in Bakterien-DNS aufgeklärt. 23
3. Eiweiß Soweit wir wissen, ist alles organismische Leben auf der Erde an Ei-weiße gebunden oder doch funktional mit Eiweißen sehr eng verknüpft. Eiweiße sind recht stabile makromolekulare Verbindungen, die aus langen Perioden von 20 bis etwa 25 verschiedenen a-Aminokarbonsäuren (Eiweißaminosäuren) kettenartig aufgebaut sind. Dabei sind jedoch die verschlungenen Ketten oft an verschiedenen Stellen durch Brücken (meist Disulfidbrücken) miteinander verbunden. Zur chemischen Struktur der Eiweißaminosäuren (AS): die 20 wichtigsten natürlichen AS haben die gleiche Grundstruktur: Als Reste (R) kommen in Frage: Glycin — H Valin — CH — (CH3)2 Isoleucin — CHCII2(CH3)2

Threonin — CHCH3OH Asparginsäure — CH2COO — Glutaminsäure — (CH2)2COO — Arginin — (CH2)3 NHCNH2NH Phenylalanin — CH2C(CH)- Tryptophan — CH2CHCNH C(CH)4C Cystein — CH2SH Alanin —CH3 Leucin — CH2CH(CH3)2 Serin — CH2OH Aspargin — CH2CONH2 24
Glutamin — (CH2)2CONH2 Lysin — (CH2)4NH2 Histidin — CH2(CH2)2(NH)2 Tyrosin — CH2C(CH)4COH Prolin — (CH2)3 Methionin — (CH2)2SCH3 Die Eiweißkette beginnt mit einem Aminoende (— NH2) und schließt mit einem Carboxylende (— COOH). Hinsichtlich der Aminosäuresequenz bestehen keine strengen Gesetzmäßigkeiten, und man hat somit mit jeder Kombinationsmöglichkeit zu rechnen. Das bedeutet, daß es bei einer durchschnittlichen Molekülgröße von 150 Aminosäureeinheiten pro Molekel und 20 verschiedenen Aminosäuren 20150 isomere Eiweißarten (eine 195-stellige Zahl!) gibt. Die Polypetidkette hat ähnliche wie die NS-Kette schraubenförmige Gestalt (Helix), die meist durch Wasserstoffbrücken zwischen den CO- und NH-Gruppen übereinanderliegender Aminosäuren stabilisiert ist. Auch die für die chemischen Umsetzungen wichtigen Biokatalysatorcn (Enzyme) gehören meist zu den Polypeptiden. Mitunter ist an diese Kette noch eine niedermolekulare Komponente (das Koenzym) als Wirkgruppe gebunden. Oft jedoch besteht das Enzym auch nur aus reinem Protein. Zu diesem Typ gehört das bekannte Lysozyrn, das aus 119 Aminosäurebausteinen besteht, deren genaue Reihenfolge bekannt ist. An vier Stellen ist die Kette durch Disulfidbrücken in sich verknüpft. Die Wirkung aller Enzyme ist, wie die Wirkung aller Eiweiße, von der Sequenz der Aminosäuren im Molekül abhängig. Da die Verschiedenheit der Eiweiße Organstrukturen und Organfunktionen weitgehend bestimmt, die in der Ontogenese aus der befruchteten Zelle entstehen und die Lebensfunktionen des ausgewachsenen Individuums regulieren, müssen die Informationen über die spezifischen Aminosauresequenzen im genetischen Material des Individuums, das heißt in seiner DNS, gespeichert sein. 4. Der genetische Code Es kann heute als sicher gelten, daß die Basensequenzen der Nukleinsäuren den genetischen Code enthalten und daß jeweils drei aufeinanderfolgende Basen (Basentriplett) als Chiffre für eine Aminosäure stehen. Bei vier verschiedenen Basen, die in die DNS eingebaut sind, ergeben sich insgesamt 43 = 64 Tripletts. Da die Natur zum Aufbau ihrer Proteine aber nur etwa 20 Aminosäuren verwendet, sind mehr als 40 25 Tripletts überzählig. Die überzähligen Tripletts stehen zum größten Teil jedoch auch als Chiffren für eine Aminosäure, d. h. den meisten Aminosäuren sind mehrere Chiffren zugeordnet (Degeneration des Codes). Außerdem dürfte feststehen, daß es noch einen durch ein oder mehrere Tripletts codierten formalen Begriff gibt: den Begriff ‹Kettenende›. Manche Informationsträger bestehen aus einem einzigen DNS-Molekül, auf dem eine ganze Reihe von verschiedenen Enzymen und anderen Proteinen (Aminosäuresequenzen) codiert sind (z. B. die DNS des Escherichia coli). Die Chiffren dieser verschiedenen Proteine müssen irgendwie voneinander abgegrenzt sein, sonst würden riesengroße Aminosäureketten aneinandergereiht werden. Der eigentliche Aufbau der Eiweiße geht jedoch im Regelfall nicht im Zellkern vor sich, sondern an den Ribosomen. Ribosome sind kleine Körperchen mit einem Durchmesser von etwa 0,015 nm -m Zellplasma. Sie bestehen zu einem hohen Prozentsatz aus Ribonukleinsäure (RNS), einer nahen Verwandten der DNS. RNS und DNS unterscheiden sich bezüglich ihrer chemischen Bausteine darin, daß bei der RNS der Pen-tosepart durch die Ribase (anstelle der Desoxyribose) besetzt ist und anstelle des Thymins das Uracil tritt: Die Anzahl der Ribosomen hängt von der Stoffwechsellage der Zelle ab. Zellen in Stoffwechselruhe haben wenige, solche, die sich teilen, sehr viele Ribosome. Wie kommt nun die Information, die zum Aufbau eines Proteins notwendig ist, vom Zellkern zu den Ribosomen? Zunächst wird die in der DNS enthaltene Information von einem Boten-RNS-Molekel (messenger-RNS oder mRNS)

übernommen. Ein solches mRNS-Molekül hat ein Molekulargewicht von etwa l00 000 bis 500 000. Die mRNS macht ungefähr 5 % der in der Zelle enthaltenen RNS aus. Die Übernahme geschieht durch Anlagerung entsprechender 26
Nukleotide an einen der beiden DNS-Fäden im Zellkern und dem enzymatischen Zusammenschluß dieser Nukleotide etwa in folgender Art:
Das einsträngige RNS-Molekül wandert dann aus dem Zellkern ins Cytoplasma zu einem Ribosom. Hier, an der Oberfläche eines Ribosoms, lagert sich an jeweils ein Basentriplett der mRNS ein Träger-RNS-Molekel (transfer RNS oder tRNS) an, an dessen Ende eine aktivierte Aminosäure gekoppelt ist. Die durch die tRNS in Nachbarschaft gebrachten Aminosäuren verbinden sich zu einer Kette, die schließlich zum Eiweißmolekül wird. Nach erfolgter Anlagerung löst sich die tRNS von der Aminosäure und von der mRNS. Die beiden ersten Basen des von der mRNS bestimmten Codons binden spezifisch die Seitenkette einer Aminosäure, während die dritte Base, die für viele Aminosäuretypen beliebig ist, den allen Aminosäuren gemeinsamen ‹Kopf› anlagert. Das Codelexikon der 64 Basentripletts nach dem Wissensstand vom Herbst 1968 sieht wie folgt aus: Alanin GCU GCC GCA GCG Aspargin AAU AAC Asparginsäure GAU GAG Glutaminsäure GAA GAG Histidin CAU CAC Isoieucin AUU AUC AUA Methionin AUG Phenylalanin UUU UUC Tryptophan UGG Valin GUU GUC GUA GUG Arginin CGU CGC CGA CGG AGA AGG Cystein UGU UGC Glutamin CAA GAG Glycin GGU CGC GGA GGG Leucin UUA UUG CUU CUC GUA CUG Lysin AAA AAG Prolin CCU CGC CCA CCG 27 Serin UCU UCC UCA UCC AGU AGC Threonia ACU ACC ACA ACG unbekannt? UGA (bricht vermutlich Kette ab) bei manchen Mikroorganismen: ‹Kettenende›: UAA UAG Kettenanfang: AUG (wie Methionin) Die Struktur verschiedener tRNS konnte von H. G. ZACHAU (1967), da sie meist nur Molekulargewicht um 25000 haben, aufgeklärt werden, so die der serinspezifischen tRNS mit 77 Nukleotiden. Dabei stellte sich heraus, daß außer den vier ‹klassischen› Nukleotiden 10 ‹seltene› Nukleotide vorkommen, die zumeist durch Methylisierung eines Ringkohlenstoffs, einer Aminogruppe der Nukleinbasen oder der Hydoxylgruppe der Ribose zustande kommen. Die

Strukturen der ‹seltenen› Nukleotide sind bekannt. Die tRNS enthalten Sequenzen, die den Codons (den oben erwähnten Codeworten) komplementär sind (Antikodons). Durch ihre Antikodons ‹erkennen› die tRNS die Codons der mRNS, lagern sich an und bringen so die Aminosäuren in die durch den genetischen Code geforderte Reihenfolge. Um die Funktion des Akzeptierens und Transferierens von Aminosäuren richtig erfüllen zu können, muß die tRNS außer der genannten noch eine weitere in ihrer Struktur begründete Eigenschaft haben: so muß die serinspezifische RNS so gebaut sein, daß sie nur von einem serinaktiven Enzym mit Serin beladen werden kann (und nicht von einer anderen Aminosäure, d. h. sie muß eine Enzymerkennungsregion besitzen). Für jede Aminosäure gibt es also nicht nur wenigstens einen spezifischen Codon, sondern auch ein spezifisches Enzym. Diese die Aminosäuren aktivierenden Enzyme sind ihrer Struktur nach noch weitgehend unbekannt. Sie haben ein Molekulargewicht von etwa 100 000. Die zu den geschilderten Prozessen notwendige Energie wird durch ATP geliefert. Sie verbindet sich etwa zur Ausbildung der tRNS-Molekel mit Aminosäuren zu Aminosäureadenylsäureanhydriden. Die DNS enthält also den genetischen Code. Sie bestimmt weitgehend die Ontogenese der biologischen Struktur. Sie gibt vermutlich auch Anweisung, welche Organe wie und wann auszubilden sind, sie bestimmt die Bildung und Funktion von Enzymen ... Je ‹höher› ein Lebewesen auf der Stufenleiter des Lebendigen steht (je komplizierter seine dynamische Struktur ist), um so mehr Information, um so mehr DNS wird gebraucht. Viele Viren benötigen nur Anweisungen zur identischen Ver- 28 mehrung (d. h. für die Ausbildung der dazu nötigen Enzyme, und des Ablaufs der energetischen Prozesse) und zur Herstellung einer Eiweißhülle, die das genetische Material schützt. Tatsächlich besitzen viele Lebewesen ein «zuviel» an Information, sie werden durch die DNS - wie es auf den ersten Blick scheinen möchte - überinformiert. So «wiegt» bei folgenden Lebewesen der DNS-Bestand im genetischen Apparat (Molekulargewicht in Millionen): Erreger der Polyederkrankheit der Seidenspinnerraupe 2 Krebserreger bei Hamstern (Virus SV 40) 5 Pockenvirus 80 Bakteriophagus Tz 140 Bakterien 2 500 Aspergillus (Pilz) 10 000 Drosophila (Taufliege) 22 000 Maus 1 200 000 (berechnete Werte) Mensch 2 000 000 Was folgt daraus? Ein Protein durchschnittlicher Größe besitzt 500 Aminosäuren. Für seine Bildung sind 500 Codons oder 1500 Nukleotide verantwortlich. Ein Nukleotid hat ein mittleres Molekulargewicht von 330. Das Polyedervirus (mit einsträngiger DNS) besitzt also rund 2000 Nukleotripletts in seiner DNS. Wenn für ein Enzyrn etwa 500 Tripletts notwendig sind, kann es vier Enzyme selbst bestimmen. Da eine solch kleine Zahl nicht zur Reduplikation und zum Aufbau der Eiweißhülle ausreichen dürfte, muß es wohl über seine Enzyme den enzymatischen Apparat der Wirtszelle in Anspruch nehmen, d. h. ihn spezifisch auf die Bildung von Polyederviren umorientieren. Das ebenfalls mit einer einsträngigen DNS ausgestattete SV 40 Virus besitzt schon 5000 eigene Tripletts und kann somit etwa 10 Enzyme synthetisieren. Das Pockenvirus hat eine doppelsträngige DNS, es kann demnach 40 000 Codons tragen und damit 80 Enzyme ausbilden. Für Escherichia coli errechnet man leicht 2 bis 3000 Enzyme. Nun sind bei Bakterien zu Bau und Funktion der Zelle etwa 6oo bis 8oo Arten von Molekeln notwendig, die alle enzymatisch synthetisiert werden. Da ein solches Molekel oft am Ende eines bis zu 10 Schritten bestehenden Synthesevorgangs, von denen jeder ein spezifisches Enzym fordert, steht (zur Ausbildung eines Systems, das die Zelle mit Energie versorgt, sind sogar etwa 3o Syntheseschritte erforderlich), dürfte die Zahl der bereitgestellten Enzyme ungefähr den erforderlichen entsprechen. Beim Menschen sieht die Sache aber entschieden anders aus. Die DNS erlaubt die 29 Ausbildung von 2 Millionen enzymatisch wirkender Proteine, die also etwa 200 000 verschiedene Verbindungen synthetisieren helfen könnten. Diese Zahl ist aber zu groß. Man nimmt heute allgemein an, daß nur ein Teil der in der DNS enthaltenen Information für die Steuerung der Enzymsynthese (Strukturgene) verantwortlich ist, während der andere den Zeitpunkt, die Dauer und das Ausmaß der Synthese bestimmt (Regulatorgene). M. a. W. bei mehrzelligen biologischen Strukturen wird vermutlich ein zunehmend großer Teil des genetischen Materials zur Regulation eingesetzt. Natürlich wird nicht in jeder Zelle der gesamte Genbestand enzymatisch aktiv, obgleich alle Zellen eines mehrzelligen Individuums durch Zellteilung aus einer einzigen Zelle hervorgegangen sind, und somit im Regelfall den gesamten Genbestand und damit den für ein Lebewesen charakteristischen Bestand an DNS besitzen. Hier stellt sich die Frage, wie es kommen mag, daß die eine Zelle als Leberzelle, die andere als Nervenzelle usw. ausgebildet wird. Die Antwort ist relativ einfach: in verschiedenen Zellen werden nur ganz bestimmte Anteile der DNS wirksam. Sehr viel

schwieriger ist die Antwort auf die Frage, welche Mechanismen diese Auswahl regeln. Ein Wort hat man gefunden, aber man weiß noch nicht, was an biochemischer Wirklichkeit dahinter steht: Repressoren. Sie sollen die Bildung der m-RNS an bestimmten Abschnitten der DNS unterdrücken, so daß diese Teile nicht ausgewertet werden. An Riesenchromosomen kann man diese blockierten Stellen im Elektronenmikroskop sogar recht deutlich von Stellen, an denen die rn-RNS die Information abnimmt, unterscheiden. Letztere Orte sind oft zu sogenannten ‹puffs› erweitert. Die chemische Natur der Repressoren (und die der die Repressorwirkung aufhebenden Derepressoren) ist nicht genau bekannt. Es besteht jedoch eine begründete Vermutung, daß es sich bei den Repressoren um Histon-Proteine, bei den Derepressoren um Hormone handelt. Die Wirkung der Hormone könnte also darin bestehen, «daß sie ganz bestimmte Gene aktivieren und damit genabhängige Prozesse in Gang bringen» (vgl. W. BOTSCH, Molekulargenetik, Heidelberg 1967, 44). 30 DRITTES KAPITEL
MUTABILITÄT DER FORMEN
Ein weiteres Charakteristikum des Lebendigen ist die Variabilität der Formen. Ist diese Variabilität vererbbar, spricht man von Mutabilität. Mutabilität ist eine Voraussetzung für die Stammesentwicklung (Phylogenese). Die wichtigsten der Mutabilität zu Grunde liegenden Prozesse sind Mutationen, und hier vor allem die Genmutationen.
1. Genmutation Diese Mutationen beruhen auf einer chemischen Veränderung des genetischen Materials. Solche Änderungen können durch chemische Stoffe oder etwa durch Einwirkung elektromagnetischer Strahlung auf die NS zustande kommen. Ein Beispiel mag das verdeutlichen: Wirkt auf eine bestimmte Aminosäure (Phenylalanin) ein durch ein Gen (Gen 1) verursachtes Enzym (Oxygenase) ein, wird diese zu Tyrosin. Ein anderes Gen (Gen 2) veranlaßt die Bildung des Enzyms Tyrosinase, das eine Verwandlung des Tyrosins über ein Zwischenprodukt (3,4 Dihydroxyphenylalanin) in Melanin bewirkt. Melanin ist verantwortlich für die Hautpigmentierung beim Menschen. Wird nun das Gen 2 durch chemische Veränderung daran gehindert, gerade Tyrosinase zu produzieren, wird die Melaninbildung unmöglich gemacht. Es entsteht als Mutante ein Albino. Soweit bekannt, verhindern Mutationen im Regelfall die Ausbildung eines bestimmten Enzyms und führen damit zu einem enzymaiischen Defizit im Organismus. Nur in den seltensten Fallen (man kann in etwa ausrechnen, wie selten solche Fälle sind) kommt es zur Ausbildung eines anderen wirksamen Enzyms, das dann unter Umständen zu einer neuen ‹positiven› Eigenschaft führen kann. Aber auch solche ‹positiven› Neuerwerbungen sind in den seltensten Fällen positiv im Sinne eines Selektionsvorteils der neu entstandenen Mutanten gegenüber nicht mutierten Formen. Es scheint in diesem Stadium unserer Untersuchung äußerst zweifelhaft die Phylogenese allein auf solche ‹zufälligen› mutationsauslösende Faktoren zurückzuführen. Wir werden weiter unten sehen, daß mit großer Sicherheit andere Strukturelemente anzunehmen sind, die nicht mehr rein chemisch oder physikalisch beschrieben werden können. 31 Neben den Genmutationen gibt es jedoch auch andere Möglichkeiten, das Erbgut positiv zu verändern. Da diese teilweise manipulierbar sind, gehört ihnen das besondere Interesse der biologischen Forschung.
2. Transformation Unter Transformation versteht man eine Manipulation des DNS-Bestan-des durch Einwirken einer «fremden» DNS: Ein Beispiel mag das veranschaulichen: Unter den Pneumokokken (Bakterien) sind uns pathogene (genuine Lungenentzündung und andere entzündliche oder eitrige Erkrankungen erzeugende Formen) (Pneumokokkus S) und nicht pathogene (Pneumokokkus R) Formen bekannt. Extrahiert man durch Zentrifugieren die DNS der S-Formen und läßt diese auf R-Formen einwirken, wandeln sich etwa 1 % der R-Formen zu S-Formen, d. h. sie werden pathogen. Diese Eigenschaft wird auch auf die Nachkommen der so umgestimmten R-Kokken weitervererbt. Transformationsähnliche Veränderungen des Erbgutes sind aber auch natürlich beobachtet worden. Bei manchen Bakterien wechseln Genomteile zwischen einem integrierten Status und einem freien hin und her (Episome). Im freien Zustand können sie sich u. U. selbständig vermehren und bei Zellkontakt in die Nachbarzelle ‹hinüberreplizieren› und dabei Gene von der einen in die andere Zelle mit hinübernehmen. Eigentliche Transformationen können bei verschiedenen Bakterienarten durchgeführt werden (etwa bei denen der Gattungen Haemophilus, Xanthosmonas, Streptokokkus). In diesem Zusammenhang stellt sich die Frage, ob solche genetischen Manipulationen auch bei höheren Tieren, vielleicht gar beim Menschen möglich sind. «Zur eindeutigen experimentellen Prüfung dieser Frage ist es zunächst notwendig, isolierte Zellen vielzelliger Organismen in geeigneten Nährmedien kultivieren zu können... Bereits seit längerer Zeit ist diese Voraussetzung erfüllt... Dabei hat sich gezeigt, daß menschliche Zellen durchaus in der Lage sind, zugesetzte DNS aufzunehmen... Nachdem der Nachweis möglicher Transformationen von Zellen erbracht worden ist, läßt sich nicht ausschließen, daß in naher Zukunft auch Wege gefunden werden, nicht nur isoliert kultivierte Zellen, sondern auch Zellen in Gewebeverbänden und vielzelligen Organismen zu transformieren . . . Nach allem, was wir heute wissen, ist es keine abwegige Spekulation, wenn man sich vorstellt, daß z. B. durch diese Methoden erbliche Defekte ... behoben werden könnten.» (W. Laskowski, Elemente

des Lebens, Berlin 1966, 180f.). 32
3. Transduktion Transduktion ist die Übertragung genetischen Materials durch Bakteriophagen. Diese sind - wie wir noch zeigen werden — virusähnliche Gebilde, die Bakterien befallen und zur Auflösung bringen können. Im Regelfall dringt die Phagus-DNS in das Bakteriurn ein und zwingt den Bakterienstoffwechsel, neue Phagus-DNS zu produzieren. Aber nicht immer kommt es dazu: bei sog. temperierten Phagen, die vermutlich häufiger vorkommen als virulente Phagen, dringt die Phagus-DNS in das Bakterium ein, und die Phagus-DNS lagert sich in die Bakterien-DNS ein. Das Bakteriengenom enthält jetzt auch die Erbinformation zur Produktion von Phagen. Solche Bakterien nennt man lysogen. Vermehren sie sich, vermehrt sich auch die Phagus-DNS und das veränderte Genom geht auf alle Nachkommen des Bakteriums über. In solchen lysogenen Bakterien ist also auch die Information zur Phagenbildung vorhanden, wird aber durch einen Repressor blockiert. Phagen entstehen erst dann, wenn der Repressor ausfallen sollte. Wie kommt es nun zur Transduktion? Wir wollen uns das am Beispiel zweier Salmonellenstämme veranschaulichen. Ein Bakterium vom Stamm A wird durch einen Phagus infiziert und dabei zur Auflösung gebracht. Einige der neu entstandenen Phagen nehmen geringe Teile der Bakterien-DNS mit. Befällt nun ein solcher Phagus ein lysogenes Bakterium vom Stamm B, wird außer der Phagus-DNS auch noch der DNS-Rest des lysierten Bakteriums in die Bakterien-DNS eingelagert und ins Erbgut übernommen. Der Phagus hat somit Erbinformation von einem auf ein anderes Bakterium übertragen (transduziert). Phagen transduzieren aber niemals ein ganzes Genom, sondern immer nur einzelne Gene. Bei Versuchen an Salmonellen wurde Z. B. die Fähigkeit, die Aminosäure Tryptophan zu synthetisieren, transduziert (vgl. dazu W. Botsch, a. a. O., 32-36). Transformation und Transduktion geben also grundsätzliche Möglichkeiten an die Hand, das Erbgut in bestimmter Weise zu manipulieren, indem Eigenschaften von einem «Stamm» auf den anderen übertragen werden. Die Versuche sind vorläufig noch auf Bakterien beschränkt. Damit wollen wir unsere Überlegungen zu einer biologischen Beschreibung der Aussageformen des Lebendigen abbrechen. Vor diesem empirischen Hintergrund wird es uns vielleicht möglich sein, die Frage nach der Natur des Lebendigen (was mit lebendig gemeint ist, haben wir jetzt erklärt) zu beantworten. 33 ZWEITER ABSCHNITT
NATURPHILOSOPHISCHE FRAGEN UM DIE «NATUR» DES LEBENDIGEN
Die zentrale Frage nach dem Verständnis dessen, was Leben eigentlich bedeutet, hat schon seit altersher die Gemüter bewegt. Wir wollen hier nur einige Theorien erwähnen, die noch heute zur Diskussion stehen, um uns dann für diejenige zu entscheiden, die am besten dem empirischen Befund gerecht wird. 34 ERSTES KAPITEL
DIE SCHOLASTISCHE THEORIE
These: Das Lebewesen hat wesentlich die Fähigkeit, sich aus sich selbst zu bewegen. Diese aristotelische These scheint zunächst dem Grundsatz des ARISTOTELES zu widersprechen, nach dem alles, was sich bewegt, von einem anderen bewegt wird (Phys. VII, 1; 241 b). Dazu aber schreibt er: «Was sich aus sich selbst bewegt, bewegt sich gemäß seiner Natur, z. B. ein Tier. Ein Tier bewegt sich ja aus sich selbst» (Phys. VIII, 4; 254 b) und modifiziert: «Wir können (an Lebewesen) beobachten, daß sie zuzeiten unbewegt sind, sich dann aber wieder bewegen, wie es scheint. Das muß man aber so verstehen, daß sie nur eine einzige Bewegungsart selbst zustande bringen und auch die nicht ganz selbständig. Denn nicht aus ihnen stammt die Ursache, sondern es gibt in den Lebewesen viele natürliche Bewegungen, die sich nicht aus sich veranlassen, wie z. B. Wachstum, Stoffwechsel, Atmung; das sind Bewegungen, die jedes Lebewesen ausführt, auch wenn es ruht und die von ihm aus mögliche Bewegung unterläßt oder nicht ... Wenn die Lebewesen sich nach dem Schlaf bewegen, dann immer so, daß der erste Anstoß von außen kommt ... in allen bewegt sich der erste Beweger und die Ursache ihrer Selbstbewegung» (Phys. VIII, 6; 259 b). Mit dieser merkwürdigen Ursache werden wir uns noch zu beschäftigen haben. In seinem Buch über die Seele meint ARISTOTELES: «Leben hat ein Ding, wenn es aus sich selbst Nahrung aufnimmt, wächst und schwindet. Somit ist also jeder belebte natürliche Körper Substanz (ουσία) und zwar zusammengesetzt» (II, 1; 4123). Zusammengesetzt nämlich aus Körper und Seele, die ihrerseits Substanz (ουσία), Gestalt (είδος), Form (µορφή) eines Lebewesens ist (ibd.). Die Schüler des Aristoteles folgen ihm in dieser Bestimmung des Lebendigen, so AVERROES (in Phys. VIII comm. 17 und 20). THOMAS VON AQUIN (In III Sent. d. 35, 1, 1 c; de verit. 4. 8; Summa theol. I, 18, 1 c und 3 c), SUAREZ (Disp. met. d. 24 sect. 2 n. 6), THOMAS (Pot. 10, 1 c), SUAREZ (d. 24. sect. 2 n. 6) und viele andere der von Aristoteles mittelbar oder unmittelbar beeinflußten scholastischen Philosophen verbinden mit der Selbstbewegungstheorie bis in

unsere Tage als akzidentellen Ausdruck des Lebendigseins die Fähigkeit «von, aus und nach innen» zu wirken (actio immanens). Diese actio-immanens-Theorie des 35 Lebendigen beeinflußte auch die Lehren HOBBES' (Leviathan, Einl.), SPINOZAS (Cogit. met. II, 6), CRUSIUS' (Vernunftwahrheiten § 458), KANTS (WW IV, 439) ... Die Neuscholastik sieht in der Bestimmung des Lebens als actio immanens meist eine Wesensbestimmung. Versteht man die ‹actio› als aristotelische Kategorie (vgl. WS II, 10), wird das Leben (besser das ‹Lebendigsein›) nur als Akzidenz eines materiellen Körpers gelehrt. Wir sind dagegen der Meinung, daß Leben eine (transzendentale) Eigenschaft des belebt Seienden ist, insofern es da-ist (und nicht nur insofern es so-ist). Im anderen Zusammenhang haben wir das Leben als partikuläres Transzendentale charakterisiert (WS II, 10 Anm. 14). ZWEITES KAPITEL
DIE MASCHINENTHEORIE DES LEBENDIGEN
Die Maschinentheorie des Lebendigen in ihren heutigen Gestalten geht auf DESCARTES zurück. Descartes unterschied bekanntlich zwei «Arten von Substanzen»: ausgedehnte (res extensae) und denkende (res cogitantes), die beim Menschen in einem wirkursächlichen Zusammenhang miteinander stehen. Die Seele also wirkt auf den Körper. So kommt er mit einiger Notwendigkeit zu dem Schluß: «Er wird diesen Leib für eine Maschine ansehen, die aus den Händen Gottes kommt und daher unvergleichlich besser konstruiert ist und weit wunderbarere Getriebe in sich birgt als jede Maschine, die der Mensch erfinden kann» (Methode V, 9). Da er die untermenschlichen Lebewesen nicht mit einer res cogitans ausstattet, ist er gezwungen, Pflanzen und Tiere als bloße Maschinen zu interpretieren. Den Pflanzen spricht er ein ‹Lebensprinzip› ab, da nur die Ordnung und die Bewegung der Teile ihre Funktionalität bestimme, den Tieren, weil sich sonst die Menschen in ihrem Verhalten ihnen gegenüber der größten Grausamkeiten schuldig machen würden. Der eigentliche Grund aber ist die Unfähigkeit von Pflanzen und Tieren zu ‹denken›, so daß ihnen keine «res cogitans» — und nur sie gibt es neben der res extensa — zugesprochen werden kann. Übrigens hatte schon einige Jahrzehnte zuvor ein spanischer Arzt und begeisterter Gegner des Aristoteles, Gomez PEREIRA, in seiner Schrift «Antoniana magerita» Tieren das Belebtsein abgesprochen und sie als Maschinen interpretiert. Die Lehre des DESCARTES ist jedoch nicht immer so eindeutig, wie es auf den ersten Blick erscheinen möchte. Wiederholt spricht er von «Lebensgeistern», welche die Muskeln veranlassen, die Glieder zu bewegen. Diese «Lebensgeister» (Spiritus animales) versteht er als «äußerst feine Luft» und «reinste Flammen», die im Herz erzeugt und durch das Blut verteilt werden. Heute würden wir das von DESCARTES Gemeinte chemisch interpretieren: Es wäre dann etwa von Oxydationsprozessen in Erythrozyten zu sprechen. Diese «Lebensgeister» haben mit Geist etwa soviel zu tun, wie der Weingeist. Die von Descartes vorgestellte Maschinentheorie wird heute in der Regel nicht mehr vertreten. An ihre Stelle sind Theorien getreten, die entweder alle Lebenserscheinungen aus dem physiko-chemischen Bau der Organismen erklären und neben physikalischen und chemischen Ursachen keine anderen gelten lassen wollen (extremer Mechanismus) oder bio- 37 logische Prozesse auf kybernetische Regelkreise reduzieren (Kybernetik). Im ersten Fall wird das Lebewesen als reine materielle Einheit ohne eigenes einheitsstiftendes Prinzip, im letzten in Entsprechung zur technischen Einheit betrachtet. Der Mechanismus ist historisch aus der Descartes'schen Theorie hervorgegangen und verdient deshalb, hier behandelt zu werden, die kybernetische Interpretation des Lebendigen schließt sich inhaltlich vielleicht am deutlichsten an die Maschinentheorie an.
1. Mechanistische Theorie des Lebendigen These: Das Lebewesen ist eine rein materielle Einheit ohne ein einheitsstiftendes Prinzip. Wir wollen hier ohne jeden Anspruch auf Vollständigkeit einige typische mechanische Deutungen des Lebendigen vorführen. Ernst HAECKEL hat verschiedentlich (Generelle Morphologie, Lebenswunder) die Meinung geäußert, zwischen Kristallen und niederen Lebewesen bestehe eine nahe Verwandtschaft. Der Einwand, daß Lebewesen «weiche, manchmal eiweißartig-flüssige Gebilde» seien, «während Kristalle als typische starre Körper gelten» wurde zuerst von O. LEHMANN (Flüssige Kristalle, Leipzig 1906) ausgeräumt. Als besonders geeignet erwies sich eine kristalline (wie Kristalle lichtbrechende und polarisierende) Flüssigkeit, der Parazoxyzimtsäureäthylester:
Dieser Stoff stößt bei Abkühlung innerhalb bestimmter Temperaturen (136-117° C?) plötzlich Vorwölbung aus, die sich bald zu Knospen und dann zu sich heftig windenden Schlangen ausformen. Flüssige Kristalle wachsen durch Aufnahme chemisch gleichartiger Moleküle und scheiden fremde Stoffe (z. B. Tusche) aus. Beim Wachstum bleibt die Dicke der Kristallschlangen konstant. Übersteigt die Aufnahme ein bestimmtes Maß, teilen sich die Kristalle von selbst und beginnen ein ‹Eigenleben›. Heute gilt die Theorie vorn Belebtsein auch solcher Kristalle allgemein als

überwunden. Mechanistisch in unserem Sinne ist vor allem aber auch jede Theorie des Lebendigen, welche alle Funktionen des Lebewesen auf die von der DNS gelieferten Information bzw. deren Blockierung durch Repressoren zurückführt. Doch hat diese Theorie ihre Schwierigkeiten. Vor allem ist 38 es bis zum heutigen Tag noch nicht gelungen, die Funktion der Repressoren und Gegenrepressoren auf die in der DNS angelegte Information zurückzuführen. Sie vermag also nicht oder nur mit Hilfe von Vermutungen ad hoc zu erklären, wie und warum gerade aus dieser Zelle eine Leber- aus jener Zelle eine Nervenzelle wird. Nicht erklärt werden vor allem die Ergebnisse von Transplantationsversuchen. Transplantiert man Gewebeteile eines Molchembryos auf ein anderes, dann verhält sich solch junges Gewebe orfsgemäß. Pflanzt man etwa einen embryonalen Molchschwanz in der Schultergegend eines anderen Molchembryos ein, so entwickelt sich daraus ein Vorderbein. Viele Zellen junger Gewebe sind aequipotentiell und können je nach dem Ort, an dem sie eingepflanzt werden, sehr verschiedene Funktionen übernehmen. Der Blockermechanismus der Repressoren ist also schon keineswegs mit dem Entstehen einer Zelle gegeben. Die «Lage» bestimmt, welche Repressoren wirksam werden und welche nicht. Die Lagebedingtheit der Funktion der Repressoren wird deutlich auch am Beispiel der Regenerationen. So bilden in der normalen Entwicklung Molche die Augenlinse aus der Hornhaut. Entfernt man die Linse, regeneriert der Triton aus Irisgewebe eine neue Linse. Das Irisgewebe bleibt also auch bei erwachsenen Individuen noch aequipotentiell, und paßt seine Funktion der Lage an. Ganz unerklärlich ist in einer streng mechanistischen Theorie der verschiedentlich beobachtete Tatbestand, daß Teile von Ahnentieren regeneriert werden: So bilden mitunter verwundete Krebse statt Augenstielen Antennen aus. Eine spezielle Form des Mechanismus hat die Biologie des Dialektischen Materialismus entwickelt. OPARIN löst sich nachdrücklich von primitiven mechanischen Theorien: «Es geht nicht an, wie das die Vertreter der mechanistischen Auffassung machen, alles, was nicht in den Rahmen der Physik und Chemie paßt als ... übermateriell anzusehen. Im Gegenteil, die Formen der Organisation und Bewegung der Materie können sehr verschiedenartig sein» (a. a. O., 5). «Die Besonderheit, durch die sich das Leben von allen anderen Bewegungsformen der Materie (insbesondere von anorganischen fließenden Systemen) qualitativ unterscheidet, besteht darin, daß in den lebenden Körpern viele Hunderttausende individueller chemischer Reaktionen, die in ihrer Gesamtheit den Stoffwechsel darstellen, nicht nur in bezug auf Raum und Zeit streng aufeinander abgestimmt sind, nicht nur harmonisch in der einzigartigen Ordnung der unaufhörlichen Selbsterneuerung verknüpft sind, sondern daß auch diese ganze Ordnung gesetzmäßig auf die ständige Selbsterhaltung und Selbsterzeugung des gesamten lebenden Systems als Ganzes gerichtet und äußerst vollkommen darauf abgestimmt ist, die 39 Aufgabe lösen zu können, die Existenz des Organismus unter den gegebenen Umweltbedingungen zu sichern» (a.a.O. 9,f). Oparin führt hier den Begriff der Zweckmäßigkeit ein. «Die Idealisten erblicken in dieser ‹Zweckmäßigkeit› die Erfüllung irgendwelcher höherer Bestimmungen ... Die Materialisten benutzen dagegen das Wort (aus Mangel an einem besseren) nur zur kürzeren Bezeichnung der Übereinstimmung der Organisation des gesamten lebenden Systems mit der Selbsterhaltung ... unter den gegebenen Umweltbedingungen, ferner auch zur Bezeichnung der Anpassung der Struktur einzelner Teile des lebenden Systems an die vollständige und koordinierte Verwirklichung derjenigen lebensnotwendigen Funktionen, die die erwähnten Teile im lebenden System wie im ganzen tragen» (a. a. O. 11 f). Zusammenfassend läßt sich sagen: - Leben ist eine qualitativ von anderen unterschiedene Bewegungsform der Materie, - Merkmal dieser Bewegungsform ist eine spezifische «Zweckmäßigkeit», die dem lebenden System als Ganzem im Verhältnis zu seinen Umweltbedingungen zukommt. - Diese Zweckmäßigkeit ist auch Eigenschaft der Funktionen der Teile der lebendigen Ganzheit in bezug auf die Erhaltung eben dieser Ganzheit. Gegen die klassische Maschinentheorie argumentiert OPARIN z. Teil recht dogmatisch. Während die Funktion der Maschinen im Regelfall von ihrer Konstruktion abhängt und nicht von dem Baumaterial, sei das bei Lebewesen anders, die «nach einem treffenden Ausdruck von Engels ‹Eiweißkörper› sind» (a.a.O. 18). Im Gegensatz zur statischen Grundkonstruktion der Maschine ist die Existenz eines Organismus an ständig ablaufende chemische Reaktionen gebunden, deren Stillstand zum Tod des Organismus führt. Das Prinzip der Beständigkeit ist also bei Maschinen und Organismen grundsätzlich verschieden. Gegen den Mechanismus der modernen Molekularbiologie argumentiert er gut: «Je weiter die konkrete biochemische Erforschung des Prozesses der Selbsterzeugung lebender Körper fortschreitet, umso mehr wird offenbar, daß dieser Prozeß nicht nur mit dem einen oder anderen einzelnen Stoff... verbunden ist, sondern von dem ganzen System der Organisation des lebenden Körpers bestimmt wird, das ... einen fließenden Charakter besitzt und kaum mit einer mit unveränderlicher Matrize ausgestatteten Stempelmaschine verglichen werden kann» (a. a. O., 23). Für jeden Vergleich der biologischen mit einer wie immer gearteten technischen Einheit gilt, das letztere «zwangsläufig in ihrer 40

Organisation diejenigen spezifischen Aufgaben widerspiegeln, die ihnen ihr Konstrukteur gestellt hat, sie werden stets Spuren ihres Ursprungs in sich tragen» (a. a. O. 24 f.). Damit leitet er zu einer wesentlichen Überlegung über: die der ‹Entstehung des Lebendigen», die sich grundsätzlich anders vollzieht als das Entstehen einer Maschine. Die ursprünglich mechanistische Theorie müßte eine rein zufällige Entstehung des ersten lebenden Molekels annehmen, eine Hypothese, die nach OPARIN «praktisch völlig unfruchtbar und ... theoretisch auch nicht richtig ist» (a. a. O. 27). Wir wollen bei aller Anerkennung der Thesen Oparins im Einzelnen doch nicht übersehen, daß er nicht anzugeben vermag, wodurch sich die materielle Bewegungsform ‹Leben› denn qualitativ von materiell-technischen Einheiten unterscheidet. Eine Aufzählung verschiedener Funktionsweisen beantwortet die Frage nicht. Muß nicht qualitativ anderen Funktionen auch eine qualitativ andere Struktur zugrunde liegen? Weil und insofern er diese zentrale Frage umgeht, glauben wir - bei aller Anerkennung einzelner Gedanken - seine Theorie des Lebens den mechanistischen zuzählen zu dürfen, wenn wir auch gerne zugeben, daß eine materialistische Theorie nicht schon notwendig mechanistisch sein muß, wie wir am Beispiel des Holismus zeigen werden.
2. Kybernetische Theorien des Lebendigen These: Das Lebewesen ist eine kybernetische Einheit ohne ein nicht kybernetisch zu deutendes spezifisches Prinzip. Der Terminus «Kybernetik» dürfte erstmals von AMPÈRE (Essai sur la philosophie des sciences, 1834) in die Wissenschaft eingebracht worden sein. Er nannte die Methoden der Ansteuerung eines politischen Zieles durch die Regierung cybernétique. Vermutlich ohne die Schrift Amperes zu kennen, schrieb N. WIENER 1948 ein Buch Cybernetics und bezeichnete mit diesem Wort eine ‹neue› Wissenschaft, die die allgemeine Regeltheorie mit der Theorie der Information und Informationsverarbeitung in Zusammenhang brachte. Da bis zum heutigen Tag noch keine allgemein akzeptierte Definition des Begriffes ‹Kybernetik› bekannt ist, mag ein Rückgriff auf die Sprachherkunft des Wortes das Gemeinte veranschaulichen. In jedem soziotechnischen System, etwa einem Schiff auf Fahrt, kann man wenigstens vier Instanzen voneinander unterscheiden: - den Kapitän, der das Ziel (SOLL) setzt und somit zwischen mehreren möglichen Zielen eines auswählt, 41 - den Lotsen, der den augenblicklichen Zustand (IST) ermittelt und ein Programm entwickelt, diesen in den SOLL-Zustand zu überführen. Er muß beide Zustände miteinander vergleichen und das Resultat in Befehlsform (determinierte Entscheidung) an den Steuermann weitergeben, - den Steuermann, der das Schiff nach diesen Befehlen steuert, indem er mit geringem Arbeitsaufwand die Energie des Antriebssystems zur Verwirklichung des Befehls einsetzt, - das Antriebssystem (nach H. Frank). Der jeweils neu zu bestimmende IST-Zustand wird dann dem Lotsen zu modifizierten Befehlen führen, so daß endlich der SOLL-Zustand erreicht wird. Das Wort κυβερνήτης bezeichnet im Griechischen den Steuermann. Kybernetik wäre dann die Wissenschaft, den Steuermann durch Kenntnisse des SOLL-Zustandes und der jeweiligen IST-Zustände unter Zuhilfenahme des Antriebssystems dazu zu bringen, den SOLL-Zustand zu erreichen. Wir haben damit schon einen einfachen Regelkreis kennengelernt: Einem Regelkreis wird zunächst eine Führungsgröße eingegeben. Sie wird im Regler (regulierendes Zentrum) gespeichert. Der Regler überträgt die Stellgröße mittels eines Korrekturmechanismus (Stellglied) in die Zone der Regelgröße (einem durch das SOLL konstant zu haltenden Vorgang oder Zustand). In die Zone der Regelgröße wirken nun durch Außeneinflüsse (in unserem Schiffsbeispiel sich ändernde Strömungs- oder Windverhältnisse) Faktoren ein, die wir Störgröße nennen wollen. Eine Meßeinrichtung überträgt nun den IST-Wert wieder an den Regler, der durch neue Impulse die Regelgröße dem SOLL-Wert annähert. Als biologisches Beispiel sei die Pupillenreaktion der meisten Wirbeltiere angeführt: Die Pupille stellt sich in ihrer Größe auf die Helligkeit ein. Der Lichteinfall ins Auge ist die Regelgröße, die wechselnde Helligkeit die Störgröße, das ZNS (Zentralnervensystem) enthält den Regler, die Irismuskeln sind die Stellglieder. Nun lassen sich die weitaus meisten biologischen Vorgänge als solche Regelkreisvorgänge beschreiben. Da solche Regelkreise nun auch das Geschehen in technischen kybernetischen Systemen bestimmen, hin vom Fliehkraftregler bis zum «Rechenautomaten», liegt es nahe, biologische Einheiten mit solchen technischen Einheiten nicht nur zu vergleichen, sondern auch zu identifizieren. Während der Vergleich durchaus legitim ist, ist die Identifikation wissenschaftstheoretisch äußerst bedenklich, folgt doch aus ähnlicher Funktion nicht schon eine gleiche Struktur. Nun haben wir etwas voreilig von einer Ähnlichkeit der Funktion gesprochen. Ein näheres Zuschauen zeigt, daß es damit keineswegs ganz so eindeutig bestellt ist, wie es eine universelle kybernetische Theorie 42 wahrhaben möchte. Biologische Regelkreise sind zunächst sehr viel «mechanischer» als technische. Letztere ahmen sehr viel eher menschliche Funktionen nach, denn biologische. Ich denke, daß man, bis zum Beweis des Gegenteils, soweit gehen darf zu sagen, daß ein «Computor» von der Kompliziertheit eines menschlichen ZNS behaupten würde, er könne denken und habe Selbstbewußtsein. Der Akzent liegt auf dem Wort behaupten. Man sollte diese Einschränkung aber nicht überschätzen. Wenn wir auch nur mit der Möglichkeit rechnen müssen, daß es außer dem Menschen noch andere biologische oder technische Einheiten gibt, die denken können oder Selbstbewußtsein haben,

dürfen wir diese beiden Begriffe nicht vom Selbstverständnis des Menschen her bestimmen, sondern müssen eine Definition von den Denkäußerungen und Bewußtseinsäußerungen her versuchen. Damit aber wird der Tatbestand recht problematisch: Kann doch ein solcher Computor vermutlich - er ist noch nicht gebaut, und wird auch vorläufig nicht gebaut werden können - funktional ganz dasselbe leisten wie der Mensch, d. h. glaubhaft versichern, er denke und habe Selbstbewußtsein. Ferner sei festgestellt, daß die Behauptung, Regelkreissysteme würden biologische Funktionen nur simulieren, schon den Unterschied zwischen technischer und biologischer Funktion apriori festgelegt. Diese Festlegung ist jedoch in nichts als einem ontologischem Vorurteil gegründet. Es ist durchaus wahrscheinlich, daß vergleichsweise komplexe Automaten ganz dasselbe leisten, wie ein ZNS. Ein funktionaler und struktureller (weil wir nur von Funktion legitim auf Struktur schließen können aber auch müssen) Unterschied läßt sich nach dem heutigen Stand des Wissens nicht sicher behaupten. So wollen wir dann die Hinweise im folgenden Teil dieses Kapitels nicht überbewerten - ernst zu nehmen sind sie jedoch immerhin. Ein Unterschied sei schon hier festgehalten: das Modell des Regelkreises enthält gleichsam das ‹Gehirn› seines Konstrukteurs. Doch kehren wir zurück zum Vergleich biologischer und technischer Einheiten. V. VON WEIZSÄCKER und seine Schüler haben da einen immerhin beachtenswerten Unterschied herausgefunden, den wir berücksichtigen müssen. Das biologische Geschehen ist nicht durchgängig von Regelkreismechanismen bestimmt, sondern teilweise charakterisiert durch Gestaltkreise. (Vgl. V. von Weizsäcker, Der Gestaltkreis, Theorie der Einheit von Wahrnehmen und Bewegen, 1947). Der Gestaltkreis wird im Gegensatz zum vorgegebenen Regelkreis vom Lebewesen selbst produziert, indem die Zielvorstellungen (das SOLL, das Programm) erst «unterwegs» in der Handlung selbst aufgebaut wird. Zwar setzt ein solcher Gestaltkreis einen funktionierenden Regelkreis voraus, doch läuft das Geschehen nicht nach Art eines fortlaufenden und sich zuneh- 43 mend erhellenden geschlossenen Hin und Her (das die IST-Werte zunehmend auf die SOLL-Werte ausrichtet) in kreisartiger Verbundenheit ab, sondern nach Art eines offenen Systems, bei dem zusätzliche, den endgültigen SOLL-Wert erst während des Handelns selbst aufbauende, (und nicht nur den IST-Wert feststellende) Informationen dem kybernetischen System eingegeben werden. Somit erhält biologische Leistung (im Gegensatz zur technischen) notwendig den Charakter der Improvisation, weil und insofern biologische Leistung immer in dialektischem Bezug steht zur Sinnstruktur der Situation, der Befindlichkeit (Stimmung ...) des biologischen Systems und der «unergründlichen Genialität des Könnens». Der Terminus «Gestaltkreis» ist also ein durchaus der Kybernetik verpflichteter Terminus, der nur sinnvoll in dem zeitknapp-sten Bereich angewendet werden kann, «in welchem die rückläufige Bestimmung die Kohärenz eines intentional engagierten Aktes zur Zeit seines Vollzuges garantiert» (A. Auersperg). Untersuchen wir nun einmal einige funktionale Unterschiede zwischen Regel- und Gestaltkreis: 1. Der Gestaltkreis hat eine eigene Zeitstruktur, insofern das biologische Verhalten von künftigen Eindrücken, d. h. von der Bedeutung kommender Wahrnehmung und Bewegung in einer Art Vorausschau bestimmt wird. So erfolgt etwa die Umstellung des Kreislaufs beim Leistungssportler schon vor dem Start (Blutdruck und Pulsfrequenz werden erhöht), obschon gerade dieses Beispiel der Zukunftsorientiert-heit durch in der Vergangenheit gespeicherte Information zu erklären ist. Die prospektive Regulation ist vom Ganzen der Leistung her bestimmt, so daß sie beim Kurzstreckenläufer anders aussieht als beim Langstreckenläufer. Der Gestaltkreis setzt so etwas wie eine zeitüberbrückende Vergegenwärtigung voraus. Hier wird ein Sachverhalt empirisch verständlich, der uns (WS II, 14 und 278 ff) dazu geführt hat, physikalische Zeiten von der biologischen Realzeit zu unterscheiden. Biologische Werte können nicht von uns gesetzt werden. Es gibt somit auch keine eigentlichen biologischen SOLL-Werte, sondern nur IST-Werte, die man ihrer Häufigkeit wegen statistisch (im Sinn statistischer Normalität) zu SOLL-Werten erklärt hat. 2. Technische Systeme lernen durch die Negation des Unzutreffenden. Das Lernen durch Versuch und Irrtum (instrumentale Koordinierung) kann sicherlich auch von Automaten geleistet werden, doch unterscheidet es sich vom biologischen Lernen zunächst einmal dadurch, daß dieses eine spontane, oft zufällige Suche nach erwünschten Außenweltreizen hinzufügt. Das Lernen im Gestaltkreis geschieht nicht durch Selektion des Unzutreffenden auf dem Wege des Ausschlusses, sondern 44 unter positiver Präzisierung des Verhältnisses, das sich in der Subjekt-Umwelt-Beziehung immer neu gestaltet. Technische Systeme lernen zumeist nach dem Prinzip der Minimalisierung des maximalen Mißerfolgs, um so jedes Risiko des Irrtums so weit als möglich auszuschließen (Minimax-Prinzip). (Automaten können jedoch auch zum Lernen außer einer solchen ‹reinen› Strategie gemischte Strategien verwenden, Strategien also, die mehrere reine Strategien miteinander verbinden und sie mit gewisser Wahrscheinlichkeit anwenden. Die Darstellung und kritische Analyse solcher gemischter Lernstrategien würde jedoch den Rahmen unserer Abhandlung sprengen.) Biologische Systeme können außer nach dem Minimaxprinzip auch in der ‹Überschau› des als Einheit erfahrenen Subjekt-Umwelt-Bezuges lernen. Die elektronische Schildkröte W. G. WALTERS weicht Hindernissen aus, indem sie sich ihnen wieder nähert und dann zurückweicht, eine ‹echte› Schildkröte könnte in der Erkenntnis der Subjekt-Objekt-Relation das Hindernis als Ganzheit (in. seiner ‹Gestalt›) überschauen und danach ihren Weg einrichten. Es wird wohl keine Maschine geben, die ihr zukünftiges Verhalten gemäß dem Erfolg oder Mißerfolg früheren Verhaltens

modifiziert, wenn das Verhalten durch gestalthafte Eindrücke bestimmt ist. In diesem Zusammenhang wird oft die Lernmatrix K. STEINBUCHS strapaziert. Sie hat jedoch nicht die Aufgabe tierisches Lernen zu simulieren, sondern dem Menschen bestimmte mechanisierbare Lernprozesse abzunehmen. Doch auch hier wird der charakteristische Unterschied zwischen ‹technischem› und biologischem Lernen deutlich. Biologisches Lernen beruht nicht nur auf Informationsvermehrung, sondern leistet selbst ein Mehr an Information. 3. Der Regelkreis funktioniert stets ‹logisch› und kann nur seiner Logik entsprechend geordnete logische Informationen verarbeiten. So kann ein Automat auf Mehrdeutigkeiten (Äquivokationen...) nicht sinnvoll reagieren. Er kann sie bestenfalls, wie es etwa Übersetzungsautomaten tun, hintereinander ausdrucken. In Gestaltkreisabläufen sind dagegen Mehrdeutigkeiten, ja antilogische Informationen, durchaus verarbeitbar, d. h. ein biologisches System kann sich jeweils auf eine höher-wertige Logik einrichten als die, für die es an und für sich eingerichtet ist. Während menschliches Denken in der Regel zweiwertiger Logik gehorcht, kann es dennoch einen Informationsgehalt sinnvoll verarbeiten, wenn diese Verarbeitung eine höherwertige Logik voraussetzt. Auch die Autoren, die sich dem DIAMAT verpflichtet wissen, lehnen in der Regel eine rein kybernetische Interpretation des Lebendigen ab. Zwar kann die Vielheit der Regelkreise in einem lebenden Gebilde 45 einen biologisch optimalen Zustand einregulieren, der begrenzten Schwankungen aus der Umwelt zum Trotz aufrecht erhalten wird, doch sind «Lebewesen... so komplizierte Systeme, daß einfache Regelkreise allein ... das innere Milieu nicht konstant halten können. Lebewesen sind sogenannte ultrastabile Systeme mit der Fähigkeit, trotz vielfältiger ... Störungen ein bestimmtes optimales biochemisches Fließgleichgewicht aufrecht erhalten zu können. Die ultrastabilen Systeme enthalten neben den notwendigen Rückkopplungsmechanismen noch sogenannte Stufenfunktionen, die mehrere Variable enthalten». Doch werden auch solche Stufenfunktionen als rein materielle ‹Schaltmechanismen› interpretiert. Der eigentliche Unterschied wird natürlich in der Lebensbestimmung Engels gesehen: «in den Organismen gibt es keine elektronischen, sondern spezifisch biochemische Steuer- und Regelmechanismen.» Wenn es dann weiter heißt «selbstverständlich soll ... nicht gesagt werden, daß kybernetische Erkenntnisse allein genügen, um biologische Prozesse zu verstehen» (O. Klohr, Moderne Naturwissenschaft und Atheismus, Berlin 1964, 149-151), wird man sich fragen müssen, worin nach Auffassung des Autors denn der Unterschied zwischen technischen und biologischen Funktionen besteht. Der Autor verzichtet darauf, das näher auszuführen. Wir haben im Vorhergehenden versucht, einige wichtige mechanistische Theorien über die Natur des Lebendigen vorzulegen und zugleich das Ungenügen des Ansatzes einer statischen präformierten Struktur zur Erklärung dessen, was Leben ist, hingewiesen. Daraus ergibt sich als notwendige Konsequenz die Einführung des Begriffs der dynamischen Struktur. Eine solche Struktur ist an sich denkbar als sich selbst formierend und transitorisch und «somit labil fixiert bzw. stabilisiert» (Th. Ballauf, Das Problem des Lebendigen, Bonn, 1949, 13). Solche dynamischen Strukturen sollten bestehen können, ohne ein spezifisch immaterielles Prinzip, wie es vorn Vitalismus gefordert wurde und wird. 46 DRITTES KAPITEL
VITALISMUS
These: Das Lebewesen ist eine Einheit aus Materie und einem immateriellen Lebensprinzip. Der Vitalismus hat seinen Ursprung im aristotelisch-scholastischen Hylemorphismus, seiner Lehre von metaphysischer Materie und Form, die alles materielle als deren innere ‹Ursachen› aufbauen (vgl. WS I, 23 f)! Die Lehre von der Seele, die nach ARISTOTELES alles Lebende durch-formt, sei hier kurz wiedergegeben: «Die Seele ist die erste Entelechie (πρώτη εντελέχεια) eines natürlichen Körpers, der das Vermögen besitzt zu leben» (de anima II, 1; 412 a 27) oder «die erste Entelechie eines Organismus (σώµα φυσικόν οργανικόν)» (ibd. 412 b 4). Als Entelechie des Leibes ist die Seele Form (µορφή) (ibd. 4142), Bewegungsgrund und Zweck des Ganzen. Der Leib ist um der Seele willen da (de part, anim. I, 5; 645 b 14 ff). Der Begriff der Entelechie wurde von H. DRIESCH (Philosophie des Organischen, Leipzig 4i928) wieder aufgegriffen. Er verstand darunter einen immateriellen in den Raum hineinwirkender Naturfaktor, dessen Zusammenwirken mit den physiko-chemischen Naturkräften er experimentell aufzuweisen suchte. Genauerhin spricht er der Entelechie die Steuerung der ontogenetischen Formbildung zu, während er bei den Lenkungen der organismischen Handlungen von einem «Psychoid» spricht. DRIESCH hat sich bemüht, ausgehend von den Erscheinungen der Regulation, der Vererbung, der Aktion und der Insertion, den Aufweis für die Autonomie des Organismischen zu erhärten: 1. Im Gegensatz zu technischen Einheiten vertragen die biotischen harmonisch aequipotentiellen Systeme nicht nur nach Art und Ort beliebige Entnahme von Elementen, sondern auch Zerrungen und Verlagerungen der bleibenden (a. a. O. 124). 2. In der Vererbung erweist sich die Mitose (Zellkernteilung nach erfolgter Chromosomenverdoppelung) als typisch biologischer Vorgang. Eine Maschine besitzt keine Struktur und keinen Mechanismus, der es ihr erlaubte, sich beliebig oft zu teilen (ibd. 177). 3. Die Besonderung der Aktion des Organismischen hängt von der Besonderheit aller sich auf Empfindung und Bewegung beziehender Reize ab, die den Organismus in der Vergangenheit betroffen haben.

47 Die historische Reaktionsbasis ist eine der Hauptkomponenten jeder biologischen Aktion (ibd. 243). Das zweite Charakteristikum jeder biologischen Aktion sieht er darin, daß sie stets eine einem individualisierten Reiz entsprechende Reaktion ist. Reiz und Effekt sind individuell einander zugeordnet (ibd. 248 f). 4. Die Unmöglichkeit, harmonische und komplexe Aequipotentialität mechanistisch auszuerklären, gipfelt in der Lokalisations- bzw. der Insertionsproblematik. Daß sich bei Transplantaten eine ortsbestimmte Funktion einstellt, fordere die Einführung eines neuen Kausalfaktors, der Entelechie. Insofern die Entelechie ein räumlich und zeitlich ordnendes Prinzip ist, das in Analogie zur psychischen Produktivität des Menschen (Maler gestaltet ein Bild durch räumliche Anordnung von Farben...) verstanden werden kann, spricht DRIESCH der Entelechie einen seelenartigen, psychoiden Charakter zu [Studien zur Theorie der organischen Formbildung, Acta biotheoretica 3 (1937), 72]. 5. Das Sterben des Lebewesens ist nach DRIESCH zu interpretieren als das Enden der Kontrolle, welche die individuellen aus dem überpersönlich Vitalen herausgesproßten Portionen des vitalen Agens ausübten. Wo aber kommt sie her, die Entelechie, das vitale Agens? Aus dem überpersönlich Vitalen? Wo geht sie hin? (Biologische Probleme höherer Ordnung, Leipzig 1941, 50 u. o.). Auf diese Fragen, die sich aus der «Theorie der entelechialen Autonomie» ergeben, fand DRIESCH keine zufriedenstellende Antwort, scheint doch die Annahme eines überpersonal Vitalen oder der Unsterblichkeit der Einzelentelechien kaum zu befriedigen. Der Theorie von der entelechialen Autonomie haben sich verschiedene Naturphilosophen und Biologen angeschlossen: A. WENZL, J. VON UEXKÜLL (Theoretische Biologie, Berlin 1928), O. FEYERABEND (Das organische Weltbild, Berlin 1933), R. WOLTERECK, H. CONRAD-MARTIUS. Alle haben aber charakteristische Modifikation angebracht. WENZL nimmt einen unbewußt wirkenden Handlungsplan an, den er als «psychischen Faktor» wirksam sein läßt. Er konstruiert ein Stufen-reich von Entelechien; die Entelechie des Leibes ist nicht individuell, sondern Artentelechie, diese wiederum Besonderung von Gattungsentelechien. Über allen steht die Entelechie aller Entelechien (Philosophie als Weg von den Grenzen der Wissenschaften an die Grenzen der Religion, Leipzig 1939, 93). Über die Natur der Materie schreibt er, daß sie «ihrem inneren Wesen nach ein potentieller und aktueller Wille der Natur zur extensiven Äußerung ihres intensiven und qualitativen Innen» sei (Metaphysik der Biologie heute, Leipzig 1938, 98). Sie kann 48 wegen ihrer Bezüge zum Leben nicht lebensfremd, nicht tot sein (ibd. 85). Die Entelechie dagegen «muß ihrem" Wesen nach Wille zur Verwirklichung besonderer Lebensideen sein, der sich konkret in Individuen erfüllt, eingebettet in den allgemeinen umfassenden des Lebens, der sich in der Bildung der Arten erfüllt» (ibd. 130). UEXKÜLL hat besonders das Problem der Gerichtetheit (wohl ein besserer Terminus als Planmäßigkeit oder Zielstrebigkeit, die recht anthropomorph anmuten) beschäftigt. Eine wichtige Rolle in seinen Überlegungen spielt u. a. der ARNDT-Effekt [A. Meyer-Abick und E. Arndt, Der Arndteffekt und seine Probleme, Z. f. d. gesamte NW 1 (1935/36), 457-470]: Im Zeitrafferfilm zeigt die Entwicklung eines Schleimpilzes (Dictyostelium) einen eigenartigen Effekt. Die anfangs regel- und ordnungslos wuchernden Einzelzellen geraten plötzlich in eine heftige kollektive Bewegung, ordnen sieh in Zügen und bilden ein zentralisiertes Plasmodium (vielkerniger Protoplasmakörper) aus, der von diesem Zeitpunkt an einer eigenen Gesetzmäßigkeit gehorcht. Es scheint also ein den Einzelteilchen übergeordnetes einheitliches Prinzip zu geben, das dann in keiner Weise mehr mechanistisch gedeutet werden kann. Besonders fruchtbar aber wurde UEXKÜLLS Umwelttheorie, nach der das Tier als autonomes Subjekt in einer von ihm selbst erschaffenen Umwelt aus Raum, Zeit und Inhaltsqualitäten steht. Jedem Lebewesen ordnet er eine ganz bestimmte ‹Welt› zu, die es sich selbst aus den erlebten Funktionen aufbaut. «Alles und jedes, das in den Bann der Umwelt gerät, wird umgestimmt und umgeformt, bis es zu einem brauchbaren Bedeutungsträger geworden ist, oder es wird völlig vernachlässigt.» Die Bedeutung erwächst aus der Tätigkeit, dem tätigen Umgang mit der Umwelt (Bedeutungslehre, Leipzig 1940). So kann für den Einsiedlerkrebs eine Seeanemone ein Schutzkumpan oder ein Beute-tier sein. Die Bedeutung wechselt im Wechsel der durch das Handeln bestimmten Situation. Brechen wir.an dieser Stelle einmal unsere Darstellung des Vitalismus ab, obschon noch zahlreiche Autoren und zahlreiche interessante Hypothesen zu nennen wären. Eine gute und umfassende Übersicht gibt Tu. BALLAUF (Das Problem des Lebendigen, Bonn, 1949). Viele namhafte Biologen und Philosophen halten das Vitalismus-Mechanismus-Problem nach dem heutigen Stand der Forschung für unentscheidbar. So hat E. UNGERER (Der Sinn des Vitalismus und des Mechanismus in der Lebensforschung, Leipzig 1927) darauf hingewiesen, daß die Autonomiebeweise DRIESCHS nur eine gute Wahrscheinlichkeit, nicht aber völlige Sicherheit für sich haben, da ihnen eine un- 49 abgeschlossene Theorie der Materie zugrunde liegt. Es ist nicht entscheidbar, ob es sich bei der Aufstellung DRIESCHS um eine Kennzeichnung oder eine eigentliche Erklärung des Lebendigen handelt. Auch L. VON BERTALANFFY [Zur Theorie der organ. ‹Gestalt›, Arch. Ent. Mech. 108 (1926)] versucht durch die Begründung einer ‹exakten Biologie›, die auf der Erkenntnis der «ganzheitlichen Eigengesetzlichkeit» des Organismus aufbaut, die Frage nach der Gültigkeit des Vitalismus offen zu lassen. Der Streit zwischen Vitalismus und Mechanismus ist für die meisten Biologen heute Geschichte. Es ist ein überholter

Streit, der auf falschen wissenschaftstheoretischen Voraussetzungen beruht. So sind denn im Laufe der letzten Jahrzehnte Theorien «jenseits von Vitalismus und Mechanismus» ausgebildet worden. VIERTES KAPITEL
DER HOLISMUS
These: Das Lebewesen ist eine materielle Einheit, ausgestattet mit einem spezifisch materiellen Prinzip, (einem nicht physikalischen Naturfaktor). Die Naturgegebenheiten zeigen eine Schichtung, derart, daß jede höhere Stufe durch einen Komplex von Sondergesetzen bestimmt wird. Zumeist wird zur Aufhellung dieser Schichtung der Weg von unten nach oben begangen. E. OLDEKOP (Über das hierarchische Prinzip in der Natur, Reval 1930) betont ausdrücklich, daß die jeweils höheren Stufen nicht auf ihre Komponenten zurückführbar seien. A. MITTASCH (Über katalytische Verursachung im biologischen Geschehen, Berlin 1935 u. ö.) nimmt eine biologische Ganzheitskausalität an, die sich als Führungskausalität unter Beteiligung des «Ganzen» einfüge. Das Eingreifen höherer Kausalitäten wird ermöglicht durch Unbestimmtheiten der Kausalitäten niederer Stufe. R. WOLTERECK (Allgemeine Biologie, Stuttgart 21940; Ontologie des Lebendigen, Stuttgart 1940) führt eine Vielheit «dynamischer Grund-mächte» ein, die gemäß «geltenden So-Determinanten» oder «Ideen» wirken. Dabei kann Leben aber sehr wohl als Steigerung aus Nicht-Leben hervorgegangen sein. Von philosophischer Seite versuchten vor allem N. HARTMANN (Der Aufbau der realen Welt, Berlin 1940) und W. BURKAMP (Wirklichkeit und Sinn, Berlin 1938) die Ganzheitsstufung der Wirklichkeit zu begründen. Den entgegengesetzten Weg, den Weg «von oben her» schlägt der Holismus ein. Er wurde von J. S. HALDANE (1907) und J. C. SMUTS (1925) im englischen Sprachraum begründet und von A. MEYER-ABICH im deutschen Sprachgebiet entwickelt. Ursprünglich stand bei Meyer-Abich der Gedanke im Vordergrund, die Unableitbarkeit verschiedener Wissenschaftsbereiche — insbesondere der der Physik und Biologie -nicht durch eine Physikalisierung, sondern durch eine Biologisierung aufzuheben. Physikalische Gesetze möchte er durch «Simplifikation» biologischer erklären. Doch wird dieser Versuch heute allgemein abgelehnt. Der Holismus war also anfangs eher eine (unzureichend begründete und begründbare) Wissenschaftstheorie als ein Versuch, das Wesen des Lebendigen zu verstehen. Was aber kann er zum Wesen des Lebendigen sagen? 51 «Die Natur ist keine Einheit, sie ist auch keine Vielheit, sie ist vielmehr eine echte organische Ganzheit» (Naturphilosophie, Stuttgart 1948, 43), sie ist «ein mächtiger lebendiger Weltorganismus» (ibd. 44). Nachdrücklich lehnt Meyer-Abich jeden Mechanismus ab. Es gibt eine typische biologische Energie (ibd. 132, 190) und eine typische biologische Kausalität (ibd. 160), die mit ihren mechanischen (physikalischen) Entsprechungen ontologisch nichts gemein haben. Andererseits lehnt er aber auch jeden Pluralismus, wie er durch die Einführung eines dem materiellen gegenübergestellten immateriellen Lebensprinzips erstellt wird, ab. Nach den Aufstellungen Meyer-Abichs würde sie die eine Welt in ‹geistige› und materielle Bereiche unterteilen. Er ist aber durchaus bereit, die aristotelische Entelechie als Ganzheitsprinzip gelten zu lassen (ibd. 104), Von BICHAT übernimmt er die Bestimmung: «Das Leben ist das Beieinander aller derjenigen Funktionen, die dem Tode widerstehen» (ibd. 125). Wir halten dafür, daß der Holismus als Versuch, den Mechanismus-Vitalismus-Gegensatz zu überwinden, auf einem Mißverständnis beruht. Genauerhin auf einem Mißverständnis des Vitalismus. Es sind neben geistigen (= materiefremden) und materiellen Wirklichkeiten immaterielle (materiegebundene) Prinzipien durchaus denkbar. Damit wollen wir unsere Überlegungen zur Geschichte des Problems, was Leben denn eigentlich sei, abbrechen und uns der von uns entwickelten Theorie zuwenden. 52 FÜNFTES KAPITEL
EINE INDIVIDUUM-THEORIE DES LEBENDIGEN
Wenn wir uns im folgenden für die Theorie entscheiden, nach der eine biologische Einheit, insofern sie eine biologische Einheit ist, ein Individuum sein kann, sind wir uns darüber im klaren, daß der Begriff des Individuums einer der verschwommensten Begriffe der Biologie und Naturphilosophie überhaupt ist. Wir werden weiter unten also genau zu erklären haben, was wir unter dem Begriff Individuum verstehen wollen. Dabei werden wir nicht die übliche Methode verwenden, zunächst eine Definition anzubieten und dann die biologischen Tatsachen i n diese Definition zu pressen, sondern wollen umgekehrt zunächst einmal ein möglichst breites Spektrum von Eigenschaften, die biologische Einheiten von physikalischen unterscheiden, vorstellen, um dann eventuell gesicherte Differenzen zu dem Begriff ‹Individuum› zusammenzufassen. Der Begriff Individuum wird von uns also als eine Chiffre für eine Summe von spezifischen Eigenschaften biologischer Einheiten eingeführt, die es uns erleichtert, uns zu verständigen, insofern wir nicht immer den ganzen Katalog der Differenzen aufzählen müssen. Individuum sei der Name für den noch zu erstellenden Katalog. Dabei haben wir natürlich anzugeben, warum wir gerade diesen historisch durchaus belasteten Namen wählen. Nun, das hat noch Zeit. Zunächst seien hier einmal die in der Biologie üblichen und in der

Naturphilosophie gebräuchlichen Definitionen des Begriffes Individuum vorgestellt.
1. Zum Begriff «Individuum» Wir folgen dabei in wesentlichen Punkten einer guten Arbeit von E. MAYER [Das Problem der Individualität im organischen Sein in WissWeis 30 (1967), 130-153 und 226-237)]. a) G. VON NATZMER definiert: «Dem reinen Wortsinne nach meint (der Begriff) ein Lebensganzes, das unteilbar ist und sich in Raum und Zeit sowie auch dynamisch (d. h. in allen Daseinsäußerungen) von der gesamten Umwelt und damit auch von allen anderen ‹Individuen› abhebt und unterscheidet» (Individualität in Z. f, ges. NW 1935, 305). Die Problematik dieser Definition scheint uns im Wort unteilbar zu liegen, wie noch gezeigt werden wird. L. von bertalanffy schreibt: 53 Wenn wir sagen, daß ein Fisch, ein Hund oder ein Mensch ein «Individuum» sei, meinen wir damit ungefähr, «daß er ein räumlich, zeitlich und dynamisch von allen unterschiedenes, lebendes Einzelwesen darstellt, das als solches einen bestimmten Lebenszyklus durchläuft» (Das biologische Weltbild I, Bern 1949, 55). Die Schwierigkeit dieser Definition scheint uns in dem Wort Einzelwesen zu liegen. Auch das ist noch zu zeigen. A. PORTMANN meint, «daß Tiere tätige Zentren sind, deren Erleben einen mehr oder minder großen Raum, eine verschieden lange Spanne Zeit in sich einbezieht. Man möchte sagen, daß das Tier sich seine Umgebung einräumt und daß es den Lebenslauf nach seinem Erlebnisrhythmus zeitigt. Raum und Zeit werden so mit vielen Gehalten erfüllte Glieder eines Individuums« [Das Verhalten des Tieres als Objekt der Naturwissenschaft, in Universitas 10 (1955), 277]. C. NÄGELI definiert: «Ein Individuum ist ein zu selbständigem Leben befähigtes ‹Einzelwesen›» und «unter Individuum verstehen wir ... eine einheitliche Erscheinung, welche wir nicht teilen können, ohne ihr Wesen zu vernichten, welche daher ein abgeschlossenes Ganzes mit eigentümlicher Entwicklung und eigentümlichen Beziehungen zur Außenwelt darstellt» (Die Individualität ..., Zürich 1856, 17 und 12) Wie schon NAGELI für die Pflanze, unterscheiden O. HERTWIG, R.HESSE und F. DOFLEIN das tierische Individuum in physiologischer und morphologischer Hinsicht. In physiologischer Hinsicht ist das Individuum nach O. HERTWIG «eine Lebenseinheit, die nach außen abgegrenzt und sich selbst zu erhalten imstande ist, weil sie mit den Grundfunktionen des Lebens ausgerüstet ist, mit der Funktion sich zu ernähren, und zu wachsen, sich fortzupflanzen, gegen Reize der Außenwelt irritabel zu sein und auf sie in verschiedener Art zu reagieren» (Allgemeine Biologie VI und VII, Jena 1923, 489). Das morphologische Individuum definiert er als «Formeinheit, welche zwar morphologisch, d. h. nach Aussehen, Struktur und Zusammensetzung einem physiologischen Individuum gleicht, aber nicht in physiologischer Beziehung». Es ist nicht mehr in der Lage sich selbst zu erhalten und «stellt keine selbständige Lebenseinheit mehr dar; es ist als ein abhängiger Teil in eine höhere physiologische Individualität eingegangen oder, mit anderen Worten, zu einem anatomischen Element von ihr geworden» (ibd. 490). Hesse und Doflein sprechen in diesem Zusammenhang von Individualitätsstufen (Tierbau und Tierleben I, Jena 1935, 41 f). P. JORDAN sieht auch solche Individuenhierarchien, meint dennoch aber, daß der «Zug zur Individuation ... im ganzen Reiche des Lebendigen unverkennbar» hervortrete. «Auch erhält sich deutlich die Unter- 54 teilung aller höheren Lebewesen in ihre ‹Unterindividuen›, die Zellen. Es gibt nun in der ganzen Makrophysik . . . keinerlei Erscheinung, die mit einigem Recht, ohne Willkür und Gedankenzwang, zu dieser biologischen Grunderscheinung in Parallele gestellt werden konnte» (Die Physik und das Geheimnis des organischen Lebens, Braunschweig 1947, 43 f). Der Begriff des Individuums ist jedoch keine Entdeckung der Biologie. Er wurde zuerst von Philosophen aus recht verschiedenen Gründen entwickelt. SENECA formuliert: «Individuen sind zusammenhängende Gebilde, die nicht geteilt werden können» (De prov. 5). BOETHIUS schreibt: «Individuum wird genannt, was in keiner Weise geteilt werden kann, wie Einheit oder Geist, aber auch das, was wegen seiner Festigkeit nicht geteilt werden kann, wie Stahl und das, dessen Bezeichnung den übrigen ähnlichen Seienden nicht zukommt, wie Sokrates» (In Isag.). Die scholastische Philosophie sah vor allem die Unmittelbarkeit, die Unvertauschbarkeit, die Einmaligkeit und die volle Einheit als typische Eigenschaften des Individuums. «Es ist dem Individuum zu eigen, nicht in mehreren sein zu können», «weil die substantielle oder akzidentelle Form (gemeint ist die metaphysische Form) von Natur aus in einem, nicht aber in mehreren sein kann..., weil jene an sich ungeteilt und von allen anderen unterschieden sind» (S. Theol. III, 77, 2), schreibt THOMAS VON AQUIN. Klassisch geworden ist seine Bestimmung: «Das Individuum ist in sich ununterschieden, von allen anderen aber verschieden» (ibd. I, 29, 4 c). Ganz ähnliche Gedanken finden wir auch bei Duns Scotus (vgl. Rep. Id. 25 q. 2 n. 15; In Met. VII q. 13 n. 17). Eine ausführliche Darstellung dieser scholastischen Position bringt E. Mayer (a.a.O. 113-136). Es ist jedoch zu betonen, daß die Scholastiker viel mehr den kategorial-grundsätzlichen Aspekt sehen als ein eigentliches Begreifen dessen versuchen, was Leben des Lebewesens bedeuten möchte. Heute ist diese kategoriale Betrachtung des Individuums als lebendige Einheit weitgehend aufgegeben zugunsten einer strukturanalytischen Sicht (vgl. Th. Ballauf, Das Problem des Lebendigen, Bonn 1949, 12-15 u. ö.). So wollen auch wir denn im folgenden nicht von Substanzen sprechen, wenn wir das Wort ‹biologische Einheit) gebrauchen, sondern von Strukturen, nicht als ob die Substanzphilosophie der Scholastik falsch sei; sie stellt jedoch keinen geeigneten Begriffsapparat zur Verfügung,

um das was biologische Einheit in all der möglichen Mannigfaltigkeit, die dieser Begriff einschließt, auch nur einigermaßen zureichend zu umgreifen. 55 b) Struktur sei hier verstanden als Seinstypos mit einem typischen Geltungsbereich, dem des Lebendigen nämlich. Der Ausgang von der Funktion des Lebendigen führt zum Begriff der Struktur und nicht zum Begriff der Substanz, zumindest nicht mit der gleichen Selbstverständlichkeit. Wir haben einmal gesagt, daß gute Philosophie vom Phänomen ausgehe und nicht von einer mehr oder minder apriorisch entwickelten Ontologie, und dieses Phänomen führt uns zu Strukturen, nicht aber -wenn man auf die Hilfestellung einer den Einzelwissenschaften vorgelagerten Ontologie verzichten will — zu Substanzen. «Der Funktionalismus entsteht als Gegenphilosophie gegen den klassischen Typus der Metaphysik; er etabliert sich über den Trümmern der Substanzontologie. In der geschichtlichen Faktizität stehen beide Grundmöglichkeiten des Denkens .. . gegeneinander . . . Der Funktionalismus kommt - was die Metaphysik nur schwer begreift - ohne Metaphysik aus. Er ist ein sich selbst regulierender Gedanke» (H. Rombach, Substanz, System, Struktur I, München 1965, 49). Für den Funktionalismus ist Wesen kein sinnvolles Wort mehr. «Nicht das, was die Sache ist, sondern das, was sie in ihren Relationen bewirkt, ist allein entscheidend» (Rombach, a.a.O. I, 213). Es wäre nun aber falsch, eine Struktur — uns interessiert hier ausschließlich die biologische - als Summe von Funktionen zu verstehen. Denn eine Struktur ist eigentliche Einheit, zugleich aber in ihrer Funktion Vielheit. Wir wollen also den Begriff der Struktur ‹dialektisch› begreifen. L. W. STERN schreibt einmal: «Jedes Individuum ist etwas Singuläres, ein einzig dastehendes, nirgends und niemals sonst vorhandenes Gebilde. An ihm betätigen sich wohl gewisse Gesetzmäßigkeiten, in ihm verkörpern sich wohl gewisse Typen, aber es geht nicht restlos auf in diesen Gesetzmäßigkeiten und Typen; es bleibt noch ein Plus, durch welches es sich von anderen Individuen unterscheidet, die den gleichen Gesetzen und Typen unterliegen. Und dieser letzte Wesenskern .. . bewirkt, daß das Individuum ein Dieses und ein Solches, allen anderen durchaus Heterogenes vorstellt; er ist in fachwissenschaftlichen Begriffen unausdrückbar, unclassifizierbar, incommensurabel. In diesem Sinne ist das Individuum ein Grenzbegriff, dem die theoretische Forschung zwar zustrebt, den sie aber nie erreichen kann; es ist, so könnte man sagen, die Asymptote der Wissenschaft» (Üb. Psychol. d. indiv. Diff. 1900). Die Abwendung von einem substanzontologischen Begreifen dessen, was Individuum sagt, ist hier schon deutlich ausgesprochen. Bei allem Pathos und aller Metaphysik des Textes enthält er aber auch unser Programm, nur daß wir nicht nach dem «Wesenskern» fragen (ein merkwürdiges Wort, 56 das um so mehr verschwimmt, je genauer man darüber nachdenkt), sondern der spezifischen, der Struktur als in aller Funktion einheitgebenden und -erhaltenden Wirklichkeit nachgehen wollen.
2. Eigenheiten biologischer Einheiten Wir haben im Vorhergehenden schon zahlreiche Versuche erwähnt, das Lebendige vom Nichtbelebten zu unterscheiden. Dabei wurden jedoch in der Regel biologische Einheiten physikalischen Vielheiten entgegengestellt und in ihrer Funktion voneinander unterschieden. Wir wollen einen anderen Ansatz versuchen: Deutlich ist zunächst zu unterscheiden zwischen biologischen Einheiten, technischen Einheiten (Maschinen, Computern .. .) und physikalischen Einheiten (Elementarteilchen im eigentlichen Sinne). Da wir über die Unterschiede der, physikalisch gesehen, komplexen Einheiten Lebewesen und Maschine schon ausführlich gehandelt haben, bleibt uns hier die Gegenüberstellung von physikalischen und biologischen Einheiten. Wie unterscheiden sie sich? a) Sie unterscheiden sich stets wenigstens numerisch voneinander «Einheiten» des materiellen Bereichs sind bestenfalls numerisch voneinander unterschieden, das soll heißen, bei einer Abzählung von etwa loo Elektronen unterscheidet sich das eine Elektron von einem anderen allenfalls durch die Numerierung, die es beim Durchzählen erhält. Im übrigen sind Elektronen durch ein und dieselbe Gleichung (etwa eine Wellengleichung) voll ausbeschrieben. Diese numerische Unterscheidung ist aber auch nur dann möglich, wenn die 100 Elektronen während des Zählprozesses bis hin zum Augenblick, in dem wir eine numerische Identifikation eines durchgezählten Elektrons versuchen, nicht miteinander wechselwirken. Zwischen wechselwirkenden Elektronen gilt die Unschärfebeziehung ∆ε · ∆ n › h : 2π (ε = Elektrische Feldstärke, n = Zahl der Elektronen). Versucht man also eine Messung der elektrischen Feldstärke des physikalischen Systems, in das 100 geeinzelte Elektronen hineingezählt wurden, die nun miteinander wechselwirken, und bemüht sich, den Meßfehler auf 0 herabzudrücken, wird die Zahl der Elektronen völlig unbestimmt. Wir haben uns an anderer Stelle für die Unbestimmtheit als ontologische Basis der Unschärfebeziehung entschieden (WS I, 152 f.). Das aber bedeutet, daß bei unserem Meßprozeß die Anzahl der Elektronen nicht nur unbestimmbar, sondern unbestimmt ist. Man kann also nicht mehr legitim aussagen, daß 10, 50 oder 100 57 Elektronen miteinander wechselwirken, weil keine bestimmte (und nicht nur keine bestimmbare) Zahl von Elektronen vorhanden ist! Nach den bekannten Sätzen von Spin (vgl. WS I, 148) und Statistik, gehorcht ein System von Teilchen mit

halbzahligem Spin der Fermi-Dirac-Statistik, ein Teilchen mit ganzzahligem Spin der Bose-Einstein-Statistik. Das Gemeinte wollen wir an einem einfachen Beispiel erklären: Es gelte zwei Einheiten in zwei «Feldern» anzuordnen. Nach der klassischen Statistik sind folgende vier Verteilungen möglich:
Lebewesen gehorchen wie alle makrophysikalischen Gebilde der klassischen Statistik. Gemäß der Bose-Einstein-Statistik sind - wegen der Ununterscheidbarkeit der Teilchen - nur drei Verteilungen möglich:
Zu den sogen. Bosonen zählen Photonen (Spin — 1) und Mesonen (Spin = o) sowie die hypothetischen Gravitonen (Spin = 2). Die Fermi-Dirac-Statistik fügt zu dem Moment der Ununterscheidbarkeit noch das Pauliprinzip hinzu, nach dem die die Teilchen bestimmende Wellenfunktion gegenüber der Vertauschung von Raum-, Spin- und Ladungskoordinaten (als innere Variablen) antisymmetrisch ist. Aus dieser Antisymmetrie folgt, angewandt auf unser Beispiel, daß jedes Teilchen nur verschiedene Orte besetzen kann:
Zu den Fermionen zählen Neutrinos, Leptonen (Elektronen und Myonen), Nukleonen und Hyperonen (Spin in den meisten Fällen 1/2)· Es gibt also kein System gleichartiger Elementarteilchen, das der klassischen Statistik gehorcht. b) Sie unterscheiden sich funktionell von ihrer Umgebung Im Gegenteil zu biologischen Einheiten unterscheiden sich physikalische Einheiten nicht scharf von ihrer Umgebung. Sie sind sogar mit ihrer Umgebung strukturell verbunden, während biologische Einheiten keine 58 strukturelle Einigung mit anderen biologischen Einheiten eingehen, sondern allenfalls funktionale. Der materielle Kosmos bildet eine substantielle Einheit, d.h. alle reellen Einheiten sind durch virtuelle Einheiten wie durch strukturelle Brücken miteinander verknüpft. Virtuelle Elementarteilchen (vgl. dazu WS I, 157) werden auf Grund virtueller Emission von reellen Elementarteilchen erzeugt. Solche virtuellen Prozesse sind physikalisch reale Vorgänge, für die der Energieerhaltungssatz nur unter Berücksichtigung der Unschärfebeziehung gilt. Wenn etwa ∆ E die Energie ist, die ein Neutron aufbringen müßte, um ein Meson zu erzeugen, dann ist trotz fehlender Energie diese Erzeugung möglich, wenn das Meson nach spätestens A t = (h : 2π · ∆ E) ~ 10-23 sec wieder durch Absorption «vernichtet» wird. Solche Prozesse sind natürlich — das liegt im Wesen der Unschärfebeziehung - nicht unmittelbar beobachtbar, können aber durch ihre Effekte (z. B. den Lambshift, d. h. eine Verschiebung der Energieterme eines physikalischen Systems) nachgewiesen werden. Da nun in der quantenphysikalischen Beschreibung (durch operatorenmechanische Zustandsfunktionen etwa) für virtuelle Teilchen die gleichen mathematischen Ausdrücke (vor allem Erzeugungs- oder Vernichtungsoperatoren) wie bei realen Teilchen auftreten, dürfte es gerechtfertigt sein, auch virtuelle Teilchen als physikalisch wirklich› zu betrachten. Der Sachverhalt der virtuellen Emission erlaubt nun zwei Deutungen: Entweder erlangt das virtuelle Teilchen einen eigenen Selbstand gegenüber dem emittierenden Teilchen oder es löst sich nicht eigentlich von dem emittierenden und geht auch nicht vollständig in das absorbierende ein, so daß es zwischen beiden so etwas wie eine materielle Klammer herstellt. Gegen die erstgenannte Möglichkeit spricht u. a., daß die Beugungsbilder von Röntgenquanten an Kristallgittern nur zu erklären sind, wenn das Quant mit allen Atomen des Gitters wechselwirkt. Diese Wechselwirkung erscheint im Formalismus als virtuelle Absorption und Emission des Quants durch die Gitteratome. Wäre eine solche Absorption (und damit auch die nachfolgende Emission) vollständig», konnte das Quant beim Durchgang durch das Gitter nur mit einem einzigen Gitteratom wechselwirken, da sich der Interferenzeffekt in quantenphysikalischer Beschreibung in einer einmaligen Absorption und Emission ausdrückt. Für die zweite Möglichkeit spricht das Auftreten von Übergangsstadien teilweise Absorption und Emission) beim Entstehen auch realer Teilchen. So benötigt etwa ein Atom (bei monochromatischer Abstrahlung) für die Aussendung eines Photons von einer Wellenlänge von i bis 3 m etwa 10-8 sec. Diese Wellenlänge ist immerhin so real, daß man Anfang und Ende eines ihr zugeordneten Wellenzuges mit- 59 einander zur Interferenz bringen kann. Wenn aber die Existenz eines Längenanalogons im Ansich gesichert ist, dann auch die des Zeitanalogons (wir haben in diesem Zusammenhang von mikrophysikalischer Realzeit gesprochen; vgl. WS II, 291 f.). Das aber bedeutet, daß es ein reales Übergangsstadium gibt, in dem das Quant teilweise emittiert oder absorbiert ist. Übrigens wäre bei Leugnung solcher Übergangszustände die Zahl der Feldquanten stets eindeutig bestimmt, was der schon erwähnten quantenphysikalischen Überlegung widerspricht, nach der (als Folge eines virtuellen Feldquantenaustauschs) die Zahl der Quanten unbestimmt ist. Virtuelle Teilchen bilden also materielle Brücken zwischen den realen Teilchen. Da nun aber der virtuelle Photonenaustausch ein quantenphysikalisches Charakteristikum des elektromagnetischen, der virtuelle Mesonenaustausch eines des Kernkraftfeldes und der virtuelle Gravitonenaustausch eines des Schwerefeldes ist,

andererseits aber alle elektrisch geladenen Teilchen durch elektromagnetische Felder und alle ‹schweren› Teilchen durch Gravitationsfelder verbunden sind, sind somit alle realen Teilchen durch materielle Brücken miteinander verknüpft. Man mag den materiellen Kosmos mit einer Glasschmelze vergleichen. Die festen Teile (entsprechend den realen Elementarteilchen) sind materiell verbunden durch die Schmelze (entsprechend den virtuellen Teilchen; WS II, 379 ff.). Im Gegensatz dazu sind biologische Einheiten als solche (nicht also in ihrer Materialität betrachtet, sondern in ihrer spezifischen «Biologität») nicht strukturell miteinander verschmolzen. Exkurs: Formen biologischer Einheiten In diesem Zusammenhang tauchen eine Reihe von Problemen auf, die mit der Bestimmung dessen zusammenhängen, was denn eigentlich eine biologische Einheit sei. Sicher ist damit nicht immer und mit Notwendigkeit ein Lebewesen gemeint, denn u. a. bilden mehrere Lebewesen oft zusammen eine spezifische biologische Funktionseinheit (und von solchen mit den eingangs genannten biologischen Funktionen ausgestatteten Einheiten sprechen wir hier). Wir haben bei der Behandlung des ARNDT-effektes schon ein solches Beispiel kennen gelernt, doch müssen wir uns an dieser Stelle noch mit einer Reihe anderer Fälle beschäftigen.
aa) Kolonialindividuum Den Begriff Kolonialindividuum übernehmen wir von A. KAESTNER und wollen darunter feste Verbände einzelliger Lebewesen verstehen, die mit Leistungsspezialisierungen und wenigstens partieller Aufgabe 60 der Selbständigkeit der Einzel«lebewesen» ausgestattet sind. In solchen Kolonien gibt es Ernährungszellen, Fortpflanzungszellen .. . Keine Zelle kann schließlich längere Zeit außerhalb der Kolonie leben (vgl. Lehrbuch der speziellen Zoologie I, i; Stuttgart 1954/55, 23). Das Wort Individuum sei hier noch ganz und gar unthematisch gemeint und mit keiner genaueren Definition bedacht. Einige Kolonialindividuen seien vorgestellt. Pleodorina. Die Kolonie besteht aus 32 Mitgliedern, von denen 28 teilungs- und damit fortpflanzungsfähig sind und 4 teilungsunfähigen Mitgliedern, die sonst alle Zellfunktionen zeigen, aber nach der Auflösung der Kolonie in Tochterkolonien zugrunde gehen (vgl. R. Lotze und H. Sihler, Das Weltbild der Naturwissenschaften, Stuttgart 1954, 264). Volvox globator. Das Kugeltierchen, wie es uns oft in heimischen Tümpeln begegnet, ist eine Kolonie aus bis zu 30 000 unselbständigen Mitgliedern, die regelmäßig in eine gallertartige Wand einer mit Flüssigkeit gefüllten Hohlkugel eingelagert sind. Die Einheit der Kolonie sichern fadenförmige Plasmabrücken, die den Austausch von Stoffen und die Reizleitung von Mitglied zu Mitglied sowie die Koordination der Bewegung der Geißeln der einzelnen Mitglieder besorgen, so daß eine geordnete Gesamtbewegung zustande kommt. Die Arbeitsteilung der einzelnen Mitglieder ist sehr deutlich ausgeprägt: Es gibt Zellen, die für Vermehrung, andere die für Ernährung und Bewegung zuständig sind. Die Mitglieder der Kolonie, die in der Nähe des voranschwimmenden Pols liegen, haben recht große «Augenflecken» (optische ‹Organe›), während die anderen Mitglieder diese Organellen kaum entwickeln. Die für die Vermehrung sorgenden Mitglieder der Zellkolonie werden deutlich großer als die anderen und wandern im Inneren der Kolonie. Hier entwickeln sie neue Tochterkolonien oder wandeln sich zu verschieden gearteten Geschlechtszellen, den Makrogameten (große einzellenähnliche Gebilde) und den Mikrogameten (durch Geißeln frei beweglich spermienähnliche Gebilde) um. Durch Verschmelzung eines Makro- und eines Mikrogameten wird der Grundstock zu einer neuen Kolonie gelegt. Sind eine bestimmte Zahl von solchen Tochterkolonien entstanden, platzt die Mutterkolonie, gibt ihre Töchter frei und geht selbst zugrunde (vgl H. Linder, Biologie, Stuttgart 141964, 12-14). Zoothamniiim arbuscula. Dieses Kolonialindividuum hat baumähnliche Gestalt und besteht aus zwei gestaltmäßig (topologisch) unterschiedenen Mitgliedern. Die Größeren haben einen weitgehend zurückgebildeten Wimpernapparat, der es ihnen nicht erlaubt, sich selbst zu ernähren. Sie müssen über Plasmabrücken von den Kleineren miternährt 61 werden. Die großen Zellen können jedoch ausschwärmen und neue Kolonien bilden (A. Kaestner, a. a. O. 73 f.). Die biologische Einheit ist das Kolonialindividuum selbst. Genauerhin müssen wir zwischen physiologischer und morphologischer Einheit unterscheiden. Die eigentliche biologisch-strukturelle (physiologische) Einheit setzt sich aus mehreren morphologischen (= Funktions-)Einheiten zusammen. Der Unterschied von Kolonie und klassischem Organismus ist jedoch dabei deutlich gewahrt, insofern viele Mitglieder einer Kolonie eine Selbständigkeit besitzen, die wir im Bereich von Zellen, die einen Organismus aufbauen, nicht kennen.
bb) Zellfusionen Auch hier schließen sich Zellen zu einem relativ lockeren Zellverband zusammen. Eine solche Zellfusion verhält sich zu einem Kolonialindividuum etwa wie ein Staatenbund mit weitgehender Integration zu einem Bundesstaat. Die Einheiten bestehen aus einer Plasmamasse verschiedener Größe, in der bis zu vielen Hundert Kerne in regelmäßigen Abständen eingelagert sind. Hier seien vor allem als Beispiel bestimmte Entwicklungs-Vorgänge bei den schon erwähnten Schleimpilzen erwähnt: Aus deren Fortpflanzungskörpern schlüpfen amöbenähnliche Wesen, die sich

kaum oder garnicht von einzelligen Tieren unterscheiden lassen. Jedes führt ein räumliches Sonderdasein. Von einem bestimmten Zeitpunkt an beginnen die sich turbulent bewegenden Tierchen zu ordnen, schließen sich immer enger zusammen, bis sich schließlich die gesamte Ordnung zu einer festen einheitlichen Plasmamasse, dem eigentlichen Schleimpilz, zusammenfindet, der dann neue Fruchtkörper auszubilden beginnt (vgl. G. van N atmer, Individualität, in Z. f. gesam. NW, 1935, 31 o). E. Mayer führt als Beispiel Condrioderma difforme an, bei dem sich kleine amöbenartige Zellen in großer Anzahl in ihrem Plasma - bei Bestand der Kerne - vereinen (a. a. O., 142). Ähnliches soll auch gelegentlich bei Sonnentierchen beobachtet worden sein: zwei bis 20 einzelne Tiere verschmelzen in ihrem Plasmaanteil. Treten besondere Umstände auf, ist eine Trennung nicht ausgeschlossen. (L. Seitz, Die Wirkungseinheit des Lebens, Berlin und München 1950, 9). Die Einheit solcher Zellfusionen ist lockerer als bei Zellkolonien. Sicherlich handelt es sich bei den einzelnen Zellen um morphologische Einheiten. Die Frage, in welchem Umfang man von physiologischen Einheiten sprechen kann, ist umstritten (E. Mayer, a. a. O. 143). Uns scheint jedoch nicht einsichtig zu sein, warum man nicht von einer strukturellen 62 biologischen (physiologischen) Einheit auf Zeit, die gegebenenfalls wieder zerbricht, sprechen sollte.
cc) Klassische Organismen Unter klassischen Organismen seien hier mehrzellige Gebilde verstanden, deren Zellen wenigstens teilweise funktional verschieden sind, wobei die Selbständigkeit der Zellen fast gänzlich für dauernd aufgegeben ist, so daß sie natürlich für sich nicht mehr lebensfähig sind (und somit auch keine eigentlichen morphologischen Einheiten mehr darstellen), sondern allenfalls künstlich (in vitro), d. h. unter Bedingungen wie sie in der Natur äußerst unwahrscheinlich sind, lebendig erhalten werden können. Solche klassischen Organismen sind alle höheren Pflanzen und Tiere. Wir wollen zunächst einmal von der Problematik der in vitro «lebenden» organismischen Zellen oder Zellverbänden absehen, auch seien die Fragen, die sich im Zusammenhang mit der Vermehrung, der Regeneration, der einengen Mehrlinge, der Existenz von Chimären ... für die exakte Fassung des Begriffs der biologischen Einheit stellen, zurückgestellt. Sie werden uns noch beschäftigen. Hier interessiert uns zunächst das Problem von Oberorganismen, das heißt von organismusähnlichen Verbindungen von klassischen Organismen.
dd) Oberorganismen Solche Überorganismen können sich aus zwei oder sehr vielen (bis zu einigen Tausend) klassischen Organismen zu einer biologischen Funktionseinheit zusammenfinden. Wir möchten hier also die biologische Einheit nicht ausschließlich unter dem Vorzeichen streng physiologischer Abläufe sehen, sondern ganz allgemein organismische Funktionen betrachten. Einige einfache, aber sehr verschiedene Beispiele für das Gemeinte seien angeführt. Hierher gehören vor allem einige Formen von Schmarotzertum und Symbiose. Edrolychnus schmidti. Das Weibchen dieses Tiefseefisches aus dem Karibischen Meer und dem Stillen Ozean ist etwa 7 cm lang und trägt auf seiner Unterseite Zwergmännchen, die sich in ihrem Jugendstadium am Körper des Weibchens festgebissen haben und nun mit ihm verwachsen sind. Ihre Sinneswerkzeuge sind verkümmert, ihr Maul ohne Zähne, da sie weder irgend etwas erbeuten oder verzehren wollen oder müssen: ihr Blutkreislauf hat sich dem des Weibchens verbunden. Ausschließlich die Keimdrüsen der Männchen vergrößern sich enorm (vgl. K. von Frisch, Du und das Leben, Berlin 1936, 253). Ganz ähnliche Paarbindungen finden wir bei vielen Tiefseefischen wie Linophryne argyresca (Indischer Ozean), Ceratias holbaelli (Nordatlantik), Melanocetus cirrifer. Nun, ich denke, man wird hier das Pärchen als struk- 63 turelle biologische Einheit betrachten müssen. Nicht ganz so einfach fällt die Beurteilung mancher Symbionten, etwa der Flechten. Flechten bestehen aus kugel- oder fadenförmigen chlorophyllführenden Algen, die von Pilzfäden dicht umsponnen sind. Beide Anteile tragen zur Ernährung des Ganzen bei: die Algen können assimilatorisch Zucker aufbauen, die Pilze versorgen das Ganze mit Wasser und Mineralien, liefern den Algen durch ihre Atmung das zum Assimilieren benötigte Kohlendioxyd und halten das Ganze auf der Unterlage fest. Pilze und Algen bilden natürlich ihre eigenen Fortpflanzungszellen aus, ja es gibt sogar Flechten, bei denen Pilze und Algen auch getrennt leben können. Noch etwas komplizierter ist die Symbiose bei einigen Amöben, etwa der beschälten Paulinella chromatophora. Sie umschließt stets genau zwei wurstförmige Blaualgenzellen. Vermehrt sich die Amöbe, wird jedesmal die eine der beiden Blaualgenzellen in die Tochterzelle hineingepreßt, worauf sich beide Algenzellen sogleich teilen, um die Zweiheit wieder herzustellen. Die Algen sind für das Leben der Amöbe absolut notwendig, denn sie allein liefern Nahrung, die sie verarbeiten kann. Die Algen haben also gleichsam die Rolle von Organen übernommen. Ich denke, bei solchen engen Bindungen der Symbionten aneinander, die den einen ohne den anderen nicht lebensfähig sein läßt, müssen wir wohl die symbiotische Ganzheit als biologisch-strukturelle Einheit verstehen.

ee) Tiersozietäten Überorganismen sind aber ganz besonders problematisch, wenn es sich dabei um Tiersozietäten handelt. Solche Tiersozietäten begegnen uns vor allem als Insektengemeinschaften (Bienen, Ameisen, Termiten . . .). Die Einzelwesen weisen oberflächlich betrachtet keinen «körperlichen» Zusammenhang auf, dennoch sind sie in ihren sozialen Leistungen, ihrer sozialen Funktion so eindeutig fixiert, daß ihre Leistung und Funktion und damit auch weitgehend das Einzelwesen selbst, allein im Gesamtzusammenhang des Übergeordneten Ganzen einen Sinn erhalten. Auch können einzelne Tiere nicht längere Zeit außerhalb des Gesamtverbandes leben. Trennt man sie ab, gehen sie sehr bald zugrunde, obwohl ihnen alles materiell zum Leben Notwendige reichlich angeboten wird. «Das gesamte Leben ist nach einem auf die Bedürfnisse des Ganzen eingestellten Plan geordnet, was soweit geht, daß bei Ausfall von Bienen einer bestimmten Funktion andere deren Aufgabe übernehmen können, wobei geschwundene Organe wieder aufgebaut werden können». Die Sozietät «besitzt also weitgehend die Fähigkeit zur Selbstregulierung» (R. Lotze und H. Sihler, a. a. O. 267). Daß es sich bei solchen Tiersozietäten um 64 funktionale Einheiten handelt, läßt sich somit kaum bestreiten. Ist aber die funktionale Einheit derart, daß wir als Grund der Einheit der Funktion eine Einheit der Struktur als eigentliche biologische Einheit annehmen müssen? Sind, mit anderen Worten, die funktionalen Einheiten nur dann zu erklären, wenn sie von numerisch einer Struktur gleichsam gesteuert, übereint werden? Viele Biologen neigen dazu, diese Frage zu bejahen, verhalten sich doch die «scheinbar einzelnen Tiere wie «lockere Organe* einer losen strukturellen Ganzheit» (A. Wenzl, Wissenschaft und Weltanschauung, Leipzig 21949, 317). Wie Körperzellen sind sie auf die Verbindung mit der Ganzheit (der Struktur) angewiesen. Nicht die Menge der Einzelnen macht die Sozietät aus, sondern «die innere Gliederung ihrer Gemeinschaft, ihrer Arbeitsteilung und die Unterordnung der Einzelwesen unter die Bedürfnisse und das Wohl der Gesamtheit» (K. von Frisch, a.a.O., 208). Auch die Sprachpraxis der Imker scheint die biologisch-strukturelle Einheit im ‹Bien› zu sehen: Der Bien ist klug, der Bien schwärmt, der Bien schickt seine Bienen aus (vgl. E. Mayer, a. a. O., 227). FRITZ HAHΝ spricht der Einzelbiene ausdrücklich eine «Individualität» ab: «Die Biene, die an uns vorbeifliegt, ist kein Individuum, sondern sie ist die Arbeiterin eines Bienenstaates, die sich nicht fortpflanzen kann, oder sie ist eine Königin, die nur leben kann, so lange die Dienerinnen sie füttern» (Das Buch der Natur II, Rüschlikon, Zürich 1952, 268). Rémy CHAUVIN schreibt: «Die Annahme ist berechtigt, daß es sich bei ‹der Biene› nur um eine vom Menschen vorgenommene Abstraktion handelt, daß die Insektenstaaten gar keine Gemeinschaft sind, sondern regelrechte Organismen, bei denen die einzelnen Bienen, Ameisen oder Termiten Zellen darstellen ... sie können sich vorübergehend aus dem Organismus lösen, um Nahrung einzuholen, den Gesamtverband gegen Feinde zu verteidigen.» (Tiere unter Tieren, Frankfurt und Hamburg, 1967, 13 f.). Falsch wäre es, in der Königin das Oberhaupt einer Gemeinschaft zu sehen, das die Gemeinschaft reguliert: Die «Königin erteilt keine Befehle und kontrolliert nur indirekt die Tätigkeiten des Bienenvolkes. Eher könnte man sie das Ovar des Bienenstaates nennen, ein Fortpflanzungs-organ, das nicht nur Eier erzeugt, sondern auch bestimmte Hormone, durch welche mehr oder weniger rasch die meisten Funktionen des sozialen Organismus geregelt werden» (R. Chauvin, a.a.O., 52). Bestimmte physiologische oder funktionale Reaktionen setzen eine Bienen-sozietät voraus, obwohl sie auch durchaus sinnvoll wären, wenn die Biene «für sich» wäre. Fehlt in einem Stock die Königin, entwickeln sieh Ovarien bei Arbeiterinnen nur dann richtig, wenn mehrere zusammen sind (ibd. 53). 65 Bringt man zwischen zwei Waben, einen Wachsstreifen an, der senkrecht zu ihnen steht, biegen die Bienen ihn bald zurecht, obwohl diese Situation unter normalen Umständen niemals eintritt. Dabei spielt - wie beim Wabenbau - nicht die Einzelbiene, sondern eine Bienenkette eine irgendwie ordnende Rolle (ibd. 70). Solche Bienenketten werden zwischen dem Wachsstreifen und den benachbarten Waben hergestellt. Sie stellen gleichsam die «Lochkarteninformation» für den richtigen Umbau des Wachsstreifens dar. «Ganz langsam verformten sich die Maschen nach einem Gesetz, das wir nicht zu ergründen vermochten, und schließlich stand der Streifen parallel zu den Waben. Bei diesem Anblick fiel mir unwillkürlich das Nervensystem mit seinen zahllosen, maschenartig verzweigten Fasern ein ...» (ibd. 67). CHAUVIN faßt seine Meinung so zusammen: «Ich glaube also, daß es durchaus berechtigt ist, den Bienenstaat als eine Art SuperOrganismus zu betrachten. So gesehen, werden die Holzkasten, die wir in unseren Gärten aufstellen, und aus deren Fluglöchern die Sammelbienen geschäftig ausfliegen, irgendwie ein wenig unheimlich. Jeder dieser Kästen birgt ein ziemlich großes Tier, das 4 bis 5 Kilogramm schwer ist, in den Wachswaben ein Skelett und in den Eierstöcken der Königin und den Hoden der Drohnen hermaphroditische Geschlechtsorgane besitzt... Für Atmung sorgen eine Reihe von ‹Fächelbienen›, die verbrauchte, zu feuchte oder zu warme Luft aus dem Stock heraustreiben ... Der ununterbrochene Stoffaustausch zwischen den einzelnen Bienen stellt einen sehr regen Kreislauf dar.. . Zwar gibt es keine Venen und Arterien, aber diese werden durch mündlichen Austausch von Nahrung und Sozialhormonen vollkommen ersetzt. Die Wärmeerzeugung ist eine der wichtigsten ... sozialen Funktionen (der Bienen) ... sobald sie sich zu einer Traube sammeln, vermögen sie eine recht konstante Temperatur zu erzeugen und aufrecht zu erhalten (die Biene ist wechselwarm, der Bien ein ‹Warmblütler›),.. Unter normalen Verhältnissen herrscht im Bienenstock eine Wärme von 34°, und diese Temperatur wird, wie beim Menschen, auf etwa ein halbes Grad genau eingehalten» (ibd. 85 f). Wo aber sitzt das ‹soziale› ZNS? Diese Frage läßt sich wohl biologisch kaum beantworten. Fest steht nur der

Sachverhalt: wenn es staatenbildenden Insekten «gelingt, ihre winzigen Einzelgehirne zusammenzuschließen, sie gemeinsam tätig werden zu lassen, dann erlangen sie psychisch ein ganz anderes, höheres Niveau als die solitären Insekten» (ibd. 88). Vielleicht vermag uns die Leistung der Termiten eine noch präzisere Antwort zu geben. Termiten haben einen sehr hohen Stand sozialer Organisation erreicht, obwohl sie anatomisch zu den primitivsten und 66 biohistorisch zu den ältesten Insekten gehören. Sie errichten Bauten bis zu 100 m Durchmesser (GRASSE) nach strengen architektonischen Regeln. «Können die winzigen Insekten solch gewaltige regelmäßige Bauten ohne einen Plan errichten? Ist ein solcher Plan in den einzelnen Insektengehirnen vorhanden, oder ist er dem Volk als Ganzem eingeprägt? Daß das einzelne Insekt den Plan in sich trägt ist sehr unwahrscheinlich, denn wenn wir etwas sicher wissen, dann die Tatsache, daß die Bienen, Ameisen oder Termiten als Einzelwesen ausgesprochen stupid sind. Wenn man sie von ihren Artgenossen isoliert, sind sie zu überhaupt nichts fähig und gehen nach kurzer Zeit ein... Diese Überlegung brachte midi auf die Theorie, daß im Insektenstaat eine Verbindung der einzelnen Gehirne gegeben ist, vergleichbar der Zusammenschaltung von Einzelelementen in einem Elektronengehirn» (ibd. 12.5). Und solche Einzelelemente sind für sich genommen auch außerordentlich «stupid», vermögen aber im Zusammenschluß mit vielen ihresgleichen, Erstaunliches zu leisten. Ich denke, wir werden also auch Tiersozietäten der geschilderten Art als strukturelle biologische Einheiten betrachten müssen. Wir haben also sehr verschiedene Formen biologischer Einheiten zu unterscheiden gelernt: morphologische Einheiten vermögen zwar alle charakteristischen Lebensfunktionen zu erfüllen, doch nur - jedenfalls unter natürlichen Bedingungen — im Verband mit anderen Einheiten. Nur morphologische Einheiten sind etwa Zellen oder Zellverbände klassischer Organismen, einzelne Glieder von Kolonialindividuen, vermutlich aber auch einzelne Partner bestimmter enger Symbiosen. Physiologische Einheiten dagegen können auch für sich natürlich alle wesentlichen biologischen Funktionen zeitigen und damit wenigstens für einige Zeit, für sich bestehen. Physiologische Einheiten sind etwa die klassischen Organismen, Kolonialindividuen oder einige der genannten symbiotischen Systeme. Strukturelle Einheiten sind Einheiten, bei denen sich die Einheit der Funktion (der morphologischen wie der physiologischen aber auch von sozialer) nur durch die Annahme einer höheren - eben der strukturellen — Einheit erklären lassen. Die meisten physiologischen Einheiten sind auch strukturelle Einheiten, Doch scheint es physiologische Einheiten zu geben, wie etwa die Einzelorganismen eines Bienenstaates, die keine strukturelle Einheit vorstellen. Bei der Behandlung solcher struktureller Einheiten wird sich vielleicht mancher Leser die Frage gestellt haben, ob nicht auch bestimmte soziale Gruppen von Menschen (ein Menschenstaat etwa) solche strukturellen Einheiten im biologischen Sinn darstellen. Ich möchte meinen, daß im 67 augenblicklichen Stand der Phylogenese der Menschheit solche strukturellen Einheiten noch nicht zustande gekommen sind, noch zustande kommen können. Doch ist zu beachten, daß sich seit einigen Jahrzehntausenden die Evolution der Menschheit nicht mehr im eigentlich biologischen Bereich vollzieht, sondern im sozialen. Die Menschen, die Menschheit gar, zeigt zunehmend Tendenzen zur Vergemeinschaftung, entwickelt zunehmend quasibiologische Mechanismen, die eine wachsende Vergemeinschaftung ermöglichen, ja erzwingen. Wir können also nicht ausschließen, daß die Evolution die Menschheit zu einer solchen strukturellen Einheit (in quasibiologischem Sinn) führt. Wäre eine solche radikale Vergemeinschaftung der Menschen \vunschenswert? Diese Frage ist sicherlich reichlich problematisch, vor allem, wenn man den evolutiven Trend berücksichtigt und die zunehmende Übervölkerung, die einen dichteren sozialen Zusammenschluß aus praktischen oder auch quasibiologischen Gründen erzwingen. Immerhin ist unsere Frage nicht ganz sinnlos, wenn man nach der auf den Menschen und die Menschheit zukommende Form der Vergemeinschaftung blickt. Es gibt sicher Ansätze, die vermuten lassen könnten, daß die menschliche Vergemeinschaftung einmal nach Art eines Insektenstaates zustande kommt. Symptome dafür wurden im Stalinismus oder im Nationalsozialismus signifikant. Aber noch deutlichere Symptome sprechen für eine ganz andere Form der endgültigen Vergemeinschaftung bzw. Übersozialisierung der Menschheit, läßt sich doch aufweisen, daß im Gegensatz zu Tiersozietäten die Vergemeinschaftung des Menschen nicht mit einem zunehmenden «Individualitätsverlust» Hand in Hand gehen muß, weil und insofern gerade soziales Engagement zu einer der unaufgebbaren Bedingungen der Personalisierung des Menschen gehört. Vergemeinschaftung im Menschenreich muß, wie es scheint, nicht in den grausigen Visionen Orwells enden, sondern kann durchaus Hand in Hand gehen mit zunehmender Personalisierung. Daß sie so und nicht anders abläuft, scheint in die Freiheit des Menschen gestellt. Der Überorganismus ‹Menschheit› wird aber nur dann in Verbindung mit echter Personalisierung entstehen können, wenn es uns gelingt, soziale Verhaltensweisen auszubilden, die zugleich zur Personalisierung führen. Eine solche Verhaltensweise hat - wenigstens theoretisch - das Christentum vorgestellt und stellt es immer noch vor: Es ist die der Vergemeinschaftung in Liebe. Liebe hat nicht nur soziale gemeinschaftgründende Funktion, sondern auch personalisierende. Durch Lieben wird der Mensch sowohl ver-gemeinschaftet, aber auch zugleich immer mehr Personen werden. Kehren wir nach diesem kurzen Exkurs über den Menschen und seine Zukunft wieder zurück zum Bereich eigentlich

biologischer Einheiten. 68 c) Sie sieben mit ihrer Umgebung in determinierender Wechselwirkung Physikalische Einheiten stehen in konstitutiver Wechselwirkung mit ihrer Umgebung, d. h. ein Elementarteilchen wird durch die physikalische Situation seiner Raumzeitbefindlichkeit konstituiert, während ein Lebewesen nur in determinierender Wechselwirkung mit seiner Umwelt steht. Dabei kann man durchaus soweit gehen zu behaupten, daß der Entscheid, ob sich ein Elementarteilchen als ein bestimmtes (a. h. durch bestimmte Quantenzahlen ausgestattetes) oder ein anderes bestimmtes vorstellt, ausschließlich von der Struktur der Raumzeit abhängt, in die es eingebettet ist.. Wir haben schon im Zusammenhang mit der Darstellung der allgemeinen Relativitätstheorie darauf verwiesen, daß Veränderung der metrischen Krümmung der Raumzeit mit dem Entstehen eines bestimmten Elementarteilchens zusammenfällt. Die Bootstrap Hypothese (vgl. WS I, 66) besagt zudem, daß man sich Mikroteilchen aus allen möglichen Elementarteilchen zusammengesetzt denken darf (unter anderem kann etwa ein Proton also auch aus Protonen ‹zusammengesetzt, sein), wenn nur die Summen der Quantenzahlen mit der für ein Elementarteilchen typischen identisch ist. Bei der Ununterscheidbarkeit gleichartiger miteinander wechselwirkender Bosonen kann man sogar sinnvoll sagen, daß unser Proton u. a. sich selbst als ‹Teil› enthält. Solche PseudoStrukturen sind natürlich für biologische Einheiten undenkbar. Eine biologische Einheit steht in einer ganz anderen Beziehung zu ihrer raumzeitlichen Umgebung: sie konstituiert sie nicht und wird nicht von ihr konstituiert, sondern determiniert sie und wird von ihr determiniert. Schon DARWIN hatte das Lebewesen in engstem Zusammenhang mit seinen Lebensbedingungen gesehen. Die Umwelttheorie von UEXKÜLLS wurde von K. GOLDSTEIN sehr charakteristisch erweitert: «Statisch betrachtet erscheint der Organismus wie schlechthin eingebettet und angepaßt an seine Welt, dynamisch gesehen aber steht er in Zusammenstoß und Auseinandersetzung mit ihr, er erscheint wie ein Sein in Unordnung. Die Begegnung mit der Umwelt stellt für den Organismus eine Folge von katastrophalen Erschütterungen dar und diese ergeben ein Ungleichgewicht, das überwunden werden muß. Diese Erschütterungen sind der Ausdruck der Individualität des Organismus mit dem Anderssein in der Welt, Erst im Fortschreiten von Katastrophe zu Katastrophe gewinnt er sein eigentliches Sein: als eine sich ordnende und Ordnung herstellende Struktur. Der Organismus erscheint als ein Sein in «Ordnung., sein Sinn als die erobernde Einbeziehung und Gestaltung der Welt als organisierter. Welt und Organismus verwirklichen sich gegenseitig» (Der Aufbau des Organismus, Haag 1934, 349 ff). 69 Auch im Raum des DIAMAT wird die wechselseitige determinierende Funktion der biologischen Einheit mit seiner Umwelt betont I. W. MITSCHURIN hat hier (wenn auch zur Begründung seiner inzwischen allgemein abgelehnten Theorie der Vererbung) vieles gut formuliert: «Jedes Organ, jedes Merkmal, jedes Glied, alle inneren und äußeren Teile eines Organismus sind durch die äußeren Verhältnisse seine Daseins bedingt. Die Organisation der Pflanzen in ihrer heutigen Form erklärt sich daraus, daß jedes Detail eine bestimmte Funktion erfüllt, die nur unter den gegebenen Bedingungen möglich und notwendig ist. Verändert man diese Bedingungen, so wird diese Funktion unmöglich oder unnötig, und das entsprechende Organ stirbt allmählich ab» (Kriticeskij obsor dostizenij genetiki poslednego vremini, Moskau 1948, 590). Nun, das Wort bedingt scheint uns zu viel zu sagen. Richtig aber betrachtet MITSCHURIN «den Organismus und seine Existenzbedingungen als eine komplizierte dialektische Einheit», bei der die Umwelt durchaus aktiv in Erscheinung tritt (vgl. I. T. Frolow, Kausalität und Zweckmäßigkeit in der lebenden Natur, Berlin 1963, 85 f). Dabei unterstrich Mitschurin stets die Widerstandsfähigkeit der organismischen Strukturen und ihre relative Freiheit von der äußeren Umgebung in Bezug auf die Ontogenese, tauchte jedoch in der phylogenetischen Betrachtung den Organismus allzu sehr in seine Umwelt ein (ibd. 87 f). Die physikochemische Beziehung von Umwelt und biologischer Struktur wird vielleicht am deutlichsten in der Ausfaltung des von L. VON BERTALANFFY in die Biologie eingebrachten Terminus «Fließgleichgewicht». Versteht man Stoffwechselvorgänge aufbauender Art vereinfacht als Assimilation, solche abbauender Art als Dissimilation, dann besteht die Funktionalität des Organismischen im Fließgleichgewicht zwischen beiden. In der Zelle und anderen biologischen Gebilden findet man bestimmte chemische Verbindungen, ein konstantes Verhältnis zwischen verschiedenen enzymatischen Komponenten, die jedoch in ständigem Auf- und Abbau begriffen sind. Das Fließgleichgewicht in biologischen Strukturen ist - im Gegensatz zu ähnlichen Erscheinungen bei anorganismischen Systemen - irreversibel, weil und insofern das chemische Gleichgewicht arbeitsunfähig, das biologische (offene) arbeitsfähig ist. Auf den ersten Blick scheint für die biologische Einheit der Satz von der Zunahme thermodynamischer Entropie nur mit einigen Modifikationen zu gelten (vgl. WS II, 29 ff). Biologische Systeme ‹streben› ein dynamisches Gleichgewicht zwischen Ein- und Ausfluß an. Dieses Gleichgewicht ist nur abhängig von den Reaktionskoeffizienten des Stoffwechsels, nicht vom Ausgangszustand (etwa der Ausgangsmasse, wie sie für die Endzustände geschlossener 70 Systeme - Uhren, Gefäß mit reagierenden Stoffen - entscheidend ist). Teilt man ein Fließsystem, so wachsen die Teile unabhängig von ihrer Größe automatisch) zum Gleichgewichtszustand des Muttersystems heran, sofern nur die

Umgebung und die Reaktionskoeffizienten des Stoffwechsels gleich geblieben sind. Für die Erhaltung eines Fließsystems ist es von Vorteil, wenn Regulationsverfahren eingebaut sind, die die Konzentration wichtiger Stoffe konstant halten, auch wenn die Umweltbedingungen sich ändern. Schon Einzeller besitzen solche Apparate, bei Mehrzellern wird diese Funktion oft durch ganze Organsysteme gesteuert (Nerven- und Hormonsystem). Solche Systeme arbeiten nach zum guten Teil kybernetisch zu deutenden Mechanismen (vgl. R. W. Kaplan, Probleme der Lebensentstehung in G. Herberer (Hrsg.), Die Evolution der Organismen I, Stuttgart 1967, 515f). L. VON BERTALANFEY konnte noch eine weitere typische nur der biologischen Einheit zukommende Eigenschaft in ihrem Verhältnis zur ‹Umwelt› aufzeigen: die Äquifinalität. Damit ist gemeint, daß eine organismische Einheit von sehr verschiedenen Ausgangsbedingungen (der Umwelt) den gleichen Endzustand erreichen kann [Der Organismus als physikalisches System betrachtet, in NW 28 (1940), 527 ff]. Nicht selten wurde die Lehre vom Organismus als eines im Fließgleichgewicht stehenden Systems überstrapaziert. Man folgerte, daß offenbar das System beim Austausch aller materieller Bausteine mit sich selbst identisch bleibe. Nun, das wäre immerhin möglich, nur gibt es einen solchen totalen Stoffwechsel vermutlich nicht. Zunächst einmal werden in einem höheren Organismus keineswegs alle Zellen im Laufe einer bestimmten Zeit durch andere ausgetauscht. So bleiben manche Zellen des ZNS so wie sie einmal ausgewachsen sind, bis zum Tode des Lebewesens, gleichsam für sich ‹alternd› erhalten. Haben aber nicht auch diese Zellen einen Stoffwechsel, der vielleicht ihre Teile total erneuert? FRIEDRICH FREKSA hat einige Versuche mit radioaktivem Kalzium, Phosphor, Sauerstoff, Kohlenstoff unternommen. Er ‹fütterte› einfache Lebewesen mit solchen Isotopen, ein Teil der radioaktiv markierten Moleküle wurde in die Geschlechtszellen eingebaut. Bei den Nachkommen dieser Lebewesen blieben nun diese offensichtlich in die DNS eingelagerte Atome ein Leben lang erhalten, sie unterlagen also nicht dem Stoffaustausch. Es scheint also in jedem Organismus einen bestimmten Bestand an DNS Molekeln zu geben, der die Identität des Systems mit sich selbst in allem Stoffwechsel wahrt. Jedoch gibt es durchaus An- 71 zeichen für die Erhaltung der Struktur, unabhängig vom DNS-Bestand: Auch weniger materielle Eigenschaften können für ein Lebewesen ein Leben lang erhalten bleiben. Wenn man neugeborenen Tieren (auch Menschen) makromolekulare Fremdstoffe, die meist an Eiweiße angelagert sind, etwa als «Gewebecoktail» aus fremdem Gewebe oder auch andersartige Antigène (das sind großmolekulare Fremdstoffe, die vom Organismus an sich als fremd empfunden werden und gegen die er Antikörper ausbildet) injiziert, führt diese Behandlung zu einer lebenslangen Toleranz gegenüber diesen Antigenen. So werden etwa entsprechende Transplantate nicht mehr abgestoßen. Der Organismus erhält eine zusätzliche Information, an die er sich bei der Zuführung von Stoffen, die eigentlich als Antigène wirksam werden müßten, ‹erinnert› und stößt etwa jene Transplantate nicht mehr ab. Hier eröffnet sich für die Transplantatmedizin eine wichtige Möglichkeit, selbst größere Organe zu transplantieren, ohne daß sie abgestoßen würden. Bislang läßt sich ein solches Abstoßen nur in seltenen Ausnahmefällen vermeiden, wenn man die Bildung von Antikörpern künstlich ungezielt verhütet und damit die allgemeine Widerstandskraft des Organismus auch gegen Infektionskrankheiten auslösende FremdStoffe (Viren, Bakterien ...) erheblich senkt. Ein Leben lang bleibt meist auch die von den Verhaltensforschern als Prägung bezeichnete Determinierung des Verhaltens erhalten. Eine Prägung findet immer nur in einer bestimmten sensiblen Periode statt. Für die Nachfolgereaktion der Enten liegt diese sensible Periode zwischen der 13. und 16. Stunde nach dem Schlüpfen (E. HESS). Die erworbene Kenntnis des auslösenden Objekts bleibt zeitlebens behalten, während sonst gerade das Vergessen ein wesentliches Merkmal alles Erlernens ist. Man kann zwei auf Menschen als Geschlechtspartner geprägte Wellensittiche in einem dunklen Käfig zwar dazu bringen zu. balzen und zu brüten. Zeigt man sich aber den Vögeln, balzen sie den Menschen an, die Paarbindung zerbricht und die Brut wird vernachlässigt (HELLMANN). Geprägt wird aber immer nur eine bestimmte Reaktion auf ein bestimmtes Objekt. Eine von LORENZ aufgezogene Dohle erkannte nur Menschen als Eltern- und Geschlechtskumpan an (Kumpan = Partner in einem bestimmten Funktionskreis). Sie flog jedoch mit Nebelkrähen (Flugkumpan) und nahm schließlich junge Dohlen als Kindkumpan an. Es dürfte auch beim Menschen Prägungen geben (P. GRAY, W. THORPE), doch ist das experimentelle Material hier noch recht dürftig (vgl I. Eibl-Eibesfeldt, Ethologie, in Handwörterbuch der Biologie II, Frankfurt 1966, 458-460). 72 Es scheint heute als sicher zu gelten, daß solche Prägungen nicht durch den konstanten Bestand an informierter DNS erklärt werden können. Sie scheinen also den Stoffwechsel irgendwie zu überlisten. d) Sie zeigen eine Dynamik zur Bewahrung ihrer Einheit Elementarteilchen kennen im Gegensatz zu biologischen Einheiten keine Dynamik der Funktion und des Verhaltens zur Bewahrung der Einheit. Während die biologische Einheit in ihrem Fließgleichgewicht doch die Einheit der Struktur durch den Wechsel der die Funktion mittragenden materiellen Systeme bewahrt, scheint es der physikalischen. Einheit völlig ‹gleichgültig› zu sein, wann sie zerfällt. So zerfallen, über eine große Zahl ausgemittelt, etwa die langlebigsten radioaktiven Isotopen von Kohlenstoff (14C) nach durchschnittlich 5 570 Jahren, von Stickstoff (13N) nach 10,05 Minuten, Kalium (40K) nach 1470000000 Jahren, Uran (238U) nach 4510000000 Jahren. Wir nennen diese Zeiten Halbwertzeiten. Nun scheinen solche Halbwertzeiten für das einzelne Elementarteilchen nichts oder sehr wenig

zu sagen, kann doch immerhin ein l4C-Atom nach einer Sekunde oder nach 1 000 000 Jahren zerfallen, ohne daß uns die Gründe dafür bekannt wären, also auch nicht, ob eventuell im letzteren Teilchen ein Selbsterhaltungsmechanismus stärker ausgeprägt ist, als in dem Teilchen, das nach einer Sekunde zerfällt. Ganz ähnliche Halbwertzeiten lassen sich nun als Durchschnittliche Lebenserwartungen, selbst des Menschen angeben. So haben nach der «Allgemeinen Deutschen Sterbetafel 1924/26 (Männer)» männliche Neugeborene eine ‹Halbwertzeit› von 66,4 Jahren, zehnjährige Jungen eine solche von 59,0 usw. Kennt man das Alter nicht (wie es ja bei den Elementarteilchen meist der Fall ist), kann man einen Durchschnittswert für alle lebenden männlichen Individuen errechnen. Nun aber ist der Mensch als lebendige Einheit offenbar mit einer Dynamik zur Bewahrung der Einheit der Struktur (d. h. der Erhaltung des ‹Lebens›) ausgestattet. Wie unterscheidet er sich unter der von uns angegebenen Rücksicht von einem Elementarteilchen? Wenn wir das ‹Alter› von Elementarteilchen kennen würden, müßten sich je nach dem Alter deutliche Veränderungen der ‹durchschnittlichen Lebenserwartung) ergeben, wenn sie eine Dynamik auf die Bewahrung der Einheit besäßen. Das aber ist nicht der Fall. Elementarteilchen mit unbekanntem ‹Alter› und solche mit bekanntem Alter (von denen wir also die Entstehungszeit genau kennen, wie etwa bei künstlich radioaktiv gemachten Isotopen) haben, soweit bekannt, genau die gleiche ‹mittlere Lebenserwartung), das aber ist bei Lebewesen nicht der Fall: die mittlere Lebenserwartung nimmt mit zunehmendem Alter ab. Dieser Sachverhalt ist am einfachsten dadurch zu 73 interpretieren, daß bei jugendlichen biologischen Einheiten die Selbsterhaltungstendenz effektiver ist als bei älteren biologischen Systemen, während bei physikalischen Einheiten entweder die Selbsterhaltungstendenz immer gleich wirksam bleibt oder gar nicht vorhanden ist. Nun gibt es für physikalische Einheiten keinerlei physikalische Anzeichen für die Existenz einer solchen Tendenz bzw. eines Faktors, der eine solche Tendenz bewirkt, während sie bei vielen - vor allem höheren - biologischen Einheiten als Selbsterhaltungstrieb durchaus bekannt ist. Wir dürfen somit zu Recht behaupten, daß sich biologische von physikalischen Einheiten durch eine effektive Tendenz zur Selbsterhaltung auszeichnen. Bei den vorhergehenden Überlegungen haben wir vorausgesetzt, daß ein Lebewesen sterben muß, gegen dieses Sterbenmüssen jedoch aufschiebende Struktur- und Verhaltensmechanismen entwickelt hat und tätig werden läßt. Man kann jedoch, wie schon angedeutet, den gemeinten Tatbestand von einer anderen Seite her angehen: dem des Alterns. Elementarteilchen scheinen nicht zu altern, während biologische Systeme altern können. Das Altern ist nur verständlich als zunehmende Abnahme der Effektivität der Erhaltungsmechanismen. Zudem muß nach BÜRGER streng zwischen biologischem und kalendarischem Alter unterschieden werden, beide sind individuell sehr verschieden. Ein Einwand gegen unsere Theorie mag im Hinweis auf die materielle (somatische) Unsterblichkeit mancher Tiere (entsprechendes gilt wenigstens im Prinzip für die meisten Pflanzen) gegeben sein. Materiell unsterblich sind vor allem teilbare labile Tiere, z. B. Schwämme und manche Hydrozoen, aber auch fast alle Verbandsorganismen (Kolonialtiere .. .). Bei solchen Organismen ist keine ‹Zellart› zellkonstant geworden, so kann man denn aus isolierten Zellen wieder ganze Tiere entstehen lassen, die dadurch zustande kommen, daß morphologisch undifferenzierte und indifferente totipotente Zellelemente amöboid zusammenkriechen und so einen Zellhaufen bilden, aus dem sich das neue Tier differenziert. Labile Tiere sind natürlich auch alle Einzeller, die sich ungeschlechtlich durch Teilung vermehren, wie etwa die Protozoen. Auch sie sind potentiell materiell unsterblich. Aber zeigen nicht gerade solche Einzeller (aber auch manche Metazoen, etwa Turbellarien, die sich in günstigen Kulturmedien dauernd ungeschlechtlich fortpflanzen), daß das Lebendige hier Funktionen ausgebildet hat, um den materiellen Tod von überzellulären Einheiten zu verhindern (die Zelle selbst stirbt natürlich ab, wie auch bei Verbandsorganismen die einzelne Zelle abstirbt). Es bleibt doch materiell gesehen ein Überganzes erhalten. Doch gibt es hier den «individuellen, immateriellen Tod» (J. W. HARMS), 74 gegen den offensichtlich solche Mechanismen nicht entwickelt wurden. Bei solchen Tieren stellt die phylogenetische biologische Einheit vor der ontogenetischen. Erheblich sind auch die Einwände, die von der Existenz total zellkonstanter Organismen ausgehen: So besitzt etwa das Rädertierchen Hydatina senta im Soma 959 Zellen, von denen 247 das Nervensystem, 122 die Muskulatur, 501 die Haut und ihre Bildungen und 76 Zellen den Darm bilden. Alle diese Zellen sind absolut formkonstant und nach der progressiven Phase (im Erwachsenenstadium des Tieres also) nicht mehr teilbar. Hier sind vom Tier offenbar keine strukturellen, wohl aber Verhaltungsmechanismen zur Selbsterhaltung ausgebildet worden. Der regressiven Phase (dem Altern) des Organismus als Ganzem wirken also keine besonderen strukturellen Mechanismen entgegen. Wir glauben nicht, diese Schwierigkeiten lösen zu können, möchten aber darauf verweisen, daß man typische Eigenschaften bzw. Funktionen des Lebendigen nicht auch stets in Grenzsituationen als gültig belegen können muß. Es kömmt uns bei unseren Ausführungen auf das Auffinden von durchgangigen Regeln an, die durchaus Ausnahmen zulassen. Doch haben wir mit unseren Überlegungen zum Problem des Alterns und Sterbens schon den folgenden Punkt unserer Untersuchungen berührt: e) Sie zeigen eine Dynamik zur Vervollkommnung Physikalische Einheiten kennen keine Dynamik auf eine Vervollkommnung hin, ja sie kennen keine Ontogenese. Biologische Einheiten dagegen entwickeln sich dagegen im Verlauf ihres Bestehens wenigstens zeitweilig (in der

progressiven Phase) auf ein bestimmtes ‹Ziel› hin. Ein Elektron ist entweder ein vollkommenes Elektron oder es ist keines. Ein Lebewesen wird dagegen in der genannten Phase durchaus Stadien auf eine «Vervollkommnung) hin durchlaufen. Diese ‹Stadien› wollen wir einmal des näheren untersuchen: sie sind entweder ontogenetischer (ordentlicher oder außerordentlicher) oder phylogenetischer Art. aa. Die Ontogenese (wir sehen im folgenden von der regressiven Phase der Ontogenese ab) als typische Erscheinung bei biologischen Einheiten. Die Individualentwicklung (= Ontogenese) verläuft bei mehrzelligen Tieren zumeist so, daß sich aus einer relativ einfach strukturierten befruchteten Eizelle durch eine Reihe von Zellteilungen ein zunächst noch wenig differenzierter Keim bildet, an dem dann nach und nach Organanlagen sichtbar werden, die sich zu funktionierenden Organen 75 ausbilden. Ursprünglich glaubte man, daß sich in einer Keimzelle schon das ganze zukünftige Lebewesen befinde, daß es nur noch zu wachsen brauche. Diese auf EMPEDOKLES zurückgehende Theorie (Präformationstheorie) wurde jedoch schon von ARISTOTELES in Frage gestellt, was nicht heißen soll, daß sie nicht doch noch bekannte Vertreter bis hin in die Neuzeit fand. Heute ist diese Theorie aber allgemein aufgegeben. Wir wissen, daß Keimzellen und auch noch die ersten undifferenzierten Gewebe früher Entwicklungsstadien totipotent sind, d. h. sie sind befähigt, alle Strukturen eines Organismus auszubilden. Mit dem allmählichen Verlust dieser Totipotenz im Laufe der Entwicklung setzt eine autonome, zum Teil wenigstens von den Erbfaktoren geleitete Differenzierung der Gewebe ein. Dabei haben manche Gewebe einen starken gestaltenden Einfluß auf die Nachbargewebe. Entdeckt wurden solche richtenden Einflüsse der Nachbargewebe von H. SPEMANN. u. a. bei seinen Transplantationsversuchen an Molchkeimen. So transplantierte er etwa ein Stück der oberen Urmundlippe einer frühen Molchgastrula in die künftige Bauchregion und stellte fest, daß das transplantierte Gewebe ortsgemäß zur Bauchhaut wurde. Im allgemeinen kann die Regel gelten: Was sich entwickelt, bestimmt die Umgebung, wie es sich entwickelt, bestimmt das regenerierende Gewebe. So ist z. B. bei Fröschen die ganze Epidermis (die äußere Zellschiebt der Haut) zur Linsenbildung befähigt, wenn man eine Augenblase aus einer Froschlarve herausschneidet und sie irgendwo auf der Epidermis einsetzt, entsteht eine Linse, während bei Unken nur die Kopfhaut zur Ausbildung einer Augenlinse gebracht werden kann. Die Totipotenz der Furchungszellen (Zellen, die im ersten Abschnitt der Embryonalentwicklung entstehen) mögen folgende Beispiele belegen: Wird ein kernloses Eifragment eines Seeeigels durch stark salzhaltiges Wasser künstlich zur Entwicklung angeregt, entstehen, trotz des Fehlens der Kern-DNS, sternförmige Bündel zäherer Plasmamassen, die sieh teilen und durch ‹Furchung› auf zwei ‹Zellen› verteilt werden. Solche kernlosen Gewebe können sich etwa bis zum Blastulastadium entwickeln, sterben dann aber ab. Bei dem Polypen Hydra vulg. kann sich aus jeder Zelle des 16-Zellen-Stadiums noch ein ganzer Polyp entwickeln. Ein zweigeteiltes 64-ZelIen-Stadium des Seeigels ergibt noch zwei normale Seeigellarven. Preßt man andererseits zwei 2-Zellen-Stadien (auch verschiedener Art) von Molchen 'zusammen, kann aus dem künstlichen 4-Zellen-Stadium ein neues Tier hervorgehen. Auf Grund solcher und ähnlicher Versuche entwickelte C. M. CHILD seine Gradiententheorie, nach der bestimmte Stellen jeweils eine starke 76 physiologische Aktivität entfalten und gewissermaßen das zur Differenzierung benötigte Material an sich reißen, wobei sich konkurrierende Anlagen unterdrücken können. Dabei kommt es oft zu Regulationen von der Ganzheit des Keimes her: Wird z. B. bei einem Molchembryo das Mesodermgewebe einer Beinanlage derart umgedreht, daß die Innenseite nach außen zu liegen kommt, entsteht ein seitenrichtiges Bein. Offenbar übt also die Haut einen richtenden Einfluß aus. Wir haben im vorhergehenden Text einige Fachausdrücke der Ontogenese verwandt, die noch naher erklärt werden müssen: Zunächst teilt sich ontogenetisch die Eizelle bei etwa gleichbleibender Größe durch mehrere Furchungsteilungen in mehrere Zellen (Morula). Den Abschluß dieser Entwicklungsphase bildet in der Regel eine Blastula, zu der die noch recht unspezifizierten Zellen eine Hohlkugel ausbilden. Es kann aber auch eine kompakte Zellmasse ohne Hohlraum entstehen oder eine flache Keimscheibe. Daran schließt sich die Phase der Keimblätterbildung (Gastrulation) an, bei der durch Einstülpung, Einwucherung oder Überwachsen eine äußere Zellschicht, das Ektoderm und eine innere Zellschicht, das Endoderm entsteht, später entwickelt sich ein zwischengeschaltetes Keimblatt, das Mesoderm. Nach weiteren Zellwucherungen und Faltungen bilden sich dann noch wenig differenzierte Organanlagen heraus, die zu den Primitivorganen des Embryos werden. Aus diesen werden die fertigen und funktionstüchtigen Organe des Jungtieres (vgl. B. Rensch, Biologie II, Frankfurt 1963, 72-95). Eine kausalanalytische Beschreibung der Ontogenese ist vor allem deshalb schwierig, weil es bis zum heutigen Tage noch nicht möglich war zu beweisen, daß die typische Zeitordnung der Einzelentwicklung durch eigens dafür vorhandene Gene oder durch sonstwie geartete Einflüsse der DNS gesteuert wird. Die Information über die Zeitordnung scheint vielmehr in der Komposition des gesamten genetischen Systems enthalten zu sein. Wie das aber im Einzelnen aussehen könnte, ist ein Rätsel der Biologie. Noch unvollständiger ist unser Wissen um das Zustandekommen der Raumordnung. Obwohl die Raumordnung zu den konstantesten und strengst determinierten

Elementen der gesamten Ontogenese gehört, sind die verursachenden Informationsstrukturen nahezu völlig unbekannt. Auch hier scheint der Ausweg über die Gesamtheit der Geninformation eher -ein nichts erklärender Notbehelf, denn eine brauchbare oder gar zu verifizierte Hypothese zu sein (vgl. A. Barthelmess, Grundlagen der Vererbung, in Handbuch der Biologie III, 2, Frankfurt 1966, 792). bb. Die Regeneration als typische Erscheinung biologischer Einheiten. Die meisten Lebewesen besitzen die Fähigkeit, bereits differenzierte Teile 77 ihres Organismus nach Verlust neu zu bilden, sei es als Ausheilung von Wunden (Reparation) oder als Wiederherstellung ganzer Organe (rekonstitutive Regeneration). Uns interessiert hier vor allem die letztere. Viele Süßwasserpolypen der Gattung Hydra können aus 0,5 %, manche Strudelwürmer aus etwa i %, viele Seesterne und Schlangensterne aus einem Arm ein ganzes Individuum regenerieren. Obwohl solche Regenerationen meist nur bei «niederen» Tierfamilien vorkommen (Schwämme, Schnurwürmer, Ringelwürmer, Manteltiere, Weichtiere, Spinnentiere, Krebse, Insektenlarven), regenerieren auch Wirbeltiere ganze Körperteile: Fische regenerieren Schuppen und Flossenstrahlen, Schwanzlurche und Larven von Froschlurchen Beine und Schwänze, Eidechsen einen Teil des Schwanzes, Stachelratten (ein Säuger!) das Schwänzende mit einem langen Endwirbel. Nicht regenerieren können Dauergewebe wie das Hirngewebe erwachsener Säuger und Tiere, die aus einer genau festgelegten Zahl von Zellen bestehen, wie die Rädertierchen und viele Fadenwürmer (Rensch, a. a. O. 229 f). cc. Die Phylogenese als typische Erscheinung lebendiger Einheiten. Wenn wir von der Stammesentwicklung lebendiger Einheiten sprechen, wird das Wort lebendige Einheit recht problematisch. Dennoch wollen wir aus noch zu erwähnenden Gründen daran festhalten. Die biologische Einheit ist hier wieder überorganismisch zu verstehen, die biologische Einheit ist das sich entwickelnde Phylum. Was aber berechtigt uns ein ganzes Phylum als biologische Einheit zu begreifen? Es ist das die so sehr umstrittene orthogenetische Phylogenese. Unter Orthogenese verstehen wir eine Entwicklung des Phylums auf ein Funktions-ziel hin. Sie ist etwa vergleichbar der ‹normalen› Ontogenese, die ja auch auf die Ausbildung von funktionalen Evolutionszielen ausgerichtet ist. Das Faktum der Orthogenese scheint uns unbestreitbar zu sein. Anhaltende und vorherrschende Tendenzen in der phylogenetischen Progression zeigen.etwa 90 % aller paläontologischen Funde. «Die Tendenz von Stämmen, ihre Entwicklung in nahezu gleicher Richtung während beträchtlicher Zeitabschnitte fortzusetzen, gradlinige Evolution oder Orthogenesis in einem deskriptiven Sinn, ist ein allgemeines Evolutions-phänomen», schreibt G. G. SIMPSON (Zeitmaße und Ablaufformen der Evolution, Göttingen 1951, 249). Solche Entwicklungstendenzen (/Trends,) treten oft nicht nur bei einzelnen Abstammungsreihen, sondern meist gehäuft in Bündeln naher verwandter Arten oder Gattungen auf. Sicher hat man die Orthogenese lange Zeit allzu statisch und allzu einfach gesehen. Man glaubte eine einfache Gradlinigkeit zu erkennen und wußte nichts von Schwankungen in der Evolutionsrichtung und 78 der evolutiven Geschwindigkeit. Die realen Trends verlaufen meist sehr viel unruhiger. Das Musterbeispiel bildet die Evolution der Equiden. Der Ursprung der Equiden liegt im Dunkel. Dann taucht plötzlich in Nordamerika und Europa vor etwa 40 Mio. Jahren ein fuchsgroßer Buschschlüpfer mit Laubfressergebiß, einem reptilienhaften Gehirn und raubtierähnlichen Ballenfüßen (4 Zehen vorn, drei hinten) auf: der Eohippus. 10 Mio. Jahre später hatte sich aus ihm über den Orohippus, den Epihippus und den Mesohippus in Nordamerika der Miohyppos entwickelt, während der Eohyppos in Europa ausgestorben zu sein scheint. Mit dem Mesohippos begann ein deutlicher Wachstumsbeginn. Das Großhirn überlappt das Kleinhirn (typisches Saugerhirn!), die erste Zehe des Vorderfußes ist verschwunden. Mit dem Miohippos setzt eine Aufspaltung in verschiedene Gattungen ein (Archäohippos, Parahyppos, Anchiterium):der Archäohippos wird wieder kleiner. Mit dem Anchiterium, setzt eine Sonderentwicklung ein, vor allem werden seine Nachkommen bedeutend größer. Anchiterium und Hypohippos wandern auch in die Altwelt ein, sterben hier aber bald aus. Vor etwa 15 Mio. Jahren lebte in Amerika der Meryhyppos, ein Nachkomme des Parahyppos (auch ein Nachkomme des Miohyppos und Zeitgenosse des Hypohyppos). Der Übergang vom Para- zum Meryhippos ist wieder wichtig: Der Meryhyppos besitzt im wesentlichen schon die Hirnfurchen der heutigen Wiederkäuer (Wiederkäuerhirn), die Fußballen verschwinden, die Seitenzehen erscheinen zurückgebildet, es erfolgt die Umstellung von Laub- auf Grasnahrung, im Fuß werden neue Bänderapparate zu einem Sprungmechanismus entwickelt. Vor 9 Mio. Jahren entstanden in den folgenden 800 000 Jahren aus den Nachkommen des Meryhippos zahlreiche neue Gattungen: Prothyppos, Callhippos, Nannhippos, Neohyparion.-und die Gattung Hyparion, die wieder einmal in die alte Welt auswandert und hier in ihren Nachkommen, vor allem der Gattung Stylohippos, vor etwa 600 000 Jahren ausstirbt. Bei den Gattungen Hyparion und Prothyppos verschwinden die Seitenzehen und der Sprungmechanismus wird verstärkt. In einer Nachfolgegattung des Prothippos, den Mitgliedern der Gattung Pliohippos, setzt dann wieder eine ‹Aufspaltung, der Gattungen ein, in Südamerika entwickeln sieh die Gattungen Parahippidium, Hippidium und Onohippidium, während in Nordamerika die Gattung Equus entsteht, hier aber bald ausstirbt, während ein in die Alte Welt ausgewanderter Schuh weiterlebt und die uns heute bekannten Arten (Pferd, Esel, Zebra) ausbildet. Erst durch den Menschen wurden dann Pferde und Esel wieder in die Neue Welt ‹importiert›. .

79 Vom Eohippos zum Equus verläuft die Entwicklungslinie also keineswegs ganz ungestört und gradlinig. Doch lassen sich bestimmte Entwicklungslinien fast durchgängig auf weisen: Die Praemolaren passen sich den Molaren zunehmend an (aus einem Laubfressergebiß wird ein Grasfressergebiß), die Zehenzahl reduziert sich, das Gehirn wird umgestaltet, die Schädelproportionen ändern sich, die Größe nimmt zu. Ähnliche Reihen lassen sich für die Ammoniten, die Probosciden (sukzessive Verlängerung der Nase zu einem Rüssel, Größenzunahme der oberen Schneidezähne zu Stoßzähnen, Wachstum des Gesamtkörpers) und etwa den Titanotherien (die im Verlauf des Tertiärs immer größer wurden und immer größere Nasenhörner ausbildeten) aufweisen. Was aber ist die Ursache solcher Orthogenesen? Entweder wird die Entwicklung durch eine immer gleichsinnige, auf zunehmende Adaptation gerichtete Selektion (Orthoselektion) besorgt, oder die ‹richtenden› Kräfte liegen im Organismus selbst (Autogenese). Im ersten Fall wären es erststellig Umweltfaktoren, die Verlauf und Richtung der Phylogenese bestimmen, im letzteren Fall würden «Eigengesetzlichkeiten des Organismus» die Evolution richten. (Vgl. G. Osche, Grundzüge der allgemeinen Phylogenetik, in Handbuch der Biologie III, 2, 890 f). Wenn wir uns bei wenigstens einigen Fällen für die Autogenese entscheiden, dann aus einem besonderen Grund: Es gibt orthogenetische Evolutionsketten, die auf die Ausbildung eines Apparats hinzielen, der für alle Zwischenformen keinerlei Selektionsvorteil bietet (eher ist das Gegenteil der Fall, da nicht gebrauchstüchtige Organe in der Regel einen Selektionsnachteil bedeuten). Solche Erscheinungen lassen sich wohl nur als Autogenese interpretieren. WOLTERECK spricht in. diesem Zusammenhang von «evolutiver Organanalyse» und eine solche wollen wir am Beispiel des Weberschen Apparats bei den Cypriniden (karpfenförmige Fische) vorführen. Während bei den meisten Fischen die ‹Situation› der Schwimmblase, welche etwa die Tauchtiefe des Fisches angibt, nervös an das ZNS des Tieres gemeldet wird, haben die Cypriniden im Laufe ihrer Phylogenese dafür einen halbmechanischen Apparat ausgebildet. Der Webersehe Apparat besteht aus neun Elementen, fünf Knöchelchen einer ganz bestimmten Struktur und vier Bändern. Außerdem gehört dazu ein Loch in der Basiocipitale, durch welche die ‹mechanische› Information an das ZNS weitergegeben wird. Die Knöchelchen wurden aus Elementen der Rippen und der Wirbelsäule gebildet, sind also recht verschiedener Herkunft. Man wird wohl annehmen müssen, daß die genannten Elemente des Apparats auf Grund von Genmutationen nach und nach entstanden sind, dann wurde er erst funktionstüchtig, nachdem alle Teile produziert, worden waren. Gegen eine Ausbildung ‹auf einmal› spricht fol- 80 gende Überlegung: Die normale Mutationsrate pro Genlocus liege bei 10-6, die Zahl der Individuen jeder Generation bei 100 Mio. An jedem Tag entstünde eine neue Generation, das mutierende Gen habe zehn verschiedene Mutationsmöglichkeiten, von denen gerade eine die geforderte spezifische Gestalt eines der Elemente des Apparats ‹bewirke›. Dann wird nach Berechnungen von SIMPSON, der sicherlich ein ganz unverdächtiger Mechanist ist, wahrscheinlich nach 274 Mio. Jahren (unter den sehr vereinfachten Bedingungen) einmal eine gleichzeitige Mutation aller in Frage kommenden Gene eintreten, so daß der Webersche Apparat ‹auf einmal› entsteht. Nun kennt die Genetik immerhin Pleiotropenwirkungen, nach denen ein einziges Gen gleichzeitig mehrere Effekte an verschiedenen Elementen verursachen kann. Doch darf nicht übersehen werden, daß die «Pleiotropengene» in ihrer pleiotropen Wirkung nicht ganzheitsbezogen sind. Auch auf solche Weise läßt sich also die phantastische Unwahrscheinlichkeit der gleichzeitigen Ausbildung aller Elemente des Weberschen Apparats nicht erklären. Man wird nicht umhin können, einen qualitativen spezifischen Faktor anzunehmen, eben die Tatsache, «daß unter den sehr verschiedenen Möglichkeiten gerade diejenigen realisiert werden, welche für die spezifische Form des Ganzen notwendig sind. Das in plausibler Weise auf die wirklich einsichtig gewordenen Wirkungsmechanismen des organismisehen Lebens zurückzuführen, ist heute nicht möglich. Ob es einmal möglich sein wird, das zu glauben bin ich nicht imstande», meint J.KÄLIN (Struktur und Dynamik der Materie, Freiburg und München 1967, 148 f). Wir werden wohl also nicht umhin können anzunehmen, daß der Webersche Apparat im Laufe vieler Millionen Jahre nach und nach entstanden ist. Die Deutungsschwierigkeit wird dadurch aber nicht geringer; es sei denn, wir verstehen das Phylum der Cypriniden als biologische Einheit, die in ihrer Entwicklung ähnlichen Gesetzen gehorcht, wie wir ihnen bei der Vorstellung vom ontogenetischen Entwickeln einzelner Organe begegnen. Hier tritt nämlich oft etwas sehr Ähnliches auf: zunächst nicht zusammenpassende Teile werden nach und nach gebildet, so daß als Endergebnis eine funktionierende Struktur entsteht. Damit schließen wir unsere - sicher unvollständige - Aufzählung von grundsätzlich verschiedenen Strukturen und Verhaltensweisen biologischer und physikalischer Einheiten ab. Aus rein sprachtechnischen Gründen wollen wir die Differenzen derart zusammenfassen, daß wit biologische Einheiten, d. h. Einheiten, welche die genannten charakteristischen Merkmale besitzen, Individuen nennen. Wir wählen mit Absicht diesen belasteten Terminus, weil sich sehr viele Momente, die wir genannt haben, in der Philosophiegeschichte 81 in einer mehr apriorisch-ontologischen Betrachtung als dem Individuum» zugehörig erkannt wurden, hier wiederfinden: Eigenstand, Selbstand, Vollstand und die Dynamik auf die Bewahrung von Eigen- und Selbstand sowie auf die Erlangung des Vollstandes. Dabei wollen wir jedoch nicht im Gegensatz zur Sprachtradition, den Begriff

«Individuum» ontologisch als eine Vervollkommnung in der Linie der Substanz begreifen. Wir sind im Laufe unserer Darstellung einer ganzen Reihe solcher biologischen Einheiten (Einheiten, die die biologischen Grundfunktionen wie Stoffwechsel, Vermehrung, Mutabilität...) zeigen und zugleich unserer Begriffsbestimmung Individuum gehorchen, begegnet: Einzellern, ‹Kolonien› von Einzellern, die sich zu Kolonialindividuen zusammenschließen, Fusionen von Einzellern, Organismen und verschiedenen Arten von Überorganismen wie besondere Formen von Symbionten, von Tiersozietäten und von stammesgeschichtlich verbundenen ‹Phylen›. Merkwürdigerweise ist der in der Geschichte der Naturphilosophie umstrittenste Fragepunkt der der ‹Einheit› des ‹Organismus› (im klassischen Sinn) gewesen. Wir wollen dazu noch eine Anmerkung anschließen: Bekanntlich kann man einzelne Gewebeteile auch von den höchsten Organismen in vitro (auf häufig zu wechselnder oft sehr spezifischer Nährlösung oder einer entsprechenden Nährgelatine) ‹lebendig› erhalten. Das heißt, sie zeigen wesentliche biologische Merkmale des Lebendigen wie Stoffwechsel und Vermehrung. Sind sie naturphilosophisch betrachtet Individuen (das ist ja, wie aus dem Vorhergesagten wohl ersichtlich geworden ist, kein biologischer Begriff, sondern ein naturphilosophischer)? Ich denke nicht. Aber diese Antwort bedarf einer Begründung.
3. Das Individualprinzip Wenn sich Individuen in wesentlichen Merkmalen und Funktionen von materiellen Einheiten unterscheiden, muß es einen zureichenden Grund für diesen Unterschied geben. Wir wollen diesen Grund der Individualität Individualprinzip nennen. Da sich im Bereich des Materiellen als solchem ein diese spezifische Einheit begründendes Prinzip nicht funktional ausdrückt, somit also die physikalische Grundstruktur von der biologischen Grundstruktur funktional grundsätzlich unterschieden ist, wäre es nicht sinnvoll, ein solches aktuales Prinzip auch für den Bereich physikalischer Einheit als solcher anzunehmen (obwohl es durchaus sinnvoll ist, auch hier nach einheitsstiftenden Gründen zu fragen), wollen wir doch - und das ist ein wohl allgemeines philosophisches Gesetz -keine aktuellen Prinzipien da annehmen, wo sie sich weder unmittelbar 82 noch mittelbar ausdrücken. Das aber bedeutet, daß wir für die Begründung der Individualität ein spezifisches Prinzip verlangen müssen. Die uns hier interessierende Frage ist die nach den Eigenarten eines solchen Prinzips. Die entscheidende Eigenart ist jene, daß sie nicht die Funktionen physikalischer Einheiten als solcher bestimmt. Es ist somit kein physikalisches und auch von der Physik nicht zu entdeckendes Prinzip. Kann man vielleicht noch mehr darüber ausmachen? Wenn wir mit der vorwiegenden Tradition abendländischen Denkens den Begriff Materie nicht als Chiffre für alles Reale benutzen, sondern ihn auf den Objektbereich der Physik einschränken, wäre unser Prinzip also ein nichtphysikalisches (sondern ein ‹biologisches›), und damit nichtmaterielles Prinzip. Hüten wir uns jedoch, nichtmateriell mit geistig zu verwechseln: sicher ist alles Geistige - wenigstens in der von uns gewählten Begriffsbestimmung von Materie - immateriell, aber keineswegs ist alles Immaterielle schon geistig. Unter geistig wollen wir hier eine Eigenschaft von Strukturen verstehen, die auch ohne aktuelle Bindung an Materie bestehen können. So ist nach allgemeiner Ansicht im Raum monotheistischer Religionen Gott, aber auch die menschliche Seele geistig. Während Gott kein realer Bezug zur Materie zugesprochen wird, wird die menschliche Seele niemals ganz aus ihrem Bezug zur Materie entlassen, sie bleibt stets grundsätzlich und unaufhebbar auf Materie hingeordnet, doch kann diese. Hinordnung wenigstens ‹vorübergehend› als reine ideale Hinordnung bestehen, die sich nicht in Materie aktualisiert. Im Gegensatz dazu sagt (nicht geistige) Immaterialität einen notwendigen realisierten Bezug zur Materie aus. Es läge nun nahe anzunehmen, daß das Individualprinzip auf die Materie einwirke, derart, daß es durch sein Wirken typische Funktionen verursacht, die anders nicht zustande kämen. Doch würde eine solche Annahme den ontologischen Eigenstand des geforderten Prinzips in einer Weise konkretisieren, den unsere biologischen und naturphilosophischen Reflektionen in keiner Weise rechtfertigen. Offenbar bilden Individualprinzip und biologische Funktion eine nicht zu trennende strukturelle Einheit. Wir wollen also durch die Forderung der Existenz eines immateriellen Prinzips als Grund des spezifisch biologischen Funktionierens keineswegs eine zusätzliche strukturelle Vielheit in das Individuum hineintragen. Es gibt keine biologischen Fakten, die uns zu einer solchen Konsequenz nötigten. In erster Annäherung mag die aristotelische Lehre von Form und Materie eine brauchbare Vorstellung des Gemeinten vermitteln. Die 83 metaphysische Form wirkt keineswegs auf die Materie ein, sondern informiert sie. Sie ist keine dem Lebewesen, physisch betrachtet, äußerliche Funktionszentrale, sondern bestimmt das Ganze, insofern es ganz ist (gleichsam von innen her). Das Individualprinzip ist so ein dem Ganzen innerliches Prinzip, das das Ganze funktional informiert. Solche Prinzipien wurden in der scholastischen Philosophie nicht als Seiende verstanden, die an sich oder durch andere sind, sondern als Seiende, durch die einer Ganzheit ein spezifisches Sein zukommt. Nun hatte die scholastische Philosophie diese Prinzipien allzu sehr substantialisiert. So wurde das ‹Lebensprinzip› (die metaphysische Form des Lebendigen, insofern es Lebensfunktionen zeigt oder zeigen kann) als Teilsubstanz verstanden, die mit dem materiellen Prinzip eine Vollsubstanz bildet. Von einer solchen Substanzvorstellung haben wir schon nachdrücklich

distanziert (WS II, 381 f). Der Begriff Substanz ist - jedenfalls im klassischen Verständnis dieses Wortes (vgl. WS II, 337 f) - nicht auf das Individualprinzip anzuwenden. Wir erklärten das Individuum als eine Struktur, die sich durch ihre Funktion spezifisch von der anderer Strukturen unterscheidet. Somit wäre das Individualprinzip als Strukturelement (der Begriff der Struktur scheint das Aufgebautsein aus verschiedenen oder gleichartigen Elementen zu implizieren) zu charakterisieren. Die Elemente einer solchen Struktur müssen jedoch nicht aufeinander wirken, es genügt eine innere Information und ein solches inneres Informationszentrum, um von Strukturelementen zu sprechen. Die vom Individualprinzip ausgehenden ‹Impulse› sind demnach nicht als eigentliche Funktionen zu denken, sondern als Funktionselemente, die jede typische biologische Funktion mit aufbauen. Die klassische Substanzontologie nahm eine ganze Hierarchie von substantiellen Formen an, deren niederste möglicherweise für den Bereich der Chromosomen anzusetzen wäre. So schreibt E. WASMANN, daß man eine gewisse Individualität schon den Chromosomen des Zellkerns zuerkennen müsse, da sie zu jeder Zeit und durch lange Generationen ihre Selbständigkeiten bewahren, «indem sie stets wieder in bestimmter Zahl und in bestimmter Gestalt der Zellteilungen zum Vorschein kommen und sich innerhalb dieser Grenzen durch Spaltung und Wachstum gewissermaßen selbständig fortpflanzen und entwickeln» (Die moderne Biologie und die Entwicklungstheorie, Freiburg 31906, 190). TH. BOVERI hat eine ganze Theorie der Chromosomenindividualität aufgestellt, und von ihnen als «elementarsten Organismen», «selbständigen elementaren Lebewesen», die in den Zellen ihre selbständige Existenz führen, gesprochen (Ergebnisse über die Konstitution der chromatischen Substanz 84 des Zellkerns, Jena 1904, 4-23). Doch hat WASMANN schon sehr genau zwischen Individuen und biologischen Einheiten unterschieden. Er meint, daß die Chromosomen «nicht biologische Elementareinheiten innerhalb der Zelle» sind, sondern nichts als «wesentliche morphologische und physiologische Bestandteile der Zelle». Gleichsam wie an einem Dogma hing er und seine Zeit der Meinung an, daß die Zellen «tatsächlich die niedersten Elementarteileinheiten des organischen Lebens sind» (ibd. 170 und 190 ff). Der Begriff Individuum scheint in dieser Gegenüberstellung von WASMANN recht metaphysisch interpretiert worden zu sein. Das würde u. a. heißen, daß Viren als nicht zelluläre Strukturen dogmatisch aus dem Reich des Lebendigen verbannt würden. Andererseits stimmt es wirklich, daß Zellen kleinste biologische Einheiten sind, denen schon der Charakter der Individualität zukommt? Sicher gibt es einzellige Gebilde, vor allem die Einzeller - Protophyten und Protozoen -, denen man mit einigen Schwierigkeiten alle Eigenschaften zusprechen .mag, die wir unter dem Begriff «Individuum» zusammengefaßt haben. Wie aber steht es mit der Individualität einer von einem mehrzelligen Organismus abgetrennten Zelle (oder eines Zellverbandes), die nicht mehr in der Lage ist, ein ‹Lebewesen› im herkömmlichen Sinn zu regenerieren, sondern nur noch Stoffwechselvorgänge zeigt, und sich etwa nach Verdoppelung ihres Genoms in vitro fortpflanzen kann, ohne daß eine irgendwie auf die Erhaltung oder die Ausbildung eines Vollstandes (es fehlt also jede irgendwie geartete Ontogenese im eigentlichen Sinn) gerichtete Funktion möglich wäre? Solche Zellen wären biologische Gebilde (denn sie erfüllen die biologischen Grundfunktionen des Lebendigen), nicht aber Individuen. Die Frage nach der Existenz eines Individualprinzips läßt also in diesem Fall relativ einfach entscheiden: Es gibt keine Gründe ein solches anzunehmen. Ich denke, man könnte grundsätzlich alle Funktionen eines solchen Gebildes auf biochemische Prozesse zurückführen. Dennoch bleibt es uns nicht erspart anzuerkennen, daß wir mit dem Begriff «Individuum- sehr verschiedene Gebilde bezeichnen. Es war einmal üblich, von Ordnungen der Individuen zu sprechen. Das wollen wir - um es vorweg zu sagen - nicht tun. Man unterschied: Individuen erster Ordnung seien die Protozoen und Protophyten; Individuen zweiter Ordnung die klassischen vielzelligen Organismen; Individuen dritter Ordnung die aus eigentlichen Zellfunktionen entstandenen Gebilde und die Tiersozietäten. Vermutlich hätte man unsere phylogentischen Einheiten als Individuen vierter Ordnung zu bezeichnen. Diese merkwürdige Einteilung wurde vermutlich von O. HERTWIG (Allgemeine Biologie, Jena 1923, 502 f) in die Biologie eingebracht. 85 E. MAYER hat diese Theorie philosophisch aufgearbeitet: er spricht von metaphysischen Individualitätsstufen, die er entweder nach metaphysischen Seins- oder Ursächlichkeitsstufen aufbaut (a. a. O. 231-237). Dem wollen wir hier nicht folgen, vor allem dann nicht, wenn man die Vielfalt der Individuen auf Hierarchien von metaphysischen Formen zurückführt, die in die jeweils höhere Individualitätsstufe eingehen bzw. in ihr untergehen, so daß sie im Einzelfall, etwa dem der abgetrennten Gewebe oder des Todes des Organismus, noch ein begrenztes und bedingtes Eigenleben fortsetzen können. Wie schon gesagt, möchten wir solche Gebilde nicht als Individuen verstehen. Mann kann aber auch die Individualitätsstufen verstehen nach dem Grad und der Intensität der Verwirklichung der Individualitätsmerkmale. Das scheint uns eine recht vernünftige Unterscheidung zu sein. Viren würden dann die unterste Stufe besetzen, Pflanzen (ein- oder mehrzellige) die zweite, Tiere die dritte. Wir wollen jedoch nicht von Wesensunterschieden sprechen, denn die Grenzen zwischen den einzelnen Stufen können durchaus fließend sein. In einem einzigen Fall scheint uns der Ausdruck «Überindividuum» angebracht zu sein: im Fall phylogenetischer Einheiten nämlich. Jedes Mitglied der Familie der Cypriniden ist ein eigentliches Individuum, weil und insofern es alle Eigenschaften, die als Merkmale im Begriff Individuum enthalten sind, besitzt. Das Phylum der Cypriniden,

phylogenetisch betrachtet, bildet dann eine diese Art von eigentlicher Individualität umgreifende und übergreifende Form von Individualität. Hier wäre das Individualprinzip als eigentliche Überstruktur zu verstehen.
4. Struktur Wir haben zur Erklärung der Eigenart des Individualprinzips den Begriff ‹Struktur› eingeführt. Er bedarf noch einiger Erläuterung. Heute versteht man unter Struktur im naturwissenschaftlichen Raum eine regelmäßige oder gesetzmäßige Anordnung von ‹Teilen› zu einem funktionalen Ganzen. Dabei ist der Begriff ‹Struktur› durchaus in einem weiteren Sinne verstanden, als wir ihn eingeführt haben. Elementarteilchen etwa besitzen keine Individualität, wohl aber eine Struktur, die sich in Meßergebnissen ausdrückt. Nicht ein ungeordnetes Chaos von Meßergebnissen bildet eine physikalische Struktur, sondern eine nach mathematischen oder geometrischen oder sonstwie gearteten Gesichtspunkten aufgebaute Ordnung solcher Daten. So wäre also etwa ein Elektron nichts als eine aus Meßergebnissen gebildete Struktur, für die bestimmte, aus der. Diracgleichung abgeleitete, Gesetze bestimmend 86 sind (vgl. A. March, Die physikalische Erkenntnis und ihre Grenzen. Braunschweig 1955). Eine solche Struktur bildet dann das System Elektron. Uns beschäftigen hier nicht diese Strukturen, sondern solche, die sich als funktionale Einheit vorstellen. Dabei unterscheiden wir mit der modernen Strukturpsychologie zwischen Struktur und Phänomen, die bekanntlich von S. FREUD noch nicht genügend auseinandergehalten wurden. Die Struktur liegt dem Phänomen zugrunde. Sie drückt sich im Phänomen aus. Da sich die Naturwissenschaften - also auch die Biologie - ihrer Beobachtungs- und Aussageintention nach auf Phänomene beschränken, ist der Begriff Struktur der Naturphilosophie zuzuordnen. Uns beschäftigen hier dynamische Strukturen, d. h. wir verstehen die Struktur von der Funktion, die immer phänomenale Funktion ist, her. Da es nun recht unterschiedliche biologische Funktionen gibt, die von einem Individuum geleistet werden, müssen wir diesen Funktionen Gliedstrukturen (F. Krüger) zusprechen, die aber ihrerseits durchaus wieder aus sehr unterschiedenen Strukturelementen aufgebaut sein können. Mit einigen erläuternden Gedanken wollen wir unsere Betrachtungen über die naturphilosophische Eigenart des Lebendigen abschließen, obwohl noch eine ganze Reihe von interessanten Fragen zu behandeln wären: Sind locker verbundene Metazoen (Hydoidpolypenstöcke, Korallenstöcke, Vergesellschaftungen von Moostierchen) oder fester verbundene Metazoen (Stöcke von Staatsquallen oder von Siphonophoren, das sind frei im Meer treibende Polypenstöcke) Individuen? .Scheinprobleme dagegen haben wir mit Absicht nicht behandelt: was geschieht mit dem Individualprinzip bei Zellteilungen, die zu neuen Individuen führen? Solche Probleme stellen sich nur in einer naturphilosophischen Betrachtung, die auf einer Substanzontologie aufruht. Nur dann, wenn man sich das Individualprinzip irgendwie hypostasiert denkt, es irgendwie abhebt von der Einheit der Funktion, es als reinen Funktionsgrund begreift (und nicht als Individualgrund), entsteht ein ernsthaftes Problem. Unserer Meinung nach entstehen solche Individualprinzipien mit einer Struktureinheit, wenn sie sich als Funktionseinheit ausspricht, neu. Woraus sie entstehen, was die Bedingungen und was die Ursachen ihres Entstehens sind, wird uns im zweiten Teil unserer Untersuchungen beschäftigen. Eins sei aber schon hier angemerkt: wegen ihrer Immaterialität sind Individualprinzipien sicher nicht teilbar oder zusammensetzbar, sie können nur neu entstehen oder untergehen. Ihre Teilung würde, wenn sie möglich wäre, eine Aufgabe der Struktur und damit den Tod des Lebewesens bedeuten. 87 Teilbarkeit setzt ein additives Zusnmmengesetztsein der Struktur aus Gliedstrukturen voraus. Und gerade ein solches Zusammengesetztsein schließt der moderne naturwissenschaftliche Strukturbegriff aus: weder Elektronen noch sonstige Elementarteilchen noch Individuen sind teilbar, sondern es entstehen, etwa beim Zerfall eines Elementarteilchens, zwei oder mehrere Elementarteilchen neu, es entsteht bei physischer Teilung ein Lebewesen neu. Das ist etwas ganz anderes, als wenn man einen Ziegelstein in zwei kleinere Einheiten zerschlägt, die ihrerseits durchaus wieder Ziegelsteine sein können. Ein Ziegelstein ist in unserem - durchaus eingeschränkten Sinne - keine Struktur, sondern ein Konglomerat, eine bloße (künstliche oder, im Fall eines Bruchsteins, natürliche) Anhäufung ungeordneter Teile, nicht grundsätzlich verschieden von einem Haufen Sand. TEILHARD DE CHARDIN nennt Strukturen echte Komplexitäten. Vielleicht ist hier der Ort, etwas bei der Theorie der echten Komplexitäten des großen Franzosen zu verweilen. Auch er kennt so etwas wie Individualprinzipien, es sind das Conscience (Bewußtsein) und Complexité (Komplexität). Solche Prinzipien sind keineswegs unmittelbar naturwissenschaftlich erhebbar, obwohl es auch Bewußtsein und Komplexität als empirische Gegebenheiten gibt. Die Verwechslung von empirischer Komplexität (im Sinne von echter Komplexität verstanden) und empirischem Bewustsein (wie es uns etwa am ausgeprägtesten im Menschen begegnet) mit beiden, als Prinzipien verstanden, hat zu den üblichen Fehlinterpretationen Teilhards geführt. Er selbst hat das von ihm Gemeinte mehrmals an seinem berühmten und doch so wenig beachteten Ellipsenbeispiel erklärt (vgl. Der Mensch im Kosmos, München 1959, 306; Die Entstehung des Menschen, München 1961, 129, Comment je vois §§ 2, 18 ff; Der Ort der Technik in: Die lebendige Macht der Evolution, Olten 1967,39). Um terminologisch verständlich zu bleiben, wollen wir einmal empirische Komplexität als Komplexität I bezeichnen, die Komplexität als Prinzip - in unserer Ellipse als Brennpunkt F, dargestellt - sei Komplexität II, das Bewußtsein als empirisches Datum sei Bewußtsein I, und als Prinzip (F2) werde es Bewußtsein II genannt. Jede Struktur (vom

Elementarteilchen bis zum Menschen) hat einen bestimmten Ort auf der Peripherie der Ellipse: ein Elementarteilchen ganz in der Nähe von F,, der Mensch ganz in der Nahe von F2. Die Nähe zu einem der Brennpunkte gibt den charakteristischen Einfluß der beiden Brennpunkte (= Prinzipien) auf die Struktur (und damit auch auf die Funktion) des Gebildes an. Die Evolution des Kosmos (Kosmogenese) geschieht längs der Ellipse, dabei verändern sich jedoch auch die Brennpunkte: F1 wan- 88 dert nach F2. Damit wird die Evolution irreversibel, denn der Prinzipienbereich unterliegt einer immanenten Gesetzmäßigkeit, die von den Geschehnissen, die sich in der Zone des Empirischen abspielen, nicht unmittelbar betroffen ist. F, und F2 sind als Individualprinzipien - wenn wir einmal diesen von TEILHARD nicht gebrauchten Terminus verwenden dürfen - auch Motor der Evolution vom Elementarteilchen zum Menschen: genauerhin von Punkt A zum Punkt Ω die beide Christus repräsentieren. A den Schöpfer, Ω den Vollender. Dabei wird in dem Augenblick, in dem F, sich mit F, vereinigt, F2 als Ω (jetzt im Sinne eines Prinzips verstanden) wirksam. Die Ellipse wird zum Kreis, um dessen Mittelpunkt Ω alles geordnet ist, die Evolution hat ein Ende.
Die Annahme zweier Individualprinzipien im dialektischen Bezug ist ein Charakteristikum der Naturphilosophie TEILHARDS. Wir haben uns mit Absicht nicht zu dieser Auffassung bekannt, nicht weil wir sie für falsch halten, sondern weil sie uns nicht genügend am biologisch-empirischen Material erhebbar scheint. Erst wenn man die Gesamtwirklichkeit, vor allem das Phänomen Mensch mit in die naturphilosophischen Überlegungen hineinnimmt, wird man der Konzeption TEILHARDS folgen dürfen, vielleicht gar müssen. Da wir uns aber auf das Phänomen biologisches Leben› beschränken, können wir sehr viel weniger über die Natur des Individualprinzips ausmachen. Doch in einem wesentlichen 89 Punkt wissen wir uns mit TEILHARD einig: jedem Phänomen dieser Welt ordnet er eine eigene Ellipse zu und dabei entpuppen sich die beiden Individualprinzipien keineswegs als physisch separate Strukturen, sondern als das Innen jeder Struktur, das nicht wirkursächlich das Ganze führt, sondern es gleichsam von innen (als innere Ursache) führt, informiert. Es sei hier noch ein Gedanke angefügt, den wir mit Absicht bislang aus unseren Überlegungen ausgeklammert haben; die Frage nach dem Verhältnis von Lebewesen und Substanz. Wir wollen Substanz nicht als Seiendes, das ist, sondern als Prinzip (Seiendes, wodurch etwas ist) verstehen, wie wir es in Anlehnung an antike Theorien schon getan haben (vgl. WS II, 381 f). Substanz ist in unserem Verständnis das, wodurch ein Seiendes an und für sich da ist. Die Scholastik pflegte dieses Prinzip mit dem Terminus ‹Sein› zu beschreiben. Weil und insofern nun aber ein Lebewesen, insofern es da ist, belebt ist (und nicht nur insofern es so ist) können wir das Leben als das, wodurch es da ist, als seine Substanz bezeichnen. Zur Substanz des Lebewesens gehört also durchaus auch sein Individualprinzip. Wenn die Formulierung nicht so mißverständlich wäre, könnte man durchaus von der «Substantialität des Individualprinzips» sprechen, denn das Lebendige ist nicht Substanz, aber es hat Substanz, weil und insofern es da ist. Und weil und insofern es als Lebendiges nur da ist durch ein Individualprinzip, ist dieses substantiell. Mit viel Umsicht und einiger Mühe haben wir im Vorhergehenden einen ganz wesentlichen Unterschied zwischen biologischen und physikalischen Einheiten zu übersehen versucht. Es ist das der Unterschied des Ursprungs. Und mit dieser sicherlich bedeutsamsten aller Differenzen wollen wir uns jetzt im zweiten Teil unserer Überlegungen beschäftigen. 90 ZWEITER TEIL

DER URSPRUNG DES LEBENDIGEN
91 ERSTER ABSCHNITT
EMPIRISCHE DATEN ZUR FRAGE NACH DER ENTSTEHUNG DES LEBENS
Die Herkunft des Lebendigen ist sicherlich eines der ältesten Rätsel, welches die Natur dem Menschen aufgab. Wie eigentlich entsteht ein Lebewesen? Wie kam das Leben auf diese Erde? Diese beiden Fragen sollen uns jetzt beschäftigen. Zunächst aber und vor allem die Frage nach dem ersten Entstehen des Lebendigen auf unserer Erde. Die Beantwortung der ersten Frage wird uns helfen, die zweite recht zu verstehen. Wir wollen nicht die Frage behandeln, ob es auch auf anderen Planeten unseres oder irgendeines anderen Sonnensystems Lebewesen gibt und welche chemische Struktur diesem Leben zugrunde liegen könnte oder müßte, denn über all das wissen wir nichts. Und worüber man nichts weiß, sollte man bekanntlich schweigen. Wir wollen auch hier jede Art von Kosmozoentheorie ausschließen, nicht weil es unmöglich sei, daß primitivste Lebewesen einmal den Weg durch den Weltraum hin zu unserer Erde gefunden haben, sondern weil diese Hypothese die Frage der Lebensentstehung nicht beantwortet, sondern nur auf irgend ein anderes Gestirn verschiebt, und damit naturphilosophisch ziemlich uninteressant ist. In einem ersten Abschnitt seien zunächst wieder einige Daten zusammengestellt, die uns empirische Auskünfte über die Entstehung des ersten Lebewesens und über seine Beschaffenheit geben können, um dann in einem zweiten Abschnitt die damit aufgeworfenen philosophischen (und vielleicht auch theologischen) Fragen zu beantworten. 93 ERSTES KAPITEL
DIE CHEMISCHE EVOLUTION
Wir wissen heute, daß dem Auftreten des ersten Lebewesens auf dieser Erde eine Zeit chemischer Evolution vorangegangen ist: Unter «chemischer Evolution» wollen wir eine Phase der vorbiologischen Entwicklung verstehen, genauer: die Phase der Erdgeschichte, in der die physikalischen Bedingungen gegeben waren, primitivste Lebewesen zu erhalten bis hin zum ersten Auftreten der ersten Lebewesen (Eobionten). Die ältesten bekannt gewordenen Lebewesen (z. B. Stromatolithen von Bulawayo, Mikroorganismen der Fig-Tree-Serie) haben ein Alter von etwa 3 Milliarden Jahren, ja die von A. E. J. ENGEL neuerdings in Ouverwacht (Transvaal) gefundenen kugelförmigen Fossilien scheinen noch einige Jahrhunderttausende älter zu sein. Da zumindest die Organismen der Fig-Tree-Serie schon verhältnismäßig hochdifferenziert sind, schätzt man gemeinhin die Dauer der vorausgegangenen biologischen Evolution auf 1 Milliarde Jahre. Nimmt man an, daß die Erde seit etwa 4,5 Milliarden Jahren, physikalisch gesehen, Lebewesen Heimat sein konnte, bleiben für die chemische Evolution noch etwa 500 Millionen Jahre. Als die präbiologische Evolution begann, mußte die Atmosphäre der Erde fast sauerstofffrei (weniger als 0,02 % O,) gewesen sein, da hoch-molekulare organische Verbindungen in Gegenwart von Sauerstoff degenerieren. Zu dieser Zeit wird die Erde ihre erste Atmosphäre aus Wasserstoff und (im bescheidenen Umfang) Helium weitgehend verloren haben. Eine zweite Atmosphäre aus den Wasserstoffverbindungen der häufigsten Elemente der stellaren Materie dürfte entstanden sein: Wasserdampf (H2O), Ammoniak (NH3) und Methan (CH4), vermischt mit Spuren von reinem Stickstoff (N2) und Kohlenrnonoxyd (CO) sowie Resten der primären Atmosphäre. In dieser Frühzeit haben neben der Sonnenstrahlung kosmische Strahlen, elektrische Entladungen und radioaktive Prozesse der erdnahen Atmosphäre Energien zugeführt, welche die Verbindungen der Uratmosphäre sicher teilweise umstrukturierten. Diese Prozesse im Labor nachzuahmen, ist seit 1953 Ziel zahlreicher Versuche. Damals simultierte S. L. MILLER auf Veranlassung seines Lehrers UREY eine Uratmosphäre von Methan, Ammoniak, Wasser und Wasserstoff und schickte blitzartige Entladungen hindurch. Als Reaktionsprodukte erhielt er u. a. 94 Glykolsäure (CH2OHCOOH),Milchsäure (CH3CHOHCOOH), Ameisensäure (HCOOH), Essigsäure (CH3COOH), Bernsteinsäure (CH2COOH) und eine Reihe von Aminosäuren. Dabei dürfte das Glycin (= Glycoll, die einfachste Aminocarbonsäure) über folgende Zwischenstufen entstanden sein:

Mit blitzähnlichen elektrischen Entladungen arbeiteten auch C. ΜΑTHEWS und R. E. MOSER (1966), die sie auf eine wasserfreie Uratmosphäre einwirken ließen. Sie synthetisierten dabei ein Polyaminomomotril, das sich in thermischer Nachreaktion mit Wasser zu Polygly (—COCH2NH—)n hydrolisierte. Versuche mit elektrischen Entladungen sind grundsätzlich auf G beschränkt und diese Beschränkung bedingt, daß die Molekulargewicht der entstehenden Verbindungen 300 kaum überschreiten können. Höher gewichtige Moleküle kommen in der Regel erst bei thermischen Nachreaktionen zu Stande. So liefen denn seit 1956 in Frankfurt (K. DOSE und B. RAJEWSKY) Versuche, die statt mit blitzartigen Entladungen ionisierender Strahlung arbeiteten. Die Ergebnisse ähnelten anfangs den Millerschen Entladungsversuchen. Inzwischen konnte die Möglichkeit einer strahlenchemischen Bildung von Peptiden unter präbiologischen Bedingungen nachgewiesen werden (1957, 1965)· Die einzelnen Schritte die zur Bildung des Diglycins führten, kann man durch Bruttogleichungen etwa wie folgt zusammenfassen:
95 Die durch ionisierende Strahlen erzeugten Verbindungen besitzen eine hohe Reaktionsfähigkeit. Das ist aber auch von Nachteil, da organische Moleküle von Molekulargewichten über 500 mit größerer Wahrscheinlichkeit zerstört als polymerisiert werden. Durch ionisierende Strahlung wurden also in der präbiologischen Entwicklung wahrscheinlich viele relativ niedermolekulare Verbindungen synthetisiert. Ein ähnlicher Effekt dürfte dem Einfluß von ultravioletter Strahlung zuzusprechen sein, da in diesen Strahlungsbereich nur Strahlen von Wellenlängen unter 2000 A von der Uratmosphäre absorbiert wurden. Bei einer weiteren photochemischen Umsetzung der Primärprodukte entstehen eine Reihe von interessanten hochmolekularen Verbindungen. Vor allem seien hier die Versuche von PONNAMPERUMA (1963) genannt. Er erhielt u. a. aus Adenin, Ribose und Wasser Adenosin, aus Adenin, Ribose und Äthylmetaphosphat ATP und aus HCOH und H2O Ribose und Desoxyribose, Verbindungen also, die für den Aufbau von Nukleinsäuren wesentlich sind. Im gleichen Jahr gelang SZUTKA bei Verwendung von Strahlen (X = 2537 A) die Erzeugung von porphyrinähnlichen Verbindungen aus Pyrrol (NH(CH)4), Wasser und Aldehyden. Chlorophyll, Hämoglobin sind Porphyrinderivate. Von besonderem Interesse sind auch seit 1960 (S. W. Fox) laufende Versuche, durch vulkanische Einflüsse die Entstehung von hochmolekularen Eiweiß- und Nukleinsäurebausteinen zu erklären. Fox erhielt bei Temperaturen um 1000° C aus Methan, Wasserdampf und Ammoniak verschiedene Aminosäuren, als er die Dämpfe dieser ‹Uratmosphäre› über auf etwa 1000° C erhitztes Kieselgel, Tonerde (Al2O3), Lava usw. leitete. Nun ist es sehr gut möglich, daß im Verlauf der präbiologischen Entwicklung in Bereichen vulkanischer Tätigkeit Seen austrockneten. Dabei bildeten sieh zunächst konzentrierte Lösungen organischer und anorganischer Verbindungen. Nach völligem Austrocknen werden bei möglichem Kontakt mit heißer Lava recht gute Voraussetzungen für die Ausbildung hochpolymerer Verbindungen aus den niedermolekularen Verbindungen des Sediments bestanden haben. Möglicherweise haben anorganische Polyphosphate, die bei Temperaturen um 300° C aus einfachen Phosphaten entstehen, bei verschiedenen Prozessen als Energieträger eine Rolle gespielt (Fox). Auch organische Polyphosphate, die sich etwa aus Äthern und anorganischen Polyphosphaten bilden können, mögen eine ähnliche Rolle gespielt haben. Die auf den ersten Blick erfolgreichsten Simulationsversuche wurden 1962 von SCHRAMM bei Temperaturen um 60° C mit Arginin, Desoxynukleosidphosphat und Glukose in wasserfreier Äthylmetaphosphatlösung durchgeführt. Schramm erhielt eine Reihe hochpolymerer Ver- 96 bindungen. Es ist jedoch kaum wahrscheinlich, daß auf der primitiven Erde jemals solche Bedingungen bestanden haben sollten. Sehr viel wahrscheinlicher reichet ten sich in Bereichen vulkanischer Tätigkeit unter Ionenaustauschreaktionen der erhitzten Mineralien u. a. auch wasserfreie Gemische organischer Verbindungen (Aminosäuren...) an. Solche ‹trockenen› Aminosäuren können in Gegenwart von 150° C -180°C heißer Lava in einigen Stunden hochpolymere, eiweißähnIiche Verbindungen mit Molekulargewichten von über 5000 aufbauen. Wegen ihrer großen Ähnlichkeit mit Proteinen nennt man solche Verbindungen Proteinoide. Sie lassen sich z. B. aus in Simulationsversuchen erhaltenen Reaktionsgemischen durch Gelfiltrationen als homogene Fraktionen isolieren. Proteinoide können wie eigentliche Proteine hydrolytitsch in Aminosäuren gespalten oder durch proteolytische Enzyme abgebaut werden. Auch zeigten verschiedene Proteinoidfraktionen enzymatische Aktivität (Hydrolase-, Decarboxylase-, Transaminase-, Oxidase-Aktivitäten). Auch hier waren die Versuche von S. W. Fox (1965) richtungsweisend. Daneben gibt es jedoch beachtenswerte biologische Unterschiede zwischen Proteinen und Proteinoiden. Letztere führen etwa - wenn sie einem höheren Organismus injiziert werden - nicht zur Ausbildung von Antikörpern. Sieht man einmal von den ziemlich konstruierten Simulationsversuchen Schramms ab, gibt es für die Polykondensation

von Nukleotiden zu einer präbiologischen Nukleinsäure keine Belege aus Simulationsversuchen. 1965 hat zwar eine chilenische Forschergruppe unter A. E. AQUILERA durch Gammabestrahlung verdünnter Lösungen der Uridylsäure Polyuridylsäure erhalten. Doch sind solche Versuche weder genügend überprüft, noch zeigen sie eine der natürlichen Eiweißsynthese vergleichbare Leistung. Damit stellt sich uns die Frage, ob in der präbiologischen Evolution Nukleinsäuren und Proteine gleichzeitig an der Bildung primitiver biologischer Strukturen teilgenommen haben. Dafür scheint unsere Erfahrung im Umgang mit primitivsten Lebewesen oder quasibiologischen Strukturen (einige Viren) zu sprechen. Sie zeigen einen deutlichen biologischen Primat der Nukleinsäure. Ein zeitlicher Primat der Nukleinsäure ist aber, nach dem heutigen Stand unseres aus Simultationsversuchen stammendes Wissen, für die ersten biologischen Strukturen auszuschließen. Ein weiterer Schritt gelang E. ITO und J. L. STROMINGER, die (1960) zeigen konnten, daß das Prinzip der Ordnung und der Informationsübertragung bei der biologischen Eiweißsynthese (grundsätzlich wenigstens) primitiver Lebewesen auch ohne die Kontrolle durch eine Nuklein- 97 säurematrix aufrecht erhalten werden kann. Auch die thermische Bildung von Proteinoiden erfolgt nicht durch eine willkürliche oder ‹zufällige› Aneinanderreihung beliebiger Aminosäuren. Hier werden Ordnungsprinzipien erkennbar, wie sie uns ganz ähnlich aus dem Bereich der Eiweiße bekannt sind. Aber sie werden durch die Struktur der benachbarten Aminosäuren und den Einfluß der anorganischen Umgebung (wenigstens teilweise) bestimmt (W. GROTH und H. VON WEYSENHOFF). 98 ZWEITES KAPITEL
PROTEINOIDE UND MIKROSPHÄREN
Im Laufe der Simulationsversuche mit thermischer Kondensation gelang es, Proteinoide mit einem Überschuß an (der basischen Aminosäure) Lysin herzustellen, die somit, wie die meisten natürlichen Proteine, schwach basisch reagierten. Bei der Untersuchung der quantitativen Zusammensetzung von Proteinoiden stellte man fest, daß in den Proteinoiden keineswegs die gleichen Mischungsverhältnisse von Aminosäuren vorkamen wie im Ausgangsgemisch. Das Verhältnis glich sehr viel mehr dem der natürlichen Proteine: bestimmte Aminosäuren und Aminosäure-Sequenzen werden deutlich bevorzugt. Auch der häufige Einbau bestimmter Aminosäuren am Amino- oder Säureende entsprach ziemlich genau der Struktur natürlicher Proteine. Das aber bedeutet, daß - wie schon erwähnt - auch ohne die DNS-Information bestimmte Strukturen in Aminosäurepolymeren auftreten können. Mit den Proteinen haben die Proteinoide ferner gemeinsam, daß sie enzymatisch wirken und ähnliche Funktionen übernehmen wie die Proteinenzyme: G. KRAMPITH und S. W. Fox zeigten, daß Glykose über Glycoronsäure von Proteinoiden zu Kohlendioxyd abgebaut wird. G. KRAMPITH und H. HARDENBECK beobachteten die Spaltung von Brenztraubensäure (CH3—CO — COOH) in Essigsäure (CH3—COOH) und Kohlendioxyd bei Anwesenheit von Proteinoiden, eine Reaktion, die in biologischen Strukturen von großer Bedeutung ist. D. ROHLFING konnte p-Nitrophenylacetat in Gegenwart saurer Proteinoide hydrolisieren und in Gegenwart basischer Proteinoide Oxalacetat in Brenztraubensäure umwandeln. Die Proteinoide wären also durchaus in der Lage gewesen, einen Urstoffwechsel in Gang zu bringen. Besonders interessant ist die Fähigkeit der Proteinoide, sich zu größeren Aggregaten zusammenzulagern: Kühlt man eine heiße Lösung von Proteinoiden ab, entsteht eine Trübung und schließlich ein Niederschlag, der sich unter dem Mikroskop als aus kugel- oder fadenförmigen kokkengroßen Mikrosphären aufgebaut entpuppt. Diese Mikrosphären zeigen manche Eigenschaften frühvitaler Strukturen: sie lassen sich zentrifugieren, ohne zusammenzufließen, sie zeigen osmotische Eigenschaften, haben eine Doppelmembran, durch die, bei rascher Änderung des Säure-wertes der umgebenden Lösung, ohne Verletzung Stoffe in die Mikrosphäre eindringen können. Auch haben sie die katalytische Wirkung 99 mancher Zellen: Mikrosphären, die Zinkionen enthalten, spalten Phosphatgruppen von ATP ab und setzen Energie frei. Wir haben in den Mikrosphären organismische Einheiten gefunden, die in Laborversuchen entstehen, ohne daß biologisch entstandene Verbindungen zu Hilfe genommen werden müßten. Andererseits wäre es verfehlt, Mikrosphären als Eubionten zu bezeichnen, erfüllen sie doch neben dem eiweißartigen Aufbau keines der von uns genannten Lebenskriterien. Vor allem kennen sie wegen des Fehlens des genetischen Materials keine identische Reduplikation, wenn man von Knospung und Teilung nach der Art flüssiger Kristalle absieht. So ist es nicht verwunderlich, daß die biochemische Forschung das Problem der Synthese eines primitiven reduplikativen Systems anders anzugehen versucht als durch Simulation der Zustände der frühen Erde und ihrer Atmosphäre. Eine entsprechende Technik wurde von der Molekularbiologie als Rekombinationsmethode entwickelt. Bei Experimenten solcher Art werden bestimmte Zellfragmente isoliert und unter geeigneten Bedingungen mit nicht biologisch entwickelten Polymeren oder Mikromolekülen, etwa Aminosäuren oder Nukleotiden in Verbindung gebracht. S. SPIEGELMANN konnte zeigen, daß eine Virusnukleinsäure mit Hilfe des Syntheseapparates der Wirtszelle - und nur mit Hilfe eines solchen -ein Enzym (Replicase) produziert, das dann wiederum die Virusnukleinsäure kopiert. Isoliert

man die Replicase von den befallenen Zellen, ist eine Virusvermehrung in vitro möglich. SPIEGELMANN arbeitet mit RNS-Viren (die RNS ist Träger der genetischen Information) Q-Beta und MS 2, mit denen er Bakterien (Escherichia coli) infizierte. Die entstehende Replicase isolierte er. Sie besorgte in vier Stunden die Synthese großer Mengen spezifischer RNS, die in Zellen eingebracht wieder richtige Viren erzeugte (vgl. Seite 147). Eine zweite Theorie über die Entstehung der ersten Lebewesen wurde vor allem in der Sowjetunion entwickelt: die Koazervattheorie. 100 DRITTES KAPITEL
KOAZERVATE
Der Holländer G. BUNGENBERG DE JONG unternahm wohl als erster Versuche mit Koazervaten. Solche Koazervate lassen sich unter Laboratoriumsbedingungen leicht herstellen: einfache Lösungen verschiedener Proteine oder anderer hochmolekularer Stoffe vermischt man bei Zimmertemperatur und einem bestimmten - meist nicht sehr hohen - pH-Wert in einem wässerigen Medium miteinander. Dann bilden sich nach einiger Zeit mikroskopisch sichtbare Tropfchen aus, die deutlich voneinander und dem umgebenden Medium unterschieden sind. BUNGENBERG DE JONG nahm an, daß bei der Bildung dieser «Koazervate» genannten Tröpfchen elektrostatische und «Hydrationskräfte» eine Rolle spielen, doch sind quantitative Untersuchungen darüber noch nicht abgeschlossen. Bei der Bildung von Eiweißkoazervaten findet ein teilweiser Übergang von Lipoiden (fettartigen Verbindungen) und Polysacchariden in die Grenzfläche der Eiweißmolekel und die Ausbildung von Molekülschichten mit veränderten strukturmechanischen Eigenschaften statt. Auch können Eiweißkoazervate eine innere Struktur aufweisen, die recht stabil sein kann. In Verbindung mit einer bestimmten Oberflächenorganisation und einer relativ stabilen inneren Struktur können Koazervate die Fähigkeit entwickeln, Stoffe ihrer Umgebung zu absorbieren. Selbst wenn ihre Konzentration im Medium i:100000 nicht übersteigt, werden viele organische Verbindungen fast vollständig von dun Koazervaten aufgenommen (Oparin, a. a. O. 63 f). A. L OPARIN hält solche Koazervate für die wahrscheinlichste Ausgangsform für die Organisation praevitaler Systeme. Als Gründe führt er an: 1. Die primär im Urozean gebildeten hochmolekularen organischen Verbindungen (Zuckerpolymere, Aminosäuren, Nuklcotide) dürften sich in kolloidchemischer Hinsicht kaum von den uns heute bekannten Verbindungen unterscheiden, d. h. sie bilden in wässriger Lösung Koazervate. 2. Da die Konzentration solcher Verbindungen in Teilen des Urozeans wahrscheinlich bei 1 :1000 gelegen haben dürfte, wird es wohl kaum einen anderen so wirkungsvollen Weg zur Konzentration eiweißähnlicher und anderer hochmolekularer Stoffe gegeben haben als den der Koazervatbildung. 101 3. Koazervate sondern sich nicht vollkommen von ihrem Medium ab, sondern wechselwirken mit dem Medium (Stoffaustausch). Damit ist eine wichtige Voraussetzung für eine praevitale Evolution erfüllt. 4. Das Protoplasma biologischer Systeme stellt in kolloidchemischer Hinsicht ein sehr komplexes Koazervat dar. Im Gegensatz zu den im Urozean entstandenen Koazervaten bleibt die dem ‹lebenden› Protoplasma eigene Koazervatstruktur nur solange erhalten, wie in ihm biochemische Prozesse (enzymatischer Natur) in meist großer Zahl ablaufen (Oparin, ibd. 64 f). Ein entscheidender Schritt in der Theorie OPARINS ist die nun folgende Ausbildung von Koazervaten mit immanenten chemischen Prozessen. Die chemischen Prozesse können entweder unabhängig von der Art des umgebenden Mediums verlaufen, dann müßte das Koazervat früher oder später zugrunde gehen, oder sie können bei aller primitiven ‹Individualität›, die ihnen OPARIN wegen ihrer nicht absoluten Bindung an die Umgebung zuspricht (a. a. O. 75), in Übereinstimmung» mit der Umgebung stattfinden, dann wäre die Beständigkeit des Koazervats weitgehend gesichert. Es entstanden so vorwiegend solche Systeme, die durch die ständig in ihnen ablaufenden chemischen Prozesse der Selbsterzeugung zur Selbsterhaltung auf Grund einer ständigen Wechselwirkung mit der Umwelt befähigt waren (ibd. 75 f). Ein weiterer Schritt auf eigentlich biologische Strukturen hin war der des Wachstums. Unter Wachstum versteht OPARIN die Vergrößerung der Masse auf Kosten der Stoffe im umgebenden Medium. Die für die Selbsterhaltung und das Wachstum nötigen Energien wurden dem Koazervat «hauptsächlich durch relativ energiereiche organische Verbindungen, die in den Tropfen gelangten und dort irgendwelchen chemischen Reaktionen unterworfen wurden» geliefert (ibd. 76). Die ‹Vermehrung› der Koazervate mag anfangs durch mechanische Teilung (Brandung, Wellenschlag) zustande gekommen sein. Für höher entwickelte Koazervate (dynamisch-stationäre Systeme) ist aber eine Vermehrung auch durch innere Ursachen möglich. Hier verweist OPARIN auf die Möglichkeit, daß durch Hydrolyse hochmolekularer Verbindungen der osmotische Druck im Koazervat so groß wurde, daß die Wände zerrissen (ibd. 77). Ein dritter Schritt auf die Ausbildung eigentlicher biologischer Strukturen hin wurde getan, als die Koazervate nicht nur dynamisch beständiger, sondern auch dynamischer wurden, d. h. Stoffwechselprozesse schneller und sicherer ausführten (ibd. 77 f). Sie entwickelten dabei ein konstantes System von Wiederholungen der in Wechselbeziehungen stehenden und miteinander koordinierten, zu einem einheitlichen Netz verbun-

102 denen, Reaktionen und damit die für biologische Systeme charakteristische Fähigkeit zur Selbsterneuerung aus. Von diesem Zeitpunkt an hält OPARIN die Koazervattröpfchen für belebt (ibd. 78). «Das von uns entworfene Bild der gesetzmäßigen Evolution auf dem Wege zur Entstehung des Lebens ist nicht nur hypothetisch. Es basiert auf dem Studium der chemischen Mechanismen, die wir heute in den lebenden Körpern und ihren isolierten Bruchstücken finden, aber auch der Untersuchung des Baus künstlicher Systeme, wie z. B. der komplexen Koazervattropfen, die Fermenteiweiß und andere Komplexe und einfache Katalysatoren enthalten» (ibd. 81). Die Koazervattheorie OPARINS mußte sich im Westen wie im Osten heftige Kritik gefallen lassen. Sie wird von ‹westlichen› Biologen zumeist abgelehnt, weil sie, obzwar schon durchaus als Modell brauchbar, doch nicht die praevitale Evolution verstehen lehrt, insofern sie stets von Verbindungen rezenter Organismen ausgeht (K. DOSE). Eine Frage aber ist allen diesen Theorien gemeinsam: Stammen alle Lebewesen von einem einzigen «Urlebewesen» ab, oder ist das Leben mehrmals auf der Erde entstanden? 103 VIERTES KAPITEL
ABSTAMMUNG VON EINEM EOBIONTEN
Alle entsprechend untersuchten Organismen besitzen gemeinsame chemische Merkmale, die es nahelegen anzunehmen, daß alle Lebewesen von einem Eobionten abstammen, wie schon ERNST HAECKEL (1866) vermutete. 1. Alle biologisch gebildeten Aminosäuren sind L-Isomere. Sie haben also die Konfiguration des L-Glycerinaldehyds, Die übliche Schreibweise charakterisiert das L-Glycerinaldehyd (gegenüber dem R-Glycerinalde-hyds) wie folgt:
Mit Ausnahme des Glycocolls haben alle Aminosäuren ein assymetrisches C-Atom, d. h. ein C-Atom, das so mit vier verschiedenen Atomen oder Atomgruppen verbunden ist, daß durch dieses C-Atom keine Symrnetfieebene gelegt werden kann. Solche Verbindungen sind optisch aktiv, d. h. sie haben die Eigenschaft, die Schwingungsebene von linear polarisiertem Licht zu drehen. Die meisten biologisch gebildeten Aminosäuren sind linksdrehend. 2. Alle biologisch gebildeten Zucker (Pentosen, Hexosen . . .) sind R-Isomere, d. h. sie haben die Konfiguration des R-Glycerinaldehyds. Bei technischen Synthesen von optisch aktiven Verbindungen entstehen, da beide Formen gleichen Energiegehalt haben, beide also die gleichen (chemischen) Chancen haben, immer razemische Gemische (d. h. gleich-gewichtige Gemische von R- und L-Formen). Aber nur eine Form ist physiologisch wirksam. So können Bakterien oder Schimmelpilze mit nur einer Form eines razemischen Gemischs etwas «anfangen». Nur sie kann angegriffen und zerstört werden. Das beruht darauf, daß ihre Enzyme, die selbst optisch aktiv sind, nur mit entsprechenden Verbindungen «reagieren». 104 Nicht selten versucht man die Bildung ausschließlich einer Form auf den Einfluß polarisierten Lichts bei der Entstehung der frühen Verbindungen zurückzuführen. Es ist tatsächlich durch Versuche belegt, daß bei Einstrahlung polarisierten Lichtes bevorzugt eine Komponente synthetisiert wird. Nicht erklären aber kann diese Hypothese, warum gerade L-Aminosäuren und R-Kohlehydrate ausgebildet wurden (also verschieden strukturierte Verbindungen). Zum ändern werden öfters chemische Struktur und optische Aktivität verwechselt. Die Gleichheit der biogenen Verbindungen bezieht sich keineswegs auf die Gleichgerichtetheit der optischen Aktivität, sondern auf die der chemischen Struktur. L-Verbindungen können durchaus rechtsdrehend und R-Verbindungen linksdrehend sein. 3. Von den (43)20 möglichen genetischen Codes (die bestimmt sind durch die Tripletts von 4 Nukleotidarten und 20 Aminosäuren) ist im biologischen Bereich nur eine Gruppe von 43 realisiert, wenn man einmal von geringen, wohl sekundär entstandenen, Variationen absieht, obgleich, soweit heute bekannt, die vorhandenen 43 Tripletts auf Grund ihrer Molekularstruktur oder Ladungskonfiguration nicht gegenüber den anderen bevorzugt sind. Es läßt sich natürlich nicht ausschließen, daß mehrere Eobionten entstanden. Einer aber nur kann sich zu der uns bekannten Form des Lebens entwickelt haben. Manche von ihnen waren sicher recht instabile Gebilde, andere werden sich bezüglich ihrer Stabilität aber kaum von der des am Anfang unserer Lebenslinie stehenden Eobionten unterschieden haben. Hier mag Zufall im Spiel gewesen sein. Man kann sich natürlich fragen, wie das Leben auf der Erde aussehen würde, wenn der Eobiont, der am Anfang alles Lebens gestanden hat, anders konstruiert gewesen wäre. Diese Frage wäre auch interessant für eine Spekulation über biologische Strukturen auf anderen Himmelskörpern. Die Beantwortung dieser Frage ist aber nach dem bisherigen

Stand unseres biologischen Wissens nicht möglich. 105 FÜNFTES KAPITEL
DIE LABORSYNTHESE DES LEBENDIGEN
Daß eine Laborsynthese des Lebendigen in absehbarer Zeit gelingen wird, läßt sich heute kaum mehr bezweifeln. Es sei hier ein Experimentalentwurf vorgestellt, der vielleicht schon recht bald realisiert werden wird: Wie wir schon erwähnten, kann man das Genom des Q-Beta Phagen (ein sehr kleiner Phage, der Escherichia coli befällt) in vitro durch eine biologisch gewonnene RNS Replicase vermehren. Anscheinend trägt jeder RNS-Faden des Phagen u. a. auch die Information (d. h. das Gen) für die Bildung der Replicase. Gelingt es nun, die Q-Beta-RNS in vitro zur Proteinsynthese zu verwenden, bei der sie als mRNS fungiert, so würde u. a. auch RNS Replicase entstehen. Diese könnte dann ihrerseits zur in vitro Vermehrung von Q-Beta-RNS dienen. Vereinigt man beide Systeme, entstünde jene gegenseitige Koppelung von Protein- und Nukleinsäuresynthese, die uns der entscheidende Schritt zum Leben zu sein scheint: Das RNS-Genom würde durch die Replicase gebildet und diese unter Steuerung durch jenes .entstehen. Als ‹Nahrung› wären diesem an sich chemischen System die RNS-Bausteine, die tRNS-Aminosäure sowie auch Ribosomen mit eventuell zugehörigen Übersetzungsenzymen zuzusetzen. Der ‹flüssige› Organismus aus RNS und Replicase würde sich dann identisch vermehren, leistete Stoffwechselfunktionen und würde auch gelegentlich mutieren (R. W. KAPLAN). Wir werden auf diese und noch weitreichendere Versuche im Schlußkapitel dieses Buches zu sprechen kommen. Wichtig ist jedoch die Feststellung, daß die ‹Nahrung› noch aus biologischen Systemen gewonnen würde, wenn sie auch selbst nicht mehr ‹belebt› wäre, d. h. wir hätten sicher kein Modell unseres Eobionten synthetisiert. Von einer Totalsynthese, im Sinne eines ausschließlichen Ausgangs von anorganismisch gebildeten Bausteinen, sind wir noch sehr weit entfernt. Doch wird man bei einer Synthese, die von ‹toten›, organismisch gebildeten Bausteinen, ausgeht, durchaus von einer echten ‹Laborsynthese› des Lebendigen sprechen dürfen. Eine ‹Vitalisierung› der Proteinoide ist dazu sicher nicht erforderlich. Wir haben im Vorhergehenden oft von Viren gesprochen. Es ist wohl angebracht, etwas Genaueres von diesen merkwürdigen Gebilden zu berichten. 106
EXKURS: VIREN
Viren können sehr wohl als Modelle primitivster Lebewesen gelten, wenn auch nicht als Modell des Eobionten, von dem wahrscheinlich alle auf der Erde lebenden biologischen Strukturen herstammen. Wichtig für das Problem des Lebendigen sind die Viren jedoch für die Nachahmung der Biogenese im Labor geworden, Als Entdecker der Viren [das Wort kommt von dem lateinischen virus (sing, tantum, n.) = Schleim, Gift] gilt der russische Botaniker D. IWANOWSKY, der 1892 der Kaiserlichen Akademie der Wissenschaften in Petersburg berichtete, daß der Saft von mosaikkranken Tabakpflanzen infektiös sei, obwohl der Krankheitserreger nicht von Bakterienfiltern zurückgehalten werde. Um 1900 kam dann das Wort filtrierbares Virus in Gebrauch zur Bezeichnung für Erreger von Infektionskrankheiten, die nicht durch die üblichen Methoden der Bakterienisolierung (Filtrieren etc.) dargestellt, nicht im Lichtmikroskop sichtbar gemacht und nicht in vitro gezüchtet werden können. Rein größenmäßig schließen die Viren die Lücke zwischen kleinen Bakterien und großen Molekülen. Das gleiche gilt für die Molekulargewichte. Das größte bekannte Molekulargewicht hat ein Protein, das Haemocyanin des Hummers, mit 6 700 000. Wir können hier nicht die vielen hundert bekannten Viren vorstellen, sondern wollen uns auf zwei beschränken, das Tabakmosaikvirus (TMV) und den T-Phagus.
1. Das Tabakmosaikvirus Das TMV ruft auf Blättern infizierter Pflanzen eine typische Fleckung hervor. Es ist auf der ganzen Welt verbreitet und befällt vor allem Nachtschattengewächse. Wir kennen heute 27 Familien des TMV mit vielen Unterarten. 1935 gelang es Wendel STANLEY vom Viruslaboratorium in Berkeley, das TMV durch fraktionierte Ammoniumsulfatfällung rein in parakristalliner Form aus dem Saft infizierter Tabakpflanzen darzustellen. Im folgenden Jahr zeigte F. C. BAWDEN, daß das Virus aus reinem Eiweiß und RNS besteht. Durch Erhitzen auf etwa 90° C in 5%iger Trichloressigsäure über 3o min kann man die Eiweißkomponente denaturieren und die RNS rein darstellen. 5,1'% des TMV besteht aus RNS. Die Basen verteilen sich nach Untersuchungen von R. MARKHAM und K. M. SMITH (1953) wie folgt: 107 Adenin: 31%, Guanin: 29,25%, Cytosin: 15,5%, Uracil: 24%. Der Rest besteht aus 2300 gleichartigen Proteinmolekülen mit einem Molekulargewicht von je 17000. Die wichtigsten Aminosäuren (in Mol-Prozent) sind: Aspargin (10,5-12,5 %), Serin (8,5-11 %), Alanin (9,2-10,5 %)› Valin (8-11 %), Glutaminsäure (8,5-10,5 %), Thryptophan (9-12 %), Prolin (4,8-5,7 %), Isoleucin u. Leucin (12,3-14 %). Die Zusammensetzung der Eiweißhülle

schwankt also bei verschiedenen Stämmen z.T. erheblich (einige Extremalweite sind möglicherweise auch durch Meßfehler bedingt). G. BRAUNITZER (1954) bestimmte das eine Ende der Polypetidkette: Glutaminsäure-Isoleucin-Prolin. Das andere Ende bildet stets Tryptophan. Unter dem Elektronenmikroskop erscheint das TMV als Stäbchen von etwa 300 nm Länge und einem Durchmesser von 18nm. Es hat ein Molekulargewicht von etwa 40 Millionen. Das TMV ist gegenüber Schwankungen der Säurewerte ziemlich unempfindlich und stabil zwischen pH 2 und pH 8. .Bei höheren pH-Werten wird das Viruseiweiß gespalten. Senkt man den pH-Wert wieder, so treten ab pH 6,5 die Bruchstücke wieder geregelt zusammen, umschließen sogar mitunter wieder die RNS und bilden intakte Viren. Dabei bleibt die RNS völlig unbeteiligt, denn auch das reine Viruseiweiß reagiert unter den gleichen Bedingungen ebenso: es entstehen leere Eiweißhülsen. Die Hülsen haben einen inneren Durchmesser von 4 nm. Sie werden je Rundung von 9 Proteinmolekülen spiralig aufgebaut. Der Proteinspirale entspricht im Innern eine RNS-Spirale aus etwa 6000 Nukleotiden. Die Vermehrung des TMV geschieht ausschließlich in lebenden Zellen, nicht weil es bestimmte schwer zu erfüllende Bedingungen an den Nährboden stellt, sondern weil ihm bestimmte Fähigkeiten abgehen.. Es fehlt ihm die Fähigkeit zu eigenem Stoffwechsel bzw. zur Energiegewinnung, die für diesen Stoffwechsel erforderlich ist. Ein TMV vermehrt sich auch nicht eigentlich, sondern wird von der erkrankten Zelle gebildet: es wird in die lebende Zelle ein zellfremder übermachtiger genetischer Code eingeschleust, der den Zellstoffwechsel so umorientiert, daß er Virus-NS aus den vorhandenen Bausteinen bildet. Träger des Code ist die RNS, die, auch wenn sie isoliert in gesunde Zellen gebracht wird, diese zur Bildung von intakten TMV zwingt (A. GIERER und G. SCHRAMM 1956). Doch verliert die reine RNS ihr infizierendes Vermögen nach etwa 48 Stunden fast völlig. Der Eiweißmantel hat also die wesentliche Funktion einer Schutzhülle. Die chemische Veränderung eines einzigen Nukleotids der RNS-Kette kann, aber muß nicht, Vermehrungsunfähigkeit bedeuten. 108 Von den beiden bekanntesten TMV-Stämmen (vulgäre und dahlemense) mit sehr unterschiedlichen Eigenschaften (vulgäre ist etwa auf Phaseolus vulgans nekrotisch und dahlemense symptomlos), kennt man Gelbstämme: flavum bzw. luridum. Von flavum leitet sich necans ab, von diesem revirescens. Diese Formen («Arten») sind eigentliche Mutanten der Ursprungsart, entstanden durch Änderungen der Basenverteilung in der RNS. Alle genannten Arten unterscheiden sich auch serologisch und elektrophoretisch voneinander. Diese Unterscheidbarkeit zeigt eine Differenzierung in der Anordnung der Aminosäuren der Eiweißhülle an. Während sich bei den beiden Wildstammen acht Aminosäuren voneinander unterscheiden, wird beim Übergang flavum - necans nur ein Alanin gegen ein Phenylalanin, beim Übergang necans - revirescens Asparginsäure gegen Alanin ausgetauscht. Solche Mutanten können auch künstlich erzeugt werden. Erwähnenswert sind die Versuche von H. SCHUSTER und G. SCHRAMM (1958). Sie behandelten die isolierte RNS mit salpetriger Säure. Dabei wurden einzelne Basen verändert: Cytosin in Uracil, Adenin in das normalerweise nicht vertretene Hypoxanthin. Die quantitative Auswertung ergab, daß die chemische Änderung nur eines Nukleotids zu veränderlichen erblichen Eigenschaften (= Mutanten) führt (K. W. MUDRY und A. GIERER, 1958).
2. T-Phagen Bakteriophagen sind Bakterien befallende und auflösende Viren. Sie wurden erstmals 1915 von F. W. TWORT beschrieben. Da man glaubte, durch Phagen bakterielle Infektionskrankheiten bekämpfen zu können, nahm sich zunächst die medizinisch orientierte Mikrobiologie der Phagusforschung an. Bis 1925 sind über 600 wissenschaftliche Arbeiten über Phagen erschienen. Sehr gut untersucht sind die Phagen der T-Serie der Coliphagen. Sie umfaßt wenigstens sieben Stämme. T2, T4 und T6 zeigen unter dem Elektronenmikroskop die gleiche Gestalt: sie ähneln in etwa einer Kaulquappe.. Auch serologisch sind sie nahe miteinander verwandt. Eine zweite Gruppe serologisch verwandter T-Phagen, T3 und T7, ist wesentlich kleiner. Sie haben einen runden Kopf und einen kaum erkennbaren Schwanz. T1 und T5 sind weder untereinander noch mit Phagen der beiden anderen Gruppen zu kreuzen. Sie ähneln - abgesehen von ihrer Größe - der erstgenannten Gruppe. Wie alle Viren, benötigen auch T-Phagen einige Zeit, um in der Bakterienzelle neue Viren auszubilden. Sie haben eine minimale Latenzzeit, die zwischen 13 (T1, T3, T7) und 25 min (T6) liegt. 109 Die T-Phagen befallen alle den B-Stamm des Darmbakteriums Escherichia coli. Wie alle Phagen bestehen sie aus Eiweiß und DNS. Die DNS ist knäulartig im Phageninnern gelagert. Der Nukleinsäureanteil beträgt bei T-Phagen 35 bis 50 %. Während die DNS der ungradzahligen Phagenstämme aus klassischen Basen aufgebaut ist, ersetzen die gradzahligen das Cytosin durch 5-Oxymethylcytosin. Sie bilden eine Base, die in Escherichia coli nicht vorzukommen scheint. Da weder Phagus noch Colibakterium aktiv beweglich sind, geht der Absorption der Phagen-DNS ein zufälliges Zusammentreffen von Phagus und Bakterium voraus. Trifft ein Phagus auf ein gegen es nicht resistentes Bakterium, heftet es sich an. Diese Anheftung ist eine Oberflächenreaktion und wird genauso bei leeren Bakterienhüllen und leeren Phagenhüllen beobachtet. Die dabei wirksame Verbindung ist ein Lipopolysaccharid der Bakterienmembran (W. F. GOEBEL und M. A. JESAITS 1952). Es stellt offenbar die Haftstelle für den Phagus dar. Die Rezeptoren sind für die verschiedenen Phagusarten typisch. Da nun solche Rezeptoren nicht nur an

Bakterienoberflächen, sondern auch etwa an der Oberfläche roter Blutkörperchen vorkommen, heften sich Phagen mitunter an solchen völlig ungeeigneten Gebilden an. Doch löst sich das Virus schon ziemlich bald aus einer solchen Falle, indem es die Rezeptorsubstanz einfach mit eigenen Mitteln zerstört. Die Haftstelle des Phagus ist an der Spitze des Schwanzes lokalisiert. Die dünnen Eiweißschichten werden an dieser Stelle bei der Anheftung offenbar abgewickelt und halten den Phagus am Bakterium fest. Ein dicker Stab, der im Schwanz des Phagus verborgen ist, stößt dann in das Innere des Bakteriums ein. Bei der Anheftung des T2 konnten aus der Bakterienmembran abgestoßene Bruchstücke beobachtet werden, die wahrscheinlich unter dem Einfluß eines Phagenenzyms losgelöst werden. Durch Experimente mit durch 35S markierten Eiweißmolekeln bzw. durch mit 32P markierter DNS konnte gezeigt werden, daß der Phagus seine gesamte DNS in das Bakterium ‹einspritzt›. Außerdem werden wie A. D. HERSHEY (1957) zeigen konnte, ein Polypeptid und zwei Polyamine zusammen mit der DNS in die Wirtszelle injiziert. Doch scheiden sie wohl als Träger genetischer Informationen aus. Nach der Infektion wird der Stoffwechsel der Bakterienzelle völlig umgestellt. Zwar laufen Eiweiß- und DNS-Synthesen weiter ab, doch werden nunmehr andere Stoffe gebildet als zuvor: Zahlreiche Enzyme werden nicht mehr nachgeliefert und statt dessen wird neuartiges Eiweiß synthetisiert, das zwar noch nicht die serologischen Eigenschaften des Phageneiweißes besitzt, aber an der Synthese des Phageneiweißes beteiligt ist. Schon nach drei Minuten p. i. ist dieser 110 Prozeß aufweisbar. Acht Minuten p. i. ist die typische Phagen-DNS nachweisbar. Bakterien-DNS wird nicht mehr synthetisiert, die vorhandene umgebaut. 10 Minuten p. i. lassen sich nach künstlichem Aufbrechen der Zelle die ersten fertigen Phagen beobachten. Zu diesem Zeitpunkt ist schon soviel Phagen-DNS gebildet worden, daß sie zur Bildung von 50 Phagen ausreicht. Diese DNS ist wahrscheinlich in einem ‹pool› zunächst gesammelt (E. KELLENBERGER, 1955). 25 Minuten p. i. ist der pool fast vollständig geleert. Neben dem typischen Hülleneiweiß der Phagen und der Phagen-DNS wird auch in reichlicher Menge das Enzym, das schon bei der Infektion die Zellmembran auflöste, hergestellt. Es hat jetzt die wichtige Aufgabe, die Zellmembran des Bakteriums aufzulösen und die neuen Phagen zu entlassen. Wird eine Bakterienzelle von Phagen infiziert, deren DNS durch Bestrahlung mit UV-Licht inaktiviert wurde, können bei Infektion der Zelle durch mehrere Phagen dennoch neue Phagen ausgebildet werden. Das läßt sich nur durch Zusammenwirken aller in das Bakterium abgegebenen Phagus-DNS erklären (S. E. LURIA und R. DULBECCO, 1949). Für ein solches Zusammenwirken der verschiedenen DNS spricht auch der Tatbestand, daß bei Infektionen durch zwei Phagen, die sich in zwei Merkmalpaaren voneinander unterscheiden, diese Merkmale bei den Tochterphagen ausgetauscht werden können. Bei der Beantwortung der Frage, ob Viren belebt seien, wird der Tatbestand erheblich, daß es Gruppen von Mikroorganismen gibt, die sich zwanglos zwischen die großen Viren und eigentlichen Zellorganismen einordnen lassen. Hierher gehören vor allem Rickettsien und Cysticeten. Rickettsien wurden schon 1901 von REED und CAROL entdeckt. Zu ihnen zählen die Erreger des 5-Tage-Fiebers und des Fleckfiebers (R. PROWAZEKI). Sie sind obligate Parasiten auf Läusedärmen oder auf Eihäuten lebender Embryonen. Ihre Größe entspricht der großen Viren (1200 X 250 nm). Strukturell ähneln sie Bakterien: sie haben eine eigentliche Zellmembran und vervollständigen sich während der Vermehrung zu einer Zelle. Die Vermehrung geschieht durch Verdoppelung (Zellteilung). Da es sich um kolloidale heterogene Gebilde von zellulärer Struktur handelt und sie mit den Bakterien u. a. die Art des Wachstums und der Vermehrung gemeinsam haben, handelt es sich nach allgemeiner Überzeugung um Eubionten. Als Cysticeten faßt man einen Komplex von Gebilden zusammen, die wie alle Eubionten, sowohl DNS als auch RNS führen, im übrigen aber recht verschieden sein können. Die bekanntesten sind der Seyffertsche Saprobiont und die Waidlawschen A- und B-Organismen. Der Saprobiont lebt frei im Kompost, hat eine Größe von 400 bis 600 nm, ist 111 bläschenförmig strukturiert, kennt ein Wachstum von 150 nm auf 600 nm und zerfällt nach Beendigung der Wachstumsphase in 50 bis l00 kleine Teile. Die A- und B-Organismen leben frei in Abwässern und unterscheiden sich - bei sonst ähnlichen Eigenschaften - von Saprobionten nur durch ihre geringere Größe (150-500 nm). Von einer eigentlichen zellulären Struktur kann bei beiden nicht die Rede sein, da eine Membran als Zellabschluß fehlt, die sonst bei allen Einzellern obligat ist. Auch die zelltypische Vermehrung durch Verdoppelung («Zweiteilung») fehlt. Dennoch dürfte es sich um Eobionten handeln. Vor allem die lebenstypische Art des Wachstums läßt die Entscheidung nicht allzu schwer werden. Wir haben hier einen Beleg dafür, daß die Vermehrung durch multiple Vervielfältigung, wie sie uns aus dem Bereich der Viren bekannt ist, auch bei Eobionten vorkommt. Daß obligater Parasitismus nicht unbedingt gegen eine Belebung spricht, zeigen uns die Rickettsien. Für eine endgültige Beurteilung des ‹belebt› bei Viren ist die Frage nach ihrer phylogenetischen Herkunft zu beantworten: leiten sie sich von vermehrungsfähigen Zellbestandteilen her, dann wird man die Frage nach dem «belebt» wohl verneinen müssen, es sei denn, man sieht in der Vermehrung nach dem Modell der NS-Matrizen das entscheidende Lebenskriterium und würde mit I. HAHN (1948) schon die Gene für belebt halten. Das scheint biologisch kaum gerechtfertigt zu sein, handelt es sich dabei doch - an sich betrachtet - um eine Art organischer Autokatalyse (W. TROLL). Stammen Viren aber von Lebewesen ab, die sich strukturell auf Grund eines totalen Zellparasitismus weitgehend

zurückgebildet haben, fällt die Antwort schon wesentlich schwerer. Man wird wohl nicht umhin können, sie auf Grund ihrer Phylogenese als lebend zu bezeichnen. Welche von den beiden Theorien ist nun die wahrscheinlichere? Gegen die erste Theorie spricht, daß spontan auftretende Virosen noch niemals einwandfrei beobachtet werden konnten. Scheinbar spontane Virosen (z. B. Ektrornelie der Maus oder Herpex simplex) konnten zumeist auf latente Infektitonen zurückgeführt werden (H. G. FRIEDRICH-FREKSA). Auch die Reindarstellung des hypothetischen Erregers etwa der Roux-Sarkome, die durch Teer ausgelöst werden, ist bislang noch nicht gelungen. Ganz das Gleiche gilt für zahlreiche andere als Krebserreger vermutete spontan entstehende Viren. Gegen diese Theorie spricht ferner, daß manche Viren in sehr verschiedenen Lebewesen vermehrungsfähig sind, wie etwa der Erreger des Gelbfiebers in Mücken und Warmblütlern oder das Aster-yellow-Virus in Astern und Heu- 112 schrecken. Dennoch scheint W. TROLL dieser Theorie den Vorzug zu geben. Wir halten mit H. G. FRIEDRICH-FREKSA die zweite Theorie, wenigstens für die DNS-Viren (Phagen u. a.) für wahrscheinlicher. Diese Viren scheinen phylogenetisch (in einer Weiterentwicklung» etwa über ein rickettsienähnliches Stadium) von obligat schmarotzenden Eubionten (oder deren phylogenetischen Vorstufen?) abzustammen. Doch bleibt grundsätzlich festzustellen, daß die Mannigfaltigkeit von Formen, wie sie im Virusbegriff zusammengefaßt werden, auf sehr verschiedene Weisen entstanden sein kann. 113 ZWEITER ABSCHNITT
NATURPHILOSOPHISCHE FRAGEN ZUR ENTSTEHUNG DES LEBENDIGEN
Wir haben schon weiter oben eine Reihe von philosophischen Vorentscheidungen genannt, die unsere Frage nach der Herkunft des Lebens trivial erscheinen lassen. Um nur die wesentlichsten zu nennen: - der Hylezoismus, nach dem die Materie aktuell belebt ist, - der Panpsychismus, nach dem die Materie aktuell beseelt ist, - der Materialismus, nach dem das Leben nichts als eine Bewegungsform der Materie ist. Und endlich neben diesen philosophischen Monismen eine von Theologie beeinflußte philosophische Theorie: - der extreme Kreatianismus, nach dem das Leben durch einen eigenen, zeitlich von dem der Materie unterschiedenen, Schöpfungsakt Gottes geschaffen wurde. Es lohnt sich kaum, auf die Widerlegung dieser Theorien allzu viel Geduld zu verwenden. Sie sind alle entweder wissenschaftstheoretisch äußerst fragwürdig oder stehen, wie der Materialismus, im Gegensatz zu dem bislang von uns Gesicherten. Gegen Panpsychismus und Hylezoismus ließe sich etwa argumentieren, daß die Fähigkeit des Lebewesens Lebensfunktionen zu zeigen, in der inneren Dynamik des Lebendigen als Lebendigem selbst beruht (ist doch Leben an Individualität gebunden, Individualität aber impliziert eine spezifische Dynamik auf die Bewahrung des Selbstandes und die Erreichung des Vollstandes hin). Wenn solche Funktionen grundsätzlich nicht gezeigt werden, besteht wissenschaftstheoretisch keinerlei Grund anzunehmen, daß sie gezeigt werden können, vor allem, wenn wir den Begriff des Lebewesens nicht abhängig machen von einem aktuell immer und stets Lebensfunktionen zeigen, sondern es als Einheit bestimmen, die wenigstens unter günstigen Bedingungen Lebensfunktionen zeigen kann. So ist etwa eine Bakterienspore durchaus belebt, obwohl sie u. U. jahrelang keine der von uns genannten biologischen Lebenskriterien zeigt. Sie kann sie aber - unter günstigen Bedingungen - sehr eindrucksvoll zeigen. Wir wollen also nicht schon physikalischen Einheiten die Möglichkeit, Lebensfunktionen zu zeigen, zusprechen, weil sie sie eben noch niemals zeigten, gleich unter welchen Bedingungen und Umständen sie beobachtet wurden. 114 Gegen den extremen Kreatianismus, wie er bis zum heutigen Tage noch in manchen Köpfen herumspukt, wäre auf die Unwissenschaftlichkeit einer Methode hinzuweisen, außerweltliche (etwa schöpferische) Eingriffe dann anzunehmen, wenn auch eine innenweltliche Deutung möglich ist. Die Aufgabe dieses theologischen Sparsamkeitsprinzips (vgl. WS I, 13) bedeutet die Aufgabe aller Weltwissenschaften. Sie führt mit einiger logischen und historischen Stringenz zum Okkasionalismus, wie er u.a. von den MUTAKALLIMUN, A.VICEBRON oder GEULINCX vertreten wurde, zur Leugnung letztlich jeder weltimmanenten Ursachlichkeit. Die Welt stellt dann nur mehr Gelegenheiten (occasiones) für göttliches Wirken zur Verfügung. Da sich nun aber, wie wir zeigen werden, durchaus Möglichkeiten, die Entstehung des Lebendigen auf weltimmanente Ursächlichkeiten zurückzuführen, anbieten, ist diese Deutung unbedingt vorzuziehen. Die Position des extremen Kreatianismus - angewandt auf die Herkunft des Lebendigen - ist eine, wenigstens im christlichen Raum, ziemlich späte Neuerwerbung. Bis hin ins 19. Jahrhundert wurde mit fast dogmatischem Eifer eine These vertreten, die eine Entstehung des Lebendigen aus dem Vermögen der Materie, ohne ein aktuelles schöpferisches Eingreifen Gottes, besagte. Als dann die Biologie die Möglichkeit der Urzeugung bezweifelte, zog, mit gewohnter Verzögerung, die allzu naturwissenschaftsgläubig gewordene Theologie nach. Seit dem ausgehenden 19. Jahrhundert wurde mit einem ebenso quasidogmatischen Eifer die Unmöglichkeit der Urzeugung vertreten. Eine recht gelegen kommende Anschauung, da sich aus dieser Unmöglichkeit leicht so etwas wie ein Gottesbeweis konstruieren

läßt, der auch von schlichten Gemütern eingesehen werden kann. Ein trauriges Wechselspiel von apologetischem Nutzen und dogmatischem Begründen begann, Sicher erlaubt eine wissenschaftlich begründete Meditation über das Phänomen Leben ein sicheres Erkennen Gottes. Das aber hat mit einem Beweis nichts, mit einem Aufweis nur wenig zu tun. Es dürfte immerhin nützlich sein, unseren philosophisch-systematischen Überlegungen um die Entstehung des Lebendigen einige historische Informationen voranzustellen, nicht um zu zeigen, daß die von uns vertretene Ansicht die der Kirche in ihren ersten 1800 Jahren war, sondern, weil auch eine moderne Position durch eine Denktradition getragen wird und nur aus ihr heraus voll verständlich werden kann. 115 ERSTES KAPITEL
DIE MEINUNG DER «KIRCHENVÄTER»
Wenn wir hier das Wort Kirchenväter benutzen, dann nicht im eingeschränkten Sinne der modernen Theologie. Wir meinen damit die Kirchenschriftsteller der ersten christlichen Jahrhunderte. Hier wollen wir von der theologischen Bedeutung der Aussagen der Kirchenväter einmal absehen, sondern sie als Philosophen und Theologen vorstellen, die sich darüber Gedanken gemacht haben, wie der biblische Schöpfungsbericht zu interpretieren sei. Zunächst einmal sei dieser Text vorgestellt, den wir ebenso nicht in seiner theologischen sondern historischen Relevanz begreifen wollen: «Dann sprach Gott: ‹Die Erde mache junges Grün sprossen, samen-tragende Pflanzen und Bäume, die je nach ihrer Art Früchte mit Samen darin auf der Erde tragen›. Und es ward so: die Eri-le veranlaßte, daß junges Grün hervorging... Dann sprach Gott: ‹Die Erde mache lebende Wesen nach ihren Arten, Vieh, Kriechtiere und wilde Landtiere, je nach ihren Arten›. Und so ward es» (Gen 1. 11 f und 24). Einer meiner Studenten (W. HÜBINGER) hat sich ausführlich in einer recht guten Arbeit mit der frühchristlichen Interpretation des Textes über das Werden vor allem der Pflanzen beschäftigt (Die Lehre von der Erschaffung des Lebens bei den frühen Kirchenvätern im Anschluß an Gen 1. 11 f). Zunächst sei ein Einwand ausgeräumt: Die jüdische Priestertradition (ihr entstammt unser Text) hat vermutlich Pflanzen nicht als belebte Wesen betrachtet. Dieser Einwand würde sicher erheblich sein, wenn der Hagiograph nicht für die Erschaffung der Tiere ganz die gleiche Formulierung verwandt hätte. Viele der von uns angeführten Kirchenväter sind sicherlich beeinflußt von der Religionsphilosophie PHILOS des Juden, der um 25 v. Chr. in Alexandrien als Sohn einer jüdischen Priesterfarnilie geboren wurde. Wir wollen also zunächst einmal seine Lehre von der Schöpfung des Lebendigen vorstellen:
a) Philo der Jude In seinem Buch «De opificio mundi» interpretiert er den Genesisbericht: Die geschaffene Welt existiert auf Grund eines (numerisch eines) Schöp- 116 fungsaktes. Im Hintergrund dieser und ähnlicher Aussagen mag auch eine Aussage des Buches Jesus Sirach (18.i) stehen, die man schon recht früh - falsch interpretierend - so las: «Gott hat alles auf einmal geschaffen». Die sechs Tage bedeuten nach PHILO einzig eine Ordnung der göttlichen Schöpfungsideen. PHILO spricht von der Schwangerschaft der Erde und vergleicht die Entstehung des Lebens mit der Niederkunft einer Mutter. Doch ist letztlich der Grund des Entstehens des Lebens ein göttlicher Befehl, der sich die Erde so einrichten ließ, daß sie in rechtem Zeitpunkt aus sich heraus und aus ihrem Vermögen das Leben hervorbrachte. Damit begann eine Art ewigen Kreislaufs: Das Leben ist ewig. Es ging hervor aus dem Vermögen der Erde, sinkt wieder in es hinein und entsteht aus ihr, ewig neu. So liegt Leben jenseits der Grenzen erfahrbarer Kontingenz. Leben und Ewigkeit werden einander ganz eng zugeordnet (a. a. O. n. 13 §§ 42 ff).
b) Hippolyt von Rom († 266) HIPPOLYT ist einer der frühesten Kirchenschriftsteller. Da er Gegenpapst zu Calixtus war, galt er als Schismatiker, und so gerieten seine Schriften bald in Vergessenheit. Es sind uns von seinem Genesiskommentar nur Bruchstücke erhalten. Eines dieser Fragmente lautet: «Am ersten Tage nämlich schuf Gott alles, was er schuf. An den folgenden Tagen gestaltete (immutavit) er das, was er am ersten Tage geschaffen hatte» (PG X, 586). Das Leben in allen seinen Ausdrucksformen wurde also in der Vorstellungswelt HIPPOLYTS nur durch ein ordnendes Gestalten der Materie hervorgebracht.
c) Origines (185-254) Auch ORIGINES lehrte die simultane Schöpfung. Danach gibt es kein eigentliches Neuentstehen mehr. In den Homilien zur Genesis verwendet er den Begriff ‹Befehl› (iussus) als Gegenbegriff zu ‹Hören› (auditus). Außer den Himmelskörpern und dem Menschen wurde alles andere durch die «auf den Befehl Gottes horchende Erde hervorgebracht» (PG XU, 149-155). In seiner Schrift gegen CELSUS schreibt er: «Ist nämlich die Welt geschaffen worden ... , so verdanken die ersten Menschen nicht notwendig ihr Dasein der Verbindung von Mann und Frau,

sondern zeugenden Kräften (logoi spermntikoi), die sich der Erde verbanden» (PG XI, 731). Vermutlich werden ganz ähnliche Worte auch in den Predigten über das Buch Genesis gestanden haben, sie wurden dann aber in der Überarbeitung durch RUFINUS entsprechend korrigiert, da vermutlich schon damals diese Lehre von der Erschaffung des Menschen nicht als rechtgläubig akzeptiert wurde. Ausdrücklich zurück- 117 gewiesen wurde sie aber erst durch Pius XII, in seiner Enzyclica Humani generis (1950) (D 3896), sieht man einmal von der Verurteilung durch den Pariser Bischof Etienne TEMPIËR (1277) (vgl. MAN-DONNET, Siger de Brabant II, Louvain 1908, 182) ab. Die Lehre von den logoi spermatikoi wurde von ZENON von Kition in die Philosophie eingebracht. Sie spielt bei Augustinus und den mittelalterlichen Philosophen (rationes séminales) eine bedeutende Rolle. .
d) Basilius der Große (330-379) «Jenes gesprochene Wort (Gen 1.11) und jener erste Befehl erging wie ein Gesetz an die Natur und haftete der Erde an und übertrug ihr die Kraft zu zeugen und Frucht zu bringen in Zukunft... Die Erde bringt durch sich selbst hervor... und benötigt keine Hilfe von andersher» (PG XXIX, 95) «Dieser Befehl haftet an der Erde und sie hört nicht auf, dem Schöpfer zu dienen . .. Manche Lebewesen erhalten, wie ersichtlich auch jetzt noch ihr Leben von der Erde selbst» (ibd. 190). Dieser Text ist so klar, daß es kein kleinliches Deuteln gibt. Ob allerdings eine Simultanschöpfung damit ausgesagt wird, mag zweifelhaft sein. Unzweifelhaft ist dagegen, daß BASILIUS, einer der bedeutendsten Theologen des christlichen Altertums über den Verdacht der Heräsie erhaben ist, was man keineswegs von allen Theologen und Philosophen der christlichen Frühzeit sagen kann, denn Denken war schon damals eine nicht ganz ungefährliche Angelegenheit. ...
e) Jobannes Chrysostomus (344-407) vermeidet, soweit als möglich, wenn er von der Entstehung der nicht-menschlichen Geschöpfe schreibt, den Ausdruck geschaffen. Er benutzt bevorzugt das Wort grundgelegt (PG LUI, 71). Von ihm spätestens stammt die fatale Formulierung, daß Sinn ihres Daseins sei, für den Menschen (propter hominem) zu sein; fatal, es sei denn, man versteht diese Aussage in dem Sinne, daß sich «unser geistiges Sein ... dauernd aus den unzähligen Kräften der faßbaren Welt ernährt» (Teilhard de Chardin, Der Göttliche Bereich, Ölten und Freiburg 31963, 40). Ähnliche Interpretationen unseres Genesistextes finden sich bei THEODORET (393-466), für den die Ursachen des Entstehen des Lebendigen nicht die unbeseelten Elemente (elementa inanimata), sondern unsichtbare Kräfte (invisibiles virtutes) sind, die auf Gottes Geheiß hin wirksam werden (PC LXXX, 90). PROKOP von Gaza (460-530) läßt die Pflanzenwelt nach einem Gesetz, das Gott zuvor (praecedente impera- 118 trice voce divina) der Erde mitgegeben hat, entstanden sein (PG lxxxvii a, 79 ff). Schauen wir uns jetzt einmal bei den lateinischen Kirchenvätern um:
f) Tertullian (160 - nach 220) TKRTULLIAN sagte sich wegen zu laxer Praktiken seiner Glaubensgenossen von der romischen Kirche los (205), doch sind uns mehr als 3o seiner Schriften erhalten. Auch er läßt das Leben aus dem Vermögen der Erde hervorgehen. Licht und trockenes Land sind die Bedingungen seines Hervorgehens. Zuvor aber war dieses Vermögen der Materie eingeschaffen worden (vgl. PL II, 1097, doch ist der Verfasser dieser Schrift ungewiß). Auch VICTORINUS MARTYR (f 304), der als erster Exeget der lateinischen Kirche gilt, lehrte in seinem Buch «De Fabrica mundi», daß das Leben aus den zeugenden Kräften von Erde und Wasser hervorging. Die gesamte lebende Natur — mit Ausnahme des Menschen - erhielt ihr Leben aus dem Vermögen der materiellen Natur (PL V, 305 ff).
g) Ambrosius (339-397) Nach AMBROSIUS ist das Wort Gottes, das gleichsam als Befehl den Entstehungsprozeß der belebten Natur auslöste, ein Naturgesetz, dem die materielle Welt bis an das Ende aller Zeiten gehorcht. Die Erde selbst soll lebende Wesen hervorbringen, ohne daß sie dabei irgendeiner Hilfe von außen bedürfe. So leistet denn die Erde, deren Ziel es ist, vollkommen und ganz gut zu sein, ihren Beitrag zum Willen des Schöpfers: sie gibt aus eigenem Vermögen Leben (PL XIV, 166).
h) Augustinus (354-430) Alle Wesensformen — AUGUSTINUS steht in manchen seiner Ansichten dem Hylemorphismus nahe - sind zusammen mit der Materie von Gott aus dem Nichts geschaffen worden, und zwar zum größten Teil nicht aktuell, sondern als «Keimkräfte» (rationes séminales) der Materie eingegeben worden, die sie im rechten Augenblick zur Entfaltung bringt. Auch alles Belebte wurde durch rationes séminales der Materie eingeschaffen und gelangt unter dem Einfluß äußerer Ursachen zur Entfaltung: «Die Arten der Lebewesen des Meeres und der Erde waren ... in der Erde.. . ihrem

Vermögen nach (potentialiter) angelegt... So bringt denn die Erde auf das Wort Gottes hin hervor, was sie, ehe diese hervorgingen, schon in all ihrer Vielzahl empfangen hatte» (PL XXXIV, 326). Damit wollen wir die Aufzählung der Lehren früher Kirchenväter abschließen. Die Aufzählung derer, die eine Erschaffung des Lebendigen seinem aktiven Vermögen nach, das der Materie eingegeben wurde 119 (creatio virtualis vitae) lehrten, ließe sich fortsetzen. Man kann sagen, daß offensichtlich die ersten fünf christlichen Jahrhunderte es für eine geoffenbarte Wahrheit hielten, daß Gott das Leben nicht durch einen eigenen Schöpfungseingriff geschaffen habe, sondern zugleich mit der Materie, in dem er der Materie rationes séminales, virtutes ad vitam ... einschuf. 120 ZWEITES KAPITEL
DIE SCHOLASTISCHE PHILOSOPHIE
Die Lehre der mittelalterlichen Scholastik nährt sich, was ihre Aussagen über den Ursprung des Lebens betrifft, aus zwei recht verschiedenen Quellen: AUGUSTiNus und ARISTOTELES. Die Ansicht des Augustinus haben wir schon kurz skizziert, die des Aristoteles erwähnt. Die Erwähnung bedarf einer Ergänzung.
a) Aristoteles (384-322) Die klassische Definition der Materie (materia prima) des ARISTOTELES in seiner Metaphysik lautet: «Unter Materie verstehe ich dasjenige, was durch sieh selbst weder Etwas (τι) ist, weder eine Quantität (πόσον); noch irgendetwas anderes, wodurch das Seiende bestimmt wird, denn es muß etwas geben, von dem alle diese ausgesagt werden können, und dessen Sein, von dem dieser Aussagen verschieden ist ... Die Materie ist also das Letzte an sich, das nichts, keine Quantität und auch nichts anderes ist, ja nicht einmal deren Negationen, denn auch diese sind auf ihre Weise ein Akzidens» (Mph. IV, 3 n. 4; 1029 a 20 ff). Von diesem, für unsere Überlegungen reichlich unergiebigen, Materiebegriff ist der der aristotelischen Physik abzuheben: «Wir unterscheiden Materie und Gestaltlosigkeit (στέρεσις)· Die Materie ist per accidens Nichtseiendes, die Gestaltlosigkeit aber ist Nichtseiendes per se. Die Materie ist angenähert schon Substanz (ουσία), die Gestaltlosigkeit aber in keiner Weise» (Phys. I, 9 n. r; 192 a). «Materie nenne ich das erste, das einem jedem Subjekt (ύποκείµενον) zugrunde liegt, in dem etwas nicht nur per accidens eingelagert ist, oder das, in das hinein etwas verschwindet, wenn es zugrunde geht» (ibd. n. 4; 192 a 30-33). So kann er dem, wie schon berichtet, sagen: «Es entstehen in der Erde und im Wasser Tiere und Pflanzen, weil in der Erde Feuchtigkeit, im Wasser Lebensluft und überall seelische Wärme ist. So ist in gewissem Sinn alles voller Seele ...» (de generat. anim. III, 11; 62 a). Der Hintergrund dieser modifizierten Theorie von Materie in der Physik ist der für ARISTOTELES gesicherte Tatbestand der Urzeugung (generatio aequivoca): «Allgemein gilt von den Tieren, was auch von den Pflanzen zu sagen ist: die einen entstehen aus dem Samen anderer Pflanzen, die anderen von selbst aus einem geeigneten Stoff... So entstehen auch 121 die Tiere teils von Tieren, teils von selbst und nicht von Artverwandten» (de nat. anim. V, 1; 5393). Doch auch die Gestirne gelten ihm als beseeelte Wesen: «Wir reden aber über die Sterne immer nur als von Körpern ... ganz ohne Seele. Man muß sie sich jedoch als beteiligt denken am Leben und Handeln ... Dieses Handeln hat man sich vorzustellen wie bei belebten Pflanzen und Tieren» (de coelo II, 12; 292). Wir haben über diese Ansicht schon referiert (vgl. WS I, 12). Obwohl sie von Bischof Templer 1277 (ah 92. der verurteilten Sätze) verworfen wurde, konnte sie sich bis ins 17. Jahrhundert hinein fast allgemein halten. Auch die Lehre von der permanenten Urzeugung war allgemeines Gedankengut der abendländischen Philosophie bis hinein ins 19. Jahrhundert, erst dann wurde sie von den Naturwissenschaften so gründlich in Frage gestellt, daß man, über das Ziel hinausschießend, jede Form der Urzeugung abzulehnen müssen glaubte.
b) Albertus Magnus (1193-1280) Die lateinischen Aristoteliker standen vor der kaum zu meisternden Schwierigkeit, die beiden Materiedefinitionen des Aristoteles in Einklang zu bringen. Sie bemerkten meist nicht, wie manche Aristotelesinterpreten selbst unserer Tage, daß ARISTOTELES von zwei ganz verschiedenen Dingen spricht: Sein Materiebegriff ist äquivok! Diese Schwierigkeit wuchs ins Unlösbare, wenn sie nach der Art des Enthaltenseins der Formen (der des Lebendigen vor allem) in der Materie fragten. Von diesem Dilemma zeugte schon die Lehre ALBERTS: «Wir lehren nach der vollendeten Doktrin der Peripathetiker, daß die Form als konfuse Wesenheit (per essentiam) in der Materie anwesend ist... Man kann nicht sagen, daß ein Erzeugendes (agens) das Wesen der Form erzeugt, weil diese schon besteht, es handelt vielmehr nur in Bezug auf das Sein der Form» (In Phys. I tr. 3 c. 15). «Jede Form aber, die in der Materie ist, war zuvor im Erstbeweger (in Gott)» (ibd. c. 17). Mit den meisten der Aristotelesschüler unterscheidet ALBERT zwischen Wesen und Sein eines Seienden. Beide sind nach scholastischer, von ARISTOTELES inaugurierter Lehre (vgl. Anal. Post. II, 7 n. 2; 92 b), das Seiende

konstituierende Prinzipien. Sein und Wesen - oft metaphysisch real unterschieden und unterscheidbar gedacht - spielen auch in der Lehre Alberts eine hervorragende Rolle. Das Wesen der metaphysischen Form (eines Lebewesens etwa) ist als konfuses von Gott in die Materie eingesenkt, das Sein der Form wird von einer handelnden Ursache hervorgebracht: Genauerhin müßte man wohl sagen mithervorgebracht, da die Ursache nicht primär eine metaphysische Form, sondern 122 ein Seiendes, das nach hylemorphistischer Lehre seinerseits wieder metaphysisch aus Materie und realer Form zusammengesetzt ist, erzeugt. Die Form erhält ihr Sein also mit dem Seienden. Die scholastische Tradition interpretierte die Sätze ALBERTS oft so, als ob er ein aktuelles Enthaltensein der Formen in der Materie angenommen habe. Das ist nach dem Ausgeführten offensichtlich falsch. Eine Form, die nur ihrem Wesen (nicht aber ihrem Sein) nach in die Materie vorgegeben ist, kann allenfalls virtuell, d. h. ihrem aktiven Vermögen nach (vgl. dazu In Phys. II tr. 2 c. 6) in der Materie angelegt sein.
c) Thomas von Aquin (1225-1274) Die Lehre des großen Aquinaten gibt sich schon differenzierter: «Die Formen praexistieren potentiell in der Materie ... Sie entstehen nicht neu, sondern sind (mit der Materie) mit geschaffene» (Summa Theol. I q. 45 a. 8c). Seiende, deren Form substantiell ist (dazu gehören die Lebewesen, deren Form ja von den Scholastikern meist als substantielle Lebensform verstanden wurde), bedürfen zur Aktualisation dieser Form der Kraft natürlicher Prinzipien» (ibd. III q. 753. 6 ad 1), sie können also nicht aus der Dynamik der Form selbst hervorgebracht werden. Was aber sind solche natürlichen Prinzipien? «Zum Hervorbringen unvollkommener Tiere genügt ein allgemeines agens, nämlich die Kräfte der Gestirne ... Doch kann man nicht sagen, daß ihre Formen durch ein von ihnen unterschiedenes agens geschaffen würden. Zum Hervorbringen vollkommener Tiere aber genügt kein allgemeines agens, sondern es ist ein eigenes erforderlich, das univoce hervorbringt» (ibd. I. q. 45, a. 8 ad 3). Nach der Lehre des THOMAS sind also die Formen des Lebendigen potentiell in der Materie enthalten und so miterschaffen. Handelt es sich dabei um reine Dispositionen (potentiae passivae) oder um eigentliche Vermögen der Materie (potentiae activae; virtutes?). Die Feststellung, daß zu ihrer Realisation eine Ursache, seien es die Sternkräfte (bei niederen Tieren) oder die Eltern (bei höheren Tieren), notwendig ist, kann diese Frage kaum beantworten. Eine Entscheidung mag die Lehre Thomas' von den «rationes séminales» erleichtern: Diese rationes séminales können in verschiedenen Ordnungen beobachtet werden: zunächst einmal sind sie ... hauptsachlich und ursprünglich im Wort Gottes als ideale Inhalte (rationes ideales), dann aber sind sie in den Elementen der Welt, in die hinein sie zugleich mit ihr geschaffen wurden, wie in allgemeine Ursachen, drittens aber sind sie in dem, das im Laufe der Zeit aus den allgemeinen Ursachen hervorging, wie in dieser Pflanze und in diesem Tier und endlich sind 123 sie im Samen, der von Pflanze und Tier erzeugt wird» (ibd. q. 115 a 2 c). Nachdrücklich betont er, daß die rationes séminales sowohl aktive als auch passive Prinzipien der ‹Zeugung› seien. Wann aber sind sie aktive, wann passive Prinzipien? Sicher sind sie in diesem Lebewesen und in seinem Samen aktive Vermögen. Wurden sie aber in die Elemente dieser Welt nicht doch nur als Dispositionen, als reine Anlagen eingeschaffen? THOMAS beruft sich bei der Darstellung seiner Lehre von den rationes séminales auf AUGUSTINUS. Wenn diese Berufung nicht nur der Abstützung der eigenen These durch eine anerkannte Autorität dienen soll, wird man wohl zurecht annehmen dürfen, daß er sie wie Augustinus als eigentliche Vermögen verstand. Einen klareren Text fanden wir im Kommentar zu den Büchern des Aristoteles zur Metaphysik: Die Lebewesen «... entstehen aus der Materie, die ein Prinzip enthält, das dem aktiven Vermögen (virtus activa) des Samens ähnlich ist, aus dem das Lebensprinzip (anima) solcher Tiere hervorgeht. Und wie das Vermögen (virtus), das im Samen ist, von der Vollseele (anima compléta) des Tieres und von dem Vermögen eines Himmelskörpers herrührt, so kommt das Vermögen, das in der faulenden ein Tier zeugenden Materie ist, allein vom Himmelskörper, in dem als Vermögen alle erzeugten Formen wie in einem aktiven Prinzip enthalten sind» (a.a.O. VII n. 1457). THOMAS nimmt also zwar nicht in der Materie als solcher, sondern in «faulender Materie» ein aktives Vermögen, Lebewesen hervorzubringen, an, doch ist dieses nicht ursprünglich, sondern stammt von den belebten Himmelskörpern. Diese Lehre ist sicher recht unbefriedigend. Dennoch ist sie die bestdurchdachte der ganzen mittelalterlichen Scholastik. Will man einerseits an einem virtuellen Enthaltensein der Lebensformen (Individualprinzipien) in der Materie und andererseits an der klassischen Substanzontologie festhalten, wird man mit einiger Notwendigkeit die Aktualisation dieser Formen (d. h. doch ihre Substanzwerdung) auf ein anderes, schon zuvor substantiell Seiendes zurückführen müssen, das, da eine Wirkung niemals seinsmächtiger sein kann als ihre Ursache, schon belebt ist. Die These von der Ursächlichkeit belebter Sternkörper ist also bei Thomas keine astrologische Spielerei, sondern eine bittere Notwendigkeit, in die hinein ihn seine Substanzontologie zwang. Wir werden also entweder die Theorie von dem virtuellen Enthaltensein der Lebensformen oder die Substanzontologie (zugunsten einer Strukturontologie etwa) aufgeben müssen, wenn wir uns von den merkwürdigen Sternseelen trennen wollen. Da scheint uns das geringste Opfer die Aufgabe der Substanzontologie als einer das kategorial-Seiende durchherrschenden Ontologie zu sein. 124

Der recht eigenwillige Schüler des Thomas, AEGIDIUS ROMANUS (1245-1316), versucht in seinem Sentenzenkommentar die Ansicht seines Meisters zu modifizieren: «Gott legte die rationes séminales in die Materie, damit sie dauernd Dinge hervorbringe ... solange es Gott beliebt. Deshalb aber dürfen die rationes séminales keine bestimmte l Unordnung auf eine bestimmte Form haben, weil sie, nachdem die bestimmte Form durch die rationes séminales aus der Materie hervorgebracht wurde, und diese Form zugrunde gegangen ist, sonst keine weitere ähnliche hervorbringen könnten. Daher sind die rationes séminales nur in gewisser Allgemeinheit auf die Formen hingeordnet» (In Sent. II p. 2 d. 18 q. 2 a. 1). Über den philosophischen Wert dieses Textes läßt sich streiten. Über die Art des Enthaltenseins der Formen in der Materie sagt er nichts aus. Aber das Ursachenproblem wird vielleicht etwas glücklicher gelöst: die rationes séminales, gering an Quantität, reich aber an aktivem Vermögen, sind selbst weder Form, noch aktives Vermögen noch passiv in Dingen, sondern eigentlich Seiende (habitudines),die verursachend die Form aus der Materie hervorbringen, selbst aber nicht in das Lebewesen einzugehen scheinen.
d) Johannes Duns Scotus (1270-1308) Ausdrücklich betont auch DUNS SCOTUS, daß die rationes séminales ein actu Seiendes sind und als solche eigentlicher Grund dafür, daß etwas entsteht (In Sent. II d. 18 q. unica). Aber auch er erklärt sich nicht über die Art des Enthaltenseins der Formen in der Materie. Einer seiner Schüler ist da entschiedener: PETRUS AUREOLI meint, daß die Formen teilweise (unvollkommen) in der Materie enthalten seien und erst durch die Wirksamkeit eines agens vervollkommnet werden (In Sent. II. d. 18, q. 1 a. 2). Er dürfte sich wohl von allen Scholastikern des [3. und 14. Jahrhunderts am klarsten für ein aktuelles Enthaltensein der Formen in der Materie ausgesprochen haben. Wir wollen die Jahrhunderte des nun folgenden Schulgezänks übergehen und einen Philosophen und Theologen des 16. Jahrhunderts vorstellen, der, zum Teil unter dem Einfluß der Renaissance, neue Ideen entwickelte:
e) Dominicas Soto (1494-1560) «Die Form ist in der Potentialität der Materie enthalten, was nichts anderes besagt, als daß sie im Werden wie im Sein von der Materie als ihrer rezeptiven und passiven Ursache abhängt . . . wie stets, wenn ein Seiendes in einem anderen ist, ist es in ihm wie in einer Ursache ... So war die Welt in Gottes Schöpfungsmacht (virtus productiva),. ehe sie 125 wurde... So ist fast wie Früchte im Samen ... etwas in der Potenz der Materie, doch nicht auf die Weise einer Wirk-, sondern einer Materialursache ... Weil nun aber alle Formen in solcher Abhängigkeit von der Materie entstehen, sagt man sehr richtig, sie seien in der Potenz der Materie und werden aus ihr hervorgebracht. Jede Form ... kommt aus der Potentialität der Materie (edncitur ex potentialitate materiae)» (Summa theol. I q. 76 a. 4 c). Das gilt nach SOTO für alle Formen, außer für die menschliche Seele. «Aus der Potentialität der Materie herausgeführt werden heißt actu das werden, was zuvor in der Potenz der Materie war» (In Phys. I, 4, text. 78). Nach den Ausstellungen SOTOS scheint, da die Materie als passive Potenz gedacht wird, die Form nur ihrer passiven Potenz (gleichsam als Disposition der Materie) nach in der Materie enthalten zu sein. Dagegen betont er jedoch nachdrücklich: «daß die Form vorher potentiell in der Materie war und zwar in gewisser Hinsicht ihrem aktiven Vermögen nach, zwar nicht so, daß die Materie aktiv wirkt, doch so, insofern sie der natürliche Träger ist, durch dessen Wandlung das Seiende hervorgebracht wird» (ibd.). Die Frage nach dem aktiven Vermögen der Form wird also nicht behandelt, doch wird der Materie eine recht schwache und recht uneigentliche Aktivität zugesprochen, insofern sie zwar nicht zur Form strebe «wie Ähnliches zu Ähnlichem, sondern wie Unvollkommenes zu aktueller Vollkommenheit» (ibd.). Historisch ist die Lehre SOTOS von einiger Bedeutung, weil er eine Formulierung verwandte, die später zu einer Art von scholastischem Grundsatz wurde, wenn es galt, die Entstehung und das Vergehen von Lebewesen zu erklären: Die Form wird aus der Potentialität herausgeführt und in die Potentialität der Materie zurückgeführt. Wie wenig diese Theorie tatsächlich leistet, mögen die Ausführungen Sotos zeigen. 126 DRITTES KAPITEL
STIMMEN DER NEUZEIT ZUR ENTSTEHUNG DES LEBENDIGEN
a) PARAZELSUS (1493-1541) schreibt: «Nicht nur das, was sich bewegt ist lebend, wie Menschen, Tiere, Würmer der Erde . . ., sondern ebenso alle körperlichen Dinge» (de nat. rerum IV; ed. Aschner III, 924). Bei DESCARTES lesen wir: «... zum Verständnis der Natur der Pflanzen und Menschen ist es weit besser (als eine aktuelle Schöpfung aller Arten), ihre allmähliche Entstehung ans dem Samen zu beobachten so, wie Gott sie zu Beginn der Welt geschaffen hat» (Princ. III, 45). b) G. W. LEIBNIZ (1648-1716) meint: «Ich nehme an, daß die Lebensprinzipien in der ganzen Natur verbreitet und daß sie unsterblich sind, du sie ja unteilbare Substanzen oder Einheiten sind, während Körper Vielheiten sind ... Es gibt in meinem System keinen Teil der Materie, in dem nicht eine unendliche Anzahl von organischen und beseelten Körpern ... enthalten sind. Man darf jedoch nicht sagen, daß jeder Teil der Materie beseelt ist, so wenig wie man einen Teich voller Fische darum einen beseelten Körper nennt, weil jeder Fisch beseelt ist» (Betr. ü. d. Lebensprinzipien). Die Reihe der Philosophen und Biologen, die die Existenz einer Urzeugung (sei es einer permanenten oder einer

einmaligen) annahmen oder noch annehmen, ließe sieh fast beliebig verlängern. Noch um die Mitte des 19. Jahrhunderts behaupteten namhafte Biologen wie H. G. RRONN, K. E. VON BAER, K. VON NÄGELI eine noch sieh unserer Zeit abspielende Urzeugung (vgl. W. Zimmermann, Evolution, Freiburg und München 1953, 503). Daran änderten grundsätzlich auch nichts jene klassischen Versuche, das Gegenteil zu beweisen. Der Florentiner FR. REDI (1628-1698) bedeckte Fleischbehälter mit Gaze und stellte sie in die Sonne. Fliegen kamen und legten ihre Eier auf dem Gewebe ab. Im Fleisch selbst entstanden keine Maden. REDIS Untersuchungen führten W. HARVEY (1578-1657) zu einer Aussage, die bis zum heutigen Tage noch von manchen Biologen wie ein Dogma ge-handhabt wird: Omne vivum ex ovo (alles Leben kommt aus der Eizelle). Doch kam schon HARVEY selbst dieser Satz so phantastisch vor, daß er bis zu seinem Tod an der Möglichkeit einer Urzeugung festhielt. Als dann A. VAN LEEUWUNHOEK (1632-1723) die ersten Mikroskope baute, entdeckte man eine neue Welt, die Welt der Infusorien. Und 127 diese Kleinstlebewesen schienen aus Luft, Wasser oder Fleischbrühe zu entstehen. Manche Theologen jubelten, lasen sie doch in der HL Schrift einen Bericht, nach dem Bienen aus dem Leichnam eines Löwen entstanden (Richt 13.8) und sahen darin die Möglichkeit der Urzeugung als geoffenbarte Wahrheit ausgesagt. Doch ihr Jubel kam etwas zu früh. L. PASTEUR (1822-1895) füllte Glaskolben mit Fleischbrühe, bestieg den Gletscher des Montblanc, ließ die hier praktisch keimfreie Luft in den Kolben einströmen und schmolz ihn zu. In dem Kolben entstanden keine Lebewesen. Dieser Erfolg ließ seine Gegner nicht ruhen. JOLLY wiederholte den Versuch mit einem Heuaufguß und der Luft des Pic du Midi in den Pyrenäen. Und in seinem Kolben wimmelte es bald von Lebewesen. Heute wissen wir, daß in Heuaufgüssen Infusorien leben, die Temperaturen von 100° überstehen. JOLLY und seine Zeit hatte davon keine Ahnung und so blieb die Urzeugungstheorie in der Diskussion, obwohl R. VIRCHOW das Harveysche Axiom weiter präzisierte: omnis vita ex cellula (Alles Leben aus einer Zelle). Heute wissen wir, daß Urzeugung ein sehr viel komplizierterer Vorgang ist, als man es sich von den Anfängen der abendländischen Philosophie bis hin zur Mitte des 2o. Jahrhunderts vorstellte. Komplizierte biochemische Prozesse, die nachzuahmen uns heute noch nicht gelungen ist, sind zu leisten. Wir glauben nicht, daß die Natur sie mehrmals zu leisten in der Lage war. Sollte aber noch einmal ein Lebewesen aus unbelebter Materie neu entstehen, müßte das in absolut steriler Umgebung geschehen, sind doch schon die chemischen Vorformen eines solchen Gebildes beliebte Nahrung für zahlreiche Kleinlebewesen. Eine solch sterile Umgebung ist außer in unseren Laboratorien nur noch auf anderen Sternen zu finden; auf der Erde jedenfalls seit einigen Milliarden Jahren nicht mehr. 128 VIERTES KAPITEL
DIE ENTSTEHUNG EINES LEBEWESENS DURCH FORTPLANZEN (GENERATIO UNIVOCA)
Unter Fortpflanzung wollen wir hier ein biologisches Geschehen verstehen, bei dem ein neues Lebewesen unter Beteiligung eines oder mehrerer Lebewesen ähnlicher Struktur und Funktion entsteht. Die Fortpflanzung geschieht durch Absonderung von Teilkomponenten jugendlichen Charakters vom Elternorganismus. Meist sind solche Teilkomponenten ziemlich klein (Sporen, Knospen, Geschlechtszellen). Die zur Fortpflanzung dienenden Zellen sind Glieder von Ketten dauernd teilungsfähiger totipotenter Zellen. Eine solche Kette nennt man Keimbahn, die einen kontinuierlichen ‹unsterblichen› Lebensfaden vorstellt, an dem die einzelnen Lebewesen mit ihren differenzierten (und damit sterblichen) Geweben hintereinander aufgereiht sind. In vielen Fallen ist die Fortpflanzung ungeschlechtlich (Agamogonie). So vermehren sich Einzeller meist durch einfache Zellverdopplung. Mitunter findet erst eine mehrfache Kernverdoppelung und dann ein gleichzeitiger Zerfall in mehrere Zellen statt (Foraminiferen, Sporozoen). Manche vielzelligen Tiere (aber auch einige Einzeller, wie die Suctorien) vermehren sich ungeschlechtlich durch Knospung, wobei es mitunter zur Stockbildung kommt (Schwämme, Korallen, Moostierchen). Doch schon recht früh in der Stammesgeschichte hat sich die geschlechtliche Fortpflanzung entwickelt, die schon in Vorstufen bei Bakterien, dann aber bei allen Klassen der Protozoen auftritt. Charakteristisch für die geschlechtliche Fortpflanzung ist die Befruchtung, d.h. die Verschmelzung von zwei Geschlechtszellen, die meist von zwei verschiedenen Lebewesen geliefert werden. So wie bei der Knospung totipotente Körperzellen zu einer Entwicklung eines neuen Lebewesens führen, vermögen auch manche totipotente Keimzellen ohne Befruchtung zur Entwicklung zu kommen. Parthenogenese ist üblich bei Saugwürmern, Fadenwürmern, Rädertierchen, Wasserflöhen, Schildläusen, manchen Käfern und Hautflüglern (Bienen, Wespen, Ameisen). Selbst bei manchen Wirbeltieren läßt sich künstlich die Entwicklung unbefruchteter Eier anregen (z.B. Frösche). Oft ist Parthenogenese und geschlechtliche Vermehrung mit einem (Generationswechsel verbunden (Saugwürmer, Rädertierchen, Wasser- 129 flöhe, Blattläuse, Gallwespen, Moostierchen) (vgl. B. Rensch, a. a. O. 135 ff).

Wir wollen uns bei folgenden Überlegungen auf die geschlechtliche Fortpflanzung beschränken, doch alles Gesagte gilt in ganz ähnlicher Weise auch für alle Formen ungeschlechtlicher Vermehrung. Die meisten höheren Lebewesen entstehen obligat durch geschlechtliche Vermehrung, genauerhin durch (oder nach) Verschmelzung der Kerne von Samen- und Eizelle. Uns interessieren hier die Fragen, ob es sich bei der Verschmelzung um eine Verursachung oder nur um eine Bedingung für die Entstehung eines neuen Lebewesens handelt und woher das neue Individualprinzip kommt. Ausgeschlossen haben wir schon die Möglichkeit, daß das Individualprinzip des neuen Individuums durch Teilung von den elterlichen Individualprinzipien entsteht und sich aus ihnen zusammensetzt. Die Gameten sind zwar biologische Gebilde, aber keinesfalls Individuen. Das gilt auch für die Samenzellen, die keineswegs die biologischen und naturphilosophischen Forderungen erfüllen, die wir an ein Individuum stellen. Wir möchten ihre Funktion durchaus auf chemische oder physikalische Mechanismen zurückführen. Ganz anders aber das neue Lebewesen: mit der Kernverschmelzung kommt ein Prozeß in Gang, der zu einer Struktur führt, welche die typischen Charakterika der Individualität erfüllt, so daß wir gezwungen sind, nach dem Grund dieser Individualität zu fragen. Da stoßen wir wieder auf unser Individualprinzip, das scheinbar aus dem Nichts aufgetaucht ist. Wir haben uns schon entschlossen keinen Schöpfungseingriff Gottes anzunehmen, wenn eine ‹natürliche› Erklärung möglich ist. Und da bietet sich als einzige an: das Individualprinzip kommt aus der Materie, obwohl es vorher nicht in ihr enthalten war, haben wir uns doch schon aus guten Gründen gegen den Hylezoismus entschieden. Dieser Nebensatz «obwohl es zuvor nicht in ihr enthalten war» bedarf natürlich einer Erläuterung: Grundsätzlich bieten sich, wenn wir die Existenz eines Individualprinzips voraussetzen, drei Möglichkeiten an: 1. das Individualprinzip ist aktuell in der Materie vorhanden. Das würde bedeuten, daß es auch wirksam werden müßte, sind doch Strukturen ohne mögliche Funktionen sinnlose Gebilde. Materie müßte also Lebensfunktionen zeigen. Das aber tut sie als solche nicht. Also ist sie nicht aktuell mit einem Individualprinzip ausgestattet. 2. das Individualprinzip ist potentiell in der Materie vorhanden. Das würde bedeuten, daß die Materie die passive Fähigkeit hätte, unter bestimmten Bedingungen und beim Wirksamwerden äußerer Ursachen (Gott, Sterngeister ...) ein Individualprinzip zu empfangen. 130 3. das Individualprinzip ist virtuell in der Materie vorhanden. Das würde bedeuten, daß die Materie mit einem durchaus nichtmateriellen Vermögen begabt ist, unter bestimmten Umständen Individuen und mit ihnen ein aktuelles Individualprinzip hervorzubringen. Diese Bedingung wäre in unserem Fall die Kernverschmelzung zweier einander entsprechender Gameten. Wir wollen uns hier für die letzte der genannten Möglichkeiten entscheiden, nicht nur weil die erste zum Hylezoismus und die zweite zum exzessiven Kreatianismus führt, sondern weil sie sich harmonisch in ein einheitliches Weltbild einordnet. Greifen wir hier wieder unsere Theorie vom Apeirischen als dem allgemeinen und universellen ontologischen Welthintergrund auf (vgl. WS II, 382). Es ist das, wodurch alles ist. Schon ANAXIMANDER dachte sich diesen Urstoff als mit einer bewegenden Kraft begabt. Er enthält auch das Vermögen, Lebendiges hervorzubringen (vgl. Aristoteles, Phys. III c. 4; 203 b 10). Zur näheren Erklärung und weiteren Entfaltung unserer Theorie wollen wir die Methode der Entgegnung wählen. Wir stellen eine Folge von Bedenken gegen unsere Theorie vor und versuchen sie zu erklären, indem wir diese Bedenken - mehr oder weniger erfolgreich - zurückweisen. 131 FÜNFTES KAPITEL
THEOLOGISCHE BEDENKEN
Natürlich wird der Theologe manche Einwürfe machen wollen: Wird doch die von uns mit einigen Modifikationen aufgegriffene Lehre von der virtuellen Schöpfung (vom Verfasser schon verschiedentlich dargestellt, so in Leben woher?, München 1958, 28; Unsere Welt, München 1959, 214 f) seit etwa 100 Jahren (aber erst seitdem) als fast ketzerisch abgetan. Es bedarf wohl keiner weiteren Erklärung, daß die theologischen Quellen sehr viel eher — wie wir gezeigt haben - für als gegen die Annahme unserer Theorie sprechen. Dennoch versteckt sich hinter der theologischen Frage ein durchaus berechtigtes Anliegen: führt nicht die Annahme einer bloßen Schöpfung dem aktiven Vermögen nach vielleicht zu einer materialistischen Auffassung von der Welt? Ich denke, daß dieser Vorwurf leicht entkräftet werden kann mit dem Hinweis auf die Immaterialität unseres Individualprinzips. Leben ist eben nicht nur eine spezifisch neue Form der Funktionalität (bzw. eine neue Bewegungsform, wie der DIAMAT meint) der Materie. Der spezifisch neue Funktionskomplex des Lebendigen kann nur in einem wissenschaftstheoretisch unzulässigen Monismus, der die Methode nicht optimal dem Objekt und seinen Funktionen, sondern einer Ideologie anzupassen sucht, auf ausschließliche Materiestrukturen zurückgeführt werden. Eine dem Objekt angepaßte Methode führt, wie selbstverständlich, zu einer neuen, grundsätzlich immateriellen Gliedstruktur, weil und insofern wesentlich neue Funktionen nur in wesentlich anderen Gesamtstrukturen begründet sein können. Die Gesamtstruktur des Lebendigen enthält also außer den materiellen Elementen noch wenigstens (und wie aus denkökonomischen Prinzipien folgt: genau) ein immaterielles Element, das Individualprinzip.

Doch - wie gesagt - ist der theologische Einwand, der uns veranlaßt, unsere Theorie von der Immaterialität des Individualprinzips zu wiederholen, relativ unerheblich. Sehr viel erheblicher ist eine theologische Frage, die sich — wie es in der Theologie nun einmal üblich ist — mit reinen Möglichkeiten (potentia absoluta) befaßt: Hätte Gott die Materie auch ohne ein Vermögen auf Leben hin schaffen können? Die Frage ist nicht ganz leicht zu beantworten: Sicher ist sie zu verneinen, wenn man jeden Bezug zum Lebenden aus der Schöpfungsabsicht Gottes ausschlösse. 132 Die Materie als Materie, in unserem Verständnis des Materiebegriffes natürlich, nicht dem des DIAMAT und nicht dem TEILHARDS DE CHARDIN, der bekanntlich «als Materie die Gesamtheit der Dinge, der Energien und der Geschöpfe» versteht, «die uns umgeben, sofern sie für uns greifbar, fühlbar und - im theologischen Sinn des Wortes - natürlich sind», wie er in seinem Buch über den göttlichen Bereich (116) schreibt, ist an und für sich sinnleer. Und Gott kann nichts Sinnleeres schaffen. Ja noch mehr: Gott kann nichts Geringeres schaffen als etwas, das so vollständig auf ihn hingeordnet ist, daß es ihm möglich ist, aus der Unsichtbarkeit der Allgegenwart in die Sichtbarkeit der Geschöpflichkeit einzubrechen. TEILHARD hat das Gemeinte mit dem aus der Geophysik entlehnten Begriff der Emergenz charakterisiert. Ähnlich wie sich eine vulkanische Insel aus der Unsichtbarkeit eines schon langst und immer schon Vorhandenen in die Sichtbarkeit erhebt und schließlich zu einer vulkanischen Eruption kommt, so bricht Gott, gleichsam das Leben und das Bewußtsein emergierend vor sich herschiebend in die Sichtbarkeit ein. Ohne diese letzte Emergenz wäre alles, was sie vorbereitet oder ermöglicht, sinnlos: Materie, Leben, Bewußtsein. Doch auch hier wollen wir TEILHARD nicht ganz folgen, wir glauben, daß es sich beim ersten Auftreten des Lebens auf unserer Erde nicht nur um eine Emergenz, nicht nur um ein Sichtbarwerden, ein «Auftauchen» eines schon längst in Unsichtbarkeit aktuell Vorhandenen handelt, sondern nehmen an, daß mit dem Leben nicht nur etwas Neues auf der Erde sichtbar wird, sondern auch etwas Neues entstanden ist, das zuvor nur dem aktiven Vermögen nach vorhanden war. TEILHARD ist ganz sicher kein Panpsychist oder ein Hylezoist, sondern sehr viel mehr - wenn man schon unbedingt ein Etikett möchte, um es seinem System aufzukleben - ein christlicher Enpantheist. In gewisser Weise ist er Monist. Aber er lehrt keinen Monismus des Lebendigen oder des Bewußtseins, sondern einen durchaus christlichen Monismus, nach dem Gott in allen Dingen ist und wirkt, hin auf seine Erscheinung in Schöpfung. Hätte Gott also die Materie ohne Hinordnung auf das Lebendige schaffen können, ohne ihr wenigstens die passive Fähigkeit mitzugeben, in einem von Anfang an geplanten Schöpfungseingriff, Leben mitzuteilen? Ich denke diese Frage bejahen, hieße die gesamte christliche Schöpfungstheologie in Frage stellen. Sehr viel schwerwiegender als die theologischen, scheinen uns einige philosophische Bedenken gegen unsere Anschauung vom virtuellen Enthaltensein der Individualprinzipien in der Materie zu sein. 133 SECHSTES KAPITEL
PHILOSOPHISCHE BEDENKEN
So merkwürdig es scheinen mag: gerade scholastisch orientierte Philosophen glauben aus philosophischen Gründen unsere Ansicht von dem virtuellen Enthaltensein der Individualprinzipien (Lebensprinzipien. Formen des Lebendigen) in der Materie und somit auch der der Schöpfung des Lebens in Gestalt eines der Materie mitgegebenen Vermögens, eben diesen virtuellen Individualprinzipien, nicht folgen zu können, obwohl sie in einer durchgehenden abendländischen Tradition steht, die auch von der Scholastik übernommen wurde. Ihre Einwände lauten: a) Ist das virtuell in der Materie enthaltene Individualprinzip substantiell oder ein Akzidenz der Materie? Im ersten Fall wäre es keine Eigenschaft der Materie, sondern müßte, wenn überhaupt, ein schon aktuell Seiendes sein. Im zweiten Fall wäre es eine Eigenschaft der Materie, aber nichts mehr. Da nun aus akzidentellem Sein nichts Substantielles werden kann, wäre eine Aktualisation in die Substantialität hinein unmöglich. Wie schon gesagt halten wir den Gegensatz Substanz - Akzidenz nicht für geeignet naturwissenschaftliche Strukturen zu beschreiben und in ihrer Funktion zu erfassen. Die mit dem Vermögen begabte Materie, unter bestimmten Umständen Lebensfunktionen zu zeigen, d. h. ein Individualprinzip hervorzubringen, ist eine Struktur, die u. a. diese Eigenschaft (als Strukturelement verstanden und nicht als Akzidens) besitzt. Sie könnte sie - wenigstens in rein gedanklicher Konstruktion - auch nicht besitzen: ihre physikalischen und chemischen Funktionen im Kleinen (also etwa in der Einzelbeobachtung oder im Experiment) blieben die gleichen. Nicht ganz dasselbe würde gelten, wenn man unter bestimmten Umständen ihr ungestörtes Verhalten im Großen (über viele Jahrmillionen und weite Räume) beobachten konnte. Ist doch das ihr als Strukturelement mitgegebene Vermögen durchaus funktionabel, oder drängt doch als aktives Prinzip in die Funktion und erscheint in diesem Drängen funktionabel. Hier liegt nun eine zweite Schwierigkeit verborgen: b) Wie kann etwas, das nicht oder noch nicht aktuell ist, etwas bewirken (als Wirkursache tätig werden)? Diese Frage ist um so bedeutsamer, als wir die Funktion des Individualprinzips als Mitfunktion in 134 einiger Entsprechung zur aristotelischen Formursache charakterisiert haben. Diese Darstellung bedarf einer Ergänzung.

Wie vermutlich auch die menschliche Seele, obschon durchaus in ihrer Funktion der aristotelischen Formursache ähnlich, vermutlich wirkursächlich tätig werden kann - jedenfalls ließen sich so parapsychologische Effekte am bequemsten erklären (bedeutet doch wissenschaftliches Erklären Aufhellen der wirkursächlichen Zusammenhänge) - nehmen wir an, daß die Gliedstruktur «Individualprinzip» durchaus auch wirkursächlich tätig werden kann. Das gilt vor allem für die Vollerklärung ontogenetischer und phylogenetischer Effekte. Das Individualprinzip formt nicht nur Einheit der Struktur, sondern wirkt sie auch, wenn auch stets und immer in funktionaler Verbindung mit Materie: das soll heißen, daß sie wirkursächlich tätig wird als strukturelle Einheit mit Materie, die von ihm informiert wird. Eine solche Information haben wir durchaus auch anzunehmen für die, durch ein immaterielles Vermögen zur Hervorbringung von Individualprinzipien begabte Materie. Auch die virtus ad vitam, das Vermögen hin auf Leben, wird als die Materie informierendes Prinzip mit Materie wirkursächlich funktional und bewirkt so die chemische Evolution. c) Was ist nun Ursache und was Bedingung für die Aktualisation des Vermögens der Materie, ein Individualprinzip hervorzubringen? Nun, die Bedingungsfragen haben wir schon abgehandelt. Im Regelfall ist bei höheren Lebewesen Bedingung die Verschmelzung von entsprechenden Gameten, versteht man doch unter (notwendiger) Bedingung etwas, das zwar zum Entstehen eines Seienden erforderlich ist (weil ohne sie etwa die Ursache nicht wirken kann), selbst aber keinen positiven Einfluß auf das Werden des Seienden ausübt, wohl aber sein So weitgehend bestimmen kann. Was aber ist die Ursache für die Entstehung eines Individuums, die einen positiven Einfluß auf das Werden des Seienden ausübt und in es eingeht? Die Ursache ist das jetzt, nachdem die notwendigen Bedingungen erfüllt sind, effizient aus der Virtualität in die Aktualität drängende Individualprinzip. Die Ursache für die Aktualisation des Individual-Prinzips liegt in dem noch nicht aktualisierten Prinzip selbst. Das Gemeinte mag ein Beispiel erklären. Nach scholastischer (und nicht nur scholastischer) Ansicht ist der Intellekt eines Menschen eine solche virtuelle Anlage. Er drängt, kraft eigener Dynamik - obwohl noch nicht aktualisiert - in die Aktualisation. Sind die äußeren Bedingungen erfüllt, eine bestimmte psychische Reife, ein entsprechender Erkenntnisinhalt, wird das zunächst reine (aktive) Vermögen Intellekt 134 in die Funktion aktualisiert. Dabei wäre es falsch, ihm als aktiven Vermögen jede wirkursächliche Dynamik absprechen zu wollen. Er drängt - wie gesagt - in die Aktualisation, indem er wirkend die Voraussetzungen schaffen hilft und aktualisiert sich dann auf Grund seiner Aktivität selbst. Es ist also Ursache für die Aktualisation das virtuelle Prinzip selbst, nicht insofern es Potenz, sondern insofern es aktive Potenz ist. Ganz ähnlich denken wir uns das Tätigwerden des reinen, aber aktiven Vermögens der Materie, unter gegebenen Umständen (Bedingungen), Lebensfunktionen zu zeigen, die symptomatisch sind für die Überführung des aktiven Vermögens in die Aktualität. d) Noch ein weiteres Problem wird in diesem Zusammenhang immer wieder angeführt: Wie kann etwas seinsmäßig Niedrigeres (die reine Materie) etwas wesentlich Höheres enthalten (das aktive Vermögen, Individualprinzipien hervorzubringen)? Zunächst einmal möchten wir darauf verweisen, daß wir die Begriffe höher und niedriger für unangebracht halten. Wenn man sie anders versteht als komplizierte und höher komplizierte Struktur, enthalten sie ein wertendes Moment, das durch nichts berechtigt ist, es sei denn durch eine Substanzontolo-gie, die im Individualprmzip einen ‹höheren› weil immateriellen substantiellen Baustein der Gesamtrealität sieht. Damit ist unsere Frage eigentlich schon beantwortet: Bestimmte materielle Strukturen vermögen immaterielle Strukturen einzulagern, die zu einer einheitlichen durch die immaterielle Struktur informierte und mitverursachte Funktion führen oder doch führen können. Ernster zu nehmen ist schon folgende Frage und das in ihr enthaltene Problem: e) Besitzt jedes Elementarteilchen dieses immaterielle Vermögen, auch wenn es mit Sicherheit nicht mehr für den Erstaufbau einer lebendigen Struktur in Frage kommt? In dieser Frage sind eine Reihe von Teilfragen enthalten, die wir uns nacheinander vornehmen wollen. - Ist - wie etwa TEILHARD DE CHARDIN anzunehmen scheint - nach der Biogenese, der Erstentstehung des Lebens, die Materie steril geworden? Ist ihr Vermögen, Lebewesen hervorzubringen, erloschen? Wir denken, daß man diese Frage ruhig verneinen darf. Man kann schlechterdings nicht einsehen, daß sich die gesamte Materie des Kosmos als diese (als mit dem genannten Vermögen begabte) durch die Aktualisation und aktuelle Information einiger weniger Molekel (oder war es nur eines?) durch ein Individualprinzip erschöpft habe, bringt doch Materie bis zum heutigen Tage noch täglich viele Milliarden Mal Individualprinzipien in die Aktualität (bei der Fortpflanzung von Lebewesen)! - Besitzt jedes Elementarteilchen das Vermögen, wenn es mit anderen 136 Elementarteilchen zusammen eine entsprechende makrophysikalische Struktur bilden kann, Lebensfunktionen zu zeigen? Diese Frage ist sicher ernstzunehmender als alle anderen bislang an unsere Theorie gestellten: Welches ist die kleinste Einheit, die ein immaterielles Vermögen der behandelten Art tragen kann oder wirklich trägt? Sind es Elementarteilchen? Wir erinnern uns, daß Elementarteilchen zunächst einmal nichts anderes als strukturierte Beobachtungsergebnisse sind, die einer bestimmten Differentialgleichung gehorchen, wobei das Strukturieren durch den beobachtenden Menschen geschieht. Wir haben schon versucht, von solchen strukturierten Erscheinungen auf Strukturen im Ansich zurückzuschließen (WS II, 343 ff), indem wir als Bedingung der Möglichkeit solcher subjektiver Strukturierung eine - wenn uns auch außer eben dieser nicht sehr viel mehr über das Elementarteilchen bekannt ist —

objektive Struktur von uns unbekannten Eigenschaften, als Elementarteilchen a« sich definierten. Gerne geben wir zu, daß es sich dabei um eine naturphilosophische Überlegung und eine naturphilosophische Aussage handelt, und daß ein Elementarteilchen physikalisch betrachtet sehr viel eher mit der erwähnten Differentialgleichung (etwa der Diracgleichung) identisch ist, mit einer mathematischen Funktion also, als mit irgendeinem Gebilde an sich. Immerhin konnten wir das Elementarteilchen auf Grund einer philosophischen Überlegung als objektive Struktur behaupten, und in solche objektive Strukturen lassen sich noch andere objektive Strukturen einlagern, die form- oder/und wirkursächlich mit der vorgegebenen Struktur verbunden sind. Dennoch glauben wir nicht schon Elementarteilchen als solchen bzw. den ihnen entsprechenden Strukturen als solchen irgendein immaterielles Vermögen zusprechen zu dürfen. Der letzte Träger des immateriellen Vermögens ist weniger ein einzelner Ausdruck des Apeirischen als das Apeirische selbst. Und dieses Apeirische drückt sich in einem virtuell vitalen Aspekt eben erst bei makro-physikalischen Strukturen aus. Erst diese Strukturen zeigen unter bestimmten Umständen in ihrer Wirkursächlichkeit die Anwesenheit eines virtuellen oder aktuellen Individualprinzips. Dazu kommt noch, daß im Bereich der Elementarteilchen das CPT-Invarianztheorem mit mehr oder weniger großer Einschränkung gültig ist (WS II 17 ff). Das bedeutet, daß wir ihnen nicht Zeitlichkeit und damit Ursächlichkeit in der Weise, wie wir sie im Makrophysikalischen (auf Grund statistischer Ausmittlung der möglichen zeitlichen Rückläufigkeit) kennen, zusprechen können. Zeitlichkeit aber und Ursächlichkeit sind notwendige Bedingungen, daß sich auch nur eine Dynamik hin auf das Leben zeigt. Und wir waren übereingekommen, das, was 137 sich in keiner Weise zeigen kann, als nicht existent anzunehmen, selbst auf die Gefahr hin, daß wir ihm damit Unrecht tun. Aber auch diese Überlegung mag als Symptom dafür gelten, daß das Apeirische sich erst in makrophysikalischen Strukturen, diese virtuell oder aktuell belebend, spezifisch ausdrückt. 138 SIEBTES KAPITEL
BIOLOGISCHE BEDENKEN
Wir haben unsere Theorie vom «Wesen» und Werden des Lebendigen immer schon durch biologische Bedenken und Einwände modifiziert und in diesem Modifizieren entwickelt. Dennoch bleiben da einige biologische Fakten, die sich so gar nicht in unsere Vorstellung von der Natur des Lebendigen einzufügen scheinen.
1. Die fremddienliche Zweckmäßigkeit Manche Individuen haben Funktionen ausgebildet, die nicht ihrem eigenen (auch wenn man das Individuum phylogenetisch versteht) Nutzen dienen, sondern fremden. Das bekannteste Beispiel ist die Entwicklung des Vermögens zur Ausbildung von Pflanzengallen zum Nutzen von Bakterien, Pilzen oder Tieren. Diese Gallen entstehen durch chemische Beeinflussung der Wirtspflanze durch den Parasiten. Doch muß diese Beeinflussung auf eine entsprechende Anlage treffen, und diese Anlage wurde offenbar nur zum Nutzen des Gastes ausgebildet. Vor allem Galläpfel stellen äußerst zweckmäßig funktionierende Neubildungen dar. Wählen wir von der Vielzahl der Gallen, die zur Ausbildung eines eigenen Wissenschaftszweiges, der Cecidologie, geführt haben, nur eine einzige aus: Die Gallwespe Dryophanta folii legt in das Innere der Blattnerven von Eichenblättern' ein Ei ab. Es entsteht ein Vegetationskegel, der schließlich die Larve ganz einhüllt. Auf der Unterseite des Blattes bilden sich auf den Blattnerven große rötlich-grün gefärbte Galläpfel. Diese Gallen besitzen eine derbe Epidermis, ein Assimilationsgewebe und eine hohlkugelförmige Schicht von Festigungsgewebe. Die an die Larvenkammer anschließenden Zellen enthalten Reservestoffe und dienen der Larve als Nährgewebe. Aus dieser Folii-Galle schlüpft im Herbst ein weibliches Insekt aus, das dann seine (stets unbefruchteten) Eier in eine Eichenknospe, genau auf den Vegetationskegel, ablegt. Im Frühling hüllt dann der Kegel das Ei ein und bildet eine hochdifferenzierte Knospengalle, die im Juni fertige Männchen und Weibchen entläßt. Es gibt Gallen, die nach Fertigentwicklung der Larven sich durch abspringende Deckel genau im rechten Augenblick öffnen und so die 139 junge Gallwespe ins Freie lassen (vgl. S. Strugger, Biologie l, Frankfurt 1962, 123 f). «Dieses Beispiel zeigt, wie Tier und Pflanze in erstaunlicher und kaum begreiflicher Art aneinander angepaßt sind» (ibid.). Wichtig ist in diesem Zusammenhang, daß die Eiche durch die Fähigkeit, Gallen auszubilden, wie erwähnt, keinen entsprechenden Selektionsvorteil hat. Obwohl manche Gallen die Pflanze vor Tierfraß zu schützen scheinen, ist der Schaden, der der Pflanze durch Entzug wichtiger Aufbaustoffe zugefügt wird, wesentlich größer, so daß phylogenetisch die Entstehung der Anlage zur Ausbildung von Gallen kaum nach dem darwinistischen Schema: Überbevölkerung - Mutation -Selektion - Isolation erklärt werden kann. Begegnen wir hier nicht einem phylogenetischen Überindividuum, das nicht nur das Phylum Eiche, sondern auch das Phylum Gallwespe umfaßt? Bedeutet das nicht eine Ausdehnung unseres Individuumbegriffs ins Uferlose? Müssen wir nicht ein Überindividualprinzip annehmen, das nicht nur die Phylogenese der Eiche steuert, sondern auch die der

Gallwespe, so daß sie entsprechende chemische Auslöser für die Ausbildung der Gallen entwickeln kann? Ich muß gestehen, daß ich auf diese Frage keine Antwort kenne. Es scheint jedenfalls, daß wir dem Apeirischen Steuerungsmechanismen zusprechen müssen, die durch den Begriff des Individualprinzips nicht erfaßt werden.
2. Blindheit des Instinkts Viele Tiere haben im Verlauf ihrer Phylogpnese komplizierte Verhal-tensmechanismen ausgebildet, die ebenfalls nur gezwungen durch das darwinistische Schema erklärt werden können. Bei den niederen Tieren ist das Verhalten vor allem durch Reßexe bestimmt, d. h. durch erblich festgelegte, von einem Außenreiz ausgelöste, immer gleichbleibende durchschnittlich vorteilhafte Reaktionen. Daneben bildeten sich auch Taxien aus, d. h. durch die Reizrichtung bestimmte Einstellungseinrichtungen der Bewegung. Reflexe und Taxien sind wesentliche Elemente des Instinktverhaltens, wenn sie es auch nicht voll erklären. Den Außenreiz, der instinktbegründete Verhaltensweisen auslost, nennt man Schlüsselreiz oder einfach Auslöser (reaktionsauslösender Reiz). Dem Auslöser muß eine Stimmung entsprechen, d. h. ein Stau reaktionsspezifischer Energien im ZNS. So unterscheidet man Hunger-, Paarungs-, Eiablagestimmungen. Symptom für eine solche Stimmung ist oft ein ungeordnetes Umherlaufen, Fliegen oder Schwim- 140 men (Appetenzverhalten). Durch allzu häufiges Auftreten von Schlüsselreizen kann sich die reaktionspezifische Energie erschöpfen. Durch übernormale Auslöser wird die Reaktion des Tieres verstärkt. So bevorzugt etwa eine Silbermöve zur Bebrütung ein größeres und kontrastreicher angemaltes Attrappenei vor ihren eigenen Eiern, selbst wenn sie sich gar nicht darauf setzen kann. Die Starrheit mancher instinktgegebener Verhaltensweisen erweist sich im Leerlauf: so versucht ein Wüstenfuchs, der ein Stück Fleisch in einem Zimmerboden vergraben will (ein in der Wüste u.U. sehr sinnvolles Verhalten), mit der Schnauze immer wieder ‹imaginären› Sand über das Fleisch zu schütten. Übernormaler Stau der reaktionsspezifischen Energie, wie sie etwa in Konfliktsituationen angereichert werden kann, führt zu Übersprungshandlungen: das sind nicht situationsgerechte Instinktreaktionen, wie wir sie auch beim Menschen etwa als Verlegenheitshandlungen beobachten können (überflüssige Putzgesten wie Kopfkratzen) (vgl. B. Keusch, a. a. O., 297 ff). «Es ist also ein großer Irrtum zu glauben, daß jede Instinkthandlung, deren arterhaltende Leistung z. B. der Nahrungsaufnahme dient, unbedingt vom Hunger verursacht werden müsse» (K. Lorenz, Das sogenannte Böse, Wien 1965, 124). «In allen Fallen aber trägt die . . . befohlene Aktivität nicht den Charakter des ‹Reflexes›, das heißt, die gesamte Organisation der Instinktbewegung verhält sich nicht wie eine Maschine, die, solange sie nicht gebraucht wird, unbegrenzte Zeit passiv brachliegt und ‹wartet›, bis jemand auf den Knopf des Anlassers drückt» (ibd. 129). Zumeist wird eine auf ein bestimmtes Ziel ausgerichtete Handlung durch eine ganze Reihe von Instinkten, eine Instinkthierarchie, bestimmt. Ein einfaches Beispiel mag das zeigen: LEYHAUSEN gab jagdfreudigen Katzen eine Maus nach der anderen und beobachtete die Teilhandlungen. Nach und nach fielen dabei bestimmte instinktgesteuerte Teilhandlungen aus. Zunächst stellte die gesättigte Katze das Fressen ein. Sie tötete noch einige Mäuse und ließ sie liegen. Dann erlosch auch der Zwang zum Tötungsbiß. Die Katze beschlich und haschte nur noch nach Mäusen. Als auch die Bewegungsweisen des Fangens erschöpft waren, belauerte sie nur noch Mäuse, vor allem solche, die weit entfernt waren, jene, die ihr über die Vorderpfoten liefen, ließ sie unbeachtet. Bei solchen Versuchen läßt sich abzählen, wann eine bestimmte reaktionsspezifische Energie, die einem eigenen instinktiven Verhalten (Fressen, Töten, Fangen, Belauern) zugeordnet ist, erschöpft ist (vgl. ibd., 31). 141 Zudem scheint der Instinkt oft (nicht immer) blind zu sein, d. h. das Tier hat keinerlei Einsicht in das Zweckmäßige seiner Handlungen: Eine Languedoc-Grabwespe sucht Springschrecken, um sie durch einen sehr gezielten Stich in das ZNS zu lahmen und so das für die Nahrung ihrer Larven unbedingt unverfaulte Fleisch zu konservieren. Dabei transportiert sie ihre Beute an einem der langen Fühler zu ihrem ‹Nest›. Schneidet man dem gelähmten Beutetier beide Fühler ab, wird es von der Springschnecke nicht wiedererkannt, obwohl ein Transport an einem der Beine genauso praktikabel wäre. Sie läßt ihre Beute liegen (vgl. J. H. Fahre, Aus der Wunderwelt der Instinkte, Meisenheim 1950, 43). Allerdings ist der Instinkt nicht immer ganz so unplastisch wie im vorgenannten Fall. Es paßt sich etwa ein Bienenvolk durchaus auch im Bereich instinktgebundener Verhaltensweisen veränderten Situationen an. Als RÖSCH aus einem Bienenstock alle Jungbienen herausnahm, leisteten die zurückgebliebenen Altbienen (die auf Grund ihres Instinktverhaltens als Sammelbienen fungierten), die Aufgaben der Jungbienen (Brutpflege) mit. Die schon verkümmerten Futterdrüsen begannen wieder zu wachsen. Nach zahlreichen Versuchen kam LINDAUER zu dem Schluß, daß «die Zahl der Bienen, die sich mit bestimmten Aufgaben befassen .. . einzig und allein von den Bedürfnissen des Bienenvolkes» abhängt (vgl. R.. Chauvin, a. a. O., 25 f). DARCHEN konnte zeigen, daß die Bienen ihre Wabengröße und Form den Umständen anpassen. CHAUVIN schreibt dazu, «daß einige althergebrachte Anschauungen über die Instinktgebundenheit der Tiere den Tatsachen nicht entsprechen... Wir haben es mit einer Sozialgruppe zu tun, die Schwierigkeiten «erkennt» und die gestellten Probleme durch Versuche und Retuschen selbst dann zu lösen versucht, wenn es offenbar keine Lösung gibt» (ibd. 74). Nun kann man diese Erscheinung plastischen Instinkts sicher erklären, wenn man die Bienensozietät als ein

Individuum versteht. Die Instinktgebundenheit kann so weitgehend gelockert erscheinen, daß man versucht ist, der Sozietät. wenn schon keine Intelligenz, so doch «Tätigkeiten höherer Ordnung» als es die instinktgebundenen der Einzelbienen sind, zuzuschreiben. Doch nun zu unserem Problem: Einzelne Momente, die wir im Begriff ‹Individuum› zusammengefaßt haben, scheinen nicht mehr zu stimmen: Das Individualprinzip scheint i. zu Leistungen zu befähigen, die eine Intelligenz voraussetzen und 2. Leistungen zu verhindern, die eigentlich ganz selbstverständlich wären. Werden wir genötigt, dem Apeirischen eine eigentliche Einsicht in den Sinn zweckmäßigen Han- 142 delns zuzuschreiben die dann unter Umständen durch physiologisch bedingt Blockierungen nicht immer realisiert werden können? Auch diese Frage lässt sich – wie mir scheint - nach dem Stand unseres heutigen Wissens nicht sicher beantworten. Jedenfalls scheint ein Rückgriff auf eine dem tierischen Individualprinzip übergeordnete oder vorgeordnete Instanz notwendig zu sein, manche instinktgebundene Verhaltensweisen zu erklären. 143 ACHTES KAPITEL
DIE ENTSTEHUNG DES LEBEWESENS DURCH URZEUGUNG
Vielleicht hat mancher Leser bei den Versuchen theologische, philosophische und biologische Schwierigkeiten unseres Lösungsansatzes zum Begreifen der Natur und der Entstehung der Lebewesen aufzulösen, das Thema aus dem Blick verloren. Wir haben im vierten Kapitel die Theorie vertreten, und wie wir hoffen erhärtet, daß das Individualprinzip aus einem aktiven Vermögen der Materie herausgeführt wird, wenn die normalen Bedingungen (etwa der Zeugung) gegeben sind. Wir haben die Zeugung als Bedingung für das Aktualisieren einer aktiven, der Materie innewohnenden Anlage verstanden, nicht aber als deren Ursache. Ist es nun unmöglich, andere Bedingungen als die normalen 7.u erstellen, die gleichsam ‹künstlich› zur Aktualisierung des Individual-Prinzips führen? Dabei wollen wir uns vergegenwärtigen, daß die Bedingungen im Normalfall durchaus nur chemischer und physikalischer Art sind, und somit dem Belebtsein der Erzeuger dieser Bedingungen eine durchaus zweitrangige Rolle zukommt. Wenn aber die Bedingungen nicht eigentlich biologischer (jetzt sei dieser Terminus einmal naturphilosophisch verstanden), sondern chemischer und physikalischer Art sind, ist es durchaus denkbar, daß wegen anders gearteter chemischer und physikalischer Prozesse als jener, die etwa bei der Kernverschmelzung statthaben, die Aktualisierung des Individualprinzips geschehen kann. Wir haben im empirischen Abschnitt den Leser an die Schwelle einer solchen uneigentlichen Zeugung (generatio aequivoca) herangeführt. Philosophisch gesehen, scheint uns kein wesentlicher Unterschied . zwischen der biogenen und abiogenen «Zeugung» eines Lebewesens zu bestehen. Die Problematik ist also nicht philosophischer, sondern rein biologisch-technischer Art. Ich denke, daß heute kaum ein Biologe daran zweifelt, daß, wenn es einmal gelänge, Phagen-DNS künstlich (abiogen) zu erzeugen und sie in ein entsprechendes Bakterium einzubringen, dieses Bakterium natürliche Phagen produziert. Im Prinzip abiogen wäre schon ein DNS-Faden erzeugt, wenn es gelingt, einen natürlich gebildeten Faden chemisch so zu teilen, daß die Bruchstücke für sich sicher nicht in der Lage sind, die genetische Funktion der Zelle im Sinne der Neuproduktion von Phagen umzuinformieren und ihn dann in einem 145 geeigneten chemischen Verfahren wieder in seiner ursprünglichen chemischen Struktur zu synthetisieren. Gemeinhin wendet man gegen solche Überlegungen ein, daß Phagen, weil und insofern sie ihre Vermehrungsfähigkeit ‹aus eigener Kraft, verloren haben (oder vielleicht niemals besaßen), keine Lebewesen seien. Nun ist obligates Schmarotzertum, wenn es zur Bereitung stoffwechselgerechter Verbindungen führt, kein Grund, einem solchen Schmarotzer Belebtsein abzusprechen. Warum sollte aber dann ein obligates Schmarotzertum in Bezug auf die für die Umstrukturierung und Vermehrung der DNS notwendigen Energien (die Phagen entnehmen bekanntlich diese Energien dem Zellstoffwechsel ihres Gastes), ein Kriterium für das Unbelebtsein eines Gebildes sein? Vor allem wenn feststeht, daß Phagen, wenn auch keinen eigenen Energiestoffwechsel, so doch erste Anzeichen eines Baustoffwechsels zeigen. Ferner entwickeln sich Phagen über nichtinfektiöse Zwischenstufen und deuten damit so etwas wie eine Ontogenese an. Zudem können Phagen durchaus mutieren oder Erbanlagen nach dem crossing-over-Mechanismus austauschen, wenn auch solche Mutationen nur in den Wirtszellen möglich sind. Wir haben also recht gute Gründe, Phagen (wenigstens die DNS-haltigen) für belebt zu halten, obwohl im Grenzbereich natürlich eine Entscheidung, ob eine Funktion durch ein aktualisiertes Individual-Prinzip informiert ist (eigentliche biologische Funktion) oder durch ein virtuelles (uneigentliche biologische Funktion) kaum sicher zu begründen ist. Im Dezember 1967 konnte man selbst in Boulevardblättern lesen, es sei einem Team amerikanischer Wissenschaftler gelungen, eine biologisch aktive (das muß nicht schon das Zeigen von eigentlichen Funktionen sagen) DNS künstlich

zu synthetisieren. Unter Leitung von R. KORNBERG synthetisierte das Team die im ΦΧ 174-Phagus enthaltene DNS. Sie war, obgleich in einem zellfreien System aufgebaut, biologisch aktiv und verhielt sich also genauso wie die DNS natürlicher φ Χ 174 Phagen. Dieser Phagus ist übrigens recht klein. Er enthält, bei einem Durchmesser von 30 nm und einem Molekulargewicht von 6,2 X 106 etwa 25 % NS. Ein DNS-Faden hat nun aber ein Molekulargewicht von etwa 1,6 X 106 (SINHEIMER), so daß ziemlich sicher ist, daß unser Phagus nur einen einzigen Faden besitzt. Schon vor gut 10 Jahren haben KORNBERG und seine Mitarbeiter das zur DNS-Synthese nötige Enzymsystem aus lebenden Zellen gewonnen. Es gelang ihnen damals unter bestimmten Voraussetzungen (vor allem müssen alle vier Nukleotidarten angelagert an ATP und eine hochmolekulare DNS als Muster vor- 146 handen sein), aus diesem zellfreien System DNS zu synthetisieren, die in ihrer Struktur im wesentlichen mit der der eingebrachten DNS übereinstimmt. Doch ein wichtiger Mangel haftete diesen Versuchen an: die neu synthetisierte DNS war nicht biologisch aktiv! S. SPIEGELMANN (und sein Team) konnte - wie schon berichtet - als erster einen kleinen Phagus replizieren, indem er dessen NS in ein aus Enzymen und energiereichen Nukleotiden bestehendes zellfreies System einbrachte. Doch gehörte dieser Phagus zu den RNS-Phagen, deren ‹Belebtheit› recht umstritten ist. KORNJBERG wählte sich ein sehr einfaches DNS-Phagus aus (eben das φ Χ 174)- Mitte 1967 gelang es, aus dem Bakterium Escherichia coli eine hochgradig gereinigte DNS-Polymerase (das für die DNS-Synthese verantwortliche Enzym) zu gewinnen, mit deren Hilfe sich die Phagen-DNS ohne Fehler in vitro replizierte. Aber wieder war die DNS biologisch inaktiv! Wo lag der Fehler? In fünf Laboratorien der USA hatte man schon im Vorjahr herausgefunden, daß bei der natürlichen Bildung infektiöser Phagen neben der DNS-Polymerase noch ein anderes Enzym auftrat, das «Verbindungsenzym» (polynucleotide joining enzyme). Dieses Enzym katalysiert die Verknüpfung des Phosphatrestes der einen Seite der Nukleotidkette mit dem OH-Ende der anderen Seite, bewirkt also einen Ringschluß der Kette. Inzwischen hatte man auch herausgefunden, wie der Vervielfältigungsmechanismus von φ Χ 174 bei einer Vermehrung in vivo (in der lebenden Zelle) abläuft: Der infizierende einsträngige und ringförmig geschlossene DNS (+)-Strang bildet in der Zelle zunächst eine komplementäre Matrix ((-)-Strang) in Gestalt eines ebenfalls geschlossenen Ringes aus. Dieser Doppelring bildet die replikative Form der Phagus-DNS. Löst man die Phagus-DNS aus den Zellen, erweisen sich sowohl der Doppelring, als auch die (weil nur durch Wasserstoffbrücken miteinander verbundene) leicht zu lösende Einzelstränge als infektiös. Damit war erklärt, warum der erste Strang, obwohl er fehlerfrei repliziert wurde, nicht infektiös war: Er war nicht ringförmig geschlossen, sondern eine offene Kette. Bringt man nun neben der Polymerase noch das Verbindungsenzym in die zellfreie Lösung ein, entsteht die replikative Form der Phagus-DNS. Der neuentstehende (künstliche) (-)-Strang unterscheidet sich in keiner Weise von dem natürlichen (-)-Strang. Er, wie auch der Doppelstrang sind infektiös. Wenn man ihn also in ein entsprechendes zellfreies System bringt, bildet er, selbst synthetisch gebildet, synthetische (+)-Stränge und damit die replikative Form der DNS des Virus aus, die vollständig künstlich synthetisiert ist. 147 Es handelt sich bei solchen Versuchen und Ergebnissen natürlich nicht um eine Simulation der ersten Biogenese. Alle erwähnten Versuche setzen schon die Existenz von bislang nur in lebenden Systemen gebildeten Enzymen und Mutter-DNS voraus Handelt es sich um eine erste Laborsynthese des Lebendigen? Die Beantwortung der Frage hängt davon ab, ob man sich entschließen kann, den φ Χ 174-Phagus als aktuell belebt anzusehen. Wir haben auf die Schwierigkeit, die einer solchen Annahme entgegensteht, schon verwiesen: es fehlt den Phagen ein eigenes Enzymsystem als Voraussetzung für einen Energiestoffwechsel. J. HAAS versucht seine sicher dogmatische These, daß eine Laborsynthese des Lebendigen unmöglich sei, mit dem Hinweis zu erhärten, daß die Phagus-DNS genau so wenig ‹belebt› sei wie Vitamine, Enzyme und Hormone [vgl. Künstliches Leben, in Stimmen der Zeit 181 (1968),405]. Das ist aber ganz offensichtlich nicht der Fall, da der Phagus, außer dem genannten, alle anderen wesentlichen Lebenskriterien erfüllt. Das aber gilt nicht für Vitamine, Enzyme und Hormone. Auch die künstlich erzeugte DNS (sie mag 5 oder 6 Gene enthalten) veranlaßte u. a. die Bildung einer ganz normalen Proteinhülle und vermag all das, was natürliche Phagen-DNS leistet. Man mag zu diesen Versuchen stehen, wie man will: Ein wichtiger Schritt zumindest auf dem Wege zur Laborbiosynthese ist beschriften. Es scheint kein allzu weiter Weg mehr zu sein, bis sie einmal in einer Weise gelingen wird, die keinen Raum für irgendeinen vernünftigen Zweifel übrig läßt. Es würden also in anderer als der normalen Weise Bedingungen geschaffen, die es den virtuell in der Materie enthaltenen Individualprinzipien ermöglichten, in die Aktualität auszubrechen. Das wäre ein Triumph unserer Naturwissenschaften. Ein Triumph aber auch unseres naturphilosophischen Bemühens um die Erkenntnis des «Wesens» der Materie, von deren Außen wir so manches zu wissen glauben, so viel immerhin, daß wir kaum noch wagen, etwas Sicheres darüber zu sagen (und um soweit zu kommen, muß man schon allerhand wissen), von deren Innen wir aber bislang kapitulieren mußten. Dieses geheimnisvolle Innen beginnt sicher schon heute, zwar noch recht zögernd, sich auch der Naturwissenschaft zu erschließen. 148

LITERATURHINWEISE
Zeitschriften Vor allem folgende Zeitschriften behandeln das vorgelegte Thema regelmäßig in allgemeinverständlicher Form: - Die Umschau, Halbmonatsschrift über die Fortschritte in Wissenschaft und Technik, Umschau Verlag, Frankfurt (seit 1900)) - Kosmos, Franckh'sche Verlagshandlung, Stuttgart (seit 1904)
Gesamtdarstellungen Handbuch der Biologie, hrsg. von Fr. Gessner, Konstanz 1965 Die Evolution der Organismen I, hrsg. von G. Heberer, Stuttgart 31967 Biologie l, hrsg. von S. Strugger (Fischer Lexikon 27), Frankfurt 1962 Biologie II, hrsg. von B. Rensch (Fischer Lexikon 28), Frankfurt 1963
Einzeldarstellungen Ballauf, Th., Das Problem des Lebendigen, Bonn 1949 Botsch, W., Molekulargenetik, Heidelberg 1967 Chauvin, R., Tiere unter Tieren, Frankfurt 1967 Christian, P., Gesetzlichkeit und Leistungsgrenzen biologischer Regelsysteme in N. A. Luyten Red.), Struktur und Dynamik der Materie, Freiburg 1967 Fabre, J. H., Aus der Wunderwelt der Instinkte, Meisenheim 1950 Frank, H. (Hrsg.), Kybernetik, Frankfurt 61966 Friedrich-Freksa, H., Die stammesgeschichtliche Stellung der Virusarten, in Heberer, G., Die Evolution der Organismen, Stuttgart 1954 Frieling, H., Das Leben, Stuttgart 1949 Hesse, G., Der Lebensbegriff bei den Klassikern der Naturforschung, Jena 1943 Jordan, P., Die Physik und das Geheimnis des organischen Lebens, Braunschweig 1947 Laskowski, W., Elemente des Lebens, Berlin 1966 Mayer, E.. Das Problem der Individualität im organischen Sein in Wissenschaft und Weißheit 30 (1967), 130 ff Meyer-Abich, A., Naturphilosophie auf neuen Wegen, Stuttgart 1948 Nardi, Fr., Organismus und Gestalt, München 1942 Nelter, H., Theoretische Biochemie, Berlin 1959 Oparin, A. L, Das Leben, seine Natur, Herkunft und Entwicklung, Jena 1963 Portmann, A., Probleme des Lebens, Basel 1949 Schulz, ]., Die Maschinentheorie des Lebens, Leipzig 1929 Weidel, W., Virus und Molekularbiologie, Heidelberg 1964 Wessel, H., Kausalität und Zweckmäßigkeit in der lebenden Natur, Berlin 1963 Wieland, T h. und Pfleider, G., Molekularbiologie, Frankfurt 1967 149