lbop-gen3 - alliance française halifax · populations: exercices d'application, simulations...
TRANSCRIPT

LBOP-Gen3
Génétique des populations et génétique
quantitative
Joël Cuguen
Laboratoire de Génétique et Evolution des Populations Végétales
(UMR CNRS 8016), Bureau 101, Bâtiment SN2
Université de Lille 1
59655 Villeneuve d'Ascq Cedex
Tel: 03 20 43 40 24
Fax: 03 20 43 69 79
Email: [email protected]
http://www.univ-lille1.fr/gepv

Thèmes de recherche du laboratoire:
Biologie évolutive et génétique des populations
• Stérilité mâle cytoplasmique et gynodioecie.
• Evolution des systèmes d'auto incompatibilité
• Evolution des traits d'histoire de vie (floraison, durée de vie,
dispersion)
• Tolérance aux métaux lourds
• Flux géniques entre formes cultivées et sauvages: risques
agronomiques et environnementaux associés à la diffusion des
OGM.
• Biologie de la conservation d'espèces menacées

Enseignements de LBOP Gen3
Cours
• Analyse génétique des caractères à variation continue: la génétique
quantitative, 8 heures, Joël Cuguen
• Génétique des populations approfondie: 18 heures, Xavier
Vekemans (responsable de l'UE)
TD/TP
• Séances de TD/TP de génétique quantitative et de génétique des
populations: exercices d'application, simulations sur ordinateur

Ouvrages de référence
• Griffiths, AJF, Gelbart, WM, Miller, JH & Lewontin, RC 2001. Analyse Génétique Moderne. De Boeck Université
• Ridley, M 1997 Evolution Biologique, De Boeck
• Ressources pédagogiques sur le site du laboratoire: http://www.univ-lille1.fr/gepv
• Réseau d’enseignement en Génétique (GENET)http://www.univ-tours.fr/genet/

Ouvrages de référence (spécialisés)
• Falconer, D. S. and T. F. C. Mackay (1996). Introduction to Quantitative Genetics. London, Prentice Hall.
• Kearsey, M. J. and H. S. Pooni (1996). The Genetical Analysis of Quantitative Traits, Chapman & Hall.
• Lynch, M. and B. Walsh (1998). Genetics and Analysis of Quantitative Traits, Sinauer Associates Inc.
• Verrier E., Brabant P., Gallais A. (2001). Faits et concepts de base en génétique quantitative. Polycopié INA Paris-Grignon, 134 p.http://www.inapg.inra.fr/dsa/uvf/GQ/GQintro.htm
• Crédits: P. Touzet; C. Dillmann & F. Hospital; E. Verrier, Ph. Brabant & A. Gallais…

Génétique des caractères à variation continue:
Introduction
• Essor de la Génétique: dissection du phénotype en caractèresunitaires simples (Mendel 1865, Bateson 1902,…)
• Phénotype à variation discontinue ("discrète"), binaire, qualitative; ex. type sauvage / mutant
– Couleur des yeux, groupes sanguins, etc…
• Mais il existe aussi des caractères "complexes", à variation phénotypique continue, quantitative
– Ex: caractères agronomiques, masse, taille, production, maladies, traits adaptatifs
Génétique quantitative: génétique des caractères issus d’une mesure

Génétique des caractères à variation continue
Domaines d'applications multiples
• Génétique et amélioration des plantes et des productions animales:
sélection artificielle (dirigée par l'homme)
• Médecine: génétique des maladies multifactorielles
• Ecologie évolutive: génétique des caractères adaptatifs, étude de
l'évolution et de la sélection naturelle

Plan du cours de
génétique des caractères à variation continue
• Introduction
• Exemples de caractères à variation continue
• Les questions spécifiques
• Le modèle multifactoriel infinitésimal de Fisher
• La décomposition de la variation phénotypique
• L'héritabilité
• La sélection
• Les outils statistiques
• L'estimation des paramètres génétiques
• La dissection biométrique des caractères quantitatifs
• Les QTL: gènes impliqués dans la variation

Les caractères à variation continue

Quelques exemples de caractères à variation
continue
• Caractères biométriques
– Taille des individus, poids, croissance
– Pression artérielle, taux de cholestérol, glycémie
– Nombre de soies de l'abdomen de la drosophile
– Nombre de facettes oculaires
• Caractères agronomiques
– Taille de portée chez les animaux
– Teneur en huile chez le Maïs
– Nombre de grains par épi de Blé
– Date de floraison chez le Blé
• Maladies multifactorielles / maladies "monogéniques"
– Diabète de type II,
– Prédisposition à l'obésité
• Caractères adaptatifs
– Précocité floraison, fertilité, tolérance facteurs du milieu

Exemples de caractères quantitatifs
La distribution de la variation suit une loi normale
tiré de Dillmann et Hospital, 2002

Distribution de la taille d'étudiants de l'université
du Connecticut

Soies abdominales de la Drosophile
Le nombre de soies abdominales est un caractère très étudié car facile à
phénotyper et répondant bien à la sélection
tiré de Hartl, 1994
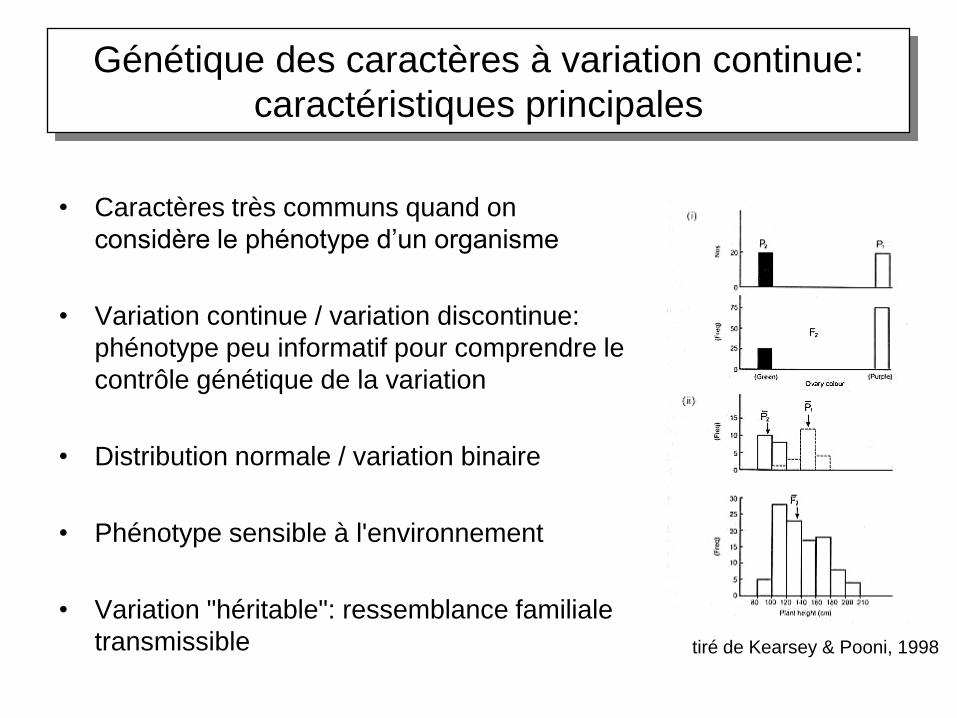
Génétique des caractères à variation continue:
caractéristiques principales
• Caractères très communs quand on
considère le phénotype d’un organisme
• Variation continue / variation discontinue:
phénotype peu informatif pour comprendre le
contrôle génétique de la variation
• Distribution normale / variation binaire
• Phénotype sensible à l'environnement
• Variation "héritable": ressemblance familiale
transmissible tiré de Kearsey & Pooni, 1998

Génétique des caractères à variation continue:
les questions spécifiques
• Description de la variation quantitative: outils statistiques adaptés
• Modèle statistique de prédiction de l’évolution des caractères quantitatifs sous l'influence de la sélection naturelle ou dirigée par l'homme
– La ressemblance entre individus apparentés
– L'origine génétique de la variation quantitative: le modèle multifactoriel infinitésimal
• Décomposer la variation phénotypique: la notion d'héritabilité
• L'utilisation de l'héritabilité pour la sélection
• L'identification des facteurs génétiques: la recherche des QTL: analyse du déterminisme génétique de la variation des caractères
– Nombre de gènes impliqués
– Répartition dans le génome
– Effets des gènes et caractérisation

Le modèle multifactoriel infinitésimal de Fisher

Controverse mutationnistes / biométriciens
• William Bateson (1861-1926), généticien éminent, jugeait incompatibles les lois de Mendel et la variation héréditaire des caractères à variation continue, s'opposant en cela vigoureusement à la théorie darwinienne de l'évolution graduelle
• Karl Pearson (1857-1936), statisticien non moins éminent et darwinien convaincu, pensait que les applications des lois de Mendel étaient limitées et n'expliquaient pas la variation des caractères à variation continue

Le modèle infinitésimal de Fisher (1918)
• Annoncé par Yule (1906), East (1910)
• Fisher, R. A. (1918). "The Correlation Between Relatives on the Supposition of Mendelian Inheritance." Philosophical Transactions of the Royal Society of Edinburgh 52: 399-433.
• La ressemblance entre individus apparentés résulte du partage d'allèles en commun
• La variation observée pour les caractères quantitatifs résulte:
– de l'action combinée d'un grand nombre de gènes (infini) à hérédité "mendélienne" individuelle
– de l'influence de "l'environnement"

Sir Ronald Aylmer Fisher (1890-1962)
Généticien et statisticien
• 1890: Born in East Finchley, London.
• 1909: Student at Gonville and Caius College, Cambridge.
• 1917: Married Ruth E. Guiness.
• 1919: Started work as a statistician at Rothamsted Experimental Station.
• 1933: Chair of Eugenics at University College, London.
• 1943: Balfour Professor of Genetics, Cambridge University.
• 1957: President of Gonville and Caius College.
• 1962: Died Adelaide, South Australia.

Bases génétiques et environnementales de la
variation quantitative
tiré
de
Str
ickb
erg
er, 1
98
8
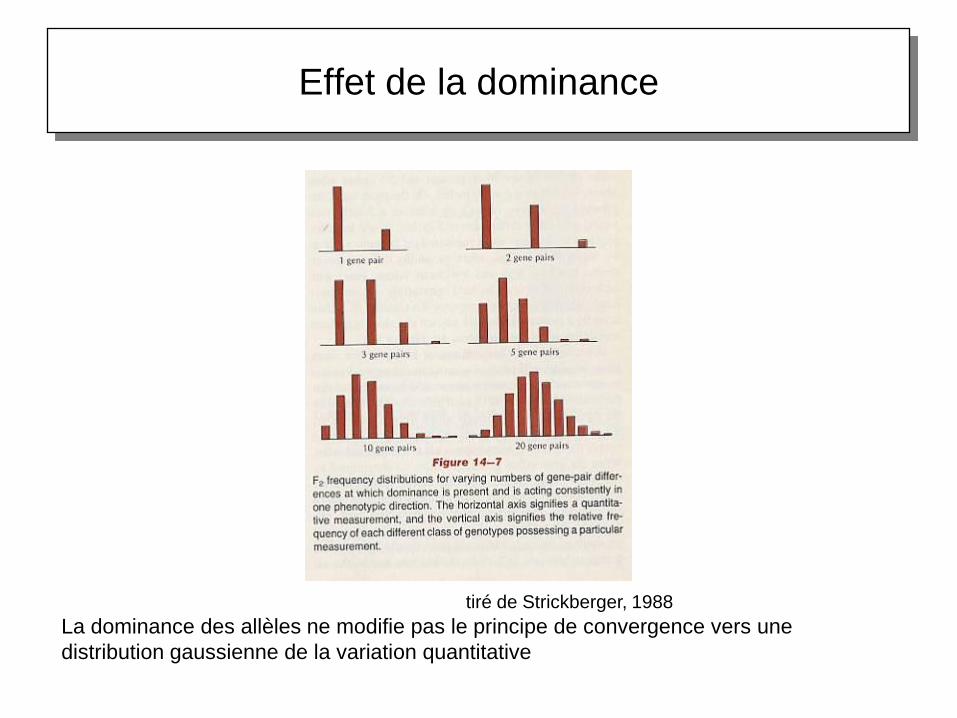
Effet de la dominance
La dominance des allèles ne modifie pas le principe de convergence vers une
distribution gaussienne de la variation quantitative
tiré de Strickberger, 1988

Normalité des caractères quantitatifs

Normalité des caractères quantitatifs
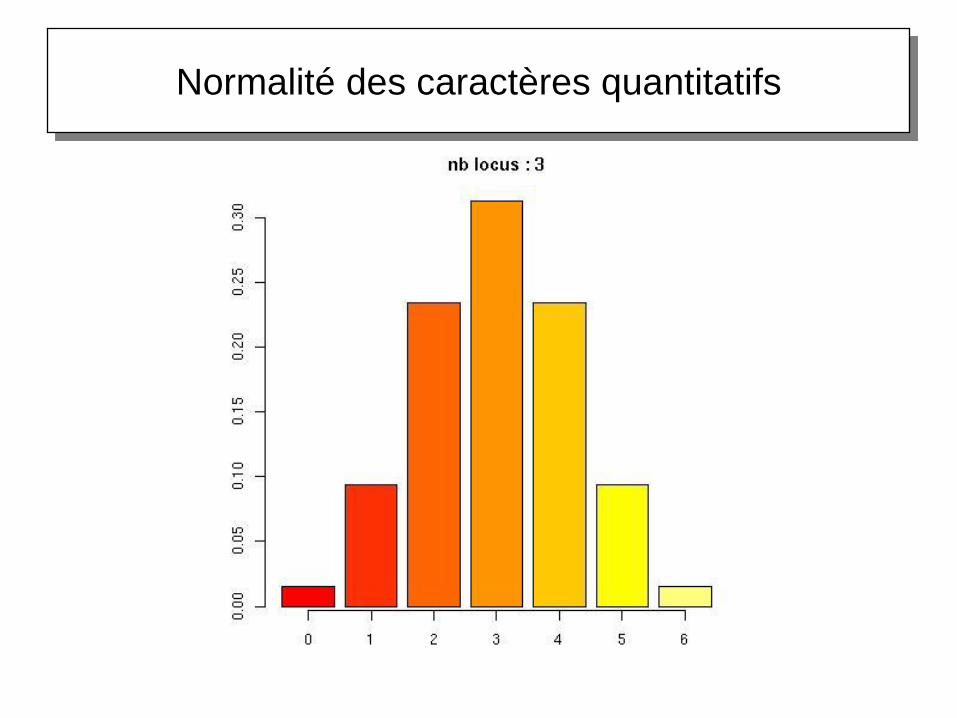
Normalité des caractères quantitatifs

Normalité des caractères quantitatifs

Normalité des caractères quantitatifs
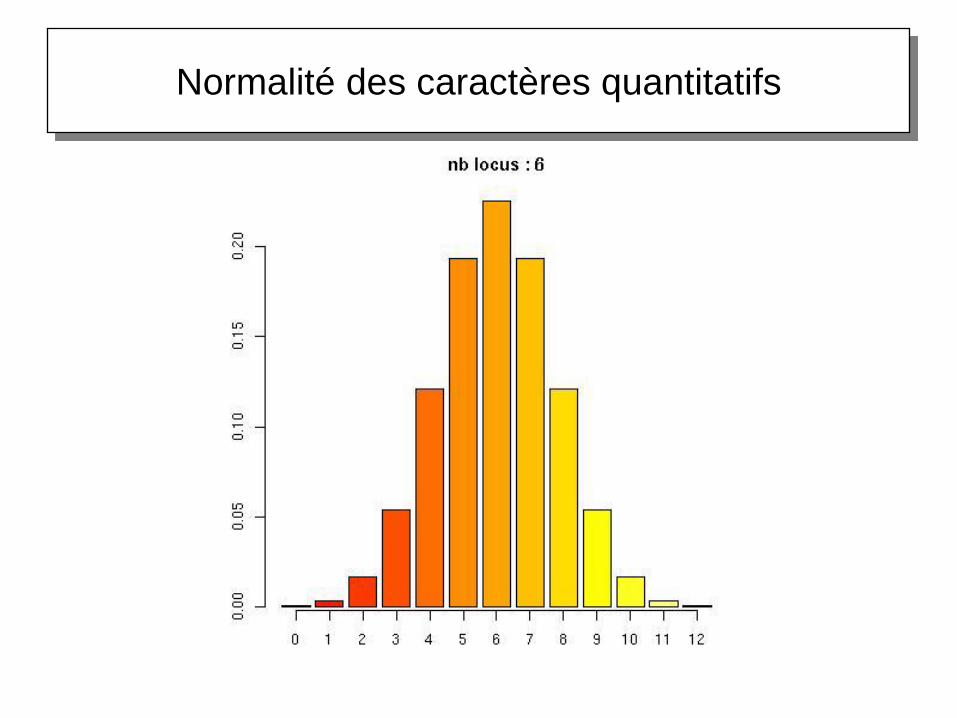
Normalité des caractères quantitatifs
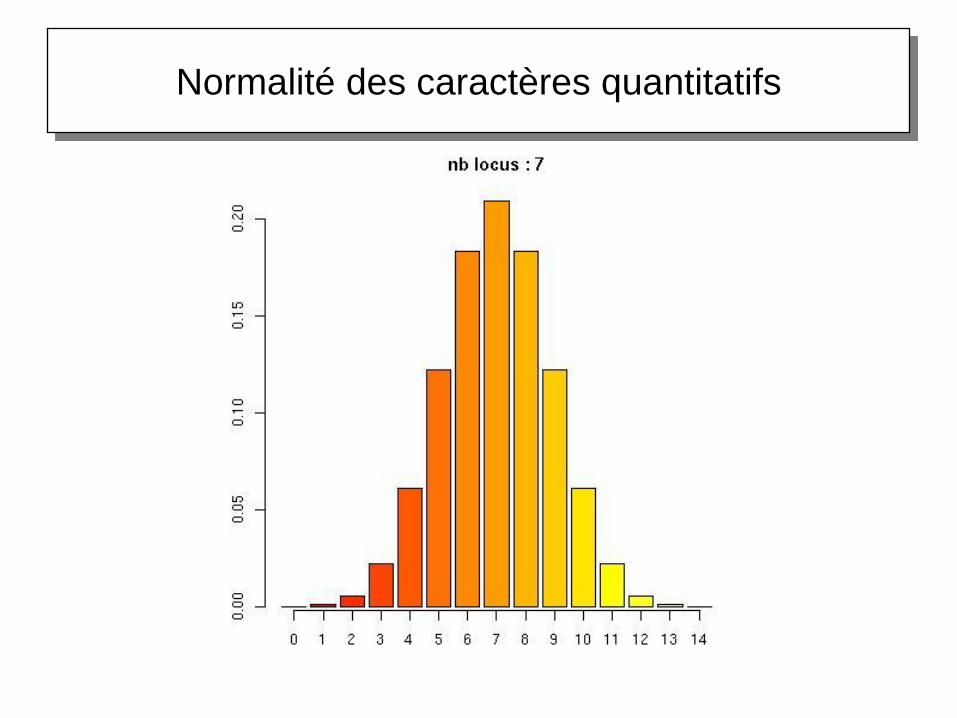
Normalité des caractères quantitatifs
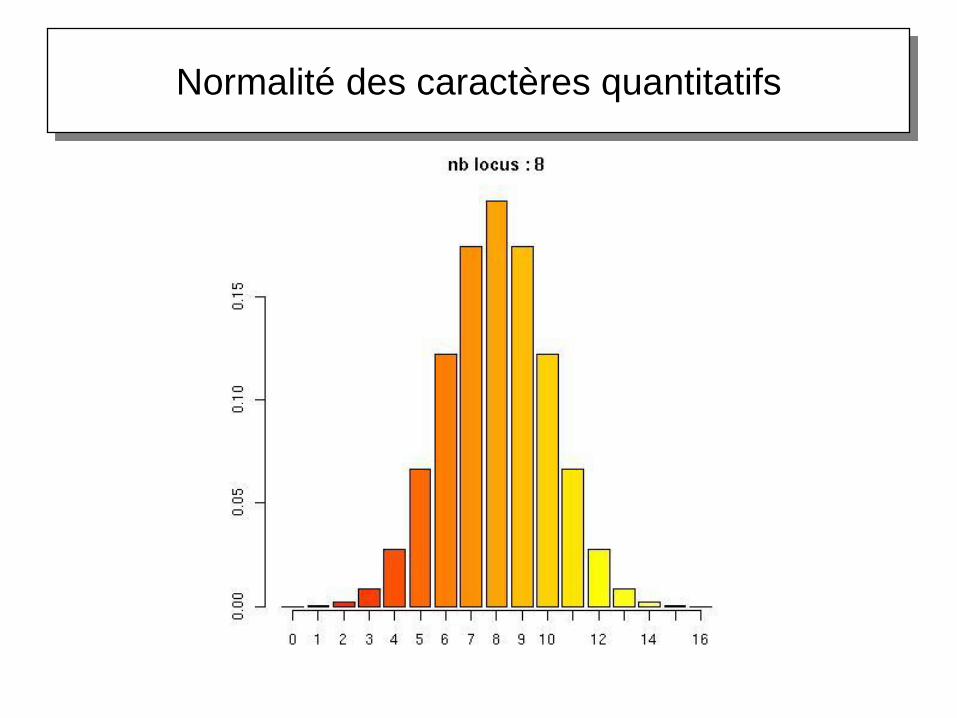
Normalité des caractères quantitatifs

Normalité des caractères quantitatifs

Normalité des caractères quantitatifs
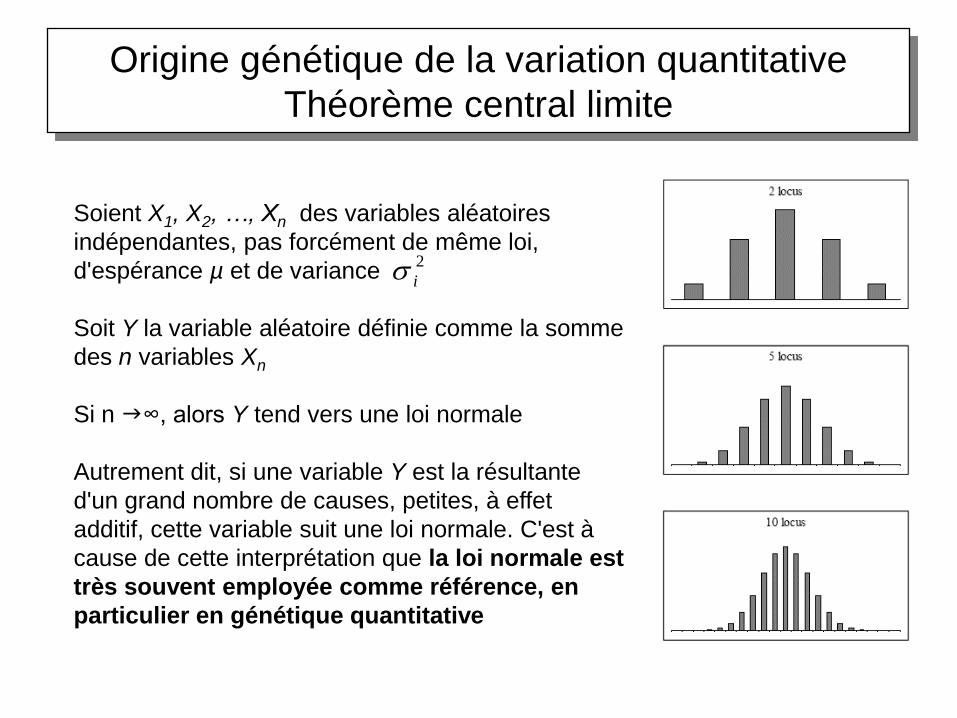
Origine génétique de la variation quantitative
Théorème central limite
Soient X1, X2, …, Xn des variables aléatoires
indépendantes, pas forcément de même loi,
d'espérance µ et de variance
Soit Y la variable aléatoire définie comme la somme
des n variables Xn
Si n ∞, alors Y tend vers une loi normale
Autrement dit, si une variable Y est la résultante
d'un grand nombre de causes, petites, à effet
additif, cette variable suit une loi normale. C'est à
cause de cette interprétation que la loi normale est
très souvent employée comme référence, en
particulier en génétique quantitative
2
i

Origine génétique de la variation quantitative
La normalité de la distribution de la variation
quantitative résulte de l’action combinée de
nombreux gènes et de l’impact de
l’environnement

Relation phénotype - génotype
• Soient 3 gènes présentant 2 allèles chacun, R1, R2 et R3 codominants impliqués dans la coloration de l'enveloppe du grain de blé. Les allèles R1, R2 et R3 confèrent une production de pigments accrue.
• R1=R=R3=1,
• b1=b2=b3=0
• On peut donc distinguer 7 phénotypes selon la dose d'allèle R en proportion variables dans une F2 :
Nombre
d'allèles R0/6 1/6 2/6 3/6 4/6 5/6 6/6
Proportion en
F21/64 6/64 15/64 20/64 15/64 6/64 1/64

Contrôle de la coloration des grains de blé
tiré de Bernard et al, 1993

Relations nombre de gènes – nombre de
génotypes
Le nombre de génotypes est une fonction exponentielle du nombre de locus

Essor et limites de la génétique quantitative
statistique « classique »
• Le modèle infinitésimal de Fisher fournit une excellente explication de la base génétique de la variation quantitative
• Une conséquence négative du modèle est cependant de prédire l'impossibilité d'identifier les gènes et leurs allèles puisqu'il sont en nombre infini et donc qu'ils n'ont chacun qu'un effet infinitésimal
• Suivant Kempthorne (1957), cité dans Verrier, Brabant et Gallais 2001), on peut dire que
« l'objectif de la génétique quantitative est de développer des modèles statistiques pour l'expression phénotypique, en face d'une impossibilité partielle d'identification des génotypes et des facteurs de l'environnement »
• Vrai jusqu’à l’avènement de la génétique moléculaire et de la génomique mais il est maintenant possible d’identifier les gènes concernés

Principe fondamental de la génétique quantitative:
décomposition du phénotype
• La valeur phénotypique d'un individu dépend de sa valeur génotypique et d'un effet dû à l'environnement
Phénotype = génotype + environnement
P = G + E
• G: valeur génotypique
• E: non transmis à la descendance

Décomposition de la valeur génotypique
Signification de A, D, I
• Dans une espèce diploïde à reproduction sexuée, la valeur génotypique d'un individu dépend d'une composante additive, d'un résidu de dominance et d'un résidu d'interaction entre les gènes (épistasie)
G = A + D + I
• Lors de la reproduction sexuée, la contribution d'un parent se limite à la transmission d'un gamète haploïde issu de méiose et résultant d'une recombinaison. Les interactions génotypiques sont rompues et chaque allèle est transmis indépendamment
En conséquence:
– A: La valeur génétique additive est transmise à la descendance
– D: Le résidu de dominance n'est pas transmis à la descendance dans un gamète haploïde
– I: Le résidu d'interaction épistatique n'est pas transmise à la descendance dans un gamète haploïde

La décomposition de la variation d'un caractère
quantitatif
• Dans une population
Var(P) = Var(G) + Var(E)
• Var(P) : variance phénotypiqueVar(G) : variance génotypiqueVar(E) : variance environnementale
Var(G) = Var(A) + Var(D) + Var(I)
• Var(A) : variance additiveVar(D) : variance de dominanceVar(I) : variance d'interaction entre loci (épistasie)

Partition de la variance du poids des nouveau-nés
chez l'Homme
Cause de la variation % du total
Génétique
Additive 15
Non additive 1
Sex 2
Total génotypique 18
Environnement
Génotype maternel 20
Environnement maternel général 18
Environnement maternel immédiat 6
Age de la mère 1
Rang de naissance 7
Cause non identifiée 30
Total environnement 82
tiré de Falconer & Mackay, 1996

Le paramètre fondamental de la génétique
quantitative: l’héritabilité

La notion d'héritabilité
•Définition de l'héritabilité: définition populationnelle et non
individuelle!
Proportion de la variation phénotypique d'une population qui est
d'origine génétique: Elle mesure les contributions relatives des
différences génétiques et des différences dues à l'environnement
dans la variation phénotypique totale des individus
•Au sens large:
•Au sens strict:
EG
G
P
G
VV
V
V
VH
2
EIDA
A
EG
A
P
A
VVVV
V
VV
V
V
Vh
2

Partition de la variance et héritabilité
chez Drosophila melanogaster
Nb. de soies
abdominales
Longueur
du thorax
Taille
des
ovaires
Nb. d'œufs
pondus /
4 jours
Phénotypique
(totale)
Var(P)100 100 100 100
Additive Var(A) 52 43 30 18
Non additive Var(D) + Var(I) 9 6 35 44
Environnementale Var(E) 39 51 35 38
H20.61 0.49 0.65 0.62
h20.52 0.43 0.30 0.18
tiré de Falconer & Mackay, 1996

Remarques importantes sur la notion d'héritabilité
• L'héritabilité est utilisée pour séparer les composantes génétiques et environnementales de la variation phénotypique
• La variance génétique est due à des différences entre génotypes: Il n'y a pas de variance génétique dans une population génétiquement homogène
• La variance environnementale est due à des différences dans l'environnement de vie, de croissance et de développement des individus
• L'héritabilité au sens large est une évaluation relative de la variation génétique présente, incluant les effets de dominance et d'épistasie
• L'héritabilité au sens strict est une évaluation relative de la variation génétique dite additive, transmissible, accessible à la sélection naturelle ou artificielle (dirigée par l'homme)
• L'héritabilité d'un caractère est une notion relative à une populationétudiée dans un milieu donné: ce n'est pas une mesure absolue pour un caractère et une espèce.
• Pour une population donnée, l'héritabilité évolue au cours du temps en fonction des changements de l'environnement, de l'effet de la sélection, de la perte ou du gain de diversité génétique (mutation, migration)…

Exemples de valeurs d'héritabilité pour des espèces
animales et végétales domestiquées
tiré de Hartl, 1994

Héritabilité de divers traits chez l'Homme
tiré de Hartl, 1994
L'estimation de l'héritabilité chez l'homme est rendue plus
difficile du fait de la présence d'environnement commun
entre les apparentés

Classification des caractères en fonction de leur
héritabilité
En amélioration génétique, on distingue trois principales catégories de caractères selon la valeur de l'héritabilité:
• Caractères à variation peu héritable (h2 <0.2): essentiellement des caractères liés aux aptitudes de reproduction et de viabilité des jeunes. D'une part, ces caractères sont très sensibles aux conditions de milieu (Var(E) très élevée) et, d'autre part, des phénomènes génétiques non additifs (notamment des interactions de dominance) induisent probablement d'importantes variations sur ces caractères (Var(D) élevée).
• Caractères à variation moyennement héritable (0,2 < h2 < 0,4): essentiellement des caractères liés à l'intensité d'une production.
• Caractères à variation fortement héritable (h2 > 0,4): essentiellement des caractères liés aux caractéristiques qualitatives des produits, notamment leur composition, qui sont beaucoup moins sensibles aux variations liées au milieu que les précédents ou bien des caractères qui n'ont pas fait l'objet d'une sélection intense.

La réponse à la sélection naturelle ou dirigée
par l’homme

Différents modes de sélection
tiré de Kearsey & Pooni, 1998

Sélection directionnelle par troncature
tiré de Hartl, 1994

Héritabilité et réponse à la sélection
tiré de Hartl et Clark, 1997

Héritabilité et réponse à la sélection
• "Breeder’s equation" ou équation de prédiction de la réponse à la
sélection
R=h2S
• S = différentiel de sélection = µs – µ où µs est la moyenne de la
sous-population sélectionnée et u est la moyenne de la population
totale
• R = réponse à la sélection = µ' – µ où et µ' la moyenne dans la
génération fille
• Sans héritabilité, pas de réponse à la sélection et par conséquent
aucune possibilité de changement génétique ni d'évolution!
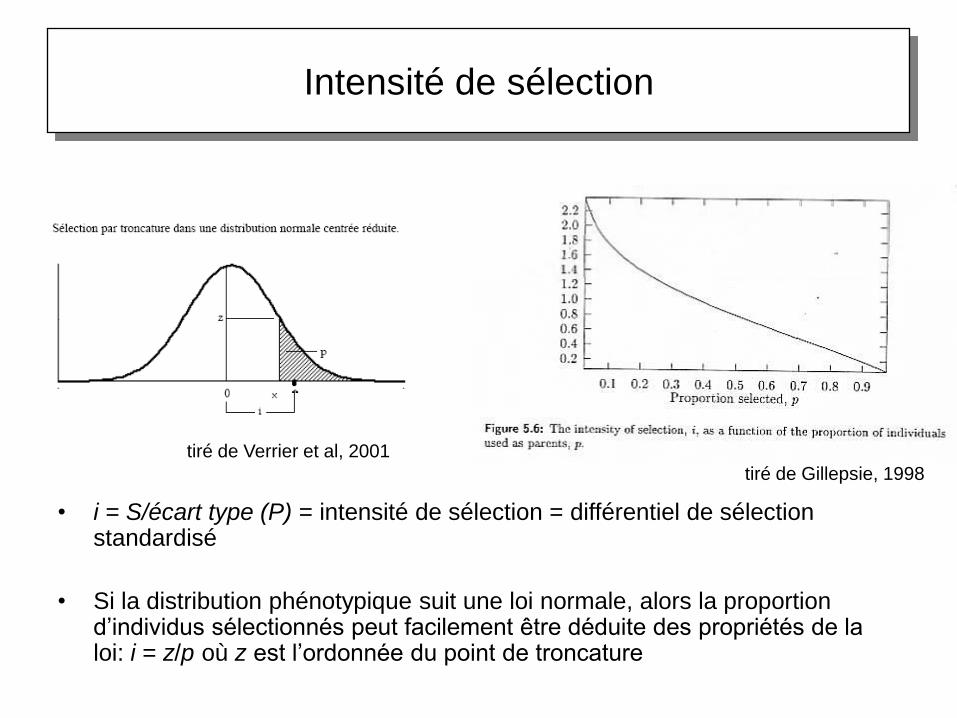
Intensité de sélection
tiré de Gillepsie, 1998
tiré de Verrier et al, 2001
• i = S/écart type (P) = intensité de sélection = différentiel de sélection standardisé
• Si la distribution phénotypique suit une loi normale, alors la proportion d’individus sélectionnés peut facilement être déduite des propriétés de la loi: i = z/p où z est l’ordonnée du point de troncature

Validation expérimentale
Sélection du poids des graines chez le Haricot
tiré de Hartl, 1994

Validation expérimentale
Réponse à la sélection chez la Drosophile

« Illinois Long Term Selection Corn Experiment »
Dudley & Lambert, 1992.
Validation expérimentale
Les réponses à long terme à la sélection artificielle
Le cas du maïs
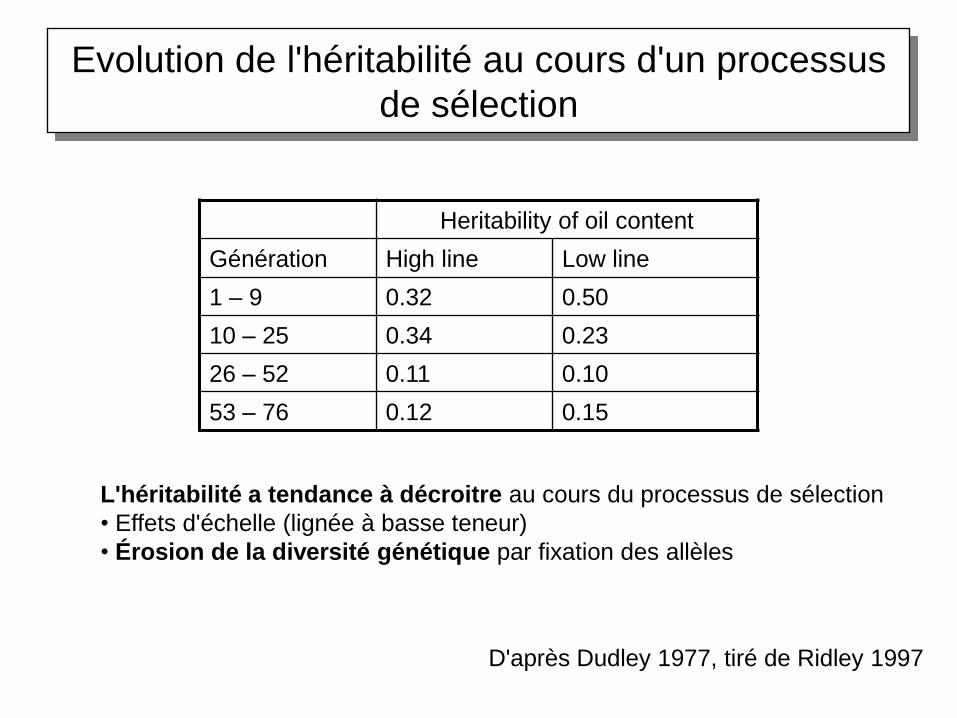
Evolution de l'héritabilité au cours d'un processus
de sélection
Heritability of oil content
Génération High line Low line
1 – 9 0.32 0.50
10 – 25 0.34 0.23
26 – 52 0.11 0.10
53 – 76 0.12 0.15
D'après Dudley 1977, tiré de Ridley 1997
L'héritabilité a tendance à décroitre au cours du processus de sélection
• Effets d'échelle (lignée à basse teneur)
• Érosion de la diversité génétique par fixation des allèles

Réponse à la sélection:
effet de la pêche sur la taille des saumons
Tiré de USHERB
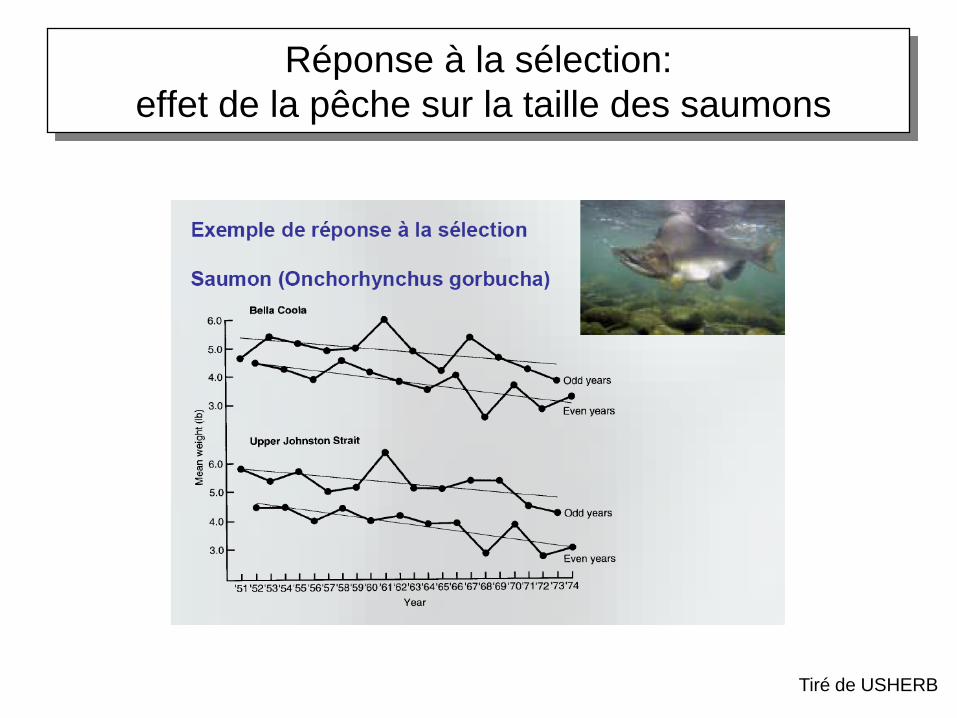
Réponse à la sélection:
effet de la pêche sur la taille des saumons
Tiré de USHERB

Estimation des paramètres génétiques et de
l'héritabilité

Les outils statistiques
• Description des distributions
• La moyenne
• la variance
• la corrélation
• la régression
• L'analyse de la variance: ANOVA
• La régression linéaire multiple

La variance: mesure de la variation
• Variance: mesure standard de la variation, c'est-à-dire de la
dispersion autour de la valeur moyenne
Var(x) = Moy[(xi - µx)2]
• Propriété: additivité des variances
Var(x + y) = Var (x) + Var(y) si les variables x et y sont
indépendantes
Donc
Var(P) = Var (G) + Var(E)
Var(G) = Var (A) + Var(D) + Var(I)

La covariance: mesure de la covariation
et de la ressemblance entre apparentés
• Covariance: mesure de l'association entre deux variables, c'est-à-dire de leur covariation
Cov(x,y) = Moy[(xi - µx)(yi - µy)]
• Propriété: La covariance et la variance sont étroitement liées. En particulier
Cov(x,x) = Var(x)
• Les valeurs phénotypiques P1 et P2 de deux individus apparentés (parent/descendant, frères, jumeaux) sont généralement proches car ils partagent à la fois des gènes et un environnement en commun
• En conséquence
Cov(P1,P2) = Cov(G1,G2) + Cov(E1,E2)
Sous réserve d'indépendance entre génotype et environnement

Ressemblances dues à un environnement
commun
Cov(E1,E2)
• Effets maternels
• Environnement familial
• …
• La randomisation permet de contrôler en partie ces effets
• En génétique humaine, on utilise dans la mesure du possible des
jumeaux ou des enfants séparés à la naissance afin d'éliminer la
covariance phénotypique due à un environnement commun

Ressemblances dues à l'apparentement
• G = effet moyen des gènes paternels + effet moyen des gènes maternels + effets d'interactions entre les gènes
• Sachant que G = A + D + I, et sous réserve d'indépendance entre A, D et I
Cov(G1,G2) = Cov(A1,A2) + Cov(D1,D2) + Cov(I1,I2)
= Var(A) + Var(D) + Var(I)
• Pour un locus donné, deux individus apparentés peuvent partager 0, 1 ou 2 allèles avec une probabilité πi (i = 0, 1 ou 2)
• P. ex. dans une population non consanguine, un parent et son descendant partagent systématiquement 1 allèle pour chaque gène, deux vrais jumeaux ont leurs deux allèles en commun et deux frères peuvent avoir 0, 1 ou 2 allèles en commun en probabilité respectivement de ¼, ½, ¼
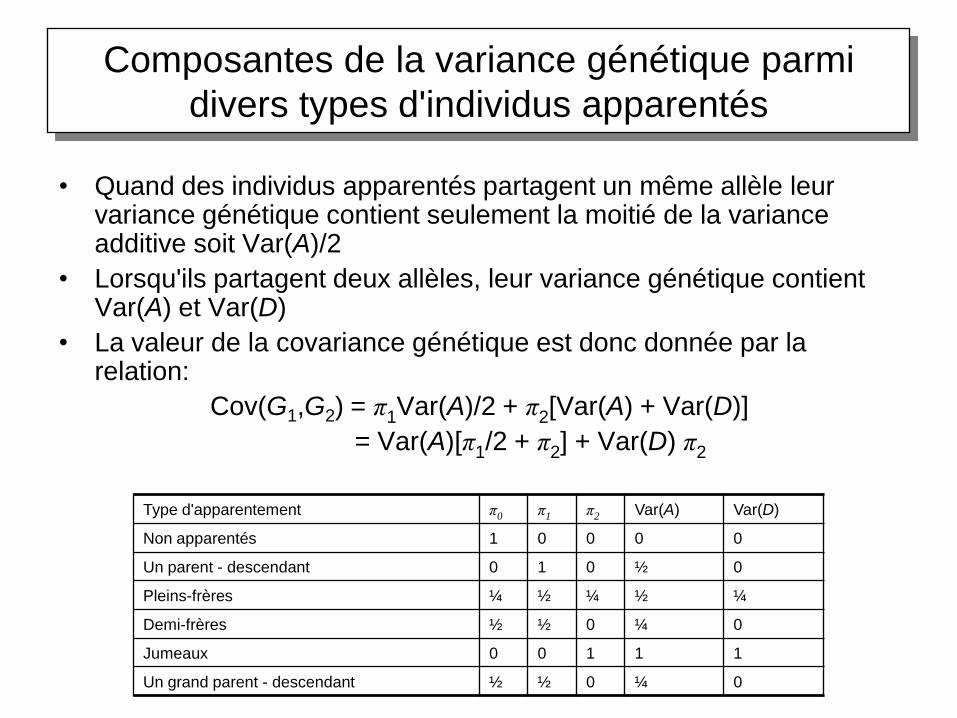
Composantes de la variance génétique parmi
divers types d'individus apparentés
• Quand des individus apparentés partagent un même allèle leur variance génétique contient seulement la moitié de la variance additive soit Var(A)/2
• Lorsqu'ils partagent deux allèles, leur variance génétique contient Var(A) et Var(D)
• La valeur de la covariance génétique est donc donnée par la relation:
Cov(G1,G2) = π1Var(A)/2 + π2[Var(A) + Var(D)]
= Var(A)[π1/2 + π2] + Var(D) π2
Type d'apparentement π0 π1 π2 Var(A) Var(D)
Non apparentés 1 0 0 0 0
Un parent - descendant 0 1 0 ½ 0
Pleins-frères ¼ ½ ¼ ½ ¼
Demi-frères ½ ½ 0 ¼ 0
Jumeaux 0 0 1 1 1
Un grand parent - descendant ½ ½ 0 ¼ 0

Ressemblances dues à l'apparentement et
estimation des paramètres génétiques
• L'estimation des paramètres génétiques se base sur la comparaison entre la ressemblance théorique entre apparentés et la ressemblance mesurée expérimentalement pour le caractère considéré
Apparentement Covariance Régression (b) ou corrélation (t)
Descendant et un seul parent ½ Var(A) b = ½ Var(A) / Var(P)
Descendant et la moyenne des parents ½ Var(A) b = ½ Var(A) / ½ Var(P)=Var(A) / Var(P)
Demi-frères ¼ Var(A) t = ¼ Var(A ) / Var(P)
Pleins frères ½ Var(A) + ½ Var(D) t = [½ Var(A) + ½ Var(D)]/Var(P)

Estimation de l'héritabilité:
distribution de la longueur de la fleur de tabac (East 1916)
tiré de Verrier et al, 2001

Le modèle à un gène
• Valeurs des génotypes incluant les effets additifs (a) et de
dominance (d)
Parent2
A2A2
Parent 1
A1A1
Moyenne des parents
m = 1/2(A1A1 + A2A2)
-a +a
d
F1 (A1A2)

Le modèle à un gène: les moyennes
Après centrage sur la moyenne des parents:
• Moyenne du caractère chez les parents P1 et P2:
+a et -a respectivement
• Moyenne du caractère dans la F1: d
• Moyenne du caractère dans une F2:
1/4(-a) + 1/2(d) + 1/4(+a) = 1/2d
• dans une F3: 1/4(-a) + 1/2(1/2d) + 1/4(+a) = 1/4ddans une Fn: (1/2)n-1d
• Moyenne du caractère dans un BC1.1: 1/2(a) + 1/2(d)
• Moyenne du caractère dans un BC1.2: 1/2(-a) + 1/2(d)
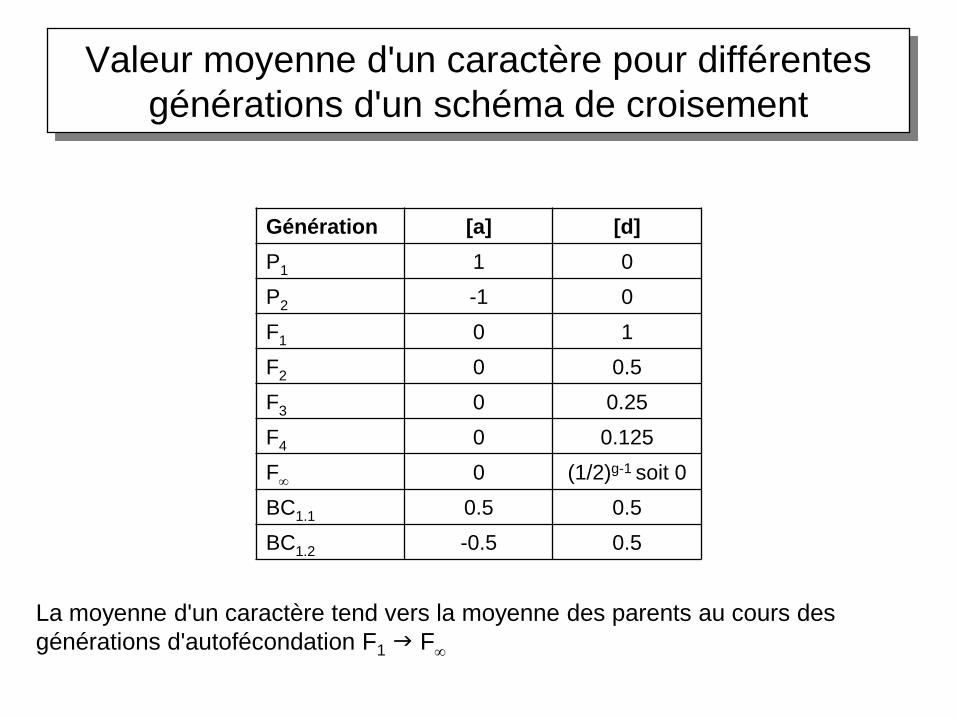
Valeur moyenne d'un caractère pour différentes
générations d'un schéma de croisement
Génération [a] [d]
P1 1 0
P2 -1 0
F1 0 1
F2 0 0.5
F3 0 0.25
F4 0 0.125
F 0 (1/2)g-1 soit 0
BC1.1 0.5 0.5
BC1.2 -0.5 0.5
La moyenne d'un caractère tend vers la moyenne des parents au cours des
générations d'autofécondation F1 F

L'hétérosis pour la taille
chez le tabac
tiré de Kearsey & Pooni, 1998
L'évolution de la moyenne
au cours des générations
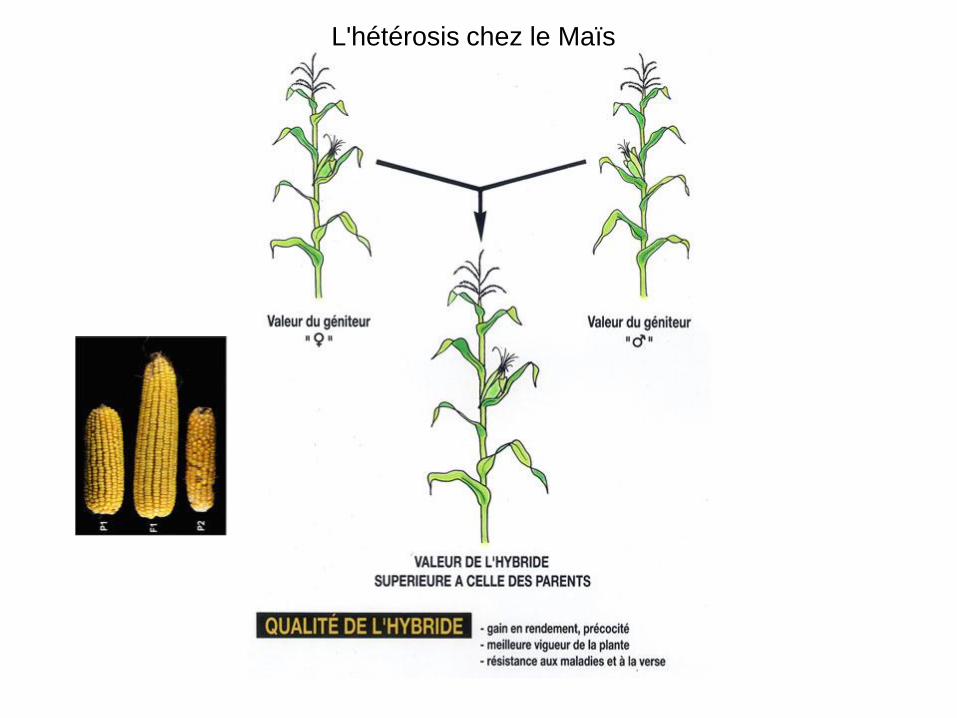
L'hétérosis chez le Maïs

L'hétérosis chez le porc

Le modèle à un gène: variance génétique d'un
caractère dans une F2
• La variance est calculée selon la formule:
• Var(G) dans une F2
Var(G) = 1/4(-a - 1/2d)2 + 1/2(d - 1/2d)2 + 1/4(a - 1/2d)2
= 1/4(a2 + ad +1/4d2) + 1/2(1/4d2) + 1/4(a2 - ad +1/4d2)= 1/2a2 + 1/4d2
• a2 + d2 représentent respectivement les composantes de la variance dues à l'additivité des effets alléliques et aux effets de dominance entre allèles.
• La démonstration pour un seul gène se généralise aisément à plusieurs gènes en supposant l'indépendance des effets des différents gènes (pas d'épistasie).
i
ii xxVar2

Exemple :
Héritabilité de la longueur de la fleur chez le tabac
Composantes de la variance a2 d2 ad E Variance estimée
P1 0 0 0 1 48
P2 0 0 0 1 32
F1 0 0 0 1 46
F2 1/2 1/4 0 1 130.5
BC1.1 1/4 1/4 1/2 1 85.5
BC1.2 1/4 1/4 -1/2 1 98.5
BC1.1 + BC1.2 1/2 1/2 2 184
1. E = 1/3(48 + 32 + 46) = 42
2. F2 – E = 1/2a2 + 1/4d2 = 130.5 – 42 = 88.5
BC1.1+ BC1.2 – 2E = 1/2a2 + 1/2d2 = 184 – 84 = 100
3. 1/2a2 + 1/4d2 = 88.5
1/2a2 + 1/2d2 = 100
4. 1/2a2 = 77, donc a2 = 154, et d2 = 46

Héritabilité de la longueur de la fleur chez le tabac
• En conséquence,
• Les composantes de la variance calculées à partir de la F2 sont:
Var(A) = 1/2a2 = 77.0
Var(D) = 1/4d2 = 11.5
Var(E) = E = 42
Var(P) = 130.5
55.0154
46soit
2
2
a
d
a
d

Héritabilité de la longueur de la fleur chez le tabac
• L'héritabilité au sens strict h2 estimée dans la F2 est égale à 0.59, alors que l'héritabilité au sens large H2 est de 0.68.
• En revanche, les héritabilités estimées dans les BC sont de 0.42 et 0.54 respectivement! Ce point illustre l'importance de la diversité génétique dans la notion d'héritabilité (à la limite, l'héritabilité calculée dans une F1 est nulle car la variance génotypique est nulle!).
• L'héritabilité est donc une notion très relative: Elle dépend complètement du polymorphisme de la population concernée, des interactions d'expression entre allèles, et bien sûr de l'importance de l'impact des facteurs environnementaux sur l'expression du caractère.

La pente de la droite de régression parent-enfant est une estimation de l’héritabilité du
caractère
h2=VA/VP
Estimation de l'héritabilité:
La régression parent-enfant
tiré de Dillmann et Hospital, 2002

S
R
h2 = VA/VP
= R/S
R=h2.S
Parents Parents
sélectionnés
Descendants
La régression parent-enfant et la réponse à une
génération de sélection
tiré de Dillmann et Hospital, 2002
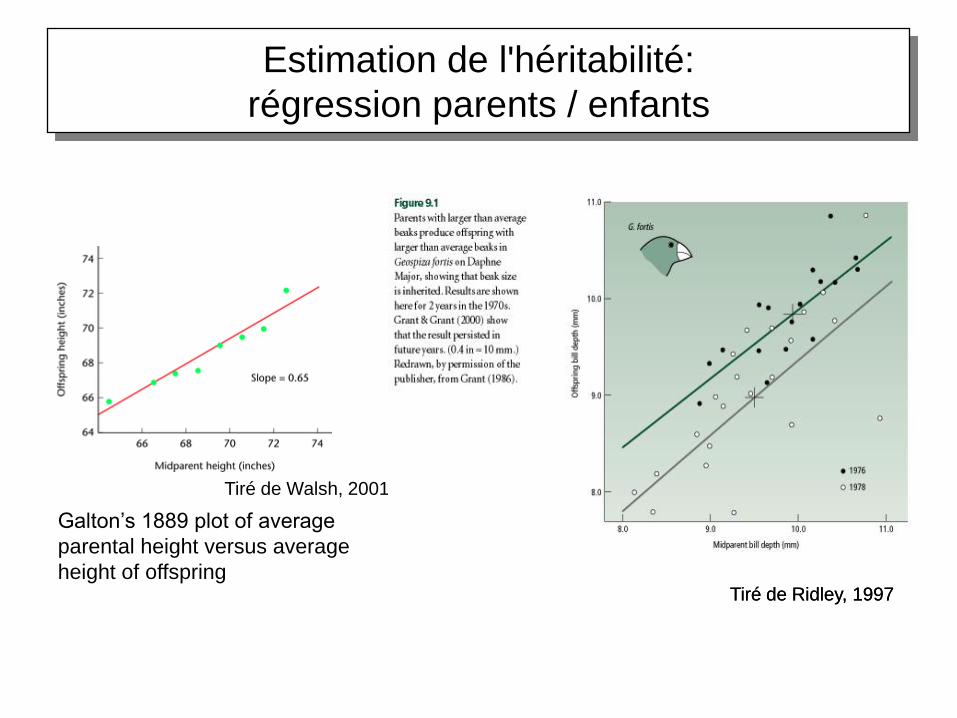
Estimation de l'héritabilité:
régression parents / enfants
Galton’s 1889 plot of average
parental height versus average
height of offspring
Tiré de Walsh, 2001
Tiré de Ridley, 1997Tiré de Ridley, 1997

Héritabilité de la date de ponte chez les Mésanges
bleues
Nuage de Points (Feuille de données1 10v*15c)
Descendants = 14.9142+0.8746*x
110 112 114 116 118 120 122 124 126
Parents
106
108
110
112
114
116
118
120
122
124
126
128
Descendants
Parents:Descendants: r² = 0.8619
h2=0.875: forte héritabilité et donc potentiel de réponse à la sélection élevé
Remarque: Attention aux biais possibles dus à des valeurs aberrantes!
Question: les oiseaux peuvent-ils ajuster génétiquement leur date de ponte en fonction
des changements climatiques attendus? Le cas des mésanges bleues
Tiré de USHERB

La variation quantitative dans les populations

La variation quantitative dans les populations
• Plus de deux allèles en ségrégation pour chaque locus?
• Des fréquences alléliques quelconques
• Régime de reproduction plus ou moins panmictique?
• Déséquilibre de liaison entre allèles dus à des facteurs historiques

Le modèle à un gène:
cas d'une population panmictique
Moyenne et variance d'un caractère dans une population panmictique
Soit p la fréquence de A2 (A+) et q la fréquence de A1 (A-)
Moyenne = p2(a) + 2pq(d) + q2(-a)
= (p2 – q2)a + 2pq(d)
sachant que (p2 – q2) = (p – q) (p + q)
Moyenne = (p – q)a + 2pqd
Var(G) = moyenne des carrés – carré de la moyenne
Var(G) = [p2a2 + 2pqd2 + q2(-a)2] – [(p – q)a + 2pqd]2
= 2pq[a +d(q-p)]2 + [2pqd]2
= Var(A) + Var(D)
Dans le cas d'une F2 où p = q = ½
la moyenne vaut ½ d et
la variance vaut ½ a2 + ¼ d2
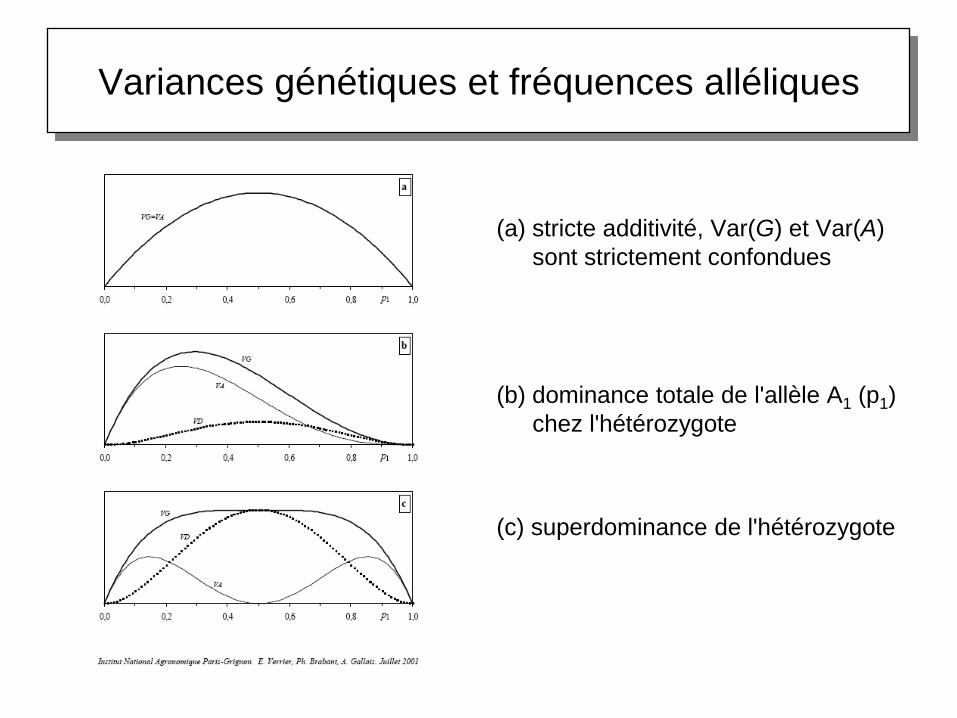
Variances génétiques et fréquences alléliques
(a) stricte additivité, Var(G) et Var(A)
sont strictement confondues
(b) dominance totale de l'allèle A1 (p1)
chez l'hétérozygote
(c) superdominance de l'hétérozygote

L'architecture génétique des caractères complexes
• Combien de gènes impliqués?
• Où sont-ils localisés?
• Quels sont leurs effets

L'architecture génétique des caractères
complexes
L'approche biométrique

La précocité de floraison est sous le contrôle de nombreux gènes, localisés sur
différents chromosomes, avec des interactions épistatiques
Déterminisme génétique de la date d’épiaison
chez le blé
tiré de Dillmann et Hospital, 2002

L'architecture génétique des caractères
complexes
La recherche de QTL

La notion de QTL (Quantitative Trait Locus)
• On appelle QTL, un gène polymorphe dont la variation allélique
influe sur la variation phénotypique d'un caractère quantitatif
• Les avancées de la génomique structurale et fonctionnelle
permettent de les:
– détecter
– cartographier
– caractériser
• On utilise le polymorphisme de marqueurs du génome afin de
détecter et localiser les QTL

• Il faut disposer d’une carte génétique de l’espèce étudiée
• On effectue le génotypage pour des marqueurs bien répartis sur la
carte
• On réalise le phénotypage pour le caractère au sein de familles
recombinantes
• On recherche la liaison génotype au marqueur / phénotype au
caractère quantitatif pour localiser la région qui contient le QTL: il
faut donc plutôt parler de QTR (Quantitative Trait Region)
Principe de la recherche de QTL

Principe de la recherche de QTL:
lignées recombinantes
• On choisit des parents qui diffèrent pour le caractère et on fabrique des lignées recombinantes qui ségrégent à la fois pour le phénotype quantitatif et pour des marqueurs moléculaires répartis dans le génome
• On recherche les associations entre phénotype quantitatif et marqueurs moléculaires
• Quand on détecte une association phénotype quantitatif / génotype pour un marqueur, on a détecté la présence d'un QTL au voisinage du marqueur

Principe de la recherche de QTL
association marqueur / QTL
On utilise la
liaison génétique
entre marqueur et
QTL pour
localiser le QTL

Méthodes de détection de la variation génétique
On utilise de plus en plus des marqueurs génétiques très polymorphes:
• Microsatellites: VNTR (Variable Number of Tandem Repeats)

Méthodes de détection de la variation génétique
• SNPs (Single Nucleotide Polymorphisms): très nombreux dans le
génome (1 site / 100bp), bien répartis dans le génome
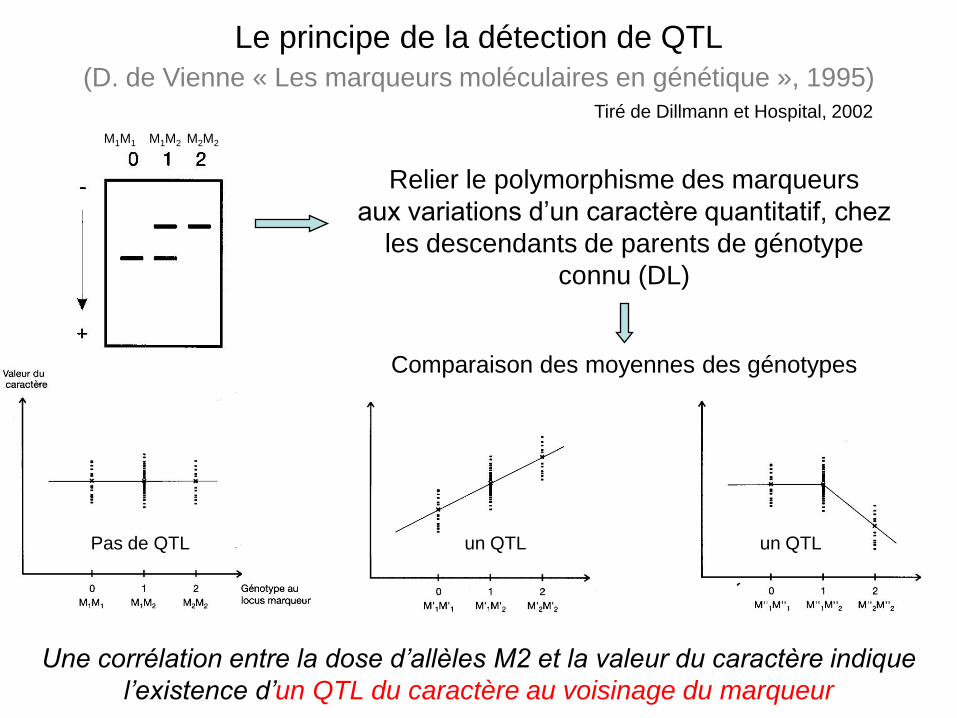
M1M1 M1M2 M2M2
Relier le polymorphisme des marqueurs
aux variations d’un caractère quantitatif, chez
les descendants de parents de génotype
connu (DL)
Une corrélation entre la dose d’allèles M2 et la valeur du caractère indique
l’existence d’un QTL du caractère au voisinage du marqueur
Le principe de la détection de QTL
(D. de Vienne « Les marqueurs moléculaires en génétique », 1995)
Pas de QTL
Comparaison des moyennes des génotypes
un QTL un QTL
Tiré de Dillmann et Hospital, 2002

Principe de la recherche de QTL:
association phénotype - marqueur

Exemples de détection de QTL

QTL de prédisposition à l'obésité chez l'homme
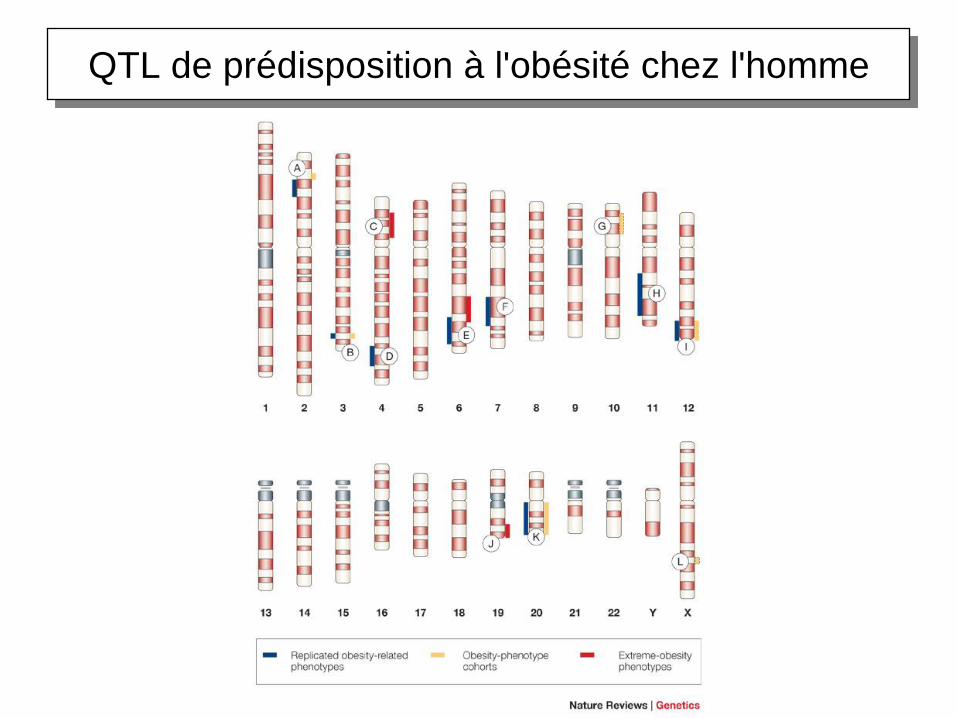
QTL de prédisposition à l'obésité chez l'homme

QTL de prédisposition à la schizophrénie

Des QTL aux gènes: un long chemin semé
d'embûches
• QTL / QTR: Quantitative Trait Loci / Quantitative Trait Régions : on localise des régions chromosomiques plutôt que des gènes
• Ex A. thaliana, 585 cM, 125 Mb, soit 213 kb et ~ 50 gènes par cM
• Clonage positionnel est très fastidieux, et hasardeux
• Approche gène candidat, basée sur la localisation et la fonction supposée de gènes
• Génétique d'association entre polymorphisme nucléotidique et phénotype: SNPs
• Validation expérimentale par transgénèse (RNAi)

The end