lec physio regulate heart contractility
TRANSCRIPT

8/11/2019 Lec Physio Regulate Heart Contractility
http://slidepdf.com/reader/full/lec-physio-regulate-heart-contractility 1/12
R e g u l a t i o n o f a r d i a c M u s c le o n t r a c ti li ty
A R N O L D M . K A T Z
From the Depa rtment o f Physiology, College of Physicians and Surgeons, Columb ia
University, New York. Dr. K atz s present address is the Dep artment o f Medicine,
The University of Chicago
ABSTRACT Th e hear t s phys i o log i ca l per fo rmance , un l i ke t ha t o f ske l e ta l
musc l e , is r egu l a t ed p r i mar i l y b y var ia t i ons i n t he con t r ac t i l e fo rce deve l op ed
by t he i nd i v i dua l m yocard i a l f i ber s. In an a t t em pt t o i den t ify the bas is fo r t he
charac t e r i s ti c p roper t i es o f my ocard i a l con t r ac t i on , t he i nd i v i dua l card i ac con -
t r ac t i l e p ro t e i n s and t he i r behav i o r i n con t r ac t i l e model s i n v i t ro have been
exami ned . Th e l ow sho r ten i ng ve l oc i t y o f hear t musc l e appear s t o r e f lec t t he
w e a k A T P a s e a c t iv i ty o f c a r d ia c m y o s in , b u t t h i s e n z y m a t i c a c t i v i ty p r o b a b l y
does no t de t e rm i ne ac t i ve s t a t e in t ens it y . Qua n t i f i ca t i on o f t he e f f ec ts o f Ca ++
upon card i ac ac t omyos i n suppor t s t he v i ew t ha t myocard i a l con t r ac t i l i t y can
be mod i f i ed by changes i n t he am oun t o f ca l c i um re l eased du r i ng exc i t a t i on -
con t r ac t i on coup l i ng . Ex change o f i n t r ace l lu l a r K + wi t h N a + der i ved f rom t he
ex t r ace l l u l a r space a l so cou l d enhance myocard i a l con t r ac t i l i t y d i r ec t l y , as
h i gh l y pu r i f ied ca rd i ac ac t omyos i n i s s ti mu l a t ed w hen K + is r ep l aced by a n
e q u i m o l a r a m o u n t o f N a + . O n t h e o t h e r h a n d , c a r d i a c g l yc o s id e s a n d c a t e -
cho l ami nes , agen t s wh i ch g rea t l y i ncrease t he c on t r ac t i li t y o f the i n t ac t he ar t ,
were found t o be wi t hou t s i gn i f i can t ac t i ons upon h i gh l y pu r i f i ed r econs t i t u t ed
card i ac ac t omyos i n .
R e g u l a t i o n o f c o n t r a c t i o n i n m a m m a l i a n c a r d i a c a n d s k e le t a l m u s c l e , w h i c h
h a v e m a n y c o m m o n b i o c h e m i c a l a n d m o r p h o l o g i c a l f e a tu r e s , is a c h i e v e d b y
d i f f e r e n t p h y s i o l o g i c a l m e c h a n i s m s . R a p i d t r a in s o f s t i m u l i d e l i v e r e d t o a
s k e le t a l m u s c l e f ib e r th r o u g h i ts m o t o r n e r v e e n h a n c e t e n s io n d e v e l o p m e n t b y
c a u s i n g t h e s u m m a t i o n o f i n d i v i d u a l c o n t r a c t i l e r e s p o n s e s o r , in i ts f u l l y
d e v e l o p e d fo r m , a p o w e r f u l t e ta n i c c o n t r a c t io n . I n t h e h e a r t , o n t h e o t h e r
h a n d , f u s io n o f c o n tr a c t i le r e s p on s e s d o e s n o t o c c u r b e c a u s e t h e r e f r a c t o r y
p e r i o d p e r s is t s a l m o s t t o t h e e n d o f t h e a c t i v e s t a te . R e c r u i t m e n t o f i n c r e a s i n g
n u m b e r s o f a c t i v e m o t o r u n i ts p l a y s a m a j o r r o l e i n a u g m e n t i n g t h e t e n s io n
d e v e l o p e d b y a s k e l e t a l m u s c l e , w h e r e a s a l l f i b e rs o f t h e h e a r t , w h i c h i s a
f u n c t i o n a l s y n c y t i u m , a r e n o r m a l l y a c t i v a t e d d u r i n g e a c h c a r d i a c s y s t o l e .
T h e t e n s i o n g e n e r a t e d b y b o t h c a r d i a c a n d s k e l e t a l m u s c l e f ib e r s i s i n f lu -
e n c e d b y i n it i a l f ib e r l e n g t h ( t h e le n g t h - t e n s i o n r e l a t i o n s h i p o f s k e l e t a l
m u s c l e , a n d S t a r l i n g s l a w o f t h e h e a r t ) , b u t t h e r e l a t i v e l y le s s c o m p l i a n t
m y o c a r d i a l f i b e r a p p e a r s t o o p e r a t e w i t h s h o r te r s a r c o m e r e s t h a n d o e s t h a t
x85
The Journal of General Physiology
Published July 1, 1967

8/11/2019 Lec Physio Regulate Heart Contractility
http://slidepdf.com/reader/full/lec-physio-regulate-heart-contractility 2/12

8/11/2019 Lec Physio Regulate Heart Contractility
http://slidepdf.com/reader/full/lec-physio-regulate-heart-contractility 3/12
A RN OLD M . K A TZ Regulation of ardiac Muscle ontractility
I 8 7
Tropomyosin
No dif ferences between the phys iochemical proper t ies and amino acid com-
pos it ions of cardiac a nd skeletal t ropomyosins t ropomyosin B) have bee n
found, and there is no evidence that the hear t contains s ignif icant amounts
of t ropomyosin A paramyosin) 22). Ma mm ali an skeletal t ropomyosin has
a dual act ion upon the Mg++-act ivated recons t i tuted actomyosin made f rom
skeletal musc le 23). At low ionic strengths tropo myo sin stimulates the M g ++-
act ivated ATPase act ivi ty and abbreviates the clear ing phase that precedes
superprecipitation, whereas at high ionic strengths this protein inhibits this
ATP ase act ivity and delays superprecipi ta t ion 23). These pheno me na have
not been extens ively s tudied in the case of the cardiac contract i le proteins ,
T BLE I
C O M P A R I S O N O F T H E A T P A S E A C T I V I T I E S O F R A B B I T
R E D S K E L E T A L , W H I T E S K E L E T A L , A N D C A R D I A C M Y O S I N S
TPase ctivity
Cation White skeletal Red skeletal Cardiac
btmole P¢ min mg ~nole Pi min mg IJtnole Pi min mg
2 . 0 m M M g + + 0 . 0 0 9 7 4 0 . 0 0 0 7 * 0 . 0 0 2 3 4 0 . 0 0 0 5 0 . 0 0 2 4 4 0 . 0 0 0 4
1 .0 r n~ Ca ++ 0 .600 - + - 0 .038 O . 1304 - 0 .00 5 O . 1064 - 0 .016
R e a c t i o n s w e r e c a r r i e d o u t a t 2 5 ° C i n 1 0 r r ~ A T P , 0 . 1 M K C 1 , a n d 1 .0 mM T r i s a c e t a t e a t p H 6 . 8.
F r o m K a t z e t a l . ( 21 ) .
* S t a n d a r d e r r o r o f t h e m e a n .
but prel iminary f indings indicate that cardiac t ropomyosin, l ike i t s skeletal
homo logue, st imulates actomyosin at low ionic s t rengths Katz, A. M. U n-
published observations).
yosin
The over-al l molecular dimens ions of cardiac and whi te skeletal myos ins are
s imilar 24-26) , as are several solvent- induced changes in secondary and
ter t iary s t ructure 27). On the other hand, di f ferences have been found in
cysteine con ten t 20), sensit ivit ies to proteoly tic enzym es 28, 29), im mu no -
logical speci f ic i ty 30, 31) , and th e amin o acid sequence aroun d on e of the
cysteine residues 32).
The lower ATPase ac t iv i ty o f r abb i t ca rd iac myos in , compared to tha t o f
rab bit skeletal myosin, was f irst repo rted in 1942 by Bailey 33), bu t the
myosins examined in this pioneer ing s tudy contained unknown amounts of
act in , which was then an unrecognized protein . Subsequent compar isons of
more highly pur i f ied rab bi t skeletal myos ins wi th m yosins obtained f rom the
hear ts of larger mamm als dogs , cat t le , sheep) conf i rmed the lower ATP ase
Published July 1, 1967

8/11/2019 Lec Physio Regulate Heart Contractility
http://slidepdf.com/reader/full/lec-physio-regulate-heart-contractility 4/12
88
COMPARATIVE ASPECTS OF MUSCULAR CONTRACTION
act iv i t ies of the la t ter . T he poss ibi li ty tha t these f indings repres ented a species
d i f f e rence , r a the r th an a t rue d i f f e rence be tween c a rd iac a nd ske le ta l m yos ins ,
was r a i s ed by the f ind ing tha t dog ske le ta l m yos in had on ly s l igh t ly h igher
AT Pase ac t iv i ty than dog ca rd iac m yos in (26) . However , a ca re fu l s tudy o f
h igh ly pur i f i ed r abb i t ca rd iac and ske le ta l m yos ins (20) c onf i rm ed Ba i ley s
ea r l i e r f ind ing. T h is d i s c repanc y was reso lved by the f ind ing tha t the AT Pas e ac -
t iv i ty of rab bi t red skeletal myosin was less tha n t hat of white skeletal myos in
(34 , 35 ) , a conc lus ion sugges ted independen t ly by h i s tochem ica l s tud ies o f
these m usc le types in m an (36) , and by the obse rva t ion tha t dog ske le ta l
m usc le i s com po sed p r inc ipa l ly o f r ed f ibe rs . E xam ina t io n o f m yos ins pur i f i ed
from red skeletal , white skeletal , and cardiac muscle , a l l f rom the rabbi t ,
r evea led the AT P ase ac t iv i ti e s o f r ed ske le tal and ca rd iac m yos ins to be
s imilar to each other , bu t less than that of white skeletal myo sin (T able I ) .
TABLE I I
CALCIUM S ENS ITIVITIES OF THE IN ITIAL
M g -ACTIVATED ATP AS E ACTIVITY OF
R E C O N S T I T U T E D C A R D I A C A C T O M Y O S I N S
ATPase activity
Actomyosin made with 0.1 mu CaCl2 0.25 mu EGTA
Tropomyosin-free actin
Actin-tropomyosin complex
mote P~/min/mg ~nole Pg/min/mg
0.0077 0.0066
0.0142 0.0081
Results obtained with 0.5 mg/ml myosin and either 0.125 mg/ml tropomyosin-free actin or
0.167 mg/ ml of the ac tin- tropo myos in compl ex in 1.0 mM MgA TP, 0.10 M KC1, and 20 mM
histidine, pH 6.8, at 25°C. The ATPase activity of the myosin alone in the presence of EGTA
was 0.0039 t tmole Pi /min /mg.
Com par i son o f the dyn am ic cons tan t s o f r ed a nd whi te ske le ta l m usc le
func t ion sugges ts tha t m yos in AT P ase ac t iv i ty m ay de te rm ine Vm~x, the
m ax im al shor ten ing ve loc i ty o f the un loaded m usc le . In r ed ske le ta l m usc le
Vm~ is app rox ima tely 1~ tha t of white skeletal muscle (37, 38), a ra t io s imilar
to th a t s een be tween th e m yos in AT P ase ac t iv i t ie s o f these two m usc le types
(21 , 34) . On the o the r ha nd , i t i s un l ike ly tha t the A T Pas e ac t iv i ty o f the
con t r ac t i l e p ro te ins in f luences P0, the m ax im al fo rce -gene ra t ing ab i l ity o f the
muscle , becau se the te tanic tens ions per u ni t of cross-sect ional area of red
and white skeletal muscle do not d if fer s ignif icant ly (38) .
W hi le p rec i s e quan t i f i ca t ion o f the dynam ic cons tan t s o f ca rd iac m usc le
func t ion i s d i f f i cu l t (39 ) , bo th the shor ten ing ve loc i ty and fo rce -genera t ing
capac i ty o f the m y oca rd iu m are le ss than in whi te ske le ta l m usc le (8) . T he
la rge m i to chon dr ia l co n ten t o f hea r t m usc le (40) , wh ich s ign i fi can t ly r educes
the f r ac t ion o f c ross - sec t iona l a r ea oc cup ied by the m yo f i l am en ts , appear s
to be pa r t ly r espons ib le fo r the low tens ion genera ted by the m yocard ium .
Published July 1, 1967
8/11/2019 Lec Physio Regulate Heart Contractility
http://slidepdf.com/reader/full/lec-physio-regulate-heart-contractility 5/12
A R N O ~ M . K A T Z
Regulation of Cardiac Muscle Contractility
189
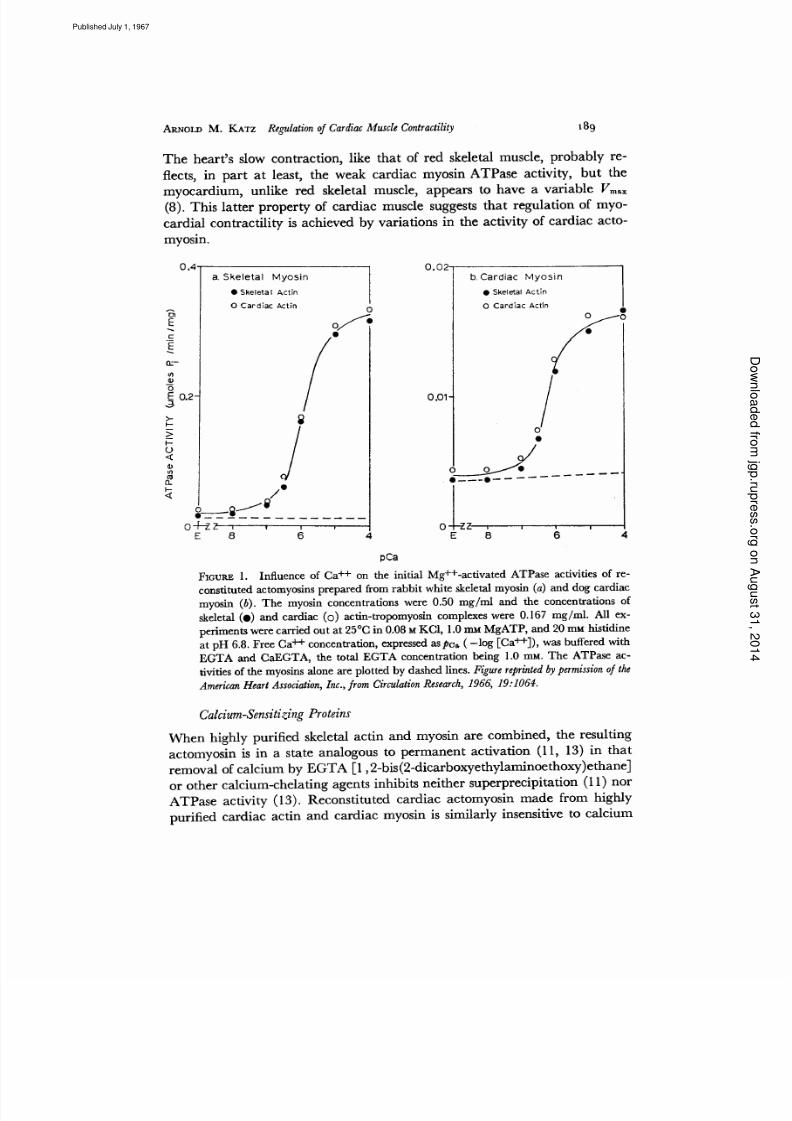
T h e h e a r t s s l o w c o n t r a c t i o n , l i k e t h a t o f r e d s k e l e t a l m u s c l e , p r o b a b l y r e -
f l e e ts , i n p a r t a t l e a s t, t h e w e a k c a r d i a c m y o s i n A T P a s e a c t i v i t y , b u t t h e
m y o c a r d i u m , u n l i k e r e d s k e l e t a l m u s c l e , a p p e a r s t o h a v e a v a r i a b l e V m ~x
( 8 ). T h i s l a t te r p r o p e r ty o f c a r d i a c m u s c l e s u g ge s ts t h a t r e g u l a t i o n o f m y o -
c a r d i a l c o n t r a c t i l i t y i s a c h i e v e d b y v a r i a t i o n s i n t h e a c t i v i t y o f c a r d i a c a c t o -
m y o s i n .
6
E
E
n .
E 0 . 2 -
..B-
> -
I - -
I .-
L~
.<
g_
I
0 , 4 0 , 0 2 -
a . S k e l e t a l M y o s i n
• S k e l e t a l Act~n
Cardiac AcUn
/
/ •
O - ~ - Z 7 , ,
E 4
0 o 0 1 -
b . C a r d i a c M y o s i n
• Skeletal Ac tin
0 Card iac Ac t in
o
E g 4
p C a
F IG U RE 1. I n f l u e n c e o f C a + + o n t h e i n i t i a l M g + + - a c t i v a t e d A T P a s e a c t i v i t i e s o f r e -
c o n s t i t u t e d a c to m y o s i n s p r e p a r e d f r o m r a b b i t w h i t e sk e l e ta l m y o s i n a ) a n d d o g c a r d i a c
m y o s i n b ) . T h e m y o s i n c o n c e n t r a t io n s w e r e 0 . 50 m g / m l a n d t h e c o n c e n t r a t io n s o f
s k e le t a l O ) a n d c a r d i a c o ) a c t i n - t r o p o m y o s i n c o m p l e x e s w e r e 0 . 16 7 m g / m l . A l l e x-
p e r i m e n t s w e r e c a r r i e d o u t a t 2 5 °1 2 i n 0 . 0 8 M K C I , 1 .0 m _~ M g A T P , a n d 2 0 m M h i s t i d i n e
a t p H 6 .8 . F r e e C a + + c o n c e n t r a ti o n , e xp r e s se d a s p e a - l o g [ C a + + ] ) , w a s b u f f e re d w i t h
E G T A a n d 1 2 a E G T A , t h e t o t a l E G T A c o n c e n t r a t i o n b e in g 1 .0 r a M . T h e A T P a s e a c -
t i v i ti e s o f t h e m y o s i n s a l o n e a r e p l o t t e d b y d a s h e d l i n e s. Figu re reprinted by permission of the
American Heart Association Inc. from Circulation Research 1 966 19:1064.
Calcium Sensitizing Proteins
W h e n h i g h l y p u r i f i e d s k e l e t a l a c t i n a n d m y o s i n a r e c o m b i n e d , t h e r e s u l t i n g
ac t om yos i n i s i n a s t a t e ana l ogou s to pe rm an en t ac t i va t i on 11, 13 ) i n t ha t
r e m o v a l o f c a l c iu m b y E G T A [ 1 , 2 -b i s 2 - d i c a rb o x y e t h y la m i n o e t h o x y )e t h a n e ]
o r o t h e r c a l c i u m - c h e l a t i n g a g e n t s in h i b i ts n e i t h e r s u p e r p r e c i p i ta t i o n 1 1) n o r
A T P a s e a c t i v i ty 1 3 ). R e c o n s t i t u t e d c a r d i a c a c t o m y o s i n m a d e f r o m h i g h l y
p u r i f ie d c a r d i a c a c t i n a n d c a r d i a c m y o s i n i s s i m i l a rl y in s e n si ti v e to c a l c i u m
Published July 1, 1967

8/11/2019 Lec Physio Regulate Heart Contractility
http://slidepdf.com/reader/full/lec-physio-regulate-heart-contractility 6/12
I 9 o
C O M P R T I V E S P E C T S O F M U S C U L R C O N T R G T I O N
r e m o v a l ( T a b l e I I ) . C a l c i u m s e ns i ti v it y c a n b e r e s t o re d t o b o t h c a r d i a c a n d
s k el e ta l a c t o m y o s i n b y n a t i v e t r o p o m y o s i n ( 41 ), o r b y p r e p a r i n g t h e a c t i n
u n d e r c o n d i t i o n s t h a t c a u s e a t r o p o m y o s i n - c o n t a i n i n g p r o t e i n c o m p l e x t o b e
e x t r a c t e d f r o m t h e a c e t o n e - d r i e d m u s c l e p o w d e r a l o n g w i t h t h e a c t i n ( T a b l e
I I ) ( 13 , 42 , 4 3 ). T h i s p r o t e i n c o m p l e x , i s o l at e d b y v i r t u e o f i ts b i n d i n g t o
F - a c t i n , i n h i b i t s t h e a c t i v a t i o n o f m y o s i n b y a c t i n a t l o w f r e e C a + + c o n c e n t r a -
t i o n s ( 1 3 ). T h e C a + + s e n s it iv i ti e s c o n f e r r e d b y t h e t r o p o m y o s i n - c o n t a i n i n g
p r o t e i n c o m p l e x e s e x t r a c t e d f r o m c a r d i a c a n d w h i t e s k e le t al m u s c l e s a r e t h e
same (42) (F ig . 1 ) .
C A R D I A C A C T O M Y O S I N
T h e m e c h a n i s m s r es p o n s ib l e fo r t h e c o n t r o l o f m y o c a r d i a l c o n t r a c t il i ty c a n b e
s t u d i e d i n h i g h l y p u r i f i e d c a r d i a c a c t o m y o s i n s y s t e m s , w h i c h p r e s e r v e m a n y
T B L E I I I
C O M P A R I S O N O F T H E M Y O C A R D I A L C A L C I U M U P T A K E D U R I N G
A P O S I T I V E R A T E S T A I R C A S E A N D T H E C A L C I U M R E Q U I R E D
T O P R O D U C E A S IM I L A R I N C R E A S E I N C A R D I A C
A C T O M Y O S I N A T P A S E A C T I V I T Y
M y o s i n c o n t e n t o f th e h e a r t
S e n s i t iv i t y o f a c t o m y o s i n A T P a s e t o c h a n g i n g
l e v el o f b o u n d c a l c i u m
E s t i m a t e d c h a n g e i n t o t a l m y o c a r d i a l c a l c i u m
n e e d e d t o e f f e c t a 4 0 i n c r e a s e in a c t o m y o s i n
A T P a s e
C a l c i u m u p t a k e a c c o m p a n y i n g 4 0 c o n t r a c -
t i l i t y i n c r e a s e
5 0 -8 0 ~ m o l e s / k g w e t w e i g h t
A n i n c r ea s e o f 0 .3 m o l e c a l c i u m p e r m o l e o f
m y o s i n e ff e c ts a 4 0 i n c r e a s e i n a c t i v i t y
1 5 - 2 4/ ~ m o l e s/ k g w e t w e i g h t
2 0 t ~ m o l e s / k g w e t w e i g h t
f e a t u r e s o f t h e c o n t r a c t i l e a p p a r a t u s o f t h e in t a c t h e a r t . S u c h a c t o m y o s i n s
c a n b e m a d e t o s u p e r p r e c i p i t a t e , a n i n v i t ro f o r m o f c o n t r a c t i o n ( 4 4 ) ; t h e y
h y d r o l y z e A T P , w h i c h a p p e a r s t o p r o v i d e t h e i m m e d i a t e s o u r ce o f e n e r g y
f o r m u s c u l a r c o n t r a c t i o n ( 4 5 ) ; a n d t h e i r a c t i v i t y i s m o d i f i e d b y l o w c o n c e n -
t r a ti o n s o f c a l c i u m i o n i n a m a n n e r t h a t a p p e a r s a n a l o g o u s t o t h e p h y s i o lo g i -
c a l r o l e o f t h i s i o n i n e f f e c ti n g e x c i t a t i o n - c o n t r a c t i o n c o u p l i n g ( 42 , 4 3 , 4 6 ) .
B e c a u s e c h a n g e s i n m y o c a r d i a l c o n t r a c t i l i t y a r e p a r a l l e l e d b y c h a n g e s i n
V max, a n d b e c a u s e a d i r e c t r e l a t i o n s h i p b e t w e e n V m ,x a n d t h e A T P a s e a c t i v i t y
o f t h e c o n t r a c t i l e p r o t e i n s p r o b a b l y e x i st s ( 47 ) , e n h a n c e d c a r d i a c a c t o m y o s i n
A T P a s e a c t i v i t y i n v i t r o m i g h t b e a n a l o g o u s t o t h e a c c e l e r a t e d s h o r t e n i n g
v e l o c i t y t h a t a c c o m p a n i e s i n c r e a s e d c o n t r a c t i li t y i n v i v o. T w o i o n i c m e c h a -
n i s m s t h a t c o u l d b r i n g a b o u t s u c h c h a n g e s in t h e i n t a c t h e a r t h a v e b e e n
id e n t i f i e d .
V a r i a t io n s i n t h e a m o u n t o f c a l c i u m r el e as e d d u r i n g e x c i t a t io n - c o n t r a c t io n
Published July 1, 1967
8/11/2019 Lec Physio Regulate Heart Contractility
http://slidepdf.com/reader/full/lec-physio-regulate-heart-contractility 7/12
ARNO~ M KATZ
Regulat ion of ardiac Muscle ontractility
[ [
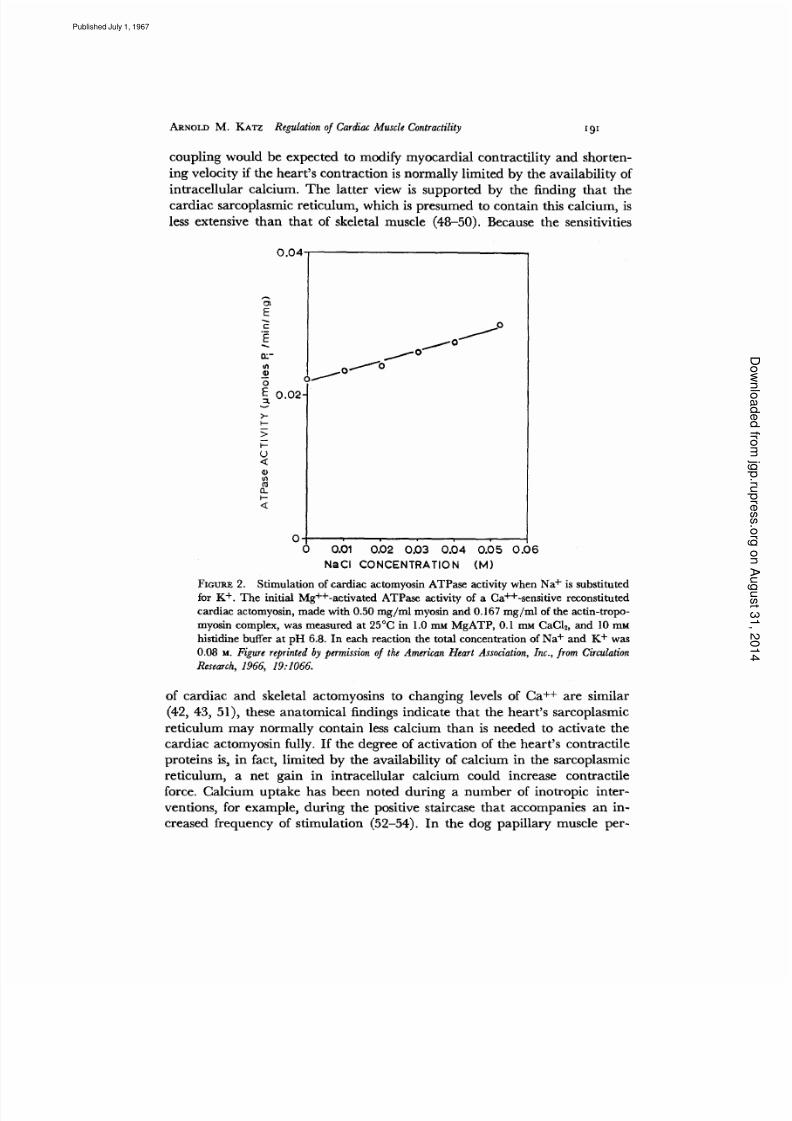
coupl ing would be expected to modify myocardial contract i l i ty and shor ten-
ing veloci ty i f the hea r t s con tract ion is normal ly l imi ted by the avai labi l i ty of
int racel lular calcium. The la t ter view is suppor ted by the f inding that the
cardiac sarcoplasmic ret iculum, w hich is presume d to con tain this calcium, is
less extensive tha n tha t of skeletal muscle (48-50). Because the sensit ivit ies
0 . 0 4
S
E
£=
n ' -
t/I
0
E
- i
v
>-
I - -
I-.-
<
Q_
I,-,-
<
0 . 0 2 -
/ o /O / °
/ o / °
o o ol 0;o2 o b3 o 64 o bs 0 o6
N a C , C O N C E N T R A T I O N ( M )
FIoum~ 2. Stimulation of cardiac actomyosin ATPase activity when Na is substituted
for K . The initial Mg -activated ATPase activity of a Ca -sensitive reconstituted
cardiac actomyosin, made with 0.50 mg/ ml myosin and 0.167 mg /ml of the actin-tropo-
myosin complex, was measured at 25°C in 1.0 rr~ MgATP, 0.1 mM CaCI~, and 10 rnM
histidine buffer at pH 6.8. In each reaction the total concentration of Na and K was
0.08 M. Figure reprinted by permission of the American Heart Association Inc. from Circulation
Research 1966 19:1066.
of cardiac and skeletal actomyosins to ch angin g levels of Ca ++ are s imi lar
(42, 43, 51), these ana tom ical f indings indicate th at t he hea rt s sarcop lasmic
ret iculum may normal ly contain less calcium than is needed to act ivate the
cardiac actomyosin fully . I f the degree of act ivat ion of the hear t s contract i le
proteins is, in fact , l imi ted by the avai labi l i ty of calcium in the sarcoplasmic
ret iculum, a net gain in int racel lular calcium could increase contract i le
force. Calcium uptake has been noted dur ing a number of inotropic inter -
vent ions , for example, dur ing the pos i t ive s ta i rcase that accompanies an in-
creased f requency of s t imulat ion (52-54) . In the dog pa pi l lary muscle per -
Published July 1, 1967
8/11/2019 Lec Physio Regulate Heart Contractility
http://slidepdf.com/reader/full/lec-physio-regulate-heart-contractility 8/12
8/11/2019 Lec Physio Regulate Heart Contractility
http://slidepdf.com/reader/full/lec-physio-regulate-heart-contractility 9/12
A~Notz~ :M. KATZ
Regulat ion of ardiac Muscle ontractility
I93
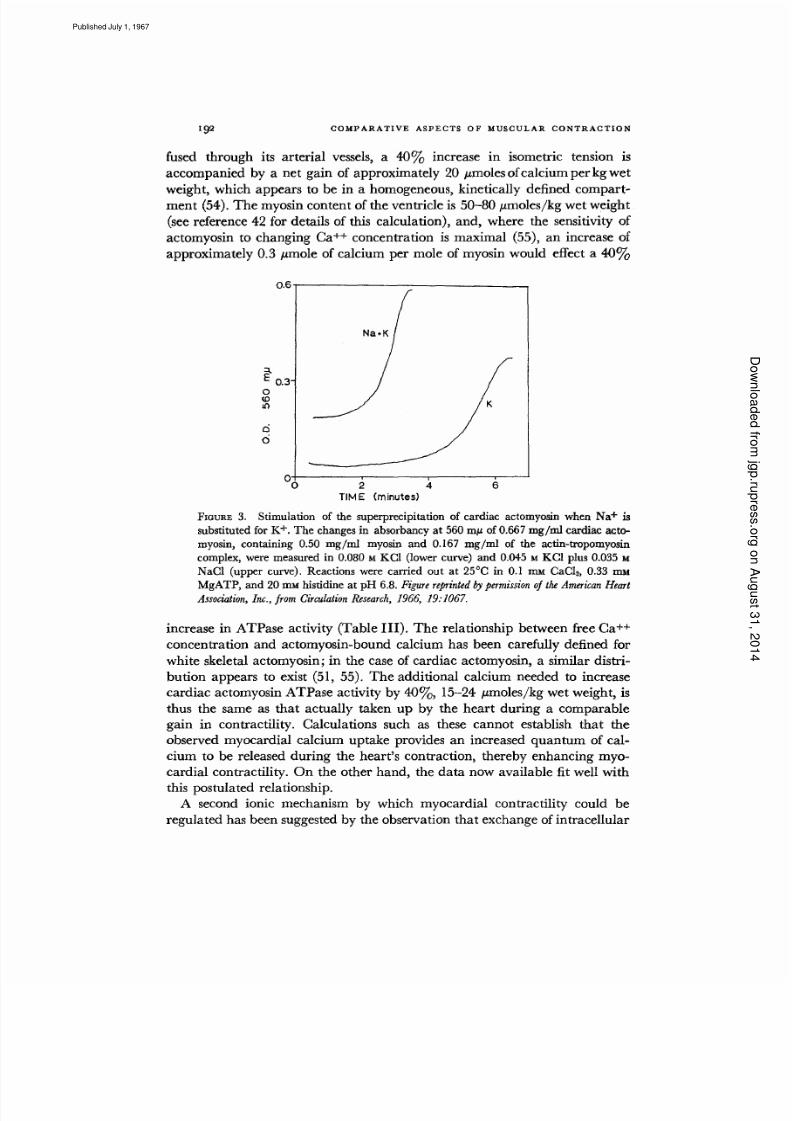
K÷ w ith extracellular N a + accompanie s a var iety of interventions which in-
crease myocardial contractility (56-59). The possibility that the cardiac myo-
fibril is more active in the presence of Na + than K + is supported by the find-
ing that both ATPase activity (Fig. 2) and superprecipitation (Fig. 3) of
highly purified cardiac actomyosin are stimulated when K + is replaced by an
equim olar am ou nt of Na + (42). T he relatively small effects of Na+-K + ex-
change on act omyosin are seen only when large quantit ies of Na + are sub-
stituted for K +, however, so tha t the physiological significance of this phe -
nomenon remains uncertain.
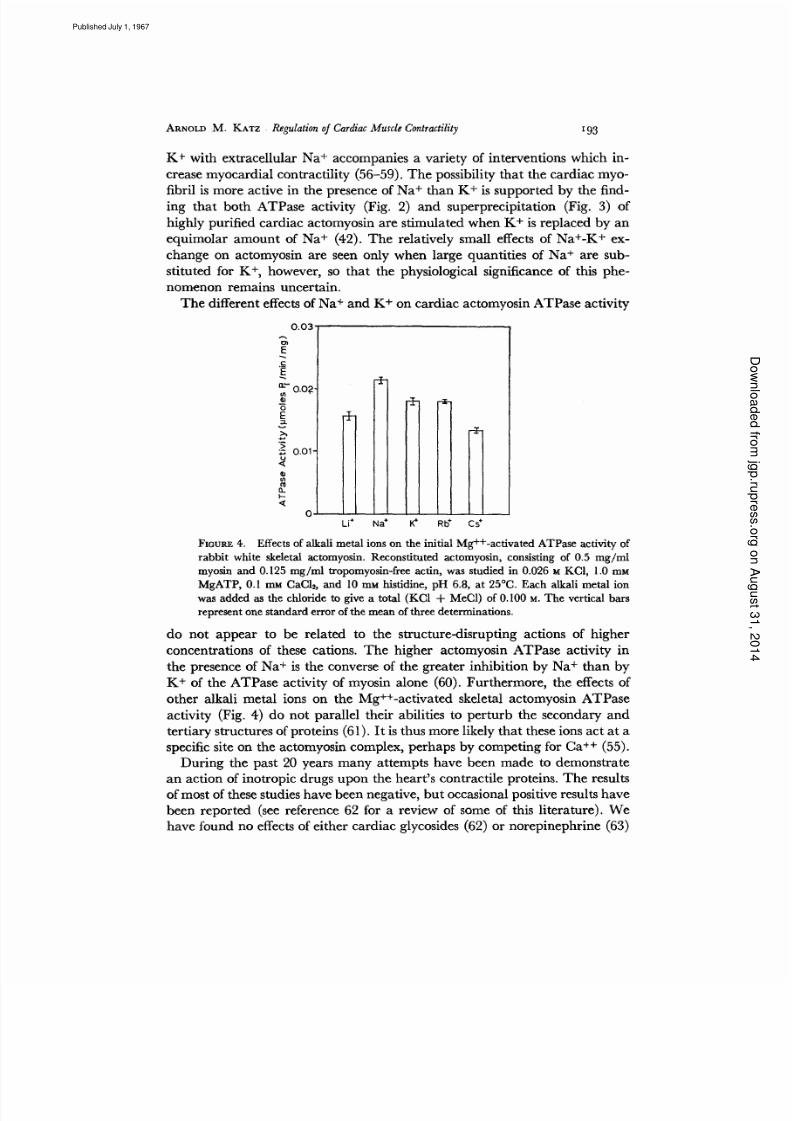
The different effects of Na ÷ and K+ on cardiac actomyosin ATPase activity
0 . 0 3
c
~
o.o~-
G
E
=L
. ~ 0 . 0 1 -
ii1
o
t
÷
L i ÷ Na ÷ I (
b÷ C s +
FXOURE 4. Effectsof alkali metal ions on the initial Mg++-activated ATPase activity of
rabbit white skeletal actomyosin. Reconstituted actomyosin, consisting of 0,5 mg/ml
myosin and 0.125 mg/ml tropomyosin-freeact.in, was studied in 0.026 ~t KCI, 1.0 mM
MgATP, 0.I mM CaCI~, and 10 ram histidine, pH 6.8, at 25°C. Each alkali metal ion
was added as the chloride to give a total (KC1 + MeC1) of 0.I00 M. The vertical bars
represent one standard error of the mean of three determinations.
do not app ear to be related to the structure-disru pting actions of higher
concentra tions of these cations. The higher actomyo sin ATPas e activ ity in
the presence of Na + is the converse of the grea ter inhibitio n by Na + than by
K + of the ATPas e a ctivity of myosin alone (60). Furt herm ore, the effects of
other alkali metal ions on the Mg++-activat ed skeletal actomyo sin ATP ase
activity (Fig. 4) do not parallel their abilities to perturb the secondary and
terti ary structures of proteins (61). It is thus more likely that these ions act at a
specific site on the actomyosin complex, perhaps by compet ing for C a ++ (55).
During the past 20 years many attempts have been made to demonstrate
an action of inotropic drugs up on the heart s contractile proteins. The results
of most of these studies have been negative, b ut occasional positive results have
been reported (see reference 62 for a review of some of this literature). We
have found no effects of either car diac glycosides (62) or nor epine phrine (63)
Published July 1, 1967

8/11/2019 Lec Physio Regulate Heart Contractility
http://slidepdf.com/reader/full/lec-physio-regulate-heart-contractility 10/12
x94
C OM P R T IVE S P E C T S OF M US C UL R C ONT R C T ION
upon the highly purified, calcium-sensitive, reconstituted actomyosin. While
these negative findings do not exclude such a direct action of these drugs,
they are in accord with physiological evidence suggesting that these agents
act indirectly upon the contracti le proteins. Most significant is the finding
that, after maxim al contracti l i ty is achieved by any of a num ber of agents or
ionic manipulations, additional inotropic interventions of a different class
bring about no further enha ncem ent of myocard ial contracti li ty 64).
Miss Doris I. Repke, M iss Barbara R . Cohen, Mrs. Ka thleen D. Clarke, Mrs. B onnie B. Rubin,
and Miss Anne Sand ers participated in these studies.
This investigation was supported by Research Grants HE-085 15 and HE-05741 from the U.S.
Public Health Service and 65-G-61 from the American Heart Association.
The author is an Established Investigator of the American Heart Association.
R E F E R E N C E S
1. SPOTmTZ, H. M., E. H. SONNF.NBLICK,an d D. SPIRO. 1966.
Circulation Res. 18:49
2. G oRDo n, A. M . , A . F . HtrxL~.y, and F . J . JULIAN. 1966. J. Physiol., London).
184:170.
3. BURCH, G. E ., C. T . RAY an d J. A. CRONVICH. 1952. Circulation. 5:504.
4. BURTON, A. 1957. Am. Heart J. 54:801.
5. KATZ, A. M. 1965. Circulation. 32:871.
6. WIn tERS, C. J. 1927. a . Pharmacol. Exp tl. Therap. 30:217 .
7. ABBOTT, B. C., an d W . F. H . M . MOMMAERTS. 1959.
J. Gen. Physiol.
42:533 .
8. SONN~.NBLmK,E. H. 1962. Federation Proc. 21:975.
9. SANDOW, A. 1964.
Arch. Phys. Med. Rehabit.
45:62 .
10. SANDOW, A. 1965. Pharmaol. Rev. 17:265.
11. EBb,sin, S., and F. EB ASaL 1964. O. Biochem. Tokyo). 55:604 .
12. Mtr~LX.~R, H. 1966. Biochem. Z. 345:300 .
13. K ATZ, A. M. 1966.
J. Biol. Chem.
241:1522.
13 a. EBASHI, S., an d A. KODAMA. 1966.
J. Biochem. Tokyo ).
60:733.
14. AZUMA, N., an d S. WATANABE. 1965. o . Biol. Chem. 240:3852 .
15. PERRY, S~ V., V. DAvis, a nd D. HA , TaR. 1966.
Biochem. J.
99:1c.
16. KATZ, A. M ., a nd E. J. HALL. 1963. Circulation Res. 13:187.
17. KATZ, A. M ., an d M . E. CARSTEN. 1963. Circulation Res. 13:474.
18. KATZ, A. M ., an d J. B. MAXWELL. 1964. Circulation Res. 14:345.
19. KA TZ, A. M . 1963. Biochim. Biophys. Acta. 71:397.
20. BXRAN,, , M ., E. GAETJENS, K . BARXNY, an d E. KARP. 1964. Arch. Biochem. Bio-
phys. 106:280.
21. KATZ, A. M ., D. I. R~.Pr,~, and B. B. Rum N. 1966. Circulation Res. 19:611.
22. KATZ, A. M ., a n d R . P . CONVaRS~.. 1964.
Circulation Res.
15:194.
23. KATZ, A. M. 1964. J. Biol. Chem. 239:3304.
24. GERa~.LY, J . , M . A. GOUWA, and H. KO H~ R. 1956.
Circulation.
14:940.
25. DA vis, J. O. , W . R. CARROLL, M . TRAPASSO, an d N. A. YANI,:OPOULOS. 1960.
J. Clin. Invest. 39:1463.
26. MUELLER, H., J . FRANZEN, R. V. RmF., an d R. E. OLSON. 1964. O . Biol. Chem.
239:1447.
Published July 1, 1967

8/11/2019 Lec Physio Regulate Heart Contractility
http://slidepdf.com/reader/full/lec-physio-regulate-heart-contractility 11/12
ARNOLD M. KATZ
Regulationof Cardiac Muscle Contractility
~95
27. KAY
C. M., W. A. GREEN, and K. OlKAWA. 1964.
Arch. Biochem. Biophys.
108:89.
28. GF.ROELY,J. 1959.
Ann. N. Y. Acad . Sci.
72:538.
29. MtmLLER,H., M. TrmmEl~, and R. E. OtSON. 1964. J. Biol. Chem. 239:2153.
30. EBERT,J. D. 1953.
Proc. Nat l. A cad. Sci. U. S.
39:333.
31. FXNCK,H. 1965.
Biochim. Biophys. Acta.
111:231.
32. STRACm~.R,A. 1965.
In
Muscle. W. M. Paul, E. E. Daniel, C. M. Kay, and G.
Monckton, editors. Pergamon Press, Oxford. 85.
33. BAILEYK. 1942.
Biochem. J.
36:121.
34. SmDET., J. C., F. A. SPatTER, M. M. THOMPSON, and J . GERGEL Z. 1964.
Biochem.
Biophys. Res. Commun.
17:662.
35. BARANy, M., K. B~R~N¥, T. R~CKARD, and A. VOLPE. 1965.
Arc h. Biochem.
Biophys.
109:185.
36. ENOS, W. K. 1962.
Neurology.
12:778.
37. CLOSE, R. 1964.
J. Physiol., London).
173:74.
38. WEms, J. B. 1965.
J. Physiol., London).
178:252.
39. BRAD (, A. J. 1966. J. Physiol., London). 184:560.
40. STENOER, R. J ., and D. SPmo. 1961.
Am. J. Med.
30:653.
41. EBASHI S. H. IWAKURA H. NAKAJIMA R. NAKAMURA and Y. Ool. 1966.
Biochem. Z.
345:201.
42. KATZ, A. M., D. I. t~r , rd~, and B. R. Com~N. 1966.
Circulation Res.
19:1062.
43. KATZ, A. M. , and D. I. REPm~. 1966.
Science.
152:1242.
44. SZF.NT-GY6RO,n, A. 1947. Chemistry of Muscular Contraction. Academic Press,
Inc., New York.
45. Im~AnTE,A. A., and R. E. DAVmS. 1965.
J. Biol. Chem.
240:3996.
46. FANBURC,B., R. M. FINm~L, and A. MARTONOSI. 1964.
J. Biol. Chem.
239:2298.
47. BARAN , ,M. 1967.
J. Gen. Physiol.
50 (6, Pt. 2): 197.
48. PORTER, K. R., and G. E. PALAD~.. 1957.
J. Biophys. Biochem. Cytol.
3:269.
49. FAWCETT, D. W. 1961.
Circulation.
24:236.
50. NELSON, P. A., and E. S. BF.NSON. 1963. J. Cell Biol. 16:297.
51. FANBURC, B. 1964.
Federation Proc.
23:922.
52. NmDEROF.Rrd~, R. 1962.
J. Physiol., London).
167:515.
53. WINECRAD,S., and A. SHANES. 1962.
J. Gen. Physiol.
45:371.
54. LANCe.R, G. A. 1965.
Circulation Res.
17:78.
55. WP.BgR, A., and R. H~gz. 1963.
J. Biol. Chem.
238:599.
56. HAJDU, S. 1953.
Am. J. Physiol.
174:371.
57. Koch-WEst.g, J., and J. R. BLINKS. 1963.
Pharmacol. Rev.
15:601.
58. SAR~OFF, S. J., J. P. Gi~.Mom~,J. H. MITCH~.LL,and J . P. REM~.NS~ ~Sg. 1963.
Am. J. Med.
34:440.
59. LAN~ER, G. A., and A. J. BRADY. 1966.
Circulation Res.
18:164.
60. WARPdSN,J. C., L. STOW~tNO, and M. F. MOgA~ES. 1966.
J. Biol. Chem.
241:309.
61. VON HmP~L, P. H., and K . Y. WONO. 1964. Science. 145:577.
62. KATZ, A. M. 1966.
J. Pharmacol. Exptl. Therap.
154:558.
63. KATZ, A. M. 1967.
Am. J. Physiol.
212:39.
64. KAVA~R, F., V. J. FIs~Eg, and J . H. S~JCK~.¥. 1965.
Bull. N. Y. Acad. Med.
41:592.
Published July 1, 1967

8/11/2019 Lec Physio Regulate Heart Contractility
http://slidepdf.com/reader/full/lec-physio-regulate-heart-contractility 12/12
96
C O M P R T I V E S P E C T S O F M U S C U L R C O N T R C T I O N
iscussion
Dr Sandow:
I t h ink , Dr . Ka tz , t ha t you have shor t -changed ske le t a l musc le in
s t a t ing tha t i t has l i t t l e capac i ty for hav ing i t s cont rac t i l i t y a l t e red . I t depends on
wha t you do w i th i t . For examp le , t he z inc ion , which has a depres s ive e f fec t on h ea r t
musc le , can grea t ly pro long the ac t ive s ta t e of ske le ta l musc le an d so cause i t t o
produce a much b igge r tw i t ch ou tput .
Now , this di fference between h eart and skele tal muscle ra ises some ques t ions as
to mechani sm. T he ava i l ab le ev idence ind ica te s tha t t h i s i s fundam enta l ly a pro blem
involv ing d i f fe rence in membrane func t ion , which , t hough ve ry in t e res t ing , w i l l no t
be discussed further s ince our interes t here is essent ia l ly in the contract i le process .
Dr Katz: I ce r t a in ly agree . I a m aware of these find ings , bu t I d idn t m ent ion
them because they a re proba bly of l i tt l e physio log ica l impor t anc e . My main po in t i s
that changes in cardia c muscle contract i l i ty, unl ike those which occ ur in skeleta l
muscle , are paral le led by chang es in Vm~,. Thi s s t rongly sugges ts that these func-
t iona l ly impo r t an t va r i a t ions in myoc ard ia l cont rac t i l i t y a re due to changes in the
exten t o f ac t iva t ion of the hea r t s co nt rac t i le p ro te ins .
Published July 1, 1967