lectins and their application to clinical microbiologycmr.asm.org/content/3/3/197.full.pdf ·...
TRANSCRIPT

CLINICAL MICROBIOLOGY REVIEWS, JU1y 1990, P. 197-218 Vol. 3, No. 30893-8512/90/030197-22$02.00/0Copyright © 1990, American Society for Microbiology
Lectins and Their Application to Clinical MicrobiologyMALCOLM SLIFKINl* AND R. J. DOYLE2
Microbiology Section, Department of Laboratory Medicine, Allegheny General Hospital, Pittsburgh, Pennsylvania 15212,1and Department of Microbiology and Immunology, University of Louisville Health Sciences Center,
Louisville, Kentucky 402922
OVERVIEW OF LECTINS........................................................... 197APPLICATION OF LECTINS IN CLINICAL MICROBIOLOGY ..................................................200
Gram-Positive Bacteria........................................................... 200Streptococci ........................................................... 200Lactobacilli ........................................................... 201Staphylococci ........................................................... 201Bacillus spp............................................................202Listeria monocytogenes......................................................... 203
Gram-Negative Bacteria ........................................................... 204Neisseria spp............................................................204Campylobacter (Helicobacter) spp...........................................................205Legionella spp............................................................205Enterobacteriaceae ......................................................... 205Pseudomonas spp............................................................205
Mycobacteria ........................................................... 206Mycology.......................................................... 206Parasitology .......................................................... 207Virology ........................................................... 210
USE OF LECTINS IN EPIDEMIOLOGIC INVESTIGATIONS ......................................................210APPLICATION OF LECTINS IN SEROLOGY ........................................................... 212APPLICATION OF LECTINS IN BLOOD GROUP DETERMINATIONS ........................................212CONCLUDING REMARKS ........................................................... 212ACKNOWLEDGMENTS ........................................................... 213LITERATURE CITED ........................................................... 214
OVERVIEW OF LECTINS
The term lectin is derived from the latin word legere, topick, choose, or select (17), as a generic name for a sub-stance that has the capacity to bind analogously to anantibody but is not induced within the immune system byantigenic stimulation (31).
Several definitions of lectins have been stated by investi-gators during the last 10 years. The definition of Goldsteinand his associates (61), however, represents the currentlyaccepted meaning of the word lectin. A lectin is a carbohy-drate-binding protein or glycoprotein of nonimmune originwhich agglutinates cells or precipitates glycoconjugates orboth. Lectins contain at least two carbohydrate-bindingsites. The specificity of a lectin is generally presented interms of the monosaccharide(s) or simple oligosaccharidethat inhibits the lectin-associated reaction. Lectins may bederived from plant, microbial, or animal sources and may besoluble or membrane bound. In some instances, carbohy-drate-associated enzymes that have multiple combining siteswill agglutinate or precipitate glycoconjugates and thus maybe considered as lectins (61). Lectins may also bind tononcarbohydrate ligands (31, 32, 90) and have been shown toreact with nuclei (90). The binding of some lectins tohydrophobic residues is well known.Some plant lectins may agglutinate various blood groups
of erythrocytes (19, 30, 99) and are therefore called phyto-hemagglutinins. Lectins are also referred to in the literature
* Corresponding author.
as agglutinins, phytoagglutinins, and protecting (20, 31, 90,107).Most common lectins are derived from plant seeds, bark,
tubers, pulp, bulbs, and leaves. Substances, however, thatselectively bind carbohydrates and interact with procaryoticand eucaryotic cells have been found in other biologicalsources, including bacteria, invertebrates, and mammaliantissues (31, 51, 59, 60, 69, 90). Plant and animal lectins maybe classified into groups on the basis of their carbohydrate-binding specificities. Table 1 provides a summary of thesources and specificities of lectins most commonly used inmicrobiology. Generally, lectins are classified in five groupson the basis of their affinity for (i) glucose/mannose; (ii)galactose and N-acetyl-D-galactosamine; (iii) N-acetylglu-cosamine; (iv) L-fucose, and (v) sialic acids (31). Binding isreversible and noncovalent with simple or complex carbo-hydrate conjugates, whether free in solution or on cellsurfaces (20, 107). Glycoconjugate-containing surfaces arecalled lectin receptors.The specificity of lectins is generally defined on the basis
of hapten inhibition tests in which various sugars or saccha-rides are tested for their capacity to inhibit hemagglutinationof erythrocytes. The binding of many lectins can be effec-tively inhibited with more than one carbohydrate. All lectinmolecules possess two or more carbohydrate-binding sites, aproperty essential to their ability to agglutinate cells or toreact with complex carbohydrates (165). The combiningsizes of some lectins may accommodate up to five or moresugar residues (43, 165). Many multibranched oligosaccha-
197
on Septem
ber 5, 2018 by guesthttp://cm
r.asm.org/
Dow
nloaded from
198 SLIFKIN AND DOYLE
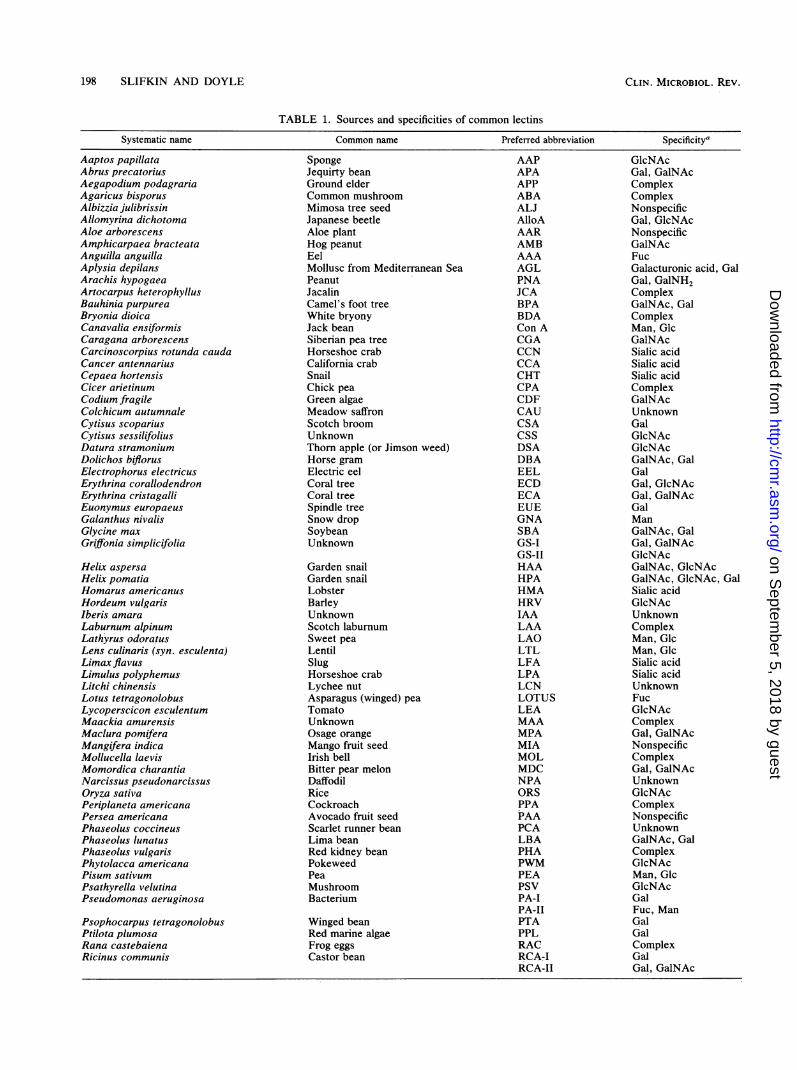
TABLE 1. Sources and specificities of common lectins
Systematic name Common name Preferred abbreviation Specificity'
Aaptos papillataAbrus precatoriusAegapodium podagrariaAgaricus bisporusAlbizzia julibrissinAllomyrina dichotomaAloe arborescensAmphicarpaea bracteataAnguilla anguillaAplysia depilansArachis hypogaeaArtocarpus heterophyllusBauhinia purpureaBryonia dioicaCanavalia ensiformisCaragana arborescensCarcinoscorpius rotunda caudaCancer antennariusCepaea hortensisCicer arietinumCodium fragileColchicum autumnaleCytisus scopariusCytisus sessilifoliusDatura stramoniumDolichos biflorusElectrophorus electricusErythrina corallodendronErythrina cristagalliEuonymus europaeusGalanthus nivalisGlycine maxGriffonia simplicifolia
Helix aspersaHelix pomatiaHomarus americanusHordeum vulgarisIberis amaraLaburnum alpinumLathyrus odoratusLens culinaris (syn. esculenta)Limax flavusLimulus polyphemusLitchi chinensisLotus tetragonolobusLycoperscicon esculentumMaackia amurensisMaclura pomiferaMangifera indicaMollucella laevisMomordica charantiaNarcissus pseudonarcissusOryza sativaPeriplaneta americanaPersea americanaPhaseolus coccineusPhaseolus lunatusPhaseolus vulgarisPhytolacca americanaPisum sativumPsathyrella velutinaPseudomonas aeruginosa
Psophocarpus tetragonolobusPtilota plumosaRana castebaienaRicinus communis
SpongeJequirty beanGround elderCommon mushroomMimosa tree seedJapanese beetleAloe plantHog peanutEelMollusc from Mediterranean SeaPeanutJacalinCamel's foot treeWhite bryonyJack beanSiberian pea treeHorseshoe crabCalifornia crabSnailChick peaGreen algaeMeadow saffronScotch broomUnknownThorn apple (or Jimson weed)Horse gramElectric eelCoral treeCoral treeSpindle treeSnow dropSoybeanUnknown
Garden snailGarden snailLobsterBarleyUnknownScotch laburnumSweet peaLentilSlugHorseshoe crabLychee nutAsparagus (winged) peaTomatoUnknownOsage orangeMango fruit seedIrish bellBitter pear melonDaffodilRiceCockroachAvocado fruit seedScarlet runner beanLima beanRed kidney beanPokeweedPeaMushroomBacterium
Winged beanRed marine algaeFrog eggsCastor bean
AAPAPAAPPABAALJAlloAAARAMBAAAAGLPNAJCABPABDACon ACGACCNCCACHTCPACDFCAUCSACSSDSADBAEELECDECAEUEGNASBAGS-IGS-IIHAAHPAHMAHRVIAALAALAOLTLLFALPALCNLOTUSLEAMAAMPAMIAMOLMDCNPAORSPPAPAAPCALBAPHAPWMPEAPSVPA-IPA-ILPTAPPLRACRCA-IRCA-Il
GlcNAcGal, GalNAcComplexComplexNonspecificGal, GlcNAcNonspecificGalNAcFucGalacturonic acid, GalGal, GalNH2ComplexGalNAc, GalComplexMan, GlcGalNAcSialic acidSialic acidSialic acidComplexGalNAcUnknownGalGlcNAcGlcNAcGalNAc, GalGalGal, GlcNAcGal, GalNAcGalManGaINAc, GalGal, GaiNAcGlcNAcGalNAc, GlcNAcGalNAc, GlcNAc, GalSialic acidGlcNAcUnknownComplexMan, GlcMan, GlcSialic acidSialic acidUnknownFucGlcNAcComplexGal, GalNAcNonspecificComplexGal, GalNAcUnknownGlcNAcComplexNonspecificUnknownGalNAc, GalComplexGlcNAcMan, GlcGlcNAcGalFuc, ManGalGalComplexGalGal, GalNAc
CLIN. MICROBIOL. REV.
on Septem
ber 5, 2018 by guesthttp://cm
r.asm.org/
Dow
nloaded from

TABLE 1. (continued)
Systematic name Common name Preferred abbreviation Specificity'
Robinia pseudoacacia Black locust RPA UnknownSambucus nigra Elderberry SNA GalSarcophaga perigrina Flesh fly SAP ComplexSarothamnus scoparius Unknown SRA Gal, FucSolanum dulcamara Woody nightshade SDA UnknownSolanum tuberosum Potato STA (GIcNAc)25Soja hipspida Bean, very similar to soybean SOH GalNAc, GalSophora japonica Japanese pagoda tree SJA GaINAc, GalTridacna crocea Mollusc TCA GalTrichosanthes kinlouii Chinese herb TKA GalTrifolium repens White clover RTA ComplexTriticum vulgaris Wheat WGA (GIcNAc)1-3Tulipa gesneriana Tulip TUL UnknownUlex europaeus Gorse (or furze) UEA-I Fuc
UEA-II (GlcNAc)2Ulva lectura Green marine alga ULL FucUrtica dioica Stinging nettle URD (GlcNAc)2Vicia cracca Common vetch VCA GalNAcVicia faba Fava (broad) bean VFA Man, GlcVicia graminea Unknown VGA GalVicia villosa Hairy vetch VVA GaINAcVigna radiata Mung bean VGR GalViscum album Mistletoe VAA Gal, GalNAWisteria floribunda Japanese wisteria WFA GalNAc, Gal
a Specificities determined by hemagglutination inhibition, equilibrium dialysis, fluorescence quenching, other. Specificities do not reflect contribution ofpenultimate residues or peptides. Table derived from references 43, 58, 60, 87, 97, and 143 and commercial literature of E-Y laboratories and Sigma ChemicalCo. Gal, Galactose; Fuc = fucose; Glc = glucose; GalNAc, N-acetylgalactosamine; GlcNAc, N-acetylglucosamine; Man, mannose.
rides exhibit stronger lectin-binding reactivities than linearones, because of cooperative binding effects of lectin andcarbohydrate complexes (165). Some lectins possess dual ormultiple affinities for different disaccharides. Binding mayinvolve hydrophobic and hydrogen bonds, while electro-static forces are rarely involved.
Lectins have been purified from crude aqueous, buffer, orsaline extracts of various tissues by standard methods asso-ciated with protein chemistry. These include ammoniumsulfate or ethanol precipitation and the use of affinity chro-matography (20, 107).The biological role of lectins is quite speculative (52; 60).
Lectins may be involved in sugar transport or carbohydratestorage. Some lectins may be associated with the binding ofsymbiotic rhizobia to form root nodules. Because of theirrole in adhesion and agglutination, lectins have been consid-ered to be important in both symbiotic and pathogenicinteractions between some microorganisms and hosts. Themicrobial lectins may play an essential role in mediatingadhesion to surfaces colonized by the microorganism. Thepresence of lectins in food products such as the tomato, forexample, is common (104). The tomato lectin binds tomucosal cells and resists denaturation by acid and by pro-teolytic enzymes (52, 90).The hemagglutinating activity of plant extracts was first
reported in 1889-1889 by Stillmark, who investigated thecastor bean agglutinin Ricinus communes (cited in reference16). Other investigators established lectins as agglutininsspecific for blood groups of erythrocytes (15, 17, 20, 31, 66,67, 107). Due to their multifaceted biological properties,lectins were later developed by cell biologists as probes toinvestigate cell surface structures and functions. Interactionwith lectins can be used to obtain independent informationabout the presence of specific carbohydrates, the configura-tion of anomeric linkages, and the location or position ofcarbohydrate residues in polysaccharide molecules.
Lectin-cell binding can elicit a wide variety of biological
phenomena. Some lectins induce mitosis in resting lympho-cytes and some agglutinate neoplastic transformed cells, butnot their healthy nonmalignant counterparts, whereas othersagglutinate healthy nonmalignant cells (20, 31, 69). Lectinshave been used to fractionate animal cells, including B and Tlymphocytes (107), and to demonstrate changes in cellsurface architecture following virus infection or parasiteinfection (31). These extremely versatile tools have beenapplied as fluorescent and electron microscopic probes andin gel diffusion assays. Antisera to many of the lectins havebeen produced and labeled with markers for use in indirectstudies on lectin binding. Immobilized lectins used for af-finity chromatography have an advantage over many otherpurification techniques because elution can be pursued witha relatively inexpensive monosaccharide, and the glycopro-tein to be purified is generally not subjected to harsh ordenaturing conditions.Some lectins are potent toxins and have been applied as
therapeutic agents. The lectins ricin and abrin have beencoupled to specific monoclonal antibodies and applied asimmunotoxins in cancer therapy. Lectins have been used ascarriers for the delivery of chemotherapeutic agents. Lectinsare significant reagents for investigating cell surface recep-tors in bacteria, protozoa, and higher organisms. The inter-actions of plant lectins with microorganisms have beenapplied for the typing of bacteria, fungi, and protozoa.Lectins are useful for characterizing bacterial cell compo-hents and for detecting bacteriophage receptors (52, 90, 91,107).Many bacteria contain surface-associated lectins that en-
able these organisms to adhere to surfaces (37). Bacteriallectins resemble plant lectins in carbohydrate specificity,relative thermostability, divalent cation requirements, andother properties and may be used for some of the samepurposes as other lectins. Several bacterial species possesssurface lectins that yield coaggregation responses with cer-tain yeast cells (98, 99).
VOL. 3, 1990 LECTINS 199
on Septem
ber 5, 2018 by guesthttp://cm
r.asm.org/
Dow
nloaded from

200 SLIFKIN AND DOYLE
The unique property of lectins to bind noncovalently tosimple sugars and polysaccharides has attracted interest inmicrobial taxonomy. Lectins have a role in the clinicallaboratory identification and taxonomic classification ofmany microorganisms. Because lectins are generally mono-clonal proteins and because they possess a spectrum ofspecificities and molecular weights, they are substantialtools for diagnostic microbiology applications.One of the distinct advantages of applying lectins in
microbiology is that cellular or surface receptor sites can bepartially characterized by hapten inhibition studies (31, 87).Unlike the production of antisera, which requires pretreat-ment of microorganisms for antigen preparation, injection ofthe microorganism and glyconjugate into animals, such asrabbits, and absorption of antisera to eliminate nonspecificantibody reactions (144), lectins are simple to use. Whenconjugated to a histochemical label such as fluorescein,peroxidase, or colloidal gold, lectins may be used as histo-chemical probes to identify and localize specific carbohy-drate residues in microorganisms by light or electron micros-copy, as well as by blotting methods.
In 1936, Summer and Howell (152) reported that conca-navalin A agglutinated various Mycobacterium spp. Fromthat time, the momentum of lectin use in clinical microbiol-ogy began.
In bacteria, numerous surface structures bind lectins, suchas peptidoglycan, teichoic acids, lipopolysaccharides, andcapsular materials. Similarly, fungi exhibit a diversity ofpotential lectin-reactive sites, including various polysaccha-rides, chitin, and glycoproteins. Protozoa and metazoa havesurface glycoproteins. In addition, glycoproteins and enve-lope proteins of some viruses may bind lectins. Plant lectinsusually interact with nonreducing carbohydrate termini, butpenultimate residues and their nearest neighbors influencethe affinity between the protein and carbohydrate functionalgroups. The large number of potential lectin-reactive sites onmicrobial surfaces enhances the prospect of using lectins asdiagnostic reagents. Even though several lectins possess thesame specificities, it does not follow that each of them willinteract with a given microbial structure. The nature of lectinreceptors on microbial-viral surfaces is determined by acombination of factors such as penultimate residues, hydro-phobicity, and accessibility.
This article reviews the use of lectins as diagnostic re-agents for the identification, taxonomy, and epidemiologicmonitoring of microbial and viral agents. Overviews con-cerning the applications of lectins are presented for thecommon areas of clinical microbiology. The review is in-tended to stimulate a reexamination of published diagnosticapplications of lectins as well as the development of newapplications of lectins as diagnostic probes in clinical micro-biology.
APPLICATIONS OF LECTINS INCLINICAL MICROBIOLOGY
Gram-Positive Bacteria
Streptococci. The use of lectins for investigating virulencefactors, surface structures, and identification of streptococciis well established. Lectins have been used to identifybeta-hemolytic streptococci, especially group C strepto-cocci. The albumen gland of the edible snail, Helix pomatia,contains an agglutinin that specifically binds to group Ahuman erythrocytes with a specificity directed to terminalnonreducing N-acetyl-D-galactosamine residues (83, 123,
158). Because group C beta-hemolytic streptococci have asimilar terminal aminocarbohydrate as part of their groupcarbohydrate, it was considered that the agglutinin wouldreact with them.
In early investigations, crude extracts containing Helixpomatia lectin were shown to identify group C specificallywithout agglutination responses from either group A or Gstreptococci (81-83). The lectins from Dolichos biflorus (82,123) and Wisteria floribunda (116) also agglutinate group Cbeta-hemolytic streptococci.A test that used three fluorogenic 4-methylumbelliferyl
substrates and the lectin from Dolichos biflorus provided arapid, simple, and specific means to identify group A, B, G,F, and G streptococci (146). The use of the Dolichos bifloruslectin confirmed its specificity for identifying group C strep-tococci. In another test, crude extracts of Dolichos bifloruswere coupled to polystyrene particles with a spacer arm toyield an effective lectin-latex reagent that agglutinated groupC streptococcal antigens prepared as nitrous acid, autoclave,or enzyme extracts (147). Group C streptococcal isolatesfrom horses and cattle agglutinated with lectins from Doli-chos biflorus and Helix pomatia. Some strains requiredboiling for 30 min at pH 2.0 to yield an agglutination reactionwith these lectins (136).The lectin from the snail Cepaea hortensis agglutinated all
group B streptococcal strains containing the sialic acidtype-specific polysaccharide. Group B streptococci lackingthe type-specific polysaccharide, as well as strains of otherstreptococcal groups and many other bacteria, includingspecies containing sialic acid, were not agglutinated (162).Complement was necessary for phagocytosis of group Bstreptococci after sialic acid was blocked on the surface oflectin-treated cells (71). Cepaea hortensis lectin promotedthe phagocytosis of serotype Ia group B streptococci in theabsence of opsonic antibodies. Therefore, blocking of thesialic acid by the sialic acid-specific lectin from Cepaeahortensis promotes phagocytosis of a group B Streptococcussp. type Ia (71). Group B strains lacking the polysaccharidetype antigens and containing only protein antigen ic, R, or Xdid not react with this lectin (162). This test may be ofdiagnostic value since the latter strains have been associatedwith neonatal human infections (7).The lectin of the tomato, Lycopersicon esculentum, or the
potato, Solanum tuberosum (Table 1), can be passivelycoupled to amide-modified polystyrene spheres and used asa detection reagent for the specific identification of group Bstreptococci grown in selective or nonselective Todd-Hewittbroth for 5 h (144). In this medium, group B streptococciproduce excess group antigens. Therefore, the Lycopersiconesculentum-latex reagent agglutinates when reacted withclarified broth cultures. These lectins did not agglutinate inbroths in which group A, C, D, F, or G streptococci weregrown. False-negative agglutination reactions occurredwhen the reagent was reacted directly with serotypes Ia andlb of group B streptococci grown on Columbia sheep bloodagar (144). The lectin from the related solanaceous plantDatura stramonium (Jimson weed) did not agglutinate groupB streptococci, perhaps because it does not react withdimers or trimers of N-acetyl-D-glucosamine. Although therelevant antigenic determinants of group B polysaccharideappear to be associated with rhamnose-glucitol units orterminal rhamnose on side chains associated with N-acetyl-glucosamine (121, 122), it is possible that these lectins caninteract with other determinants on group B organisms. Thelectin from the snail Cepaea hortensis has a specificity for
CLIN. MICROBIOL. REV.
on Septem
ber 5, 2018 by guesthttp://cm
r.asm.org/
Dow
nloaded from

VOL. 3, 1990
MAJY3
Hi he s is **I-
FIG 1. Interaction between group B st iT ad Pr s cue Lyc ci
FIG. 1. Interaction between group B streptococci and fluorescein-conjugated Lysopersicon esculentum lectin.
sialic acid-containing polysacharides (162), but tomato andpotato lectins may also have low affinities for sialic acids.The tomato and potato lectin reagents should be of value
for their ability to help diagnose group B streptococcalcolonization in women and infants. These lectins could beused with Todd-Hewitt broth containing antibiotics for theselective isolation and detection of group B streptococci(88).A fluorescein-conjugated Lycopersicon lectin reagent la-
beled group B streptococcal cells (Fig. 1). No fluorescencewas detected when the other serogroups of streptococciwere examined with this fluorescein-conjugated reagent(144).Loach egg lectin extract from the fish Misgurnus anguil-
licaudatus was found to be inhibited best with L-rhamnoseand L-mannose (132). This observation suggests that thislectin might be useful in distinguishing streptococci fromother bacteria. However, loach lectin also reacts weaklywith galactose residues.
Lectins have been used to study attachment of non-beta-hemolytic streptococci to host cell surfaces. One inves-tigation with the agglutinin (possibly a tannin) extracted fromthe seeds of the avocado, Persea americana, indicated thatattachment of Streptococcus mutans to tooth surfaces isdependent on nonglucan factors (149). Pretreatment ofStreptococcus mutans with this agglutinin reduced adhesionof the streptococcus independent of sucrose concentration(150). Furthermore, the Persea agglutinin agglutinatedStreptococcus mutans cells (150). No visible agglutination,
however, occurred when extracts of this seed were mixedwith polymers found on bacterial cell surfaces, such asteichoic acids and dextrans (97).
Peroxidase-conjugated asparagus pea lectin was used toidentify fucosyl residues in the cell walls of Enterococcusfaecalis with Western blotting (2). In addition, by usingvarious lectin probes, such as wheat germ agglutinin, con-canavalin A, and soybean agglutinin, the presence of N-acetylneuraminyl, glucosyl, or mannosyl and N-acetylgalac-tosaminyl residues on the surface of this bacterial specieswas determined.
In other studies, crude polysaccharide from Streptococcuspneumonia type 12 was purified with concanavalin A (24).Wheat germ agglutinin and the hemagglutinin from Ricinuscommunis seeds precipitated with pneumococcal S-14polysaccharide (50).
Lactobacilhi. Various bacterial polysaccharides are precip-itated by concanavalin A, which interacts specifically withbacterial cell walls containing glycosidic residues associatedwith teichoic acid. Accordingly, bacterial teichoic acids fromcell walls containing a-glucopyranosyl residues, such asfrom Lactobacillus plantarum and Lactobacillus buchneri,precipitated in the presence of concanavalin A when testedby gel diffusion (5).
Staphylococci. Concanavalin A was shown to interact withextracted teichoic acid derived from Staphylococcus aureusand S. epidermidis (125). That investigation emphasized theusefulness of this lectin as a relatively simple means todetermine the anomeric configuration of the carbohydrate
LECTINS 201
on Septem
ber 5, 2018 by guesthttp://cm
r.asm.org/
Dow
nloaded from
202 SLIFKIN AND DOYLE
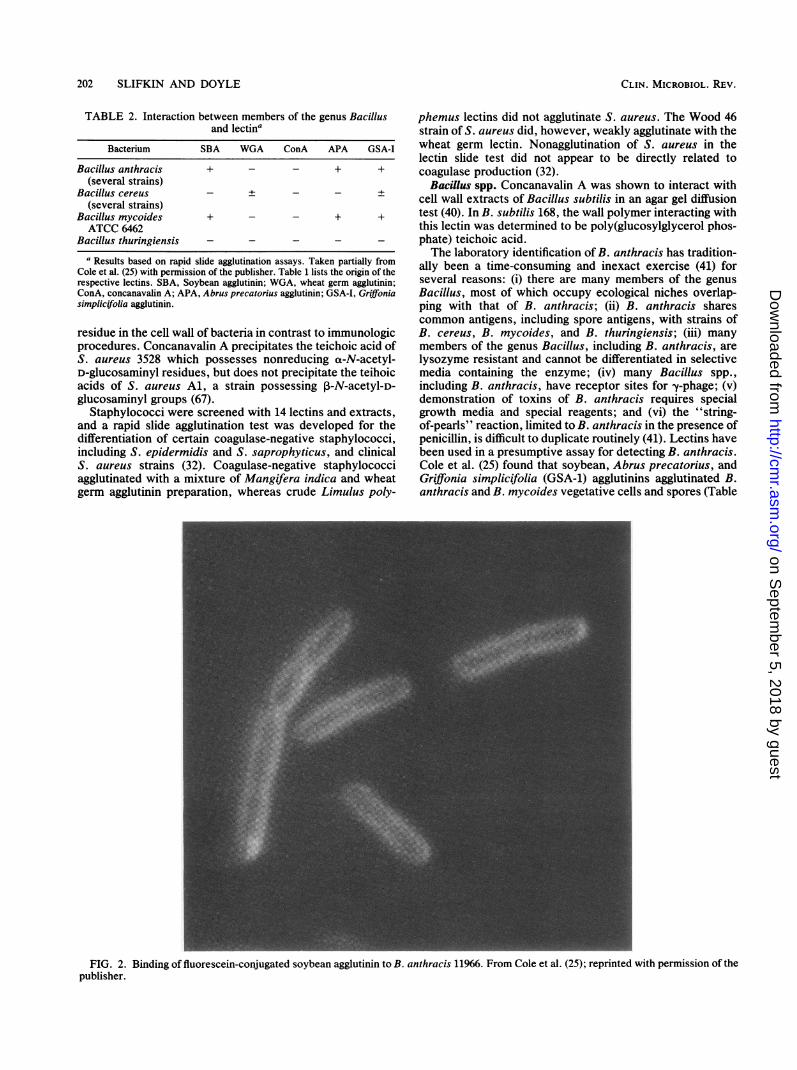
TABLE 2. Interaction between members of the genus Bacillusand lectina
Bacterium SBA WGA ConA APA GSA-I
Bacillus anthracis + - - + +(several strains)
Bacillus cereus - + - - +(several strains)
Bacillus mycoides + - - + +ATCC 6462
Bacillus thuringiensis - - - - -a Results based on rapid slide agglutination assays. Taken partially from
Cole et al. (25) with permission of the publisher. Table 1 lists the origin of therespective lectins. SBA, Soybean agglutinin; WGA, wheat germ agglutinin;ConA, concanavalin A; APA, Abrus precatorius agglutinin; GSA-I, Griffoniasimplicifolia agglutinin.
residue in the cell wall of bacteria in contrast to immunologicprocedures. Concanavalin A precipitates the teichoic acid ofS. aureus 3528 which possesses nonreducing oa-N-acetyl-D-glucosaminyl residues, but does not precipitate the teihoicacids of S. aureus Al, a strain possessing P-N-acetyl-D-glucosaminyl groups (67).
Staphylococci were screened with 14 lectins and extracts,and a rapid slide agglutination test was developed for thedifferentiation of certain coagulase-negative staphylococci,including S. epidermidis and S. saprophyticus, and clinicalS. aureus strains (32). Coagulase-negative staphylococciagglutinated with a mixture of Mangifera indica and wheatgerm agglutinin preparation, whereas crude Limulus poly-
phemus lectins did not agglutinate S. aureus. The Wood 46strain of S. aureus did, however, weakly agglutinate with thewheat germ lectin. Nonagglutination of S. aureus in thelectin slide test did not appear to be directly related tocoagulase production (32).
Bacillus spp. Concanavalin A was shown to interact withcell wall extracts of Bacillus subtilis in an agar gel diffusiontest (40). In B. subtilis 168, the wall polymer interacting withthis lectin was determined to be poly(glucosylglycerol phos-phate) teichoic acid.The laboratory identification of B. anthracis has tradition-
ally been a time-consuming and inexact exercise (41) forseveral reasons: (i) there are many members of the genusBacillus, most of which occupy ecological niches overlap-ping with that of B. anthracis; (ii) B. anthracis sharescommon antigens, including spore antigens, with strains ofB. cereus, B. mycoides, and B. thuringiensis; (iii) manymembers of the genus Bacillus, including B. anthracis, arelysozyme resistant and cannot be differentiated in selectivemedia containing the enzyme; (iv) many Bacillus spp.,including B. anthracis, have receptor sites for -y-phage; (v)demonstration of toxins of B. anthracis requires specialgrowth media and special reagents; and (vi) the "string-of-pearls" reaction, limited to B. anthracis in the presence ofpenicillin, is difficult to duplicate routinely (41). Lectins havebeen used in a presumptive assay for detecting B. anthracis.Cole et al. (25) found that soybean, Abrus precatorius, andGriffonia simplicifolia (GSA-1) agglutinins agglutinated B.anthracis and B. mycoides vegetative cells and spores (Table
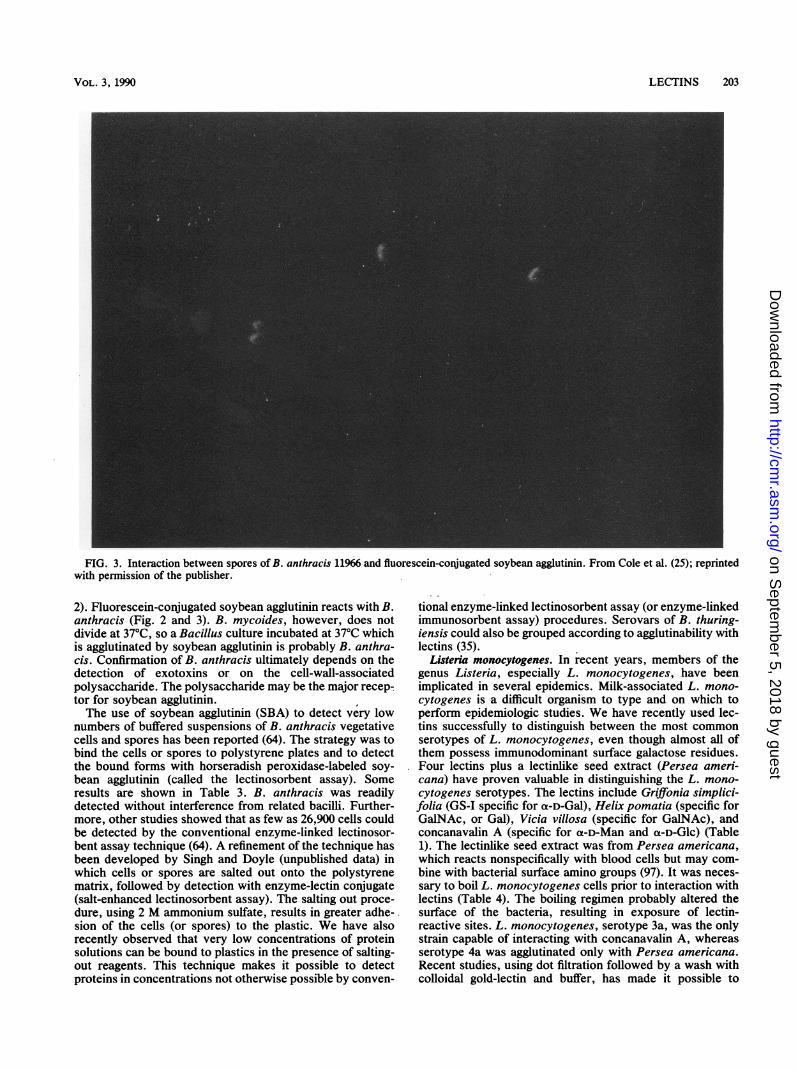
FIG. 2. Binding of fluorescein-conjugated soybean agglutinin to B. anthracis 11966. From Cole et al. (25); reprinted with permission of thepublisher.
CLIN. MICROBIOL. REV.
on Septem
ber 5, 2018 by guesthttp://cm
r.asm.org/
Dow
nloaded from
LECTINS 203
FIG. 3. Interaction between spores of B. anthracis 11966 and fluorescein-conjugated soybean agglutinin. From Cole et al. (25); reprintedwith permission of the publisher.
2). Fluorescein-conjugated soybean agglutinin reacts with B.anthracis (Fig. 2 and 3). B. mycoides, however, does notdivide at 37°C, so a Bacillus culture incubated at 37°C whichis agglutinated by soybean agglutinin is probably B. anthra-cis. Confirmation of B. anthracis ultimately depends on thedetection of exotoxins or on the cell-wall-associatedpolysaccharide. The polysaccharide may be the major recep-tor for soybean agglutinin.The use of soybean agglutinin (SBA) to detect very low
numbers of buffered suspensions of B. anthracis vegetativecells and spores has been reported (64). The strategy was tobind the cells or spores to polystyrene plates and to detectthe bound forms with horseradish peroxidase-labeled soy-bean agglutinin (called the lectinosorbent assay). Someresults are shown in Table 3. B. anthracis was readilydetected without interference from related bacilli. Further-more, other studies showed that as few as 26,900 cells couldbe detected by the conventional enzyme-linked lectinosor-bent assay technique (64). A refinement of the technique hasbeen developed by Singh and Doyle (unpublished data) inwhich cells or spores are salted out onto the polystyrenematrix, followed by detection with enzyme-lectin conjugate(salt-enhanced lectinosorbent assay). The salting out proce-dure, using 2 M ammonium sulfate, results in greater adhe-sion of the cells (or spores) to the plastic. We have alsorecently observed that very low concentrations of proteinsolutions can be bound to plastics in the presence of salting-out reagents. This technique makes it possible to detectproteins in concentrations not otherwise possible by conven-
tional enzyme-linked lectinosorbent assay (or enzyme-linkedimmunosorbent assay) procedures. Serovars of B. thuring-iensis could also be grouped according to agglutinability withlectins (35).
Listeria monocytogenes. In recent years, members of thegenus Listeria, especially L. monocytogenes, have beenimplicated in several epidemics. Milk-associated L. mono-cytogenes is a difficult organism to type and on which toperform epidemiologic studies. We have recently used lec-tins successfully to distinguish between the most commonserotypes of L. monocytogenes, even though almost all ofthem possess immunodominant surface galactose residues.Four lectins plus a lectinlike seed extract (Persea ameri-cana) have proven valuable in distinguishing the L. mono-cytogenes serotypes. The lectins include Griffionia simplici-folia (GS-I specific for a-D-Gal), Helix pomatia (specific forGalNAc, or Gal), Vicia villosa (specific for GalNAc), andconcanavalin A (specific for oL-D-Man and a-D-Glc) (Table1). The lectinlike seed extract was from Persea americana,which reacts nonspecifically with blood cells but may com-bine with bacterial surface amino groups (97). It was neces-sary to boil L. monocytogenes cells prior to interaction withlectins (Table 4). The boiling regimen probably altered thesurface of the bacteria, resulting in exposure of lectin-reactive sites. L. monocytogenes, serotype 3a, was the onlystrain capable of interacting with concanavalin A, whereasserotype 4a was agglutinated only with Persea americana.Recent studies, using dot filtration followed by a wash withcolloidal gold-lectin and buffer, has made it possible to
VOL. 3, 1990
on Septem
ber 5, 2018 by guesthttp://cm
r.asm.org/
Dow
nloaded from
204 SLIFKIN AND DOYLE
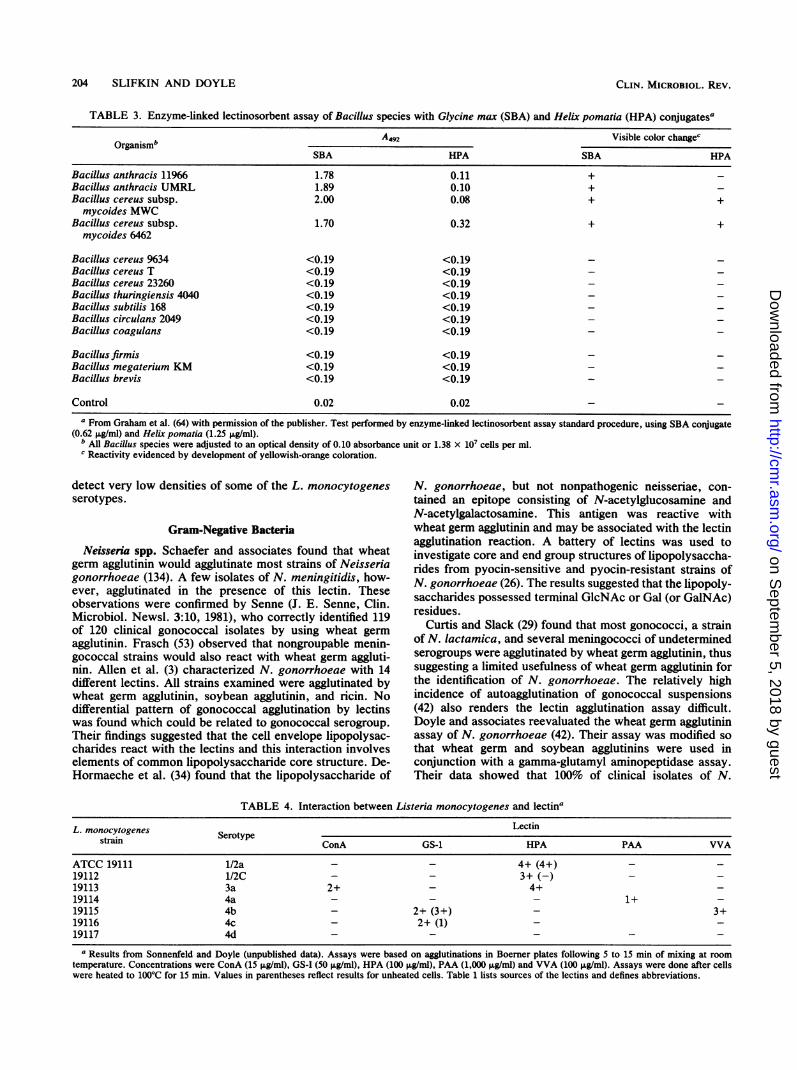
TABLE 3. Enzyme-linked lectinosorbent assay of Bacillus species with Glycine max (SBA) and Helix pomatia (HPA) conjugatesa
A4s2 Visible color changeOrganismb__
SBA HPA SBA HPA
Bacillus anthracis 11966 1.78 0.11 +Bacillus anthracis UMRL 1.89 0.10 +Bacillus cereus subsp. 2.00 0.08 + +
mycoides MWCBacillus cereus subsp. 1.70 0.32 + +
mycoides 6462
Bacillus cereus 9634 <0.19 <0.19Bacillus cereus T <0.19 <0.19 - -
Bacillus cereus 23260 <0.19 <0.19Bacillus thuringiensis 4040 <0.19 <0.19Bacillus subtilis 168 <0.19 <0.19Bacillus circulars 2049 <0.19 <0.19Bacillus coagulans <0.19 <0.19
Bacillusfirmis <0.19 <0.19 - -
Bacillus megaterium KM <0.19 <0.19Bacillus brevis <0.19 <0.19
Control 0.02 0.02a From Graham et al. (64) with permission of the publisher. Test performed by enzyme-linked lectinosorbent assay standard procedure, using SBA conjugate
(0.62 ,ug/ml) and Helix pomatia (1.25 ,u&glml).b All Bacillus species were adjusted to an optical density of 0.10 absorbance unit or 1.38 x 107 cells per ml.c Reactivity evidenced by development of yellowish-orange coloration.
detect very low densities of some of the L. monocytogenesserotypes.
Gram-Negative Bacteria
Neisseria spp. Schaefer and associates found that wheatgerm agglutinin would agglutinate most strains of Neisseriagonorrhoeae (134). A few isolates of N. meningitidis, how-ever, agglutinated in the presence of this lectin. Theseobservations were confirmed by Senne (J. E. Senne, Clin.Microbiol. Newsl. 3:10, 1981), who correctly identified 119of 120 clinical gonococcal isolates by using wheat germagglutinin. Frasch (53) observed that nongroupable menin-gococcal strains would also react with wheat germ aggluti-nin. Allen et al. (3) characterized N. gonorrhoeae with 14different lectins. All strains examined were agglutinated bywheat germ agglutinin, soybean agglutinin, and ricin. Nodifferential pattern of gonococcal agglutination by lectinswas found which could be related to gonococcal serogroup.Their findings suggested that the cell envelope lipopolysac-charides react with the lectins and this interaction involveselements of common lipopolysaccharide core structure. De-Hormaeche et al. (34) found that the lipopolysaccharide of
N. gonorrhoeae, but not nonpathogenic neisseriae, con-tained an epitope consisting of N-acetylglucosamine andN-acetylgalactosamine. This antigen was reactive withwheat germ agglutinin and may be associated with the lectinagglutination reaction. A battery of lectins was used toinvestigate core and end group structures of lipopolysaccha-rides from pyocin-sensitive and pyocin-resistant strains ofN. gonorrhoeae (26). The results suggested that the lipopoly-saccharides possessed terminal GlcNAc or Gal (or GalNAc)residues.
Curtis and Slack (29) found that most gonococci, a strainof N. lactamica, and several meningococci of undeterminedserogroups were agglutinated by wheat germ agglutinin, thussuggesting a limited usefulness of wheat germ agglutinin forthe identification of N. gonorrhoeae. The relatively highincidence of autoagglutination of gonococcal suspensions(42) also renders the lectin agglutination assay difficult.Doyle and associates reevaluated the wheat germ agglutininassay of N. gonorrhoeae (42). Their assay was modified sothat wheat germ and soybean agglutinins were used inconjunction with a gamma-glutamyl aminopeptidase assay.Their data showed that 100% of clinical isolates of N.
TABLE 4. Interaction between Listeria monocytogenes and lectina
L. monocytogenes Lectinstrain Serotype LciConA GS-1 HPA PAA VVA
ATCC 19111 1/2a - - 4+ (4+)19112 1/2C - - 3+ (-) - -
19113 3a 2+ - 4+19114 4a - - - 1+19115 4b - 2+ (3+) 3+19116 4c - 2+ (1)19117 4d - - - -
a Results from Sonnenfeld and Doyle (unpublished data). Assays were based on agglutinations in Boerner plates following 5 to 15 min of mixing at roomtemperature. Concentrations were ConA (15 p1g/ml), GS-I (50 1Lg/ml), HPA (100 Fig/ml), PAA (1,000 Fg/ml) and VVA (100 SLg/ml). Assays were done after cellswere heated to 1000C for 15 min. Values in parentheses reflect results for unheated cells. Table 1 lists sources of the lectins and defines abbreviations.
CLIN. MICROBIOL. REV.
on Septem
ber 5, 2018 by guesthttp://cm
r.asm.org/
Dow
nloaded from
LECTINS 205
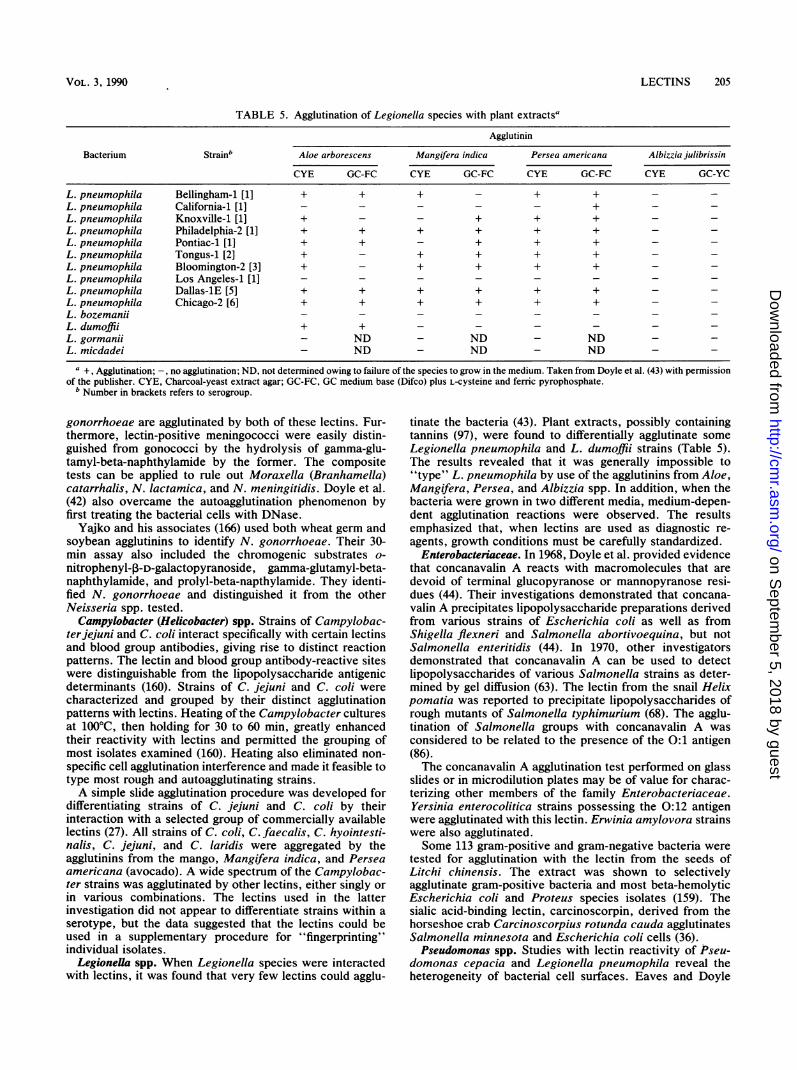
TABLE 5. Agglutination of Legionella species with plant extractsa
Agglutinin
Bacterium Strainb Aloe arborescens Mangifera indica Persea americana Albizzia julibrissin
CYE GC-FC CYE GC-FC CYE GC-FC CYE GC-YC
L. pneumophila Bellingham-1 [1] + + + - + + -
L. pneumophila California-1 [1] - - - - - + -
L. pneumophila Knoxville-1 [1] + - - + + + -
L. pneumophila Philadelphia-2 [1] + + + + + + -
L. pneumophila Pontiac-1 [1i + + - + + + -
L. pneumophila Tongus-1 [2] + - + + + + -
L. pneumophila Bloomington-2 [3] + - + + + + -
L. pneumophila Los Angeles-1 [1iL. pneumophila Dallas-iE [5] + + + + + +L. pneumophila Chicago-2 [6] + + + + + +L. bozemaniiL. dumoffii + +L. gormanii - ND - ND - NDL. micdadei - ND - ND - ND -
a +, Agglutination; -, no agglutination; ND, not determined owing to failure of the species to grow in the medium. Taken from Doyle et al. (43) with permissionof the publisher. CYE, Charcoal-yeast extract agar; GC-FC, GC medium base (Difco) plus L-cysteine and ferric pyrophosphate.
b Number in brackets refers to serogroup.
gonorrhoeae are agglutinated by both of these lectins. Fur-thermore, lectin-positive meningococci were easily distin-guished from gonococci by the hydrolysis of gamma-glu-tamyl-beta-naphthylamide by the former. The compositetests can be applied to rule out Moraxella (Branhamella)catarrhalis, N. lactamica, and N. meningitidis. Doyle et al.(42) also overcame the autoagglutination phenomenon byfirst treating the bacterial cells with DNase.Yajko and his associates (166) used both wheat germ and
soybean agglutinins to identify N. gonorrhoeae. Their 30-min assay also included the chromogenic substrates o-nitrophenyl-,3-D-galactopyranoside, gamma-glutamyl-beta-naphthylamide, and prolyl-beta-napthylamide. They identi-fied N. gonorrhoeae and distinguished it from the otherNeisseria spp. tested.
Campylobacter (Helicobacter) spp. Strains of Campylobac-terjejuni and C. coli interact specifically with certain lectinsand blood group antibodies, giving rise to distinct reactionpatterns. The lectin and blood group antibody-reactive siteswere distinguishable from the lipopolysaccharide antigenicdeterminants (160). Strains of C. jejuni and C. coli werecharacterized and grouped by their distinct agglutinationpatterns with lectins. Heating of the Campylobacter culturesat 100°C, then holding for 30 to 60 min, greatly enhancedtheir reactivity with lectins and permitted the grouping ofmost isolates examined (160). Heating also eliminated non-specific cell agglutination interference and made it feasible totype most rough and autoagglutinating strains.A simple slide agglutination procedure was developed for
differentiating strains of C. jejuni and C. coli by theirinteraction with a selected group of commercially availablelectins (27). All strains of C. coli, C. faecalis, C. hyointesti-nalis, C. jejuni, and C. laridis were aggregated by theagglutinins from the mango, Mangifera indica, and Perseaamericana (avocado). A wide spectrum of the Campylobac-ter strains was agglutinated by other lectins, either singly orin various combinations. The lectins used in the latterinvestigation did not appear to differentiate strains within aserotype, but the data suggested that the lectins could beused in a supplementary procedure for "fingerprinting"individual isolates.
Legionella spp. When Legionella species were interactedwith lectins, it was found that very few lectins could agglu-
tinate the bacteria (43). Plant extracts, possibly containingtannins (97), were found to differentially agglutinate someLegionella pneumophila and L. dumoffii strains (Table 5).The results revealed that it was generally impossible to"type" L. pneumophila by use of the agglutinins from Aloe,Mangifera, Persea, and Albizzia spp. In addition, when thebacteria were grown in two different media, medium-depen-dent agglutination reactions were observed. The resultsemphasized that, when lectins are used as diagnostic re-agents, growth conditions must be carefully standardized.
Enterobacteriaceae. In 1968, Doyle et al. provided evidencethat concanavalin A reacts with macromolecules that aredevoid of terminal glucopyranose or mannopyranose resi-dues (44). Their investigations demonstrated that concana-valin A precipitates lipopolysaccharide preparations derivedfrom various strains of Escherichia coli as well as fromShigella flexneri and Salmonella abortivoequina, but notSalmonella enteritidis (44). In 1970, other investigatorsdemonstrated that concanavalin A can be used to detectlipopolysaccharides of various Salmonella strains as deter-mined by gel diffusion (63). The lectin from the snail Helixpomatia was reported to precipitate lipopolysaccharides ofrough mutants of Salmonella typhimurium (68). The agglu-tination of Salmonella groups with concanavalin A wasconsidered to be related to the presence of the 0:1 antigen(86).The concanavalin A agglutination test performed on glass
slides or in microdilution plates may be of value for charac-terizing other members of the family Enterobacteriaceae.Yersinia enterocolitica strains possessing the 0:12 antigenwere agglutinated with this lectin. Erwinia amylovora strainswere also agglutinated.Some 113 gram-positive and gram-negative bacteria were
tested for agglutination with the lectin from the seeds ofLitchi chinensis. The extract was shown to selectivelyagglutinate gram-positive bacteria and most beta-hemolyticEscherichia coli and Proteus species isolates (159). Thesialic acid-binding lectin, carcinoscorpin, derived from thehorseshoe crab Carcinoscorpius rotunda cauda agglutinatesSalmonella minnesota and Escherichia coli cells (36).Pseudomonas spp. Studies with lectin reactivity of Pseu-
domonas cepacia and Legionella pneumophila reveal theheterogeneity of bacterial cell surfaces. Eaves and Doyle
VOL. 3, 1990
on Septem
ber 5, 2018 by guesthttp://cm
r.asm.org/
Dow
nloaded from
206 SLIFKIN AND DOYLE
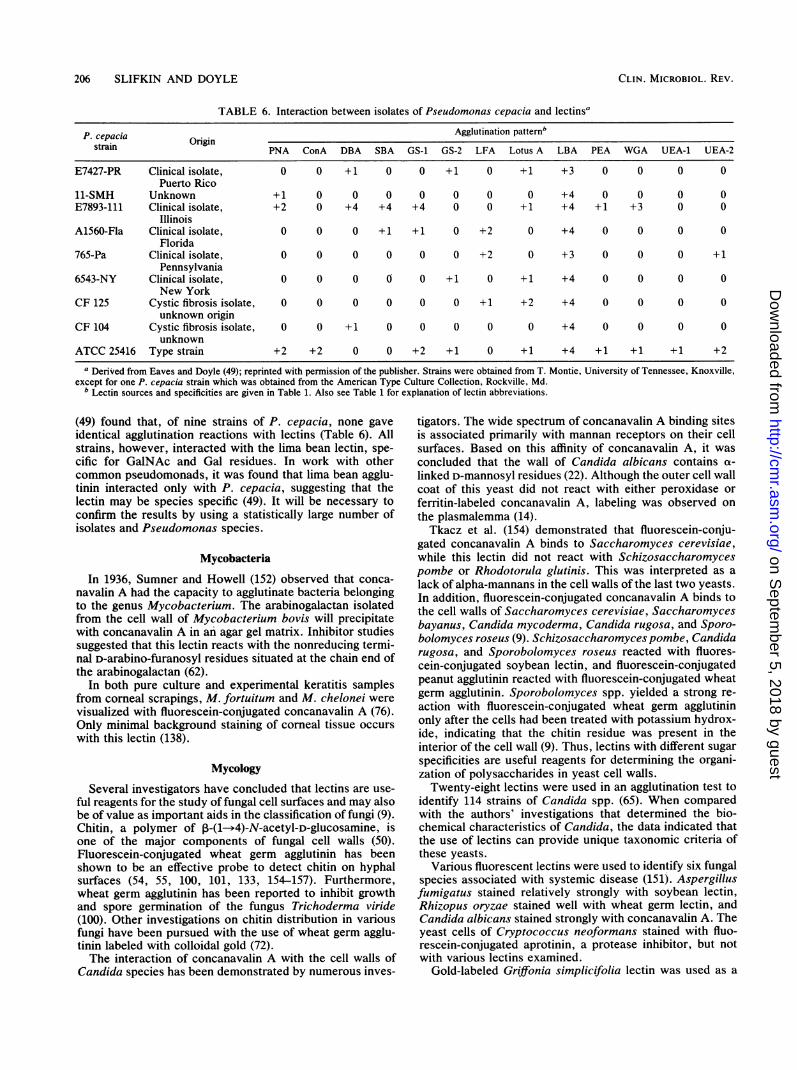
TABLE 6. Interaction between isolates of Pseudomonas cepacia and lectinsa
P. cepacia OrgnAgglutination patternsstrain Orlgln PNA ConA DBA SBA GS-1 GS-2 LFA Lotus A LBA PEA WGA UEA-1 UEA-2
E7427-PR Clinical isolate, 0 0 +1 0 0 +1 0 +1 +3 0 0 0 0Puerto Rico
11-SMH Unknown +1 0 0 0 0 0 0 0 +4 0 0 0 0E7893-111 Clinical isolate, +2 0 +4 +4 +4 0 0 +1 +4 +1 +3 0 0
IllinoisA1560-Fla Clinical isolate, 0 0 0 +1 +1 0 +2 0 +4 0 0 0 0
Florida765-Pa Clinical isolate, 0 0 0 0 0 0 +2 0 +3 0 0 0 +1
Pennsylvania6543-NY Clinical isolate, 0 0 0 0 0 +1 0 +1 +4 0 0 0 0
New YorkCF 125 Cystic fibrosis isolate, 0 0 0 0 0 0 +1 +2 +4 0 0 0 0
unknown originCF 104 Cystic fibrosis isolate, 0 0 +1 0 0 0 0 0 +4 0 0 0 0
unknownATCC 25416 Type strain +2 +2 0 0 +2 +1 0 +1 +4 +1 +1 +1 +2
a Derived from Eaves and Doyle (49); reprinted with permission of the publisher. Strains were obtained from T. Montie, University of Tennessee, Knoxville,except for one P. cepacia strain which was obtained from the American Type Culture Collection, Rockville, Md.
b Lectin sources and specificities are given in Table 1. Also see Table 1 for explanation of lectin abbreviations.
(49) found that, of nine strains of P. cepacia, none gaveidentical agglutination reactions with lectins (Table 6). Allstrains, however, interacted with the lima bean lectin, spe-cific for GalNAc and Gal residues. In work with othercommon pseudomonads, it was found that lima bean agglu-tinin interacted only with P. cepacia, suggesting that thelectin may be species specific (49). It will be necessary toconfirm the results by using a statistically large number ofisolates and Pseudomonas species.
Mycobacteria
In 1936, Sumner and Howell (152) observed that conca-navalin A had the capacity to agglutinate bacteria belongingto the genus Mycobacterium. The arabinogalactan isolatedfrom the cell wall of Mycobacterium bovis will precipitatewith concanavalin A in an agar gel matrix. Inhibitor studiessuggested that this lectin reacts with the nonreducing termi-nal D-arabino-furanosyl residues situated at the chain end ofthe arabinogalactan (62).
In both pure culture and experimental keratitis samplesfrom corneal scrapings, M. fortuitum and M. chelonei werevisualized with fluorescein-conjugated concanavalin A (76).Only minimal background staining of corneal tissue occurswith this lectin (138).
Mycology
Several investigators have concluded that lectins are use-ful reagents for the study of fungal cell surfaces and may alsobe of value as important aids in the classification of fungi (9).Chitin, a polymer of ,-(1->4)-N-acetyl-D-glucosamine, isone of the major components of fungal cell walls (50).Fluorescein-conjugated wheat germ agglutinin has beenshown to be an effective probe to detect chitin on hyphalsurfaces (54, 55, 100, 101, 133, 154-157). Furthermore,wheat germ agglutinin has been reported to inhibit growthand spore germination of the fungus Trichoderma viride(100). Other investigations on chitin distribution in variousfungi have been pursued with the use of wheat germ agglu-tinin labeled with colloidal gold (72).The interaction of concanavalin A with the cell walls of
Candida species has been demonstrated by numerous inves-
tigators. The wide spectrum of concanavalin A binding sitesis associated primarily with mannan receptors on their cellsurfaces. Based on this affinity of concanavalin A, it wasconcluded that the wall of Candida albicans contains ot-linked D-mannosyl residues (22). Although the outer cell wallcoat of this yeast did not react with either peroxidase orferritin-labeled concanavalin A, labeling was observed onthe plasmalemma (14).Tkacz et al. (154) demonstrated that fluorescein-conju-
gated concanavalin A binds to Saccharomyces cerevisiae,while this lectin did not react with Schizosaccharomycespombe or Rhodotorula glutinis. This was interpreted as alack of alpha-mannans in the cell walls of the last two yeasts.In addition, fluorescein-conjugated concanavalin A binds tothe cell walls of Saccharomyces cerevisiae, Saccharomycesbayanus, Candida mycoderma, Candida rugosa, and Sporo-bolomyces roseus (9). Schizosaccharomyces pombe, Candidarugosa, and Sporobolomyces roseus reacted with fluores-cein-conjugated soybean lectin, and fluorescein-conjugatedpeanut agglutinin reacted with fluorescein-conjugated wheatgerm agglutinin. Sporobolomyces spp. yielded a strong re-action with fluorescein-conjugated wheat germ agglutininonly after the cells had been treated with potassium hydrox-ide, indicating that the chitin residue was present in theinterior of the cell wall (9). Thus, lectins with different sugarspecificities are useful reagents for determining the organi-zation of polysaccharides in yeast cell walls.
Twenty-eight lectins were used in an agglutination test toidentify 114 strains of Candida spp. (65). When comparedwith the authors' investigations that determined the bio-chemical characteristics of Candida, the data indicated thatthe use of lectins can provide unique taxonomic criteria ofthese yeasts.
Various fluorescent lectins were used to identify six fungalspecies associated with systemic disease (151). Aspergillusfumigatus stained relatively strongly with soybean lectin,Rhizopus oryzae stained well with wheat germ lectin, andCandida albicans stained strongly with concanavalin A. Theyeast cells of Cryptococcus neoformans stained with fluo-rescein-conjugated aprotinin, a protease inhibitor, but notwith various lectins examined.
Gold-labeled Griffonia simplicifolia lectin was used as a
CLIN. MICROBIOL. REV.
on Septem
ber 5, 2018 by guesthttp://cm
r.asm.org/
Dow
nloaded from

LECTINS 207
probe to detect galactomannan on the cell walls of Schizo-saccharomyces pombe by scanning and transmission elec-tron microscopy (72). The lectin from the brown alga Fucusvesiculosus agglutinates Candida guilliermondii cells. Can-dida krusei cells weakly agglutinated in the presence of thislectin, whereas other species of Candida did not agglutinate(28).The patterns of staining of fungal cell walls with fluores-
cein-conjugated lectins are generally consistent with theestablished cell wall chemistries of the respective fungiexamined. For example, fluorescein-conjugated wheat germagglutinin, and the lectin from the asparagus pea Lotustetragonolobus, weakly stain Candida albicans (151). Thesereactions may be due to the presence of relatively smallamounts of N-acetylglucosamine and other cell wall residuesfor which these lectins have an affinity.The cell wall of Cryptococcus neoformans contains man-
nan and has no internal residues to which concanavalin Acan bind. Thus, in histologic sections, this fungus did notstain with fluorescein-conjugated concanavalin A (8, 9). Thecastor bean lectin (Ricinus communes), with its affinitytowards P-D-galactopyranose termini, was reported to labelthe cytoplasm and nuclei of the macrophages enveloping theyeast cells. However, the yeast did not label with this lectin.Doyle (unpublished observations) has determined that Ser-ratia marcescens lectin can differentiate Cryptococcus fromRhodotorula spp. When Rhodutorula spp. are heated to100'C, they become agglutinable by a Serratia marcescensstrain.
In tissue sections, fluorescein-conjugated wheat germ ag-glutinin was effective in labeling Rhizopus oryzae and fluo-rescein-conjugated concanavalin A, Ricinus communes, andwheat germ agglutinin stained Blastomyces dermatitidis(151). The binding of the wheat germ agglutinin was associ-ated with the presence of chitin in these organisms, whereasthe other lectins may stain components of galactomannans inthe Blastomyces cell. Soybean lectin was shown to beeffective in labeling Paracoccidiodes brasiliensis and As-pergillus fumigatus in tissue sections (151).The data just described indicate the value of using appro-
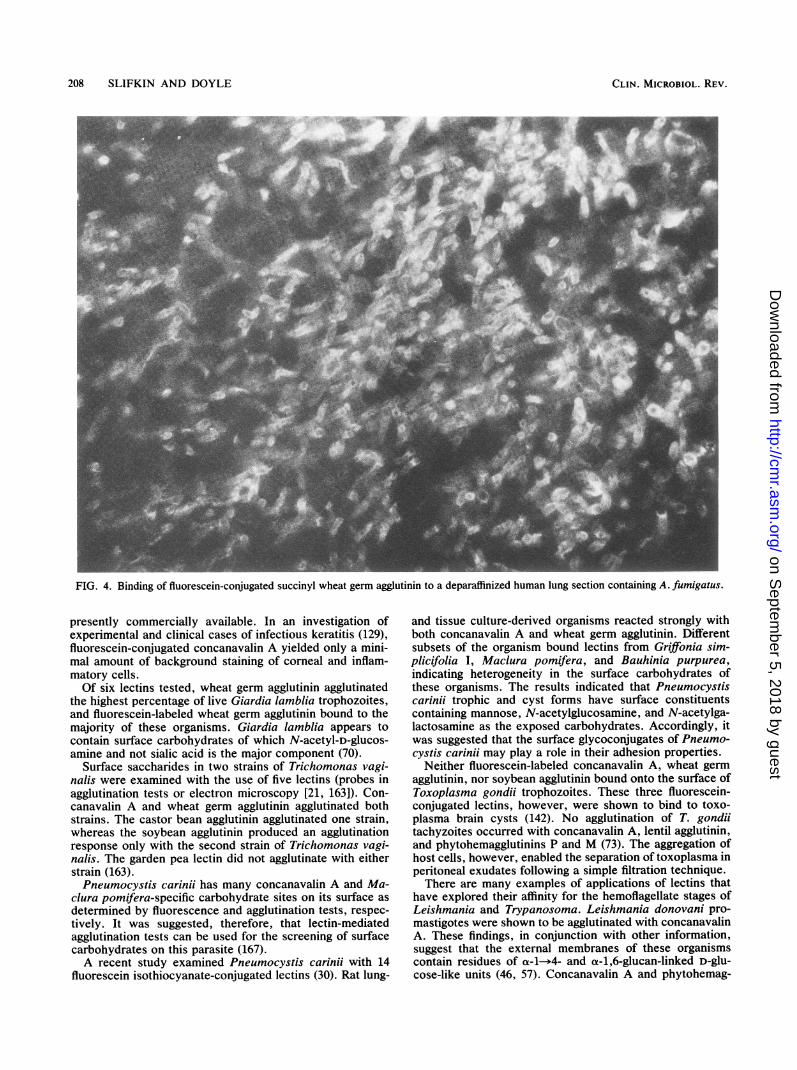
priate fluorescein-labeled lectins for detecting fungi in his-tologic sections of surgical and postmortem material. Theresults correlate with the known chemistry of the fungalwalls. Karayannopoulou et al. (79) used a battery of 21fluoresceinated lectins to examine tissue imprints, frozensections, and sections of paraffin-embedded human tissuesto determine which lectins were effective as diagnosticreagents for the microscopic diagnosis of fungal diseases.More lectins sustained fungi in frozen sections and imprints,indicating that paraffin embedding alters the reactivity offungal cellular glycoconjugates. Only receptors for wheatgerm, concanavalin A, and pokeweed mitogen were leftintact in the paraffin-embedded tissue. In tissue sectionsincubated with wheat germ agglutinin and concanavalin A,fungi were not easily discernible due to high, nonspecificbackground reactivity. In sections of lung, liver, and braintissue treated with pokeweed mitogen and succinylatedwheat germ agglutinin, lower background in deparaffinizedsections was observed. Although some background reactiv-ity was seen in bronchial, gastrointestinal, and renal tissue,fungi could be easily detected with the fluorescein-conju-gated lectins (Fig. 4). Succinylation of wheat germ agglutininreduces its affinity for sialic acid, while not affecting itsbinding to N-acetylglucosamine (102).The feasibility of applying fluorescein-conjugated lectins
to detect and differentiate fungi commonly involved in
ophthalmic mycoses was examined (128). Concanavalin Abrightly stained Candida albicans, Aspergillus fumigatus,and Fusarium solani. Fluorescein-conjugated wheat-germagglutinin and Ulex europaeus agglutinin stained only thelast two filamentous fungi. This was thought to be related tothe similar cell wall carbohydrate compositions of these twofungi (128). A panel of fluorescein-labeled lectins was used toobserve and differentiate a relatively wide spectrum of fungiassociated with ophthalmic infections (131). Concanavalin Abrightly stained all fungi examined. This lectin was consid-ered useful for the rapid visualization of fungi in ophthalmicspecimens due to its affinity for the mannose and a-D-glucans associated with some fungal cell walls (129). Severalgalactose-binding lectins were effective in staining Mucorspp., Rhizopus spp., and Acremonium killiense (131).Robin et al. (130) used a panel of fluorescein-conjugated
lectins and rapidly identified Candida albicans, Aspergillusfumigatus, and Fusarium solani grown in culture as well aspresent in tissue samples and fixed histopathologic speci-mens taken from experimental mycoses. Ulex europaeusagglutinin and wheat germ agglutinin were effective in dif-ferentiating Aspergillus and Fusarium from Candida spe-cies. A case of human keratomycosis was reported in whichthe use of fluorescein-conjugated concanavalin A was usedto visualize Candida albicans in scrapings from a cornealinfiltrate (130). The results allowed the rapid initiation ofappropriate anti-Candida therapy. This report illustrated thepotential for the clinical use of fluorescein-conjugated lectinsfor the detection of fungal pathogens associated with ocularinfections.
Parasitology
A large body of data exists on the interaction of lectinswith a relatively broad spectrum of parasites ranging fromthe protozoa through the metazoa. Although concanavalin Awas used by many investigators as a lectin probe for theseorganisms, many other lectins have been shown to be ofvalue in the study of cell surfaces and the identification anddifferentiation of the parasites. In some instances virulenceof parasitic protozoa appears to be related to their surfaceproperties, as revealed by interactions with lectins. Thus,several investigators have deemed important the comparisonof surface saccharides of parasites known to differ in theirvirulence traits.
It has been conjectured that the virulence of the tropho-zoite form of Entamoeba histolytica may depend, in part, onits surface properties. Data have been presented indicatingthat only strains isolated from cases of amoebic dysenteryagglutinate with concanavalin A (96); strains isolated fromasymptomatic cases of amoebic dysentery, however, do notagglutinate with this lectin.Two staining reagents, calcofluor white and fluorescein-
conjugated concanavalin A, have been reported to identifythe presence of Acanthamoeba organisms. Calcofluor white,which binds to chitin and cellulose, is used to visualize fungiin clinical specimens (129). Calcofluor has been used todetect Acanthamoeba spp. (128), but only the cyst stage ofthis organism reacts with the reagent. Surface glycoconju-gates containing either D-glucose or D-mannose are probablythe receptors for concanavalin A on these amoebae (129). Itappears that fluorescein-conjugated concanavalin A is moreeffective for the detection of Acanthamoeba spp. than cal-cofluor. Although immunofluorescent antibodies have beenreported to stain all stages of these amoebae with excellentspecificity (129), no antibodies against these organisms are
VOL. 3, 1990
on Septem
ber 5, 2018 by guesthttp://cm
r.asm.org/
Dow
nloaded from
208 SLIFKIN AND DOYLE
FIG. 4. Binding of fluorescein-conjugated succinyl wheat germ agglutinin to a deparaffinized human lung section containing A. fumigatus.
presently commercially available. In an investigation ofexperimental and clinical cases of infectious keratitis (129),fluorescein-conjugated concanavalin A yielded only a mini-mal amount of background staining of corneal and inflam-matory cells.Of six lectins tested, wheat germ agglutinin agglutinated
the highest percentage of live Giardia lamblia trophozoites,and fluorescein-labeled wheat germ agglutinin bound to themajority of these organisms. Giardia lamblia appears tocontain surface carbohydrates of which N-acetyl-D-glucos-amine and not sialic acid is the major component (70).
Surface saccharides in two strains of Trichomonas vagi-nalis were examined with the use of five lectins (probes inagglutination tests or electron microscopy [21, 163]). Con-canavalin A and wheat germ agglutinin agglutinated bothstrains. The castor bean agglutinin agglutinated one strain,whereas the soybean agglutinin produced an agglutinationresponse only with the second strain of Trichomonas vagi-nalis. The garden pea lectin did not agglutinate with eitherstrain (163).Pneumocystis caring has many concanavalin A and Ma-
clura pomifera-specific carbohydrate sites on its surface asdetermined by fluorescence and agglutination tests, respec-tively. It was suggested, therefore, that lectin-mediatedagglutination tests can be used for the screening of surfacecarbohydrates on this parasite (167).A recent study examined Pneumocystis carinii with 14
fluorescein isothiocyanate-conjugated lectins (30). Rat lung-
and tissue culture-derived organisms reacted strongly withboth concanavalin A and wheat germ agglutinin. Differentsubsets of the organism bound lectins from Griffonia sim-plicifolia I, Maclura pomifera, and Bauhinia purpurea,indicating heterogeneity in the surface carbohydrates ofthese organisms. The results indicated that Pneumocystiscarinji trophic and cyst forms have surface constituentscontaining mannose, N-acetylglucosamine, and N-acetylga-lactosamine as the exposed carbohydrates. Accordingly, itwas suggested that the surface glycoconjugates of Pneumo-cystis carinji may play a role in their adhesion properties.
Neither fluorescein-labeled concanavalin A, wheat germagglutinin, nor soybean agglutinin bound onto the surface ofToxoplasma gondii trophozoites. These three fluorescein-conjugated lectins, however, were shown to bind to toxo-plasma brain cysts (142). No agglutination of T. gondiitachyzoites occurred with concanavalin A, lentil agglutinin,and phytohemagglutinins P and M (73). The aggregation ofhost cells, however, enabled the separation of toxoplasma inperitoneal exudates following a simple filtration technique.There are many examples of applications of lectins that
have explored their affinity for the hemoflagellate stages ofLeishmania and Trypanosoma. Leishmania donovani pro-mastigotes were shown to be agglutinated with concanavalinA. These findings, in conjunction with other information,suggest that the external membranes of these organismscontain residues of a-1-*4- and a-1,6-glucan-linked D-glu-cose-like units (46, 57). Concanavalin A and phytohemag-
CLIN. MICROBIOL. REV.
on Septem
ber 5, 2018 by guesthttp://cm
r.asm.org/
Dow
nloaded from

LECTINS 209
glutinin reacted with Leishmania donovani promastigotes toyield a variety of agglutination patterns, including somatic-somatic, flagellar-somatic, and flagellar-flagellar agglutina-tions (46).
Pathogenic and noninfective strains of Leishmania enriet-tii were distinguished by their different cell surface carbohy-drates in reactions with concanavalin A (77, 138). This lectinwas shown to be of use in distinguishing between thepromastigotes of A1 and A2 subserotypes of Leishmaniaenriettii, whereas the lectin from Ricinus communis wasused to discern differences between the same subserotypes.Soybean lectin differentiated between the A and the Bserotypes. Various subserotypes of Leishmania enriettiiwere differentiated by peanut lectin (77). Leishmania brazil-iensis, however, does not agglutinate in the presence ofRicinus communis lectin but agglutinates with concanavalinA. Furthermore, agglutination was not observed with anystage of this organism with either soybean or wheat germagglutinin (33). Two glycoproteins in promastigotes of Leish-mania braziliensis were identified, using electrophoretic andlectin immunoblotting techniques (105). The cellular locationof these glycoproteins was determined by direct fluorescentconjugated lectin binding (33).Wheat germ agglutinin agglutinates the promastigotes of
Leishmania donovani and Leishmania enriettii (48, 117), butcompared with concanavalin A it only weakly agglutinatesLeishmania mexicana (18). In addition, wheat germ aggluti-nin does not agglutinate the promastigotes of Leishmaniabraziliensis (33). Thus, the nature of the lectin binding mayvary with the species of Leishmania. Peanut agglutinin waseffective in the separation of infective from noninfectiveforms of Leishmania major (124). Concanavalin A will reactwith Leishmania tropica (56).
Wisteriafloribunda lectin, peanut agglutinin, and soybeanagglutinin differentiated leishmanial isolates associated withvisceral, post-Kala-azar dermal, and cutaneous leishmania-sis, respectively, in a lectin-mediated agglutination test thatwas scored by microscopic examination of the reaction (57).Bloodstream and culture forms of Trypanosoma lewisi
were specifically agglutinated with the plant lectins conca-navalin A, soybean agglutinin, wheat germ agglutinin, andfucose-binding protein. The cumulative results of this inves-tigation indicated that ligands similar or identical to a-D-mannose, N-acetylgalactosamine, and N-acetylglucosamineand to a-L-fucose are constituents in the extracellular sur-face coat of this protozoan (47).Trypanosoma cruzi and T. cruzi-like strains can be easily
distinguished from Trypanosoma rangeli by an indirectimmunofluorescence test, using the lectin from the spongeAaptos papillata (19). This procedure was considered usefulas a diagnostic test. Accordingly, T. cruzi can be distin-guished from the nonpathogenic trypanosomes T. rangeliand T. conorhini with various radioisotopic labeled lectins.Tomato lectin and crude extracts of peanut were useful in
distinguishing T. conorhini from T. cruzi and T. rangeli.Tomato lectin, Sophora japonica, and Bauhinea purpureabound specifically with T. cruzi. Vicia villosa reacts with T.rangeli, whereas peanut, Ulex, and Lotus lectins bind withT. conorhini. Freshly squeezed tomato juice, after filtrationin gauze cloth and adjustment to pH 7.0, agglutinated T.cruzi epimastigotes, whereas the culture forms of T. rangeliand T. conorhini did not agglutinate (36).
Schottelius and Muller (140) established a method todifferentiate among culture forms of Trypanosma cruzi, T.rangeli, and T. conorhini with the application of lectins. T.rangeli was agglutinated by the D-mannose-specific lectins
concanavalin A and Pisum sativum. T. cruzi and T.conorhini, however, were distinguished by the lectin fromTridacna crocea.
T. cruzi reacts with Ricinus communis and Glycine maxlectins coupled with fluorescein or colloidal gold, whereasculture forms of T. rangeli do not (95). Investigations haveindicated that certain fluorescein-conjugated lectins are ofpractical use for specific detection of T. cruzi in the feces ofthe vector Rhodnius prolitus, thereby eliminating the prob-lems of false-positive results by fluorescence tests for T.cruzi (95). Epimastigote forms in many T. cruzi culturestocks were separated intraspecifically into two groups byagglutination reactions with 15 lectins (141). One groupreacted with Triticum vulgaris and Aaptos papillata lectins,and the second group was agglutinated by Arachis hypogaeaand Aaptos papillata lectins. Antigenic determinants corre-lated with these lectin reactions and glycoconjugate types.
Cell surface carbohydrates of various T. cruzi stages wereexamined by agglutination and lectin-binding assays (118).Trypomastigotes obtained from the blood of infected miceand from culture differ in their surface carbohydrates. Spe-cific receptors for wheat germ agglutinin, Helix pomatia,Sophora japonica, and Bandeiraea simplicifolia lectin IIwere found only in cultured epimastigotes, whereas peanutagglutinin sites were found only in cultured amastigotestages. Phaseolus vulgaris lectin had receptors for blood-stream trypanomastigotes and amastigotes, while Wisteriafloribunda hemagglutinin reacted predominately with cultureforms of T. cruzi.
Stock cultures of T. cruzi were agglutinated by the lectinsfrom Ricinus communis and Soja hipspida. T. cruzi but notT. rangeli was labeled by these lectins coupled with fluores-cein or colloidal gold. Only T. cruzi was agglutinated by thesialic acid-specific lectins from Aaptos papillata and Limaxflavus, and some stocks were also agglutinated by lectinfrom Triticum vulgaris. The lectins from Triticum vulgarisand Arachis hypogaea appear to be useful for intraspecificdifferentiation of T. cruzi. When a certain stock of T. cruzireacts with the Arachis lectin, it does not react with theTriticum lectin. These lectin reactions appear to be corre-lated with certain geographical distributions of the trypano-somes (139).
Epimastigotes of T. cruzi derived from culture agglutinatein the presence of low concentrations of concanavalin A.Blood forms (trypomastigotes) of this protozoan, however,do not agglutinate with concanavalin A, suggesting thatmembrane differences of T. cruzi stages might be related todifferent pathogenic properties (4). The insect Triatomainfestans is the host to T. cruzi and Blastocrithidia tri-atomae. Although the latter parasite does not infect humans,it may interfere with the identification of T. cruzi in thevector. The use of lectins to discriminate between these twoparasites has been suggested. T. cruzi binds to gold-labeledsoybean lectin, revealing N-acetylgalactosamine or galac-tose residues or both. On the other hand, Blastocrithidiatriatomae cell membranes bind with gold-labeled wheatgerm lectin, specific for N-acetylglucosamine (168).Both concanavalin A and wheat germ agglutinin were
reported to bind to the adult stage of Schistosoma mansoni(12). Receptors for concanavalin A and wheat germ aggluti-nin have been detected on the surfaces of all parasitic stagesof Schistosoma mansoni, whereas surface binding sites forthe fucose-specific lectin, Ulex europaeus, and soybeanagglutinin are absent. Little or no binding of these lectinswas observed on the free-living cercarial stages (103).
VOL. 3, 1990
on Septem
ber 5, 2018 by guesthttp://cm
r.asm.org/
Dow
nloaded from

210 SLIFKIN AND DOYLE
Virology
The unique property of lectins to bind noncovalently tosimple sugars and therefore to polysaccharides and glyco-conjugates has attracted the interest of virologists. In virol-ogy, lectins have been used for detection of viral glycopro-teins in purified and infected cells, as well as for viralpurification. Lectin studies have revealed information aboutthe structure of viral glycoproteins, structures important intheir pathogenicity. A significant contribution of lectin use invirology has been in the development of unique diagnosticmethods that yield specific identification of viral agents.
Purified influenza virus yields macroscopically visibleflocculation when mixed with concanavalin A (80). Wheninfluenza virus is treated with a proteolytic enzyme, theglycoprotein spikes of the virus are released. These treatedviral particles no longer agglutinate with this lectin, but willflocculate in the presence of N-acetylgalactosamine-asso-ciated lectins, such as Dolichos biflorus or Helix pomatia.Other viruses, including arboviruses, vesicular stomatitisvirus, paramyxoviruses, leukoviruses, and hepatitis B virus,also agglutinate with concanavalin A.Pea lectin did not bind to WSN influenza virus or New-
castle disease virus but did react with Sendai (parainfluenzaI) and B/Lee influenza viruses (85). Glycoproteins of Sendaivirus were isolated on a column of Helix pomatia lectin-Sepharose 6MB as a preparative step for the study of viralglycoproteins (10). This method can be used to select spe-cific protein classes and to isolate them in an intact form. Forexample, glycoprotein preparations of Rous sarcoma viruswere obtained for amino acid sequence analysis by Lensculinaris lectin-affinity chromatography (74). Fluorescein-conjugated Helix pomatia lectin was used to detect parain-fluenza I virus on the surface of the cell membrane of an
infected lymphoblastoid cell line (11). The presence of theglycoproteins was detectable 12 and 24 h after inoculation.
Various organelle-specific lectins were used to detectrapidly by light microscopy the intracellular location ofmembrane glycoproteins of Semliki Forest virus transport-defective mutants (78). When the cells were infected with a
temperature-sensitive mutant of this virus at a restrictivetemperature, cytoplasmic immunofluorescence was ob-served when fluorescein-conjugated concanavalin A was
applied to infected human embryonal fibroblasts. This sug-gested that the viral glycoproteins were arrested in theendoplasmic reticulum. When the cultures were incubated at28°C in the presence of cycloheximide and treated withrhodamine-conjugated wheat germ lectin, the arrested gly-coproteins appeared to be associated with the Golgi complexand plasma membrane.The potato lectin (Solanum tuberosum) and wheat germ
lectin were useful for the detection of glycoproteins of theplant rhabdoviruses potato yellow dwarf virus and eggplantmottled dwarf virus propagated in plants of Nicotiana rus-
tica (1). Lectin affinity for the viral proteins was examinedafter electrophoretic separation and transfer to nitrocellulosemembranes. No differences in the lectin-binding propertieswere observed for different viral glycoproteins of the tworhabdoviruses or between the various serotypes of eachvirus. Carbohydrate sequences common to viral glycopro-teins of these viruses may be present. The lectin assay forrhabdovirus glycoproteins in plants was reported to be about8 to 16 times less sensitive than assays with virus-specificantibodies.
Fluorescein-conjugated concanavalin A was effective indetection of Newcastle disease virus in BHK cells (120).
Viruses that possess envelope glycoproteins have beenshown to interact with the lectins concanavalin A and wheatgerm agglutinin with concomitant agglutination of the virus-infected host cells (20). Intracellular transport of SemlikiForest viral glycoproteins (78) and the insertion of Newcas-tle disease virus into the cell membrane (120) were observedwith the application of fluorescein-coupled lectins.
Biotinylated lectins, in conjunction with an avidin-biotin-peroxidase complex, were used to detect glycoproteins incells infected with prototype members of the avian herpes-virus groups, Marek's disease virus serotypes 1 and 2, andturkey herpesvirus (94).Herpes simplex virus types 1 and 2 (HSV-1 and HSV-2)
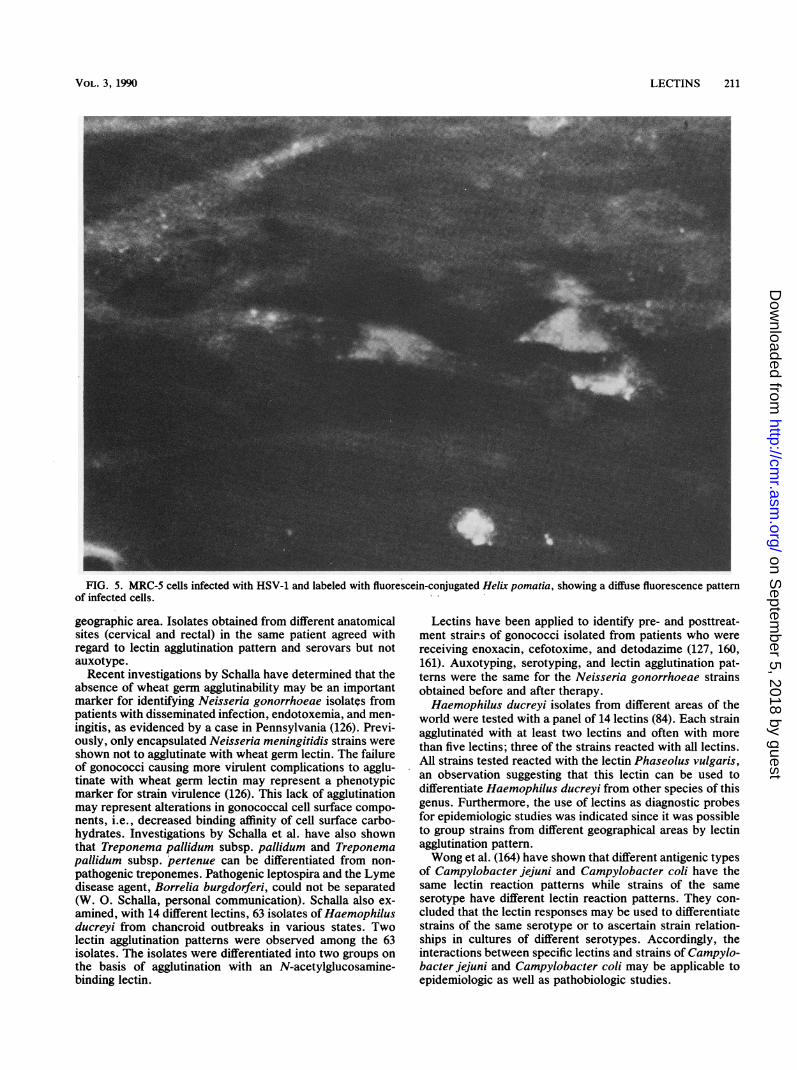
induce glycoproteins that bind to the N-acetylgalactosamine-specific Helix pomatia lectin (109-115). The glycoproteins ofHSV-infected cells have been purified by immunosorbenttechniques and by H. pomatia affinity chromatography ofthe protease-digested glycoproteins (92, 115). Fluorescein-conjugated H. pomatia lectin was shown to be as effective asfluorescein-conjugated monoclonal antibody reagents for thedetection and differentiation of HSV-1 and HSV-2 in MRC-5cell culture. Cells infected with HSV-1 generally displayed apattern of nongranular or diffuse fluorescence, whereas cellsinfected with HSV-2 were identified by the production offluorescent grains or flecks (145). In contrast to the immu-nofluorescence method, the serotypes of HSV were differ-entiated with only one fluorescein-conjugated H. pomatiareagent in MRC-5 cell cultures (Fig. 5 and 6).Concanavalin A was shown to block specifically adsorp-
tion of the bacteriophage binding sites of Bacillus subtilispossessing alpha-glucosylated teichoic acids in the cell walls(6) associated with teichoic acids. It was suggested that theapplication of this lectin might be useful as a means tocorrelated bacteriophage and serologic typing of staphylo-cocci (6).
USE OF LECTINS IN EPIDEMIOLOGICINVESTIGATIONS
Lectins have the ability to bind to a wide variety ofmicrobial substances containing carbohydrates. The pro-teins have been shown to be of value in the detection ofintrastrain variations in their respective cell wall carbohy-drate residues. Schalla and associates have shown thatlectins are useful in the epidemiological characterization ofNeisseria gonorrhoeae (135). They grouped Neisseria gon-orrhoeae isolates from various geographical regions basedon patterns of agglutination reactions with 14 lectins. A totalof 101 isolates with known nutritional requirements (auxo-types), plasmid content, and serovars were used in thatinvestigation. Among the 101 isolates tested, 24 differentagglutination groups were demonstrated. Of the organismstested, 55% were located within 3 of the 24 groups, and 86%of the isolates reacted with lectins derived from Trichosan-thes kinlowii, Griffonia simplicifolia, peanut, soybean, po-tato, and wheat germ. There were to be no differences inlectin groups that could be related to the sex of the patients.In one set of geographic isolates, one lectin group wasconfined to isolates of one particular serogroup, an associa-tion not seen for the other geographic areas. Some of theisolates with identical plasmid content, auxotype, and sero-var could be differentiated on the basis of lectin agglutinationpatterns, while other gonococcal isolates were identical byall test criteria.
In other investigation, Schalla et al. (135), examined 40Neisseria gonorrhoeae isolates obtained from the same
CLIN. MICROBIOL. REV.
on Septem
ber 5, 2018 by guesthttp://cm
r.asm.org/
Dow
nloaded from
LECTINS 211
FIG. 5. MRC-5 cells infected with HSV-1 and labeled with fluorescein-conjugated Helix pomatia, showing a diffuse fluorescence patternof infected cells.
geographic area. Isolates obtained from different anatomicalsites (cervical and rectal) in the same patient agreed withregard to lectin agglutination pattern and serovars but notauxotype.Recent investigations by Schalla have determined that the
absence of wheat germ agglutinability may be an importantmarker for identifying Neisseria gonorrhoeae isolates frompatients with disseminated infection, endotoxemia, and men-ingitis, as evidenced by a case in Pennsylvania (126). Previ-ously, only encapsulated Neisseria meningitidis strains wereshown not to agglutinate with wheat germ lectin. The failureof gonococci causing more virulent complications to agglu-tinate with wheat germ lectin may represent a phenotypicmarker for strain virulence (126). This lack of agglutinationmay represent alterations in gonococcal cell surface compo-nents, i.e., decreased binding affinity of cell surface carbo-hydrates. Investigations by Schalla et al. have also shownthat Treponema pallidum subsp. pallidum and Treponemapallidum subsp. pertenue can be differentiated from non-pathogenic treponemes. Pathogenic leptospira and the Lymedisease agent, Borrelia burgdorferi, could not be separated(W. 0. Schalla, personal communication). Schalla also ex-amined, with 14 different lectins, 63 isolates of Haemophilusducreyi from chancroid outbreaks in various states. Twolectin agglutination patterns were observed among the 63isolates. The isolates were differentiated into two groups onthe basis of agglutination with an N-acetylglucosamine-binding lectin.
Lectins have been applied to identify pre- and posttreat-ment strains of gonococci isolated from patients who werereceiving enoxacin, cefotoxime, and detodazime (127, 160,161). Auxotyping, serotyping, and lectin agglutination pat-terns were the same for the Neisseria gonorrhoeae strainsobtained before and after therapy.Haemophilus ducreyi isolates from different areas of the
world were tested with a panel of 14 lectins (84). Each strainagglutinated with at least two lectins and often with morethan five lectins; three of the strains reacted with all lectins.All strains tested reacted with the lectin Phaseolus vulgaris,an observation suggesting that this lectin can be used todifferentiate Haemophilus ducreyi from other species of thisgenus. Furthermore, the use of lectins as diagnostic probesfor epidemiologic studies was indicated since it was possibleto group strains from different geographical areas by lectinagglutination pattern.Wong et al. (164) have shown that different antigenic types
of Campylobacter jejuni and Campylobacter coli have thesame lectin reaction patterns while strains of the sameserotype have different lectin reaction patterns. They con-cluded that the lectin responses may be used to differentiatestrains of the same serotype or to ascertain strain relation-ships in cultures of different serotypes. Accordingly, theinteractions between specific lectins and strains of Campylo-bacter jejuni and Campylobacter coli may be applicable toepidemiologic as well as pathobiologic studies.
VOL. 3, 1990
on Septem
ber 5, 2018 by guesthttp://cm
r.asm.org/
Dow
nloaded from
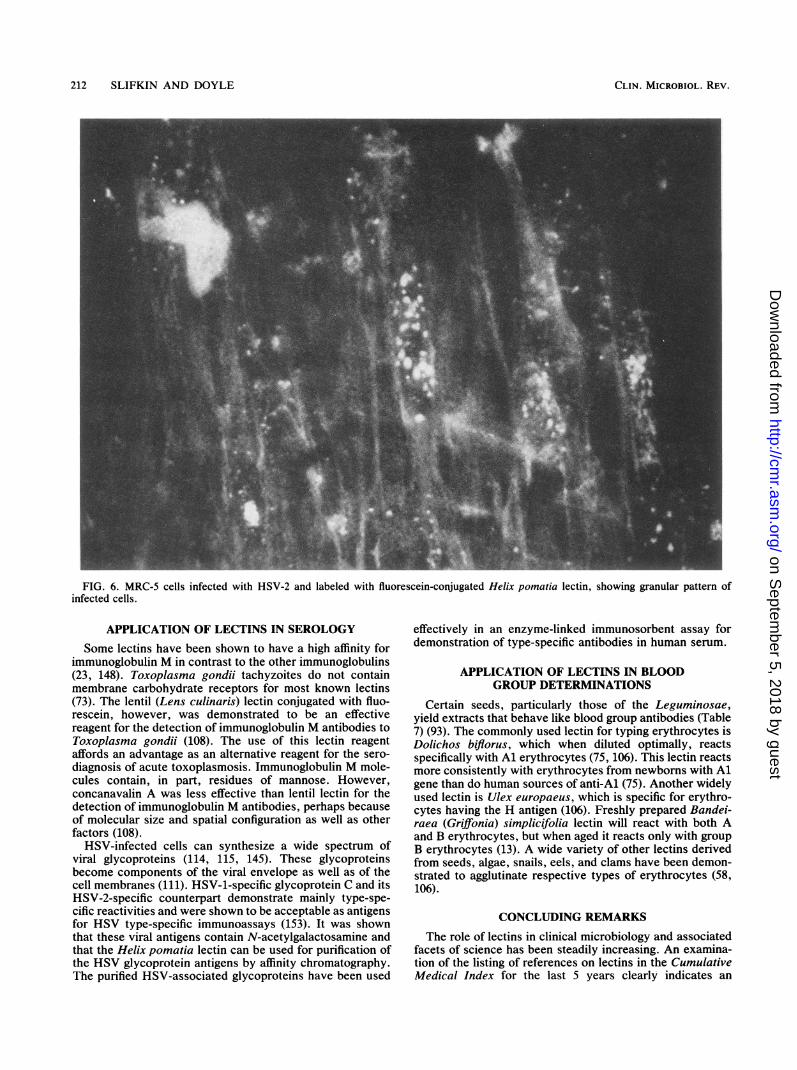
212 SLIFKIN AND DOYLE
FIG. 6. MRC-5 cells infected with HSV-2 and labeled with fluorescein-conjugated Helix pomatia lectin, showing granular pattern ofinfected cells.
APPLICATION OF LECTINS IN SEROLOGYSome lectins have been shown to have a high affinity for
immunoglobulin M in contrast to the other immunoglobulins(23, 148). Toxoplasma gondii tachyzoites do not containmembrane carbohydrate receptors for most known lectins(73). The lentil (Lens culinaris) lectin conjugated with fluo-rescein, however, was demonstrated to be an effectivereagent for the detection of immunoglobulin M antibodies toToxoplasma gondii (108). The use of this lectin reagentaffords an advantage as an alternative reagent for the sero-diagnosis of acute toxoplasmosis. Immunoglobulin M mole-cules contain, in part, residues of mannose. However,concanavalin A was less effective than lentil lectin for thedetection of immunoglobulin M antibodies, perhaps becauseof molecular size and spatial configuration as well as otherfactors (108).
HSV-infected cells can synthesize a wide spectrum ofviral glycoproteins (114, 115, 145). These glycoproteinsbecome components of the viral envelope as well as of thecell membranes (111). HSV-1-specific glycoprotein C and itsHSV-2-specific counterpart demonstrate mainly type-spe-cific reactivities and were shown to be acceptable as antigensfor HSV type-specific immunoassays (153). It was shownthat these viral antigens contain N-acetylgalactosamine andthat the Helix pomatia lectin can be used for purification ofthe HSV glycoprotein antigens by affinity chromatography.The purified HSV-associated glycoproteins have been used
effectively in an enzyme-linked immunosorbent assay fordemonstration of type-specific antibodies in human serum.
APPLICATION OF LECTINS IN BLOODGROUP DETERMINATIONS
Certain seeds, particularly those of the Leguminosae,yield extracts that behave like blood group antibodies (Table7) (93). The commonly used lectin for typing erythrocytes isDolichos biflorus, which when diluted optimally, reactsspecifically with Al erythrocytes (75, 106). This lectin reactsmore consistently with erythrocytes from newborns with Algene than do human sources of anti-Al (75). Another widelyused lectin is Ulex europaeus, which is specific for erythro-cytes having the H antigen (106). Freshly prepared Bandei-raea (Griffionia) simplicifolia lectin will react with both Aand B erythrocytes, but when aged it reacts only with groupB erythrocytes (13). A wide variety of other lectins derivedfrom seeds, algae, snails, eels, and clams have been demon-strated to agglutinate respective types of erythrocytes (58,106).
CONCLUDING REMARKS
The role of lectins in clinical microbiology and associatedfacets of science has been steadily increasing. An examina-tion of the listing of references on lectins in the CumulativeMedical Index for the last 5 years clearly indicates an
CLIN. MICROBIOL. REV.
on Septem
ber 5, 2018 by guesthttp://cm
r.asm.org/
Dow
nloaded from

LECTINS 213
TABLE 7. Some blood-group-specific lectinsa
Lectin Blood group specificity
Anguilla anguilla Anti-O(H)Arachis hypogaea Anti-TCystisus sessilifolius Anti-O(H) and anti-A2Dolichos biflorus Anti-AlGriffonia simplicifolia isolectin IA4 Anti-AGriffonia simplicifolia isolectin IB4 Anti-BHelix pomatia Anti-ALotus tetragonolobus Anti-O(H)Moluccella laevis Anti-ATnPhaseolus limensis Anti-APtilota plumosa Anti-BSaliva sclarea Anti-TnUlex europeus Anti-O(H)Ulva lectuca Anti-O(H)Vicia cracca Anti-AVicia graminea Anti-NVicia villosa Anti-A, + Tn
a Adapted from E-Y Laboratories circulars and Sigma Chemical Co.circulars.
increased use of lectins in scientific research. Due to theirgeneral abilities to bind specific glycoconjugates, lectins ofplant and animal origin have been used effectively withvarious microorganisms to correlate virulence with theirsurface properties. Investigators have shown that lectinbinding sites can shed light not only on the fundamentalquestion of the antigenicity of an organism, but also onselection of host environments by the organism.
Lectins are versatile reagents that can be applied in certaininstances to provide definitive identification and strain char-acterization of a particular infectious agent. Relatively sim-ple agglutination reactions can be used to detect the bindingactivity of a given lectin in many instances. However, thelectins are amenable to respective conjugation with a varietyof signals such as fluorescein, colloidal gold, ferritin, andradioisotopic markers. The soluble lectins can be bound topolystyrene spheres. Table 8 summarizes some of the meth-ods that have been used to study microorganism-lectininteractions.
Lectins may bind with receptors associated with theirhighest affinity. Accordingly, increasing incubation periodsof a lectin-organism mixture may increase the possibility forsecondary lectin reactivity. Thus, the use of long incubationtimes may be necessary in some instances to demonstratespecificity.To demonstrate the usefulness of a lectin in clinical
microbiology, it may first be necessary to use empiricaltesting procedures. For example, if one were interested indetermining whether a lectin or a series of lectins was ofvalue in the identification of group A streptococci, it wouldbe necessary initially to test those lectins known to haveaffinity for the primary receptor associated with the strepto-coccal surface. Thus, lectins with these specificities wouldbe selected first and tested, for example, by an agglutinationprocedure. A lectin with an established primary affinity forthe relevant antigenic determinant associated with group Astreptococci, however, may well interact with other sub-strates on cells of this serogroup. Published lectin affinitiesshould be considered only as an approximate guide of theirreactivity(s) (31). It is therefore obligatory to test othergroups of beta-hemolytic streptococci with a lectin found toreact with group A streptococcal organisms. It is importantto be aware that the nominal monosaccharide specificity of a
TABLE 8. How lectins have been used in clinical microbiology
Method Description
Direct agglutination Soluble lectins bind to a microbial surfaceIndirect agglutination Latex spheres are passively sensitized
with lectinsEnzyme-linked Lectin may bind to plastic or lectin may
lectinosorbent be used to complex with plastic-boundassay microbe or antigen
WELLA Western blot modified so a lectin canbind to a macromolecule in a gel
SELLA Salt-enhanced enzyme-linkedlectinosorbent assay; ammonium sulfatepromotes binding of lectin or proteinantigen to polystyrene
Fluorescence Fluorescent lectin is used to detectglycoconjugates or microbes
Colloidal gold Lectin-colloidal gold mixture detects lowdensities of microbes or low concn ofglycoconjugates
Enzyme assays Lectin is coupled to enzyme, such as -galactosidase or a peroxidase
Lectin-biotin Biotin-conjugated lectin may be detectedby avidin-enzyme conjugate
Isotope-lectin Lectin may be derivatized to contain 3H,14C, 131I, or 125i
Lectin in Lectin may detect one kind of bacterium,combination with but conventionalfluorogenic or fluorogenic/chromogenic substrate maychromogenic detect closely related bacteriasubstrate
lectin does not necessarily reflect on the complete nature ofthe carbohydrate that has the highest affinity for that lectin(31).When lectins are used for gaining diagnostic or surface
structure data, a set protocol must be applied that first allowsa given isolate to be placed in a particular group such as byGram-staining affinity or be characterized into a specificgenus. Thus, a lectin that reacts with group A streptococcibut not with isolates of other beta-hemolytic streptococcalserogroups could possibly react with an isolate that isEscherichia coli.The use of lectins in clinical microbiology is associated
with an extremely broad spectrum of advantages that renderthem of profound value in routine diagnostic procedures.Some of these significant advantages are (i) stability, (ii) theiractivity in small concentrations, (iii) the commercial avail-ability of many lectins, and (iv) their ability to probe subtlesurface structural differences between various isolates. Thecost effectiveness of these unique naturally occurring cy-tochemical-histochemical tools further emphasizes theirvalue as real and potential diagnostic reagents for the clinicalmicrobiologist.
This review article as well as other reviews on lectins (20,38, 60, 61, 69, 87, 89, 98, 99, 119, 143, 165; R. J. Doyle,Biotechs, E-Y Labs Inc., 1987; R. J. Doyle and K. F. Keller,Clin. Microbiol. Newsl. 8:157, 1986; R. J. Doyle and M.Slifkin, ASM News 55:655, 1989) should provide the readerwith the foundation needed to use published protocols or todevelop methods that use lectins for specific applications inclinical microbiology.
ACKNOWLEDGMENTSWork in the laboratory of R.J.D. relative to lectin-bacteria inter-
actions has been supported by the March of Dimes (Ohio Valley
VOL. 3, 1990
on Septem
ber 5, 2018 by guesthttp://cm
r.asm.org/
Dow
nloaded from

214 SLIFKIN AND DOYLE
Chapter), the International Life Sciences Institute, the U.S.Army R and D Command, and the National Institute for DentalResearch.We thank K. Shakanina and N. Kalinin of the Gamaleya Institute
and V. Lakhtin of the A. N. Bakh Institute for discussions.
LITERATURE CITED
1. Adam, G., P. Heegard, T. C. B0g-Hansen, and K. W. Mundry.1987. Lectins as probes for the assay of rhabdovirus infectionsin plants. J. Virol. Methods 17:263-275.
2. Aitchison, E. J., P. A. Lambert, and I. D. Farrell. 1986.Antigenic composition of an endocarditis-associated isolate ofStreptococcus faecalis and identification of its glycoproteinantigens by ligand blotting with lectins. J. Med. Microbiol.21:161-167.
3. Allen, P. Z., M. C. Connelly, and M. A. Apicella. 1980.Interactions of lectins with Neisseria gonorrhoeae. Can. J.Microbiol. 26:468-474.
4. Alves, M. J. M., and W. Colli. 1974. Agglutination of Trypa-nosoma cruzi by concanavalin A. J. Protozool. 21:575-578.
5. Archibald, A. R., and H. E. Coapes. 1971. The interaction ofconcanavalin A with teichoic acids and bacterial walls. Bio-chem. J. 123:665-667.
6. Archibald, A. R., and H. E. Coapes. 1972. Blocking of bacte-riophage receptor sites by concanavalin A. J. Gen. Microbiol.73:581-585.
7. Baker, C. J., and F. F. Barrett. 1974. Group B streptococcalinfections in infants: the importance of various serotypes. J.Am. Med. Assoc. 230:1158-1161.
8. Barkai-Golan, R., D. Mirelman, and N. Sharon. 1978. Studieson growth inhibition by lectins of penicillia and aspergilla.Arch. Microbiol. 116:119-124.
9. Barkai-Golan, R., and N. Sharon. 1978. Lectins as a tool for thestudy of yeast cell walls. Exp. Mycol. 2:110-113.
10. Bartzett, R., and F. J. Denaro. 1988. Detection of parainfluenzaI glycoproteins on cell membranes by means of lectin cy-tochemistry. J. Histotechnol. 11:91-92.
11. Bartzatt, R., and M. Martin. 1986. Isolation of glycoproteins ofparainfluenza I (Sendai) by Helix pomatia lectin column.Biosci. Rep. 6:591-595.
12. Bennet, J. L., and J. L. Seed. 1977. Characterization andisolation of concanavalin A binding sites from the epidermis ofS. mansoni. J. Parasitol. 63:250-288.
13. Bird, G. W. G. 1977. Lectins, p. 459-473. In D. Seligson (ed.),CRC handbook series in clinical laboratory science, sect. D.CRC Press,Inc., Cleveland.
14. Boiler, J., M. Durr, and A. Wienken. 1976. Asymmetricdistribution of concanavalin A binding sites on yeast plasma-lemma and vacuolar membrane. Arch. Microbiol. 109:115-118.
15. Boyd, W. C. 1954. Erythrocyte agglutinins, p. 755-839. In H.Neurath and K. Bailey (ed.), The proteins, vol. 2, part B.Academic Press, Inc., New York.
16. Boyd, W. C. 1963. The lectins: their present status. Vox Sang.8:1-32.
17. Boyd, W. C., and E. Shapleigh. 1954. Specific precipitatingactivity of plant agglutinins (lectins). Science 119:419.
18. Bray, R. S. 1983. Leishmania mexicana mexicana: attachmentand uptake of promastigotes to and by macrophages in vitro. J.Protozool. 30:314-322.
19. Bretting, H., and J. Schottelius. 1978. Immunofluoreszenzmik-roskopische Unterscheidung zwischem T. cruzi-, T. cruzi-like stammen, T. conorhini und T. rangeli mit dem Protektindes Schwammes Aaptos papillata. Z. Parasitenkd. 57:213-219.
20. Brown, J. C., and R. C. Hunt. 1972. Lectins. Int. Rev. Cytol.52:277-349.
21. Cappuccinelli, P., I. Cagliani, and G. Cavallo. 1975. Involve-ment of a surface concanavalin A-binding glycoprotein in theadhesion of Trichomonas vaginalis to substrates. Experientia31:1157-1159.
22. Cassone, A., E. Mattia, and L. Boldrini. 1978. Agglutination of
blastospores oF Candida albicans by concanavalin A and itsrelationship with the distribution of mannan polymers andthe ultrastructure of the cell wall. J. Gen. Microbiol. 105:263-273.
23. Chatelain, C., A. Lemoine, J. M. Dugouyon, M. Clanc, and P.Rouge. 1982. Interaction of polyclonal and monoclonal humanimmunoglobulins G with various lectins (Concanavalin A;lentil and pea lectins), p. 393-402. In T. C. B0g-Hansen (ed.),Lectins, biology, biochemistry, clinical biochemistry. Walterde Gruyter, New York.
24. Cifonelli, J. A., P. Rebers, M. B. Perry, and J. K. F. Jones.1966. The capsular polysaccharide of pneumococcus type XII,SXII. Biochemistry 5:3066-3072.
25. Cole, H. B., J. W. Ezzell, K. F. Keller, and R. J. Doyle. 1984.Differentiation of Bacillus anthracis and other Bacillus speciesby use of lectins. J. Clin. Microbiol. 19:48-53.
26. Connelly, M. C., and P. Z. Allen. 1983. Chemical and immu-nochemical studies on lipopolysaccharides from pyocin 103-sensitive and -resistant Neisseria gonorrhoeae. Carbohydr.Res. 120:171-186.
27. Corbel, M. J., and K. P. W. Gill. 1987. Lectin agglutination ofthermophilic Campylobacter species. Vet. Microbiol. 15:163-173.
28. Criado, M. T., and C. M. Ferreros. 1983. Selective interactionof a Fucus vesiculosus lectin-like mucopolysaccharide withseveral Candida species. Ann. Microbiol. (Paris) 134A:149-154.
29. Curtis, G. D. W., and M. P. E. Slack. 1981. Wheat-germagglutination of Neisseria gonorrhoeae. A laboratory investi-gation. Br. J. Vener. Dis. 57:252-255.
30. Cushion, M. T., J. A. DeStefano, and P. D. Walzer. 1988.Pneumocystis carinii: surface reactive carbohydrates detectedby lectin probes. Exp. Parasitol. 67:137-147.
31. Damjanov, I. 1987. Biology of disease. Lectin cytochemistryand histochemistry. Lab. Invest. 57:5-20.
32. Davidson, S. K., K. F. Keller, and R. J. Doyle. 1982. Differen-tiation of coagulase-positive and coagulase-negative staphylo-cocci by lectins and plant agglutinins. J. Clin. Microbiol.15:547-553.
33. Dawidowicz, K., A. G. Hernandez, R. B. Infante, and J. Corvit.1985. The surface membrane of Leishmania. I. The effects oflectins on different stages of Leishmania braziliensis. J. Para-sitol. 61:950-953.
34. deHormaeche, R. D., C. Burdell, H. Chong, D. W. Taylor, andP. Wildy. 1986. Definition of a virulence-related antigen ofNeisseria gonorrhoeae with monoclonal antibodies and lec-tins. J. Infect. Dis. 153:535-546.
35. DeLucca, A. J., 2nd. 1984. Lectin grouping of Bacillus thur-ingiensis serovars. Can. J. Microbiol. 30:1100-1104.
36. deMiranda Santos, I. K. F., and M. E. A. Pereira. 1984. Lectirtsdiscriminate between pathogenic and nonpathogenic SouthAmerican Trypanosoma. Am. J. Trop. Med. Hyg. 33:839-844.
37. Dietz, E., G. Bohmer, J. Solter, and G. Uhlenbruck. 1988.Adhesion of bacteria and tumors: inhibition by lectin blocking,p. 27-33. In T. C. B0g-Hansen and D. L. J. Freed (ed.),Lectins-biology, biochemistry, clinical biochemistry, vol. 6.Sigma Chemical Co., St. Louis.
38. Dorai, D. T., S. Srimal, S. Mohan, B. L. Bachhawat, and T. S.Balgenesh. 1982. Recognition of 2-keto-3-deoxyoctonate inbacterial cells and lipopolysaccharides by the sialic acid bind-ing lectin from horseshoe crab Carcinoscorpuis rotunda cau-dia. Biochem. Biophys. Res. Commun. 104:141-147.
39. Doyle, R., and K. Keller. 1984. Lectins in diagnostic microbi-ology. Eur. J. Clin. Microbiol. 3:4-9.
40. Doyle, R. J., and D. C. Birdsell. 1982. Interaction of concana-valin A with the cell wall of Bacillus subtilis. J. Bacteriol.109:652-658.
41. Doyle, R. J., K. F. Keller, and J. W. Ezzell. 1985. Bacillus,p. 211-215. In E. H. Lennette, A. Balows, W. J. Hausler, Jr.,and H. J. Shadomy (ed.), Manual of clinical microbiology,4th ed. American Society for Microbiology, Washington,D.C.
CLIN. MICROBIOL. REV.
on Septem
ber 5, 2018 by guesthttp://cm
r.asm.org/
Dow
nloaded from

LECTINS 215
42. Doyle, R. J., F. Nedjat-Haiem, K. F. Keller, and C. F. Frasch.1984. Diagnostic value of interactions between members of thefamily Neisseriaceae and lectins. J. Clin. Microbiol. 19:383-387.
43. Doyle, R. J., F. Nedjat-Haiem, R. D. Miller, and K. F. Keller.1982. Interaction between plant agglutinins and Legionellaspecies. J. Clin. Microbiol. 15:973-975.
44. Doyle, R. J., E. E. Woodside, and C. W. Fishel. 1968. Protein-polyelectrolyte interactions. The concanavalin A precipitinreaction with polyelectrolytes and polysaccharide derivatives.Biochem. J. 106:35-40.
45. Drake, D., K. G. Taylor, A. S. Bleiweis, and R. J. Doyle. 1988.Specificity of the glucan-binding lectin of Streptococcus crice-tus. Infect. Immun. 56:1864-1872.
46. Dwyer, D. M. 1974. Lectin binding saccharides of a parasiticprotozoan. Science 184:471-473.
47. Dwyer, D. M. 1976. Cell surface saccharides of Trypanosomalewisi. II. Lectin-mediated agglutination and fine-structurecytochemical detection of lectin-binding sites. J. Cell Sci.22:1-19.
48. Dwyer, D. M. 1977. Leishmania donovani: surface membranecarbohydrates of promastigotes. Exp. Parasitol. 41:341-358.
49. Eaves, D. J., and R. J. Doyle. 1988. Surface characteristics ofPseudomonas cepacia. Microbios 53:119-128.
50. Ebisu, S., J. Lonngren, and I. J. Goldstein. 1977. Interaction ofpneumococcal S-14 polysaccharide with lectins from Ricinuscommunes, Triticum vulgaris and Bandeiraea simplicifolia.Carbohydr. Res. 58:187-191.
51. Eshdat, Y., I. Ofek, Y. Yashouv-Gan, N. Sharon, and D.Mirehnan. 1978. Isolation of a mannose-specific lectin fromEscherichia coli and its role in the adherence of the bacteria toepithelial cells. Biochem. Biophys. Res. Commun. 85:1551-1560.
52. Etzler, M. E. 1986. Distribution and function of plant lectins, p.371-435. In I. E. Liener, N. Sharon, and I. J. Goldstein (ed.),The lectins, properties, functions, and applications in biologyand medicine. Academic Press, Inc., Orlando, Fla.
53. Frasch, E. C. 1980. Role of lipoplysaccharide in wheat germagglutinin-mediated agglutination of Neisseria meningitis andNeisseria gonorrhoeae. J. Clin. Microbiol. 12:498-501.
54. Galun, M., A. Braun, A. Frensdorf, and E. Galun. 1976.Hyphal walls of isolated lichen fungi: autoradiographic local-ization of precursor incorporation and binding of fluorescein-conjugated lectins. Arch. Microbiol. 108:9-16.
55. Galun, M., D. Malki, and E. Galun. 1981. Visualization ofchitin-wall formation in hyphal tips and anastomases of Diplo-dia natalensis by fluorescein-conjugated wheat-germ agglutininand [3H] N-acetyl-D-glucosamine. Arch. Microbiol. 130:105-110.
56. Gardiner, P. R., and D. M. Dwyer. 1983. Radioiodination andidentification of externally diseased membrane components ofLeishmania tropica. Mol. Biochem. Parasitol. 8:283-295.
57. Ghosh, D. K., A. K. Ghosh, and A. Bhattacharya. 1987.Differentiation of Leishmania strains of Indian origin by lectin-mediated aglutination. Trop. Med. Parasitol. 38:331-332.
58. Gilboa-Garber, N., R. Citronbaum, C. Levene, and R. Sela.1988. H-blood group detection by the L-fucose binding lectinsof the green marine alga Ulva lactuca. Dev. Comp. Immunol.12:695-705.
59. Glick, J., and N. Garber. 1985. Effects of growth phase andboiling of enteropathogenic Escherichia coli strains on theirinteraction with Pseudomonas aeruginosa lectins. Microbios43:73-86.
60. Goldstein, I. J., and C. E. Hayes. 1978. The lectins: carbohy-drate-binding proteins of plants and animals. Carbohydr.Chem. Biochem. 35:601-615.
61. Goldstein, I. J., R. C. Hughes, M. Monsigny, T. Osawa, and N.Sharon. 1980. What should be called a lectin? Nature (London)285:66.
62. Goldstein, I. J., and A. Misaki. 1970. Interaction of concana-valin A with an arabinogalactan from the cell wall of Myco-bacterium bovis. J. Bacteriol. 103:422-425.
63. Goldstein, I. J., and A. M. Staub. 1970. Interaction of conca-
navalin A with polysaccharides of Salmonellae. Immunochem-istry 7:315-319.
64. Graham, K., K. Keller, J. Ezzell, and R. Doyle. 1984. Enzyme-linked lectinosorbent assay (ELLA) for Bacillus anthracis.Eur. J. Clin. Microbiol. 3:210-212.
65. Guillot, J., M. Scandariato, and M. Coulet. 1974. Essai d'ap-plication du pouvoir agglutinant des lectins a 1 etude du genreCandida. Ann. Microbiol. (Paris) 125:489-500.
66. Hammarstrom, S., and E. A. Kabat. 1969. Purification andcharacterization of a blood group A reactive hemagglutininfrom the snail Helix pomatia and a study of its combining site.Biochemistry 8:2696-2705.
67. Hammarstrom, S., and E. A. Kabat. 1971. Studies on speci-ficity and binding properties of the blood group A reactivehemagglutinin from Helix pomatia. Biochemistry 10:1684-1692.
68. Hammarstrom, S., A. A. Lindberg, and E. S. Robertson. 1972.Precipitation of lipopolysaccharides from rough mutants ofSalmonella typhimurium by an A-hemagglutinin from Helixpomatia. Eur. J. Biochem. 25:274-282.
69. Hart, D. A. 1980. Lectins in biological systems: applications tomicrobiology. Am. J. Clin. Nutr. 33:2416-2425.
70. Hill, D. R., E. R. Hewlett, and R. D. Pearson. 1981. Lectinbinding by Giardia lamblia. Infect. Immun. 34:733-738.
71. Holm, S. E., A. Bergholm, D. Wager, and M. Wagner. 1985. Asialic-acid-specific lectin from Cepaea hortensis that promotesphagocytosis of a group-B, type la, streptococcal strain. J.Med. Microbiol. 19:317-323.
72. Horisberger, M., and J. Rosset. 1977. Localization of a-galactomannan on the surface of Schizosaccharomyces pombecells by scanning microscopy. Arch. Microbiol. 112:123-126.
73. Hoshino-Shimizu, S., J. R. Mineo, and M. E. Camargo. 1980.Lectin used in the purification process of Toxoplasma gondiitachyzoites. J. Parasitol. 66:989-991.
74. Hunter, E., E. Hill, M. Hardwick, A. Brown, D. E. Schwartz,and R. Tizard. 1983. Complete sequence of the Rous sarcomavirus env gene: identification of structural and functionalregions of its product. J. Virol. 46:920-936.
75. Issitt, P. D. 1985. Applied blood group serology, 3rd ed.Montgomery Scientific Publications, Miami.
76. Jackson, M., R. Chan, A. Y. Matoba, and J. B. Robin. 1989.The use of fluorescein-conjugated lectins for visualizing atyp-ical mycobacteria. Arch. Ophthalmol. 107:1206-1209.
77. Jacobson, R. L., G. M. Slutzky, C. L. Greenblatt, and L. F.Schnur. 1982. Surface resection of leishmania. I. Lectin-mediated agglutination. Ann. Trop. Med. Parasitol. 76:45-52.
78. Kaariainen, L., I. Virtanen, J. Saraste, and S. Keranen. 1983.Transport of virus membrane glycoproteins, use of tempera-ture-sensitive mutants and organelle-specific lectins. MethodsEnzymol. %:453-465.
79. Karayannopoulou, G., J. Weiss, and I. Damjanov. 1988. Detec-tion of fungi in tissue sections by lectin histochemistry. Arch.Pathol. Lab. Med. 112:746-748.
80. Klenk, H. D., H. Becht, and R. Rott. 1984. Reaction of virusesand virus-infected cells with heterophile agglutinins. Ann.N.Y. Acad. Sci. 234:355-367.
81. Kohler, W., and 0. Prokop. 1967. Agglutination von Strep-tokokken der Gruppe C durch ein Agglutinin aus Helixpomatia. Z. Immunitaetsforch. Allerg. Klin. Immunol. 133:50-53.
82. Kohler, W., and 0. Prokop. 1967. Agglutinationsversuchean Streptokokken mit dem Phytagglutinin aus Dolichos biflo-rus. Z. Immunitaetsforch. Allerg. Klin. Immunol. 133:171-175.
83. Kohler, W., 0. Prokop, and 0. Kuhnemund. 1973. Routineidentification of group-C streptococci by means of an aggluti-nin (protectin) from the albumen gland of the edible snail, Helixpomatia. J. Med. Microbiol. 6:127-130.
84. Korting, H. C., D. Abeck, A. P. Johnson, R. C. Ballard, D.Taylor-Robinson, and 0. Braun-Falco. 1988. Lectin typing ofHaemophilus ducreyi. Eur. J. Clin. Microbiol. Infect. Dis.
VOL. 3, 1990
on Septem
ber 5, 2018 by guesthttp://cm
r.asm.org/
Dow
nloaded from

216 SLIFKIN AND DOYLE
7:678-680.85. Krah, D. L., and P. W. Choppin. 1988. Selective inhibition of
WSN influenza virus haemolysis by pea lectin. J. Gen. Virol.69:717-722.
86. LeMinor, L., P. Tournier, and A. M. Chalon. 1973. Agglutin-abilite par la concanavaline a de divers bacilles a gram negatif.Etude chez les Salmonella de la correlation avec la structureantigenique 0. Ann. Microbiol. (Paris). 124A:467-476.
87. Liener, I. E., N. Sharon, and I. J. Goldstein (ed.). 1986. Thelectins: properties, functions and applications in biology andmedicine. Academic Press, Inc., Orlando, Fla.
88. Lim, D. V., W. J. Morales, A. F. Walsh, and D. Kazanis. 1986.Reduction of morbidity and mortality rates for neonatal groupB streptococcal disease through early diagnosis and chemopro-phylaxis. J. Clin. Microbiol. 23:489-492.
89. Lis, H., and N. Sharon. 1977. Lectins: their chemistry andapplication to immunology, p. 429-529. In M. Sela (ed.), Theantigens, vol. 4. Academic Press, Inc., New York.
90. Lis, H., and N. Sharon. 1986. Application of lectins, p. 293-370. In I. E. Liener, N. Sharon, and I. J. Goldstein (ed.), Thelectins: properties, functions and applications in biology andmedicine. Academic Press, Inc., Orlando, Fla.
91. Lis, H., and N. Sharon. 1986. Lectins as molecules and tools.Annu. Rev. Biochem. 55:35-67.
92. Lundstrom, M., S. Olfsson, S. Jeansson, E. Lycke, R. Datema,and J. Mansson. 1987. Host cell-induced difference in 0-glycosylation of herpes simplex virus gC-1. I. Structures ofnonsialylated HPA- and PNA-binding carbohydrates. Virology161:385-394.
93. Makela, D. 1957. Studies in hemagglutinins of leguminosaeseeds. Ann. Med. Exp. Fenn 35(Suppl. 11):1-130.
94. Malkinson, M., V. Orgad, and Y. Becker. 1986. Use of lectinsto detect and differentiate subtypes of Marek's disease virusand turkey herpes virus glycoproteins in tissue culture. J.Virol. Methods 19:129-133.
95. Marinkelle, C. J., G. A. Vallejo, J. Schottelius, F. Guhl, and N.de Sanchez. 1986. The affinity of the lectins Ricinus communisand Glycine maxima to carbohydrates on the cell surface ofvarious forms of Trypanosoma cruzi and Trypanosomarangeli, and the application of these lectins for the identifica-tion of T. cruzi in the feces of Rhodnius prolixus. Acta Trop.43:215-223.
96. Martinez-Palomo, A., A. Gonzales-Robles, and M. de la Torre.1973. Selective agglutination of pathogenic strains of Entamoe-ba histolytica induced by concanavalin A. Nature (London)New Biol. 245:186-187.
97. Meade, N. A., R. H. Staat, S. D. Langley, and R. J. Doyle. 1980.Lectin-like activity from Persea americana. Carbohydr. Res.78:349-363.
98. Mirelman, D. (ed.). 1986. Microbial lectins and agglutinins:properties and biological activity. John Wiley & Sons, Inc.,New York.
99. Mirelman, D., G. Altmann, and Y. Eshdat. 1980. Screening ofbacterial isolates for mannose-specific lectin activity by agglu-tination of yeasts. J. Clin. Microbiol. 11:328-331.
100. Mirelman, D., E. Galon, N. Sharon, and R. Lotan. 1975.Inhibition of fungal growth by wheat germ agglutinin. Nature(London) 236:414-417.
101. Molano, J., B. Bowers, and E. Cabib. 1980. Distribution ofchitin in the yeast cell wall. An ultrastructural and chemicalstudy. J. Cell Biol. 85:199-212.
102. Monsigny, M., A. C. Sene, R. Maget-Dana, and F. Delmotte.1980. Sugar-lectin interactions: how does wheat-germ aggluti-nin bind sialoglycoconjugates? Eur. J. Biochem. 104:147-153.
103. Murrell, K. D., D. W. Taylor, W. E. Vannier, and D. A. Dean.1978. Schistosoma mansoni: analysis of surface membranecarbohydrates using lectins. Exp. Parasitol. 46:247-255.
104. Nachbar, M. S., D. J. Oppenheim, and J. D. Thomas. 1980.Lectins in the U.S. diet. Isolation and characterization of alectin from the tomato (Lycopersicon esculentum). J. Biol.Chem. 225:2056-2061.
105. Nagakura, K., H. Tachibana, Y. Kaneda, and T. Nakae. 1986.Leishmania braziliensis: localization of glycoproteins in pro-
mastigotes. Exp. Parasitol. 61:335-342.106. Nance, S. T. 1986. Serology of the ABH and Lewis blood group
systems, p. 57-81. In M. E. Wallace and F. L. Gibbs (ed.),Blood group systems: ABH and Lewis. Associated BloodBanks, Arlington, Va.
107. Nicolson, G. L. 1974. The interactions of lectins with animalcell surfaces. Int. Rev. Cytol. 39:89-190.
108. Nojimoto, I. T. I., S. Hoshino-Shimizu, T. K. Hagasse-Suga-hara, and M. E. Camargo. 1987. Lectins for the detection of1gM antibodies to T. gondii in the diagnosis of acute toxoplas-mosis by immunofluorescence test. Rev. Inst. Med. Trop. SaoPaulo 29:354-360.
109. Norrild, B. 1980. Immunochemistry of herpes simplex virusglycoprotein. Curr. Top. Microbiol. Immunol. 90:67-106.
110. Norrild, B., S. L. Shore, T. L. Cromeans, and A. J. Nahmias.1980. Participation of three major glycoprotein antigens ofherpes simplex virus type 1 early in the infectious cycle asdetermined by antibody-dependent cell-mediated cytotoxicity.Infect. Immun. 28:38-44.
111. Olofsson, S., S. Jeansson, and E. Lycke. 1981. Unusual lectin-binding properties of a herpes simplex virus type 1-specificglycoprotein. J. Virol. 38:564-570.
112. Olofsson, S., M. Lundstrom, S. Jeansson, and E. Lycke. 1985.Different populations of herpes simplex virus glycoprotein Cdiscriminated by the carbohydrate-binding characteristics ofN-acetylgalactosamine specific lectins (soybean and Helixpomatia). Arch. Virol. 86:121-128.
113. Olofsson, S., M. Lundstrom, H. Marsden, S. Jeansson, and A.Vahlne. 1986. Characterization of a herpes simplex virus type2-specified glycoprotein with affinity for N-acetylgalac-tosamine-specific lectins and its identification as g92K or gG. J.Gen. Virol. 67:737-744.
114. Olofsson, S., B. Norrild, B. Andersen, L. Pereira, S. Jeansson,and E. Lycke. 1983. Populations of herpes simplex virusglycoprotein gC with and without affinity for the N-acetyl-galactosamine specific lectin of Helix pomatia. Arch. Virol.76:25-30.
115. Olofsson, S., I. Sjoblom, M. Lundstrom, S. Jeansson, and E.Lycke. 1983. Glycoprotein C of herpes simplex virus type 1:characterization of O-linked oligosaccharides. J. Gen. Virol.64:2735-2747.
116. Ottensooser, F., Y. Nakamizo, M. Sato, Y. Miyamoto, and K.Takizawa. 1974. Lectins detecting group C streptococci. 9:971-973.
117. Pardoe, G. I., R. Hahn, H. J. Jaquet, and W. Wojnarowski.1975. Immunochemistry of Leishmania enriettii surface anti-gen. Experientia 31:723-730.
118. Pereira, M. E. A., M. A. Loures, F. Villalta, and A. F. B.Andrade. 1980. Lectin receptors as markers for Trypanosomacruzi. Developmental stages and a study of the interaction ofwheatgerm agglutinin with sialic acid residues on epimastigotecells. J. Exp. Med. 152:1375-1392.
119. Pistole, T. G. 1981. Interaction of bacteria and fungi withlectins and lectin-like substances. Annu. Rev. Microbiol. 35:85-112.
120. Poste, G., and P. Reeve. 1984. Increased mobility and redistri-bution of concanavalin A receptors on cells infected withNewcastle disease virus. Nature (London) 247:469-471.
121. Pritchard, D. G., J. E. Coligan, S. E. Speed, and B. M. Gray.1981. Carbohydrate fingerprints of streptococcal cells. J. Clin.Microbiol. 13:89-92.
122. Pritchard, D. G., B. M. Gray, and H. C. Dillon, Jr. 1984.Characterization of the group-specific polysaccharide of groupB streptococcus. Arch. Biochem. Biophys. 235:385-392.
123. Prokop, D., and W. Kohler. 1967. Agglutinations reactions envon Mikroorganismen mit Helix pomatia Eiweiss Drusenex-trakt. Z. Immunitaetsforsch. Allerg. Klin. Immunol. 133:176-179.
124. Ready, P. D., and D. F. Smith. 1988. Peanut lectin agglutinationand isolation of infective forms of Leishmania major. Trans. R.Soc. Trop. Med. Hyg. 82:418.
125. Reeder, N. J., and R. D. Ekstedt. 1971. Study of the interactionof concanavalin A with staphylococcal teichoic acids. J. Im-
CLIN. MICROBIOL. REV.
on Septem
ber 5, 2018 by guesthttp://cm
r.asm.org/
Dow
nloaded from

LECTINS 217
munol. 106:334-340.126. Rice, R. J., W. 0. Schalla, J. W. Biddle, W. E. DeWitt, Y.
JeanLouis, M. S. McGlahn, G. Perkins, C. 0. Pasquariello, R.Swenson, E. Albrutyn, M. Goldberg, and W. L. Whittington.1986. Phenotypic characterization of Neisseria gonorrhoeaeisolated from three cases of meningitis. J. Infect. Dis. 153:362-365.
127. Rice, R. J., W. 0. Schalla, W. L. Whittington, J. W. Biddle, Y.JeanLouis, W. E. DeWitt, and S. E. Thompson. 1985. In-vestigation of Neisseria gonorrhoeae causing disseminatedinfection, endotoxemia, and meningitis and identification ofa possible virulence marker, p. 61-65. In G. K. Schoolnik,G. F. Brooks, S. Falkow, C. E. Frasch, J. S. Knapp,J. A. McCutchan, and S. A. Morse (ed.), The pathogenicneisseriae, American Society for Microbiology, Washington,D.C.
128. Robin, J. B., R. C. Arffa, I. Avni, and N. A. Rao. 1986. Rapidvisualization of three common fungi with fluorescein-conju-gated lectins. Invest. Ophthalmol. Vis. Sci. 27:500-506.
129. Robin, J. B., R. Chan, and B. R. Anderson. 1988. Rapidvisualization of Acanthamoeba using fluorescein-conjugatedlectins. Arch. Ophthalmol. 106:1273-1276.
130. Robin, J. B., S. Nielson, and M. D. Trousdale. 1986. Fluores-cein-conjugated lectin identification of a case of human kerat-omycosis. Am. J. Ophthalmol. 102:797-798.
131. Robin, J. B., L. Schmi0dt, T. Haimov, S. A. Nielsen, and J.Salazar. 1988. Fluorescein-conjugated lectin visualization ofinfections microorganisms, p. 485-489. In H. Caranaga (ed.),Proceedings of the Third World Congress on the Cornea.Raven Press, New York.
132. Sakakibara, F., G. Takayanagi, and H. Kawauchi. 1981. AnL-rhamnose-binding lectin in the eggs of Misgurnos anguilli-caudatus. Yakugaku-Zasshi 101:918-925. (In Japanese.)
133. Santos, E., J. R. Villanueva, and R. Sentandreu. 1978. Theplasma membrane of Saccharomyces cerevisiae. Isolation andsome properties. Biochim. Biophys. Acta 508:39-54.
134. Schaefer, R. L., K. F. Keller, and R. J. Doyle. 1979. Lectins indiagnostic microbiology: use of wheat germ agglutinin for thelaboratory identification of Neisseria gonorrhoeae. J. Clin.Microbiol. 10:669-672.
135. Schalla, W. D., R. J. Rice, J. W. Biddle, Y. Jean Louis, S. A.Larsen, and W. L. Whittington. 1985. Lectin characterizationof gonococci from an outbreak caused by penicillin-resistantNeisseria gonorrhoeae. J. Clin. Microbiol. 22:481-483.
136. Schalla, W. D., W. L. Whittington, R. J. Rice, and S. A.Larsen. 1985. Epidemiological characterization of Neisseriagonorrhoeae by lectins. J. Clin. Microbiol. 22:379-382.
137. Schaufuss, P., C. Lammler, and H. Blobel. 1986. Rapid differ-entiation of streptococci isolated from cows with mastitis. J.Clin. Microbiol. 24:1098-1099.
138. Schottelius, J. 1987. Selective lectin reactions of two stocks ofLeishmania enriettii with differing pathogenicity. Parasitol.Res. 73:1-8.
139. Schottelius, J., and H. Muhlpfordt. 1984. Carbohydrates asmarkers for Trypanosoma cruzi and Trypanosoma rangeli.Hoppe-Seyler's Z. Physiol. Chem. 365:1061.
140. Schottelius, J., and V. Muller. 1984. Interspecific differentia-tion of Trypanosoma cruzi, Trypanosoma conorhini and Try-panosoma rangeli by lectin in combination with complementlysis. Acta Trop. 41:29-38.
141. Schottelius, J., and G. Uhlenbruck. 1983. Comparative studiesof Trypanosoma cruzi and T. cruzi stocks from differentSouth American countries using lectins. J. Parasitenkd. 69:727-736.
142. Sethi, K. K., A. Rahman, B. Pelster, and H. Brandis. 1977.Search for the presence of lectin-binding sites on Toxoplasmagondii. J. Parasitol. 63:1076-1080.
143. Sharon, N., and H. Lis. 1989. Lectins. Chapman and Hall,Ltd., London.
144. Slifkin, M., and R. Cumbie. 1987. Identification of group Bstreptococcal antigen with lectin-bound polystyrene particles.J. Clin. Microbiol. 25:1172-1175.
145. Slifldn, M., and R. Cumbie. 1989. Rapid detection of herpes
simplex virus with fluorescein-labeled Helix pomatia lectin. J.Clin. Microbiol. 27:1036-1039.
146. Sliflin, M., and G. M. Gil. 1983. Rapid biochemical tests forthe identification of groups A, B, C, F, and G streptococci fromthroat cultures. J. Clin. Microbiol. 18:29-32.
147. Sliflin, M., and G. M. Gil. 1984. Identification of group Cstreptococcal antigen extracts with lectin-bound polystyreneparticles. J. Clin. Microbiol. 19:83-84.
148. Spengler, G. A., and R. M. Weber. 1981. Interactions of PHAwith human normal serum proteins, p. 231-240. In T. C.B0g-Hansen (ed.), Lectins, biology, biochemistry, clinicalbiochemistry. Walter de Gruyter, New York.
149. Staat, R. H., R. J. Doyle, S. D. Langley, and R. P. Suddick.1978. Modification of in vitro adherence of Streptococcusmutans by plant lectins. Adv. Exp. Med. Biol. 107:639-647.
150. Staat, R. H., S. D. Longley, and R. J. Doyle. 1980. Streptococ-cus mutans adherence: presumptive evidence for protein-mediated attachment followed by glucan-dependent cellularaccumulation. Infect. Immun. 27:675-681.
151. Stoddart, R. W., and B. M. Herbertson. 1978. The use offluorescein-labelled lectins in the detection and identification offungi pathogenic for man: a preliminary study. J. Med. Micro-biol. 11:315-324.
152. Sumner, J. B., and S. F. Howell. 1936. The identification of thehemagglutinin of the jack bean with concanavalin A. J. Bacte-riol. 32:227-237.
153. Svennerholm, B., S. Olofsson, S. Jeansson, A. Vahlneand, andE. Lycke. 1984. Herpes simplex virus type-selective enzyme-linked immunosorbent assay with Helix pomatia lectin-purifiedantigens. J. Clin. Microbiol. 19:235-239.
154. Tracz, J. S., E. B. Cybolska, and J. 0. Lampson. 1971. Specificstaining of wall mannan in yeast cells with fluorescein-conju-gated concanavalin A. J. Bacteriol. 105:1-5.
155. Tkacz, J. S., and J. 0. Lampen. 1972. Wall replication inSaccharomyces species: use of fluorescein-conjugated conca-navalin A to reveal the site of mannan insertion. J. Gen.Microbiol. 72:243-247.
156. Tronchin, G., D. Poulain, J. Herbaut, and J. Biguet. 1981.Localization of chitin in the cell wall of Candida albicans bymeans of wheat germ agglutinin. Fluorescence and ultrastruc-tural studies. Eur. J. Cell Biol. 26:121-128.
157. Tronchin, G., D. Poulain, J. Herbaut, and J. Biguet. 1981.Cytochemical and ultrastructural studies of Candida albicans.II. Evidence for a cell wall coat using concanavalin A. J.Ultrastruct. Res. 75:50-59.
158. Uhlenbruck, G., and 0. Prokop. 1966. An agglutinin from Helixpomatia which reacts with terminal N-acetyl-D-galactosamine.Vox Sang. 11:519.
159. Uppal, J. B. 1985. Use of Litchi chinesis lectins (agglutinins) indiagnostic microbiology. J. Pak. Med. Assoc. 35:110-111.
160. Van der Willigen, A. H., J. C. S. Van Der Hoek, J. H. T.Wagenvoort, H. J. A. Van Vliet, B. Van Klingeren, W. 0.Schalla, J. S. Knapp, T. Van Joost, M. F. Michel, and E. Stolz.1987. A comparative double-blind study of 200 and 400 mg ofenoxacin given orally in the treatment of acute uncomplicatedurethral gonorrhea in males. Antimicrob. Agents Chemother.31:535-538.
161. Van der Willigen, A. H., J. C. S. Van Der Hoek, J. H. T.Wagenroort, H. J. A. Van Vliet, B. Van Klingeren, W. 0.Schalla, J. W. Knapp, T. Van Joost, M. F. Michel, and E. Stolz.1987. Randomized comparative study of 0.5 and 1g of cefodi-zime (HR 221) versus 1 g of cefotaxime for acute uncompli-cated urogenital gonorrhea. Antimicrob. Agents Chemother.32:426-429.
162. Wagner, M. 1982. Agglutination of bacteria by a sialic-specificlectin of the snail Cepaea hortensis. Acta Histochem. 71:35-39.
163. Warton, A., and B. M. Honisberg. 1980. Lectin analysis ofsurface saccharides in two Trichomonas vaginalis strains dif-fering in pathogenicity. J. Protozool. 27:410-419.
164. Wong, K. H., S. K. Skelton, and J. C. Feeley. 1985. Inter-action of Campylobacter jejuni and Campylobacter coli withlectins and blood group antibodies. J. Clin. Microbiol. 22:134-135.
VOL. 3, 1990
on Septem
ber 5, 2018 by guesthttp://cm
r.asm.org/
Dow
nloaded from

218 SLIFKIN AND DOYLE
165. Wu, A. M., S. Sugii, and A. Herp. 1988. A guide for carbohy-drate specificities of lectins. Adv. Exp. Med. Biol. 228:819-847.
166. Yajko, D. M., A. Chu, and W. K. Hadley. 1984. Rapidconfirmatory identification of Neisseria gonorrhoeae with lec-tins and chromogenic substrates. J. Clin. Microbiol. 19:380-382.
167. Yoshikawa, H., T. Tegashi, and Y. Yoshida. 1987. Detection of
CLIN. MICROBIOL. REV.
surface carbohydrates on Pneumocystis carinii by fluorescein-conjugated lectins. Parasitol. Res. 74:43-49.
168. Zimmerman, D., W. Peters, and G. A. Schaub. 1987. Differ-ences in binding of lectin-gold conjugates by Trypanosomacruzi and Blastocrithidia triatomae (Trypanosomatidae) in theintestine of Triatoma infestans (Reduviidae) Parasitol. Res.74:5-10.
on Septem
ber 5, 2018 by guesthttp://cm
r.asm.org/
Dow
nloaded from