lecture 4: short- distance transport of nutrients · 2/16/2016 5 2. cell wall pores primary cell...
TRANSCRIPT
2/16/2016
1
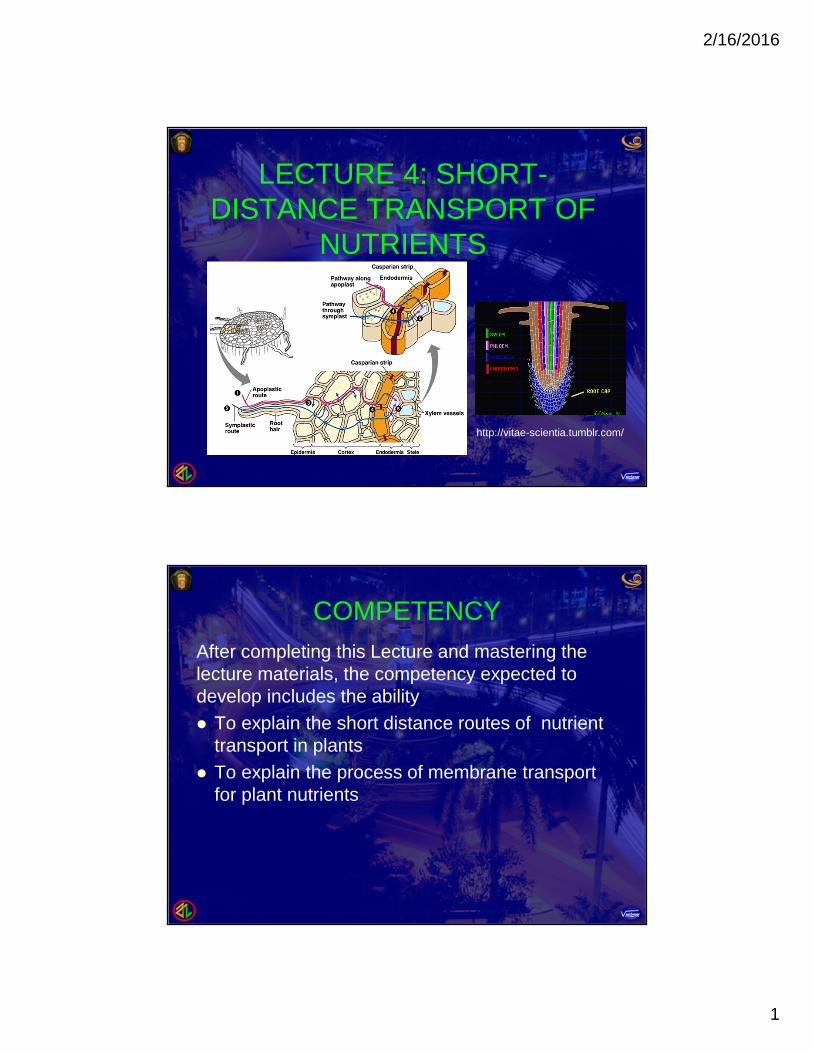
LECTURE 4: SHORT-DISTANCE TRANSPORT OF
NUTRIENTS
http://vitae-scientia.tumblr.com/
COMPETENCYAfter completing this Lecture and mastering thelecture materials, the competency expected todevelop includes the ability To explain the short distance routes of nutrient
transport in plants To explain the process of membrane transport
for plant nutrients

2/16/2016
2
LECTURE FLOW Transport Routes Symplast pathway Apoplast pathway TransmembranePore sizeWFS & DFSCEC of Roots
Membrane transportMembrane Characteristics Downhill and Uphill transport Proton pumps
TRANSPORT PATHWAY
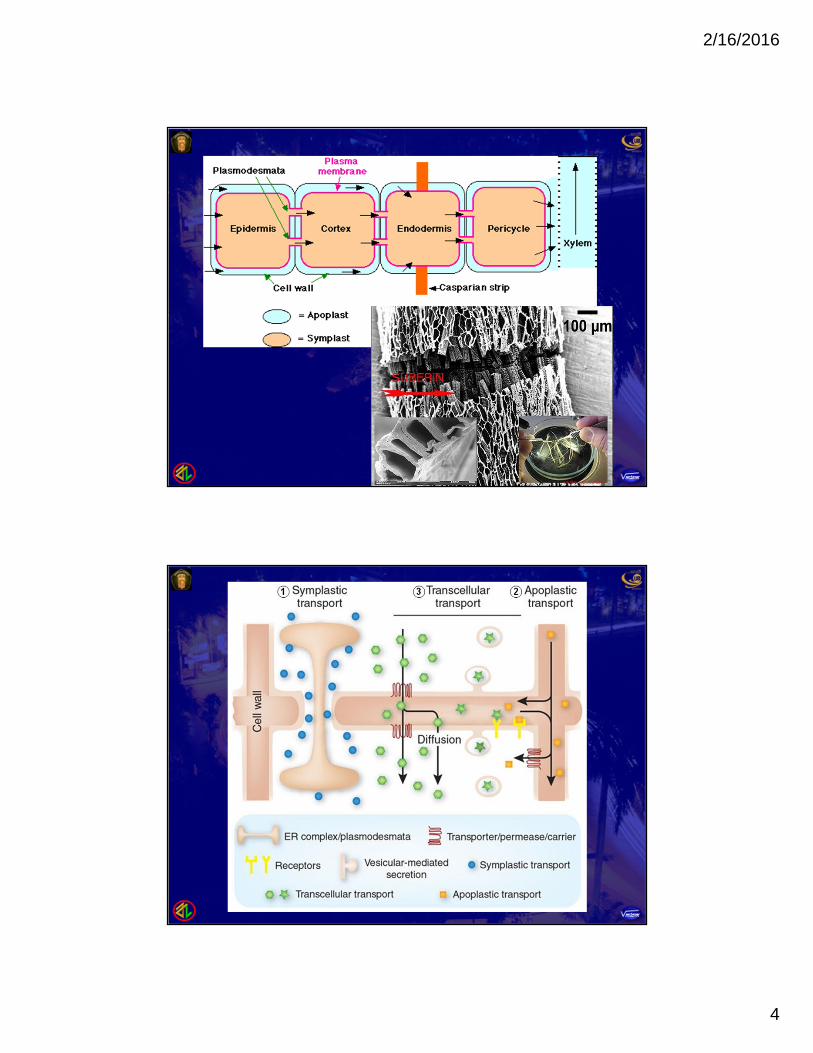
1. TRANSPORT ROUTES Three routes are available for lateral transport, the
movement of water and solutes from one location toanother within plant tissues and organs.a. Symplast routeb. Apoplast routec. Transmembrane route
a. The Symplast route is the route via thesymplast that requires only one crossing of aplasma membrane. After entering one cell, solutes and water move from
cell to cell via plasmodesmata.
2/16/2016
3
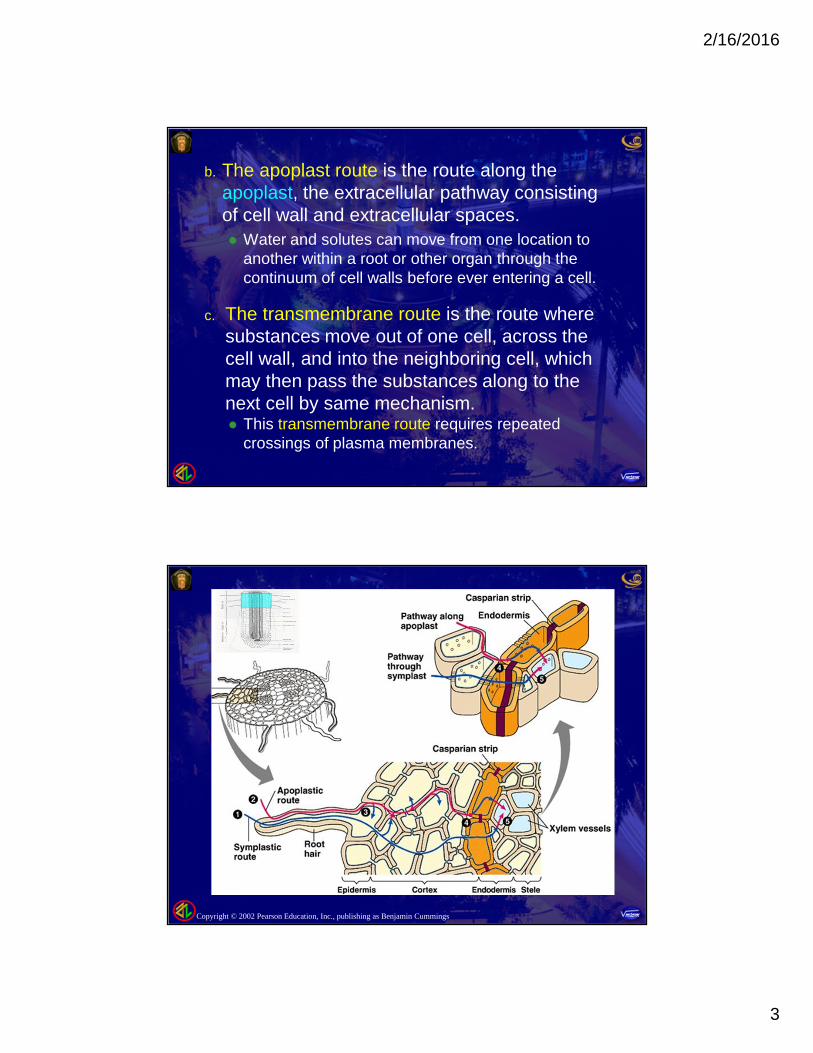
b. The apoplast route is the route along theapoplast, the extracellular pathway consistingof cell wall and extracellular spaces. Water and solutes can move from one location to
another within a root or other organ through thecontinuum of cell walls before ever entering a cell.
c. The transmembrane route is the route wheresubstances move out of one cell, across thecell wall, and into the neighboring cell, whichmay then pass the substances along to thenext cell by same mechanism. This transmembrane route requires repeated
crossings of plasma membranes.
Copyright © 2002 Pearson Education, Inc., publishing as Benjamin Cummings
2/16/2016
4
2/16/2016
5
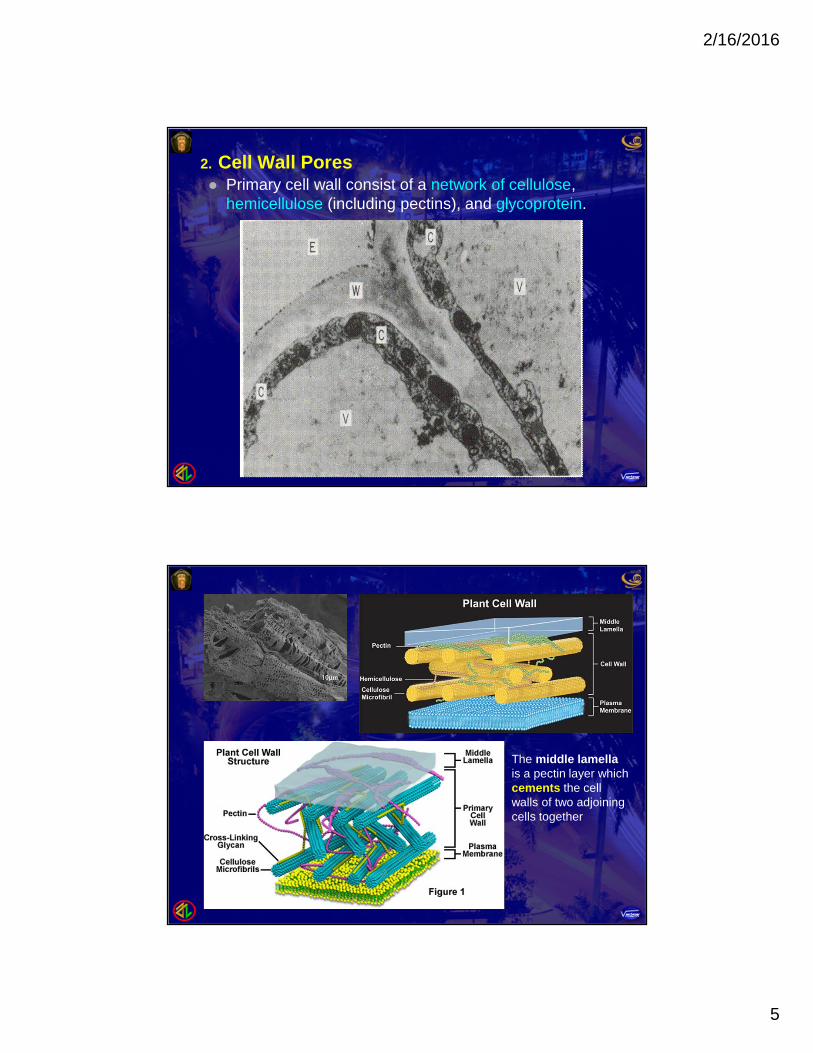
2. Cell Wall Pores Primary cell wall consist of a network of cellulose,
hemicellulose (including pectins), and glycoprotein.
The middle lamellais a pectin layer whichcements the cellwalls of two adjoiningcells together
2/16/2016
6
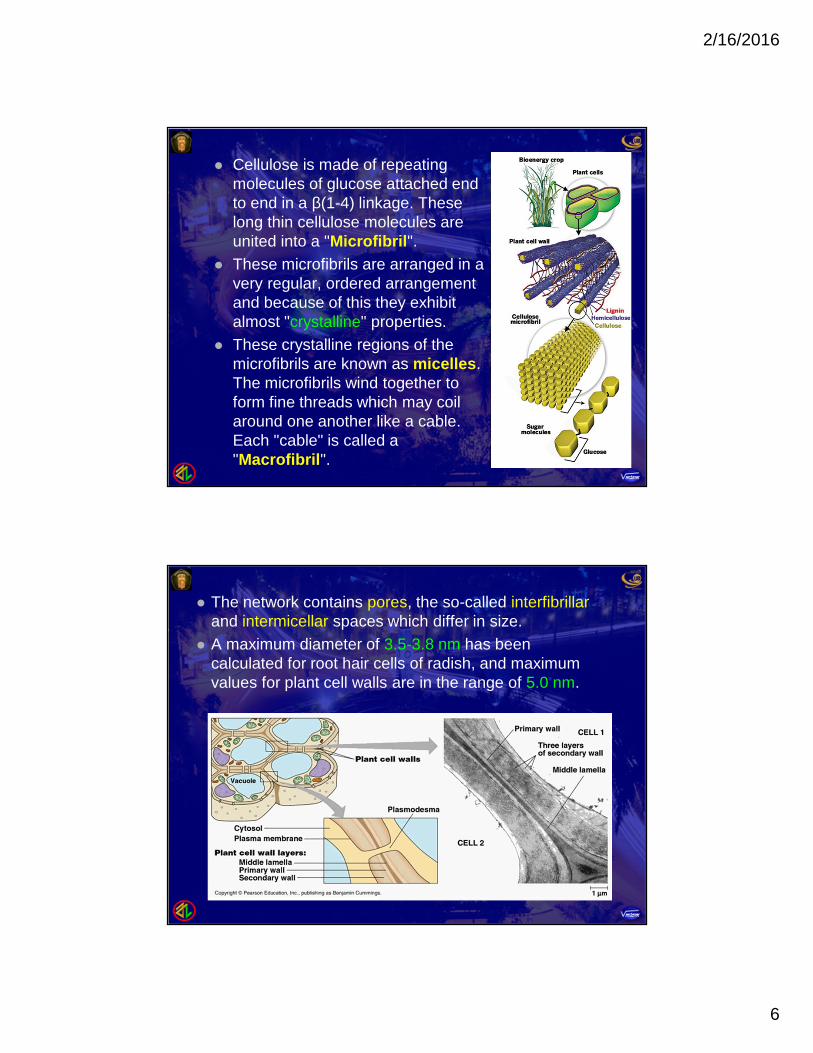
Cellulose is made of repeatingmolecules of glucose attached endto end in a β(1-4) linkage. Theselong thin cellulose molecules areunited into a "Microfibril".
These microfibrils are arranged in avery regular, ordered arrangementand because of this they exhibitalmost "crystalline" properties.
These crystalline regions of themicrofibrils are known as micelles.The microfibrils wind together toform fine threads which may coilaround one another like a cable.Each "cable" is called a"Macrofibril".
The network contains pores, the so-called interfibrillarand intermicellar spaces which differ in size.
A maximum diameter of 3.5-3.8 nm has beencalculated for root hair cells of radish, and maximumvalues for plant cell walls are in the range of 5.0 nm.

2/16/2016
7
Materials Diameter (nm)Rhizodermal cell wall 500-3000Cortical cell wall 100-200Pores in cell wall <5Sucrose 1.0Hydrated ions• K+ 0.66• Ca2+ 0.82
The dimensions of hydrated ions such as K+ and Ca2+
are small compared with the diameter of these pores,thus the pores themselves should not restrict themovement of these ions within the free space
The volume of roots available for the passive transport(free space) is about 10% of total space in youngroots
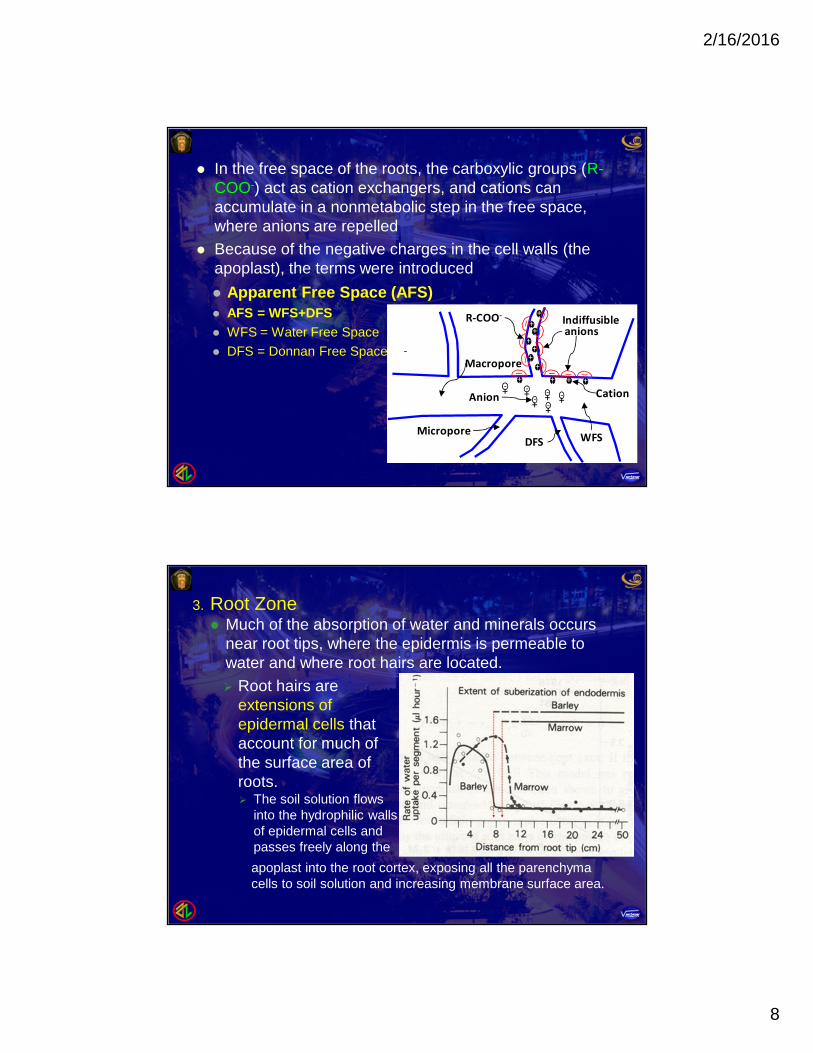
In the free space of the roots, the carboxylic groups (R-COO-) act as cation exchangers, and cations canaccumulate in a nonmetabolic step in the free space,where anions are repelled
Plant species differ considerably in their cation exchangecapacity (CEC), that is the number of cation exchangesites in their cell walls
Plant species CEC (meq/100 gdry weight)
Wheat 23Maize 29Bean 54
Tomato 62CEC = Cation-Exchange Capacity
As a rule, the CEC ofdicotyledonous speciesis much higher thanthat ofmonocotyledonousspecies
2/16/2016
8
In the free space of the roots, the carboxylic groups (R-COO-) act as cation exchangers, and cations canaccumulate in a nonmetabolic step in the free space,where anions are repelled
Because of the negative charges in the cell walls (theapoplast), the terms were introduced
-
++++
++++++++
++ ++ ++ ++
++
-+-+
-+-+
-+-+ CationAnion
Macropore
MicroporeDFS WFS
Indiffusibleanions
R-COO-
Apparent Free Space (AFS) AFS = WFS+DFS WFS = Water Free Space DFS = Donnan Free Space
3. Root Zone Much of the absorption of water and minerals occurs
near root tips, where the epidermis is permeable towater and where root hairs are located. Root hairs are
extensions ofepidermal cells thataccount for much ofthe surface area ofroots. The soil solution flows
into the hydrophilic wallsof epidermal cells andpasses freely along theapoplast into the root cortex, exposing all the parenchymacells to soil solution and increasing membrane surface area.

2/16/2016
9
1. Chemical Composition Membranes are typically composed of two classes of
compounds: protein and lipid. Carbohydrate compriseonly a minor fraction of membrane
Its framework consists of a double layer ofphospholipids.
PLASMA MEMBRANE
The major types ofproteins are tightlycoiled, rod-shaped,fibrous proteins, andthe more compact,globular-shapedintegral protein andpheripheral proteins
Lipid utama dari Membran: phospholipid,glucolipid & sulfolipidLipid utama lain : Sterol khususnyacholesterol pada khewan, dan b-sistosterol pada tanaman

2/16/2016
10
2. Permeability Because the cell membrane is mostly lipid, it only
allows lipid-soluble substances (e.g. oxygen, carbondioxide, and steroids) to go through. Water-solublesubstances (e.g glucose, amino acids, ions, andwater) need the help of proteins transporters
Permeability (cm.s-1)
Therefore, the main sites of selectivity in the uptake ofcations and anions as well as solutes in general arelocated in the plasma membrane of individual cells

2/16/2016
11
This can be seen that most of the Ca2+ (45Ca) taken upwithin 30 min (influx) is still readily exchangeable (efflux)and is almost certainly located in the AFS
In contrast, only a minor fraction o0f the K+ (42K) is readilyexchangeable within the 30-min period, most of the K+
having already beentransported acrossthe membranesinto the cytoplasmand vacuoles(“inner space”)
Downhill & Uphill transport
3. Passive and Active Transport
from Knox, Ladiges & Evans

2/16/2016
12
4. Proton pumps Proton pumps play a central role in transport across
plant membranes The most important active transporter in the plasma
membrane of plant cells is the proton pump.
Copyright © 2002 Pearson Education, Inc., publishing as Benjamin Cummings
1. It hydrolyzes ATP anduses the releasedenergy to pumphydrogen ions (H+)out of the cell.
2. This creates a protongradient because theH+ concentration ishigher outside the cellthan inside.
2/16/2016
13
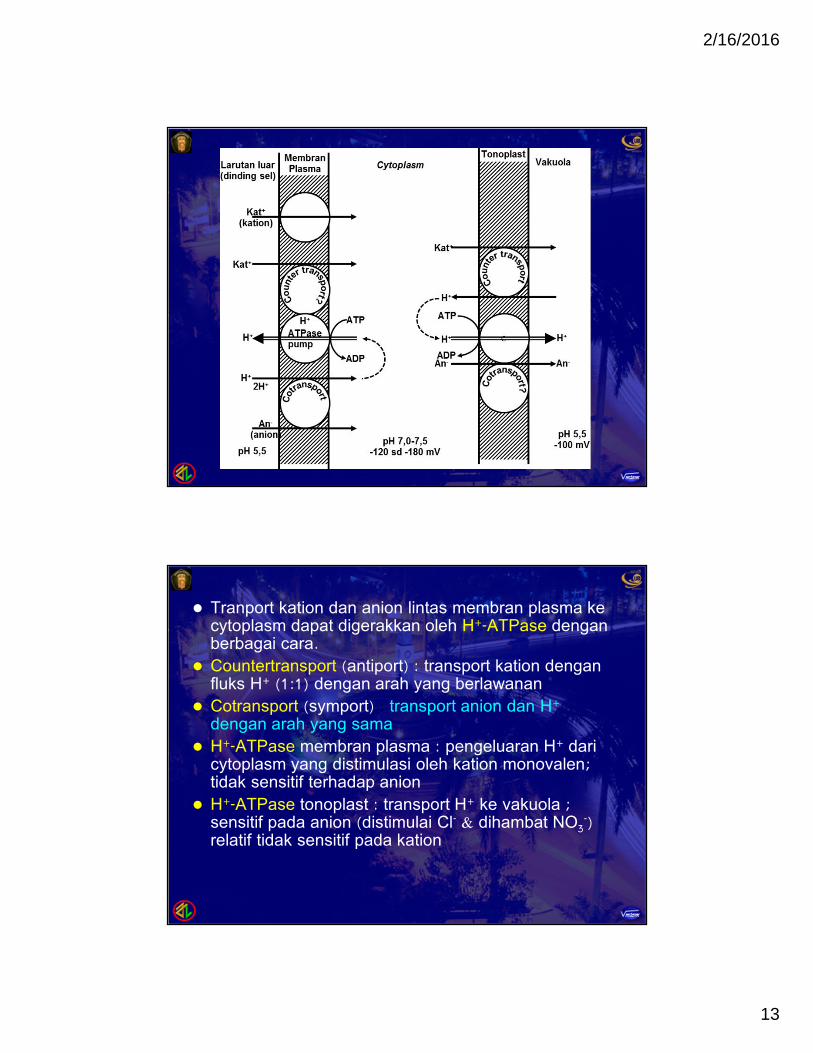
Tranport kation dan anion lintas membran plasma kecytoplasm dapat digerakkan oleh H+-ATPase denganberbagai cara.
Countertransport (antiport) : transport kation denganfluks H+ (1:1) dengan arah yang berlawanan
Cotransport (symport) : transport anion dan H+
dengan arah yang sama H+-ATPase membran plasma : pengeluaran H+ dari
cytoplasm yang distimulasi oleh kation monovalen;tidak sensitif terhadap anion
H+-ATPase tonoplast : transport H+ ke vakuola ;sensitif pada anion (distimulai Cl- & dihambat NO3
-)relatif tidak sensitif pada kation
2/16/2016
14
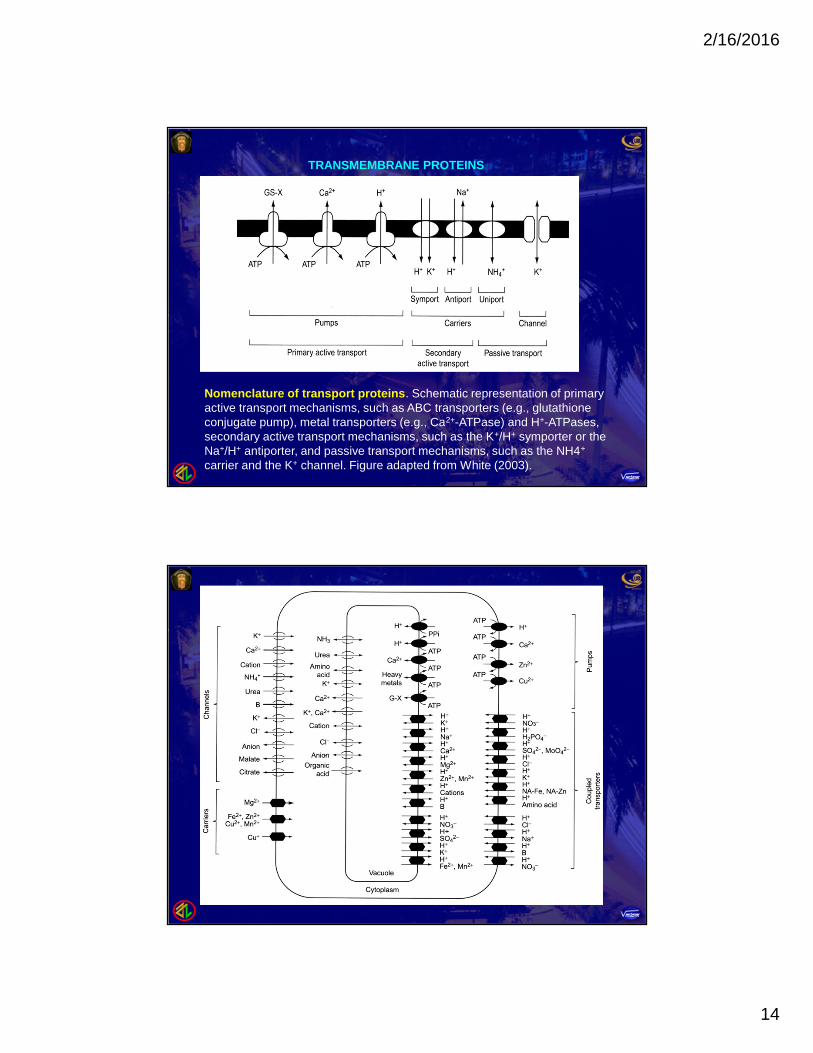
Nomenclature of transport proteins. Schematic representation of primaryactive transport mechanisms, such as ABC transporters (e.g., glutathioneconjugate pump), metal transporters (e.g., Ca2+-ATPase) and H+-ATPases,secondary active transport mechanisms, such as the K+/H+ symporter or theNa+/H+ antiporter, and passive transport mechanisms, such as the NH4+
carrier and the K+ channel. Figure adapted from White (2003).
TRANSMEMBRANE PROTEINS
2/16/2016
15
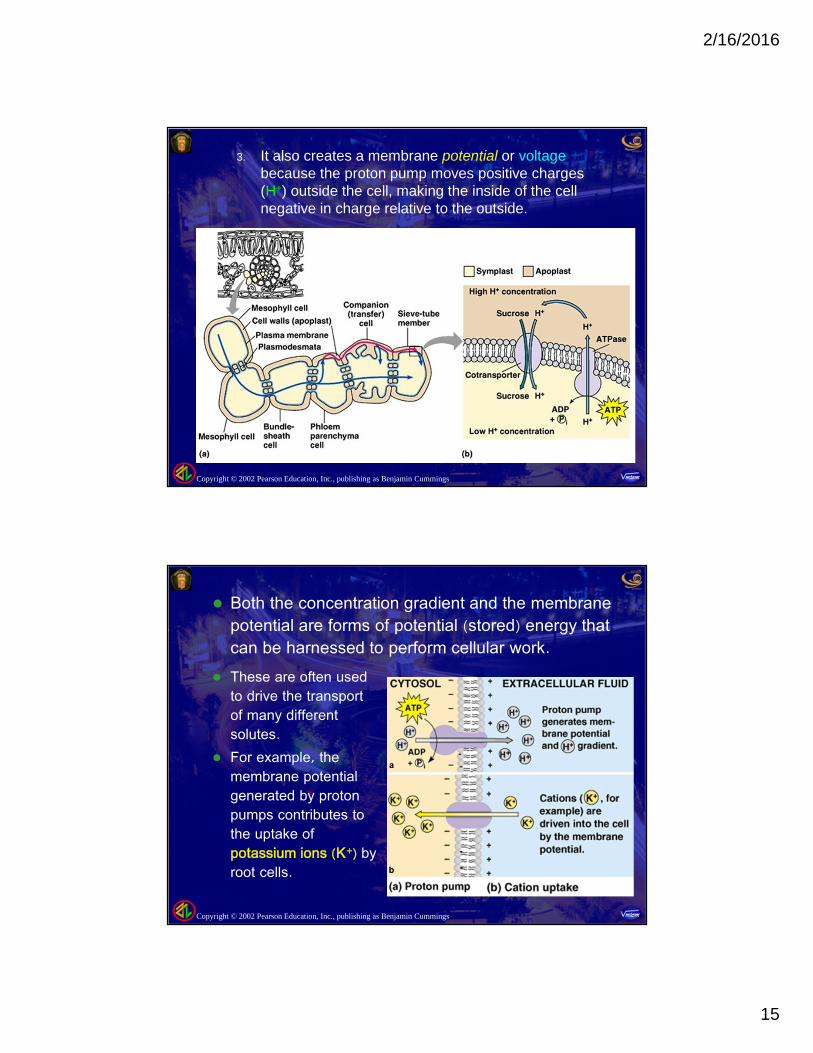
3. It also creates a membrane potential or voltagebecause the proton pump moves positive charges(H+) outside the cell, making the inside of the cellnegative in charge relative to the outside.
Copyright © 2002 Pearson Education, Inc., publishing as Benjamin Cummings
Both the concentration gradient and the membranepotential are forms of potential (stored) energy thatcan be harnessed to perform cellular work.
Copyright © 2002 Pearson Education, Inc., publishing as Benjamin Cummings
These are often usedto drive the transportof many differentsolutes.
For example, themembrane potentialgenerated by protonpumps contributes tothe uptake ofpotassium ions (K+) byroot cells.

2/16/2016
16
The proton gradientalso functions incotransport, in whichthe downhillpassage of onesolute (H+) iscoupled with theuphill passage ofanother, such asNO3
- or sucrose.
Copyright © 2002 Pearson Education, Inc., publishing as Benjamin Cummings
The role of protons pumps in transport is aspecific application of the general mechanismcalled chemiosmosis, a unifying principle incellular energetics. In chemiosmosis, a transmembrane proton gradient
links energy-releasing processes to energy-consuming processes. The ATP synthases that couple H+ diffusion to ATP
synthesis during cellular respiration and photosynthesisfunction somewhat like proton pumps.
However, proton pumps normally run in reverse, usingATP energy to pump H+ against its gradient.
Copyright © 2002 Pearson Education, Inc., publishing as Benjamin Cummings
2/16/2016
17
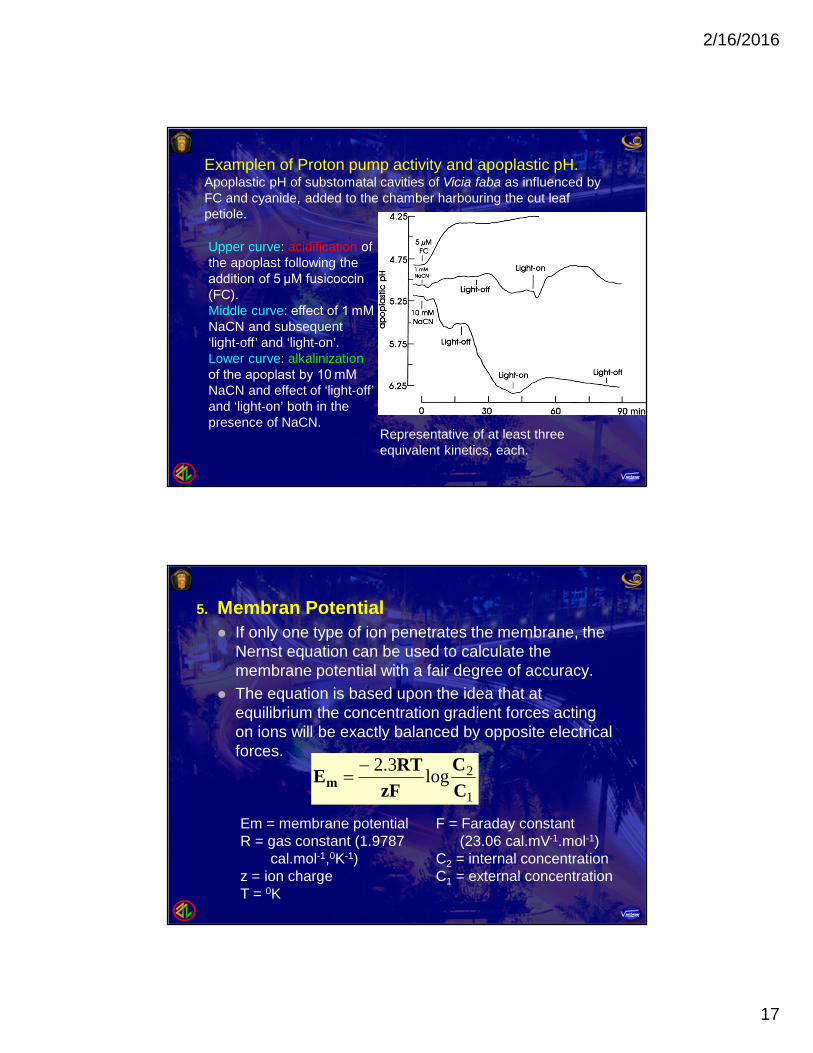
Examplen of Proton pump activity and apoplastic pH.Apoplastic pH of substomatal cavities of Vicia faba as influenced byFC and cyanide, added to the chamber harbouring the cut leafpetiole.
Upper curve: acidification ofthe apoplast following theaddition of 5 μM fusicoccin(FC).Middle curve: effect of 1 mMNaCN and subsequent‘light‐off’ and ‘light‐on’.Lower curve: alkalinizationof the apoplast by 10 mMNaCN and effect of ‘light‐off’and ‘light‐on’ both in thepresence of NaCN.
Representative of at least threeequivalent kinetics, each.
5. Membran Potential If only one type of ion penetrates the membrane, the
Nernst equation can be used to calculate themembrane potential with a fair degree of accuracy.
The equation is based upon the idea that atequilibrium the concentration gradient forces actingon ions will be exactly balanced by opposite electricalforces.
1
2log3.2
CC
zFRT
Em
Em = membrane potentialR = gas constant (1.9787
cal.mol-1,0K-1)z = ion chargeT = 0K
F = Faraday constant(23.06 cal.mV-1.mol-1)
C2 = internal concentrationC1 = external concentration

2/16/2016
18
A simple calculation of the Nernst potential E can be made as afunction of H+ concentration in two compartments as follows:
E = (RT/zF ) ln (C0/Ci)where R is the gas constant, T is the absolute temperature, z is thecharge of the ion, and F is the Faraday constant. C0 and Cirepresent the ion concentrations outside and inside the cellrespectively (considered here as two compartments).
Since H+ is the monovalent cation, the value of z would be 1 (z = 1).The numerical values of the constants R, F, and T at 30°C (303 K)can be substituted, and after converting from natural logarithm tolog10 (x2.303), we obtain:
E = 60 log (C0/Ci). Suppose the H+ concentration across the membrane is 10–5 M (pH
5.0) and 10–7 M (pH 7.0) as shown in Figure 2, then, E = 60 log(10–5
M/10–7 M) =log (100), and therefore, the membrane potential insidethe cell is –120 mV.
1
2
1
2 log06.59log06.23*1
25273987.1*3.2
CC
CC
Em
For z = 1 or 2 at 250C,
1
2
1
2 log53.29log06.23*2
25273987.1*3.2
CC
CC
Em

2/16/2016
19