leishmania accepted - ec.asm.org · 58 dimensional gel electrophoresis (2-de) increases from four...
TRANSCRIPT

1
Internal and surface-localized MSP of Leishmania and their differential release from 1
promastigotes 2
3
Chaoqun Yao 1,2*, John E. Donelson 3,4 and Mary E. Wilson 1,2,4,5,6 4
5
6
Departments of Internal Medicine1, Biochemistry3, Microbiology5, and Epidemiology6, Program 7
in Molecular Biology4, University of Iowa, Iowa City, IA 52242; VA Medical Center2, Iowa 8
City, IA 52246 9
10
11
*Corresponding author: Chaoqun Yao, 471 EMRB, Department of Internal Medicine, University 12
of Iowa, Iowa City, IA 52242. Tel: (319)335-6807; Fax: (319)353-4565; E-mail: chaoqun-13
15
16
Running Title: Major surface protease of Leishmania 17
ACCEPTED
Copyright © 2007, American Society for Microbiology and/or the Listed Authors/Institutions. All Rights Reserved.Eukaryotic Cell doi:10.1128/EC.00073-07 EC Accepts, published online ahead of print on 10 August 2007
on Decem
ber 2, 2018 by guesthttp://ec.asm
.org/D
ownloaded from

2
Abstract 18
MSP (major surface protease), also called GP63, is a virulence factor of Leishmania spp. 19
protozoa. There are three pools of MSP, located either internally within the parasite, anchored to 20
the surface membrane, or released into the extracellular environment. The regulation and 21
biological functions of these MSP pools are unknown. Herein we investigated the trafficking 22
and extrusion of surface versus internal MSPs. Virulent L. chagasi undergo a growth-associated 23
lengthening in the T½ of surface-localized MSP, but this did not occur in the attenuated L5 24
strain. The release of surface-localized MSP was enhanced in a dose-dependent manner by 25
MβCD, which chelates membrane cholesterol/ergosterol. Furthermore, incubation of 26
promastigotes at 37oC with Matrigel matrix, a soluble basement membrane extract of EHS 27
tumor cells, stimulated release of internal MSP but not surface-located MSP. Taken together, 28
these data indicate that MSP subpopulations in distinct cellular locations are released from the 29
parasite under different environmental conditions. We hypothesize that the internal MSP with its 30
lengthy T½ does not serve as a pool for promastigote surface MSP in the sand fly vector, but that 31
it instead functions as an MSP pool ready for a quick release upon inoculation of metacyclic 32
promastigotes into mammals. We present a model in which these different MSP pools are 33
released under distinct life cycle-specific conditions. 34
ACCEPTED
on Decem
ber 2, 2018 by guesthttp://ec.asm
.org/D
ownloaded from

3
The digenetic protozoa of Leishmania spp. shuttle between an extracellular promastigote 35
form in the sand fly vector and an intracellular amastigote form in mammalian hosts, including 36
humans. In the sand fly, the avirulent procyclic promastigotes develop to the virulent metacyclic 37
organisms, a process termed metacyclogenesis that can be mimicked by in vitro cultivation of 38
logarithmic to stationary phase promastigotes (36). Leishmania causes 1.5 to 2 million new 39
cases of human leishmaniasis annually, with manifestations ranging from self-healing cutaneous 40
sores to life-threatening visceral leishmaniasis (8, 9). Among a handful of well-characterized 41
Leishmania virulence factors is the major surface protease (MSP), also called GP63. MSP plays 42
several important roles during Leishmania spp. infection of mammals, including: (i) enhancing 43
promastigote phagocytosis by macrophages; (ii) facilitating promastigote evasion of 44
complement-mediated lysis; and (iii) promoting amastigote survival in the phagolysosomes of 45
macrophages [see (45) for a review]. There is also evidence suggesting that in the sand fly MSP 46
plays a role in the early-stage development of promastigotes (16), and contributes to 47
promastigote adhesion in the guts and salivary glands [see (37) for a review]. 48
MSP is encoded by a family of highly conserved genes organized in a tandem array. 49
MSP genes (MSPs) and homologues have been found in all Leishmania spp. studied to date, as 50
well as in other trypanosomatids, including the monoxenous insect parasite Crithidia and the 51
extracellular mammalian parasite Trypanosoma brucei (11, 13, 45). The number of MSPs in 52
individual trypanosomatids ranges from seven in L. major, to dozens in L. braziliensis, to 53
hundreds in T. cruzi (12, 28, 39, 41). At least 18 MSPs are present in L. chagasi, the causative 54
protozoan of visceral leishmaniasis in Latin America (33, 35). During in vitro promastigote 55
growth of virulent strain L. chagasi from logarithmic to stationary phase, MSP protein 56
abundance increases 14 fold. Concomitantly, the number of MSP isoforms observed on two-57
ACCEPTED
on Decem
ber 2, 2018 by guesthttp://ec.asm
.org/D
ownloaded from

4
dimensional gel electrophoresis (2-DE) increases from four to eleven (46-48). In this publication 58
we will use “MSP” when referring to properties of all MSP isoforms, and “MSPs” or “MSP 59
isoforms” when referring to the different MSP isoforms. 60
In addition to surface MSP, we and other groups have independently found that MSP is 61
released into the extracellular medium from Leishmania spp. and other trypanosomatids (7, 10, 62
18, 26, 46). In addition, a subpopulation of internal MSPs has been detected and appears to be 63
stable for several days (42, 47). Collectively, data generated from several laboratories, including 64
our own, demonstrate the existence of three subpopulations, i.e., surface-localized MSP, internal 65
MSP and released MSP. 66
We hypothesize that these three MSP subpopulations are separately trafficked through 67
the cell to interact with the environment, and that internal MSP serves as a pool ready for rapid 68
release after inoculation of metacyclic promastigote into mammalian skin. We previously 69
showed that the half life (T½) of surface-localized 63-kDa MSP in virulent L. chagasi increases 70
75% during promastigote growth from logarithmic to stationary phase (47). In the current report, 71
we demonstrate that this growth-associated regulation of surface-localized MSP T½ diminished 72
in the attenuated L5 L. chagasi strain. Furthermore, we report that the membrane lipid disruption 73
reagent methyl-β-cyclodextrin (MβCD) enhanced the release of surface-localized MSP into the 74
extracellular medium, whereas the internal MSP was released only after environmental exposure 75
to an in vitro extracellular matrix modeling basement membrane, but only at the elevated 76
temperature characteristic of a mammalian host. These data suggest that the different MSP pools 77
are regulated independently and play distinct functions during the life cycle of Leishmania spp. 78
A model illustrating the potential relevance of these findings during the parasite life cycle is 79
presented. 80
ACCEPTED
on Decem
ber 2, 2018 by guesthttp://ec.asm
.org/D
ownloaded from

5
Materials and Methods 81
Parasites. A Brazilian strain of L. chagasi (MHOM/BR/00/1669) was continuously passaged, by 82
intracardiac injection of amastigotes, in golden hamsters to maintain its virulence. Amastigotes 83
were isolated from the spleens of infected hamsters and transformed to promastigotes at 26oC, in 84
hemoflagellate-modified minimal essential medium with 10% heat-inactivated fetal calf serum 85
(HOMEM; reagents from GIBCO, Rockville, MO). Virulent promastigotes were passaged 86
weekly in HOMEM, and used within five passages. The attenuated L5 strain of L. chagasi has 87
been continuously cultured in vitro in HOMEM for over nine years (43). Strain L5 differs from 88
the virulent strain in several respects including (i) less abundant MSP and the expression of only 89
MSPL genes (6, 43, 48), (ii) a shorter and simpler lipophosphoglycan (27), and (iii) reduced 90
virulence for rodent models (43). In some experiments, virulent promastigotes were spread on 91
semi-solid M199-agar plates to obtain clonal cells (19). A total of 124 clones were established in 92
two independent experiments. Promastigote cultures were started at a cell density of 1 × 106 93
cell/ml at day 0 of cultivation. Logarithmic and stationary phase promastigotes were collected 94
between days 2-4 and 6-9, respectively, with phases defined according to cell density and 95
morphology as previously described (49). 96
Chemicals and antibodies. Sulfo-NHS-Biotin, streptavidin agarose beads, and growth factor 97
reduced Matrigel™ matrix were purchased from Pierce (Rockford, IL), Sigma (St. Louise, MO), 98
and BD Biosciences (Bedford, MA), respectively. MβCD, protein G-agarose beads and Promix 99
containing [35S]-methionine and -cysteine were bought from Sigma, CalBioChem (San Diego, 100
CA) and Amersham Pharmacia Biotech (Piscataway, NJ), respectively. Polyclonal rabbit and 101
sheep antisera to MSP were raised against purified L. chagasi MSP as previously described (43). 102
Monoclonal antibody to α-tubulin (AB-1) was purchased from Oncogene (San Diego, CA). 103
ACCEPTED
on Decem
ber 2, 2018 by guesthttp://ec.asm
.org/D
ownloaded from

6
Peroxidase conjugated anti-rabbit, anti-sheep, and anti-mouse antisera were purchased from 104
CalBioChem, Kirkegaard & Perry Laboratories (Gaitherburg, MA) and Bio-Rad Laboratories 105
(Richmond, CA), respectively. 106
Metabolic labeling and surface biotinylation. These procedures were conducted using 107
previously published protocols (47). Briefly, the promastigotes were pulsed in Hank’s balanced 108
salt solution (HBSS, GIBCO) with Promix for 0.5 h, followed by surface biotinylation for an 109
additional 0.5 h in Sulfo-NHS-Biotin. Samples were taken between 0 and 72 h of “chase” in 110
serum-free, bovine serum albumin-free medium. Both cells and cell-free spent medium were 111
collected. Newly synthesized MSP, localized either on the cell surface or intracellularly, was 112
isolated by streptavidin-affinity purification and immunoprecipitated from the streptavidin-113
cleared fraction, respectively, and detected by autoradiography as previously described (47, 48). 114
The efficiency of pull-down via biotin/streptavidin was routinely monitored by peroxidase 115
conjugated ExtrAvidin (Sigma) and ECL™ western blotting detection reagents (Amersham). In 116
contrast to the streptavidin pull-down fractions, the streptavidin-cleared fractions exhibited no 117
detectable signals. 118
MβCD treatment of promastigotes. Promastigotes were washed twice by centrifugation in 119
HBSS, and incubated for 48 h at 2 × 107 cells/ml in freshly prepared MβCD in RPMI 1640 120
(GIBCO) ranging in concentration from 0 to 15 mM. All conditions were done in triplicate. To 121
monitor cell viability, MβCD-treated or control (0 mM MβCD) promastigotes were 122
metabolically labeled in HBSS with Promix for 0.5 h, and triplicate samples were assayed by a 123
liquid scintillation analyzer for incorporation of radioisotope after total proteins were 124
precipitated with trichloroacetic acid as described (3). The relative [35S]-amino acid 125
incorporation in the presence of MβCD was compared to that in control (untreated) 126
ACCEPTED
on Decem
ber 2, 2018 by guesthttp://ec.asm
.org/D
ownloaded from

7
promastigotes. A ratio of 1.0 indicated that MβCD had no effect on promastigote viability. To 127
investigate whether membrane lipid chelation reagent MβCD affects the release of surface-128
localized MSP, stationary phase promastigotes were incubated in either 0 or 15 mM MβCD for 3 129
h after surface biotinylation. Spent medium was collected and concentrated as previously 130
described (48). Both biotinylated proteins and the internal MSP were analyzed. 131
MSP release into Matrigel™ matrix. Stationary phase promastigotes in the first passage after 132
being converted from amastigotes isolated from hamsters were surface biotinylated. Triplicate 133
samples of cells were suspended to a density of 2 × 108 cells/ml, in ice-cold HBSS (100 µl) plus 134
Matrigel™ matrix (200 µl). Cultures were then incubated at either room temperature (RT) or 135
37oC for 1 to 3 h. Matrigel™ matrix solidified under these conditions. The mixtures were 136
transferred to 4oC overnight to liquefy the matrix, and promastigote cells were separated from 137
the liquefied Matrigel™ matrix by centrifugation. Biotinylated proteins and the non-biotinylated 138
MSP were collected from both the whole cellular lysate and the liquefied Matrigel™ matrix. 139
Electrophoresis and protein detection. SDS-polyacrylamide gel electrophoresis and western 140
blotting were conducted as described (46). Autoradiogram was achieved by exposing X-ray MR 141
films (Kodak, Rochester, NY). Samples analyzed by 2-DE were separated in the first dimension 142
by isoelectric focusing (IEF) in Immobiline™ Drystrips pH 4-7 (Amersham), and in the second 143
dimension according to size, in 7.5% SDS-polyacrylamide gels (48). In the case of cellular 144
lysates of individual clones, the samples were separated in Immobiline™ Dryplates pH 4-7 145
(Amersham), after which proteins were transferred to a nitrocellulose membrane and MSP was 146
detected by western blotting. 147
ACCEPTED
on Decem
ber 2, 2018 by guesthttp://ec.asm
.org/D
ownloaded from

8
Results 148
MSP expression in clonal L. chagasi lines. Virulent stationary phase L. chagasi promastigotes 149
contain at least eleven isoforms of MSP according to 2-DE immunoblots. However, there have 150
been no documented differences in the function or localization of these MSP isoforms. At least 151
some of the different MSP isoforms are derived from different MSPs (47, 48). We approached 152
the question of differential function by carefully characterizing MSP isoforms. Our prior work 153
documents MSP isoforms in uncloned L. chagasi populations (47, 48); Fig. 1 shows 2-DE MSP 154
profiles in clonal L. chagasi cell lines. Clonal lines were derived from a stationary phase 155
promastigote population on semi-solid M199-agar plates. Proteins were separated by IEF and 156
transferred to nitrocellulose membranes, and MSPs were detected by immunoblotting with 157
polyvalent antiserum against MSP. An MSP profile similar to that of the entire population was 158
observed for all 124 clones examined; representative examples shown in Fig. 1. Eight bands 159
between isoelectric points (pI) 5.8 and 6.7 were detected, along with an additional three bands 160
between pI 4.8 and 5.2. Each of these bands formed a spot in the second dimension, as shown in 161
immunoblots of 24 representative clones (Fig. 1). As we anticipated but thought it important to 162
investigate, we did not detect clonal variation in MSP expression during growth in vitro. 163
Surface-localized MSP isoforms are differently regulated in attenuated and virulent 164
strains. The MSP proteins of L. chagasi promastigotes are found in three cellular locations, i.e. 165
internal, surface, and released into the environment (46, 47). The cellular distribution of MSP 166
proteins changes during in vitro growth. We previously showed that an increase in total 167
promastigote MSP content during “in vitro metacyclogenesis” is associated with a decrease in 168
the rate of MSP release into the environment. To study the population of MSP released we 169
contrasted MSP synthesis in, and loss from, virulent promastigotes compared to a non-virulent 170
ACCEPTED
on Decem
ber 2, 2018 by guesthttp://ec.asm
.org/D
ownloaded from

9
attenuated line of parasites (L5) that does not undergo changes in total MSP content during 171
metacyclogenesis (Fig. 2). For the purposes of comparison we contrasted the 63-kDa MSP, an 172
isoform that is expressed in both promastigote lines. 173
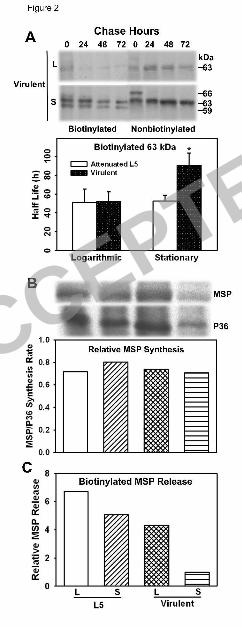
First, the rate of MSP synthesis in both virulent and attenuated lines, in both logarithmic 174
and stationary growth phases, was almost identical (Fig. 2B). L5 or virulent L. chagasi 175
promastigotes were metabolically labeled with [35S]-methionine, MSPs were 176
immunoprecipitated, and newly synthesized MSPs were detected by autoradiogram. Cytosolic 177
P36, which is constitutively expressed (22, 47), was used as a control. The ratio of MSP to P36 178
remained constant in logarithmic and stationary phase promastigotes of both the L5 and virulent 179
strains. 180
Secondly, the T½ of cellular surface MSP was longer in stationary virulent promastigotes 181
than in logarithmic virulent promastigotes, coinciding with its increased abundance in stationary 182
virulent promastigotes. In contrast, surface MSP was lost at a uniform rate in the growth phases 183
of attenuated L5 parasites (Fig. 2A). Both virulent and attenuated promastigotes were surface 184
labeled by biotinylation during logarithmic or stationary phase of growth, and chased over the 185
next 72 h. Immunoblotting was used to confirm that the indicated bands were indeed MSPs (not 186
shown). The surface MSP of virulent L. chagasi promastigotes had a shorter T1/2 (52 h) when 187
labeled in logarithmic growth phase than MSP proteins labeled in stationary phase (90 h; Fig. 188
2A). Whether this is due to the predominant MSP isoforms synthesized in the different growth 189
stages, or to other factors inherent in the growth phase of parasites cannot be determined (46, 190
47). In contrast, the T½ of surface-localized 63-kDa MSP in the attenuated L5 strain of L. 191
chagasi promastigotes remained unchanged during growth from logarithmic (51 h) to stationary 192
phase (52 h). Indeed, the MSP T½ was almost identical to that of logarithmic growth-phase 193
ACCEPTED
on Decem
ber 2, 2018 by guesthttp://ec.asm
.org/D
ownloaded from

10
virulent strain promastigotes (52 h) (Fig. 2A). In contrast to surface MSP, the internal MSP of 194
both virulent and attenuated L5 strains remained stable throughout promastigote growth [Fig. 2A 195
and (47)]. 196
Thirdly, the mechanism differentiating the T½ of surface MSP in virulent as opposed to 197
attenuated L5 promastigotes was a difference in the rate of MSP shedding into the medium. 198
Surface MSP was labeled by biotinylation in both L5 and virulent strain parasites. Parasites 199
were then incubated in fresh medium, and surface biotinylated MSPs were detected by western 200
blotting of the streptavidin bead-purified fraction of the spent media after 48 h incubation. A 201
minimum of four fold more MSPs was found in the spent media of both logarithmic and 202
stationary phase of L5 strain and the logarithmic phase of the virulent strain than the stationary 203
phase of the virulent strain (Fig. 2C). Collectively, these data indicate the increase in surface-204
localized MSP in the stationary phase virulent promastigotes is associated with a decrease in the 205
rate of shedding into the environment, compared to logarithmic phase virulent promastigotes. 206
There is not similar growth phase-dependent regulation of MSP in the L5 attenuated strain of L. 207
chagasi. 208
MβCD enhances release of the surface-localized MSP isoforms. The unique retention of 209
surface MSP by virulent stationary phase promastigotes could be due to its association with 210
surface lipid-containing membrane domains. MβCD depletes lipid rafts from the plasma 211
membranes of a variety mammalian cells by chelating and transiently removing membrane 212
cholesterol (14, 17, 21, 24, 31, 40). Based on the hypothesis that differential association of MSP 213
with membrane lipids could account for its release by logarithmic promastigotes and retention by 214
stationary promastigotes, we reasoned that membrane lipid disruption with MβCD could enhance 215
MSP release from the Leishmania membrane. In replicate experiments, virulent L. chagasi 216
ACCEPTED
on Decem
ber 2, 2018 by guesthttp://ec.asm
.org/D
ownloaded from

11
promastigotes were treated with 0, 5, 10 or 15 mM MβCD for 48 h. A dose-dependent 217
augmented release of MSP into the extracellular medium was observed. Specifically, control 218
cells (0 mM MβCD) released about 35% of MSP, whereas the cells in 15 mM MβCD released 219
~80% of MSP into the extracellular medium (Fig. 3A and B). 220
To eliminate the possibility that the enhanced MSP release was due to a toxic effect of 221
MβCD on promastigotes, the rate of promastigote protein synthesis was measured in the absence 222
or presence of MβCD under the experimental conditions. Comparable levels of [35S]-223
radioisotopes were incorporated into newly synthesized proteins of untreated or MβCD-treated 224
cells (Fig. 3C), indicating that MβCD treatment of promastigotes under these conditions is not 225
detrimental to the cells. Furthermore, the growth curves of untreated or MβCD-treated cells in 226
HOMEM were similar (data not shown). Thus, perturbing the plasma membrane lipid-227
containing domains of stationary phase promastigotes accelerates MSP release into the 228
extracellular medium, although it does not appear to harm the promastigotes in culture. 229
To test the hypothesis that disruption of membrane lipid-containing domains with MβCD 230
only promotes release of surface-localized MSP, stationary phase promastigotes were treated 231
with 15 mM MβCD for 3 h after surface biotinylation. Control cells were treated identically but 232
received no MβCD. Spent medium was collected, from which surface-biotinylated proteins were 233
isolated by streptavidin affinity purification. Internal MSP was purified by immunoprecipitation 234
from the streptavidin-cleared fraction of the spent medium. Immunoblots were used to assay for 235
the presence of MSP. As showed in Fig. 3D, non-biotinylated, internal MSP was undetectable in 236
extracellular medium. In contrast, biotinylated, surface MSP was 34.6±12.8% (n=3) more 237
abundant in the spent media of the MβCD-treated cells than controls. Furthermore, no 238
cytoskeletal β-tubulin and cytosolic P36 markers were detected by immunoblotting in the clear 239
ACCEPTED
on Decem
ber 2, 2018 by guesthttp://ec.asm
.org/D
ownloaded from

12
fraction of the same spent media post biotin-streptavidin affinity purification and MSP 240
immunoprecipitation (Fig. 3D, and data not shown), which eliminates the possibility that MSP 241
release is due to cell lysis. These data indicate that MβCD enhances release of only surface-242
localized MSP, a result consistent with the possibility that MSP stabilization in the surface 243
membrane requires an association with cholesterol/ergosterol-containing lipid domains. 244
Release of internal MSP isoforms is stimulated by the Matrigel™ matrix, specifically at 245
37oC. Although surface MSP can be artificially released by disrupting membrane lipid domains, 246
the natural evolution of stationary promastigotes in the sand fly is to a cellular state that retains 247
surface MSP. Metacyclic promastigotes are inoculated by sand flies into mammalian tissues, 248
whereupon they initially encounter an elevated temperature and components of extracellular 249
mammalian environment. We investigated whether MSP would be released under conditions 250
that mimic the in vivo setting. First we tested whether the highest mammalian body temperature 251
encountered by the parasite, i.e., 37oC, would stimulate internal MSP release. Stationary phase 252
promastigotes were metabolically labeled, surface biotinylated, and subsequently incubated at 253
37oC for 24 h to test for release of surface versus internal MSP. Similarly treated control 254
promastigotes were incubated at RT. Surface and internal MSP were immunoprecipitated from 255
the streptavidin bead-enriched or -cleared cellular lysates and detected by autoradiography. 256
Under these conditions, there was no detectable change in internal versus surface MSP in the 257
promastigotes after 24 h at higher temperature (data not shown). These data suggest that 258
temperature increase to 37oC is by itself insufficient to stimulate internal MSP release. 259
We then incubated stationary phase promastigotes in the Matrigel™ matrix at 37oC to test 260
whether this combination would stimulate the release of internal MSP. Matrigel matrix is a 261
soluble basement membrane extract of EHS tumor cells, which has been used to study the 262
ACCEPTED
on Decem
ber 2, 2018 by guesthttp://ec.asm
.org/D
ownloaded from

13
metastasis of cancer cells (29, 34). One prominent feature of this matrix is that it is a liquid at 263
4oC but it gels at RT and above, forming a reconstituted basement membrane. Consequently, 264
when promastigotes are incubated in the matrix at 37oC, this setting experimentally mimics the 265
site of sand fly inoculation into a mammalian host. 266
Stationary phase promastigotes were surface biotinylated prior to incubation in either 267
Matrigel matrix or HBSS. Promastigotes incubated in HBSS released surface MSP but little or 268
no internal MSP into the extracellular medium at RT. Neither MSP form, either surface or 269
internal, was substantially released at 37oC (Fig. 4A and B). In contrast, incubation of 270
promastigotes in the Matrigel matrix for 3 h at 37oC stimulated release of mostly internal MSP 271
(Fig. 4A and B). This effect was enhanced by a longer (3 vs. 1 h) incubation time. Strikingly, 272
the effect of Matrigel on release of internal MSP was significantly lower at RT, whereas more 273
surface MSP was released under these conditions (Fig. 4A and B). Furthermore, the level of 274
total internal MSP was significantly higher in parasites incubated in Matrigel™ compared to 275
HBSS, although there was no change in internal MSP when parasites were incubated at RT 276
versus 37oC (Fig. 4C). Hence, it is very unlikely that the specific release of internal MSP 277
stimulated by a combination of Matrigel™ matrix and 37oC was due to leakiness of intracellular 278
content from damaged promastigotes, even though we can not formally eliminate this possibility 279
at this time. Overall, these results lead us to conclude that surface MSP is released at RT and 280
that this release is inhibited at 37oC, whereas internal MSP is released in response to the presence 281
of Matrigel matrix, specifically at 37oC (Fig. 4). 282
ACCEPTED
on Decem
ber 2, 2018 by guesthttp://ec.asm
.org/D
ownloaded from

14
Discussion 283
MSPs are among the most abundant cellular proteins in promastigotes of all Leishmania 284
spp. studied to date. Indeed, in L. mexicana, MSPs account for 1% and 0.1% of total proteins in 285
promastigotes and amastigotes, respectively (1). Promastigote cell-associated MSP is 286
predominantly attached to the cell surface by glycosylphosphatidylinositol anchors (4, 5). 287
However, our laboratory and others have observed that as much as one-third of the cell-288
associated MSP is located intracellularly, as determined by a combination of surface 289
biotinylation, immunoelectron microscopy and cytofluorometry (42, 47). Furthermore, the 290
internal MSP in L. chagasi is so stable that no reduction in abundance is detected for up to six 291
days using pulse-chase analysis (47). We hypothesized that the surface-localized and internal 292
MSPs are regulated separately via different mechanisms. Furthermore, the role and origin of the 293
MSP released by promastigotes into extracellular medium has as yet been uncharacterized. In 294
the current study we showed using MβCD that the decreased release of surface MSP by the 295
virulent stationary promastigotes is associated with the content of membrane lipids, since 296
MβCD-mediated removal of cholesterol/ergosterol specifically enhanced the release of surface-297
localized MSP into extracellular medium. This likely reflects changes in the promastigote 298
membrane during metacyclogenesis, in that a lipid-rich membrane retaining MSP in metacyclic 299
parasites could promote retention of high surface levels of this virulence factor. In contrast, 300
exposure to conditions mimicking mammalian tissue with Matrigel at 37oC stimulated the 301
release of internal, but not surface, MSP. These data demonstrate for the first time that the 302
surface-localized and internal MSPs are trafficked out of the promastigote cell in response to 303
different external stimuli. 304
ACCEPTED
on Decem
ber 2, 2018 by guesthttp://ec.asm
.org/D
ownloaded from

15
Phenotypic variation has been found in isoforms of a 235-kDa rhpoptry protein between 305
clones of Plasmodium yoelli parasites. This protein is encoded by a multigene family of ~50 306
genes and may be involved in the selection of red blood cells for invasion by merozoites (2, 15, 307
20, 30, 38). Because there are at least 11 MSP isoforms in stationary phase virulent L. chagasi 308
promastigotes (47, 48), we hypothesized that similar variation between L. chagasi parasites 309
could yield clonal isolates that express one or a few MSPs. However, we were not able to 310
document clonal variation in MSP expression by cells expanded from individual clones, using 2-311
DE immunoblotting. This does not prove that all parasite clones express all MSP isoforms, or 312
that individual parasite clones cannot express only one or a few MSP isoforms in vivo. 313
Nonetheless, according to our ability to detect we tentatively conclude that at least some L. 314
chagasi parasites are able to express the majority of MSP isoforms when derived from a single 315
cloned cell. 316
The three MSP classes of mRNAs (MSPL, MSPS and MSPC) in L. chagasi are post-317
transcriptionally regulated. In the case of MSPL mRNA this regulation is known to occur 318
specifically at the level of mRNA stability (6, 32, 44). Regarding MSP regulation at the protein 319
level, we showed herein that the measurable rate of MSP synthesis was very similar throughout 320
promastigote growth in vitro from logarithmic to stationary phase, consistent with our earlier 321
report (47). Therefore, the growth-associated 14-fold increase in the abundance of cell-322
associated MSP must be post-translationally regulated. An increase in protein stability, 323
associated with decreased MSP shedding, accounts for a 5-fold increase (47). We show in the 324
current study that internal pool of MSP is extremely stable throughout growth of both attenuated 325
L5 and virulent parasite strains. Consequently, internal MSP appears not to be affected by the 326
growth-associated regulation of MSP stability. We also demonstrate that the T½ of surface-327
ACCEPTED
on Decem
ber 2, 2018 by guesthttp://ec.asm
.org/D
ownloaded from

16
localized 63-kDa MSP in the attenuated strain is similar to that of the lower MSP-expressing, 328
logarithmic phase promastigotes of the virulent strain, regardless of the growth phase (Fig. 2). 329
One plausible explanation for this difference between attenuated and virulent strains during 330
growth is the different rates of MSP shedding. We documented that the rate of MSP shedding by 331
stationary phase virulent promastigotes is slower than that of logarithmic phase virulent 332
promastigotes, and that MSP shedding by L5 attenuated promastigotes is more rapid than 333
virulent L. chagasi in all growth phases (see Fig. 2C). 334
The biochemical mechanisms by which Leishmania spp. promastigotes regulate MSP 335
release are not well understood. Released MSPs have similar electrophoretic mobility to their 336
cell-associated counterparts (46). At least some surface-localized L. amazonensis MSP is 337
released through autoproteolytic activity as showed by site-specific mutation and inhibition by a 338
zinc chelator (26). We previously determined that released MSP does not bind to a antibody 339
against the cross-reactive determinant, suggesting it is not released by a PI-PLC (46) similar to 340
the released MSP of L. amazonensis. Released L. amazonensis MSP does not contain 341
ethanolamine suggesting it lacks a GPI membrane anchor (26). Data generated in this report 342
using lipid chelation suggests that the decreased release of MSP from stationary virulent 343
promastigotes is due to remodeling of the surface membrane such that MSP is retained in 344
association with lipids. We cannot rule out the additional possibility that there may also be 345
recycling and degradation of MSP as a means of decreasing cellular levels of MSP, but this has 346
yet to be tested. 347
In addition to the above evidence that MSP release by virulent promastigotes requires a 348
specific membrane lipid composition, we approached the mechanisms by which Leishmania spp. 349
promastigotes release MSP using a model of in vivo conditions. The Matrigel™ matrix contains 350
ACCEPTED
on Decem
ber 2, 2018 by guesthttp://ec.asm
.org/D
ownloaded from

17
laminin, collagen IV, entacin, heparin sulfate proteoglycan, growth factors, collagenases, and 351
other undefined components. We demonstrated here that a combination of this matrix and 352
mammalian body temperature is sufficient to stimulate internal MSP release. We suggest that 353
the mechanism of MSP release in mammalian tissue differs from release in promastigote axenic 354
culture. Whether this reflects differences between MSP trafficking in the sand fly versus the 355
mammalian hosts is not clear. 356
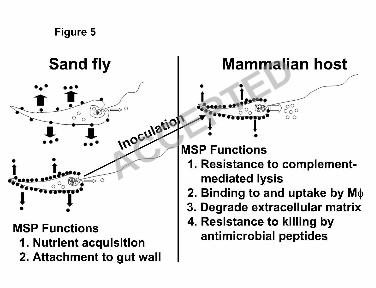
The goal of the current study was to address how the three MSP subpopulations (surface, 357
internal and released) are regulated during metacyclogenesis, and in response to the mammalian 358
host environment. A model for MSP regulation in the different promastigote environments is 359
illustrated in Fig. 5. In this model MSP is abundantly released by the dividing, procyclic 360
promastigotes in the sand fly gut, as simulated by logarithmic growth of L. chagasi in culture. 361
This released MSP might be related to the nutrient requirements of Leishmania in the insect gut 362
environment, where residual mammalian blood from the sand fly meal is a main source of 363
nutrients. Indeed, it has been shown that down-regulation of MSP in L. amazonensis reduces the 364
parasites’ early development in sand flies (16). As procyclic promastigotes develop to 365
metacyclic promastigotes, the rate of released surface-localized MSP decreases and the 366
abundance of surface-localized MSP increases (47). Our data suggest this increase is due to 367
association of metacyclic MSP with lipid-containing membrane domains. Internal MSP is not 368
released during metacyclogenesis. However, after inoculation into mammalian subcutaneous 369
tissue by a sand fly vector, metacyclic promastigotes encounter a temperature increase, host 370
extracellular matrix, and innate immune mechanisms such as complement, antimicrobial peptides 371
and phagocytotic cells. In response to these stimuli, promastigotes could release internal MSP 372
into mammalian tissue. It is thus logical to consider the possibility that the surface-localized 373
ACCEPTED
on Decem
ber 2, 2018 by guesthttp://ec.asm
.org/D
ownloaded from

18
MSP plays a role in the promastigotes’ evasion of complement-mediated killing and their 374
phagocytosis/internalization by macrophages and other cells. Internal MSP, on the other hand, 375
may play a role in the degradation of extracellular matrix components such as collagen IV and 376
fibronectin, as suggested in a prior report on an L. amazonensis (25). As such, it could facilitate 377
promastigote migration toward cells such as macrophages, dendritic cells and fibroblasts that are 378
favorable for parasite entry and long-term survival. Thus, it is likely that the many isoforms of 379
MSP protease facilitate parasite survival through different mechanisms in the diverse host and 380
vector environments encountered by the parasite. 381
ACCEPTED
on Decem
ber 2, 2018 by guesthttp://ec.asm
.org/D
ownloaded from

19
Acknowledgments 382
CY was supported by a Veterans’ Affairs Merit Review Entry grant and a Veterans’ 383
Affairs Merit Review grant. Other support for this work included NIH R01 grants AI32135 and 384
AI059451 (MEW and JED), NIH R01 grants AI045540, AI048822 and AI067874 (MEW), and 385
Merit Review and Gulf War RFA grants from the Department of Veterans’ Affairs (MEW). 386
References 387
1. Bahr, V., Y. D. Stierhof, T. Ilg, M. Demar, M. Quinten, and P. Overath. 1993. 388
Expression of lipophosphoglycan, high-molecular weight phosphoglycan and glycoprotein 389
63 in promastigotes and amastigotes of Leishmania mexicana. Mol. Biochem. Parasitol. 390
58:107-121. 391
2. Bayele, H. K., and K. N. Brown. 2007. Plasmodium yoelii: combinatorial expression of 392
variants of the 235 kDa rhoptry antigen during infection. Exp. Parasitol. 116:354-360. 393
3. Bonifacino, J. S. 1998. Metabolic laleling with amino acids, p. 10.18.11-10.18.10. In F. M. 394
Ausubel, R. Brent, R. E. Knigston, D. D. Moore, J. G. Seidman, J. A. Smith, and K. Struhl 395
(ed.), Current Protocols in Molecular Biology, vol. 2. John Wiley & Sons, Inc. 396
4. Bordier, C. 1981. Phase separation of integral membrane proteins in Triton X-114 solution. 397
J. Biol. Chem. 256:1604-1607. 398
5. Bordier, C., R. J. Etges, J. Ward, M. J. Turner, and M. L. Cardoso de Almeida. 1986. 399
Leishmania and Trypanosoma surface glycoproteins have a common glycophospholipid 400
membrane anchor. Proc. Natl. Acad. Sci. USA 83:5988-5991. 401
6. Brittingham, A., M. A. Miller, J. E. Donelson, and M. E. Wilson. 2001. Regulation of 402
GP63 mRNA stability in promastigotes of virulent and attenuated Leishmania chagasi. Mol. 403
Biochem. Parasitol. 112:51-59. 404
ACCEPTED
on Decem
ber 2, 2018 by guesthttp://ec.asm
.org/D
ownloaded from

20
7. Cuervo, P., L. Saboia-Vahia, F. Costa Silva-Filho, O. Fernandes, E. Cupolillo, and D. 405
E. J. JB. 2006. A zymographic study of metalloprotease activities in extracts and 406
extracellular secretions of Leishmania (Viannia) braziliensis strains. Parasitology 132:177-407
185. 408
8. Desjeux, P. 1992. Human leishmaniases: epidemiology and public health aspects. Wld. 409
Hlth. Statist. Quart. 45:267-275. 410
9. Desjeux, P. 2004. Leishmaniasis: current situation and new perspectives. Comp. Immunol. 411
Microbiol. Infect. Dis. 27:305-318. 412
10. Elias, C. G., F. M. Pereira, B. A. Silva, C. S. Alviano, R. M. Soares, and A. L. Santos. 413
2006. Leishmanolysin (gp63 metallopeptidase)-like activity extracellularly released by 414
Herpetomonas samuelpessoai. Parasitology 132:37-47. 415
11. El-Sayed, N. M., and J. E. Donelson. 1997. African trypanosomes have differentially 416
expressed genes encoding homologues of the Leishmania GP63 surface protease. J. Biol. 417
Chem. 272:26742-26748. 418
12. El-Sayed, N. M., P. J. Myler, D. C. Bartholomeu, D. Nilsson, G. Aggarwal, A. N. Tran, 419
E. Ghedin, E. A. Worthey, A. L. Delcher, G. Blandin, S. J. Westenberger, E. Caler, G. 420
C. Cerqueira, C. Branche, B. Haas, A. Anupama, E. Arner, L. Aslund, P. Attipoe, E. 421
Bontempi, F. Bringaud, P. Burton, E. Cadag, D. A. Campbell, M. Carrington, J. 422
Crabtree, H. Darban, J. F. da Silveira, P. de Jong, K. Edwards, P. T. Englund, G. 423
Fazelina, T. Feldblyum, M. Ferella, A. C. Frasch, K. Gull, D. Horn, L. Hou, Y. Huang, 424
E. Kindlund, M. Klingbeil, S. Kluge, H. Koo, D. Lacerda, M. J. Levin, H. Lorenzi, T. 425
Louie, C. R. Machado, R. McCulloch, A. McKenna, Y. Mizuno, J. C. Mottram, S. 426
Nelson, S. Ochaya, K. Osoegawa, G. Pai, M. Parsons, M. Pentony, U. Pettersson, M. 427
ACCEPTED
on Decem
ber 2, 2018 by guesthttp://ec.asm
.org/D
ownloaded from

21
Pop, J. L. Ramirez, J. Rinta, L. Robertson, S. L. Salzberg, D. O. Sanchez, A. Seyler, R. 428
Sharma, J. Shetty, A. J. Simpson, E. Sisk, M. T. Tammi, R. Tarleton, S. Teixeira, S. 429
Van Aken, C. Vogt, P. N. Ward, B. Wickstead, J. Wortman, O. White, C. M. Fraser, K. 430
D. Stuart, and B. Andersson. 2005. The genome sequence of Trypanosoma cruzi, etiologic 431
agent of Chagas disease. Science 309:409-415. 432
13. Etges, R. 1992. Identification of a surface metalloproteinase on 13 species of Leishmania 433
isolated from humans, Crithidia fasciculata, and Herpetomonas samuelpessoai. Acta Trop. 434
50:205-217. 435
14. Foster, L. J., C. L. De Hoog, and M. Mann. 2003. Unbiased quantitative proteomics of 436
lipid rafts reveals high specificity for signaling factors. Proc. Natl. Acad. Sci. USA 437
100:5813-5818. 438
15. Gruner, A. C., G. Snounou, K. Fuller, W. Jarra, L. Renia, and P. R. Preiser. 2004. The 439
Py235 proteins: glimpses into the versatility of a malaria multigene family. Microbes Infect. 440
6:864-873. 441
16. Hajmova, M., K. P. Chang, B. Kolli, and P. Volf. 2004. Down-regulation of gp63 in 442
Leishmania amazonensis reduces its early development in Lutzomyia longipalpis. Microbes 443
Infect. 6:646-649. 444
17. Hao, M., S. Mukherjee, and F. R. Maxfield. 2001. Cholesterol depletion induces large 445
scale domain segregation in living cell membranes. Proc. Natl. Acad. Sci. USA 98:13072-446
13077. 447
18. Jaffe, C. L., and D. M. Dwyer. 2003. Extracellular release of the surface metalloprotease, 448
gp63, from Leishmania and insect trypanosomatids. Parasitol. Res. 91:229-237. 449
ACCEPTED
on Decem
ber 2, 2018 by guesthttp://ec.asm
.org/D
ownloaded from

22
19. Kapler, G. M., C. M. Coburn, and S. M. Beverley. 1990. Stable transfection of the human 450
parasite Leishmania major delineates a 30-kilobase region sufficient for extrachromosomal 451
replication and expression. Mol. Cell. Biol. 10:1084-1094. 452
20. Khan, S. M., W. Jarra, and P. R. Preiser. 2001. The 235 kDa rhoptry protein of 453
Plasmodium (yoelii) yoelii: function at the junction. Mol. Biochem. Parasitol. 117:1-10. 454
21. Kojro, E., G. Gimpl, S. Lammich, W. Marz, and F. Fahrenholz. 2001. Low cholesterol 455
stimulates the nonamyloidogenic pathway by its effect on the alpha -secretase ADAM 10. 456
Proc. Natl. Acad. Sci. USA 98:5815-5820. 457
22. Liu, X., and K. P. Chang. 1994. Identification by extrachromosomal amplification and 458
overexpression of a zeta-crystallin/NADPH-oxidoreductase homologue constitutively 459
expressed in Leishmania spp. Mol. Biochem. Parasitol. 66:201-210. 460
23. Macdonald, M. H., C. J. Morrison, and W. R. McMaster. 1995. Analysis of the active 461
site and activation mechanism of the Leishmania surface metalloproteinase GP63. Biochim. 462
Biophys. Acta 1253:199-207. 463
24. Marwali, M. R., J. Rey-Ladino, L. Dreolini, D. Shaw, and F. Takei. 2003. Membrane 464
cholesterol regulates LFA-1 function and lipid raft heterogeneity. Blood 102:215-222. 465
25. McGwire, B. S., K. P. Chang, and D. M. Engman. 2003. Migration through the 466
extracellular matrix by the parasitic protozoan Leishmania is enhanced by surface 467
metalloprotease gp63. Infect. Immun. 71:1008-1010. 468
26. McGwire, B. S., W. A. O'Connell, K. P. Chang, and D. M. Engman. 2002. Extracellular 469
release of the glycosylphosphatidylinositol (GPI)-linked Leishmania surface 470
metalloprotease, Gp63, is independent of GPI phospholipolysis: implications for parasite 471
virulence. J. Biol. Chem. 277:8802-8809. 472
ACCEPTED
on Decem
ber 2, 2018 by guesthttp://ec.asm
.org/D
ownloaded from

23
27. Miller, M. A., S. E. McGowan, K. R. Gantt, M. Champion, S. L. Novick, K. A. 473
Andersen, C. J. Bacchi, N. Yarlett, B. E. Britigan, and M. E. Wilson. 2000. Inducible 474
resistance to oxidant stress in the protozoan Leishmania chagasi. J. Biol. Chem. 275:33883-475
33889. 476
28. Peacock, C. S., K. Seeger, D. Harris, L. Murphy, J. C. Ruiz, M. A. Quail, N. Peters, E. 477
Adlem, A. Tivey, M. Aslett, A. Kerhornou, A. Ivens, A. Fraser, M. A. Rajandream, T. 478
Carver, H. Norbertczak, T. Chillingworth, Z. Hance, K. Jagels, S. Moule, D. Ormond, 479
S. Rutter, R. Squares, S. Whitehead, E. Rabbinowitsch, C. Arrowsmith, B. White, S. 480
Thurston, F. Bringaud, S. L. Baldauf, A. Faulconbridge, D. Jeffares, D. P. Depledge, S. 481
O. Oyola, J. D. Hilley, L. O. Brito, L. R. Tosi, B. Barrell, A. K. Cruz, J. C. Mottram, D. 482
F. Smith, and M. Berriman. 2007. Comparative genomic analysis of three Leishmania 483
species that cause diverse human disease. Nat. Genet. 39:839-847. 484
29. Porter, K. E., N. A. Turner, D. J. O'Regan, and S. G. Ball. 2004. Tumor necrosis factor 485
alpha induces human atrial myofibroblast proliferation, invasion and MMP-9 secretion: 486
inhibition by simvastatin. Cardiovasc. Res. 64:507-515. 487
30. Preiser, P. R., W. Jarra, T. Capiod, and G. Snounou. 1999. A rhoptry-protein-associated 488
mechanism of clonal phenotypic variation in rodent malaria. Nature 398:618-622. 489
31. Pucadyil, T. J., P. Tewary, R. Madhubala, and A. Chattopadhyay. 2004. Cholesterol is 490
required for Leishmania donovani infection: implications in leishmaniasis. Mol. Biochem. 491
Parasitol. 133:145-152. 492
32. Purdy, J. E., J. E. Donelson, and M. E. Wilson. 2005. Regulation of genes encoding the 493
major surface protease of Leishmania chagasi via mRNA stability. Mol. Biochem. Parasitol. 494
142:88-97. 495
ACCEPTED
on Decem
ber 2, 2018 by guesthttp://ec.asm
.org/D
ownloaded from

24
33. Ramamoorthy, R., J. E. Donelson, K. E. Paetz, M. Maybodi, S. C. Roberts, and M. E. 496
Wilson. 1992. Three distinct RNAs for the surface protease gp63 are differentially 497
expressed during development of Leishmania donovani chagasi promastigotes to an 498
infectious form. J. Biol. Chem. 267:1888-1895. 499
34. Ravanko, K., K. Jarvinen, J. Helin, N. Kalkkinen, and E. Holtta. 2004. Cysteine 500
cathepsins are central contributors of invasion by cultured adenosylmethionine 501
decarboxylase-transformed rodent fibroblasts. Cancer Res. 64:8831-8838. 502
35. Roberts, S. C., K. G. Swihart, M. W. Agey, R. Ramamoorthy, M. E. Wilson, and J. E. 503
Donelson. 1993. Sequence diversity and organization of the msp gene family encoding gp63 504
of Leishmania chagasi. Mol. Biochem. Parasitol. 62:157-171. 505
36. Sacks, D. L. 1989. Metacyclogenesis in Leishmania promastigotes. Exp. Parasitol. 69:100-506
103. 507
37. Santos, A. L., M. H. Branquinha, and C. M. D'Avila-Levy. 2006. The ubiquitous gp63-508
like metalloprotease from lower trypanosomatids: in the search for a function. An. Acad. 509
Bras. Cienc. 78:687-714. 510
38. Snounou, G., W. Jarra, and P. R. Preiser. 2000. Malaria multigene families: the price of 511
chronicity. Parasitol. Today 16:28-30. 512
39. Victoir, K., J. Arevalo, S. De Doncker, D. C. Barker, T. Laurent, E. Godfroid, A. 513
Bollen, D. Le Ray, and J. C. Dujardin. 2005. Complexity of the major surface protease 514
(msp) gene organization in Leishmania (Viannia) braziliensis: evolutionary and functional 515
implications. Parasitology 131:207-214. 516
ACCEPTED
on Decem
ber 2, 2018 by guesthttp://ec.asm
.org/D
ownloaded from

25
40. von Tresckow, B., K. J. Kallen, E. P. von Strandmann, P. Borchmann, H. Lange, A. 517
Engert, and H. P. Hansen. 2004. Depletion of cellular cholesterol and lipid rafts increases 518
shedding of CD30. J. Immunol. 172:4324-4331. 519
41. Voth, B. R., B. L. Kelly, P. B. Joshi, A. C. Ivens, and W. R. McMaster. 1998. 520
Differentially expressed Leishmania major gp63 genes encode cell surface leishmanolysin 521
with distinct signals for glycosylphosphatidylinositol attachment. Mol. Biochem. Parasitol. 522
93:31-41. 523
42. Weise, F., Y. D. Stierhof, C. Kuhn, M. Wiese, and P. Overath. 2000. Distribution of GPI-524
anchored proteins in the protozoan parasite Leishmania, based on an improved 525
ultrastructural description using high-pressure frozen cells. J. Cell Sci. 113 Pt 24:4587-4603. 526
43. Wilson, M. E., K. K. Hardin, and J. E. Donelson. 1989. Expression of the major surface 527
glycoprotein of Leishmania donovani chagasi in virulent and attenuated promastigotes. J 528
Immunol 143:678-684. 529
44. Wilson, M. E., K. E. Paetz, R. Ramamoorthy, and J. E. Donelson. 1993. The effect of 530
ongoing protein synthesis on the steady state levels of Gp63 RNAs in Leishmania chagasi. J. 531
Biol. Chem. 268:15731-15736. 532
45. Yao, C., J. E. Donelson, and M. E. Wilson. 2003. The major surface protease (MSP or 533
GP63) of Leishmania sp. Biosynthesis, regulation of expression, and function. Mol. 534
Biochem. Parasitol. 132:1-16. 535
46. Yao, C., K. G. Leidal, A. Brittingham, D. E. Tarr, J. E. Donelson, and M. E. Wilson. 536
2002. Biosynthesis of the major surface protease GP63 of Leishmania chagasi. Mol. 537
Biochem. Parasitol. 121:119-128. 538
ACCEPTED
on Decem
ber 2, 2018 by guesthttp://ec.asm
.org/D
ownloaded from

26
47. Yao, C., J. Luo, C. Hsiao, J. E. Donelson, and M. E. Wilson. 2005. Internal and surface 539
subpopulations of the major surface protease (MSP) of Leishmania chagasi. Mol. Biochem. 540
Parasitol. 139:173-183. 541
48. Yao, C., J. Luo, P. Storlie, J. E. Donelson, and M. E. Wilson. 2004. Multiple products of 542
the Leishmania chagasi major surface protease (MSP or GP63) gene family. Mol. Biochem. 543
Parasitol. 135:171-183. 544
49. Zarley, J. H., B. E. Britigan, and M. E. Wilson. 1991. Hydrogen peroxide-mediated 545
toxicity for Leishmania donovani chagasi promastigotes. Role of hydroxyl radical and 546
protection by heat shock. J. Clin. Invest. 88:1511-1521. 547
ACCEPTED
on Decem
ber 2, 2018 by guesthttp://ec.asm
.org/D
ownloaded from

27
Figure Legends 548
Figure 1. Homogeneity of MSP expression in clonal lines of L. chagasi. Single clones were 549
selected from M199 plates prepared from virulent L. chagasi promastigote populations. After 550
expanding, total cell lysates of stationary phase promastigote clones were subjected to IEF (A) or 551
2-DE (B) and transfer to nitrocellulose membranes. MSP proteins were detected by 552
immunoblotting. (A) immunoblot of total cellular proteins separated by IEF in a lysate of 553
stationary phase cells from the uncloned parental population (lane 1) or from four representatives 554
from a total of 124 clones (lanes 2-5). (B) a representative of the 2-DE MSP profiles conducted 555
on two dozen individual clones. Eleven previously described MSP isoforms from uncloned 556
promastigotes are labeled for reference (47, 48). 557
Figure 2. Surface-localized MSPs of attenuated versus virulent L. chagasi are differentially 558
regulated. (A) Example of a pulse-chase experiment demonstrating the rates of surface or 559
internal MSP loss from virulent L. chagasi promastigotes. Cultured promastigotes on day 3 (L) 560
or day 7 (S) of in vitro growth were metabolically labeled with [35S]-methionine + -cysteine and 561
surface biotinylated, followed by a chase without radioactivity for up to 72 h. Biotinylated 562
surface proteins were separated from the nonbiotinylated internal proteins using streptavidin 563
beads, and internal MSP was immunoprecipitated from bead-cleared fractions. Autoradiograms 564
are shown. The 66-kDa isoform seen at 0 h in the nonbiotinylated samples is most likely MSP 565
containing its propeptide (23). The T½ of the 63-kDa surface biotinylated MSP was determined 566
by a linear regression of f(x)=a+bx, using densitometric analysis of the bands. Graphic results 567
shown are the average ± SD of densitometric measurements of four independent experiments. 568
*P<0.01 by paired student t-test. (B) Rates of MSP synthesis in attenuated L5 versus virulent L. 569
chagasi were measured during logarithmic (L) or stationary (S) phase growth were determined 570
ACCEPTED
on Decem
ber 2, 2018 by guesthttp://ec.asm
.org/D
ownloaded from

28
by metabolically labeling promastigote proteins with [35S]-methionine + -cysteine and 571
immunoprecipitation with polyclonal antisera to MSP or P36. The ratio of MSP to P36 was 572
standardized to virulent stationary phase promastigotes, which was at one arbitrary unit. Data 573
shown are representative of two independent experiments. (C) Release of surface-localized MSP 574
into the extracellular medium. Attenuated L5 or virulent L. chagasi promastigotes during 575
logarithmic (L) or stationary (S) phase growth were surface biotinylated and resuspended in 576
fresh medium. Extracellular medium was collected 48 h later. MSP was detected in the 577
streptavidin-bead pull-down fraction by western blotting. Shown is the relative MSP abundance 578
standardized to stationary phase virulent promastigotes. One of two independent experiments is 579
shown. 580
Figure 3. MβCD enhances a dose-dependent release of surface-localized MSP. (A and B) 581
Virulent promastigotes on different days of growth were treated with the indicated concentration 582
of MβCD (0-15 mM) for 48 h. Filtered supernatants and cells were subjected to SDS-583
polyacrylamide gel electrophoresis. MSP was detected by immunoblotting. One of three 584
independent experiments, each of which was conducted in triplicate, is shown. (A) Extracellular 585
(E) and cellular (C) MSP content in parasites incubated with the indicated MβCD concentration 586
are shown. (B) The abundance of extracellular MSP in the medium of cells shown in panel (A) 587
was quantified by densitometric analysis. Values represent the percentage ([extracellular/total 588
MSP] × 100%) for each concentration. Statistical analysis on the 0 versus 10 or 15 mM MβCD: 589
*, p<0.05; **, p<0.01. (C) MβCD effect on total incorporation of [35S]-methionine + -cysteine 590
into newly synthesized proteins. MβCD-treated cells were metabolically labeled and newly 591
synthesized total protein was monitored as described in Materials and Methods. Values 592
represent the total incorporation of radioisotope into the newly synthesized proteins of treated 593
ACCEPTED
on Decem
ber 2, 2018 by guesthttp://ec.asm
.org/D
ownloaded from

29
parasites relative to the controls (0 mM MβCD). One of two independent experiments, with 594
three replicate conditions, is shown. (D) MβCD treatment enhances release of surface-localized 595
MSP into the extracellular medium. Stationary phase virulent L. chagasi promastigotes were 596
surface biotinylated in HBSS and incubated for 3 h at RT in the absence (0 mM) or presence (15 597
mM) of MβCD, as described in Materials and Methods. Biotinylated (S) and nonbiotinylated (I) 598
MSPs were isolated from filtered spent-medium with streptavidin beads and 599
immunoprecipitation, respectively. MSP was detected by immunoblotting. To monitor for cell 600
lysis, the cytosolic protein P36 was measured in the cleared fraction post biotin-streptavidin 601
affinity and MSP immunoprecipitation by immunoblot. Total cell lysates (cell) were included. 602
One of three independent replicate experiments is shown. 603
Figure 4. Release of internal MSP. (A) Stationary phase promastigotes in the first passage after 604
isolation from hamsters were incubated in HBSS (control, 3 h) or the Matrigel matrix for 1 or 605
3 h after surface-biotinylation at either 37oC or RT. After transfer to 4oC, parasites were 606
removed by centrifugation. Streptavidin beads were used to isolate biotinylated surface MSP 607
from both the whole cell lysate and the extracellular medium. The internal MSP was purified 608
from streptavidin-bead cleared fractions by immunoprecipitation, and was detected by western 609
blotting. Each condition was performed in triplicate. (B-C) Quantitation of released and internal 610
MSP. The band intensity of the extracellular (B) and internal (C) fractions for each experiment 611
in (A) was determined by densitometric analysis, and the average and standard deviation of the 612
triplicate samples were determined. Results from one of three experiments are presented. 613
Figure 5. A model of MSP release and possible functions during metacyclogenesis in the sand 614
fly vector and inoculation into the mammalian hosts. Solid and open circles represent surface-615
localized and internal MSP, respectively. Arrows show MSP being released into the 616
ACCEPTED
on Decem
ber 2, 2018 by guesthttp://ec.asm
.org/D
ownloaded from

30
extracellular environment. The width of the arrow is proportional to the amount of released 617
MSP. In the sand fly panel, the upper diagram depicts MSP release from procyclic (logarithmic 618
growth phase) promastigotes and the lower diagram depicts release from their metacyclic 619
(stationary phase) counterparts. In the mammalian host, a metacyclic promastigote is depicted. 620
MØ: macrophage. 621
ACCEPTED
on Decem
ber 2, 2018 by guesthttp://ec.asm
.org/D
ownloaded from