lineage choice and differentiation in mouse embryos and embryonic stem cells
TRANSCRIPT

Review
Lineage choice and differentiation in mouse embryos andembryonic stem cells
David A.F. Loebel, Catherine M. Watson, R. Andrea De Young, and Patrick P.L. Tam*Embryology Unit, Children’s Medical Research Institute, Locked Bag 23, Wentworthville, NSW 2145 Australia
Received for publication 21 February 2003, revised 21 May 2003, accepted 11 June 2003
Abstract
The use of embryonic stem (ES) cells for generating healthy tissues has the potential to revolutionize therapies for human disease orinjury, for which there are currently no effective treatments. Strategies for manipulating stem cell differentiation should be based onknowledge of the mechanisms by which lineage decisions are made during early embryogenesis. Here, we review current research into thefactors influencing lineage differentiation in the mouse embryo and the application of this knowledge to in vitro differentiation of ES cells.In the mouse embryo, specification of tissue lineages requires cell–cell interactions that are influenced by coordinated cell migration andcellular neighborhood mediated by the key WNT, FGF, and TGF� signaling pathways. Mimicking the cellular interactions of the embryoby providing appropriate signaling molecules in culture has enabled the differentiation of ES cells to be directed predominately towardparticular lineages. Multistep strategies incorporating the provision of soluble factors known to influence lineage choices in the embryo,coculture with other cells or tissues, genetic modification, and selection for desirable cell types have allowed the production of ES cellderivatives that produce beneficial effects in animal models. Increasing the efficiency of this process can only result from a betterunderstanding of the molecular control of cell lineage determination in the embryo.© 2003 Elsevier Inc. All rights reserved.
Keywords: FGF; WNT; BMP; Stem cell; Germ cell; Differentiation; Lineage; Pluripotent; Mouse; Embryo
Introduction
There is considerable interest in the therapeutic applica-tions of pluripotent cells derived from embryos or adulttissues. Consequently, much effort is being directed towardunderstanding the mechanisms for maintenance of the plu-ripotent state and the pathways leading to lineage specifi-cation. Control of these mechanisms may enable the ma-nipulation of pluripotent cells for the development of cell-and tissue-based replacement therapies for damaged ordysfunctional tissues. The ability to manipulate the differ-entiation of embryo-derived pluripotent cells requires athorough understanding of the processes of lineage commit-ment and differentiation of embryonic cells in vivo. In thisreview, we outline the current state of knowledge of themolecular and cellular basis of the determination and dif-
ferentiation of tissue lineages in the early mouse embryoand describe the application of this knowledge to in vitroembryonic stem (ES) cell differentiation.
Around the time of implantation, the mouse embryo iscomposed of three distinct cell types (Fig. 1A and B): thetrophectoderm, the primitive endoderm, and the inner cellmass (ICM). The trophectoderm gives rise to the chorionand the trophoblast, which differentiates into the giant cells,the cytotrophoblast and syncytiotrophoblast of the placenta.Primitive endoderm forms the visceral and parietalendoderm that lines the yolk sac cavity and the exocoelom.The ICM is organized into a pluripotent epithelial layer (theepiblast) from which embryonic and other extraembryonictissues are derived (Gardner and Beddington, 1988).
Gastrulation is a major milestone in early postimplanta-tion development. During gastrulation, pluripotent epiblastcells are allocated to the three primary germ layers of theembryo (ectoderm, mesoderm, and definitive endoderm)and the extraembryonic mesoderm of the yolk sac and
* Corresponding author. Fax: �61-2-96872120.E-mail address: [email protected] (P.P.L. Tam).
R
Available online at www.sciencedirect.com
Developmental Biology 264 (2003) 1–14 www.elsevier.com/locate/ydbio
0012-1606/$ – see front matter © 2003 Elsevier Inc. All rights reserved.doi:10.1016/S0012-1606(03)00390-7
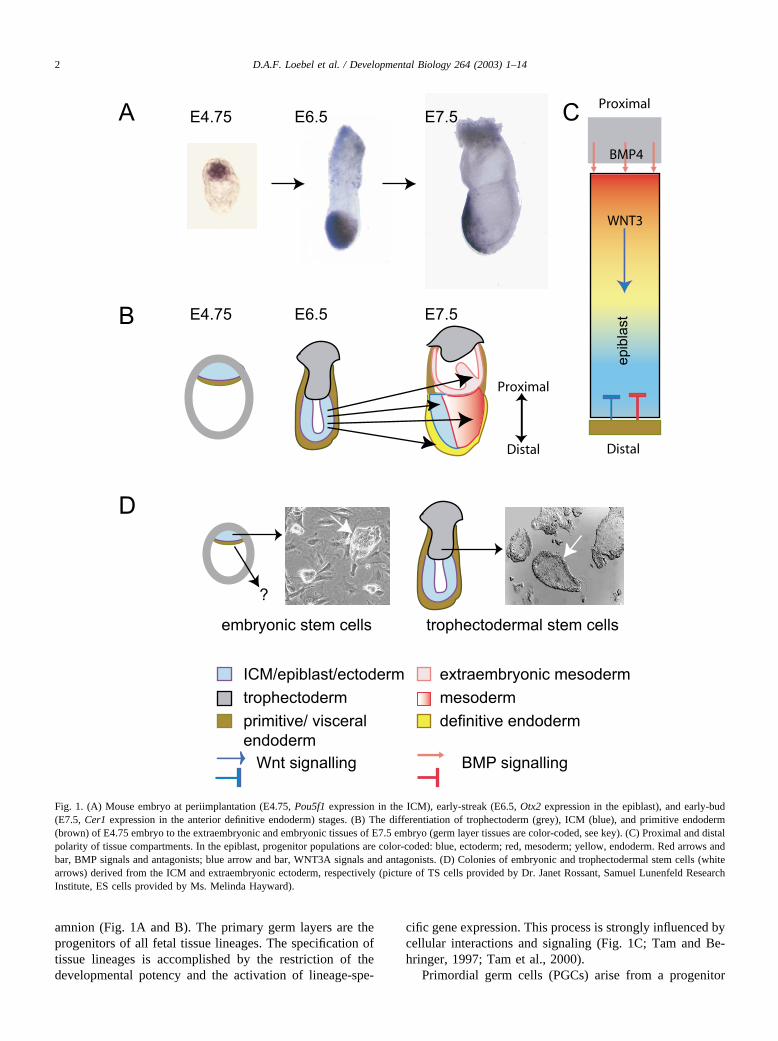
amnion (Fig. 1A and B). The primary germ layers are theprogenitors of all fetal tissue lineages. The specification oftissue lineages is accomplished by the restriction of thedevelopmental potency and the activation of lineage-spe-
cific gene expression. This process is strongly influenced bycellular interactions and signaling (Fig. 1C; Tam and Be-hringer, 1997; Tam et al., 2000).
Primordial germ cells (PGCs) arise from a progenitor
Fig. 1. (A) Mouse embryo at periimplantation (E4.75, Pou5f1 expression in the ICM), early-streak (E6.5, Otx2 expression in the epiblast), and early-bud(E7.5, Cer1 expression in the anterior definitive endoderm) stages. (B) The differentiation of trophectoderm (grey), ICM (blue), and primitive endoderm(brown) of E4.75 embryo to the extraembryonic and embryonic tissues of E7.5 embryo (germ layer tissues are color-coded, see key). (C) Proximal and distalpolarity of tissue compartments. In the epiblast, progenitor populations are color-coded: blue, ectoderm; red, mesoderm; yellow, endoderm. Red arrows andbar, BMP signals and antagonists; blue arrow and bar, WNT3A signals and antagonists. (D) Colonies of embryonic and trophectodermal stem cells (whitearrows) derived from the ICM and extraembryonic ectoderm, respectively (picture of TS cells provided by Dr. Janet Rossant, Samuel Lunenfeld ResearchInstitute, ES cells provided by Ms. Melinda Hayward).
2 D.A.F. Loebel et al. / Developmental Biology 264 (2003) 1–14

population in the epiblast around E5.5 and give rise to themature spermatozoa and ova of adults. The germ cell lin-eage is unusual because it differentiates into a highly spe-cialized cell type, but must retain the totipotent property thatis required by the zygote of the next generation. The pro-genitor cells of this lineage respond selectively to specificsignals promoting differentiation into PGCs. They are thensequestered into the base of the allantois to avoid beingswept up into the dynamic embryonic milieu of gastrulation.Eventually, they migrate to the genital ridges and displaysex-specific differentiation before maturing into gametes.
Embryonic stem (ES) cells can be derived in vitro fromthe ICM of the periimplantation embryo (Fig. 1D). Like theICM and the early epiblast, ES cells are capable of differ-entiating into a wide range of fetal tissues in culture (Ros-sant and McKerlie, 2001) or in chimeric embryos and caneven form the entire fetus (Nagy et al., 1993). However,significant contribution to either the trophectoderm or vis-ceral endodermal lineages by ES cells has not been demon-strated.
Embryonic germ cell (EG) lines can be established frompremigratory (E8.0 and E8.5), migratory (E9.5), and post-migratory PGCs that have arrived at the genital ridge(E11.5–E12.5). EG cells are pluripotent and can contributeto embryonic cell lineages in chimeras (Labosky et al.,1994). Many of the cellular specializations of the PGC, suchas pseudopodia, disappear in culture, and EG cells resembleES cells in morphology regardless of the stage of develop-
ment from which they were established (reviewed inMcLaren and Durcova-Hills, 2001).
Trophectodermal stem (TS) cells (Fig. 1D), with themolecular properties of diploid trophoblast cells, may beisolated from the blastocyst or extraembryonic ectoderm ofperiimplantation embryos. TS cells have a restricted differ-entiation potential: they can form giant cells in vitro(Tanaka et al., 1998; Yan et al., 2001) and have only beenobserved to contribute to trophectodermal cell lineages invivo. Whereas ES cells can be induced to become TS cells(Niwa et al., 2000), TS cells have not been shown to acquirethe lineage potential of ES cells. As yet, there are nopublished reports of stem cells derived from the third celltype, the primitive endoderm of the embryo, but this re-mains an exciting possibility.
Maintaining pluripotency
Pluripotency of embryonic cells refers to their ability todifferentiate into cell types of many tissue lineages. Under-standing the molecular basis of pluripotency and the con-ditions required to maintain this state in the early embryo isvital for an understanding of the maintenance of pluripotentcells in culture.
The transcription factor OCT4, encoded by Pou5f1, is anessential regulator of preimplantation mouse developmentand is expressed in pluripotent cells: oocytes, the ICM, early
Fig. 2. Establishment of the germ-cell lineage in prestreak to late streak mouse embryos: showing the signaling and mediating molecules that influence (A)priming, (B) specification and localization of cells derived from the proximal epiblast, and (C) commitment of the PGCs prior to and during gastrulation(Lawson et al., 1986, 1999; Tremblay et al., 1995, 2001; Hayashi et al., 1997, 2002; Tsang et al., 1999, 2001; Chang et al., 1999; Ying et al., 2000; Yingand Zhao, 2001; Mukhopadhyay et al., 2001, 2003; Chang and Matzuk, 2001; Saitou et al., 2002).
3D.A.F. Loebel et al. / Developmental Biology 264 (2003) 1–14

preimplantation embryos, primitive ectoderm, PGCs, andES cells (Niwa, 2001). OCT4 is essential for the formationof the ICM in vivo (Nichols et al., 1998). Epiblast celldifferentiation is accompanied by repression of Pou5f1,which is mediated by the binding of germ cell nuclear factor(GCNF) to the proximal promoter of Pou5f1. This results inthe restriction of Pou5f1 expression to the germ cell lineage(Fuhrmann et al., 2001). In vitro studies of ES cells haveshown that developmental regulation is effected by varyingOCT4 expression levels. Thus, repression of Pou5f1 in-duces trophectodermal differentiation, and elevated Pou5f1expression induces endodermal and mesodermal differenti-ation, whereas intermediate Pou5f1 levels are required forthe maintenance of pluripotency of ES cells in vitro (Niwaet al., 2000).
OCT4 can act as a repressor or activator of transcriptiondepending on the presence of its cofactors (Niwa, 2001).SOX2 is a potential coactivator for OCT4 (Ambrosetti et al.,1997). Candidate targets of OCT4 and SOX2 include Fgf4and Hand1, which are associated with maintenance anddifferentiation of trophoblasts (Niwa, 2001; Avilion et al.,2003). The developmental defects displayed by embryoslacking Sox2 function suggest that both Sox2 and Pou5f1 areimportant for correct specification of the ICM, the primitiveendoderm, and the trophectoderm (Avilion et al., 2003).
In culture, ES cells remain undifferentiated in the pres-ence of the cytokine LIF. LIF acts via a receptor comprisingthe LIF-specific subunit LIFR� and gp130 and can be trans-duced by the JAK-STAT signaling pathway. STAT3 seemsto be the principal component of the signaling process in EScells (Raz et al., 1999). However, an uncharacterized EScell renewal factor (ESRF) can maintain the pluripotency ofES cells without activating Stat3 (Niwa et al., 1998).Stat3�/� embryos develop until E6.0 and then rapidly de-generate at gastrulation (Takeda et al., 1997). Until ESRF ischaracterized, it will remain unclear whether both Stat3-dependent and -independent pathways are critical for main-taining pluripotency in the embryo.
ES cells can differentiate into early primitive ectoderm-like (EPL) cells when cultured in conditioned medium(Rathjen et al., 1999). EPL cells are a step closer to ecto-dermal commitment and, unlike ES cells, EPL cells areunable to contribute to tissues in chimeric embryos (Rathjenet al., 1999; Pelton et al., 2002). EPL cells resemble theepiblast of the embryo in that they continue to expressPou5f1, and the differences in gene expression between ESand EPL cells recapitulate the differences between the ICMand epiblast. EPL cell-derived embryoid bodies are predis-posed to differentiation to mesoderm (Lake et al., 2000), butcan be redirected toward a neural fate by continued expo-sure to conditioned medium (Rathjen et al., 2002). With-drawal of conditioned medium from EPL cell culture resultsin reversion to an ES cell-like state, including the potentialfor in vitro differentiation and the ability to contribute totissues in chimeric mice (Rathjen et al., 1999; Lake et al.,2000).
Establishment of the germ line is a multistep process
Specification of primordial germ cells begins in the epi-blast of periimplantation embryos. PGCs continue to ex-press Pou5f1 and can restore the totipotent propertiesunique to germ cells during gametogenesis. The formationof the PGC population from proximal epiblast progressesthrough multiple stages, requiring BMP signaling, cell–cellinteraction, and cell movement (Fig. 2). BMP4 signalingfrom the extraembryonic ectoderm primes proximal epiblastcells to be capable of attaining either a germ cell or allantoicfate.
BMP2, �4, and �8B, produced by the extraembryonictissues in the vicinity of the proximal epiblast of the pre-streak embryo, are necessary for specification of germ cells(Fig. 2A and B). Bmp4�/� mice have no PGCs, whereasBmp4�/� mice have reduced numbers, indicating thatBMP4 acts in a dose-dependent manner to induce PGCs(Lawson et al., 1999). Bmp2�/� null embryos have reducednumbers of PGCs and Bmp4�/�; Bmp2�/� double heterozy-gotes have fewer PGCs than either Bmp4�/� or Bmp2�/�
embryos. Bmp2 and Bmp4 therefore exert an additive effecton PGC specification. In contrast, Bmp2 and Bmp8b do notdisplay any additive effects (Ying and Zhao, 2001). Ho-modimers of BMP4 and BMP8B (Ying et al., 2000) actsynergistically to induce PGCs in the epiblast in vitro (Yinget al., 2001). Coculture of epiblast and visceral endoderm inthe presence of BMP4 leads to stronger expression of TissueNonspecific Alkaline Phosphatase (TNAP), a germ cellmarker, in the epiblast cells adjacent to visceral endodermrelative to cultures lacking BMP4. Although it has not beentested whether the visceral endoderm in the coculture pro-vides a source of BMP2, the data are nevertheless consistentwith a role for combined BMP2 and BMP4 action in theinduction of PGCs (Pesce et al., 2002). Thus, in vivo and invitro data show that BMP2, �4, and �8B proteins all playa role in PGC induction.
Close proximity of the extraembryonic ectoderm to theepiblast during a narrow window of development is essen-tial for germ cell specification in vivo. Distal epiblast cellsform PGCs when placed adjacent to extraembryonicectoderm in vivo (Tam and Zhou, 1996) or in culture(Yoshimizu et al., 2001). The interaction with extraembry-onic ectoderm is only required for PGC production by theepiblast of E5.5 embryos but not by E6.0 epiblast, and thetissue requirement can be replaced by BMP4 (Fig. 2A;Yoshimizu et al., 2001).
Smad1 and Smad5 are important for germ cell specifica-tion and act downstream of BMP receptors in the germ cellprecursors. Smad1 is expressed in epiblast and visceralendoderm during gastrulation and is expressed in about 25%of proximal epiblast cells. Only a minority of these cellscoexpress Smad5 or �8, but it is postulated that these cellsare the PGC progenitors (Hayashi et al., 2002). Smad1�/�
embryos fail to develop an allantois and have very lownumbers of PGCs, whereas heterozygotes are similar to wild
4 D.A.F. Loebel et al. / Developmental Biology 264 (2003) 1–14

type embryos (Tremblay et al., 2001). Smad5�/� embryosalso have very few or no PGCs, whereas Smad5�/� havefewer PGCs than wild type embryos, showing that Smad5 isalso involved in germ cell specification in a dose-dependentmanner similar to BMP4 (Fig. 2B; Chang et al., 2001).
Interactions within a community of epiblast cells may beimportant for germ cell specification (Fig. 2B). Induction ofPGCs is more efficient in epiblast fragments containing atleast 40 cells or in closely packed epiblast explants onfeeder layers than in dissociated epiblast cells (Yoshimizu etal., 2001; Pesce et al., 2002). fragilis/mil1, fragilis2/mil2,and fragilis3/mil3 are members of a gene family encodinginterferon-induced transmembrane proteins, which in micecomprises five members (Saitou et al., 2002; Tanaka andMatsui, 2002). This gene family is conserved in other spe-cies and is thought to be involved in homotypic cell adhe-sion and extension of cell cycle time. fragilis is highlyexpressed in the proximal epiblast and gradually becomesrestricted to a group of cells in the posterior PS coincidentwith the site where PGCs arise and TNAP and also fragilis2and �3 are expressed. fragilis expression can be induced indistal epiblast cells by coculturing them with extraembry-onic ectoderm. Its expression is reduced in Bmp4�/� andabsent in Bmp4�/� embryos, indicating a dose-dependentinduction of fragilis by BMP4 (Saitou et al., 2002). Thus,fragilis, fragilis2, and fragilis3 are potentially mediators ofcell–cell interactions in PGC specification in the mouse(Lange et al., 2003).
PGCs must be localized to the base of the allantoisduring gastrulation to avoid a somatic cell fate. The pro-gressive shift in the location of fragilis-expressing epiblastcells is consistent with the general pattern of cell movementduring gastrulation followed by isolation of the progenitorPGC population to the base of the allantois, where theirproliferation is reduced relative to the somatic cells that passthrough the PS (Tam and Snow, 1981; Saitou et al., 2002).Bmp2 is expressed just prior to and during gastrulation inthe visceral endoderm overlying the ectoderm–epiblastboundary in the region where the PS forms. Its expressionpattern also makes it a candidate for a localization signal,together with Bmp4 for the PGC progenitors (Fig. 2B; Yingand Zhao, 2001; Fujiwara et al., 2001). Cells expressingfragilis congregate in the posterior PS and allantois ofE7.0–E7.5 late streak stage embryos. At this site, some ofthe fragilis-expressing cells begin to express stella (alsoreferred to as PGC7 and Dppa3; Saitou et al., 2002). Ex-pression of stella, which encodes a putative chromosomalorganization or RNA processing factor, may mark the emer-gence of a committed PGC population (Saitou et al., 2002;Sato et al., 2002).
Specification of the germ line is a multistep process (Fig.2). Progenitors for PGCs in the epiblast are initially primedby BMP4 signals from the extraembryonic ectoderm atE5.5. The specification of the PGCs is influenced by theaction of BMPs and SMADs and by the interactions be-tween cells, which may be mediated by the product of the
fragilis gene. PGC progenitors need to be in the right placeat the right time to respond to local signals. Commitment tothe germ-cell lineage is accomplished by spatial sequestra-tion of the PGCs, reinforcement by localized signals, andinduction of germ line-specific gene expression.
Specification of the primary germ layer tissues
BMP signaling is essential for the choice betweenectodermal and mesodermal fates
BMP signaling is required to induce mesoderm forma-tion in vivo (Fig. 3, red area). Most Bmp4�/� embryos failto gastrulate and form no mesoderm (Winnier et al., 1995).Loss of Bmpr1 and Bmpr2b function also results in reducedepiblast proliferation and no formation of mesoderm (Wil-son et al., 1993; Mishina et al., 1995). The similarities in thephenotypes of Bmp4, Bmpr1, and Bmpr2 mutants suggestthat BMP4 signals predominantly through heterodimers ofthese receptors (Wilson et al., 1993; Mishina et al., 1995).BMP signaling activates receptor regulated SMADS 1, 5,and 8, which interact with the common partner SMAD4 toregulate gene expression (Miyazawa et al., 2002). Smad4 isrequired for epiblast proliferation, gastrulation, and meso-derm formation (Yang et al., 1998; Sirard et al., 1998).
For correct patterning and differentiation of the embryo,the influence of BMP signaling must be modulated byantagonistic molecules. The BMP antagonists Chordin andNoggin are expressed in the mouse organizer and promotedifferentiation of ectodermal lineages (Fig. 3, blue area).Chordin and Noggin have overlapping functions duringembryogenesis and are required for anterior neural differ-entiation in the embryo (Bachiller et al., 2000).
BMP4 is able to influence ES cells to choose a meso-dermal rather than neurectodermal fate in culture (Fig. 4,red area). Treatment of ES cells with BMP4 inhibits neuraldifferentiation and represses the neurectodermal markerPax6 and upregulates T in a concentration-dependent man-ner, independent of cell survival or proliferation (Johanssonand Wiles, 1995; Wiles and Johansson, 1997; Finley et al.,1999). BMP4 induces differentiation of cells resemblingposterior–ventral embryonic mesoderm, whereas dorsal–an-terior mesodermal cell types are induced by another TGF�-related molecule, Activin A (Johansson and Wiles, 1995).The effectiveness of BMP4 in altering cell fate is limited toa period prior to the onset of neural differentiation in culture(Finley et al., 1999). Under different conditions, BMP4promotes the differentiation of ES cells and EPL cells tosurface ectoderm at the expense of neuroectoderm, under-lining the properties of BMP4 as a suppressor of neurecto-dermal differentiation (Fig. 4, blue area; Kawasaki et al.,2000; Rodda et al., 2002). However, there is no directevidence for a role for BMP4 in surface ectoderm differen-tiation in mouse embryos (Fig. 3).
BMP antagonists promote the acquisition of an ectoder-
5D.A.F. Loebel et al. / Developmental Biology 264 (2003) 1–14

mal fate by ES cells (Fig. 4, blue area). Exposure of ES cellsto recombinant Noggin, which counteracts the mesoderm-inducing effects of BMP4 (Finley et al., 1999), or transfec-tion with a Noggin-encoding plasmid promotes the differ-entiation of neurectodermal cells (Gratsch and O’Shea,2002). Exposure to Chordin results in reduced differentia-tion, with some cells retaining ES cell characteristics and asmall number differentiating to mesenchymal cell types(Gratsch and O’Shea, 2002). Disruption of SMAD4-medi-ated BMP/TGF� signal transduction enhances the ability ofES cells to adopt a neural fate (Tropepe et al., 2001), whichis consistent with the suppression of neural differentiationby BMP signaling.
Induction and differentiation of the neurectodermallineages
Initial specification of neurectoderm from the ectodermof the gastrulating embryo is accomplished by inductiveinteractions with nascent mesoderm and definitive
endoderm. In tissue recombination and coculture experi-ments, anterior ectoderm can be induced to express neuralmarkers (Otx2 and En1) by the prospective anterior (headand axial) mesoderm and foregut endoderm. Inductive in-teractions are also required to maintain expression of neuralgenes. In contrast, the posterior mesoderm and endodermsuppresses the expression of neural markers in the anteriorectoderm (Ang and Rossant, 1993; Ang et al., 1994). How-ever, the nature of the inductive and suppressive activity isnot known.
The Sry-related HMG box-containing transcription fac-tor Sox1 is expressed during early neurogenesis, becomingprogressively downregulated as cells exit from mitosis(Pevny et al., 1998; Wood and Episkopou, 1999). Sox1 isupregulated in response to retinoic acid (RA)-induced neu-ral differentiation in cultured ectodermal cells, and whenectopically expressed, can substitute for treatment with RA.This suggests that Sox1 plays a role in initially directingectodermal cells toward a neural fate in response to externalsignals (Pevny et al., 1998).
Fig. 3. Specification and differentiation of germ layer derivatives in mouse embryos. Ectodermal (blue), mesodermal (red), and definitive endodermal (orange)lineages derived from the epiblast (brown) are shown, with the signaling pathways and transcription factors indicated in green. The question mark signifiesthe lack of direct evidence for involvement of BMP4 in surface ectoderm differentiation in mouse embryos (Harrison et al., 1995, 1999; Ahlgren et al., 1996,1997).
6 D.A.F. Loebel et al. / Developmental Biology 264 (2003) 1–14
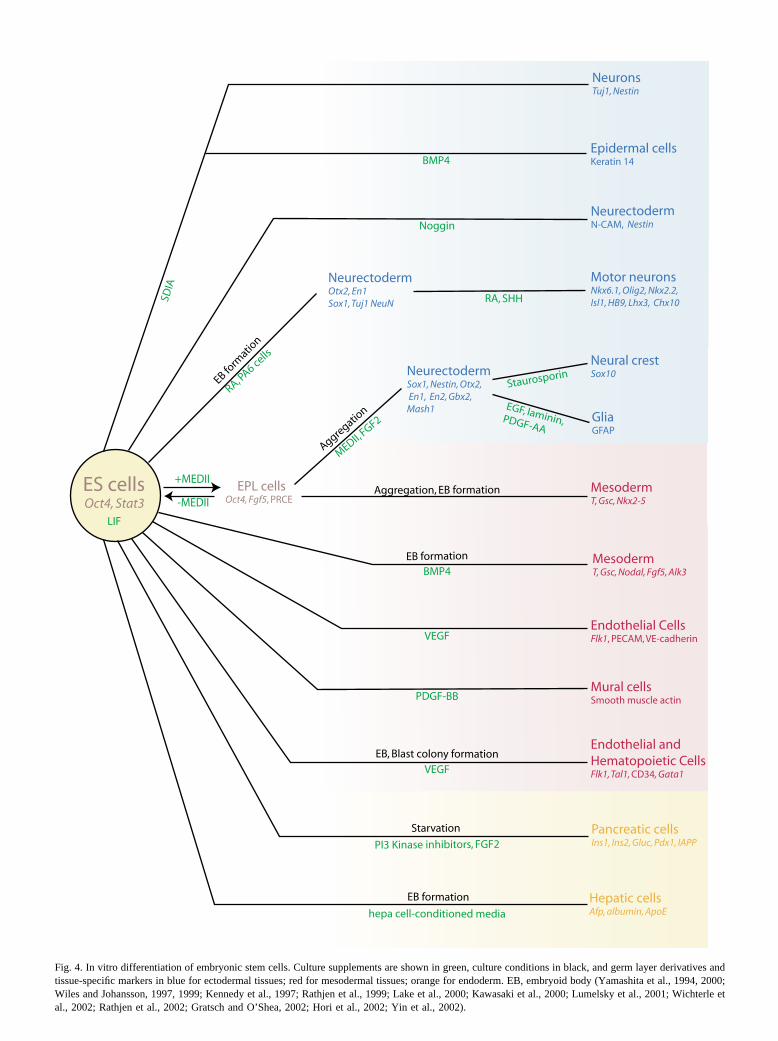
Fig. 4. In vitro differentiation of embryonic stem cells. Culture supplements are shown in green, culture conditions in black, and germ layer derivatives andtissue-specific markers in blue for ectodermal tissues; red for mesodermal tissues; orange for endoderm. EB, embryoid body (Yamashita et al., 1994, 2000;Wiles and Johansson, 1997, 1999; Kennedy et al., 1997; Rathjen et al., 1999; Lake et al., 2000; Kawasaki et al., 2000; Lumelsky et al., 2001; Wichterle etal., 2002; Rathjen et al., 2002; Gratsch and O’Shea, 2002; Hori et al., 2002; Yin et al., 2002).

Expression of posterior Hox genes and SHH activity areassociated with differentiation of ventral motor neurons inthe caudal neural tube. In the embryo, neurectoderm ini-tially has a rostral character and requires the action of RA toimpart the caudal characteristics of spinal cord, presumablyby the activation of the Hox genes specific to posteriorneural tube. Subsequently, graded SHH signaling, mediatedby the transcriptional repressor Gli3, is required to inducemotor neuron progenitors (Fig. 3, blue area; Persson et al.,2002).
Several groups have reported the generation of neurec-todermal lineages from ES cells by the provision of factors,including FGF, SHH, and RA, that affect neural differenti-ation in vivo. Using Sox1-GFP expression as a marker, Yinget al. (2003) demonstrated that differentiation of ES cells toneurectodermal lineages could be accomplished withoutembryoid body formation. Addition of FGF4 stimulated EScell differentiation to neurectodermal cells, and inhibition ofFGF signaling abolished the differentiation of pluripotentcells to neurectoderm (Ying et al., 2003). Treating embryoidbodies with RA and either a HH agonist or recombinantSHH allows the differentiation of ES cells to motor neuronsin vitro, presumably by mimicking the conditions that pro-mote the differentiation of motor neurons in vivo (Fig. 4,blue area; Wichterle et al., 2002). Multistep strategies in-volving the application of signaling molecules (FGF8 andSHH), and the upregulation of Nurr1, a neuronal differen-tiation-related transcription factor (Kim et al., 2002) orselection for cells that express Nestin (Nishimura et al.,2003) have enabled the generation of dopaminergic neuronsin vitro.
Molecular control of mesodermal differentiation
WNT3A signaling induces cells from the epiblast toadopt a paraxial mesodermal rather than a neurectodermalfate after gastrulation (Yoshikawa et al., 1997). InWnt3a�/� embryos, epiblast cells ingressing into the PS arediverted to a neurectodermal fate instead of forming meso-dermal cells. Consequently, ectopic structures resemblingneural tubes are formed in place of the somites (Yoshikawaet al., 1997).
Regulation of genes acting downstream of WNT3A sig-naling requires the transcription factors Lef1 and Tcf1.Mouse mutants lacking both Lef1 and Tcf1 develop ectopicneural tubes and are defective in paraxial mesoderm differ-entiation (Galceran et al., 1999). WNT3a signaling, viaLef1/Tcf1, maintains expression of the mesodermal tran-scription factor T in presumptive paraxial mesoderm(Yamaguchi et al., 1999). Maintenance of T expression canbe rescued by a constitutively active LEF1-�-catenin fusionprotein (Galceran et al., 2001).
The WNT signaling pathway interacts with the FGFpathway to induce mesoderm differentiation (Fig. 3, redarea). Fgfr1 is required for expression of T and Tbx6 (Chap-man and Papaioannou, 1998; Ciruna and Rossant, 2001). In
chimeric embryos, ectopic neural tubes form entirely fromFgfr1�/� cells, which fail to fully traverse the PS and takeon an ectodermal fate (Ciruna et al., 1997; Yoshikawa et al.,1997). The phenotypes of Fgf8 and Fgfr1 mutant micesuggest a role in the migration of nascent mesoderm awayfrom the streak, possibly by repression of E-cadherin(Yamaguchi et al., 1994; Ciruna et al., 1997; Sun et al.,1999; Ciruna and Rossant, 2001). E-cadherin prevents theinteraction of LEF/TCF proteins with the transcriptionalcoactivator �-catenin, disrupting WNT signaling. Seques-tering E-cadherin rescues expression of a T-lacZ reporter,presumably due to the restoration of the WNT signalingpathway (Ciruna and Rossant, 2001). However, expressionof Tbx6 is independent of Wnt3a (Yamaguchi et al., 1999).The potential involvement of FGF signaling in directingcells down a mesodermal pathway rather than a neurecto-dermal pathway has been demonstrated by the ability ofexogenous FGF2 to induce the formation of mesoderm inexplants of anterior epiblasts in culture (Burdsal et al.,1998).
Antagonism of WNT signaling promotes neural differ-entiation in culture. The WNT antagonist Sfrp2 is expressedduring neural differentiation of embryoid bodies. Whentransfected into ES cells, Sfrp2 enhances neuronal differen-tiation (Aubert et al., 2002).
The earliest mesodermal derivatives to differentiate arethe endothelial and hematopoietic cells of the blood islandsin the yolk sac. The extraembryonic mesoderm from whichthe vascular and hematopoietic progenitors arise is amongstthe earliest to pass through the PS (Kinder et al., 1999). Ithas been demonstrated that in vitro, a single population ofcells can act as precursors of both hematopoietic and endo-thelial lineages (Robb and Begley, 1996; Yamashita et al.,2000).
Scl is required for differentiation of hematopoietic lin-eages. Yolk sac hematopoiesis fails in Scl�/� embryos(Shivdasani et al., 1995; Robb et al., 1995), and Scl isnecessary for contribution to any primitive or definitivehematopoietic lineage in the mouse (Porcher et al., 1996).Scl is also expressed in yolk sac endothelium, but is notnecessary for its differentiation.
Scl is necessary for the early stages of in vitro differen-tiation of mesoderm to hematopoietic lineages and actsduring a limited time window (Elefanty et al., 1997; Rob-ertson et al., 2000; Endoh et al., 2002). Loss of Scl affectsgenes specifically expressed during hematopoiesis, but doesnot affect the expression of early mesodermal markers orendothelial markers (Elefanty et al., 1997).
Signaling from the endoderm via vascular endothelialgrowth factor (VEGF) is required for endothelial and he-matopoietic differentiation of mesoderm (Fig. 3, red area;Carmeliet et al., 1996; Shalaby et al., 1997; Fong et al.,1999). Vegf�/� or Vegf�/� embryos die before birth withdisrupted blood vessel formation (Carmeliet et al., 1996;Damert et al., 2002). Targeting of the VEGF receptors Flt1and Flk1 confirms the role of VEGF signaling in the correct
8 D.A.F. Loebel et al. / Developmental Biology 264 (2003) 1–14

differentiation of endothelial lineages (Fong et al., 1995,1999; Shalaby et al., 1995). Flt1 regulates the differentia-tion of mesoderm to hemangioblast/angioblasts (Fong et al.,1999), whereas Flk1 is required cell autonomously for cor-rect maturation of endothelial cells and migration of pre-cursor cells (Shalaby et al., 1995, 1997). A role for VEGFin differentiation of hematopoietic precursors has beenshown by the generation of mice homozygous for a hypo-morphic allele of VEGF (Damert et al., 2002). Flk1 maymediate this function since loss of Flk1 activity also impairshematopoiesis (Shalaby et al., 1995, 1997).
VEGF signaling can also influence the differentiation ofhematopoietic and endothelial lineages in vitro (Fig. 4, redarea). Blast cell colonies, which are derived from embryoidbodies and give rise to hematopoietic cells, are inducedfollowing treatment with VEGF (Kennedy et al., 1997). Inculture, VEGF encourages ES cell-derived Flk1-expressingcells to differentiate into endothelial cell types. Treatmentwith platelet-derived growth factor (PDGF) BB induces thesame cells to differentiate into smooth muscle cells. Fur-thermore, by treating cells with VEGF in collagen gels,three-dimensional vascular structures can be formed (Ya-mashita et al., 2000).
Hedgehog signaling has the ability to promote the earlydifferentiation of endothelial and hematopoietic precursorsin tissue explants (Dyer et al., 2001). However, this does notfully reflect the situation in vivo, as the initial stages ofendothelial and hematopoietic differentiation can occur inthe absence of IHH or its receptor, SMO (Byrd et al., 2002).
SMAD-mediated signal transduction is also implicatedin differentiation of heart tissue from mesodermal precur-sors. Expression of the cardiac-specific transcription factorNkx2-5 requires an enhancer element containing SMADbinding sites (Lien et al., 2002), suggesting that Nkx2-5, likeits Drosophila orthologue tinman, may be regulated byBMP signaling. This agrees with previous findings thattransient exposure of chick explants to BMP2 or �4 incombination with FGF2 or �4 induces contractile cardiacmuscle differentiation and Nkx2-5 expression in nonprecar-diac mesoderm (Ladd et al., 1998; Barron et al., 2000).
SHH and WNT signaling are also important for furtherdifferentiation of paraxial mesoderm derivatives (Fig. 3, redarea). All skeletal muscles in the body of the mouse arederived from the somites, which arise by segmentation ofthe paraxial mesoderm on either side of the neural tube andnotochord. Wnt3a activates myogenic transcription factorsMyoD and Myogenin in embryonal carcinoma cells (Ridge-way et al., 2000). Expression of the early myogenic tran-scription factor Myf5 is reduced in the early dermomyotomeof Wnt1�/�;Wnt3�/� embryos but later increases to normallevels, suggesting that WNT signaling is compensated forby other factors in later development (Ikeya and Takada,1998). Myf5 is regulated by SHH signaling, via GLI tran-scription factor binding to an SHH responsive promoter(Gustafsson et al., 2002). However, there is also evidence
that the primary function of SHH in muscle development isas a survival or proliferation factor (Kruger et al., 2001).
Definitive endoderm differentiation requires FGF, TGF�and WNT signalling
At present, the data relating to the roles of signalingmolecules in definitive endoderm differentiation are frag-mentary. In zebrafish and Xenopus, a pathway of interactingfactors responding to Nodal-related signaling molecules hasbeen established, involving SMAD signal transduction andpaired-like homeodomain transcription factors acting up-stream of SOX transcription factors to induce endoderm(Alexander and Stainier, 1999; Poulain and Lepage, 2002).
There is some evidence that similar pathways may op-erate in the mouse (Fig. 3, orange area). In chimeric mice,Smad2�/� ES cells are not able to contribute to the defin-itive endoderm lineages, indicating that Smad-mediatedTGF� signaling is needed for the initial specification of thedefinitive endoderm lineage (Weinstein et al., 1998;Waldrip et al., 1998; Tremblay et al., 2000). The phenotypesof Sox17�/� and Mixl1�/� mice suggest that both transcrip-tion factors are required for the differentiation but not forformation of the definitive endoderm (Hart et al., 2002;Kanai-Azuma et al., 2002). Mixl1 is apparently essential forthe proper allocation of endoderm precursors from mesen-doderm progenitors during gastrulation. Loss of Sox17 re-sults in a severe reduction in the amount of definitiveendoderm. Sox17-null definitive endoderm has reduced vi-ability and proliferation, resulting in occupation of the gutby visceral endoderm-like cells. Testing the tissue potencyof the null-mutant ES cells in chimeric embryos revealsmore dramatic effects of the loss-of-function of these fac-tors. Sox17�/� and Mixl1�/� ES cells fail to contribute tothe gut endoderm in competition with the wild type embry-onic cells (Hart et al., 2002; Kanai-Azuma et al., 2002).
Germ layer explant experiments show that differentiationof definitive endoderm requires soluble factors originatingfrom the mesoderm and ectoderm layers during gastrulation(Fig. 3, orange area). FGF4 alone is sufficient initiate dif-ferentiation, but additional factors provided by interactingtissues are necessary for survival of the induced endoderm(Wells and Melton, 2000). FGF signaling is also involvedlater during organogenesis, where exposure of ventral fo-regut endoderm to FGF from the cardiac mesoderm inhibitsventral pancreas, the default fate, to liver formation (Deut-sch et al., 2001). Embryos lacking �-catenin in the anteriorPS, node, and notochord are depleted of definitiveendoderm, although the visceral endoderm is displaced nor-mally. In chimeric embryos, precardiac mesoderm popu-lates the endodermal layer, and ectopic hearts form in�-catenin-null embryos. This suggests the downregulationof �-catenin is required to direct cells in the PS to choose anendodermal rather than mesodermal fate (Lickert et al.,2002) and implicates the WNT-responsive intracellularpathway in the formation of the definitive endoderm. How-
9D.A.F. Loebel et al. / Developmental Biology 264 (2003) 1–14

ever, the signaling molecule is as yet unidentified, and thereis no direct evidence for the involvement of WNT.
Manipulation of gene expression in ES cells has revealedroles for several other transcription factors in endodermdifferentiation. Tcf2�/� cells are severely impaired in theirability to contribute to the gut in chimeric mice, indicatingan additional requirement for Tcf2 in definitive endodermformation (Barbacci et al., 1999; Coffinier et al., 1999).Overexpression of Foxa2 in ES cells induces genes nor-mally expressed in endoderm-derived tissues such as theliver, gut, and lung. However, genes involved in lateendoderm differentiation are not induced (Levinson-Dush-nik and Benvenisty, 1997).
As yet there are no reports of directed in vitro differen-tiation of ES cells to early gut endoderm by manipulation ofculture conditions with conditioned media or soluble fac-tors. However, the production of hepatic progenitors andpancreatic islet cells, which are both later endodermal de-rivatives, by directed in vitro differentiation of ES cells hasbeen reported (Fig. 4, orange area; Lumelsky et al., 2001;Hori et al., 2002; Yin et al., 2002).
Potential for therapeutic applications
For in vitro differentiated ES cells to be useful in humanstem cell therapies, the cells must be capable of contributingto target tissues of a recipient, without the presence ofundifferentiated cells precipitating the formation of terato-mas. This requires homogeneous populations of differenti-ated cells. In the embryo, the decision of a pluripotent cellto progress toward a particular lineage requires the con-certed efforts of inductive or inhibitory signals from tissuesin the immediate vicinity and the ability of the cell tointerpret these signals. Replication of these conditions invitro can be achieved by controlling the delivery of similarcombinations of growth factors to the ES cells.
To date it has not been possible to generate truly homo-geneous populations of differentiated cells purely by themanipulation of culture conditions. In the majority of cases,the generation of nearly homogeneous population of cellshas been accomplished by the selection of the desired celltype on the basis of expression of lineage-specific markersor transgenes, or ectopic expression of lineage-specific tran-scription factors. However, enriching specific cell types bythese means is less attractive for therapeutic applicationsbecause it involves genetic modifications, which may havean undesirable and unpredictable outcome and which mayimpair the ability of cells to undergo the full repertoire oflineage differentiation.
Despite difficulties in obtaining homogeneous popula-tions, functional derivatives of all three primary germ layershave been successfully produced from ES cells. Differenti-ated dopamine-producing neurons were found to incorpo-rate into the striatum and improve performance in behav-ioral tests when grafted into mice that phenocopy
Parkinson’s disease (Kim et al., 2002; Nishimura et al.,2003). ES cell-derived motor neurons have been grafted intochick embryo spinal cords and are shown to survive anddifferentiate correctly in vivo (Wichterle et al., 2002). Flk1-expressing, ES cell-derived vascular progenitor cells wereable to differentiate into endothelial cells, after grafting intotumor-bearing mice. The grafted cells were incorporatedinto the developing vasculature of the tumor, demonstratingan ability to contribute to adult neovascularization (Yurugi-Kobayashi et al., 2003). Pancreatic cells, produced by cul-ture in selective media and supplemented with FGF2,formed islet-like structures and, upon transplantation intodiabetic mice, improved survival and body weight mainte-nance (Lumelsky et al., 2001). In vitro derived hepatocyteprogenitors were found to undergo proper differentiationinto parenchymal cells in the liver of recipient animals(Ishizaka et al., 2002; Yin et al., 2002).
The encouraging outcome of the above experiments in-dicates that directed differentiation of ES cells is possibleand may be translated into therapeutic applications. How-ever, the need to rigorously validate the fidelity of celldifferentiation by marker analysis has been highlighted bycontroversy over the ability of in vitro derived cells thatresemble pancreatic islets to synthesize appreciable quanti-ties of insulin (Rajagopal et al., 2003). In addition, the needto eliminate undesirable cell types is illustrated by the ob-servation of tumor formation in recipient animals, probablydue to the presence of undifferentiated stem cells in thetransplanted material, despite barely detectable levels ofPou5f1 expression (Nishimura et al., 2003).
Therefore, a more thorough understanding of the pro-cesses that drive tissue differentiation in the embryo isessential for improving the current techniques for directedES cell differentiation. Better control of in vitro cell differ-entiation may therefore necessitate the replication of the invivo situation that favors specific lineage development. Inparticular, it is necessary to reconstitute the interactionsamong cells within a community and between cell popula-tions in vivo. Further complications are the multiple effectsof single factors and the synergistic and antagonistic effectsof combinations of factors. Understanding the complex tis-sue interactions and interdependence of molecular mecha-nisms of lineage choices in vivo presents a significant chal-lenge, but the outcome of embryological investigations willbe crucial to the realization of stem cell-based therapies.
Acknowledgments
We thank Melinda Hayward for providing the ES cellsand Janet Rossant for the picture of TS cells (Fig. 1D), andBruce Davidson, Leon McQuade, and Peter Rowe for com-ments on the manuscript. Our work is supported by theNational Health and Medical Research Council (NHMRC)of Australia and Mr James Fairfax. P.P.L.T. is a NHMRCSenior Principal Research Fellow.
10 D.A.F. Loebel et al. / Developmental Biology 264 (2003) 1–14

Note added in proof. Two recent papers have describedtranscription factor, Nanog, that can maintain ES cells in anundifferentiated state independent of the LIF/STAT3 path-way (Chambers et al. (2003) Cell 113, 643–55; Mitsui et al.(2003) Cell 113, 631–42).
References
Ahlgren, U., Jonsson, J., Edlund, H., 1996. The morphogenesis of thepancreatic mesenchyme is uncoupled from that of the pancreatic epi-thelium in IPF1/PDX1-deficient mice. Development 122, 1409–1416.
Ahlgren, U., Pfaff, S.L., Jessell, T.M., Edlund, T., Edlund, H., 1997.Independent requirement for ISL1 in formation of pancreatic mesen-chyme and islet cells. Nature 385, 257–260.
Alexander, J., Stainier, D.Y., 1999. A molecular pathway leading toendoderm formation in zebrafish. Curr. Biol. 9, 1147–1157.
Ambrosetti, D.C., Basilico, C., Dailey, L., 1997. Synergistic activation ofthe fibroblast growth factor 4 enhancer by Sox2 and Oct-3 depends onprotein-protein interactions facilitated by a specific spatial arrangementof factor binding sites. Mol. Cell. Biol. 17, 6321–6329.
Ang, S.L., Conlon, R.A., Jin, O., Rossant, J., 1994. Positive and negativesignals from mesoderm regulate the expression of mouse Otx2 inectoderm explants. Development 120, 2979–2989.
Ang, S.L., Rossant, J., 1993. Anterior mesendoderm induces mouse En-grailed genes in explant cultures. Development 118, 139–149.
Aubert, J., Dunstan, H., Chambers, I., Smith, A., 2002. Functional genescreening in embryonic stem cells implicates Wnt antagonism in neuraldifferentiation. Nat. Biotechnol. 20, 1240–1245.
Avilion, A.A., Nicolis, S.K., Pevny, L.H., Perez, L., Vivian, N., Lovell-Badge, R., 2003. Multipotent cell lineages in early mouse developmentdepend on SOX2 function. Genes Dev. 17, 126–140.
Bachiller, D., Klingensmith, J., Kemp, C., Belo, J.A., Anderson, R.M.,May, S.R., McMahon, J.A., McMahon, A.P., Harland, R.M., Rossant,J., De Robertis, E.M., 2000. The organizer factors Chordin and Nogginare required for mouse forebrain development. Nature 403, 658–661.
Barbacci, E., Reber, M., Ott, M.O., Breillat, C., Huetz, F., Cereghini, S.,1999. Variant hepatocyte nuclear factor 1 is required for visceralendoderm specification. Development 126, 4795–4805.
Barron, M., Gao, M., Lough, J., 2000. Requirement for BMP and FGFsignaling during cardiogenic induction in non-precardiac mesoderm isspecific, transient, and cooperative. Dev. Dyn. 218, 383–393.
Burdsal, C.A., Flannery, M.L., Pedersen, R.A., 1998. FGF-2 alters the fateof mouse epiblast from ectoderm to mesoderm in vitro. Dev. Biol. 198,231–244.
Byrd, N., Becker, S., Maye, P., Narasimhaiah, R., St Jacques, B., Zhang,X., McMahon, J., McMahon, A., Grabel, L., 2002. Hedgehog is re-quired for murine yolk sac angiogenesis. Development 129, 361–372.
Carmeliet, P., Ferreira, V., Breier, G., Pollefeyt, S., Kieckens, L., Gertsen-stein, M., Fahrig, M., Vandenhoeck, A., Harpal, K., Eberhardt, C.,Declereq, C., Pawling, J., Moons, L., Collen, D., Risau, W., Nagy, A.,1996. Abnormal blood vessel development and lethality in embryoslacking a single VEGF allele. Nature 380, 435–439.
Chang, C., Holtzman, D.A., Chau, S., Chickering, T., Woolf, E.A.,Holmgren, L.M., Bodorova, J., Gearing, D.P., Holmes, W.E., Brivan-lou, A.H., 2001. Twisted gastrulation can function as a BMP antago-nist. Nature 410, 483–487.
Chang, H., Huylebroeck, D., Verschueren, K., Guo, Q., Matzuk, M.M.,Zwijsen, A., 1999. Smad5 knockout mice die at mid-gestation due tomultiple embryonic and extraembryonic defects. Development 126,1631–1642.
Chang, H., Matzuk, M.M., 2001. Smad5 is required for mouse primordialgerm cell development. Mech. Dev. 104, 61–67.
Chapman, D.L., Papaioannou, V.E., 1998. Three neural tubes in mouseembryos with mutations in the T-box gene Tbx6. Nature 391, 695–697.
Ciruna, B., Rossant, J., 2001. FGF signaling regulates mesoderm cell fatespecification and morphogenetic movement at the primitive streak.Dev. Cell 1, 37–49.
Ciruna, B.G., Schwartz, L., Harpal, K., Yamaguchi, T.P., Rossant, J., 1997.Chimeric analysis of fibroblast growth factor receptor-1 (Fgfr1) func-tion: a role for FGFR1 in morphogenetic movement through the prim-itive streak. Development 124, 2829–2841.
Coffinier, C., Thepot, D., Babinet, C., Yaniv, M., Barra, J., 1999. Essentialrole for the homeoprotein vHNF1/HNF1 beta in visceral endodermdifferentiation. Development 126, 4785–4794.
Damert, A., Miquerol, L., Gertsenstein, M., Risau, W., Nagy, A., 2002.Insufficient VEGFA activity in yolk sac endoderm compromiseshaematopoietic and endothelial differentiation. Development 129,1881–1892.
Deutsch, G., Jung, J., Zheng, M., Lora, J., Zaret, K.S., 2001. A bipotentialprecursor population for pancreas and liver within the embryonicendoderm. Development 128, 871–881.
Dyer, M.A., Farrington, S.M., Mohn, D., Munday, J.R., Baron, M.H.,2001. Indian hedgehog activates hematopoiesis and vasculogenesis andcan respecify prospective neurectodermal cell fate in the mouse em-bryo. Development 128, 1717–1730.
Elefanty, A.G., Robb, L., Birner, R., Begley, C.G., 1997. Hematopoietic-specific genes are not induced during in vitro differentiation of scl-nullembryonic stem cells. Blood 90, 1435–1447.
Endoh, M., Ogawa, M., Orkin, S., Nishikawa, S., 2002. SCL/tal-1-depen-dent process determines a competence to select the definitive hemato-poietic lineage prior to endothelial differentiation. EMBO J. 21, 6700–6708.
Finley, M.F., Devata, S., Huettner, J.E., 1999. BMP-4 inhibits neuraldifferentiation of murine embryonic stem cells. J. Neurobiol. 40, 271–287.
Fong, G.H., Rossant, J., Gertsenstein, M., Breitman, M.L., 1995. Role ofthe Flt-1 receptor tyrosine kinase in regulating the assembly of vascularendothelium. Nature 376, 66–70.
Fong, G.H., Zhang, L., Bryce, D.M., Peng, J., 1999. Increased hemangio-blast commitment, not vascular disorganization, is the primary defect inflt-1 knock-out mice. Development 126, 3015–3025.
Fuhrmann, G., Chung, A.C., Jackson, K.J., Hummelke, G., Baniahmad, A.,Sutter, J., Sylvester, I., Scholer, H.R., Cooney, A.J., 2001. Mousegermline restriction of Oct4 expression by germ cell nuclear factor.Dev. Cell 1, 377–387.
Fujiwara, T., Dunn, N.R., Hogan, B.L., 2001. Bone morphogenetic protein4 in the extraembryonic mesoderm is required for allantois develop-ment and the localization and survival of primordial germ cells in themouse. Proc. Natl. Acad. Sci. USA 98, 13739–13744.
Galceran, J., Farinas, I., Depew, M.J., Clevers, H., Grosschedl, R., 1999.Wnt3a�/�-like phenotype and limb deficiency in Lef1(�/�)Tcf1(�/�) mice. Genes Dev. 13, 709–717.
Galceran, J., Hsu, S.C., Grosschedl, R., 2001. Rescue of a Wnt mutation byan activated form of LEF-1: regulation of maintenance but not initia-tion of Brachyury expression. Proc. Natl. Acad. Sci. USA 98, 8668–8673.
Gardner, R.L., Beddington, R.S., 1988. Multi-lineage “stem” cells in themammalian embryo. J. Cell Sci. Suppl. 10, 11–27.
Gratsch, T.E., O’Shea, K.S., 2002. Noggin and chordin have distinctactivities in promoting lineage commitment of mouse embryonic stem(ES) cells. Dev. Biol. 245, 83–94.
Gustafsson, M.K., Pan, H., Pinney, D.F., Liu, Y., Lewandowski, A., Ep-stein, D.J., Emerson Jr., C.P., 2002. Myf5 is a direct target of long-range Shh signaling and Gli regulation for muscle specification. GenesDev. 16, 114–126.
Harrison, K.A., Thaler, J., Pfaff, S.L., Gu, H., Kehrl, J.H., 1999. Pancreasdorsal lobe agenesis and abnormal islets of Langerhans in Hlxb9-deficient mice. Nat. Genet. 23, 71–75.
Harrison, S.M., Dunwoodie, S.L., Arkell, R.M., Lehrach, H., Beddington,R.S., 1995. Isolation of novel tissue-specific genes from cDNA librar-
11D.A.F. Loebel et al. / Developmental Biology 264 (2003) 1–14

ies representing the individual tissue constituents of the gastrulatingmouse embryo. Development 121, 2479–2489.
Hart, A.H., Hartley, L., Sourris, K., Stadler, E.S., Li, R., Stanley, E.G.,Tam, P.P., Elefanty, A.G., Robb, L., 2002. Mixl1 is required for axialmesendoderm morphogenesis and patterning in the murine embryo.Development 129, 3597–3608.
Hayashi, H., Abdollah, S., Qiu, Y., Cai, J., Xu, Y.Y., Grinnell, B.W.,Richardson, M.A., Topper, J.N., Gimbrone Jr., M.A., Wrana, J.L., Falb,D., 1997. The MAD-related protein Smad7 associates with the TGF-beta receptor and functions as an antagonist of TGFbeta signaling. Cell89, 1165–1173.
Hayashi, K., Kobayashi, T., Umino, T., Goitsuka, R., Matsui, Y., Kita-mura, D., 2002. SMAD1 signaling is critical for initial commitment ofgerm cell lineage from mouse epiblast. Mech. Dev. 118, 99.
Hori, Y., Rulifson, I.C., Tsai, B.C., Heit, J.J., Cahoy, J.D., Kim, S.K., 2002.Growth inhibitors promote differentiation of insulin-producing tissuefrom embryonic stem cells. Proc. Natl. Acad. Sci. USA 99, 16105–16110.
Ikeya, M., Takada, S., 1998. Wnt signaling from the dorsal neural tube isrequired for the formation of the medial dermomyotome. Development125, 4969–4976.
Ishizaka, S., Shiroi, A., Kanda, S., Yoshikawa, M., Tsujinoue, H.,Kuriyama, S., Hasuma, T., Nakatani, K., Takahashi, K., 2002. Devel-opment of hepatocytes from ES cells after transfection with the HNF-3beta gene. FASEB J. 16, 1444–1446.
Johansson, B.M., Wiles, M.V., 1995. Evidence for involvement of activinA and bone morphogenetic protein 4 in mammalian mesoderm andhematopoietic development. Mol. Cell. Biol. 15, 141–151.
Kanai-Azuma, M., Kanai, Y., Gad, J.M., Tajima, Y., Taya, C., Kurohmaru,M., Sanai, Y., Yonekawa, H., Yazaki, K., Tam, P.P., Hayashi, Y., 2002.Depletion of definitive gut endoderm in Sox17-null mutant mice. De-velopment 129, 2367–2379.
Kawasaki, H., Mizuseki, K., Nishikawa, S., Kaneko, S., Kuwana, Y.,Nakanishi, S., Nishikawa, S.I., Sasai, Y., 2000. Induction of midbraindopaminergic neurons from ES cells by stromal cell-derived inducingactivity. Neuron 28, 31–40.
Kennedy, M., Firpo, M., Choi, K., Wall, C., Robertson, S., Kabrun, N.,Keller, G., 1997. A common precursor for primitive erythropoiesis anddefinitive haematopoiesis. Nature 386, 488–493.
Kim, J.H., Auerbach, J.M., Rodriguez-Gomez, J.A., Velasco, I., Gavin, D.,Lumelsky, N., Lee, S.H., Nguyen, J., Sanchez-Pernaute, R., Bank-iewicz, K., McKay, R., 2002. Dopamine neurons derived from embry-onic stem cells function in an animal model of Parkinson’s disease.Nature 418, 50–56.
Kinder, S.J., Tsang, T.E., Quinlan, G.A., Hadjantonakis, A.K., Nagy, A.,Tam, P.P., 1999. The orderly allocation of mesodermal cells to theextraembryonic structures and the anteroposterior axis during gastru-lation of the mouse embryo. Development 126, 4691–4701.
Kruger, M., Mennerich, D., Fees, S., Schafer, R., Mundlos, S., Braun, T.,2001. Sonic hedgehog is a survival factor for hypaxial muscles duringmouse development. Development 128, 743–752.
Labosky, P.A., Barlow, D.P., Hogan, B.L., 1994. Mouse embryonic germ(EG) cell lines: transmission through the germline and differences inthe methylation imprint of insulin-like growth factor 2 receptor (Igf2r)gene compared with embryonic stem (ES) cell lines. Development 120,3197–3204.
Ladd, A.N., Yatskievych, T.A., Antin, P.B., 1998. Regulation of aviancardiac myogenesis by activin/TGFbeta and bone morphogenetic pro-teins. Dev. Biol. 204, 407–419.
Lake, J., Rathjen, J., Remiszewski, J., Rathjen, P.D., 2000. Reversibleprogramming of pluripotent cell differentiation. J. Cell Sci. 113, 555–566.
Lange, U., Saitou, M., Western, P., Barton, S., Surani, M., 2003. TheFragilis interferon-inducible gene family of transmembrane proteins isassociated with germ cell specification in mice. BMC Dev. Biol. 3, 1.
Lawson, K.A., Dunn, N.R., Roelen, B.A., Zeinstra, L.M., Davis, A.M.,Wright, C.V., Korving, J.P., Hogan, B.L., 1999. Bmp4 is required for
the generation of primordial germ cells in the mouse embryo. GenesDev. 13, 424–436.
Lawson, K.A., Meneses, J.J., Pedersen, R.A., 1986. Cell fate and celllineage in the endoderm of the presomite mouse embryo, studied withan intracellular tracer. Dev. Biol. 115, 325–339.
Levinson-Dushnik, M., Benvenisty, N., 1997. Involvement of hepatocytenuclear factor 3 in endoderm differentiation of embryonic stem cells.Mol. Cell. Biol. 17, 3817–3822.
Lickert, H., Kutsch, S., Kanzler, B., Tamai, Y., Taketo, M.M., Kemler, R.,2002. Formation of multiple hearts in mice following deletion of�-catenin in the embryonic endoderm. Dev. Cell 3, 171–181.
Lien, C.L., McAnally, J., Richardson, J.A., Olson, E.N., 2002. Cardiac-specific activity of an Nkx2-5 enhancer requires an evolutionarilyconserved Smad binding site. Dev. Biol. 244, 257–266.
Lumelsky, N., Blondel, O., Laeng, P., Velasco, I., Ravin, R., McKay, R.,2001. Differentiation of embryonic stem cells to insulin-secreting struc-tures similar to pancreatic islets. Science 292, 1389–1394.
McLaren, A., Durcova-Hills, G., 2001. Germ cells and pluripotent stemcells in the mouse. Reprod Fertil. Dev. 13, 661–664.
Mishina, Y., Suzuki, A., Gilbert, D.J., Copeland, N.G., Jenkins, N.A.,Ueno, N., Behringer, R.R., 1995. Genomic organization and chromo-somal location of the mouse type I BMP-2/4 receptor. Biochem. Bio-phys. Res. Commun. 206, 310–317.
Miyazawa, K., Shinozaki, M., Hara, T., Furuya, T., Miyazono, K., 2002.Two major Smad pathways in TGF-beta superfamily signalling. GenesCells 7, 1191–1204.
Mukhopadhyay, M., Shtrom, S., Rodriguez-Esteban, C., Chen, L., Tsukui,T., Gomer, L., Dorward, D.W., Glinka, A., Grinberg, A., Huang, S.P.,Niehrs, C., Izpisua Belmonte, J.C., Westphal, H., 2001. Dickkopf1 isrequired for embryonic head induction and limb morphogenesis in themouse. Dev. Cell 1, 423–434.
Mukhopadhyay, M., Teufel, A., Yamashita, T., Agulnick, A.D., Chen, L.,Downs, K.M., Schindler, A., Grinberg, A., Huang, S.P., Dorward, D.,Westphal, H., 2003. Functional ablation of the mouse Ldb1 gene resultsin severe patterning defects during gastrulation. Development 130,495–505.
Nagy, A., Rossant, J., Nagy, R., Abramow-Newerly, W., Roder, J.C., 1993.Derivation of completely cell culture-derived mice from early-passageembryonic stem cells. Proc. Natl. Acad. Sci. USA 90, 8424–8428.
Nichols, J., Zevnik, B., Anastassiadis, K., Niwa, H., Klewe-Nebenius, D.,Chambers, I., Scholer, H., Smith, A., 1998. Formation of pluripotentstem cells in the mammalian embryo depends on the POU transcriptionfactor Oct4. Cell 95, 379–391.
Nishimura, F., Yoshikawa, M., Kanda, S., Nonaka, M., Yokota, H., Shiroi,A., Nakase, H., Hirabayashi, H., Ouji, Y., Birumachi, J., Ishizaka, S.,Sakaki, T., 2003. Potential use of embryonic stem cells for the treat-ment of mouse parkinsonian models: improved behavior by transplan-tation of in vitro differentiated dopaminergic neurons from embryonicstem cells. Stem Cells 21, 171–180.
Niwa, H., 2001. Molecular mechanism to maintain stem cell renewal of EScells. Cell Struct. Funct. 26, 137–148.
Niwa, H., Burdon, T., Chambers, I., Smith, A., 1998. Self-renewal ofpluripotent embryonic stem cells is mediated via activation of STAT3.Genes Dev. 12, 2048–2060.
Niwa, H., Miyazaki, J., Smith, A.G., 2000. Quantitative expression ofOct-3/4 defines differentiation, dedifferentiation or self-renewal of EScells. Nat. Genet. 24, 372–376.
Pelton, T.A., Sharma, S., Schulz, T.C., Rathjen, J., Rathjen, P.D., 2002.Transient pluripotent cell populations during primitive ectoderm for-mation: correlation of in vivo and in vitro pluripotent cell development.J. Cell Sci. 115, 329–339.
Persson, M., Stamataki, D., te, W.P., Andersson, E., Bose, J., Ruther, U.,Ericson, J., Briscoe, J., 2002. Dorsal–ventral patterning of the spinalcord requires Gli3 transcriptional repressor activity. Genes Dev. 16,2865–2878.
Pesce, M., Gioia, K.F., De Felici, M., 2002. Derivation in culture ofprimordial germ cells from cells of the mouse epiblast: phenotypic
12 D.A.F. Loebel et al. / Developmental Biology 264 (2003) 1–14

induction and growth control by Bmp4 signalling. Mech. Dev. 112,15–24.
Pevny, L.H., Sockanathan, S., Placzek, M., Lovell-Badge, R., 1998. A rolefor SOX1 in neural determination. Development 125, 1967–1978.
Porcher, C., Swat, W., Rockwell, K., Fujiwara, Y., Alt, F.W., Orkin, S.H.,1996. The T cell leukemia oncoprotein SCL/tal-1 is essential for de-velopment of all hematopoietic lineages. Cell 86, 47–57.
Poulain, M., Lepage, T., 2002. Mezzo, a paired-like homeobox protein isan immediate target of Nodal signalling and regulates endoderm spec-ification in zebrafish. Development 129, 4901–4914.
Rajagopal, J., Anderson, W.J., Kume, S., Martinez, O.I., Melton, D.A.,2003. Insulin staining of ES cell progeny from insulin uptake. Science299, 363.
Rathjen, J., Haines, B.P., Hudson, K.M., Nesci, A., Dunn, S., Rathjen,P.D., 2002. Directed differentiation of pluripotent cells to neural lin-eages: homogeneous formation and differentiation of a neurectodermpopulation. Development 129, 2649–2661.
Rathjen, J., Lake, J.A., Bettess, M.D., Washington, J.M., Chapman, G.,Rathjen, P.D., 1999. Formation of a primitive ectoderm like cell pop-ulation, EPL cells, from ES cells in response to biologically derivedfactors. J. Cell Sci. 112, 601–612.
Raz, R., Lee, C.K., Cannizzaro, L.A., d’Eustachio, P., Levy, D.E., 1999.Essential role of STAT3 for embryonic stem cell pluripotency. Proc.Natl. Acad. Sci. USA 96, 2846–2851.
Ridgeway, A.G., Petropoulos, H., Wilton, S., Skerjanc, I.S., 2000. Wntsignaling regulates the function of MyoD and myogenin. J. Biol. Chem.275, 32398–32405.
Robb, L., Begley, C.G., 1996. The helix–loop–helix gene SCL: implicatedin T-cell acute lymphoblastic leukaemia and in normal haematopoieticdevelopment. Int. J. Biochem. Cell Biol. 28, 609–618.
Robb, L., Lyons, I., Li, R., Hartley, L., Kontgen, F., Harvey, R.P., Metcalf,D., Begley, C.G., 1995. Absence of yolk sac hematopoiesis from micewith a targeted disruption of the scl gene. Proc. Natl. Acad. Sci. USA92, 7075–7079.
Robertson, S.M., Kennedy, M., Shannon, J.M., Keller, G., 2000. A tran-sitional stage in the commitment of mesoderm to hematopoiesis requir-ing the transcription factor SCL/tal-1. Development 127, 2447–2459.
Rodda, S.J., Kavanagh, S.J., Rathjen, J., Rathjen, P.D., 2002. Embryonicstem cell differentiation and the analysis of mammalian development.Int. J. Dev. Biol. 46, 449–458.
Rossant, J., McKerlie, C., 2001. Mouse-based phenogenomics for model-ling human disease. Trends Mol. Med. 7, 502–507.
Saitou, M., Barton, S.C., Surani, M.A., 2002. A molecular programme forthe specification of germ cell fate in mice. Nature 418, 293–300.
Sato, M., Kimura, T., Kurokawa, K., Fujita, Y., Abe, K., Masuhara, M.,Yasunaga, T., Ryo, A., Yamamoto, M., Nakano, T., 2002. Identifica-tion of PGC7, a new gene expressed specifically in preimplantationembryos and germ cells. Mech. Dev. 113, 91–94.
Shalaby, F., Ho, J., Stanford, W.L., Fischer, K.D., Schuh, A.C., Schwartz,L., Bernstein, A., Rossant, J., 1997. A requirement for Flk1 in primitiveand definitive hematopoiesis and vasculogenesis. Cell 89, 981–990.
Shalaby, F., Rossant, J., Yamaguchi, T.P., Gertsenstein, M., Wu, X.F.,Breitman, M.L., Schuh, A.C., 1995. Failure of blood-island formationand vasculogenesis in Flk-1-deficient mice. Nature 376, 62–66.
Shivdasani, R.A., Mayer, E.L., Orkin, S.H., 1995. Absence of blood for-mation in mice lacking the T-cell leukaemia oncoprotein tal-1/SCL.Nature 373, 432–434.
Sirard, C., de la Pompa, J.L., Elia, A., Itie, A., Mirtsos, C., Cheung, A.,Hahn, S., Wakeham, A., Schwartz, L., Kern, S.E., Rossant, J., Mak,T.W., 1998. The tumor suppressor gene Smad4/Dpc4 is required forgastrulation and later for anterior development of the mouse embryo.Genes Dev. 12, 107–119.
Sun, X., Meyers, E.N., Lewandoski, M., Martin, G.R., 1999. Targeteddisruption of Fgf8 causes failure of cell migration in the gastrulatingmouse embryo. Genes Dev. 13, 1834–1846.
Takeda, K., Noguchi, K., Shi, W., Tanaka, T., Matsumoto, M., Yoshida,N., Kishimoto, T., Akira, S., 1997. Targeted disruption of the mouse
Stat3 gene leads to early embryonic lethality. Proc. Natl. Acad. Sci.USA 94, 3801–3804.
Tam, P.P., Behringer, R.R., 1997. Mouse gastrulation: the formation of amammalian body plan. Mech. Dev. 68, 3–25.
Tam, P.P., Goldman, D., Camus, A., Schoenwolf, G.C., 2000. Early eventsof somitogenesis in higher vertebrates: allocation of precursor cellsduring gastrulation and the organization of a meristic pattern in theparaxial mesoderm. Curr. Top. Dev. Biol. 47, 1–32.
Tam, P.P., Snow, M.H., 1981. Proliferation and migration of primordialgerm cells during compensatory growth in mouse embryos. J. Embryol.Exp. Morphol. 64, 133–147.
Tam, P.P., Zhou, S.X., 1996. The allocation of epiblast cells to ectodermaland germ-line lineages is influenced by the position of the cells in thegastrulating mouse embryo. Dev. Biol. 178, 124–132.
Tanaka, M., Kasahara, H., Bartunkova, S., Schinke, M., Komuro, I., Ina-gaki, H., Lee, Y., Lyons, G.E., Izumo, S., 1998. Vertebrate homologsof tinman and bagpipe: roles of the homeobox genes in cardiovasculardevelopment. Dev. Genet. 22, 239–249.
Tanaka, S.S., Matsui, Y., 2002. Developmentally regulated expression ofmil-1 and mil-2, mouse interferon-induced transmembrane protein likegenes, during formation and differentiation of primordial germ cells.Gene Expr. Patterns 2, 297–303.
Tremblay, K.D., Dunn, N.R., Robertson, E.J., 2001. Mouse embryos lack-ing Smad1 signals display defects in extra-embryonic tissues and germcell formation. Development 128, 3609–3621.
Tremblay, K.D., Hoodless, P.A., Bikoff, E.K., Robertson, E.J., 2000.Formation of the definitive endoderm in mouse is a Smad2-dependentprocess. Development 127, 3079–3090.
Tremblay, K.D., Saam, J.R., Ingram, R.S., Tilghman, S.M., Bartolomei,M.S., 1995. A paternal-specific methylation imprint marks the allelesof the mouse H19 gene. Nat. Genet. 9, 407–413.
Tropepe, V., Hitoshi, S., Sirard, C., Mak, T.W., Rossant, J., van Der, K.D.,2001. Direct neural fate specification from embryonic stem cells: aprimitive mammalian neural stem cell stage acquired through a defaultmechanism. Neuron 30, 65–78.
Tsang, T.E., Khoo, P.L., Jamieson, R.V., Zhou, S.X., Ang, S.L., Behringer,R., Tam, P.P., 2001. The allocation and differentiation of mouse pri-mordial germ cells. Int. J. Dev. Biol. 45, 549–555.
Tsang, T.E., Kinder, S.J., Tam, P.P., 1999. Experimental analysis of theemergence of left–right asymmetry of the body axis in early postim-plantation mouse embryos. Cell Mol. Biol. 45, 493–503.
Waldrip, W.R., Bikoff, E.K., Hoodless, P.A., Wrana, J.L., Robertson, E.J.,1998. Smad2 signaling in extraembryonic tissues determines anterior–posterior polarity of the early mouse embryo. Cell 92, 797–808.
Weinstein, M., Yang, X., Li, C., Xu, X., Gotay, J., Deng, C.X., 1998.Failure of egg cylinder elongation and mesoderm induction in mouseembryos lacking the tumor suppressor smad2. Proc. Natl. Acad. Sci.USA 95, 9378–9383.
Wells, J.M., Melton, D.A., 2000. Early mouse endoderm is patterned bysoluble factors from adjacent germ layers. Development 127, 1563–1572.
Wichterle, H., Lieberam, I., Porter, J.A., Jessell, T.M., 2002. Directeddifferentiation of embryonic stem cells into motor neurons. Cell 110,385–397.
Wiles, M.V., Johansson, B.M., 1997. Analysis of factors controlling pri-mary germ layer formation and early hematopoiesis using embryonicstem cell in vitro differentiation. Leukemia 11 (Suppl. 3), 454–456.
Wiles, M.V., Johansson, B.M., 1999. Embryonic stem cell development ina chemically defined medium. Exp. Cell Res. 247, 241–248.
Wilson, V., Rashbass, P., Beddington, R.S., 1993. Chimeric analysis of T(Brachyury) gene function. Development 117, 1321–1331.
Winnier, G., Blessing, M., Labosky, P.A., Hogan, B.L., 1995. Bone mor-phogenetic protein-4 is required for mesoderm formation and pattern-ing in the mouse. Genes Dev. 9, 2105–2116.
Wood, H.B., Episkopou, V., 1999. Comparative expression of the mouseSox1, Sox2 and Sox3 genes from pre-gastrulation to early somitestages. Mech. Dev. 86, 197–201.
13D.A.F. Loebel et al. / Developmental Biology 264 (2003) 1–14

Yamaguchi, T.P., Bradley, A., McMahon, A.P., Jones, S., 1999. A Wnt5apathway underlies outgrowth of multiple structures in the vertebrateembryo. Development 126, 1211–1223.
Yamaguchi, T.P., Harpal, K., Henkemeyer, M., Rossant, J., 1994. fgfr-1 isrequired for embryonic growth and mesodermal patterning duringmouse gastrulation. Genes Dev. 8, 3032–3044.
Yamashita, H., Ten Dijke, P., Franzen, P., Miyazono, K., Heldin, C.H.,1994. Formation of hetero-oligomeric complexes of type I and type IIreceptors for transforming growth factor-beta. J. Biol. Chem. 269,20172–20178.
Yamashita, J., Itoh, H., Hirashima, M., Ogawa, M., Nishikawa, S., Yurugi,T., Naito, M., Nakao, K., Nishikawa, S., 2000. Flk1-positive cellsderived from embryonic stem cells serve as vascular progenitors. Na-ture 408, 92–96.
Yan, J., Tanaka, S., Oda, M., Makino, T., Ohgane, J., Shiota, K., 2001.Retinoic acid promotes differentiation of trophoblast stem cells to agiant cell fate. Dev. Biol. 235, 422–432.
Yang, Y., Guillot, P., Boyd, Y., Lyon, M.F., McMahon, A.P., 1998.Evidence that preaxial polydactyly in the Doublefoot mutant is dueto ectopic Indian Hedgehog signaling. Development 125, 3123–3132.
Yin, Y., Lim, Y.K., Salto-Tellez, M., Ng, S.C., Lin, C.S., Lim, S.K., 2002.AFP(�), ESC-derived cells engraft and differentiate into hepatocytesin vivo. Stem Cells 20, 338–346.
Ying, Q.L., Stavridis, M., Griffiths, D., Li, M., Smith, A., 2003. Conver-sion of embryonic stem cells into neuroectodermal precursors in ad-herent monoculture. Nat. Biotechnol. 21, 183–186.
Ying, Y., Liu, X.M., Marble, A., Lawson, K.A., Zhao, G.Q., 2000. Re-quirement of Bmp8b for the generation of primordial germ cells in themouse. Mol. Endocrinol. 14, 1053–1063.
Ying, Y., Qi, X., Zhao, G.Q., 2001. Induction of primordial germ cellsfrom murine epiblasts by synergistic action of BMP4 and BMP8Bsignaling pathways. Proc. Natl. Acad. Sci. USA 98, 7858–7862.
Ying, Y., Zhao, G.Q., 2001. Cooperation of endoderm-derived BMP2 andextraembryonic ectoderm-derived BMP4 in primordial germ cell gen-eration in the mouse. Dev. Biol. 232, 484–492.
Yoshikawa, Y., Fujimori, T., McMahon, A.P., Takada, S., 1997. Evidencethat absence of Wnt-3a signaling promotes neuralization instead ofparaxial mesoderm development in the mouse. Dev. Biol. 183, 234–242.
Yoshimizu, T., Obinata, M., Matsui, Y., 2001. Stage-specific tissue andcell interactions play key roles in mouse germ cell specification. De-velopment 128, 481–490.
Yurugi-Kobayashi, T., Itoh, H., Yamashita, J., Yamahara, K., Hirai, H.,Kobayashi, T., Ogawa, M., Nishikawa, S., Nishikawa, S.I., Nakao, K.,2003. Effective contribution of transplanted vascular progenitor cellsderived from embryonic stem cells to adult neovascularization inproper differentiation stage. Blood 101, 2675.
14 D.A.F. Loebel et al. / Developmental Biology 264 (2003) 1–14