lipid droplets and cellular lipid metabolism • lipid droplets and cellular lipid metabolism 691....
TRANSCRIPT

BI81CH28-WaltherFarese ARI 3 May 2012 14:22
Lipid Droplets and CellularLipid MetabolismTobias C. Walther1 and Robert V. Farese Jr.21Department of Cell Biology, Yale University School of Medicine, New Haven,Connecticut 06520; email: [email protected] Institute of Cardiovascular Disease, Department of Medicine, and Departmentof Biochemistry and Biophysics, University of California, San Francisco, California 94158;email: [email protected]
Annu. Rev. Biochem. 2012. 81:687–714
First published online as a Review in Advance onApril 13, 2012
The Annual Review of Biochemistry is online atbiochem.annualreviews.org
This article’s doi:10.1146/annurev-biochem-061009-102430
Copyright c⃝ 2012 by Annual Reviews.All rights reserved
0066-4154/12/0707-0687$20.00
Keywordsorganelle, energy metabolism, triacylglycerol, membranes, obesity,oil, fat
AbstractAmong organelles, lipid droplets (LDs) uniquely constitute a hydropho-bic phase in the aqueous environment of the cytosol. Their hydrophobiccore of neutral lipids stores metabolic energy and membrane compo-nents, making LDs hubs for lipid metabolism. In addition, LDs areimplicated in a number of other cellular functions, ranging from pro-tein storage and degradation to viral replication. These processes arefunctionally linked to many physiological and pathological conditions,including obesity and related metabolic diseases. Despite their impor-tant functions and nearly ubiquitous presence in cells, many aspects ofLD biology are unknown. In the past few years, the pace of LD in-vestigation has increased, providing new insights. Here, we review thecurrent knowledge of LD cell biology and its translation to physiology.
687
Ann
u. R
ev. B
ioch
em. 2
012.
81:6
87-7
14. D
ownl
oade
d fro
m w
ww
.ann
ualre
view
s.org
by H
arva
rd U
nive
rsity
on
10/2
8/14
. For
per
sona
l use
onl
y.

BI81CH28-WaltherFarese ARI 3 May 2012 14:22
Contents1. INTRODUCTION . . . . . . . . . . . . . . 6882. GENERAL PROPERTIES OF
LIPID DROPLETS . . . . . . . . . . . . . 6892.1. Historical Aspects of Lipid
Droplets . . . . . . . . . . . . . . . . . . . . . . 6892.2. Lipid Droplets Are Found in
Most Cells . . . . . . . . . . . . . . . . . . . . 6892.3. Lipid Droplets Separate a
Hydrophobic Phase from theAqueous Cytosol . . . . . . . . . . . . . . 689
3. LIPID DROPLETS SERVEMULTIPLE FUNCTIONS INCELLS . . . . . . . . . . . . . . . . . . . . . . . . . . 691
4. LIPID DROPLETS AREANALOGOUS TOLIPOPROTEINS. . . . . . . . . . . . . . . . 692
5. LIPID DROPLETFORMATION. . . . . . . . . . . . . . . . . . . 6925.1. Models of Lipid Droplet
Formation . . . . . . . . . . . . . . . . . . . . 6925.2. Identifying Genes in Lipid
Droplet Formation . . . . . . . . . . . . 6946. LIPID DROPLETS GROW BY
EXPANSION OR BYCOALESCENCE. . . . . . . . . . . . . . . . 694
7. LIPID DROPLET PROTEINS . . 6957.1. Specific Proteins Localize to
the Surfaces of LipidDroplets . . . . . . . . . . . . . . . . . . . . . . 695
7.2. Protein Signals that MediateLipid Droplet Targeting . . . . . . . 696
7.3. Cellular Pathways Involved inTargeting Proteins to LipidDroplets . . . . . . . . . . . . . . . . . . . . . . 698
7.4. Removal of Lipid DropletProteins . . . . . . . . . . . . . . . . . . . . . . 698
8. LIPID DROPLETS INTERACTWITH OTHER CELLULARORGANELLES . . . . . . . . . . . . . . . . . 699
9. LIPID DROPLET MOVEMENTWITHIN CELLS . . . . . . . . . . . . . . . 699
10. LIPIDS IN LIPID DROPLETSARE MOBILIZED BYLIPASES . . . . . . . . . . . . . . . . . . . . . . . . 70010.1. Lipid Droplet Catabolism
by Lipolysis . . . . . . . . . . . . . . . . . . . 70010.2. The Fate of Hydrolyzed
Lipids . . . . . . . . . . . . . . . . . . . . . . . . 70110.3. Lipid Droplet Catabolism
by Autophagy . . . . . . . . . . . . . . . . . 70111. LIPID DROPLETS FIGURE
PROMINENTLY INPHYSIOLOGY ANDDISEASE . . . . . . . . . . . . . . . . . . . . . . . . 70111.1. Lipid Droplets and Lipid
Storage in Tissues . . . . . . . . . . . . . 70111.2. Excessive Lipid Droplet
Storage and Disease . . . . . . . . . . . 70311.3. Lipodystrophies and Too
Few Lipid Droplets . . . . . . . . . . . . 70411.4. Lipid Droplets and
Cancer . . . . . . . . . . . . . . . . . . . . . . . . 70411.5. Lipid Droplets and
Lactation . . . . . . . . . . . . . . . . . . . . . 70412. LIPID DROPLETS ARE
TARGETS OF INDUSTRIALOIL PRODUCTION . . . . . . . . . . . . 705
Lipid droplet (LD):the cytoplasmicorganelle composed ofa hydrophobic core ofneutral lipids boundedby a phospholipidmonolayer and specificproteins
1. INTRODUCTIONLipid droplets (LDs) are dynamic cytoplasmicorganelles found nearly ubiquitously in cells.They are linked to many cellular functions,including lipid storage for energy generationand membrane synthesis, viral replication, andprotein degradation. LD biology is connectedto myriad physiological processes, metabolic
diseases, and oil production. Interest in LDbiology has surged in recent years, reflectingrecognition of the importance of this relativelyunderstudied organelle and opportunities tounravel questions concerning LD biology.
Here, we focus primarily on recent dis-coveries in LD biology, building on otherreviews (1–3), and highlight many areas of LD
688 Walther · Farese
Ann
u. R
ev. B
ioch
em. 2
012.
81:6
87-7
14. D
ownl
oade
d fro
m w
ww
.ann
ualre
view
s.org
by H
arva
rd U
nive
rsity
on
10/2
8/14
. For
per
sona
l use
onl
y.

BI81CH28-WaltherFarese ARI 3 May 2012 14:22
Phase: a region of asystem in which theproperties, measuredin terms of orderparameters, areuniform. In anemulsion, two liquidphases are mixed
Neutral lipid: anonpolar, hydrophobiclipid
Sterol ester (SE): anester of cholesterol (ora related sterol) and afatty acid moiety; aneutral lipid
biology where knowledge is incomplete orcontroversial.
2. GENERAL PROPERTIESOF LIPID DROPLETS
2.1. Historical Aspectsof Lipid Droplets
LDs were identified by light microscopy as cel-lular organelles in the nineteenth century. Formany years, LDs often were called liposomes.In the late 1960s, a method was developed togenerate vesicles in vitro, also called liposomes,and these vesicles assumed the name. Sincethen, LDs have been referred to as LDs, lipidbodies, fat bodies, oil bodies, spherosomes, oradiposomes. In the past decade, the field settledprimarily on the name LDs.
For years, LDs were largely ignored in cellbiology research, presumably because theywere perceived as inert lipid globules with littlefunctional relevance. In 1991, seminal workby Constantine Londos and coworkers (4)identified a protein, perilipin, that specificallylocalized to LD surfaces, and this opened thedoor to mechanistically studying the organelle.More recently, research on LD biology has ac-celerated dramatically. This reflects increasedinterest in the basic biology of prevalentmetabolic diseases linked to LDs (e.g., obesity)and technological advances, which have shednew light on LDs and highlighted manyunanswered basic questions.
2.2. Lipid Droplets Are Foundin Most CellsNearly all cells have LDs or the capacityto form them. Several bacteria store lipidsin LDs, including predominantly the actino-mycetes group (e.g., Mycobacteria, Rhodococcus,Streptomyces, and Nocardia) (5).
Among eukaryotes, LDs are easily detectedin budding yeast (Saccharomyces cerevisiae), facil-itating genetic screens for altered LD morphol-ogy (6, 7). In higher eukaryotes, many cells havesome LDs, and culturing cells with fatty acids
(FAs) stimulates LD formation. Some cells,such as adipocytes or hepatocytes, exhibit manyLDs at baseline owing to active lipid synthesisand storage. LD abundance varies dynamicallyin cells. For example, in S. cerevisiae, LDs arefound most prominently during the stationaryphase, and catabolism of LDs in the exponen-tial phase is coordinated with an increased needfor phospholipids during cell division (8). Also,the number of LDs increases up to severalfoldin yeast with cellular stress (9, 10) and in cellsof some cancers (11). The connections of LDsto these states are poorly understood.
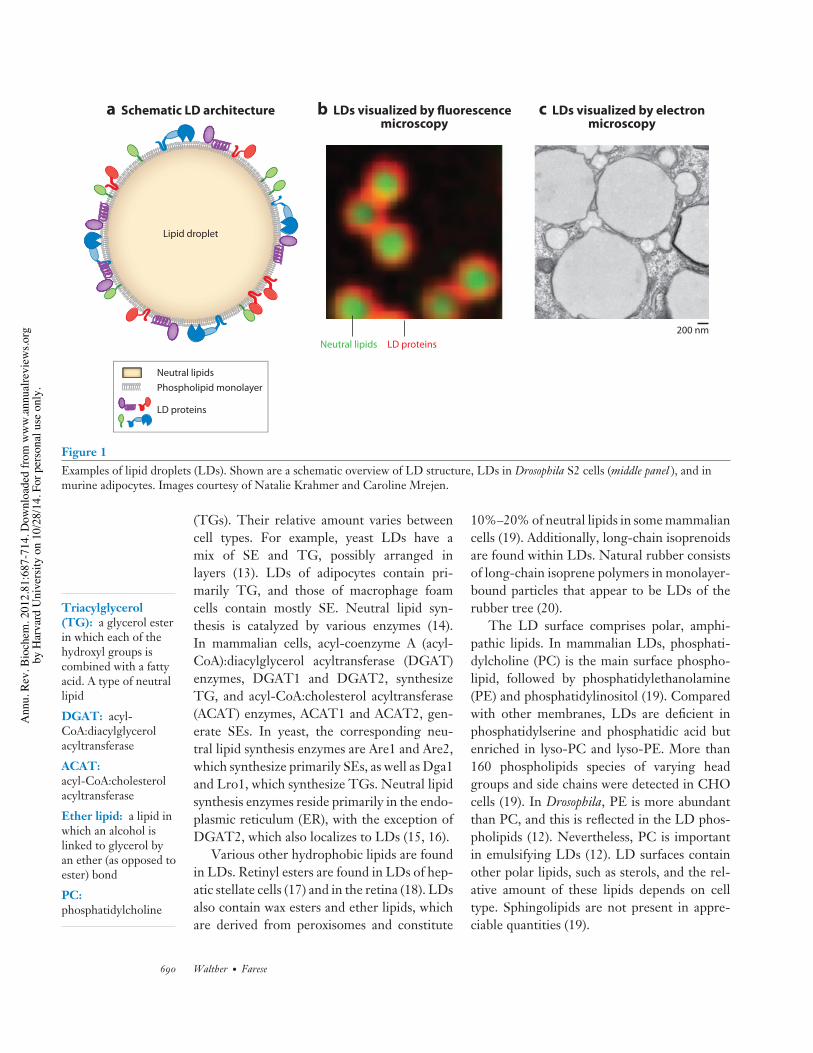
LD number and size in different cell typesor between individual cells of a population dif-fer considerably. Many cells have small LDs(100–200-nm diameters), whereas LDs in whiteadipocytes have diameters up to 100 µm andfill almost the entire cytoplasm. With diame-ters from 100 nm to 100 µm, the correspond-ing surface area and volume for LDs vary by106 and 109, respectively. Many LDs are visiblewith light microscopy (Figure 1 and the sidebartitled Imaging of Lipid Droplets).
LD size can change rapidly. Oleate load-ing of Drosophila S2 cells increases the LDmean diameter nearly threefold within hours,corresponding to an almost 30-fold volumeincrease (12). LDs also grow rapidly duringadipogenesis, when cells increase their capacityto synthesize lipids de novo. In contrast, LDsshrink within hours of culturing cells withlimited nutrients.
2.3. Lipid Droplets Separate aHydrophobic Phase from theAqueous CytosolAmong cellular organelles, LDs are uniquelycomposed of an organic phase of neutral lipids.This hydrophobic core is separated from theaqueous cytosol by a monolayer of surface phos-pholipids. The cytoplasm contains an emulsionof LDs in the cytosol. The LD phase of theemulsion provides a large interface for interac-tions with amphipathic molecules.
The primary neutral lipids of the LD coreare sterol esters (SEs) and triacylglycerols
www.annualreviews.org • Lipid Droplets and Cellular Lipid Metabolism 689
Ann
u. R
ev. B
ioch
em. 2
012.
81:6
87-7
14. D
ownl
oade
d fro
m w
ww
.ann
ualre
view
s.org
by H
arva
rd U
nive
rsity
on
10/2
8/14
. For
per
sona
l use
onl
y.
BI81CH28-WaltherFarese ARI 3 May 2012 14:22
200 nm
Lipid droplet
a Schematic LD architecture b LDs visualized by fluorescencemicroscopy
c LDs visualized by electronmicroscopy
LD proteinsNeutral lipids
Phospholipid monolayer
LD proteins
Neutral lipids
Figure 1Examples of lipid droplets (LDs). Shown are a schematic overview of LD structure, LDs in Drosophila S2 cells (middle panel ), and inmurine adipocytes. Images courtesy of Natalie Krahmer and Caroline Mrejen.
Triacylglycerol(TG): a glycerol esterin which each of thehydroxyl groups iscombined with a fattyacid. A type of neutrallipid
DGAT: acyl-CoA:diacylglycerolacyltransferase
ACAT:acyl-CoA:cholesterolacyltransferase
Ether lipid: a lipid inwhich an alcohol islinked to glycerol byan ether (as opposed toester) bond
PC:phosphatidylcholine
(TGs). Their relative amount varies betweencell types. For example, yeast LDs have amix of SE and TG, possibly arranged inlayers (13). LDs of adipocytes contain pri-marily TG, and those of macrophage foamcells contain mostly SE. Neutral lipid syn-thesis is catalyzed by various enzymes (14).In mammalian cells, acyl-coenzyme A (acyl-CoA):diacylglycerol acyltransferase (DGAT)enzymes, DGAT1 and DGAT2, synthesizeTG, and acyl-CoA:cholesterol acyltransferase(ACAT) enzymes, ACAT1 and ACAT2, gen-erate SEs. In yeast, the corresponding neu-tral lipid synthesis enzymes are Are1 and Are2,which synthesize primarily SEs, as well as Dga1and Lro1, which synthesize TGs. Neutral lipidsynthesis enzymes reside primarily in the endo-plasmic reticulum (ER), with the exception ofDGAT2, which also localizes to LDs (15, 16).
Various other hydrophobic lipids are foundin LDs. Retinyl esters are found in LDs of hep-atic stellate cells (17) and in the retina (18). LDsalso contain wax esters and ether lipids, whichare derived from peroxisomes and constitute
10%–20% of neutral lipids in some mammaliancells (19). Additionally, long-chain isoprenoidsare found within LDs. Natural rubber consistsof long-chain isoprene polymers in monolayer-bound particles that appear to be LDs of therubber tree (20).
The LD surface comprises polar, amphi-pathic lipids. In mammalian LDs, phosphati-dylcholine (PC) is the main surface phospho-lipid, followed by phosphatidylethanolamine(PE) and phosphatidylinositol (19). Comparedwith other membranes, LDs are deficient inphosphatidylserine and phosphatidic acid butenriched in lyso-PC and lyso-PE. More than160 phospholipids species of varying headgroups and side chains were detected in CHOcells (19). In Drosophila, PE is more abundantthan PC, and this is reflected in the LD phos-pholipids (12). Nevertheless, PC is importantin emulsifying LDs (12). LD surfaces containother polar lipids, such as sterols, and the rel-ative amount of these lipids depends on celltype. Sphingolipids are not present in appre-ciable quantities (19).
690 Walther · Farese
Ann
u. R
ev. B
ioch
em. 2
012.
81:6
87-7
14. D
ownl
oade
d fro
m w
ww
.ann
ualre
view
s.org
by H
arva
rd U
nive
rsity
on
10/2
8/14
. For
per
sona
l use
onl
y.

BI81CH28-WaltherFarese ARI 3 May 2012 14:22
IMAGING OF LIPID DROPLETS
Given their size (0.5 µm to tens of microns), many LDs are readily detected by light mi-croscopy (see Figure 1, middle panel). Their refracting nature makes them identifiable withphase-contrast imaging. Several hydrophobic dyes can be used to image LDs. Oil red O isvisible by light microscopy and typically used to stain fixed samples. Nile red and BODIPYare concentrated in LDs and fluoresce upon stimulation. They provide a robust signal inliving or fixed cells. Care must be taken to avoid artifacts. BODIPY appears to be more spe-cific for staining LDs; Nile red also stains endocytic vesicles. Additionally, BODIPY-taggedFAs can stain LDs, as the tagged FAs in many cases are taken up by cells and esterified andincorporated into neutral lipids.
Because LDs are closely associated with other organelles, it can be difficult to distin-guish them from surrounding membranous organelles. This limitation was addressed bythe recent development of “super high-resolution” microscopy that overcomes the clas-sical resolution limits of light microscopy, in some cases improving resolution by a fac-tor of 10 (i.e., from ∼250 nm to ∼25 nm) (166). In one approach, exemplified by pho-toactivated localization microscopy or stochastic optical reconstruction microscopy, thesequential photoactivation of single fluorescent molecules allows for the precise deter-mination of their position by fitting a Gaussian curve to the resulting signal, leading tohigh-resolution reconstructed images. In a complementary approach (stimulated emissiondepletion microscopy), the resolution limit is increased further by depleting the excitedstate of fluorophores around a narrow focus of light, thus restricting molecules that emit asignal.
Several approaches based on Raman spectroscopy are also used to image LDs. Mostprominently, coherent anti-Stokes Raman scattering microscopy uses inelastic scatteringof photons interacting with matter to detect molecule-specific vibrations and resultingblue-shifted light (167). These approaches also can quantify lipids.
EM methods are also used to analyze LDs (see Figure 1, right panel), including trans-mission EM and freeze-fracture EM. Difficulties of fixing and staining lipids during samplepreparation often cause LDs to appear as empty spheres in conventional EMs. Fixation andsectioning problems may lead to artifacts, but improved protocols can avoid these problems(89, 168).
3. LIPID DROPLETS SERVEMULTIPLE FUNCTIONSIN CELLSForemost, LDs are intracellular lipid reser-voirs, providing building blocks for membranesor substrates for energy metabolism. Packag-ing highly reduced, hydrophobic lipids, suchas TGs, in a phase without water provides themost efficient form of energy storage. For ex-ample, an average non-obese person stores upto 2,500 kJ of metabolic energy in glycogen, but
>500,000 kJ in TGs of adipocyte LDs, enoughto run ∼30 marathons.
LDs also serve as organizing centers for syn-thesizing specific lipids. TGs, for example, aresynthesized in the ER and at LDs (15). Otherenzymes of lipid synthesis (e.g., ergosterol) lo-calize to LDs, suggesting links between lipidsynthesis pathways and LDs (21).
LDs have been linked to protein storage(22). For example, during embryogenesisin Drosophila, histones localize to LDs until
www.annualreviews.org • Lipid Droplets and Cellular Lipid Metabolism 691
Ann
u. R
ev. B
ioch
em. 2
012.
81:6
87-7
14. D
ownl
oade
d fro
m w
ww
.ann
ualre
view
s.org
by H
arva
rd U
nive
rsity
on
10/2
8/14
. For
per
sona
l use
onl
y.

BI81CH28-WaltherFarese ARI 3 May 2012 14:22
HCV: hepatitis Cvirus
Lipoprotein: a lipid-protein particle thatcirculates in thebloodstream anddistributes lipidsthroughout the body
needed for rapid nuclear division associatedwith embryo segmentation (23). LDs may alsotemporarily store unfolded membrane proteinsbefore proteasomal degradation.
LDs are involved in hepatitis C virus (HCV)assembly. During viral replication, the HCVcore protein is cleaved from the precursor viralpolypeptide and binds to LDs (reviewed in Ref-erence 24) via amphipathic helices. New HCVvirions are assembled in LDs and ER mem-branes, where viral RNA is packaged with cap-sid proteins. Mature viruses are secreted as partof a lipoprotein-virus particle. LDs may pro-vide a location for HCV core proteins untilviral assembly or until cells degrade the over-expressed protein. Notably, HCV core proteinlocalization to LDs requires DGAT1 activity,and HCV core proteins bind to DGAT1 in vitro(25). Moreover, blocking DGAT1 activity di-minishes viral replication (25).
4. LIPID DROPLETS AREANALOGOUS TO LIPOPROTEINSLDs share structural features with plasmalipoproteins. Both contain neutral lipid coresencased in polar lipid monolayers. Addition-ally, both particles are decorated with spe-cific surface proteins, often possessing amphi-pathic α-helices. However, LDs (with diame-ters of 100 nm up to 100 microns) are generallymuch larger than lipoproteins [diameters rangefrom <20 nm (e.g., high-density lipoprotein) to∼500 nm (e.g., chylomicrons)].
Although the formation of both LDs andlipoproteins is linked to neutral lipid synthesisin the ER, their physiological functions differ:LDs primarily store lipids, and lipoproteinsdistribute lipids in the body. Also, only afew cell types (e.g., hepatocytes, enterocytes,and yolk sac endodermal cells) express therequired proteins, such as apolipoprotein (apo)B and the microsomal TG transfer protein, forlipoprotein assembly, whereas most cells makeLDs. Thus, lipoproteins may have evolved byadapting LDs to secretion.
How cells regulate the fate of newlysynthesized neutral lipids—storage in LDsversus secretion on lipoproteins—is mostly
unknown. Secretory cells may store lipids inLDs only after exceeding the capacity to as-semble and secrete lipoproteins. Alternatively,secretion may be activated when a storagethreshold is achieved. Specific LD proteins[including cell death–inducing DFF45-likeeffector (CIDE) proteins] may direct the LDpool of TG toward lipoprotein formation (26).Secretion of TG via lipoproteins is thought toinvolve their hydrolysis at LDs and resynthesisin the ER (27).
5. LIPID DROPLET FORMATION
5.1. Models of LipidDroplet Formation
Many organelles self-replicate. However, LDslikely form de novo. In bacteria, LDs form bylipid synthesis in the cell-delimiting membrane(28). In eukaryotes, LDs may arise primarilyfrom the ER, where the enzymes that synthe-size neutral lipids reside (14). In yeast geneti-cally engineered to lack LDs, induction of LDformation shows they invariably arise from orclose to the ER (7, 29). LDs appear to remainin contact with the ER once formed, and pro-teins that associate with both compartmentsmove between them (29). However, light mi-croscopy, with limited resolution, cannot deter-mine if such proteins reside directly on the LDsurface or in ER membranes in close apposition.Many studies employing electron microscopy(EM) suggest a tight assocation of LDs andthe ER. In mammalian cells, such studies showmembrane cisternae, which could be connectedto the ER, in close proximity to LDs (30, 31).Also, LDs in hepatocytes are tightly associatedwith ER membrane cisternae, marked by lumi-nal apo B100 protein, and these cisternae maybe continuous with LDs (31).
Despite these findings, the molecular mech-anisms of LD formation are not understood.How does a monolayer-coated LD arise froma bilayer membrane? Because neutral lipid syn-thesis enzymes reside in the ER, the products ofthese enzymes might occupy space between themembrane bilayers, forming a lens of neutral
692 Walther · Farese
Ann
u. R
ev. B
ioch
em. 2
012.
81:6
87-7
14. D
ownl
oade
d fro
m w
ww
.ann
ualre
view
s.org
by H
arva
rd U
nive
rsity
on
10/2
8/14
. For
per
sona
l use
onl
y.

BI81CH28-WaltherFarese ARI 3 May 2012 14:22
TG, SE
DAGCholesterol
FA-CoA
TG synthesis onlipid droplets
Lipid droplet fusion
Triacylglycerol
PC to balancesurface expansion
DGATACAT
Lipid droplet formation Lipid droplet growth
ERlumen
Lipid droplet
Cytoplasm
Figure 2Models of lipid droplet (LD) formation and expansion. For formation, the budding model is shown, with theLD either remaining attached to the endoplasmic reticulum (ER) or detached. The neutral lipid synthesisenzymes, ACAT or DGAT, catalyze the formation of lipids that fill the core. For lipid droplet expansion, themodels of local synthesis of triacylglycerol (TG) at the LD and of LD fusion (coalescence) are shown.Abbreviations: DAG, diacylglycerol; FA-CoA, fatty acyl-coenzyme A; SE, sterol ester;PC, phosphatidylcholine.
lipids. In vitro studies with ER microsomesfrom sunflower (Helianthus annuus) found for-mation of small liquid lenses of 60-nm diameterin the ER bilayer that, similar to LDs, containfreely mobile TG and recruit oleosins (32). It isunclear, however, how neutral lipids are orga-nized into nascent LDs, whether this happens atspecific locations in the ER, or whether specificproteins are involved. In mammalian cells, per-ilipin3/Tip47 is recruited to the ER during lipidstorage (33) and was suggested to be requiredfor LD formation (34). However, many or-ganisms (e.g., yeast) have no perilipin 3/Tip47ortholog, and many cells appear not to expressit (e.g., Drosophila S2 cells), so this mechanismalone cannot mediate LD formation.
Several models of LD formation have beenproposed (1). They include (a) ER budding,where LDs grow from the ER bilayer and re-main connected or bud off (Figure 2); (b) bi-celle formation in which an entire lipid lens isexcised from the ER (35); (c) vesicular budding
in which a bilayer vesicle forms, followed by fill-ing of the bilayer intramembranous space withneutral lipids (36); and (d ) an “eggcup” model inwhich a LD grows within a concave depressionof the ER through transport of neutral lipidsfrom the ER (37). The latter transfer process re-sembles a model for prokaryotes (28). All mod-els posit LDs forming toward the cytosolic faceof the ER membrane. However, cells, such ashepatocytes, also secrete neutral lipids into theER lumen (38), and LDs could be derived fromluminal origins. Such a model was recently pro-posed for yeast (29).
Which model is correct? Several obstaclesprevent an easy answer. Most cells have LDs,complicating identification of nascent LDs, andsystems of induced LD formation in mammalsare lacking. Also, the LD size during the ini-tial stages of formation is below the resolu-tion of light microscopy. Thus, combinationsof inducible systems with super high-resolutionlight microscopy and EM will likely be required
www.annualreviews.org • Lipid Droplets and Cellular Lipid Metabolism 693
Ann
u. R
ev. B
ioch
em. 2
012.
81:6
87-7
14. D
ownl
oade
d fro
m w
ww
.ann
ualre
view
s.org
by H
arva
rd U
nive
rsity
on
10/2
8/14
. For
per
sona
l use
onl
y.

BI81CH28-WaltherFarese ARI 3 May 2012 14:22
Lipodystrophy: apathological conditioncharacterized byseverely diminished orabsent adipose tissue
Lipolysis: theregulated breakdownof complex neutrallipids, such astriacylglycerols orsterol esters, into theircomponents
to gain further insights (see the sidebar titledImaging of Lipid Droplets). Alternatively, char-acterizing a system for in vitro LD formationfrom ER membranes might allow moleculardissection of different steps of formation.
5.2. Identifying Genes in LipidDroplet FormationTo identify genes involved in LD formation,several studies utilized genome-wide screenswith LD morphology as a phenotypic readout.Three screens in S. cerevisiae (6, 7, 39) identi-fied gene deletions leading to different LD mor-phologies. Notably, deleting the yeast orthologof seipin (FLD1) caused abnormal LDs. Seipinis of particular interest in LD biogenesis. It isa multimeric ER membrane protein (40) whosedeficiency in humans results in lipodystrophy.Surprisingly, there was otherwise little overlapbetween genes identified in the yeast screens,and no single-gene deletion was found to causethe complete absence of LDs.
Genome-wide screens to find genes regu-lating LD morphology have been performedby RNAi in Drosophila cells (41, 42). In onescreen employing oleate loading of cells, over200 genes were identified. Interestingly, theybinned into five phenotypic classes (41). In an-other screen, over 800 genes showed an effecton LD accumulation (42). Among overlappinghits of both screens knockdowns of proteasomegenes yielded fewer LDs, knockdowns of theARF/COPI vesicular transport machinery gavelarge and disperse LDs, and knockdowns ofgenes related to phospholipid synthesis led tovery large LDs. The latter two classes showeddefects in lipolysis, indicating functional conse-quences of these knockdowns. The ARF/COPImachinery functions in retrograde transport ofproteins from the Golgi apparatus to the ER,but its role in lipolysis is uncertain. Adiposetriglyceride lipase (ATGL), or its Drosophilahomolog Brummer, fails to target to LDs incells depleted of ARF/COPI machinery (30,43). The link between phospholipid synthesisgenes and LD size is discussed below.
Several systematic screens of genes in wholeorganisms also focused on lipid storage. Screensin Caenorhabditis elegans (44) and Drosophila (45)revealed a plethora of genes involved in fat stor-age. Although many of these genes participatein feeding behavior and energy expenditure,some may be involved directly with LDs. Othersurveys identified proteins that might be im-portant in LD formation. These include fat-inducing transcript (FIT) proteins, which areER proteins that bind TG and have been im-plicated LD assembly (46, 47). FIT2 overex-pression results in more LDs and knockdownin fewer LDs and TG, but there is no evidencethat FIT proteins affect DGAT activity (47).How FIT proteins organize TG into LDs isunclear. Phospholipases might also be requiredfor LD biogenesis (48).
6. LIPID DROPLETS GROWBY EXPANSION ORBY COALESCENCETo accommodate more TG, cells form newLDs and grow existing ones (Figure 2). Addingneutral lipids to existing LDs requires local syn-thesis or transfer from the ER.
Neutral lipids might be synthesized lo-cally at LDs, particularly during lipid loading.DGAT2, which catalyzes the final step of TGsynthesis and is normally found in the ER, lo-calizes to LDs during oleate loading (16, 49), asdoes its yeast ortholog Dga1 (29). It is unclearwhether more proximal steps in TG synthesisalso localize to LDs. In contrast to TG synthe-sis, sterol esterification likely occurs primarilyin the ER because ACAT enzymes reside in theER and do not localize to LDs (50).
The synthesis of neutral lipids in the ERnecessitates their transfer to LDs, throughmembrane connections or via interorganelletransport by transfer proteins. Although TGtransfer proteins move TG from the ER bilayerto nascent lipoproteins in the ER lumen, nosuch mechanism is known for cytosolic LDs.
LDs may also grow by coalescence(Figure 2) (51). Coalescence of neutral LDsminimizes the phase boundary area and free
694 Walther · Farese
Ann
u. R
ev. B
ioch
em. 2
012.
81:6
87-7
14. D
ownl
oade
d fro
m w
ww
.ann
ualre
view
s.org
by H
arva
rd U
nive
rsity
on
10/2
8/14
. For
per
sona
l use
onl
y.

BI81CH28-WaltherFarese ARI 3 May 2012 14:22
energy of the emulsion. Thus, the challenge forcells is to prevent coalescence and maintain LDsas isolated entities. LD emulsions are stabilizedby surface proteins (e.g., oleosins) or surfac-tants (e.g., PC) (12), which prevent coalescenceby lowering surface tension. In contrast, accu-mulation of fusogenic lipids (e.g., phosphatidicacid) might induce LD coalescence (39).
During rapid expansion, LD diameters in-crease more than threefold within hours—anearly tenfold increase in surface area. Thus,cells need to synthesize and transport largeamounts of phospholipids to expanding LD sur-faces. Among phospholipids, PC is key for coat-ing LDs and preventing their coalescence. PCcan emulsify artificial LDs in vitro and pre-vent coalescence (12). During expansion, LDsbecome PC deficient and CTP:phosphocholinecytidylyltransferase enzymes, which catalyzethe rate-limiting step in PC biosynthesis, areactivated by binding to LDs (12). The de novosynthesized PC is transported to the expand-ing LD surface through unknown mechanisms.In this manner, de novo synthesis of PC is ac-tivated to maintain adequate PC levels at LDsurfaces.
PC to coat the surfaces of LDs is also synthe-sized from lyso-PC and fatty acyl-CoA precur-sors by lyso-PC acyltransferase enzymes, whichfunction in the Lands cycle of phospholipid re-modeling of FA moieties. Lyso-PC acyltrans-ferase 1 and 2 enzymes localize to and are activeon LDs (49). They likely function in remodel-ing the surface phospholipids rather than pro-viding net PC synthesis, as they cannot yieldincreased PC unless lyso-PC is provided.
In Drosophila cells, LD fusion happens rarelyexcept under specific conditions, such as PCdeficiency (12, 41). PC deficiency likely rendersLDs prone to coalesce. A recent report foundhomotypic LD fusion to occur rarely in murineembryonic fibroblasts or NIH-3T3 cells undernormal conditions (52), although higher ratesof fusion were also reported (51). However,numerous pharmacological agents stimulatedLD fusion in different cell types. Theseincluded propranolol and other drugs, albeit atsupraphysiological concentrations, which may
trigger fusion by inserting into and disruptingLD surface monolayers (52). Fusion was rela-tively slow, occurring over seconds to minutes.The volumes of fusing LDs were conserved,and surface areas were decreased, suggestingexcess surface lipids are removed during fusion.
Proteins might be involved in catalyz-ing LD fusion. One such protein is afat-specific protein of 27 kDa [FSP27, orcell death-inducing DFF45-like effector C(Cidec)]. In adipocytes, FSP27 is expressed asa peroxisome proliferator-activated receptor-γ(PPARγ)-regulated gene and promotes the for-mation of unilocular LDs (53–55). Mice lackingFsp27/Cidec have multilocular white adipocytes,implying the protein forms large LDs (53, 55,56). If Fsp27 is expressed ectopically in fi-brobasts, LD sizes increase. The mechanism isunclear but may include promoting LD fusionor inhibiting lipolysis (53–55, 57, 58).
Because SNARE proteins (soluble N-ethylmaleimide-sensitive factor attachment re-ceptor proteins) mediate homotypic fusion ofbilayer-bounded vesicles during cellular traf-ficking, they are attractive candidates for LDfusion. In studies performed in NIH 3T3cells, knockdowns of genes encoding SNAP23,syntaxin-5, and VAMP4 were reported to de-crease the rate of LD fusion (59). However,it is unclear how SNARE proteins would me-diate fusion of monolayer-bound vesicles, andmore studies to address SNARE involvementare needed.
7. LIPID DROPLET PROTEINS
7.1. Specific Proteins Localize to theSurfaces of Lipid Droplets
LDs are characterized by specific proteins ontheir surfaces. Analyses of LD proteomes indifferent organisms yielded diverse lists of LD-associated proteins (21, 60–65). The relativelyeasy purification of LDs (by centrifugation)facilitates proteomic analyses, but their closeassociation with other organelles, particularlythe ER, confounds such analyses. Thus, it isoften unclear which proteins are genuine LD
www.annualreviews.org • Lipid Droplets and Cellular Lipid Metabolism 695
Ann
u. R
ev. B
ioch
em. 2
012.
81:6
87-7
14. D
ownl
oade
d fro
m w
ww
.ann
ualre
view
s.org
by H
arva
rd U
nive
rsity
on
10/2
8/14
. For
per
sona
l use
onl
y.
BI81CH28-WaltherFarese ARI 3 May 2012 14:22
proteins. In addition, researchers may overlookLD proteins because they have other well-known functions. For example, histones wereunexpectedly found by LD proteomics and sub-sequently confirmed to transiently target LDsin Drosophila embryos (23). LDs may similarlytransiently store other proteins that otherwisemight aggregate. For example, α-synuclein, aParkinson’s disease–associated protein proneto self-aggregation, localizes to LDs (66).
Some data on LD protein localization can bemisleading. By fluorescence microscopy, manyproteins reportedly localize to LDs upon oleateloading of cells. These studies assume that acharacteristic ring-like appearance of proteinssurrounding LDs indicates targeting to the LD-delimiting surface monolayer. In ultrastructuralstudies, however, membranous structures oftenaccumulate juxtaposed to LDs, so some pro-teins might localize to LD-associated mem-branes rather than the LD surface itself (30).Indeed, many reports show no overlap betweenthe LD signal (e.g., from neutral lipid staining)and the LD-encircling protein rings. Becauselight microscope resolution is ∼250 nm, such
proteins might actually localize to membranesat a distance from the LD surface.
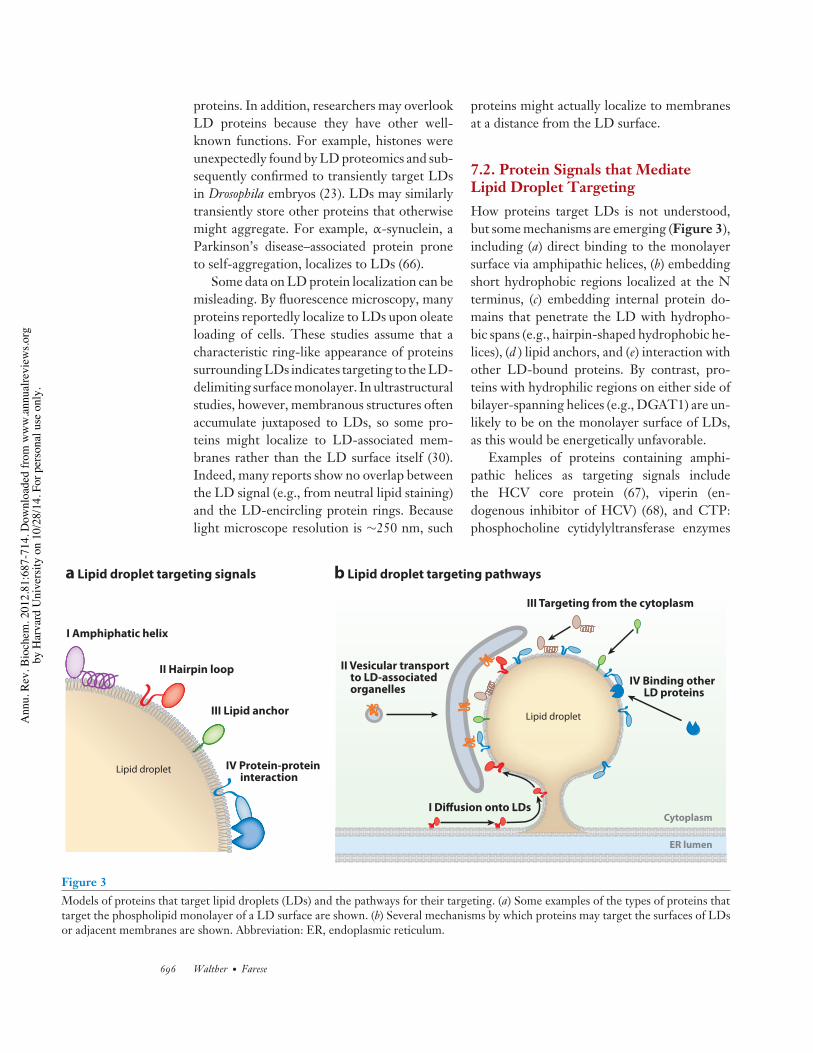
7.2. Protein Signals that MediateLipid Droplet TargetingHow proteins target LDs is not understood,but some mechanisms are emerging (Figure 3),including (a) direct binding to the monolayersurface via amphipathic helices, (b) embeddingshort hydrophobic regions localized at the Nterminus, (c) embedding internal protein do-mains that penetrate the LD with hydropho-bic spans (e.g., hairpin-shaped hydrophobic he-lices), (d ) lipid anchors, and (e) interaction withother LD-bound proteins. By contrast, pro-teins with hydrophilic regions on either side ofbilayer-spanning helices (e.g., DGAT1) are un-likely to be on the monolayer surface of LDs,as this would be energetically unfavorable.
Examples of proteins containing amphi-pathic helices as targeting signals includethe HCV core protein (67), viperin (en-dogenous inhibitor of HCV) (68), and CTP:phosphocholine cytidylyltransferase enzymes
ER lumen
IV Binding other LD proteins
I Diffusion onto LDs
III Targeting from the cytoplasm
II Vesicular transport to LD-associated organelles
III Lipid anchor
I Amphiphatic helix
II Hairpin loop
a Lipid droplet targeting signals b Lipid droplet targeting pathways
Cytoplasm
Lipid droplet
Lipid droplet IV Protein-protein interaction
Figure 3Models of proteins that target lipid droplets (LDs) and the pathways for their targeting. (a) Some examples of the types of proteins thattarget the phospholipid monolayer of a LD surface are shown. (b) Several mechanisms by which proteins may target the surfaces of LDsor adjacent membranes are shown. Abbreviation: ER, endoplasmic reticulum.
696 Walther · Farese
Ann
u. R
ev. B
ioch
em. 2
012.
81:6
87-7
14. D
ownl
oade
d fro
m w
ww
.ann
ualre
view
s.org
by H
arva
rd U
nive
rsity
on
10/2
8/14
. For
per
sona
l use
onl
y.

BI81CH28-WaltherFarese ARI 3 May 2012 14:22
(12, 41). How amphipathic helices distinguishthe LD surface monolayer from other cellularmembranes is unclear. CTP:phosphocholinecytidylyltransferase 1 binds to and is activatedby PC-deficient LD surfaces (12), one of fewcases where a specific lipid composition of theLD surface mediates amphipathic helix recruit-ment. Another example is recruitment of per-ilipin3/Tip47 to the ER with diacylglycerol(DAG) accumulation, which was suggested tocoordinate DAG buildup with LD protein re-cruitment and LD formation (33).
For other organelles, specific lipids (e.g.,phosphoinositides, phosphatidylserine, andphosphatidic acid) mediate protein targetingto membranes. No lipid signals are knownto target proteins to LDs, but it is possiblebecause the lipid composition of the LDsurface differs from that of the ER (19, 69).
Several proteins (e.g., AAM-B, UBXD8,or ALDI) (70, 71) possess an N-terminal,hydrophobic, short domain (minimally ∼28amino acids) that is necessary and sufficient forLD targeting. These proteins localize to theER under some conditions (e.g., in the absenceof LDs), suggesting they first are inserted intothe ER and subsequently transported to LDs.Their domain structure and the molecularmechanism(s) to target LDs are unknown.
Some proteins (e.g., caveolin, oleosins, and17-hydroxysteroid dehydrogenases) have an in-ternal hydrophobic domain of variable lengththat likely forms a hairpin structure that couldbe integrated into a phospholipid bilayer or LDmonolayer. For plant oleosins, members of thisclass, the topology and some requirements havebeen elucidated. Oleosins, which account for upto 8% of seed protein, are important for stor-ing oils in seeds. LD targeting of oleosins ini-tiates with their synthesis at the ER (72, 73),followed by targeting to LDs. A hairpin con-tains a central proline knot (three prolines in12 amino acids) that is essential for LD target-ing and might induce a sharp bend between twoadjacent hydrophobic protein segments (74).The hairpin domain might thermodynamicallyfavor LD localization (i.e., the space betweenthe ER membrane bilayer may be limiting).
Alternatively, proteins recognizing such hair-pins may be in involved in LD localization.
In mammalian cells, caveolins contain a sim-ilar hairpin motif. Caveolins primarily local-ize to caveolae at the plasma membrane butalso to LDs. Initially, LD targeting of cave-olin was observed in cells treated with brefeldinA, which blocks membrane trafficking, or withcaveolin-1 mutants (75–77). However, target-ing occurs under physiological conditions (e.g.,during lipid loading) (78, 79). At the plasmamembrane, caveolin topology includes a centralhairpin membrane anchor and adjacent lipid-binding domains. For LDs, the topology ofcaveolin is unknown, but it may be similar. Thehydrophobic sequences of AAM-B, ALDI, orDGAT2 may also form hairpins. For enzymes,such as DGAT2, it has yet to be demonstratedthat the protein localizes directly on the LDsurface (versus the adjacent ER), and, if so,whether it is active at this location.
Several different LD targeting mecha-nisms exist for perilipins (perilipin1, per-ilipin2/ADRP/adipophilin, perlipin3/Tip47,perilipin4/S3-12, and perilipin5/OXPAT),which are the first identified specific LDmarker proteins (80). Perilipins are notessential for LD formation (e.g., yeast lackperilipins) (34) but are important for regulatinglipid metabolism at LDs. In mammals, severalperilipins are expressed ubiquitously (e.g., per-ilipin2/ADRP and perilipin3/Tip47), whereasothers are expressed specifically in certain celltypes (e.g., perilipin1 in white adipocytes andperilipin5/OXPAT in highly oxidative celltypes).
Adipocytes express high levels of perilipin1,which is involved in the regulation of lipolysis.Perilipin1 contains a combination of domainsinteracting with LDs. Particularly importantare three C-terminal hydrophobic stretches,which may penetrate LDs (81). Additionalamphipathic stretches of the protein likely in-teract with the LD surface. Similarly, both N-and C-terminal regions of perilipin2/ADRPcontribute to LD binding (82, 83). Perilipin2/ADRP localization to LDs may also involveARF1/COPI (84).
www.annualreviews.org • Lipid Droplets and Cellular Lipid Metabolism 697
Ann
u. R
ev. B
ioch
em. 2
012.
81:6
87-7
14. D
ownl
oade
d fro
m w
ww
.ann
ualre
view
s.org
by H
arva
rd U
nive
rsity
on
10/2
8/14
. For
per
sona
l use
onl
y.

BI81CH28-WaltherFarese ARI 3 May 2012 14:22
Perilipin3/Tip47 may bind to LDs likeapolipoprotein E (apo E) binds to lipoproteins.Its C-terminal four-helix bundle and α/β-domain (85) are similar to the N-terminal four-helix bundle of apo E. Apo E binds lipoproteinsby opening its four-helix bundle and exposinghydrophobic, amphipathic sequences to thelipid surface (86). Perilipin3/Tip47 might usea similar mechanism. Unlike perilipin1 andperilipin2/ADRP, which are unstable whennot bound to LDs, perilipin3/Tip47 is foundin the cytoplasm when LDs are absent (87).
As an alternative to protein segments me-diating LD interactions, lipid modifications ofproteins may serve as anchors to the LD sur-face. Rab18, a small GTPase, localizes to LDs,where it mediates ER interactions (88, 89). Byanalogy to other Rabs with C-terminal lipid an-chors, Rab18 might target to LDs by a prenyla-tion anchor combined with protein-protein in-teractions. Unlike other Rabs, Rab18 containsone, rather than two, lipid modification.
Proteins might also localize to LDs by in-teracting with LD-bound proteins. Regulatedrecruitment of hormone-sensitive lipase (HSL)to LDs is an example. Under basal conditions,HSL is mostly cytosolic, and access to LDs isrestricted by perilipin1. Upon hormonal stim-ulation, perilipin1 is phosphorylated, which re-cruits HSL to LDs.
7.3. Cellular Pathways Involved inTargeting Proteins to Lipid DropletsPathways targeting proteins to LDs are lessunderstood than the protein signals. Severalmechanisms may be involved, including directrecruitment from the cytosol (Figure 2). Forsome membrane proteins, vesicular traffickingmay be important. The ARF/COPI machin-ery, which mediates vesicular trafficking fromthe Golgi apparatus to the ER, is requiredfor normal LD turnover and localization ofthe major TG lipase, ATGL, or its Drosophilahomolog, brummer, to LDs (30, 90). A frac-tion of ATGL colocalized with ER-exit sites(ER membrane domains dedicated to forma-tion of secretory vesicles) and the expression of
a dominant-negative Sar1 (GTPase that is re-quired for vesicles to leave the ER exit sites)inhibited ATGL targeting to LDs (30). ATGLrequires a C-terminal hydrophobic stretch forLD localization (91) and may behave biochem-ically as a membrane-associated protein (30).Thus, ATGL may contain a hydrophobic se-quence (e.g., hairpin loop) targeted to the ERand trafficked to LDs. From EM studies, cis-ternal structures around LDs could represent aLD-target compartment for vesicular traffick-ing (30). It is unclear how ATGL or similarproteins would move from this compartmentto the LD surface.
Physical bridges may connect LDs andthe ER (29, 31, 92). Such bridges may allowmembrane-bound proteins, such as those withhairpins, to diffuse from the bilayer to the LDsurface. Data from yeast support this model; flu-orescently labeled LD proteins, such as Dga1,rapidly exchange with other membrane pools(29).
7.4. Removal of Lipid Droplet ProteinsMechanisms to remove LD proteins arelikely similar to those for membrane pro-teins. Endocytosed membrane proteins orautophagic vesicles are degraded in lysosomes.ER membrane proteins can be degraded byproteasomes (ER-associated degradation).Both mechanisms may degrade LD proteins.LDs are delivered to autophagosomes (93), andsome LD proteins are modified by ubiquitin forproteasomal degradation (e.g., some perilipins)(94–100). Additionally, a nonbiased screenidentified proteasome components as requiredfor normal LD morphology (41).
Molecular links between LDs and protea-somal degradation were recently identified.Among LD proteins, ancient ubiquitous pro-tein 1 (Aup1), located in the ER or directly onLDs, binds ubiquitin E2 ligase, Ube2g2 (101,102). The ubiquitin E3 ligase, spartin/SPG20,localizes to LDs, and its depletion or overex-pression leads to LD accumulation, perhapsfrom the altered turnover of LD proteins,such as perilipin2/ADRP (97, 100). These find-
698 Walther · Farese
Ann
u. R
ev. B
ioch
em. 2
012.
81:6
87-7
14. D
ownl
oade
d fro
m w
ww
.ann
ualre
view
s.org
by H
arva
rd U
nive
rsity
on
10/2
8/14
. For
per
sona
l use
onl
y.

BI81CH28-WaltherFarese ARI 3 May 2012 14:22
ings imply that a specific machinery degradesLD-associated proteins, but the details areunknown.
Some ER-associated degradation substratesmay transiently localize to LDs (e.g., HMG-CoA reductase when proteasomal degrada-tion was inhibited) (103). However, at least inyeast, LDs are not required for ER-associateddegradation (104).
8. LIPID DROPLETS INTERACTWITH OTHER CELLULARORGANELLESLDs interact with other organelles, includ-ing the ER, endosomes, mitochondria, andperoxisomes. Freeze-fracture imaging showedER membranes in mammalian cells are of-ten wrapped around LDs in a shape resem-bling an eggcup (37). Because interactionsbetween membrane-bound organelles are im-plicated in lipid exchange, apposition of LDswith the ER may facilitate lipid transfer be-tween them. Oxysterol-binding proteins lo-calize to LDs (105), where they might beinvolved in lipid trafficking or neutral lipidmetabolism. In steroidogenic cells, ER-LD in-teractions might function in the synthesis orcatabolism of steroid hormones (106). In yeast,ER-LD junctions at least partly reflect interme-diates of LD formation (6, 7, 29, 40), and seipinmay participate in this process.
Mechanisms mediating ER association withLDs are unknown, but the small GTPase Rab18may be involved (88, 89). Because Rab18 is re-cruited to LDs late, its localization likely doesnot reflect a role during LD formation. Rab18localization is mutually exclusive with LD lo-calization of perilipin 2/ADRP or a dominant-negative mutant of caveolin (88, 89), suggestinga specialized function. Indeed, Rab18 localiza-tion to LDs in cultured adipocytes is related tolipolysis stimulation (89).
Sometimes endosomal structures appear toenwrap LDs (89). These interactions may beimportant for delivering LDs to lysosomes byautophagy in macrophages to generate choles-terol for ABCA1-dependent efflux (107). Rab5
was detected on purified LDs and implicatedin the interaction of early endosomes with LDs(108).
LDs also associate with mitochondria (109)and peroxisomes (110). These interactions maychannel FAs liberated from lipolysis to sitesof oxidation. Supporting this notion, exercisetraining increases the number of LDs and theircontacts with mitochondria in skeletal muscle(111).
9. LIPID DROPLET MOVEMENTWITHIN CELLSLDs are usually dispersed and move in smalloscillations. However, in mammalian cells,LD movement is sometimes coordinated. Per-ilipin1 expression leads to LD clustering, whichis reversed upon perilipin1 phosphorylation(112). Similarly, when Drosophila cells (whichlack perilipin1) synthesize TG for a prolongedtime (e.g., during 24 h of oleate loading), newlyformed LDs eventually cluster into organizedsuperstructures (41). Increased clustering is alsoobserved when genes involved in protein syn-thesis are depleted from S2 cells (41). Whatdetermines this clustering and its purpose areunknown.
LD distribution in the cell may be partiallydetermined by active transport via micro-tubules. LDs move directionally in axons ofAplysia by uncharacterized mechanisms (113).LDs in the Drosophila embryonal syncitiummove synchronously from the periphery toa central location and back again. Althoughthe function of this movement is unclear, itsdisruption leads to altered transparency of theembryo, providing a system to screen for mu-tants of LD transport machinery (114). Suchstudies revealed that LDs move bidirectionallyalong microtubules with dynein and kinesinmotors, which pull LDs in opposite directions.Net movement depends on the relative ac-tivities of the motors. The LD protein Halointeracts with kinesin1 as part of a complex thatincludes the regulatory Klar protein and theperilipin-like protein Lsd2 (115). The complexappears to link LDs to microtubules and to
www.annualreviews.org • Lipid Droplets and Cellular Lipid Metabolism 699
Ann
u. R
ev. B
ioch
em. 2
012.
81:6
87-7
14. D
ownl
oade
d fro
m w
ww
.ann
ualre
view
s.org
by H
arva
rd U
nive
rsity
on
10/2
8/14
. For
per
sona
l use
onl
y.

BI81CH28-WaltherFarese ARI 3 May 2012 14:22
regulate kinesin1’s activity during LD trans-port. Microtubules have also been implicatedin LD clustering in mammalian cells, but sup-porting evidence is limited. Interestingly, theHCV core protein promotes clustering of LDsaround microtubule organizing centers (116).
10. LIPIDS IN LIPID DROPLETSARE MOBILIZED BY LIPASES
10.1. Lipid Droplet Catabolismby Lipolysis
In lipolysis, lipids are hydrolyzed to liber-ate FAs. Cells storing TG use lipolysis togenerate energy and membrane lipids. Inmulticellular organisms, energy storage occurspredominantly in adipose tissue, and mostof the knowledge concerning lipolysis comesfrom studies in adipocytes, where FAs arehydrolyzed sequentially by ATGL, HSL, andmonoacylglycerol lipase. During lipolysis, LDsshrink as core lipids are catabolized. With de-creased volume, the surface area contracts. It isunknown if excess proteins and phospholipidsare resorbed into the ER or degraded.
ATGL contains an N-terminal domain withsimilarity to patatin, a plant acyl-hydrolase ex-pressed highly in the potato tuber, which con-tains the catalytic activity. It is unknown whichof the TG fatty acyl-esters are hydrolyzed byATGL. Among the potential DAG species pro-duced, only sn-1,2-DAG is known to act as a sig-naling molecule, and sn-1,3-DAG is not likelyto be an optimal substrate for re-esterificationby DGAT enzymes.
The ATGL N terminus also interacts withCGI-58, which has a hydrolase fold and is anATGL activator. The activation mechanism isunknown, but it requires interaction of bothproteins and CGI-58 with LDs (117). G0S2may inhibit ATGL, although its function is un-clear (118). Interaction of ATGL and perilipin1regulates lipolysis. In mammalian adipocytes,lipolysis is initiated by hormones that triggerβ-adrenergic stimulation to activate protein ki-nase A (PKA). In unstimulated cells, CGI-58is bound to LD-localized perilipin1. Perilipin1
restricts basal lipolysis by sequestering CGI-58or by sterically shielding LDs from lipolysis, orby a combination of both (119, 120). Upon acti-vation, perilipin1 is phosphorylated by cAMP-dependent PKA at Ser492 and Ser517, releasingCGI-58 to bind and activate ATGL (121).
ATGL’s interaction with CGI-58 alsotargets the lipase to LDs, where it is activated(122, 123). Similarly, interactions of ATGLand perilipin5/OXPAT (but not other perilip-ins) recruit ATGL to LDs, but in this case, theinteraction limits lipolysis (122, 123). Becauseperilipin5 is expressed mostly in oxidativetissues, perilipin5’s interaction with ATGLmight be used in those tissues to regulateATGL differently than it does in others.Perilipin5/OXPAT interacts with CGI-58,possibly forming high-molecular-weight as-semblies (122). These data suggest a complexinterplay among ATGL, perilipins, and CGI-58 to mediate LD targeting and lipolysis (122).
HSL catalyzes the second step of lipolysisand is also regulated by hormones. PKA acti-vation leads to HSL phosphorylation at mul-tiple sites, of which Ser650 (human HSL) isparticularly important (124). Phosphorylationincreases HSL lipolytic activity twofold. Thismechanism is combined with regulated recruit-ment of HSL to LDs (125). Specifically, PKA-phosphorylated perilipin1 (at Ser81, Ser222,and Ser276, particularly) recruits HSL from thecytosol to LDs (125). With HSL phosphoryla-tion, recruitment to LDs increases HSL activity100-fold (126).
In the final lipolysis step, monoacylglycerolis hydrolyzed by monoacylglycerol lipase andpossibly by HSL (127). No evidence indicatesthe monoacylglycerol lipase–catalyzed reactionoccurs at LDs or is regulated by hormones.
Lipolysis in nonadipocyte cells is not aswell understood, but similar steps must exist.Unfortunately, sequence homology cannotpredict functions in proteins with hydrolasemotifs. Adiponurtrin/PNPLA3, another mem-ber of the patatin-like family, is expressed inliver, and a human adiponurtrin/PNPLA3variant (I148M) is associated with nonal-coholic fatty liver disease (128). However,
700 Walther · Farese
Ann
u. R
ev. B
ioch
em. 2
012.
81:6
87-7
14. D
ownl
oade
d fro
m w
ww
.ann
ualre
view
s.org
by H
arva
rd U
nive
rsity
on
10/2
8/14
. For
per
sona
l use
onl
y.

BI81CH28-WaltherFarese ARI 3 May 2012 14:22
adiponurtrin/PNPLA3 knockout mice do nothave LD and TG accumulation in liver (129,130), so the biochemical and physiologicalroles of this protein are unknown.
In addition to TG, other lipids are hy-drolyzed at LD surfaces by phospholipases andother enzymes. For example, anandamide andother N-acylethanolamines traffic to and arecatabolized at LDs (131).
10.2. The Fate of Hydrolyzed LipidsFAs liberated from TGs in LDs have one ofseveral fates. They can be re-esterified to TG,used for β-oxidation to generate energy, usedas building blocks for membrane lipid synthe-sis, or used as cofactors for cell signaling, orexported.
In adipocytes, many FAs generated bylipolysis are re-esterified by DGAT or MGATenzymes to TG, although the percentagedrops with fasting (132). It is unclear whetherre-esterification occurs in the ER or at LD sur-faces. Because DGAT enzymes use activatedFAs bound to CoA, re-esterification includesgenerating fatty acyl-CoA esters by long-chainacyl-CoA synthetases (ACSLs) in a reactionthat requires the input of cellular energy.Among ACSL proteins, ACSL3 and -4 arerecruited to LDs specifically during lipolysis,and ACSL1 constitutively localizes there (62,65). Specific ACSLs, therefore, may act atLD surfaces as part of the re-esterificationcycle. The function of TG turnover in thisfutile cycle is unknown but may fine-tunelipolysis and preserve stores of essentialFAs.
FAs liberated from LD hydrolysis can be uti-lized directly for β-oxidation. Contact sites be-tween LDs and mitochondria and peroxisomesmay directly transfer FAs (discussed above). FAsmay also be used to synthesize membrane lipids.Like re-esterification, this reaction requires FAactivation by attaching a CoA moiety, possiblyat LDs (65, 133). FAs liberated by ATGL alsoserve as endogenous activators of PPARα, asshown by lack of PPARα signaling in hearts ofATGL-deficient mice (134).
10.3. Lipid Droplet Catabolismby AutophagyLipids in LDs can also be degraded by lipaseswithin lysosomes after delivery by macroau-tophagy. Like lipolysis, macroautophagy is in-duced during nutrient deprivation and respondsto hormonal signals. In macroautophagy ofLDs (or macrolipophagy), an LD, whole or inpart, is engulfed by a membrane bilayer withthe activated PE-modified LC3-II protein. Theresulting autophagosome containing an LD isdelivered to the lysosome where acid lipases lib-erate FAs from TG. This last step recapitulatesthe fate of lipoprotein-derived TG deliveredto lysosomes via endocytosis. The relative con-tributions of LD autophagy to LD catabolismare unclear and may vary among tissues. LDautophagy was first reported in hepatocytes,where expression levels of ATGLs and HSLsare relatively low (93). Hepatic-specific inacti-vation of autophagy leads to hepatic steatosis,consistent with a function of macroautophagyin hepatic lipid metabolism. The molecu-lar events that trigger LD autophagy areunknown.
11. LIPID DROPLETS FIGUREPROMINENTLY IN PHYSIOLOGYAND DISEASEWe cannot review in detail the metabolic andphysiological processes that involve lipid stor-age and LDs. Instead, we focus on describingLDs in various cell types and present examplesof how LDs are linked to physiology and dis-ease. Other reviews address medical aspects ofLD biology (135).
11.1. Lipid Droplets and LipidStorage in Tissues11.1.1. Adipose tissue. Some insects andmost vertebrates have highly specialized whiteadipocytes and adipose tissue. In most inverte-brates, TG is distributed throughout the body,often in connective tissue that fills gaps betweenother tissues, or in enterocytes of C. elegans. In
www.annualreviews.org • Lipid Droplets and Cellular Lipid Metabolism 701
Ann
u. R
ev. B
ioch
em. 2
012.
81:6
87-7
14. D
ownl
oade
d fro
m w
ww
.ann
ualre
view
s.org
by H
arva
rd U
nive
rsity
on
10/2
8/14
. For
per
sona
l use
onl
y.

BI81CH28-WaltherFarese ARI 3 May 2012 14:22
flies, fat is located predominantly in a centrallylocated fat body. In vertebrates, adipose tissueis distributed to distinct regions and regulatedby factors, such as hormones. With the evolu-tionary advent of homeothermy (in birds andmammals), adipose tissue gained a function asa subcutaneous insulator (136).
Adipocytes are the most highly specializedcell type for storing lipids in LDs, and inwhite adipocytes, a single large LD frequentlyoccupies most of the cytoplasm. Human fatcells can store vast quantities of energy in LDs,primarily as TG. Adipocyte LDs also storecholesterol esters and fat-soluble vitamins. Asexpected, the adipocyte gene expression profilereflects the high flux of lipid synthesis, storage,and turnover. Adipocyte fat content is coupledto leptin expression, the main adipocyte-derived hormone that regulates long-termenergy homeostasis (e.g., by regulating foodintake) (137).
Brown adipocytes in mammals catabolizelipids in mitochondria that are uncoupled fromoxidative phosphorylation to generate heat.LDs are prominent but smaller and more nu-merous than those in white adipocytes. Thestudy of brown adipocytes is likely to shed lighton interactions among LDs, mitochondria, andFA oxidation.
11.1.2. Liver. Next to adipose tissue, liver hasthe greatest capacity to store lipids in LDs.At basal conditions, the fraction of the cy-toplasm occupied by LDs in hepatocytes issmaller than in adipoctyes, but they can holdmassive amounts of TG, as illustrated by foiegras. In humans, abnormal LD accumulation iscalled hepatic steatosis.
LDs in hepatocytes are typically multiloc-ular, and their formation and consumption aredynamic in normal physiology. For example,during overnight fasting in mice, large amountsof FAs are mobilized from TG stores in whiteadipose tissue and taken up by the liver to formLDs, generating a fatty liver. Hepatocytesexpress DGATs and ACATs, and the lipidcomposition of LDs partly reflects substrateavailability. The liver also has large numbers
of stellate cells that store retinol (vitamin A) asretinyl esters in LDs (17).
11.1.3. Small intestine. LDs are dynamic,prominent organelles in intestinal enterocytes(138). They reflect the enormous capacity ofsmall intestine to absorb fat from a meal. Ente-rocytes have a large surface area and significantability to synthesize and store TGs. The intes-tine absorbs over 95% of the TG in a typicalmeal, leading to rapid formation of LDs in en-terocytes until TGs are exported from the cellas part of chylomicrons. This process typicallytakes minutes to hours after a meal. Many as-pects of this process remain unclear.
11.1.4. Yolk sac. During embryonic develop-ment in many animals, the yolk sac endodermfunctions like the liver and intestine to holdlipids from the mother that are destined for thedeveloping embryo (139). As in the intestinalor liver cells, similar processes of fat uptake,storage in LDs, and export on apoB-containinglipoproteins occur in the yolk sac (139).
11.1.5. Skeletal muscle. Oxidative (type I,slow-twitch) muscles have a high rate ofoxidative metabolism and are rich in mi-tochondria and LDs. In obese individuals,intramyocellular TG accumulation in type Imuscles is associated with insulin resistance.However, in highly trained athletes, TG storesin LDs in oxidative muscle are increasedwithout negative consequences. This “athlete’sparadox” has been linked to DGAT1 function(140, 141). LDs in oxidative muscle are oftenclose to mitochondria (111, 142). Maintainingcoupling of lipid storage with consumptionof lipids for fuel appears to be important forefficient energy utilization.
11.1.6. Adrenal cortex. LDs are prominentin the adrenal cortex. They store large amountsof SEs, presumably as cholesterol for steroidhormone synthesis. In fact, the yellow colorof adrenal glands is due to lipid storage, andmice lacking ACAT1, and thus cholesterol es-ters and LDs, have pale-colored adrenal glands.
702 Walther · Farese
Ann
u. R
ev. B
ioch
em. 2
012.
81:6
87-7
14. D
ownl
oade
d fro
m w
ww
.ann
ualre
view
s.org
by H
arva
rd U
nive
rsity
on
10/2
8/14
. For
per
sona
l use
onl
y.

BI81CH28-WaltherFarese ARI 3 May 2012 14:22
Surprisingly, adrenocortical hormone synthe-sis in ACAT1 knockout mice is normal. Likeadrenocortical cells, other steroidogenic cells,such as Leydig cells of the testes, often haveprominent LDs. Additionally, LDs are foundin oocytes.
11.1.7. Macrophages. Macrophages storelipids in LDs (e.g., after phagocytosis of mod-ified lipoproteins in the arterial wall) un-til they are exported. Macrophages expressACAT and DGAT enzymes, and the con-tent of their LDs reflects their exposure tosubstrates. Macrophage foam cells store largeamounts of cholesterol from lipoproteins, suchas low-density lipoproteins. Macrophages uti-lize LDs to store arachidonic acid, the precur-sor for bioactive lipids (e.g., prostaglandins andleukotrienes) in inflammation.
11.2. Excessive Lipid DropletStorage and DiseaseMany metabolic diseases are characterized byexcessive lipid storage in LDs. In these dis-eases, lipids may exceed the cellular capacityto store them and to buffer against their toxiceffects. Unesterified lipids, such as cholesterolor FAs and their derivatives, can trigger in-flammatory responses that result in tissue dam-age, fibrosis, scarring, and potentially organfailure.
11.2.1. Obesity and diabetes as diseases ofexcessive lipid droplets. Obesity is a state ofexcess lipid storage and LDs in the adipose tis-sue, often accompanied by lipid deposition andexcessive LDs in nonadipose tissue, which causelipotoxicity or tissue dysfunction. Whetherobesity results in complications, such as type 2diabetes mellitus, may relate to the capacityof LDs in adipocytes or macrophages to storelipids. In transgenic mice, increasing adipocytecapacity to store TG results in obesity withoutdiabetes (143, 144), thereby uncoupling thetwo. Similarly, increasing DGAT1 expres-sion selectively in macrophages avoids manymetabolic consequences of obesity (145). The
LD capacity of macrophages to store neutrallipids therefore might profoundly influencepathophysiology. Taken together, LD-targetedstrategies for therapies for pathologies associ-ated with obesity can be aimed at blocking lipidabsorption and LD formation in the small intes-tine, preventing influx into the system; catabo-lizing, rather than storing, excess lipids throughoxidation; and increasing LD capacity inadipocytes or macrophages to store toxic lipidsand prevent associated inflammation and tissuedamage.
Obesity is often accompanied by hepaticsteatosis or LD accumulation in the liver, af-fecting millions of people. Many individualsprogress to hepatic inflammation (called non-alcoholic steatohepatitis), and some go on todevelop fibrosis and liver failure. Possibly, thecapacity of hepatocytes to store lipids in LDsis exceeded, promoting inflammation. Hepaticsteatosis is also strongly associated with hep-atic insulin resistance, although the causativemechanism is unclear. Proposed contributingmechanisms include alterations in insulin sig-naling pathways mediated by bioactive lipids,ER stress, and activation of inflammation sig-naling (146). A common theme, however, re-lates to limited hepatocyte capacity to secreteor store lipids, thereby leading to lipid excessand liver dysfunction.
11.2.2. Atherosclerosis and macrophagelipid droplets. Like obesity, atherosclerosis isassociated with excessive deposition of lipidsin tissues, in this instance cholesterol, derivedfrom apo B-containing lipoproteins in arterialwalls. Excess cholesterol is taken up and es-terified by macrophages. Cholesterol esters inturn are stored in LDs until they can be mo-bilized and effluxed to high-density lipopro-teins, which are transported to the liver forclearance of cholesterol in bile. Macrophagesfull of cholesterol ester–containing LDs arecalled foam cells. LDs protect macrophagefoam cells from toxicity resulting from ex-cess free cholesterol. Deletion of ACAT1,the primary enzyme responsible for esterifyingcholesterol in macrophages, in mice does not
www.annualreviews.org • Lipid Droplets and Cellular Lipid Metabolism 703
Ann
u. R
ev. B
ioch
em. 2
012.
81:6
87-7
14. D
ownl
oade
d fro
m w
ww
.ann
ualre
view
s.org
by H
arva
rd U
nive
rsity
on
10/2
8/14
. For
per
sona
l use
onl
y.

BI81CH28-WaltherFarese ARI 3 May 2012 14:22
prevent atherosclerosis and even exacerbatesit (147). Additionally, lack of cholesterol es-terification in macrophages results in depo-sition of large amounts of free cholesterolin the skin and brains of ACAT1 knockoutmice that were crossed into a hyperlipidemicgenetic background (147). These studies high-light the lipid-buffering function of LDs andhow storage of potentially toxic lipids preventslipotoxicity.
11.2.3. The heart and lipid droplets. Al-though FAs are a fuel for heart muscle, few largeLDs are normally present in cardiac myocytes.With lipid overload, however, LDs accumulateand heart dysfunction may occur. In murinemodels, overexpression of ACSL1 in the heartresults in cardiomyopathy and severe heart dys-function (148). Interestingly, this can be res-cued by targeting DGAT1 overexpression tothe heart, which esterifies the excess fatty acyl-CoAs to store them as TG in LDs (149). In con-trast, ATGL deletion causes lipid accumulationin the heart and severe cardiomyopathy, whichresults in death of mice (150). Intriguingly, thisphenotype can be rescued by treatment with aPPARα agonist (134), which likely increases fatoxidation in the tissue.
11.3. Lipodystrophies and Too FewLipid DropletsAbsence or deficiency of white adipose tissue,or lipodystrophy (reviewed in Reference 151),results in an LD deficiency. Genetic causes forlipodystrophy include mutations in genes thatencode for an acylglycerol-phosphate acyl-transferase (AGPAT2), seipin (BCSL2), cave-olin (CAV1) for generalized lipodystrophy, andlamin A/C (LMNA), peroxisome proliferator-activated receptor-γ (PPARG), Akt2/proteinkinase B (AKT2), and endoprotease Face-1(ZMPSTE24) for partial lipodystrophy. Thepathogenesis of lipodystrophies is not under-stood for all causes but can relate to impair-ments in the development of white adipose tis-sue; reduced ability to synthesize glycerolipidsand TGs; or an apparent deficiency in LD
formation, as in lipodystrophy related to theabsence of seipin (BSCL2). When LDs ofwhite adipose tissue are insufficient to storeTGs, the liver assumes this role, and massivehepatic steatosis often results. Associatedleptin deficiency results in many metabolicabnormalities, including lipid deposition innonadipose tissue and insulin resistance. As aconsequence, leptin replacement has emergedas a therapy for some lipodystrohies.
11.4. Lipid Droplets and CancerMetabolism in cancer cells, including lipidmetabolism (11), has received increased inter-est for therapeutic interventions. Most can-cer cells upregulate FA synthesis, presumablyto provide FAs for membrane proliferation.Some tumor cells and associated inflammatorycells have prominent LDs (11). The mechanismfor LD accumulation in cancer cells is unclearbut could relate to increased FA synthesis orimpairments in FA oxidation.
11.5. Lipid Droplets and LactationLDs are prominent in mammary epithelial cellsof lactating mammary glands, where they essen-tially serve as a secreted organelle, transferringlipid nutrients and energy from mother to child.In this process, LDs are secreted from the apicalside of epithelial cells (reviewed in Reference152), and at least three proteins are involved.Butyrophilin, a transmembrane protein, local-izes to the apical plasma membrane. Xanthineoxidoreductase, an enzyme oxidizing purinesand other molecules, has a role in the envelop-ment of LDs by the plasma membrane (153).Perilipin2/ADRP may act as a LD-specificanchor. The disruptions of butyrophilin andxanthine oxidoreductase genes resulted inimpaired lactation (153, 154). Interestingly,the lactation defect in perilipin2/ADRP genedisruption was not severe, most likely becausea truncated protein was still produced fromthe allele (155). Disruption of DGAT1 alsoimpairs lactation in homozygous females (156),which relates to impaired development of the
704 Walther · Farese
Ann
u. R
ev. B
ioch
em. 2
012.
81:6
87-7
14. D
ownl
oade
d fro
m w
ww
.ann
ualre
view
s.org
by H
arva
rd U
nive
rsity
on
10/2
8/14
. For
per
sona
l use
onl
y.

BI81CH28-WaltherFarese ARI 3 May 2012 14:22
mammary gland and lack of LDs for milkproduction (157).
12. LIPID DROPLETS ARETARGETS OF INDUSTRIALOIL PRODUCTIONLike other organisms, plants store TG. In par-ticular, seeds store oil, and different speciesstore varying amounts of oil in LDs (often re-ferred to as oil bodies) in different parts of theseed. Most plants, including soybean (Glycinemax), sunflower (Helianthus annuus), safflower(Carthamus tinctorius), and Brasicaceae, store oilin the embryo itself. In other plants, such ascastor bean (Ricinus communis), oil accumulatesmostly in the triploid endosperm surround-ing the embryonic tissue. In most of thesespecies, the TG provides metabolic energy forthe seedling upon germination. Some species,such as oat grains, fuse large numbers of LDsin the endosperm, and this oil attracts fruitpredators that disperse seeds. The FA compo-sition of TG in seeds differs widely with vari-ous chain length and levels of desaturation andis genetically controlled. The synthesis of TGis mediated by enzymes analogous to those inmammalian cells, including GPATs, AGPATs,lipin (PA hydrolase), and DGATs. In additionto DGAT1 and DGAT2, an additional DGATenzyme was recently purified from the cytosolof peanuts (Arachis hypogaea) (158). A poly-morphism of DGAT in maize (Zea mays) in-creases oil content substantially (159). As anadditional complexity in plants, TG generationinvolves cross talk between four different com-partments: the cytosol for glycerol production,plastids for the production of FAs, mitochon-dria for the production of very-long-chain FAsin some plants, and the ER for LD (oil body)formation.
LD production in plants is part of a highlyregulated differentiation program, and manyTG synthetic enzymes are under combinatorialcontrol of several transcription factors, includ-ing activators and repressors (160). Most likely
from this highly integrated process, attemptsto increase oil production in seeds by overex-pressing TG synthesis enzymes have thus farbeen disappointing. However, overexpressionof Arabidposis thaliana DGAT in tobacco plantsincreased TG content of leaves 20-fold, provid-ing a potential strategy for biofuel production(161).
When seeds germinate, they mobilize TGin oil bodies, using a lipase (Sdp1) similarto ATGL (162). Additional proteins, such assteroleosins, regulate lipolysis and β-oxidationduring germination (163). In EMs, plant LDsappear to contact glyoxisomes, where FAs maybe transported for β-oxidation (164).
Similar to animal LDs, plant oil bodies aredecorated with specific proteins. Most notably,oleosins prevent the coalescence of oil bodiesand shield them from lipolysis (165), with thelatter function similar to that of mammalianperilipins.
Industries have targeted biodiesel produc-tion from crops, such as palm or oilseed rape.However, increasing production of these plantsin which only a fraction is used for oil presentschallenges because it puts them in competitionfor arable land with food crops. Thus, TGproduction in algae, or other microorganisms,for biofuel has received increasing attention.To generate biofuels, algae must be grownand harvested, and the lipids extracted andprocessed. Considerable effort has gone intoidentifying suitable species for cultivationin open ponds or photobioreactors and intooptimizing other production steps. To beeconomcially and environmentally viable, bio-fuel production from algae must have a positivelife-cycle analysis, meaning a positive balanceof energy yield compared with the energyrequired for biofuel generation. This is chal-lenging, and thus efforts to optimize growthsystems and conditions are underway. Nev-ertheless, engineering of metabolism in algaetoward oil production (e.g., by overexpressingDGAT or acetyl-CoA carboxylase enzymes) ispromising.
www.annualreviews.org • Lipid Droplets and Cellular Lipid Metabolism 705
Ann
u. R
ev. B
ioch
em. 2
012.
81:6
87-7
14. D
ownl
oade
d fro
m w
ww
.ann
ualre
view
s.org
by H
arva
rd U
nive
rsity
on
10/2
8/14
. For
per
sona
l use
onl
y.

BI81CH28-WaltherFarese ARI 3 May 2012 14:22
SUMMARY POINTS
1. LDs are ubiquitous organelles in most cells and most organisms.
2. LDs primarily store lipids for metabolic energy and for membrane components. Theyare also implicated in protein storage and degradation and in complex processes, such asviral replication.
3. LDs represent an emulsion of a hydrophobic phase of neutral lipids, such as TGs and SEs,within the aqueous cytosol. Individual LDs are stabilized by a monolayer of phospholipidsinto which specific proteins are embedded. The molecular architectures of LDs andlipoproteins are similar.
4. LDs are believed to be formed at the ER, where neutral lipids are produced, and tobud toward the cytoplasm. They may remain attached to the ER or detach. LDs mayoriginate from other cell membranes. The mechanism for LD biogenesis is unclear.
5. LDs grow by expansion or coalescence (fusion). The molecular details of these processesare being unraveled. PC is important as a surfactant and to prevent coalescence.
6. Specific proteins target LDs through several general targeting motifs. The mechanismof how proteins traffic to LDs remains mostly unknown.
7. Lipids are mobilized from LDs through the sequential actions of lipases. In adiposetissue, this process is highly regulated. LDs may also be catabolized via macroautophagy.
8. LDs in different mammalian tissues have important functions in energy and membranehomeostasis. In adipose tissue, but also in liver and intestine, LDs store TGs for energygeneration. Macrophages utilize LDs to store and clear surplus cholesterol from theperiphery of the body. Excess or failure to form LDs leads to lipotoxicity and metabolicdiseases, including obesity, lipodystrophy, or atherosclerosis.
9. LDs are the cellular organelles that store oils in plants that are used for agriculturalproduction of food oils and biofuels.
FUTURE ISSUES
1. How are LDs formed? Is the ER the only place of LD formation in eukaryotes? Whatmechanisms are involved in their organization and budding reactions from the ERmembrane?
2. How are proteins targeted to LDs? Are there one or multiple targeting mechanisms?What is their relationship to secretory trafficking? How are LD proteins degraded?
3. How do LDs interact with other organelles, such as mitochondria, peroxisomes, and theER? What are the functions of these interactions?
4. What are the mechanisms of lipolysis? How do lipases access neutral lipids within LDs?What are the fates and functions of lipolysis intermediates and products? What are therelative contributions of lipolysis and macroautophagy to catabolism of LDs?
5. What determines the capacities of cells to store lipids and form LDs? What molecularmechanisms mediate lipotoxicity in different tissues?
706 Walther · Farese
Ann
u. R
ev. B
ioch
em. 2
012.
81:6
87-7
14. D
ownl
oade
d fro
m w
ww
.ann
ualre
view
s.org
by H
arva
rd U
nive
rsity
on
10/2
8/14
. For
per
sona
l use
onl
y.

BI81CH28-WaltherFarese ARI 3 May 2012 14:22
6. How can agricultural organisms be engineered to produce large amounts of desired lipidsin an easy, accessible, and energy-efficient manner?
DISCLOSURE STATEMENTThe authors are not aware of any affiliations, memberships, funding, or financial holdings thatmight be perceived as affecting the objectivity of this review.
ACKNOWLEDGMENTSWe acknowledge the many scientists who contributed to the knowledge in the field, some ofwhich we were unable to cite owing to space constraints. In particular, we salute the pioneeringcontributions of cell biologists Constantine Londos and Richard Anderson, who helped to launchthe field of LD biology. We thank current and former members of the Walther and Fareselaboratories for their many contributions to the ideas contained in the review, Sylvia Richmondfor assistance with manuscript preparation, and Gary Howard for editorial assistance.
LITERATURE CITED
1. Farese RV Jr, Walther TC. 2009. Lipid droplets finally get a little R-E-S-P-E-C-T. Cell 139:855–602. Fujimoto T, Parton RG. 2011. Not just fat: the structure and function of the lipid droplet. Cold Spring
Harb. Perspect. Biol. 3:a0048383. Thiele C, Spandl J. 2008. Cell biology of lipid droplets. Curr. Opin. Cell Biol. 20:378–854. Greenberg AS, Egan JJ, Wek SA, Garty NB, Blanchette-Mackie EJ, Londos C. 1991. Perilipin, a major
hormonally regulated adipocyte-specific phosphoprotein associated with the periphery of lipid storagedroplets. J. Biol. Chem. 266:11341–46
5. Alvarez HM, Steinbuchel A. 2002. Triacylglycerols in prokaryotic microorganisms. Appl. Microbiol.Biotechnol. 60:367–76
6. Fei W, Shui G, Gaeta B, Du X, Kuerschner L, et al. 2008. Fld1p, a functional homologue of humanseipin, regulates the size of lipid droplets in yeast. J. Cell Biol. 180:473–82
7. Szymanski K, Binns D, Bartz R, Grishin N, Li W, et al. 2007. The lipodystrophy protein seipin is foundat endoplasmic reticulum lipid droplet junctions and is important for droplet morphology. Proc. Natl.Acad. Sci. USA 104:20890–95
8. Kurat C, Wolinski H, Petschnigg J, Kaluarachchi S, Andrews B, et al. 2009. Cdk1/Cdc28-dependentactivation of the major triacylglycerol lipase Tgl4 in yeast links lipolysis to cell-cycle progression.Mol. Cell. 33:53–63
9. Yamamoto K, Takahara K, Oyadomari S, Okada T, Sato T, et al. 2010. Induction of liver steatosisand lipid droplet formation in ATF6α-knockout mice burdened with pharmacological endoplasmicreticulum stress. Mol. Biol. Cell 21:2975–86
10. Fei W, Wang H, Fu X, Bielby C, Yang H. 2009. Conditions of endoplasmic reticulum stress stimulatelipid droplet formation in Saccharomyces cerevisiae. Biochem. J. 424:61–67
11. Bozza PT, Viola JP. 2010. Lipid droplets in inflammation and cancer. Prostaglandins Leukot. Essent. FattyAcids 82:243–50
12. Krahmer N, Guo Y, Wilfling F, Hilger M, Lingrell S, et al. 2012. Phosphatidylcholine synthesis forlipid droplet expansion is mediated by localized activation of CTP:phosphocholine cytidylyltransferase.Cell Metab. 14:504–15
13. Czabany T, Wagner A, Zweytick D, Lohner K, Leitner E, et al. 2008. Structural and biochemicalproperties of lipid particles from the yeast Saccharomyces cerevisiae. J. Biol. Chem. 283:17065–75
www.annualreviews.org • Lipid Droplets and Cellular Lipid Metabolism 707
Ann
u. R
ev. B
ioch
em. 2
012.
81:6
87-7
14. D
ownl
oade
d fro
m w
ww
.ann
ualre
view
s.org
by H
arva
rd U
nive
rsity
on
10/2
8/14
. For
per
sona
l use
onl
y.

BI81CH28-WaltherFarese ARI 3 May 2012 14:22
14. Buhman KK, Chen HC, Farese RV Jr. 2001. The enzymes of neutral lipid synthesis. J. Biol. Chem.276:40369–72
15. Kuerschner L, Moessinger C, Thiele C. 2008. Imaging of lipid biosynthesis: How a neutral lipid enterslipid droplets. Traffic 9:338–52
16. Stone SJ, Levin MC, Zhou P, Han J, Walther TC, Farese RV Jr. 2009. The endoplasmic reticulumenzyme DGAT2 is found in mitochondria-associated membranes and has a mitochondrial targetingsignal that promotes its association with mitochondria. J. Biol. Chem. 284:5352–61
17. Blaner WS, O’Byrne SM, Wongsiriroj N, Kluwe J, D’Ambrosio DM, et al. 2009. Hepatic stellate celllipid droplets: a specialized lipid droplet for retinoid storage. Biochim. Biophys. Acta 1791:467–73
18. Orban T, Palczewska G, Palczewski K. 2011. Retinyl ester storage particles (retinosomes) from theretinal pigmented epithelium resemble lipid droplets in other tissues. J. Biol. Chem. 286:17248–58
19. Bartz R, Li WH, Venables B, Zehmer JK, Roth MR, et al. 2007. Lipidomics reveals that adiposomesstore ether lipids and mediate phospholipid traffic. J. Lipid Res. 48:837–47
20. Cornish K, Wood DF, Windle JJ. 1999. Rubber particles from four different species, examined by trans-mission electron microscopy and electron-paramagnetic-resonance spin labeling, are found to consist ofa homogeneous rubber core enclosed by a contiguous, monolayer biomembrane. Planta 210:85–96
21. Athenstaedt K, Zweytick D, Jandrositz A, Kohlwein S, Daum G. 1999. Identification and characterizationof major lipid particle proteins of the yeast Saccharomyces cerevisiae. J. Bacteriol. 181:6441–48
22. Welte MA. 2007. Proteins under new management: Lipid droplets deliver. Trends Cell Biol 17:363–6923. Cermelli S, Guo Y, Gross SP, Welte MA. 2006. The lipid-droplet proteome reveals that droplets are a
protein-storage depot. Curr. Biol. 16:1783–9524. Herker E, Ott M. 2011. Unique ties between hepatitis C virus replication and intracellular lipids. Trends
Endocrinol. Metab. 22:241–4825. Herker E, Harris C, Hernandez C, Carpentier A, Kaehlcke K, et al. 2010. Efficient hepatitis C virus
particle formation requires diacylglycerol acyltransferase-1. Nat. Med. 16:1295–9826. Ye J, Li JZ, Liu Y, Li X, Yang T, et al. 2009. Cideb, an ER- and lipid droplet-associated protein, mediates
VLDL lipidation and maturation by interacting with apolipoprotein B. Cell Metab. 9:177–9027. Dolinsky VW, Gilham D, Alam M, Vance DE, Lehner R. 2004. Triacylglycerol hydrolase: role in
intracellular lipid metabolism. Cell. Mol. Life Sci. 61:1633–5128. Waltermann M, Hinz A, Robenek H, Troyer D, Reichelt R, et al. 2005. Mechanism of lipid-body
formation in prokaryotes: How bacteria fatten up. Mol. Microbiol. 55:750–6329. Jacquier N, Choudhary V, Mari M, Toulmay A, Reggiori F, Schneiter R. 2011. Lipid droplets are
functionally connected to the endoplasmic reticulum in Saccharomyces cerevisiae. J. Cell Sci. 124:2424–3730. Soni K, Mardones G, Sougrat R, Smirnova E, Jackson C, Bonifacino J. 2009. Coatomer-dependent
protein delivery to lipid droplets. J. Cell Sci. 122:1834–4131. Ohsaki Y, Cheng J, Suzuki M, Fujita A, Fujimoto T. 2008. Lipid droplets are arrested in the ER membrane
by tight binding of lipidated apolipoprotein B-100. J. Cell Sci. 121:2415–2232. Lacey DJ, Beaudoin F, Dempsey CE, Shewry PR, Napier JA. 1999. The accumulation of triacylglycerols
within the endoplasmic reticulum of developing seeds of Helianthus annuus. Plant J. 17:397–40533. Skinner JR, Shew TM, Schwartz DM, Tzekov A, Lepus CM, et al. 2009. Diacylglycerol enrichment of
endoplasmic reticulum or lipid droplets recruits perilipin 3/TIP47 during lipid storage and mobilization.J. Biol. Chem. 284:30941–48
34. Bulankina AV, Deggerich A, Wenzel D, Mutenda K, Wittmann JG, et al. 2009. TIP47 functions in thebiogenesis of lipid droplets. J. Cell Biol. 185:641–55
35. Ploegh HL. 2007. A lipid-based model for the creation of an escape hatch from the endoplasmic reticu-lum. Nature 448:435–38
36. Walther TC, Farese RV Jr. 2009. The life of lipid droplets. Biochim. Biophys. Acta 1791:459–6637. Robenek H, Hofnagel O, Buers I, Robenek MJ, Troyer D, Severs NJ. 2006. Adipophilin-enriched
domains in the ER membrane are sites of lipid droplet biogenesis. J. Cell Sci. 119:4215–2438. Hamilton RL, Wong JS, Cham CM, Nielsen LB, Young SG. 1998. Chylomicron-sized lipid particles
are formed in the setting of apolipoprotein B deficiency. J. Lipid Res. 39:1543–5739. Fei W, Shui G, Zhang Y, Krahmer N, Ferguson C, et al. 2011. A role for phosphatidic acid in the
formation “supersized” lipid droplets. PLoS Genet. 7:e1002201
708 Walther · Farese
Ann
u. R
ev. B
ioch
em. 2
012.
81:6
87-7
14. D
ownl
oade
d fro
m w
ww
.ann
ualre
view
s.org
by H
arva
rd U
nive
rsity
on
10/2
8/14
. For
per
sona
l use
onl
y.

BI81CH28-WaltherFarese ARI 3 May 2012 14:22
40. Binns D, Lee S, Hilton CL, Jiang QX, Goodman JM. 2010. Seipin is a discrete homooligomer.Biochemistry 49:10747–55
41. Guo Y, Walther TC, Rao M, Stuurman N, Goshima G, et al. 2008. Functional genomic screen revealsgenes involved in lipid-droplet formation and utilization. Nature 453:657–61
42. Beller M, Riedel D, Jansch L, Dieterich G, Wehland J, et al. 2006. Characterization of the Drosophilalipid droplet subproteome. Mol. Cell. Proteomics 5:1082–94
43. Beller M, Sztalryd C, Southall N, Bell M, Jackle H, et al. 2008. COPI complex is a regulator of lipidhomeostasis. PLoS Biol. 6:e292
44. Ashrafi K, Chang FY, Watts JL, Fraser AG, Kamath RS, et al. 2003. Genome-wide RNAi analysis ofCaenorhabditis elegans fat regulatory genes. Nature 421:268–72
45. Pospisilik JA, Schramek D, Schnidar H, Cronin SJ, Nehme NT, et al. 2010. Drosophila genome-wideobesity screen reveals hedgehog as a determinant of brown versus white adipose cell fate. Cell 140:148–60
46. Gross DA, Snapp EL, Silver DL. 2010. Structural insights into triglyceride storage mediated by fatstorage-inducing transmembrane (FIT) protein 2. PLoS ONE 5:e10796
47. Kadereit B, Kumar P, Wang WJ, Mirnada D, Snapp EL, et al. 2008. Evolutionarily conserved genefamily important for fat storage. Proc. Natl. Acad. Sci. USA 105:94–99
48. Gubern A, Barcelo-Torns M, Casas J, Barneda D, Masgrau R, et al. 2009. Lipid droplet biogenesisinduced by stress involves triacylglycerol synthesis that depends on group VIA phospholipase A2. J. Biol.Chem. 284:5697–708
49. Moessinger C, Kuerschner L, Spandl J, Shevchenko A, Thiele C. 2011. Human lysophosphatidylcholineacyltransferases 1 and 2 are located in lipid droplets where they catalyze the formation of phosphatidyl-choline. J. Biol. Chem. 286:21330–39
50. Khelef N, Buton X, Beatini N, Wang H, Meiner V, et al. 1998. Immunolocalization of acyl-coenzymeA:cholesterol O-acyltransferase in macrophages. J. Biol. Chem. 273:11218–24
51. Bostrom P, Rutberg M, Ericsson J, Holmdahl P, Andersson L, et al. 2005. Cytosolic lipid droplets increasein size by microtubule-dependent complex formation. Arterioscler. Thromb. Vasc. Biol. 25:1945–51
52. Murphy S, Martin S, Parton RG. 2010. Quantitative analysis of lipid droplet fusion: inefficient steadystate fusion but rapid stimulation by chemical fusogens. PloS One 5:e15030
53. Puri V, Konda S, Ranjit S, Aouadi M, Chawla A, et al. 2007. Fat-specific protein 27, a novel lipid dropletprotein that enhances triglyceride storage. J. Biol. Chem. 282:34213–18
54. Keller P, Petrie JT, De Rose P, Gerin I, Wright WS, et al. 2008. Fat-specific protein 27 regulates storageof triacylglycerol. J. Biol. Chem. 283:14355–65
55. Nishino N, Tamori Y, Tateya S, Kawaguchi T, Shibakusa T, et al. 2008. FSP27 contributes to efficientenergy storage in murine white adipocytes by promoting the formation of unilocular lipid droplets.J. Clin. Investig. 118:2808–21
56. Puri V, Virbasius JV, Guilherme A, Czech MP. 2008. RNAi screens reveal novel metabolic regulators:RIP140, MAP4k4 and the lipid droplet associated fat specific protein (FSP) 27. Acta Physiol. 192:103–15
57. Jambunathan S, Yin J, Khan W, Tamori Y, Puri V. 2011. FSP27 promotes lipid droplet clustering andthen fusion to regulate triglyceride accumulation. PLOS One 6:e28614
58. Gong J, Sun Z, Wu L, Xu W, Schieber N, et al. 2011. Fsp27 promotes lipid droplet growth by lipidexchange and transfer at lipid droplet contact sites. J. Cell Biol. 195:953–63
59. Bostrom P, Andersson L, Rutberg M, Perman J, Lidberg U, et al. 2007. SNARE proteins mediate fusionbetween cytosolic lipid droplets and are implicated in insulin sensitivity. Nat. Cell Biol. 9:1286–93
60. Zweytick D, Athenstaedt K, Daum G. 2000. Intracellular lipid particles of eukaryotic cells. Biochim.Biophys. Acta 1469:101–20
61. Liu P, Ying Y, Zhao Y, Mundy DI, Zhu M, Anderson RG. 2004. Chinese hamster ovary K2 cell lipiddroplets appear to be metabolic organelles involved in membrane traffic. J. Biol. Chem. 279:3787–92
62. Fujimoto Y, Itabe H, Sakai J, Makita M, Noda J, et al. 2004. Identification of major proteins in the lipiddroplet-enriched fraction isolated from the human hepatocyte cell line HuH7. Biochim. Biophys. Acta1644:47–59
63. Umlauf E, Csaszar E, Moertelmaier M, Schuetz GJ, Parton RG, Prohaska R. 2004. Association ofstomatin with lipid bodies. J. Biol. Chem. 279:23699–709
www.annualreviews.org • Lipid Droplets and Cellular Lipid Metabolism 709
Ann
u. R
ev. B
ioch
em. 2
012.
81:6
87-7
14. D
ownl
oade
d fro
m w
ww
.ann
ualre
view
s.org
by H
arva
rd U
nive
rsity
on
10/2
8/14
. For
per
sona
l use
onl
y.

BI81CH28-WaltherFarese ARI 3 May 2012 14:22
64. Wu CC, Howell KE, Neville MC, Yates JR III, McManaman JL. 2000. Proteomics reveal a link betweenthe endoplasmic reticulum and lipid secretory mechanisms in mammary epithelial cells. Electrophoresis21:3470–82
65. Brasaemle DL, Dolios G, Shapiro L, Wang R. 2004. Proteomic analysis of proteins associated with lipiddroplets of basal and lipolytically stimulated 3T3-L1 adipocytes. J. Biol. Chem. 279:46835–42
66. Cole NB, Murphy DD, Grider T, Rueter S, Brasaemle D, Nussbaum RL. 2002. Lipid droplet bindingand oligomerization properties of the Parkinson’s disease protein α-synuclein. J. Biol. Chem. 277:6344–52
67. Barba G, Harper F, Harada T, Kohara M, Goulinet S, et al. 1997. Hepatitis C virus core protein showsa cytoplasmic localization and associates to cellular lipid storage droplets. Proc. Natl. Acad. Sci. USA94:1200–5
68. Hinson ER, Cresswell P. 2009. The antiviral protein, viperin, localizes to lipid droplets via its N-terminalamphipathic alpha-helix. Proc. Natl. Acad. Sci. USA 106:20452–57
69. Tauchi-Sato K, Ozeki S, Houjou T, Tagushi R, Fujimoto T. 2002. The surface of lipid droplets is aphospholipid monolayer with a unique fatty acid composition. J. Biol. Chem. 277:44507–12
70. Zehmer JK, Bartz R, Liu P, Anderson RG. 2008. Identification of a novel N-terminal hydrophobicsequence that targets proteins to lipid droplets. J. Cell Sci. 121:1852–60
71. Turro S, Ingelmo-Torres M, Estanyol JM, Tebar F, Fernandez MA, et al. 2006. Identification andcharacterization of associated with lipid droplet protein 1: A novel membrane-associated protein thatresides on hepatic lipid droplets. Traffic 7:1254–69
72. Napier JA, Stobart AK, Shewry PR. 1996. The structure and biogenesis of plant oil bodies: the role ofthe ER membrane and the oleosin class of proteins. Plant Mol. Biol. 31:945–56
73. Qu R, Wang S-M, Lin Y-H, Vance VB, Huang AHC. 1986. Characteristics and biosynthesis of mem-brane proteins of lipid bodies in the scutella of maize (Zea mays L.). Biochem. J. 235:57–65
74. Abell BM, Holbrook LA, Abenes M, Murphy DJ, Hills MJ, Moloney MM. 1997. Role of the prolineknot motif in oleosin endoplasmic reticulum topology and oil body targeting. Plant Cell 9:1481–93
75. Pol A, Luetterforst R, Lindsay M, Heinoo S, Ikonen E, Parton RG. 2001. A caveolin dominant negativemutant associates with lipid bodies and induces intracellular cholesterol imbalance. J. Cell Biol. 152:1057–70
76. Fujimoto T, Kogo H, Ishiguro K, Tauchi K, Nomura R. 2001. Caveolin-2 is targeted to lipid droplets,a new “membrane domain” in the cell. J. Cell Biol. 152:1079–85
77. Ostermeyer A, Paci J, Zeng Y, Lublin D, Munro S, Brown D. 2001. Accumulation of caveolin in theendoplasmic reticulum redirects the protein to lipid storage droplets. J. Cell Biol. 152:1071–78
78. Pol A, Martin S, Fernandez MA, Ingelmo-Torres M, Ferguson C, et al. 2005. Cholesterol and fatty acidsregulate dynamic caveolin trafficking through the Golgi complex and between the cell surface and lipidbodies. Mol. Biol. Cell 16:2091–105
79. Le Lay S, Hajduch E, Lindsay MR, Le Liepvre X, Thiele C, et al. 2006. Cholesterol-induced caveolintargeting to lipid droplets in adipocytes: a role for caveolar endocytosis. Traffic 7:549–61
80. Greenberg AS, Egan JJ, Wek SA, Garty NB, Blanchette-Mackie EJ, Londos C. 1991. Perilipin, a majorhormonally regulated adipocyte-specific phosphoprotein associated with the periphery of lipid storagedroplets. J. Biol. Chem. 266:11341–46
81. Subramanian V, Garcia A, Sekowski A, Brasaemle DL. 2004. Hydrophobic sequences target and anchorperilipin A to lipid droplets. J. Lipid Res. 45:1983–91
82. McManaman JL, Zabaronick W, Schaack J, Orlicky DJ. 2003. Lipid droplet targeting domains ofadipophilin. J. Lipid Res. 44:668–73
83. Targett-Adams P, Chambers D, Gledhill S, Hope RG, Coy JF, et al. 2003. Live cell analysis and targetingof the lipid droplet-binding adipocyte differentiation-related protein. J. Biol. Chem. 278:15998–6007
84. Nakamura N, Fujimoto T. 2003. Adipose differentiation-related protein has two independent domainsfor targeting to lipid droplets. Biochem. Biophys. Res. Commun. 306:333–38
85. Hickenbottom SJ, Kimmel AR, Londos C, Hurley JH. 2004. Structure of a lipid droplet protein: thePAT family member TIP47. Structure 12:1199–207
86. Hatters DM, Peters-Libeu CA, Weisgraber KH. 2006. Apolipoprotein E structure: insights into function.Trends Biochem. Sci. 31:445–54
710 Walther · Farese
Ann
u. R
ev. B
ioch
em. 2
012.
81:6
87-7
14. D
ownl
oade
d fro
m w
ww
.ann
ualre
view
s.org
by H
arva
rd U
nive
rsity
on
10/2
8/14
. For
per
sona
l use
onl
y.

BI81CH28-WaltherFarese ARI 3 May 2012 14:22
87. Wolins NE, Quaynor BK, Skinner JR, Schoenfish MJ, Tzekov A, Bickel PE. 2005. S3–12, adipophilin,and TIP47 package lipid in adipocytes. J. Biol. Chem. 280:19146–55
88. Ozeki S, Cheng J, Tauchi-Sato K, Hatano N, Taniguchi H, Fujimoto T. 2005. Rab18 localizes to lipiddroplets and induces their close apposition to the endoplasmic reticulum-derived membrane. J. Cell Sci.118:2601–11
89. Martin S, Driessen K, Nixon SJ, Zerial M, Parton RG. 2005. Regulated localization of Rab18 tolipid droplets: effects of lipolytic stimulation and inhibition of lipid droplet catabolism. J. Biol. Chem.280:42325–35
90. Beller M, Bulankina AV, Hsiao HH, Urlaub H, Jackle H, Kuhnlein RP. 2010. PERILIPIN-dependentcontrol of lipid droplet structure and fat storage in Drosophila. Cell Metab. 12:521–32
91. Schweiger M, Schoiswohl G, Lass A, Radner FP, Haemmerle G, et al. 2008. The C-terminal regionof human adipose triglyceride lipase affects enzyme activity and lipid droplet binding. J. Biol. Chem.283:17211–20
92. Wanner G, Formanet H, Theimer RR. 1981. The ontogeny of lipid bodies (spherosomes) in plant cells.Planta 151:109–23
93. Singh R, Kaushik S, Wang Y, Xiang Y, Novak I, et al. 2009. Autophagy regulates lipid metabolism.Nature 458:1131–35
94. Xu G, Sztalryd C, Lu X, Tansey JT, Gan J, et al. 2005. Post-translational regulation of adiposedifferentiation-related protein by the ubiquitin/proteasome pathway. J. Biol. Chem. 280:42841–47
95. Masuda Y, Itabe H, Odaki M, Hama K, Fujimoto Y, et al. 2006. ADRP/adipophilin is degraded throughthe proteasome-dependent pathway during regression of lipid-storing cells. J. Lipid Res. 47:87–98
96. Xu G, Sztalryd C, Londos C. 2006. Degradation of perilipin is mediated through ubiquitination-proteasome pathway. Biochim. Biophys. Acta 1761:83–90
97. Hooper C, Puttamadappa SS, Loring Z, Shekhtman A, Bakowska JC. 2010. Spartin activates atrophin-1-interacting protein 4 (AIP4) E3 ubiquitin ligase and promotes ubiquitination of adipophilin on lipiddroplets. BMC Biol. 8:72
98. Nian Z, Sun Z, Yu L, Toh SY, Sang J, Li P. 2010. Fat-specific protein 27 undergoes ubiquitin-dependentdegradation regulated by triacylglycerol synthesis and lipid droplet formation. J. Biol. Chem. 285:9604–15
99. Edwards TL, Clowes VE, Tsang HT, Connell JW, Sanderson CM, et al. 2009. Endogenous spartin(SPG20) is recruited to endosomes and lipid droplets and interacts with the ubiquitin E3 ligases AIP4and AIP5. Biochem. J. 423:31–39
100. Eastman SW, Yassaee M, Bieniasz PD. 2009. A role for ubiquitin ligases and Spartin/SPG20 in lipiddroplet turnover. J. Cell Biol. 184:881–94
101. Spandl J, Lohmann D, Kuerschner L, Moessinger C, Thiele C. 2011. Ancient ubiquitous protein 1(AUP1) localizes to lipid droplets and binds the E2 ubiquitin conjugase G2 (Ube2g2) via its G2 bindingregion. J. Biol. Chem. 286:5599–606
102. Klemm EJ, Spooner E, Ploegh HL. 2011. The dual role of ancient ubiquitous protein 1 (AUP1) in lipiddroplet accumulation and ER protein quality control. J. Biol. Chem. 286:37602–14
103. Hartman IZ, Liu P, Zehmer JK, Luby-Phelps K, Jo Y, et al. 2010. Sterol-induced dislocation of 3-hydroxy-3-methylglutaryl coenzyme A reductase from endoplasmic reticulum membranes into the cy-tosol through a subcellular compartment resembling lipid droplets. J. Biol. Chem. 285:19288–98
104. Olzmann JA, Kopito RR. 2011. Lipid droplet formation is dispensable for endoplasmic reticulum-associated degradation. J. Biol. Chem. 286:27872–74
105. Hynynen R, Suchanek M, Spandl J, Back N, Thiele C, Olkkonen VM. 2009. OSBP-related protein 2 is asterol receptor on lipid droplets that regulates the metabolism of neutral lipids. J. Lipid Res. 50:1305–15
106. Yokoi Y, Horiguchi Y, Araki M, Motojima K. 2007. Regulated expression by PPARα and unique lo-calization of 17β-hydroxysteroid dehydrogenase type 11 protein in mouse intestine and liver. FEBS J.274:4837–47
107. Ouimet M, Franklin V, Mak E, Liao X, Tabas I, Marcel YL. 2011. Autophagy regulates cholesterol effluxfrom macrophage foam cells via lysosomal acid lipase. Cell Metab. 13:655–67
108. Liu P, Bartz R, Zehmer JK, Ying YS, Zhu M, et al. 2007. Rab-regulated interaction of early endosomeswith lipid droplets. Biochim. Biophys. Acta 1773:784–93
www.annualreviews.org • Lipid Droplets and Cellular Lipid Metabolism 711
Ann
u. R
ev. B
ioch
em. 2
012.
81:6
87-7
14. D
ownl
oade
d fro
m w
ww
.ann
ualre
view
s.org
by H
arva
rd U
nive
rsity
on
10/2
8/14
. For
per
sona
l use
onl
y.

BI81CH28-WaltherFarese ARI 3 May 2012 14:22
109. Blanchette-Mackie EJ, Scow RO. 1983. Movement of lipolytic products to mitochondria in brownadipose tissue of young rats: an electron microscope study. J. Lipid Res. 24:229–44
110. Binns D, Januszewski T, Chen Y, Hill J, Markin VS, et al. 2006. An intimate collaboration betweenperoxisomes and lipid bodies. J. Cell Biol. 173:719–31
111. Tarnopolsky MA, Rennie CD, Robertshaw HA, Fedak-Tarnopolsky SN, Devries MC, Hamadeh MJ.2007. Influence of endurance exercise training and sex on intramyocellular lipid and mitochondrialultrastructure, substrate use, and mitochondrial enzyme activity. Am. J. Physiol. Regul. Integr. Comp.Physiol. 292:R1271–78
112. Marcinkiewicz A, Gauthier D, Garcia A, Brasaemle DL. 2006. The phosphorylation of serine 492 ofperilipin A directs lipid droplet fragmentation and dispersion. J. Biol. Chem. 281:11901–9
113. Savage MJ, Goldberg DJ, Schacher S. 1987. Absolute specificity for retrograde fast axonal transportdisplayed by lipid droplets originating in the axon of an identified Aplysia neuron in vitro. Brain Res.406:215–23
114. Welte MA, Gross SP, Postner M, Block SM, Wieschaus EF. 1998. Developmental regulation of vesicletransport in Drosophila embryos: forces and kinetics. Cell 92:547–57
115. Shubeita GT, Tran SL, Xu J, Vershinin M, Cermelli S, et al. 2008. Consequences of motor copy numberon the intracellular transport of kinesin-1-driven lipid droplets. Cell 135:1098–107
116. Boulant S, Douglas MW, Moody L, Budkowska A, Targett-Adams P, McLauchlan J. 2008. Hepatitis Cvirus core protein induces lipid droplet redistribution in a microtubule- and dynein-dependent manner.Traffic 9:1268–82
117. Gruber A, Cornaciu I, Lass A, Schweiger M, Poeschl M, et al. 2010. The N-terminal region of com-parative gene identification-58 (CGI-58) is important for lipid droplet binding and activation of adiposetriglyceride lipase. J. Biol. Chem. 285:12289–98
118. Yang X, Lu X, Lombes M, Rha GB, Chi Y-I, et al. 2010. The G0/G1 switch gene 2 regulates adiposelipolysis through association with adipose triglyceride lipase. Cell Metab. 11:194–205
119. Brasaemle DL, Rubin B, Harten IA, Gruia-Gray J, Kimmel AR, Londos C. 2000. Perilipin A increasestriacylglycerol storage by decreasing the rate of triacylglycerol hydrolysis. J. Biol. Chem. 275:38486–93
120. Souza SC, de Vargas LM, Yamamoto MT, Lien P, Franciosa MD, et al. 1998. Overexpression of perilipinA and B blocks the ability of tumor necrosis factor α to increase lipolysis in 3T3-L1 adipocytes. J. Biol.Chem. 273:24665–69
121. Subramanian V, Rothenberg A, Gomez C, Cohen AW, Garcia A, et al. 2004. Perilipin A mediates thereversible binding of CGI-58 to lipid droplets in 3T3-L1 adipocytes. J. Biol. Chem. 279:42062–71
122. Granneman JG, Moore HP, Mottillo EP, Zhu Z, Zhou L. 2011. Interactions of perilipin-5 (Plin5) withadipose triglyceride lipase. J. Biol. Chem. 286:5126–35
123. Wang H, Bell M, Sreenevasan U, Hu H, Liu J, et al. 2011. Unique regulation of adipose triglyceridelipase (ATGL) by perilipin 5, a lipid droplet-associated protein. J. Biol. Chem. 286:15707–15
124. Krintel C, Osmark P, Larsen MR, Resjo S, Logan DT, Holm C. 2008. Ser649 and Ser650 are the majordeterminants of protein kinase A-mediated activation of human hormone-sensitive lipase against lipidsubstrates. PLoS ONE 3:e3756
125. Wang H, Hu L, Dalen K, Dorward H, Marcinkiewicz A, et al. 2009. Activation of hormone-sensitivelipase requires two steps, protein phosphorylation and binding to the PAT-1 domain of lipid dropletcoat proteins. J. Biol. Chem. 284:32116–25
126. Schweiger M, Schreiber R, Haemmerle G, Lass A, Fledelius C, et al. 2006. Adipose triglyceride lipaseand hormone-sensitive lipase are the major enzymes in adipose tissue triacylglycerol catabolism. J. Biol.Chem. 281:40236–41
127. Taschler U, Radner FP, Heier C, Schreiber R, Schweiger M, et al. 2011. Monoglyceride lipase deficiencyin mice impairs lipolysis and attenuates diet-induced insulin resistance. J. Biol. Chem. 286:17467–77
128. He S, McPhaul C, Li JZ, Garuti R, Kinch L, et al. 2010. A sequence variation (I148M) in PNPLA3associated with nonalcoholic fatty liver disease disrupts triglyceride hydrolysis. J. Biol. Chem. 285:6706–15
129. Chen W, Chang B, Li L, Chan L. 2010. Patatin-like phospholipase domain-containing 3/adiponutrindeficiency in mice is not associated with fatty liver disease. Hepatology 52:1134–42
130. Basantani MK, Sitnick MT, Cai L, Brenner DS, Gardner NP, et al. 2011. Pnpla3/adiponutrin deficiencyin mice does not contribute to fatty liver disease or metabolic syndrome. J. Lipid Res. 52:318–29
712 Walther · Farese
Ann
u. R
ev. B
ioch
em. 2
012.
81:6
87-7
14. D
ownl
oade
d fro
m w
ww
.ann
ualre
view
s.org
by H
arva
rd U
nive
rsity
on
10/2
8/14
. For
per
sona
l use
onl
y.

BI81CH28-WaltherFarese ARI 3 May 2012 14:22
131. Kaczocha M, Glaser ST, Chae J, Brown DA, Deutsch DG. 2010. Lipid droplets are novel sites ofN-acylethanolamine inactivation by fatty acid amide hydrolase-2. J. Biol. Chem. 285:2796–806
132. Leibel RL, Hirsch J, Berry EM, Gruen RK. 1985. Alterations in adipocyte free fatty acid re-esterificationassociated with obesity and weight reduction in man. Am. J. Clin. Nutr. 42:198–206
133. Fujimoto Y, Itabe H, Kinoshita T, Homma KJ, Onoduka J, et al. 2007. Involvement of ACSL in localsynthesis of neutral lipids in cytoplasmic lipid droplets in human hepatocyte HuH7. J. Lipid Res. 48:1280–92
134. Wolkart G, Schrammel A, Dorffel K, Haemmerle G, Zechner R, Mayer B. 2011. Cardiac dysfunction inadipose triglyceride lipase deficiency: Treatment with a PPARα agonist. Br. J. Pharmacol. 165:380–89
135. Greenberg AS, Coleman RA, Kraemer FB, McManaman JL, Obin MS, et al. 2011. The role of lipiddroplets in metabolic disease in rodents and humans. J. Clin. Investig. 121:2102–10
136. Gesta S, Tseng YH, Kahn CR. 2007. Developmental origin of fat: tracking obesity to its source. Cell131:242–56
137. Friedman JM. 2009. Leptin at 14 y of age: an ongoing story. Am. J. Clin. Nutr. 89:S973–79138. Zhu J, Lee B, Buhman KK, Cheng JX. 2009. A dynamic, cytoplasmic triacylglycerol pool in enterocytes
revealed by ex vivo and in vivo coherent anti-Stokes Raman scattering imaging. J. Lipid Res. 50:1080–89139. Farese RV Jr, Cases S, Ruland SL, Kayden HJ, Wong JS, et al. 1996. A novel function for apolipoprotein
B: Lipoprotein synthesis in the yolk sac is critical for maternal-fetal lipid transport in mice. J. Lipid Res.37:347–60
140. Liu L, Zhang Y, Chen N, Shi X, Tsang B, Yu YH. 2007. Upregulation of myocellular DGAT1 aug-ments triglyceride synthesis in skeletal muscle and protects against fat-induced insulin resistance. J. Clin.Investig. 117:1679–89
141. Schenk S, Horowitz JF. 2007. Acute exercise increases triglyceride synthesis in skeletal muscle andprevents fatty acid-induced insulin resistance. J. Clin. Investig. 117:1690–98
142. Shaw CS, Jones DA, Wagenmakers AJM. 2008. Network distribution of mitochondria and lipid dropletsin human muscle fibres. Histochem. Cell Biol. 129:65–72
143. Chen HC, Stone SJ, Zhou P, Buhman KK, Farese RV Jr. 2002. Dissociation of obesity and impairedglucose disposal in mice overexpressing acyl coenzyme A:diacylglycerol acyltransferase 1 in white adiposetissue. Diabetes 51:3189–95
144. Franckhauser S, Munoz S, Pujol A, Casellas A, Riu E, et al. 2002. Increased fatty acid re-esterification byPEPCK overexpression in adipose tissue leads to obesity without insulin resistance. Diabetes 51:624–30
145. Koliwad SK, Streeper RS, Monetti M, Cornelissen I, Chan L, et al. 2010. DGAT1-dependent triacyl-glycerol storage by macrophages protects mice from diet-induced insulin resistance and inflammation.J. Clin. Investig. 120:756–67
146. Nagle CA, Klett EL, Coleman RA. 2009. Hepatic triacylglycerol accumulation and insulin resistance.J. Lipid Res. 50(Suppl.):S74–79
147. Accad M, Smith SJ, Newland DL, Sanan DA, King LE Jr, et al. 2000. Massive xanthomatosis and alteredcomposition of atherosclerotic lesions in hyperlipidemic mice lacking acyl CoA:cholesterol acyltrans-ferase 1. J. Clin. Invest 105:711–19
148. Chiu H-C, Kovacs A, Ford DA, Hsu F-F, Garcia R, et al. 2001. A novel mouse model of lipotoxiccardiomyopathy. J. Clin. Investig. 107:813–22
149. Liu L, Shi X, Bharadwaj KG, Ikeda S, Yamashita H, et al. 2009. DGAT1 expression increases hearttriglyceride content but ameliorates lipotoxicity. J. Biol. Chem. 284:36312–23
150. Haemmerle G, Lass A, Zimmermann R, Gorkiewicz G, Meyer C, et al. 2006. Defective lipolysis andaltered energy metabolism in mice lacking adipose triglyceride lipase. Science 312:734–37
151. Garg A, Agarwal AK. 2009. Lipodystrophies: disorders of adipose tissue biology. Biochim. Biophys. Acta1791:507–13
152. McManaman JL, Russell TD, Schaack J, Orlicky DJ, Robenek H. 2007. Molecular determinants of milklipid secretion. J. Mammary Gland Biol. Neoplasia 12:259–68
153. Vorbach C, Scriven A, Capecchi MR. 2002. The housekeeping gene xanthine oxidoreductase is necessaryfor milk fat droplet enveloping and secretion: gene sharing in the lactating mammary gland. Genes Dev.16:3223–35
www.annualreviews.org • Lipid Droplets and Cellular Lipid Metabolism 713
Ann
u. R
ev. B
ioch
em. 2
012.
81:6
87-7
14. D
ownl
oade
d fro
m w
ww
.ann
ualre
view
s.org
by H
arva
rd U
nive
rsity
on
10/2
8/14
. For
per
sona
l use
onl
y.

BI81CH28-WaltherFarese ARI 3 May 2012 14:22
154. Ogg SL, Weldon AK, Dobbie L, Smith AJ, Mather IH. 2004. Expression of butyrophilin (Btn1a1) inlactating mammary gland is essential for the regulated secretion of milk-lipid droplets. Proc. Natl. Acad.Sci. USA 101:10084–89
155. Russell TD, Palmer CA, Orlicky DJ, Bales ES, Chang BH, et al. 2008. Mammary glands of adipophilin-null mice produce an amino-terminally truncated form of adipophilin that mediates milk lipid dropletformation and secretion. J. Lipid Res. 49:206–16
156. Smith SJ, Cases S, Jensen DR, Chen HC, Sande E, et al. 2000. Obesity resistance and multiple mecha-nisms of triglyceride synthesis in mice lacking DGAT. Nat. Genet. 25:87–90
157. Cases S, Zhou P, Schillingford J, Wiseman B, Fish J, et al. 2004. Development of the mammary glandrequires DGAT1 expression in stromal and epithelial tissues. Development 131:3047–55
158. Saha S, Enugutti B, Rajakumari S, Rajasekharan R. 2006. Cytosolic triacylglycerol biosynthetic pathwayin oilseeds. Molecular cloning and expression of peanut cytosolic diacylglycerol acyltransferase. PlantPhysiol. 141:1533–43
159. Zheng P, Allen WB, Roesler K, Williams ME, Zhang S, et al. 2008. A phenylalanine in DGAT is a keydeterminant of oil content and composition in maize. Nat. Genet. 40:367–72
160. Baud S, Lepiniec L. 2010. Physiological and developmental regulation of seed oil production. Prog. LipidRes. 49:235–49
161. Andrianov V, Borisjuk N, Pogrebnyak N, Brinker A, Dixon J, et al. 2010. Tobacco as a productionplatform for biofuel: Overexpression of Arabidopsis DGAT and LEC2 genes increases accumulation andshifts the composition of lipids in green biomass. Plant Biotechnol. J. 8:277–87
162. Eastmond PJ. 2006. SUGAR-DEPENDENT1 encodes a patatin domain triacylglycerol lipase that initiatesstorage oil breakdown in germinating Arabidopsis seeds. Plant Cell 18:665–75
163. Graham IA. 2008. Seed storage oil mobilization. Annu. Rev. Plant. Biol. 59:115–42164. Hayashi Y, Hayashi M, Hayashi H, Hara-Nishimura I, Nishimura M. 2001. Direct interaction between
glyoxysomes and lipid bodies in cotyledons of the Arabidopsis thaliana ped1 mutant. Protoplasma 218:83–94165. Schmidt MA, Herman EM. 2008. Suppression of soybean oleosin produces micro-oil bodies that aggre-
gate into oil body/ER complexes. Mol. Plant 1:910–24166. Toomre D, Bewersdorf J. 2010. A new wave of cellular imaging. Annu. Rev. Cell Dev. Biol. 26:285–314
714 Walther · Farese
Ann
u. R
ev. B
ioch
em. 2
012.
81:6
87-7
14. D
ownl
oade
d fro
m w
ww
.ann
ualre
view
s.org
by H
arva
rd U
nive
rsity
on
10/2
8/14
. For
per
sona
l use
onl
y.

BI81-FrontMatter ARI 9 May 2012 7:26
Annual Review ofBiochemistry
Volume 81, 2012Contents
PrefacePreface and Dedication to Christian R.H. Raetz
JoAnne Stubbe ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣xi
Prefatories
A Mitochondrial OdysseyWalter Neupert ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ 1
The Fires of LifeGottfried Schatz ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣34
Chromatin, Epigenetics, and Transcription Theme
Introduction to Theme “Chromatin, Epigenetics, and Transcription”Joan W. Conaway ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣61
The COMPASS Family of Histone H3K4 Methylases:Mechanisms of Regulation in Developmentand Disease PathogenesisAli Shilatifard ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣65
Programming of DNA Methylation PatternsHoward Cedar and Yehudit Bergman ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣97
RNA Polymerase II Elongation ControlQiang Zhou, Tiandao Li, and David H. Price ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ 119
Genome Regulation by Long Noncoding RNAsJohn L. Rinn and Howard Y. Chang ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ 145
Protein Tagging Theme
The Ubiquitin System, an Immense RealmAlexander Varshavsky ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ 167
Ubiquitin and Proteasomes in TranscriptionFuqiang Geng, Sabine Wenzel, and William P. Tansey ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ 177
v
Ann
u. R
ev. B
ioch
em. 2
012.
81:6
87-7
14. D
ownl
oade
d fro
m w
ww
.ann
ualre
view
s.org
by H
arva
rd U
nive
rsity
on
10/2
8/14
. For
per
sona
l use
onl
y.

BI81-FrontMatter ARI 9 May 2012 7:26
The Ubiquitin CodeDavid Komander and Michael Rape ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ 203
Ubiquitin and Membrane Protein Turnover: From Cradle to GraveJason A. MacGurn, Pi-Chiang Hsu, and Scott D. Emr ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ 231
The N-End Rule PathwayTakafumi Tasaki, Shashikanth M. Sriram, Kyong Soo Park, and Yong Tae Kwon ♣ ♣ ♣ 261
Ubiquitin-Binding Proteins: Decoders of Ubiquitin-MediatedCellular FunctionsKoraljka Husnjak and Ivan Dikic ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ 291
Ubiquitin-Like ProteinsAnnemarthe G. van der Veen and Hidde L. Ploegh ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ 323
Recent Advances in Biochemistry
Toward the Single-Hour High-Quality GenomePatrik L. Stahl and Joakim Lundeberg ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ 359
Mass Spectrometry–Based Proteomics and Network BiologyAriel Bensimon, Albert J.R. Heck, and Ruedi Aebersold ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ 379
Membrane Fission: The Biogenesis of Transport CarriersFelix Campelo and Vivek Malhotra ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ 407
Emerging Paradigms for Complex Iron-Sulfur Cofactor Assemblyand InsertionJohn W. Peters and Joan B. Broderick ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ 429
Structural Perspective of Peptidoglycan Biosynthesis and AssemblyAndrew L. Lovering, Susan S. Safadi, and Natalie C.J. Strynadka ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ 451
Discovery, Biosynthesis, and Engineering of LantipeptidesPatrick J. Knerr and Wilfred A. van der Donk ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ 479
Regulation of Glucose Transporter Translocationin Health and DiabetesJonathan S. Bogan ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ 507
Structure and Regulation of Soluble Guanylate CyclaseEmily R. Derbyshire and Michael A. Marletta ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ 533
The MPS1 Family of Protein KinasesXuedong Liu and Mark Winey ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ 561
The Structural Basis for Control of Eukaryotic Protein KinasesJane A. Endicott, Martin E.M. Noble, and Louise N. Johnson ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ 587
vi Contents
Ann
u. R
ev. B
ioch
em. 2
012.
81:6
87-7
14. D
ownl
oade
d fro
m w
ww
.ann
ualre
view
s.org
by H
arva
rd U
nive
rsity
on
10/2
8/14
. For
per
sona
l use
onl
y.

BI81-FrontMatter ARI 9 May 2012 7:26
Measurements and Implications of the Membrane Dipole PotentialLiguo Wang ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ 615
GTPase Networks in Membrane TrafficEmi Mizuno-Yamasaki, Felix Rivera-Molina, and Peter Novick ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ 637
Roles for Actin Assembly in EndocytosisOlivia L. Mooren, Brian J. Galletta, and John A. Cooper ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ 661
Lipid Droplets and Cellular Lipid MetabolismTobias C. Walther and Robert V. Farese Jr. ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ 687
Adipogenesis: From Stem Cell to AdipocyteQi Qun Tang and M. Daniel Lane ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ 715
Pluripotency and Nuclear ReprogrammingMarion Dejosez and Thomas P. Zwaka ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ 737
Endoplasmic Reticulum Stress and Type 2 DiabetesSung Hoon Back and Randal J. Kaufman ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ 767
Structure Unifies the Viral UniverseNicola G.A. Abrescia, Dennis H. Bamford, Jonathan M. Grimes,
and David I. Stuart ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ 795
Indexes
Cumulative Index of Contributing Authors, Volumes 77–81 ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ 823
Cumulative Index of Chapter Titles, Volumes 77–81 ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ 827
Errata
An online log of corrections to Annual Review of Biochemistry articles may be found athttp://biochem.annualreviews.org/errata.shtml
Contents vii
Ann
u. R
ev. B
ioch
em. 2
012.
81:6
87-7
14. D
ownl
oade
d fro
m w
ww
.ann
ualre
view
s.org
by H
arva
rd U
nive
rsity
on
10/2
8/14
. For
per
sona
l use
onl
y.

ANNUAL REVIEWSIt’s about time. Your time. It’s time well spent.
ANNUAL REVIEWS | Connect With Our ExpertsTel: 800.523.8635 (US/CAN) | Tel: 650.493.4400 | Fax: 650.424.0910 | Email: [email protected]
New From Annual Reviews:Annual Review of Statistics and Its Application=VS\TL�����6USPUL�1HU\HY`��������O[[W!��Z[H[PZ[PJZ�HUU\HSYL]PL^Z�VYN
Editor: Stephen E. Fienberg, Carnegie Mellon UniversityAssociate Editors: Nancy Reid, University of Toronto
Stephen M. Stigler, University of ChicagoThe Annual Review of Statistics and Its Application aims to inform statisticians and quantitative methodologists, as well as all scientists and users of statistics about major methodological advances and the computational tools that HSSV^�MVY�[OLPY�PTWSLTLU[H[PVU��0[�^PSS�PUJS\KL�KL]LSVWTLU[Z�PU�[OL�ÄLSK�VM�Z[H[PZ[PJZ��PUJS\KPUN�[OLVYL[PJHS�Z[H[PZ[PJHS�\UKLYWPUUPUNZ�VM�UL^�TL[OVKVSVN ��HZ�^LSS�HZ�KL]LSVWTLU[Z�PU�ZWLJPÄJ�HWWSPJH[PVU�KVTHPUZ�Z\JO�HZ�IPVZ[H[PZ[PJZ�and bioinformatics, economics, machine learning, psychology, sociology, and aspects of the physical sciences.
*VTWSPTLU[HY`�VUSPUL�HJJLZZ�[V�[OL�ÄYZ[�]VS\TL�^PSS�IL�H]HPSHISL�\U[PS�1HU\HY`�������TABLE OF CONTENTS:
��What Is Statistics? Stephen E. Fienberg��A Systematic Statistical Approach to Evaluating Evidence
from Observational Studies, David Madigan, Paul E. Stang, Jesse A. Berlin, Martijn Schuemie, J. Marc Overhage, Marc A. Suchard, Bill Dumouchel, Abraham G. Hartzema, Patrick B. Ryan
��The Role of Statistics in the Discovery of a Higgs Boson, David A. van Dyk
��Brain Imaging Analysis, F. DuBois Bowman��Statistics and Climate, Peter Guttorp��Climate Simulators and Climate Projections,
Jonathan Rougier, Michael Goldstein��Probabilistic Forecasting, Tilmann Gneiting,
Matthias Katzfuss��Bayesian Computational Tools, Christian P. Robert��Bayesian Computation Via Markov Chain Monte Carlo, 9HK\�=��*YHP\��1LɈYL`�:��9VZLU[OHS
��Build, Compute, Critique, Repeat: Data Analysis with Latent Variable Models, David M. Blei
��Structured Regularizers for High-Dimensional Problems: Statistical and Computational Issues, Martin J. Wainwright
��High-Dimensional Statistics with a View Toward Applications in Biology, Peter Bühlmann, Markus Kalisch, Lukas Meier
��Next-Generation Statistical Genetics: Modeling, Penalization, and Optimization in High-Dimensional Data, Kenneth Lange, Jeanette C. Papp, Janet S. Sinsheimer, Eric M. Sobel
��Breaking Bad: Two Decades of Life-Course Data Analysis in Criminology, Developmental Psychology, and Beyond, Elena A. Erosheva, Ross L. Matsueda, Donatello Telesca
��Event History Analysis, Niels Keiding��:[H[PZ[PJHS�,]HS\H[PVU�VM�-VYLUZPJ�+5(�7YVÄSL�,]PKLUJL��
Christopher D. Steele, David J. Balding��Using League Table Rankings in Public Policy Formation:
Statistical Issues, Harvey Goldstein��Statistical Ecology, Ruth King��Estimating the Number of Species in Microbial Diversity
Studies, John Bunge, Amy Willis, Fiona Walsh��Dynamic Treatment Regimes, Bibhas Chakraborty,
Susan A. Murphy��Statistics and Related Topics in Single-Molecule Biophysics,
Hong Qian, S.C. Kou��Statistics and Quantitative Risk Management for Banking
and Insurance, Paul Embrechts, Marius Hofert
Access this and all other Annual Reviews journals via your institution at www.annualreviews.org.
Ann
u. R
ev. B
ioch
em. 2
012.
81:6
87-7
14. D
ownl
oade
d fro
m w
ww
.ann
ualre
view
s.org
by H
arva
rd U
nive
rsity
on
10/2
8/14
. For
per
sona
l use
onl
y.