lipidy, metabolismus lipidů a biologické membrányvacha/vyuka/biochemie/06_lipidy.pdf · trans...
TRANSCRIPT

Lipidy, metabolismus
lipidů a biologické
membrány

Lipidy: strukturně rozlišné látky
• Nerozpustné ve vodě
• Dobře rozpustné v nepolárních
rozpouštědlech
Funkce:
• Zásobní látky
• Strukturní molekuly
• Signální, kofaktory enzymů, pigmenty

Klasifikace lipidů
Založená na struktuře a funkci
• Lipidy, které obsahují mastné kyseliny
– Zásobní lipidy a membránové lipidy
• Lipidy, které neobsahují mastné kyseliny
cholesterol, terpeny, karotenoidy, ...

Lipidy bez mastných kyselin
Steroidy (cholesterol), terpeny, karotenoidy
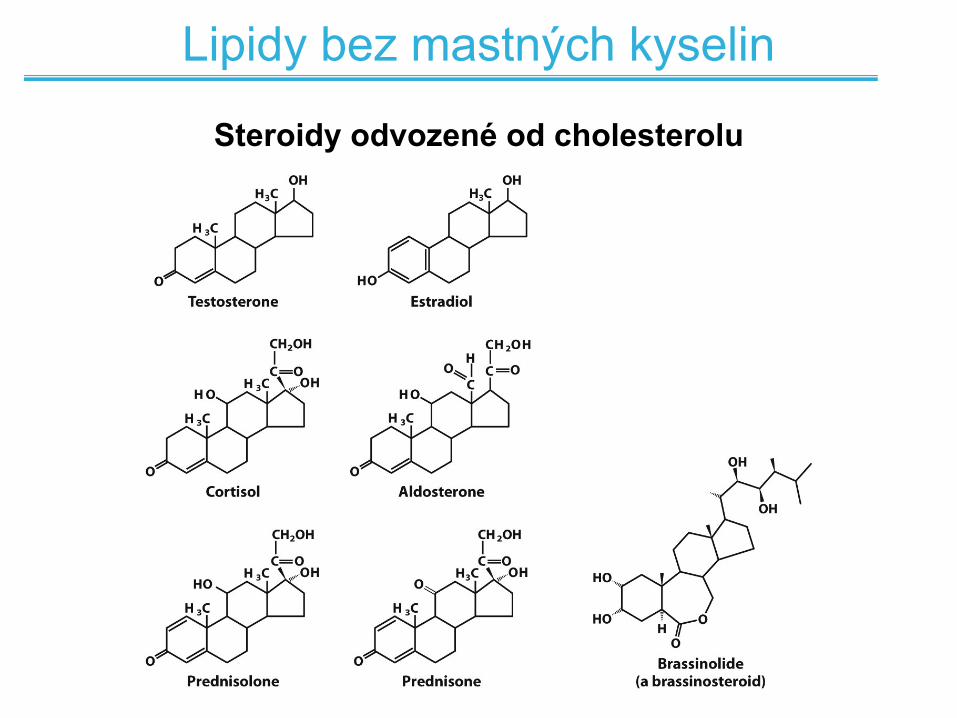
Steroidy odvozené od cholesterolu
Lipidy bez mastných kyselin
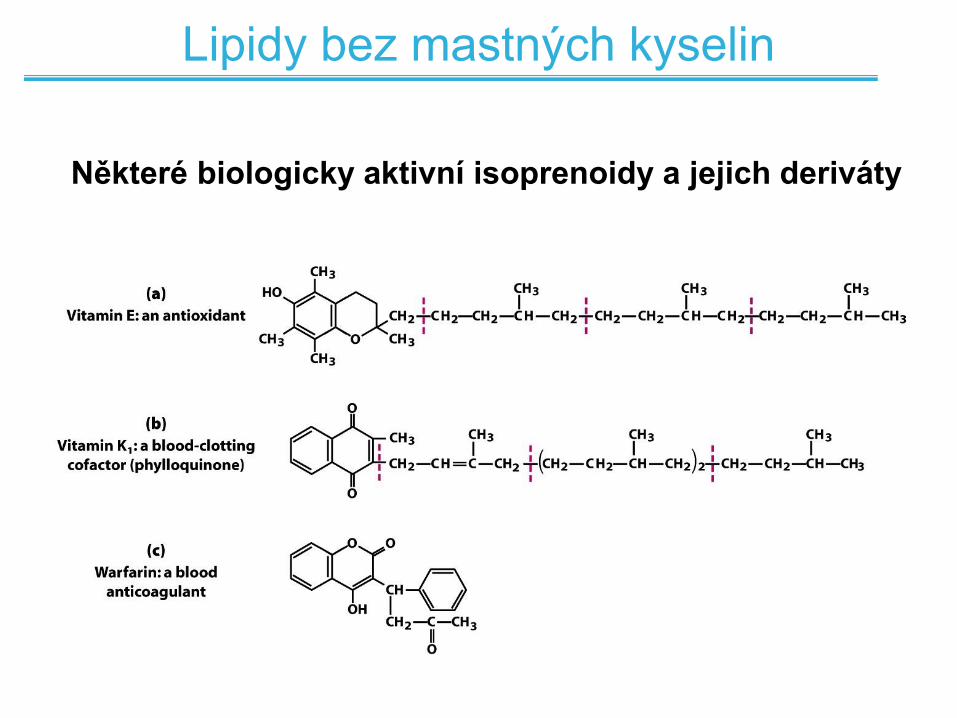
Některé biologicky aktivní isoprenoidy a jejich deriváty
Lipidy bez mastných kyselin
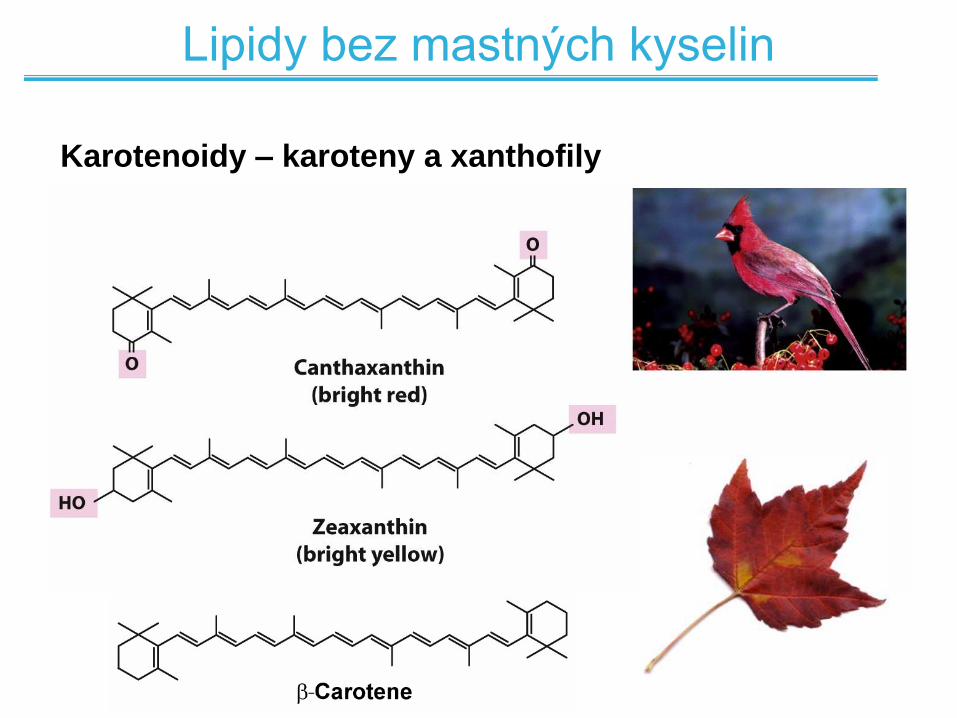
Karotenoidy – karoteny a xanthofily
Lipidy bez mastných kyselin
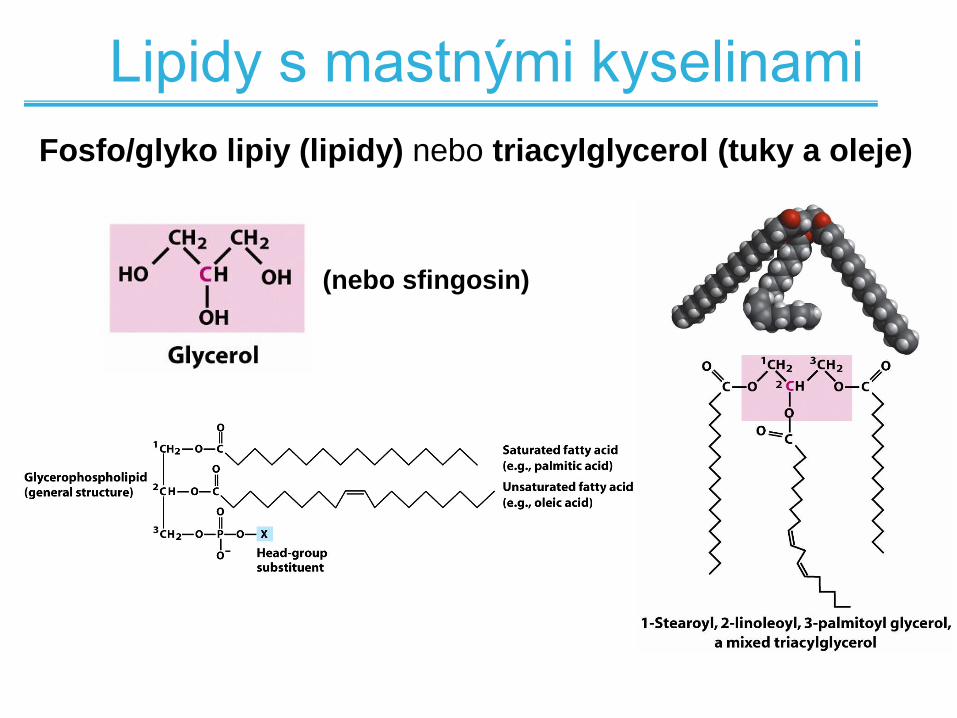
Lipidy s mastnými kyselinami
Fosfo/glyko lipiy (lipidy) nebo triacylglycerol (tuky a oleje)
(nebo sfingosin)

Mastné kyseliny
• Karboxylové kyseliny s uhlíkovým řetězcem s 4 až 36 uhlíky
• Takřka všechny přírodní mastné kyseliny mají sudý počet
uhlíků
• Většina je nevětvená
• Nasycené: neobsahují žádnou dvojnou vazvu
• (Mono)Nenasycené: jedna dvojná vazba na alkylovém
řetězci
• Polynenasycené: více dvojných vazeb na alkylovém řetězci
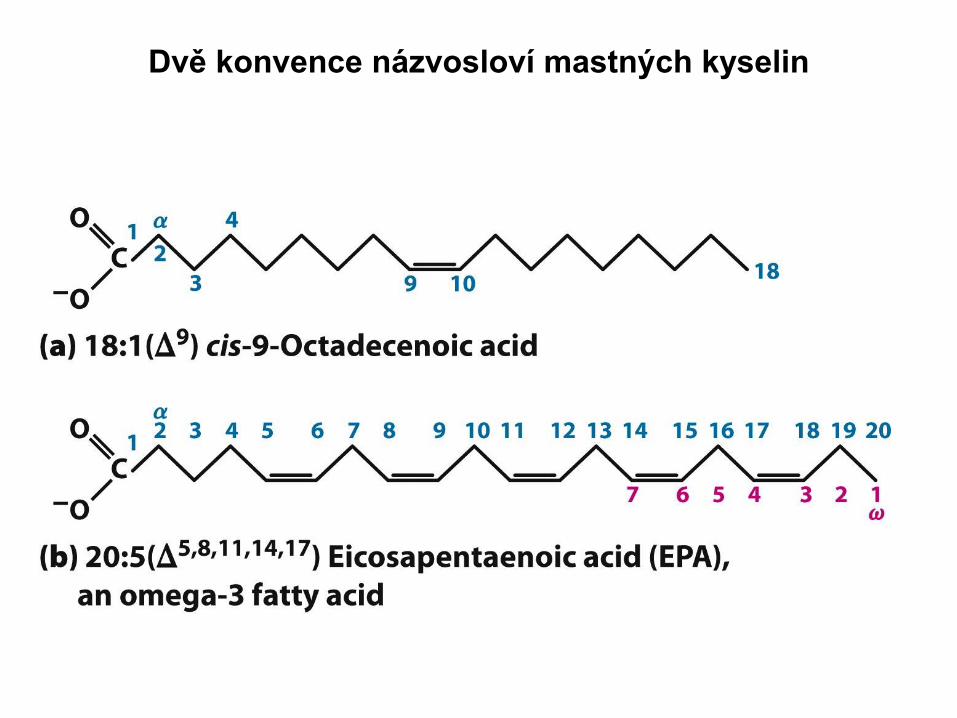
Dvě konvence názvosloví mastných kyselin
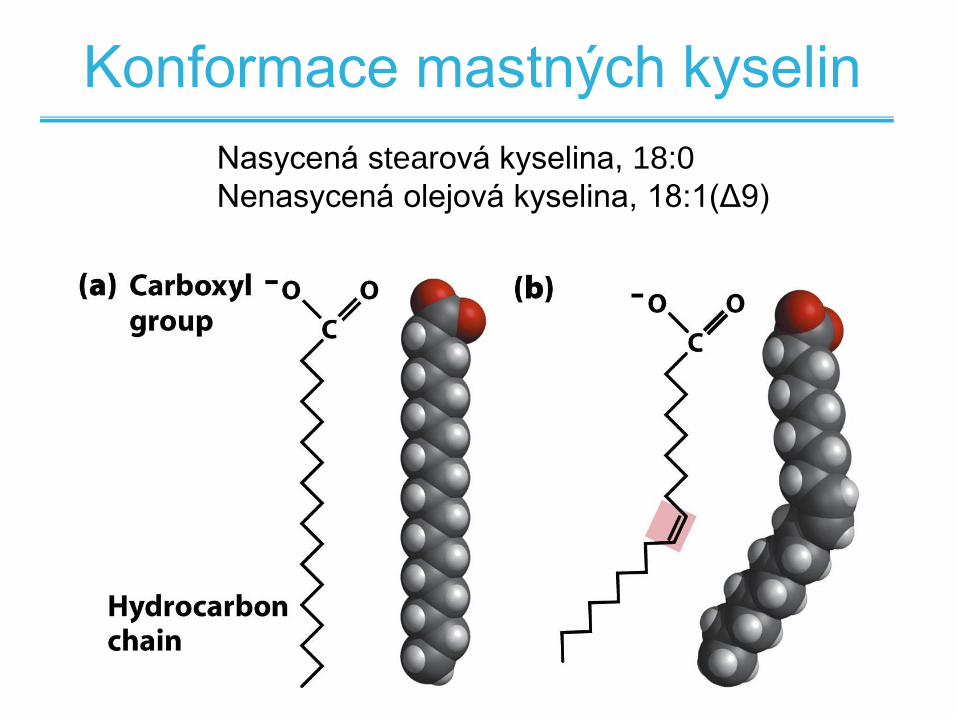
Nasycená stearová kyselina, 18:0
Nenasycená olejová kyselina, 18:1(Δ9)
Konformace mastných kyselin

Konformace mastných kyselin
• Saturované mají tendenci zaujímat přímou
konformaci
• Dvojná vazba v přírodních nenasycených
mastných kyselinách je v cis konfiguraci
• Tato konfigurace způsobuje ohyb řetězce
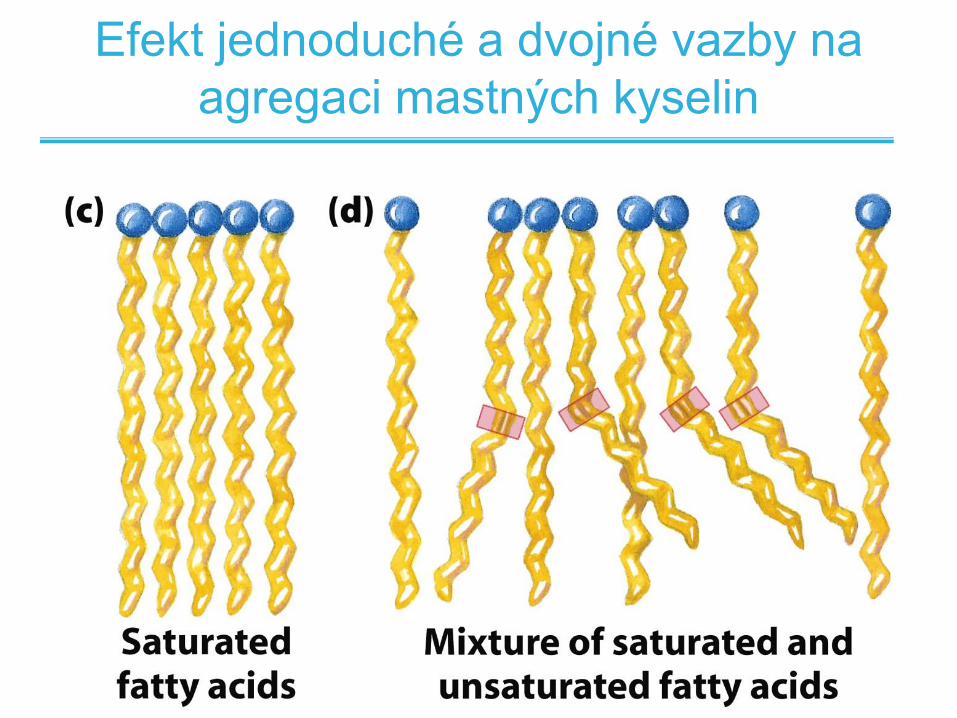
Efekt jednoduché a dvojné vazby na
agregaci mastných kyselin
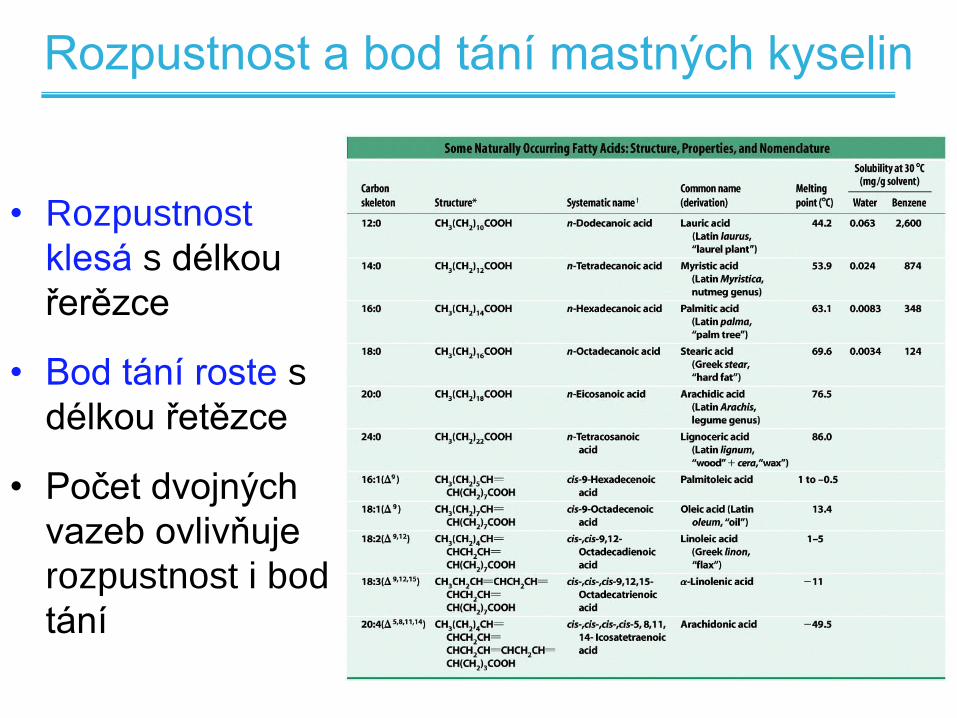
Rozpustnost a bod tání mastných kyselin
• Rozpustnost
klesá s délkou
řerězce
• Bod tání roste s
délkou řetězce
• Počet dvojných
vazeb ovlivňuje
rozpustnost i bod
tání

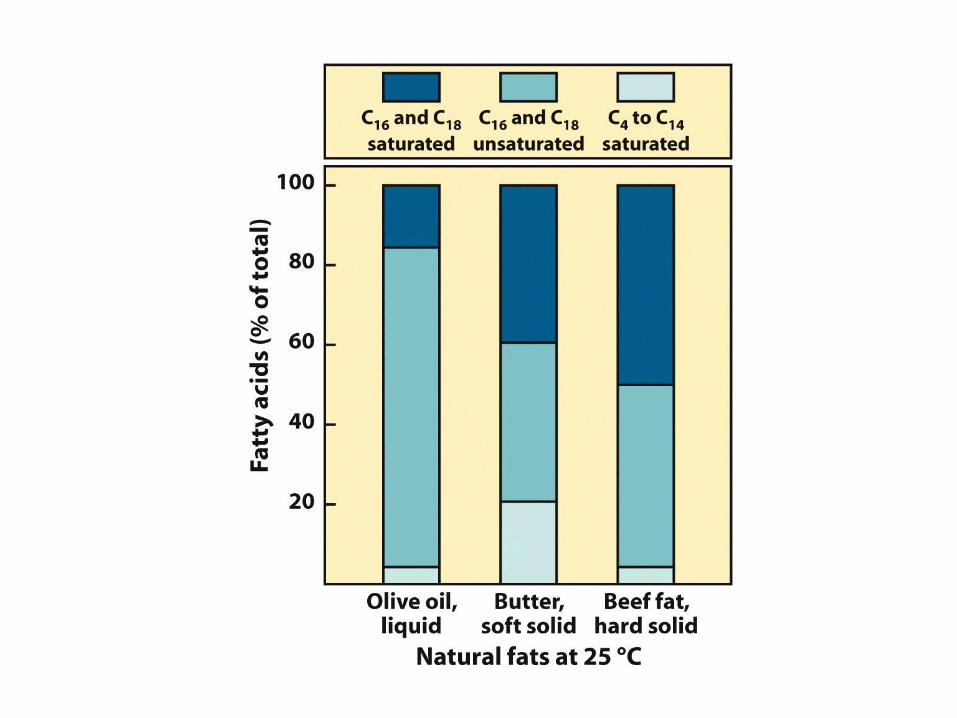
Bod tání a dvojná vazba
• Saturované mastné kyseliny se samoorientují ve
velmi organizované pravidelné struktury
• Nenasycené mastné kyseliny tvoří díky cis ohybu
méně kompaktní a méně organizované struktury
• Je potřeba méně tepelné energie k rozrušení
neorganizované struktury nenasycených mastných
kyselin: nižší bod tání
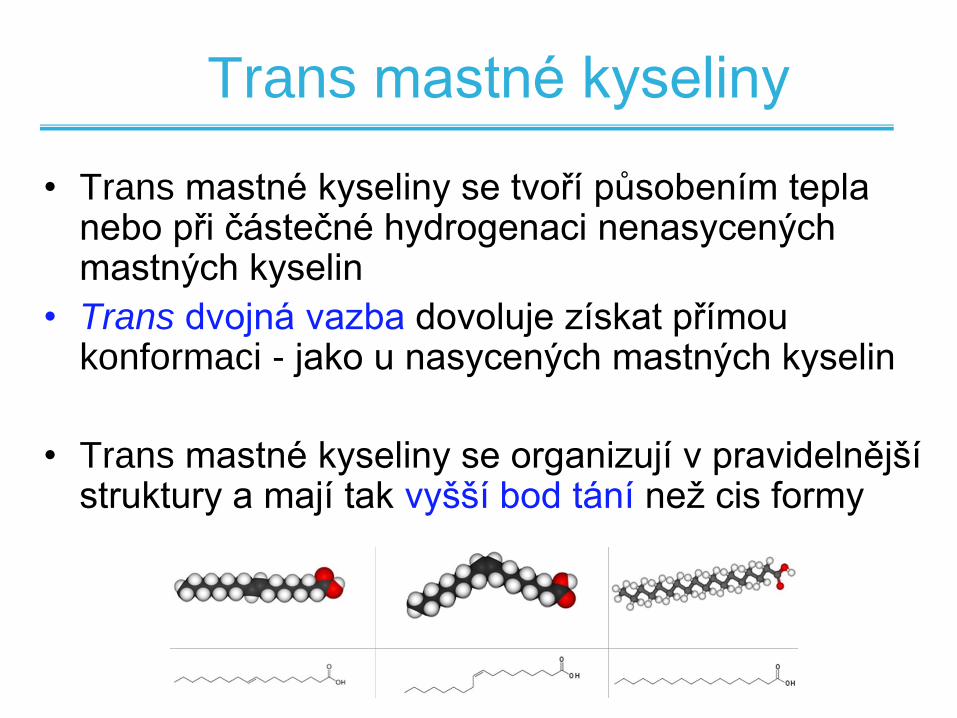
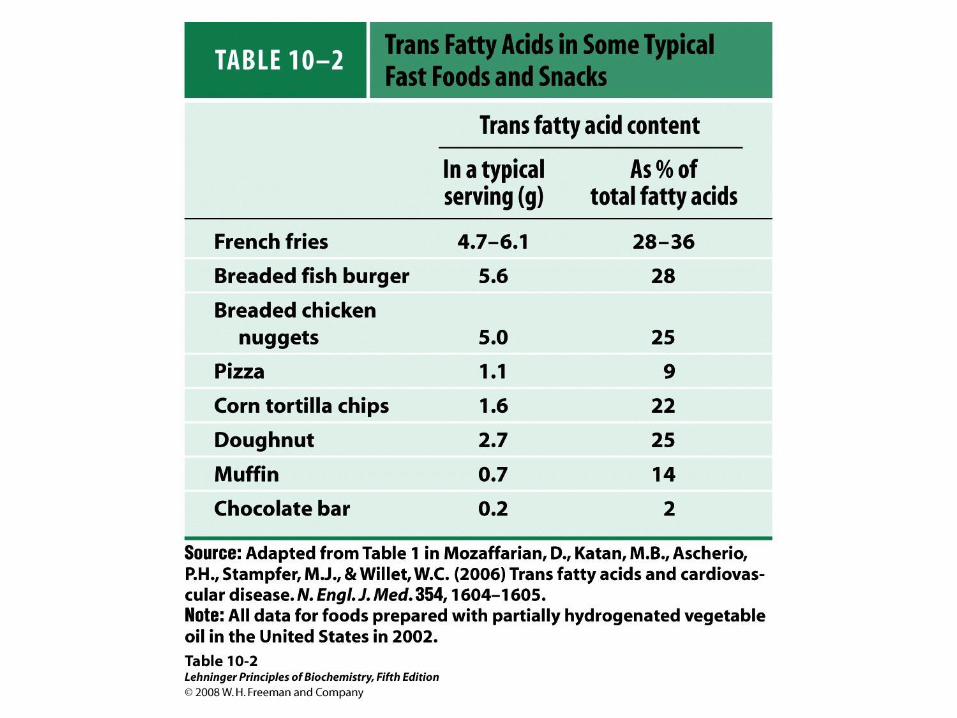
Trans mastné kyseliny
• Trans mastné kyseliny se tvoří působením tepla nebo při částečné hydrogenaci nenasycených mastných kyselin
• Trans dvojná vazba dovoluje získat přímou konformaci - jako u nasycených mastných kyselin
• Trans mastné kyseliny se organizují v pravidelnější struktury a mají tak vyšší bod tání než cis formy

• Konzumace trans mastných kyselin vede k
nebezpečí vzniku kardiovaskulárních chorob
– Vyhnout se hydrogenovaným rostlinným olejům
– Vyhnout se přepalovaným tukům
Trans mastné kyseliny
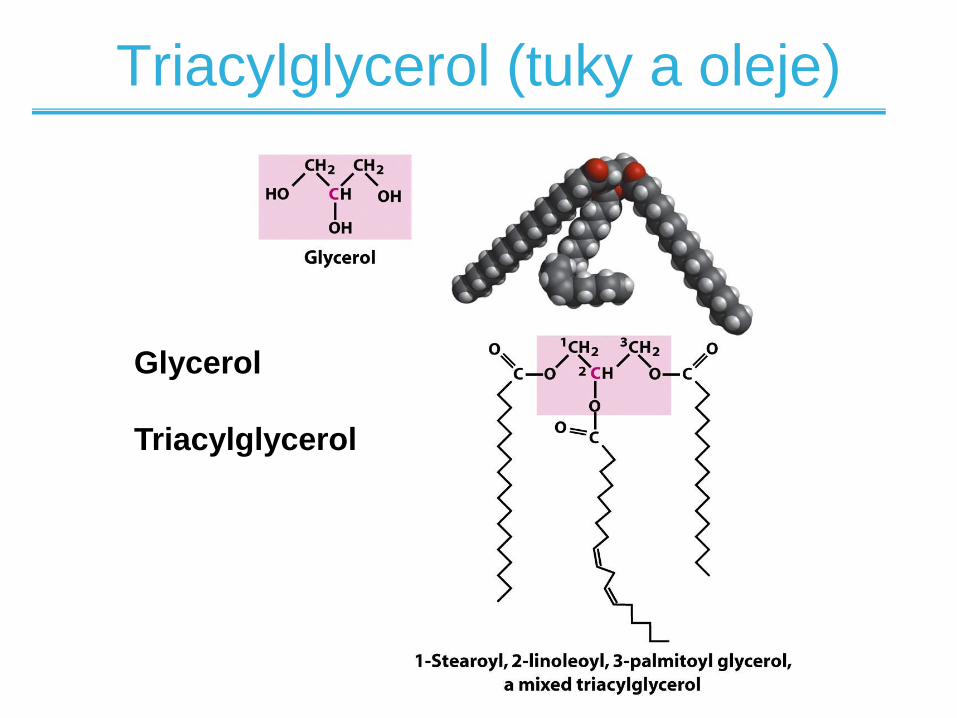
Glycerol
Triacylglycerol
Triacylglycerol (tuky a oleje)

Triacylglycerol (tuky a oleje)
• Většina mastných kyselin je v biologických
systémech vázaná ve formě triacylglycerolů
• Tuhé se nazývají tuky
• Tekuté se nazývají oleje
• Triacylglyceroly jsou primárně zásobními lipidy
(tělesný tuk)
• Triacylglyceroly jsou lehčí než voda: tuky a oleje
plavou na vodě

Tuky a oleje jako zásoba energie
• Výhoda tuků a olejů oproti polysacharidům:
– Mastné kyseliny obsahují více energie na uhlík protože
jsou více redukované
– Mastné kyseliny váží méně vody díky tomu, že jsou
nepolární
• Glukóza a glykogen jsou na rychlou a krátkodobou
energetickou potřebu – rychlá dodávka energie
• Tuky a oleje jsou na dlohodobou (měsíce) energetickou
potřebu - dobrá zásoba, pomalá dodávka
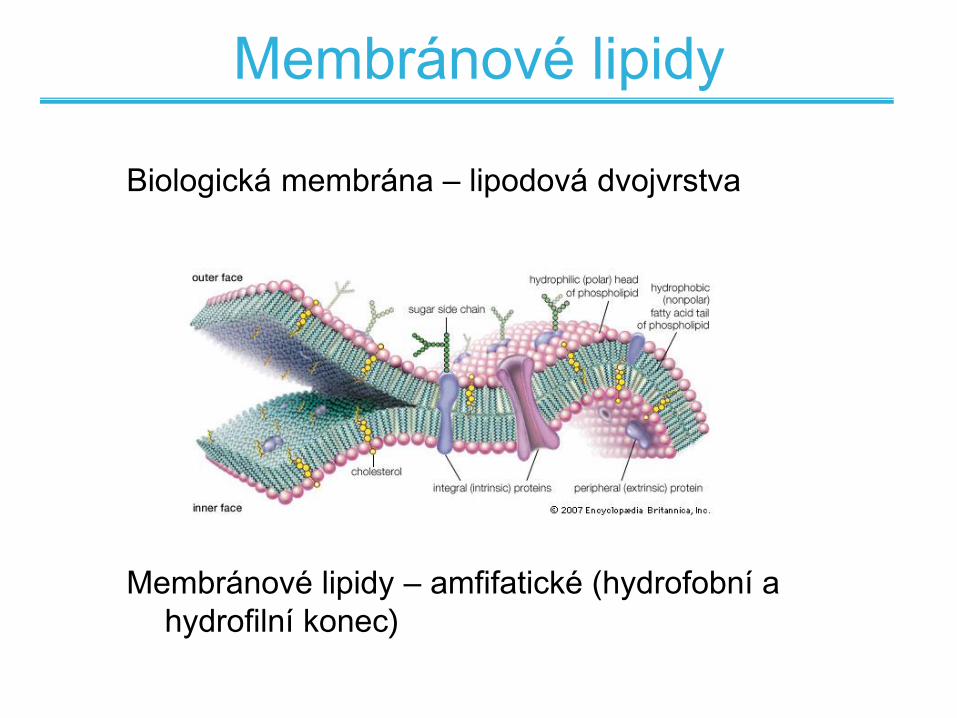
Membránové lipidy
Biologická membrána – lipodová dvojvrstva
Membránové lipidy – amfifatické (hydrofobní a
hydrofilní konec)

• Glycerolfosfolipidy – fosfolipidy
• Glycerolglykolipidy – glykolipidy, rostlinné lipidy
• Sulfolipidy – sirné glykolipidy
• Tetraetherové lipidy – membránové lipidy archaeí
• Sfingolipidy
• Steroly – další součást biologických membrán
Membránové lipidy
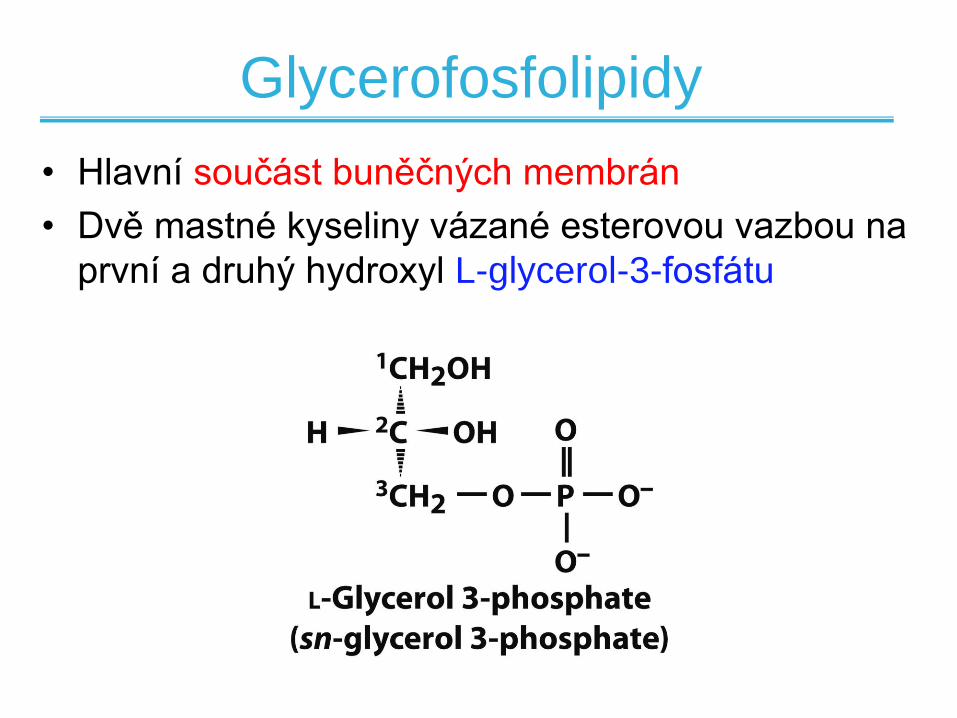
Glycerofosfolipidy
• Hlavní součást buněčných membrán
• Dvě mastné kyseliny vázané esterovou vazbou na
první a druhý hydroxyl L-glycerol-3-fosfátu
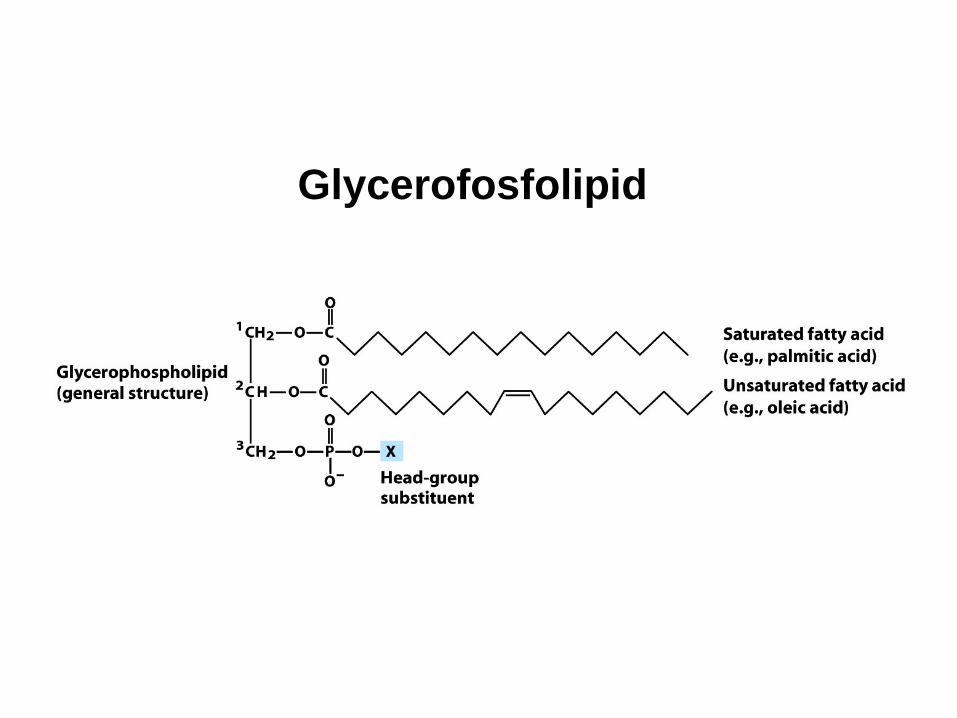
Glycerofosfolipid

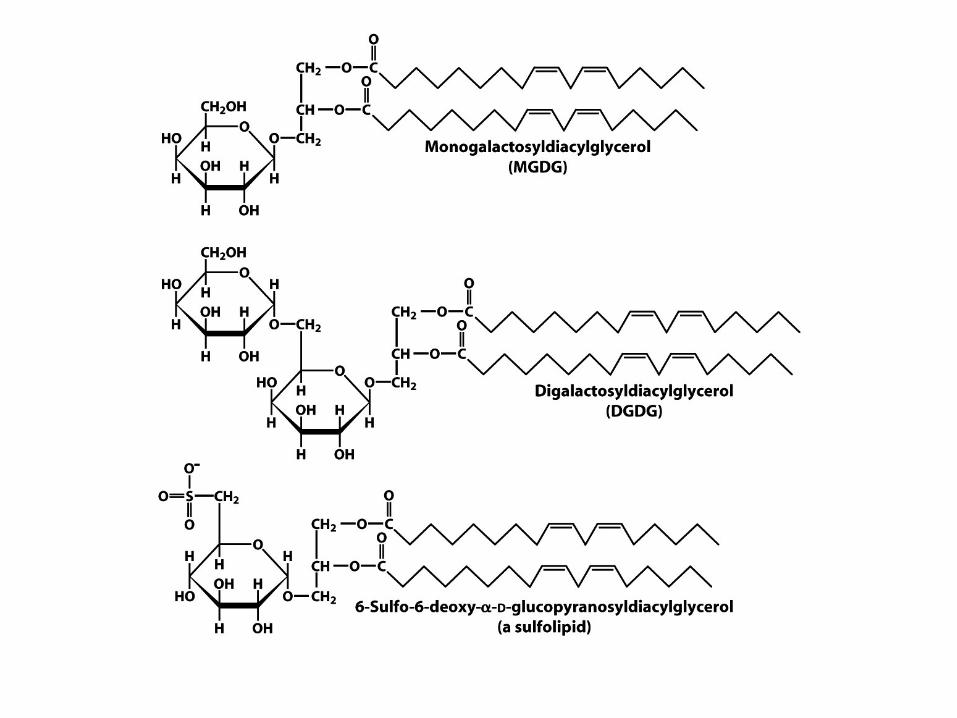
Glykolipidy a sulfolipidy
• Lipidy rostlinných organel chloroplastů
• Obsahují cukry
• Nejčastější galaktosyldiacylglycerol
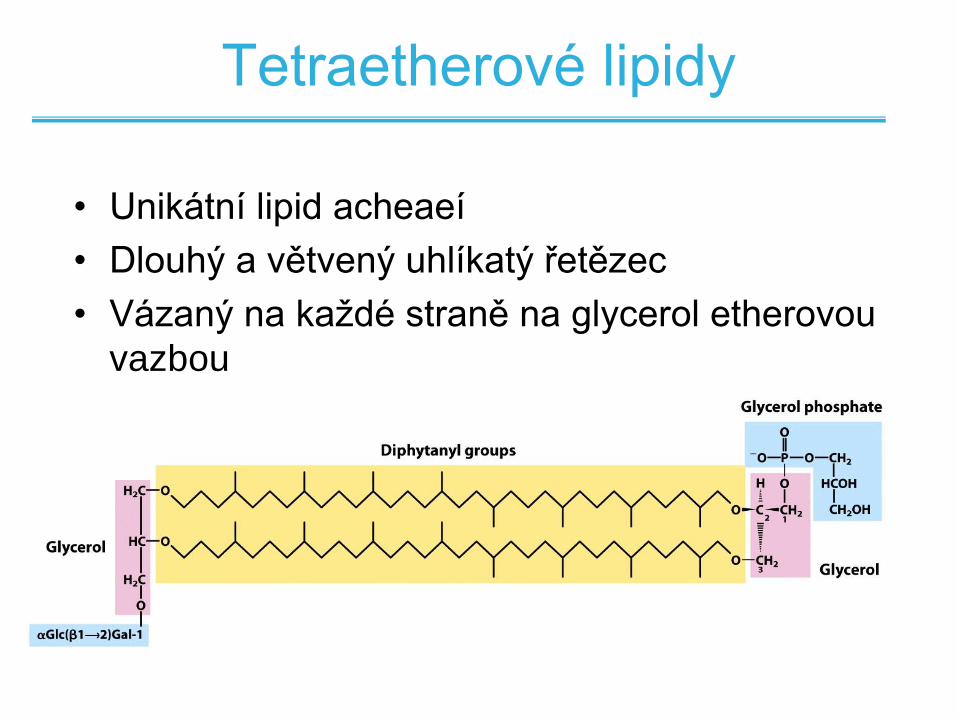
Tetraetherové lipidy
• Unikátní lipid acheaeí
• Dlouhý a větvený uhlíkatý řetězec
• Vázaný na každé straně na glycerol etherovou
vazbou

Sfingolipidy
• Zákaldní kostrou je sfingosin NE glycerol
• Sfingosin je aminoalkohol s dlouhým (18)
uhlíkatým řetězcem
• Mastné kyseliny jsou vázány ke sfingosinu
amidovou vazbou, ne esterovou jak je to u
běžných lipidů
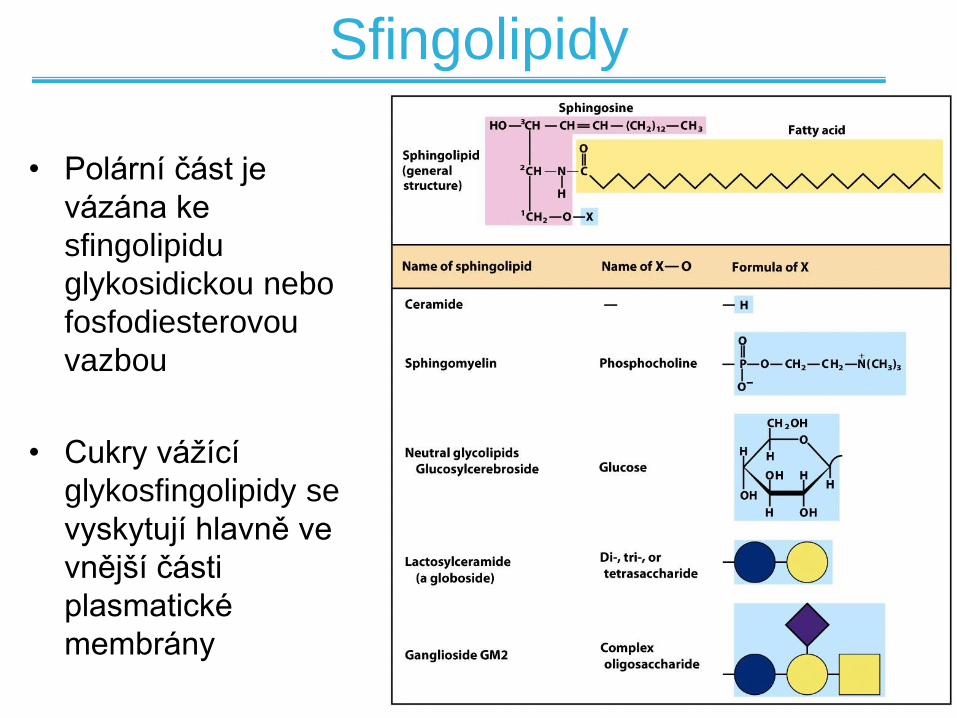
Sfingolipidy
• Polární část je
vázána ke
sfingolipidu
glykosidickou nebo
fosfodiesterovou
vazbou
• Cukry vážící
glykosfingolipidy se
vyskytují hlavně ve
vnější části
plasmatické
membrány
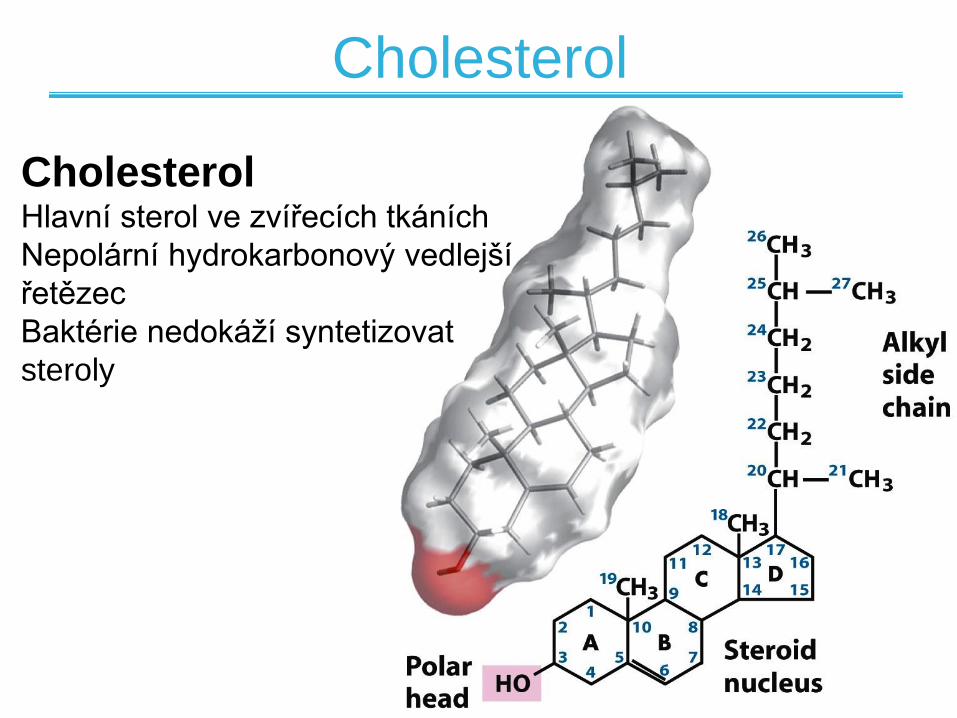
CholesterolHlavní sterol ve zvířecích tkáních
Nepolární hydrokarbonový vedlejší
řetězec
Baktérie nedokáží syntetizovat
steroly
Cholesterol


Biologické membrány a
membránový transport
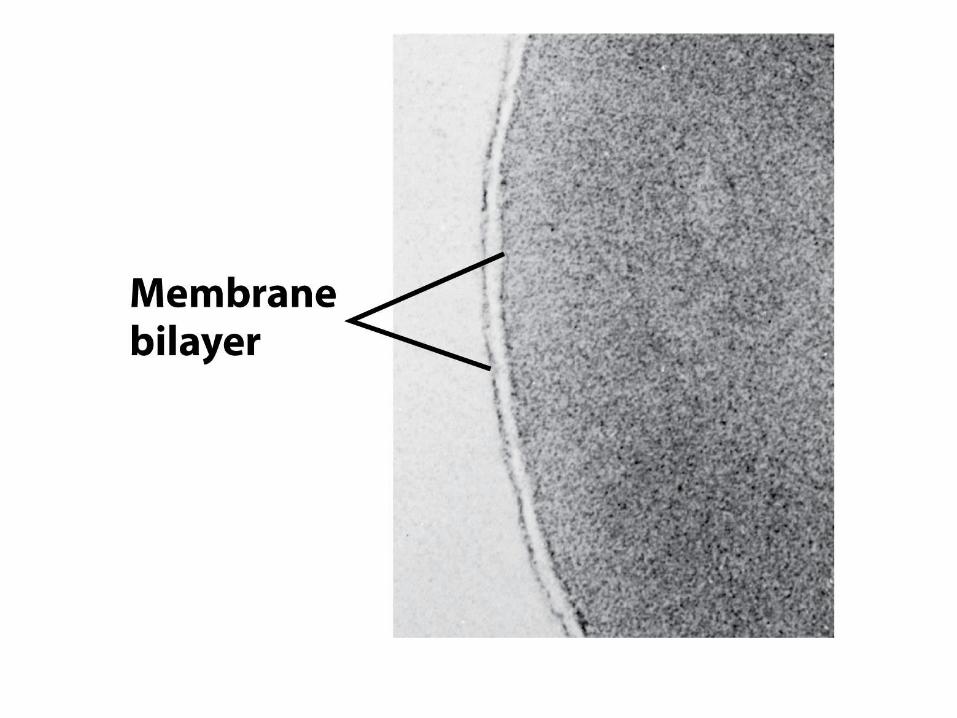
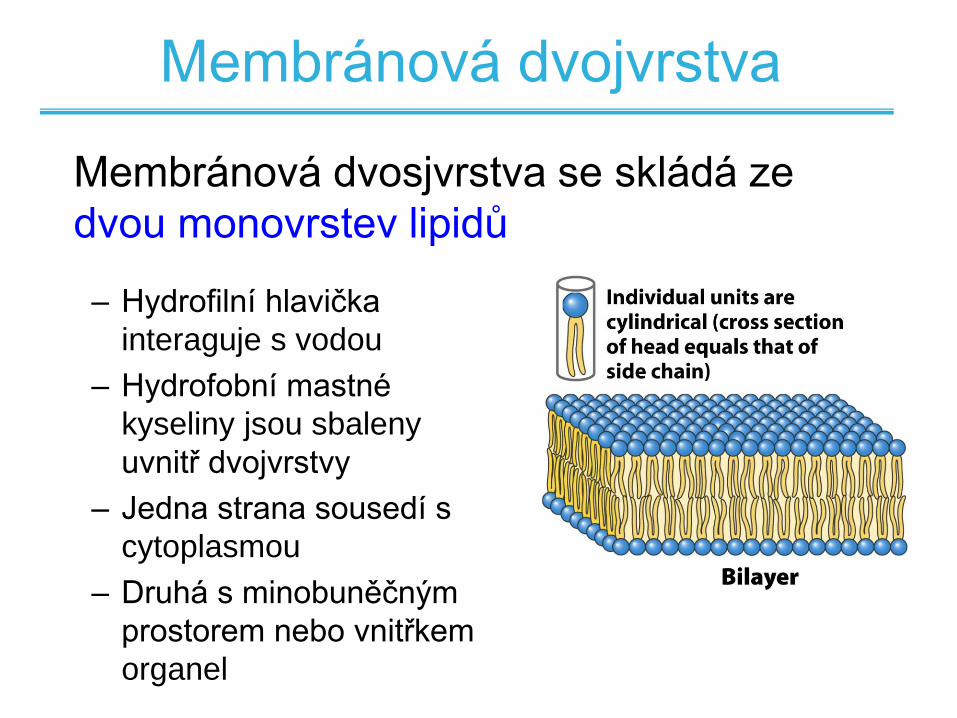
Membránová dvojvrstva
– Hydrofilní hlavička
interaguje s vodou
– Hydrofobní mastné
kyseliny jsou sbaleny
uvnitř dvojvrstvy
– Jedna strana sousedí s
cytoplasmou
– Druhá s minobuněčným
prostorem nebo vnitřkem
organel
Membránová dvosjvrstva se skládá ze
dvou monovrstev lipidů

Funkce biologické membrány
• Určuje hranici buňky nebo organely
• Umožňuje import a export látek
– Selektivní import živin …
– Selektivní export odpadu, toxinů … (e.g. antibiotics)
• Zadržování metabolitů v buňce
• Příjem vnějších signálů a přenos informace dovnitř buňky
• Kompartmentace v rámci buňky, organely, sekreční váčky ...
– Oddělení energii produkujících a energii spotřebovávajících reakci
– Oddělení proteolytických enzymů od okolí
• Vznik a přenos nervových signálů
• Akumulace energie ve formě protonového gradientu a
tvorba ATP

• Vlastnosti hydrofilních částí lipidů určují povrchové vlastnosti membrány
• Vlastnosti hydrofobní částí lipidů (složení mastných kyselin) určuje fyzikální vlastnosti membrány
• Různé organismy mají různé lipidové složení membrán
• Různé tkáně mají rozdílné lipidové složení membrán
• Různé organely mají rozdílné lipidové složení membrán
Složení biologických membrán

Lipidové složení se liší mezi organismy, tkáněmi a pletivy i mezi tkáněmi jednoho organismu:
• Druh lipidů
• Poměr mezi množstvím lipidů a proteinů
• Množství a typy sterolových lipidů
Složení biologických membrán

Plasmtická a organelové membrány krysích hepatocytů
Rozdílné složení plasmatické
membrány a membrán organel

Rozdílné lipidové složení
monovrstev lipidové dvojvrstvy
• Lipidová dvojvrstva je
asymetrická
• Vnější strana je obvykle
positivně nabitá
• Např. fosfatidylserin na
vnější straně má
specifickou funkci:
– Aktivuje srážení krve
– Označuje buňky k destrukci

Organismy mění složení svých
biologických membrán
• Vlastnosti membrán jsou dány poměrem nasycených a
nenasycených mastných kyselin v lipidech
• Pro zachování stálé fluidity mohou buňky měnit lipidové
složení membrán a např. zvyšovat množství saturovaných
mastných kyselin při vyšší teplotě
Složení mastných kyselin membrány E. coli při různé teplotě

Funkce sterolů v eukariotních
membránách
• Zvyšují tuhost biologických membrán
• Zvyšují propustnost biologických
membrán
– Cholesterol u živočichů
– Fytosteroly u rostlin
– Ergosteroly u hub

Typy membránových proteinů
• Integrální membránové
proteiny
• Procházejí lipidovou
dvojvrstvou
• Periferální
membránové proteiny
• vázané k povrchu membrány
nevazebnými interakcemi
(vodíkové vazby,
elektrostaticky) nebo
kovalentní vazba na lipid
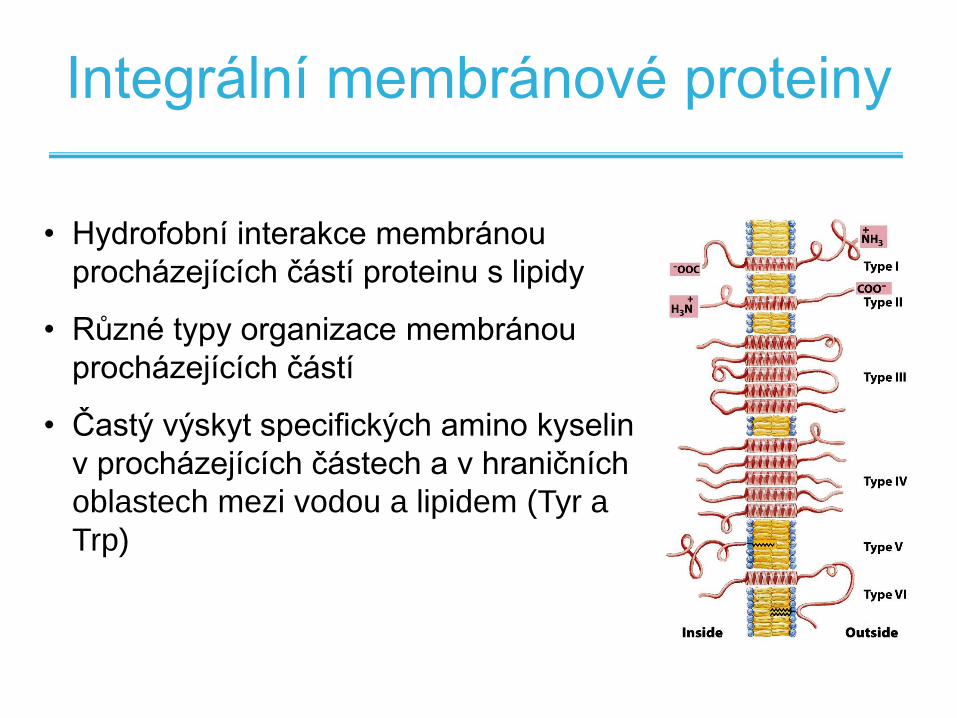
Integrální membránové proteiny
• Hydrofobní interakce membránou
procházejících částí proteinu s lipidy
• Různé typy organizace membránou
procházejících částí
• Častý výskyt specifických amino kyselin
v procházejících částech a v hraničních
oblastech mezi vodou a lipidem (Tyr a
Trp)
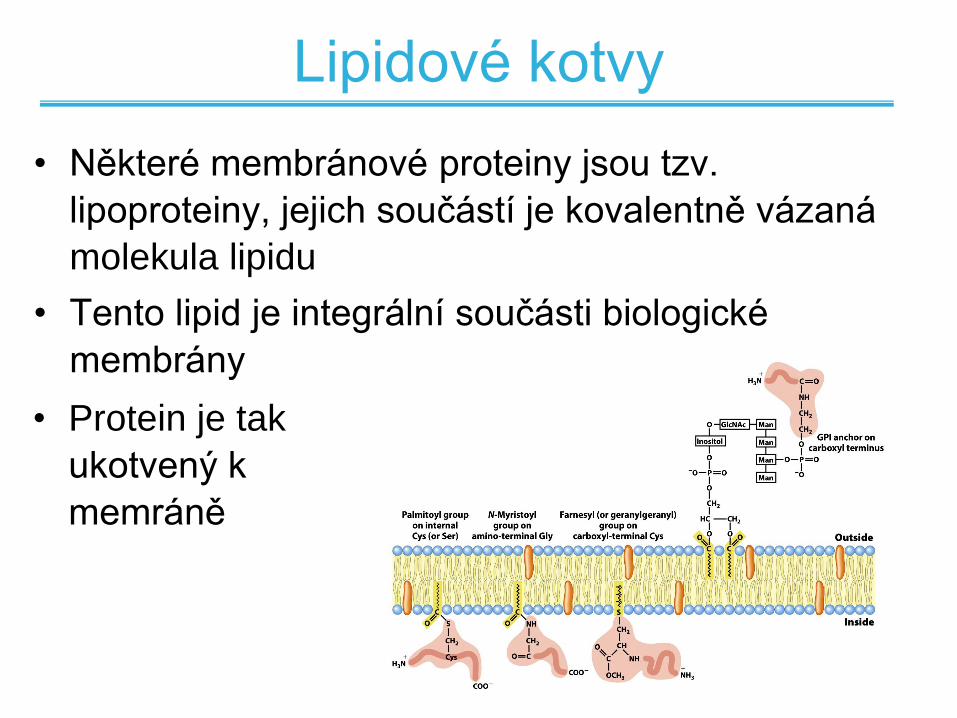
Lipidové kotvy
• Některé membránové proteiny jsou tzv.
lipoproteiny, jejich součástí je kovalentně vázaná
molekula lipidu
• Tento lipid je integrální součásti biologické
membrány
• Protein je tak
ukotvený k
memráně
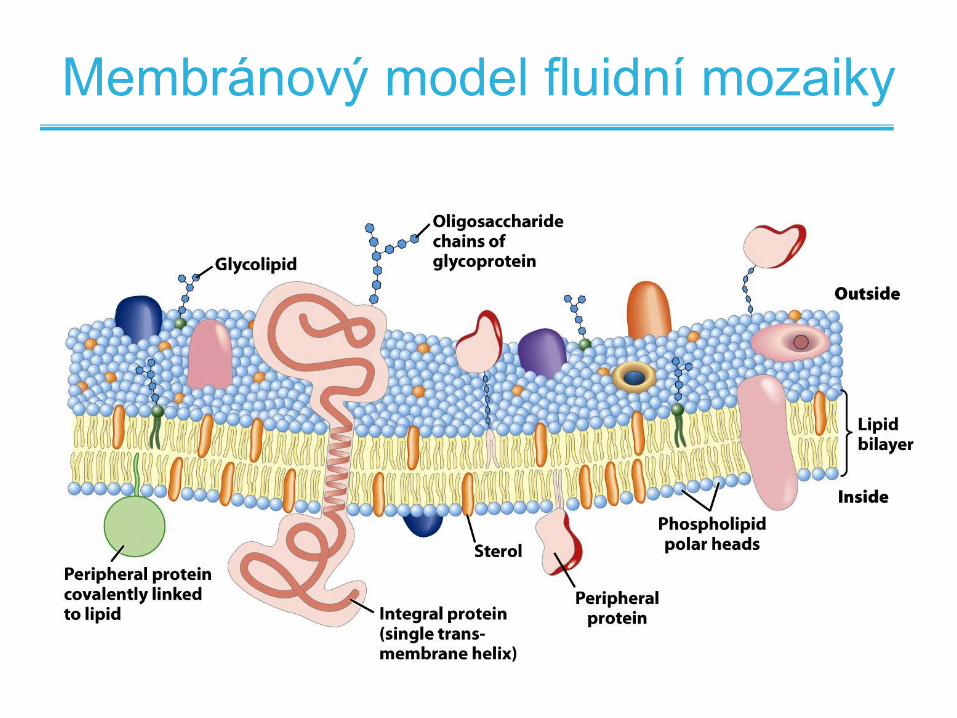
Membránový model fluidní mozaiky

• Lipidy tvoří viskózní 2D prostředí (rozpouštědlo),
ve kterém jsou ukotveny proteiny a rozpuštěny
různé další látky
• Integrální proteiny jsou pevně svázané s
membránou a protínají ji
• Periferní proteiny jsou vázány volněji a mohou být
snadno odstraněny
– Nekovalentní vazba
– Kovalentní vazba na membránové lipidy
Membránový model fluidní mozaiky

Dynamika biologických membrán

Fyzikální vlastnosti membrán
• Dynamické a flexibilní struktury
• Existují v různých fázích, dochází s fázovému
přechodu
• Nepropustné pro větší a polární částice a
ionty
• Propustné pro malé polární částice a
nepolární látky

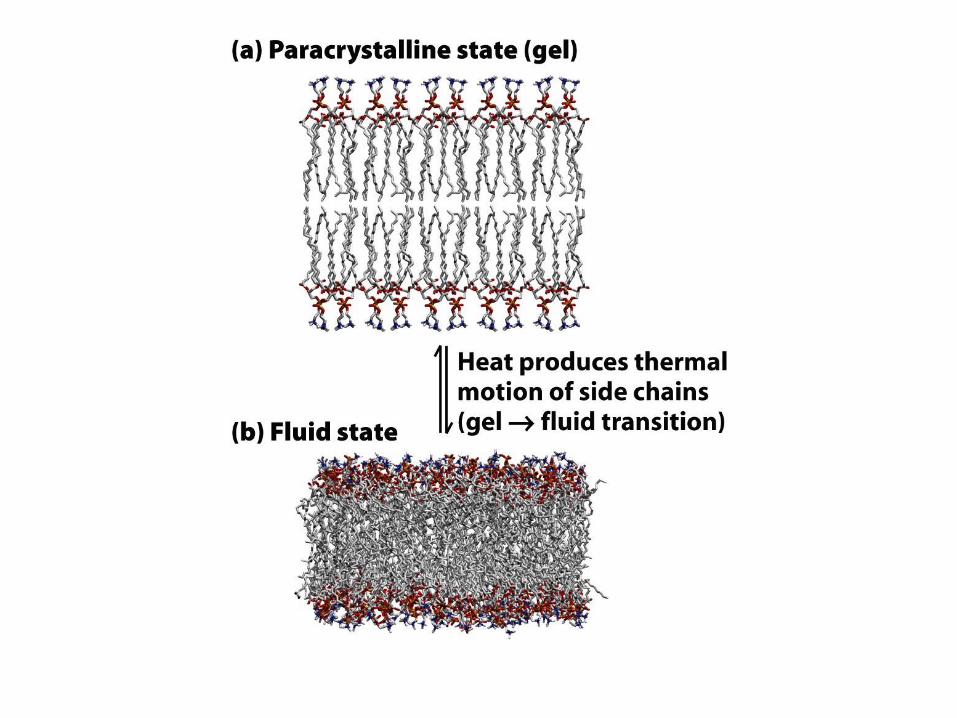
Fáze membrán
• Závisí na složení a teplotě, existují dvě fáze
biologických membrán:
– Gelová fáze: jednotlivé molekuly se volně
laterálně nepohybují
– Tekutá fáze: jednotlivé molekuly se volně
pohybují membránou
• Ohřev/ochlazení membrány - fázová změna
• Za fyzilologických podmínek jsou membrány v
tekutém stavu
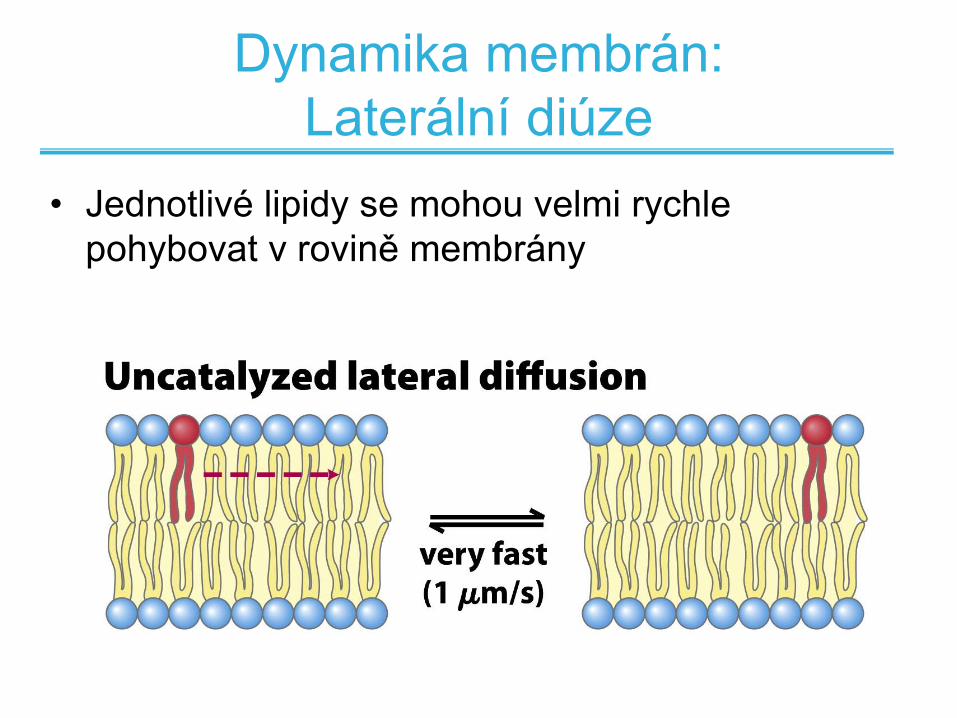
Dynamika membrán:
Laterální diúze
• Jednotlivé lipidy se mohou velmi rychle
pohybovat v rovině membrány
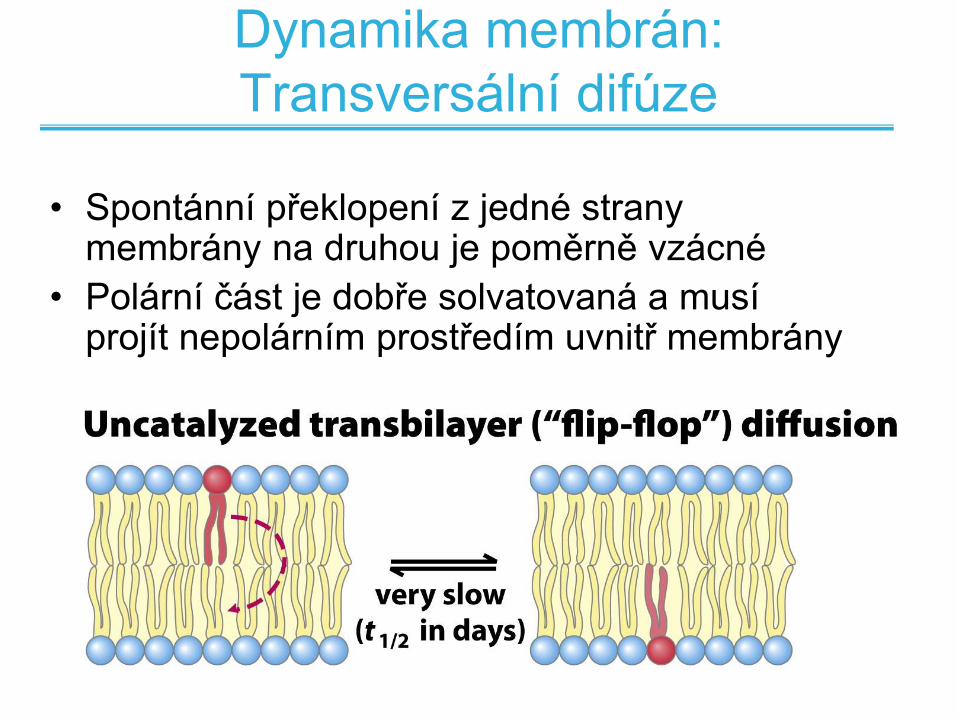
Dynamika membrán:
Transversální difúze
• Spontánní překlopení z jedné strany membrány na druhou je poměrně vzácné
• Polární část je dobře solvatovaná a musí projít nepolárním prostředím uvnitř membrány

Dynamika membrán: Flipázy
• Speciální enzymy – flipázy – katalyzují
difúzi/překlopení mezi jednotlivými vrstvami
• Některé flipázy využívají energii ATP k přenosu
lipidů proti koncentračnímu gradientu
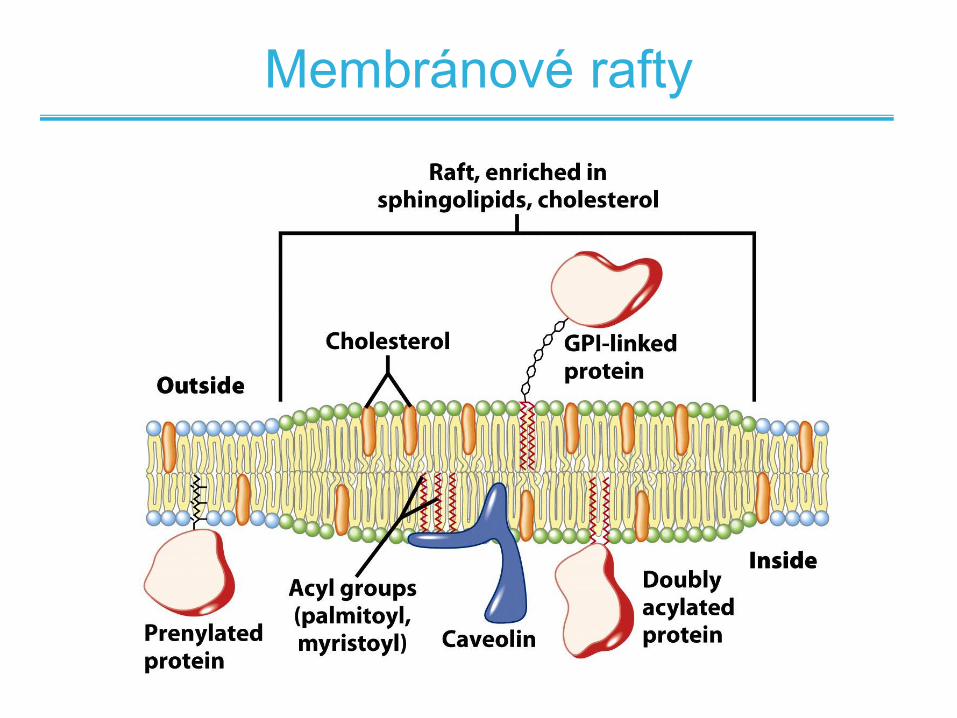
Membránové rafty

Membránové rafty
• Distribuce lipidů v membránových vrstvách není
náhodná
• Některé regiony obsahují klastry glykosfingolipidů
s delšími mastnými kyselinami
• Tyto části jsou organizovanější a obsahují
speciální proteiny
• Takové oblasti – membránové rafty – dovolují
agregaci či funkční shlukování součástí
biologických membrán (proteinů)

Fúze membrán
• Membráy mohou splývat mezi sebou bez jejich
přerušení/porušení
• Fúze může být spontánní nebo
zprostředkovaná proteiny
• Příklady zprostředkované fúze jsou
– Vstup viru chřipky do buňky
– Vypuštění neurotransmiterů nervovými synapsemi
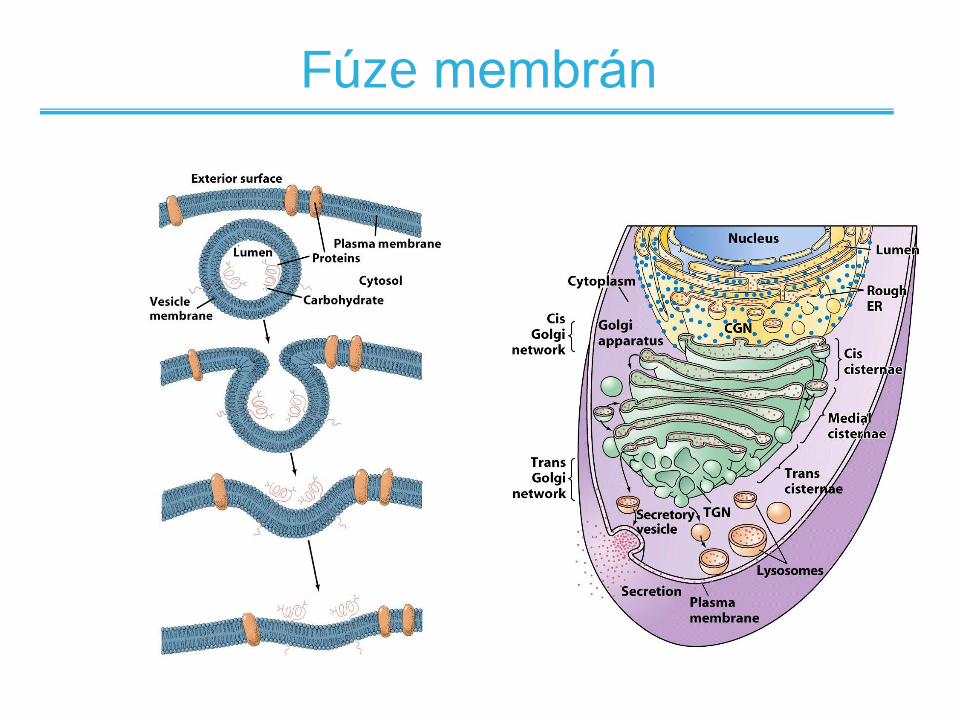
Fúze membrán

Membránový
transport
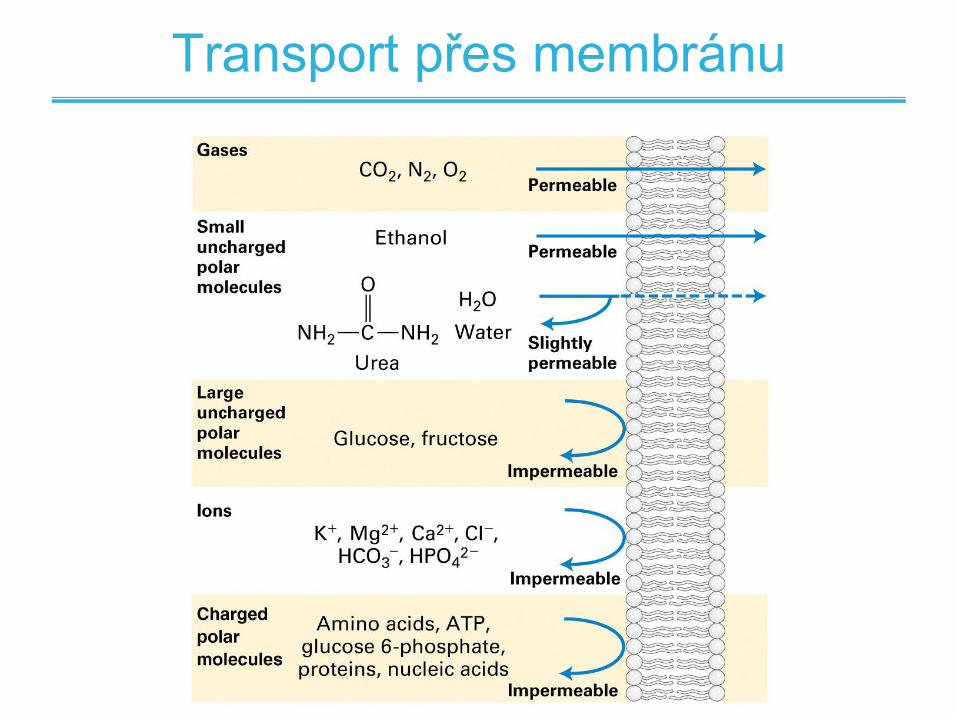
Transport přes membránu

Transport přes membránu
• Některé látky mohou přímo difundovat biologickou membránou
• Pasivní difúze - pouze plyny a malé nenabité hydrofobní molekuly
• Difúze polárních molekul vyžaduje desolvataci a tudíž je nutné překonání vyšší aktivační bariéry
• Usnadněný transport přes membránu pomocí proteinových přenašečů – membránových transportérů –alternativní cesta difúze
• Aktivní transport – transport za pomoci energie, spotřeba ATP
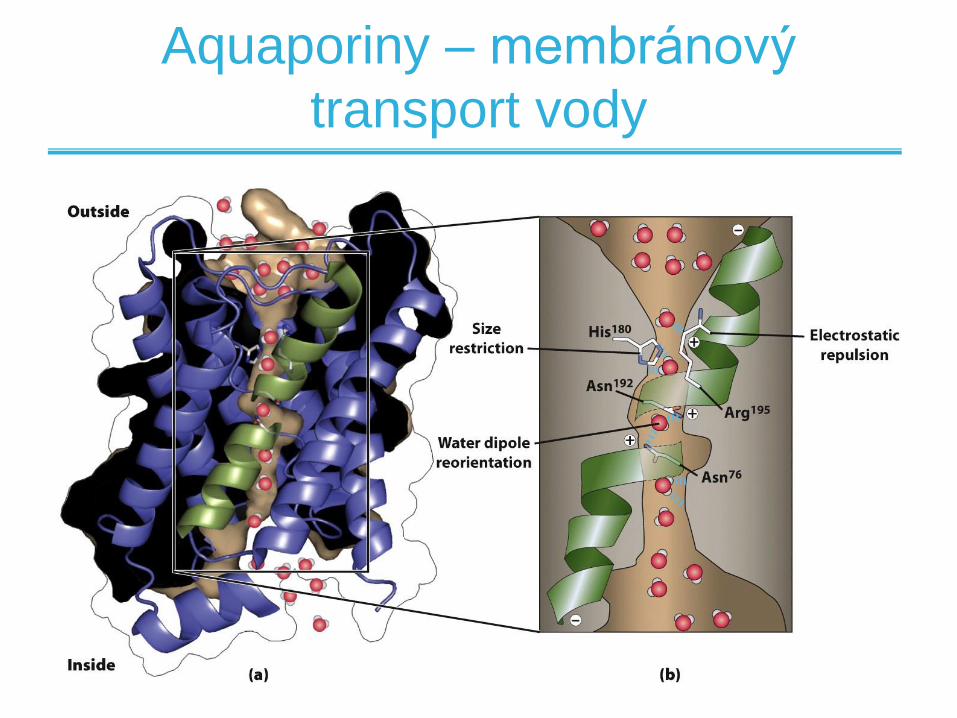
Aquaporiny – membránový
transport vody
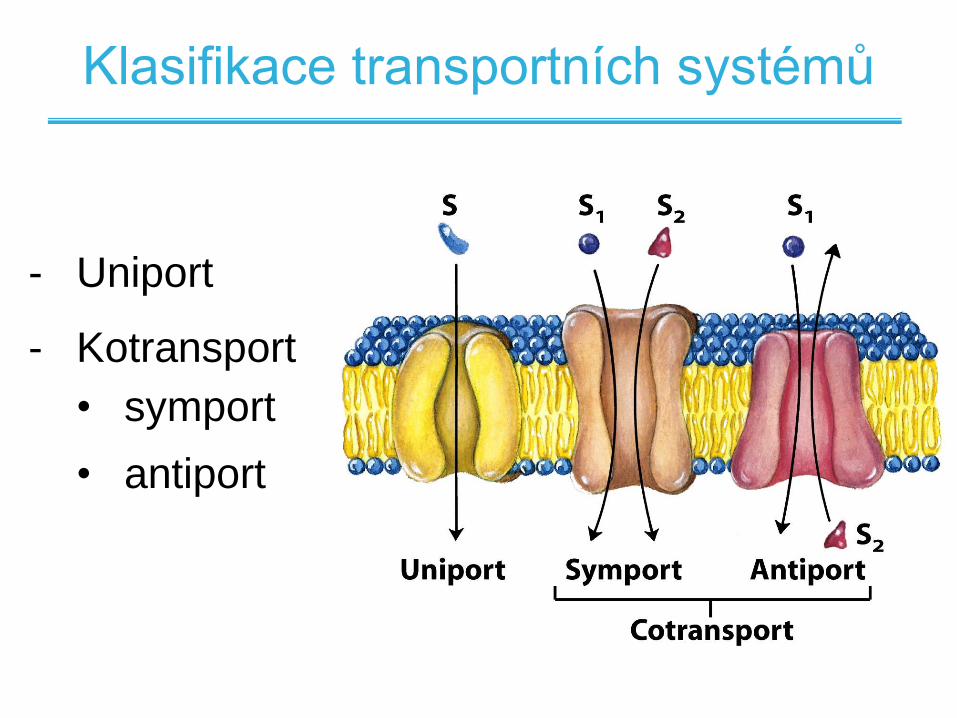
Klasifikace transportních systémů
- Uniport
- Kotransport
• symport
• antiport
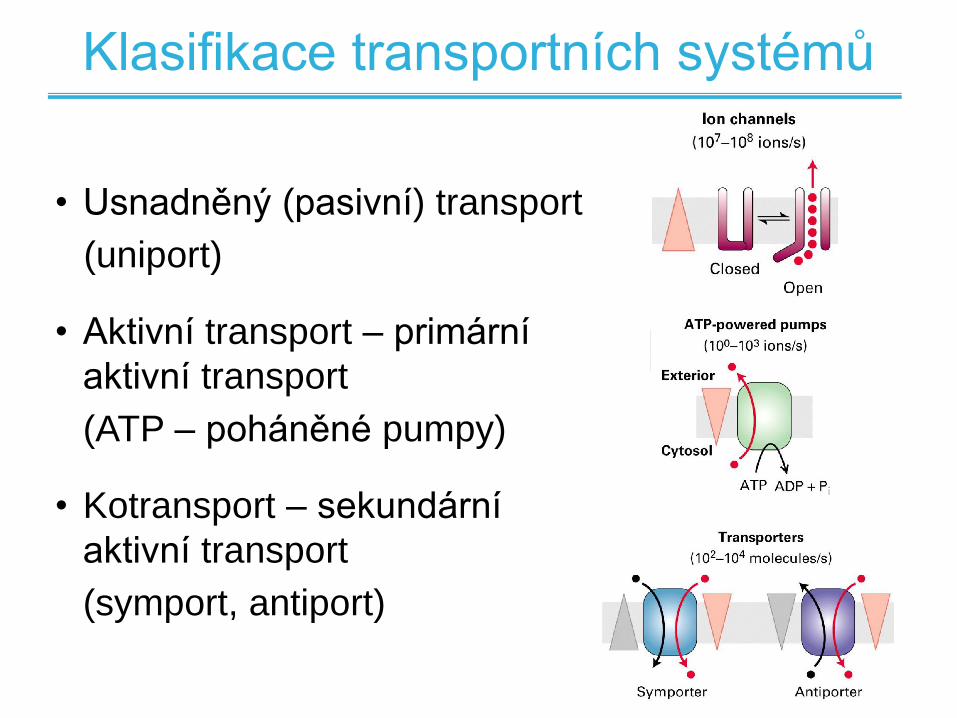
• Usnadněný (pasivní) transport
(uniport)
• Aktivní transport – primární
aktivní transport
(ATP – poháněné pumpy)
• Kotransport – sekundární
aktivní transport
(symport, antiport)
Klasifikace transportních systémů
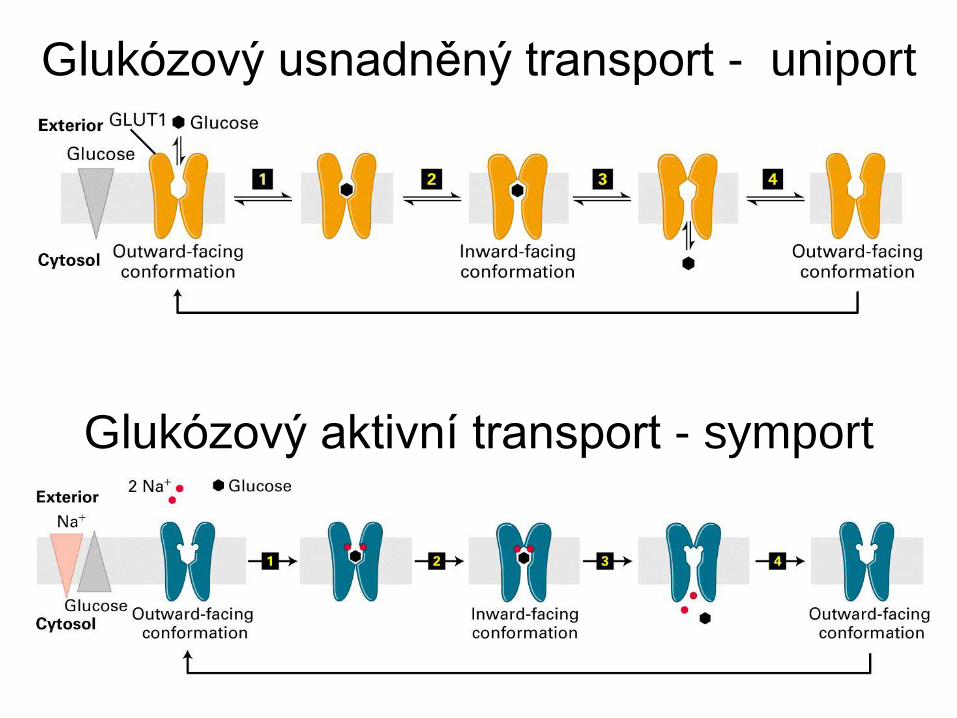
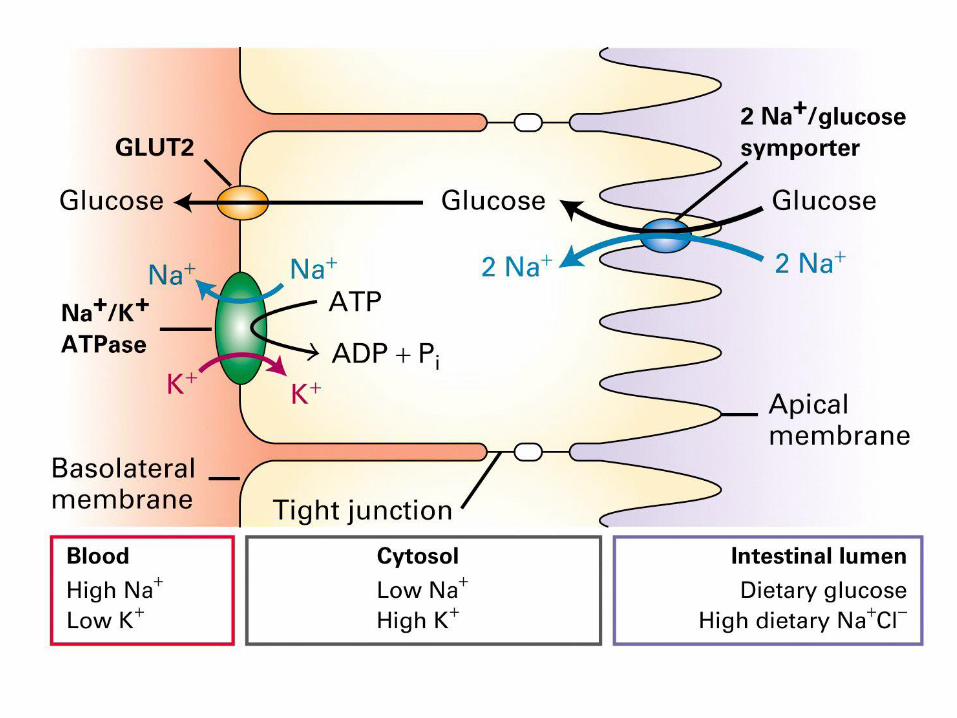
Glukózový usnadněný transport - uniport
Glukózový aktivní transport - symport
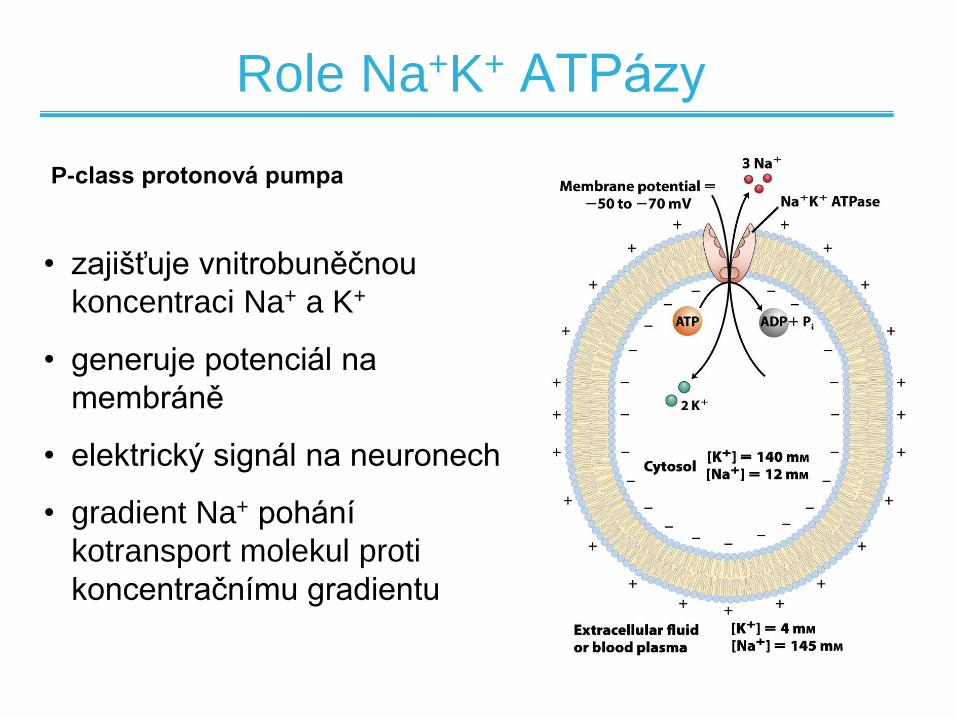
Role Na+K+ ATPázy
• zajišťuje vnitrobuněčnou
koncentraci Na+ a K+
• generuje potenciál na
membráně
• elektrický signál na neuronech
• gradient Na+ pohání
kotransport molekul proti
koncentračnímu gradientu
P-class protonová pumpa

Metabolismus lipidů
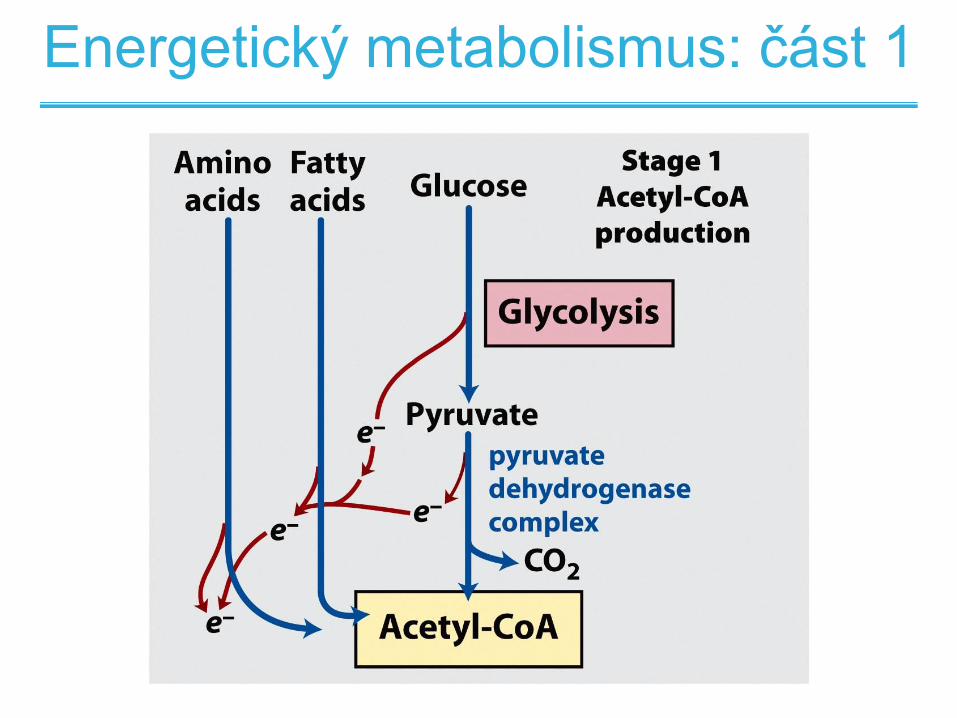
Energetický metabolismus: část 1
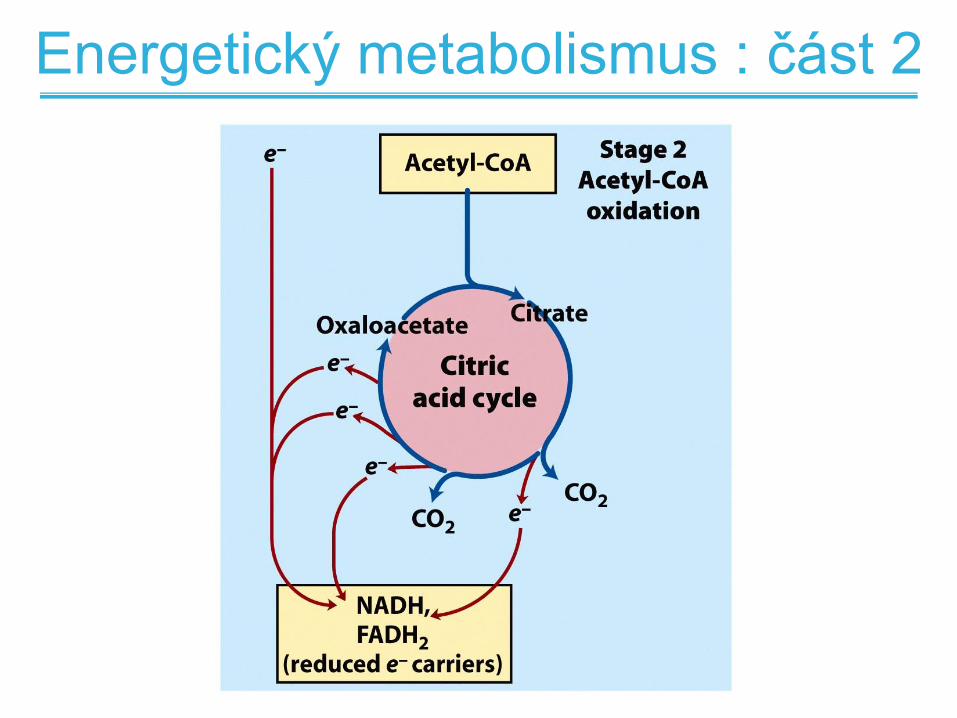
Energetický metabolismus : část 2
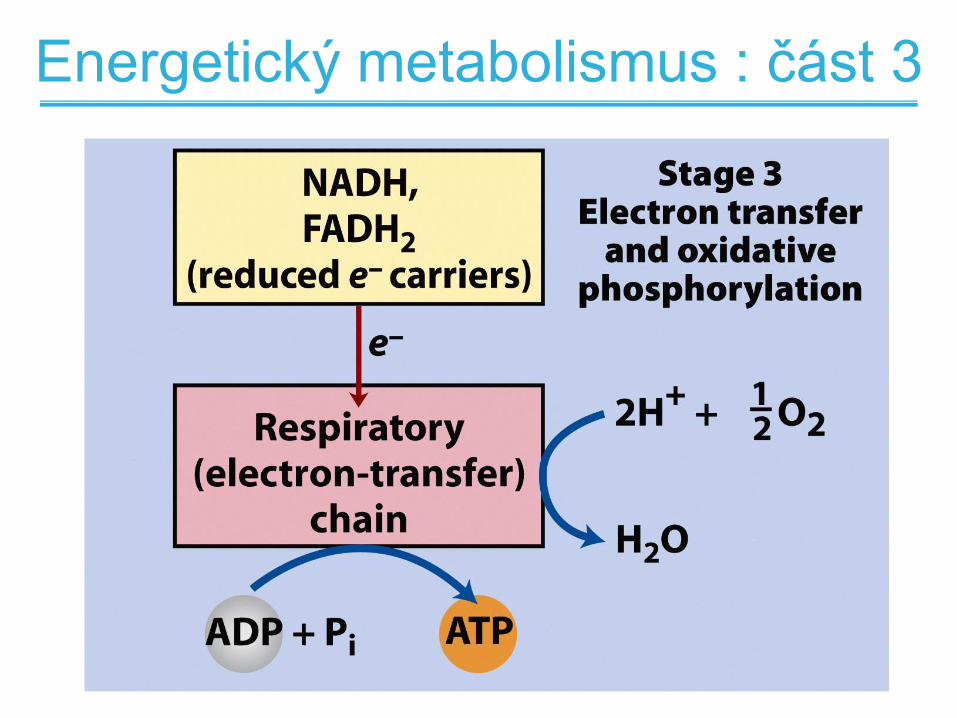
Energetický metabolismus : část 3

Katabolismus lipidů

Oxidace mastných kyselin – hlavní
zdroj energie mnoha organismů
• Asi jedna třetina naší celkové energetické potřeby
je kryta oxidací triacylglycerolů
• Přibližně 80% energetických potřeb srdce a jater
savců je pokryto oxidací mastných kyselin
• Řada hibernujících organismů, jako např. grizzly
závisí výhradně na tuku jako zdroji energie

Tuk je účinný zdroj energie
• Výhoda tuků oproti cukrům:
– Mastné kyseliny obsahují více energie na uhlík
protože jsou redukovanější
– Mastné kyseliny nevážou vodu lepší sladování
• Glukóza a glykogen – rychlý a krátkodobý zdroj
energie
• Mastné kyseliny – pomalejší ale dlouhodobější
zdroj energie
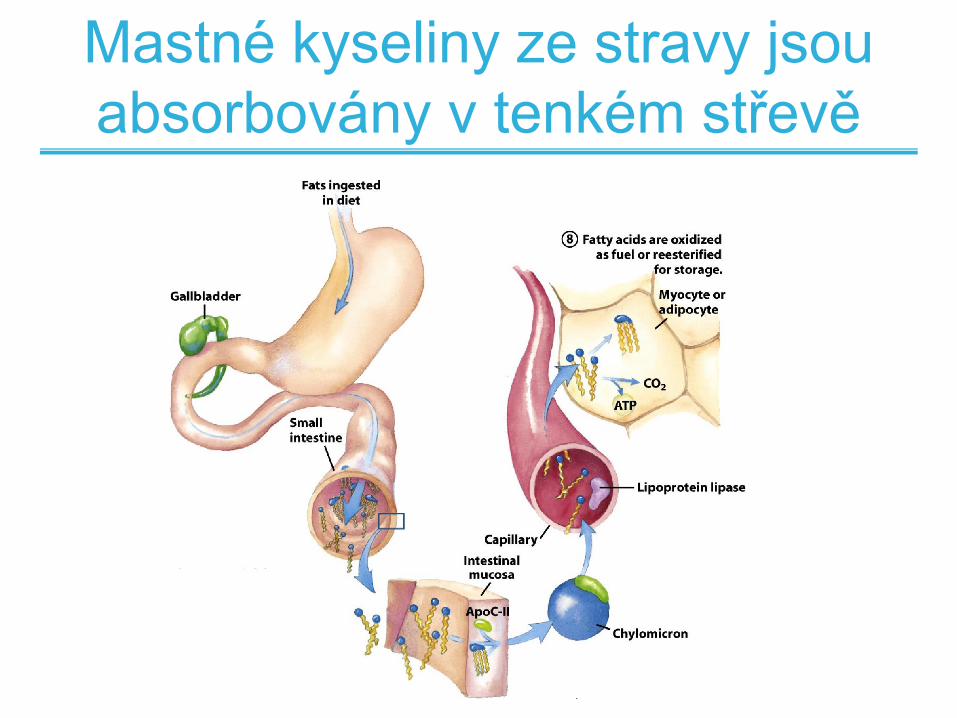
Mastné kyseliny ze stravy jsou
absorbovány v tenkém střevě

Hydrolýza tuků na mastné
kyseliny a glycerol
• Hydrolýza tuků je katalyzována lipázami
• Triacylglyceroly jsou rozloženy na mastné kyseliny a glycerol
a transportovány přes střevní stěnu
• Poté jsou zpět přeměněny na triacylglyceroly a přenášny ve
formě částic - chylomikronů
• V kapilárních cévách je triacylglycerol opět rozložen masté
kyseliny a glycerol a tak transpotovány do buňky
• Mastné kyseliny jsou oxidovány nebo opět re-esterifikovány
na triacylglycerol a skladovány
Adipocyty – skladování tuků
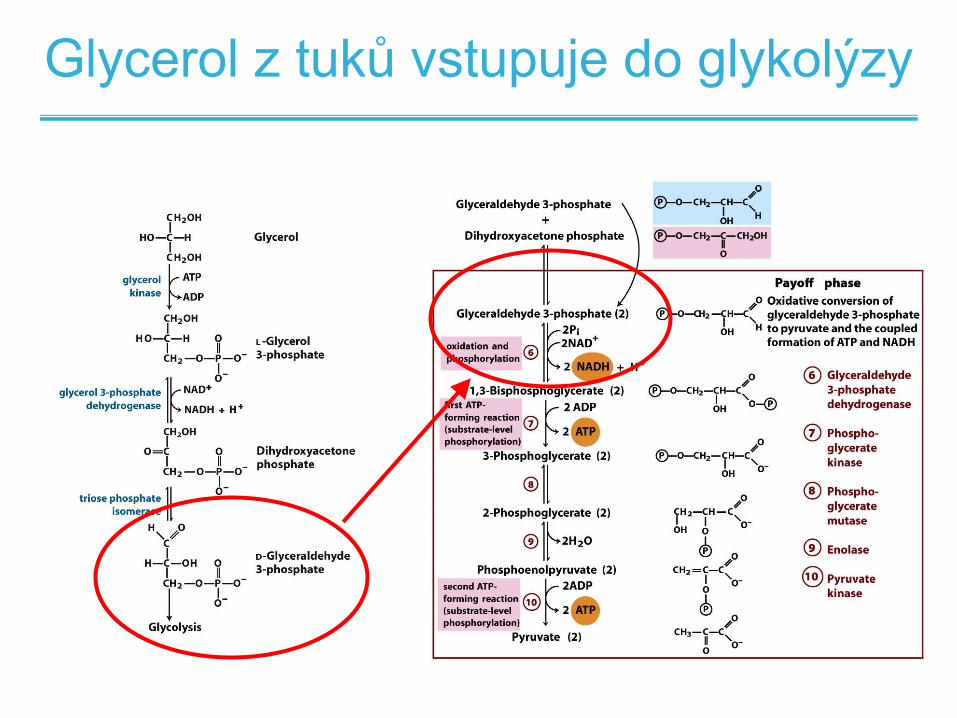
Glycerol z tuků vstupuje do glykolýzy

• Glycerol kináza – aktivace glycerolu za spotřeby ATP
• Následné reakce glykolýzy vyprodukují 2 molekuly
ATP - dostatečné pokrytí potřeb
• Pouze velmi omezené anaerobní štěpení tuků
Glycerol z tuků vstupuje do glykolýzy

Mastné kyseliny jsou tranportovány
do mitochondrií
• Tuky jsou štěpeny na mastné kyseliny a glycerol v
cytoplazmě
• -oxidace mastných kyselin se vyskytuje v
mitochondriích
• Krátké (< 12 uhlíků) mastné kyseliny difundují
volně přes mitochondriální
• Delší jspu transportovány pomocí acyl-karnitin /
karnitinového transporteru
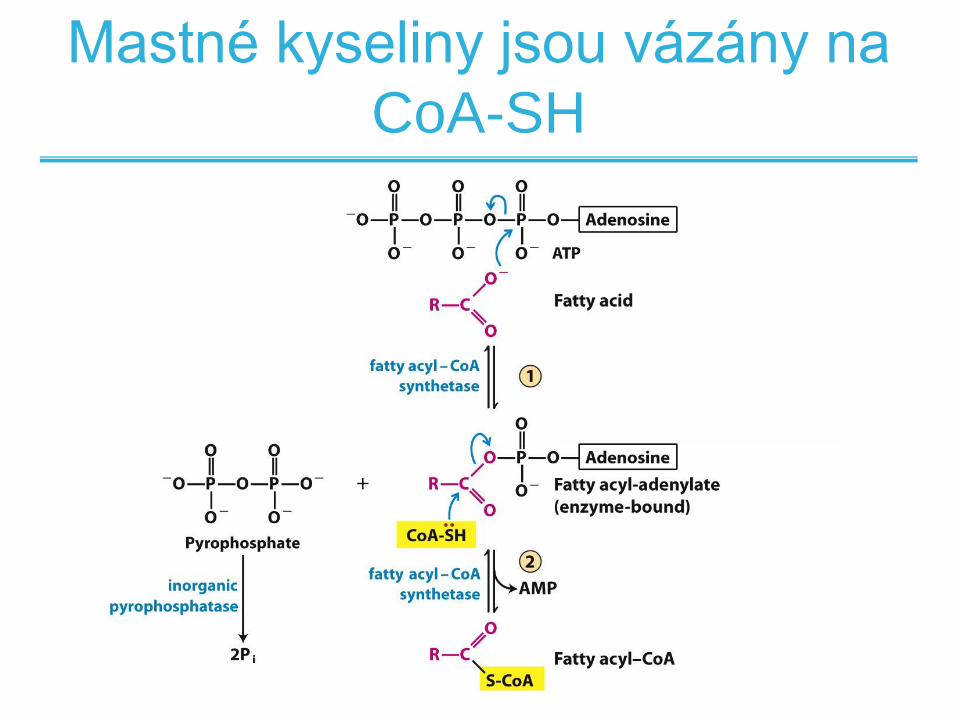
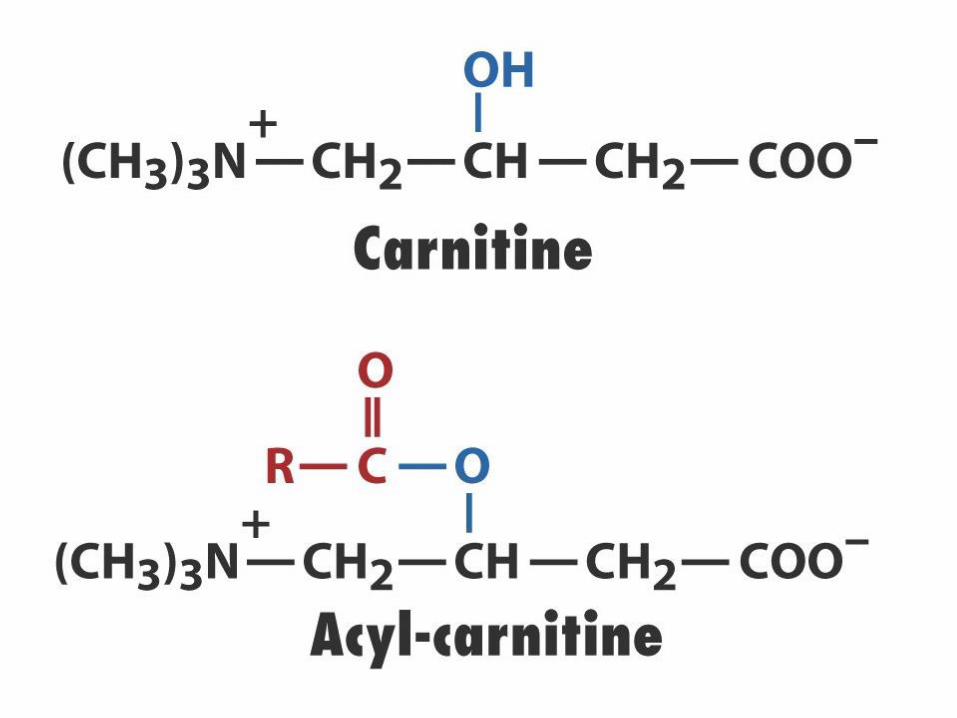
Mastné kyseliny jsou vázány na
CoA-SH
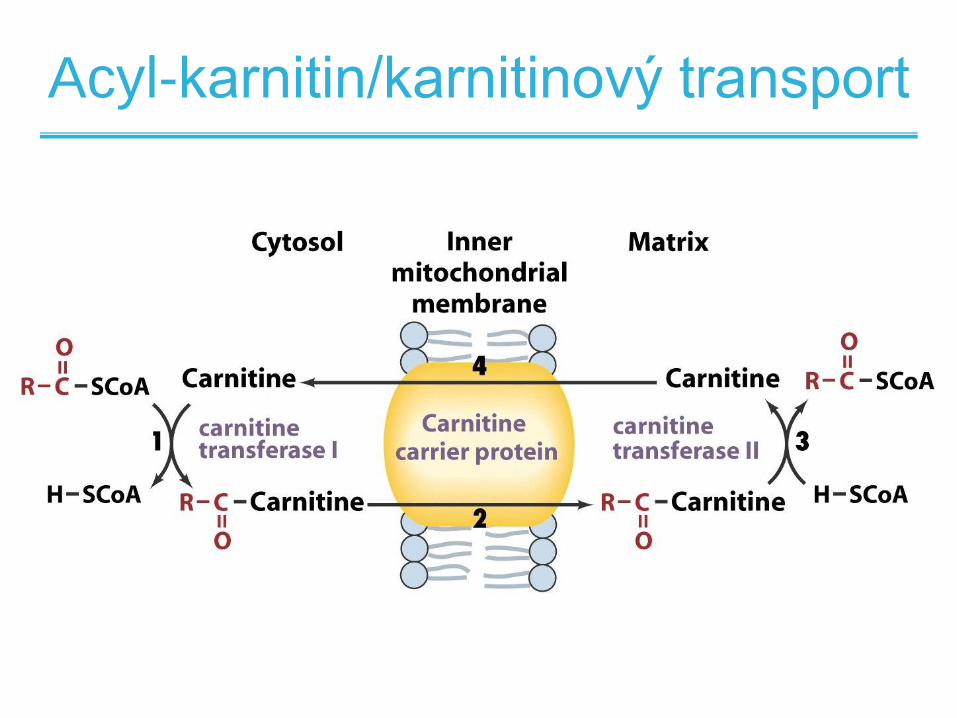
Acyl-karnitin/karnitinový transport

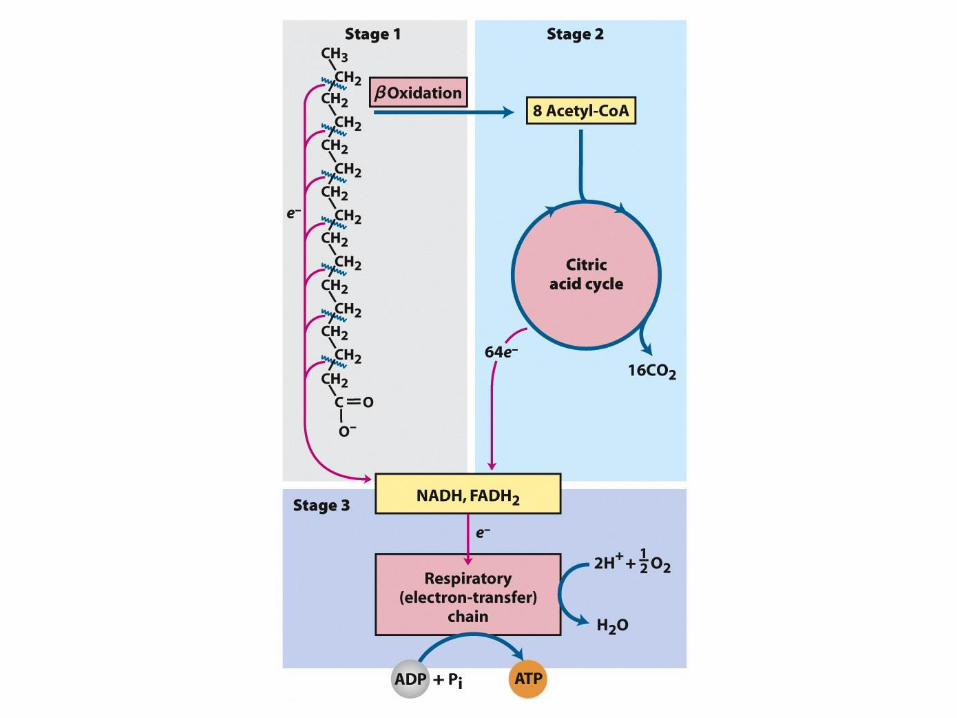
Oxidace mastných kyselin

Stupně oxidace mastných kyselin
• 1. stupeň oxidativní stěpení mastných kyselin
na dvouuhlíkaté jednoty vázané v acetyl-CoA
za současné produkce NADH
• 2.stupeň oxidace acetyl-CoA na CO2 v
citrátovém cyklu za současné produkce
NADH a FADH2
• 3. stupeň produkce ATP z NADH a FADH2 v
dýchacím řetězci
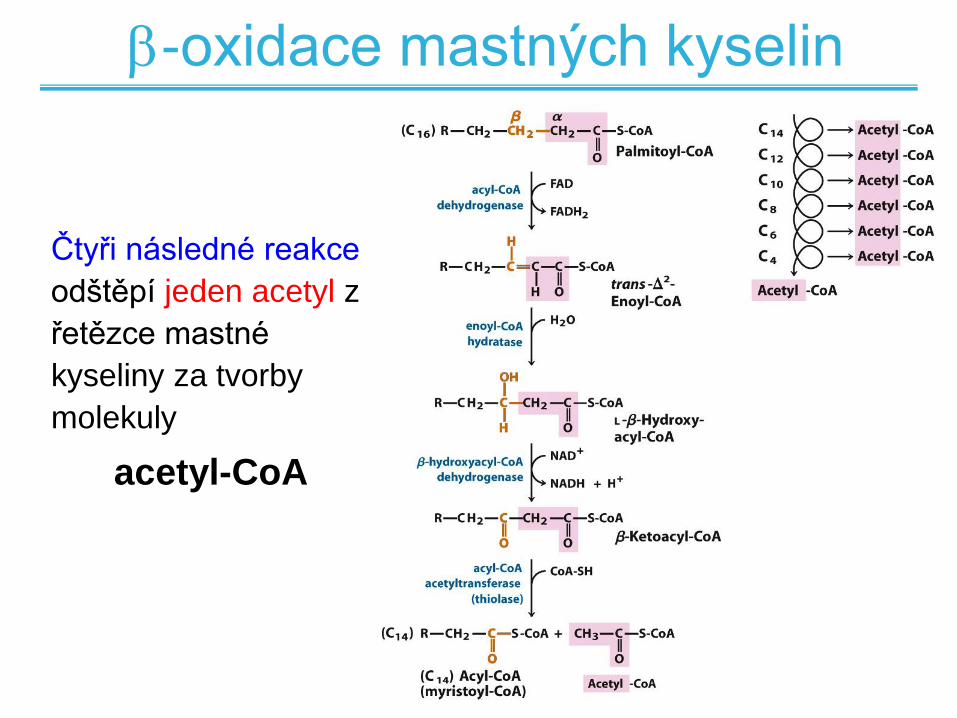
-oxidace mastných kyselin
Čtyři následné reakce
odštěpí jeden acetyl z
řetězce mastné
kyseliny za tvorby
molekuly
acetyl-CoA

1. reakce -oxidace
dehydrogenace alkanu na alken

1. reakce -oxidace
dehydrogenace alkanu na alken
• Katalyzovánarůznými isoformami acyl-CoA
dehydrogenázy (AD) na vnitřní mitochondriální
membráně
– Dlouhé řetězce AD (12-18 uhlíků)
– Střední řetězce AD (4-14 uhlíků)
– Krátké řetězce AD (4-8 uhlíků)
• Reakce podobná sukcinát dehydrogenáze v
citrátovém cyklu

FAD kofactor
• FAD podstupuje 2-elektronové redukci
• Elektrony z vázaného FADH2 jsou
předány na elektron-přenášející
flavoprotein (ETF)
• ETF přenášejí elektrony do
mitochondriálního dýchacího řetězce
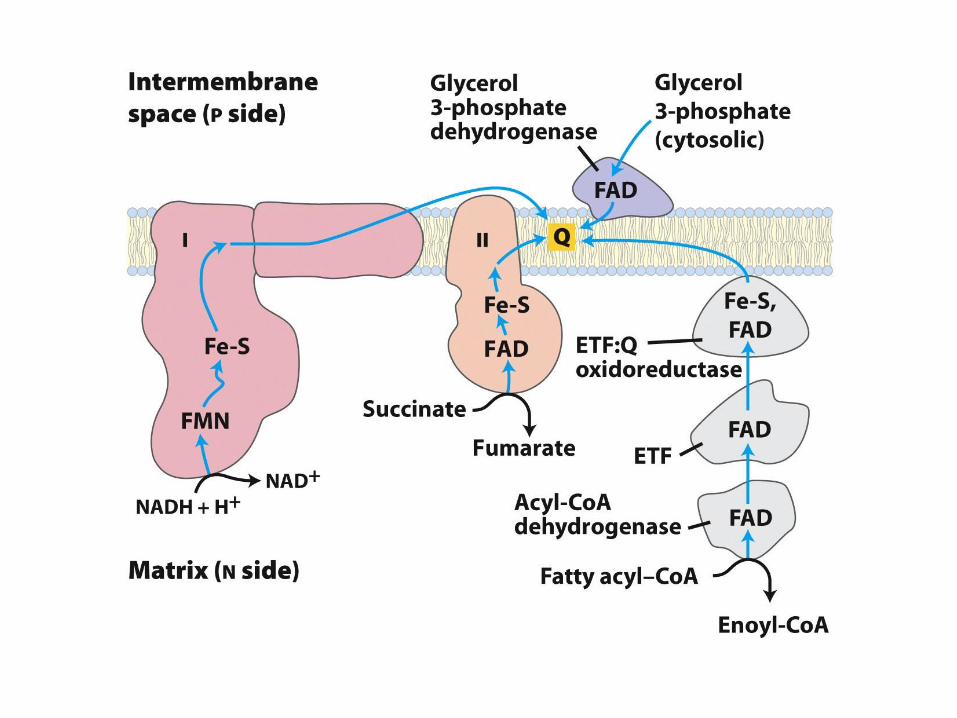

Trifunkční protein (TFP)• Následné tři reakce jsou katalyzované
multienzymovým komplexem na vnitřní mitochondriální membráně
• Zpracovává mastné kyseliny s řetězcem nad 12 uhlíků
• Kratší řetězce jsou oxidovány rozpustnými enzymy v matrix
• Trifunkční proteinový komplex je hetero-oktamer
– Čtyři podjednotky - vazba k membráně
• enoyl-CoA hydratázová aktivita (2)
• -hydroxyacyl-CoA dehydrogenázová aktivita (3)
– Čtyři podjednotky
• thiolázová aktivita (4)
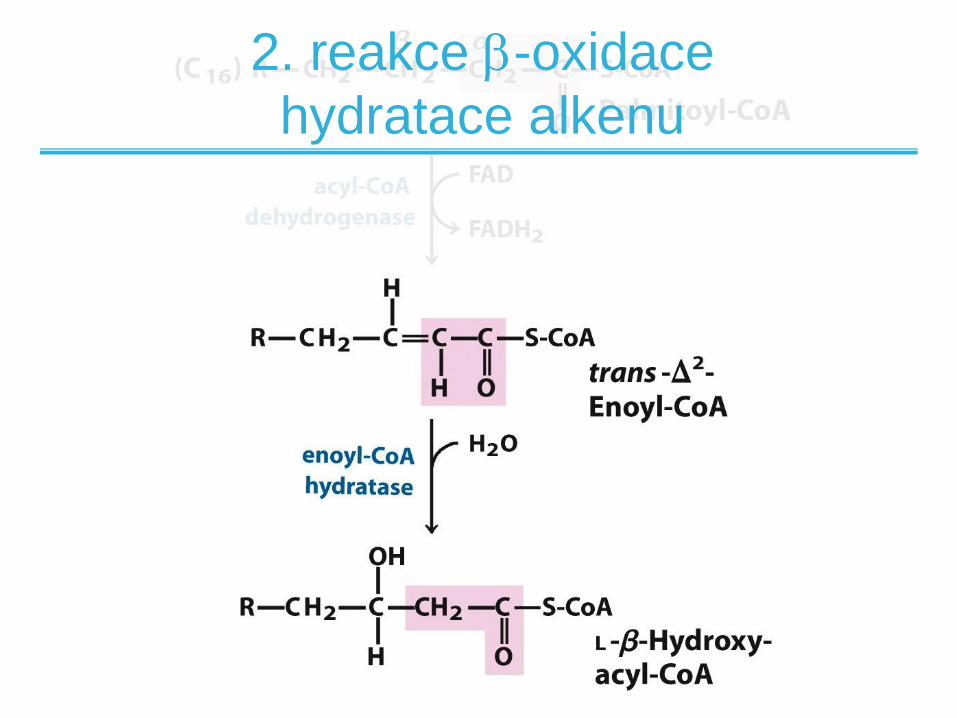
2. reakce -oxidace
hydratace alkenu

2. reakce -oxidace
hydratace alkenu
• Katalyzována dvěma izoformami enoyl-CoA
hydratázy:
– Rozpustná hydratáza na krátné řetězce
– Membránově vázaná na dlouhé řetězce (TFP)
• Váže vodu na dvojnou vazbu za vzniku alkoholu
• Analogické k fumarázové reakci v citrátovém
cyku

3. reakce -oxidace
dehydrogenace alkoholu

3. reakce -oxidace
dehydrogenace alkoholu
• Katalyzována -hydroxyacyl-CoA dehydrogenázou
• Enzym využívá NAD+ kofaktor
• Produkuje NADH, které vstupuje do dýchacího
řetězce
• Analogická malát dehydrogenázové reakci v
citrátovém cyklu
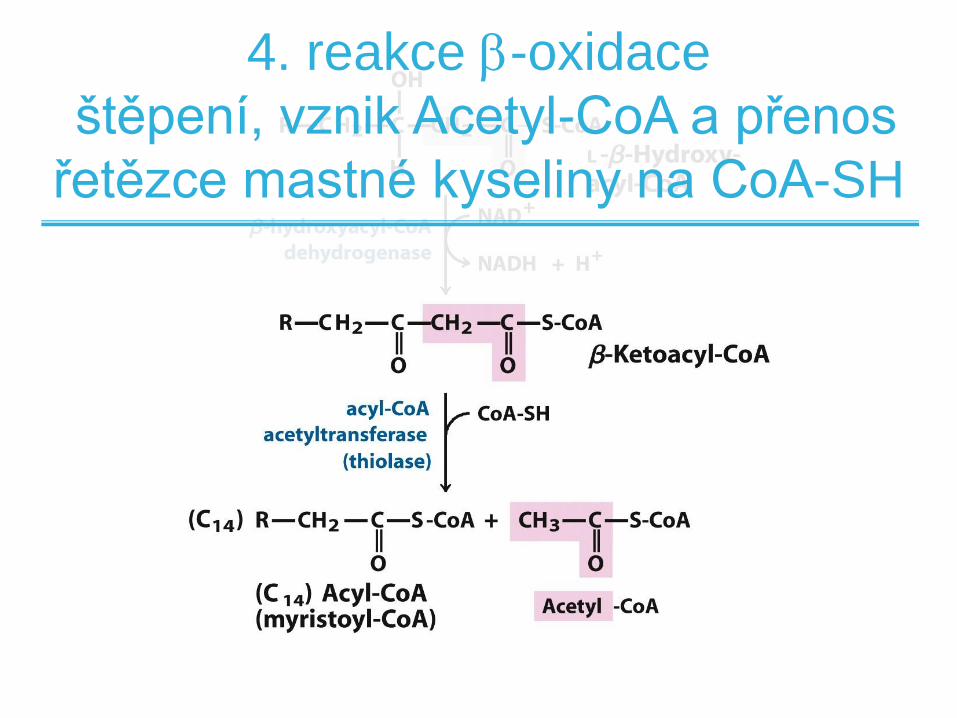
4. reakce -oxidace
štěpení, vznik Acetyl-CoA a přenos
řetězce mastné kyseliny na CoA-SH

4. reakce -oxidace
štěpení a transfer
• Katalyzováno acyl-CoA acetyltransferázou
(thiolázou)
• Reakcí je thiolýza uhlík-uhlíkové vazby


Energetická bilance
katabolismu mastných kyselin
• Palmitová kyselina C16 – sedminásobné
opakování cyklu čtyř reakcí – vznik:
– 8 Acetyl-CoA
– 7 FADH2 (v každém cyklu 1) – do dýchacího řetězce
– 7 NADH (v každém cyklu 1) – do dýchacího řetězce
• Acetyl-CoA vstupuje do citrátového cyklu kde je
dále oxidován na CO2
– za vzniku GTP (1), NADH (3) a FADH2 (1)

NADH a FADH2
zdroj energie pro tvorbu ATP
Výtěžek ATP na celkovou oxidaci jedné molekuly kyseliny palmitové
na CO2 a H2O

Oxidace nenasycených
mastných kyselin
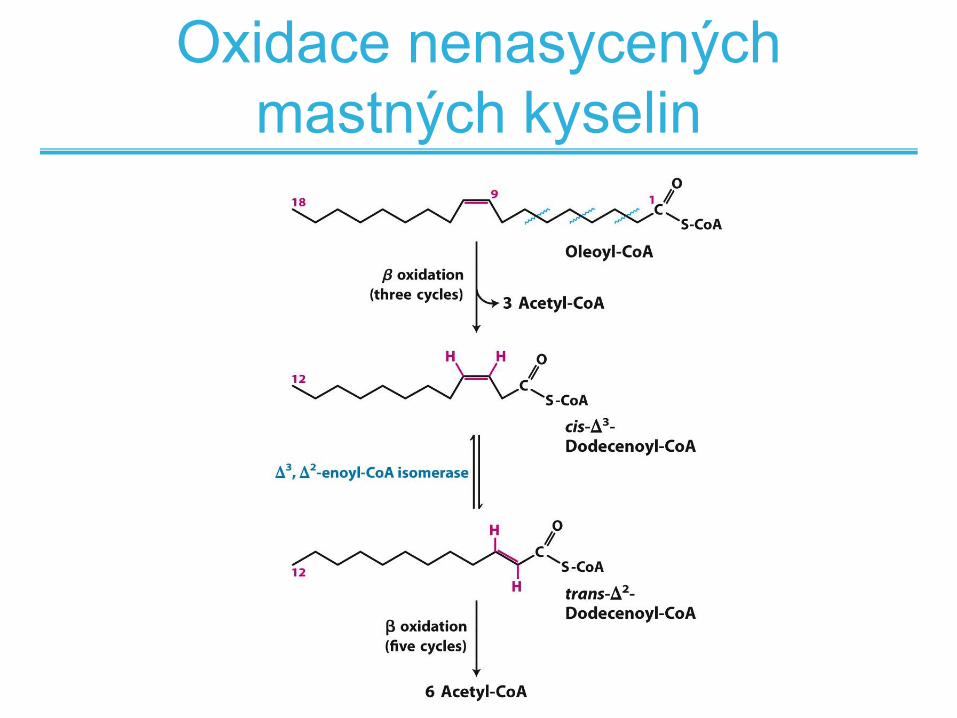
Oxidace nenasycených
mastných kyselin

Ketononá tělíska
(Ketone Bodies)

Tvorba ketonových tělísek
• Vstup acetyl-CoA do citrátového cykly vyžaduje
přítomnost oxalacetátu
• Za nedostatku oxalacetátu je acetyl-CoA
přeměňován na ketonová tělíska
• První krok je opak posledního kroku -oxidace:
thiolázová reakce spojí dvě acetátové jednotky


Ketonová tělíska vznikají v játrech
• Produkce ketonových tělísek se zvyšuje během hladovění
• Ketonová tělíska – vylučovány z jater do krve
• Ostatní orgány mohou využívat ketonová tělíska jako zdroj energie
• Příliš vysoké hladiny acetoacetátu a -hydroxybutyrátu snižují pH krve – ohrožení zdraví a života


Biosyntéza lipidů

Lipidy a jejich funkce v
organismech
• Zásoba energie
• Tvorba membrán
• Ukotvení membránových proteinů
• Kofaktory enzymů
• Signální molekuly
• Pigmenty
• Detergenty
• Transportní funkce
• Antioxidanty

Odbourávání a syntéza mastných
kyselin jsou dva odlišné procesy
• Odbourávání mastných kyselin
– produkce acetyl-CoA
– redukčních ekvivalentů NADH, FADH2
– lokalizace: mitochondrie
• Syntéza mastných kyselin
– vyžaduje malonyl-CoA a acetyl-CoA
– redukce za využití elektronů z NADPH
– lokalizace: cytosol u zvířat, chloroplasty rostlinách

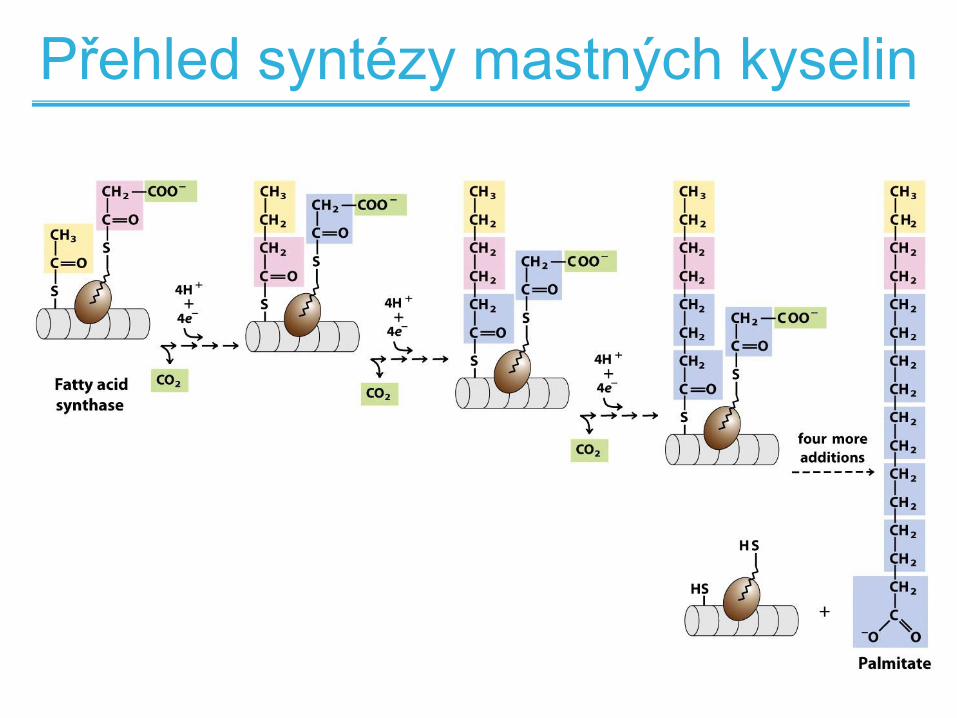
Přehled syntézy mastných kyselin
• Mastné kyseliny jsou tvořeny postupným
přidáváním dvouuhlíkatých acetátových
jednotek
• Acetát přichází z aktivované molekuly
malonyl-CoA
• Následně dochází k redukci karbonylu
acetátu na methylen
Přehled syntézy mastných kyselin

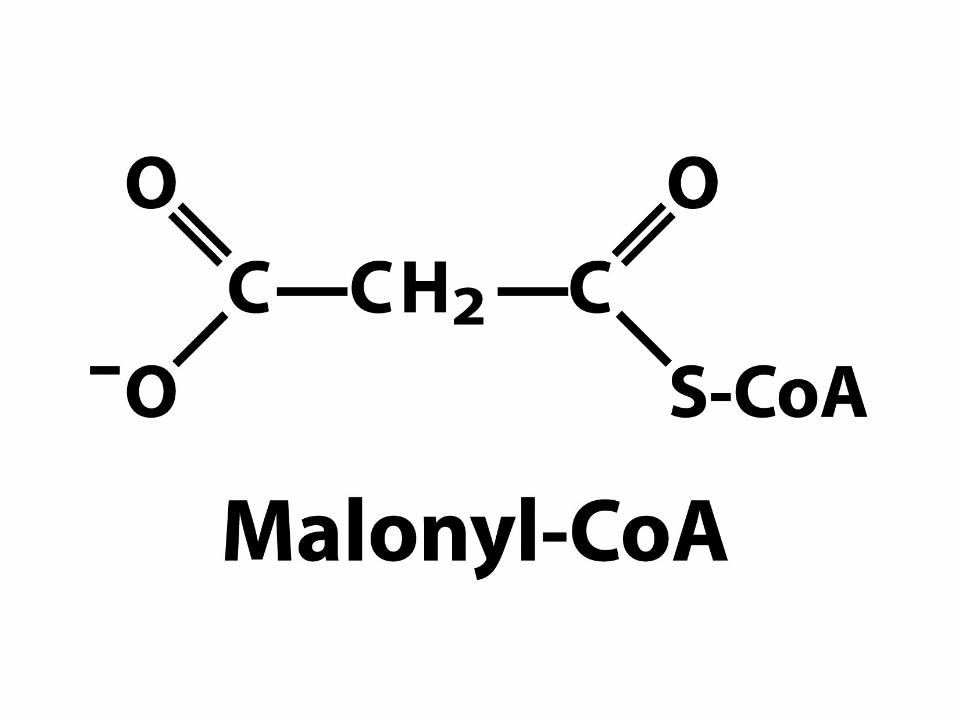
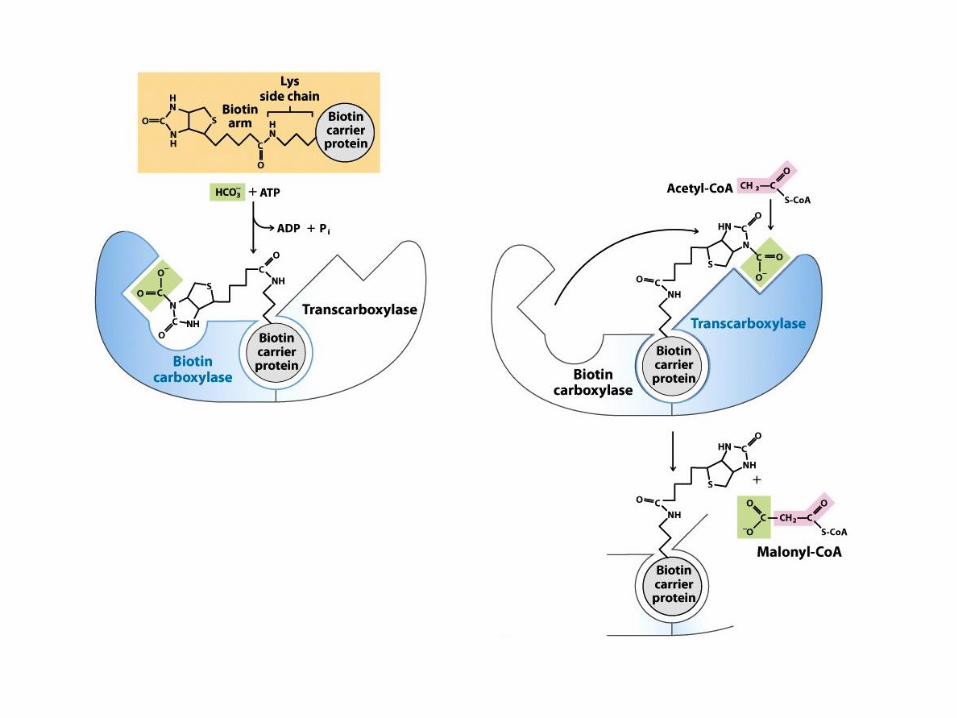
Syntéza Malonyl-CoA
• Tříuhlíkatý prekurzor syntézy mastných kyselin
vzniká z acetyl-CoA a CO2
• Reakce je katalyzována acetyl-CoA karboxylázou
(ACC)
• ACC je bifunkční enzum
– Biotin karboxyláza
– Transkarboxyláza
• ACC obsahuje biotin, koenzym s funkcí
přenašeče CO2
– Biotin přesmykává mezi dvěma aktivními centry
enzymového komplexu

Syntéza mastných kyselin
• Cílem je navázat dvouuhlíkatý zbytek z malonyl-CoA na
rostoucí řetězec mastné kyseliny a redukovat jej
• Reakce zahrnuje čtyři enzymově katalyzované kroky
– Kondenzace rostousího řetězce s acetátem z malonyl-CoA
– Redukce karbonylu acetátu na hydroxyl
– Dehydratace alkoholu na trans-alken (vznik dvojné vazby)
– Redukce alkenu na alkan

Komplex enzymu syntetizujícího
mastné kyseliny
• Syntézy mastných kyselin se účastní dva thioly
– Thiol z 4-fosfopantetheinu v Acyl Carrier Protein
– Thiol z cysteinu proteinu Syntázy mastných kyselin

Připojení dvouuhlíkatého řetězce
Acetyl kondenzuje s
malonylem za vzniku
acetoacetylu
vázeného na ACP
za současné
dekarboxylace
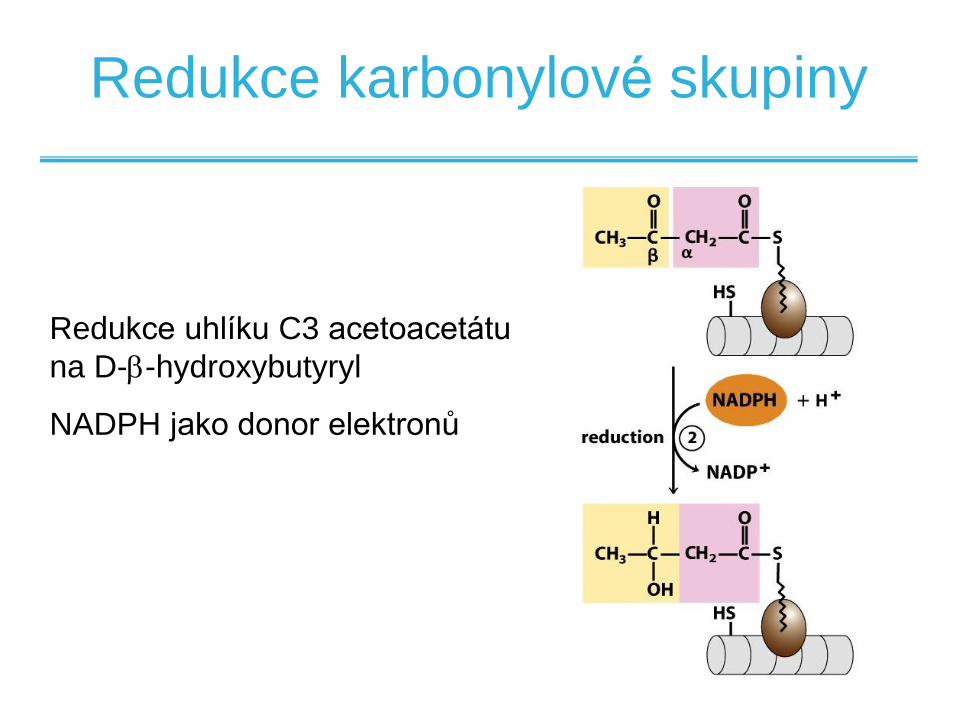
Redukce karbonylové skupiny
Redukce uhlíku C3 acetoacetátu
na D--hydroxybutyryl
NADPH jako donor elektronů
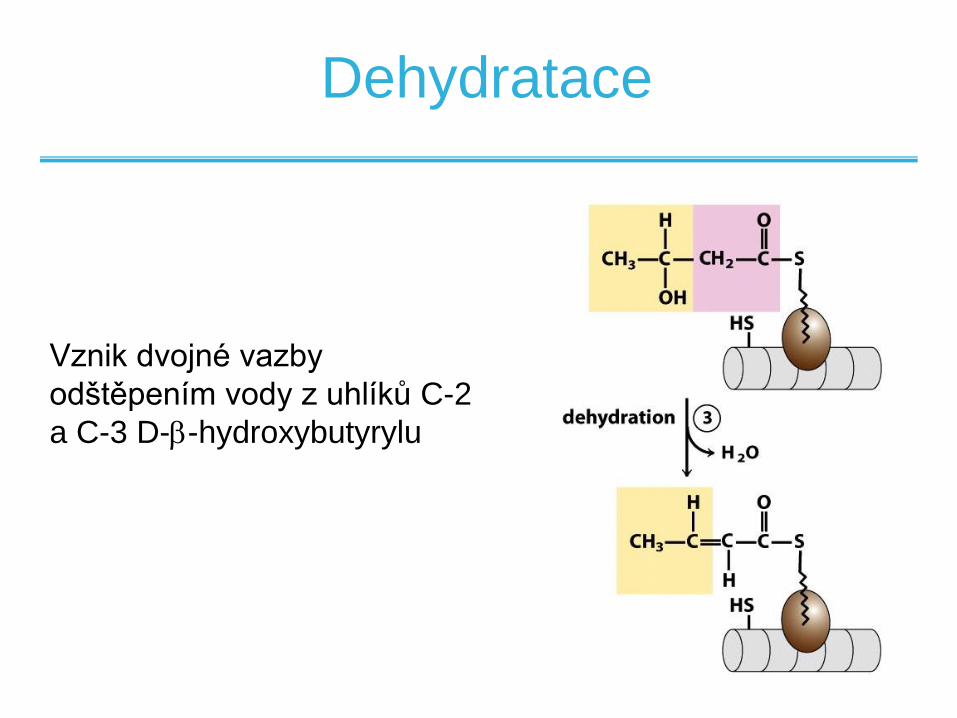
Dehydratace
Vznik dvojné vazby
odštěpením vody z uhlíků C-2
a C-3 D--hydroxybutyrylu

Druhá redukce
Závěrem je redukovaná
(saturovaná) dvojná
vazba za tvorby acyl-
ACP
NADPH je donorem
elektronů

• Acylová skupina je přenesena z ACP na
Syntázu mastných kyselin
• ACP nově váže další malonyl z malonyl-
CoA
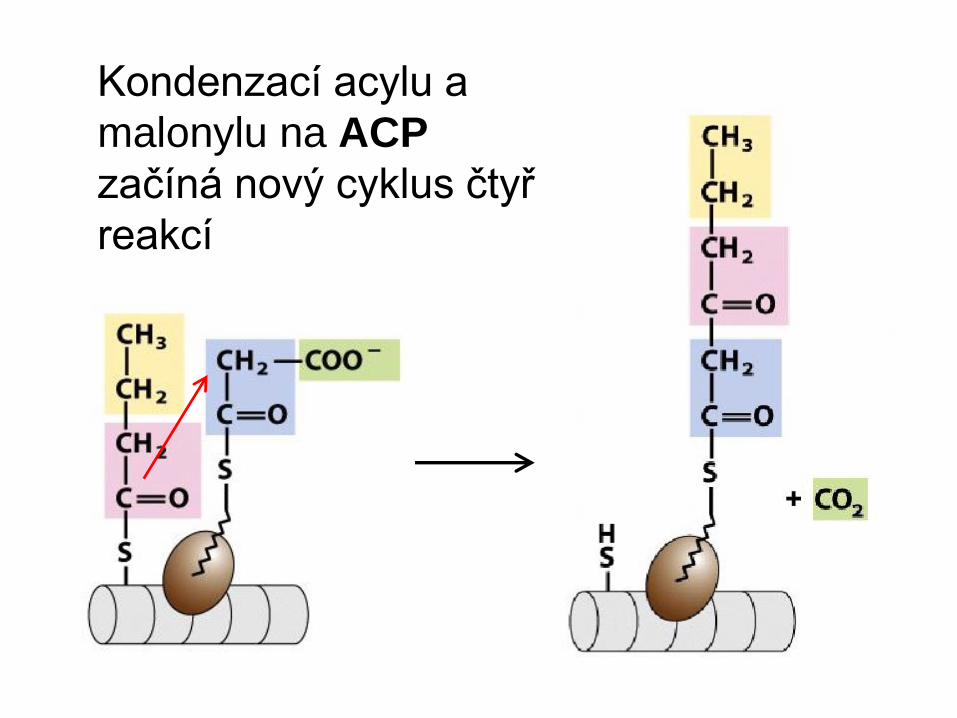
Kondenzací acylu a
malonylu na ACP
začíná nový cyklus čtyř
reakcí

Shrnutí syntézy mastných kyselin
• Cílem je postupné prodlužování uhlíkového řetězce
mastné kyseliny vždy o jeden dvouuhlíkatý zbytek
• Reakce zahrnuje čtyři kroky
– Kondenzace rostousího řetězce s acetátem z malonyl-CoA
– Redukce karbonylu acetátu na hydroxyl
– Dehydratace alkoholu na trans-alken (vznik dvojné vazby)
– Redukce alkenu na alkan
• Rostoucí řetězec je vázaný k enzymu thioesterovou
vazbou
• Při kondenzaci je řetězec přenesen na ACP
• Po druhém redukčním kroku je řetězec přenesen zpět na
Syntázu mastných kyselin
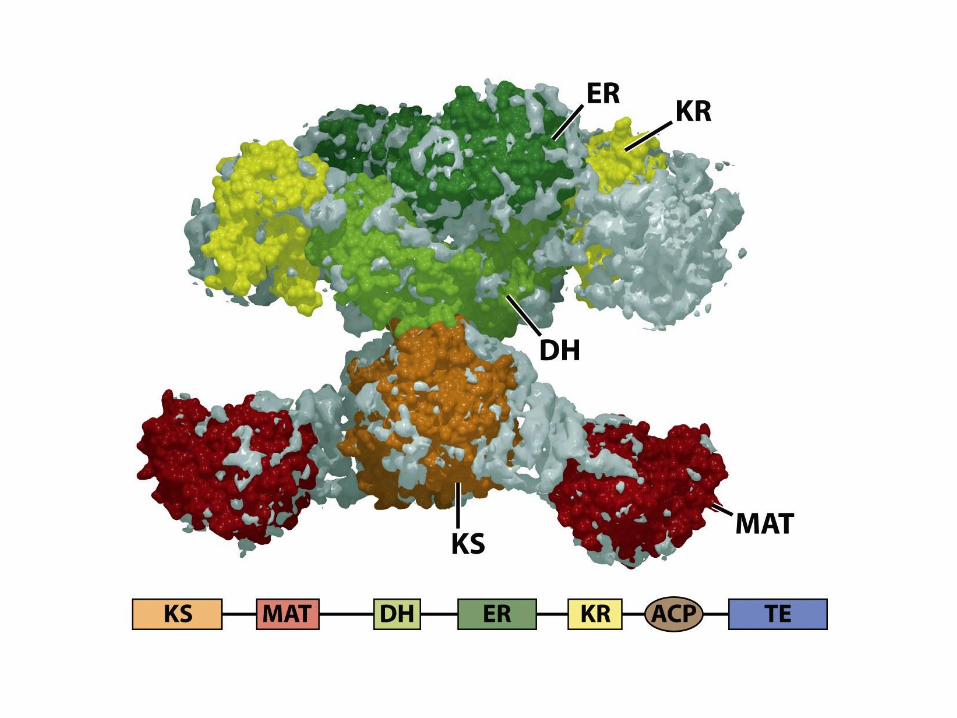

Enzymatic Activities in Fatty
Acid Synthase• Kondenzace s acetátem
– -ketoacyl-ACP syntáza (KS)
• Redukce karbonylu na hydroxyl
– -ketoacyl-ACP reduktáza (KR)
• Dehydratace alkoholu na alken
– -hydroxyacyl-ACP dehydratáza (DH)
• Redukce alkenu na alkan
– enoyl-ACP reduktáza (ER)
• Přenos řetězce
– Malonyl/acetyl-CoA ACP transferáza
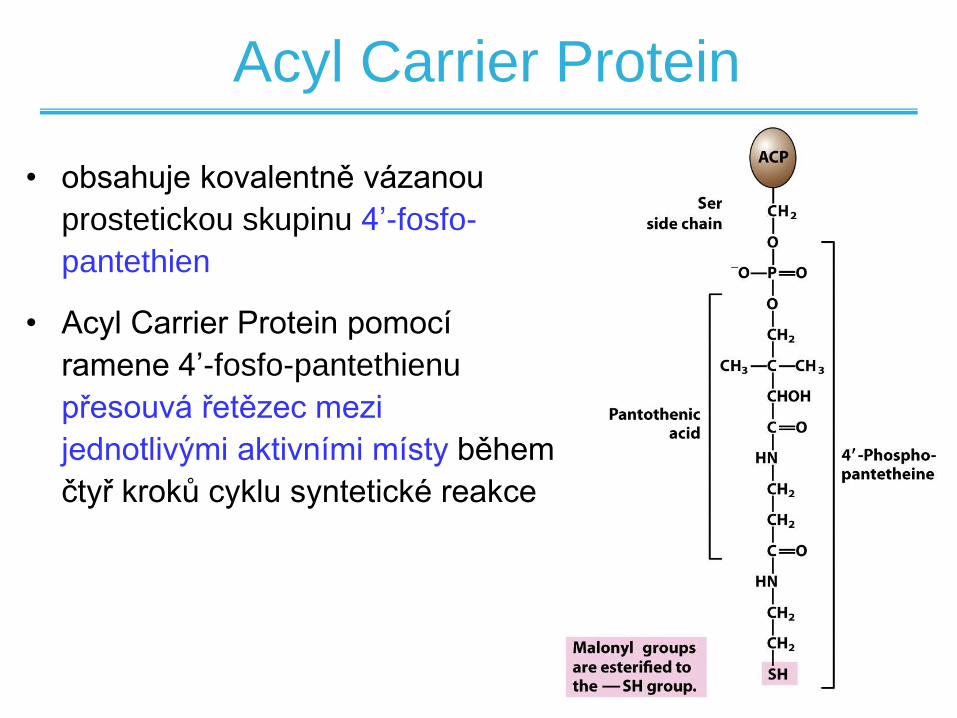
Acyl Carrier Protein
• obsahuje kovalentně vázanou
prostetickou skupinu 4’-fosfo-
pantethien
• Acyl Carrier Protein pomocí
ramene 4’-fosfo-pantethienu
přesouvá řetězec mezi
jednotlivými aktivními místy během
čtyř kroků cyklu syntetické reakce
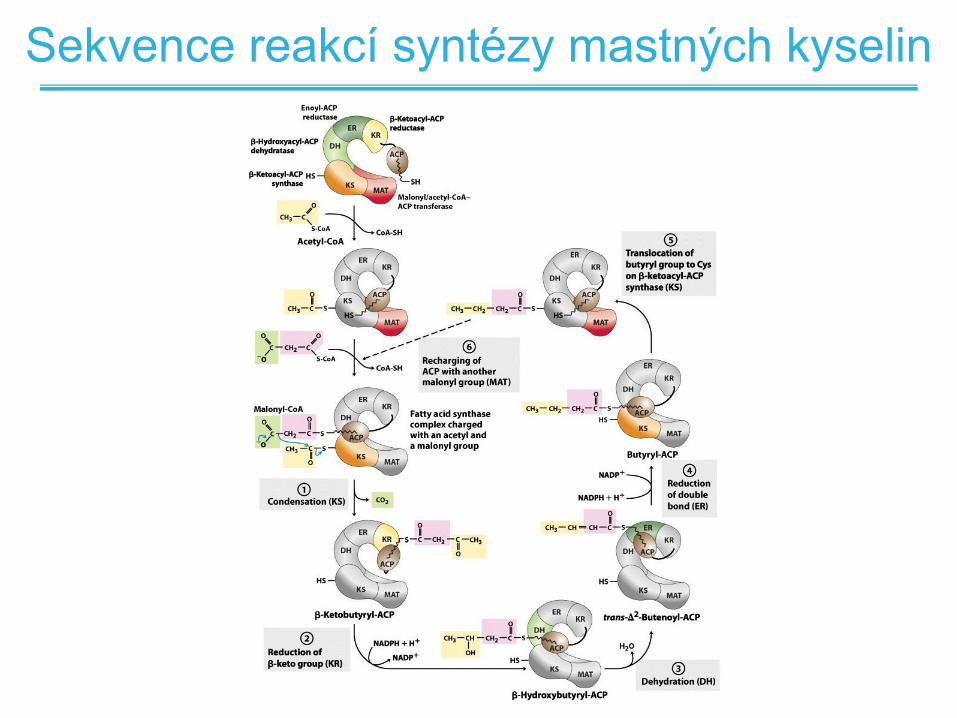
Sekvence reakcí syntézy mastných kyselin

Lokalizace matabolismu mastných kyselin
Katabolismus – Oxidace mastných kyselin
• zvířata - Mitochondrie
• rostliny – Peroxisomy
Anabolismus – Biosyntéza mastných kyselin
• zvířata – Cytosol
• rostliny - Chloroplasty

Syntéza nenasycených mastných kyselin
• Zvířata mohou snadno zavést jednu dvojnou vazbu do
palmitové a stearové kyseliny
• Obratlovci ale nedokáží zavést další dvojnou vazbu mezi
uhlík C10 a methylový konec
• Takové mastné kyseliny, linolová a -linolenová,
přijímáme v potravě jako esenciální mastné kyseliny
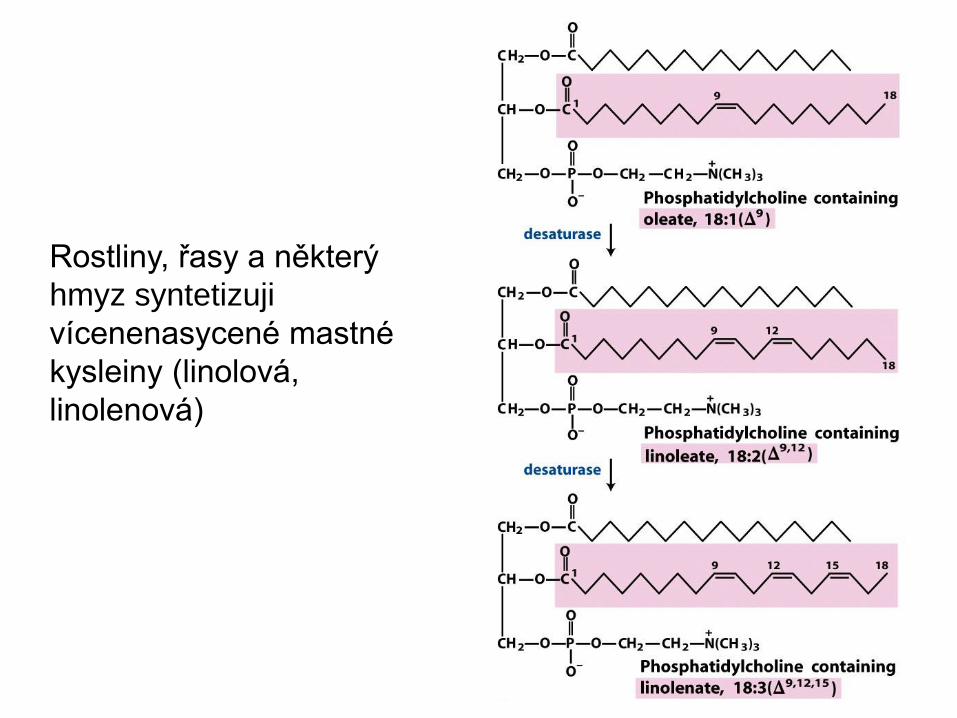
Rostliny, řasy a některý
hmyz syntetizuji
vícenenasycené mastné
kysleiny (linolová,
linolenová)
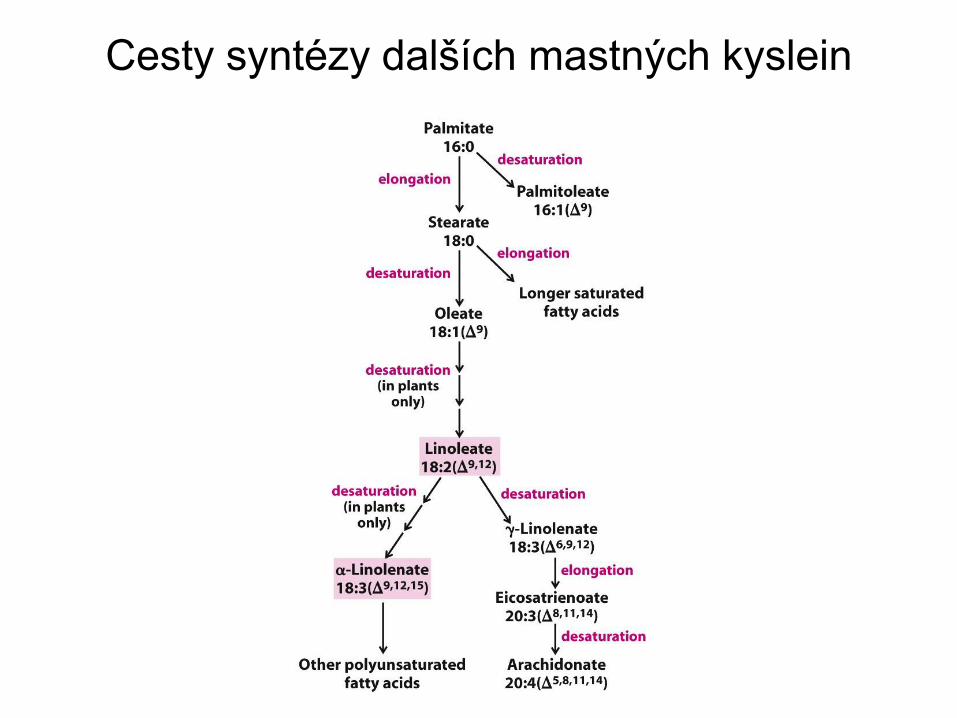
Cesty syntézy dalších mastných kyslein
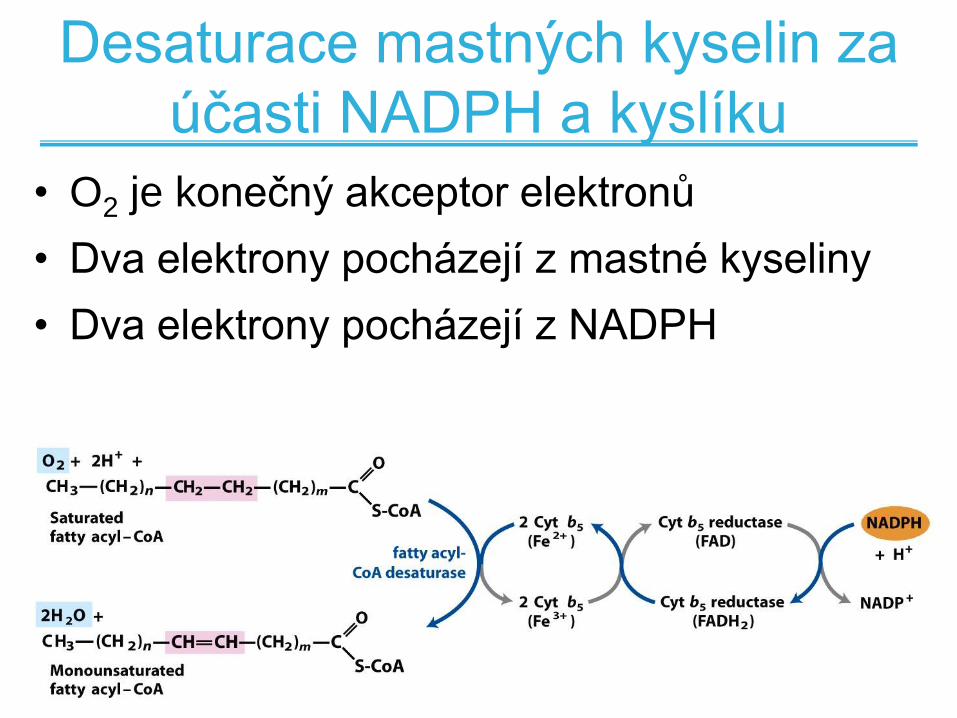
Desaturace mastných kyselin za
účasti NADPH a kyslíku
• O2 je konečný akceptor elektronů
• Dva elektrony pocházejí z mastné kyseliny
• Dva elektrony pocházejí z NADPH
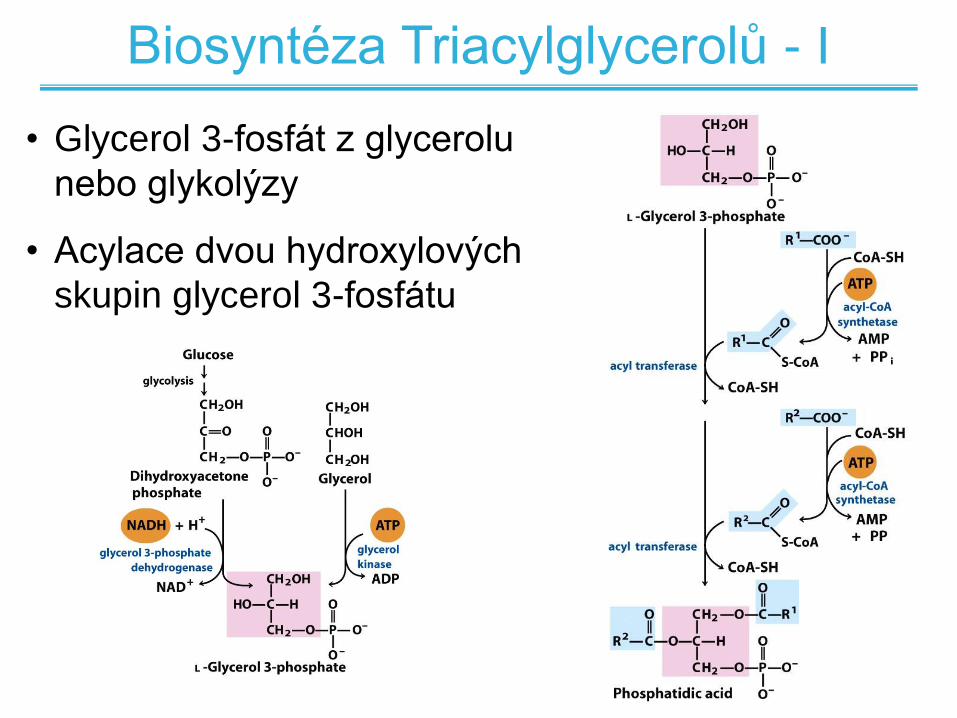
Biosyntéza Triacylglycerolů - I
• Glycerol 3-fosfát z glycerolu
nebo glykolýzy
• Acylace dvou hydroxylových
skupin glycerol 3-fosfátu
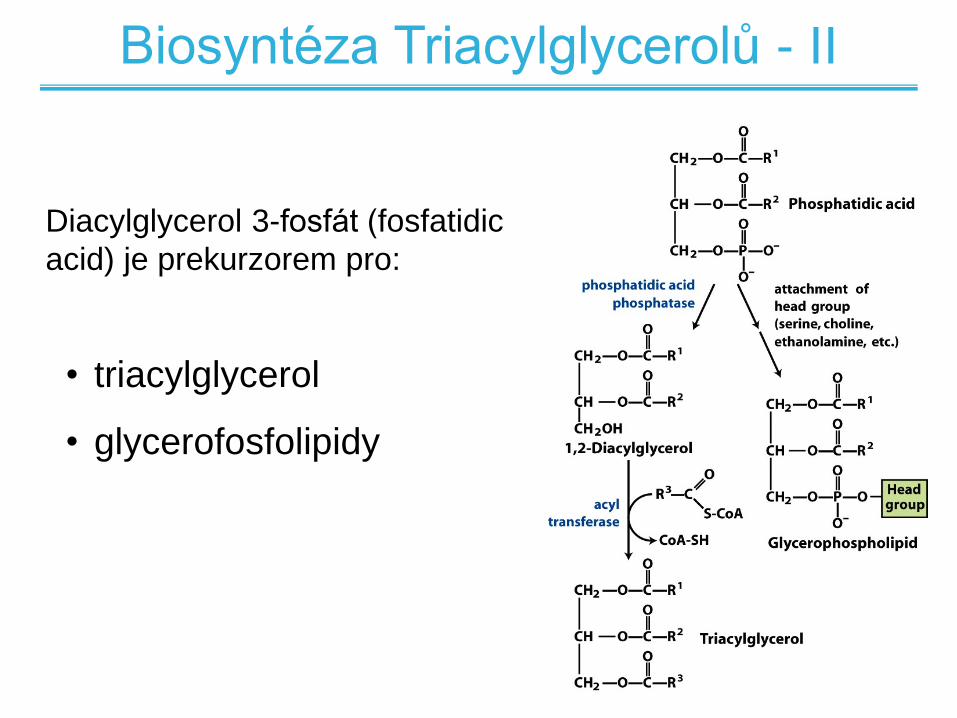
Biosyntéza Triacylglycerolů - II
Diacylglycerol 3-fosfát (fosfatidic
acid) je prekurzorem pro:
• triacylglycerol
• glycerofosfolipidy

- Rozdělení a klasifikace lipidů
- Mastné kyseliny a jejich vlastnosti
- Membránove lipidy
- Biologické membrány funkce, složení, dynamika a vlastnosti
- Membránové proteiny, fluidní model membrány
- Membránový transport
- Metabolismus lipidů, vstřebávání a degradace
- Ketonová tělíska
- Biosyntéza lipidů
Co musím znát
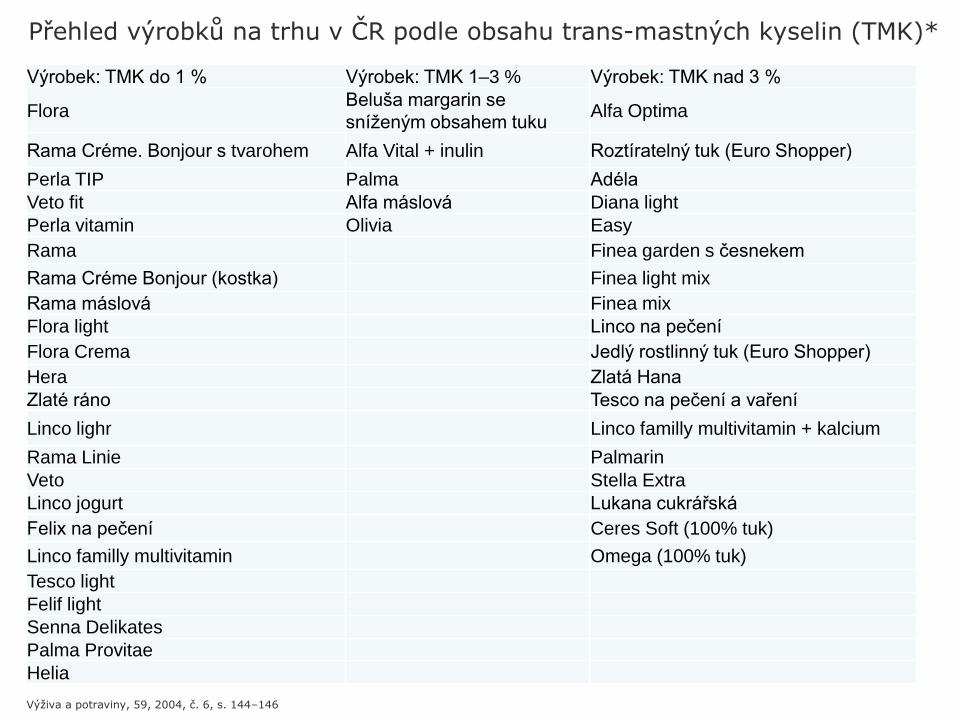
Výrobek: TMK do 1 % Výrobek: TMK 1–3 % Výrobek: TMK nad 3 %
FloraBeluša margarin se
sníženým obsahem tukuAlfa Optima
Rama Créme. Bonjour s tvarohem Alfa Vital + inulin Roztíratelný tuk (Euro Shopper)
Perla TIP Palma Adéla
Veto fit Alfa máslová Diana light
Perla vitamin Olivia Easy
Rama Finea garden s česnekem
Rama Créme Bonjour (kostka) Finea light mix
Rama máslová Finea mix
Flora light Linco na pečení
Flora Crema Jedlý rostlinný tuk (Euro Shopper)
Hera Zlatá Hana
Zlaté ráno Tesco na pečení a vaření
Linco lighr Linco familly multivitamin + kalcium
Rama Linie Palmarin
Veto Stella Extra
Linco jogurt Lukana cukrářská
Felix na pečení Ceres Soft (100% tuk)
Linco familly multivitamin Omega (100% tuk)
Tesco light
Felif light
Senna Delikates
Palma Provitae
Helia
Přehled výrobků na trhu v ČR podle obsahu trans-mastných kyselin (TMK)*
Výživa a potraviny, 59, 2004, č. 6, s. 144–146