lipopolysaccharide activated phosphatidylcholine-specific phospholipase c and induced il-8 and mcp-1...
TRANSCRIPT

Lipopolysaccharide ActivatedPhosphatidylcholine-SpecificPhospholipase C and InducedIL-8 and MCP-1 Production inVascular Endothelial CellsLU ZHANG,1,2 HAI YING LI,1,2 HUI LI,3 JING ZHAO,1,2 LE SU,1,2 YUN ZHANG,2
SHANG LI ZHANG,1,2 AND JUN YING MIAO1,2*1Institute of Developmental Biology, School of Life Science, Shandong University, Jinan, China2The Key Laboratory of Cardiovascular Remodeling and Function Research,
Chinese Ministry of Education and Chinese Ministry of Health, Shandong University, Qilu Hospital, Jinan, China3Department of Internal Medicine, College of Medicine, East Tennessee State University, Johnson City, Tennessee
Secretion of proinflammatory cytokines by lipopolysaccharide (LPS) activated vascular endothelial cells (VECs) contributes substantiallyto the pathogenesis of several inflammatory diseases such as atherosclerosis and septic shock. However, the mechanisms involved in thisprocess are not well understood. Here, we investigated the role of phosphatidylcholine-specific phospholipase C (PC-PLC) in LPS-induced IL-8 and MCP-1 production in VECs. The results showed that LPS elevated the level of PC-PLC and the production of IL-8 andMCP-1 in Human umbilical vein vascular endothelial cells (HUVECs). Blocking the function of PC-PLC by exploiting the neutralizationantibody of PC-PLC or tricyclodecan-9-yl-xanthogenate (D609), an inhibitor of PC-PLC, significantly inhibited LPS-induced production ofIL-8 andMCP-1 inHUVECs. Furthermore, the in vivo experimental results showed that the levels of PC-PLC, IL-8, andMCP-1 in the aorticendothelium and serum were increased in mice injected with LPS. The increased levels of these molecules were also inhibited by thetreatment with D609. The data suggested that blocking PC-PLC function significantly inhibited LPS-induced IL-8 andMCP-1 production incultured HUVECs and in vivo. PC-PLCmight be a potential target for therapy in inflammation associated-diseases such as atherosclerosis.J. Cell. Physiol. 226: 1694–1701, 2011. � 2010 Wiley-Liss, Inc.
Vascular endothelial cells (VECs) are the first cells exposed toinflammatory mediators circulating in the bloodstream duringinflammation (Gonzalez and Selwyn, 2003). Several studieshave postulated an association between infectious agents andchronic cardiovascular diseases such as atherosclerosis(Fong, 2000). One such agent that targets the endothelium islipopolysaccharide (LPS), a unique glycolipid that makes up theouter leaflet of the outer membrane of Gram-negative bacteria(Reyes and Chang, 2005). The Bruneck study has provided thefirst epidemiologic evidence that circulating LPS constitutesa strong risk factor for the development of atherosclerosis(Wiedermann et al., 1999). LPS induces production of pro-inflammatory chemokines by VECs, which further impacts onatherogenesis (Roth et al., 2007). The endothelial cell-derivedchemokines IL-8 andMCP-1 have been implicated in this processmost strongly (Libby, 2002). In addition, the high level of LPS is amajor cause of Gram-negative septic shock. During septicemia,secretion of IL-8 by LPS-activated VECs plays a critical role ininflammatory responses (Baggiolini et al., 1997). LPS-induced IL-8and MCP-1 production in VECs contributes substantially to thepathogenesis of several inflammatory diseases such asatherosclerosis and septic shock. However, the mechanismsinvolved in this process are not understood completely.
Recently we found that suppression of PC-PLC inhibitedprogressionof preexisting atheroma andpromoted lesion stabilityin apoE�/�mice, suggesting that PC-PLCplays an important role inatherosclerosis (Zhang et al., 2010). It was also reported thatPC-PLC was required for LPS-mediated macrophage activation(Cuschieri et al., 2006). Although accumulating evidence suggeststhe implication of PC-PLC in inflammatory processes, very fewreports have focused on its expression and role in IL-8 andMCP-1production of VECs stimulated by LPS.
Today, a great challenge in our knowledge of this importantenzyme is linked to the fact that no PC-PLC isoforms have yetbeen cloned from mammalian sources, nor has the mechanismby which the enzyme regulates cellular responses been fullyelucidated (Cecchetti et al., 2007). Therefore, exploiting itsinhibitor D609 and neutralization antibody and performing theactivity assay have been the strategies for the study of PC-PLCfunctions in mammalian cells. It has been reported that thepolyclonal antibody from rabbit raised against bacterial (Bacilluscereus) PC-PLC possesses proved selective cross-reactivity
Lu Zhang and Hai Ying Li contributed equally to this work.
Contract grant sponsor: National 973 Research Project;Contract grant number: 2011CB503906.Contract grant sponsor: State Program of National Natural ScienceFoundation of China for Innovative Research Group;Contract grant number: 81021001.Contract grant sponsor: National Natural Science Foundation ofChina;Contract grant numbers: 90813022, 31070735.Contract grant sponsor: The Science and TechnologyDevelopmental Project of Shandong Province;Contract grant numbers: 2008GG10002014, 2008GG10002034.
*Correspondence to: Jun Ying Miao, Institute of DevelopmentalBiology, School of Life Science, Shandong University, Jinan 250100,China. E-mail: [email protected]
Received 15 September 2010; Accepted 12 October 2010
Published online in Wiley Online Library(wileyonlinelibrary.com), 10 November 2010.DOI: 10.1002/jcp.22500
ORIGINAL RESEARCH ARTICLE 1694J o u r n a l o fJ o u r n a l o f
CellularPhysiologyCellularPhysiology
� 2 0 1 0 W I L E Y - L I S S , I N C .

against mammalian PC-PLC (Spadaro et al., 2008). So it ispossible to investigate the changes of PC-PLC at protein level inmammalian cells (Ramoni et al., 2004; Fantuzzi et al., 2008;Spadaro et al., 2008). Furthermore, specific binding inhibitiontests indicated that this antibody was directed against thecatalytic site of a mammalian 66 kDa PC-PLC isoform andspecifically inhibited the enzyme activity (Ramoni et al., 2004;Spadaro et al., 2008). We have used the antibody to neutralizePC-PLC activity in our previous study (Zhang et al., 2010). Inthis study, we used all the three methods to validate the role ofPC-PLC in LPS-induced IL-8 and MCP-1 production in VECs.
Materials and MethodsCell culture and treatment
Human umbilical vein vascular endothelial cells (HUVECs) wereobtained in our laboratory by using themethod of Jaffe et al. (1973).The cells were cultured on gelatin-coated plastic dishes in M199medium (Gibco, Grand Island, NY) supplemented with 20% fetalbovine serum (Hyclone Lab, Inc., Tauranga, New Zealand) and70 ng/ml FGF-2 in a humidified incubator at 378Cwith 5% CO2 andused for experiments at not greater than passage 10. WhenHUVECswere grown to 80% confluency, the cells were stimulatedwith LPS (E. coli serotype 055:B5; Sigma–Aldrich, Spruce Street, St.Louis, MO as described by Nandi et al. (2010)) in the presence of0.5% FBS. D609 (Acros Organics co.), rabbit anti-PC-PLCpolyclonal Abs (was obtained, purified, and characterized aspreviously described (Clark et al., 1986; Ramoni et al., 2004))treatments were performed 1 h prior cell exposure to LPS.
Immunofluorescence
After treatment, the cells were incubated with anti-PC-PLC(dilution 1:200) Abs or anti-IL-8 Abs (200mg/ml, dilution 1:100,Santa Cruz Biotechnology, Santa Cruz, CA) overnight at 48C. Thencells were incubated with Alexa Fluor 488 goat anti-rabbit IgG(dilution 1:200) for 1 h at room temperature and then rinsed in1� PBS. Negative control was added with the respectivenonimmune IgGs (Santa Cruz Biotechnology). The samples werephotographed under a laser scanning confocal microscope (LSCM;Carl Zeiss LSM510, Wiesbaden, Germany). Theimmunofluorescence techniques allow semi-quantitativeevaluation of protein expression (Babcock, 1999). PC-PLC and IL-8levels were quantified using the Leica confocal software andexpressed as relative fluorescence intensity per cell.
Western blot analysis
HUVECs with various treatments were lysed in protein lysis buffer(1% SDS in 25mM Tris–HCl, pH 7.5, 4mM EDTA, 100mM NaCl,1mM PMSF, 10mg/ml leupeptin, and 10mg/ml soybean trypsininhibitor). The protein concentration of cells was determined withthe Coomassie brilliant blue protein assay. Protein samples (40mg/lane) were loaded on a 12% SDS–polyacrylamide gel, separated,and electrophoretically transferred to a nitrocellulose membrane.The membrane was incubated in TBST (10mM Tris–HCl, pH 7.6,and 150mM NaCl, 0.1% Tween-20) containing 2% (w/v) BSA and2% (w/v) skim milk powder for 12 h at 48C. Subsequently themembrane was probed with the primary antibody for 90min atroom temperature, and then incubated with the second antibodyfor 1 h and washed with TBST. Then the membrane was soaked inNi-enhanced 3,30-diaminobenzidine (DAB) solution (total volume:20ml, 0.1M Tris–HCl, pH 7.6, 18ml, DAB 12mg, 0.3%CoCl2 2ml,30% H2O2 20ml) until the protein strip could be visualized. Therelative quantity of proteinswas analyzed byQuantity one softwareand normalized to b-actin levels.
Animal treatment
Male C57BL/6J mice (8 weeks old) were obtained fromDepartment of Laboratory Animal Science, Peking University
Health Science Center (PUHSC), China. C57BL/6J mice wererandomly divided into the following groups. LPS group: micereceived a single intraperitoneal injection of LPS at 6mg/kg. Inseparate experiments, blood samples were obtained from mice at3, 6, 12, and 24 h after injection. D609þ LPS group: mice werepretreated with D609 by intraperitoneal injection with 100ml ofD609 (10mg/kg) dissolved in sterile PBS (pH 7.0). After 15min,mice received an intraperitoneal injection of LPS at 6mg/kg. D609group: mice were intraperitoneally injected with 100ml of D609(10mg/kg) dissolved in PBS. At 3, 6, 12, and 24 h after injection, twomice per time point were killed and blood was collected in each ofthe above-mentioned groups. Control group: animals from thesame batch of C57BL/6J (n¼ 6) mice only receiving PBS treatmentserved as controls. These experiments were repeated three times.Serum was placed at �808C until assayed. Thoracic aortae wererapidly removed after perfusion with ice-cold PBS and immersed inoptimal cutting temperature (OCT) embedding medium (Tissue-Tek). Serial 7-mm-thick cryo-sections were collected every threesections and mounted on poly-D-lysine-coated slides. The animalexperimental protocols in this study were in accordance with theGuide for theCare andUse of Laboratory Animals published by theUS National Institutes of Health (NIH Publication No. 85-23,revised 1996) and was approved by the Animal Care Committee ofShandong University.
Double immunofluorescence
The serial sections were used for double immunofluorescenceanalysis with the following antibodies: anti-mouse endothelial cellmAbs (CD31/PECAM-1, 200mg/ml, dilution 1:200, Santa CruzBiotechnology), rabbit polyclonal anti-PC-PLC Abs (dilution1:200), and anti-MCP-1 Abs (200mg/ml, dilution 1:200, BosterBiological Technology, Wuhan, China). Negative control wasadded with the respective nonimmune IgGs (Santa CruzBiotechnology). After incubation with the appropriate FITC-conjugated and TRITC-conjugated secondary antibodies (dilution1:200, Zhongshan Biological Technology, Beijing, China), thesections were observed using LSCM and all images were analyzedusing the Leica confocal software.
PC-PLC activity assay
For inhibition of PC-PLC activity, 5mg/ml of D609 was selectedbased on our previous experiment results (Zhang et al., 2010).Relative changes of PC-PLC activity were determined in serum andwhole-cell lysates using the method of Wu et al. (1997). PC-PLCactivity was calculated by the formula below:
PC-PLC ðnmol=min=mg proteinÞ¼ ðAsample=min�Ablank=minÞ � F
where F¼ 1,000�V total/(V sample� sample proteinconcentration); Asample or Ablank¼ change in absorbance;V¼ volume in ml.
Enzyme-linked immunosorbent assay (ELISA)
The amount of IL-8 or MCP-1 secreted into the culture mediumwas determined by ELISA using the human IL-8 or MCP-1 kit (R&DSystems), respectively. Mouse KC IL-8 or MCP-1 in serum wasdetermined by ELISA using the mouse IL-8 (USCNLIFE) or MCP-1kit (R&D Systems, Minneapolis, MN), respectively. This assay wasperformed according to the manufacturer’s instructions, and thedata were expressed relative to a standard curve prepared for IL-8and MCP-1.
Statistical analysis
All experiments were performed in duplicate and repeated atleast three times. The results were expressed as the means� SE.
JOURNAL OF CELLULAR PHYSIOLOGY
P C - P L C , I L - 8 , A N D M C P - 1 I N V E C S 1695

Statistical analysis was performed by t-test and differences atP< 0.05 were considered statistically significant.
ResultsLPS increased PC-PLC level in HUVECs
HUVECs were exposed to LPS at various concentrations(0, 0.01, 0.05, 0.1, 0.5, and 1mg/ml) for 12 h. PC-PLC levelincreased in a concentration-dependent manner and peaked ata concentration of LPS 0.1mg/ml (Fig. 1A). Thus, all subsequentexperiments were performed using 0.1mg/ml of LPS in vitro.Western blot analysis allowed detection of a distinct PC-PLCisoform with apparent Mr¼ 66 kDa in total cell lysates of allinvestigated HUVECs, in agreement with previous reports inothermammalian cells (Ramoni et al., 2004; Fantuzzi et al., 2008;Spadaro et al., 2008). The intensity of the PC-PLC band at66 kDa was much higher in LPS treated HUVECs comparedwith normal cells (P< 0.01). D609 at 5mg/ml had no effect onthe high level of PC-PLC in LPS-treated HUVECs (Fig. 1B).
LPS induced PC-PLC relocalization in cytoplasm andstimulated its enzymatic activity in HUVECs
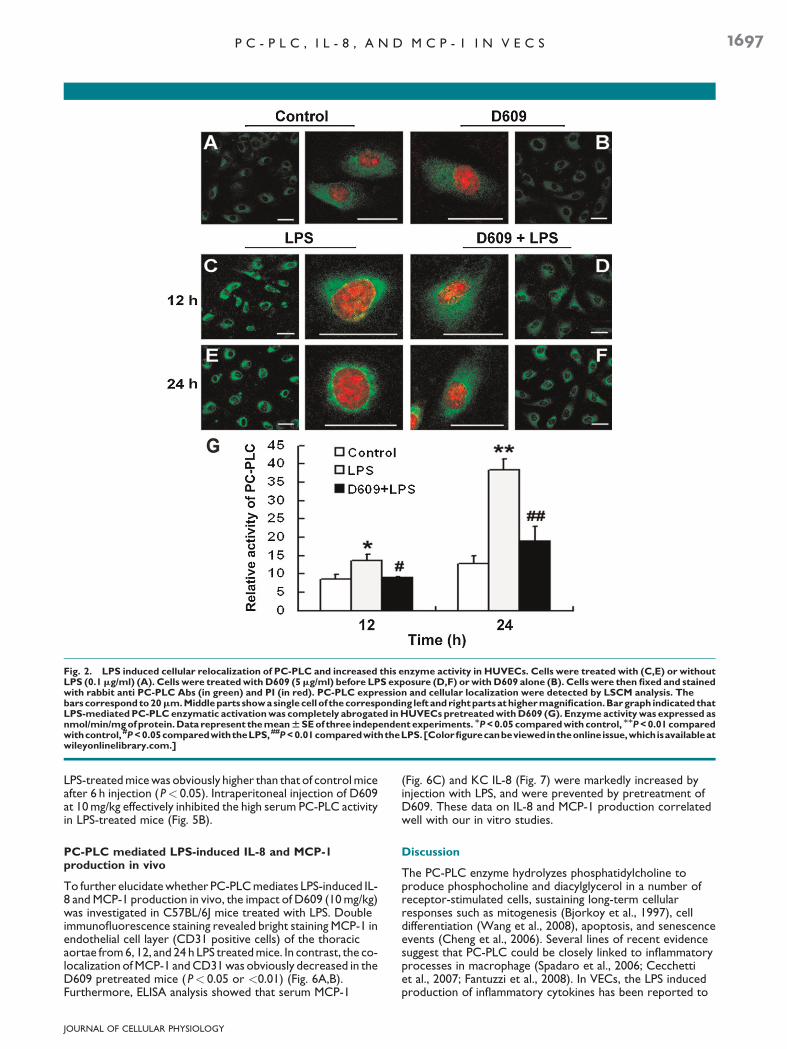
LSCM analysis showed that PC-PLC was expressed in normalHUVECs with a wide distribution throughout cytoplasm(Fig. 2A). Upon 12 h stimulation of HUVECswith LPS, a markedaccumulation of this enzyme mainly confined to a perinuclearregion was clearly detected (Fig. 2C). This effect was retainedafter 12 h (Fig. 2E). Although D609 did not significantly affectPC-PLC distribution in normal cells (Fig. 2B), it inhibited theenzyme accumulation of perinuclear region in LPS-treated cells(Fig. 2D,F).We next investigatedwhether LPS-induced PC-PLCtranslocation to the perinuclear region was associated withchanges in the activity of this enzyme. As shown in Figure 2G,in keeping with the observed effect on PC-PLC translocationto the perinuclear region, a marked increase of enzymaticactivation was observed at 12 h upon LPS exposure and thiseffect continued to 24 h (P< 0.05 or <0.01). LPS-mediated
PC-PLC enzymatic activation was completely abrogated inHUVECs pretreated with D609 (5mg/ml).
Inhibition of PC-PLC activity attenuated LPS-inducedIL-8 and MCP-1 production in HUVECs
Immunofluorescence assay showed that pretreatment withD609 or anti-PC-PLC Abs (40 or 80mg/ml) inhibited LPS-induced IL-8 production in HUVECs (Fig. 3A,B), and thiseffect was confirmed by Western blot experiments (Fig. 3D).Moreover, we evaluated the involvement of PC-PLC in LPS-mediated IL-8 release by ELISA. Pretreatment with D609 oranti-PC-PLC Abs significantly (P< 0.01 or <0.05) inhibitedLPS-induced IL-8 release from HUVECs, thus confirming theinhibition of functional IL-8 production (Fig. 3C).
ELISA assay also indicated that inhibition of PC-PLC byD609or anti-PC-PLC Abs (40 or 80mg/ml) suppressed the LPS-induced MCP-1 production (Fig. 4). Nonspecific IgG had noeffect on the production of IL-8 and MCP-1.
LPS increased PC-PLC level in aortic endothelial cellsand its enzymatic activity in serum of C57BL/6J mice
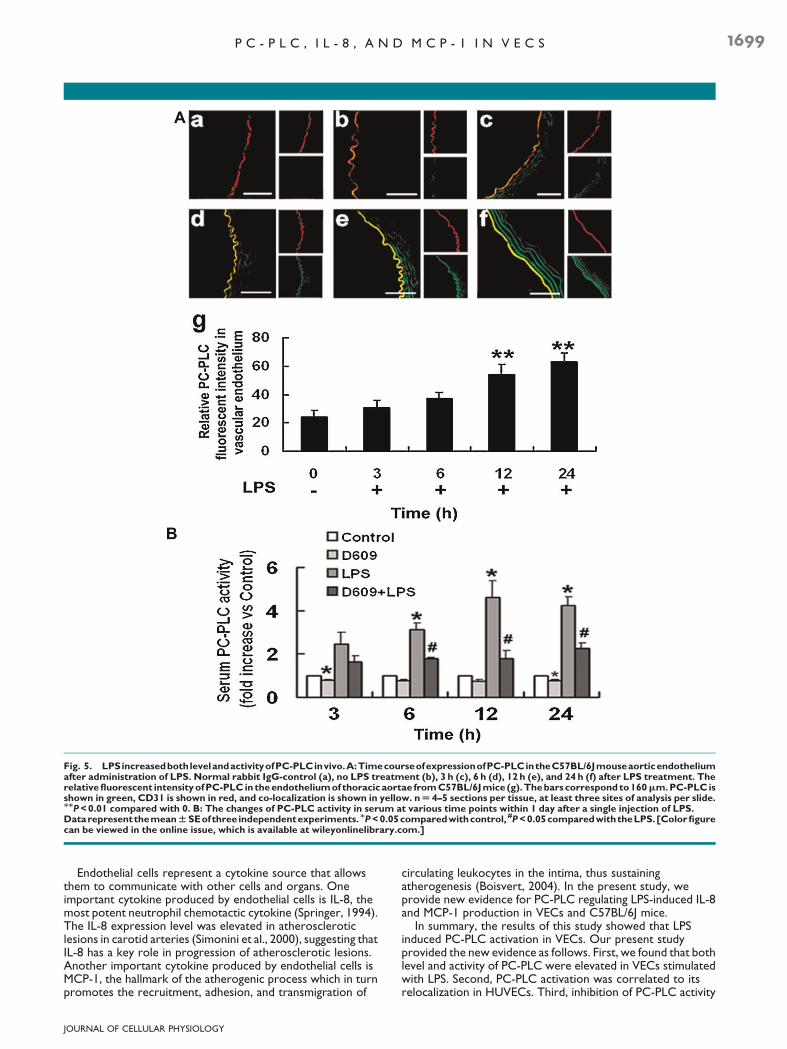
To determine if the increase in PC-PLC level observed incultured HUVECs occurs in vivo, we injected sterile PBS or LPS(6mg/kg) into the peritoneal cavity of C57BL/6J mice. The doseof LPS was chosen, as it induced a pronounced systemicinflammatory reaction, yet did not cause changes in animalbehavior or early mortality. In the absence of treatment withLPS, PC-PLC was expressed at low levels in aortic endothelialcells (Fig. 5A-b), norwere changes found in 3 h post-LPS animals(Fig. 5A-c). However, by 6 h after receiving LPS, strongimmunoreactivity was detected for PC-PLC in the endotheliumof the thoracic aortae of C57BL/6J mice (Fig. 5A-d). Theincrease of PC-PLC levels in aortic endothelial cells were moreprominent at 12 and 24 h (Fig. 5A-e,f) after the LPS injection.PC-PLC enzymatic activity was also measured in all serumsamples. As shown in Figure 5B, the mean value of serumPC-PLC activity of control mice was normalized to 1.0. In thisarbitrary unit scale, the mean value of serum PC-PLC activity of
Fig. 1. LPS increased the level of PC-PLC inHUVECs. A: HUVECswere stimulatedwith various concentrations of LPS (0.01–1mg/ml) for 12h.We observed that LPS increased PC-PLC level in a dose-dependent manner (a–f). The bars correspond to 20mm. Median values of PC-PLCfluorescence intensity in HUVECs (g). Data represent the meanWSE of three independent experiments. MP<0.05 compared with control,MMP<0.01 compared with control. B: The bands of proteins fromWestern blot assay and PC-PLC levels were normalized to the levels of b-actin.Data represent the meanWSE of three independent experiments. MP<0.05 compared with control, MMP<0.01 compared with control.
JOURNAL OF CELLULAR PHYSIOLOGY
1696 Z H A N G E T A L .
LPS-treatedmicewas obviously higher than that of controlmiceafter 6 h injection (P< 0.05). Intraperitoneal injection of D609at 10mg/kg effectively inhibited the high serum PC-PLC activityin LPS-treated mice (Fig. 5B).
PC-PLC mediated LPS-induced IL-8 and MCP-1production in vivo
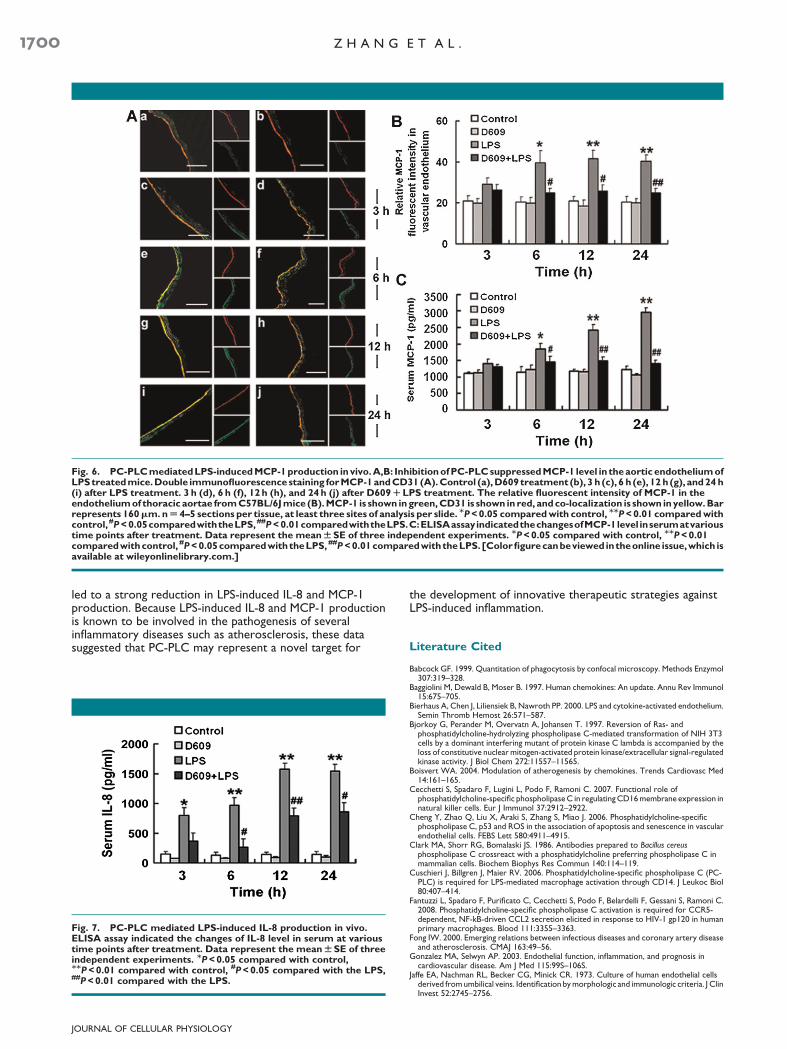
To further elucidatewhether PC-PLCmediates LPS-induced IL-8 andMCP-1 production in vivo, the impact of D609 (10mg/kg)was investigated in C57BL/6J mice treated with LPS. Doubleimmunofluorescence staining revealed bright staining MCP-1 inendothelial cell layer (CD31 positive cells) of the thoracicaortae from6, 12, and 24 h LPS treatedmice. In contrast, the co-localization ofMCP-1 andCD31was obviously decreased in theD609 pretreated mice (P< 0.05 or <0.01) (Fig. 6A,B).Furthermore, ELISA analysis showed that serum MCP-1
(Fig. 6C) and KC IL-8 (Fig. 7) were markedly increased byinjection with LPS, and were prevented by pretreatment ofD609. These data on IL-8 and MCP-1 production correlatedwell with our in vitro studies.
Discussion
The PC-PLC enzyme hydrolyzes phosphatidylcholine toproduce phosphocholine and diacylglycerol in a number ofreceptor-stimulated cells, sustaining long-term cellularresponses such as mitogenesis (Bjorkoy et al., 1997), celldifferentiation (Wang et al., 2008), apoptosis, and senescenceevents (Cheng et al., 2006). Several lines of recent evidencesuggest that PC-PLC could be closely linked to inflammatoryprocesses in macrophage (Spadaro et al., 2006; Cecchettiet al., 2007; Fantuzzi et al., 2008). In VECs, the LPS inducedproduction of inflammatory cytokines has been reported to
Fig. 2. LPS induced cellular relocalization of PC-PLC and increased this enzyme activity in HUVECs. Cells were treated with (C,E) or withoutLPS (0.1mg/ml) (A). Cells were treatedwith D609 (5mg/ml) before LPS exposure (D,F) orwith D609 alone (B). Cells were then fixed and stainedwith rabbit anti PC-PLC Abs (in green) and PI (in red). PC-PLC expression and cellular localization were detected by LSCM analysis. Thebarscorrespondto20mm.Middleparts showasinglecellof thecorresponding left andrightpartsathighermagnification.Bargraph indicated thatLPS-mediatedPC-PLCenzymatic activationwas completely abrogated inHUVECs pretreatedwithD609 (G). Enzyme activitywas expressed asnmol/min/mgofprotein.Data represent themeanWSEof three independentexperiments. MP<0.05 comparedwith control, MMP<0.01comparedwithcontrol, #P<0.05comparedwiththeLPS,##P<0.01comparedwiththeLPS.[Colorfigurecanbeviewedintheonlineissue,whichisavailableatwileyonlinelibrary.com.]
JOURNAL OF CELLULAR PHYSIOLOGY
P C - P L C , I L - 8 , A N D M C P - 1 I N V E C S 1697

contribute significantly to the inflammatory process (Bierhauset al., 2000). However, the involvement of PC-PLC in thisprocess is poorly understood.
It must be mentioned here that D609 is the only availablecompound acting as a competitive PC-PLC inhibitor till now(Jaffe et al., 1973; Bjorkoy et al., 1997; Spadaro et al., 2006;Cecchetti et al., 2007; Fantuzzi et al., 2008). However, D609was reported to affect PC-PLD activity in NIH 3T3 fibroblasts(Kiss and Tomono, 1995). It is noteworthy that 5mg/mL D609has been demonstrated specifically inhibited PC-PLC enzymaticactivity and did not affect PC-PLD activity in HUVECs in ourprevious studies (Zhang et al., 2010). Our data suggestedthat PC-PLC activation and cellular localization are the integralpart of a strictly regulated machinery of cell function control,indicating that the mechanisms of PC-PLC activation andtranslocation may be coupled in HUVECs. Western blotanalysis showed the intensity of PC-PLC band was muchhigher in LPS treated HUVECs compared with normal cells.Furthermore, intraperitoneal injection of LPS increased PC-PLC level in aortic endothelial cells and its enzymatic activity inserum of C57BL/6J mice, suggesting that LPS actually activatedVECs through increasing both level and activity of PC-PLC.
Fig. 3. Inhibition of PC-PLC activation blocked LPS-induced IL-8 production inHUVECs.A: Effect of D609 or anti-PC-PLCAbs on LPS-inducedIL-8 production in HUVECs by immunofluorescence assay combined with LSCM. After pretreatment, the cells were cultured without or withLPS in basal M199mediumwith 0.5% FBS for 24 h. Control (a), LPS (b), D609 (c), D609RLPS (d), Nonspecific IgG (80mg/ml, e), IgG80RLPS (f),anti-PC-PLCAb (40mg/ml, g), anti-PC-PLCAb40RLPS (h), anti-PC-PLCAb (80mg/ml, i), and anti-PC-PLCAb80RLPS (j). Thebars correspondto20mm.B:Median valuesof IL-8fluorescence intensity inHUVECs.Data represent themeanWSEof three independentexperiments. MMP<0.01compared with control, #P<0.05 compared with the LPS, ##P<0.01 compared with the LPS. C: IL-8 secretion from HUVECs was measuredby ELISA. Data represent the meanWSE of three independent experiments. MMP<0.01 compared with control, #P<0.05 compared with theLPS, ##P<0.01 compared with the LPS. D: Western blot analysis of the level of IL-8 in cell extracts. IL-8 levels were normalized to the levels ofb-actin. Data represent the meanWSE of three independent experiments. MP<0.05 compared with control, #P<0.05 compared with the LPS.[Color figure can be viewed in the online issue, which is available at wileyonlinelibrary.com.]
Fig. 4. ELISA assay indicated that inhibition of PC-PLC by D609 oranti-PC-PLC Abs suppressed MCP-1 secretion from HUVECs. Datarepresent the meanWSE of three independent experiments.MMP<0.01 compared with control, #P<0.05 compared with the LPS,##P<0.01 compared with the LPS.
JOURNAL OF CELLULAR PHYSIOLOGY
1698 Z H A N G E T A L .
Endothelial cells represent a cytokine source that allowsthem to communicate with other cells and organs. Oneimportant cytokine produced by endothelial cells is IL-8, themost potent neutrophil chemotactic cytokine (Springer, 1994).The IL-8 expression level was elevated in atheroscleroticlesions in carotid arteries (Simonini et al., 2000), suggesting thatIL-8 has a key role in progression of atherosclerotic lesions.Another important cytokine produced by endothelial cells isMCP-1, the hallmark of the atherogenic process which in turnpromotes the recruitment, adhesion, and transmigration of
circulating leukocytes in the intima, thus sustainingatherogenesis (Boisvert, 2004). In the present study, weprovide new evidence for PC-PLC regulating LPS-induced IL-8and MCP-1 production in VECs and C57BL/6J mice.
In summary, the results of this study showed that LPSinduced PC-PLC activation in VECs. Our present studyprovided the new evidence as follows. First, we found that bothlevel and activity of PC-PLC were elevated in VECs stimulatedwith LPS. Second, PC-PLC activation was correlated to itsrelocalization in HUVECs. Third, inhibition of PC-PLC activity
Fig. 5. LPSincreasedbothlevelandactivityofPC-PLCinvivo.A:TimecourseofexpressionofPC-PLCintheC57BL/6Jmouseaorticendotheliumafter administration of LPS. Normal rabbit IgG-control (a), no LPS treatment (b), 3 h (c), 6 h (d), 12 h (e), and 24 h (f) after LPS treatment. Therelativefluorescent intensityofPC-PLC in theendotheliumof thoracicaortae fromC57BL/6Jmice (g).Thebarscorrespondto160mm.PC-PLC isshown in green, CD31 is shown in red, and co-localization is shown in yellow. nU 4–5 sections per tissue, at least three sites of analysis per slide.MMP<0.01 compared with 0. B: The changes of PC-PLC activity in serum at various time points within 1 day after a single injection of LPS.DatarepresentthemeanWSEofthree independentexperiments.MP<0.05comparedwithcontrol, #P<0.05comparedwiththeLPS.[Colorfigurecan be viewed in the online issue, which is available at wileyonlinelibrary.com.]
JOURNAL OF CELLULAR PHYSIOLOGY
P C - P L C , I L - 8 , A N D M C P - 1 I N V E C S 1699
led to a strong reduction in LPS-induced IL-8 and MCP-1production. Because LPS-induced IL-8 and MCP-1 productionis known to be involved in the pathogenesis of severalinflammatory diseases such as atherosclerosis, these datasuggested that PC-PLC may represent a novel target for
the development of innovative therapeutic strategies againstLPS-induced inflammation.
Literature Cited
Babcock GF. 1999. Quantitation of phagocytosis by confocal microscopy. Methods Enzymol307:319–328.
Baggiolini M, Dewald B, Moser B. 1997. Human chemokines: An update. Annu Rev Immunol15:675–705.
Bierhaus A, Chen J, Liliensiek B, Nawroth PP. 2000. LPS and cytokine-activated endothelium.Semin Thromb Hemost 26:571–587.
Bjorkoy G, Perander M, Overvatn A, Johansen T. 1997. Reversion of Ras- andphosphatidylcholine-hydrolyzing phospholipase C-mediated transformation of NIH 3T3cells by a dominant interfering mutant of protein kinase C lambda is accompanied by theloss of constitutive nuclear mitogen-activated protein kinase/extracellular signal-regulatedkinase activity. J Biol Chem 272:11557–11565.
Boisvert WA. 2004. Modulation of atherogenesis by chemokines. Trends Cardiovasc Med14:161–165.
Cecchetti S, Spadaro F, Lugini L, Podo F, Ramoni C. 2007. Functional role ofphosphatidylcholine-specific phospholipaseC in regulatingCD16membrane expression innatural killer cells. Eur J Immunol 37:2912–2922.
Cheng Y, Zhao Q, Liu X, Araki S, Zhang S, Miao J. 2006. Phosphatidylcholine-specificphospholipase C, p53 and ROS in the association of apoptosis and senescence in vascularendothelial cells. FEBS Lett 580:4911–4915.
Clark MA, Shorr RG, Bomalaski JS. 1986. Antibodies prepared to Bacillus cereusphospholipase C crossreact with a phosphatidylcholine preferring phospholipase C inmammalian cells. Biochem Biophys Res Commun 140:114–119.
Cuschieri J, Billgren J, Maier RV. 2006. Phosphatidylcholine-specific phospholipase C (PC-PLC) is required for LPS-mediated macrophage activation through CD14. J Leukoc Biol80:407–414.
Fantuzzi L, Spadaro F, Purificato C, Cecchetti S, Podo F, Belardelli F, Gessani S, Ramoni C.2008. Phosphatidylcholine-specific phospholipase C activation is required for CCR5-dependent, NF-kB-driven CCL2 secretion elicited in response to HIV-1 gp120 in humanprimary macrophages. Blood 111:3355–3363.
Fong IW. 2000. Emerging relations between infectious diseases and coronary artery diseaseand atherosclerosis. CMAJ 163:49–56.
Gonzalez MA, Selwyn AP. 2003. Endothelial function, inflammation, and prognosis incardiovascular disease. Am J Med 115:99S–106S.
Jaffe EA, Nachman RL, Becker CG, Minick CR. 1973. Culture of human endothelial cellsderived fromumbilical veins. Identification bymorphologic and immunologic criteria. J ClinInvest 52:2745–2756.
Fig. 6. PC-PLCmediatedLPS-inducedMCP-1production invivo.A,B: InhibitionofPC-PLCsuppressedMCP-1 level in theaortic endotheliumofLPStreatedmice.Double immunofluorescencestaining forMCP-1andCD31(A).Control (a),D609treatment (b), 3 h (c), 6 h (e), 12 h(g), and24 h(i) after LPS treatment. 3 h (d), 6 h (f), 12 h (h), and 24h (j) after D609RLPS treatment. The relative fluorescent intensity of MCP-1 in theendotheliumof thoracic aortae fromC57BL/6Jmice (B).MCP-1 is shown ingreen,CD31 is shown in red, andco-localization is shown inyellow.Barrepresents 160mm.nU 4–5 sections per tissue, at least three sites of analysis per slide. MP<0.05 comparedwith control, MMP<0.01 comparedwithcontrol,#P<0.05comparedwiththeLPS,##P<0.01comparedwiththeLPS.C:ELISAassayindicatedthechangesofMCP-1level inserumatvarioustime points after treatment. Data represent the meanWSE of three independent experiments. MP<0.05 compared with control, MMP<0.01comparedwithcontrol, #P<0.05comparedwiththeLPS,##P<0.01comparedwiththeLPS. [Colorfigurecanbeviewed intheonline issue,which isavailable at wileyonlinelibrary.com.]
Fig. 7. PC-PLC mediated LPS-induced IL-8 production in vivo.ELISA assay indicated the changes of IL-8 level in serum at varioustime points after treatment. Data represent the meanWSE of threeindependent experiments. MP<0.05 compared with control,MMP<0.01 compared with control, #P<0.05 compared with the LPS,##P<0.01 compared with the LPS.
JOURNAL OF CELLULAR PHYSIOLOGY
1700 Z H A N G E T A L .

Kiss Z, TomonoM. 1995. Compound D609 inhibits phorbol ester-stimulated phospholipaseD activity and phospholipase C-mediated phosphatidylethanolamine hydrolysis. BiochimBiophys Acta 1259:105–108.
Libby P. 2002. Inflammation in atherosclerosis. Nature 420:868–874.Nandi D, Mishra MK, Basu A, Bishayi B. 2010. Protective effects of interleukin-6 inlipopolysaccharide (LPS)-induced experimental endotoxemia are linked to alteration inhepatic anti-oxidant enzymes and endogenous cytokines. Immunobiology 215:443–451.
Ramoni C, Spadaro F, Barletta B, Dupuis ML, Podo F. 2004. Phosphatidylcholine-specificphospholipase C in mitogen-stimulated fibroblasts. Exp Cell Res 299:370–382.
Reyes CL, Chang G. 2005. Structure of the ABC transporter MsbA in complex with ADPvanadate and lipopolysaccharide. Science 308:1028–1031.
RothGA,Moser B, Roth-Walter F, GiaconaMB, Harja E, Papapanou PN, Schmidt AM, Lalla E.2007. Infectionwith a periodontal pathogen increasesmononuclear cell adhesion to humanaortic endothelial cells. Atherosclerosis 190:271–281.
Simonini A, MoscucciM,Muller DW, Bates ER, Pagani FD, BurdickMD, Strieter RM. 2000. IL-8 is an angiogenic factor in human coronary atherectomy tissue. Circulation 101:1519–1526.
Spadaro F,Cecchetti S, SanchezM,AusielloCM, Podo F, RamoniC. 2006. Expression and roleof phosphatidylcholine-specific phospholipase C in human NK and T lymphocyte subsets.Eur J Immunol 36:3277–3287.
Spadaro F, Ramoni C, Mezzanzanica D, Miotti S, Alberti P, Cecchetti S, Iorio E, Dolo V,Canevari S, Podo F. 2008. Phosphatidylcholine-specific phospholipase C activation inepithelial ovarian cancer cells. Cancer Res 68:6541–6549.
Springer TA. 1994. Traffic signals for lymphocyte recirculation and leukocyte emigration: Themultistep paradigm. Cell 76:301–314.
Wang N, Sun C, Huo S, Zhang Y, Zhao J, Zhang S, Miao J. 2008. Cooperation ofphosphatidylcholine-specific phospholipase C and basic fibroblast growth factor in theneural differentiation of mesenchymal stem cells in vitro. Int J Biochem Cell Biol 40:294–306.
Wiedermann CJ, Kiechl S, Dunzendorfer S, Schratzberger P, Egger G, Oberhollenzer F,Willeit J. 1999. Association of endotoxemia with carotid atherosclerosis andcardiovascular disease: Prospective results from the Bruneck Study. J Am Coll Cardiol34:1975–1981.
Wu X, Lu H, Zhou L, Huang Y, Chen H. 1997. Changes of phosphatidylcholine-specificphospholipase C in hepatocarcinogenesis and in the proliferation and differentiation of ratliver cancer cells. Cell Biol Int 21:375–381.
Zhang L, Zhao J, Su L, Huang B, Wang L, Su H, Zhang Y, Zhang S, Miao J. 2010. D609 inhibitsprogression of preexisting atheroma and promotes lesion stability in apolipoprotein e�/�
mice: A role of phosphatidylcholine-specific phospholipase in atherosclerosis. ArteriosclerThromb Vasc Biol 30:411–418.
JOURNAL OF CELLULAR PHYSIOLOGY
P C - P L C , I L - 8 , A N D M C P - 1 I N V E C S 1701