liu & krieger page 1 in vitro assay of purified sr-bi highly purified
TRANSCRIPT

Liu & Krieger page 1 In vitro assay of purified SR-BI
Highly Purified Scavenger Receptor Class B, Type I Reconstituted into Phosphatidylcholine
/Cholesterol Liposomes Mediates High Affinity HDL Binding and Selective Lipid Uptake
Bin Liu¥*, and Monty Krieger§
From the Department of Biology, Massachusetts Institute of Technology,Cambridge, MA, 02139 USA,
Running title: In vitro assay of purified SR-BI
This work was supported by grant HL52212 from the National Institutes of Health.
¥ Postdoctoral Fellow of the National Institutes of Health, Heart Lung and Blood Institute*Current address: NeoGenesis Pharmaceuticals, Inc. Cambridge, MA 02139
§ To whom correspondence should be addressed:
Monty Krieger
Room 68-483
Biology Department
Massachusetts Institute of Technology
Cambridge, MA 02139
tel: 617-253-6793
fax: 617-258-5851; e-mail: [email protected]
Copyright 2002 by The American Society for Biochemistry and Molecular Biology, Inc.
JBC Papers in Press. Published on July 10, 2002 as Manuscript M204265200 by guest on February 14, 2018
http://ww
w.jbc.org/
Dow
nloaded from

Liu & Krieger page 2 In vitro assay of purified SR-BI
SUMMARY
The murine class B, type I scavenger receptor mSR-BI is a high and low density
lipoprotein (HDL and LDL) receptor that mediates selective uptake of cholesteryl
esters. Here we describe a reconstituted phospholipid/cholesterol-liposome assay of
the binding and selective uptake activities of SR-BI derived from detergent
solubilized cells. The assay, employing lysates from epitope tagged receptor (mSR-
BI-t1) expressing mammalian and insect cells, recapitulated many features of SR-BI
activity in intact cells, including high affinity and saturable 125I-HDL binding,
selective lipid uptake from [3H]cholesteryl ether-labeled HDL, and poor inhibition
of HDL receptor activity by LDL. The novel properties of a mutated receptor
(Q402R/Q418R: normal LDL binding but loss of most HDL binding) were reproduced
in the assay, as was the ability of the SR-BI homologue CD36 to bind HDL but not
mediate efficient lipid uptake. In this assay, essentially homogeneously pure mSR-
BI-t1 - prepared by single-step immunoaffinity chromatography - mediated high
affinity HDL binding and efficient selective lipid uptake from HDL. Thus, SR-BI-
mediated HDL binding and selective lipid uptake are intrinsic properties of the
receptor that do not require the intervention of other proteins or specific cellular
structures or compartments.
by guest on February 14, 2018http://w
ww
.jbc.org/D
ownloaded from

Liu & Krieger page 3 In vitro assay of purified SR-BI
INTRODUCTION
The LDL receptor pathway for the delivery of lipoprotein cholesterol to cells involves
clathrin coated pit-mediated endocytosis and subsequent lysosomal degradation of the
entire LDL particle (1). Almost 20 years ago a strikingly different mechanism for the
cellular uptake of lipoprotein cholesterol, called selective cholesterol uptake, was
identified during the analysis of HDL metabolism in vivo (2,3). Selective cholesterol
uptake from HDL and other lipoproteins does not involve endocytosis and subsequent
degradation of the entire lipoprotein particle (2,3, reviewed in (4) and (5)). In the case of
HDL, the lipoprotein binds to the cell surface, transfers its cholesteryl esters to the cell,
and then the lipid-depleted HDL particle dissociates from the cell and can re-enter the
circulation.
The HDL receptor SR-BI (scavenger receptor, class B, type I) was the first cell
surface receptor to be shown to mediate physiologically relevant selective lipid uptake
((6-8), reviewed in (5)). In vivo studies have established that SR-BI critically influences
HDL structure and metabolism and apparently plays an important role in the transport of
cholesterol from peripheral tissues to the liver for recycling or biliary excretion (5,7-12).
This probably accounts for the ability of SR-BI to protect against atherosclerosis in
murine models (9,13-15). Recent studies have established that expression of SR-BI in
mice is normally required for red blood cell development (16) and female fertility (9,17),
and can prevent the development of myocardial infarctions, cardiac dysfunction and
premature death in apoE-deficient mice (18).
by guest on February 14, 2018http://w
ww
.jbc.org/D
ownloaded from

Liu & Krieger page 4 In vitro assay of purified SR-BI
In vitro studies have shown that SR-B can bind and mediate lipid uptake from
LDL as well as HDL (6,19-22). Strikingly, HDL competes efficiently for LDL binding,
whereas LDL is a poor competitor of HDL binding (6,23). SR-BI also can facilitate the
efflux of unesterified cholesterol from cultured cells (24,25), although the physiologic
significance of this is not certain. Several studies support the proposal (26) that SR-BI-
mediated transport of lipids between cells and lipoproteins involves two sequential steps:
1) productive lipoprotein binding and 2) binding-dependent, yet distinct, SR-BI-mediated
lipid transfer (25-29). It has not yet been determined if SR-BI-mediated selective lipid
uptake occurs only at the cell surface, or in some intracellular compartment followed by
retroendocytosis (secretion) of the lipid-depleted lipoprotein, or both ((30-32), reviewed
in (4) and (5)).
A particularly important question regarding the mechanism of SR-BI activity has been:
does SR-BI require the participation of another protein(s) to mediate either ligand
binding, lipid transport, or both, or are these activities autonomous properties of SR-BI
(independent of other proteins)? This question has arisen, in part, because of the multiple
and complex activities of SR-BI (5). A direct approach for studying the autonomous
properties of SR-BI and its mechanism of action is to examine in an in vitro system the
activity of the receptor purified away from other proteins. Here we describe an in vitro
reconstituted liposome assay for SR-BI-mediated ligand binding and selective lipid
uptake that reflects many of the characteristics of SR-BI activity in intact cells. This
assay, effective with total cell lysates as well as highly purified protein, was used to show
that, in the absence of other proteins, SR-BI can bind HDL and LDL and mediate
efficient selective cholesteryl ether uptake from HDL.
by guest on February 14, 2018http://w
ww
.jbc.org/D
ownloaded from

Liu & Krieger page 5 In vitro assay of purified SR-BI
EXPERIMENTAL PROCEDURES
Materials
Human high density lipoprotein (HDL), human low density lipoprotein (LDL),
125I-labeled HDL (125I-HDL), 125I-labeled LDL (125I-LDL), 1,1’-dioctadecyl-3,3,3’,3’-
tetramethylindocarbocyanime perchlorate (DiI)-labeled HDL (DiI-HDL), and human
lipoprotein deficient serum (HLPDS) were prepared as previously described (6,19). The
phCD36 expression vector (33) was a generous gift from B. Seed (Massachusetts General
Hospital). The KKB-1 antibody was a generous gift from Karen Kozarsky (25). The 1D4
antibody was obtained from the ATCC. The mixture of complete protease inhibitors was
purchased from Roche. All other reagents were purchased from standard suppliers or
obtained as indicated below. Cell culture supplies were purchased from GIBCO/BRL,
Irvine Scientific or JRH Biosciences. The peptide TETSQVAPA was prepared in the
biopolymer lab at MIT and was a gift from the G. Khorana laboratory.
[3H]CE-HDL Preparation
[3H]Cholesteryl ether ([3H]CE)-labeled HDL ([3H]CE-HDL) was prepared
according to the procedure of Rodrigueza et al. (34) with minor modifications. The
labeled [3H]CE-HDL was isolated by ultracentrifugation (225,000xg), filtered through a
0.22 µm membrane and the protein concentration was determined using the method of
Lowry et al. (35).
by guest on February 14, 2018http://w
ww
.jbc.org/D
ownloaded from

Liu & Krieger page 6 In vitro assay of purified SR-BI
Construction of Expression Vectors
Mammalian expression vectors: mSR-BI cDNA was amplified from pmSR-BI (6) by
PCR using the primers BL5,
5’GACACTGGTACCGATATCACGCGGACATGGGCGGCAGCTCCAG-3’, and BL3,
5’CTGTCACTCGAGGTCGACTTAGGCAGGCGCCACTTGGCTGGTCTCTGTTAGC
TTGGCTTCTTGCAGCACCGTG-3’ that resulted in a cDNA encoding a full length
mSR-BI protein that contained a 9-amino acid (TETSQVAPA) C-terminal extension
representing the C-terminus of bovine rhodopsin as an epitope tag. The protein product of
this cDNA is designated ‘mSR-BI-t1’. The PCR product was treated with restriction
endonucleases Kpn1 and XhoI, and the KpnI/XhoI fragment was cloned into the
mammalian expression vector pcDNA3(+) (Invitrogen) that includes an internal
neomycin resistance marker for selection in mammalian cells. The ligation product was
used to transform E. coli cells and the plasmid DNAs of selected clones were isolated and
sequenced. One of these with the expected sequence was designated pmSR-BI-t1. The
mSR-BI-t1 cDNA was then reconstructed into an expression plasmid (36) using a SalI
and EcoRV restriction fragment from pmSR-BI-t1. This plasmid was designated
pACmSR-BI-t1 and was used for the generation of the stable cell line HEK[mSR-BI-t1]
(see below).
Sf21 expression vectors: Three receptors were expressed in sf21 cells at high levels using
the Bac-To-Bac baculovirus expression system (GIBCO/BRL). These were mSR-BI-t1, a
double substitution mutant of mSR-BI-t1 (arginines in place of the glutamines at
positions 402 and 418 (designated ‘402R/418R’)) that retains most of the LDL but little
by guest on February 14, 2018http://w
ww
.jbc.org/D
ownloaded from

Liu & Krieger page 7 In vitro assay of purified SR-BI
of the HDL receptor activity of the parent receptor (23), and human CD36. Briefly,
pFastBac donor plasmids, were constructed by ligation of the restriction endonucleases
XhoI and EcoRI-digested fragments of pmSR-BI-t1 or the PCR products for CD36 or
mutant mSR-BI-t1 into XhoI/StuI-treated pFastBac vector (GIBCO-BRL). The template
and PCR primers for CD36 were: phCD36 (33), 5’
CGAGGATATCGGCAAGAAACAGGTGC – 3’ and 5’
GCAGCTCGAGGTCACAAGTACATC – 3’. The template and PCR primers for the
mutant SR-BI-t1 402R/418R were: VM54 (23),
5’GACACTGGTACCGATATCACGCGGACATGGGCGGCAGCTCCAG-3’, and
5’CTGTCACTCGAGGTCGACTTAGGCAGGCGCCACTTGGCTGGTCTCTGTTAGC
TTGGCTTCTTGCAGCACCGTG-3’. Recombinant bacmids were generated by
transformation of DH10Bac E. coli cells with the cloned donor plasmids, selection of
antibiotic (kanamycin/tetracycline/gentamicin) resistant colonies, and isolation of
plasmid DNA from the cultures of the selected colonies (37).
Mammalian Cell Culture and Transfection
The mammalian cell lines HEK293S ((38), gift from P. Reeves and G. Khorana)
and COS M6 were maintained attached to the substratum in medium A (Dulbecco’s
modified Eagle’s medium: Ham’s F12 medium (1:1) supplemented with 50 units/ml
penicillin, 50 µg/ml streptomycin, 2 mM glutamine (Gibco BRL) and 10% fetal bovine
serum (JRH Biosciences)) in a humidified 95% air, 5% CO2 incubator. For suspension
culture, HEK293S cells were grown with constant gentle stirring at 20-40 rpm in spinner
bottles in medium B (HBGro medium (Irvine Scientific) supplemented with 50 units/ml
by guest on February 14, 2018http://w
ww
.jbc.org/D
ownloaded from

Liu & Krieger page 8 In vitro assay of purified SR-BI
penicillin, 50 µg/ml streptomycin, 2 mM glutamine and 10% fetal bovine serum) at 37 °C
in a humidified incubator in 5% CO2/95% air. For suspension growth, cells were
inoculated at 1.2x105 cells/ml in 500 ml of medium B and incubated for 5-7 days without
changing the medium. Cells were harvested either after reaching confluence on dishes by
treatment with trypsin/EDTA (JRH Biosciences) or after 6 days of growth in suspension.
For transient transfections, 1.5 x 106 COS M6 cells were seeded in 10-cm dishes
in medium A without antibiotics and grown at 37oC overnight. The cells were treated
with 10 µg per plate of DNA (pmSR-BI-t1 or the ‘empty’ vector pcDNA3 without an
expression cassette insert as a control) using the LIPOFECTAMINE (GIBCO/BRL)
method according to the manufacturer’s recommendations. The cells were grown in
medium A for an additional two days and then receptor activity assays were performed.
Briefly, 24 hours post-transfection, the cells were washed with PBS, harvested with
trypsin/EDTA and plated in 1 ml/well of medium A at 150,000 cells/well in 24-well
plates. The receptor activity of the cells was analyzed 48 hours post-transfection.
A stable cell line expressing high levels of mSR-BI-t1, HEK[mSR-BI-t1] (clone
7), was established as follows: HEK293S cells were transfected with the pACmSR-BI-t1
vector using the LIPOFECTAMINE method and maintained for 2 weeks in medium A
supplemented with 0.8 mg/ml G418. Individual colonies were isolated by screening for
their abilities to take up the fluorescent dye DiI from DiI-HDL (10 µg protein/ml, 2 hrs at
37°C) with an inverted fluorescence microscope and by flow cytometry as previously
described (26).
by guest on February 14, 2018http://w
ww
.jbc.org/D
ownloaded from

Liu & Krieger page 9 In vitro assay of purified SR-BI
Insect (sf) Cell Culture and Baculovirus Expression
The sf21 cells (gift from K. Cha, E. Getmanova and G. Khorana), were
maintained in SFM II medium (GIBCO/BRL) in suspension at 27oC in a humidified
incubator and passaged when the viable cell count reached 2 x 106 cells/ml. The
recombinant bacmid DNAs described above, encoding mSR-BI-t1, the 402R/418R
mutant form of mSR-BI-t1 and CD36, were used to transfect sf21 cells in T25 flasks
using the Bac-to-Bac expression system (GIBCO/BRL) according to the manufacturer’s
recommendations. After growth of the transfected sf21 cells for 5 days at 27oC in Grace’s
medium, the media were collected as the recombinant baculovirus particle stocks and
used for infection of sf21 cells (procedures carried out according to the manufacturer’s
recommendations). The infected cells were grown in Grace’s medium for 72-168 hours
and harvested by centrifugation. High level expression of the proteins was verified by
SDS-PAGE and immunoblotting analysis of cell extracts prepared as described below
using polyclonal anti-SR-BI antibody KKB-1 or anti-CD36 antibody (PharminGen).
Cells also were infected with an ‘empty’ vector to permit generation of receptor-free cell
extract controls. Cell lysates were prepared (see below) and used for immunoblot
analysis and liposome reconstitution studies.
Determination of Receptor Cell Surface Expression and Function in Intact Mammalian
Cells
Cell surface expression: Transiently transfected COS cells were incubated with
polyclonal antisera (KKB-1) against mSR-BI, washed, incubated with fluorescein
by guest on February 14, 2018http://w
ww
.jbc.org/D
ownloaded from

Liu & Krieger page 10 In vitro assay of purified SR-BI
(FITC)-conjugated goat anti-rabbit IgG (Amersham), washed again, harvested with
trypsin and receptor cell surface expression was determined by flow cytometry as
previously described (23).
Cell function analysis: The 125I-HDL binding assay was performed as previously
described (6). The [3H]CE-HDL cell association assay was similar to the 125I-HDL
binding assay, except for the following changes: the NaOH cell lysis step was preceded
by the addition of 1 ml of isopropanol at room temperature for 30 minutes to extract the
incorporated [3H]cholesteryl ether from the cells. Radioactivity in the isopropanol extract
was measured using a liquid scintillation analyzer (Packard Instrument Co., Meriden,
CT). The amount of [3H]cholesteryl ether associated with the cells (or liposomes in the
assay described below) is expressed as the equivalent amount of [3H]CE-HDL protein
(ng). This standard method of presenting the data (e.g., see (26)) permits a direct
comparison of the relative amounts of 125I-HDL binding and lipid uptake from [3H]CE-
HDL and clearly indicates the extent of selectivity of the lipid uptake process.
Preparation of Total Cell Lysates
HEK[mSR-BI-t1] and untransfected HEK293S cells were plated at 1-2x105
cells/plate in 10-cm plates and grown in 15 ml of medium A and grown for 3-4 days.
When the cells were confluent, the plates were washed twice with PBS, and the cells
were harvested by scraping with a rubber policeman and concentrated into a pellet by
centrifugation at 4oC at 1460xg for 20 minutes in a SS34 rotor in a Sorvall RC-5B
centrifuge (DuPont Instruments). The cell pellets were dissolved by adding 1 ml of lysis
buffer A (1.5% (w/v) octyl glucoside and protease inhibitors (1x) in PBS) per 0.1 gr (wet
by guest on February 14, 2018http://w
ww
.jbc.org/D
ownloaded from

Liu & Krieger page 11 In vitro assay of purified SR-BI
weight) pellet and incubating at 4oC for 30 minutes with gentle mixing. The lysates were
clarified by centrifugation at 11,951xg using a SS34 rotor for 20 minutes at 4oC, and the
supernatants were collected and used as total cell lysates. Protein concentration was
determined by the DC protein assay (BioRad) and was typically 1-2 mg/ml. The same
procedure was used to prepare lysates from Sf 21 cells.
Immunoaffinity Purification of mSR-BI-t1
mSR-BI-t1 was purified by immunoaffinity chromatography using a modification
of the procedure of Reeves et al. (38). Briefly, 1.5 gram (wet weight) of HEK[mSR-BI-
t1] cells were incubated with 13 ml of lysis buffer A at 4oC for 30 min. The solution was
clarified by centrifugation (4oC, 11,951xg, 20 minutes), and filtration (0.45 µM Durapore
membrane (Millipore)). The subsequent purification steps were carried out at room
temperature. Five ml of the extract were loaded onto a 0.3 ml anti-C-terminal 1D4
antibody-Sepharose column, which was pre-equilibrated with lysis buffer A. After
washing with 50 column-volumes of lysis buffer A, and 10 column-volumes of column
buffer A (10 mM Tris (pH 6.0), 1.5% octyl glucoside and protease inhibitors), bound
proteins were eluted with 2 ml of elution buffer (column buffer A containing 100 µM of
the nonapeptide epitope (TETSQVAPA), 0.3 ml fractions). Samples (10 µl) were
fractionated by SDS-PAGE and analyzed by either silver staining or immunoblotting
with the anti-SR-BI antibody KKB-1. The purified protein was stored in the elution
buffer at –20oC.
by guest on February 14, 2018http://w
ww
.jbc.org/D
ownloaded from

Liu & Krieger page 12 In vitro assay of purified SR-BI
Liposome Preparation
The preparation of multilamelar vesicles was carried out according to Schneider
et al. (39) with minor modifications. Briefly, egg yolk L-a-phosphatidylcholine (egg PC)
(Avanti) and cholesterol (Sigma) (molar ratio of 5:1) in ether were dried and resuspended
at 2 mg/ml in 50 mM Tris buffer (pH 6.0). Then 0.5 ml of the suspension and 2.4 ml of
buffer B (50 mM Tris, pH 6.0, 150 mM NaCl, 2 mM CaCl2) containing either 100 µg of
detergent solubilized cell lysate or 2 µg of detergent solubilized purified mSR-BI-t1 were
mixed, precipitated by 0.6 volumes of ice cold acetone and the precipitate recovered by
centrifugation (4oC at 30,596xg, 20 min.). The liposomes were resuspended in 300 µl of
buffer C (20 mM Tris pH 8.0, 1 mM CaCl2, 20 mM NaCl and protease inhibitors).
Filter Binding Assay
Liposomes (8 µl) were diluted with 12 µl of assay buffer (20 mM Tris pH 8.0, 2
mM CaCl2, 80 mg/ml fat-free BSA (Sigma), and protease inhibitors (1x)) and the
indicated amounts of 125I- HDL or [3H]CE-HDL (usually 10 µg protein/ml) in the
presence (single incubations) or absence (duplicate incubations) of a 40-fold excess of
unlabeled HDL. The mixture was incubated at 37oC for 2 hours unless otherwise noted,
and the liposomes with bound 125I-HDL were isolated by filtration (39) using 0.45 µM
nitrocellulose membranes and a multifilter filtration manifold (Millipore, Milford, MA).
Briefly, the filters in the manifold were moistened with wash buffer (20 mM Tris pH 8.0,
50 mM NaCl, 20 µM CaCl2 and 1 mg/ml fatty acid-free BSA), the assay mixture was
filtered at room temperature and then the filters were washed three times with wash
buffer. The amounts of 125I bound were measured with a gamma counter (LKB-Wallac,
by guest on February 14, 2018http://w
ww
.jbc.org/D
ownloaded from

Liu & Krieger page 13 In vitro assay of purified SR-BI
Finland). To measure 3H, the filters were added to 4 ml of Hydrofluor scintillation fluid
(National Diagnostics) and radioactivity was measured using a liquid scintillation
analyzer (Packard Instrument Co., Meriden, CT).
by guest on February 14, 2018http://w
ww
.jbc.org/D
ownloaded from

Liu & Krieger page 14 In vitro assay of purified SR-BI
RESULTS
The two goals of this study were 1) to develop an in vitro reconstituted liposome
assay of SR-BI-mediated ligand binding and lipid uptake and 2) to use the assay to
determine if highly purified SR-BI required the cooperation of other proteins to mediate
HDL binding and selective lipid uptake or if it could do so independently of other
proteins. To address the later goal, we developed a modification of the method of Reeves
et al. (38), developed for the analysis of rhodopsin, to isolate essentially homogeneously
pure and functional mSR-BI containing an exogenous epitope tag.
Comparison of Binding and Uptake Activities of Cells Transfected with Plasmid DNA of
Wild Type and Tagged mSR-BI
To facilitate isolation of pure mSR-BI protein for reconstitution into liposomes,
we slightly modified the approach described by Reeves, Thurmond and Khorana for the
generation and purification of recombinant bovine rhodopsin from transfected
mammalian cells (38). Reeves et al. expressed bovine rhodopsin in HEK293S cells and
purified the detergent solubilized protein using monoclonal antibody affinity purification
and epitope peptide elution (38). The anti-rhodopsin antibody 1D4 recognizes the C-
terminal 9-aa peptide from rhodopsin (40). For the synthesis and purification of mSR-BI
we constructed a mammalian cell expression vector, pmSR-BI-t1, that encodes a
chimeric protein (mSR-BI-t1) containing the full-length mSR-BI and, at its C-terminus,
the C-terminal epitope-tag TETSQVAPA from bovine rhodopsin. To determine if epitope
tagging interfered with the activity of the receptor, we transiently transfected COS M6
cells with expression vectors encoding either wild type mSR-BI (COS[mSR-BI]),
by guest on February 14, 2018http://w
ww
.jbc.org/D
ownloaded from

Liu & Krieger page 15 In vitro assay of purified SR-BI
epitope-tagged mSR-BI-t1 (COS[mSR-BI-t1]) or an empty vector (pcDNA) control
(COS[control]), and measured the two best defined activities of SR-BI (5): binding of
125I-HDL and uptake of cholesteryl ether from [3H]CE-HDL. The values for receptor-
specific binding or lipid uptake are defined as the differences between total binding or
uptake and the values determined in the presence of a 40-fold excess of unlabeled ligand
(nonspecific values). The lipid uptake data are presented as [3H]CE-HDL protein
equivalents (ng of [3H]CE-HDL protein that contain the amount of [3H]cholesteryl ether
associated with the cells). Figure 1 shows that the specific 125I-HDL binding (panel A) or
[3H]cholesteryl ether uptake (panel C) values for mSR-BI and mSR-BI-t1 were similar.
The relative levels of surface expression of the wild-type and tagged receptors were
determined using an anti-SR-BI antibody (KKB-1) and quantitative flow cytometry (25).
There was a somewhat higher level expression of the tagged receptor (1.2-fold). When
the 125I-HDL binding and [3H]cholesteryl ether uptake values for the wild-type receptor
were normalized to account for this difference, the binding and lipid uptake curves for the
two receptors were virtually identical (Figure 1B & 1D). The ratios of the maximal
[3H]CE-HDL uptake/125I-HDL binding, expressed as equivalent ng of HDL protein/mg of
cell protein, were 47 and 44, respectively, clearly showing that both receptors mediated
selective lipid uptake. Therefore, the epitope tagging at the C-terminus of mSR-BI did
not appear to alter the key activities of mSR-BI in transfected COS cells, and this tagged
mSR-BI should be useful for the isolation of purified SR-BI (see below) to study its
activity in a reconstituted system.
by guest on February 14, 2018http://w
ww
.jbc.org/D
ownloaded from

Liu & Krieger page 16 In vitro assay of purified SR-BI
Reconstituted Liposome Assay for the Cell-free Analysis of SR-BI Function
We have developed a reconstituted liposome filter binding assay for measuring
the binding of 125I-HDL and the uptake of [3H]cholesteryl ether from [3H]CE-HDL
mediated by SR-BI derived from detergent solubilized cultured cells based on the LDL
receptor binding assay of Schneider et al. (39) (see Experimental Procedures). In brief,
phosphatidylcholine/cholesterol vesicles were prepared using detergent solubilized whole
cell lysates or detergent solubilized purified receptor protein. After incubation with
radiolabeled lipoprotein (standard conditions: 5-10 µg protein/ml of lipoprotein for 2 hrs
at 37°C, variations indicated below), the liposome associated radioactivity was
determined by ultrafiltration and counting. Standard controls included: preparation of
liposomes either with lysates from cells that do not express recombinant receptor or
without added cell lysate or purified receptor; and incubation with a 40-fold excess of
unlabeled lipoprotein to compete for the specific binding of the ligands to the receptor.
The nonspecific background binding of 125I-HDL varied from 19-50% of the total binding
and appeared to depend critically on the quality of the preparation, e.g., extent of
radiolytic decomposition of the HDL, which depends on the age and specific activity of
the preparations and can occur rapidly (41). The absolute values for binding and lipid
uptake constants (apparent Kd and Bmax (binding maximum) and Umax (uptake
maximum)) also varied somewhat between different receptor preparations and depended
on the quality of the radiolabeled lipoprotein (41).
by guest on February 14, 2018http://w
ww
.jbc.org/D
ownloaded from

Liu & Krieger page 17 In vitro assay of purified SR-BI
Validation of the Reconstituted Liposome Assay
The reconstituted liposome assay was initially validated using cell lysates from
insect cells (Sf21) expressing high levels of tagged receptor (not shown) due to infection
with a baculovirus encoding mSR-BI-t1. Previous studies have shown that insect cells
can express on their surfaces functional mammalian SR-BI (42,43). Figure 2A shows the
ligand concentration dependence of the binding of 125I-HDL (squares) and the uptake of
[3H]cholesteryl ether from [3H]CE-HDL (circles) by mSR-BI-t1-containing insect cell
lysate-reconstituted liposomes (open symbols) and control liposomes prepared with
lysates from cells infected with baculovirus without the insertion (control, filled
triangles). Specific 125I-HDL binding to the mSR-BI-t1-containing liposomes (open
squares) was high affinity (apparent Kd ~21 µg protein/ml, similar to that seen in mSR-
BI expressing mammalian cells (6,41)) and saturable (Bmax ~15 ng protein/assay),
whereas there was very little specific binding to the mSR-BI-t1-negative control
liposomes (filled triangles). Similar results were obtained for the uptake of
[3H]cholesteryl ether from [3H]CE-HDL (open circles, apparent Kd ~11 µg protein/ml,
Umax ~106 ng protein/assay); the maximal value for lipid uptake was seven-times
greater than that for binding, indicating selective uptake.
Figure 3 shows the time dependence of 125I-HDL binding (open squares) and
[3H]cholesteryl ether uptake from [3H]CE-HDL (open circles) by mSR-BI-t1-containing
insect cell lysate-reconstituted liposomes at 37°C. The 125I-HDL binding reached a steady
state after about 1 hour, while the [3H]cholesteryl ether uptake increased until
approximately three hours of incubation. Similar differences in the kinetics of SR-BI-
by guest on February 14, 2018http://w
ww
.jbc.org/D
ownloaded from

Liu & Krieger page 18 In vitro assay of purified SR-BI
mediated HDL binding and lipid uptake have been observed in intact cultured cells (6).
There was little binding or lipid uptake by the receptor-negative control liposomes (solid
triangles). Figure 4 shows the temperature dependence of 125I-HDL binding and
[3H]cholesteryl ether uptake from [3H]CE-HDL (10 µg protein/ml) by mSR-BI-t1-
containing insect cell lysate-reconstituted liposomes. The value for 125I-HDL binding at
0°C was somewhat lower (79%) than that at 37°C (100%), whereas there was a more
substantial reduction in [3H]cholesteryl ether uptake at 0°C (25%) compared to that at
37°C (100%). It has previously been noted that SR-BI-mediated binding of HDL (23,34)
and selective uptake (34) are lower at 4°C than at 37°C and that the temperature
sensitivity of selective uptake is greater than that of HDL binding ((34), T. Nieland, T.
Kirchhausen and M. Krieger, unpublished observations).
To determine if the liposome assay could be used for recombinant SR-BI
produced by mammalian cells, we transfected HEK293S cells with a mammalian
expression vector for mSR-BI-t1 (pACmSR-BI-t1) and isolated a clone expressing high
levels of the receptor, HEK[mSR-BI-t1] (clone 7). Detergent solubilized lysates of these
cells were reconstituted into liposomes and the binding of 125I-HDL and the uptake of
[3H]cholesteryl ether from [3H]CE-HDL were measured. Figure 5A shows results similar
to those seen using extracts from the insect cells, i.e., high affinity HDL binding
(apparent Kd 8.5 µg protein/ml, Bmax 12.8 ng HDL protein/assay) and selective lipid
uptake (apparent Kd 29.4 µg protein/ml, Umax 220.9 ng HDL protein/assay).
by guest on February 14, 2018http://w
ww
.jbc.org/D
ownloaded from

Liu & Krieger page 19 In vitro assay of purified SR-BI
The specificity of the receptor’s activity in mSR-BI-t1-containing mammalian cell
lysate-reconstituted liposomes was examined by comparing the abilities of unlabeled
HDL and LDL to inhibit [3H]cholesteryl ether uptake from [3H]CE-HDL (10 µg
protein/ml). Previous studies of SR-BI expressed in intact cells established that, although
LDL can bind to SR-BI with high affinity, LDL is a poor inhibitor of HDL binding to
SR-BI (6,25). Figure 5B shows that, as is the case for intact cells, HDL was an effective
inhibitor of mSR-BI-t1-mediated lipid uptake activity from [3H]CE-HDL, while LDL was
less effective. The [3H]cholesteryl ether uptake from [3H]CE-HDL (10 µg protein/ml) by
mSR-BI-t1-containing mammalian cell lysate-reconstituted liposomes was inhibited by
the polyclonal anti-SR-BI antibody KKB-1 to the same extent as by excess unlabeled
HDL (Figure 5C), whereas uptake was not inhibited by control pre-immune antibody.
Control untransfected HEK293S cell lysate-reconstituted liposomes exhibited essentially
no [3H]cholesteryl ether uptake activity.
Although the above data support the validity of the liposome assay, we further
tested the assay by determining the receptor activities of liposomes reconstituted with
insect cell lysates containing in place of mSR-BI-t1 either a mutant form of mSR-BI-t1 or
human CD36, another class B scavenger receptor (5,19). We have isolated a set of mutant
mSR-BIs which exhibit altered ligand binding properties when expressed in transfected
mammalian cells (23,25). One of these has a double substitution of arginines for the
glutamines at positions 402 and 418 (designated ‘402R/418R’). This 402R/418R mutant
is as effective as wild-type mSR-BI in functioning as an LDL receptor in transfected
mammalian cells - mediating high affinity LDL binding, uptake of metabolically active
by guest on February 14, 2018http://w
ww
.jbc.org/D
ownloaded from

Liu & Krieger page 20 In vitro assay of purified SR-BI
cholesterol from LDL, and efflux of cholesterol to LDL; however, it has lost most of the
corresponding HDL receptor activity exhibited by the wild-type receptor (23). Lysates
from insect cells expressing mSR-BI-t1, the 402R/418R mutant form of mSR-BI-t1, or no
recombinant protein (control) were reconstituted into liposomes and the abilities of the
liposomes to bind 125I-HDL or 125I-LDL were determined. Figure 6B shows that, as
expected, the specific binding of 125I-LDL (5 µg protein/ml) to the 402R/418R mutant
(stippled bar) was similar to that of mSR-BI-t1 (open bar) and substantially greater than
that of the receptor-free control (filled bar). In contrast, Figure 6A shows that the binding
of 125I-HDL (5 µg protein/ml) to the 402R/418R mutant was much lower than that of
mSR-BI-t1 and was not significantly different from that of the receptor-free control.
Because the binding specificities of the whole insect lysate reconstituted liposomes
reflected the specificities of the corresponding intact mammalian cells, it seems likely
that the mechanism of SR-BI-mediated lipoprotein binding in the lysate reconstituted
liposomes is similar to that in intact mammalian cells.
CD36 is a class B scavenger receptor that is structurally similar to SR-BI and
shares a number of ligand binding activities (19,26,42,44). For example, CD36 binds
HDL with an affinity similar to that of mSR-BI (26,27,42); however, CD36 cannot
mediate efficient selective uptake of cholesterol from HDL to cells (26,27). Figure 2B
shows that CD36-containing insect cell lysate-reconstituted liposomes bound 125I-HDL
(filled squares) at a level comparable to that of the corresponding mSR-BI-t1-containing
liposomes (Figure 2A, open squares), Bmax ~15 ng protein/assay (apparent Kd 30.3 µg
protein/ml). In contrast, the CD36-containing liposomes exhibited almost no
by guest on February 14, 2018http://w
ww
.jbc.org/D
ownloaded from

Liu & Krieger page 21 In vitro assay of purified SR-BI
[3H]cholesteryl ether uptake from [3H]CE-HDL (Figure 2B, filled circles, Umax ~3.5 ng
protein/assay) compared to that of mSR-BI-t1-containing liposomes (Figure 2A, open
circles, Umax ~106.3 ng protein/assay). Thus, the lipid uptake activities of mSR-BI-t1
and CD36 in whole insect lysate reconstituted liposomes reflected their activities when
expressed on the surfaces of intact mammalian cells.
Taken together the data in Figures 2-6 show that the whole cell lysate/liposome
assay recapitulated many key features of cellular SR-BI-mediated HDL receptor activity:
1)125I-HDL binding and [3H]cholesteryl ether uptake from [3H]CE-HDL were high
affinity and saturable (6), 2) binding reached a steady state more rapidly than lipid uptake
(6), 3) LDL was a poor inhibitor of HDL binding and lipid uptake (6,25), 4) lipid transfer
was specifically inhibited by an anti-SR-BI blocking antibody (25), 5) lipid uptake was
substantially more temperature sensitive than binding ((34), T. Nieland, T. Kirchhausen
and M. Krieger, unpublished observations) and 6) lipid transfer occurred via selective
uptake (6). Furthermore, in this assay the activities of a mutant form of SR-BI
(402R/418R) and the homologue CD36 also recapitualted those in intact cells. Therefore,
the reconstituted liposome system provides a valid assay for the HDL binding and lipid
uptake activities of SR-BI in detergent solubilized, whole cell lysates.
One-step Immunoaffinity Purification of mSR-BI-t1
To examine the function of SR-BI using the reconstituted liposome assay in the
absence of other proteins, we isolated highly purified mSR-BI-t1 from HEK[mSR-BI-t1]
cells using a modified version of the rhodopsin purification of Reeves et al. (38). The
by guest on February 14, 2018http://w
ww
.jbc.org/D
ownloaded from

Liu & Krieger page 22 In vitro assay of purified SR-BI
receptor was purified from octyl glucoside solubilized cells by immunoaffinity
chromatography using an antibody to its C-terminal rhodopsin peptide epitope tag.
Results of a typical purification are shown in Figure 7, in which specimens obtained
throughout the procedure were fractionated by SDS-PAGE and visualized by silver
staining of the gel. Both the starting cell lysate (lane 1) and the initial column flow
through (material not retained by the column, lane 2) were highly complex protein
mixtures. After washing the column so that no additional protein was detected by silver
staining (lane 3), we eluted bound material with the rhodopsin C-terminal peptide and
recovered virtually homogeneously pure mSR-BI-t1 (lanes 4-8). The electrophoretic
mobility of the bulk of the purified material corresponded to 82 kDa, as expected from
previous studies (6). Immunoblotting of a replicate gel with anti-mSR-BI KKB-1
antibody established that the major protein band detected by silver staining, as well as the
very low abundance minor bands (e.g. see lane 5), was either mSR-BI-t1 or minor
proteolytic or aggregated forms of mSR-BI-t1 (not shown). Immunoblotting also
revealed that a very small amount of mSR-BI-t1 was present in the column flow through
and wash fractions (corresponding to lanes 2 and 3, not shown). We estimate from
quantitative immunoblotting that the overall recovery of purified mSR-BI-t1 from the
lysate was approximately 80%. The yield of mSR-BI-t1 was 100–150 µg/liter of
suspension cell culture.
Activity of Purified mSR-BI-t1 Reconstituted into Liposomes
With the availability of pure mSR-BI-t1 and a fully validated in vitro
reconstituted liposome assay, we were able to address the main question of this study:
by guest on February 14, 2018http://w
ww
.jbc.org/D
ownloaded from

Liu & Krieger page 23 In vitro assay of purified SR-BI
could mSR-BI, independently of any other protein, mediate HDL binding and selective
lipid uptake? Figure 8 shows the results of an experiment in which we measured as a
function of ligand concentration 125I-HDL binding to (open squares) and [3H]cholesteryl
ether uptake from [3H]CE-HDL by (open circles) liposomes reconstituted with the
immunoaffinity purified receptor. Figure 8 (open squares) shows that specific 125I-HDL
binding was high affinity (apparent Kd of 11.9 µg protein/ml) and saturable (Bmax ~16.6
ng protein/assay). The maximal binding, corrected for the amount of protein incorporated
in the liposomes was 311 ng protein/µg liposome protein, a value 65-fold higher than the
corresponding value for binding of 125I-HDL to liposomes reconstituted with HEK[mSR-
BI-t1] whole cell lysate (Bmax of 4.8 ng protein/µg liposome protein). Figure 8 (open
circles) shows the specific [3H]cholesteryl ether uptake from [3H]CE-HDL was also high
affinity and saturable. For [3H]cholesteryl ether uptake, the apparent Kd was 13.8 µg
protein/ml and the Umax was 134 ng protein/assay or 2,500 ng protein/µg liposome
protein. The relative amount of lipid uptake was substantially greater than that of binding,
with a ratio of the maximal [3H]CE-HDL uptake/125I-HDL binding of 8, clearly showing
that the pure receptor mediated selective lipid uptake from HDL. The HDL binding and
lipid uptake activities of the mSR-BI-t1-containing liposomes increased linearly with the
amount of mSR-BI-t1 incorporated into the liposomes (data not shown).
We conducted three additional experiments to determine if the selective lipid
uptake activity of the purified receptor reconstituted into liposomes exhibited
characteristics similar to those of SR-BI in intact cells. First, we compared the abilities of
unlabeled HDL and LDL to inhibit 125I-HDL binding and [3H]cholesteryl ether uptake
by guest on February 14, 2018http://w
ww
.jbc.org/D
ownloaded from

Liu & Krieger page 24 In vitro assay of purified SR-BI
from [3H]CE-HDL (10 µg protein/ml). Figure 9 shows that excess unlabeled HDL (open
squares and circles) effectively inhibited 125I-HDL binding (panel A) and [3H]cholesteryl
ether uptake (panel B), whereas unlabeled LDL (filled squares and circles) did not. These
results were similar to those observed using liposomes reconstituted with mSR-BI-t1-
containing mammalian cell lysates (Figure 5B) and mSR-BI expressed in intact
mammalian cells (6,25). We did note that the extent of LDL competition for 125I-HDL
binding and [3H]cholesteryl ether uptake from [3H]CE-HDL varied somewhat from
experiment to experiment (maximum % inhibition of 8-20% for binding and 0-25% for
uptake), perhaps reflecting the effects that small changes in the state of the HDL (e.g.,
oxidation) can have on lipoprotein binding affinities (41). Second, we tested the anti-SR-
BI antibody KKB-1 specific inhibition of [3H]CE-HDL uptake. Figure 10 shows that the
KKB-1 antibody inhibited the [3H]cholesteryl ether uptake by mSR-BI-t1-reconstituted
liposomes, but the control antibody from preimmune serum did not. These results were
similar to those obtained with the mSR-BI-t1 mammalian cell lysate-reconstituted
liposomes (Figure 5C) and mSR-BI transfected intact mammalian cells (25). Third, we
examined the temperature dependence of 125I-HDL binding to (Figure 11A) and
[3H]cholesteryl ether uptake from [3H]CE-HDL (Figure 11B) at 37°C (open bars) or 0°C
(shaded bars). The lower temperature slightly lowered the125I-HDL binding (4.1 vs 3.3 ng
protein/assay at 37oC and 0 °C, respectively, 19.5% reduction), whereas lipid uptake was
substantially reduced at the lower temperature (35.7 vs 14.6 ng protein/assay at 37oC and
0°C, respectively, 59.1% reduction). These findings were consistent with those obtained
with mSR-BI-t1-containing cell lysates reconstituted liposomes (Figure 4) and intact cells
((34), and T. Nieland, T. Kirchhausen and M. Krieger, unpublished observations). Thus,
by guest on February 14, 2018http://w
ww
.jbc.org/D
ownloaded from

Liu & Krieger page 25 In vitro assay of purified SR-BI
SR-BI needed no additional protein co-factors to allow it to mediate HDL binding and
selective cholesteryl ether uptake.
DISCUSSION
We have developed an in vitro reconstituted liposome assay to measure the ligand
binding and lipid transport activities of detergent solubilized forms of the HDL receptor
SR-BI. An epitope-tagged form of the recombinant receptor (mSR-BI-t1) in detergent
solubilized whole insect or mammalian cell lysates, or mSR-BI-t1 purified by
immunoaffinity chromatography essentially to homogeneity, can be assayed using this
liposome system. The liposome assay recapitulated many features of the HDL receptor
activity of SR-BI expressed in intact cells (5): 1) 125I-HDL binding and [3H]cholesteryl
ether uptake from [3H]CE-HDL were high affinity and saturable, 2) binding reached a
steady state more rapidly than lipid uptake, 3) LDL, although a tight binding ligand of the
receptor, was a poor inhibitor of HDL binding and lipid uptake from HDL, 4) lipid
transfer was specifically inhibited by an anti-SR-BI blocking antibody, 5) lipid uptake
was substantially more temperature sensitive than HDL binding, and 6) lipid transfer
occurred via selective uptake. Furthermore, the novel binding properties of a doubly
mutated receptor (Q402R and Q418R, normal LDL binding with the loss of HDL binding
(23)) were reproduced in the liposome assay, as was the ability of the SR-BI homologue
CD36 to bind HDL but not mediate efficient lipid uptake (26,27,42). It should be possible
to use this assay to explore in detail many features of the mechanism underlying the
complex ligand binding and lipid transport activities of SR-BI. It should be noted that,
by guest on February 14, 2018http://w
ww
.jbc.org/D
ownloaded from

Liu & Krieger page 26 In vitro assay of purified SR-BI
prior to and after the discovery of SR-BI and its role as an HDL receptor for selective
lipid uptake, several groups reported that intact – not solubilized - membranes isolated
from adipocyte, steroidogenic or hepatic tissues or cells could mediate selective lipid
uptake (32,45-49).
The first question regarding the mechanism of SR-BI activity that we addressed
using this assay was: does SR-BI require the cooperation of other proteins to mediate
HDL binding and selective lipid uptake, or are these activities autonomous properties of
the receptor (independent of other proteins)? Several observations raised the possibility
that the complex activities mediated by SR-BI might require the intervention or
collaboration of other proteins. For example, SR-BI can be found in specialized
membrane microdomains, including caveolar-like domains under certain conditions in
some cultured cells (50) and microvillar channels in mammalian steroidogenic cells in
vivo (51,52). Indeed, SR-BI expression in cultured cells can induce the formation of
microvillar channel-like structures (43) and plays a role in the formation and/or stability
of microvillar channels in steroidogenic cells in vivo (53,54). In addition, SR-BI has
been shown to bind, via its C-terminal cytoplasmic tail, to a multiple PDZ domain-
containing scaffold protein called CLAMP (55). The only unequivocal way to determine
if other proteins were essential for key SR-BI activities was to examine the activity of the
receptor purified away from other proteins. The liposome assay together with the highly
efficient immunoaffinity purification of an epitope tagged form of the receptor permitted
an unequivocal answer to this question. Essentially homogeneously pure mSR-BI-t1
incorporated into phosphatidylcholine/cholesterol liposomes did mediate high affinity
by guest on February 14, 2018http://w
ww
.jbc.org/D
ownloaded from

Liu & Krieger page 27 In vitro assay of purified SR-BI
and saturable binding of 125I-HDL that was accompanied by efficient selective uptake of
[3H]cholesteryl ether from [3H]CE-HDL. These findings do not address the question of
whether or not other proteins can or do modulate SR-BI’s intrinsic ability to mediate
HDL binding to and selective lipid uptake by intact cells. Neither do they address the
role, if any, of lipoprotein internalization by cells in selective uptake. They do, however,
establish that SR-BI itself has the capacity to function as an HDL receptor for selective
lipid uptake in liposomes without the required intervention of other proteins or cellular
structures or compartments. Therefore these results suggest that SR-BI itself is primarily
responsible for the lipid transfer step during SR-BI-mediated selective lipid uptake in
vivo.
Acknowledgements: We thank M. Penman and S. Xu for help with lipoprotein
preparations and advice regarding assays, T. Nieland and W. Evans for assistance, and X.
Gu, B. Trigatti, W. Schneider, A. Rigotti, K. Cha, E. Getmanova and C. Bruel, S. Bell, F.
Yang, G. Paradis, and R. Cook for advice and generously providing reagents. We are
especially grateful to P. Reeves and G. Khorana who very generously provided advice,
access to equipment and assistance in adapting to SR-BI the procedure for the
purification of rhodopsin developed by them and their colleagues, and to Karen Kozarsky
for providing the KKB-1 antibody used in these studies.
by guest on February 14, 2018http://w
ww
.jbc.org/D
ownloaded from

Liu & Krieger page 28 In vitro assay of purified SR-BI
REFERENCES
1. Brown, M. S., and Goldstein, J. L. (1986) Science 232, 34-472. Glass, C., Pittman, R. C., Weinstein, D. B., and Steinberg, D. (1983) Proc NatlAcad Sci U S A 80, 5435-93. Stein, Y., Dabach, Y., Hollander, G., Halperin, G., and Stein, O. (1983) BiochimBiophys Acta 752, 98-105.4. Pieters, M. N., Schouten, D., and Van Berkel, T. J. (1994) Biochim Biophys Acta1225, 125-34.5. Krieger, M. (1999) Annu Rev Biochem 68, 523-586. Acton, S., Rigotti, A., Landschulz, K. T., Xu, S., Hobbs, H. H., and Krieger, M.(1996) Science 271, 518-207. Kozarsky, K. F., Donahee, M. H., Rigotti, A., Iqbal, S. N., Edelman, E. R., andKrieger, M. (1997) Nature 387, 414-78. Rigotti, A., Trigatti, B. L., Penman, M., Rayburn, H., Herz, J., and Krieger, M.(1997) Proc Natl Acad Sci U S A 94, 12610-59. Trigatti, B., Rayburn, H., Vinals, M., Braun, A., Miettinen, H., Penman, M.,Hertz, M., Schrenzel, M., Amigo, L., Rigotti, A., and Krieger, M. (1999) Proc Natl AcadSci U S A 96, 9322-710. Mardones, P., Quinones, V., Amigo, L., Moreno, M., Miquel, J. F., Schwarz, M.,Miettinen, H. E., Trigatti, B., Krieger, M., VanPatten, S., Cohen, D. E., and Rigotti, A.(2001) J Lipid Res 42, 170-8011. Sehayek, E., Ono, J. G., Shefer, S., Nguyen, L. B., Wang, N., Batta, A. K., Salen,G., Smith, J. D., Tall, A. R., and Breslow, J. L. (1998) Proc Natl Acad Sci U S A 95,10194-912. Ji, Y., Wang, N., Ramakrishnan, R., Sehayek, E., Huszar, D., Breslow, J. L., andTall, A. R. (1999) J Biol Chem 274, 33398-40213. Kozarsky, K. F., Donahee, M. H., Glick, J. M., Krieger, M., and Rader, D. J.(2000) Arterioscler Thromb Vasc Biol 20, 721-714. Ueda, Y., Gong, E., Royer, L., Cooper, P. N., Francone, O. L., and Rubin, E. M.(2000) J Biol Chem 275, 20368-7315. Arai, T., Wang, N., Bezouevski, M., Welch, C., and Tall, A. R. (1999) J BiolChem 274, 2366-7116. Holm, T. M., Braun, A., Trigatti, B. L., Brugnara, C., Sakamoto, M., Krieger, M.,and Andrews, N. C. (2002) Blood 99, 1817-2417. Miettinen, H. E., Rayburn, H., and Krieger, M. (2001) J Clin Invest 108, 1717-2218. Braun, A., Trigatti, B. L., Post, M. J., Sato, K., Simons, M., Edelberg, J. M.,Rosenberg, R. D., Schrenzel, M., and Krieger, M. (2002) Circ Res 90, 270-27619. Acton, S. L., Scherer, P. E., Lodish, H. F., and Krieger, M. (1994) J Biol Chem269, 21003-920. Calvo, D., Gomez-Coronado, D., Lasuncion, M. A., and Vega, M. A. (1997)Arterioscler Thromb Vasc Biol 17, 2341-921. Stangl, H., Hyatt, M., and Hobbs, H. H. (1999) J Biol Chem 274, 32692-822. Swarnakar, S., Temel, R. E., Connelly, M. A., Azhar, S., and Williams, D. L.(1999) J Biol Chem 274, 29733-9
by guest on February 14, 2018http://w
ww
.jbc.org/D
ownloaded from

Liu & Krieger page 29 In vitro assay of purified SR-BI
23. Gu, X., Lawrence, R., and Krieger, M. (2000) J Biol Chem 275, 9120-3024. Jian, B., de la Llera-Moya, M., Ji, Y., Wang, N., Phillips, M. C., Swaney, J. B.,Tall, A. R., and Rothblat, G. H. (1998) J Biol Chem 273, 5599-60625. Gu, X., Kozarsky, K., and Krieger, M. (2000) J Biol Chem 275, 29993-3000126. Gu, X., Trigatti, B., Xu, S., Acton, S., Babitt, J., and Krieger, M. (1998) J BiolChem 273, 26338-4827. Connelly, M. A., Klein, S. M., Azhar, S., Abumrad, N. A., and Williams, D. L.(1999) J Biol Chem 274, 41-728. Liu, T., Krieger, M., Kan, H. Y., and Zannis, V. I. (2002) J Biol Chem 277,21576-84.29. de la Llera-Moya, M., Rothblat, G. H., Connelly, M. A., Kellner-Weibel, G.,Sakr, S. W., Phillips, M. C., and Williams, D. L. (1999) J Lipid Res 40, 575-80.30. Reaven, E., Chen, Y. D., Spicher, M., and Azhar, S. (1984) J Clin Invest 74,1384-97.31. Silver, D. L., Wang, N., Xiao, X., and Tall, A. R. (2001) J Biol Chem 276, 25287-9332. Rinninger, F., Jaeckle, S., and Pittman, R. C. (1993) Biochim Biophys Acta 1166,275-83.33. Oquendo, P., Hundt, E., Lawler, J., and Seed, B. (1989) Cell 58, 95-101.34. Rodrigueza, W. V., Thuahnai, S. T., Temel, R. E., Lund-Katz, S., Phillips, M. C.,and Williams, D. L. (1999) J Biol Chem 274, 20344-5035. Lowry, O. H., Rosebrough, N. J., Farr, A. L., and Randall, R. J. (1951) J BiolChem 193, 265-7536. Velan, B., Kronman, C., Ordentlich, A., Flashner, Y., Leitner, M., Cohen, S., andShafferman, A. (1993) Biochem J 296, 649-56.37. Sambrook, J., Fritsch, E. F., and Maniatis, T. (1989) Molecular cloning : alaboratory manual, 2nd Ed., Cold Spring Harbor Laboratory Press, Cold Spring Harbor,N.Y.38. Reeves, P. J., Thurmond, R. L., and Khorana, H. G. (1996) Proc Natl Acad Sci US A 93, 11487-9239. Schneider, W. J., Goldstein, J. L., and Brown, M. S. (1980) J Biol Chem 255,11442-740. Molday, R. S., and MacKenzie, D. (1983) Biochemistry 22, 653-60.41. Liadaki, K. N., Liu, T., Xu, S., Ishida, B. Y., Duchateaux, P. N., Krieger, J. P.,Kane, J., Krieger, M., and Zannis, V. I. (2000) J Biol Chem 275, 21262-7142. Calvo, D., Gomez-Coronado, D., Suarez, Y., Lasuncion, M. A., and Vega, M. A.(1998) J Lipid Res 39, 777-8843. Reaven, E., Leers-Sucheta, S., Nomoto, A., and Azhar, S. (2001) Proc Natl AcadSci U S A 98, 1613-844. Rigotti, A., Acton, S. L., and Krieger, M. (1995) J Biol Chem 270, 16221-445. Parkes, J. G., and Angel, A. (1990) Biochem Cell Biol 68, 870-9.46. Shi, X. Y., Azhar, S., and Reaven, E. (1992) Biochemistry 31, 3230-6.47. Rinninger, F., Jaeckle, S., Greten, H., and Windler, E. (1993) Biochim BiophysActa 1166, 284-99.48. Morrison, J. R., Silvestre, M. J., and Pittman, R. C. (1994) J Biol Chem 269,13911-8.
by guest on February 14, 2018http://w
ww
.jbc.org/D
ownloaded from

Liu & Krieger page 30 In vitro assay of purified SR-BI
49. Werder, M., Han, C. H., Wehrli, E., Bimmler, D., Schulthess, G., and Hauser, H.(2001) Biochemistry 40, 11643-5050. Babitt, J., Trigatti, B., Rigotti, A., Smart, E. J., Anderson, R. G., Xu, S., andKrieger, M. (1997) J Biol Chem 272, 13242-951. Reaven, E., Nomoto, A., Leers-Sucheta, S., Temel, R., Williams, D. L., andAzhar, S. (1998) Endocrinology 139, 2847-5652. Rigotti, A., Edelman, E. R., Seifert, P., Iqbal, S. N., DeMattos, R. B., Temel, R.E., Krieger, M., and Williams, D. L. (1996) J Biol Chem 271, 33545-953. Williams, D. L., Wong, J. S., and Hamilton, R. L. (2002) J Lipid Res 43, 544-9.54. Azhar, S., Nomoto, A., and Reaven, E. (2002) J Lipid Res 43, 861-71.55. Ikemoto, M., Arai, H., Feng, D., Tanaka, K., Aoki, J., Dohmae, N., Takio, K.,Adachi, H., Tsujimoto, M., and Inoue, K. (2000) Proc Natl Acad Sci U S A 97, 6538-43
by guest on February 14, 2018http://w
ww
.jbc.org/D
ownloaded from

Liu & Krieger page 31 In vitro assay of purified SR-BI
FIGURE LEGENDS
Figure 1. 125I-HDL binding (A,B) and [3H]CE uptake from [3H]CE-HDL (C,D) by
COS cells expressing wild type mSR-BI or the C-terminal epitope tagged mSR-BI-
t1. COS M6 cells were transiently transfected with expression vectors for mSR-BI, mSR-
BI-t1 (mSR-BI with a rhodopsin epitope tag incorporated as a C-terminal extension), or
the control ‘empty’ vector pcDNA. Two days later, the cells were incubated for two
hours at 37oC with the indicated concentrations of either 125I-HDL or [3H]CE-HDL in the
presence (single determinations) or absence (duplicate determinations) of a 40-fold
excess of unlabeled HDL. Specific 125I-HDL binding (A) and [3H]CE uptake (C) were
determined as described in Experimental Procedures. The relative levels of cell surface
expression of mSR-BI and mSR-BI-t1 were determined using the polyclonal anti-mSR-
BI antibody KKB-1 by flow cytometry as described in Experimental Procedures. The
values for the binding and lipid uptake by the mSR-BI expressing cells were corrected to
account for the 1.2-fold difference in surface expression relative to that of the mSR-BI-t1
expressing cells and are shown in panels C and D.
Figure 2. 125I-HDL binding and [3H]CE-HDL uptake by liposomes reconstituted
with insect (sf21) cell lysates . Sf21 cells were infected with baculoviruses encoding
mSR-BI-t1 (open symbols, panel A), human CD36 (filled squares and circles, panel B) or
no receptor (empty virus, filled triangles, both panels) and grown at 27oC for 6 days. Cell
lysates in 1.5% octyl glucoside lysis buffer were reconstituted into liposomes as
described in Experimental Procedures. The liposomes were incubated with 125I-HDL or
[3H]CE-HDL at the indicated concentrations at 37oC for 3 hours in the presence (single
by guest on February 14, 2018http://w
ww
.jbc.org/D
ownloaded from

Liu & Krieger page 32 In vitro assay of purified SR-BI
determinations) or absence (duplicate determinations) of a 40-fold excess of unlabeled
HDL, isolated and washed by filtration, and the amounts of specific125I-HDL binding and
[3H]cholesteryl ether uptake were determined as described in Experimental Procedures.
Error bars represent the range of variation in duplicate determinations. The nonspecific
background values for 125I-HDL binding ranged between 19-28% (mSR-BI-t1) and 29-
45% (CD36) of the total binding. The values for the no receptor-lysates were very low
and overlap for specific125I-HDL binding and [3H]cholesteryl ether uptake (broken lines).
Panel C shows with an expanded scale the binding of 125I-HDL to the liposomes
containing mSR-BI-t1 (open squares) and CD36 (filled squares).
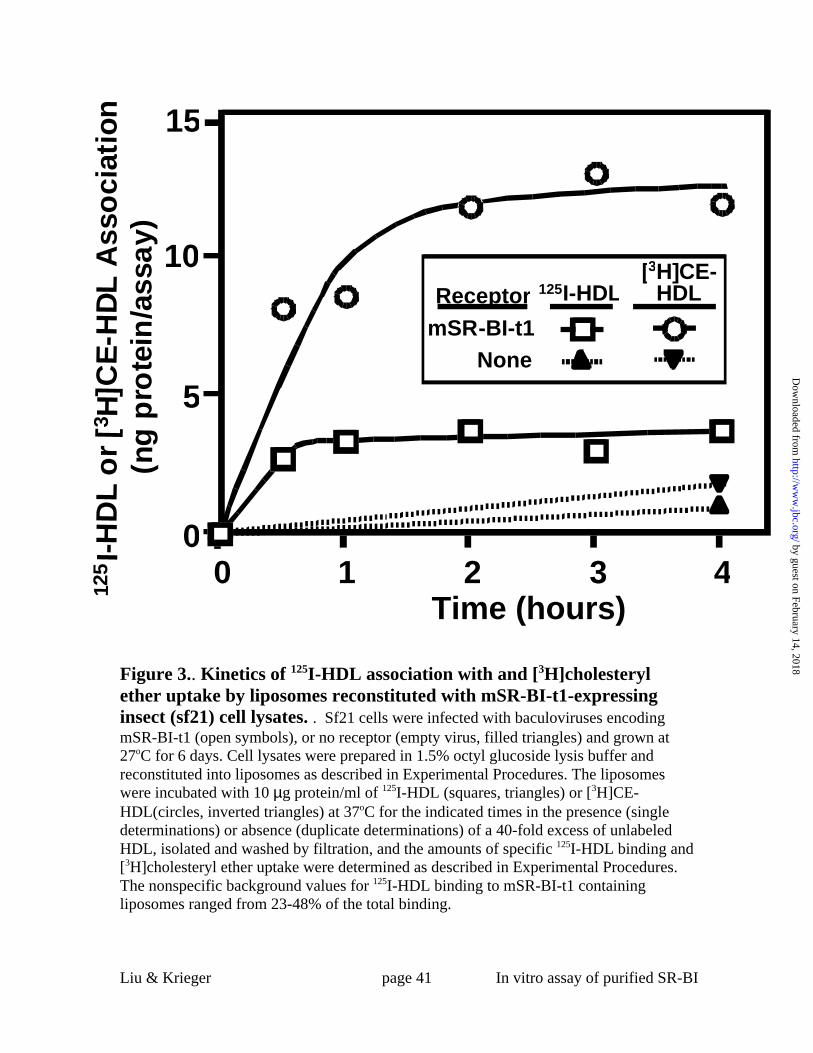
Figure 3.. Kinetics of 125I-HDL association with and [3H]cholesteryl ether uptake by
liposomes reconstituted with mSR-BI-t1-expressing insect (sf21) cell lysates. . Sf21
cells were infected with baculoviruses encoding mSR-BI-t1 (open symbols), or no
receptor (empty virus, filled triangles) and grown at 27oC for 6 days. Cell lysates were
prepared in 1.5% octyl glucoside lysis buffer and reconstituted into liposomes as
described in Experimental Procedures. The liposomes were incubated with10 µg
protein/ml of 125I-HDL (squares, triangles) or [3H]CE-HDL(circles, inverted triangles) at
37oC for the indicated times in the presence (single determinations) or absence (duplicate
determinations) of a 40-fold excess of unlabeled HDL, isolated and washed by filtration,
and the amounts of specific 125I-HDL binding and [3H]cholesteryl ether uptake were
determined as described in Experimental Procedures. The nonspecific background values
for 125I-HDL binding to mSR-BI-t1 containing liposomes ranged from 23-48% of the total
binding.
by guest on February 14, 2018http://w
ww
.jbc.org/D
ownloaded from

Liu & Krieger page 33 In vitro assay of purified SR-BI
Figure 4.. Temperature dependence of 125I-HDL association with and [3H]cholesteryl
ether uptake by liposomes reconstituted with mSR-BI-t1-expressing insect (sf21) cell
lysates. . Sf21 cells were infected with baculoviruses encoding mSR-BI-t1 (bars), or no
receptor (empty virus, not shown) and grown at 27oC for 6 days. Cell lysates were
prepared in 1.5% octyl glucoside lysis buffer and reconstituted into liposomes as
described in Experimental Procedures. The liposomes were incubated with10 µg
protein/ml of 125I-HDL (A) or [3H]CE-HDL (B) at 37oC for 2 hours in the presence
(single determinations) or absence (duplicate determinations) of a 40-fold excess of
unlabeled HDL, isolated and washed by filtration, and the amounts of specific 125I-HDL
binding and [3H]cholesteryl ether uptake were determined as described in Experimental
Procedures. The 100% of control values for binding and lipid uptake at 37°C were 3.0
and 13.0 ng protein/assay, respectively. The no receptor control lysates binding and lipid
uptake activities at 37°C were 26% and 9% of those of the mSR-BI-t1-containing lysates
(not shown). The nonspecific background values for 125I-HDL binding were 48% (37°C)
and 30% (0°C) of the total binding. Error bars represent the range of variation in
duplicate determinations.
Figure 5. 125I-HDL association with and [3H]cholesteryl ether uptake by liposomes
reconstituted with HEK[mSR-BI-t1] or control HEK cell lysates. . HEK[mSR-BI-t1]
and control untransfected HEK293S cells were grown in culture and lysates were
prepared in 1.5% octyl glucoside lysis buffer and reconstituted into liposomes as
described in Experimental Procedures.
by guest on February 14, 2018http://w
ww
.jbc.org/D
ownloaded from

Liu & Krieger page 34 In vitro assay of purified SR-BI
Panel A: The mSR-BI-t1-containing liposomes were incubated with the indicated
concentrations of 125I-HDL (squares) or [3H]CE-HDL (circles) at 37oC for 2 hours in the
presence (single determinations) or absence (duplicate determinations) of a 40-fold
excess of unlabeled HDL, isolated and washed by filtration, and the amounts of specific
125I-HDL binding and [3H]cholesteryl ether uptake were determined as described in
Experimental Procedures. In the same experiment, the background values for binding
and uptake by control (untransfected HEK293S cell-derived) liposomes at 10 µg
protein/ml of labeled lipoprotein were 45% and 32% of those of mSR-BI-t1-containing
liposomes (not shown). The nonspecific background values for 125I-HDL binding at 2.5,
5, 10, 25 and 100 µg protein/ml were 10%, 13%, 25%, 32% and 70% of total binding,
respectively. Inset: expanded scale for 125I-HDL binding.
Panel B: The mSR-BI-t1-containing liposomes were incubated in duplicate with 10 µg
protein/ml of [3H]CE-HDL at 37oC for 2 hours in the presence of the indicated amounts
of unlabeled lipoprotein competitors, HDL (open squares) or LDL (filled squares),
isolated and washed by filtration, and the amounts of [3H]cholesteryl ether uptake were
determined. In the same experiment, the background values for uptake by control
(untransfected HEK293S cell-derived) liposomes in the absence of competitor or the
presence of 400 µg protein/ml of either HDL or LDL were 7.7, 7.1 or 6.7 ng
protein/assay, respectively (not shown).
Panel C: The mSR-BI-t1-containing (open bars) or control (filled bars) liposomes were
incubated in triplicate at 37oC for 2 hours with 10 µg protein/ml of [3H]CE-HDL in the
absence (None) or presence of either 400 µg protein /ml of unlabeled HDL, 50 µg/ml of
the blocking antibody KKB-1 or 50 µg/ml of control antibody from preimmune serum.
by guest on February 14, 2018http://w
ww
.jbc.org/D
ownloaded from

Liu & Krieger page 35 In vitro assay of purified SR-BI
The liposomes were isolated and washed by filtration, and the amounts of [3H]cholesteryl
ether uptake were determined as described in Experimental Procedures. The 100% of
control value for lipid uptake in the absence of inhibitors was 19.1 ng protein/assay.
Error bars represent the range of variations in the triplicate determinations.
Figure 6. 125I-HDL and 125I-LDL binding by liposomes reconstituted with insect
(sf21) cell lysates containing mSR-BI-t1 (open bars), the 402R/418R mutant of mSR-
BI-t1 (stippled bars), or no recombinant receptor (filled bars) . Sf21 cells were
infected with baculoviruses encoding mSR-BI-t1 (open bars), the 402R/418R mutant of
mSR-BI-t1(light gray bars) or no receptor (empty virus, dark filled bars) and grown at
27oC. Cell lysates were prepared in 1.5% octyl glucoside lysis buffer and reconstituted
into liposomes as described in Experimental Procedures. The liposomes were incubated
with 5 µg protein/ml of 125I-HDL or 125I-LDL in the presence (single determinations) or
absence (duplicate determinations) of a 40-fold excess of the corresponding unlabeled
lipoprotein at 37oC for 2 hours, isolated and washed by filtration, and the amounts of
specific125I-HDL (A) and 125I-LDL (B) binding were determined as described in
Experimental Procedures. The 100% of control values for 125I-HDL and 125I-LDL binding
were 6.99 and 6.95 ng protein/assay, respectively. The nonspecific background values
for 125I-HDL binding were 23% (mSR-BI-t1) and 25% (402R/418R mutant) of the total
binding. Error bars represent the range of variations in duplicate determinations. * The
range of variation was ~50%.
by guest on February 14, 2018http://w
ww
.jbc.org/D
ownloaded from

Liu & Krieger page 36 In vitro assay of purified SR-BI
Figure 7. Immunoaffinity purification of mSR-BI-t1 from HEK[mSR-BI-t1] cell
lysates. HEK[mSR-BI-t1] cells were grown in suspension culture, lysed, and the lysates
subjected to immunoaffinity chromatrography using the 1D4 monoclonal anti-C-terminal
epitope tag antibody. The bound protein was eluted from the column with the peptide
epitope as described in Experimental Procedures. Samples of the cell lysate, column
flow through (unbound material), column wash, and peptide-eluted fractions (10 µl of
each fraction) were fractionated by 10% SDS-PAGE and the proteins in the gel were
visualized using a BioRad silver staining kit. The mobilities of molecular weight
standards are indicated on the left.
Figure 8. 125I-HDL binding and [3H]CE-HDL uptake by liposomes reconstituted
with immunoaffinity purified mSR-BI-t1. Immunoaffinity purified mSR-BI-t1 isolated
from HEK[mSR-BI-t1] cells was reconstituted into liposomes as described in
Experimental Procedures. Control liposomes without added receptor were prepared in
parallel. The liposomes were incubated with the indicated concentrations of 125I-HDL
(squares or triangles) or [3H]CE-HDL (circles or inverted triangles) at 37oC for 3 hours in
the presence (single determinations) or absence (duplicate determinations) of a 40-fold
excess of unlabeled HDL, isolated and washed by filtration, and the amounts of
specific125I-HDL binding and [3H]cholesteryl ether uptake were determined as described
in Experimental Procedures. The binding and lipid uptake values for the control (no
receptor) liposomes (filled triangles, partially obscured by the squares) determined at
labeled lipoprotein concentrations of 10 µg protein/ml were 0.8 and 0.9 ng/assay,
respectively. Error bars represent the range of variations in duplicate determinations. The
by guest on February 14, 2018http://w
ww
.jbc.org/D
ownloaded from

Liu & Krieger page 37 In vitro assay of purified SR-BI
nonspecific background values for 125I-HDL binding at 2.5, 10, 25, 70 and 100 µg
protein/ml were 20%, 24%, 26%, 30%, 42% and 50% of total binding, respectively.
Inset: expanded scale for 125I-HDL binding.
Figure 9. Unlabeled HDL and LDL inhibition of 125I-HDL binding and [3H]CE-HDL
uptake by liposomes reconstituted with immunoaffinity purified mSR-BI-t1.
Immunoaffinity purified mSR-BI-t1 isolated from HEK[mSR-BI-t1] cells was
reconstituted into liposomes as described in Experimental Procedures. Control liposomes
without added receptor were prepared in parallel. The liposomes were incubated in
duplicate with 5 µg protein/ml of 125I-HDL (A) or 10 µg protein/ml of [3H]CE-HDL (B)
in the presence of the indicated concentrations of unlabeled HDL (open symbols) or LDL
(filled symbols) at 37oC for 2 hours, isolated and washed by filtration, and the amounts of
125I-HDL binding and [3H]cholesteryl ether uptake were determined as described in
Experimental Procedures. The values for the control (no receptor) liposomes (triangles)
determined in the absence of competitor or in the presence of 400 µg protein/ml of HDL
or LDL were: 125I-HDL binding, 1.98, 0.94 and 1.25 ng/assay, respectively; and [3H]CE
uptake 5.03, 5.34 and 3.93 ng/assay, respectively.
Figure 10. Blocking antibody (KKB-1) inhibition of [3H]CE-HDL uptake by
liposomes reconstituted with immunoaffinity purified mSR-BI-t1. Immunoaffinity
purified mSR-BI-t1 isolated from HEK[mSR-BI-t1] cells was reconstituted into
liposomes as described in Experimental Procedures. Control liposomes without added
receptor were prepared in parallel. The liposomes were incubated in triplicate at 37oC for
by guest on February 14, 2018http://w
ww
.jbc.org/D
ownloaded from

Liu & Krieger page 38 In vitro assay of purified SR-BI
2 hours with 10 µg protein/ml of [3H]CE-HDL in the absence (-) or presence (+) of
unlabeled HDL (400 µg protein/ml), KKB-1 antibody (50 µg/ml), or preimmune IgG (50
µg/ml), isolated and washed by filtration, and the amounts of 125I-HDL binding and
[3H]cholesteryl ether uptake were determined as described in Experimental Procedures.
The 100% of control value for lipid uptake by the receptor containing liposomes in the
absence of inhibitors was 39.9 ng protein/assay. Error bars represent the range of
variations in triplicate determinations.
Figure 11. Temperature dependence of 125I-HDL association with and
[3H]cholesteryl ether uptake by liposomes reconstituted with immunoaffinity
purified mSR-BI-t1. Immunoaffinity purified mSR-BI-t1 isolated from HEK[mSR-BI-
t1] cells was reconstituted into liposomes as described in Experimental Procedures. The
liposomes were incubated with 10 µg protein/ml of 125I-HDL (A) or [3H]CE-HDL (B) at
37 oC (open bars) or 0oC (filled bars) for 2 hours in the presence (single determinations)
or absence (duplicate determinations) of a 40-fold excess of unlabeled HDL, isolated and
washed by filtration, and the amounts of specific 125I-HDL binding and [3H]cholesteryl
ether uptake were determined as described in Experimental Procedures. The 100% of
control values for binding and lipid uptake at 37°C were 4.1 and 35.7 ng protein/assay,
respectively. The specific values for the control (no receptor) liposomes determined at
37°C were: 125I-HDL binding, 28%; and [3H]CE uptake, ~0% (not shown). The
nonspecific background values for 125I-HDL binding were 24% (37°C) and 22% (0°C) of
the total binding. Error bars represent the range of variations in duplicate determinations.
by guest on February 14, 2018http://w
ww
.jbc.org/D
ownloaded from

Liu & Krieger page 39 In vitro assay of purified SR-BI
Figure 1. 125I-HDL binding (A,B) and [3H]CE uptake from [3H]CE-HDL (C,D) byCOS cells expressing wild type mSR-BI or the C-terminal epitope tagged mSR-BI-t1. COS M6 cells were transiently transfected with expression vectors for mSR-BI, mSR-BI-t1 (mSR-BI with a rhodopsin epitope tag incorporated as a C-terminal extension), or the control ‘empty’ vectorpcDNA. Two days later, the cells were incubated for two hours at 37oC with the indicatedconcentrations of either 125I-HDL or [3H]CE-HDL in the presence (single determinations) or absence(duplicate determinations) of a 40-fold excess of unlabeled HDL. Specific 125I-HDL binding (A) and[3H]CE uptake (C) were determined as described in Experimental Procedures. The relative levels of cellsurface expression of mSR-BI and mSR-BI-t1 were determined using the polyclonal anti-mSR-BIantibody KKB-1 by flow cytometry as described in Experimental Procedures. The values for thebinding and lipid uptake by the mSR-BI expressing cells were corrected to account for the 1.2-folddifference in surface expression relative to that of the mSR-BI-t1 expressing cells and are shown inpanels B and D.
0
4
8
12
16
0 25 50 75 100 0 25 50 75 100 125
0
0.1
0.2
0.3
0 25 50 75 100
A. 125I-HDL
0 25 50 75 100 125
B. 125I-HDL Corrected12
5 I-H
DL
Cel
lA
ssoc
iati
on
(ng
HD
L p
rote
in/
gce
ll p
rote
in)
125I-HDL ( g protein/ml)
[3H]CE-HDL ( g protein/ml)
[3H
]CE
-HD
L C
ell
Ass
ocia
tion
(ng
HD
L p
rote
in/
gce
ll p
rote
in)
C. [3H]CE-HDL D. [3H]CE-HDL Corrected
COS[mSR-BI]COS[mSR-BI-t1]COS[control]
125I-HDL[3H]CE-
HDLCells
by guest on February 14, 2018http://w
ww
.jbc.org/D
ownloaded from

Liu & Krieger page 40 In vitro assay of purified SR-BI
Figure 2. 125I-HDL binding and [3H]CE-HDL uptake by liposomesreconstituted with insect (sf21) cell lysates . Sf21 cells were infected withbaculoviruses encoding mSR-BI-t1 (open symbols, panel A), human CD36 (filled squaresand circles, panel B) or no receptor (empty virus, filled triangles, both panels) and grownat 27oC for 6 days. Cell lysates in 1.5% octyl glucoside lysis buffer were reconstitutedinto liposomes as described in Experimental Procedures. The liposomes were incubatedwith 125I-HDL or [3H]CE-HDL at the indicated concentrations at 37oC for 3 hours in thepresence (single determinations) or absence (duplicate determinations) of a 40-foldexcess of unlabeled HDL, isolated and washed by filtration, and the amounts ofspecific125I-HDL binding and [3H]cholesteryl ether uptake were determined as describedin Experimental Procedures. Error bars represent the range of variation in duplicatedeterminations. The nonspecific background values for 125I-HDL binding ranged between19-28% (mSR-BI-t1) and 29-45% (CD36) of the total binding. The values for the noreceptor-lysates were very low and overlap for specific125I-HDL binding and[3H]cholesteryl ether uptake (broken lines). Panel C shows with an expanded scale thebinding of 125I-HDL to the liposomes containing mSR-BI-t1 (open squares) and CD36(filled squares).
None
mSR-BI-t1CD36
[3H]CE-HDL125I-HDLReceptor
0
20
40
60
80
100
0 20 40 60 80
A. mSR-BI-t1 lysate
0 20 40 60 80125 I
-HD
L o
r [3
H]C
E-H
DL
Ass
oci
atio
n (
ng
HD
L p
rote
in/a
ssay
)
g protein/ml)Lipoprotein (
B. CD36 lysate
0
5
10
15
0 20 40 60 80
C. I-HDL binding comparison
125
CD36
mSR-BI-t1
by guest on February 14, 2018http://w
ww
.jbc.org/D
ownloaded from
Liu & Krieger page 41 In vitro assay of purified SR-BI
Figure 3.. Kinetics of 125I-HDL association with and [3H]cholesterylether uptake by liposomes reconstituted with mSR-BI-t1-expressinginsect (sf21) cell lysates. . Sf21 cells were infected with baculoviruses encodingmSR-BI-t1 (open symbols), or no receptor (empty virus, filled triangles) and grown at27oC for 6 days. Cell lysates were prepared in 1.5% octyl glucoside lysis buffer andreconstituted into liposomes as described in Experimental Procedures. The liposomeswere incubated with 10 µg protein/ml of 125I-HDL (squares, triangles) or [3H]CE-HDL(circles, inverted triangles) at 37oC for the indicated times in the presence (singledeterminations) or absence (duplicate determinations) of a 40-fold excess of unlabeledHDL, isolated and washed by filtration, and the amounts of specific 125I-HDL binding and[3H]cholesteryl ether uptake were determined as described in Experimental Procedures.The nonspecific background values for 125I-HDL binding to mSR-BI-t1 containingliposomes ranged from 23-48% of the total binding.
125I-H
DL
or
[3H
]CE
-HD
L A
sso
ciat
ion
(ng
pro
tein
/ass
ay)
Time (hours)
0
5
10
15
0 1 2 3 4
mSR-BI-t1None
[3H]CE-HDL125I-HDLReceptor
by guest on February 14, 2018http://w
ww
.jbc.org/D
ownloaded from

Liu & Krieger page 42 In vitro assay of purified SR-BI
Figure 4.. Temperature dependence of 125I-HDL association with and[3H]cholesteryl ether uptake by liposomes reconstituted with mSR-BI-t1-expressing insect (sf21) cell lysates. . Sf21 cells were infected withbaculoviruses encoding mSR-BI-t1 (bars), or no receptor (empty virus, not shown) andgrown at 27oC for 6 days. Cell lysates were prepared in 1.5% octyl glucoside lysis bufferand reconstituted into liposomes as described in Experimental Procedures. The liposomeswere incubated with 10 µg protein/ml of 125I-HDL (A) or [3H]CE-HDL (B) at 37oC for 2hours in the presence (single determinations) or absence (duplicate determinations) of a40-fold excess of unlabeled HDL, isolated and washed by filtration, and the amounts ofspecific 125I-HDL binding and [3H]cholesteryl ether uptake were determined as describedin Experimental Procedures. The 100% of control values for binding and lipid uptake at37°C were 3.0 and 13.0 ng protein/assay, respectively. The no receptor control lysates’binding and lipid uptake activities at 37°C were 26% and 9% of those of the mSR-BI-t1-containing lysates (not shown). The nonspecific background values for 125I-HDL bindingwere 48% (37°C) and 30% (0°C) of the total binding. Error bars represent the range ofvariation in duplicate determinations.
037
125 I
-HD
L o
r [3 H
]CE
-HD
LA
sso
ciat
ion
(% o
f ac
tivi
ty a
t 37o
C)
0
25
50
75
100
37 0Temperature (oC)
A. 125I-HDL B. [3H]CE-HDL
by guest on February 14, 2018http://w
ww
.jbc.org/D
ownloaded from

Liu & Krieger page 43 In vitro assay of purified SR-BI
Figure 5. 125I-HDL association with and [3H]cholesteryl ether uptake byliposomes reconstituted with HEK[mSR-BI-t1] or control HEK cell lysates. .HEK[mSR-BI-t1] and control untransfected HEK293S cells were grown in culture and lysates were reconstituted into liposomes.Panel A: The mSR-BI-t1-containing liposomes were incubated with the indicated concentrations of 125I-HDL (squares) or [3H]CE-HDL (circles) at 37oC for 2 hours in the presence (single determinations) or absence (duplicate determinations) of a 40-fold excessof unlabeled HDL, isolated and washed by filtration, and the amounts of specific 125I-HDL binding and [3H]cholesteryl ether uptakewere determined as described in Experimental Procedures. In the same experiment, the background values for binding and uptakeby control (untransfected HEK293S cell-derived) liposomes at 10 µg protein/ml of labeled lipoprotein were 45% and 32% of thoseof mSR-BI-t1-containing liposomes (not shown). The nonspecific background values for 125I-HDL binding at 2.5, 5, 10, 25 and 100µg protein/ml were 10%, 13%, 25%, 32% and 70% of total binding, respectively. Inset: expanded scale for 125I-HDL binding.Panel B: The mSR-BI-t1-containing liposomes were incubated in duplicate with 10 µg protein/ml of [3H]CE-HDL at 37oC for 2hours in the presence of the indicated amounts of unlabeled lipoprotein competitors, HDL (open squares) or LDL (filled squares),isolated and washed by filtration, and the amounts of [3H]cholesteryl ether uptake were determined. In the same experiment, thebackground values for uptake by control (untransfected HEK293S cell-derived) liposomes in the absence of competitor or thepresence of 400 µg protein/ml of either HDL or LDL were 7.7, 7.1 or 6.7 ng protein/assay, respectively (not shown).Panel C: The mSR-BI-t1-containing (open bars) or control (filled bars) liposomes were incubated in triplicate at 37oC for 2 hourswith 10 µg protein/ml of [3H]CE-HDL in the absence (None) or presence of either 400 µg protein /ml of unlabeled HDL, 50 µg/mlof the blocking antibody KKB-1 or 50 µg/ml of control antibody from preimmune serum. The liposomes were isolated and washedby filtration, and the amounts of [3H]cholesteryl ether uptake were determined. The 100% of control value for lipid uptake in theabsence of inhibitors was 19.1 ng protein/assay. Error bars represent the range of variations in the triplicate determinations.
by guest on February 14, 2018http://w
ww
.jbc.org/D
ownloaded from

Liu & Krieger page 44 In vitro assay of purified SR-BI
Figure 6. 125I-HDL and 125I-LDL binding by liposomes reconstitutedwith insect (sf21) cell lysates containing mSR-BI-t1 (open bars), the402R/418R mutant of mSR-BI-t1 (stippled bars), or no recombinantreceptor (filled bars). Sf21 cells were infected with baculoviruses encoding mSR-
BI-t1 (open bars), the 402R/418R mutant of mSR-BI-t1(light gray bars) or no receptor(empty virus, dark filled bars) and grown at 27oC. Cell lysates were prepared in 1.5%octyl glucoside lysis buffer and reconstituted into liposomes as described in ExperimentalProcedures. The liposomes were incubated with 5 µg protein/ml of 125I-HDL or 125I-LDLin the presence (single determinations) or absence (duplicate determinations) of a 40-foldexcess of the corresponding unlabeled lipoprotein at 37oC for 2 hours, isolated andwashed by filtration, and the amounts of specific125I-HDL (A) and 125I-LDL (B) bindingwere determined as described in Experimental Procedures. The 100% of control valuesfor 125I-HDL and 125I-LDL binding were 6.99 and 6.95 ng protein/assay, respectively. Thenonspecific background values for 125I-HDL binding were 23% (mSR-BI-t1) and 25%
(402R/418R mutant) of the total binding. Error bars represent the range of variations induplicate determinations. * The range of variation was ~50%.
mSR-BI-t1Q402R/Q418R
None
B. 125I-LDL
*12
5I-H
DL
or
125I-L
DL
Bin
din
g(%
of m
SR
-BI-t
1 ac
tivi
ty)
0
25
50
75
100
mSR-BI-t1Q402R/Q418R
None
A. 125I-HDL
by guest on February 14, 2018http://w
ww
.jbc.org/D
ownloaded from

Liu & Krieger page 45 In vitro assay of purified SR-BI
Figure 7. Immunoaffinity purification of mSR-BI-t1 from HEK[mSR-BI-t1] cell lysates. HEK[mSR-BI-t1] cells were grown in suspension culture, lysed,and the lysates subjected to immunoaffinity chromatography using the 1D4 monoclonalanti-C-terminal epitope tag antibody. The bound protein was eluted from the columnwith the peptide epitope as described in Experimental Procedures. Samples of the celllysate, column flow through (unbound material), column wash, and peptide-elutedfractions (10 µl of each fraction) were fractionated by 10% SDS-PAGE and the proteinsin the gel were visualized using a BioRad silver staining kit. The mobilities of molecularweight standards are indicated on the left.
Pept
ide-
Elut
ed
Frac
tions
Cell L
ysat
eFl
ow T
hrou
ghW
ash
200
11697
66
45
31
kDa
mSR-BI-t1
1 2 3 4 5 6 7 8 by guest on February 14, 2018http://w
ww
.jbc.org/D
ownloaded from

Liu & Krieger page 46 In vitro assay of purified SR-BI
Figure 8. 125I-HDL binding and [3H]CE-HDL uptake by liposomesreconstituted with immunoaffinity purified mSR-BI-t1. Immunoaffinitypurified mSR-BI-t1 isolated from HEK[mSR-BI-t1] cells was reconstituted intoliposomes as described in Experimental Procedures. Control liposomes without addedreceptor were prepared in parallel. The liposomes were incubated with the indicatedconcentrations of 125I-HDL (squares or triangles) or [3H]CE-HDL (circles or invertedtriangles) at 37oC for 3 hours in the presence (single determinations) or absence(duplicate determinations) of a 40-fold excess of unlabeled HDL, isolated and washed byfiltration, and the amounts of specific125I-HDL binding and [3H]cholesteryl ether uptakewere determined as described in Experimental Procedures. The binding and lipid uptakevalues for the control (no receptor) liposomes (filled triangles, partially obscured by thesquares) determined at labeled lipoprotein concentrations of 10 µg protein/ml were 0.8and 0.9 ng/assay, respectively. Error bars represent the range of variations in duplicatedeterminations. The nonspecific background values for 125I-HDL binding at 2.5, 10, 25,70 and 100 µg protein/ml were 20%, 24%, 26%, 30%, 42% and 50% of total binding,respectively. Inset: expanded scale for 125I-HDL binding.
by guest on February 14, 2018http://w
ww
.jbc.org/D
ownloaded from

Liu & Krieger page 47 In vitro assay of purified SR-BI
Figure 9. Unlabeled HDL and LDL inhibition of 125I-HDL binding and[3H]CE-HDL uptake by liposomes reconstituted with immunoaffinitypurified mSR-BI-t1. Immunoaffinity purified mSR-BI-t1 isolated from HEK[mSR-BI-t1] cells was reconstituted into liposomes as described in Experimental Procedures.Control liposomes without added receptor were prepared in parallel. The liposomes wereincubated in duplicate with 5 µg protein/ml of 125I-HDL (A) or 10 µg protein/ml of[3H]CE-HDL (B) in the presence of the indicated concentrations of unlabeled HDL (opensymbols) or LDL (filled symbols) at 37oC for 2 hours, isolated and washed by filtration,and the amounts of 125I-HDL binding and [3H]cholesteryl ether uptake were determined asdescribed in Experimental Procedures.
125 I
-HD
L B
ind
ing
or
[3H
]CE
-HD
L U
pta
ke(n
g p
rote
in/a
ssay
)
0
15
30
45
60
0 100 200 300 400 5000
2
4
6
8
10
0 100 200 300 400 500
A. 125I-HDL B. [3H]CE-HDL
HDL LDL
mSR-BI-t1
None
Receptor
Lipoprotein ( g protein/ml)
by guest on February 14, 2018http://w
ww
.jbc.org/D
ownloaded from
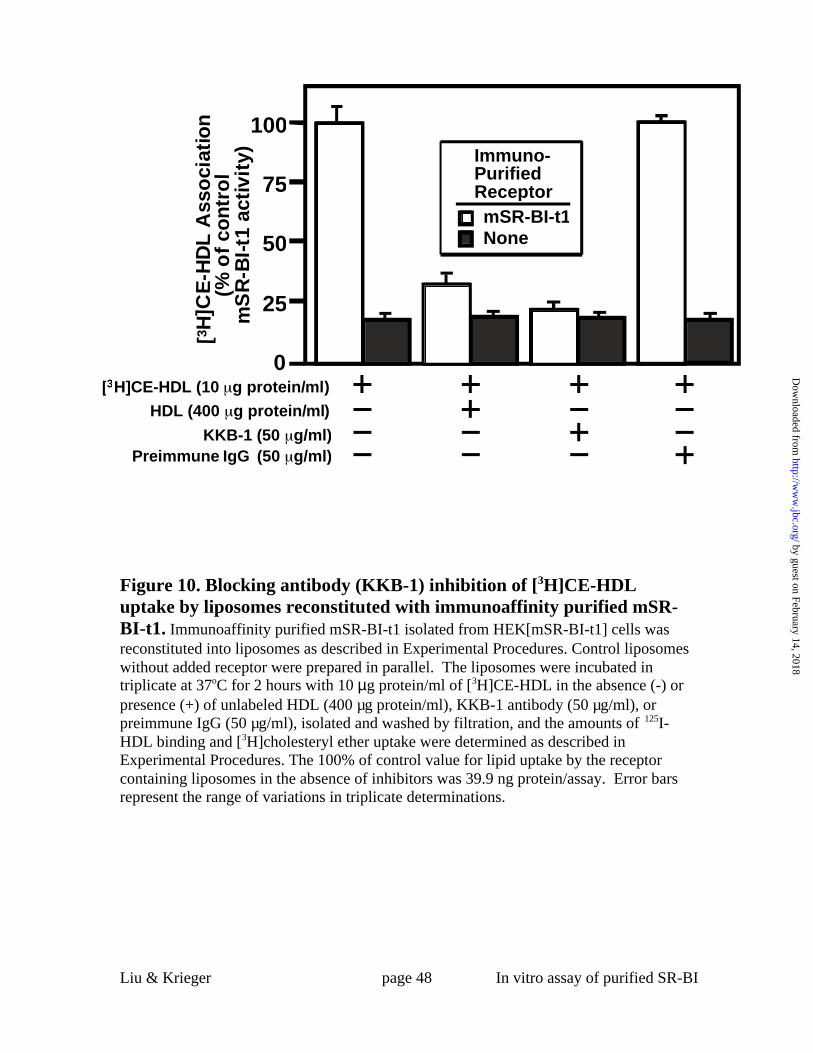
Liu & Krieger page 48 In vitro assay of purified SR-BI
Figure 10. Blocking antibody (KKB-1) inhibition of [3H]CE-HDLuptake by liposomes reconstituted with immunoaffinity purified mSR-BI-t1. Immunoaffinity purified mSR-BI-t1 isolated from HEK[mSR-BI-t1] cells wasreconstituted into liposomes as described in Experimental Procedures. Control liposomeswithout added receptor were prepared in parallel. The liposomes were incubated intriplicate at 37oC for 2 hours with 10 µg protein/ml of [3H]CE-HDL in the absence (-) orpresence (+) of unlabeled HDL (400 µg protein/ml), KKB-1 antibody (50 µg/ml), orpreimmune IgG (50 µg/ml), isolated and washed by filtration, and the amounts of 125I-HDL binding and [3H]cholesteryl ether uptake were determined as described inExperimental Procedures. The 100% of control value for lipid uptake by the receptorcontaining liposomes in the absence of inhibitors was 39.9 ng protein/assay. Error barsrepresent the range of variations in triplicate determinations.
(% o
f co
ntr
ol
mS
R-B
I-t1
act
ivit
y)
[3H
]CE
-HD
L A
sso
ciat
ion
0
25
50
75
100
[3H]CE-HDL (10 g protein/ml)
HDL (400 g protein/ml)
KKB-1 (50 g/ml)Preimmune IgG (50 g/ml)
NonemSR-BI-t1
Immuno-PurifiedReceptor
by guest on February 14, 2018http://w
ww
.jbc.org/D
ownloaded from

Liu & Krieger page 49 In vitro assay of purified SR-BI
Figure 11. Temperature dependence of 125I-HDL association with and[3H]cholesteryl ether uptake by liposomes reconstituted withimmunoaffinity purified mSR-BI-t1. Immunoaffinity purified mSR-BI-t1isolated from HEK[mSR-BI-t1] cells was reconstituted into liposomes as described inExperimental Procedures. The liposomes were incubated with 10 µg protein/ml of 125I-HDL (A) or [3H]CE-HDL (B) at 37 oC (open bars) or 0oC (filled bars) for 2 hours in thepresence (single determinations) or absence (duplicate determinations) of a 40-foldexcess of unlabeled HDL, isolated and washed by filtration, and the amounts of specific125I-HDL binding and [3H]cholesteryl ether uptake were determined as described inExperimental Procedures. The 100% of control values for binding and lipid uptake at37°C were 4.1 and 35.7 ng protein/assay, respectively. The specific values for the control(no receptor) liposomes determined at 37°C were: 125I-HDL binding, 28%; and [3H]CEuptake, ~0% (not shown). The nonspecific background values for 125I-HDL binding were24% (37°C) and 22% (0°C) of the total binding. Error bars represent the range ofvariations in duplicate determinations.
B. [3H]CE-HDL
0
25
50
75
100
37
125 I
-HD
L o
r [3 H
]CE
-HD
LA
sso
ciat
ion
(% o
f ac
tivi
ty a
t 37o
C)
Temperature (°C)00 37
A. 125I-HDL
by guest on February 14, 2018http://w
ww
.jbc.org/D
ownloaded from

Bin Liu and Monty Kriegerselective lipid uptake
phosphatidylcholine /cholesterol liposomes mediates high affinity HDL binding and Highly purified scavenger receptor class B, type I reconstituted into
published online July 10, 2002J. Biol. Chem.
10.1074/jbc.M204265200Access the most updated version of this article at doi:
Alerts:
When a correction for this article is posted•
When this article is cited•
to choose from all of JBC's e-mail alertsClick here
by guest on February 14, 2018http://w
ww
.jbc.org/D
ownloaded from