live-cell imaging of endogenous ras-gtp shows predominant ras activation at the plasma membrane and...
TRANSCRIPT

Lt
SD
a
ARRAA
KYcSGNF
1
wadwccnTmiTts1pp(
UI
1h
The International Journal of Biochemistry & Cell Biology 45 (2013) 384– 394
Contents lists available at SciVerse ScienceDirect
The International Journal of Biochemistry& Cell Biology
journa l h o me page: www.elsev ier .com/ locate /b ioce l
ive-cell imaging of endogenous Ras-GTP shows predominant Ras activation athe plasma membrane and in the nucleus in Saccharomyces cerevisiae
erena Broggi, Enzo Martegani, Sonia Colombo ∗
epartment of Biotechnology and Biosciences, University of Milano-Bicocca, Milan, Italy
r t i c l e i n f o
rticle history:eceived 1 June 2012eceived in revised form 2 October 2012ccepted 25 October 2012vailable online 2 November 2012
eywords:eastAMP/PKA pathwaymall G proteins
a b s t r a c t
Ras proteins function as a point of convergence for different signalling pathways in eukaryotes and areinvolved in many cellular responses; their different subcellular locations could regulate distinct func-tions. To investigate the localization of active Ras in vivo in Saccharomyces cerevisiae, we expressed aprobe consisting of a GFP fusion with a trimeric Ras binding domain of Raf1 (eGFP-RBD3), which bindsRas-GTP with a much higher affinity than Ras-GDP. Our results show that in wild type cells active Rasaccumulates mainly at the plasma membrane and in the nucleus during growth on medium containingglucose, while it accumulates mainly in mitochondria in wild type glucose-starved cells and relocalizesto the plasma membrane and to the nucleus upon addition of this sugar. A similar pattern is observedin a strain deleted in the CYR1 gene indicating that the absence of adenylate cyclase does not impair the
pa2utrientluorescence microscopy
localization of Ras-GTP. Remarkably, in a gpa2�, but not in a gpr1� mutant, active Ras accumulates ininternal membranes and mitochondria, both when cells are growing on glucose medium or are starved,indicating that Gpa2, but not Gpr1 is required for the recruitment of Ras-GTP at the plasma membraneand in the nucleus. Moreover, deletion of both HXK1 and HXK2 also causes a mitochondrial localization
alizesgges
of the probe, which reloca centromeric plasmid, su
. Introduction
In the yeast Saccharomyces cerevisiae the Ras/cAMP/PKA path-ay is involved in the regulation of cell growth in response to
vailable nutrients and in the adaptation to glucose (Thevelein ande Winde, 1999; Busti et al., 2010). This pathway controls targetshich are involved in processes required for the shift from glu-
oneogenic to fermentative growth, in the breakdown of storagearbohydrates, in stress resistance, growth control and determi-ation of life span (Pedruzzi et al., 2003; Reinders et al., 1998;hevelein and de Winde, 1999; Wei et al., 2008). It also affectsorphogenesis and development, including pseudohyphal and
nvasive growth and sporulation capacity (Lengeler et al., 2000).he cyclic AMP is synthesized by adenylate cyclase, which is con-rolled by two different systems: the G-protein-coupled receptorystem (GPCR), acting through the G-protein Gpa2 (Colombo et al.,998; Nakafuku et al., 1988) and the Ras small G proteins. Ras
roteins activity is strictly controlled by two classes of regulatoryroteins: Cdc25 and Sdc25 guanine nucleotide exchange factorsGEFs) (Broach, 1991), which stimulate GDP-GTP exchange on Ras,∗ Corresponding author at: Department of Biotechnology and Biosciences,niversity of Milano-Bicocca, Piazza della Scienza 2, 20126, Milan,
taly. Tel.: +39 0264483551; fax: +39 0264483565.E-mail address: [email protected] (S. Colombo).
357-2725/$ – see front matter © 2012 Elsevier Ltd. All rights reserved.ttp://dx.doi.org/10.1016/j.biocel.2012.10.013
to the plasma membrane and to the nucleus upon expression of HXK2 onting that this kinase is involved in the proper localization of active Ras.
© 2012 Elsevier Ltd. All rights reserved.
and Ira1 and Ira2 GTPase activating proteins (GAPs), which promotethe intrinsically low Ras GTPase activity.
Using a sensitive GTP-loading assay for Ras2 we previouslyshowed that the increase in the level of cAMP after addition of glu-cose to glucose-starved cells is accompanied by a rapid increasein the level of Ras2-GTP/total Ras2 (Rudoni et al., 2001; Colomboet al., 2004). This assay measures only the “when” of activationand does not report anything about the “where” of signalling.Since Ras proteins transduce signals from the surface of the cell,it was generally considered that they localize and function onlyat the plasma membrane. Conversely, data provided by the yeastGFP fusion localization database (Ghaemmaghami et al., 2003)and recently published data (Belotti et al., 2011, 2012) suggestthat Ras2, Cyr1, Cdc25 and the Ira proteins are mainly localizedin internal membranes and only marginally at the plasma mem-brane level. Recent data have shown that mammalian Ras isoformsare differentially distributed within cell surface signalling nan-oclusters and on endomembranous compartments. In particular,Augsten et al. (2006) performed Ras-GTP imaging in mammaliancells using a probe consisting of a GFP fusion with a trimeric Rasbinding domain of Raf1 (eGFP-RBD3), which binds Ras-GTP witha much higher affinity than Ras-GDP, as a reporter for Ras activa-
tion. They could show that Golgi-located endogenous Ras is notunder the control of growth factors and argue for the plasmamembrane as the predominant site of agonist-induced Ras acti-vation.
S. Broggi et al. / The International Journal of Biochemistry & Cell Biology 45 (2013) 384– 394 385
Table 1List of yeast strains used in this study.
Strains Genotype Source/Ref.
W303-1A MATa ade2-1 can1-100 his3-11,15 leu2-3112 trp1-1 ura3-1 Thomas and Rothstein (1989)KT98 I MATa W303 expressing a fusion of Nup49 protein with the fluorescent protein tdimer Sheff and Thorn (2004)W303-1A [YCpRAS2Val19] Colombo et al. (2004)GG104 MATa W303 with pde2::TRP1, cyr1::KanMX2, msn2::HIS3, msn4::TRP1 Roosen et al. (2005)GG104 cdc25� GG104 with cdc25::URA3 This workYSH327 MATa W303 with hxk1::HIS3 hxk2::LEU2 de Winde et al. (1996)YSH297 MATa W303 with hxk1::HIS3 de Winde et al. (1996)YSH310 MATa W303 with hxk2::LEU2 de Winde et al. (1996)PM731 MATa W303 with gpa2::URA3 Colombo et al. (1998)LK42/RL5 MATa W303 with gpr1::LEU2 Kraakman et al. (1999)
iattttgioohli
2
2
SsIpfu(c
2
tbepEfSESpl
Kp
RGW29 MATa W303 with gpa2::LEU2 gpr1::KanMX4JT2083 MATa W303 with hxk1::HIS3, hxk2::TRP1
JT2083 [YCpHXK2]
In this work, in order to investigate the localization of active Rasn S. cerevisiae, the eGFP-RBD3 fusion was expressed in wild typend mutant strains in the cAMP/PKA pathway. Our results showhat the probe localizes essentially to the plasma membrane ando the nucleus of cells growing in fermentable carbon sources andhat some components of the cAMP/PKA pathway play a role inhe localization of active Ras. In particular, our data show that in apa2�, but not in a gpr1� strain, the eGFP-RBD3 probe accumulatesn internal membranes and mitochondria. Deletion of both the hex-kinases HXK1 and HXK2 also causes a mitochondrial localizationf Ras-GTP in glucose growing cells. The expression of HXK2 in thexk1� hxk2� mutant restores the nuclear and plasma membraneocalization of the probe, pointing to Hxk2 as the kinase involvedn the proper localization of active Ras.
. Experimental procedures
.1. Yeast strains and growth conditions
S. cerevisiae strains used in this study are listed in Table 1.ynthetic complete media (SD) containing the appropriate carbonource (2%), 6.7 g/l YNB w/o aminoacids (Becton and Dickinsontalia, Buccinasco) and the proper selective drop-out CSM (Com-lete Synthetic Medium, supplied by BIO101, CA, USA) was usedor the immunofluorescence experiments. The cell density of liq-id cultures grown at 30 ◦C was determined with a Coulter Countermodel Z2; Beckman Coulter, Cassina de Pecchi) on mildly soni-ated, diluted samples.
.2. Plasmids
To obtain pYX212-eGFP-RBD3 and pYX242-eGFP-RBD3 we usedhe following strategy. The peGFP-C2 construct (kindly providedy I. Rubio University of Jena, Germany) expressing the fusionGFP-RBD3 (Augsten et al., 2006) was digested with NheI, and theYX expression vectors, pYX212 and pYX242, were digested withcoRI. Filling in of the 3′ recessing ends was done using the Klenowragment of DNA polymerase I (New England Biolabs, MA, Unitedtates). The blunted DNA was then purified using the JETSORB Gelxtraction Kit (Genomed GmbH, Lönne, Gernany) and digested withalI. Finally, the fragment containing the fusion eGFP-RBD3 wasurified using the JETSORB Gel Extraction Kit and ligated with the
inearized vectors.Plasmid pmt-RFP was a kind gift of C.W. Gourlay (University of
ent). The mtRFP sequence is inserted in the centromeric plasmidYX142, under the control of the TPI promoter.
J. Thevelein (KU Leuven)J. Thevelein (KU Leuven)This work
2.3. Fluorescence microscopy
Cells were grown at 30 ◦C until exponential phase. Whenindicated, they were incubated with the mitochondrial marker rho-damine B hexyl ester perchlorate (Molecular Probes, Eugene, OR)100 nM final concentration for about 5–10 min before imaging. Sub-sequently, 20 �l of cells suspension were seeded on concanavalinA (Sigma–Aldrich, Milano, Italy)-coated cover glass for 10 min. Thecover glass was washed four times using the proper medium andput on top of a Thoma chamber. Images were acquired with aNikon Eclipse E600 microscope equipped with a 60X, 1.4 oil Plan-Apochromat objective and a standard FITC filter set (Nikon, EX450-490, DM 505, BA 520) for GFP fluorescence and a Cy3 filter setfor tdimer, RFP and rhodamine fluorescence. Images were recordeddigitally using a Leica DC 350F camera and processed using AdobePhotoshop (Adobe Systems, Inc.).
Determination of the spatio-temporal localization of Ras-GTPin vivo in single cells after addition of either 100 mM glucose or100 mM fructose was performed as following. Cells were grownin medium containing 2% glucose at 30 ◦C till exponential phase,collected by centrifugation and resuspended in 25 mM MES buffer,pH 6 (5 × 107 cells/ml) for at least 1 h. When indicated, cells weretreated with rhodamine B hexyl ester perchlorate as describedabove. Subsequently, 20 �l of glucose-starved cells were seeded onconcanavalin A-coated cover glass for 10 min. The cover glass waswashed four times using 1 ml of 25 mM MES buffer, pH 6 and puton top of a Thoma chamber. Images were acquired before and afteraddition of glucose with a Nikon Eclipse E600 microscope. Glucosewas added by pipetting, between the cover glass and the ThomaChamber, 20 �l of 3% glucose dissolved in 25 mM MES buffer, pH6. To avoid bleaching, the fluorescence images were acquired for1 s every minute for at least 10 min and the shutter was kept offin the meantime. The time series of images was then processedusing Adobe Photoshop (Adobe Systems, Inc.) and ImageJ software.The percentage of cells showing different subcellular localization ofthe fluorescent probe was calculated using the following criteria.Cells were divided into five classes, depending on the localizationof the probe: (1) nuclei; (2) plasma membrane; (3) nuclei + plasmamembrane; (4) mitochondria; (5) widespread (or diffuse). Cellsbelonging to each class were counted and expressed as a percent-age on the total number of fluorescent cells. Approximately 300cells were scored and error bars represent the standard deviationof 3 independent experiments. Examples of cells for each class areshown in Fig. 3.
For confocal microscopy, a Leica TCS SP2 confocal microscopeequipped with an inverted Leica DMIRE2 microscope and a PL APO
63× oil immersion objective (numerical aperture: 1.4) was used.An average of 10 optical sections were acquired for every singlecell (Z scan: 0.9 �m) and representative single optical sections wereshown.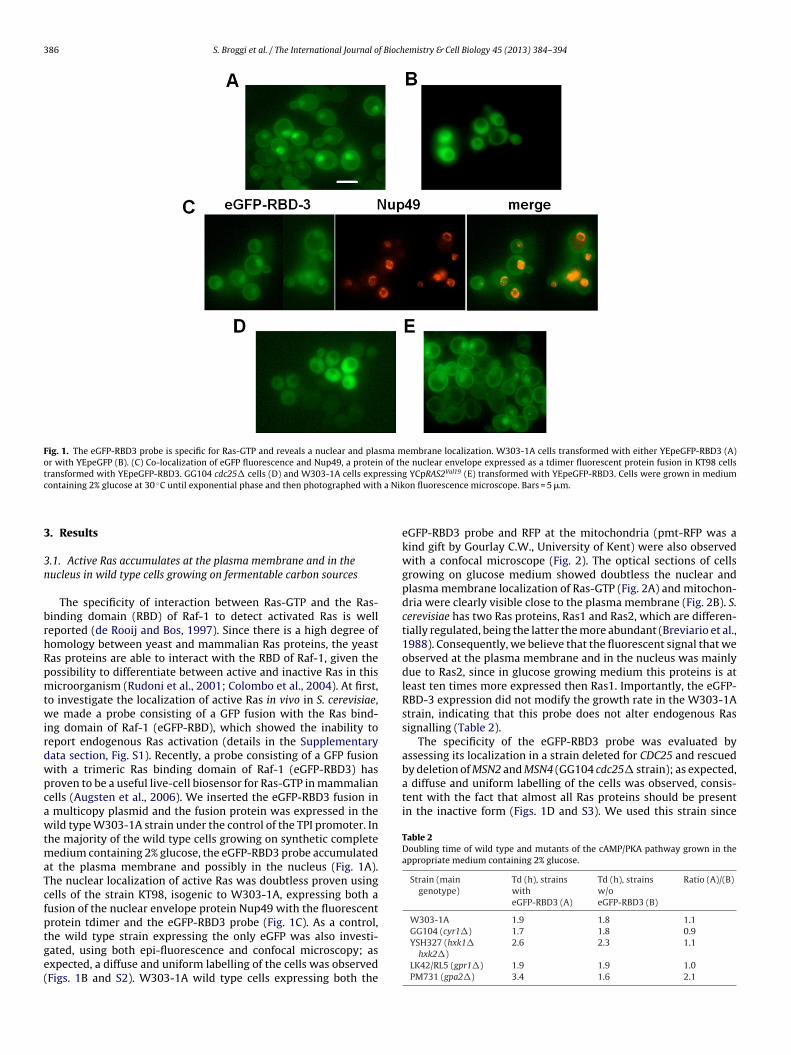
386 S. Broggi et al. / The International Journal of Biochemistry & Cell Biology 45 (2013) 384– 394
Fig. 1. The eGFP-RBD3 probe is specific for Ras-GTP and reveals a nuclear and plasma membrane localization. W303-1A cells transformed with either YEpeGFP-RBD3 (A)or with YEpeGFP (B). (C) Co-localization of eGFP fluorescence and Nup49, a protein of the nuclear envelope expressed as a tdimer fluorescent protein fusion in KT98 cellst ressin Val19
c a Nik
3
3n
brhRpmtwirdwpcawtmaTcfptge(
by deletion of MSN2 and MSN4 (GG104 cdc25� strain); as expected,a diffuse and uniform labelling of the cells was observed, consis-tent with the fact that almost all Ras proteins should be presentin the inactive form (Figs. 1D and S3). We used this strain since
Table 2Doubling time of wild type and mutants of the cAMP/PKA pathway grown in theappropriate medium containing 2% glucose.
Strain (maingenotype)
Td (h), strainswitheGFP-RBD3 (A)
Td (h), strainsw/oeGFP-RBD3 (B)
Ratio (A)/(B)
W303-1A 1.9 1.8 1.1GG104 (cyr1�) 1.7 1.8 0.9
ransformed with YEpeGFP-RBD3. GG104 cdc25� cells (D) and W303-1A cells expontaining 2% glucose at 30 ◦C until exponential phase and then photographed with
. Results
.1. Active Ras accumulates at the plasma membrane and in theucleus in wild type cells growing on fermentable carbon sources
The specificity of interaction between Ras-GTP and the Ras-inding domain (RBD) of Raf-1 to detect activated Ras is welleported (de Rooij and Bos, 1997). Since there is a high degree ofomology between yeast and mammalian Ras proteins, the yeastas proteins are able to interact with the RBD of Raf-1, given theossibility to differentiate between active and inactive Ras in thisicroorganism (Rudoni et al., 2001; Colombo et al., 2004). At first,
o investigate the localization of active Ras in vivo in S. cerevisiae,e made a probe consisting of a GFP fusion with the Ras bind-
ng domain of Raf-1 (eGFP-RBD), which showed the inability toeport endogenous Ras activation (details in the Supplementaryata section, Fig. S1). Recently, a probe consisting of a GFP fusionith a trimeric Ras binding domain of Raf-1 (eGFP-RBD3) hasroven to be a useful live-cell biosensor for Ras-GTP in mammalianells (Augsten et al., 2006). We inserted the eGFP-RBD3 fusion in
multicopy plasmid and the fusion protein was expressed in theild type W303-1A strain under the control of the TPI promoter. In
he majority of the wild type cells growing on synthetic completeedium containing 2% glucose, the eGFP-RBD3 probe accumulated
t the plasma membrane and possibly in the nucleus (Fig. 1A).he nuclear localization of active Ras was doubtless proven usingells of the strain KT98, isogenic to W303-1A, expressing both ausion of the nuclear envelope protein Nup49 with the fluorescentrotein tdimer and the eGFP-RBD3 probe (Fig. 1C). As a control,
he wild type strain expressing the only eGFP was also investi-ated, using both epi-fluorescence and confocal microscopy; asxpected, a diffuse and uniform labelling of the cells was observedFigs. 1B and S2). W303-1A wild type cells expressing both theg YCpRAS2 (E) transformed with YEpeGFP-RBD3. Cells were grown in mediumon fluorescence microscope. Bars = 5 �m.
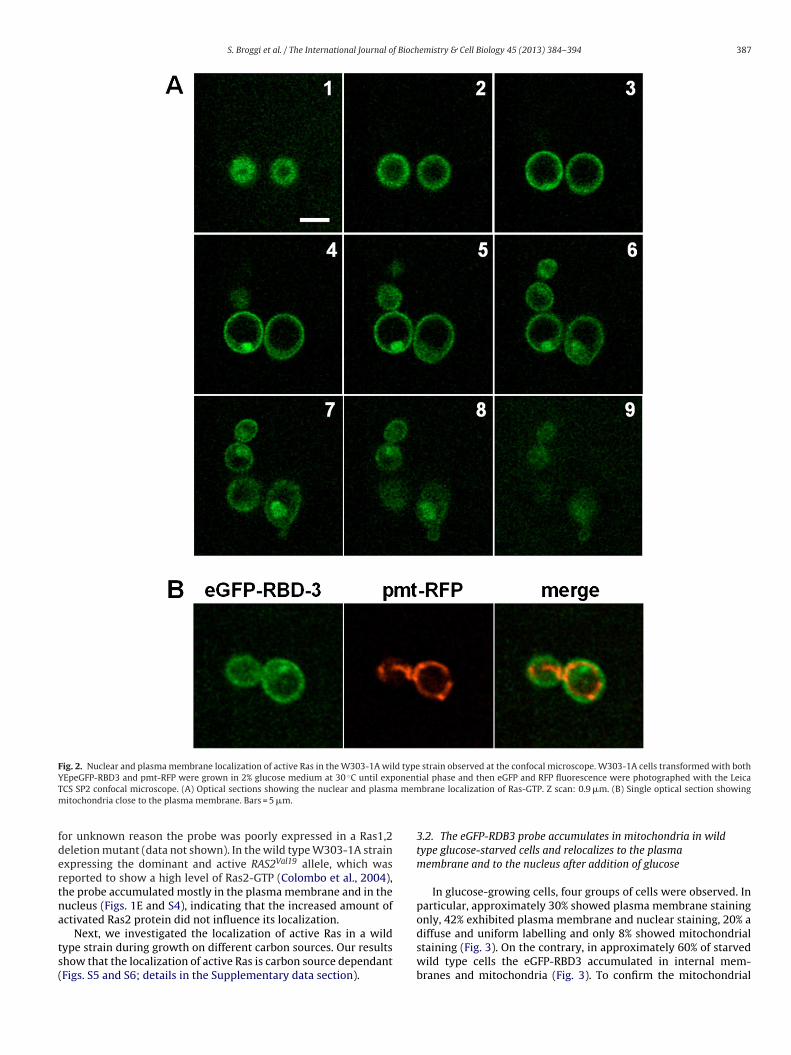
eGFP-RBD3 probe and RFP at the mitochondria (pmt-RFP was akind gift by Gourlay C.W., University of Kent) were also observedwith a confocal microscope (Fig. 2). The optical sections of cellsgrowing on glucose medium showed doubtless the nuclear andplasma membrane localization of Ras-GTP (Fig. 2A) and mitochon-dria were clearly visible close to the plasma membrane (Fig. 2B). S.cerevisiae has two Ras proteins, Ras1 and Ras2, which are differen-tially regulated, being the latter the more abundant (Breviario et al.,1988). Consequently, we believe that the fluorescent signal that weobserved at the plasma membrane and in the nucleus was mainlydue to Ras2, since in glucose growing medium this proteins is atleast ten times more expressed then Ras1. Importantly, the eGFP-RBD-3 expression did not modify the growth rate in the W303-1Astrain, indicating that this probe does not alter endogenous Rassignalling (Table 2).
The specificity of the eGFP-RBD3 probe was evaluated byassessing its localization in a strain deleted for CDC25 and rescued
YSH327 (hxk1�hxk2�)
2.6 2.3 1.1
LK42/RL5 (gpr1�) 1.9 1.9 1.0PM731 (gpa2�) 3.4 1.6 2.1
S. Broggi et al. / The International Journal of Biochemistry & Cell Biology 45 (2013) 384– 394 387
Fig. 2. Nuclear and plasma membrane localization of active Ras in the W303-1A wild type strain observed at the confocal microscope. W303-1A cells transformed with bothY ◦ onentT a memm
fdertna
ts(
EpeGFP-RBD3 and pmt-RFP were grown in 2% glucose medium at 30 C until expCS SP2 confocal microscope. (A) Optical sections showing the nuclear and plasmitochondria close to the plasma membrane. Bars = 5 �m.
or unknown reason the probe was poorly expressed in a Ras1,2eletion mutant (data not shown). In the wild type W303-1A strainxpressing the dominant and active RAS2Val19 allele, which waseported to show a high level of Ras2-GTP (Colombo et al., 2004),he probe accumulated mostly in the plasma membrane and in theucleus (Figs. 1E and S4), indicating that the increased amount ofctivated Ras2 protein did not influence its localization.
Next, we investigated the localization of active Ras in a wildype strain during growth on different carbon sources. Our resultshow that the localization of active Ras is carbon source dependantFigs. S5 and S6; details in the Supplementary data section).
ial phase and then eGFP and RFP fluorescence were photographed with the Leicabrane localization of Ras-GTP. Z scan: 0.9 �m. (B) Single optical section showing
3.2. The eGFP-RDB3 probe accumulates in mitochondria in wildtype glucose-starved cells and relocalizes to the plasmamembrane and to the nucleus after addition of glucose
In glucose-growing cells, four groups of cells were observed. Inparticular, approximately 30% showed plasma membrane stainingonly, 42% exhibited plasma membrane and nuclear staining, 20% a
diffuse and uniform labelling and only 8% showed mitochondrialstaining (Fig. 3). On the contrary, in approximately 60% of starvedwild type cells the eGFP-RBD3 accumulated in internal mem-branes and mitochondria (Fig. 3). To confirm the mitochondrial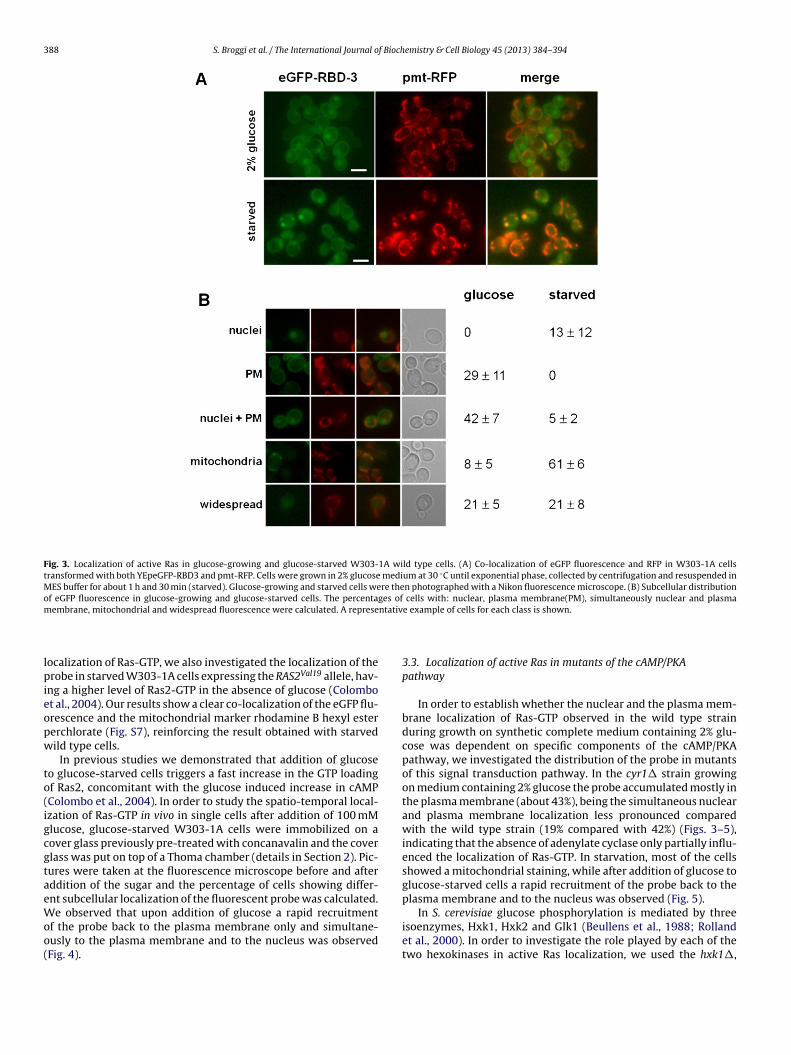
388 S. Broggi et al. / The International Journal of Biochemistry & Cell Biology 45 (2013) 384– 394
Fig. 3. Localization of active Ras in glucose-growing and glucose-starved W303-1A wild type cells. (A) Co-localization of eGFP fluorescence and RFP in W303-1A cellstransformed with both YEpeGFP-RBD3 and pmt-RFP. Cells were grown in 2% glucose medium at 30 ◦C until exponential phase, collected by centrifugation and resuspended inMES buffer for about 1 h and 30 min (starved). Glucose-growing and starved cells were then photographed with a Nikon fluorescence microscope. (B) Subcellular distributionof eGFP fluorescence in glucose-growing and glucose-starved cells. The percentages of cells with: nuclear, plasma membrane(PM), simultaneously nuclear and plasmam ntativ
lpieopw
to(igcgtaeWoo(
In S. cerevisiae glucose phosphorylation is mediated by threeisoenzymes, Hxk1, Hxk2 and Glk1 (Beullens et al., 1988; Rolland
embrane, mitochondrial and widespread fluorescence were calculated. A represe
ocalization of Ras-GTP, we also investigated the localization of therobe in starved W303-1A cells expressing the RAS2Val19 allele, hav-
ng a higher level of Ras2-GTP in the absence of glucose (Colombot al., 2004). Our results show a clear co-localization of the eGFP flu-rescence and the mitochondrial marker rhodamine B hexyl estererchlorate (Fig. S7), reinforcing the result obtained with starvedild type cells.
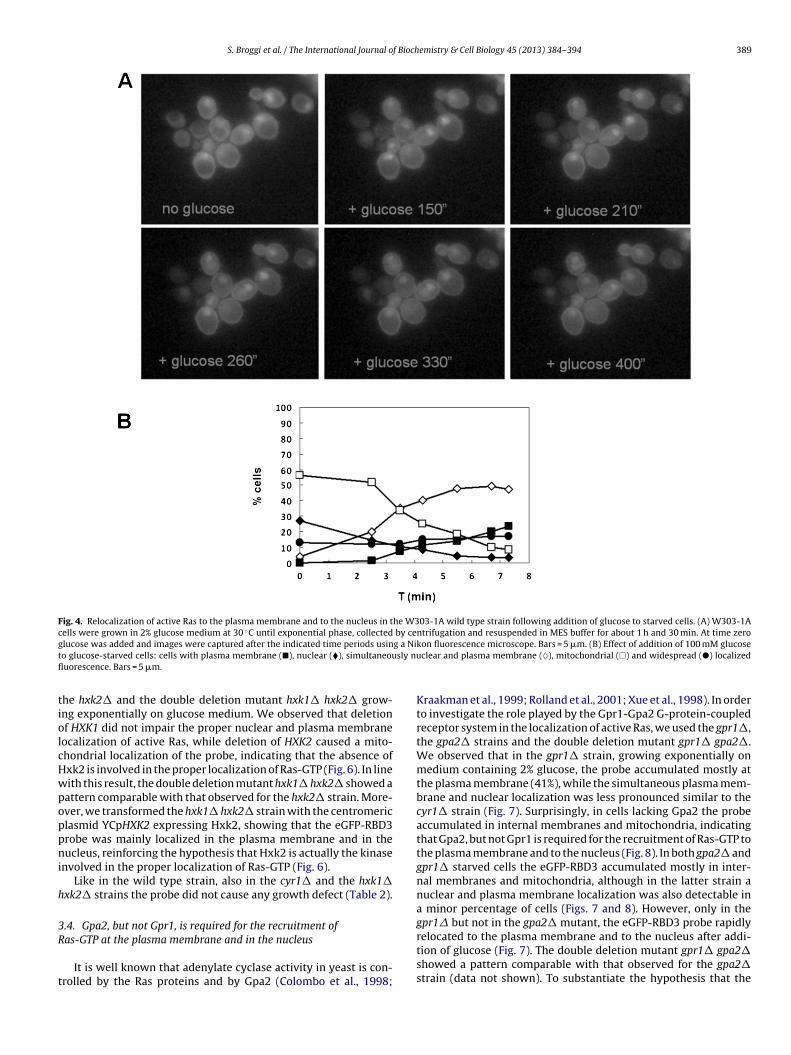
In previous studies we demonstrated that addition of glucoseo glucose-starved cells triggers a fast increase in the GTP loadingf Ras2, concomitant with the glucose induced increase in cAMPColombo et al., 2004). In order to study the spatio-temporal local-zation of Ras-GTP in vivo in single cells after addition of 100 mMlucose, glucose-starved W303-1A cells were immobilized on aover glass previously pre-treated with concanavalin and the coverlass was put on top of a Thoma chamber (details in Section 2). Pic-ures were taken at the fluorescence microscope before and afterddition of the sugar and the percentage of cells showing differ-nt subcellular localization of the fluorescent probe was calculated.e observed that upon addition of glucose a rapid recruitment
f the probe back to the plasma membrane only and simultane-
usly to the plasma membrane and to the nucleus was observedFig. 4).e example of cells for each class is shown.
3.3. Localization of active Ras in mutants of the cAMP/PKApathway
In order to establish whether the nuclear and the plasma mem-brane localization of Ras-GTP observed in the wild type strainduring growth on synthetic complete medium containing 2% glu-cose was dependent on specific components of the cAMP/PKApathway, we investigated the distribution of the probe in mutantsof this signal transduction pathway. In the cyr1� strain growingon medium containing 2% glucose the probe accumulated mostly inthe plasma membrane (about 43%), being the simultaneous nuclearand plasma membrane localization less pronounced comparedwith the wild type strain (19% compared with 42%) (Figs. 3–5),indicating that the absence of adenylate cyclase only partially influ-enced the localization of Ras-GTP. In starvation, most of the cellsshowed a mitochondrial staining, while after addition of glucose toglucose-starved cells a rapid recruitment of the probe back to theplasma membrane and to the nucleus was observed (Fig. 5).
et al., 2000). In order to investigate the role played by each of thetwo hexokinases in active Ras localization, we used the hxk1�,
S. Broggi et al. / The International Journal of Biochemistry & Cell Biology 45 (2013) 384– 394 389
Fig. 4. Relocalization of active Ras to the plasma membrane and to the nucleus in the W303-1A wild type strain following addition of glucose to starved cells. (A) W303-1Acells were grown in 2% glucose medium at 30 ◦C until exponential phase, collected by centrifugation and resuspended in MES buffer for about 1 h and 30 min. At time zerog g a Nit sly nufl
tiolcHwpoppni
h
3R
t
lucose was added and images were captured after the indicated time periods usino glucose-starved cells: cells with plasma membrane (�), nuclear (�), simultaneouuorescence. Bars = 5 �m.
he hxk2� and the double deletion mutant hxk1� hxk2� grow-ng exponentially on glucose medium. We observed that deletionf HXK1 did not impair the proper nuclear and plasma membraneocalization of active Ras, while deletion of HXK2 caused a mito-hondrial localization of the probe, indicating that the absence ofxk2 is involved in the proper localization of Ras-GTP (Fig. 6). In lineith this result, the double deletion mutant hxk1� hxk2� showed aattern comparable with that observed for the hxk2� strain. More-ver, we transformed the hxk1� hxk2� strain with the centromericlasmid YCpHXK2 expressing Hxk2, showing that the eGFP-RBD3robe was mainly localized in the plasma membrane and in theucleus, reinforcing the hypothesis that Hxk2 is actually the kinase
nvolved in the proper localization of Ras-GTP (Fig. 6).Like in the wild type strain, also in the cyr1� and the hxk1�
xk2� strains the probe did not cause any growth defect (Table 2).
.4. Gpa2, but not Gpr1, is required for the recruitment of
as-GTP at the plasma membrane and in the nucleusIt is well known that adenylate cyclase activity in yeast is con-rolled by the Ras proteins and by Gpa2 (Colombo et al., 1998;
kon fluorescence microscope. Bars = 5 �m. (B) Effect of addition of 100 mM glucoseclear and plasma membrane (♦), mitochondrial (�) and widespread (�) localized
Kraakman et al., 1999; Rolland et al., 2001; Xue et al., 1998). In orderto investigate the role played by the Gpr1-Gpa2 G-protein-coupledreceptor system in the localization of active Ras, we used the gpr1�,the gpa2� strains and the double deletion mutant gpr1� gpa2�.We observed that in the gpr1� strain, growing exponentially onmedium containing 2% glucose, the probe accumulated mostly atthe plasma membrane (41%), while the simultaneous plasma mem-brane and nuclear localization was less pronounced similar to thecyr1� strain (Fig. 7). Surprisingly, in cells lacking Gpa2 the probeaccumulated in internal membranes and mitochondria, indicatingthat Gpa2, but not Gpr1 is required for the recruitment of Ras-GTP tothe plasma membrane and to the nucleus (Fig. 8). In both gpa2� andgpr1� starved cells the eGFP-RBD3 accumulated mostly in inter-nal membranes and mitochondria, although in the latter strain anuclear and plasma membrane localization was also detectable ina minor percentage of cells (Figs. 7 and 8). However, only in thegpr1� but not in the gpa2� mutant, the eGFP-RBD3 probe rapidly
relocated to the plasma membrane and to the nucleus after addi-tion of glucose (Fig. 7). The double deletion mutant gpr1� gpa2�showed a pattern comparable with that observed for the gpa2�strain (data not shown). To substantiate the hypothesis that the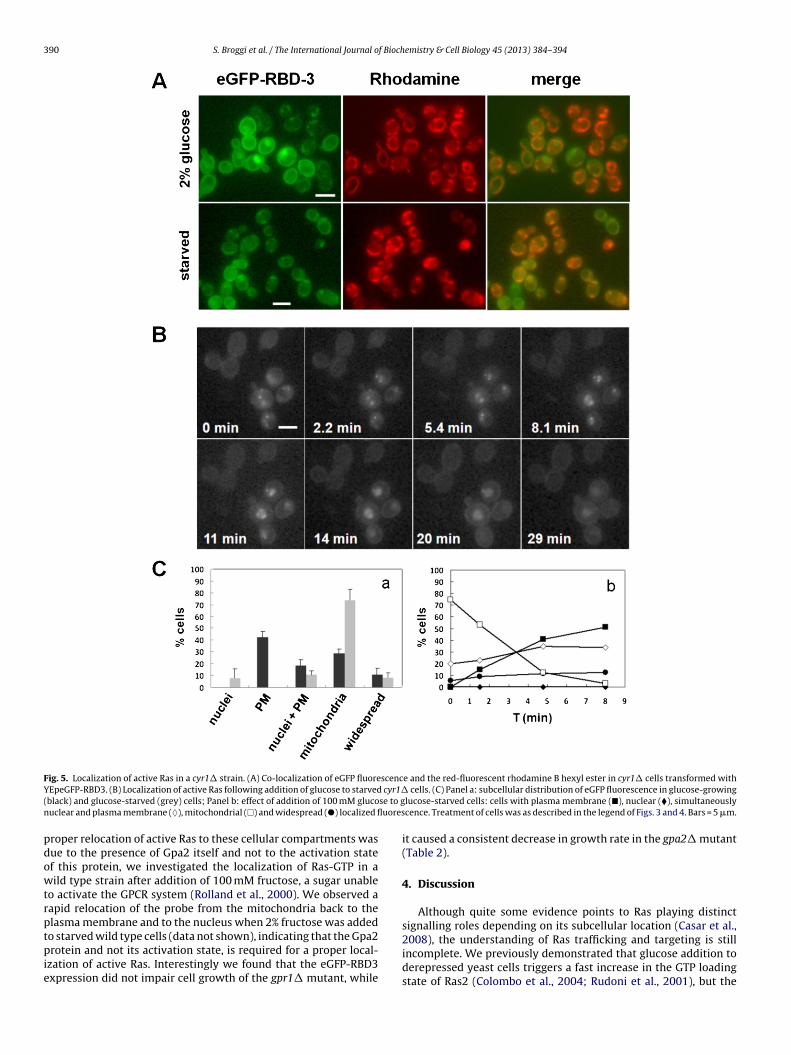
390 S. Broggi et al. / The International Journal of Biochemistry & Cell Biology 45 (2013) 384– 394
Fig. 5. Localization of active Ras in a cyr1� strain. (A) Co-localization of eGFP fluorescence and the red-fluorescent rhodamine B hexyl ester in cyr1� cells transformed withY cyr1�( se to gn uores
pdowtrptpie
EpeGFP-RBD3. (B) Localization of active Ras following addition of glucose to starvedblack) and glucose-starved (grey) cells; Panel b: effect of addition of 100 mM glucouclear and plasma membrane (♦), mitochondrial (�) and widespread (�) localized fl
roper relocation of active Ras to these cellular compartments wasue to the presence of Gpa2 itself and not to the activation statef this protein, we investigated the localization of Ras-GTP in aild type strain after addition of 100 mM fructose, a sugar unable
o activate the GPCR system (Rolland et al., 2000). We observed aapid relocation of the probe from the mitochondria back to thelasma membrane and to the nucleus when 2% fructose was added
o starved wild type cells (data not shown), indicating that the Gpa2rotein and not its activation state, is required for a proper local-zation of active Ras. Interestingly we found that the eGFP-RBD3xpression did not impair cell growth of the gpr1� mutant, while
cells. (C) Panel a: subcellular distribution of eGFP fluorescence in glucose-growinglucose-starved cells: cells with plasma membrane (�), nuclear (�), simultaneouslycence. Treatment of cells was as described in the legend of Figs. 3 and 4. Bars = 5 �m.
it caused a consistent decrease in growth rate in the gpa2� mutant(Table 2).
4. Discussion
Although quite some evidence points to Ras playing distinctsignalling roles depending on its subcellular location (Casar et al.,
2008), the understanding of Ras trafficking and targeting is stillincomplete. We previously demonstrated that glucose addition toderepressed yeast cells triggers a fast increase in the GTP loadingstate of Ras2 (Colombo et al., 2004; Rudoni et al., 2001), but the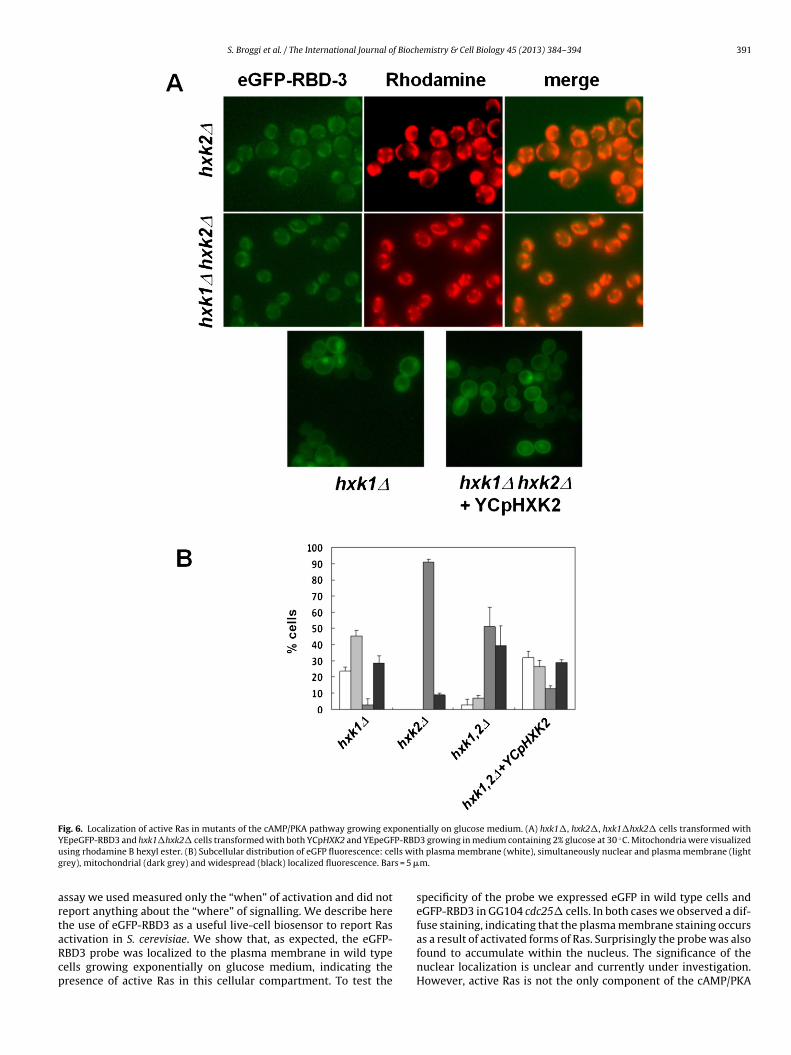
S. Broggi et al. / The International Journal of Biochemistry & Cell Biology 45 (2013) 384– 394 391
Fig. 6. Localization of active Ras in mutants of the cAMP/PKA pathway growing exponentially on glucose medium. (A) hxk1�, hxk2�, hxk1�hxk2� cells transformed withY P-RBD ◦
u lls witg s = 5 �
artaRcp
EpeGFP-RBD3 and hxk1�hxk2� cells transformed with both YCpHXK2 and YEpeGFsing rhodamine B hexyl ester. (B) Subcellular distribution of eGFP fluorescence: cerey), mitochondrial (dark grey) and widespread (black) localized fluorescence. Bar
ssay we used measured only the “when” of activation and did noteport anything about the “where” of signalling. We describe herehe use of eGFP-RBD3 as a useful live-cell biosensor to report Ras
ctivation in S. cerevisiae. We show that, as expected, the eGFP-BD3 probe was localized to the plasma membrane in wild typeells growing exponentially on glucose medium, indicating theresence of active Ras in this cellular compartment. To test the3 growing in medium containing 2% glucose at 30 C. Mitochondria were visualizedh plasma membrane (white), simultaneously nuclear and plasma membrane (lightm.
specificity of the probe we expressed eGFP in wild type cells andeGFP-RBD3 in GG104 cdc25� cells. In both cases we observed a dif-fuse staining, indicating that the plasma membrane staining occurs
as a result of activated forms of Ras. Surprisingly the probe was alsofound to accumulate within the nucleus. The significance of thenuclear localization is unclear and currently under investigation.However, active Ras is not the only component of the cAMP/PKA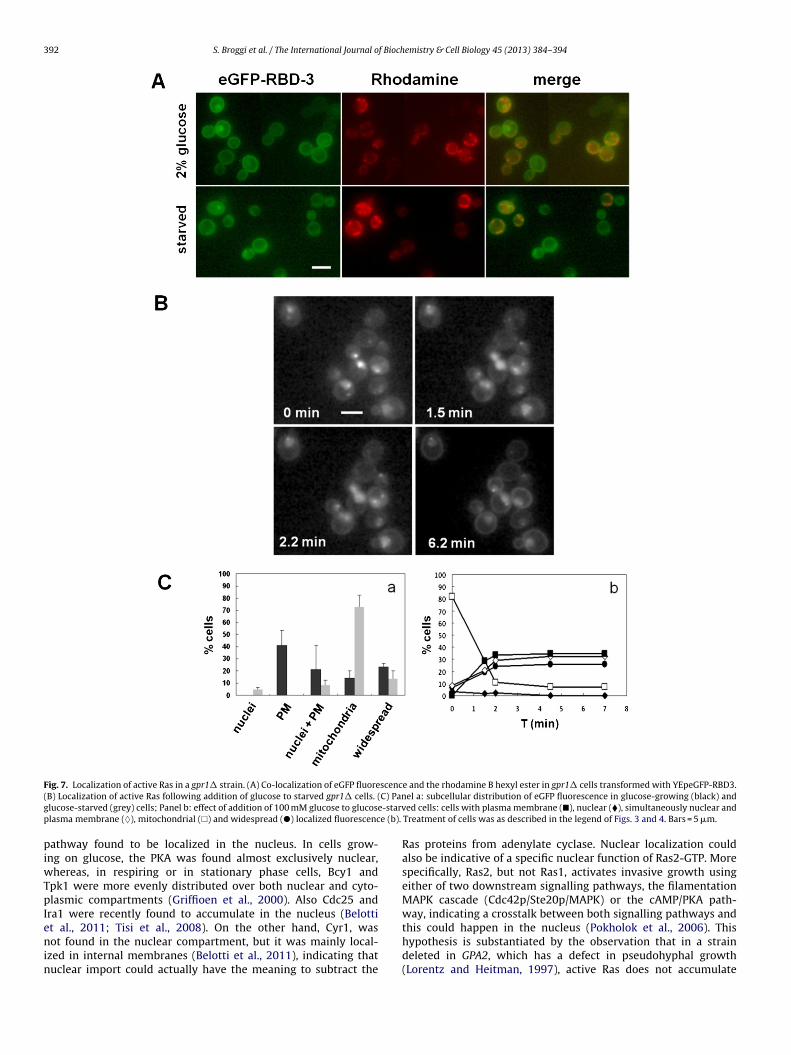
392 S. Broggi et al. / The International Journal of Biochemistry & Cell Biology 45 (2013) 384– 394
Fig. 7. Localization of active Ras in a gpr1� strain. (A) Co-localization of eGFP fluorescence and the rhodamine B hexyl ester in gpr1� cells transformed with YEpeGFP-RBD3.( C) Pang -starvp e (b).
piwTpIenin
B) Localization of active Ras following addition of glucose to starved gpr1� cells. (lucose-starved (grey) cells; Panel b: effect of addition of 100 mM glucose to glucoselasma membrane (♦), mitochondrial (�) and widespread (�) localized fluorescenc
athway found to be localized in the nucleus. In cells grow-ng on glucose, the PKA was found almost exclusively nuclear,
hereas, in respiring or in stationary phase cells, Bcy1 andpk1 were more evenly distributed over both nuclear and cyto-lasmic compartments (Griffioen et al., 2000). Also Cdc25 and
ra1 were recently found to accumulate in the nucleus (Belottit al., 2011; Tisi et al., 2008). On the other hand, Cyr1, was
ot found in the nuclear compartment, but it was mainly local-zed in internal membranes (Belotti et al., 2011), indicating thatuclear import could actually have the meaning to subtract the
el a: subcellular distribution of eGFP fluorescence in glucose-growing (black) anded cells: cells with plasma membrane (�), nuclear (�), simultaneously nuclear and
Treatment of cells was as described in the legend of Figs. 3 and 4. Bars = 5 �m.
Ras proteins from adenylate cyclase. Nuclear localization couldalso be indicative of a specific nuclear function of Ras2-GTP. Morespecifically, Ras2, but not Ras1, activates invasive growth usingeither of two downstream signalling pathways, the filamentationMAPK cascade (Cdc42p/Ste20p/MAPK) or the cAMP/PKA path-way, indicating a crosstalk between both signalling pathways andthis could happen in the nucleus (Pokholok et al., 2006). This
hypothesis is substantiated by the observation that in a straindeleted in GPA2, which has a defect in pseudohyphal growth(Lorentz and Heitman, 1997), active Ras does not accumulate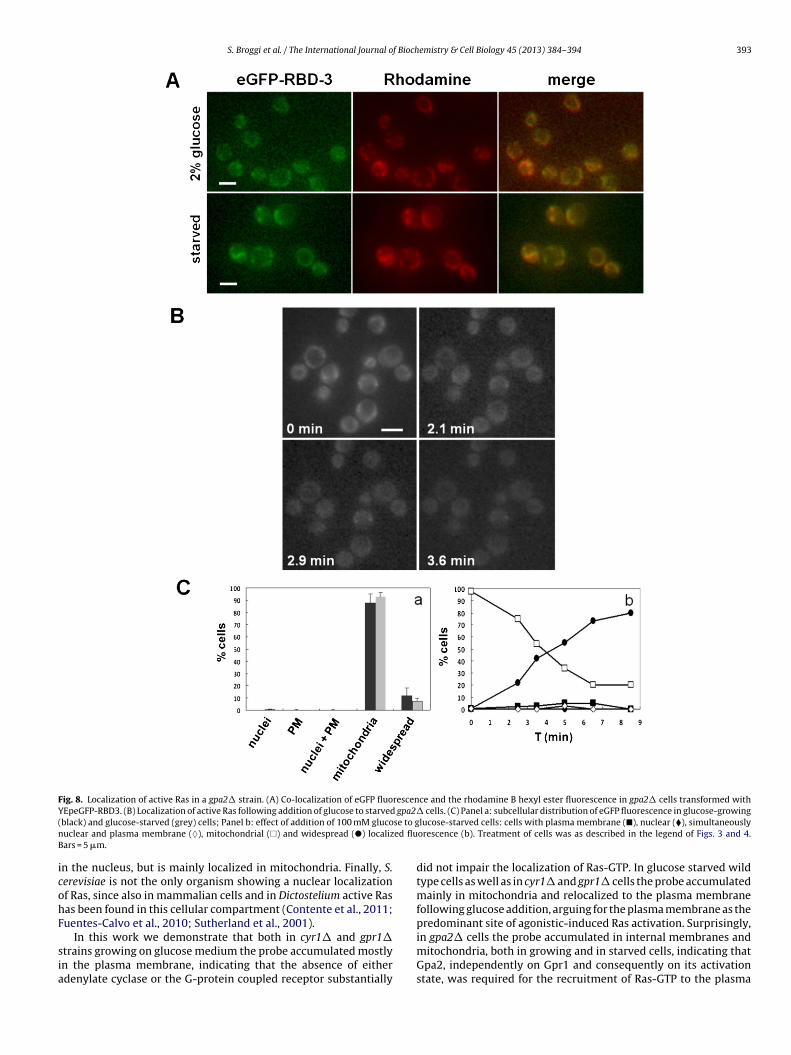
S. Broggi et al. / The International Journal of Biochemistry & Cell Biology 45 (2013) 384– 394 393
Fig. 8. Localization of active Ras in a gpa2� strain. (A) Co-localization of eGFP fluorescence and the rhodamine B hexyl ester fluorescence in gpa2� cells transformed withY gpa2�( se to gn ed fluB
icohF
sia
EpeGFP-RBD3. (B) Localization of active Ras following addition of glucose to starvedblack) and glucose-starved (grey) cells; Panel b: effect of addition of 100 mM glucouclear and plasma membrane (♦), mitochondrial (�) and widespread (�) localizars = 5 �m.
n the nucleus, but is mainly localized in mitochondria. Finally, S.erevisiae is not the only organism showing a nuclear localizationf Ras, since also in mammalian cells and in Dictostelium active Rasas been found in this cellular compartment (Contente et al., 2011;uentes-Calvo et al., 2010; Sutherland et al., 2001).
In this work we demonstrate that both in cyr1� and gpr1�trains growing on glucose medium the probe accumulated mostlyn the plasma membrane, indicating that the absence of eitherdenylate cyclase or the G-protein coupled receptor substantially
cells. (C) Panel a: subcellular distribution of eGFP fluorescence in glucose-growinglucose-starved cells: cells with plasma membrane (�), nuclear (�), simultaneouslyorescence (b). Treatment of cells was as described in the legend of Figs. 3 and 4.
did not impair the localization of Ras-GTP. In glucose starved wildtype cells as well as in cyr1� and gpr1� cells the probe accumulatedmainly in mitochondria and relocalized to the plasma membranefollowing glucose addition, arguing for the plasma membrane as thepredominant site of agonistic-induced Ras activation. Surprisingly,
in gpa2� cells the probe accumulated in internal membranes andmitochondria, both in growing and in starved cells, indicating thatGpa2, independently on Gpr1 and consequently on its activationstate, was required for the recruitment of Ras-GTP to the plasma
3 f Bioch
mbv(ptocmrmcRtsmcpitRcmemi
A
hRpvffs
A
f2
R
A
B
B
B
B
B
B
C
C
94 S. Broggi et al. / The International Journal o
embrane and to the nucleus. This result points to an uncouplingetween Gpa2 and Gpr1, uncoupling which has also been pre-iously reported using a global transcriptional analysis approachZaman et al., 2009). Moreover, we show that the nuclear and thelasma membrane localization of Ras-GTP observed in the wildype strain during growth on glucose medium was also dependentn hexokinase 2, since in the absence of this protein the fluores-ence was mainly located in mitochondria. The significance of theitochondrial localization of the probe is unclear. However other
eports have found Ras associated with mitochondria, both in mam-als and lower eukaryotes. Leadsham et al. (2008) showed that
ells lacking Whi2p exhibit an aberrant accumulation of activatedas2 at the mitochondria in response to nutritional depletion. Inhis mutant, the failure to address Ras2 to the vacuole and con-equently the failure to shut down Ras signalling would lead toitochondrial dysfunction, the accumulation of damaging ROS and
ell death. Disruption of class C VPS genes, encoding for proteinslaying a role in endosome and vacuole membrane fusion, results
n mitochondrial defects and an accumulation of total Ras pro-eins on mitochondrial membranes (Wang and Deschenes, 2006).as proteins have been shown to fractionate with the outer mito-hondrial membrane also in wild-type cells, pointing to a role ofitochondria in the nonclassical pathway of Ras trafficking (Dong
t al., 2003; Wang and Deschenes, 2006). The functional involve-ent of Ras proteins in mitochondrial function is currently under
nvestigation.
cknowledgements
We thank Ignacio Rubio (Friedrich Schiller-University Jena) foris kind gift of the peGFP-C2 construct expressing the fusion eGFP-BD3 and C.W. Gourlay (University of Kent) for his kind gift oflasmid pmt-RFP. We thank J. Winderickx and J. Thevelein, Uni-ersity of Leuven, Belgium and K. Thorn, Harvard University, USAor the gift of strains. We thank A. Villa (University Milano-Bicocca)or her kind help with the confocal microscope. This work wasupported by FAR grant to E.M. and S.C. and UniCellSys.
ppendix A. Supplementary data
Supplementary data associated with this article can beound, in the online version, at http://dx.doi.org/10.1016/j.biocel.012.10.013.
eferences
ugsten M, Pusch R, Biskup C, Rennert K, Wittig U, Beyer K, et al. Live-cell imagingof endogenous Ras-GTP illustrates predominant Ras activation at the plasmamembrane. EMBO Reports 2006;7:46–51.
elotti F, Tisi R, Paiardi C, Groppi S, Martegani E. PKA-dependent regulation of Cdc25RasGEF localization in budding yeast. FEBS Letters 2011;585:3914–20.
elotti F, Tisi R, Paiardi C, Rigamonti M, Groppi S, Martegani E. Localizationof Ras signaling complex in budding yeast. Biochimica et Biophysica Acta2012;1823:1208–16.
eullens M, Mbonyi K, Geerts L, Gladines D, Detremerie K, Jans AW, et al. Studieson the mechanism of the glucose-induced cAMP signal in glycolysis and glucoserepression mutants of the yeast Saccharomyces cerevisiae. European Journal ofBiochemistry 1988;172:227–31.
reviario D, Hinnebusch AG, Dhar R. Multiple regulatory mechanisms control theexpression of the RAS1 and RAS2 genes of Saccharomyces cerevisiae. EMBO Jour-nal 1988;7:1805–13.
roach J. RAS genes in Saccharomyces cerevisiae: signal transduction in search of apathway. Trends in Genetics 1991;7:28–33.
usti S, Coccetti P, Alberghina L, Vanoni M. Glucose signalling-mediated coordina-tion of cell growth and cell cycle in Saccharomyces cerevisiae. Sensors (Basel)2010;10(6):6195–240.
asar B, Arozarena I, Sanz-Moreno V, Pinto A, Agudo-Ibanez L, Marais R, et al. Rassubcellular localization defines ERK1/2 substrate specificity through distinct uti-lization of scaffold proteins. Molecular and Cellular Biology 2008;29:1338–53.
olombo S, Ma P, Cauwenberg L, Winderickx J, Crauwels M, Teunissen A, et al.Involvement of distant G-proteins, Gpa2 and Ras, in glucose- and intracellular
emistry & Cell Biology 45 (2013) 384– 394
acidification-induced cAMP signalling in the yeast Saccharomyces cerevisiae.EMBO Journal 1998;17:3326–41.
Colombo S, Ronchetti D, Thevelein JM, Winderickx J, Martegani E. Activation stateof the Ras2 protein and glucose-induced signalling in Saccharomyces cerevisiae.Journal of Biological Chemistry 2004;279:46715–22.
Contente S, Yeh TA, Friedman RM. H-Ras localizes to cell nuclei and varies with cellcycle. Genes Cancer 2011;20:1–7.
de Rooij J, Bos JL. Minimal Ras-binding domain of Raf-1 can be used as an activation-specific probe for Ras. Oncogene 1997;14:623–5.
de Winde JH, Crauwels M, Hohmann S, Thevelein JM, Winderickx J. Differentialrequirement of the yeast sugar kinases for sugar sensing in establishing thecatabolite-repressed state. European Journal of Biochemistry 1996;241:633–43.
Dong X, Mitchell DA, Lobo S, Zhao L, Bartels DJ, Deschenes RJ. Palmytoilation andplasma membrane localization of Ras2p by a nonclassical trafficking pathwayin Saccharomyces cerevisiae. Molecular and Cellular Biology 2003;23:6574–84.
Fuentes-Calvo I, Blázquez-Medela AM, Santos E, López-Novoa JM, Martínez-SalgadoC. Analysis of k-ras nuclear expression in fibroblasts and mesangial cells. PLoSOne 2010;5:e8703.
Ghaemmaghami S, Huh W, Bower K, Howson R, Belle A, Dephoure N, et al. Globalanalysis of protein expression in yeast. Nature 2003;425:737–41.
Griffioen G, Anghileri P, Imre E, Baroni MD, Ruis H. Nutritional control of nucle-ocytoplasmatic localization of cAMP-dependent protein kinase catalytic andregulatory subunits in Saccharomyces cerevisiae. Journal of Biological Chemistry2000;275:1449–56.
Kraakman L, Lemaire K, Ma P, Teunissen AW, Donaton MC, Van Dijck P, et al. A Sac-charomyces cerevisiae G-protein coupled receptor Gpr1, is specifically requiredfor glucose activation of the cAMP pathway during the transition to growth onglucose. Molecular Microbiology 1999;32:002–1012.
Leadsham JE, Miller K, Ayscough KR, Colombo S, Martegani E, Sudbery P, et al.Whi2p links nutritional sensing to actin-dependent Ras-cAMP-PKA regulationand apoptosis in yeast. Journal of Cell Science 2008;122:706–15.
Lengeler KB, Davison RC, D’Souza C, Harashima T, Shen W, Wang P, et al. Signal trans-duction cascades regulating fungal development and virulence. Microbiologyand Molecular Biology Reviews 2000;64:746–85.
Lorentz MC, Heitman J. Yeast pseudohyphal growth is regulated by Gpa2, a G protein� homolog. EMBO Journal 1997;16:7008–18.
Nakafuku M, Obara T, Kaibuchi K, Miyajima I, Miyajima A, Itoh H, et al. Isolationof a second yeast Saccharomyces cerevisiae gene (GPA2) coding for guaninenucleotide-binding regulatory protein: studies on its structure and possiblefunctions. Proceedings of the National Academy of Sciences of the United Statesof America 1988;85:1374–8.
Pedruzzi I, Dubouloz F, Cameroni E, Wanke V, Roosen J, Winderickx J, et al. TOR andPKA Signalling pathways converge on the protein kinase Rim15 to control entryinto G0. Molecular Cell 2003;12:1607–13.
Pokholok DK, Zeitlinger J, Hannett NM, Reynolds DB, Young RA. Activated signaltransduction kinases frequently occupy target genes. Science 2006;28:533–6.
Reinders A, Burckert N, Boller T, Wiemken A, De Virgilio C. Saccharomyces cerevisiaecAMP dependent protein kinase controls entry into stationary phase throughthe Rim15p protein kinase. Genes and Development 1998;12:2943–55.
Rolland F, de Winde JH, Lemaire K, Boles E, Thevelein JM, Winderickx J. Glucose-induced cAMP signalling in yeast requires both a G-protein coupled receptorsystem for extracellular glucose detection and a separable hexose kinase-dependent sensing process. Molecular Microbiology 2000;38:348–435.
Rolland F, Wanke V, Cauwenberg L, Ma P, Boles E, Vanoni M, et al. The role of hexosetransport and phosphorylation in cAMP signalling in the yeast Saccharomycescerevisiae. FEMS Yeast Research 2001;1403:1–13.
Roosen J, Engelen K, Marchal K, Mathys J, Griffioen G, Cameroni E, et al. PKA andSch9 control a molecular switch important for the proper adaptation to nutrientavailability. Molecular Microbiology 2005;55:862–80.
Rudoni S, Colombo S, Coccetti P, Martegani E. Role of guanine nucleotides in theregulation of the Ras/cAMP pathway in Saccharomyces cerevisiae. Biochimica etBiophysica Acta 2001;1538:181–9.
Sheff MA, Thorn KS. Optimized cassettes for fluorescent protein tagging in Saccha-romyces cerevisiae. Yeast 2004;21:661–70.
Sutherland BW, Spiegelman GB, Weeks G. A Ras subfamily GTPase shows cell cycle-dependent nuclear localization. EMBO Reports 2001;2:1024–8.
Thevelein JM, de Winde JH. Novel sensing mechanisms and targets for the cAMP-protein kinase A pathway in the yeast Saccharomyces cerevisiae. MolecularMicrobiology 1999;33:904–18.
Thomas BJ, Rothstein R. Elevated recombination rates in transcriptionally activeDNA. Cell 1989;56:619–30.
Tisi R, Belotti F, Paiardi C, Brunetti F, Martegani E. The budding yeast RasGEFCdc25 reveals an unexpected nuclear localization. Biochimica et Biophysica Acta2008;1783:2363–74.
Wang G, Deschenes RJ. Plasma membrane localization of Ras requires class C Vpsproteins and functional mitochondria in Saccharomyces cerevisiae. Molecular andCellular Biology 2006;26:3243–55.
Wei M, Fabrizio P, Hu J, Ge H, Cheng C, Li L, et al. Life span extension by calorierestriction depends on Rim15 and transcription factors downstream of Ras/PKA,Tor and Sch9. PLoS Genetics 2008;4:0139–49.
Xue Y, Montserrat B, Hirsch JP. GPR1 encodes a putative G protein-coupled receptor
that associates with the Gpa2p G� subunit and functions in a Ras-independentpathway. EMBO Journal 1998;17:1996–2007.Zaman S, Lippman SI, Schneper L, Slonim N, Broach JR. Glucose regulates tran-scription in yeast through a network of signalling pathways. Molecular SystemsBiology 2009;5:245.