localization of dna protein-binding sites in the proximal and distal
TRANSCRIPT

THE JOURNAL OF BIOLOGICAL CHEMISTRY 0 1990 by The American Society for Biochemistry and Molecular Biology, Inc.
Vol. 265, No. 6, Issue of February 25, pp. 3382-3391,199O Printed in V. S. A.
Localization of DNA Protein-binding Sites in the Proximal and Distal Promoter Regions of the Mouse cu-Fetoprotein Gene*
(Received for publication, October 17,1989)
Dong-Er Zhang, Peter R. Hoyt& and John Papaconstantinou$ From the Department of Human Biological Chemistry and Genetics, University of Texas Medical Branch, Galveston, Texas 77550
DNase I footprinting assays were performed to iden- tify the binding sites for putative trans-acting factors involved in the control of a-fetoprotein (AFP) gene expression using mouse AFP promoter fragments (-839 to +56) and nuclear protein extracts from fetal, newborn, and adult livers and from brain and kidney. Our studies have shown that with nuclear protein from adult mouse liver, there are 14 protected regions in the AFP promoter up to -839 base pairs (bp). Region I (-82 to -43) was protected by at least three different factors, one of which is CCAAT-bindinglenhancer- binding protein. This region is highly conserved in the mouse, rat, and human AFP genes and has been shown previously to be essential for the regulation of tissue- specific expression in mouse. Differences in DNase I protection with fetal, newborn, and adult nuclear pro- teins have been observed in the proximal promoter region (up to -202 bp) and in regions further upstream (up to -839 bp). Significant differences among liver, kidney, and brain nuclear protein-binding sites have also been observed. In these studies, we have mapped the fetal and adult nuclear protein-binding sites of the cis-acting DNA sequences of the mouse AFP proximal promoter (up to -200) and have identified specific protein-binding sites in the distal promoter (-200 to -839). We have also identified the sites of the AFP promoter which bind nuclear proteins from highly dif- ferentiated tissues in which AFP is not expressed.
a-Fetoprotein is an oncofetal protein whose regulatory characteristics encompass activation in the early stages of fetal liver development (l), repression in the newborn (2-5), and reactivation in the adult during liver regeneration (2-4) and chemical hepatocarcinogenesis (6-8). The gene is also expressed in yolk sac, fetal gut, and fetal kidney cells (9-16) during specific stages of development, but the major site of
* This investigation was supported by United States Public Health Service Grants CA31472-04 and CAI7701-09 awarded by the National Cancer Institute, Department of Health and Human Services (to J. P. and to the University of Texas Medical Branch Cancer Center, respectively) and by the Zelda Zinn Casper Foundation Grant (awarded to J. P.). The costs of publication of this article were defrayed in part by the payment ofpage charges. This article must therefore be hereby marked “aduertisement” in accordance with 18 U.S.C. Section 1734 solely to indicate this fact.
The nucleotide sequence(s) reported in this paper has been submitted to the GenBankTM/EMBL Data Bank with accession number(s) 505246.
SPresent address: Biology Div., Oak Ridge National Laboratory, P.O. Box Y, Oak Ridge, TN 37830.
J To whom correspondence should be addressed: Dept. of Human Biological Chemistry and Genetics, The University of Texas Medical Branch, F-43, Galveston, TX 77550.
synthesis is the fetal liver, in which the gene is transcribed throughout the period of fetal liver development. These reg- ulations occur primarily at the level of transcription (5) and provide an excellent model for studying the molecular mech- anisms of both tissue-specific and developmental regulation of eukaryotic gene expression.
In recent studies, the cis-acting elements of the rodent and human AFP’ genes have been identified by transient expres- sion assay (17-22) and by insertion of various expression vectors into mice as stable transgenes (23, 24). These studies have demonstrated that liver-specific expression as well as repression can be conferred by cis-acting DNA sequences localized up to 1 kilobase upstream of the transcription initi- ation site and/or within the coding region of the gene. Similar studies with other liver-specific serum protein genes such as albumin (25-30), cut-antitrypsin (31-34), transthyretin (35), and fibrinogen (36) genes have demonstrated that sequences in their proximal promoter regions are highly conserved and that these sequences are the potential binding sites for trans- acting factors that are both liver specific, such as HNF-1 and C/EBP (30,34,36-41), and ubiquitous, such as NF-1 (42-45) and NF-Y (46, 47). For example, purified HNF-1 has been shown to interact with promoter regions from albumin, AFP, (Y- and P-fibrinogen, cY1-antitrypsin, and transthyretin (36,37, 41, 48). The promoter of each of these genes contains a common DNA motif that is recognized by HNF-1. Another liver-specific nuclear protein, C/EBP, which was initially identified as a protein that binds to the viral enhancer core element (49,50) and to certain CCAAT-containing sequences (51), recognizes several different DNA sequence motifs (48). This protein also binds to several sites in the albumin pro- moter (30, 38, 39) and to the promoter of other liver-specific genes such as transthyretin (42) and cY1-antitrypsin (35, 36, 41). These nuclear proteins have been shown to function as positive transcription factors with the albumin and al-anti- trypsin promoters in in vitro transcription assays. These studies suggest that certain trans-acting factors should bind to conserved sequences in the AFP promoter and function in its liver-specific regulation. The AFP gene is unique, however, because its regulation requires positive trans-acting factors in the fetal liver and negative trans-acting factors in the adult liver. We have conducted experiments designed to identify potential trans-acting protein factors of the mouse AFP pro-
’ The abbreviations used are: AFP, a-fetoprotein; C/EBP, CCAAT- binding/enhancer-binding protein; HNF-1, hepatocyte nuclear factor 1; DBP, albumin promoter site D-binding protein; GR, glucocorticoid receutor: GRE, ducocorticoid receptor element; NF-1, nuclear factor 1; NF-Y; nuclear factor Y; NP-Ii1 and NP-IV, nuclear proteins of adult heuatocvte that bind to regions III and IV. respectively, of the AFP promo&r; EGTA, [ethylenebis(oxyethylenenitrilo)]tetraacetic acid; HEPES, 4-(2-hydroxyethyl)-1-piperazineethanesulfonic acid; ds, double-stranded.
3382
by guest on February 12, 2018http://w
ww
.jbc.org/D
ownloaded from

DNA Protein-binding Sites of Mouse AFP Promoter
moter in the fetal, newborn, and adult mouse liver. Our experiments are also designed to determine whether cis-acting sequences, shown previously to be essential for tissue-specific AFP expression, are the binding sites for these proteins.
MATERIALS AND METHODS
Construction of Plasmids and Labeling of DNA Fragments-All four plasmids are derived from pSVori-MAF-11’ whose insert extends from XbaI (-839) to Sau3AI (+56) of the AFP promoter region inserted into the XbaI and BamHI sites of the pUC19 multiple cloning site in the pSVori plasmid. pSVori-MAF-11 was digested with MboII/ EcoRI. To prepare pAFP-1, the smaller MboII/EcoRI fragment (+56 to -202) was inserted into SmaI/EcoRI sites of pSVori after the Mb011 site was blunt ended with T4 DNA polymerase. To prepare pAFP-2 (-104 to -369), the pSVori-MAF-li was digested with EcoRI. deleted with Ba131 to -104. then dieested with NdeI, and blunt ended with T4 DNA polymerase. The-NdeI/BalSl fragment was inserted into SmaI site of pSVori with NdeI end close to the Hind111 site and Ba131 end close to the EcoRI site. To prepare pAFP- 3 (-360 to -609), the pSVori-MAF-11 was digested with EcoRI, deleted with Ba131 to -360, blunt ended with T4 DNA polymerase, then digested with PuuII and HindIII. The PuuIIIBal31 fragment was inserted into SmaI site of pSVori with the PvuII end adjacent to the Hind111 site and Ba131 end adiacent to the EcoRI site. To nrepare pAFP-4 (-610 to -839), the H&dIII/PuuII fragment was im&ted into HindIII/SmaI-digestedpSVori. These plasmids were constructed and purified using standard protocols (52,53).
To label DNA fragments used for DNase I footprinting and gel retardation studies, the pAFP series of plasmids were either 3’ end labeled at the EcoRI site with [(u-~‘P]~ATP and [~Y-~‘P]TTP (coding strands) or at the Hind111 site with [o(-s’P]dATP, [L~-~‘P]TTP, dGTP, and dCTP (noncoding strands) using the Klenow fragment of DNA polymerase I. Labeled inserts were separated from the plasmid vector by polyacrylamide gel (5%), eluted from gel slices in-O.3 M sodium acetate, oH 7.2. and purified bv passina through Elutip-d columns.
Preparation of N&ear Extracts-Nuclear extracts-were prepared from the liver, kidney, and brain of young adult (2-4 months old), newborn (l-2 days after birth), and fetal (18-19 days of gestation) C3H/He mice, essentially as described by Gorski et al. (54) with some modifications. Briefly, tissues were cut and rinsed in saline A (20 InM Tris-HCl, pH 7.0, 137 mM NaCl, and 5 mM KCl) and homogenized in buffer A (0.25 M sucrose, 15 mM Tris-HCl, pH 7.9, 60 mM KCl, 15 mM NaCl, 2 mM EDTA, 0.5 mM EGTA, 0.15 mM spermine, 0.5 mM spermidine, 1 mM dithiothreitol, 0.4 mM phenylmethylsulfonyl fluo- ride, 2 mM benzamidine, and 1 pg/ml each of antipain, chymostatin, leupeptin, and pepstatin A). The homogenate was mixed with 2 volumes of buffer B (buffer A with 2.3 M sucrose), layered on top of buffer C (buffer A with 1.8 M sucrose), and centrifuged at 25,000 rpm for 60 min at 0 “C in a SW 27 rotor. The nuclear pellets were resuspended in buffer D (100 mM KCl, 10 mM Tris-HCl, pH 8.0, 2 mM MgCl,, 0.1 mM EDTA. 1 mM dlthiothreitol. 0.4 mM nhenvlmeth- ylsulfonyl fluoride, 2 mM' benzamidine, 1 pg/ml each-of antipain, chymostatin, leupeptin, and pepstatin A). The suspension was ex- tracted with 0.10 volume of 4 M (NH&SO4 on a rotator for 30 min and then centrifuged at 30,000 rpm for 45 min in an SW 40 rotor. The protein in the supernatant was precipitated by 0.3 g/ml (NH&SO1, pelleted, resuspended in buffer E (20 mM HEPES, pH 7.8, 10% glycerol, 100 mM KCl, 0.2 mM EDTA, 0.5 mM dithiothreitol, 0.5 mM phenylmethvlsulfonvl fluoride, 2 mM benzamidine. 1 &ml each of antipain, chymosta& leupeptin and pepstatin A), and di- alyzed against buffer E for 4 h. The dialysates were microcentrifuged for 5 min to remove the precipitates. Protein concentrations were determined by the method of Bradford (55). We routinely obtain approximately 1 mg of nuclear protein/adult mouse liver. The protein extracts were frozen in liquid nitrogen and stored at -90 “C!.
DNase I Footprinting-DNase I footprint assays were adpated from the method of Jones et al. (56) and Lichtsteiner et al. (38). DNA- binding reactions were carried out in a 20- or 40-~1 volume with approximately 1 ng of end-labeled DNA fragment, 4000 ng of unla- beled ds-poly(d1 . dC), and variable amounts-of protein (as-indicated in the figure legends) in a final buffer concentration of 10 mM HEPES (pH 7.8), 30 mM KCl, 12% glycerol, 5 mM MgCl,, 0.5 mru dithiothre- itol, 0.1 mM EDTA, and 0.2 mM phenylmethylsulfonyl fluoride.
*J. P. Rabek, P. R. Hoyt, D-E. Zhang, and J. Papaconstantinou, manuscript in preparation.
Reactions were incubated for 60 min on ice followed by a 60-s digestion at room temperature with 0.4-4 ~1 of a freshly diluted DNase I solution (10 pg/ml). DNase I stock solutions were prepared by dissolving the lyophilized enzyme (Worthington) at 1 mg/ml in ice-cold water. Small aliquots of this stock solution were kept frozen at -90 “C. The DNase I dilution buffer contains 10 mM HEPES (pH 7.8), 25 mM CaCL. The DNase I digestion reactions were stopped by the addition of 100 ~11 of stop mixture (20 mM EDTA, 0.5% sodium dodecyl sulfate, 20 !iM Tris,~pH 7.5, 75 ‘pg/ml sheared.salmon sperm DNA, 100 rg/ml proteinase K). The samples were incubated at 60 ‘C for 30 min, extracted with phenol:chloroform:sample in a ratio of l:l:l, and precipitated with 2 volumes of ethanol. The DNA pellets were dried and resuspended in sequencing stop buffer (0.1% xylene, 0.1% bromphenol blue, 10 mM Na*EDTA, 95% deionized formamide), incubated at 95 “C for 3 min, and loaded on a 6% polyacrylamide, 8.3 M urea sequencing gel. The gels were dried and autoradiographed overnight at -90 “C with a Du Pont Cronex intensifying screen.
RESULTS
Localization of Sites in the Mouse AFP Proximal Promoter Which Bind Adult Liver Nuclear Proteins (the Repressed AFP Gene)-cis-Acting DNA sequences of the mouse AFP pro- moter which are essential for tissue-specific and develop- mental regulation have been mapped to within 1 kilobase of the AFP promoter region (17-20,23, 24). Godbout et al. (18) have shown by deletion analysis that the region between -85 and -52 is essential for tissue-specific AFP expression. Dele- tion of this essential region results in virtually complete extinction of the transcriptional activity in human hepatoma (Hep G2) cells (18). Studies with transgenic animals have shown that the signals that direct postnatal repression of the AFP gene are included in 1 kilobase of 5’-flanking DNA (24) and in a portion of the structural gene (61). Since the inter- action of cis-acting sequences with transacting factors is the proposed mechanism for these regulatory processes, we have conducted DNase I protection assays to map the regions of the mouse AFP promoter which bind nuclear proteins from fetal, newborn, and adult mouse liver and to determine whether the protein-binding sites correlate with the regions identified by deletion analysis to be of functional importance.
A restriction map of the mouse AFP promoter, up to -839, the TATA box, and transcription initiation site are shown in Fig. 1. The location of probes (AFP-I, 2, 3, and 4) used for DNase I protection assays is also shown. To identify nuclear protein-binding sites of the AFP proximal and distal promoter regions and to establish a standard map as a basis for com- parison between fetal liver and nonliver cell proteins, we performed DNase I protection analyses using adult liver nu- clear proteins (Fig. 2). The probes were 3’ end labeled either on the coding or noncoding strand. The AFP promoter se-
H E AFP-4
FIG. 1. A restriction map of the 5’-flanking region of the AFP gene. Indicated above the partial restriction enzyme map are the TATA box and the site of transcription initiation. The probes used in DNase I footprint analyses are shown below the map. E and H represent, respectively, the EcoRI and Hind111 sites at the ends of the probes.
by guest on February 12, 2018http://w
ww
.jbc.org/D
ownloaded from

3384 DNA Protein-binding Sites of Mouse AFP Promoter
1 la -mo-= - - zo- lb
‘- IF -z,., me m - 1 ‘b -mo- “, - rb- ,a ?- -50. - -- LL 1 II
FIG. 2. DNase protection of the mouse AFP promoter by adult liver nuclear proteins. AFP DNA fragments were 3’ end labeled either at the EcoRI sites (panels A, C, E, and G), which rep- resent the coding strand, or at the Hind111 sites (panels B, D, F, and H), which represent the noncoding strand, and incubated with adult mouse liver nuclear protein in the presence of 4000 ng of ds-poly(dI.dC). Lanes G and G + A are the probe DNAs subjected to guan- osine and guanosine + adenine-specific Maxam-Gilbert cleavage, respectively. The numbers at the top of each lane depict the pg of nuclear protein in the incubation mixture. Numbers on the left sides of each panel indicate the nucleo- tides upstream (-) or downstream (+) of the site of transcription initiation (+l). Sequences protected from DNase I diges- tion are indicated by brackets, and the Roman numerals refer to the entire pro- tected region. Sites of enhanced cleavage (hypersensitive sites) are indicated with arrows
- -. i -- ---
-IX- - -
II I e--
.-- w
- -
.i-
. -
e-e -
- 202
AFP-1 +56 -369
AFP-2 -104
E F
0 J PPZ --SW. _-__ . ---a-
--z 4 1 XIV -*oo- = -7 - --.-- ----a _ ._ ___--- - --- - _..-_ -_. __-- em- - --_- -830 - --- - ---- --- _ _.._m- ;;I _^.. -= -E
a
-630- ---. ZL- em--
-3p- a--- E -- --a- _. --a-
_ ---- _-..
:w ._e..-
-839 -610
AFP-4 -609
AFP-3
- 360
quence is shown in Fig. 3, and the protected regions are protein concentrations, nucleotides -63 to -82 within region indicated by solid lines either above the sequence (coding Ib were protected. The sequence of this region, TAT- strand data) or under the sequence (noncoding strand data). GTTTGCTCA, is similar to sites D (TATGATT) and F
DNase I protection analysis indicates that region I (-82 to (TATGTTT) in the mouse albumin promoter (30, 38). These -43) binds more than one protein (Fig. 2, A and B). At low sequences are reported to interact with C/EBP, which binds
by guest on February 12, 2018http://w
ww
.jbc.org/D
ownloaded from

DNA Protein-binding Sites of Mouse AFP Promoter 3385
-859 TTAGCAAACA TGTCTGGACC TCTAGACACA CAGACTCTGA CACGCCAACG AATCGTTTGT ACAGACCTGG AGATCTGTGT GTCTGAGACT GTGCGGTTGC
XIV XIII -809 TCTGAGTTCT AGTTTCGATA CGCACTGGGA AGTTTTAAAA GTTTTCCATC
AGACTCAAGA TCAAAGCTAT GCGTGACCCT TCAAAATTTT CAAAAGGTAG
XII -759 AACTCTMTG TGTAGAGAM TGGAAA CTAT CATAGACTCT ACGACATTGA
TTGAGATTAC ACATCTCTTT ACCPTTGATA GTATCTGAGA TGCTGTAACT
-709 GGGTGAAGGT ATGAGTGMG CAmCTTAGG GTCAGAAGTA TGTCAGTGCC CCCACTTCCA TACTCACTTC GTGAGAATCC CAGTCTTCAT ACAGTCACGG
-559 CATTTGTTGC TGTTAGCATC ATCATCTTAG GGCTTGAGAG GATGTTGCAG GTAAACAACG ACAATCGTAG TAGTAGAATC CCGAACTCTC CTACAACGTC
XI -609 CTGACCCATG CACCTGTGAC ATAICATATGG MTTATTCTT TGGCACATAA
GACTGGGTAC GTGGACACTG TATGTATACC TTAATAAGAA ACCGTGTATT
X -559 AATTAGAATG GGAGCTGGCT CATCAGGTTT TGTGCTGTM GTTTTSTATG
TTAATCTTAC CCTCGACCGA GTAGTCCAAA ACACGACATT CAAAAGATAC
-509 TTAAACCAGA TGCGATACAC TAAATAAAAT MAATATACT TGACCGATGG MTTTGGTCT ACGCTATGTG ATTTATTTTA TTTTATATGA ACTGGCTACC
-150-
A <
U&F01 234
IX mm
-459 TTTTGAGCGA AATAATAACT GGATAATCAA GAAATATATC CCACTAATGA -2oo- -- AAAACTCGCT TTATTATTGA CCTATTAGTT CTTTATATAG GGTGATTACT &‘;..Erf
- -----.._
-409 ATAGCCTGAA CTACTGAAAC MTTTGTTCA GTGCCTAGCA TATGGTGTGC TATCGGACTT GATGACTTTG TTAAACAAGT CACGGATCGT ATACCACACG
VIII -359 ATTTTATTAT TTCTTTCMA AAGAATGTAT TTGGAGTTAC ATAGTMGTC
T-TAATA AAGAAAGTTT TTCTTACATA AACCTCAATG TATCATTCAG
VII -309 TGCTACCTTT TCTTTATGGC TATATCTATG TCTTATGTTG AGATGAATGA
ACGATGGAAA AGAAATACCG ATATAGATAC AGAATACAAC TCTACTTACT
VI -259 ATTATTCTTC AGGGGAAATA ATCTATTTGA ACAGTTTAGA TGGTGAAGAA
TAATAAGMG TCCCCTTTAT TAGATAAACT TGTCAAATCT ACCACTTCTT
V IV -209 CATTTGCAGC ATTTGCAAGA TTTTTTTCCA CTCTGAAGTG GTCTTTGTCC
GTAAACGTCG TAAACGTTCT AMAAAAGGT GAGACTTCAC CAGMACAGG
Ill -159 TTGAACATAG GATACAAGTG ACCCCTGCTC TGTTAATTAT TGGCAAATTG
AACTTGTATC CTATGTTCAC TGGGGACGAG ACAATTAATA ACCGTTTAAC
II I -109 CCTAACTTCA ACGTAAGGAA ATAGAGTCAT ATGTTTGCTC ACTGAAGGTT
GGATTGAAGT TGCATTCCTT TATCTCAGTA TACAAACGAG TGACTTCCM
-59 ACTAGTTAAC AGGCATCCCT TAAACAGGAT AT-GGAC TTCAGCAGGA TGATCAATTG TCCGTAGGGA ATTTGTCCTA TATTTTCCTG MGTCGTCCT
+1 -09 CTGCTCGAAA CATCCCACTT CCAGCACTGC CTGCGGTGAA GGAACCAGCA
GACGAGCTTT GTAGGGTGM GGTCGTGACG GACGCCACTT CCTTGGTCGA
FIG. 3. DNA sequences of the mouse AFP promoter region and localization of the adult liver nuclear protein-binding site. The regions that are protected by nuclear proteins are indicated by lines above and below the DNA sequence for the coding and noncoding strands, respectively.
to CCAAT-containing sequences (51) and the core enhancer sequence (49). To verify whether region Ib is protected by C/ EBP, an oligonucleotide containing the consensus C/EBP- binding site, and purified C/EBP (both gifts of Dr. Steven L. McKnight, Dept. of Embryology, Carnegie Institute of Wash- ington, Baltimore, MD) were used in the protection assay. The results are shown in Fig. 4. In panel A, when increasing amounts of C/EBP oligonucleotide competitor are used (lanes l-4), protection of region Ib is significantly but not completely abolished. In panel B, it is shown clearly that region Ib is fully protected by purified C/EBP (lanes 1-3). We interpret these experiments to indicate that although region Ib is a binding site for C/EBP, there are also other factor(s) present in adult mouse liver nuclei which bind to this region.
When the protein concentration is increased to 60 pg, region
-= --
+1-
-5O-
-lOO-
-150-
FIG. 4. Localization of C/EBP-binding sites in the mouse AFP proximal promoter. The DNA probe used in both panels is anticoding strand-labeled AFP-1. Panel A, the probe is subjected to DNase I digestion in the absence (lane F) and presence (lanes O-4) of 60 pg of adult mouse liver nuclear protein. The binding reactions were performed in the presence of a 0, 25 X, 50 X, 100 X, 200 X M concentration of the oligonucleotide containing the C/EBP consensus sequences in lanes O-4, respectively. All reactions included 4000 ng of the nonspecific competitor ds-poly(dI.dC). Panel B, lane F is the DNase I-digested DNA probe in the absence of any nuclear protein. Lanes Z-3 are the DNase I-digested probe in the presence of 0.5, 1, and 2 ~1 of protein solution, which contains the full-length C/EBP expressed in Escherichia coli. Each reaction also contained 4000 ng of ds-poly(dI.dC). Lane C is the control of the DNase I protection with the 50 pg of adult mouse liver nuclear extract. All the other labels are the same as in Fig. 2.
Ia, from -43 to -62, is protected (Fig. 2, A and B). This regioncontainsthesequence(-62)GTTACTAGTTAAC(-50), which is homologous to the hepatocyte nuclear factor 1 (HNF- 1)-binding consensus sequence GTTAATNATTAAC (36).
Region II (-126 to -98) contains a sequence GTTAAT- TATTGGC(-128 to -116), which is homologous to the HNF- 1 consensus site and is linked at its 3’ end with a sequence (-llS)TGGCAAATTGCCT(-107), which is homologous to the NF-1 consensus site (TGGA/,N5 GCCA). These sequence data suggest that region II could bind either HNF-1 or NF-1. Our experiments also show that C/EBP is another factor that binds to region II (Fig. 4). The protection of the 3’ end of region II with adult liver nuclear extract was partially com- peted out by the C/EBP consensus sequence (Fig. 4A, lanes 2-4). The fact that purified C/EBP also exhibits protection of region II (Fig. 4B) is very strong evidence that C/EBP is one of several factors present in adult liver nuclei which bind to this region.
Sequence analysis of region III (-146 to -131) and region IV (-169 to -152) indicates that they contain sequences (C)CTG(C)TCT (-136 to -129) at the 3’ end of region III andT(T)TGT(C)CT (-166 to -159) in region IV, which show homology to the GRE consensus sequence (A/&TGA/TTCT). The protected sequence of region III has not been identified previously as a binding site for either AFP or other serum protein gene promoters. The sequence of region IV has been
by guest on February 12, 2018http://w
ww
.jbc.org/D
ownloaded from

3386 DNA Protein-binding Sites of Mouse AFP Promoter
4 CR
v +
-2oo- IV -150- 3 IV
7 III
-lOO- -i, r-I II 1 c -. lb - .* II 1
,.-9--m - -5o- ;L
1 la
A -~ 1. -. ” *; . -_ - &i. -p
Zf - -- -f
ry;rws;gll +,- = -...
0% -- --
-- - _ g: -_ --
: - +30- :- .- --
.- -. I *
A
-250- =a
-350- -t S
- -
-I- - -
w*- - - -
- -
m LJI:
B
-
-600
-550 LX’
1 X
-500
6s:. 3 --- L1 -450- ma I’ --
z b-
-iJ- -- -
L -_ *--- E%!
‘c -zca- -
-‘F =
-4oo- ,-
-- a- -- 5 - ;T:
C
1 IX
FIG. 5. DNase I protection of the mouse AFP promoter by nuclear proteins from fetal, newborn, and adult mouse liver. DNA probes used in panels A and C are coding strand-labeled AFP-1 and AFP-3, respectively. The DNA probe used in panel B is the anticoding strand-labeled AFP-2. DNA fragments were incubated with increasing amounts of nuclear proteins from fetal, newborn, and adult mouse liver, as indicated at the top of each lane, in the presence of 4000 ng of ds-poly(d1. dC). All the other labels are the same as in Fig. 2.
reported by Guertin et al. (22) as a binding site for the glucocorticoid receptor (GR) protein in the rat AFP promoter. To verify the identity of the proteins that bind to regions III and IV, an oligonucleotide containing the GRE consensus sequence, and purified glucocorticoid receptor protein (gifts from Dr. Keith Yamomoto) were used. The results showed that the purified GR exhibits weak binding to region IV and does not bind at all to region III (data not shown). Further- more, the GRE oligonucleotide consensus sequence does not compete for either of these sites. We conclude that the pro- teins binding to regions III and IV, which we call nuclear protein III (NP-III) and nuclear protein IV (NP-IV), respec- tively, are not GR.
Region V (-177 to -202) consists of a sequence GTGGAAA, -179 to -185, which is homologous to the virus enhancer core sequence GTGGT/AT/~T/,+ A similar sequence in SV40, Moloney sarcoma virus, and polyoma virus en- hancers binds C/EBP (49,59,60). To test whether protection of region V is due to the binding of C/EBP, the oligonucleotide containing the consensus C/EBP-binding site and purified C/ EBP protein were used to perform DNase I footprinting experiments. As shown in Fig. 4A, the C/EBP-binding oligo-
mer prevents binding of the factor from adult mouse liver nuclei to the upstream part of region V. In addition, purified C/EBP also shows protection of the same upstream sequence of region V (Fig. 4B). These results suggested that C/EBP is involved in the protection of region V, but only from -183 to -202 (Fig. 3). This observation was unexpected because the sequence homologous to the virus enhancer core sequence was not fully protected by C/EBP. These data also indicate that the sequence adjacent to the enhancer core sequence is the site for C/EBP binding in region V. Since the region protected by nuclear extract is significantly larger, our results indicate that the downstream part of region V may be protected by another as yet unidentified factor.
Localization of Sites in the Mouse a-Fetoprotein Distal Pro- moter Which Bind Adult Liver Nuclear Proteins-We classify the sequences upstream of region V (from -200 to -839) as the distal promoter region. The protected sequences in this domain, with the possible exception of those in region XI, have not been identified as protein-binding sites in any liver- specific genes prior to this report. Camper and Tilghman (24), using transgenic mice, have reported that this distal promoter region may function in the postnatal repression of AFP gene
by guest on February 12, 2018http://w
ww
.jbc.org/D
ownloaded from
DNA Protein-binding Sites of Mouse AFP Promoter
3 lb -250 -
I- _ .- -- - - _ .;-- 1
- -- - . .
-120- ‘_ M ..-
I I
II . -_
.I ) . .
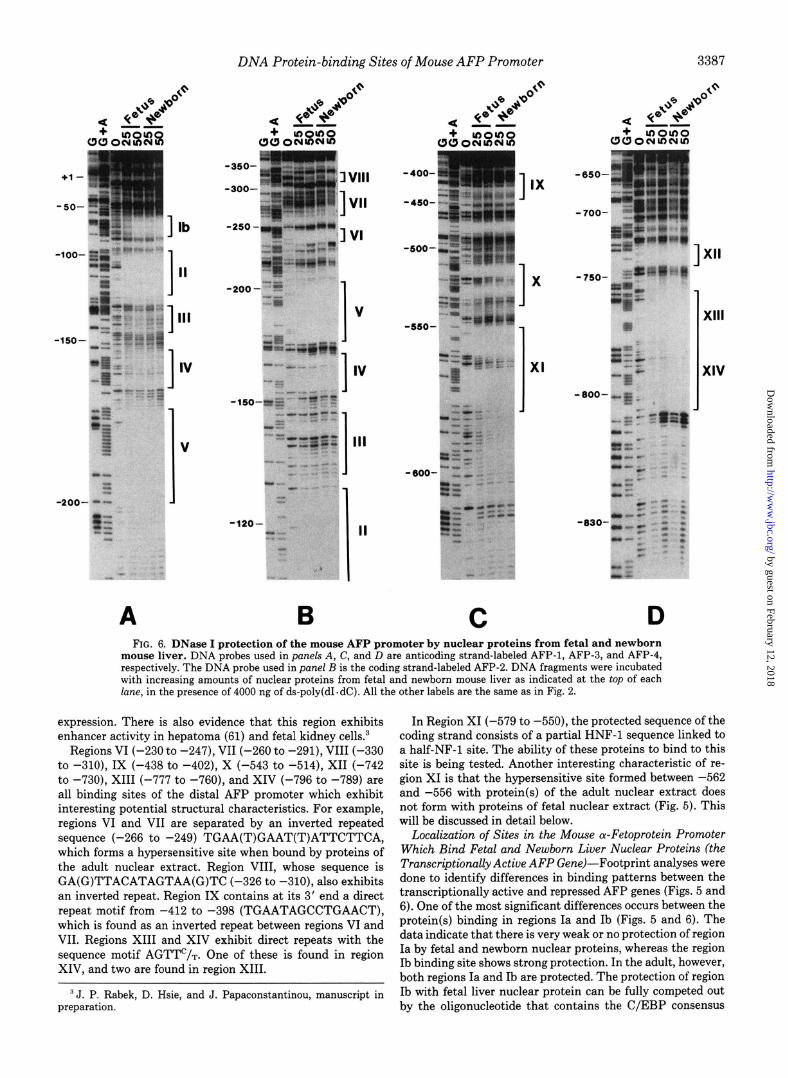
A B C D FIG. 6. DNase I protection of the mouse AFP promoter by nuclear proteins from fetal and newborn
mouse liver. DNA probes used in pat& A, C, and D are anticoding strand-labeled AFP-1, AFP-3, and AFP-4, respectively. The DNA probe used in panel B is the coding strand-labeled AFP-2. DNA fragments were incubated with increasing amounts of nuclear proteins from fetal and newborn mouse liver as indicated at the top of each lane, in the presence of 4000 ng of ds-poly(dI.dC). All the other labels are the same as in Fig. 2.
expression. There is also evidence that this region exhibits enhancer activity in hepatoma (61) and fetal kidney cells.3
Regions VI (-230 to -247), VII (-260 to -291), VIII (-330 to -310), IX (-438 to -402), X (-543 to -514), XII (-742 to -730), XIII (-777 to -760), and XIV (-796 to -789) are all binding sites of the distal AFP promoter which exhibit interesting potential structural characteristics. For example, regions VI and VII are separated by an inverted repeated sequence (-266 to -249) TGAA(T)GAAT(T)ATTCTTCA, which forms a hypersensitive site when bound by proteins of the adult nuclear extract. Region VIII, whose sequence is GA(G)TTACATAGTAA(G)TC (-326 to -310), also exhibits an inverted repeat. Region IX contains at its 3’ end a direct repeat motif from -412 to -398 (TGAATAGCCTGAACT), which is found as an inverted repeat between regions VI and VII. Regions XIII and XIV exhibit direct repeats with the sequence motif AGTTC/T. One of these is found in region XIV, and two are found in region XIII.
‘J. P. Rabek, D. Hsie, and J. Papaconstantinou, manuscript in preparation.
In Region XI (-579 to -550), the protected sequence of the coding strand consists of a partial HNF-1 sequence linked to a half-NF-1 site. The ability of these proteins to bind to this site is being tested. Another interesting characteristic of re- gion XI is that the hypersensitive site formed between -562 and -556 with protein(s) of the adult nuclear extract does not form with proteins of fetal nuclear extract (Fig. 5). This will be discussed in detail below.
Localization of Sites in the Mouse a-Fetoprotein Promoter Which Bind Fetal and Newborn Liver Nuclear Proteins (the Transcriptionally Active AFP Gene)-Footprint analyses were done to identify differences in binding patterns between the transcriptionally active and repressed AFP genes (Figs. 5 and 6). One of the most significant differences occurs between the protein(s) binding in regions Ia and Ib (Figs. 5 and 6). The data indicate that there is very weak or no protection of region Ia by fetal and newborn nuclear proteins, whereas the region Ib binding site shows strong protection. In the adult, however, both regions Ia and Ib are protected. The protection of region Ib with fetal liver nuclear protein can be fully competed out by the oligonucleotide that contains the C/EBP consensus
by guest on February 12, 2018http://w
ww
.jbc.org/D
ownloaded from
3388 DNA Protein-binding Sites of Mouse AFP Promoter
lla J]lb
1 V
-2oo- J -. -_ == - --_ w-e
--- --
A
-250- w 4Bwm.-..
5 1 VI --- - iilm+- -s.-._ _
B
-6OO-
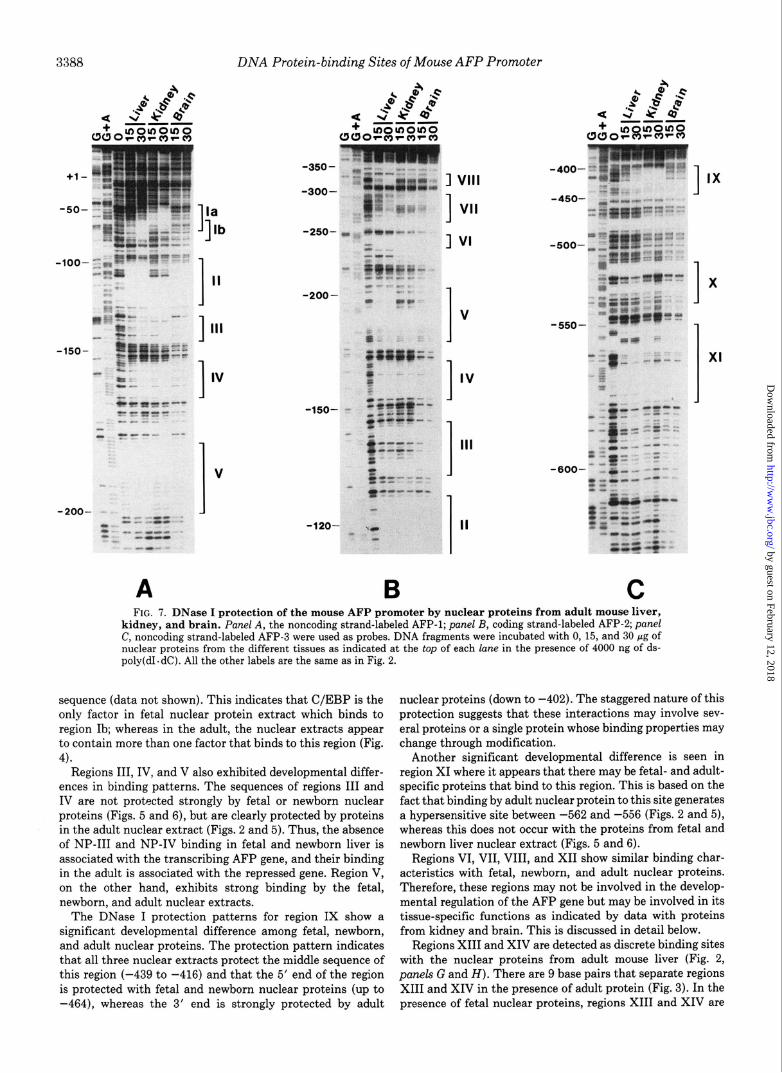
C FIG. 7. DNase I protection of the mouse AFP promoter by nuclear proteins from adult mouse liver,
kidney, and brain. Panel A, the noncoding strand-labeled AFP-1; panel B, coding strand-labeled AFP-2; panel C, noncoding strand-labeled AFP-3 were used as probes. DNA fragments were incubated with 0, 15, and 30 pg of nuclear proteins from the different tissues as indicated at the top of each lane in the presence of 4000 ng of ds- poly(dI. dC). All the other labels are the same as in Fig. 2.
sequence (data not shown). This indicates that C/EBP is the only factor in fetal nuclear protein extract which binds to region Ib; whereas in the adult, the nuclear extracts appear to contain more than one factor that binds to this region (Fig. 4).
Regions III, IV, and V also exhibited developmental differ- ences in binding patterns. The sequences of regions III and IV are not protected strongly by fetal or newborn nuclear proteins (Figs. 5 and 6), but are clearly protected by proteins in the adult nuclear extract (Figs. 2 and 5). Thus, the absence of NP-III and NP-IV binding in fetal and newborn liver is associated with the transcribing AFP gene, and their binding in the adult is associated with the repressed gene. Region V, on the other hand, exhibits strong binding by the fetal, newborn, and adult nuclear extracts.
The DNase I protection patterns for region IX show a significant developmental difference among fetal, newborn, and adult nuclear proteins. The protection pattern indicates that all three nuclear extracts protect the middle sequence of this region (-439 to -416) and that the 5’ end of the region is protected with fetal and newborn nuclear proteins (up to -464), whereas the 3’ end is strongly protected by adult
nuclear proteins (down to -402). The staggered nature of this protection suggests that these interactions may involve sev- eral proteins or a single protein whose binding properties may change through modification.
Another significant developmental difference is seen in region XI where it appears that there may be fetal- and adult- specific proteins that bind to this region. This is based on the fact that binding by adult nuclear protein to this site generates a hypersensitive site between -562 and -556 (Figs. 2 and 5), whereas this does not occur with the proteins from fetal and newborn liver nuclear extract (Figs. 5 and 6).
Regions VI, VII, VIII, and XII show similar binding char- acteristics with fetal, newborn, and adult nuclear proteins. Therefore, these regions may not be involved in the develop- mental regulation of the AFP gene but may be involved in its tissue-specific functions as indicated by data with proteins from kidney and brain. This is discussed in detail below.
Regions XIII and XIV are detected as discrete binding sites with the nuclear proteins from adult mouse liver (Fig. 2, panels G and H). There are 9 base pairs that separate regions XIII and XIV in the presence of adult protein (Fig. 3). In the presence of fetal nuclear proteins, regions XIII and XIV are
by guest on February 12, 2018http://w
ww
.jbc.org/D
ownloaded from

DNA Protein-binding Sites of Mouse AFP Promoter 3389
fully protected (Fig. 6D) and are detected as a single protected region.
Liver Specificity of Nuclear Proteins-Nuclear extracts from liver, kidney, and brain of adult C3H/He mice were used in DNase I footprinting assays to detect tissue-specific nuclear protein binding and resulting protection patterns (Fig. 7). Because of difficulties in preparing large quantities of nuclear proteins from kidney and brain, the amounts of protein used for these experiments were only increased up to 30 pg. Liver- specific binding sites were identified as regions Ib, V, VIII, and X. As mentioned above, the liver nuclear protein that binds to regions Ib and V is C/EBP (49-51). Region VIII was also only protected by nuclear protein from the liver, indicat- ing that this may be a unique AFP-binding site. Region X is a binding site that shows liver specificity. Interestingly, this site shows its strongest binding properties with the adult nuclear proteins, indicating that it may represent an adult liver-specific nuclear protein-binding site (Figs. 5 and 6).
Sites that bind nuclear proteins from two or all three of the tissues tested but also exhibit differences in binding properties are regions Ia, II, VI, VII, IX, and XI. Kidney nuclear protein, for example, only protected region Ia (-73 to -43). There is no protection in region I when nuclear protein from brain was used. The region II binding site is protected by kidney nuclear extract from -128 to -110 and liver nuclear protein from -126 to -98, clearly demonstrating differences between the nuclear proteins of these two differentiated cell types. Only half of the NF-1 site is protected by the kidney nuclear extract, and the protection appears to extend into region III. Protec- tion by brain nuclear proteins, on the other hand, is similar to that of the liver nuclear proteins.
Regions VI and VII were protected by nuclear extracts from all three cell types, but there were differences in the pattern
-200 -150 -100 -50 tl
I I I I I
ADULT AFP
V IV III II IB IA FETAL
AFP
F E DC BA ADULT ALB
FIG. 8. A diagrammatic presentation of the adult and fetal protein-binding sites of the mouse AFP promoter and the adult mouse albumin promoter. The protected regions are drawn to scale and are depicted by Roman numerals. The proteins that bind to the promoter are defined in Footnote 1. HS localizes the hypersensitive sites that are formed in the AFP promoter in the presence of nuclear proteins. The localization of binding sites in the albumin promoter are based on data published in Refs. 3 and 38 and on unpublished data (by D-E. Zang, P. R. Hoyt, and J. Papaconstantinou).
of protection. In this case, the patterns of protection for the kidney and brain proteins were similar, whereas that for the liver was unique.
In region IX, basically the same protection pattern occurs with liver and kidney nuclear protein, but the data also indicate that the kidney may have higher levels of this factor.
Although region XI was protected by nuclear proteins from all three tissues, a DNase I-hypersensitive site was formed by liver nuclear protein and to a lesser extent by kidney nuclear extract. However, no hypersensitive site could be detected in the presence of brain nuclear proteins. These data indicate that there are differences in the liver, kidney, and brain nuclear proteins that bind to the sequences in region XI.
DISCUSSION
We have identified the regions in the mouse AFP promoter (from -1 to -839) which bind nuclear proteins from fetal, newborn, and adult mouse (C3H/He) liver. Significant simi- larities and differences between the protection maps for the transcriptionally active (fetal) and repressed (adult) gene have been demonstrated. We also detected differences among liver, kidney, and brain.
Our studies indicate that the DNA sequences of regions Ia and Ib are the binding sites for two liver-specific trans-acting factors, which we believe are HNF-1 (37) and C/EBP (49-51) (Fig. 8). These proteins are liver-specific transcription factors for the mouse albumin (38, 39, 56), cul-antitrypsin (34-37), transthyretin (36, 37, 40), and fibrinogen genes (36, 37). The localization of region Ib as the binding site for C/EBP is based on competition studies (Fig. 4) with the C/EBP oligo- nucleotide and with pure C/EBP protein. Localization of region Ia as the binding site for HNF-1 is based on experi- ments by Jose-Estanyol and Danan (57), who showed that this region of this rat AFP promoter, which is homologous to the region Ia sequence of the mouse AFP promoter, is pro- tected by protein from a rat liver nuclear extract, and this protection can be competed out by an oligonucleotide con- taining the HNF-1 consensus sequence (58). Furthermore, using purified rat HNF-1 for DNase I protection analysis, Courtois et al. (37) have shown binding to region Ia of the mouse AFP promoter. These studies support our conclusion that HNF-1 is the protein in adult mouse liver nuclear extract which results in the protection of region Ia (Fig. 2). Studies from our laboratory and others indicate that region II is a binding site for NF-1, HNF-1, and C/EBP. Sequence data suggest that NF-1 and HNF-1 could bind to this region. Jose- Estanyol and Danan (57) have shown that NF-1 oligonucle- otide sequences can compete for a protein that binds to a region in the rat AFP promoter which is homologous to region II of the mouse. Our DNase I protection analyses with nuclear proteins fractionated by heparin-agarose chromatography (data not shown) have revealed that HNF-1 binds to this region at -133 to -111, but only in the absence of other region II binding factors. Since region II protection with nuclear extracts extends from -126 to -98 and since this region does not fully include the HNF-1 sequence, we believe that the protection exhibited by nuclear extract is due to the binding of NF-1. Our experiments also show that C/EBP is a third factor that binds to region II. This diversity of binding by C/EBP, which is a proven liver-specific transcription factor, poses interesting questions concerning its role in the regulation of AFP. One of the most important of these is whether the binding properties of C/EBP are altered in the presence of other factors. Since it has been proposed that the alignment of these proteins may be essential for gene regula- tion (39), any alteration in binding due to competition by
by guest on February 12, 2018http://w
ww
.jbc.org/D
ownloaded from

DNA Protein-binding Sites of Mouse AFP Promoter
other nuclear proteins may be an important factor in this regulation.
The competition studies with C/EBP oligonucleotide and pure C/EBP protein indicate that C/EBP is the only protein in fetal liver extract which binds to region Ib, whereas similar experiments with adult mouse liver nuclear protein indicate the presence of other factors besides C/EBP which bind to region Ib (data not shown). Maire et al. (39) have proposed that site D of the mouse albumin promoter binds an adult- specific protein, DBP, as well as C/EBP. Since region Ib exhibits strong homology to albumin site D and since Ib is located in the AFP promoter at relatively the same position as site D in the albumin promoter (see Fig. 8), it is possible that DBP is one of the proteins in adult nuclear extract which binds to region Ib. On the other hand, these same experiments show that C/EBP binds very strongly to region V although the protected region does not coincide with the enhancer core consensus sequence. The enhancer core consensus sequence in region V was protected by liver nuclear protein extracts. These studies also indicate that region V may bind another protein.
Interestingly, binding of HNF-1 and C/EBP shows overlap- ping protection of regions Ia and Ib when fractionated pro- teins are used in DNase I protection assays (data not shown). Our studies have shown that binding in both regions occurs with adult nuclear extract, whereas with fetal nuclear extracts we only detect strong binding at the C/EBP site. These data indicate that HNF-1 is at a low concentration or absent in the fetal liver and that it may be an adult-specific trans-acting factor. Alternatively, HNF-1 may be present in the fetal liver in a form that cannot bind to its site in the presence of C/ EBP, thus making its detection difficult by protection assay. In this mechanism, we postulate that C/EBP binding may be replaced by other region Ib binding factors such as DBP, which is also an adult-specific protein, and that this in turn facilitates the binding of HNF-1. Functional analysis by God- bout et al. (18) has shown that sequences in region I are essential for the tissue-specific expression of AFP gene. On the basis of these studies plus our footprinting experiments, we propose that the liver-specific factor C/EBP plays a major role in the transcription of the AFP gene in the fetus. This is supported by our observation that there is no protection at the HNF-1 site in the AFP promoter when fetal or newborn mouse liver nuclear extracts are used in the protection assays. In vitro transcription analyses have demonstrated that HNF- 1, C/EBP, and DBP are positive transcription factors for the mouse albumin gene (39,54) and that HNF-1 is the dominant positive transcription factor in directing liver-specific albu- min transcription in the adult (39). Since these factors also bind to the promoter of the repressed AFP gene in the adult, the underlying basis for these differences may be in the presence of unique binding sites of the AFP promoter and/or the alignment of trans-acting factors relative to the transcrip- tion start site. A major difference between fetal and adult AFP promoter binding patterns occurs in regions III and IV. The proteins (NP-III and NP-IV) that bind to both of these regions are either not present in the fetal nuclear extracts or are in a form that cannot bind. Furthermore, our analyses detect a gradual increase in protection in the newborn, and the highest level is detected in the adult, suggesting that they are specific factors of the postnatal liver. Guertin et al. (22) have reported that this site in the rat AFP promoter binds the glucocorticoid receptor complex. However, our data indi- cate that NP-III and NP-IV are not GR and that the proteins bound to regions III and IV may be involved in the repression of the AFP gene. Although each of these regions contains
consensus sequences for glucocorticoid receptor-binding sites (GRE) and although glucocorticoid enhances the repression of the AFP gene in the newborn (64-66), our experiments indicate that the glucocorticoid receptor protein complex is not the protein that binds to these sites. Furthermore, our studies indicate that NP-III is not related to CR and that NP-IV may belong to a family of GR-related proteins because purified GR exhibits weak binding to region IV.
The protection map of the promoter of the repressed AFP gene indicates that HNF-l-, C/EBP (or DBP)-, NF-l-, re- gions III (NP-III)-, IV (NP-IV)-, and V (C/EBP)-binding sites are occupied (Fig. 8). One major difference between the albumin (active) and AFP (repressed) proximal promoters is the absence of NP-III- and NP-IV-binding sites in the albu- min promoter. This is further indication that regions III and IV may be the binding sites for proteins that function in the repression of the AFP gene in the adult. However, what the relationship of these proteins is to products of the raf locus is unknown.
Several regions in the distal promoter (from -200 to -839) exhibit significant differences in binding patterns by adult compared with fetal nuclear extracts. For example, both nu- clear extracts protect the middle sequence of region IX, whereas protection is extended upstream with fetal nuclear extract and downstream with adult. Regions XIII and XIV also show a change in their protection patterns associated with development. In the adult, these regions are clearly separated by an unprotected region of approximately 9 nucle- otides. In the fetus, however, the regions are not separated, and protection extends through XIII and XIV. Recently, the distal promoter region has been shown to have enhancer activity by chloramphenicol acetyltransferase assay in both hepatoma (61) and kidney cells.3 In addition, this region of the promoter has been reported by Camper and Tilghman (24) to function in the repression of the AFP gene in the adult hepatocyte. We believe that transacting factors binding to these regions may be involved in these regulatory activities.
Analysis of protection patterns of kidney and brain nuclear extracts provides information on tissue-specific nuclear fac- tors. The C/EBP-binding sites (regions Ib and V) are not protected by kidney or brain nuclear proteins, which is further evidence that C/EBP is a liver-specific protein. On the other hand, the kidney protein does produce protection at the HNF- 1 site. This kidney protein may be a variable HNF-1 such as that detected in dedifferentiated hepatoma cells and in lung cells (62). Also, protection of the NF-l-binding site varies between liver and kidney proteins, indicating that the kidney nuclear extract contains a different NF-1 member of the proposed NF-1 family (63). Both kidney and brain nuclear extracts contain proteins that bind to regions III and IV. Thus, in all three tissues (adult liver, kidney, and brain) in which the AFP gene is repressed, regions III and IV are strongly protected.
It has been proposed that in early liver development, the AFP and albumin genes are activated by the AFP enhancers located within the intergenic region (24) and that subsequent regulation occurs via interactions of regulatory proteins with the promoter sequences for each gene. Thus, the repression of the AFP gene at birth and continued expression of albumin are attributed to the regulatory sequences of each promoter. On the basis of our studies, we propose the following model for the independent regulation of the AFP gene. (a) The high level of expression of the AFP gene in the fetal liver is due to the combinatorial activity of C/EBP, NF-1, C/EBP, in the proximal promoter. (b) In the adult, the AFP gene is repressed due to the binding of NP-III and NP-IV between NF-1 and
by guest on February 12, 2018http://w
ww
.jbc.org/D
ownloaded from

DNA Protein-binding Sites of Mouse AFP Promoter 3391
region V (C/EBP). We propose that these factors may func- tion to prevent the formation of an active transcription com- plex. However, this does not exclude the possible involvement of the distal promoter transacting factors in this regulatory process. This would explain the positive transcription activity of HNF-1, C/EBP, and DBP with the albumin gene (39) and their association with repressed activity in the adult AFP gene.
Our studies demonstrate that the distal promoter region shows differences in protein-binding patterns between fetus and adult. Since there are indications that this region may also be associated with the postnatal repression of the gene (24), their role in developmental regulation and/or tissue- specific regulation must await further study.
27.
28.
29.
30.
31.
32.
33.
34. 35.
Babiss, L. F., Friedman, J. M., and Darnell, J. E., Jr. (1986) Mol. Cell. Biol. 6. 3798-3806
Heard, J.-M., Herbomel, P., Ott, M.-O., Mottura-Rallier, A., Weiss, M., and Yaniv, M. (1987) Mol. Cell. Biol. 7, 2425-2434
Pinkert, C. A., Ornitz, D. M., Brinster, R. L., and Palmiter, R. D. (1987) Genes & Deu. 1,268-276
Izban, M. G., and Papaconstantinou, J. (1989) J. Biol. Chem. 264,9171-9179
Kelsey, G. D., Povey, A., Bygrave, A. E., and Loveli-Badge, R. H. (1987) Genes & Deu. 1, 161-171
De Simone, V., Ciliberto, G., Hardon, E., Paonessa, G., Palla, F., Lundberg, L., and Cortese, R. (1987) EMBO J. 6, 2759-2766
Shen, R-R., Li, Y., Sifers, R. N., Wang, H., Hardick, C., Tsai, S. Y., and Woo, S. L. C. (1987) Nucleic Acids Res. 16,8399-8415
Ciliberto, G., Dente, L., and Cortese, R. (1985) Cell 41,531-540 Costa, R. H., Lai, E., and Darnell, J. E., Jr. (1986) Mol. Cell. Biol.
6,4697-4708
Acknowledgments-We are grateful to Drs. J. P. Rabek and Lee- 36. Courtois, G., Baumhueter, S., and Crabtree, G. R. (1988) Proc.
Ho Wang for their helpful discussion during the initial stages of this Natl. Acad. Sci. U. S. A. 35,7937-7941
work and to M. C. Higgins for her assistance in the preparation of 37. Courtois, G., Morgan, J. G., Campbell, L. A., and Crabtree, G. R.
the manuscript. (1987) Science 238,688-692
38. Lichsteiner. S.. Wuarin. J.. and Schibler. U. (1987) Cell 51,963-
1. 2.
3. 4.
5.
6.
7.
8.
9.
10.
11. 12.
13.
14.
15.
16.
17.
18.
REFERENCES
Abelev, G. I. (1974) Transplant. Reu. 20,3-27 Olsson, M., Lindahl, G., and Ruoslahti, E. (1977) J. Erp. Med.
145,819-827 Jalanko, H. (1979) Int. J. Cancer 24,394-397 Belayew, A., and Tilghman, S. M. (1982) Mol. Cell. Biol. 2, 1427-
1435 Tilghman, S. M., and Belayew, A. (1982) Proc. Natl. Acad. Sci.
U. S. A. 79,5254-5257 Ruoslahti, E., and Seppala, M. (1979) Adu. Cancer Res. 29,275-
346 Watanabe, A., Miyazaki, M., and Taketa, K. (1976) Int. J. Cancer
17,518-524 Mifflin, R. C., Moller, P. C., and Papaconstantinou, J. (1988)
Somatic Cell Mol. Genet. 14,553-566 Janzen, R. G., Andrews, G. K., and Tamaoki, T. (1982) Dev. Biol.
SO, 18-23 Dziadek, M., and Adamson, E. D. (1978) J. Embryol. Exp. Mor-
phol. 43,289-313 Dziadek, M. A., and Andrews, G. K. (1983) EMBO J. 2,549-554 Scott, R. W., Vogt, T. S., Croke, M. E., and Tilghman, S. M.
(1984) Nature 310, 562-576 Selten, G. C. M., Princin, H. M. G., Selten-Versteegen, A-M. E.,
Mol-Backx, P. B. M., and Yap, S. H. (1982) Biochim. Biophys. Acta 699, 131-137
Sellem, C. H., Frain, M., Erdos, T., and Sala-Trepat, J. M. (1984) Deu.Biol. 102,51-60
Cote, G. J., and Chiu, J-F. (1984) Biochem. Biophys. Res. Com- mun. 120,677-685
Nahon, J.-L., Tratner, I., Poliard, A., Presse, F., Poiret, M., Gal, A., Sala-Trepat, J. M., Legres, L., Feldmann, G., andBernuau, D. (1988) J. Biol. Chem. 263. 11436-11442
Tilghman, S. M. (1985) in Oxford Surueys in Eukaryotic Genes (Dawkins, R., and Ridley, M., eds) Vol. 2, pp. 160-206, Oxford University Press, London
54.
55. 56.
Gorski, K., Carneiro, M., and Schibler, U. (1986) Cell 47, 767- 776
Bradford, M. M. (1976) Anal. Biochem. 72, 248-254 Jones, K. A., Yamamoto, K. R., and Tjian, R. (1985) Cell 42,
559-572 Godbout, R. T., Ingram, R., and Tilghman, S. M. (1986) Mol. 57 Jose-Estanvol. M.. and Danan. J.-L. (1988) J. Biol. Chem. 263.
Cell. Biol. 6, 477-487 -..
39.
40.
41.
42.
43.
44.
45.
46.
47.
48.
49.
50. 51.
52.
53.
19. Widen, S. G., and Papaconstantinou, J. (1986) Proc. Natl. Acad. Sci. U. S. A. 83, 8196-8200
20. Widen, S. G., and Papaconstantinou, J. (1987) Mol. Cell. Biol. 7, 2606-2609
21. Muglia, L., and Rothman-Denes, L. B. (1986) Proc. Natl. Acad. Sci. U. S. A. 85, 2548-2552
22. Guertin, M., LaRue, H., Bernier, L., Wrang, O., Chevrette, M., Gingras, M.-C., and Belanger, L. (1988) Mol. Cell. Biol. 8, 1398-1407
23. Krumlauf, R., Hammer, R. E., Tilghman, S. M., and Brinster, R. L. (1985) Mot. Cell. Biol. 5, 1639-1648
24. Camper, S. A., and Tilghman, S. M. (1989) Genes & Deu. 3,537- 546
25. Ott, M.-O., Sperling, L., Herbomel, P., Yaniv, M., and Weiss, M. C. (1984) EMBO J. 3, 2505-2510
26. Friedman, M. M., Babiss, L. F., Clayton, D. F., and Darnell, J, E., Jr. (1986) Mol. Cell. Biol. 6, 3791-3797
58.
59.
60. 61.
62.
63.
64.
65.
66.
973 Maire, P., Wuarin, J., and Schibler, U. (1989) Science 244,343-
346 Costa, R. H., Grayson, D. R., and Darnell, J. E., Jr. (1989) Mol.
Cell. Biol. 9, 1415-1425 Cereghini, S., Blumenfeld, M., and Yaniv, M. (1988) Genes &
Deu. 2,957-974 Rosenfeld, P. J., and Kelly, T. J. (1986) J. Biol. Chem. 261,
1398-1408 Jones, K. A., Kadonaga, J. T., Rosenfeld, P. J., Kelly, T. J., and
Tjian, R. (1987) Cell 48, 79-89 Santoro, C., Mermod, N., Andrews, P. C., and Tjian, R. (1988)
Nature 334.218224 Cereghini, S., Raymondjean, M., Carranca, A. G., Herbomel, P.,
and Yaniv, M. (1987) Cell 50,627-638 Dorn, A., Bollekens, J., Staub, A., Benoist, C., and Mathis, D.
(1987) Cell 50,863-872 Raymondjean, M., Cereghini, S., and Yaniv, M. (1989) Proc. Natl.
Acad. Sci. U. S. A. 35,757-761 Monaci, P., Nicosia, A., and Cortese, R. (1988) EMBO J. 7,2075-
2087 Johnson, P. F., Landschulz, W. H., Graves, B. J., and McKnight,
S. L. (1987) Genes & Dev. 1, 133-146 Shaul, Y., and Ben-Levy, R. (1987) EMBO J. 6,1913-1920 Graves, B. J., Johnson, P. F., and McKnight, S. L. (1986) Cell
44,565-576 Maniatis, T., Fritsch, E. F., and Sambrook, J. (1982) Molecular
Cloning: A Laboratory Manual, Cold Spring Harbor Laboratory, Cold Spring Harbor, NY
Davis, L. G., Dibner, M. D., and Battey, J. F. (1986) Basic Methods in Molecular Biology, Elsevier Science Publishing Co., New York
10865-lO-87i I
Jose-Estanyol, M., Poliard, A., Fioret, F., and Danan, J.-L. (1989) Eur. J. Biochem. 181.761-766
Weiher, H., Konig, M., and Grass, P. (1981) Science 219, 626- 631
Jones, N., Rigby, P., and Ziff, E. (1988) Genes & Dev. 2, 267-281 Molne, M., Hovart, C., Szpirer, J., and Szpirer, C. (1989) Nucleic
AcidsRes. 17.3447-3457 Baumhueter, S., Courtois, G., and Crabtree, G. R. (1988) EMBO
J. 7,2485-2493 Paonessa, G., Gounari, F., Frank, R., and Cortese, R. (1988)
EMBO J. 7,3115-3123 Belanger, L., Hamel, D., Lachance, L., Dufour, D., Trembly, M.,
and Gagnon, P. M. (1975) Nature 256,657-659 Commer, P., Schwartz, C., Tracy, S., Tamaoki, T., and Chiu, J-
F. (1979) Biochem. Biophys. Res. Commun. 89, 1264-1299 Guertin, M., Baril, P., Bartkowiak, J., Anderson, A., and Belan-
ger, L. (1983) Biochemistry 22,4296-4302
by guest on February 12, 2018http://w
ww
.jbc.org/D
ownloaded from

D E Zhang, P R Hoyt and J Papaconstantinouregions of the mouse alpha-fetoprotein gene.
Localization of DNA protein-binding sites in the proximal and distal promoter
1990, 265:3382-3391.J. Biol. Chem.
http://www.jbc.org/content/265/6/3382Access the most updated version of this article at
Alerts:
When a correction for this article is posted•
When this article is cited•
to choose from all of JBC's e-mail alertsClick here
http://www.jbc.org/content/265/6/3382.full.html#ref-list-1
This article cites 0 references, 0 of which can be accessed free at
by guest on February 12, 2018http://w
ww
.jbc.org/D
ownloaded from