localization of nadphd-exhibiting neurons in the spinal cord of the rabbit
TRANSCRIPT

Localization of NADPHd-ExhibitingNeurons in the Spinal Cord of the Rabbit
JOZEF MARSALA,1* MARTIN MARSALA,2 IVO VANICKY,1 AND YUTAKA TAIRA3
1Institute of Neurobiology, Slovak Academy of Sciences, Kosice 4, Slovak Republic2Anesthesiology Research Laboratory, University of California–San Diego,
La Jolla California, 920933Department of Anesthesiology, Faculty of Medicine, University of the Ryukyus,
Okinawa, Japan
ABSTRACTSegmental and laminar distributions of nicotinamide adenine dinucleotide phosphate
diaphorase (NADPHd)-exhibiting neurons were examined in the rabbit spinal cord by usinghorizontal, sagittal, and transverse sections. A large number of NADPHd-positive neurons inthe spinal cord of rabbit appeared to fall into six categories (N1–N6), but others could not beclassified. Major cell groups of NADPHd-exhibiting neurons were identified in the superficialdorsal horn and around the central canal at all spinal levels and in the intermediolateral cellcolumn at thoracic and upper lumbar levels. NADPHd-exhibiting neurons of the pericentralregion were divided into a thin subependymal cell column containing longitudinally arranged,small bipolar neurons with processes penetrating deeply into the intermediolateral cellcolumn and/or running rostrocaudally in the subependymal layer. The second pericentral cellcolumn located more laterally in lamina X contains large, intensely stained NADPHd-exhibiting neurons with long dendrites radiating in the transverse plane. In the pericentralregion (lamina X), close association of NADPHd-exhibiting somata and fibers and mostlylongitudinally oriented blood vessels were detected. Neurons of the sacral parasympatheticnucleus, seen in segments S1–S3, exhibited prominent NADPHd cellular staining accompa-nied by heavily stained fibers extending from Lissauer’s tract through lamina I along thelateral edge of the dorsal horn to lamina V. A massive dorsal gray commissure, highly positivein NADPHd staining, was found in segments S1–S3. Scattered positive cells were also foundin the deeper dorsal horn, ventral horn, and white matter. Fiberlike NADPHd staining wasfound in the superficial dorsal horn and pericentral region in all the segments studied. Dense,punctate, nonsomatic NADPHd staining was detected in the superficial dorsal horn, in thepericentral region all along the rostrocaudal axis, and in the nucleus phrenicus (segmentsC4–C5), nucleus dorsalis (segments Th2–L2), Onuf ’s nucleus (segments S1–S3), and thedorsal part of the dorsal gray commissure (S1–S3). J. Comp. Neurol. 406:263–284, 1999.r 1999 Wiley-Liss, Inc.
Indexing terms: NADPHd neuronal types; segmental distribution; blood vessels; pericentral region
Recently, small, morphologically heterogeneous popula-tions of neurons containing nicotinamide adenine dinucleo-tide phosphate diaphorase (NADPHd) and/or nitric oxidesynthase (NOS), an enzyme that generates nitric oxide(NO), have been identified at different sites in the centralnervous system in a number of mammalian species includ-ing the human brain (Kowall et al., 1985, 1987; Kowall andMueller, 1988; Mizukawa et al., 1989; Bredt et al., 1990;Vincent and Kimura, 1992; Egberongbe et al., 1994; Vin-cent, 1994). Immunohistochemistry of neuronal nitric ox-ide synthase (nNOS) showed that the occurrence of thisenzyme is almost completely homotopic with the localiza-tion of neurons stained for NADPHd.
It has long been known that NADPHd-exhibiting neu-rons are present in the spinal cord, especially in thesubstantia gelatinosa, lamina X, and the int1ermediolateralcell column (Thomas and Pearse, 1964; Mizukawa et al.,1989; Blottner and Baumgarten, 1992; Bredt and
Grant sponsor: Slovak Academy of Sciences; Grant number: 2/4180/97;Grant sponsor: National Institute of Health; Grant number: NS 32794.
*Correspondence to: Jozef Marsala, M.D., Ph.D., Institute of Neurobiol-ogy, SAS, 040 01 Kosice, Soltesovej 4, Slovak Republic.E-mail: [email protected]
Received 16 February 1998; Revised 29 October 1998; Accepted 5November 1998
THE JOURNAL OF COMPARATIVE NEUROLOGY 406:263–284 (1999)
r 1999 WILEY-LISS, INC.

Snyder, 1992). However, the analysis of NADPHd-exhibit-ing and/or NOS-immunoreactive neurons in the spinalcord of different animal species (Anderson, 1992;Valtschanoff et al., 1992; Dun et al., 1993; Vizzard et al.,1994b, 1995) have shown a morphologically heterogeneouspattern of NADPHd-exhibiting neuronal pools rangingfrom bipolar, poorly branched NADPHd-exhibiting neu-rons in the superficial dorsal horn to highly differentiatedneurons in the pericentral region (lamina X), deep dorsalhorn (laminae IV–V), and dorsal gray commissure contain-ing widely branching NADPHd-exhibiting neurons (Ander-son, 1992; Valtschanoff et al., 1992; Dun et al., 1993; Lee etal., 1993; Saito et al., 1994; Vizzard et al., 1994a; Wetts andVaughn, 1994; Burnett et al., 1995; Marsala et al., 1997,1998). In general, NADPHd staining is present in neuronsand fibers in the superficial dorsal horn, in neurons aroundthe central canal at all levels of the spinal cord, in thedorsal commissure, in the sacral parasympathetic nucleus(SPN), and in the intermediolateral cell column (IML) ofthe thoracolumbar segments in the different species stud-ied. However, the lack of NADPHd staining of cells in theregion of the SPN of the cat has been reported (Vizzard etal., 1994a). Moreover, NADPHd histochemical and cholineacetyltransferase immunocytochemical analysis combinedwith vasoactive intestinal polypeptide (VIP) immunohisto-chemistry of lower lumbar and sacral segments has shownthat the distribution of NADPHd-exhibiting afferent fibersin Lissauer’s tract and the lateral collateral pathway issimilar to the distribution of visceral afferents (de Groat etal., 1978; Morgan et al., 1981) and VIP-containing afferentprojection in these areas in the cat (Vizzard et al., 1994a).
Remarkable progress has been made in understandingthe role of NO in cerebral blood flow regulation underphysiological and/or ischemic conditions, showing that NOreleased within the gray and white matters may relaxvascular smooth muscle and produce vasodilation by acti-vating soluble guanylate cyclase and increasing cGMP,thus coupling regional blood flow with neural activity(Bredt et al., 1990; Gally et al., 1990; Hope et al., 1991;Iadecola, 1985, 1993; Faraci and Brian, 1994). However,little is known about the segmental and laminar distribu-tion of NO-synthesizing neurons in the spinal cord andtheir possible role in the spinal cord blood flow control.Until now, only two experiments have been done, consist-ing of a long-term and a short-term inhibition of NOSactivity in the spinal cord (Blot et al., 1994; Yezierski et al.,1996). A long-term inhibition of NOS (Blot et al., 1994) oran intraspinal injection of NOS inhibitor (Yezierski et al.,1996) resulted in considerable damage of the spinal cordgray matter, characterized by a massive occurrence ofspinal cord infarcts at the cervical and cervicothoraciclevels followed by front leg paralysis (Blot et al., 1994) orneuronal damage and loss (Yezierski et al., 1996). Theseresults suggest that NO is a key factor in spinal cordarteriolar vasomotion and structure and that basal NOSactivity is essential in maintaining the structural integrityof spinal neurons. In accordance with these findings, aremarkably well-preserved spinal cord gray matter, lami-nae I, II, and X, regions rich in NADPHd-exhibitingneurons, were found in a model of transient spinal cordischemia in rabbit (Marsala et al., 1989, 1997) and dog(Marsala et al., 1989, 1991). However, a more detaileddescription of NADPHd-exhibiting neurons was needed fora better understanding of NO-synthesizing neurons in thespinal cord arteriolar vasomotion control and structure.
The aims of this study were to (1) describe the segmentaland laminar distributions of histochemically identifiedNADPHd-exhibiting neurons and their possible relationwith intraspinal blood vessels and (2) specify the neuronaltypes of NADPHd-exhibiting neurons and their location inthe spinal cord gray matter layers (laminae) of the rabbit.
MATERIALS AND METHODS
Tissue sampling, sectioning,and examination of sections
These experiments were performed on 12 adult rabbitsof both sexes that weighed 2.5–3.5 kg. The animals wereput into one of two groups. In the first group (n 5 9) theanimals were deeply anesthetized with pentobarbital (50mg/kg, i.v.) and perfused transcardially with saline fol-lowed by freshly prepared 4% paraformaldehyde 1 0.1%glutaraldehyde buffered with 1 M sodium phosphate, pH7.4. After perfusion fixation, the spinal cords were care-fully dissected out and stored in toto in the same fixativefor 3–4 hours. After postfixation, the spinal cord wasdivided into cervical (C1–C7), thoracic (Th1–Th13), lum-bar (L1–L7), and sacral (S1–S4) segments, and eachsegment was then divided into three small blocks compris-ing the upper, middle, and lower segmental levels, respec-tively. Because the spinal cord segment is, by definition,that part of the cord which gives rise to those root fibersthat unite to form a pair of spinal nerves (Brodal, 1969),the caudal border of a segment was defined by its mostcaudal dorsal rootlet (McKenna and Nadelhaft, 1986).Specimens were then cryoprotected in an ascending concen-tration of sucrose (15–30%) with the same phosphatebuffer and stored overnight at 4°C. Frozen horizontal,sagittal, and transverse sections (50 µm thick) were cutfrom all segments studied (Fig. 1) and processed forNADPHd activity by using a modified histochemical proce-dure (Scherer-Singler et al., 1983). In this procedure, thesections were incubated for 1 hour at 37°C in a solutioncontaining: 1 mg/ml of nitroblue tetrazolium (Sigma Chemi-
Fig. 1. Orientation figure showing three standard anatomicalplanes of cutting: horizontal (A–A), sagittal (B–B), and transverse(C–C).
264 J. MARSALA ET AL.
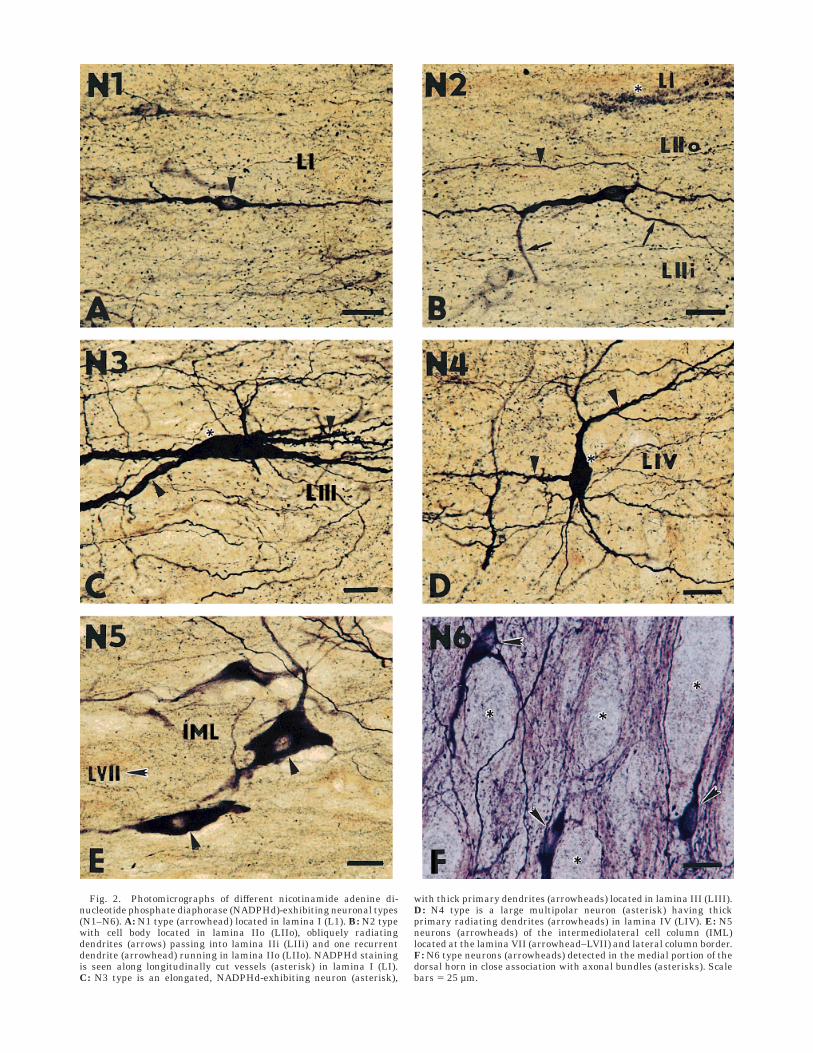
Fig. 2. Photomicrographs of different nicotinamide adenine di-nucleotide phosphate diaphorase (NADPHd)-exhibiting neuronal types(N1–N6). A: N1 type (arrowhead) located in lamina I (L1). B: N2 typewith cell body located in lamina IIo (LIIo), obliquely radiatingdendrites (arrows) passing into lamina IIi (LIIi) and one recurrentdendrite (arrowhead) running in lamina IIo (LIIo). NADPHd stainingis seen along longitudinally cut vessels (asterisk) in lamina I (LI).C: N3 type is an elongated, NADPHd-exhibiting neuron (asterisk),
with thick primary dendrites (arrowheads) located in lamina III (LIII).D: N4 type is a large multipolar neuron (asterisk) having thickprimary radiating dendrites (arrowheads) in lamina IV (LIV). E: N5neurons (arrowheads) of the intermediolateral cell column (IML)located at the lamina VII (arrowhead–LVII) and lateral column border.F: N6 type neurons (arrowheads) detected in the medial portion of thedorsal horn in close association with axonal bundles (asterisks). Scalebars 5 25 µm.
cal Co., Deisenhofen, Germany; N-6876), 0.5 mg/ml ofb-nicotinamide adenine dinucleotide phosphate (NADPH;Sigma Chemical Co.; N-1630), 0.8% Triton X-100 dissolvedin 0.1 M phosphate buffer (pH 8.0), and 1.25 mg/mlmonosodium malate (malic acid; Sigma Chemical Co.;M-1125).
NADPHd staining occurring in cells is described assomatic; the staining showing the dendritic and axonalprocesses is described as fiberlike; and dense neuropilNADPHd staining, where most of the reaction product has
a dotted appearance, is described as punctate, nonsomaticNADPHd positivity. The animals of the first group wereused only for qualitative analysis of the spinal cordNADPHd-exhibiting neuronal pool, and as a rule 15–25sections were used from each segment.
Control sections were treated in the same solutionbut without NADPH to enable testing for endogenousreduction activity in the corresponding blue formazanproduct. The sections were then rinsed in 0.1 M phosphate
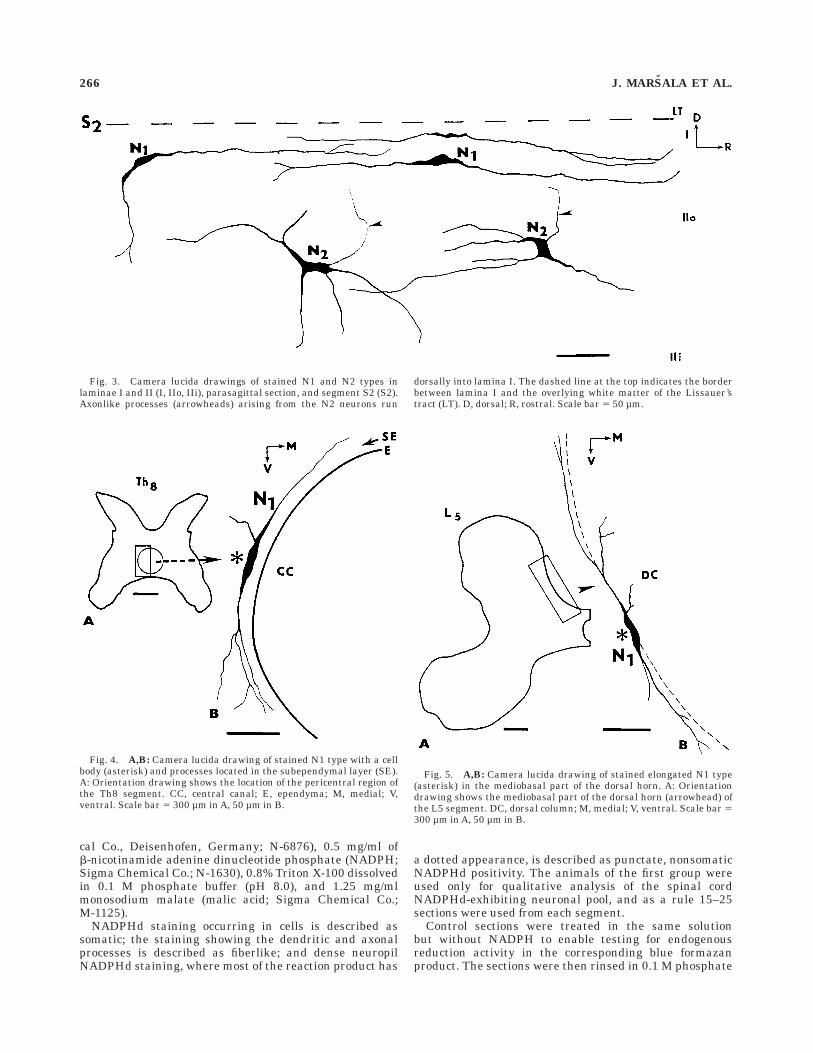
Fig. 3. Camera lucida drawings of stained N1 and N2 types inlaminae I and II (I, IIo, IIi), parasagittal section, and segment S2 (S2).Axonlike processes (arrowheads) arising from the N2 neurons run
dorsally into lamina I. The dashed line at the top indicates the borderbetween lamina I and the overlying white matter of the Lissauer’stract (LT). D, dorsal; R, rostral. Scale bar 5 50 µm.
Fig. 4. A,B: Camera lucida drawing of stained N1 type with a cellbody (asterisk) and processes located in the subependymal layer (SE).A: Orientation drawing shows the location of the pericentral region ofthe Th8 segment. CC, central canal; E, ependyma; M, medial; V,ventral. Scale bar 5 300 µm in A, 50 µm in B.
Fig. 5. A,B: Camera lucida drawing of stained elongated N1 type(asterisk) in the mediobasal part of the dorsal horn. A: Orientationdrawing shows the mediobasal part of the dorsal horn (arrowhead) ofthe L5 segment. DC, dorsal column; M, medial; V, ventral. Scale bar 5300 µm in A, 50 µm in B.
266 J. MARSALA ET AL.
buffer (pH 7.4), mounted on slides, air-dried overnight, andcoverslipped with Entellan. Some sections were stained bythe Nissl method to specify in more detail the laminardivision of the spinal cord gray matter.
Quantitative analysisof NADPHd-exhibiting neurons
In three additional adult male rabbits used for a quanti-tative assessment of the spinal NADPHd-exhibiting neu-rons, the previously described, slightly modified tissuefixation was used (Rothe et al., 1998). The animals weredeeply anesthetized with chloralhydrate (550 mg/kg bodyweight) and immediately after thoracotomy were perfusedtranscardially with 250 ml 0.1 M phosphate bufferedsaline followed by 4% freshly depolymerized paraformalde-hyde, 0.4% glutaraldehyde, and 2% sucrose in 0.1 Msodium phosphate buffer (pH 7.4), 1,200 ml for 12 minutesat room temperature. Subsequently, the spinal cordsand the dorsal and ventral roots were dissected andpostfixed in the same fixative for 3 hours. After post-fixation, an accurate anatomical identification of sevencervical (C1–C7), 13 thoracic (Th1–Th13), seven lumbar(L1–L7), and four sacral (S1–S4) segments was under-taken by using a dissection microscope. Segmentswere then cryoprotected in an ascending concentrations ofsucrose (15–30%) with the same phosphate buffer andstored overnight at 4°C. Before sectioning, each seg-
ment was longitudinally divided at the level of thedorsal median sulcus and ventral median fissure. Subse-quently, 10 transverse serial sections (30 µm thick)were cut on a Vibratome from the left half of eachC1–C7, Th1–Th13, L1–L7, and S1–S4 segments. Alto-gether, 310 transverse sections from each spinal cord wereprocessed for NADPHd activity. The right half wascut sagittally (30-µm-thick sections), and 30–40 sectionswere made from each half-segment. To improve cytoarchi-tectonic orientation and to specify in more detail thelaminar division of the spinal cord gray matter (Rexed,1954), some of the sections were counterstained withneutral red (Tolivia and Tolivia, 1985). Outlines of everysection and NADPHd-exhibiting neurons were obtained bymeasuring the major and minor axes on each cell outline.Actual sizes of each cell were obtained by converting thesedistances and applying the appropriate magnification fac-tor. The number of the N1–N6 NADPHd-exhibiting neuro-nal types are given as mean 6 S.E.M. Altogether, 37intervals between the groups of NADPHd-exhibiting neu-rons located in the IML were measured in three differentrabbits.
The use of experimental animals was approved byAnimal Care and Use Committee of Institute of Neurobiol-ogy, SAS, and all experimental procedures were conductedin compliance with the Guidelines of the NIH Guide forCare and Use of Laboratory Animals.
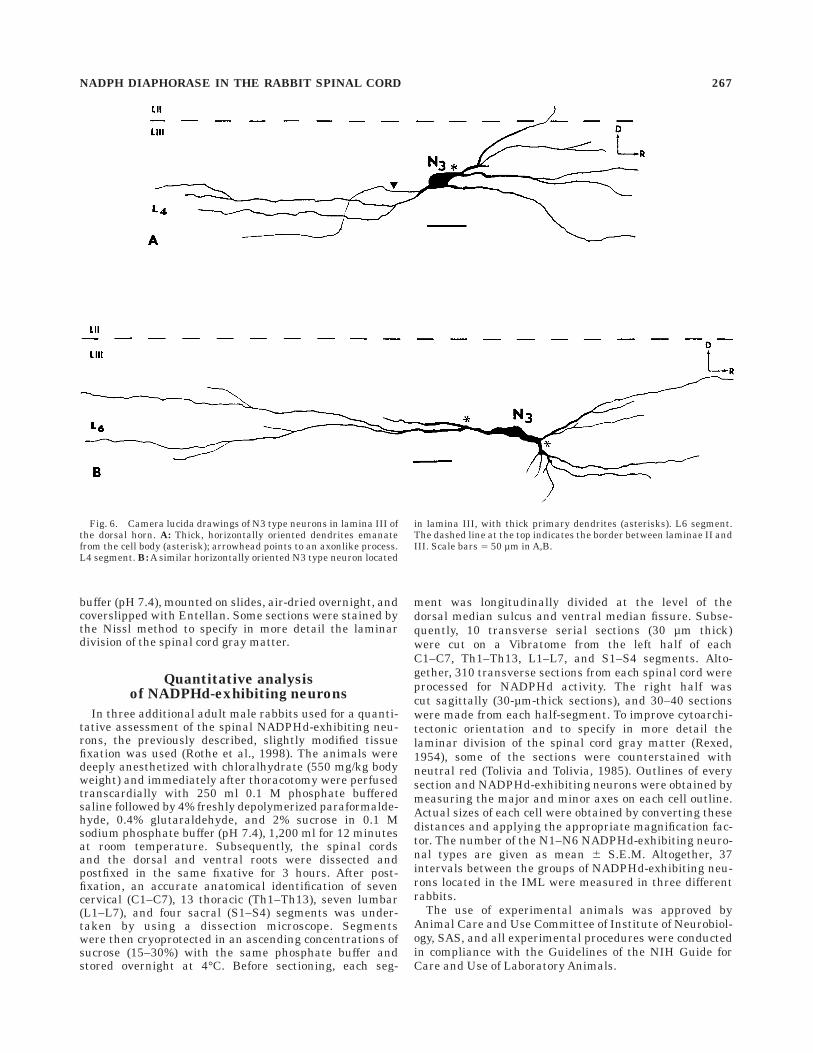
Fig. 6. Camera lucida drawings of N3 type neurons in lamina III ofthe dorsal horn. A: Thick, horizontally oriented dendrites emanatefrom the cell body (asterisk); arrowhead points to an axonlike process.L4 segment. B: A similar horizontally oriented N3 type neuron located
in lamina III, with thick primary dendrites (asterisks). L6 segment.The dashed line at the top indicates the border between laminae II andIII. Scale bars 5 50 µm in A,B.
NADPH DIAPHORASE IN THE RABBIT SPINAL CORD 267
RESULTS
Typology and quantitative analysisof NADPHd-exhibiting neurons
After considering the differences in the cell body size andshape, the number and length of primary dendrites, thepattern of secondary and tertiary dendritic arbors, thecourse and laminar orientation of the axons and axonlikeprocesses, a large number of neurons in the spinal cord ofrabbit appeared to fall into six categories, N1–N6, butothers could not be classified.
N1. This type is represented by small, elongated bipo-lar, lightly and moderately stained neurons (8–15 µm indiameter), with thick primary dendrites arising fromrostral and caudal poles of the perikarya and extending for300–500 µm in the rostrocaudal direction (Figs. 2A, 3). Thedendrites are largely confined to lamina I, with some ofmost peripheral branches penetrating obliquely into laminaIIo (Fig. 3, upper row). The terminal portion of someprimary dendrites is divided into short terminal arbors.N1 is currently seen in the outermost dorsal horn layers(laminae I–II). Similar but more elongated longitudinallyand transversally oriented bipolar neurons are seen in the
subependymal layer in the pericentral region (Fig. 4), inthe region corresponding to the internal basilar nucleus(Molander et al., 1984), oriented parallel to the medialborder of the dorsal horn (Fig. 5), in the ventralmostportion of the anterior horn and in the intermediate zone.
N2. Round, triangular, or oval neurons (8–12 µm indiameter) are more often found in layer IIo or at thelamina I–II border. Two to four obliquely running mediolat-erally compressed dendrites, seen on sagittal sections,pass into layer IIi or sometimes into lamina III (Fig. 3,lower row). The fine axonlike processes arising from theouter surface of the cell body are often seen entering layerI, thus bearing a resemblance with stalked cells, based onthe study of Golgi-stained sections (Gobel, 1975a,b, 1978),with the only interneurons in the superficial dorsal hornsending their axons into lamina I. Other N2 neuronssometimes occurring in small groups are more frequentlyfound in the deep part of the lamina II (Fig. 11A,B). Thecell bodies are small, i.e., less than 15 µm in diameter, andlie 100–200 µm from the inner border of the overlyingdorsolateral white matter. The cell bodies have largeprimary dendrites arising from their rostrocaudal poles.Dendritic ramifications give rise to secondary and long
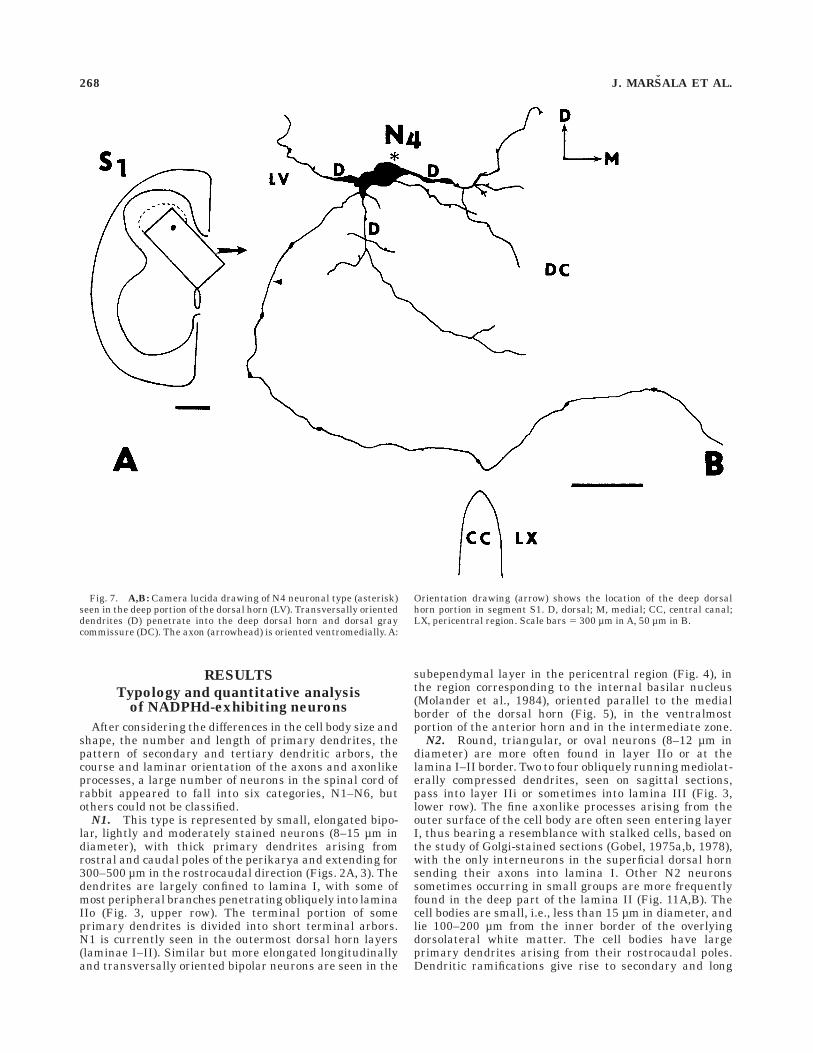
Fig. 7. A,B: Camera lucida drawing of N4 neuronal type (asterisk)seen in the deep portion of the dorsal horn (LV). Transversally orienteddendrites (D) penetrate into the deep dorsal horn and dorsal graycommissure (DC). The axon (arrowhead) is oriented ventromedially. A:
Orientation drawing (arrow) shows the location of the deep dorsalhorn portion in segment S1. D, dorsal; M, medial; CC, central canal;LX, pericentral region. Scale bars 5 300 µm in A, 50 µm in B.
268 J. MARSALA ET AL.

tertiary branches. Some of them form recurrent branches,i.e., fine arbors that proceed either rostrally or caudallyand, after a short distance, turn back toward the cell body(Fig. 2B). The existence of the recurrent dendrites (Ben-nett et al., 1982) and the propensity of these neurons toform small clusters or islands (Gobel, 1979), seen onhorizontal and sagittal sections, suggest their classifica-tion as a part of the islet cells.
N3. This is an intensely stained and elongatedNADPHd-exhibiting neuronal type (50–80 µm in diam-eter) located in lamina III (Fig. 2C). Several thick, pri-mary, longitudinally oriented spiny dendrites interdigitatewith processes of adjacent neurons in lamina III and upperlamina IV (Fig. 6A,B), thus producing dendritic trees thatare elongated in the rostrocaudal direction, otherwisecharacteristic for neurons of the spinocervical tract (Brown,1982).
N4. Large, intensely stained multipolar somata (50–70µm in diameter), with three to five long radiating spinydendrites arborizing in the deep dorsal horn layers and inthe lateral portion of the pericentral area, were foundmainly in laminae IV–V and X in all the segments studiedand in the dorsal gray commissure of S1–S4 segments(Fig. 2D). The dendritic trees extend mostly dorsoven-trally. Occasionally, some dendritic arbors reached lami-
nae II and I. The axons, or axonlike processes, whenidentified, were oriented ventrally or could be traced, evenif quite rarely, into the dorsal funiculus or after crossinginto the lateral funiculus (Fig. 7).
N5. Moderately stained, middle-sized (30–50 µmin diameter) neurons were found in the IML of the thor-acic (Th2–Th13) and lumbar (L1–L3) segments (Fig.2E) and in the SPN. Lightly stained, primary, aspinydendrites of IML neuros are organized in loosely arrangedbundles running medially, laterally, and rostrocaudally(Fig. 8).
N6. Elongated or triangular somata (15–25 µm indiameter) of this unusual neuronal type, which displayshigh NADPHd activity, were found solely in the medialportion of the dorsal horn, mainly in the cervical andlumbosacral enlargements and in the lateral reticulatedarea at the caudal medulla–cervical spinal cord transition.Intensely stained cell bodies and their di- or trichoto-mously branching processes were always found in closeassociation with rostrocaudally oriented axonal bundleslocated in the lateral reticulated area when cut sagitally oraround vertically oriented axonal bundles occurring in themedial portion of the dorsal horn when cut horizontally(Figs. 1, 9).
Fig. 8. Camera lucida drawing of N5 neuronal type of the interme-diolateral cell column of segment Th8 between the lateral part oflamina VII (LVII) and the lateral funiculus (LF). IC, nucleus intercala-
tus spinalis; ILf, nucleus intermediolateralis pars funicularis; ILp,nucleus intermediolateralis pars principalis; E, ependyma; cc, centralcanal; LX, pericentral region; R, rostral; M, medial. Scale bar 5 50 µm.
NADPH DIAPHORASE IN THE RABBIT SPINAL CORD 269

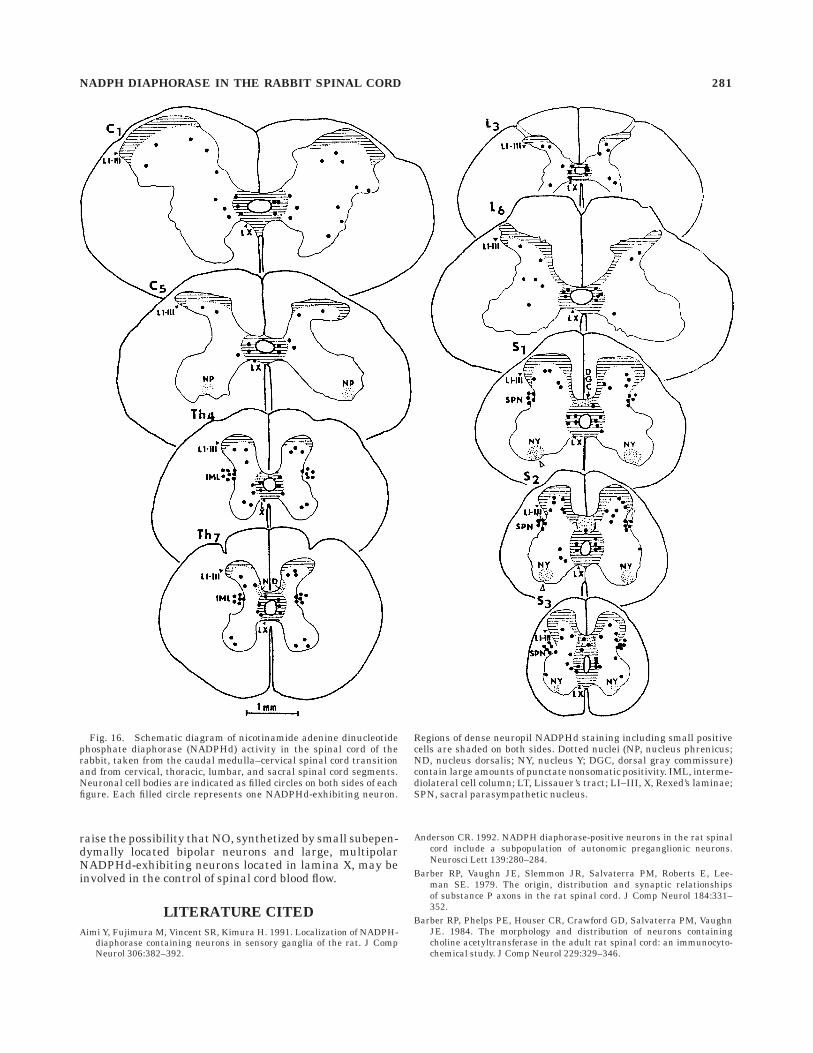
Segmental quantitative analysis of NADPHd-exhibitingneurons shows that, along with the lower cervical seg-ments, a high accumulation of NADPHd-positive neuronsoccurred in S1–S4 segments (Table 1). The percentages forthe regional distribution of identified N1–N6 types in thecervical (C1–C7), thoracic (Th1–Th13), lumbar (L1–L7),
and sacral (S1–S4) segments is 66.3%, 65.8%, 65.5%, and73.3%, respectively. Among identified NADPHd-positiveneurons, the N6 type clearly prevails (Table 2).
Laminar distribution of morphologicallyidentified NADPHd-exhibiting neurons
in the dorsal horn
Two distinctly different NADPHd-exhibiting neuronalpools, i.e., parvicellular and magnocellular, are traceablein the dorsal horn (laminae I–VI) throughout the rostrocau-dal extent of the spinal cord (Fig. 10). The parvicellularcomponent, represented by N1, N2, and N6 neuronaltypes, is almost entirely located in the superficial laminaeof the dorsal horn along with a high accumulation ofpunctate nonsomatic NADPHd positivity and fiberlikeNADPHd staining. However, considerable differences inthe density of N1, N2, and N6 types are seen across themediolateral extent of the superficial dorsal horn, mainlyin the cervical and lumbosacral enlargements. Whereaselongate bipolar (N1) and round or oval (N2) neurons arecommonly seen in the lateral and rarely in the medialportion of the superficial dorsal horn, di- or trichotomouslybranching N6 neurons occur exclusively in the medialportion of the superficial dorsal horn in close associationwith vertically or obliquely oriented axonal bundles (Fig.11A–C). More ventrally, the number of N6 neurons isgreatly reduced; instead, a dense network of NADPHd-exhibiting fibers is seen among axonal bundles (Fig.11D,E). The transition between parvi- and magnocellularlayers is quite sharp, and in upper lamina III the numberof NADPHd-exhibiting puncta is greatly reduced, as is theappearance of large, rostrocaudally oriented, intenselystained NADPHd-exhibiting neurons (N3 type) that giveoff several thick primary dendrites reaching 800–1,200 µmin the rostrocaudal direction (Fig. 10). In the lower laminaIII and in laminae IV–V, large multipolar NADPHd-exhibiting neurons (N4 type), with widely branching longprimary and secondary spiny dendrites, could be followedin the ventral and mediolateral directions. Some N4neurons resemble long projection spinothalamic or wide-dynamic-range (WDR) neurons (Mendell, 1966; Brown,1982).
Pericentral region (lamina X) and the IML
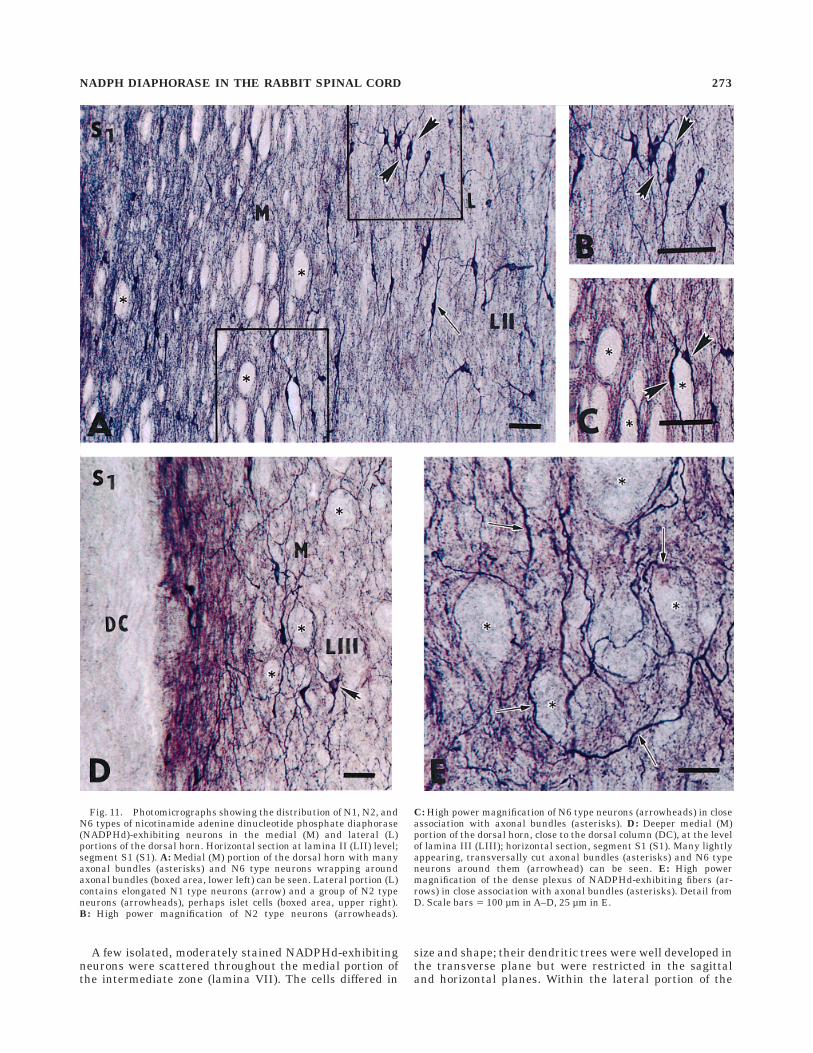
A substantial number of NADPHd-exhibiting but mor-phologically heterogeneous neurons were located aroundthe central canal (lamina X). First, parasagittal andhorizontal sections demonstrated a discontinuous columnor groupings of three to seven NADPHd-exhibiting neu-rons that were located laterally about 150–400 µm fromthe central canal (Fig. 12E,F). These neurons have thick,radiating dendrites, best seen on the transverse sections,and projecting toward the intermediate zone and theanterior horn. A second subset of NADPHd-exhibitingneurons, located in lamina X, can be described as anelongated, intensely stained bipolar somata (N1 type)occurring in the subependymal layer, 20–30 µm dorsolater-ally along the left and right borders of the central canal,mainly in the thoracic and upper lumbar segments(Fig. 12A,B). Their thin rostrally and caudally runningfibers in the subependymal layer could be followed for800–1,200 µm.
Densely clustered NADPHd-exhibiting neurons wereobserved in the IML in the thoracic (Th2–Th13) andlumbar (L1–L3) segments at the margin of the gray matterand the lateral funiculus. In horizontal and parasagittal
Fig. 9. B: Camera lucida drawing shows the N6 neuronal type(asterisks) in the medial portion of the dorsal horn in close associationwith axonal bundles (FB). A: Orientation drawing prepared fromhorizontally cut section of segment L6 shows the medial (M) andlateral (L) portions of the dorsal horn. R, rostral. Scale bars 5 300 µmin A, 50 µm in B.
270 J. MARSALA ET AL.
sections, the neurons of the IML were organized as dis-crete clusters occurring at almost equal intervals in therostrocaudal axis (Fig. 12A). In three different rabbits, themean interval measured on horizontal sections from cen-ter to center between IML clusters was 185 6 55 µm.Moderately or intensely stained NADPHd-exhibiting cellbodies had long, aspiny dendrites organized in smallbundles of variable shape and oriented in three maindirections. Medially oriented bundles proceed through theintermediate zone (lamina VII) and nucleus intercalatusand reach the subependymal layer; some of them turn atright angles rostrally or caudally and proceed in thesubependymal layer as part of the pericanalicular systemcontaining the processes of small, bipolar NADPHd-exhibiting fibers and longitudinally oriented vascular struc-tures (Fig. 12E,F). A few laterally oriented fibers penetratethe lateral funiculus, and some of them may reach the pialsurface. More dispersed, rostrocaudally running bundlesinterconnect the distinct IML groups. At the same segmen-tal level, a few NADPHd-exhibiting neurons are observedmedially within the intercalated nucleus and in the innerportion of the lateral funiculus (Fig. 8).
A close approximation of NADPHd-exhibiting somataand fibers with rostrocaudally oriented vascular struc-tures was found in the pericentral region for the entirelength of the spinal cord. The analysis of transversally and
parasagitally cut sections showed that two NADPHd-exhibiting neuronal pools participate in this approxima-tion. First, subependymally located vessels are often foundclosely apposed by small elongated or oval intensely stainedNADPHd-exhibiting somata (Fig. 13A) or by their subepen-dymally running fibers seen in close contact with thevessel wall (Fig. 13B). Moreover, some central canal clus-ter cells, known for their high NADPHd positivity, give off100–250-µm long branches running in the medial direc-tion, and some of them closely approximate subpendymalvessels (Fig. 13A). Second, more laterally located androstrocaudally oriented vessels in the pericentral regionare covered by many fine longitudinally and obliquelyoriented NADPHd-exhibiting fibers occurring in close ap-position with the vessel wall (Fig. 13C8,C). Parasagittalsections show that the central canal cluster cells are thesource of these perivascular fibers (Fig. 13D8,D).
Interestingly, a more conspicous accumulation ofNADPHd-exhibiting central canal cluster cells was foundat blood vessel bifurcation sites. However, a differentialrostrocaudal distribution pattern involving the NADPHd-exhibiting pool of large, multipolar neurons in lamina Xwas evident. We consistently found a greater number ofNADPHd-exhibiting neurons in lower lumbar and allsacral levels than in the cervical and thoracic levels. At thesame time, a rostrocaudal gradient was detected express-ing the increase of the cell body size of large NADPHd-exhibiting pericentral neurons, with the largest perikaryaand widely branching dendrites occurring in the lowersacral segments and the dorsal gray commissure.
SPN and NADPHd stainingof the Lissauer’s tract
Whereas only a few, mostly triangular, NADPHd-exhibiting neurons could be identified in the IML in L1–L4segments, the SPN which was found in segments S1–S3,
TABLE 1. Segmental Distribution of Six Identified Types (N1–N6) and Unidentified (UI) NADPHd-Exhibiting Neurons in the Spinal Cord of Rabbit1
Segment Total N1 N2 N3 N4 N5 N6 UI
C1 227.3 6 5.0 28.7 6 0.9 31.0 6 2.3 26.3 6 1.2 26.3 6 0.9 — 43.3 6 0.3 71.7 6 3.8C2 235.3 6 21.6 22.7 6 1.5 29.0 6 2.5 21.7 6 2.7 29.0 6 3.1 — 48.3 6 4.4 84.7 6 13.9C3 252.0 6 6.5 20.7 6 1.2 33.7 6 2.2 13.3 6 0.3 34.3 6 1.8 — 57.3 6 1.5 96.7 6 1.2C4 282.3 6 8.2 18.3 6 0.9 32.7 6 0.3 16.0 6 1.2 33.7 6 2.2 — 79.3 6 6.4 103.0 6 0.6C5 320.3 6 4.2 30.0 6 2.1 41.0 6 0.6 21.3 6 0.3 43.3 6 2.4 — 82.7 6 2.0 102.0 6 1.5C6 320.0 6 6.4 31.0 6 2.5 42.0 6 2.9 22.0 6 1.0 40.0 6 1.2 — 83.7 6 1.9 101.7 6 1.5C7 388.3 6 8.4 43.0 6 0.6 60.7 6 1.9 26.0 6 2.5 46.3 6 2.4 — 95.3 6 0.9 117.0 6 4.2Th1 181.7 6 10.2 16.3 6 0.3 25.7 6 1.2 7.7 6 1.3 18.3 6 1.5 — 32.0 6 1.5 81.3 6 4.7Th2 204.0 6 4.4 15.7 6 0.3 25.7 6 0.7 8.7 6 0.9 16.0 6 1.0 29.3 6 0.9 33.0 6 2.6 76.7 6 3.7Th3 175.7 6 6.7 12.3 6 0.9 20.3 6 2.2 6.3 6 0.7 16.3 6 3.3 27.7 6 0.3 32.0 6 1.5 60.3 6 4.8Th4 195.0 6 7.1 12.0 6 0.6 19.3 6 1.2 6.7 6 1.2 16.0 6 2.6 32.3 6 1.5 41.0 6 3.2 68.3 6 1.5Th5 203.3 6 7.3 9.7 6 1.5 18.7 6 1.2 7.0 6 0.6 24.0 6 3.5 31.7 6 0.3 42.6 6 4.1 69.7 6 5.3Th6 211.7 6 4.3 7.7 6 0.3 20.3 6 2.0 6.0 6 0.6 23.3 6 2.2 39.0 6 1.7 45.0 6 2.6 71.3 6 0.9Th7 213.0 6 4.0 7.3 6 0.3 18.3 6 0.9 6.7 6 0.3 24.0 6 1.5 32.7 6 1.2 52.3 6 6.7 72.0 6 1.7Th8 219.0 6 9.8 6.7 6 0.3 20.3 6 0.9 5.3 6 0.3 24.7 6 0.3 31.3 6 0.3 57.3 6 8.7 73.3 6 1.8Th9 217.7 6 2.7 6.3 6 0.9 22.3 6 1.2 5.3 6 0.3 22.3 6 0.3 33.7 6 0.3 55.3 6 1.3 72.3 6 2.0Th10 227.7 6 2.3 6.7 6 0.3 19.3 6 0.3 5.3 6 0.3 22.7 6 1.7 38.0 6 2.6 63.3 6 2.3 72.3 6 1.5Th11 229.3 6 7.4 6.0 6 0.6 21.0 6 1.5 5.7 6 0.3 24.7 6 0.7 38.0 6 2.9 64.7 6 2.8 69.3 6 1.3Th12 226.0 6 4.7 6.3 6 0.3 21.0 6 2.5 6.7 6 0.7 23.3 6 1.7 39.0 6 1.0 58.7 6 0.9 71.0 6 2.0Th13 235.0 6 7.1 6.3 6 0.3 23.0 6 0.6 6.3 6 0.3 26.7 6 2.3 41.0 6 1.2 60.0 6 3.6 72.3 6 4.7L1 218.3 6 3.2 7.3 6 1.5 21.3 6 0.3 7.3 6 0.9 23.3 6 0.7 34.0 6 1.5 54.3 6 3.8 70.7 6 2.3L2 229.0 6 6.0 8.0 6 1.5 18.0 6 0.6 6.3 6 0.9 21.7 6 0.9 37.7 6 0.9 65.3 6 1.2 71.0 6 2.6L3 227.3 6 4.8 7.7 6 1.3 21.3 6 0.3 6.3 6 0.9 23.3 6 0.7 34.7 6 2.4 61.0 6 4.4 73.0 6 1.5L4 226.0 6 2.5 7.7 6 0.3 23.0 6 0.6 6.6 6 0.3 31.3 6 1.9 8.0 6 1.7 59.7 6 3.3 89.7 6 0.3L5 228.7 6 5.2 7.3 6 0.9 25.0 6 0.6 7.0 6 1.0 30.3 6 2.2 — 67.7 6 3.8 91.3 6 2.0L6 248.3 6 5.2 8.3 6 0.9 29.0 6 2.0 14.0 6 0.6 31.3 6 0.3 — 78.0 6 3.1 87.7 6 0.3L7 305.3 6 3.3 18.0 6 1.2 38.7 6 0.9 19.7 6 2.4 48.0 6 2.3 — 86.0 6 1.5 95.0 6 1.2S1 378.7 6 11.2 30.3 6 1.2 53.7 6 0.3 31.7 6 0.9 53.3 6 1.3 34.7 6 3.7 78.7 6 4.7 96.7 6 3.9S2 379.3 6 23.1 28.3 6 0.3 55.7 6 2.4 35.0 6 0.6 59.7 6 2.3 35.7 6 4.2 75.3 6 4.1 91.0 6 2.5S3 361.7 6 6.7 24.7 6 0.9 51.0 6 2.1 32.3 6 1.2 52.3 6 2.4 34.7 6 2.2 72.0 6 1.7 96.0 6 2.6S4 298.3 6 9.9 24.0 6 0.6 40.3 6 2.0 27.0 6 3.6 41.3 6 5.5 10.0 6 1.0 64.0 6 13.1 91.7 6 2.9
1The number of nicotinamide adenine dinucleotide phosphate diaphorase (NADPHd)-exhibiting neurons was summed up from 10 sections of each cervical, thoracic, lumbar, andsacral segments of three rabbits. Data are given as means 6 S.E.M.
TABLE 2. Percentage Distribution (%) of Six Identified Types (N1–N6)and Unidentified (UI) NADPHd-Exhibiting Neurons in the C1–C7,
Th1–Th13, L1–L7, and S1–S4 Segments of the Spinal Cord1
N1 N2 N3 N4 N5 N6 UI
C1–C7 9.63 13.20 7.39 12.44 — 23.80 33.75Th1–Th13 4.51 10.11 3.12 10.26 14.94 23.01 34.18L1–L7 3.71 10.34 3.83 12.20 7.30 27.88 34.52S1–S4 7.57 14.14 8.90 14.49 7.94 20.51 26.75
1The percentage distribution was counted from the total number of nicotinamideadenine dinucleotide phosphate diaphorase (NADPHd)-exhibiting neurons in thecervical (C1–C7, n 5 2,026), thoracic (Th1–Th13, n 5 2,739), lumbar (L1–L7, n 5 1,683),and sacral (S1–S4, n 5 1,418) segments.
NADPH DIAPHORASE IN THE RABBIT SPINAL CORD 271

was composed of loosely arranged, mainly oval and triangu-lar neurons displaying moderate and intense NADPHdstaining (Fig. 14A,B). Some small bipolar and triangularNADPHd-exhibiting neurons are found within SPN and inclose approximation with NADPHd-positive fibers of thelateral collateral pathway of the Lissauer’s tract. Small,irregularly distributed groups of SPN neurons, which canbe seen more clearly on horizontal sections, are intercon-nected by densely packed dendritic bundles similar tothose identified in the IML (Fig. 14C). However, mediallyor laterally oriented dendritic bundles could not be found.A moderately stained, vertically oriented fiber bundlemore conspicuously identified in segment S2 was seenextending from Lissauer’s tract and ending among the
neurons of the SPN. More dispersed, moderately stainedcells of SPN could be followed more ventrally in the lateralpart of lamina VII. Large, irregularly shaped patches ofNADPHd activity were detected in the white matter of theLissauer’s tract, appearing as a punctate, nonsomaticNADPHd staining located intraaxonally or more often inclose association with small vessels (Fig. 14D,E).
Segmental distribution of punctate,nonsomatic NADPHd activity and its relation
to some sensitive and motor nuclei
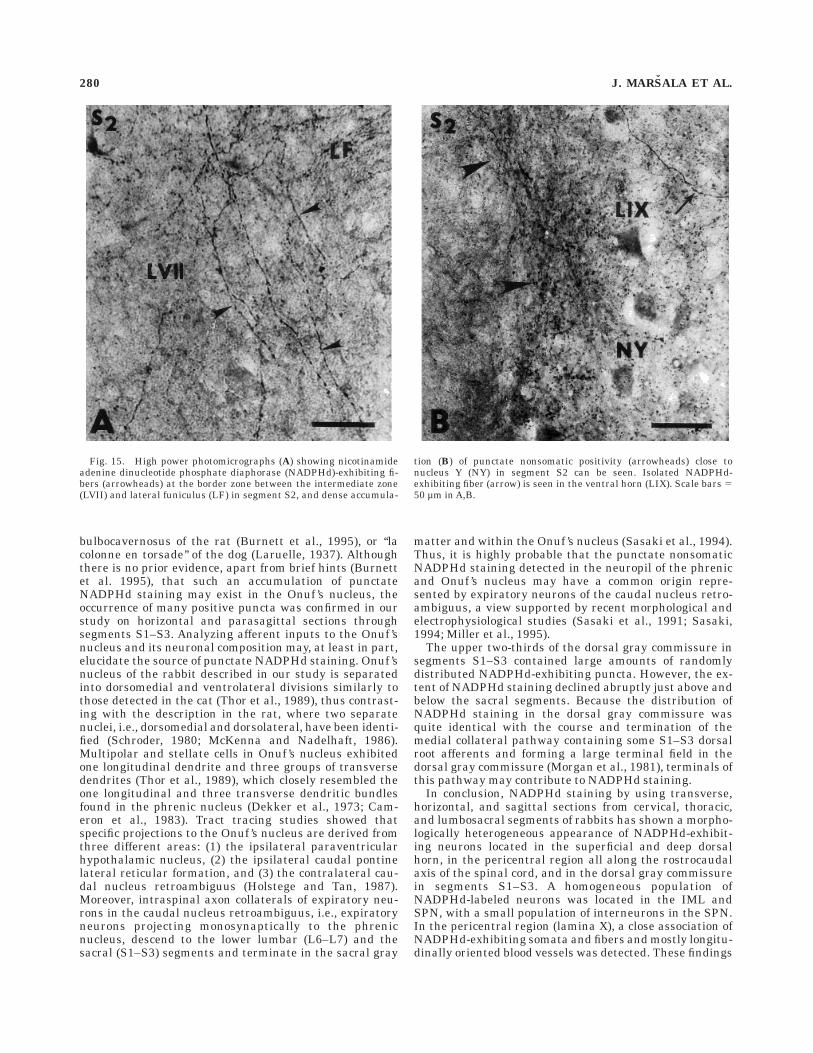
Prominent punctate, nonsomatic NADPHd staining oc-curred in the neuropil of four cytoarchitectonically well-defined nuclei. First, in the neuropil of the phrenic nucleuslocated in an intermediate position in the ventral horn ofC4–C5 segments (Ellenberger and Feldman, 1988) amongunstained cell bodies of phrenic motoneurons, intenselystained NADPHd puncta appeared (Fig. 16). Second, denseaccumulation of NADPHd-exhibiting puncta was seen inthe neuropil of the dorsal nucleus located in the ventrome-dial part of the dorsal horn (Yoss, 1953; Grant and Rexed,1958) and extending as a continuous cell column throughTh2–L2 segments (Fig. 16). Some intensely stainedNADPHd-exhibiting puncta were seen in close associationwith large sensitive neurons. Third, the neuropil of thedorsomedial and ventrolateral divisions of Onuf ’s nucleus(Onuf, 1901) or nucleus Y (Romanes, 1951), which werelocated ventromedial to the lateral motor nuclei and morecaudally found at the extreme lateral edge of the ventralhorn (Thor et al., 1989), extended rostrocaudally in S1–S3segments and had a dense accumulation of NADPHd-exhibiting puncta (Fig. 16). In some sections, mainly insegment S2, fiberlike and punctate NADPHd activity wasnoted between the SPN and Onuf ’s nucleus (Fig. 15A,B).However, no signs of NADPHd activity could be detected inthe multipolar and stellate neurons in Onuf ’s nucleus.
Dense punctate NADPHd staining and many multipolarNADPHd-exhibiting neurons were found in two-thirds ofthe dorsal gray commissure in segments S1–S3 (Fig. 16). Atransversally oriented band of the dorsal gray commissurecoalesced bilaterally with the medially located superficiallayers of the dorsal horn.
NADPHd activity in the central gray matter,ventral horn, and white matter
Occasionally, isolated NADPHd-exhibiting neurons werenoted in the inner portion of the ventral and lateralcolumns, along with NADPHd-exhibiting strandlike tractsthat penetrated deeply into the white matter. They weremost numerous in lumbar and sacral segments close to theventral commissure. In all planes of sectioning, fibersstained with different intensities were observed in thewhite matter. However, in horizontal and sagittal sections,some NADPHd-exhibiting fibers were traceable for hun-dreds of micrometers. A considerably higher number ofNADPHd-positive fibers was detected in the ventral por-tion of the dorsal column, mainly in the lower lumbar andsacral segments. However, the most prominent NADPHdfiber activity, along with a considerably enhanced stainingof vascular structures, was seen in the Lissauer’s tract inthe lower lumbar and S1–S3 segments and in a fiberbundle designated the lateral collateral pathway of theLissauer’s tract, which emerges ventrally from the Lissau-er’s tract in lamina I (Morgan et al., 1981) and passesalong the lateral edge of the dorsal horn into the SPN(Figs. 14A, 15D,E).
Fig. 10. The location of parvicellular and magnocellular nicotin-amide adenine dinucleotide phosphate diaphorase–exhibiting neuro-nal pools. Parasagittal section, segment S2 (S2); LI–LVII, LX, Rexed’slaminae; LT, Lissauer’s tract; VC, ventral column; N3 type (arrow-heads) in lamina III and N4 type (asterisks) in laminae V and X can beseen. Scale bar 5 100 µm.
272 J. MARSALA ET AL.
A few isolated, moderately stained NADPHd-exhibitingneurons were scattered throughout the medial portion ofthe intermediate zone (lamina VII). The cells differed in
size and shape; their dendritic trees were well developed inthe transverse plane but were restricted in the sagittaland horizontal planes. Within the lateral portion of the
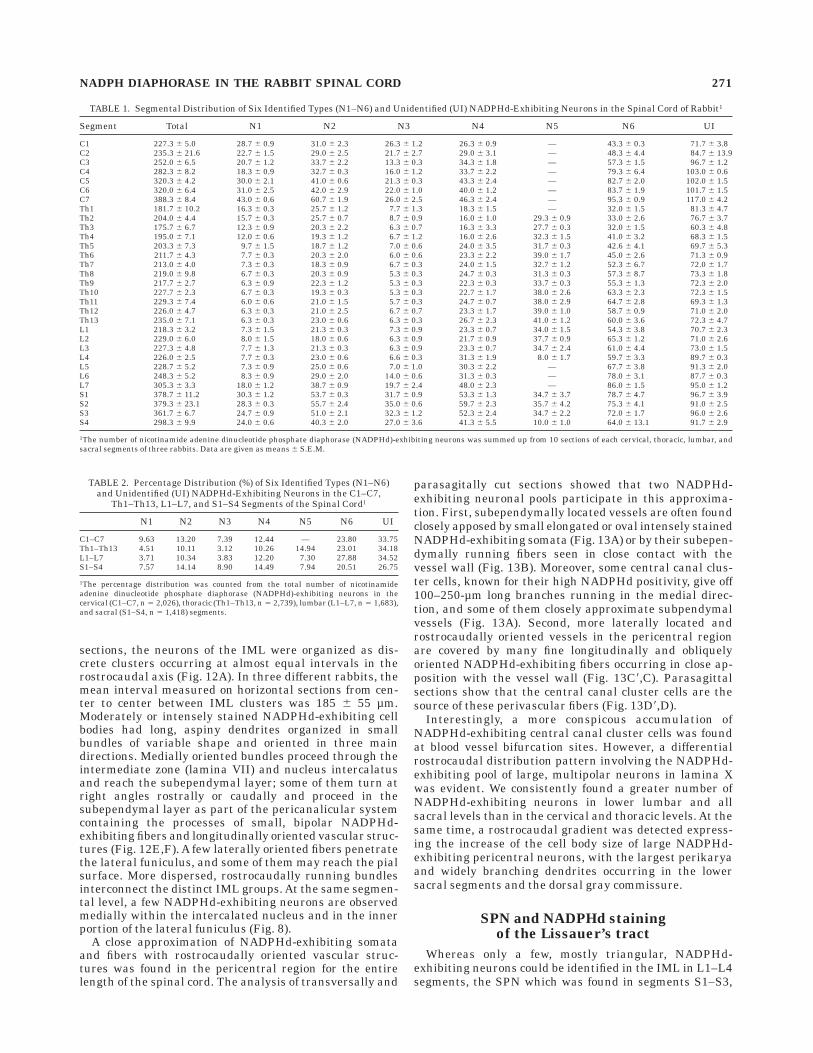
Fig. 11. Photomicrographs showing the distribution of N1, N2, andN6 types of nicotinamide adenine dinucleotide phosphate diaphorase(NADPHd)-exhibiting neurons in the medial (M) and lateral (L)portions of the dorsal horn. Horizontal section at lamina II (LII) level;segment S1 (S1). A: Medial (M) portion of the dorsal horn with manyaxonal bundles (asterisks) and N6 type neurons wrapping aroundaxonal bundles (boxed area, lower left) can be seen. Lateral portion (L)contains elongated N1 type neurons (arrow) and a group of N2 typeneurons (arrowheads), perhaps islet cells (boxed area, upper right).B: High power magnification of N2 type neurons (arrowheads).
C: High power magnification of N6 type neurons (arrowheads) in closeassociation with axonal bundles (asterisks). D: Deeper medial (M)portion of the dorsal horn, close to the dorsal column (DC), at the levelof lamina III (LIII); horizontal section, segment S1 (S1). Many lightlyappearing, transversally cut axonal bundles (asterisks) and N6 typeneurons around them (arrowhead) can be seen. E: High powermagnification of the dense plexus of NADPHd-exhibiting fibers (ar-rows) in close association with axonal bundles (asterisks). Detail fromD. Scale bars 5 100 µm in A–D, 25 µm in E.
NADPH DIAPHORASE IN THE RABBIT SPINAL CORD 273

intermediate zone, NADPHd-exhibiting somata were al-most completely lacking. In contrast, the central graymatter of the lower lumbar and sacral segments wasdensely packed with fiberlike NADPHd positivity.NADPHd-exhibiting neurons were rarely found in theventral horn, particularly in the medial portion of laminaVIII and in the ventralmost portion of lamina IX, the latterresembling Renshaw cells (Thomas and Wilson, 1965;Jankowska and Lindstrom, 1971; Willis, 1971). Moder-ately stained perikarya (10–15 µm in diameter) wereusually bipolar or spindle-shaped and were dispersedamong motoneurons that remained unstained. Both trans-verse and horizontal sections failed to demonstrateNADPHd-exhibiting neurons in the lateral spinal nucleuslocated in the dorsal part of the lateral funiculus (Gwynand Waldron, 1969).
DISCUSSION
The segmental and laminar distributions of NADPHdstaining in relation to specific neuronal types were exam-ined in the spinal cord of the rabbit. Knowledge about theanatomical distribution of NO-synthetizing somata andfibers within the spinal cord has been accrued mostly fromstudies using NADPHd histochemistry (Anderson, 1992;Bruning, 1992; Valtschanoff et al., 1992), NOS immunocy-tochemistry (Blottner and Baumgarten, 1992; Dun et al.,1993; Saito et al., 1994; Burnett et al., 1995) combinedwith choline acetyltransferase immunocytochemistry andVIP immunohistochemistry (Vizzard et al., 1994a) orcomparing differential localisation of NOS immunoreactiv-ity and NADPHd activity (Vizzard et al. 1994b). Thesegmental distribution of NADPHd-exhibiting neurons inthe rabbit is comparable to that described in the spinalcord of other species such as rat, mouse, cat, dog, andsquirrel monkey (Aimi et al., 1991; Valtschanoff et al.,1992; Dun et al., 1993; Vizzard et al., 1993a, 1994a,b; Saitoet al., 1994; Burnett et al., 1995; Marsala et al., 1998), withcertain differences noted in the superficial dorsal horn(laminae I–III) consisting in an increasing number ofNADPHd-exhibiting neurons along the rostrocaudal axisof the spinal cord and accumulation of NADPHd-exhibit-ing somata in the lower lumbar and in the sacral seg-ments. Moreover, in the lower lumbar and sacral seg-ments, a considerable increase in the cell body sizeof large, multipolar, pericentrally located (lamina X)NADPHd-exhibiting neurons was seen.
Several NADPHd-exhibiting cell types in laminae I–Vwere identified in the present study (Fig. 2A–F). Moreover,because NADPHd staining has been shown to produce aGolgi-like staining of NADPHd-exhibiting perikarya includ-ing dendrites and axons, there is an opportunity to com-pare Golgi-stained sections (Gobel, 1975a,b, 1978) withNADPHd-exhibiting neurons sharing the same morphologi-cal features. The N1 type, a small, bipolar neuron with arostrocaudally oriented cell body, is seen profusely through-out the lateral portion in laminae I and II, closely resem-bling bipolar cells with a few long, poorly branchingdendrites (Valtschanoff et al., 1992; Dun et al., 1993).Despite some morphological similarity with the ‘‘gelati-nosa cells’’ (Scheibel and Scheibel, 1968) or type A Golgi-stained spinotectal neurons (Lima and Coimbra, 1989), itseems likely that they form a part of local dorsal horncircuitry including possible participation in short ascend-ing and/or descending connections through Lissauer’s tract(Earle, 1952), known for its intense axonal NADPHd
staining in the lower lumbar and sacral segments (Vizzardet al., 1994a,b; Burnett et al., 1995). It is of substantialinterest, that, by combining three standard anatomicalplanes (Fig. 1), the NADPHd-exhibiting pool occurring inthe superficial dorsal horn, which is generally consideredto be uniformly distributed (Valtschanoff et al., 1992; Dunet al., 1993; Saito et al., 1994; Vizzard et al., 1994a,b), canbe divided into medial and lateral compartments easilyrecognizable in the cervical and lumbosacral enlargements(Figs. 9, 11A).
Whereas the lateral portion contains N1 and N2 types ofneurons evenly distributed through laminae I and II,without forming a line of cells extending mediolaterallyacross the dorsal horn as has been described in the dorsalhorn of the cat (Vizzard et al., 1994a), the medial portion ofthe gray matter is rich in the N6 type of di- or trichoto-mously branched NADPHd-exhibiting neurons, which canbe seen in close association with vertically oriented axonalbundles (Fig. 9). It seems likely that the N6 type occurs atleast in the cervical and lumbosacral enlargements in closeassociation with large bundles of afferents from the dorsalcolumn that are seen to penetrate the medial dorsal hornand to descend to medial laminae V and VI along withcollaterals of the medial collateral pathway (Morgan et al.,1981). This interpretation is supported by the periodicorganization of visceral primary afferents in the midlum-bar segments (Morgan et al., 1986). The analysis of thespatial relationship of the expression of NOS immunoreac-tivity, NADPHd histochemistry, and colocalization withJun, Fos, and Krox transcription factors in the spinal cordneurons after peripheral noxious stimulation showed thata significant increase in number of NADPHd somata andfibers could be determined in the medial part of the dorsalhorn (Herdegen et al., 1994), which is rich in the N6 type ofNADPHd-exhibiting neurons.
Fine structural characteristics of N2 neurons, closelyresembling stalked and islet cells and their strictly limitedoccurrence within lamina II, which contains five kinds ofinterneurons (Gobel, 1975a,b, 1978; Gobel et al., 1980),support the view that they represent a quite unique
Fig. 12. Photomicrographs documenting the occurrence of smallsubependymally located and nicotinamide adenine dinucleotide phos-phate diaphorase (NADPHd)-exhibiting neurons in the pericentralregion (lamina X) and their relation with the intermediolateral cellcolumn at the level of segment Th8 (A–D); horizontal section and theexistence of large intensely stained NADPHd-exhibiting neurons inlamina X; transverse section (E,F). A: Small, subependymally located(SE) neuron (asterisk) sending a laterally oriented process throughlamina X (LX) in the direction of the intermediolateral cell column(arrowheads-IML). Th8, segment Th8; CC, central canal; LC, lateralcolumn. B: Small elongated NADPHd-exhibiting somata (asterisks)and their rostrocaudally oriented processes can be seen in the subepen-dymal layer (SE) just below the ependyma lining the central canal(CC). Th8, horizontal section through segment Th8; LX, lamina X. C:Obliquely running processes (arrow) of small elongated NADPHd-exhibiting neuron in the subependymal layer (asterisk, upper right)pass through lamina X (LX) and lamina VII (LVII) and terminateamong neurons of the IML (asterisk, lower left. Th8, segment Th8; LC,lateral column. D: High power photomicrograph showing an elongatedNADPHd-exhibiting neuron (asterisk) sending off an obliquely run-ning process (arrow) passing through lamina VII (LVII); extremelyfine NADPHd-exhibiting rostrocaudally oriented fibers (arrowheads)can be seen in the subependymal layer (SE) below the ependymalining the central canal (CC). E,F: Large multipolar NADPHd-exhibiting neurons (asterisks) in the pericentral region (LX). S1,segment S1; CC, central canal. Scale bars 5 200 µm in A, 100 µm inB,C,E,F, 25 µm in D.
274 J. MARSALA ET AL.
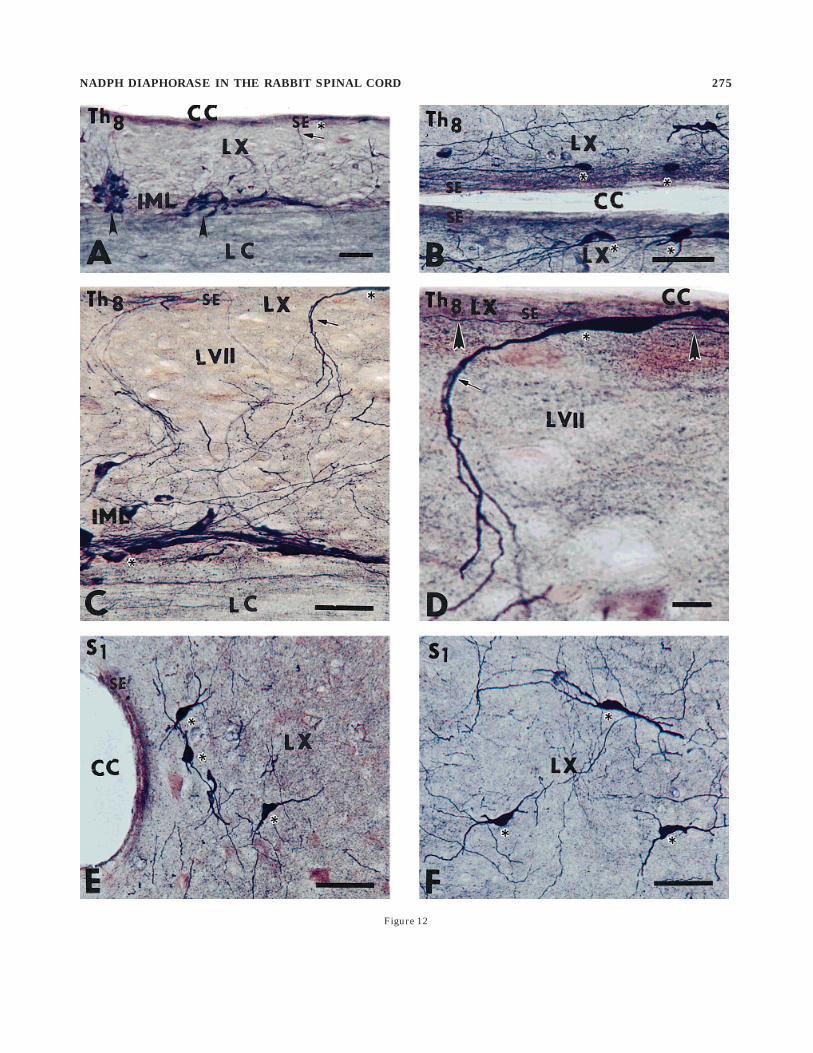
Figure 12
NADPH DIAPHORASE IN THE RABBIT SPINAL CORD 275

population of NADPHd-exhibiting interneurons participat-ing in the microcircuitry in the superficial dorsal horn(Gobel, 1979). Our finding concerning the stalked cells(Fig. 3), generally considered to be excitatory acting inter-neurons targeted at layer I projection neurons, raises thepossibility that some phenotypic diversity in the popula-tion of lamina II NADPHd-exhibiting stalked cells mayexist because some stalked cells and some of the islet cellsin the deep part of lamina II are enkephalin immunoreac-tive (Bennett et al., 1982). It is noteworthy that both thesuperficial dorsal horn layers (laminae I–II) and thelateral spinal nucleus, which receives substantial inputfrom peptidergic afferents (Ruda et al., 1986; Cliffer et al.,1988) and contains particularly high concentration ofcalbindin-D28k-like immunoreactivity (Menetrey et al.,1992), clearly differ with respect to the occurrence ofNADPHd-exhibiting neurons that are found in large num-bers in the superficial dorsal horn but that are completelylacking in the lateral spinal nucleus.
Both superficial layers are rich in punctate nonsomaticNADPHd positive staining (Fig. 10), presumed to be ofsynaptic origin and representing axonal terminals and/ordendritic spines (Saito et al., 1994). The former alternativeis supported by the existence of small, NADPHd-exhibit-ing neurons found in a high number in the lumbosacraldorsal root ganglia and projecting thin unmyelinatedfibers into laminae I and II. Whereas the finding of N1 andN2 neurons in both portions of the superficial dorsal hornmay have some relation with substance P, which is consid-ered to be a probable activator of NOS (Ross et al., 1990),the occurrence of the N6 type of di- or trichotomouslybranched NADPHd-exhibiting neurons, seen in close asso-ciation with axonal bundles, is still unclear and needsfurther study.
The N3 and N4 types of NADPHd-exhibiting neuronsoccurring in laminae III–IV were seen at all levels of thespinal cord (Figs. 2C,D, 6A,B, 7). However, a considerablyhigher number of these neurons was detected in thecervical and lumbosacral enlargements. Both types werelarger than those in the superficial laminae, and bothtypes had extremely long, rostrocaudally oriented den-dritic trees occurring in the N3 type in upper lamina IIIand long dorsoventrally and mediolaterally radiating den-drites in the N4 type. Besides differences in the cell bodysize and dendritic arborizations, some phenotypic differ-ences among NADPHd-exhibiting neurons in laminae IIand III–V are known. Whereas neurons in lamina IIexhibited only NADPHd, large multipolar neurons inlaminae III–V double labeled for choline acetyltransferaseand NADPHd clearly prevailed (Wetts and Vaughn, 1994).Moreover, the location of N3 and N4 neurons in laminaeIII–V is similar to the laminar distribution of WDRneurons (Maixner et al., 1986; Price, 1988). Because manyWDR neurons are spinothalamic, trigeminothalamic (Priceand Mayer, 1976; Price et al., 1978, 1979), dorsal columnpostsynaptic, and/or spinocervical tract neurons (Dubnerand Bennett, 1983), at least some N3 and N4 types ofNADPHd-exhibiting neurons, similar to those located inlamina X (Lee et al., 1993), may have a double function,i.e., NO synthesis and transmission of impulses throughcorresponding long ascending tracts.
The principal finding of the present study is thatNADPHd-exhibiting neurons in lamina X are closely asso-ciated with longitudinally oriented blood vessels located inthe pericentral region (Fig. 13A–D). Unlike the other
comparative studies describing the pericentral region(lamina X) as a homogeneously structured neuronalNADPHd-exhibiting accumulation (Valtschanoff et al.,1992; Dun et al., 1993; Saito et al., 1994; Vizzard et al.,1994a,b), two morphologically different NADPHd-positivepools, having a close relation with blood vessels, aredescribed in our study.
First, a subset of transversally and longitudinally ori-ented, intensely stained, bipolar NADPHd-exhibiting neu-rons representing the N1 type were identified in thesubependymal layer (Fig. 12A–D), thus topographicallyoverlapping with monoaminergic and peptidergic innerva-tion of the nucleus intercalatus pars subependymalis(Fuxe et al., 1990). Our finding describing elongatedNADPHd-exhibiting cell bodies and their branches thatsynthesize and release NO and pericentrally located vascu-lar structures indicates that, along with the bundles ofprocesses of IML neurons in the lateral funiculus accompa-nying large blood vessels and characteristic for whitematter (Valtschanoff et al., 1992), an elaborate NO-producing neuronal pool exists in the pericentral andsubependymal gray matter regions. Moreover, thin, longi-tudinally oriented processes of bipolar cells could befollowed in the subependymal layer for a long distance(Fig. 13D). Some of them then turned laterally, passedjointly with the dendritic bundles of IML cells running inthe opposite direction in the intermediate zone throughthe nucleus intercalatus spinalis (Janig and McLachlan,1987), and reached the medial border of the IML, wherethey terminated. Thus, autonomic motor neurons of IMLthat were double labeled for choline acetyltransferase andNADPHd or for choline acetyltransferase only (Wetts andVaughn, 1994), in close association with monoamine andthe neuropeptide system (Fuxe et al., 1990), are directlyinterconnected with subependymal NADPHd-exhibitingneurons. The functional significance of this interconnec-tion requires further study.
Second, another subset of pericentrally located NADPHd-exhibiting multipolar neurons was seen in parasagittaland transverse sections as a discontinuous column withmany long processes that extended mostly in the trans-verse plane (Fig. 12E,F). This finding, along with the densenetwork of fiberlike and punctate nonsomatic NADPHd
Fig. 13. Photomicrographs showing the occurrence of nicotinamideadenine dinucleotide phosphate diaphorase (NADPHd)-exhibiting neu-rons in the pericentral region and their close association with theblood vessels. A: High power photomicrograph shows a group ofintensely stained pericentral neurons (asterisk) and their processes(arrows) in lamina X (LX). A subependymally located neuron (arrow-head) is seen in close association with the blood vessel. CC, centralcanal; SE, subependymal layer. B: High power photomicrographshows the branching (arrows) of the bipolar neuron (asterisk) in thesubependymal layer (SE) close to the blood vessel (V). LX, pericentralregion. C8: Low power photomicrograph taken from the parasagittalsection of segment S1 shows many NADPHd-exhibiting neurons(asterisks) and their processes (arrowheads) in close association withthe vessel wall. C: High power photomicrograph of the same regionwith intensely stained neurons (asterisks) and their longitudinally orobliquely running branches (arrowheads) in close association with theblood vessel (V). D8: Low power photomicrophotograph taken from theparasagittal section cut through the pericentral region of S2 segment.Intensely stained cell bodies (asterisk) and their fine branches (arrow-heads) are seen in close association with the vessel wall (V). D: Highpower photomicrophotograph shows a group of cells (asterisk) and fineNADPHd-exhibiting fibers (arrowheads) in close association with theblood vessel (V). Scale bars 5 100 µm in C8,D8, 50 µm in A–D.
276 J. MARSALA ET AL.
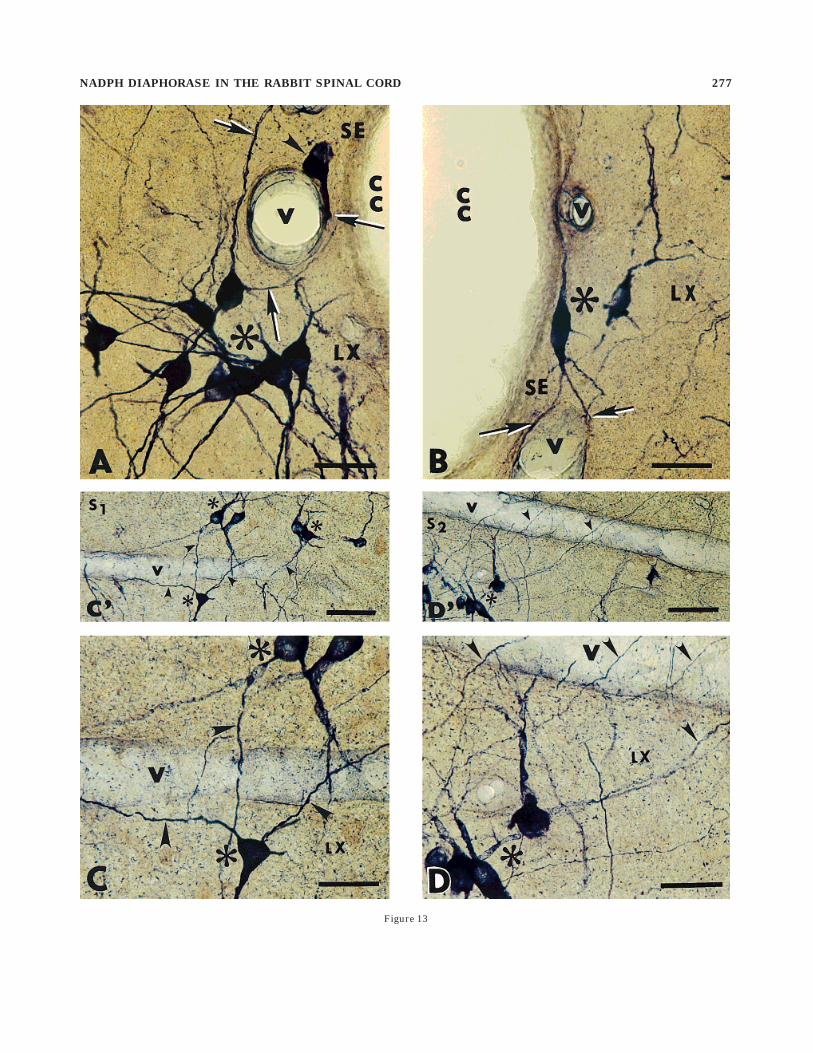
Figure 13
NADPH DIAPHORASE IN THE RABBIT SPINAL CORD 277

positivity seen in the pericentral region and the adjacentportion of the intermediate zone in lower lumbar andsacral segments, renders this region the most complicatedportion of spinal cord NADPHd positivity. Morphologicallyand functionally heterogeneous neurons of the gray mattersurrounding the central canal are the target of severalafferent inputs mediated by different transmitters such assubstance P, a peptide occurring in small-diameter dorsalroot fibers (Hokfelt et al., 1975; Jessell et al., 1978), alongwith axonal profiles and cell bodies (Barber et al., 1979;Gibson et al., 1981; Tessler et al., 1981) adjacent to thecentral canal and containing serotonin (Dahlstrom andFuxe, 1965; Oliveras et al., 1977; Steinbusch, 1981; La-Motte et al., 1982) and the endogenous opiate enkephalin(Gibson et al., 1981; Glazer and Basbaum, 1989). Althoughmany lamina X NADPHd-exhibiting neurons found in ourstudy closely resemble those receiving primary afferentinput that converges from a wide range of somatic andvisceral structures (Honda, 1985), it is at present unknownwhether they belong exclusively to NO-producing neuronsor participate, at least in part, in the formation of shortlocal circuits. However, it has been repeatedly shown thatat least some pericentral neurons, perhaps cluster cells(Barber et al., 1984; Borges and Iversen, 1986), participatein the long-projection spinothalamic, spinoreticular, spino-mesencephalic, and spinohypothalamic tracts (Kevetter etal., 1982; Mantyh, 1982; Liu, 1983; Burstein et al., 1990;Menetrey and de Pommery, 1991; Menetrey et al., 1992;Lee et al., 1993). The proportion of NADPHd-exhibitingneurons to the peptidergic neurons that form minor butdistinct subgroups in the region surrounding the centralcanal and containing bombesin-, enkephalin-, cholecystoki-nin-, and somatostatin-immunoreactive ascending tractcells found at the origin of the spinothalamic, spinomesen-cephalic, spinoreticular, and spinosolitary tracts (Leah etal., 1988) has not yet been established. Moreover, thecaudal spinal cord including lamina X and the SPN arerich in both calbindin-D28k-containing and projectioncells and, with regard to the former, they largely outnum-ber those previously reported for several neuropeptides(Leah et al., 1988; Menetrey et al., 1992).
Some findings suggest that certain species-dependentdifferences exist in the staining of the neurons in the SPN.Although the IML in the thoracic and rostral lumbarsegments exhibited positive NADPHd staining, whereasthe neurons of the SPN in the cat were not well stained andonly a few scattered cells in the region of the SPN wereNADPHd positive (Vizzard et al., 1994a), the presentfinding confirmed that the neurons of the SPN in therabbit and dog (Marsala et al., 1998) including smallinterneurons were NADPHd positive (Fig. 14A–C). How-ever, the analysis of transverse and horizontal sections ofS1–S3 segments that were processed for NADPHd histo-chemistry failed to confirm the existence of the dorsal andlateral components or of the interneuronal (‘‘gap’’) regionof the SPN that was shown by the horseradish peroxidasetracing technique (Nadelhaft et al., 1980; Morgan et al.,1981). Moreover, longitudinally oriented NADPHd-exhibit-ing processes, similar to those in the IML, were foundconnecting small groups of SPN neurons (Fig. 14B,C). Theproportion of preganglionic NADPHd-exhibiting neuronsand interneurons of the SPN is not clear, and both cellpopulations of the SPN may enhance NADPHd activityafter dorsoventral L6–S1 rhizotomy (Vizzard et al., 1993b)or after proximal colon distention (Martınez et al., 1998).
Apart from the superficial dorsal horn and pericentralregion NADPHd staining extending all along the rostrocau-dal axis and containing somatic, fiberlike, and punctatenonsomatic NADPHd positivity, there were at least fournuclei identified in the spinal cord of the rabbit, i.e.,nucleus phrenicus, nucleus dorsalis, Onuf ’s nucleus ornucleus Y (Onuf, 1901; Romanes, 1951), and dorsal graycommissure containing a high number of intensely stainedNADPHd-exhibiting puncta (Fig. 16).
As for the phrenic nucleus containing middle-sizedphrenic motoneurons clearly identified in the ventral hornof C4–C5 segments but unresponsive to NADPHd stain-ing, the explanation of the punctate NADPHd positivitymay have some relation to a monosynaptic transmission ofthe respiratory drive to phrenic motoneurons from therostroventral excitatory respiratory bulbospinal neuronsor from inhibitory acting neurons of the Botzinger complex(Cohen et al., 1974). Terminals from these descendingaxons form presumptive synaptic contacts within longitu-dinal and radial dendrite bundles and on the cell somata ofphrenic motoneurons (Ellenberger and Feldman, 1988).
The source of NADPHd-exhibiting puncta in the dorsalnucleus or the column of Clarke extending through seg-ments Th2–L2 is as yet unclear. It is not known if theprimary afferents exhibit some NADPHd activity. Theseprimary afferents originate from tendon organs, musclespindles, pressure receptors, and touch and pressure recep-tors of hairy skin (Lundberg and Oscarsson, 1960), whichare distributed in a regular segmental somatotopical pat-tern in the dorsal nucleus (Szentagothai, 1961), the fibersfrom each segment occupying an area that winds spirallyalong the column (Szentagothai and Albert, 1955). More-over, neurons situated in the gray matter of the cord,mainly those in the mediobasal part of the dorsal horn,appear to give off axons to the dorsal nucleus (Szen-tagothai, 1961). Among them, as confirmed in the presentstudy, are larger, intensely stained bipolar NADPHd-exhibiting neurons whose terminal arbors may penetrateinto the dorsal nucleus and add to its punctate NADPHdstaining.
Dense punctate nonsomatic NADPHd positivity wasfound in segments S1–S3 in a position analogous to that ofnucleus Y (Romanes, 1951), which is homologous to thehuman nucleus Y or Onuf (Onuf, 1901), the nucleus
Fig. 14. Photomicrographs depicting the appearance and locationof the sacral parasympathetic nucleus (A–C) and intense nicotinamideadenine dinucleotide phosphate diaphorase (NADPHd) staining inLissauer’s tract (D,E). A: Triangular and spindle-shaped, NADPHd-exhibiting neurons (asterisks) of the sacral parasympathetic nucleus(SPN) are seen just below the lateral collateral pathway (LCP-arrow)of Lissauer’s tract (LT), close to the ventrolateral margin of the dorsalhorn (DH). S1, transverse section through segment S1; V, longitudi-nally cut vessel. B: Small groups (asterisks) of NADPHd-exhibitingneurons of the sacral parasympathetic nucleus (SPN). S1, horizontalsection through segment S1; LC, lateral column. C: High powerphotomicrograph of dendritic bundles (arrow) located among NADPHd-exhibiting neurons of the sacral parasympathetic nucleus (SPN) closeto the lateral column (LC). S1, horizontal section through segment S1.D: The occurrence of irregularly shaped patches of NADPHd positivityin Lissauer’s tract (LT-arrow) close to the dorsolateral margin of thedorsal horn (DH) and punctate intraaxonic NADPHd staining (aster-isk) seen in some axons in Lissauer’s tract. S1, transverse sectionthrough segment S1. E: High power photomicrograph of punctate,nonsomatic NADPHd staining in Lissauer’s tract (LT-arrows). Scalebars 5 100 µm in A–D, 25 µm in E.
278 J. MARSALA ET AL.
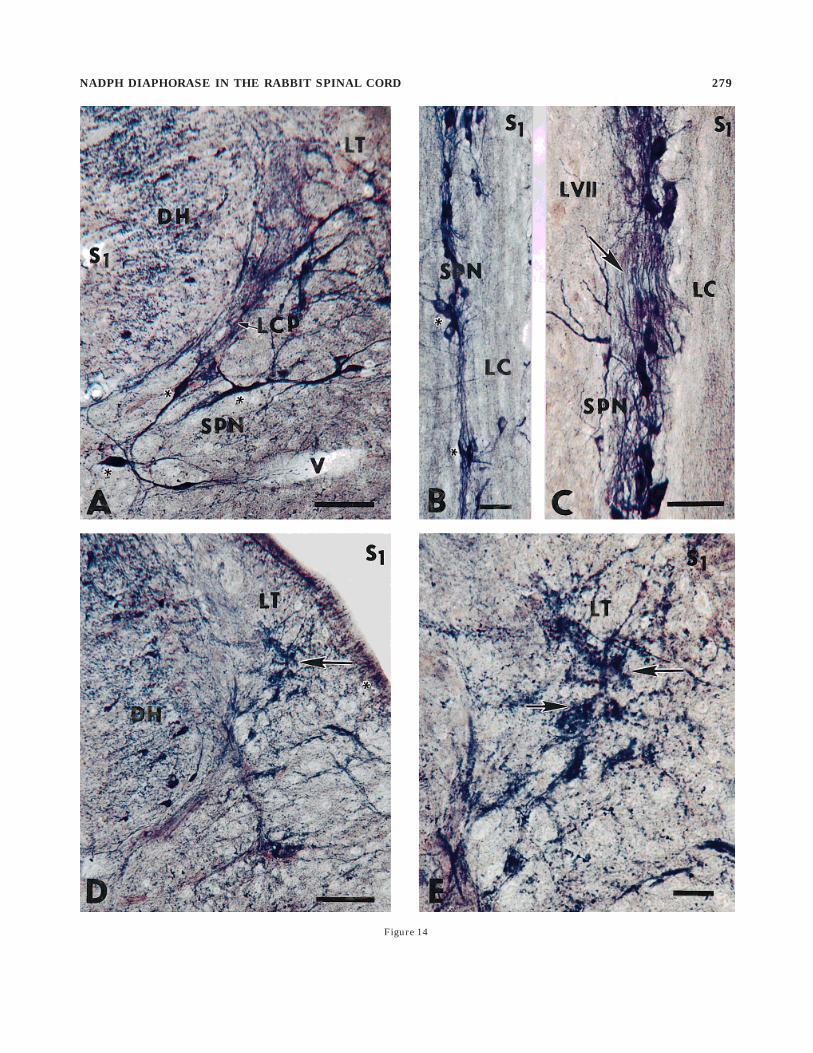
Figure 14
NADPH DIAPHORASE IN THE RABBIT SPINAL CORD 279
bulbocavernosus of the rat (Burnett et al., 1995), or ‘‘lacolonne en torsade’’ of the dog (Laruelle, 1937). Althoughthere is no prior evidence, apart from brief hints (Burnettet al. 1995), that such an accumulation of punctateNADPHd staining may exist in the Onuf ’s nucleus, theoccurrence of many positive puncta was confirmed in ourstudy on horizontal and parasagittal sections throughsegments S1–S3. Analyzing afferent inputs to the Onuf ’snucleus and its neuronal composition may, at least in part,elucidate the source of punctate NADPHd staining. Onuf ’snucleus of the rabbit described in our study is separatedinto dorsomedial and ventrolateral divisions similarly tothose detected in the cat (Thor et al., 1989), thus contrast-ing with the description in the rat, where two separatenuclei, i.e., dorsomedial and dorsolateral, have been identi-fied (Schroder, 1980; McKenna and Nadelhaft, 1986).Multipolar and stellate cells in Onuf ’s nucleus exhibitedone longitudinal dendrite and three groups of transversedendrites (Thor et al., 1989), which closely resembled theone longitudinal and three transverse dendritic bundlesfound in the phrenic nucleus (Dekker et al., 1973; Cam-eron et al., 1983). Tract tracing studies showed thatspecific projections to the Onuf ’s nucleus are derived fromthree different areas: (1) the ipsilateral paraventricularhypothalamic nucleus, (2) the ipsilateral caudal pontinelateral reticular formation, and (3) the contralateral cau-dal nucleus retroambiguus (Holstege and Tan, 1987).Moreover, intraspinal axon collaterals of expiratory neu-rons in the caudal nucleus retroambiguus, i.e., expiratoryneurons projecting monosynaptically to the phrenicnucleus, descend to the lower lumbar (L6–L7) and thesacral (S1–S3) segments and terminate in the sacral gray
matter and within the Onuf ’s nucleus (Sasaki et al., 1994).Thus, it is highly probable that the punctate nonsomaticNADPHd staining detected in the neuropil of the phrenicand Onuf ’s nucleus may have a common origin repre-sented by expiratory neurons of the caudal nucleus retro-ambiguus, a view supported by recent morphological andelectrophysiological studies (Sasaki et al., 1991; Sasaki,1994; Miller et al., 1995).
The upper two-thirds of the dorsal gray commissure insegments S1–S3 contained large amounts of randomlydistributed NADPHd-exhibiting puncta. However, the ex-tent of NADPHd staining declined abruptly just above andbelow the sacral segments. Because the distribution ofNADPHd staining in the dorsal gray commissure wasquite identical with the course and termination of themedial collateral pathway containing some S1–S3 dorsalroot afferents and forming a large terminal field in thedorsal gray commissure (Morgan et al., 1981), terminals ofthis pathway may contribute to NADPHd staining.
In conclusion, NADPHd staining by using transverse,horizontal, and sagittal sections from cervical, thoracic,and lumbosacral segments of rabbits has shown a morpho-logically heterogeneous appearance of NADPHd-exhibit-ing neurons located in the superficial and deep dorsalhorn, in the pericentral region all along the rostrocaudalaxis of the spinal cord, and in the dorsal gray commissurein segments S1–S3. A homogeneous population ofNADPHd-labeled neurons was located in the IML andSPN, with a small population of interneurons in the SPN.In the pericentral region (lamina X), a close association ofNADPHd-exhibiting somata and fibers and mostly longitu-dinally oriented blood vessels was detected. These findings
Fig. 15. High power photomicrographs (A) showing nicotinamideadenine dinucleotide phosphate diaphorase (NADPHd)-exhibiting fi-bers (arrowheads) at the border zone between the intermediate zone(LVII) and lateral funiculus (LF) in segment S2, and dense accumula-
tion (B) of punctate nonsomatic positivity (arrowheads) close tonucleus Y (NY) in segment S2 can be seen. Isolated NADPHd-exhibiting fiber (arrow) is seen in the ventral horn (LIX). Scale bars 550 µm in A,B.
280 J. MARSALA ET AL.
raise the possibility that NO, synthetized by small subepen-dymally located bipolar neurons and large, multipolarNADPHd-exhibiting neurons located in lamina X, may beinvolved in the control of spinal cord blood flow.
LITERATURE CITED
Aimi Y, Fujimura M, Vincent SR, Kimura H. 1991. Localization of NADPH-diaphorase containing neurons in sensory ganglia of the rat. J CompNeurol 306:382–392.
Anderson CR. 1992. NADPH diaphorase-positive neurons in the rat spinalcord include a subpopulation of autonomic preganglionic neurons.Neurosci Lett 139:280–284.
Barber RP, Vaughn JE, Slemmon JR, Salvaterra PM, Roberts E, Lee-man SE. 1979. The origin, distribution and synaptic relationshipsof substance P axons in the rat spinal cord. J Comp Neurol 184:331–352.
Barber RP, Phelps PE, Houser CR, Crawford GD, Salvaterra PM, VaughnJE. 1984. The morphology and distribution of neurons containingcholine acetyltransferase in the adult rat spinal cord: an immunocyto-chemical study. J Comp Neurol 229:329–346.
Fig. 16. Schematic diagram of nicotinamide adenine dinucleotidephosphate diaphorase (NADPHd) activity in the spinal cord of therabbit, taken from the caudal medulla–cervical spinal cord transitionand from cervical, thoracic, lumbar, and sacral spinal cord segments.Neuronal cell bodies are indicated as filled circles on both sides of eachfigure. Each filled circle represents one NADPHd-exhibiting neuron.
Regions of dense neuropil NADPHd staining including small positivecells are shaded on both sides. Dotted nuclei (NP, nucleus phrenicus;ND, nucleus dorsalis; NY, nucleus Y; DGC, dorsal gray commissure)contain large amounts of punctate nonsomatic positivity. IML, interme-diolateral cell column; LT, Lissauer’s tract; LI–III, X, Rexed’s laminae;SPN, sacral parasympathetic nucleus.
NADPH DIAPHORASE IN THE RABBIT SPINAL CORD 281

Bennett GJ, Ruda MA, Gobel S, Dubner R. 1982. Enkephalin immunoreac-tive stalked cells and lamina IIb islet cells in cat substantia gelatinosa.Brain Res 240:162–166.
Blot S, Arnal JF, Xu Y, Gray F, Michel JB. 1994. Spinal cord infarcts duringlong-term inhibition of nitric oxide synthase in rats. Stroke 25:1666–1673.
Blottner D, Baumgarten H. 1992. Nitric oxide synthase (NOS) containingsympathoadrenal cholinergic neurons of the rat IML-cell column:evidence from histochemistry, immunohistochemistry and retrogradelabeling. J Comp Neurol 316:45–56.
Borges LF, Iversen SD. 1986. Topography of choline acetyltransferaseimmunoreactive neurons and fibers in the rat spinal cord. Brain Res362:140–148.
Bredt DS, Snyder SH. 1992. Nitric oxide, a novel neuronal messenger.Neuron 8:3–11.
Bredt DS, Hwang PM, Snyder SH. 1990. Localization of nitric oxidesynthase indicating a neuronal role for nitric oxide. Nature 347:768–770.
Brodal A. 1969. Neurological anatomy. New York: Oxford University Press.Brown AG. 1982. The dorsal horn of the spinal cord. Q J Exp Physiol
67:193–212.Bruning G. 1992. Localization of NADPH diaphorase, a histochemical
marker for nitric oxide synthase, in the mouse spinal cord. ActaHistochem 93:397–401.
Burnett AL, Saito S, Maguire MP, Yamaguchi H, Chang TSK, Hanley DF.1995. Localization of nitric oxide synthase in spinal nuclei innervatingpelvic ganglia. J Urol 153:212–217.
Burstein R, Cliffer KD, Giesler GJ. 1990. Cells of origin of the spinohypotha-lamic tract in the rat. J Comp Neurol 291:329–344.
Cameron WE, Averill DB, Berger AJ. 1983. Morphology of cat phrenicmotoneurons as revealed by intracellular injection of horseradishperoxidase. J Comp Neurol 219:70–80.
Cliffer KD, Urca G, Elde RP, Giesler GJ Jr. 1988. Studies of peptidergicinput to the lateral spinal nucleus. Brain Res 460:356–360.
Cohen MI, Piercey MF, Gootman PM, Wolotsky P. 1974. Synaptic connec-tions between medullary inspiratory neurons and phrenic motoneuronsas revealed by cross-correlation. Brain Res 81:319–324.
Dahlstrom A, Fuxe K. 1965. Evidence for the existence of monoaminecontaining neurons in the central nervous system II. Experimentallyinduced changes in the intraneuronal amine levels. Acta Physiol Scand374(Suppl):1–36.
de Groat WC, Nadelhaft I, Morgan C, Schauble T. 1978. Horseradishperoxidase tracing of visceral efferent and primary afferent pathways inthe cat’s sacral spinal cord using benzidine processing. Neurosci Lett10:103–108.
Dekker JJ, Lawrence DG, Kuypers HGJM. 1973. The location of longitudi-nally running dendrites in the ventral horn of the cat spinal cord. BrainRes 51:319–325.
Dubner R, Bennett GJ. 1983. Spinal and trigeminal mechanisms ofnociception. Annu Rev Neurosci 6:381–418.
Dun NJ, Dun SL, Wu SY, Forstermann U, Schmidt HHHW, Tseng LF. 1993.Nitric oxide synthase immunoreactivity in the rat, mouse, cat andsquirrel monkey spinal cord. Neuroscience 54:845–857.
Earle KM. 1952. The tract of Lissauer and its possible relation to the painpathway. J Comp Neurol 96:93–111.
Egberongbe YI, Gentleman SM, Falkai P, Bogerts B, Polak JM, RobertsGW. 1994. The distribution of nitric oxide synthase immunoreactivity inthe human brain. Neuroscience 59:561–578.
Ellenberger HH, Feldman JL. 1988. Monosynaptic transmission of respira-tory drive to phrenic motoneurons from brainstem bulbospinal neuronsin rats. J Comp Neurol 269:47–57.
Faraci FM, Brian JE. 1994. Nitric oxide and the cerebral circulation. Stroke25:692–703.
Fuxe K, Tinner B, Bjelke B, Agnati LF, Verhofstad A, Steinbusch HGW,Goldstein M, Hersh L, Kalia M. 1990. Monoaminergic and peptidergicinnervation of the intermedio-lateral horn of the spinal cord. II.Relationship to preganglionic sympathetic neurons. Eur J Neurosci2:451–460.
Gally JA, Montague PR, Reek GN, Edelman GM. 1990. The NO hypothesis:Possible effects of a short-lived, rapidly diffusible signal in the develop-ment and function of the nervous system. Proc Natl Acad Sci USA87:3547–3551.
Gibson SJ, Polak JM, Bloom SR, Wall PD. 1981. The distribution of ninepeptides in rat spinal cord with special emphasis on the substantia
gelatinosa and on the area around the central canal (lamina X). J CompNeurol 201:65–79.
Glazer EJ, Basbaum AI. 1989. Leucine enkephalin localization in andaxoplasmic transport by sacral parasympathetic preganglionic neu-rons. Science 208:1479–1480.
Gobel S. 1975a. Golgi studies of the substantia gelatinosa neurons in thespinal trigeminal nucleus. J Comp Neurol 162:397–415.
Gobel S. 1975b. Neurons with two axons in the substantia gelatinosa layerof the spinal trigeminal nucleus of the adult cat. Brain Res 88:333–338.
Gobel S. 1978. Golgi studies of the neurons in layer II of the dorsal horn ofthe medulla (trigeminal nucleus caudalis). J Comp Neurol 180:395–414.
Gobel S. 1979. Neural circuitry in the substantia gelatinosa of Rolando:anatomical insights. In: Bonica JJ, Leibeskind JC, Albe-Fessard DG,editors. Advances in pain research and therapy. New York: Raven. pp175–195.
Gobel S, Falls WM, Bennett GJ, Abdelmoumene M, Hayashi H, HumphreyE. 1980. An EM analysis of the synaptic connections of horseradishperoxidase-filled stalked cells and islet cells in the substantia gelati-nosa of the adult cat spinal cord. J Comp Neurol 194:781–807.
Grant G, Rexed B. 1958. Dorsal spinal root afferents to Clarke’s column.Brain 81:567–576.
Gwyn DG, Waldron HA. 1969. Observations on the morphology of a nucleusin the dorsolateral funiculus of the spinal cord of the guinea-pig, rabbit,ferret and cat. J Comp Neurol 136:233–236.
Herdegen T, Rudiger S, Mayer B, Bravo R, Zimmermann M. 1994.Expression of nitric oxide synthase and colocalization with Jun, Fos andKrox transcription factors in spinal cord neurons following noxiousstimulation of the rat hind paw. Mol Brain Res 22:245–258.
Hokfelt T, Kellerth JO, Nilsson G, Pernow B. 1975. Experimental immuno-histochemical studies on the localization and distribution of substanceP in cat primary sensory neurons. Brain Res 100:235–252.
Holstege G, Tan J. 1987. Supraspinal control of motoneurons innervatingthe striated muscles of the pelvic floor including urethral and analsphincters in the cat. Brain 110:1323–1344.
Honda CN. 1985. Visceral and somatic afferent convergence into neuronsnear the central canal in the sacral spinal cord of the cat. J Neuro-physiol 53:1059–1078.
Hope BT, Michael GJ, Knigge KM, Vincent SR. 1991. Neuronal NADPHdiaphorase is a nitric oxide synthase. Proc Natl Acad Sci USA 88:2812–2814.
Iadecola C. 1985. Bright and dark sides of nitric oxide in ischemic injury.Trends Neurosci 20:132–139.
Iadecola C. 1993. Regulation of the cerebral microcirculation during neuralactivity: is nitric oxide the missing link? Trends Neurosci 16:206–214.
Janig W, McLachlan EM. 1987. Organization of lumbar spinal outflow todistal colon and pelvic organs. Physiol Rev 67:1332–1404.
Jankowska E, Lindstrom S. 1971. Morphological identification of Renshawcells. Acta Physiol Scand 81:428–430.
Jessell TM, Iversen LL, Cuello AC. 1978. Capsaicin-induced depletion ofsubstance P from primary sensory neurones. Brain Res 152:183–188.
Kevetter GA, Haber LH, Yezierski RP, Chung JM, Martin RF, Willis WD.1982. Cells of origin of the spinoreticular tract in the monkey. J CompNeurol 207:61–74.
Kowall NW, Mueller MO. 1988. Morphology and distribution of nicotin-amide adenine dinucleotide phosphate (reduced form) diaphorase reac-tive neurons in human brainstem. Neuroscience 26:645–654.
Kowall NW, Beal MF, Ferrante RJ, Martin JB. 1985. Topography ofnicotinamide adenine dinucleotide phosphate-diaphorase staining neu-rons in rat striatum. Neurosci Lett 59:61–66.
Kowall NW, Ferrante RJ, Beal MF, Richardson EP Jr, Sofroniew MV, CuelloAC, Martin JB. 1987. Neuropeptide Y, somatostatin and reducednicotinamide adenine dinucleotide phosphate diaphorase in the humanstriatum:a combined immunocytochemical and enzyme histochemicalstudy. Neuroscience 20:817–828.
LaMotte CC, Johus DR, de Lanerolle NC. 1982. Immunohistochemicalevidence of indolamine neurons in monkey spinal cord. J Comp Neurol206:359–370.
Laruelle L. 1937. La structure de la moelle epiniere en coupes longitudina-les. Rev Neurol 67:696–725.
Leah J, Menetrey D, de Pommery J. 1988. Neuropeptides in long ascendingspinal tract cells in the rat: evidence for parallel processing of ascendinginformation. Neuroscience 24:195–207.
282 J. MARSALA ET AL.

Lee JH, Price RH, Williams FG, Mayer B, Beitz AJ. 1993. Nitric oxidesynthase is found in some spinothalamic neurons and in neuronalprocesses that appose spinal neurons that express Fos induced bynoxious stimulation. Brain Res 608:324–333.
Lima D, Coimbra A. 1989. Morphological types of spinomesencephalicneurons in the marginal zone (lamina I) of the rat spinal cord, as shownafter retrograde labelling with cholera toxin subunit B. J Comp Neurol279:327–339.
Liu RPC. 1983. Laminar origins of spinal projection neurons to theperiaqueductal gray of the cat. Brain Res 264:118–122.
Lundberg A, Oscarsson O. 1960. Functional organization of the dorsalspino-cerebellar tract in the cat. VII. Identification of units by anti-dromic activation from the cerebellar cortex with recognition of fivefunctional subdivisions. Acta Physiol Scand 50:356–374.
Maixner W, Dubner R, Bushnell MC, Kenshalo DR Jr, Oliveras JL. 1986.Wide-dynamic-range dorsal horn neurons participate in the encodingprocess by which monkeys perceive the intensity of noxious heatstimuli. Brain Res 374:385–388.
Mantyh PW. 1982. The ascending projections to the midbrain periaqueduc-tal gray of the primate. J Comp Neurol 211:50–64.
Marsala M, Danielisova V, Chavko M, Hornakova A, Marsala J. 1989.Improvement of energy state and basic modifications of neuropathologi-cal damage in rabbits as a result of graded postischemic spinal cordreoxygenation. Exp Neurol 105:93–103.
Marsala J, Sulla I, Santa M, Marsala M, Zacharias L, Radonak J. 1991.Mapping of the canine lumbosacral spinal cord neurons by Nautamethod at the end of the early phase of paraplegia induced by ischemiaand reperfusion. Neuroscience 45:479–494.
Marsala J, Kluchova D, Marsala M. 1997. Spinal cord gray matter layersrich in NADPH diaphorase positive neurons are refractory to ischemia-reperfusion-induced injury: a histochemical and silver impregnationstudy in rabbit. Exp Neurol 145:165–179.
Marsala J, Vanicky I, Marsala M, Jalc P, Orendacova J, Taira Y. 1998.Reduced nicotinamide adenine dinucleotide phosphate diaphorase inthe spinal cord of dogs. Neuroscience 85:847–862.
Martınez V, Wang L, Mayer E, Tache Y. 1998. Proximal colon distentionincreases Fos expression in the lumbosacral spinal cord and activatessacral parasympathetic NADPHd-positive neurons in rats. J CompNeurol 390:311–321.
McKenna KE, Nadelhaft I. 1986. The organization of the pudendal nerve inthe male and female rat. J Comp Neurol 248:532–549.
Mendell LM. 1966. Physiological properties of unmyelinated fiber projec-tion to the spinal cord. Exp Neurol 16:316–332.
Menetrey D, de Pommery J. 1991. Origins of spinal ascending pathwaysthat reach central areas involved in visceroception and visceronocicep-tion in the rat. Eur J Neurosci 3:249–259.
Menetrey DJ, de Pommery J, Thomasset M, Baimbridge KG. 1992.Calbindin-D28K (CaBP28k)-like immunoreactivity in ascending projec-tions. II. Spinal projections to brain stem and mesencephalic areas. EurJ Neurosci 4:70–76.
Miller AD, Nonaka S, Siniaia MS, Jakus J. 1995. Multifunctional ventralrespiratory group: bulbospinal expiratory neurons play a role in puden-dal discharge during vomiting. J Auton Nerv Syst 54:253–260.
Mizukawa K, McGeer PL, Vincent SR, McGeer ER. 1989. Distribution ofreduced-nicotinamide-adenine-dinucleotide-phosphate diaphorase-positive cells and fibers in the cat central nervous system. J CompNeurol 279:281–311.
Molander C, Xu Q, Grant G. 1984. The cytoarchitectonic organization of thespinal cord in the rat. I. The lower thoracic and lumbosacral cord. JComp Neurol 289:375–385.
Morgan C, Nadelhaft I, de Groat WC. 1981. The distribution of visceralprimary afferents from the pelvic nerve to Lissauer’s tract and thespinal gray matter and its relationship to the sacral parasympatheticnucleus. J Comp Neurol 201:415–440.
Morgan C, de Groat WC, Nadelhaft I. 1986. The spinal distribution ofsympathetic preganglionic and visceral primary afferent neurons thatsend axons into the hypogastric nerves of the cat. J Comp Neurol243:23–40.
Nadelhaft I, de Groat WC, Morgan C. 1980. Location and morphology ofparasympathetic preganglionic neurons in the sacral spinal cord of thecat revealed by retrograde axonal transport of horseradish peroxidase.J Comp Neurol 193:265–281.
Oliveras JL, Bourgoin S, Hery F, Besson JM, Hamon M. 1977. Thetopographical distribution of serotoninergic terminals in the spinal cord
of the cat: biochemical mapping by the combined use of microdissectionand microassay procedures. Brain Res 138:393–406.
Onuf B. 1901. On the arrangement and function of the cell groups of thesacral region of the spinal cord in man. Arch Neurol Psychopathol3:387–412.
Price DD. 1988. Psychological and neural mechamisms of pain. New York:Raven.
Price DD, Mayer DJ. 1976. Neurophysiological characterization of theanterolateral quadrant neurons subserving pain in M. mulatta. Pain1:59–72.
Price DD, Hayes RL, Ruda MA, Dubner R. 1978. Spatial and temporaltransformations of input to spinothalamic tract neurons and theirrelation to somatic sensation. J Neurophysiol 41:933–947.
Price DD, Hayashi H, Dubner R, Ruda MA. 1979. Functional relationshipbetween neurons of marginal and substantia gelatinosa layers ofprimate dorsal horn. J Neurophysiol 42:1590–1608.
Rexed B. 1954. A cytoarchitectonic atlas of the spinal cord in the cat. JComp Neurol 100:297–379.
Romanes GJ. 1951. The motor cell columns of the lumbosacral spinal cord ofthe cat. J Comp Neurol 94:313–363.
Ross CA, Bredt D, Snyder SH. 1990. Messenger molecules in the cerebel-lum. Trends Neurosci 13:216–222.
Rothe F, Canzler U, Wolf G. 1998. Subcellular localization of the neuronalisoform of nitric oxide synthase in the rat brain: a critical evaluation.Neuroscience 83:259–269.
Ruda MA, Bennett GJ, Dubner R. 1986. Neurochemistry and neuralcircuitry in the dorsal horn. Prog Brain Res 66:219–268.
Saito S, Kidd GJ, Trapp BD, Dawson TM, Bredt DS, Wilson DA, Trayst-mann RJ, Snyder SH, Hanley DF. 1994. Rat spinal cord neurons containnitric oxide synthase. Neuroscience 59:447–452.
Sasaki M. 1994. Morphological analysis of external urethral and externalanal sphincter motoneurons of cat. J Comp Neurol 349:269–287.
Sasaki SI, Uchino H, Imagawa M, Miyake T, Uchino Y. 1991. Lower lumbarbranching of caudal medullary expiratory neurons of the cat. Brain Res553:159–162.
Sasaki SI, Uchino H, Uchino Y. 1994. Axon branching of medullaryexpiratory neurons in the lumbar and the sacral spinal cord of the cat.Brain Res 648:229–238.
Scheibel ME, Scheibel AB. 1968. Terminal axon patterns in cat spinal cord.II: The dorsal horn. Brain Res 9:32–58.
Scherer-Singler U, Vincent SR, Kimura H, McGeer EG. 1983. Demonstra-tion of a unique population of neurons with NADPH diaphorasehistochemistry. J Neurosci Methods 8:229–234.
Schroder HD. 1980. Organization of the motoneurons innervating thepelvic muscles of the male rat. J Comp Neurol 192:567–587.
Steinbusch HWM. 1981. Distribution of serotonin-immunoreactivity in thecentral nervous system of the rat. Cell bodies and terminals. Neurosci-ence 6:557–618.
Szentagothai J. 1961. Somatotopic arrangement of synapses of primarysensory neurones in Clarke’s column. Acta Morphol Acad Sci Hung10:307–311.
Szentagothai J, Albert A. 1955. The synaptology of Clarke’s column. ActaMorphol Acad Sci Hung 5:43–51.
Tessler A, Himes BT, Artymyshyn R, Murray M, Goldberger ME. 1981.Spinal neurons mediate return of substance P following deafferentationof cat spinal cord. Brain Res 230:263–281.
Thomas E, Pearse AGE. 1964. The solitary active cells. Histochemicaldemonstration of damage-resistant nerve cells with a TPN-diaphorasereaction. Acta Neuropathol 3:238–249.
Thomas RC, Wilson VJ. 1965. Precise localization of Renshaw cells with anew marking technique. Nature 206:211–213.
Thor KB, Morgan C, Nadelhaft I, Houston M, de Groat WC. 1989.Organization of afferent and efferent pathways in the pudendal nerve ofthe female cat. J Comp Neurol 288:263–279.
Tolivia J, Tolivia D. 1985. A new technique for differential and simultaneousstaining of nerve cells and fibers. J Neurosci Methods 13:305–311.
Valtschanoff JG, Weinberg RJ, Rustioni A. 1992. NADPH diaphorase in thespinal cord of rats. J Comp Neurol 321:209–222.
Vincent SR. 1994. Nitric oxide: a radical neurotransmitter in the centralnervous system. Prog Neurobiol 42:129–160.
Vincent SR, Kimura H. 1992. Histochemical mapping of nitric oxidesynthase in the rat brain. Neuroscience 46:755–784.
NADPH DIAPHORASE IN THE RABBIT SPINAL CORD 283

Vizzard MA, Erdman SL, de Groat WC. 1993a. Localization of NADPH-diaphorase in pelvic afferent and efferent pathways of the rat. NeurosciLett 152:72–76.
Vizzard MA, Erdman SL, de Groat WC. 1993b. The effect of rhizotomy onNADPH diaphorase staining in the lumbar spinal cord of the rat. BrainRes 607:349–353.
Vizzard MA, Erdman SL, Erickson VL, Stewart RJ, Roppolo JR, de GroatWC. 1994a. Localization of NADPH diaphorase in the lumbosacralspinal cord and dorsal root ganglia of the cat. J Comp Neurol 339:62–75.
Vizzard MA, Erdman SL, Roppolo JR, Forstermann U, de Groat WC. 1994b.Differential localization of neuronal nitric oxide synthase immunoreac-tivity and NADPH-diaphorase activity in the cat spinal cord. CellTissue Res 278:299–309.
Vizzard MA, Erdman SL, de Groat WC. 1995. Increased expresion ofneuronal nitric oxide synthase (NOS) in visceral neurons after nerveinjury. J Neurosci 15:4033–4045.
Wetts R, Vaughn JE. 1994. Choline acetyltransferase and NADPH diapho-rase are co-expressed in rat spinal cord neurons. Neuroscience 63:1117–1124.
Willis WD. 1971. The case for the Renshaw cell. Brain Behav Evol 4:5–52.Yezierski RP, Liu S, Ruenes GL, Busto R, Dietrich WD. 1996. Neuronal
damage following intraspinal injection of a nitric oxide synthaseinhibitor in the rat. J Cereb Blood Flow Metab 16:996–1004.
Yoss RE. 1953. Studies of the spinal cord. Part II. Topographic localizationwithin the ventral spino-cerebellar tract in the macaque. J CompNeurol 99:613–638.
284 J. MARSALA ET AL.