long-distance movement factor: a transport … movement factor: a transport function of ... shuttles...
TRANSCRIPT

The Plant Cell, Vol. 7, 549-559, May 1995 O 1995 American Society of Plant Physiologists
Long-Distance Movement Factor: A Transport Function of the Potyvirus Helper Component Proteinase
Stephen Cronin, Jeanmarie Verchot, Ruth Haldeman-Cahill, Mary C. Schaad, and James C. Carrington’ Department of Biology, Texas A & M University, College Station, Texas 77843
Transport of viruses from cell to cell in plants typically involves one or more viral proteins that supply dedicated move- ment functions. Transport from leaf to leaf through phloem, or long-distance transport, is a poorly understood process with requirements differing from those of cell-to-cell movement. Through genetic analysis of tobacco etch virus (TER potyvirus group), a nove1 long-distance movement factor was identified that facilitates vascular-associated movement in tobacco. A mutation in the central region of the helper component proteinase (HC-Pro), a TEV-encoded protein with previously described activities in aphid-mediated transmission and polyprotein processing, inactivated long-distance movement. This mutant virus exhibited only minor defects in genome amplification and cell-to-cell movement functions. In situ histochemical analysis revealed that the mutant was capable of infecting mesophyll, bundle sheath, and phloem cells within inoculated leaves, suggesting that the long-distance movement block was associated with entry into or exit from sieve elements. The long-distance movement defect was specifically complemented by HC-Pro supplied in trans by a transgenic host. The data indicate that HC-Pro functions in one or more steps unique to long-distance transport.
INTRODUCTION
Spread of plant viruses from sites of initial infection to dista1 sites can be conceptualized as a multistep process. First, ini- tially infected cells must support viral replication to supply infectious material for subsequent movement. Second, the vi- rus or viral genome must move from cell to cell through intercellular connections, the plasmodesmata, within the ini- tially infected leaf. Third, the virus must move through severa1 vasculature-associated cell types and enter the sieve elements, where movement occurs passively over long distances within the same leaf and between organs. Finally, the virus must exit the sieve elements and reestablish replication and cell-to-cell movement in tissues distant from the initial infection site.
Cell-to-cell transport requires the activities of movement pro- teins (MPs) encoded by viruses (Hull, 1991; Maule, 1991; Deom et al., 1992). It has been proposed that the tobamovirus and dianthovirus MPs bind directly to their respective genomes and facilitate passage through plasmodesmata (Citovsky et al., 1990, 1992; Fujiwara et al., 1993; Giesman-Cookmeyer and Lommel, 1993; Waigmann et al., 1994). Severa1 viruses, in- cluding those in the potexvirus and hordeivirus groups, encode multiple MPs, among which the required movement activities are dispersed (Petty et al., 1990; Beck et al., 1991). For some viruses, such as comoviruses, the MPs form intercellular tub- ules through which intact virions pass (van Lent et al., 1990; Kasteel et al., 1993). The bipartite geminiviruses move between cells through the coordinated activities of two MPs, one that
To whom correspondence should be addressed.
shuttles viral DNA from the nucleus to the cytoplasm and an- other that facilitates plasmodesmal transit (Noueiry et al., 1994; Pascal et al., 1994). The potyviruses, as exemplified by tobacco etch virus (TEV), differ from each of the aforementioned models in that proteins with dedicated movement functions have not been identified. The P1 protein has been proposed extensively in the literature to function as the potyviral MP, but deletions and modifications of the P1 coding sequence have little direct effect on movement (Verchot and Carrington, 1995a, 1995b). On the other hand, the cell-to-cell movement functions have been hypothesized as being performed by the capsid protein based on the results of mutational analyses (Dolja et al., 1994, 1995).
Recent evidence suggests that long-distance transport of TEV involves viral functions distinct from those necessary for cell-to-cell movement. Different domains within the capsid pro- tein (263 amino acid residues) have been shown to play specific roles in cell-to-cell and long-distance transport (Dolja et al., 1994, 1995). The core domain, which is required for assembly of the flexuous rod-shaped virions, is absolutely required for cell-to-cell movement. In contrast, the N-terminal(29 residues) and C-terminal(l8 residues) regions, which comprise surface- oriented domains not required for assembly, are dispensable for cell-to-cell transport but necessary for long-distance move- ment. Mutants that lack either terminal domain move from cell to cell in primary infection foci but are incapable of vascular- associated movement. These data suggest that the TEV cap- sid protein interacts with unique cellular and/or viral factors during cell-to-cell and long-distance transport.

550 The Plant Cell
Here, we identify the helper component proteinase (HC-Pro) of TEV as a trans-acting factor that plays a specific role in long- distance transport. Potyvirus HC-Pro is a multifunctional pro- tein that has been characterized as a helper factor for acquisition and transmission of virus by aphids (Figure 1; reviewed by Riechmann et al., 1992; Shukla et al., 1994). The helper component domain encompasses at least the N-terminal and central regions, which also contain determinants involved in genome amplification and pathogenicity (Thornbury et al., 1990; Atreya et al., 1992; Atreya and Pirone, 1993; Dolja et al., 1993; Klein et al., 1994). The C-terminal one-third of HC-Pro comprises a cysteine-type proteinase domain that catalyzes autoproteolytic cleavage between itself and the neighboring P3 protein during maturation of the viral polyprotein (Carrington et ai., 1989a, 1989b). A model in which HC-Pro provides spe- cific movement functions associated with phloem transport of virus is proposed.
ur. A 1 kb w
sp6 P1 GUS P3 CI 6 NIa NIb CP Bg
HC-Pro (459 aa) B
Nonessential for replic. or Cys His - movement 'FRN' 'IGN' 'CCCE' 345 418
L _ _ _ _ _ _ Proteinase domain I , C N
Aphid transmission
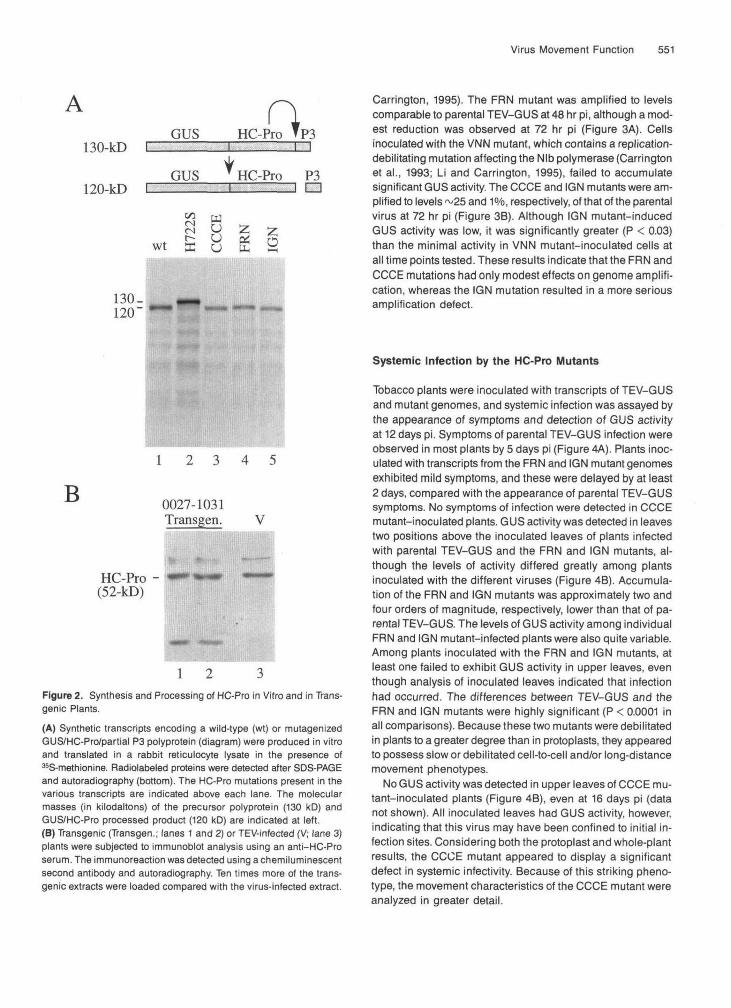
Figure 1. Diagrammatic Representation of Relevant Portions of the TEV-GUS Genome and HC-Pro.
(A) Complementary DNA representing the TEV-GUS genome was cloned downstream from a bacteriophage SP6 promoter, forming pTEV7DAN-GLH. The 5'and S'noncoding sequences and polyprotein coding region (open boxes) and GUS coding region (hatched box) are shown. The vertical lines throughout the coding region indicate se- quences coding for polyprotein cleavage sites. Proteins encoded by the TEV-GUS genome are indicated above the map. Some of the known or proposed functions of the viral proteins are as follows: P1, proteinase; HC-Pro, helper component for aphid transmission and proteinase; P3, function unknown; CI, ATPase/helicase; 6, membrane binding protein; Nla, genome-linked protein (VPg) and proteinase; Nlb, RNA-dependent RNA polymerase; CP, capsid protein. Synthetic transcripts were pro- duced using SP6 polymerase after linearization of the DNA with Bglll
(E) The HC-Pro protein is represented as a black bar. The proteinase domain within HC-Pro is shown as a thick bar; indicated are the rela- tive positions of Cys-345 and His-418 at the active site. The domain necessary for helper activity during aphid-mediated transmission is indicated near the N terminus. The positions altered by the FRN, IGN, and CCCE mutations are also indicated. C, C terminus; N, N termi- nus; aa, amino acid; replic., replication.
( W .
RESU LTS
Construction of TEV-GUS Mutants with Alterations in HC-Pro
Three different mutations, termed FRN, IGN, and CCCE, were introduced into the HC-Pro sequence of PTEV~DAN-GJH, a plasmid from which synthetic RNA transcripts representing the TEV-GUS genome are derived (Dolja et al., 1992; Carrington et al., 1993; Figure 1). Each mutation resulted in substitution of Arg-Pro-Ala for a tripeptide within the central region of HC- Pro. The tripeptides selected for substitution were among the most highly conserved sequences within the central region. The resulting viruses encoded the P-glucuronidase (GUS) reporter protein between P1 and HC-Pro. Cleavage sites rec- ognized by P1 and Nla proteinases were engineered at the sequences encoding the N and C termini of GUS, respectively, to ensure proteolytic remova1 of GUS from both P1 and HC- Pro (Dolja et ai., 1992; Carrington et al., 1993). The addition of the GUS coding sequence to the viral genome was shown to have relatively minor effects on the systemic infection capa- bilities of TEV (Dolja et al., 1992).
To determine the effects of the mutations on HC-Pro au- toproteolytic function, the sequences encoding a 130-kD GUSIHC-Prolpartial P3 polyprotein were excised from each of the parental and mutant full-length plasmids and subcloned into the vector pTL7SN.3 for in vitro transcription and transla- tion. In addition to subclones containing the three mutations in the central region of HC-Pro, a plasmid was constructed that contained a substitution of a Ser codon for the active site His- 418 codon within the proteolytic domain (termed the H722S mutation), a modification that inactivates HC-Pro processing activity (Oh and Carrington, 1989). Transcripts from each plas- mid were translated in a rabbit reticulocyte lysate system. The polyprotein containing the wild-type HC-Pro sequence under- went autoproteolysis to yield a 120-kD GUSIHC-Pro product (Figure 2A, lane l), whereas the H722S mutant 130-kD poly- protein was stable (lane 2). The polyproteins harboring the CCCE, FRN, and IGN substitutions were processed to yield 4 2 0 - k D products that comigrated with the wild-type product (Figure 2A, lanes 3 to 5), indicating that these mutations had little or no effect on HC-Pro proteolytic activity.
Replication of the HC-Pro Mutants in Protoplasts
The effects of the FRN, CCCE, and IGN mutations on genome amplification were measured. Full-length transcripts represent- ing the parental TEV-GUS and mutant genomes were used to inoculate tobacco protoplasts, and GUS activity was assayed at time intervals postinoculation (pi). GUS activity, which ac- cumulates to significant levels only in cells supporting virus replication, has proven to be an effective quantitative marker for genome amplification (Carrington et al., 1993; Dolja et al., 1994; Restrepo-Hartwig and Carrington, 1994; Li and
Virus Movement Function 551
A130-kD C
120-kD C
GUS -Pro TlHC-Pro TP3
GUS
csWt ffi
XJ
HC-Pro P3i n
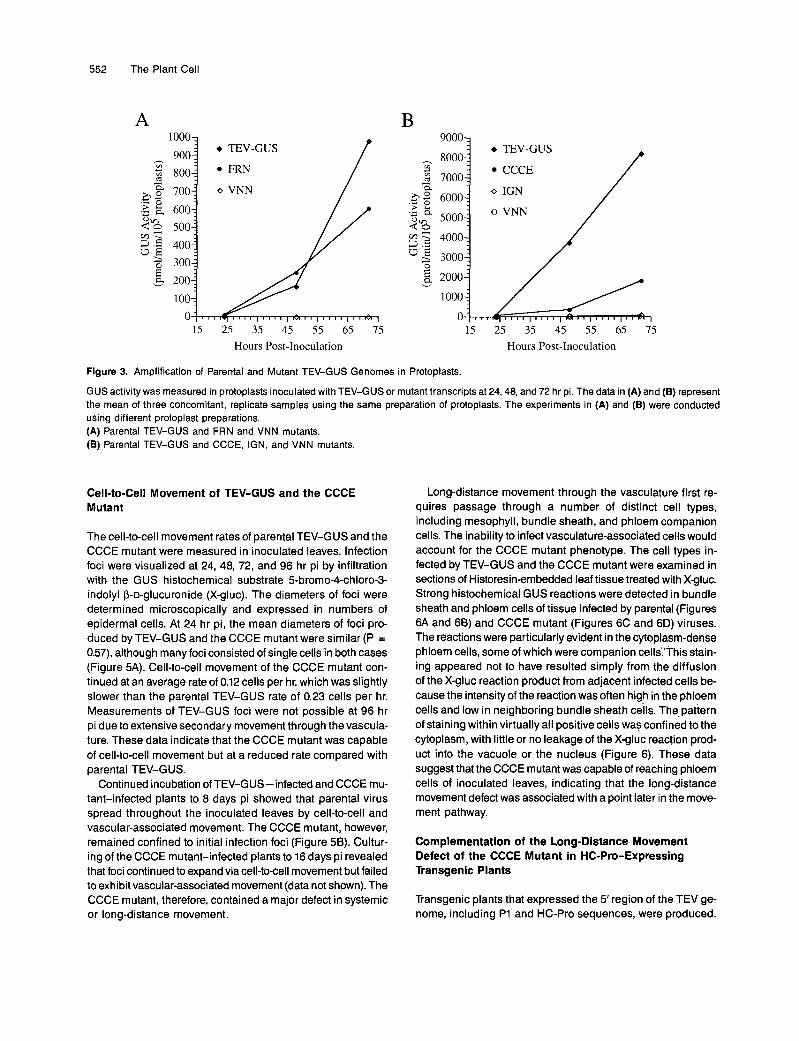
Carrington, 1995). The FRN mutant was amplified to levelscomparable to parental TEV-GUS at 48 hr pi, although a mod-est reduction was observed at 72 hr pi (Figure 3A). Cellsinoculated with the VNN mutant, which contains a replication-debilitating mutation affecting the Nib polymerase (Carringtonet al., 1993; Li and Carrington, 1995), failed to accumulatesignificant GUS activity. The CCCE and IGN mutants were am-plified to levels ~25 and 1%, respectively, of that of the parentalvirus at 72 hr pi (Figure 3B). Although IGN mutant-inducedGUS activity was low, it was significantly greater (P < 0.03)than the minimal activity in VNN mutant-inoculated cells atall time points tested. These results indicate that the FRN andCCCE mutations had only modest effects on genome amplifi-cation, whereas the IGN mutation resulted in a more seriousamplification defect.
B1 2 3 4 5
0027-1031Transgen. V
HC-Pro - — •*—(52-kD)
Figure 2. Synthesis and Processing of HC-Pro in Vitro and in Trans-genic Plants.(A) Synthetic transcripts encoding a wild-type (wt) or mutagenizedGUS/HC-Pro/partial P3 polyprotein (diagram) were produced in vitroand translated in a rabbit reticulocyte lysate in the presence of^S-methionine. Radiolabeled proteins were detected after SDS-PAGEand autoradiography (bottom), The HC-Pro mutations present in thevarious transcripts are indicated above each lane. The molecularmasses (in kilodaltons) of the precursor polyprotein (130 kD) andGUS/HC-Pro processed product (120 kD) are indicated at left.(B) Transgenic (Transgen.; lanes 1 and 2) or TEV-infected (V; lane 3)plants were subjected to immunoblot analysis using an anti-HC-Proserum. The immunoreaction was detected using a chemiluminescentsecond antibody and autoradiography. Ten times more of the trans-genic extracts were loaded compared with the virus-infected extract.
Systemic Infection by the HC-Pro Mutants
Tobacco plants were inoculated with transcripts of TEV-GUSand mutant genomes, and systemic infection was assayed bythe appearance of symptoms and detection of GUS activityat 12 days pi. Symptoms of parental TEV-GUS infection wereobserved in most plants by 5 days pi (Figure 4A). Plants inoc-ulated with transcripts from the FRN and IGN mutant genomesexhibited mild symptoms, and these were delayed by at least2 days, compared with the appearance of parental TEV-GUSsymptoms. No symptoms of infection were detected in CCCEmutant-inoculated plants. GUS activity was detected in leavestwo positions above the inoculated leaves of plants infectedwith parental TEV-GUS and the FRN and IGN mutants, al-though the levels of activity differed greatly among plantsinoculated with the different viruses (Figure 4B). Accumula-tion of the FRN and IGN mutants was approximately two andfour orders of magnitude, respectively, lower than that of pa-rental TEV-GUS. The levels of GUS activity among individualFRN and IGN mutant-infected plants were also quite variable.Among plants inoculated with the FRN and IGN mutants, atleast one failed to exhibit GUS activity in upper leaves, eventhough analysis of inoculated leaves indicated that infectionhad occurred. The differences between TEV-GUS and theFRN and IGN mutants were highly significant (P < 0.0001 inall comparisons). Because these two mutants were debilitatedin plants to a greater degree than in protoplasts, they appearedto possess slow or debilitated cell-to-cell and/or long-distancemovement phenotypes.
No GUS activity was detected in upper leaves of CCCE mu-tant-inoculated plants (Figure 4B), even at 16 days pi (datanot shown). All inoculated leaves had GUS activity, however,indicating that this virus may have been confined to initial in-fection sites. Considering both the protoplast and whole-plantresults, the CCCE mutant appeared to display a significantdefect in systemic infectivity. Because of this striking pheno-type, the movement characteristics of the CCCE mutant wereanalyzed in greater detail.
552 The Plant Cell
B A 1 0 0 0 ~ 900
$ 800 ,xg 700
.5 E 600 2% 500
O E 3 300 v 200
1 00
h
.-
5 3 400
+ TEV-GUS
FRN
o VNN
P 15 25 35 45 55 65 75
90003 EV-GUS
ICCE / h v1 c
3 ,Xg 6000 'F e
2 4000
3 3000
- R
v F& 2000
l o o o ~ 0
= + TEV-GUS
I CCCE 8000;
7000r
15 25 35 45 55 65 75
v i ü N
o VNN
Hours Post-lnoculation Hours Post-Inoculation
Figure 3. Amplification of Parental and Mutant TEV-GUS Genomes in Protoplasts.
GUS activity was measured in protoplasts inoculated with TEV-GUS or mutant transcripts at 24,48, and 72 hr pi. The data in (A) and (B) represent the mean of three concomitant, replicate samples using the same preparation of protoplasts. The experiments in (A) and (B) were conducted using different protoplast preparations. (A) Parental TEV-GUS and FRN and VNN mutants. (B) Parental TEV-GUS and CCCE, IGN, and VNN mutants.
Cell-to-Cell Movement of TEV-GUS and the CCCE Mutant
The cell-to-cell movement rates of parental TEV-GUS and the CCCE mutant were measured in inoculated leaves. lnfection foci were visualized at 24, 48, 72, and 96 hr pi by infiltration with the GUS histochemical substrate 5-bromo-4-chloro-3- indolyl P-D-glucuronide (X-gluc). The diameters of foci were determined microscopically and expressed in numbers of epidermal cells. At 24 hr pi, the mean diameters of foci pro- duced by TEV-GUS and the CCCE mutant were similar (P = 0.57), although many foci consisted of single cells in both cases (Figure 5A). Cell-to-cell movement of the CCCE mutant con- tinued at an average rate of 0.12 cells per hr, which was slightly slower than the parental TEV-GUS rate of 0.23 cells per hr. Measurements of TEV-GUS foci were not possible at 96 hr pi due to extensive secondary movement through the vascula- ture. These data indicate that the CCCE mutant was capable of cell-to-cell movement but at a reduced rate compared with parental TEV-GUS.
Continued incubation of TEV-GUS-infected and CCCE mu- tant-infected plants to 8 days pi showed that parental virus spread throughout the inoculated leaves by cell-to-cell and vascular-associated movement. The CCCE mutant, however, remained confined to initial infection foci (Figure 56). Cultur- ing of the CCCE mutant-infected plants to 16 days pi revealed that foci continued to expand via cell-to-cell movement but failed to exhibit vascular-associated movement (data not shown). The CCCE mutant, therefore, contained a major defect in systemic or long-distance movement.
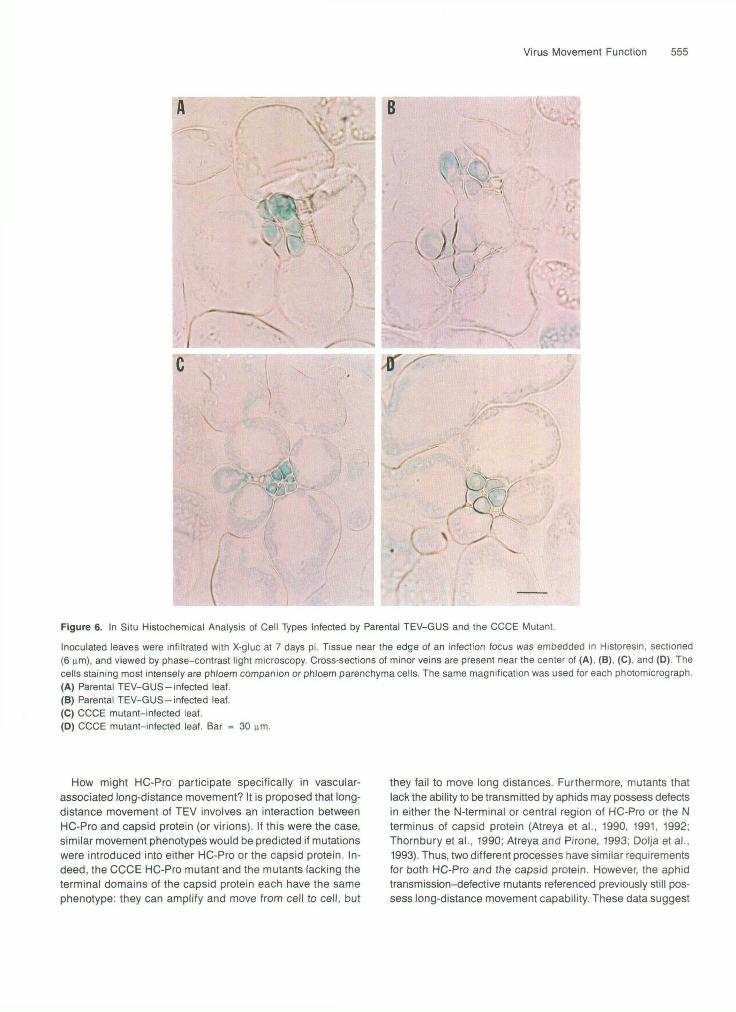
Long-distance movement through the vasculature first re- quires passage through a number of distinct cell types, including mesophyll, bundle sheath, and phloem companion cells. The inability to infect vasculature-associated cells would account for the CCCE mutant phenotype. The cell types in- fected by TEV-GUS and the CCCE mutant were examined in sections of Historesinembedded leaf tissue treated with X-gluc. Strong histochemical GUS reactions were detected in bundle sheath and phloem cells of tissue infected by parental (Figures 6A and 66) and CCCE mutant (Figures 6C and 6D) viruses. The reactions were particularly evident in the cytoplasm-dense phloem cells, some of which were companion cells.'This stain- ing appeared not to have resulted simply from the diffusion of the X-gluc reaction product from adjacent infected cells be- cause the intensity of the reaction was often high in the phloem cells and low in neighboring bundle sheath cells. The pattern of staining within virtually all positive cells was confined to the cytoplasm, with little or no leakage of the X-gluc reaction prod- uct into the vacuole or the nucleus (Figure 6). These data suggest that the CCCE mutant was capable of reaching phloem cells of inoculated leaves, indicating that the long-distance movement defect was associated with a point later in the move- ment pathway.
Complementation of the Long-Distance Movement Defect of the CCCE Mutant in HC-Pro-Expressing Transgenic Plants
Transgenic plants that expressed the 5' region of the TEV ge- nome, including P1 and HC-Pro sequences, were produced.

Virus Movement Function 553
A protein with an electrophoretic mobility identical to that of authentic HC-Pro was detected in transgenic plants using an anti-HC-Pro serum (Figure 28), indicating accurate process- ing of the transgenic polyprotein. The bulk level of accumulation of transgenic HC-Pro was -10% that of HC-Pro in TEV- GUS-infected plants.
Protoplasts from transgenic and nontransgenic plants were inoculated with full-length vira1 RNA transcripts. The relative levels of GUS activity induced by TEV-GUS and the CCCE mutant were the same in each protoplast preparation (data not shown), indicating that HC-Pro supplied by the transgenic cells had little effect on genome amplification of parental or mutant viruses at the single-cell level.
Transgenic and nontransgenic plants were inoculated with TEV-GUS and the CCCE mutant, and systemic infectivity was monitored by symptom production and GUS activity. As previ- ously described, TEV-GUS induced systemic symptoms by 5 days pi in nontransgenic plants, whereas infection by the CCCE mutant failed to result in symptoms. In contrast, both TEV-GUS and the CCCE mutant induced systemic symptoms in 1000/0 of transgenic plants, although symptoms caused by the mutant were delayed by 2 to 3 days (Figure 7A). Indistin- guishable levels of GUS activity were detected in transgenic and nontransgenic plants infected by TEV-GUS. GUS activity induced by the CCCE mutant was detected only in transgenic plants (Figure 78) and at a level reduced considerably rela- tive to that in TEV-GUS-infected plants. The same results
100 7 A o TEV-GUS
were obtained using each of two independent transgenic lines (data not shown). Systemic infection by the CCCE mutant was dueto complementation rather than to recombination between the mutant genome and the transgene because inoculation of nontransgenic plants with the virus recovered from trans- genic plants resulted in an infection phenotype identical to that of the original CCCE virus derived from transcripts.
To determine the effect of transgenic HC-Pro on cell-to-cell movement of the CCCE mutant in inoculated leaves, the in situ histochemical assay was conducted at time intervals through 72 hr pi. lnfection of transgenic plants with the CCCE mutant resulted in only modest stimulation of cell-to-cell movement (Figure 8A). Interestingly, cell-to-cell movement of TEV-GUS was suppressed partially in the transgenic plants, resulting in infection foci of sizes comparable to the CCCE mutant at each time point.
Secondary, vascular-associated movement was analyzed in situ in inoculated and noninoculated leaves of transgenic and nontransgenic plants at 13 days pi. Extensive secondary movement was detected in transgenic leaves inoculated with the CCCE mutant, whereas only individual, primary infection foci were visible in nontransgenic plants (Figure 88). Upper noninoculated leaves also exhibited GUS activity radiating from primary veins (Figure 8C), clearly showing vascular-associated movement to systemic tissues. These data indicate that HC- Pro supplied in frans complemented, at least partially, the long- distance movement defect of the CCCE mutant.
O o 1 2 3 4 5 6 7 8 9 1 0
Days Post-Inoculation
TEV- FRN IGNCCCE GUS
Figure 4. Systemic lnfection of Plants by Parenta1 TEV-GUS and Mutants.
(A) Vira1 symptoms in upper leaves were scored at daily intervals after inoculation with parental TEV-GUS or mutants containing the FRN, IGN, or CCCE mutations. The percentage of plants exhibiting symptoms is plotted. (B) GUS activity was quantified in extracts from leaves two positions above the inoculated leaves at 12 days pi. Each bar represents the mean from seven to 10 plants. Activity values from individual plants are indicated.

554 The Plant Cell
A
12H£ ^
Q 13
OO 73
c <uO
o 54-H
H 8:6-
4-
2-
» TEV-GUS
• CCCE
' ' |" ' ' |" " I1 ' ' ' I1 ' ' ' I1 ' ' ' I" ' ' I' ' ' ' I1 T rT]
15 25 35 45 55 65 75 85 95 105
Hours Post-Inoculation
B
TEV-GUS CCCE(Inoculated Xanthi leaves)
Figure 5. Cell-to-Cell Movement of Parental TEV-GUS and the CCCEMutant in Inoculated Leaves.(A) Diameters of infection foci, expressed in numbers of epidermalcells, in inoculated tobacco leaves were measured microscopically atdaily intervals postinoculation. Each point represents the mean fromat least 39 foci.(B) Infection of inoculated leaves by parental TEV-GUS or the CCCEmutant was visualized by infiltration with X-gluc at 8 days pi. Note thatparental virus spread throughout the leaf, whereas the CCCE mutantwas confined to individual infection foci.
DISCUSSION
Previous investigations revealed that cell-to-cell movement andlong-distance transport of TEV are distinct processes withdifferent requirements (Dolja et al., 1994,1995). In fact, it hasbeen suggested that these two modes of movement are uniqueprocesses in other viral systems as well (e.g., Dawson, 1992;Taliansky and Garcia-Arenal, 1995). Here, we demonstratedthat TEV encodes a nonstructural protein, HC-Pro, which pro-vides a specific frans-active function in vascular-associatedtransport.
The collective analyses of three mutants indicated that theHC-Pro long-distance movement activity is distinct from therole HC-Pro may play in genome amplification (Table 1). First,the long-distance movement-defective CCCE mutant couldamplify in protoplasts to 25% of the level of parental TEV-GUS.Based on results with other TEV-GUS mutants, such as thosewith defects in the P1 protein (Verchot and Carrington, 1995b),this level of replication within single cells is clearly not debilitat-ing to cell-to-cell or long-distance movement. Second, the IGNmutant encoded a debilitated HC-Pro that resulted in drasti-cally reduced genome amplification levels (<1% comparedwith parental virus), yet this mutant was capable of limitedlong-distance movement. The limited systemic infection capa-bility of the IGN mutant may have resulted from decreases inboth genome amplification and long-distance movement rates,although it could have been due solely to indirect effects oflimited amplification. Because the defect produced by theCCCE mutation had only a minor effect on genome amplifi-cation but a completely debilitating effect on long-distancemovement (Table 1), these two functions are genetically separ-able. Finally, the fact that HC-Pro supplied in transgenic cellscould restore long-distance movement activity to the CCCEmutant but could not stimulate genome amplification of theCCCE mutant at the single-cell level in protoplasts impliesthat the movement and replication functions of HC-Pro aredistinct.
Whereas the CCCE mutant was complemented in HC-Pro-expressing transgenic plants, it was not restored to thelevel of the parental virus. This could be due to one or morereasons. Immunoblot analysis indicated that the transgenicplants accumulated HC-Pro to levels 10% of those detectedin TEV-GUS —infected plants. The relatively low level of func-tional HC-Pro may have had a quantitative effect on thelong-distance movement function. Alternatively, the CCCE mu-tant HC-Pro accumulating in infected cells may have beenfrans-inhibitory to the complementing activity of the transgenicprotein. If the movement function of HC-Pro requires multipledomains or binding sites and the CCCE mutation affects onlyone of these putative domains, then the mutant protein mayhave partially interfered with the movement process. Suchfrans-dominant interference was proposed to explain the lackof transgenic complementation of long-distance movement ofa TEV mutant lacking the C-terminal domain of capsid protein(Dolja et al., 1995).
Virus Movement Function 555
A B
V ,
.V;..
Figure 6. In Situ Histochemical Analysis of Cell Types Infected by Parental TEV-GUS and the CCCE Mutant.
Inoculated leaves were infiltrated with X-gluc at 7 days pi. Tissue near the edge of an infection focus was embedded in Historesin, sectioned(6 urn), and viewed by phase-contrast light microscopy. Cross-sections of minor veins are present near the center of (A), (B), (C), and (D). Thecells staining most intensely are phloem companion or phloem parenchyma cells. The same magnification was used for each photomicrograph.(A) Parental TEV-GUS-infected leaf.(B) Parental TEV-GUS-infected leaf.(C) CCCE mutant-infected leaf.(D) CCCE mutant-infected leaf. Bar = 30 nm.
How might HC-Pro participate specifically in vascular-associated long-distance movement? It is proposed that long-distance movement of TEV involves an interaction betweenHC-Pro and capsid protein (or virions). If this were the case,similar movement phenotypes would be predicted if mutationswere introduced into either HC-Pro or the capsid protein. In-deed, the CCCE HC-Pro mutant and the mutants lacking theterminal domains of the capsid protein each have the samephenotype: they can amplify and move from cell to cell, but
they fail to move long distances. Furthermore, mutants thatlack the ability to be transmitted by aphids may possess defectsin either the N-terminal or central region of HC-Pro or the Nterminus of capsid protein (Atreya et al., 1990, 1991, 1992;Thornbury et al., 1990; Atreya and Pirone, 1993; Dolja et al.,1993). Thus, two different processes have similar requirementsfor both HC-Pro and the capsid protein. However, the aphidtransmission-defective mutants referenced previously still pos-sess long-distance movement capability. These data suggest

556 The Plant Cell
A B
Figure 7.
O 2 4 6 8 1 0 1 2 Days Post-Inoculation
Complementation of the CCCE Mutant in Transgenic Plants.
TEV- mv- ccc ccc GUS GUS Xan. Tran. Xan.Tran.
Nontransgenic (Xanthi or Xan.) or HC-Pro-expressing transgenic (Transgen. or Tran.) plants were inoculated with parenta1 TEV-GUS or the CCCE (CCC) mutant. (A) Vira1 symptoms in upper leaves were scored at daily intervals after inoculation. The percentage of plants exhibiting symptoms is plotted. (e) GUS activity was quantified in extracts from leaves two positions above the inoculated leaves 12 days pi. Each bar represem the mean from five to seven plants. Activity values from individual plants are indicated.
that HC-Pro and the capsid protein interact to form active com- plexes that facilitate insect transmission and long-distance movement but that the nature of the interaction required for each process may differ.
It is further proposed that the putative HC-Pro-capsid pro- tein interaction provides a unique function in movement through the companion celllsieve element boundary. The histochemi- cal analysis indicated that the CCCE mutant could reach at least as far as the phloem companion cells of inoculated leaves. Therefore, the mutant may have been blocked at the point of entry into sieve elements or at the point of exit from sieve ele- ments back into companion cells at sites dista1 to the-initial infection. The idea that transport across the companion celll sieve element boundary requires a set of unique movement functions is attractive because evidence suggests that plasmo- desmata at this junction are quite different from plasmodesmata between mesophyll cells (Kempers et al., 1993). The plas- modesmata between mesophyll cells and between phloem cells were shown, in dye injection experiments, to differ func- tionally (Ding et al., 1992). In addition, the density of companion celllsieve element plasmodesmata per unit area is relatively low in tobacco (Van Bel and Gamalei, 1992). HC-Pro mayfacili- tate transport through these specialized plasmodesmata. We do not know whether HC-Pro also enters the sieve element along with the virion or transport complex. Entry of HC-Pro into sieve elements is clearly plausible, given that MPs from
severa1 diverse virus groups are able to move themselves through plasmodesmata (Fujiwara et al., 1993; Noueiry et al., 1994; Waigmann et al., 1994).
METHODS
Strains and Plants
A modified form of the highly aphid-transmissible strain of tobacco etch virus (TEV), the genome of which is represented as cDNA in pTEV7DA (Dolja et al., 1992), was used in this study. Nicotiana taba- cum cv Xanthi-nc was used as the host in whole-plant experiments and as the source of protoplasts. All cloning of DNA was conducted in fscherichia coli HBlO1.
Two independent transgenic tobacco lines, termed 0027-1031-19x8 and 0027-1031-1XB, that expressed the P1-helper component proteinase (HC-Pro) region of the TEV genome were produced using the leaf disc method (Horsch et al., 1985; Carrington and Freed, 1990). Progeny from the RI or R2 generations were analyzed for expression of HC-Pro by immunoblot analysis using an anti-HC-Pro serum (Carrington et al., 1990) and a chemiluminescence detection system (Amersham).
Site-Directed Mutagenesis of the HC-Pro Sequence
Three substitution mutations (FRN, IGN, and CCCE) were introduced into the HC-Pro coding sequence within the plasmid pTL7SN-

Virus Movement Function 557
E =a _ 10-
73'a.udc
8-
6-
4-
» TEV-GUSXan.o TEV-GUS Transgen.• CCCEXan.V CCCE Transgen.
15 25 35 45 55 65Hours Post-Inoculation
75
Xanthi TransgenicCCCE-Inoculated Leaves
TEV-GUS CCCESystemically Infected Transgenic Leaves
Figure 8. Movement of Parental TEV-GUS and the CCCE Mutant inInoculated Systemically Infected Leaves of Transgenic Plants.
(A) Diameters of infection foci, expressed in numbers of epidermalcells, in inoculated nontransgenic (Xanthi [Xan.]) or HC-Prc-expressingtransgenic (Transgen.) tobacco leaves were measured microscopicallyat daily intervals postinoculation. Each point represents the mean fromat least 10 foci.(B) Infection of inoculated leaves of nontransgenic (Xanthi) and HC-Pro-expressing transgenic leaves by the CCCE mutant was visual-ized by infiltration with X-gluc at 13 days pi. Note that the secondarymovement defect of the CCCE mutant was rescued in the transgenicplants.(C) Leaves Systemically infected by parental TEV-GUS and the CCCEmutant in HC-Pro-expressing transgenic plants are shown. Sites ofinfection were visualized as given in (B).
0823GJH, which contains cDNA corresponding to TEV nucleotides849 to 2332 of the TEV-GUS genome between two Xbal sites. Thisplasmid also contains the 3-glucuronidase (GUS) coding sequencebetween the P1 and HC-Pro regions as well as codons for an Nla pro-teinase cleavage site between GUS and HC-Pro (Carrington et al.,1993). For each mutant, three codons were replaced by the sequenceAGGCCTGCT, which contains a Stul site and codes for the tripeptideArg-Pro-Ala. The mutations affected Phe-182-Arg-183-Asn-184 (FRN),lle-251-Gly-252-Asn-253 (IGN), and Cys-293-Cys-294-Cys-295(CCCE). The CCCE mutant also contained a substitution of Asp forGlu-299. Mutagenesis was conducted by the oligonucleotide-directedmethod (Kunkel et al., 1987) and verified by nucleotide sequenceanalysis.
The mutations were introduced into a GUS-containing, full-lengthcDNA of the TEV genome by transfer of the Xbal-Xbal fragment frompTL7SN-0823GiH into pTEV7DAN. The resulting plasmidswere namedpTEV7DAN-GiH/FRN, pTEV7DAN-GiH/IGN, and DTEV7DAN-GIH/CCCE. These plasmids contain a bacteriophage SP6 promoter adja-cent to the TEV 5' sequence and a Bglll site adjacent to the 3' poly(A)region.
The region encoding GUS, mutagenized HC-Pro, and a 10-kD seg-ment of the P3 protein was subcloned from the full-length plasmidsdescribed previously into the vector pTL7SN.3 (Carrington and Freed,1990). These plasmidswere termed pTL7SN-1027GIH/FRN, pTL7SN-1027GiH/IGN, and DTL7SN-1027G1H/CCCE.
In Vitro Transcription and Translation
Full-length TEV transcripts were generated using SP6 RNA polymer-ase and Bglll-linearized DMA as described previously (Dolja et al., 1992).These transcripts were used for inoculation of plants and protoplasts.
Transcripts were also produced from the pTL7SN-1027GiH-basedplasmids using SP6 RNA polymerase. These transcripts were trans-lated in a rabbit reticulocyte lysate (Promega) in the presence of35S-methionine (Du Pont-New England Nuclear) for 1 hr. The radio-labeled products were subjected to electrophoresis through anSDS-10% polyacrylamide gel and detected by autoradiography.
Table 1. Properties of the FRN, IGN, and CCCE Mutants inNontransformed Plants and Protoplasts
Virus
TEV-GUSFRN mutantIGN mutantCCCE mutant
RelativeGenomeAmplification3
(O/o)
10062
0.625
Relative SystemicGUS Activity"(%)
1001.60.040.00
Cell-to-CellMovementRate
0.23 cell/hrNDC
NDC
0.1 2 cell/hra Based on GUS activity in protoplasts at 72 hr pi as shown in Figure3, using the TEV-GUS value as the 100% standard.b Based on GUS activity in upper, noninoculated leaves at 12 dayspi as shown in Figure 4, using the TEV-GUS value as the 100%standard.c Not determined.

558 The Plant Cell
lnoculation of Plants and Protoplasts
Carborundum-dusted plants were inoculated mechanically using RNA transcripts ( 4 0 wg) or inoculum prepared by grinding 1 g of virus- infected leaves in 2 mL of TE buffer (10 mM Tris-HCI, 1 mM EDTA, pH 7.6).
Protoplasts were prepared from greenhouse-grown plants and in- oculated (7.5 x 105 cells) with SP6 transcripts using the polyethylene glycol method described previously (Negrutiu et al., 1987; Carrington and Freed, 1990). lnoculations were conducted in triplicate, using a single preparation of protoplasts for each experiment. Transcripts de- rived from pTEV7DAN-GJH (the plasmid yielding TEV-GUS) and pTEV7DAN-G.1 HlVNN served as the positive and negative amplifica- tion controls, respectively. The latter contained a mutation within the Nlb polymerase sequence, resulting in an amplification-defective phenotype (Carrington et al., 1993; Li and Carrington, 1995).
GUS Activity Assays
Protoplasts (2.5 x 105) were harvested at 24, 48, and 72 hr postinocu- lation (pi), frozen at -80°C, and resuspended in GUS lysis buffer (40 mM sodium phosphate, 10 mM EDTA, 0.1% Triton X-100, 0.1% sodium lauryl sarcosine, 0.07°/o p-mercaptoethanol, pH 7.0), and GUS activity was measured using the fluorometric substrate 4-methylumbelliferyl glucuronide (Jefferson, 1987). GUS activity was also measured using the fluorometric assay in extracts from virus-infected leaves. Leaf tis- sue was ground in 5 volumes of GUS lysis buffer and clarified by centrifugation at 13,OOOg for 10 min.
GUS activity was visualized in virus-infected leaves after vacuum infiltration of 5-bromo-4-chloro-3-indolyl p-D-glucuronide (X-gluc), a colorimetric GUS substrate (Dolja et al., 1992). In some experiments, infection focus diameter (expressed in number of epidermal cells) was measured (Dolja et al., 1994). Statistical analysis of data was performed using the StatView 4.0 program (Abacus Concepts, Inc., Berkeley, CA).
Histochemistry and Microscopy
lnoculated leaves of nontransgenic tobacco plants were infiltrated with X-gluc at 7 days pi. Tissue near the edge of an infection focus was excised with a razor blade, fixed in 50 mM Pipes, pH 7.4,4% parafor- maldehyde (Electron Microscopy Sciences, Fort Washington, PA), 05% glutaraldehyde (Sigma), dehydrated through a graded acetone series, and embedded in Historesin (Reichert-Jung, Heidelberg, Germany). Sections (6-bm) were cut with glass knives using an Ultracut micro- tome and mounted in Aqua-polymount (Polysciences, Inc., Warrington, PA) on glass slides. Sections were viewed with an Olympus 6x50 microscope (Tokyo, Japan) using phase-contrast optics.
ACKNOWLEDGMENTS
We thank Kristin Kasschau for constructing one of the intermediate cloning vectors used in this study, Patricia Valdez for help with grow- ing plants, Kate VandenBosch and Gyung Hee Lee for assistance with microscopy, and Valerian Dolja for assistance and advice during this project. We also thank Kristin Kasschau for providing helpful comments on the manuscript. This research was supported by grants from the National Research lnitiative Program of the U.S. Department of
Agriculture (No. 91-373036435), the National Science Foundation (No. lBN-9158559), and the National lnstitutes of Health (No. Al27832). M.C.S. was supported by a National Research Service Award from the Na- tional lnstitutes of Health (No. A109121).
Received January 23, 1995; accepted March 16, 1995.
REFERENCES
Atreya, C.D., and Pirone, T.P. (1993). Mutational analysis of the helper component proteinase gene of a potyvirus: Effects of amino acid substitutions, deletions, and gene replacement on virulence and aphid transmissibility. Proc. Natl. Acad. Sci. USA 90, 11919-11923.
Atreya, C.D., Raccah, R., and Pirone, T.P. (1990). A point mutation in the coat protein abolishes aphid transmissibility of a potyvirus. Virology 178, 161-165.
Atreya, C.D., Atreya, P.L., Thornbury, D.W., and Pirone, T.P. (1992). Site-directed mutations in the potyvirus HC-PRO gene affect helper component activity, virus accumulation, and symptom expression in infected tobacco plants. Virology 191, 106-111.
Atreya, P.L., Atreya, C.D., and Pirone, T.P. (1991). Amino acid substi- tutions in the coat protein result in loss of insect transmissibility of a plant virus. Proc. Natl. Acad. Sci. USA 88, 7887-7891.
Beck, D.L., Guilford, P.J., Voot, D.M., Andersen, M.U., and Forster, R.L. (1991). Triple gene block proteins of white clover mosaic potex- virus are required for transport. Virology 183, 695-702.
Carrington, J.C., and Freed, D.D. (1990). Cap-independent enhance- ment of translation by a plant potyvirus 5’ nontranslated region. J. Virol. 64, 1590-1597.
Carrington, J.C., Cary, S.M., Parks, T.D., and Dougherty, W.G. (1989a). A second proteinase encoded by a plant potyvirus genome.
Carrington, J.C., Freed, D.D., and Sanders, T.C. (1989b). Autocata- lytic processing of the potyvirus helper component proteinase in Escherichia coli and in vitro. J. Virol. 63, 4459-4463.
Carrington, J.C., Freed, D.D., and Oh, C.4. (1990). Expression of potyviral polyproteins in transgenic plants reveals three proteolytic activities required for complete processing. EMBO J. 9, 1347-1353.
Carrington, J.C., Haldeman, R., Dolja, V.V., and Restrepo-Hartwig, M.A. (1993). Interna1 cleavage and trans-proteolytic activities of the VPg-proteinase (Nla) of tobacco etch potyvirus in vivo. J. Virol. 67,
Citovsky, V., Knorr, D., Schuster, G., and Zambryski, P. (1990). The P30 movement protein of tobacco mosaic virus is a single-strand nucleic acid binding protein. Cell 60, 637-647.
Citovsky, V., Wong, M.L., Shaw, A.L., Prasad, B.V.V., and Zambryski, P. (1992). Visualization and characterization of tobacco mosaic vi- rus movement protein binding to single-stranded nucleic acids. Plant Cell 4, 397-411.
Dawson, W.O. (1992). Tobamovirus-plant interactions. Virology 186, 359-367.
Deom, C.M., Lapidot, M., and Beachy, R.N. (1992). Plantvirus move- ment proteins. Cell 69, 221-224.
Ding, B., Haudenshield, J.S., Hull, R.J., Wolf, S., Beachy, R.N., and Lucas, W.J. (1992). Secondary plasmodesmata are specific
EMBO J. 8, 365-370.
6995-7000.

Virus Movement Function 559
sites of localization of the tobacco mosaic virus movement protein in transgenic tobacco plants. Plant Cell 4, 915-928.
Dolja, V.V., McBride, H.J., and Carrington, J.C. (1992). Tagging of plant potyvirus replication and movement by insertion of p-glucuroni- dase into the viral polyprotein. Proc. Natl. Acad. Sci. USA 89, 10208-10212.
Dolja, V.V., Herndon, K.L., Pirone, T.P., and Carrington, J.C. (1993). Spontaneous mutagenesis of a plant potyvirus genome after inser- tion of a foreign gene. J. Virol. 67, 5968-5975.
Dolja, V.V., Haldeman, R., Robertson, N.L., Dougherty, W.G., and Carrington, J.C. (1994). Distinct functions of capsid protein in as- sembly and movement of tobacco etch potyvirus in plants. EM60
Dolja, V.V., Haldeman-Cahill, R., Montgomery, A.E., VandenBosch, K.A., and Carrington, J.C. (1995). Capsid protein determinants in- volved in cell-to-cell movement of tobacco etch potyvirus. Virology
Fujiwara, T., Giesman-Cookmeyer, D., Ding, B., Lommel, S.A., and Lucas, W.J. (1993). Cell-to-cell trafficking of macromolecules through plasmodesmata potentiated by the red clover necrotic mosaic virus movement protein. Plant Cell 5, 1783-1794.
Giesman-Cookmeyer, D., and Lommel, S.A. (1993). Alanine scan- ning mutagenesis of a plant virus movement protein identifies three functional domains. Plant Cell 5, 973-982.
Horsch, R.B., Fry, J.E., Hoffmann, N.L., Eichholtz, D., Rogers, S.G., and Fraley, R.T. (1985). A simple and general method for transfer- ring genes into plants. Science 227, 1229-1231.
Hull, R. (1991). The movement of viruses within plants. Semin. Virol. 2, 89-95.
Jeffenon, R.A. (1987). Assaying chimeric genes in plants: The GUS gene fusion system. Plant MOI. Biol. Rep. 5, 387-405.
Kasteel, D., Wellink, J., Verver, J., van Lent, J., Goldbach, R., and van Kammen, A. (1993). The involvement of cowpea mosaic virus M RNA-encoded proteins in tubule formation. J. Gen. Virol. 74,
Kempen, R., Prior, D.A.M., Van Bel, A.J.E.,andOparka, K.J. (1993). Plasmodesmata between sieve element and companion cell of ex- trafascicular stem phloem of Cucurbita maxima permit passage of 3 kD fluorescent probes. Plant J. 4, 567-575.
Klein, RG., Klein, R.R., Rodríguez-Cerezo, E., Hunt, A.G., and Shaw, J.G. (1994). Mutational analysis of the tobacco vein mottling virus genome. Virology 204, 759-769.
Kunkel, T.A., Roberts, J.D., and Zakour, R. (1987). Rapid and effi- cient site-specific mutagenesis without phenotypic selection. Methods Enzymol. 154, 367-382.
Li, X.H., and Carrington, J.C. (1985). Complementation of tobacco etch potyvirus mutants by active RNA polymerase expressed in trans- genic cells. Proc. Natl. Acad. Sci. USA 92, 457-561.
J. 13, 1482-1491.
206, 1007-1016.
1721-1724.
Maule, A.J. (1991). Virus movement in infected plants. Crit. Rev. Plant Sci. 9, 457-473.
Negrutiu, I., Shillito, R., Potrykus, I., Biasini, G., and Sala, F. (1987). Hybrid genes in the analysis of transformation conditions, 1: Set- ting up a simple method for direct gene transfer in plant protoplasts. Plant MOI. Biol. 8, 363-373.
Noueiry, A.O., Lucas, W.J., and Gilbertson, R.L. (1994). Two pro- teins of a plant DNA virus coordinate nuclear and plasmodesmata transport. Cell 76, 925-932.
Oh, C.-S., and Carrington, J.C. (1989). ldentification of essential residues in potyvirus proteinase HC-Pro by site-directed mutagen- esis. Virology 173, 692-699.
Pascal, E., Sanderfoot, A.A., Ward, B.M., Medville, R., Turgeon, R., and Lazarowitz, S.G. (1994). The geminivirus BR1 movement protein binds single-stranded DNA and localizes to the cell nucleus. Plant Cell 6, 995-1006.
Petty, I,T.D., Flench, R., Jones, R.W., and Jackson, A.O. (1990). Iden- tification of barley stripe mosaic virus genes involved in viral RNA replication and systemic movement. EMBO J. 9, 3453-3457.
Restrepo-Hartwig, M.A., and Carrington, J.C. (1994). The tobacco etch potyvirus 6-kilodalton protein is membrane-associated and in- volved in viral replication. J. Virol. 68, 2388-2397.
Riechmann, J.L., Laín, S., and Garcia, J.A. (1992). Highlights and prospects of potyvirus molecular biology. J. Gen. Virol. 73, 1-16.
Shukla, D.D., Ward, C.W., and Brunt, A.A. (1994). The Potyviridae. (Oxford, UK: CAB International).
Taliansky, M E , and GarcíaArenal, F. (1995). Role of cucumovirus capsid protein in long-distance movement within the infected plant. J. Virol. 69, 916-922.
Thornbury, D.W., Pattenon, C.A., Dessens, J.T., and Pirone, T.P. (1990). Comparative sequences of the helper component (HC) re- gion of potato virus Y and a HC-defective strain, potato virus C. Virology 178, 573-578.
Van Bel, A.J.E., and Gamalei, Y.V. (1992). Ecophysiology of phloem loading in source leaves. Plant Cell Environ. 15, 265-270.
van Lent, J., Wellink, J., and Goldbach, R. (1990). Evidence for the involvement of the 58K and 48K proteins in the intercellular move- ment of cowpea mosaic virus. J. Gen. Virol. 7l , 219-223.
Verchot, J., and Carrington, J.C. (1995a). Debilitation of plant potyvi- rus infectivity by P1 proteinase-inactivating mutations and restoration by second-site modifications. J. Virol. 69, 1582-1590.
Verchot, J., and Carrington, J.C. (1995b). Evidence that the potyvi- rus P1 proteinase functions in trans as an accessory factor for genome amplification. J. Virol. 69, in press.
Waigmann, E., Lucas, W.J., Citovsky, V., and Zambryski, P. (1994). Evidence for protein trafficking via plasmodesmata. Proc. Natl. Acad. Sci. USA 91, 1433-1437.

DOI 10.1105/tpc.7.5.549 1995;7;549-559Plant Cell
S Cronin, J Verchot, R Haldeman-Cahill, M C Schaad and J C Carringtonproteinase.
Long-distance movement factor: a transport function of the potyvirus helper component
This information is current as of June 16, 2018
Permissions https://www.copyright.com/ccc/openurl.do?sid=pd_hw1532298X&issn=1532298X&WT.mc_id=pd_hw1532298X
eTOCs http://www.plantcell.org/cgi/alerts/ctmain
Sign up for eTOCs at:
CiteTrack Alerts http://www.plantcell.org/cgi/alerts/ctmain
Sign up for CiteTrack Alerts at:
Subscription Information http://www.aspb.org/publications/subscriptions.cfm
is available at:Plant Physiology and The Plant CellSubscription Information for
ADVANCING THE SCIENCE OF PLANT BIOLOGY © American Society of Plant Biologists