losses of biota from american aquatic communities due to acid rain
TRANSCRIPT

L O S S E S OF BIOTA FROM A M E R I C A N A Q U A T I C C O M M U N I T I E S
D U E TO ACID RAIN*
D. W. S C H I N D L E R , S. E. M. KASIAN and R. H. H E S S L E I N
Department of Fisheries and Oceans, Freshwater Institute, 501 University Crescent, Winnipeg, MN R3T 2N6, Canada
(Received March 1989)
Abstract. Models based on chemical survey data and geochemical assumptions were calibrated for areas where rates of acidification are known, then used to predict the declines in alkalinity and pH of lakes in the eastern and midwestern U.S.A. These results were combined with known acid tolerances of different taxonomic groups to estimate the extent of damage caused by acid rain to biological assemblages.
An average of over 50~/0 of the species in some taxonomic groups have probably been eliminated from lakes in the Adirondacks, Poconos-Catskills and southern New England. Moderate damage to biotic communities was predicted for lakes in central New England, and north-central Wisconsin. Damage predicted in Maine, upper Michigan, northeastern Minnesota and the remainder of the upper Great Lakes region was slight. Crustaceans, molluscs, leeches and insects were among the most severely affected groups. Among fishes, species of minnows (Cyprindae) were depleted in the most heavily acidified regions, with some declines in salmonid and centrarchid species.
Predicted damage to individual lakes in all areas was highly variable. In areas receiving highly acidic deposition, 100~/0 of the species in acid-sensitive taxonomic groups were eliminated in some lakes, while damage to other lakes was predicted to be slight.
Estimated damage varied from lake to lake within each subregion, based on chemical characteristics. The most heavily damaged lakes in the Adirondacks and Pocono-Catskills have probably lost all species of molluscs, leeches and crustaceans. On the other hand, lakes of the Midwest showed either slight increases or decreases in the richness of predicted biotic communities.
The possible ranges of original sulfate concentrations in lakes and the proportion of sulfuric acid in precipitation that liberated base cations from catchments were confined to relatively narrow limits by the model.
1. Introduction
For the past several years, Americans have expressed concern about the destruction of species of plants and animals, particularly in third-world, tropical countries (Wilson, 1985 and several papers in this conference). We share this concern, but believe that North Americans should be equally concerned about biotic impoverish- ment on this continent, where ecological communities are stressed by a wide variety of human insults. As an example, here we describe results from a model that suggests that the biological communities of softwater lakes in the American Northeast have been severely impoverished by acid rain. Despite hundreds of millions of dollars spent for ecological research, we do not know the effect of a single pollutant on a single type of American ecological community! A primary reason is that a biological inventory of North America has never been performed, despite the warnings of S. A. Forbes over a century ago that such inventories were vital to maintaining our ecosystems.
* Contribution from 'Fourth World Wilderness Congress - Acid Rain Symposium, Denver (Estes Park), Colorado', September 11-18, 1987.
Environmental Monitoring and Assessment 12: 269-285, 1989. �9 1989 Schindler, Kasian and Hesslein. For the Department of Fisheries and Oceans Government of Canada.

270 D. w. SCHINDLER ET AL.
I cannot too strongly emphasize the fact.., that a comprehensive survey of our entire natural history is absolutely essential to a good working knowledge of those parts of it which chiefly attract popular attention - that is, its edible fisheries, its injurious and beneficial insects, and its parasitic plants (Forbes, 1883).
In brief: Because we have never taken the trouble to learn what we have, we cannot know what we have lost. So far, there are no signs that this dilemma will be rectified. Little research money is spent on biological inventory, and taxonomic sciences are generally given low priority in American universities.
Studies of acid-sensitive aquatic ecosystems are not exceptions. We do not have even rudimentary biological inventories for most acid-sensitive softwaters. As usual, the few inventories that have been done have usually been directed at the largest, most beautiful members of the ecosystems: adult fishes of species that are valued for sport.
Because biological surveys have been neglected, some authors have concluded that American aquatic ecosystems have been damaged very little by acid rain (NAPAP, 1987). We do not share this belief, and consider it to be necessary to deduce by indirect methods what has probably happened to American aquatic ecosystems under the onslaught of acid rain. The remainder of this paper will describe such an attempt.
Earlier Studies of Biotic Impoverishment by Acid Rain
Both extensive surveys and ecosystem-scale experiments show a correlation between the number of species and pH (Okland and Okland 1980, 1986; Mills and Schindler, 1986). However, in the U.S.A. the study of regional biological effects of aidification has received little attention. In Norway and Sweden, thousands of lakes and streams have been surveyed for invertebrate species (Okland and Okland, 1980, 1986; Raddum, 1980; Engblom and Lingdell, 1984). In the U.K., the early surveys of Bell (1971) showed the connection between acidification and biotic impoverishment. In Canada, stream surveys done 40 years ago were recently duplicated, showing that several species of insects had disappeared and that pH values had declined by several tenths of a unit (Hall and Ide, 1987). Other Canadian work on invertebrates was reviewed by Dillon et al. (1984).
Ecosystem-level experiments have also shown that acidification eliminates sensi- tive organisms at pH values as high as 5.8 to 6.0 (Nero and Schindler, 1983; Schindler et aL, 1985; France, 1987; Mills, 1984; Mills et al., 1987). Unspectacular though these organisms may be, their demise eventually caused starvation and increased mortality among lake trout (Schindler et al., 1985; Mills et aL, 1987). Furthermore, the species assemblages present in different lakes with similar pH values do not seem to differ if the lakes have been anthropogenically or naturally acidified (Engblom and Ling- dell, 1984). Several studies have shown that the pH values and alkalinities of eastern North American ecosystems have decreased due to deposition of strong acids in the past century (Harvey, 1980; Watt er al., 1979; Watt et al., 1983; Dillon et aL, 1979, 1987; Asbury et al., 1989; Hall and Ide, 1987).

LOSSES OF BIOTA FROM AMERICAN AQUATIC COMMUNITIES
TABLE I
Changes in the number os species in various taxonomic groups in Lake 223 acidification
271
during experimental
1974-75 1979-80 1981 -83 pH 6.5 pH 5.5-5.6 pH 5.0-5.1
Phytoplankton 78 87-89 73 Littoral algae 30 19 16 (53~ Zooplankton 31 28 19 (61 To)
Cladocera 4 4 3 Calanoida 3 2 1 Cyclopoida 4 3 2 Rotifera 20 19 13
Macrobenthos Chironomidae 63-71 62-64 38-39 (55%) Other insects ? ? ? Crustacea 3 1 0 (0%) Molluscs and leeches ? ? ?
Fish 5-6 3" 3**
Total species 218 204 150 (69%)
* 2 species reproducing ** no species reproducing.
Several monitoring studies show that declines in the number of species and the acidification of aquatic ecosystems are directly correlated. In North America, the earliest of these were the observations by Harvey and his colleagues of declines in pH and number of fish species in the LaCloche Mountains of Ontario, under the influence of SO2 emissions from smelters in the Sudbury area (Beamish and Harvey, 1972; Harvey, 1980; Harvey and Lee, 1982).
Whole-lake experiments have shown results that support Harvey's observations, and extend them to other taxonomic groups. In an experimentally-acidified lake (no. 223), we found that the total number of species in taxonomic groups studied was reduced from about 220 to less than 150 as the pH decreased from 6.5 to 5.0 (Table I). Increases in dissolved trace metals were very slight during this experiment, so that acid alone was responsible for the deletions (Schindler et al., 1985). Results of a second experiment, in Lake 302 south, are now partially analyzed, and so far they agree with those of Lake 223, showing that the Lake 223 results were not fortuitous (Findlay, unpubl, ms; Shearer and De Bruyn, 1987; Turner et al., 1987; Experimental Lakes staff, unpubI, data).
Indirectly .Estimating Regional Biological Damage
Regional assessments of damage to aquatic communities in North America pose a number of problems, due to the lack of either historical chemical data or regional biological surveys in the U.S.A. However, three factors made it possible for us to

272 D . w . SCHINDLER ET AL.
make indirect estimates for the northeastern U.S.A.: 1) A uniformly high-quality set of recent chemical data (Linthurst et al., 1986) made the calculation of historical pH's, and thus pH decreases possible (Schindler et al., 1989); 2) the pH tolerances of fish species are similar for the Midwest and Northeast (U.S. EPA, 1985). Most fish and invertebrate species have similar pH tolerances in different areas (Engblom and Lingdell, 1984); 3) surveys of fish species (Scott and Crossman, 1973), selected large crustaceans (Bousfield, 1958; Dadswell, 1974) and crustacean zooplankton (Edmondson, 1959; Patalas, unpubl, data) show that species assemblages are quite similar in softwaters of temperate regions of the midwestern and northeastern U.S.A., as well as in eastern Canada. Therefore, we hypothesized that average declines in species could be assessed using models based on changes in the relation- ships between various chemical specis in lakes and summaries of the pH tolerances of various taxa. We used the U.S. EPA's Eastern Lake Survey data base as the basis for our initial modelling exercise. We have published a brief summary of this work elsewhere (Schindler et al., 1989). Here we discuss the variation in the model results among lakes and regions, and treat responses to different chemical assumptions more fully.
Methods
We assumed that inputs of sulfate above natural background were all as strong acids, and that they had one of two effects: to decrease alkalinity or to increase the concentrations of calcium and magnesium. The proportion of added sulfate that caused increases in Ca + Mg was designated F:
F = (Ca + Mg) / S O 4
following the convention of Henriksen (1980). Equations were developed to estimate the original alkalinity in lakes (ANCo) by iteratively solving the equation:
ANCo = SO4(1-F) -I- ANC c
where ANCc is the currently measured alkalinity, and delta S O 4 is the increase in sulfate over background. Independent solutions by varying values of F and SO~ were obtained for two areas where alkalinity losses had been measured empirically. In the Adirondacks, Asbury et al. (1989) measured median alkalinity losses of 50 tteq L -1 over several decades. In the Experimental Lakes Area, Schindler and Ruszczynski (1983) found no average decline in alkalinity over several years. The equations calibrated for these two empirical data sets were then applied to the subregions surveyed by EPA. Estimates for original pH were then based on ANC o and theoretical relationships, corrected for organic anions by the methods of Oliver et al. (1983). Full equations and tests of this procedure are given by Schindler e t al. (1989). Once the original pH and the difference between original and contem- porary pH were calculated, the diagrams of Eilers et al. (1984) (reproduced by Mills

LOSSES OF BIOTA FROM AMERICAN AQUATIC COMMUNITIES 273
and Schindler, 1986), relating pH tolerance to relative numbers of taxa in different biotic communities, were used to calculate the degree of relative biotic impoverish- ment that had taken place in each lake. The above procedure was applied to all sampled lakes in all subregions of the Eastern Lake Survey.
We restricted our analysis to lakes where (Ca + Mg) ~< 200 eq L - l , i.e. lakes that are very acid sensitive.
Results and Discussion
Estimated median alkalinity changes in various subregions ranged from - 55 to + 2 eq L - 1. Corresponding median pH changes ranged from - 0.73 to 0.01 (Schindler et
aL, 1989, Figures 1 and 2). In brief, large declines in alkalinity and pH were calculated for Subregions 1A, 1B, and 1D, the Adirondacks, Poconos-Catskills and southern New England, respectively; moderate declines for Central New England (1C) and Upper Michigan (2B), slight declines in Maine (1E), North Central Wisconsin (2C) and northeastern Minnesota (2A), and a slight increase in the remainder of the upper Great Lakes region (2D).
However, median values for the subregions do not tell the entire story. The responses of individual lakes varied widely within a given subregion. Predicted decreases in alkalinity for individual Adirondack and Poconos-Catskill lakes ranged from insignificant to over 100 e q / L - 1 (Figure 1, panels 1A and 1B). Corresponding pH decreases were from negligible to 2.5 pH units (Figure 2, panels 1A and 1B). In areas where deposition was less acidic, the model predicted small to moderate declines in alkalinity and pH for some lakes, but no change or even slight increases in others (Figures 1 and 2). These results are expected, due to natural variation in the chemistry of lakes and the size, soils and geology of their watersheds. In all subregions, the model predicted a large variation in the response of lakes.
Enormous median losses of species were indicated for acid-sensitive taxonomic groups in heavily-acidified areas. Fewer losses occurred in moderately acidified areas, and for less sensitive groups. Median values for all taxonomic groups and subregions were discussed by Schindler et al. (1989). However, as from chemical results, there was a considerable lake to lake variation in the degree of biological damage predicted by our model. For very sensitive groups like molluscs and leeches, losses of 80-100070 of species were predicted for the most sensitive lakes in heavily acidified areas like the Adirondacks and Poconos-Catskills, while the least sensitive lakes in the same areas were predicted to have suffered almost no impoverishment. In low deposition areas of Maine and the Midwest, the model predicted that very few lakes had lost biotic diversity. In all areas, the taxonomic groups least likely to be affected by acidification were those containing larger species of fishes (Figure 3; see also Schindler et al., 1989).
The above results indicate that impoverishment of aquatic communities and food chains is likely to have been severe in some areas, even though only slight to moderate direct damage to adult sport fishes is predicted. However, in view of the effect that

274 D . W . S C F I I N D L E R E T A L .
.o
I
_~
o
-o
s~o7 ;0
I I
I I
s ~ 0 7 ~0
I I
0 ~ 0 ~ 0 ~ 0
o ~ o ~ ~ ~ o
I I
o ~ o ~ o ~ o
~ 0 7 ~0
I I
I _ _
I I
I �9 ~ L_
o ~ o ~ o ~ o
~ z
~ z ~
~ 2 N
o ~
~,~ ~ ~'~
~ ~ . ~
~ o
~'~ ~
~ z ~
~ �9 ~ ~.~ ~ e ~ N
[,~ ~ ~
�9 ~ ~ ~ 0 ~

L O S S E S O F B I O T A F R O M A M E R I C A N A Q U A T I C C O M M U N I T I E S 2 7 5
I - - - .
-.
Lr~
?
T
?
?
o
L~
_?
Q _.~
J
?
I
i '
-- L I I
"T
' I
?
gh
..=
O
r
~q
c~
O
:8
O
r~ .o
e~
c-i

276 D. W. SCHINDLER ET AL.
IOO
80
eo o
2 0
o -16o
100"
BO"
60
_, i _ - 80 60 40 20 0
R o t i f e r s , x Chonge
1A
O_1oo -so -so -40 L~o o
M o l l u s c s , x Chonge
~ -~o -'=o -4o -20 Cypr in ids , x Chonge
100
80
o~ GO
~I~ 40
20 -17 V
0
~6
100
BD
60
100
BO
60 o _z ,
"6 ~= 4o"
-111
1
. . . . , I 8 0 60 40 20 0
Crus taceons , x Change
1A
20 -68
0 , r - ~ , I ~ - t O 0 - 8 0 -6O - r - 2 O ~
Leeches. x Chonge
100' 1A
BO
_~ 60 R
~ 4 0
2 0
_' _ ' ' , 0 _ 1 oo 80 60 40 - 2 0
Perc ids, x Chonge
o
r 0
Fig. 3. The frequency distribution of modelled declines in relative numbers of species of different taxonomic groups in lakes of the Adirondacks (Subregion 1A), one of the areas most heavily damaged by acid deposition, and in Maine (subregion IE), the least damaged subregion of the Northeast. Arrows indicate medians. Schindler et al. (1989) give median values for other subregions and taxonomic groups.
food chain degradation had on lake trout, the top predator in Lake 223 (Schindler et al., 1985; Mills et aL, 1987), damage to food chains must be suspected of causing decreased survival, production, and eventual disappearance of sport fishes, though perhaps with a time-lag of several years. This question of biological steady-state has not been addressed in any assessments of damage to ecosystems or of their recovery

L O S S E S O F B I O T A F R O M A M E R I C A N A Q U A T I C C O M M U N I T I E S 277
100
80
60
"6 40
20
~
lO0
80
6O
's 40
IE
, , ~ , ,F-- -150 =60 - 4 0 - 2 0
Rotifers, �9 Change
IE
2O
- 1 0 0 - 8 0 - 6 0 - 4 0 - 2 0
Molluscs, x Change
1E 100
80
6O
3
=~ 40
20
~ -~o 40 '40 -~o Cyprinids, x Change
lOO
0
80
~ 60
~ 40
r~ 3
1E
0_4o 4~ -6'o~-;o 40 Crustaceans. x Change
100 1E
8O
0 6C
4C - -
2O
0 , , - ~ o o - 8 0 ~6o - 4 0 - 2 ~
L e e c h e s , x Change
100
- - 80
:~ 40
20
0-1(~ 0
1E
- 4 0 - 2 0
P e r c ] d s , x Change
Fig. 3, (Continued)
from acidification. Clearly, biological steady-state must follow chemical steady- state, probably with a time lag of several years.
SENSITIVITY OF THE MODEL TO CHEMICAL ASSUMPTIONS
We believe that our model is based on conservative assumptions. These were discussed in detail by Schindler et aL (1989), and will not be repeated here. Instead, we explore the sensitivity of the model to variations in values of F and SO~, because

278 D. W. SCHINDLER ET AL.
while all authors agree that inputs of sulfuric acid must either cause increases in base cations or declines in alkalinity (Henriksen, 1982; Schindler et al., 1989), controversy remains regarding the proportion of incoming acid that causes one or the other of these two responses. Likewise, estimates of the pre-acidification concentrations of sulfate in lakes vary considerably. Some authors believe that background values of sulfate were quite high, especially in areas receiving windblown dust from western agricultural areas (Gorham et al., 1984). Others believe that background sulfate concentrations must have been no more than a few microequivalents per liter (Rogalla et al., 1986), as it is in remote mountainous areas of the western USA (Landers et al., 1987). A few runs of our model illustrate that few combinations of F and original sulfates would produce results that are consistent with the observed median rates of alkalinity loss in the Adirondacks and ELA (Figure 4). For the Adirondacks, F must have values between 0.3 and 0.5. If F is smaller, background values of SO4 must have been greater than 50 lteq L - 1, which is higher than would be consistent with values in remote areas with similar geology. For the Midwest, values of Fconverge at high background sulfate concentrations, ie. background SO4 must have been at least 50 Ixeq L - 1, or alkalinity losses would have to be much larger than those observed. Such high values are consistent with the observation that much of the SO4 in Minnesota precipitation is present as neutral salts, originating from prairie soils to the west (Gorham et al., 1984). In the Midwest, the Fvalue has little influence on model results at high values of SO4o. A review of North American data (Wright, 1983) and an experimental recovery of a watershed from acidification in Norway (Wright et al., 1988) also indicate that our F values are of the right average magnitude.
I M P R O V I N G T H E M O D E L
It is not possible to obtain exact species disappearances from given lakes, for there are few historical data for the communities of individual lakes. We can, however, improve the above model by being able to better define the habitat requirements for individual species. Minns et al. (submitted) have done this for several fish species in the province of Ontario, where detailed habitat requirements have been deduced from surveys of several thousand lakes. Using such information, they were able to calculate the probability that a certain fish species would be encountered in a lake of known characteristics, or that the species had been extirpated by acidification. To perform such an exercise requires biological surveys, detailed documentation of habitat characteristics such as shoreline development, connections to other aquatic ecosystems, mean and maximum depth, and oxygen concentrations at depth, in addition to chemical data such as those obtained by EPA. While such an exercise might seem like a massive and unglamorous undertaking, it would supply a background that would allow us to assess future losses of species, whatever the cause of such losses might be.
Of course, we have no way of knowing whether the populations already lost from acidified lakes would return if we reduced acid precipitation, or even whether they
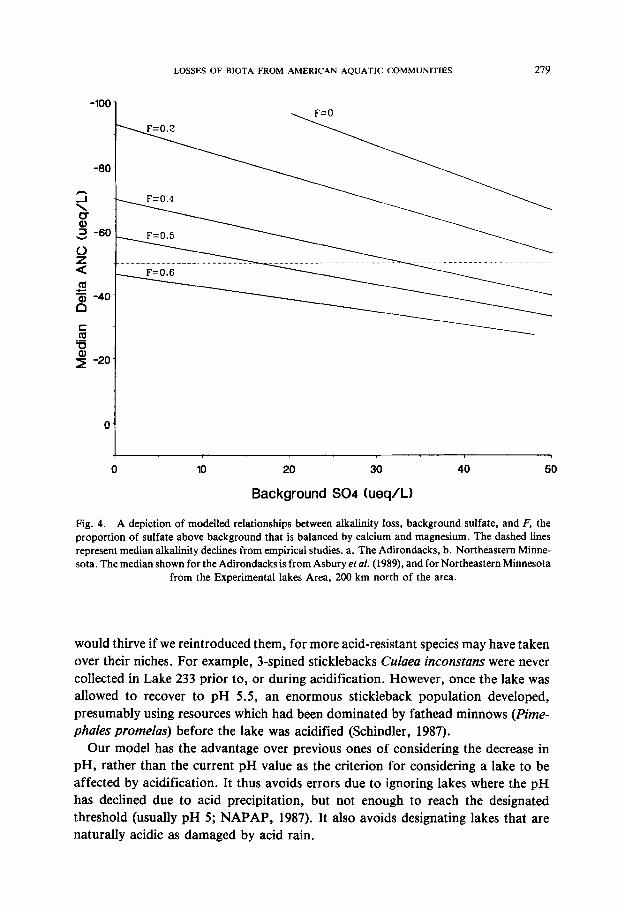
LOSSES OF BIOTA FROM AMERICAN AQUATIC COMMUNITIES 279
- 1 0 0
-80
._1
O"
-60
Z <
. . , , . . ,
r -40 a
c- to
. m
"0
:~ -20
0
0 10 20 30 40 50
Background S04 (ueq/L)
Fig. 4. A depiction of modelled relationships between alkalinity loss, background sulfate, and F, the proportion of sulfate above background that is balanced by calcium and magnesium. The dashed lines represent median alkalinity declines from empirical studies, a. The Adirondacks, b. Northeastern Minne- sota. The median shown for the Adirondacks is from Asbury et aL (1989), and for Northeastern Minnesota
from the Experimental lakes Area, 200 km north of the area.
would thirve if we reintroduced them, for more acid-resistant species may have taken over their niches. For example, 3-spined sticklebacks Culaea inconstans were never collected in Lake 233 prior to, or during acidification. However, once the lake was allowed to recover to pH 5.5, an enormous stickleback population developed, presumably using resources which had been dominated by fathead minnows (Pime- phales promelas) before the lake was acidified (Schindler, 1987).
Our model has the advantage over previous ones of considering the decrease in pH, rather than the current pH value as the criterion for considering a lake to be affected by acidification. It thus avoids errors due to ignoring lakes where the pH has declined due to acid precipitation, but not enough to reach the designated threshold (usually pH 5; NAPAP, 1987). It also avoids designating lakes that are naturally acidic as damaged by acid rain.

280
-100
-80
-60
<
~ -40
-20
D . W . S C H I N D L E R E T A L .
i , i i i
0 10 20 30 40 50
Background S04 (ueq/L)
Fig. 4. (Continued)
ARE LAKES AT BIOLOGICAL STEADY STATE 9.
One commonly-heard excuse for not rapidly controlling acidifying emissions is that lakes do not seem to be acidifying further, i.e. all of the chemical changes caused by deposition have already occurred. This may be true in some areas. However, lake acidification continues in others. Contrary to popular belief, acidification can occur even in areas where deposition has been reduced, if lakes have not reached, steady-state with high loading rates, or if base cations are becoming exhausted, as shown by Dillon et al., (1987). In other areas, increased nitric acid deposition and decreased base cation inputs in precipitation have compensated for decreases in H2SO4 to keep the acidity of precipitation from decreasing (Hedin et al., 1987; Driscoll et al., 1989). Increasing depletion of base cations from terrestrial soils has now been shown for Sweden (Hultberg, pers. comm.) and Hubbard Brook (Likens, pers. comm.). Such depletions may allow the continued acidification of lakes. In other cases, nitrate concentrations of lakes have been shown to increase dramatically in the past decade (NIVA, 1987). These changes are

LOSSES OF BIOTA FROM AMERICAN AQUATIC COMMUNITIES 281
inconsistent with the notion that lake acidity is at steady state with the acidity of
precipitation. Powerful evidence that lakes are not at steady-state also comes from paleoecologi-
cal studies. For example, in the Adirondacks, many of the lakes studied show continuing acidification (Whitehead, this conference; Charles, pers. com.; Charles
and Norton, 1986). Even after chemical equilibrium is reached, biological equilibria are expected to
lag. As our study of Lake 223, the above-referenced studies by Harvey, and studies by the Ontario Ministry of Natural Resources (Gunn, pers. comm.) show, extirpation of sport fishes may not occur for years to decades after reproduction ceases, due to the long life spans of larger fish species - up to several decades for lake trout. It therefore seems probable that many sport fish populations, if not entire commu- nities, might be recoverable if deposition is reduced soon. There are some recorded examples. Keller and Pitblado (1986) showed that acidity of some lakes in the Sudbury area had decreased enough to allow restocking of lake trout. In one case, a remnant trout population that had not reproduced for years began reproducing when pH values increased as the result of local emission controls (Gunn, pers. comm). Kelso et al. (1987) found that pH increases in the Algoma area were sufficient to allow white suckers to reinvade some lakes and reproduce. Recovery of Lake 233 to pH 5.4 allowed reproduction to resume in white sucker, pearl dace and three-spined stickleback - enough to cause the condition of emaciated lake trout populations to improve (Schindler, 1987). The entire cost of acid rain will not be known until we can assess the degree to which damaged ecosystems can be recovered, and this cost will certainly be higher if more systems reach biological steady-state, ie. all species that cannot reproduce at a given pH value have been eliminated.
SPECULATION ABOUT DAMAGE BY ACIDIFICATION TO OTHER AREAS
Results from the EPA's western lake survey indicate that there is little evidence of acidification (Landers et al. , 1987). Given the relative insensitivity of our model, it is likely that results for western areas would be similar to those in low deposition areas of the East, i.e. there would be little conclusive evidence for reductions in pH or alkalinity. Furthermore, the biota of the West is much different and more variable than that of the Northeast and Midwest. Many western ecosystems have also been stocked with exotic fish species. As a result, we did not attempt to apply our model to the western U.S.A.
In contrast, lakes of eastern Canada have biotic assemblages that are generally similar to those of the northeastern and midwestern U.S.A., although the number of species tends to decline as latitude increases. Canadian geology is even less heterogeneous than that o f the area modelled here, and deposition of strong acids in eastern Canada approaches values similar to those observed in the eastern U.S.A. However, lakes are much more numerous than in the U.S.A.: in fact, the exact number of lakes is not even known. Kelso et al. (1986) estimated that there are 700 000 lakes east of the Manitoba-Ontario border and south of 52 N latitude. If the

282 o . w . SCHINDLER ET AL.
few thousand lakes in the NAQUADAT data base are assumed to be representative, over 400 000 are acid-sensitive (Ca + Mg less than 200 eq L-1). In comparison, the U.S. regions 1 and 2 contain only 15732 lakes, with 4~ having Ca+Mg >200 eq L-~. Obviously, estimating effects of acidic deposition on Canadian lakes is a problem of enormous magnitude. Recent estimates indicate that in sensitive areas subjected to high rates of acid deposition, over 20~ of the taxa have disappeared from over 20o7o of the lakes. The affected areas are quite large, and the total damage to aquatic ecosystems is at least a hundred-fold greater than in the U.S.A. (Minns et ai., submitted).
IMPOVERISHMENT OF NORTHERN BIOTIC COMMUNITIES; A FORGOTTEN PROBLEM. 9
The number of species in a northern ecosystem is certainly much smaller than in the tropics, so that the number of species eliminated by acid rain or other human activities is probably small compared to that eliminated by tropical deforestation. However, from the standpoint of ecosystem ecology, there are other concerns. The functioning of ecosystems and food chains at northern latitudes is also dependent on fewer species than ones nearer to the Equator. As our study of Lake 223 shows, even deletions of one or two species can sometimes eliminate critical niches from northern aquatic food webs. Such deletions can also make ecosystems more vulner- able to invasion (Mooney and Drake, 1987), and more vulnerable to new stresses (Elton, 1958; Pimm, 1984). At pH values less than 5.6, a number of key biogeochemi- cal cycles and energy flows are also disrupted (Rudd et al., 1988; Turner et al., 1987 and in prep.; Schindler, in press). Altogether, when entire biotic communities and ecosystems over larger geographic areas are considered, damage from acidic precipi- tation to aquatic systems is enormous. While initial damage is confined to disappear- ance of senstitive species, at high rates of deposition there is reason for concern about the integrity of food webs and ecosystem functions. C. E. Asbury allowed us to use his results from the Adirondacks to calibrate our model.
Acknowledgements
This study was supported by the Canadian Department of Fisheries and Oceans. R. A. Linthurst and D. Landers kindly supplied a copy of the Eastern Lake Survey data, making this analysis possible.
References
Asbury, C. E., Vertucci, F. A., Mattson, M. D., and Likens, G. E.: 1989, 'Acidification of Adirondack Lakes', Environ. Sci. TechnoL 23, 362-365.
Beamish, R. J. and Harvey, H. H.: 1972, 'Acidification of the LaCioche Mountain Lakes, Ontario and Resulting Fish Mortalities', J. Fish. Res. Board Can. 29, 1131-1143.
Bell, H. L.: 1971, 'Effect of Low pH on the Survival and Emergence of Aquatic Insects', Wat. Res. 5, 313-319.
Bousfield, E. L.: 1958, 'Fresh-water Amphipod Crustaceans of Glaciated North America', Can. Field Nat. 72, 5-113.

LOSSES OF BIOTA FROM AMERICAN AQUATIC COMMUNITIES 283
Charles, D. F. and Norton, S. A.: 1986, 'Paleolimnological Evidence for Trends in Atmospheric Deposition of Acids and Metals', Ch. 9 in Acid Deposition: Long-Term Trends. National Academy Press, Washington D.C.
Dadswell, M. J.: 1974, 'Distribution, Ecology and Postglacial Dispersal of Certain Crustaceans and Fishes in Eastern Canada', Nat. Mus. Nat. Hist. Ottawa, Publ. ZooL 11, l l0.
Dillon, P. J., Yan, N. D., and Harvey, H. H.: 1984, 'Acid Deposition: Effects on Aquatic Ecosystems', CRC Reviews in Environmental Control. 13, 167-271.
Dillon, P. J., Yan, N. D., Scheider, W. D., and Conroy, N.: 1979, 'Acidic Lakes in Ontario, Canada: Characterization, Extent and Responses to Base and Nutrient Addition', Ergebn. LimnoL 13, 317-336.
Dillon, P. J., Reid, R. A., and de Grosbois, E.: 'The Rate of Acidification of Aquatic Ecosystems in Ontario, Canada', Nature 329, 45-48.
Driscoll, C. T., Likens, G. E., Hedin, L. O., Eaton, J. S., and Bormann, F. H.: 1989, 'Changes in the Chemistry of Surface Waters', Environ. Sci. Technol. 23, 137-143.
Edmondson, W. T.: 1959, Freshwater Biology. 2nd ed. Wiley, New York. Eilers, J. M., Lien, G. L., and Berg, R. G.: 1984, 'Aquatic Organisms in Acidic Environments: A
Literature Review', Wisc. Dep. Nat. Resour. Tech. Bull. 150, 18. Elton, C. S.: 1958, The Ecology o f lnvasions by Animals and Plants. Methuen, London. Engblom, E. and Lingdell, P.-E.: 1984, 'The Mapping of Short-term Acidification with the Help of
Biological pH Indicators', Rep. Inst. Freshw. Drottn. 61, 60-68. Forbes, S. A.: 1883, 'The First Food of the Common Whitefish', Bult. Illinois State Lab. Nat. Hist. 1,
95-109. France, R. L.: 1987, 'Reproductive Impairment of the Crayfish Orconectes virilis in Response to
Acidification of Lake 223', Can. J. Fish. Aquat. Sci. 44 (Suppl. 1), 97-106. Gorham, E., Martin, F. B., and Litzau, J. T.: 1984, 'Acid Rain: Ionic Correlations in the Eastern United
States, 1980-1981', Science 225, 407-409. Hall, R. J. and Ide, F. P. ' 1987, 'Changes in Aquatic Invertebrate Communities from 1937 to 1984:
Evidence of Impacts of Acidification on Stream Insects', Can. J. Fish. Aquat. Sci. 44, 1652-1657. Harvey, H. H.: 1980, 'Widespread and Diverse Changes in the Biota of North American Lakes and Rivers
Coincident with Acidification', in D. Drablos and A. Tollan (eds.), Ecological Impact o f Acid Precipitation. SNSF Project, Oslo, Norway, pp. 93-98.
Harvey, H. H. and Lee, C.: 1982, 'Historical Changes Related to Surface Water pH Changes in Canada', in R. E. Johnson (ed.), Acid Rain~Fisheries: Proceedings o f an International Symposium on Acidic Precipitation and Fishery Impacts in Northeastern North America. Am. Fish. Soc., Bethesda, MD, pp. 45-55.
Hedin, L. O., Likens, G. E., and Bormann, F. H.: 1987, 'Decrease in Precipitation Acidity Resulting From Decreased SO~-Concentration', Nature 325, 244-246.
Henriksen, A.: 1980, 'Acidification of Freshwaters - A Large Scale Titration', in D. Drablos and A. Tollan (eds.), Ecological Impact o f Acid Precipitation. SNSF Project, Norway, pp. 68-74.
Henriksen, A.: 1982, 'Susceptibility of Surface Waters to Acidification', in Proceedings, International Symposium on Acidic Rain and Fishery Impacts on Northeastern North America, Ithaca, New York, 2-5 Aug. 1981. American Fisheries Society, Bethesda, Md, pp. 103-121.
Keller, W. and Pitblado, J. R.: 1986, 'Water Quality Changes in Sudbury Area Lakes: A Comparison of Synoptic Surveys in 1974-76 and 1981-83', Water, Air, Soil Pollut. 29, 285-296.
Kelso, J. R. M., Minns, C. K., Gray, J. E. and Jones, M. L.: 1986, 'Acidification of Surface Waters in Eastern Canada and Its Relationship to Aquatic Biota', Can. Spec. Publ. Fish. Aquat. Sci. 87, 42.
Kelso, J. R. M. and Jeffries, D. S.: 1987, 'Response of Headwater Lakes to Varying Atmospheric Deposition in North-central Ontario', MS Great Lakes Fish. Res. Br., Sault Ste. Marie, ON.
Landers, D. H. et aL: 1987, 'Western Lake Survey, Phase 1 (WLS 1). Population Descriptions and Physico-chemical Relationships', U.S. Environmental Protection Agency. Environmental Research Laboratory, Corvallis, Ore. 176 pp.
Landers, D. H., Eilers, J. M., Brakke, D. F., Overton, W. S., KeUar, P. E., Silverstein, M. E., Schonbrod, R. D., Crowe, R. E., Linthurst, R. A., Omernik, J. M., Teague, S. A., Meier, E. P.: 1987, in Characte- ristics o f Lakes in the Western United States. Population Descriptions and Physico-Chemical Relation- ships; EPA-600/3-86/054a; U.S. Environmental Protection Agency: Washington, DC, Vol. I.

284 D. W. SCHINDLER El" AL.
Linthurst, R. A., Landers, D. H., Eilers, J. M., Brakke, D. F., Overton, W. F., Meier, E. P., Crowe, R. E.: 1986, 'Characteristics of Lakes in the Eastern United States. Volume 1', EPA-600/4-86/007a; U.S. Environmental Protection Agency, Washington, DC.
Mills, K. H.: 1984, 'Fish Population Responses to Experimental Acidification of a Small Ontario Lake', in G. R. Hendrey (ed.), Early Biotic Response to Advancing Lake Acidification. Acid Precipitation Series, Vol. 6, Ann Arbor Sci. Publ., Ann Arbor, MI., pp. 117-132.
Mills, K. M. and Schindler, D. W.: 1986, 'Biological Indicators of Lake Acidification', Water, Air, Soil Pollut. 30, 779-789.
Mills, K. H., Chalanchuk, S. M., Mohr, L. S., and Davies, I. J.: 1987, 'Responses of Fish Populations in Lake 223 to 8 Years of Experimental Acidification', Can. J. Fish. Aquat. Sci. 44 (Suppl. l), 114-125.
Minns, C. K., Moore, J. E., Schindler, D. W., and Jones, M. L.: 1989, 'Assessing the Potential Extent of Damage to Inland Lakes of Eastern Canada Due to Acidic Precipitation. IV. Predicting the Response of Potential Species Richness in Several Taxa of Aquatic Biota' (submitted).
Mooney, H. A. and Drake, J. A.: 1987, 'Biological Invasions', Environment 29, 10-37. NAPAP (U.S. National Atmospheric Precipitation Program): 1987, 'Executive Summary and Mid-term
Findings'. Nero, R. W. and Schindler, D. W.: 1983, 'The Decline of Mysis relicta during the Acidification of Lake
223', Can. J. Fish. Aquat. Sci. 40, 1905-1911. NIVA: 1987, 'I000 Lake Survey 1986 Norway', Norwegian Institute for Water Research, Oslo, Norway. Okland, J. and Okland, K. A.: 1980, 'pH Level and Food Organisms For Studies of 1000 Lakes in
Norway', in D. Drablos and A. Tollan (eds.), Ecological Impact of Acid Precipitation. SNSF Project, Oslo, Norway, pp. 326-327.
Okland, J. and Okland, K. A.: 1986, 'The Effects of Acid Deposition on Benthic Animals in Lakes and Streams', Experimentia 42, 471-486.
Oliver, B. G., Thurman, E. M., and Malcolm, R. L.: 1983, 'The Contribution of Humic Substances in Forested Watersheds of the Adirondack Park, New York', Geochim. et Cosmochim. Acta 47, 2031-2035.
Pimm, S. L.: 1984, 'The Complexity and Stability of Ecosystems', Nature 307, 321-326. Raddum, G. G.: 1980, 'Comparison of Benthic Invertebrates in Lakes with Different Acidity', in D.
Drablos and A. Tollan (eds.), Ecological Impact o f Acid Precipitation. SNSF Project, Oslo, Norway. Rogalla, J. A., Brezonik, P. L., and Glass, G. E.: 1986, 'Empirical Models for Lakes Acidification in
the Upper Great Lakes Region', Water, Air, Soil Pollut. 31, 95-100. Rudd, J. W. M., Kelly, C. A., Schindler, D. W., and Turner, M. A.: 1988, 'Disruption of the Nitrogen
Cycle in Acidified Lakes', Science 240, 1515-1517. Schindler, D. W.: 1987, 'The Recovery of Canadian Lakes From Acidification', in Proceedings o f
Workshop on Reversibility o f Acidification, Grimstad, Norway, 9-11 June 1986, pp. 11-22. Schindler, D. W.: 1988, 'Effects of Acid Rain on Freshwater Ecosystems', Science 239, 149-157. Schindler, D. W., Mills, K. H., Malley, D. F., Findlay, D. L., Shearer, J. A., Davies, I. J., Turner, M.
A., Linsey, G. A., and Cruikshank, D. R.: 1985, 'Long-Term Ecosystem Stress: The Effects of Years of Acidification on a Small Lake', Science (Wash., DC) 228, 1395-1401.
Schindler, D. W. and Ruszczynski, T.." 1983, 'A Test of Limnological Data From the Experimental Lakes Area, Northwestern Ontario, For Evidence of Acidification', Can. Fish. Mar. Serv. Tech. Rep. 1147, iv + 17 pp.
Schindler, D. W., Kasian, S. E. M., and Hesslein, R. H.: 1989, 'Estimating Biological Damage to Lakes of the Midwestern and Northeastern U.S.A. From Acid Rain' , Environ. Sci. TechnoL 23, 573-580.
Scott, W. B. and Crossman, E. J.: 1973, Freshwater Fishes o f Canada. Fish. Res. Board Can. Bull. 184, 966.
Shearer, J. A. and De Bruyn, E. R.: 1987, 'Phytoplankton Productivity Responses to Direct Addition of Sulfuric and Nitric Acids to the Waters of a Double-basin Lake', Water, Air, SoilPollut. 30, 695-702.
Turner, M. A., Jackson, M. B., Findlay, D. L., Graham, R. w. , De Bruyn, E. R., and Vandermeer, E. M.: 1987, 'Early Responses of Periphyton to Experimental Lake Acidification', Can. J. Fish. A quat. Sci. 44 (Suppl. 1), 135-149.
U.S. EPA: 1985, 'The Acid Deposition Phenomenon and Its Effects', EPA-600/8-83-0-6B. Watt, W. D., Scott, C. D., and Ray, S.: 1979, 'Acidification and Other Chemical changes in Halifax
County Lakes After 21 Years', Limnol. Oceanogr. 24, 1154-1161.

LOSSES OF BIOTA FROM AMERICAN AQUATIC COMMUNITIES 285
Watt, W. D., Scott, C. D., and White, W. J.: 1983, 'Evidence of Acidification of Some Nova Scotia Rivers and Impact on Atlantic Salmon, Salmo salar" Can. J. Fish. Aquat. Sci. 40, 462--473.
Wilson, E. O.: 1985, 'The Biological Diversity Crisis', Bioscience 35, 700-706. Wright, R. F.: 1983, 'Predicting Acidification of North American Lakes', NIVA Report 0-81036,
Norwegian Institute for Water Research, Oslo, Norway. Wright, R. F., Lotse, E., and Semb, A.: 1988, 'Reversibility of Acidification Shown by Whole-catchment
Experiments', Nature 334, 670--675.