low ph up‐regulates interleukin‐6 mrna in l6‐g8c5 rat
TRANSCRIPT

FASEB BioAdvances. 2021;00:1–15. | 1www.fasebbioadvances.org
Received:17July2021 | Revised:14October2021 | Accepted:22October2021
DOI:10.1096/fba.2021-00088
R E S E A R C H A R T I C L E
Low pH up- regulates interleukin- 6 mRNA in L6- G8C5 rat skeletal muscle cells independent of pH sensing by SNAT2(SLC38A2) transporters
Ziyad Aldosari1,2 | Nima Abbasian3 | Katherine Robinson4 | Alan Bevington1 | Emma Watson4
ThisisanopenaccessarticleunderthetermsoftheCreativeCommonsAttributionLicense,whichpermitsuse,distributionandreproductioninanymedium,providedtheoriginalworkisproperlycited.©2021TheAuthors.FASEB BioAdvancespublishedbyTheFederationofAmericanSocietiesforExperimentalBiology.
1DepartmentofRespiratorySciences,UniversityofLeicester,Leicester,UK2DepartmentofMedicalLaboratoriesSciences,CollegeofAppliedMedicalSciencesinAlquwayiyah,ShaqraUniversity,Riyadh,SaudiArabia3SchoolofLifeandMedicalSciences,UniversityofHertfordshire,Hatfield,UK4DepartmentofCardiovascularSciences,UniversityofLeicester,Leicester,UK
CorrespondenceZiyadAldosari,19257Alrabwah,Alquwayiyah,Riyadh,SaudiArabia.Email:[email protected]
Funding informationShaqraUniversity
AbstractExercise isknown tocreatea transient,butpotent increase in skeletalmuscleexpression of potentially anti-inflammatory myokine interleukin-6 (IL-6). Thiseffectmaybeclinically important inmanagingchronic inflammatorystates. Ithas previously been proposed that lactic acidosis following exercise promotesthis IL-6up-regulation,but themechanismof thisacidosiseffect isunknown.RatskeletalmusclecelllineL6-G8C5 hasbeenusedpreviouslytomodelmeta-boliceffectsofacidosis,sensinglowpHthroughtheresultinginhibitionofaminoacid transporter SNAT2(SLC38A2). Use of ionophore ionomycin to model theriseinintracellularCa2+concentrationoccurringincontractingmusclestronglyup-regulates IL-6 mRNAinL6-G8C5 myotubes.Thisstudyused thismodel totestthehypothesisthatlowextracellularpH(7.1)enhancesionomycin-inducedIL-6 mRNA up-regulation by inhibiting SNAT2. Incubation of L6-G8C5 myo-tubes for 6 h with 0.5 µM ionomycin at control pH (7.4) resulted in a 15-foldincreaseinIL-6 mRNAwhichwasfurtherenhanced(1.74-fold)atpH7.1.Incon-trast low pH had no significant effect on IL-6 mRNA without ionomycin, norontheIL-6 mRNAincreasethatwasinducedbycyclicstretch.EventhoughpH7.1 halvedthetransportactivityofSNAT2,alternativemethodsofSNAT2inhi-bition(JNKinhibitorSP600125;SNAT2antagonistMeAIB;orSNAT2 silencingwithsiRNA)didnotmimictheenhancingeffectoflowpHonIL-6 mRNA.Onthe contrary, JNK inhibition blunted the effect of pH 7.1 with ionomycin, buthadnoeffectatpH7.4.ItisconcludedthatlowpHpromotesCa2+/ionomycin–inducedup-regulationofIL-6 mRNAthroughanovelSNAT2-independentJNK-dependentpH-sensingpathwaynotpreviouslydescribedinthisskeletalmusclemodel.
K E Y W O R D S
interleukin-6 mRNA,JNK,pH,skeletalmuscle,SLC38A2,SNAT2

2 | ALDOSARI et al.
1 | INTRODUCTION
Inhumans,moderatephysicalexerciseisreportedtoexertsignificant anti-inflammatory effects,1 in contrast withthepro-inflammatoryeffectsofintenseexercise.2Forthisreason, there is currently considerable interest in utilis-ingmoderateexercisetherapeuticallyindiseasesthatareassociated with chronic inflammation, notably diabetesmellitus, cardiovascular disease, chronic interstitial lungdiseasesandchronickidneydisease.Animportantphys-iologicalresponsetoacuteexerciseinhealthyhumansistheresultingtransientincreaseintheconcentrationofthecytokineinterleukin-6(IL-6)locallywithinmusclewhicharisesprincipallyfromincreasedexpressionandsecretionofIL-6fromthemyocytesthemselves,resultinginasys-temic increase in the IL-6 concentration. ConsequentlyIL-6isregardedasafunctionallyimportantmyokine.3InadditiontothemetaboliceffectsofthissystemicriseinIL-6,notablyincreasedlipolysisinadiposetissue4andincreasedhepatic glucose output,5,6 there is evidence that IL-6 isanti-inflammatoryintheabsenceoftumornecrosisfactora(TNF-α)7actingprincipallybyincreasingthecirculatingconcentrationoftheanti-inflammatorycytokineinterleu-kin-108 and by increasing the concentration of interleu-kin-1receptorantagonist8andsolubleTNF-αreceptor.9Ithasbeenshown1thatresistanceexercisewithoutmuscledamagestronglyup-regulatesIL-6 mRNAexpressioninthemyocytesasanadaptiveresponse(independentofTNF-α7),leading to acute anti-inflammatory effects3 that are inmarked contrast with the pro-inflammatory effects ofIL-6 that are observed when it is released from infiltrat-ing leukocytes in conjunction withTNF-α as an integralpartoftheacuteinflammatoryresponse.1,10Thisadaptiveresponsefollowingmoderateexercisealsocontrastswiththe prolonged maladaptive IL-6 burden that is observedin chronic inflammatory diseases such as cardiovasculardisease.11
Exercise sufficient to deplete glycogen and increaseglycolyticfluxinmuscleisnecessarytoelicitastrongIL-6response,12,13andithasbeenproposedthatthelacticac-idosisresultingfromsuchexercisemaybeanimportantcontributortothisIL-6up-regulation.14Inspiteofinitialevidencetothecontraryinthespecialcaseofmitochon-drial myopathy patients,15 the most recent evidence16suggeststhat lowpHarisingfromlacticacidosismaybesufficienttostimulateIL-6expressionfollowingexercise.
It is therefore important to understand the molecu-larbasisoftheacuteeffectsofextracellularpHonskele-talmusclecells.Inpreviousstudiesofthechroniceffectsofmetabolicacidosisonmuscleproteinmetabolism, theL6-G8C5 sub-cloneoftheL6ratskeletalmusclecell lineprovedtobeausefulmodel;showingaglobaldecreaseinproteinsynthesis,17anincreaseinglobalproteolysis18and
netproteinwasting incultures thathadbeenexposed toa lowextracellularpHof7.1 forup to72 h19; consistentwith thecachexia that isobserved invivo in response tochronicmetabolicacidosisindiseasestatessuchaschronickidneydisease.20Inthisculturemodel,thechroniceffectsoflowextracellularpHweremediatedbythepHsensitiveSystem A amino acid transporters in the plasma mem-brane.21 These transporters are solute carrier proteins(SLCs)whichusetheelectrochemicalgradientofNa+ionsacross theplasmamembrane todriveactive transportofneutralaminoacidssuchasl-glutamineintothecells.22,23Key biochemical characteristics of these transporters arefirstly their strong inhibition by low extracellular pH (atleastpartlycausedbydirectprotonationoftheextracellu-larN-terminusofthetransporterprotein)24;andsecondlytheirabilitytotransportthesyntheticaminoacidmethyl-aminoisobutyrate (MeAIB).25 System A transporters areencodedbygenesoftheSLC38 solutecarrierfamily.26ThemostwidelyexpressedmemberofthisfamilyisSLC38A2—whichisalsoknownassodium-coupledneutralaminoacidtransporter2,SNAT2,SAT2,orATA2andwillbere-ferredtointheremainderofthispaperasSNAT2.Itisex-pressed in skeletal muscle27 and is the dominant SystemA transporter of L6-G8C5 cells.17,18 Inhibition of SNAT2transport activity by low extracellular pH in these cellsleadstodepletionofintracellularfreeaminoacidswhichissensedbythemTORC1 signallingcomplex,resultinginim-pairedproteinsynthesis.17HoweverlowpHactingthroughSNAT2alsoimpairsanabolicsignallingthroughPI3K/Aktresulting in enhanced proteolysis,18 probably through aso-called “transceptor” mechanism28 independent of thetransportofaminoacidsbySNAT2.Theseeffectsof lowpHonsignallingandproteinmetabolismareallmimickedbyselectivesilencingofSNAT2 geneexpressionbyusingsmallinterferingRNAs,17,18confirmingtheimportanceofSNAT2inpHsensinginthesecells.Asimilarmechanismhas been proposed to explain the bicarbonate-sensitivityof the depletion of intramuscular free amino acids andchanges in intracellular signalling that follow enduranceexerciseinpatientswithchronickidneydisease.29
The acute increase in secretion of IL-6 from skeletalmuscle following exercise is largely driven by increasedIL-6 mRNAexpressioninthemyocytesandhaspreviouslybeenmodelled invitro in theL6ratskeletalmusclecellline by applying a rise in the intracellular Ca2+ concen-tration(likethatobservedincontractingmuscle)byuseoftheionophoreionomycin.13Itwasshowninthatstudythattheionomycin-dependentsignaltoIL-6 mRNAwasmediatedbyanuclearpoolofthekinasep38 MAPK.Inview of SNAT2’s previously demonstrated ability in L6-G8C5 cells to mediate pH-dependent signalling to theubiquitin-proteasome pathway gene expression eventsthatregulateproteolysis,18thiscelllineseemsapromising

| 3ALDOSARI et al.
experimentalmodelinwhichtotestthefollowinghypoth-esis.LowextracellularpH(asinskeletalmusclefollowingexercise)up-regulates IL-6 mRNAby inhibiting thebio-logicalactivityofSNAT2(eitherbyinhibitingitsactivityasatransporteroritsabilitytoactasatransceptor).
Theexperimentalaimsofthisstudyweretherefore:
1. ToconfirmthattheL6-G8C5celllineshowsthestrongIL-6 mRNAresponsetoionomycinpreviouslyreportedin L6 cells.13
2. Todemonstratethatthiscelllineshowsup-regulationof IL-6 mRNA at low pH, consistent with recent evi-dence that acidosis can increase IL-6 expression andsecretion.16
3. To investigate (by transport inhibition or by gene si-lencingofSNAT2)theroleofthistransporterinmedi-atingpHeffectsonIL-6 mRNA.
2 | MATERIALS AND METHODS
2.1 | Materials
Tissue culture medium and supplements were obtainedfrom Invitrogen, Paisley, UK. Drugs and biochemicalswere obtained from Sigma-Aldrich unless otherwisestated. Ionomycin, P38 MAPK inhibitor SB202190 andJNKinhibitorSP600125weredissolvedindimethylsulph-oxide (DMSO)beforeaddition toexperimentalmedium.ThefinalresultingconcentrationofDMSOinthemedium(up to 0.3% vol/vol) was also added to control cultures.Drugswereapplied toculturesatpharmacologicallyac-tivefinalconcentrationsandincubationtimeswhichhadpreviously been validated for ionomycin,13 SB20219030and SP600125.31 Fluo-4 AM esterified Ca2+ indicatorwasobtainedfromFisherScientificandwasdissolvedinDMSO.ThefinalresultingconcentrationofDMSOinthemediumduringFluo-4experimentswas1%vol/vol.
3 | METHODS
3.1 | Cell culture
L6-G8C5,asub-cloneof theratskeletalmusclecell lineL632wasobtainedfromtheEuropeanCollectionofAnimalCellCultures(ref.9212111)andwasusedatpassagenum-ber 10–20. Stock cultures were periodically screened toconfirm absence of mycoplasma using an EZ-PCR my-coplasmaPCRdetectionkit (GENEFLOW)according tothe manufacturer's instructions. Cells were propagatedat37°Cundera5%CO2 atmosphereinGrowthMedium
(GM) comprising Dulbecco's Modified Eagle Medium(DMEM-Invitrogen,Paisley,UKref.11880)with5 mMD-glucoseandpyruvate,supplementedwith10 mg/Lphe-nol red (Sigma-Aldrich),100 U/mlpenicillinG,100 µg/mlstreptomycin,2 mMl-glutamineand10%vol/volheatinactivatedfoetalbovineserum(FBS).After72 hthecon-fluent myoblasts were fused to form myotubes by incu-batinginFusionMediumcomprisingMinimumEssentialMedium (MEM) (Invitrogen ref. 21090) supplementedwith 100 U/ml Penicillin G, 100 µg/ml Streptomycin,2 mM l-glutamine and 2% vol/vol FBS. Fresh FusionMediumwasaddedafter2 days.Afterafurther2 daysthemyotubeswereusedforexperimentalincubations.
For experiments that required high transfection effi-ciency (siRNA silencing) or rapid uptake of dye (Fluo-4Ca2+ analysis), unfused myoblast cultures were used asdescribedintherelevantsectionsbelow.
3.2 | Medium for experimental incubations
Experimental incubations were performed in modifiedFusionMediuminwhichFBSwasreplacedwith2%vol/volheat-inactivatedDialysedFoetalBovineSerum(DFBS,Invitrogen ref. 26400). To achieve a physiological con-trolpHof7.4undera5%CO2 atmosphere,anadditional8 mmoles of NaHCO3 was added per litre of medium.19Forculturesunderacidicconditions (pH7.1) the8 mMNaHCO3 supplementwasreplacedwithequimolarNaCltomaintainaconstantNa+concentration,and6.3 mmolesofHClper litreofmedium.ThisacidicextracellularpHof7.1waschosenbecauseitissufficienttohalvetheac-tivity of SNAT2 transporters in L6-G8C5 cells.17 DuringexercisebloodpHcanfallto7.32inhumans33andmeas-urementsinworkingdoggastrocnemiusindicatethatthecorresponding pH in muscle interstitial fluid (to whichthemuscleSNAT2transporterproteinisdirectlyexposed)canfallto7.1.34
3.3 | Cyclic stretch by vacuum suction
Myotubes on type I collagen-coated Bio-flex stretchable35 mm diameter culture plates were subjected to cyclicstretching on the vacuum manifold of a Flexercell FX-4000™ Strain Unit (Flexcell International Corporation)usingtheprotocoldescribedin.35Eachstretching/releasecyclecompriseda2-ssinewavestretchresulting in18%maximumelongation, followedbya4-s release. Ineachexperiment,parallelmyotubeculturesonFlexcellplatesweresimultaneouslyincubatedonthesamevacuumman-ifold,butwiththevacuumorificeunderthewellssealed

4 | ALDOSARI et al.
withmaskingtapetopreventingstretching.Thisservedasanegativecontrolforanynon-specificeffectsarisingfrommountingtheFlexcellplateinthevacuummanifold.
3.4 | Transport activity assay (14C MeAIB uptake measurement)
To assess the activity of SNAT2 transporters, the rateof transport of selective System A substrate α-[1-14C]-methylaminoisobutyricacid(14CMeAIB;NEN-DuPont/PerkinElmer;3.70 MBq/ml;specificradioactivity50pCi/pmol))wasmeasured in intactcells.After incubationofthecellson22 mmculturewellsinexperimentalmedia,the cells were rinsed twice with 1 ml of Hepes-bufferedsaline(HBS)comprising140 mMNaCl,2.5 mMMgSO4,5.0 mM KCl, 1.0 mM CaCl2, 10 mg/L phenol red, and20 mMHepesacid titratedwith0.5 MNaOHtopH7.1orpH7.4atroomtemperature.Then500 µlofHBSattherequired pH was added to each well. The transport ex-perimentwasstartedbytheadditionofanaliquotof14CMeAIB,sufficienttogiveafinalconcentrationof10 µM14CMeAIB(20 kBq/ml)intheculturewell.Insomecul-turewells,10 mMunlabelledMeAIB(Sigmaref.M-2383)wasalsopresent inthemediumasanegativecontrol toassessnon-specificbindingof14CMeAIBinthewell.Thecellswereincubatedwith14CMeAIBatroomtemperature(25°C)forexactly5 min.Thentransportwasimmediatelyhalted by placing the culture plate on ice. The mediumwasaspiratedandthewellswerewashedthreetimeswith1 ml of ice-cold 0.9% w/v NaCl to remove extracellularradioactivity, followedbyscrapingof thecellmonolayerin 200 µl of 0.05 M NaOH and then digestion at 70°Cfor30 min.TheresultinglysatewascountedonanLKB1219 liquid scintillationcounterwithquenchcorrectiontodeterminedisintegrationsperminute(dpm).The14C-MeAIBdpmcountinthenon-specificbindingcontrolcul-tureswassubtractedfromthecountintheotherculturestodeterminenet14C-MeAIBtransportintothecells.
3.5 | Immunoblotting and assay of IL- 6 protein
After experimental incubations, whole cell lysates wereimmediately prepared by homogenising cells in lysisbuffer36comprising10 mMbeta-glycerophosphate,1 mMEDTA,1 mMEGTA,50 mMTris-HClpH7.5,1 mMso-diumorthovanadate,50 mMsodiumfluoride,1 mMben-zamidine,0.2 mMphenylmethylsulphonylfluoride,1 µg/mlpepstatinA,1 µg/mlleupeptinhemisulphate,0.1%vol/vol beta-mercaptoethanol, and 1% vol/vol Triton X-100detergent.
Proteins in cell lysates (30 µg protein per lane) wereseparated by SDS-PAGE. To ensure uniform loading ofthelanes,totalproteininlysateswasmeasuredusingtheBioRadDetergentCompatibleproteinassay(BioRad)andthe total protein concentration was adjusted to achieve30 µgper lanebyaddinglysisbuffer.Separatedproteinswere blotted onto Hybond ECL 0.45 µm nitrocellulosemembranes (GE Healthcare).The uniformity of loadingandtransferonallmembraneswascheckedbyPonceauRedstainingandvisualinspection.Membraneswerethenprobed with primary poylclonal rabbit antibody againstPThr180/Tyr182p38(Promega)ormousemonoclonalantibodyagainst p38-α MAP Kinase (New England BioLabs) orrabbitmonoclonalantibodyagainstJNK2(NewEnglandBioLabs). Bound primary antibody was detected usinghorseradish peroxidase (HRP)–conjugated polyclonalgoat anti-rabbit or goat anti-mouse immunoglobulins(DakoCytomation)asappropriate.HRP-labelledproteinswere detected by chemiluminescence using SuperSignalWest Pico Chemiluminescent Substrate (Thermo FisherScientific). Band intensities were quantified using aChemiDoc™TouchImagingSystemwithImageLabsoft-warev5.2.1(Bio-Rad).
ForassayofsecretedIL-6protein,mediumwasretainedfromculturesattheendofexperimentalincubationsandstoredat−80°CuntilitwasassayedusingaselectiveratIL-6Enzyme-linkedImmunosorbentAssay(Bio-Techne).
3.6 | RNA techniques
After experimental incubations, total RNA was ex-tractedfromL6-G8C5cellsusingTrizol®reagent(FisherScientific).From1 µgoftotalRNA,cDNAwassynthesisedusinganAMVReverseTranscriptionSystem(Promega)accordingtothemanufacturer'sinstructions.
Real-time qPCR was performed using an AppliedBiosystems 7500 Fast Real-Time PCR System (AppliedBiosystems/Thermo Fisher Scientific), with optimisedgenespecificamplificationprimersandprobes(TaqMangeneexpressionassays,AppliedBiosystems)asfollows:
RatIL-6-assayIDnumberRn01410330_m1RatSNAT2-assayIDnumberRn00710421_m1RatTBP(TATA-boxbindingprotein)(assayIDnumber
Rn01455646_m1)wasusedasahouse-keepinggene.37
The expression of IL-6 and SNAT2 mRNA was nor-malisedtothecorrespondingTBPsignalforeachsampleandrelativeexpressionispresentedas(2−ΔΔCT).38
SNAT2 genesilencingwasconductedinL6 myoblastsas previously described17,18 using custom-synthesisedvalidated siRNA oligonucleotides (Eurogentec). It haspreviously been confirmed in L6-G8C5 myoblasts thatsiRNAsilencingofSNAT2 mRNAleadstocommensurate

| 5ALDOSARI et al.
silencing of SNAT2 protein detected by SNAT2 Westernblot(figure 4inRef.[17]).
The SNAT2 silencing siRNA (SIL) had a forward se-quence of 5’-CUGACAUUCUCCUCCUCGUdTdT whichwas directed against base position 1095 onwards in theSNAT2 gene sequence.17,18 To control for possible non-specificfunctionaleffectsofsiRNAtransfection,parallelcontrol transfections were performed using a previouslyvalidated scrambled siRNA control (SCR) (forward se-quence 5’-CGCUCAACUCUACUUGUCCdTdT) thatshared the same overall base composition as SNAT2SIL.17,18
L6 myoblastswereseededinGMat105cellsper35 mmculturewell.Afterovernightincubation,theGMwasdis-cardedand1 mloffreshGMwasaddedtothewellsandthe cells were transfected with anti-SNAT2 SIL siRNAor SCR control using a Profection Calcium PhosphateTransfection Kit (Promega). The final concentration ofsiRNAintheculturemediumwas30 nM.Thecellswereincubated in a culture incubator at 37°C under a 5%CO2 atmospherefor16 h,afterwhichthetransfectionme-diumwasaspiratedand4 mlof freshGMwasadded toeachwellandculturedforafurther24 h,afterwhichthecultureswereusedforexperimentalincubations.
3.7 | Measurement of intracellular Ca2+ using Fluo- 4 fluorescence
FreeintracellularCa2+inL6 myoblastswasassayedfluo-rometrically using the Ca2+-sensitive dye Fluo-4.39 Ca2+dependent fluorescence was measured on a NOVOstarfluorescence plate-reader (BMG LABTECH). Myoblastswere seeded at 5 × 104 cells per well on poly-D-lysinehydrobromide-treated 6 mm plastic wells in 60 µl ofGMandwereculturedfor18 hbeforecommencingdye-loadingexperiments.Afterthat,theGMwasgentlyaspi-ratedfromeachwellwhichwasthenrinsedwith200 µlofPhysiologicalSaltSolution40comprising145 mMNaCl,10 mMHepesacidtitratedwith0.5 MNaOHtopH7.4atroomtemperature,1.0 mMMgSO4,3.0 mMKCl,2.0 mMCaCl2and10 mMD-glucose.ThecellswerethenloadedwithFluo-4-AMester(FisherScientific)atafinalconcen-trationof2 μMinPSSsupplementedwithbovineserumalbumin(0.1%w/v)(PSS-BSA)41for45 minat37°Cinahumidified incubator. After that, the wells were rinsedthree timeswith200 µlofPSS-BSA,and200 µlof freshPSS-BSAwasaddedfollowedbyincubationforafurther45 minat37°CtoallowFluo-4AMtobede-esterifiedtothefreeFluo-4dye.Thefluorescenceintensitywasthenreadat37°Catanexcitationwavelengthof488 nm,de-tectingat>500 nm.CalibrationoftheFluo-4 signalwasperformed by adding a final concentration of 1.0 µM
ionomycin Ca2+ ionophore in the presence of PSS-BSAcontaining 4 mM CaCl2 to equilibrate the intracellularandextracellularconcentrationofCa2+.Thefluorescencesignalwasallowedtostabilise for10 minandthemeanstabilisedsignalwascalledFmax.Then(inthepresenceofionomycinasabove),thecellswererinsedwithPSS-BSA+2 mMEGTAtochelateCa2+andthefluorescencesignalwasagainallowedtostabilisefor10 min.Themeanstabi-lisedsignalwascalledFmin.
The intracellularCa2+ concentrationwhichyieldedafluorescencesignalFwascalculatedusingtheformula:
wherethedissociationconstantKdoftheFluo-4-Ca2+com-plexwastakentobe350 nM.42
3.8 | Statistical analysis
Data are presented from at least three independent ex-periments,andwereanalysedusingGraphPadPrism7.0.Normallydistributeddataarepresentedinfiguresasthemean ± SEMandinthetextasmeanfoldchange ± SEM,andwereanalysedusingStudent'st-testor(fordatasetsinvolving multiple comparisons) repeated measuresANOVAfollowedbyposthoc testingwithTukey'smul-tiplecomparisonstest.Non-normallydistributeddataarepresentedinfiguresaslinkeddatacurvesfortheindivid-ualexperimentsandinthetextasmedianfoldchangeandrange. These were analysed using Friedman's nonpara-metricrepeatedmeasuresANOVA,followedbyposthoctestingwithDunn'smultiplecomparisonstest.Statisticalsignificancewasacceptedatp < 0.05.
4 | RESULTS
4.1 | Ionomycin's action on IL- 6 mRNA is enhanced by low pH
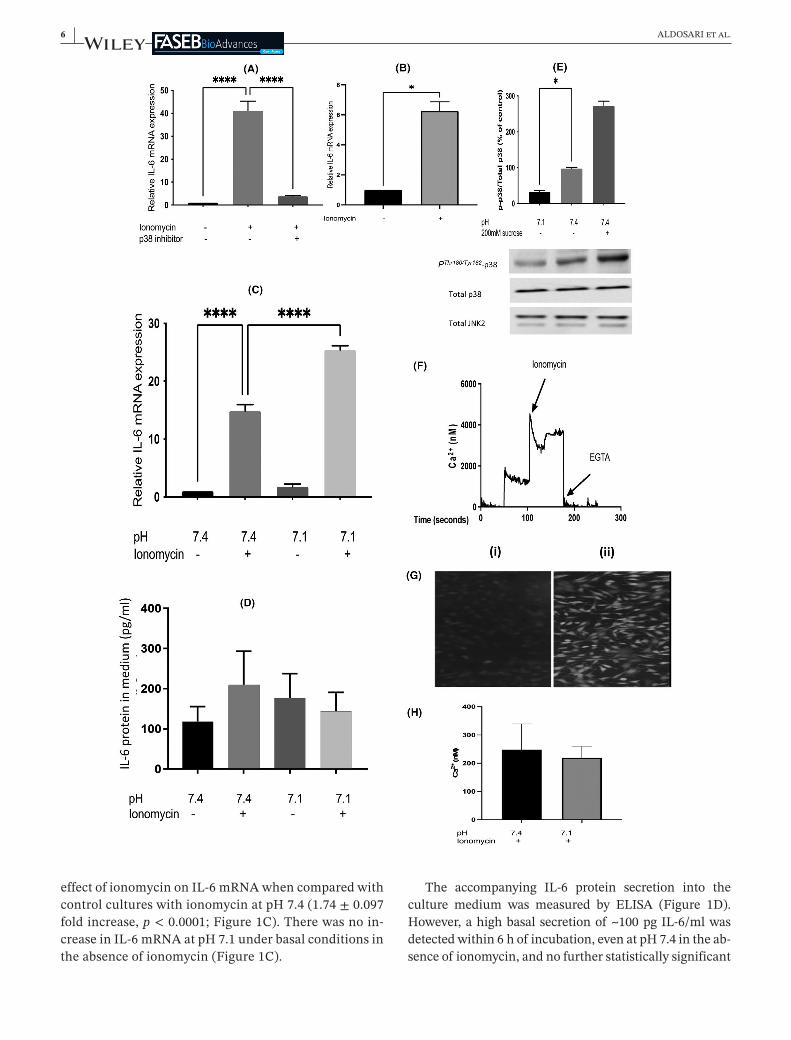
Inagreementwiththeearlierreport,13treatmentofL6-G8C5ratmyotubesfor6 hwithionomycinledtoanin-crease43inIL-6 mRNA(41 ± 6foldincrease,p < 0.0001in Figure 1A and 15 ± 1 fold increase, p < 0.0001 inFigure 1C) which was almost abolished by pharma-cological blockade of p38 MAP kinase with the selec-tive inhibitor SB202190 (0.085 ± 0.005 fold decrease,p < 0.0001)(Figure 1A).Modellingofthemetabolicef-fectsofacidosishaspreviouslybeenperformed in thispH-responsive cell line by lowering the pH of the cul-turemediumto7.1.17–19,21HereexposureoftheculturestopH7.1for6 hsignificantlyenhancedthestimulatory
[
Ca2+]
i = Kd ×((
F − Fmin)
∕(
Fmax − F))
,
6 | ALDOSARI et al.
effectofionomycinonIL-6 mRNAwhencomparedwithcontrolcultureswithionomycinatpH7.4(1.74 ± 0.097foldincrease,p < 0.0001;Figure 1C).Therewasnoin-creaseinIL-6 mRNAatpH7.1underbasalconditionsintheabsenceofionomycin(Figure 1C).
The accompanying IL-6 protein secretion into theculture medium was measured by ELISA (Figure 1D).However,ahighbasalsecretionof~100 pgIL-6/mlwasdetectedwithin6 hofincubation,evenatpH7.4intheab-senceofionomycin,andnofurtherstatisticallysignificant

| 7ALDOSARI et al.
increase was detectable above this high baseline in re-sponsetoionomycinoradeclineinpH(seeSection5).
4.2 | The effect of low pH does not arise from the pH sensitivity of p38 MAPK
ThepHsensitivityofp38 MAPKwastheninvestigatedtodeterminewhetheractivationofp38bylowpHmightex-plain thepHeffecton IL-6 mRNA.Suchactivationwasnotobserved.Onthecontrary,assessmentofPThr180/Tyr182phospho-activationofp38 MAPKby immunoblotting incultureswhichhadbeenexposedtoanextracellularpHof7.1for1 hdetectedsignificantinhibitionwhencomparedwithcontrolcultureswhichhadbeenexposedtoapHof7.4(0.35 ± 0.04folddecrease,p < 0.05)(Figure 1E).
4.3 | Stretch- induced up- regulation of IL- 6 mRNA is unaffected by low pH
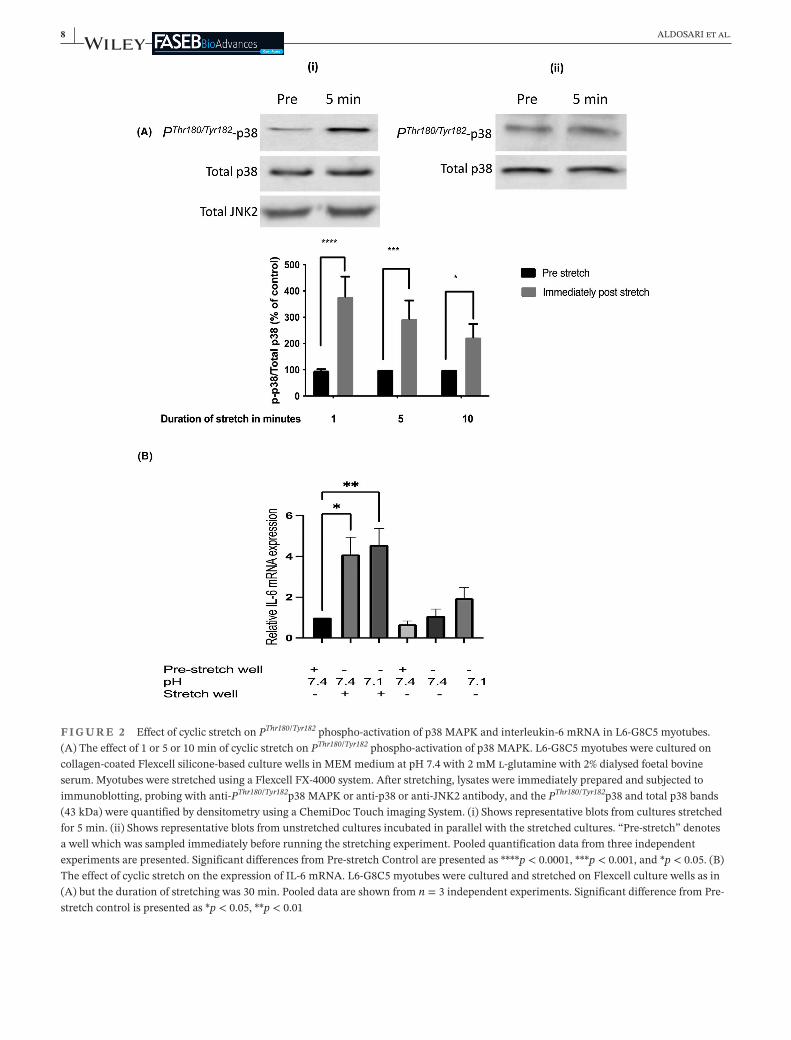
In addition to modelling with ionomycin the rise in in-tracellular Ca2+ concentration that is associated withskeletal muscle contraction (Figure 1A–C), model-ling of some of the mechanical effects of contractionwas also performed by subjecting the cells to cyclicstretch. As reported previously in rat muscle subjectedto static stretch,44 stretching led rapidly to PThr180/Tyr182phospho-activation of p38 MAPK when compared withunstretched cells which had been incubated in parallelin thesameculture incubator (3.85 ± 0.85 fold increaseafter1 minofstretch,p < 0.0001)(Figure 2A).Inagree-ment with an earlier report using mechanically loaded
myotubes45 suchstretching for30 minalso significantlyincreasedIL-6 mRNA(4.05 ± 1.15foldincrease,p < 0.05)(Figure 2B).However,unlike theeffectof ionomycin inFigure 1C, this stretch-induced increase in IL-6 mRNAwasnotenhancedbyapplyinganextracellularpHof7.1(Figure 2B).
4.4 | With ionomycin intracellular Ca2+ concentration is unaffected by extracellular pH
ApossibleexplanationforthestimulationofIL-6 mRNAexpression by low extracellular pH in the presence ofionomycininFigure 1Cis that lowpHis increasingtheintracellularCa2+concentration.AsthestimulatoryeffectoflowpHonIL-6 mRNAwasnotapparentintheabsenceofionomycin(Figure 1CandtheunstretchedculturesinFigure 2B)thismightarisebecauselowpHincreasestheabilityofionomycintocarryCa2+intothecell.However,earlierdirectmeasurementsofthepHdependenceofion-omycin'seffectasanionophoreshowedthatthisdoesnotoccur:ionomycinbecomesalessefficientionophorewhenpHislowered.46Furthermore,directmeasurementofin-tracellularCa2+inL6-G8C5 myoblastsusingthefluores-centindicatorFluo-4didnotdetectanysignificantchangeinresponsetolowpH.EventhoughtheFluo-4AMesterentered the cells and underwent de-esterification to thefluorescentdye(Figure 1G),andeventhoughthisfluores-cencewasclearlyresponsivetoCa2+loadingwithionomy-cinandtoCa2+depletionwithEGTAduringcalibration(Figure 1F); cells which had been treated with ionomy-cinas inFigure 1Cshowednorise in intracellularCa2+
F I G U R E 1 EffectoflowextracellularpHandionomycinoninterleukin-6 mRNAinL6-G8C5 myotubesandmyoblasts.(A)MyotubeswereincubatedinMEMmediumatpH7.4,containing2 mMl-glutaminewith2%dialysedfoetalbovineserumfor6 hwith0.5 µMionomycinand5 µMSB202190(p38inhibitor),andeithernodrugsasabaselinecontrol,or0.5 µMionomycinasapositivecontrol.Pooleddataareshownfromn = 3independentexperiments.****Denotesasignificantdifferencefromtheionomycinpositivecontrol(p < 0.0001).(B)Measurementswereperformedasin(A)butL6-G8C5 myoblastswereusedinsteadofmyotubes.*Denotesasignificantdifferencefromthecontrol(p < 0.05).(C)IL-6 mRNAwasmeasuredinmyotubesasin(A)butthe6 hincubationsinMEMwithorwithoutionomycinwereperformedeitherunderControlconditionsatpH7.4;orunderacidicconditionsatpH7.1.Pooleddataareshownfromn = 3independentexperiments.****Denotessignificantdifferencebetweentheconditionsshown(p < 0.0001).(D)IL-6proteinwasmeasuredinthemediumfromtheculturesin(C)attheendofthe6 hincubations.(E)Myotubeswereincubatedasin(C)for1 hatpH7.1orpH7.4,oratpH7.4with200 mMsucroseasahyper-osmoticpositivecontroltoactivatep38 MAPK.Cultureswerethenimmediatelychilledoniceandimmunoblotswereprobedwithanti-PThr180/Tyr182p38 MAPKantibodyoranti-p38oranti-JNK2antibodyandthePThr180/Tyr182p38andtotalp38bands(43 kDa)werequantifiedbydensitometryusingaChemiDocTouchImagingSystem.Pooleddataareshownfromn = 3independentexperiments,withatleastfourreplicateculturewellsineachexperiment.*DenotessignificantdifferencefromControl(p < 0.05).(F,G,H)L6 myoblastswereculturedinDMEMGrowthMediumfor18 h,rinsedwithPSSbufferandloadedwithFluo-4-AMesterfor45 minat37°Cundera5%CO2 atmosphere.CultureswerethenrinsedthreetimeswithPSS-BSA,andincubatedinfreshPSS-BSAat37°CtoallowFluo-4AMtobede-esterified.ThefluorescenceintensitywasreadonaNOVOstarplate-readeratanexcitationwavelengthof488 nm,anddetectingat>500 nmat37°C.(F)Showsthefluorescencesignalduringarepresentativecalibrationobtainedbyadditionof1 µMionomycinor2 mMEGTA.(G)(i)NegativecontrolimageofcellswithoutFluo-4 showingauto-fluorescence.(ii)ParallelimageofFluo-4 loadedcells.(H)IntracellularCa2+concentrationinculturesafterincubationfor6 hwith0.5 µMionomycinatpH7.1orpH7.4.Pooleddataareshownfromn = 3independentexperiments
8 | ALDOSARI et al.
F I G U R E 2 EffectofcyclicstretchonPThr180/Tyr182phospho-activationofp38 MAPKandinterleukin-6 mRNAinL6-G8C5 myotubes.(A)Theeffectof1or5or10 minofcyclicstretchonPThr180/Tyr182phospho-activationofp38 MAPK.L6-G8C5 myotubeswereculturedoncollagen-coatedFlexcellsilicone-basedculturewellsinMEMmediumatpH7.4with2 mMl-glutaminewith2%dialysedfoetalbovineserum.MyotubeswerestretchedusingaFlexcellFX-4000 system.Afterstretching,lysateswereimmediatelypreparedandsubjectedtoimmunoblotting,probingwithanti-PThr180/Tyr182p38 MAPKoranti-p38oranti-JNK2antibody,andthePThr180/Tyr182p38andtotalp38bands(43 kDa)werequantifiedbydensitometryusingaChemiDocTouchimagingSystem.(i)Showsrepresentativeblotsfromculturesstretchedfor5 min.(ii)Showsrepresentativeblotsfromunstretchedculturesincubatedinparallelwiththestretchedcultures.“Pre-stretch”denotesawellwhichwassampledimmediatelybeforerunningthestretchingexperiment.Pooledquantificationdatafromthreeindependentexperimentsarepresented.SignificantdifferencesfromPre-stretchControlarepresentedas****p < 0.0001,***p < 0.001,and*p < 0.05.(B)TheeffectofcyclicstretchontheexpressionofIL-6 mRNA.L6-G8C5 myotubeswereculturedandstretchedonFlexcellculturewellsasin(A)butthedurationofstretchingwas30 min.Pooleddataareshownfromn = 3independentexperiments.SignificantdifferencefromPre-stretchcontrolispresentedas*p < 0.05,**p < 0.01

| 9ALDOSARI et al.
concentration in response to an extracellular pH of 7.1whencomparedwithparallelculturesatanextracellularpHof7.4(Figure 1H).
4.5 | Inhibition or silencing of the SNAT2 transporter fails to mimic the enhancing effect of low pH on IL- 6 mRNA
TheactivityoftheSNAT2aminoacidtransporterinL6-G8C5 myotubes has been shown previously to undergoan acute ~0.5 fold inhibition when extracellular pH islowered to 7.1 (figure 1C in Ref. [17]) and this finding(0.57 ± 0.12folddecrease,p < 0.01)wasconfirmedbyus(seeFigure 3A).IfthecauseoftheincreaseinIL-6 mRNAat pH 7.1 in the presence of ionomycin in Figure 1C inthepresentpaperwasthereductionofSNAT2transportactivity by the low pH (Figure 3A), then SNAT2 inhibi-tionbyothermeanswouldalsobepredictedto increaseIL-6 mRNA. To test this, three independent methods ofSNAT2inhibitionwereused.
Firstly,itwasnotedthatpharmacologicalinhibitionofJNKMAPKwith the selective inhibitorSP600125atpH7.4 led to inhibition of the activity of the SNAT2 trans-porter (0.59 ± 0.05-fold decrease, p < 0.01) (Figure 3B)comparableinmagnitudewiththeinhibitionobservedatpH7.1(Figure 3A).Inspiteofthisinhibitionofthetrans-porter,SP600125 hadnostatisticallysignificantenhancingeffectonIL-6 mRNAinculturesatpH7.4withionomycin(Figure 3C,D).Indeed,atpH7.1withionomycin,JNKin-hibitionbySP600125 significantlybluntedtheenhancingeffectofthislowpHonIL-6 mRNA(0.52 ± 0.04foldde-crease, p < 0.0001) (Figure 3D), suggesting that the ac-tion of low pH on IL-6 mRNA (but not the stimulatorysignal fromionomycin to IL-6expression)was inhibitedbyblockadeofJNK.
Secondly, SNAT2 inhibition was also investigated bycompetitive inhibition of the transporter with a saturat-ingdoseofitsselectivesubstrateMeAIB.17,21AswithJNKinhibition, this had no statistically significant effect onIL-6 mRNAatpH7.4(Figure 3C,D)butgaveamarginallysignificantblunting(0.87 ± 0.03folddecrease,p = 0.052)oftheIL-6 mRNAresponseatpH7.1(Figure 3D).
Finally, inhibition of this transporter was also per-formed by siRNA silencing of SNAT2 gene expression(Figure 4).SuchsilencingofSNAT2ismeasurableinL6-G8C5 myotubes18butmoreefficientsilencingisobtainedinunfusedmyoblasts.17,18Itwasconfirmedthatthemyo-blasts(likethemyotubes inFigure 1A)stillshowanin-creaseinIL-6 mRNAexpressioninresponsetoionomycin(6.3 ± 0.9 fold increase,p < 0.05) (Figure 1B).Effectivesilencing of SNAT2 mRNA by siRNA oligonucleotideswasconfirmedinthemyoblastsinFigure 4A(median0.22
foldsilencing(range0.15to0.35,p = 0.019)).Whencom-pared with control cultures which had been transfectedwith scrambledcontrololigonucleotides, cultureswhichhadbeentreatedwithSNAT2-silencingsiRNAsexpressedlowerlevelsofIL-6 mRNA,atpH7.4inthepresenceof0.5 µMionomycin(median0.63folddecrease(range0.45to1.00,p = 0.0267))(Figure 4C).
Under none of the conditions tested was inhibitionof the SNAT2 transporter associated with a statisticallysignificantincreaseinIL-6 mRNA,contrarytowhathadbeenpredictedbytheoriginalhypothesis.
5 | DISCUSSION
5.1 | The culture model and its limitations
Ithasbeenreportedpreviously13thatL6ratskeletalmus-clecellstreatedwithionomycinshowastrikingincreaseinIL-6 mRNA,whichisblockedbypharmacologicalin-hibitionofp38 MAPK.Thepresentstudyconfirmedthiseffect(Figure 1A,B)bothinmyotubesandinmyoblastsofthe pH-responsive L6-G8C5 sub-clone of L6 cells whichpreviouslyshowedmarkedproteinmetabolismresponsesto low pH that are similar to those observed in skeletalmuscleinvivo.17,18TheL6-G8C5celllinethereforeseemsasuitablemodelinwhichtoinvestigatethemechanismoftherecentlyreportedroleofacidosisintheup-regulationofIL-6expressioninskeletalmuscle.16
However,apracticallimitationencounteredwiththismodel was the substantial rate of accumulation of IL-6protein in the medium (~100 pg/ml within 6 h) evenunderbasalcontrolconditions(pH7.4withoutionomy-cin).Mycoplasmascreening(seeSection3)indicatedthatthiswasnotaconsequenceofoccult infection.NoIL-6proteindatawerereportedintheoriginalstudyonIL-6expression in L6 cells on which the present study wasbased.13SuchbasalexpressionofIL-6proteinisnotob-servedinanotherrodentmyogeniccell,themouseC2C12cellline.47HoweverC2C12wasthoughttobeunsuitableforthepresentpHstudybecauseofitspreviouslydocu-mented19 rapid tendency toacidify theculturemediumowing to the cells’ extremely high rate of lactic acidoutput.
PossiblyasaresultofthehighbasalsecretionofIL-6protein observed here (Figure 1D), no further statisti-cally significant increase beyond this high baseline wasdetectedinresponsetolowpHwithorwithoutionomy-cin (Figure 1D). Furthermore it has been reported thatinsomecelltypesautocrineeffectsofIL-6 self-signallingmaylimitfurtherIL-6 secretion.48Incontrastatwo-foldstimulationofIL-6proteinsecretionhasbeenreportedin

10 | ALDOSARI et al.
F I G U R E 3 EffectoflowextracellularpHandinhibitionofJNKandofSNAT2(SLC38A2)aminoacidtransportersoninterleukin-6 mRNAinL6-G8C5 myotubes.(A)AcuteeffectoflowextracellularpH(appliedonlyduringthetransportassay)onSNAT2(SLC38A2)transporteractivityinL6-G8C5 myotubes.14C-MeAIBtransportwasassayedin5 minincubationsinHBSmediumatpH7.1or7.4.Pooleddataareshownfromsixindependentexperiments,withfivereplicateculturewellsineachexperiment.**p < 0.01versuspH7.4control.(B)Cellswerepre-incubatedfor4 hatpH7.4inMEM/2%DFBSwithJNKMAPKinhibitorSP600125(10 µM)orincontrolcultureswithoutinhibitor.The14C-MeAIBtransportratewasthenimmediatelyassayedasin(A)butatpH7.4onlyandintheabsenceoftheinhibitor.Pooleddataareshownfromn = 3independentexperiments,withfivereplicateculturewellsineachexperiment.**DenotessignificantdifferencefromControlp < 0.01.(C)Myotubeswereculturedfor6 hinMEMmediumatpH7.4with2 mMl-glutamineand2%dialysedfoetalbovineserum,eitherwithnodrugsasabaselinecontrolorwith0.5 µMionomycinwithandwithoutdrugs.Thedrugstestedwere10 µMJNKinhibitor(SP600125)and10 mMMeAIB.Pooleddataareshownfromn = 3independentexperiments.Significantdifferencesbetweenconditionsareshownas***p < 0.001,****p < 0.0001.(D)IL-6 mRNAwasmeasuredafter6-hincubationsasin(C)butthepHoftheMEMmediumwaseitheracontrolpHof7.4oranacidicpHof7.1.Pooleddataareshownfromn = 3independentexperiments.Significantdifferencesbetweenconditionsareshownas*p < 0.05,****p < 0.0001

| 11ALDOSARI et al.
responsetolacticacidtreatmentwithin6 hinelectricallystimulated human myotubes.16 However, as the IL-6 se-cretion data were reported in that study as fold changeratherthanabsoluteconcentrations inthemedium,it isnotpossibletocomparethebasalsecretionratetherewiththatinthepresentstudy.
Thereason(s)forthesedifferencesinIL-6outputbetweenculturemodelsareunknown.InfutureworkintheL6-G8C5cell line it would be of interest to determine whether thebasalrateofIL-6outputdeclinesafter6 handwhethertheIL-6 mRNAeffectsdescribedhereareultimatelyreflectedinIL-6proteinexpressioniflongerincubationsareperformed.
F I G U R E 4 EffectofsiRNAsilencingofexpressionoftheSNAT2(SLC38A2)aminoacidtransporteroninterleukin-6 mRNAinL6-G8C5 myoblasts.(A)L6 myoblastswereculturedinDMEMgrowthmediumfor24 handthentransfectedwith30 nMscrambledcontrolsiRNA(Scr)orsilencinganti-SNAT2 siRNA(Sil)for16 h.Negativecontrolcultureswerealsotreatedwithtransfectionagentonly,ornoadditions.Themediumwasthenreplacedwithgrowthmediumfor24 handincubatedforafurther6 hinMEMmediumatpH7.4with2 mMl-glutaminewith2%dialysedfoetalbovineserum.Pooleddataareshownfromn = 4independentexperiments.(B,C)L6 myoblastsweretransfectedortreatedwithnegativetransfectioncontrolsasin(A)butthesubsequent6-hincubationsinMEMatpH7.4wereperformedeither(B)inabsenceofionomycin,or(C)with0.5 µMionomycin.Pooleddataareshownfromn = 4independentexperiments

12 | ALDOSARI et al.
Afurtherlimitationpreviouslynotedwiththissponta-neously fusedL6-G8C5 myotubemodel is that themag-nitudeofthepHsensitivityofproteinmetabolisminthecultureswassensitivetomodestchangesinpassagenum-ber and myotube fusion/differentiation (figure 6 in Ref.[49]).SuchvariabilityinthemagnitudeofthepHsensitiv-itywasalsoobservedhere(Figure 1Cvs.Figure 3D),andthemagnitudeoftheresponsetoionomycinalsoshowedconsiderablevariation(Figure 1Avs.Figure 1C).Inprin-ciple this problem can be overcome by driving myotubefusion and differentiation to a consistently very highlevelusing thedifferentiatingagents insulin-likegrowthfactor-I(IGF-I)andretinoicacid(RA)asdescribedprevi-ously.50HoweverapplyingIGF-IandRAabolishesthepHsensitivityoftheculturesandadministrationofglucocor-ticoidisthenrequiredtorestorepHsensitivity.49InviewofthepossibleconfoundingeffectsofglucorticoidonIL-6expression51,52;glucocorticoid,IGF-IandRAwerenotap-pliedinthepresentstudy.
5.2 | Mechanism of the effect of low pH
Theup-regulationofIL-6 mRNApreviouslyobservedwithionomycininL6cells13wasshownheretobestronglyen-hancedbyalowpHof7.1comparedwithacontrolpHof7.4.Incontrast,nostatisticallysignificanteffectoflowpHwasdemonstratedunderbasalconditionsintheabsenceof ionomycin (Figure 1C), and the related phenomenonof an increase in IL-6 mRNA that was observed follow-ingcyclicstretchshowednodependenceonpHoverthispHrange(Figure 2B).ViewedtogethertheseresultsimplythatsynergismbetweenCa2+andlowpHmayberequiredtoexplaintheregulationofIL-6 mRNAunderconditionsrelevanttolacticacidosisinexercise.Itshouldbeempha-sisedhoweverthat(asshowninFigure 1H)thisapparentinteractionbetweentheeffectsofionomycinandpHdidnotarisefromartefactualenhancementbylowpHofion-omycin'sefficiencyasanionophore.Furthermore,itwasobserved here that the enhancing effect of low pH withionomycinonIL-6 mRNAwasmorethanhalvedbyJNKinhibition (Figure 3D), whereas JNK inhibition had nosignificanteffectinthepresenceofionomycinatthecon-trolpHof7.4(Figure 3C).(Thisisunlikelytoarisefromfailure of the JNK inhibitor to enter the cells at pH 7.4because this inhibitorwaseffective in inhibitingSNAT2transporteractivityatthispH(Figure 3B).)ThissuggeststhereforethatJNKmayhavearolehereinmediatingtheeffectoflowpH,inadditiontoitswell-documentedroleinmediating responses toCa2+ throughCa2+-induced JNKactivation.53–59
ThisJNK-dependentmechanismbywhichlowpHin-creases IL-6 mRNA in L6-G8C5 cells in the presence of
ionomycinisclearlynotthewell-documentedinhibitionof SNAT2 by low pH which has previously been showntoexertpotentandfunctionallyimportanteffectsonpro-tein metabolism in this cell line.17,18 Rapidly decreasingthe activity of SNAT2 transporters by lowering the pH(Figure 3A)wasseenheretobeaccompaniedbyenhance-mentofIL-6 mRNA(Figure 1CandFigure 3D),whereassilencingofSNAT2withsiRNA(Figure 4C),competitiveinhibitionwithMeAIBatlowpH(Figure 3D)orindirectinhibition of transport activity by blocking JNK at lowpH (Figure 3D) were all associated with a decrease inIL-6 mRNA.Thissuggeststhereforethat,whilecouplingbetweenSNAT2activityandIL-6expressionmayexist,itoperatesintheoppositedirectionfromthatoriginallyhy-pothesisedinthepresentstudy.
5.3 | Future work
AlimitationintheSNAT2 siRNAsilencingexperimentsdescribedherewastheobservationinFigure 4Bthatthescrambled control siRNA increased IL-6 mRNA whencompared with cultures treated with transfection agentalone. A possible explanation is that L6 myoblasts (likeothermusclecells60)expressToll-likereceptor3(TLR3),anon-specificsensorofdouble-strandedRNAwhichmayrespond directly to the scrambled control siRNA and isaknownactivatorofIL-6expression.61InfutureworkitwouldthereforebeofinteresttocircumventtheneedfordssiRNAsbyapplyinganalternativetechniquetosilenceSNAT2 geneexpression,forexampleusingviralvectorstoperformstableintegrationofshRNAs.62
It has been reported that electrical pulse stimulation(EPS) of C2C12 myotubes increases IL-6 mRNA andprotein expression in these myotubes, through a JNK-dependent pathway.63 In view of the observation here(Figure 3D) that the effect of low pH on ionomycin-inducedIL-6 mRNAwasdependentonJNK,itwouldbeinterestinginfuturetostudytheeffectoflowpHonEPS-inducedIL-6expression.
Finally, in view of the physiological importance ofIL-6inexercisingskeletalmuscle,3theidentityofthepHsensormediating theup-regulationof IL-6 mRNAthatwasobservedheremeritsfurtherinvestigation.Itshouldbe emphasised however that the sensor responsible forthiseffectinL6-G8C5cellsmustbeprobingextracellularpH,because thecytosolicpH inL6-G8C5cells isunaf-fectedbyafallinextracellularpHfrom7.5to7.1(table5inRef.[19]).ApHsensingmechanismcapableofsensinglacticacidosisandregulatingreleaseofacytokineonatimescaleofminutestohoursduringexercisemayneedto be distinct from the SNAT2 pH sensing mechanismbecausethelatteroperatesonalongertimescale(hours

| 13ALDOSARI et al.
todays)andfunctionsinthesensingofchronicacidosis(for example in starvation ketoacidosis64) culminatinginthereleaseoffreeaminoacidsfrommuscleprotein.18(The pH-dependent mechanism described here mightalsoplayadistinctroleinmodulatingthepreviouslyde-scribedsignallingfromIL-6tolipolysis4andhepaticglu-coseoutput5.)Intheregulationofcytokineexpressionintheimmunesystemandinflammationbiology,G-proteincoupledreceptorssuchasOGR1 havebeenshowntobefunctionallyimportantsensorsofextracellularpHlinkedtoIL-6expressionandCa2+mobilisation65andmayactthroughJNKactivation.66Thismayexplainthesuppres-sive effect of JNK inhibition on IL-6 mRNA at pH 7.1in Figure 3D. However, whether such receptors are ex-pressedinskeletalmuscle(albeit transientlyduringex-ercise),andwhethertheyplayaroleinsensingoflacticacidosisand in regulating IL-6expression inexercisingskeletalmuscle,remainstobedetermined.
ACKNOWLEDGEMENTSZA thanksShaqraUniversity,KingdomofSaudiArabiaforsponsoringhis research throughout thisproject.TheauthorsthankMrJeremyBrownfortechnicalassistance;Dr Gary Willars and Dr Heider Qassam for assistancewith the Fluo-4 fluorescence measurements; and DrJonathonWilletsforassistancewiththecycliccellstretchexperiments.
CONFLICT OF INTERESTTheauthorsdeclarethattheyhavenoconflictsofinterestarisingfromthecontentsofthisarticle.
AUTHORS’ CONTRIBUTIONSZiyad Aldosari, Nima Abbasian, Katherine Robinson,AlanBevingtonandEmmaWatsoncontributedtostudydesign,draftingtextandfigures,andrevisionofthepaper.Ziyad Aldosari, Nima Abbasian, Alan Bevington andEmma Watson performed experiments. Ziyad AldosariandAlanBevingtoncontributedtodataanalysis.Allau-thorsapprovedthefinalmanuscript.
ORCIDZiyad Aldosari https://orcid.org/0000-0001-5385-7567Nima Abbasian https://orcid.org/0000-0002-6906-2563
REFERENCES 1. Steensberg A, Keller C, Starkie RL, Osada T, Febbraio MA,
PedersenBK.IL-6andTNF-αexpressionin,andreleasefrom,contracting human skeletal muscle. Am J Physiol- Endocrinol Metabol.2002;283(6):E1272-E1278.
2. PeakeJM,NeubauerO,DellaGattaPA,NosakaK.Muscledam-age and inflammation during recovery from exercise. J Appl Physiol.2016;122(3):559-570.
3. PetersenAM,PedersenBK.Theanti-inflammatoryeffectofex-ercise.J Appl Physiol.2005;98(4):1154-1162.doi:10.1152/japplphysiol.00164.2004
4. VanHallG,SteensbergA,SacchettiM,etal.Interleukin-6stim-ulateslipolysisandfatoxidationinhumans.J Clin Endocrinol Metabol.2003;88(7):3005-3010.
5. TsigosC,PapanicolaouDA,KyrouI,DefensorR,MitsiadisCS,ChrousosGP.Dose-dependenteffectsof recombinanthumaninterleukin-6onglucoseregulation.J Clin Endocrinol Metabol.1997;82(12):4167-4170.
6. StouthardJM,RomijnJA,VanderPollT,etal.Endocrinologicandmetaboliceffectsofinterleukin-6inhumans.Am J Physiol- Endocrinol Metabol.1995;268(5):E813-E819.
7. PetersenAMW,PedersenBK.Theanti-inflammatoryeffectofexercise.J Appl Physiol.2005;98(4):1154-1162.
8. SteensbergA,FischerCP,KellerC,MøllerK,PedersenBK.IL-6enhancesplasmaIL-1ra,IL-10,andcortisol inhumans.Am J Physiol- Endocrinol Metabol.2003;285(2):E433-E437.
9. Tilg H, Dinarello CA, Mier JW. IL-6 and APPs: anti-inflammatory and immunosuppressive mediators. Immunol Today.1997;18(9):428-432.
10. StarkieR,OstrowskiSR,JauffredS,FebbraioM,PedersenBK.Exercise and IL-6 infusion inhibit endotoxin-induced TNF-αproductioninhumans.FASEB J.2003;17(8):884-886.
11. Ridker PM, Rifai N, Stampfer MJ, Hennekens CH. Plasmaconcentrationof interleukin-6and theriskof futuremyocar-dial infarction among apparently healthy men. Circulation.2000;101(15):1767-1772.
12. Keller C, Steensberg A, Pilegaard H, et al. Transcriptionalactivation of the IL-6 gene in human contracting skele-tal muscle: influence of muscle glycogen content. FASEB J.2001;15(14):2748-2750.
13. Chan MS, McGee SL,Watt MJ, Hargreaves M, Febbraio MA.Alteringdietarynutrientintakethatreducesglycogencontentleadstophosphorylationofnuclearp38MAPkinaseinhumanskeletalmuscle:associationwithIL-6genetranscriptionduringcontraction.FASEB J.2004;18(14):1785-1787.
14. OstrowskiK,RohdeT,ZachoM,AspS,PedersenB.Evidencethatinterleukin-6isproducedinhumanskeletalmuscleduringprolongedrunning.J Physiol.1998;508(3):949-953.
15. SteensbergA,VissingJ,PedersenBK.LackofIL-6productionduringexerciseinpatientswithmitochondrialmyopathy.Eur J Appl Physiol.2001;84(1):155-157.
16. Hojman P, Brolin C, Nørgaard-Christensen N, et al. IL-6 re-lease from muscles during exercise is stimulated by lactate-dependentproteaseactivity.Am J Physiol- Endocrinol Metabol.2019;316(5):E940-E947.
17. EvansK,NasimZ,BrownJ,etal.Acidosis-sensingglutaminepumpSNAT2 determines amino acid levels and mammalian target ofrapamycinsignallingtoproteinsynthesisinL6musclecells.J Am Soc Nephrol.2007;18(5):1426-1436.doi:10.1681/ASN.2006091014
18. EvansK,NasimZ,BrownJ,etal.InhibitionofSNAT2bymeta-bolicacidosisenhancesproteolysisinskeletalmuscle.J Am Soc Nephrol.2008;19(11):2119-2129.doi:10.1681/ASN.2007101108
19. BevingtonA,BrownJ,PrattA,MesserJ,WallsJ.Impairedgly-colysisandproteincatabolisminducedbyacidinL6ratmusclecells.Eur J Clin Invest.1998;28(11):908-917.
20. Reaich D, Channon S, Scrimgeour C, Daley S, WilkinsonR, Goodship T. Correction of acidosis in humans with CRF

14 | ALDOSARI et al.
decreasesproteindegradationandaminoacidoxidation.Am J Physiol- Endocrinol Metabol.1993;265(2):E230-E235.
21. BevingtonA,BrownJ,ButlerH,etal.ImpairedsystemAaminoacidtransportmimicsthecataboliceffectsofacidinL6cells.Eur J Clin Invest.2002;32(8):590-602.
22. Menchini RJ, Chaudhry FA. Multifaceted regulation of thesystemAtransporterSlc38a2suggestsnanoscaleregulationofaminoacidmetabolismandcellularsignaling.Neuropharmacol.2019;161:107789.
23. Bhutia YD, Ganapathy V. Glutamine transporters in mam-malian cells and their functions in physiology and cancer.Biochimica et Biophysica Acta (BBA)- Molecular Cell Res.2016;1863(10):2531-2539.
24. BairdFE,Pinilla-Tenas JJ,OgilvieWL,GanapathyV,HundalHS, Taylor PM. Evidence for allosteric regulation of pH-sensitiveSystemA(SNAT2)andSystemN(SNAT5)aminoacidtransporter activity involving a conserved histidine residue.Biochem J.2006;397(2):369-375.
25. ChristensenHN,OxenderDL,LiangM,VatzKA.TheuseofN-methylationtodirecttherouteofmediatedtransportofaminoacids.J Biol Chem.1965;240(9):3609-3616.
26. Schiöth HB, Roshanbin S, Hägglund MG, Fredriksson R.EvolutionaryoriginofaminoacidtransporterfamiliesSLC32,SLC36 and SLC38 and physiological, pathological and thera-peuticaspects.Mol Aspects Med.2013;34(2–3):571-585.
27. YaoD,MackenzieB,MingH,etal.AnovelsystemAisoformmediating Na+/neutral amino acid cotransport. J Biol Chem.2000;275(30):22790-22797.doi:10.1074/jbc.M002965200
28. HundalHS,TaylorPM.Aminoacidtransceptors:gatekeepersofnutrientexchangeandregulatorsofnutrientsignaling.Am J Physiol Endocrinol Metab.2009;296(4):E603-E613.doi:10.1152/ajpendo.91002.2008
29. WatsonEL,KosmadakisGC,SmithAC,etal.Combinedwalk-ing exercise and alkali therapy in patients with CKD4–5 reg-ulates intramuscular free amino acid pools and ubiquitin E3ligaseexpression.Eur J Appl Physiol.2013;113(8):2111-2124.
30. NemotoS,XiangJ,HuangS,LinA.InductionofapoptosisbySB202190throughinhibitionofp38βmitogen-activatedproteinkinase.J Biol Chem.1998;273(26):16415-16420.
31. BennettBL,SasakiDT,MurrayBW,etal.SP600125,ananthra-pyrazoloneinhibitorofJunN-terminalkinase.Proc Natl Acad Sci.2001;98(24):13681-13686.
32. Yaffe D. Retention of differentiation potentialities duringprolonged cultivation of myogenic cells. Proc Natl Acad Sci.1968;61(2):477-483.
33. SumiD,KojimaC,KasaiN,GotoK.Theeffectsofenduranceexerciseinhypoxiaonacid-basebalanceandpotassiumkinet-ics:arandomizedcrossoverdesigninmaleenduranceathletes.Sports Medicine- Open.2018;4(1):1-8.
34. SteinhagenC,HircheH,NestleH,BovenkampU,HosselmannI.TheinterstitialpHoftheworkinggastrocnemiusmuscleofthedog.Pflügers Archiv.1976;367(2):151-156.
35. GotoK,OkuyamaR,SugiyamaH,etal.Effectsofheatstressandmechanicalstretchonproteinexpressioninculturedskel-etalmusclecells.Pflügers Archiv.2003;447(2):247-253.
36. Gomez E, Powell ML, Greenman IC, Herbert TP. Glucose-stimulated protein synthesis in pancreatic beta-cells parallelsanincreaseintheavailabilityofthetranslationalternarycom-plex(eIF2-GTP.Met-tRNAi)andthedephosphorylationofeIF2alpha.J Biol Chem.2004;279(52):53937-53946.
37. KelleherAR,GordonBS,KimballSR,JeffersonLS.Changesin REDD1, REDD2, and atrogene mRNA expression areprevented in skeletal muscle fixed in a stretched positionduring hindlimb immobilization. Physiological Reports.2014;2(2):e00246.
38. PfafflMW.Anewmathematicalmodelforrelativequantifica-tioninreal-timeRT–PCR.Nucleic Acids Res.2001;29(9):e45.
39. Heding A, Elling C, SchwartzT. Novel method for the studyofreceptorCa2+signallingexemplifiedbytheNK1receptor.J Recept Signal Transduction.2002;22(1–4):241-252.
40. Klintman D, Li X, Santen S, Schramm R, Jeppsson B,Thorlacius H. p38 mitogen-activated protein kinase-dependent chemokine production, leukocyte recruitment,and hepatocellular apoptosis in endotoxemic liver injury.Ann Surg. 2005;242(6):830-839. doi:10.1097/01.sla.0000189132.86878.f7
41. Liu A, Yang J, Gonzalez FJ, Cheng GQ, Dai R. Biphasic reg-ulation of intracellular calcium by gemfibrozil contributes toinhibitingL6myoblastdifferentiation:implicationsforclinicalmyotoxicity.Chem Res Toxicol.2010;24(2):229-237.
42. Yamasaki-Mann M, Demuro A, Parker I. cADPR stimu-lates SERCA activity in Xenopus oocytes. Cell Calcium.2009;45(3):293-299.
43. Sylow L, Kleinert M, Richter EA, Jensen TE. Exercise-stimulated glucose uptake—regulation and implications forglycaemic control. Nat Rev Endocrinol. 2017;13(3):133-148.doi:10.1038/nrendo.2016.162
44. Boppart MD, Hirshman MF, Sakamoto K, Fielding RA,Goodyear LJ. Static stretch increases c-Jun NH2-terminal ki-nase activity and p38 phosphorylation in rat skeletal muscle.Am J Physiol- Cell Physiol.2001;280(2):C352-C358.
45. Juffer P, Bakker AD, Klein-Nulend J, Jaspers RT. Mechanicalloadingbyfluidshearstressofmyotubeglycocalyxstimulatesgrowth factor expression and nitric oxide production. Cell Biochem Biophys.2014;69(3):411-419.
46. StilesMK,CraigME,GunnellS,PfeifferD,TaylorR.Thefor-mation constants of ionomycin with divalent cations in 80%methanol/water.J Biol Chem.1991;266(13):8336-8342.
47. CseriK,SzentesiP,CsernochL.IL-6productionofC2C12cellsisenhanced in thepresenceofmacrophagesandpravastatin.Gen Physiol Biophys.2021;40(4):307-315.
48. Verboogen DR, Revelo NH,Ter Beest M, van den Bogaart G.Interleukin-6 secretion is limited by self-signaling in endo-somes.J Mol Cell Biol.2019;11(2):144-157.
49. PickeringW,ChengM-K,BrownJ,ButlerH,WallsJ,BevingtonA. Stimulation of protein degradation by low pH in L6G8C5skeletal muscle cells is independent of apoptosis but de-pendent on differentiation state. Nephrol Dial Transplant.2003;18(8):1466-1474.
50. Elsner P, Quistorff B, Hermann TS, Dich J, Grunnet N.RegulationofglycogenaccumulationinL6myotubesculturedunder optimized differentiation conditions. Am J Physiol- Endocrinol Metabol.1998;275(6):E925-E933.
51. WaageA,SlupphaugG,ShalabyR.GlucocorticoidsinhibittheproductionofIL6frommonocytes,endothelialcellsandfibro-blasts.Eur J Immunol.1990;20(11):2439-2443.
52. DittrichA,KhouriC,SackettSD,etal.Glucocorticoidsincreaseinterleukin-6–dependent gene induction by interfering withtheexpressionofthesuppressorofcytokinesignaling3feed-backinhibitor.Hepatology.2012;55(1):256-266.

| 15ALDOSARI et al.
53. SunW,WuX,GaoH,etal.CytosoliccalciummediatesRIP1/RIP3 complex-dependent necroptosis through JNK activationand mitochondrial ROS production in human colon cancercells.Free Radic Biol Med.2017;108:433-444.
54. BrnjicS,OlofssonMH,HavelkaAM,LinderS.Chemicalbiol-ogysuggestsaroleforcalciumsignalinginmediatingsustainedJNKactivationduringapoptosis.Mol BioSyst.2010;6(5):767-774.
55. KimJ,SharmaRP.Calcium-mediatedactivationofc-JunNH2-terminalkinase(JNK)andapoptosisinresponsetocadmiuminmurinemacrophages.Toxicol Sci.2004;81(2):518-527.
56. Saxena M, Busca A, Pandey S, Kryworuchko M, Kumar A.CpGprotectshumanmonocyticcellsagainstHIV-Vpr–inducedapoptosis by cellular inhibitor of apoptosis-2 through thecalcium-activatedJNKpathway inaTLR9-independentman-ner.J Immunol.2011;187(11):5865-5878.
57. Arthur JM, Lawrence MS, Payne CR, Rane MJ, McLeish KR.Thecalcium-sensingreceptorstimulatesJNKinMDCKcells.Biochem Biophys Res Comm.2000;275(2):538-541.
58. Dhanasekaran DN, Reddy EP. JNK-signaling: a multi-plexing hub in programmed cell death. Genes & Cancer.2017;8(9-10):682-694.
59. HuangY,LiX,WangY,WangH,HuangC,LiJ.Endoplasmicreticulumstress-inducedhepaticstellatecellapoptosisthroughcalcium-mediated JNK/P38 MAPK and Calpain/Caspase-12pathways.Mol Cell Biochem.2014;394(1-2):1-12.
60. SchreinerB,VossJ,WischhusenJ,etal.Expressionoftoll-likereceptorsbyhumanmusclecellsinvitroandinvivo:TLR3ishighlyexpressedininflammatoryandHIVmyopathies,medi-atesIL-8release,andup-regulationofNKG2D-ligands.FASEB J.2006;20(1):118-120.
61. MelkamuT,KitaH,O’GradySM.TLR3activationevokesIL-6secretion, autocrine regulation of Stat3 signaling and TLR2
expression in human bronchial epithelial cells. J Cell Comm Signaling.2013;7(2):109-118.
62. Taxman DJ, Moore CB, Guthrie EH, Huang MT-H. Short Hairpin RNA (shRNA): Design, Delivery, and Assessment of Gene Knockdown. RNA Therapeutics.Springer;2010;139-156.
63. WhithamM,ChanMHS,PalM,etal.Contraction-inducedin-terleukin-6 gene transcription in skeletal muscle is regulatedby c-Jun terminal kinase/activator protein-1. J Biol Chem.2012;287(14):10771-10779.
64. Bevington A, Poulter C, Brown J, Walls J. Inhibition of pro-tein synthesis by acid in L6 skeletal muscle cells: analogieswith the acute starvation response. Miner Electrolyte Metab.1998;24(4):261-266.
65. IchimonjiI,TomuraH,MogiC,etal.ExtracellularacidificationstimulatesIL-6productionandCa2+mobilizationthroughproton-sensingOGR1receptors inhumanairwaysmoothmusclecells.Am J Physiol- Lung Cellular Mol Physiol.2010;299(4):L567-L577.
66. Maeyashiki C, Melhem H, Hering L, et al. Activation of pH-sensingreceptorOGR1(GPR68)inducesERstressviatheIRE1α/JNKpathwayinanintestinalepithelialcellmodel.Sci Rep.2020;10(1):1-12.
How to cite this article:AldosariZ,AbbasianN,RobinsonK,BevingtonA,WatsonE.LowpHup-regulatesinterleukin-6mRNAinL6-G8C5ratskeletalmusclecellsindependentofpHsensingbySNAT2(SLC38A2)transporters.FASEB BioAdvances.2021;00:1–15.doi:10.1096/fba.2021-00088