lower cretaceous dasyclad algae from the padurea craiului massif
TRANSCRIPT

ACTA PALAEONTOLOGICA ROMANIAE V. 2 (1999), P. 53-72
53
LOWER CRETACEOUS DASYCLAD ALGAE FROM THE PADUREA CRAIULUI MASSIF (NORTHERN APUSENI MOUNTAINS, ROMANIA)
IOAN I. BUCUR1
Abstract. The Lower Cretaceous deposits in Pădurea Craiului bear a relatively rich assemblage of dasyclad algae belonging to the Valanginian-Aptian interval. Most of the species illustrated in the present paper were sampled from the Blid Formation (“Lower Pachyodont Limestone”) . Several paleontological remarks on some less known species as well as on a few species with uncertain systematic position are presented. The folowing new combinations are also suggested: Anisoporella? cretacea (DRAGASTAN, 1967) nov. comb., Anisoporella? jurassica (ENDO, 1961) nov. comb. Falsolikanella(?) silvaeregis (BUCUR, 1981) nov. comb., Milanovicella pejovicae (RADOICIČ, 1975) nov. comb. and Similiclypeina paucicalcarea (CONRAD, 1970) nov.comb., and a new species, Neomizzia dacica n.sp. is described, the genus Neomizzia LEVY, 1966 being validated by designation of a lectotype.
Keywords: systematics, dasyclad algae, Lower Cretaceous, Pădurea Craiului (Apuseni Mountains), Romania.
1 Babeş-Bolyai University, Department of Geology-Paleontology Str. M. Kogălniceanu nr.1, 3400 Cluj-Napoca, Romania
INTRODUCTION. STRATIGRAPHICAL SETTING
The Pădurea Craiului Massif is situated in north-western Romania, being part of the Apuseni Mountains (Fig. 1 ). From a structural point of view the sedimentary deposits in this massif are part of the Bihor-Pădurea Craiului unit. Their age ranges from Permian to Upper Cretaceous (PATRULIUS, in IANOVICI et al., 1976). The Cretaceous deposits follow a sedimentary gap due to the lifting of the region at the end of the Upper Jurassic. At the base they consist of bauxitic rocks. Within the succession corresponding to the Lower Cretaceous, the following lithostratigraphical units have been delimitated (PATRULIUS, in IANOVICI et al., 1976; DRAGASTAN et al., 1986, 1988, 1989; BUCUR et al., 1993; BUCUR & COCIUBA, 1996, 1998; COCIUBA, 1999) (Fig.2):
1. The Blid Formation (DRAGASTAN et al., 1986, 1988)
Following the Upper Jurassic sedimentation gape, this formation comprises two members (COCIUBA, 1999):
1.a. The Dobreşti Member, which has at the base bauxitiferous rocks or lies directly on Upper Jurassic limestones. They had initially a lacustrine character, changing subsequently into a brackish and finally normal marine one and were known as “Characean- and Gastropod-bearind limestones”. Characeans, abounding in the lower part of these limestones have been descibed by DRAGASTAN et al. (1966). In characean- and gastropod bearing limestones several levels abounding in Salpingoporella anullata CAROZZI as well as some levels containing small foraminifera, Montsalevia salevensis (CHAROLLAIS et al.) included, also occur. The age of the Dobresti Member is most probably Valanginian-Hauterivian.
1.b. The Coposeni Member, frequently known in literature as “Lower Pachyodont Limestone” (PATRULIUS in IANOVICI et al., 1976), is considered to be of Barremian age. The foraminifera assemblege occuring in the lower part of these limestones is dominated by Paracoskinolina? jourdanensis (FOURY & MOULLADE) (BUCUR et al., 1993).
2. The Ecleja Formation (PATRULIUS, in IANOVICI et al., 1976), a succession of gray, partly siltic marls to marly siltites, includes also two lithologicaly diferent members (COCIUBA, 1999): The Gugu Breccia Member (PATRULIUS et al., 1982) and the Valea
Bobdei Limestone Member (Cociuba, 1999) (Fig.2). The latter member is also known in literature as “Middle Pachyodont Limestone”. These deposits are considered to be of Early Aptian age (early Bedoulian) based on Palorbitolina lenticularis (BLUMENBACH) and Palaeodictyoconus arabicus (HENSON), occurring together in the Valea Bobdei Limestone.
3. The Valea Măgurii Limestone Formation (COCIUBA, 1999) lies on the Ecleja marls and ends with a discontinuity surface, sometimes with feruginous crusts. It bears Palorbitolina lenticularis and Orbitolinopsis div. sp., pointing to a Bedoulian (probably late Bedoulian) age.
4. The Vârciorog Formation (COCIUBA, 1999) is a new name for the so called “Glauconitic sandstone and the upper pachyodont limestone”. In the limestone
Figure 1 – Location of the study area in the teritory of the
Apuseni Mountains.

I.I. BUCUR
54
intercalations, Mesorbitolina texana (ROEMER) is present. The Vârciorog Formation is considered of Gargasian-Albian age.
Remarks on the Dobreşti Member A few additional remarks on the Dobreşti Member
are needed here. The age of its limestones is highly controversial. Dragastan et al. (1966) identified in the Characean-bearing Limestones an assemblage bearing Atopochara trivolvis, species that could account for the Barramian-Aptian age. Nevertheless, in the caption to plate I (Dragastan et al., 1966), the deposits bearing Atopochara trivolvis are regarded as Hauterivian in age, while the text (p. 24) states that: “afin de pouvoir attribuer une valeure stratigraphique aux formes citées, il sera nécessaire de faire une étude des microfaciès du Jurassique supérieur et du Crétacé inférieur de la région…” (“in order to be able to assign stratigraphic value to the forms quoted, it will be necessary to perform a study on the microfacies from the Upper Jurassic to the Lower Cretaceous interval in the region”). Dragastan et al. (1988, 1989) also assigned
the Characean- and Gastropod-bearing Limestones to the Barremian, based on their identification of the assemblage (Dragastan et al., 1966). In several other papers (Bucur et al., 1993; Bucur & Cociuba, 1996; 1998), I have argued that the Characean- and Gastropod-bearing Limestones may be Valanginian-Hauterivian in age, due to the presence of the following species: Salpingoporella annulata, S. katzeri, Montsalevia salevensis, Meandrospira favrei and Haplophragmoides joukowskyi. In a recent paper, Dragastan (1999) questions the accuracy of identification of the four species illustrated by Bucur & Cociuba (1996) as follows: “The so called «Valanginian-Hauterivian» assemblage containing Salpingoporella annulata, S. katzeri, Montsalevia salevensis and Haplophragmoides joukowskyi, found by Bucur & Cociuba (1996) in the Gastropod limestone, corresponds in fact to a longitudinal subaxial section through a Gastropod shell (the first species) and to Salpingoporella urladanasi (the second species), a Barremian-Early Aptian taxon; as concerns the two foraminifera species, they are not conclusive and do not belong to the taxa assigned by the authors (Neagu, pers. comun.)” The Salpingoporella annulata specimen, regarded by Dragastan (1999) as a gastropod, is illustrated in the present paper in plate I, fig. 1 and plate VI, figs. 1,2. For better comparison, besides the tangential-longitudinal section characteristic for Salpingoporella annulata (“avec un tracé en zigzag rejoignant les ramifications phloïophores dilatées verticalement”, BERNIER, 1984) in Pl.I, fig. 1 and Pl. VI, figs. 1 and 2, I am also presenting a few similar sections taken from Carozzi (1953, figs. 3-6) (Pl. VI, fig. 6 in this paper), Sartoni & Crescenti (1962, Pl. XLIV, fig. 4) (Pl. VI, fig. 4 in this paper), Jafrezzo (1973, Pl. I, fig. 11) (Pl. VI, fig. 3 in this paper).
The Salpingoporella katzeri specimen, regarded by Dragastan (1999) as a S. urladanasi specimen, is also illustrated in the present paper (Pl. I, fig.2 and text-fig.3) together with other speciments of the same species (Pl. I, figs. 5,6; Pl. VI, figs. 8-10,13). The specimens presented reveal two characteristics typical for S. katzeri: (1) the polygonal (even hexagonal) shape of ramifications in external tangential section (Pl. I, fig. 5) and (2) the short, narrow and excentrically situated connected pore, at the level of the lower ramification rim, characteristic depicted by Sokac (1996). On the other hand, the ramifications in S. urladanasi reveal in tangential section a typically quadrangular shape, being highly flattened transversally (Conrad et al., 1977).
The two algal species occur in the Characean- and Gastropod-bearing Limestones together with Montsalevia salevensis and rare specimens of Haplophragmoides joukowskyi and Meandrospira favrei. For a better emphasis on the accuracy of identification of the two foraminifer species mentioned by Dragastan (1999), they are illustrated in Pl. VI, figs. 19 and 23 together with copies from the original paper (Charollais et al., 1966) on the same scale (Pl. VI, figs. 21, 22, 26-31). Other specimens of these species, collected from Valanginian-Hauterivian deposits in Padurea Craiului, have also been illustrated before (Bucur & Cociuba, 1998, Pl. I, figs. 1-9).
The presence of the four species within the Characean- and Gastropod-bearing Limestones (the Dobresti Member, Cociuba, 1999) is obvious. Figure 3 presents the sequence of deposits overlying the bauxite lens 68 at Cornet, with the exact location of samples and the microfacies sequence.
Figure 2 – General succession of the Lower Creataceousdeposits from Pădurea Craiului Mountains. 1 – bauxitic rocks; 2 – limestones; 3 – breccia; 4 – marls and shales; 5 – sandstones.

LOWER CRETACEOUS DASYCLAD ALGAE FROM THE PADUREA CRAIULUI MASSIF
55
Figure 3 – Succession of Lower Cretaceous deposits above the bauxite lens 68 (Cornet), showing the alternanting Characean- and Dasyclad bearing limestones. 1 – limestone; 2 – breccia and levels with black pabbles; 3 – bauxite; 4 – rudists; 5 – small gastropods. A, B, C, E, F – Characean –bearing biomicrites; D, G, H, I, J – Dasycladalean-bearing biomicrites: Salpingoporella annulata (D, I, J), Salpingoporella katzeri (G, J), Praturlonella nerae (H).

I.I. BUCUR
56
What could be debatable here, I think, is the biostratigraphic significance of these species. The most recent deposits bearing S. annulata have been assigned to the Hauterivian (Luperto Sinni & Masse, 1984; Masse & Santenac, 1987; Chiocchini et al., 1988; Chiocchini et al., 1994; Schindler & Conrad, 1994; Claps et al., 1996), although the species is present, within the Lower Cretaceous, mainly in Berriasian-Valanginian deposits. S. katzeri, initially reported from Berriasian-Valanginian deposits (Conrad & Radoicic, 1978), has also been assigned to the Hauterivian (Luperto Sinni & Masse, 1984; Masse & Sentenac, 1987). Besides the two Salpingoporella species, the sections analysed also bear Clypeina parasolkani, a species also reported from the entire Berriasian-Hauterivian interval (Farinacci & Radoicic, 1991; Bucur, unpublished data). Montsalevia salevensis and Haplophragmoides joukowskyi have been generally identified in Valanginian deposits but have also been reported from Hauterivian deposits (Masse, 1976; Jaffrezo, 1980; Bucur, 1988; Altiner, 1991). Moreover, the first orbitolinids characteristic for the Lower Barremian (possibly even the upper part of the Hauterivian), belonging to species Paracoskinolina? jourdanensis, occur in the column of the Lower Cretaceous deposits in the area about 40 to 70 m over the Characean- and Gastropod-bearing Limestones (approximately the terminal part of the first third of the Coposeni Member limestones).
The synthesis of these data has led me to the conclusion that the assemblage may occur within deposits ranging in age from the Valanginian to the Hauterivian. This statement comes into conflict with that of Dragastan et al. (1988, 1989), according to whom the Characean-bearing Limestones bear Atopochara trivolvis and this species accounts for their Barremian-Aptian age. The accuracy of identification of the species Atopochara trivolvis, illustrated by Dragastan et al. (1966) and presented in this paper in Pl. VI, fig. 7 is not being questioned, although Schudack (written information, 1998) states that: “…it is not possible to determine charophyte species in thin section. Therefore, no stratigraphic data can be derived from thin sections”. It is worth mentioning here that, according to the same author (Schudack, 1993), Atopochara trivolvis is present in the Lower Cretaceous within the Berriasian-Albian interval, as a sequence of subspecies and assemblages. In order to accurately assign any of these subspecies to a certain age, it is necessary to undertake a thorough study of rock-detached specimens of characeae and to correlate the resulting information with data provided by the micropaleontological analysis of dasyclad algae and foraminifer assemblages present in the limestones immediately overlying the limestones bearing characeae or inserted between them. Such a study is in progress. It is likely that the deposits overlying the bauxite and underlying the Barremian rudist-bearing limestones (the Coposeni Member, 1999) may have had a complex history, which may have led to the formation of characean-bearing limestones both in the Valanginian and the Hauterivian, or even the Lower Barremian. Several discontinuities in the above mentioned sequence, as well as the presence of certain limestones bearing Valanginian - Hauterivian microflora and microfauna, inserted among levels of characea-bearing limestones or immediately overlying them, suggest such an evolution.
CALCAREOUS ALGAE ASSEMBLAGES
All calcareous algae presented in this study were sampled from the Dobreşti Member, the Coposeni Member as well as from various levels corresponding to the “Middle an Upper Pachyodont Limestones” (Valea Bobdei Member, Valea Măgurii Formation and limestone intercalations in the Vârciorog Formation). The Dobreşti limestone Member comprises, ocasionaly, several levels rich in Salpingoporella anullata CAROZZI (Pl. I, figs. 1-3; Pl.VI, figs.1, 2, 5), S. katzeri CONRAD & RADOICIČ (Pl. I, figs. 4-6; Pl.VI, figs.8-10, 13), Clypeina parasolkani FARINACCI & RADOICIČ and rare soecimens of Praturlonella nerae (DRAGASTAN, BUCUR & DEMETER) (Pl.5, fig.12). The richest algal assemblage was found in the Coposeni limestone Member: Acroporella Radoicicae PRATURLON (Pl. III, fig. 9), Actinoporella sp. (Pl. III, figs. 14, 15, 20), Clypeina ? maslovi (PRATURLON) (Pl. III, fig. 16), Clypeina marteli EMBERGER (Pl. III, figs. 17, 21), Cylindroporella cf. barnesii JOHNSON (Pl. II, figs. 13, 16), Cylindroporella ? elliptica BAKALOVA (Pl. II, figs. 12, 20), Cylindroporella cf. lyrata MASSE & LUPERTO SINNI (Pl. II, figs. 14, 19), Cylindroporella cf. pedunculata (JAFFREZO, POISSON & AKBULUT) (Pl. II, figs. 15, 18, 21), ?Dissocladella sp. (Pl. IV, fig. 18), Falsolikanella danilovae (RADOICIČ) (Pl. V, figs. 1-11, 13, 14), Falsolikanella ? silvaeregis (BUCUR) (Pl. III, figs. 8, 19), Milanovicella pejovicae (RADOICIČ) (Pl. V, fig. 18), ?Milanovicella sp. (Pl. V, figs. 15-17), Neomeris sp. (Pl. III, fig. 13), Neomizzia dacica n. sp. (Pl. III, figs. 7, 11, 12, 18), Salpingoporella cf. biokovensis SOKAC & VELIC (Pl. I, figs. 17, 22), Salpingoporella exilis DRAGASTAN (Pl. I, figs. 18-20), Salpingoporella genevensis (CONRAD) (Pl. I, figs. 12, 13), Salpingoporella melitae RADOICIČ (Pl. I, figs. 7-8), Salpingoporella muehlbergii (LORENZ) (Pl. I, figs. 9-11), Salpingoporella patruliusi BUCUR (Pl. II, figs. 1-10), Salpingoporella cf. piriniae CARRAS & RADOICIČ (Pl. I, fig. 16, 21), Salpingoporella sp. (Pl. I, figs. 14, 15), Similiclypeina conradi BUCUR (Pl. IV, figs. 9 - 14, 16, 17), Similiclypeina paucicalcarea (CONRAD) nov. comb. (Pl. IV, fig. 7), Similiclypeina cf. somalica (CONRAD, PEYBERNES & MASSE) (Pl. IV, Figs. 15, 19).
The limestones in Valea Măgurii and Vârciorog Formations bear a relatively poor algal microflora: Anisoporella? cretacea DRAGASTAN (Pl. III, figs. 1-6), Bakalovaella elitzae (BAKALOVA) (Pl. II, Fig. 11), Neomeris cretacea STEINMANN (Pl. III, fig. 10) and Terquemella sp. (Pl. II, fig. 17). The microproblematic “Coptocampylodon” (Carpathoporella) fontis PATRULIUS is also very frequent.
PALEONTOLOGICAL REMARKS ON SOME CALCAREOUS ALGAE
Genus Anisoporella BOTERON, 1961 Anisoporella? cretacea (DRAGASTAN, 1967) BUCUR,
1995 emend. Pl. III, figs. 1-6
1967 - Pseudoepimastopora cretacea n. sp. - DRAGASTAN, p. 448, Pl. IV, figs. 32, 33, 37-39. 1969 - Pseudoepimastopora cretaacea DRAGASTAN - JOHNSON, Pl. V4, figs. 1-5. 1973 - Neomeris pfendere KONISHI & EPIS - BAKALOVA, p. 83, Pl. III, fig. 5. 1974 - Pseudoepimastopora cretacea DRAGASTAN - CHRISCHEV & BAKALOVA, p. 67, Pl. III, fig. 1.

LOWER CRETACEOUS DASYCLAD ALGAE FROM THE PADUREA CRAIULUI MASSIF
57
1975 - Pseudoepimastopora cretacea DRAGASTAN - DRAGASTAN, p. 65, Pl. LI, fig. 2. ?1975 - Harlanjohnsonella sp. - DRAGASTAN, p. 68, Pl. LXXV, fig. 3. 1978 - Pseudoepimastopora cretacea DRAGASTAN - BAKALOVA, p. 8, Pl. III, fig. 7. 1978 - Neomeris pfendere KONISHI & EPIS - BAKALOVA, p. 7, Pl. III, fig. 8. ?1980 - Harlanjohnsonella sp. - DRAGASTAN, Pl. I3, fig. 7. ?1982 - Gyroporella lukicae n. sp. - SOKAC, p. 37, Pl. I, II. 1992 - Epimastoporella cretacea (DRAGASTAN) - BUCUR, p. 451, Pl. IV, figs. 1-11. 1994 - Epimastoporella cretacea (DRAGASTAN) - BUCUR, p. 48, Pl. V, figs. 14-22. 1995 – Anisoporella cretacea (DRAGASTAN) – BUCUR, p. 87, pl.IV, figs.8, 9, 12, 13 1996 – Anisoporella cretacea (DRAGASTAN) – BUCUR & COCIUBA, p. 43, pl. I, fig.9.
Remarks: The alga described by DRAGASTAN (1967) from the Lower Cretaceous deposits of the Padurea Craiului Massif was ignored by BASSOULET at al. (1978) in the synthesis on Jurassic and Cretaceous algae, which includes only the description of one Mezozoic species assigned to the genus Pseudoepimastopora: P. jurassica ENDO, 1960. BUCUR (1992) assigned the species P. cretacea to the genus Epimastoporella ROUX, 1979, with an emended diagnosis. The specimens subsequently found in the Lower Cretaceous deposits from the Padurea Craiului Mountains, in the area mentioned by DRAGASTAN (1967) as type locality, prove that we have, in fact, to deal with an alga showing ramifications with an euspondyle arrangement (Pl. III, figs. 1, 3) and a vesiculiform shape (Pl. III, figs. 1, 4-6) . The superficial tangential sections show a round shape , the poligonal configuration (BUCUR,1992) being a result of pressures due to the dense arrangement within the verticils.
The above mentioned remarks require a series of changes as follows:
1. The alga belongs probably to the genus Anisoporella BOTTERON., 1961 (vesiculiform ramifications with an euspondyle arrangement ), thus we have to consider the new combination A?. cretacea (DRAGASTAN, 1967). It differs from Triassic species of Anisoporella in having only one row of ramifications in each verticil, thus the question mark.
2. The diagnosis of the species needs the following emendation: " Cylindrical, unsegmented thallus, with a wide axial cavity bordered by a relatively thin calcareous wall. The latter is penetrated by pores (branches) displayed in an euspondyle arrangement. The shape of branches is vesiculiform in longitudinal section and round or polygonal (due to reciprocal pressures of the branches) in tangential section . Due to thallus fragility the alga occurs mostly as fragments ."
Dimensions (according to DRAGASTAN, 1967; BUCUR, 1992 and new data in this study) :
L > 5 mm; D = 0.92 – 1.92 mm; d = 0,47 – 1.38 mm; e = 0.14 – 0.39 mm. p = 0.07-0.18 mm; l = 0.15 – 0.19 mm; h = 0.12 – 0.18 mm.
3. As shown by SENOWBARI-DARYAN et al. (1994), Gyroporella lukicae SOKAC and VELIC, 1982 belongs to the genus Anisoporella. Consequently, the mentioned autors suggested the new combination A. lukicae (SOKAC & VELIC, 1982). Moreover, comparing the general shape of the thallus, the shape and arrangement of ramifications and the ratio between the main dimensional parameters from Gyroporella lukicae SOKAC and VELIC, 1982, and Pseudoepimastopora cretacea DRAGASTAN, 1967 (see also BUCUR, 1992 and the data in the present study), we consider that both
forms may belong to the same species (though G. lukicae has generaly larger dimensions of the thallus). In this case A. lukicae is a junior synonim of A? cretacea.
4. GRANIER & DELOFFRE (1993) consider P. cretacea DRAGASTAN, 1967 as nomen nudum, owing to the attribution of the species to the non valid genus Pseudoepimastopora Endo, 1960. When the inventory made by GRANIER and DELOFFRE (1993) was published the species P. cretacea had been already validated by its transfer to the genus Epimastoporella Roux 1979 (BUCUR,1992). However, it is considered a valid species by the new transfer, suggested here, to the genus Anisoporella? BOTTERON, 1961.
5. ENDO (1961) described Pseudoepimastopora jurassica n.sp. , considered by GRANIER & DELOFFRE (1993) as nomen nudum. The species was subsequently illustrated in several papers: BOUROULLEC & DELOFFRE (1970, Pl. 7, figs. 1, 2 as Epimastopora sp.), DRAGASTAN (1975, Pl. XXIII, fig. 2; Pl. XXVIII, fig. 3; Pl. .XXXVII, fig. 3), SOTAK (1987, Pl. VII, fig. 8, as Linoporella? capriotica), FARINACCI & RADOICIČ (1991, Pl. I2, figs. 1-9 as Gyroporella sp., Gyroporellas aff. lukicae and "Pseudoepimastopora-Epimastopora" group), SOTAK & MISIK (1993, Pl. I, fig. 5). This alga belongs probably, in our opinion, to the genus Anisoporella BOTTERON, 1961. We also suggest the new combination Anisoporella? jurassica (ENDO, 1961). The specimens described and figured as Anisoporella sp. (p.228, Pl. IV, figs. 9, 10, 12, 13) and Epimastoporella jurassica (ENDO, 1961) nov. comb. (p. 228, Pl. V, fig. 9) by SENOWBARY-DARYAN at al. (1994) belong in fact to Anisoporella? jurassica (ENDO, 1961) nov. comb.
Genus Actinoporella (GÜMBEL in ALTH, 1881)
Actinoporella sp. Pl. III, figs. 14, 15, 20
Remarks: The three identified and figured specimens fit the dimensional parameters indicated for Actinoporella podolica (ALTH) by CONRAD et al. (1974). Yet, there still are two morphological features which make the mentioned species different: 1.) the existance of an "excrescence" (corona) only in the lower part of the ramifications (Pl. III, figs. 14, 15)) and 2.) the reciprocal touch of the ramifications within a verticil only in the basal part (Pl. III, fig.20).
Dimensions: D = 1.17 – 1.55 mm; d = 0.37 – 0.48 mm; l = 0.45-0.69 mm; p = 0.10 – 0.25 mm; w = 10 -12.
Genus Cylindroporella JOHNSON, 1954
Cylindroporella cf. barnesii JOHNSON, 1954 Pl. II, figs. 13, 16
Cylindroporella cf. pedunculata (JAFFREZO et al., 1980)
Pl. II, figs. 15, 18, 21 Remarks: The two algae occur associated, their
distinction being based mainly on dimensional criteria (Tab. 1). Specimens resembling Cylinroporella cf. barnesii were illustrated by LUPERTO SINNI & MASSE (1984, Pl. III, figs. 5, 6, 10), LUPERTO SINNI & MASSE (1986, Pl. II, figs. 2, 3), KUSS & CONRAD (1991, Fig. 2/12-16 (as Cylindroporella aff. barnesii). Cylindroporella pedunculata, initially described from the Western Taurides (Turkey) under the name Pseudoepimastopora pedunculata (JAFFREZO et al., 1980), and considered to be a senior synonym of the species Korkyrella ivanovici SOKAC, 1981 (MASSE & LUPERTO SINNI, 1989), was also reported by CONRAD & PEYNERNES (1976, figs. 7a-d as

I.I. BUCUR
58
Cylindroporella cf. barnesii), MASSE (1976, Pl. III, fig. 14; Pl. IV, fig. 1, as Cylindroporella aff. arsici), PEYBERNES & CONRAD (1979, Pl. II, fig. 4, as Cylindroporella barnesii), JAFFREZO et al. (1982, Pl. I, figs. 6, 7, as "Dasycladale"), LUPERTO SINNI & MASSE (1984, Pl. 34, figs. 1-8, as Cylindroporella sp.), LUPERTO SINNI & MASSE (1986, Pl. II, figs. 1, 3, as Cylindroporella sp.), MANCINELLI (1992, Pl. V, figs. 1-4, as Cylindroporella ivanovici), BODROGI et al.(1993, Pl. III, figs. 5, 6), LUPERTO SINNI & MASSE (1993, Pl. II, figs. 12-15), SOTAK & MISIK (1993, Pl. V, figs. 5, 6), BODROGI et al. (1994, Pl. I0, figs. 1-3), BUCUR (1994, Pl. III, figs. 13-21, as Korkyrella cf. ivanovici). GRANIER and DELOFFRE (1993) assign the alga described by JAFFREZO et al. (1980) to the genus Epimastoporella.
Morphologically, Cylindroporella barnesii and Cylindroporella pedunculata are similar, a fact already mentioned by CONRAD (1982). Along with some dimensional differences ( C. pedunculata has larger general dimensions), the age of the deposits where they were identified was a significant factor for their assignment by different authors to one or the other species. Thus, the specimens identified in Albian deposits have been assigned rather to C. barnesii (REY et al., 1977; GRANIER, 1988; KUSS & CONRAD, 1991; MANCINELLI, 1992; SCHINDLER & CONRAD, 1994).
However, there are still many problems to clear regarding the group of cylindroporelliform dasyclads. In our opinion, Cylindroporella barnesii and C. pedunculata, belonging to the same morphological type, represent most probably one species.
Cylindroporella cf. lyrata MASSE & LUPERTO SINNI,
1989 Pl. II, figs. 14, 19
1989 - Cylindroporella lyrata n. sp. - MASSE & LUPERTO SINNI, p. 32, Pl. I, 2. ?1994 - Cylindroporella? lyrata MASSE & LUPERTO SINNI - BOGROGI et al., p. 247, Pl. I6, figs. 1-5, 8-10.
Remarks: Some rare specimens of dasyclads identified in the Blid Limestone may be assigned to Cylindroporella lyrata MASSE and LUPERTO SINNI. This cannot be done with certainty due to the absence of more adequate sections. The affiliation of the specimens figured by BODROGI et al. (1994) to C. lyrata is also doubtful. The authors noted some differences, yet they probably considered them to have a generic significance ( this explains the question mark following the name of the genus). Some specimens seem to have a single type of primary ramifications, relatively large, ending with a bush of short secondary ramifications (Pl. I6, figs. 3 and 8 in BODROGI et al., 194). Uncertainties may be also expressed concerning the specimens figured by SCHLAGINTWEIT (1991, Pl. I9, figs. 6-13, 15, 16) as Cylindroporella lyrata.
On the other hand, BODROGI et al. (1994, p. 247) assert that: "In Voralberg erscheint gleichzeitig mit den Quarzgöttungen die Kaklalge Cylindroporella? lyrata
MASSE & LUPERTO SINNI, die im Villany Gebiet nicht vorkommt. Hingegen zeigt sich in den Apuseni-Bergen Salpingoporella patruliusi BUCUR (BUCUR, 1985), die dem vorerwühten Taxon sehr nahestehen dürfte bzw. damit identisch sein könnte.("Along with the quartz share there occurs in Voralberg the calcareous alga C. lyrata MASSE and L. SINNI, which doesn`t occur in the Villany region. On the other hand, in the Apuseni Mountains occurs S. patruliusi BUCUR (BUCUR, 1985), which must be closely related to the above mentioned taxon, to which it might be even identical)
A similarity of S. patruliusi and C. lyrata is totally superficial, eventually referring to certain cross sections through less preserved specimens of C. lyrata. Otherwise, S. patruliusi is a clearly individualised species, characterised by a narrower axial cavity and long branches, of one type and order, phloioforous and with euspondyle arrangement (= genus Salpingoporella) (see BUCUR, 1985, Pl. I-III; BUCUR, 1992, Pl. II; BUCUR, 1994, Pl. X, figs. 23, 24, and the present paper, Pl. II, figs.1-10). Cylindroporella lyrata, although having a narrow axial cavity, shows two types of primary branches (sterile and fertile) disposed alternatively within the same verticil as well as secondary branches, at the distal end of the sterile branches (= genus Cylindroporella ) (see MASSE & LUPERTO SINNI, 1989, Pl. I, II). Thus, the two algae are clearlly distinct and easy to distinguish.
Genus Falsolikanella GRANIER, 1987
Falsolikanella danilovae (RADOICIČ, 1975) Pl. V, figs. 1-11, 13, 14
Remarks: This alga, frequent in the carbonate platform deposits of the mesogean area during the Hauterivian-Aptian interval, was for many years a debated subject as regarding generic assignment. Its affiliation to the genus Likanella MILANOVIC was questioned from the very beginning. Subsequently, it has been assigned to the genus Seliporella SARTONI & CRESCENTI (SOKAC & VELIC, 1978) and Praturlonella BARATTOLO (BARATTOLO, 1978). GRANIER & TRABOLD (1995) have recently suggested the transfer of this species to the genus Falsolikanella GRANIER (see also GRANIER et al., this volume). The specimens identified in Padurea Craiului sustain this assignment. In oblic and longitudinal sections (Pl. V, figs. 3, 6, 9-11, 14) or deep tangential sections (Pl. V, fig. 13) we might notice that several ramnifications occur attached in the form of a bush to a common vestibule connected by a pore to the axial cavity. And, as a matter of fact, this type of branch-arrangement corresponds to the diagnosis of the genus Falsolikanella GRANIER, 1987.
Falsolikanella(?) silvaeregis (BUCUR, 1981)
Pl. III, fig. 8 1981 - Pseudoactinoporella silvaeregis n. sp. - BUCUR, p. 151, Pl. I, figs. 1,3,6; Pl.2, figs. 1-4 (designated holotype in pl.I, fig.1). non 1981 - Pseudoactinoporella silvaeregis n. sp. - BUCUR, Pl. I, figs. 2, 4, 5; Pl. II, fig. 5. 1985 - Pseudoactinoporella silvaeregis BUCUR - BUCUR, p. 83, Pl. IV, fig. 1. 1992 - Pseudoactinoporella? silvaeregis BUCUR - BUCUR, p. 450, Pl. III, figs. 1-11. 1994 - Pseudoactinoporella? silvaeregis BUCUR - BUCUR, p. 154, Pl. VIII, fig. 18.
Remarks: The assignment of this alga to the genus Pseudoactinoporella CONRAD 1970 has been already questioned by BUCUR (1992). GRANIER (1994) has
Table 1 - Comparative dimensions of Cylinroporella cf. barnesii JOHNSON and Cylindroporella cf. pedunculata JAFFREZO et al.
Cylindroporella cf. barnesii
Cylindroporella cf. pedunculata
D 0,42 - 0,57 0,79 - 0,88 d 0,065 - 0,12 0,15 - 0,18
p (fertile) 0,12 - 0,18 0,15 - 0,21 w ~ 6 ~ 8

LOWER CRETACEOUS DASYCLAD ALGAE FROM THE PADUREA CRAIULUI MASSIF
59
recently suggested, yet with question-mark, a transfer of the alga to the genus Actinoporella (in the new combination Actinoporella? silvaeregis). The author specified that the assignment is provisory as the alga might be also assigned to Falsolikanella GRANIER 1987. Unfortunately, no attention has been paid to our observations regarding the generic assignment of the species, where we mention that the species "probably displays a two-order ramification. The specimen in Pl. III, fig. 7 seemingly exibits a short globulous primary branch (marked with arrow) splitting in two phloiophorous branches aranged in a vertical plane" (BUCUR, 1992, p. 451). The same specimen is illustrated here in Pl. III, fig. 8 . If we assimilate the globulous primary ramification to a vestibule considering that the two large phloiophorous secondary branches build a bush, we are able to assign the species in discussion to the genus Falsolikanella GRANIER 1987. However, the material we have at the moment doesn't offer the possibility of a detailed morphological analysis. As a consequence, the assignment of the species to the genus Falsolikanella is doubtful and we further consider that this alga might belong to a new genus.
Genus Milanovicella GRANIER & BERTHOU, 1994
Milanovicella pejovicae (RADOICIČ, 1975) Pl. V, fig. 18
1969 - Clypeina pejovici sp. nov. - RADOICIČ, p. 71, Figs. 1-3. 1975 - Clypeina pejovicae orth. mut. - RADOICIČ, p. 277 (designation of a lectotype). 1978 - Likanella pejovicae RADOICIČ nov. comb. - BASSOULLET et al., p. 145, Pl. V, figs. 1, 2.
Remarks: GRANIER & DELOFFRE (1993) assign this alga to the genus Clypeina. But, the original figures prove that we are dealing with an alga having double verticiles (RADOICIČ, 1969, Fig. 1a, Figs. 2h, 2i). On the basis of this specific feature the alga has been assigned by BASSOULET et al. (1978) to the genus Likanella MILANOVIC 1960. As the alga has double verticiles and phoiophorous ramifications we should rather assign it to the genus Milanovicella GRANIER & BERTHOU, 1994.
Genus Neomizzia LEVY, 1966
Remarks: GRANIER & DELOFFRE (1993) consider the genus Neomizzia LEVY 1966 as non valid, as for the type species Neomizzia elongata LEVY, 1966 several specimens have been assigned as "holotype". In fact, we do not have to take into account Pl. I, fig. 1, as mentioned by GRANIER & DELOFFRE (1993), this figure representing a reconstruction of the alga, but Pl. I, fig. 2, a section through the sample KS-2:466, 3 m, sample assigned by LEVY (1966) as holotype. Aiming to eliminate this error of nomenclature in order to validate the species created by LEVY (1966) and also the genus Neomizzia, we designate as lectotype of the species N. elongata LEVY the specimen in the center of the photography, constituted of a succession of three articles from Pl. I, fig. 2 in LEVY (1966).
Neomizzia dacica n. sp. Pl. III, figs.7, 11, 12, 18
Origin of the name : After the Dacians, ancient inhabitants of the present Romanian teritorry before the Roman conquest during the ruling of emperor Traian.
Holotype: The specimen in Pl. III, fig. 11, sample 184-Aconi, thin section 184i, N.I.1674 deposited in the collection I.I. Bucur, Geology Department of " Babes-Bolyai " University, Cluj-Napoca.
Paratype: Specimen figured in Pl. III, fig. 12, sample 184-Aconi, thin section 184c, N.I.1668, deposited in the same collection.
Type locality: Right slope of the Ticasului Valley, source tributary of the Poieni Valley, about 500 m upstream from the confluence with the Pestiselului Valley. Coordinates: x = 4'609'360; y = 5'301'040 (Padurea Craiului, Apuseni Mountains, Romania).
Type level: Barremian Blid Formation, Coposeni Member.
Diagnosis: Articulated thallus. Small-barrel-shaped articles, with ellipsoidal axial cavity and phloiophorous ramifications in an espondile arrangement.
Dimensions: H = 0.60 – 0.71 mm; D = 0.52 – 0.85 mm; d = 0.27 – 0.39 mm; l = 0.13 – 0.24 mm; p = 0.10 – 0.15 mm;
Remarks: Neomizzia dacica n. sp. has been identified only as dissociated articles within the sediment.The shape of these articles, including the elliptical shape of the axial cavity, indicates their affiliation to an alga with a thallus made up of a succession of such articles, respectivelly to the genus Neomizzia. A reconstruction of the alga , on the basis of the articles figured in Pl. III, figs. 7,11,12,18 is given in Fig. 3. N. dacica n.sp. differs from N. elongata LEVY 1966 in the shape of the articles which is much more elongated, cylindrical, with a slight constriction in the articulation area in the case of N. elongata. The new species differs from N. filipescui DRAGASTAN 1978 (transferred to the new genus Bancilaporella by
Figure 4 – Reconstruction of the thallus of Neomizzia dacica n. sp. starting from the specimens illustrated in Pl.II, figs.7, 11, 12, 18.

I.I. BUCUR
60
Dragastan in this volume) in the variable diameter of the central cavity and in a clearer delimitation of the articles.
Genus Pseudoactinoporella CONRAD, 1970
emend. CONRAD & PEYBERNES, 1976 Pseudoactinoporella fragilis CONRAD, 1970
Pl. IV, figs. 1-6 1968 - Actinoporella podolica (ALTH) - BOUROULLEC & DELOFFRE, p. 229, Pl. V, figs. 1-3, 5-11. 1970 - Pseudoactinoporella fragilis n. gen., n.sp. - CONRAD, p. 66, text-fig. 4, Pl. I, figs.1-3, Pl. II, figs. 1-3, ?4, Pl. VIII, fig. 4. non 1970 - Pseudoactinoporella fragilis n. gen., n. sp. - CONRAD, Pl. I, fig. 4 (=Rajkaella). 1972 - Pseudoactinoporella fragilis CONRAD - FOURCADE et al., p. 240, Pl. III, figs. 7-9. 1976 - Pseudoactinoporella fragilis CONRAD - CONRAD & PEYBERNES, p. 188, Figs. 12a, b, d-g. 1979 - Pseudoactinoporella fragilis CONRAD - PEYBERNES et al., p. 182, Pl. III, fig. 7 1980 - Pseudoactinoporella fragilis CONRAD - ARNAUD-VANNEAU, Pl. I11, figs. 1, 2. 1980 - Pseudoactinoporella fragilis CONRAD - JAFFREZO, p. 331, Pl. XII, figs. 1, 2 (Further synonymy list!) 1980-81 - Pseudoactinoporella fragilis CONRAD - BUCUR, p. 57, Pl. IV, figs. 1, 2. 1982 - Pseudoactinoporella fragilis CONRAD - BUCUR et al., p. 36, Pl. XI, fig. 3. 1983 - Triploporella? aff. marsicana var. adriatica SOKAC & NIKLER - SOTAK et al., p. 334, Pl. 6, fig. 2 (pars). 1993 - PSEUDOACTINOPORELLA FRAGILIS CONRAD - MASSE, p. 318, Pl. II, fig. 15. 1993 - Pseudoactinoporella fragilis CONRAD - SOTAK & MISIK, p. 403, Pl. III, figs. 1-11. 1994 - Pseudoactinoporella fragilis CONRAD - BUCUR, p. 153, Pl. VIII, figs. 9-17. N.F.1994 - Actinoporella fragilis (CONRAD) n. comb. - GRANIER, p. 15 (Further synonymy list!).
Remarks: The existence of the genus Pseudoactinoporella CONRAD, 1970 has been recently denied by GRANIER (1994). The autor suggested the transfer of the species to the genus Actinoporella in the new combination A. fragilis (CONRAD 1970). He follows in fact a suggestion made by JAFFREZO (1980), who considered the small secondary ramifications pointed out by FOURCADE et al. (1972), ramifications which actually led to the emendation of the genus and the species (CONRAD & PEYBERNES, 1976), as elements of an upper corona of Actinoporella. In my opinion, there are two essential features accounting for the difference between the genera Pseudoactinoporella and Actinoporella:
1. The general shape of the Pseudoactinoporella thallus differs from the one of Actinoporella in the presence of a lower cylindrical part with a relatively constant diameter and an upper part where the thallus widens ( it flares out, the diameter of the axial cavities being relatively constant, suggesting a club-shaped outline ). The enlargement is mainly achieved by the elongation of the ramifications in the upper part of the thallus, where they also display a stronger inclination to the axial cavity (Pl.4, figs. 2,4; see also FOURCADE et al., 1972, Pl. III, fig. 9; CONRAD & PEYBERNES, 1976, Fig. 12d; PEYBERNES et al. 1979, Pl. III, fig. 7; SOTAK & MISIK, 1993, Pl. III, figs. 1, 2, 6). The general shape of the thallus represents one of the main classification criteria of dasyclads, from genus to subfamily level (BASSOULLET et al., 1977). As the genus Actinoporella has by definition a cylindrical thallus (CONRAD et al., 1974; GRANIER, 1994), the delimitation of the two genera is naturally established.
2. The short ramifications attached closely to the proximal extremity of the branches, noticed by
FOURCADE et.al. (1972) and used as basis for the emendation of the genus (CONRAD & PEYBERNES, 1976) , cannot be considered an upper corona in our opinion. They are not only an upper expansion of a vestibule, as in the case of Actinoporella, but they represent secondary ramifications, probably having the role of fertile ampules, as suggested by FOURCADE et.al. (1972). This interpretation is also supported by the spherical shape of the small ramifications attached to the base of the large ramification by a short and narrow peduncle (Pl. IV, figs. 3-5, arrows), a fact also noticed by FOURCADE et.al.,1972, p. 240. Consequently, the peduncle joining the branches and the axial cavity might be considered a primary ramification, giving rise to two secondary ramifications: a large, phloiophorus one, evolved approximately on the same direction with the primary ramification, and a small, spherical one, developed as a small fertile ampule on the upper part of the primary ramification and attached at its distal end by a short and narrow peduncle.
Such a developing of ramifications reminds rather of the genera Montiella MORELET & MORELLET, 1992 and Bakalovaella BUCUR, 1993, the general shape of the thallus being again a morphological feature of delimitation.
Considering this interpretation, we may wonder what biological necessity might have required these fertile ampules in the case of an alga with large branches which may have housed the organs of reproduction.The answer to this question is difficult, if not impossible. But, we can indicate at least three examples of dasyclads having far smaller fertile ampules than the primary or secondary ramifications: Cymopolia eochoristosporica ELLIOTT, 1968; Iodotella koradae (DIENI & MASSARI, 1983) PARENTE, 1997 and Iodotella deloffrei (TRAGELEHN, BUCUR & SYLVESTER, 1995 nom. nud.) (as Neomeris (Larvaria) deloffrei in TRAGELEHN et al., 1995).
On the basis of the above mentioned remarks, we consider the genus Pseudoactinoporella a distinct taxonomic entity.
Genus Similiclypeina BUCUR, 1993 Similiclypeina conradi BUCUR, 1993
Pl. IV, figs. 9-14, 16, 17 Remarks: Initially described from the Barremian
deposits of the Resita - Moldova Noua zone (Southern Carpathians) (BUCUR, 1993), this alga has been recovered in the Blid Formation (Coposeni Member) from Padurea Craiului in many relatively well preserved specimens.
Similiclypeina paucicalcarea (CONRAD 1970)
nov. comb. Pl. IV, fig. 7
1970 - Heteroporella? paucicalcarea n. sp. - CONRAD, p. 68, text-fig. 5, Pl. III, IV. 1973 - Heteroporella? paucicalcarea CONRAD - JAFFREZO, p. 80, Pl. III, figs. 9-11, 16. 1976 - Heteroporella? paucicalcarea CONRAD - CONRAD & PEYBERNES, p. 185, Fig. 10a, b, Fig. 13a. 1976 - Heteroporella? paucicalcarea CONRAD - MASSE, p. 127, Pl. IV, fig. 7. 1976 - Heteroporella? paucicalcarea CONRAD - PEYBERNES, Pl.XXIV, figs. 14-16. 1980 - Heteroporella(?) paucicalcarea CONRAD - ARNAUD-VANNEAU, Pl. II1, figs. 3-5. 1980 - Heteroporella paucicalcarea CONRAD - JAFFREZO, p. 252, Pl. XIX, figs. 1-4. 1993 - Heteroporella(?) paucicalcarea CONRAD - BUCUR et al., p. 38, Pl. II, fig. 4.

LOWER CRETACEOUS DASYCLAD ALGAE FROM THE PADUREA CRAIULUI MASSIF
61
1993 - Heteroporella? paucicalcarea CONRAD - MASSE, p. 316, Pl. II, fig. 1. 1994 - Heteroporella? paucicalcarea CONRAD - BUCUR, p. 152, Pl. VI, fig. 13.
Remarks: This alga has been tentatively assigned to the genus Heteroporella, yet it does not correspond either to the diagnosis of the genus Heteroporella PRATURLON, 1966 amend. OTT, 1968 or to the diagnosis presented by GRANIER et.al.(1994), the latter authors considering the genus as monospecific, restricted to the type species, Heteroporella lepina PRATURLAN, 1966. The presence of sterile primary ramifications, signaled by CONRAD (1970), is disputable in our opinion . However, their presence is not further mentioned by CONRAD & PEYBERNES (1976), where the authors present only data on some fertile ampules (fertile blisters).
We consider that the alga exhibits only one type of primary ramifications, joint in their proximal part and, usually, free in the distal part, with successive verticiles touching each other. Or, all these features correspond to the diagnosis of the genus Similiclypeina BUCUR 1993. Consequently, we suggest the transfer of the species under discussion to this genus.
CONCLUSIONS
The dasyclad algae assemblage from the Lower Cretaceous deposits of the Padurea Craiului Mountains, comprises species found mainly in the Coposeni Member of the Blid Limestone Formation (= “Lower Pachyodont Limestone”). Several species have also been identified in the Dobreşti limestone Member of the same Formation (“Characean- and Gastropod- bearing Limestones”), Valea Măgurii Formation and Vârciorog Formation (“Middle Pachyodont Limestone” and “Upper Pachyodont Limestone”) . We present several remarks on the species with uncertain sytematic position as well
as on some poorly known species. Thus, Pseudoepimastopora cretacea DRAGASTAN, 1967 has been tentatively transferred to the genus Anisoporella BOTTERON, 1961. Likarella? danilovae RADOICIČ, 1975 has been assigned to the genus Falsolikanella GRANIER, 1987, following GRANIER's suggestion (1995) and in agreement with GRANIER et al. (this volume). We also transfer, for the time being, Pseudoactinoporella silvaeregis BUCUR, 1981 to the same genus. Further, we consider the genus Pseudoactinoporella CONRAD, 1970 as valid. Heteroporella? paucicalcarea CONRAD, 1970 is transfered to the genus Similiclypeina BUCUR, 1993. We describe a new species of the genus Neomizzia LEVY, 1966, Neomizzia dacica n.sp., the genus being validated by the designation of a lectotype of the type species Neomizzia elongata LEVY, 1966.
Acknowledgements This paper was presented at the 6th International
Symposium on Fossil Algae (Ankara, September 1995) and was supposed to be published in the symposium volume. As five years have passed and there is no sign that the symposium volume is going to be issued, I have updated the information in the paper and have decided to publish it in the present volume.
Special thanks are due to the Alexander von Humboldt Foundation (Bonn) and to Professor Erik Flügel (Erlangen) for the financial support and for the technical facilities respectively, offered during the preparation of the present study. Part of the work was also supported from the CNCSIS grants 21/85/1998 and 84 BM
Professor Ovidiu Dragastan read a first version of the manuscript and made some valuable remarks.
REFERENCES ALTINER D. (1991) – Microfossil biostratigraphy (mainly
Foraminifers) of the Jurassic-Lower Cretaceous carbonate successions in North-Western Anatolia (Turkey). In: Farinacci A., Ager D.V. & Nicosia U. (eds.), “Geology and Paleontology of Western Pontides, Turkey”. Geologica Romana, XXVII, p.167-213, 7 figs., 16 pls., Roma
ARNAUD-VANNEAU A. (1980) - Micropaléontologie, paléoécologie et sédimentologie d'une plate-forme carbonatée de la marge passive de la Téthys: l'Urgonien du Vercors septentrional et de la Chartreuse (Alpes occidentales). Géologie Alpine, Mém. 11, 874 p., 254 figs., 115 pls., Grenoble.
BAKALOVA D. (1973) - Calcareous algae from the Lower Cretaceous in northern Bulgaria (in Bulgarian, with English summary). Bulg. Acad. Sci., Bull. Geol. Inst., ser. Paleontol., XXII, p. 81-90, 3 pls, Sofia.
BAKALOVA D. (1978) - Algues calcaires de la Formation de Brestnica (Tithonique supérieur-Barrémien supérieur). (In Bulgarian, with French abstract). Rev. Bulg. Geol. Soc., XXXIX/1, p. 3-16, 4 pls., Sofia.
BARATTOLO F. (1978) - Su di un nuova dasicladacea (alghe verdi) nel Paleocene dell'Appennino meridionale. Bull. Soc. Natur. Napoli, LXXXVII, p. 1-76, 18 figs., 1 tabl., 19 pls., Napoli.
BASSOULLET J.P., BERNIER, P., CONRAD M.A., DELOFFRE R., JAFFREZO M. (1978) - Les algues dasycladales du Jurassique et du Crétacé. Geobios, Mém. spéc. 2, 330 p., 40 pls., Lyon.
BASSOULLET J.P., BERNIER, P., DELOFFRE P., GENOT P., JAFFREZO M., POIGNANT A.F., SEGONZAC G. (1977) - Classification criteria of fossil dasycladales. In: FL�GEL E. (ed.) Fossil Algae, Springer Verlag, p.154-166, Berlin.
BODROGI I., BONA J., LOBITZER H. (1994) - Vergleichende Untersuchung der Foraminiferen- und Kalkalgen-Assoziationen der Urgon-Entwiklung des Schrattenkalks in Voralberg (Şsterreich) und der Nagyharsany Kalkstein Formation des Villany-Gebirges (Ungarn). Jubiläumschrift 20 Jahre Geol. zusammen Arbeit Şsterreich-Ungarn, 2, p.225-283, 12 figs., 2 tabl., 17 pls., Wien & Becs.
BODROGI I., CONRAD M.A., LOBITZER H. (1993) - Lower Cretaceous Dasycladales from the Villany zone, Southwest Hungary. Biogeographical significance. In: BARATTOLO et al. (eds.) Studies on benthic fossil algae; Boll. Soc. Paleontol. Ital., Spec. vol. 1, p.59-68, 2 figs., 3 pls., Modena.
BOUROULLEC J., DELOFFRE R. (1968) - Les algues du Néocomen d'Aquitaine. Bull. Centre Rech. Pau-SNPA, 2/2, p.213-261, 3 figs., 9 tabl., 6 pls., Pau.
BOUROULLEC J., DELOFFRE R. (1970) - Les algues du Jurassique aquitain. Bull. Centre Rech. Pau-SNPA, 4/1, p.79-127, 3 figs., 11 pls., Pau.
BUCUR I. (1980-1981) - Algues calcaires du Crétacé inférieur des Monts Pădurea Craiului. Nimphaea, Folia Naturae Bihariae, VIII-IX, p.53-68, 6 pls., Oradea.
BUCUR I. (1981) - Pseudoactinoporella silvaeregis n. sp. in the Lower Cretaceous limestones from Pădurea Craiului (Apuseni Mountains). Rev. Roum. Géol., Géophys., Géogr., Géologie, 25, p. 151-153, 2 pls., Bucureşti.
BUCUR I. I. (1985) - A new dasycladacean alga in the Urgonian limestones from Pădurea Craiului (Apuseni Mountains): Salpingoporella patruliusu n. sp. Rev. Roum. Géol., Géophys., Géogr., Géologie, 29, p.81-84, 1 fig., 4 pls., Bucureşti.

I.I. BUCUR
62
BUCUR I.I. (1988) – Les foraminifères du Crétacé inférieur (Berriasien-Hauterivien) de la zone de Reşiţa-Moldova Nouă (Carpathes Méridionales, Roumanie). Remarques biostratigraphiques. Revue de Paléobiologie, vol.spec.2 (Benthos ’86), p.379-389, 2 figs., 2 pls., Genève.
BUCUR I. I. (1992) - Revised description of some dasyclad species from the Romanian Lower Cretaceous. Rev. Paléobiol., 11/2, p.447-461, 1 fig., 5 pls., Genève.
BUCUR I. I. (1994) - Algues calcaires de la zone de Reşiţa-Moldova Nouă (Carpathes Méridionales, Roumanie). Rev. Paléobiol., 13/1, p.147-209, 4 figs., 22 pls., Genève.
BUCUR I.I. (1995) – Algues calcaires dans les dépôts du Jurassique supérieur-Crétacé inférieur des Monts Pădurea Craiului. Sudii şi Comunicări, Muzul Bistriţa-Năsăud, 1, p.79-89, 6 pls., Bistriţa.
BUCUR I. , BĂDĂLUŢĂ A., POPESCU O. (1982) - Date noi privind biostratigrafia depozitelor jurasice şi cretacice din partea mediană a zonei Reşiţa (Banat). D.S. Inst. Geol. Geofiz., LXVI/4 (1979), p.21-51, ( figs., 14 pls., Bucureşti.
BUCUR I. I., COCIUBA I., COCIUBA M. (1993) - Microfacies and microfossils in the Upper Jurassic-Lower Cretaceous limestones in the southern part of the Pădurea Craiului Mountains. Rom. J. Stratigraphy, 75, p.33-40, 3 figs., 9 pls., Bucureşti.
BUCUR I.I. & COCIUBA I. (1996) – Microbiostratigraphic markers in the Lower Cretaceous deposits from Pădurea Craiului (Northern Apuseni Mountains). Anuarul I.G.R., 69, suppl.1, p.40-43, 2 pls., Bucureşti.
BUCUR I.I. & COIUBA I. (1998) – La plate-forme carbonatée du Cretacé inférieur des Monts Pădurea Craiului (Monts Apuseni, Roumanie). Biostratigraphie et configuration. Studia Univ. Babeş-Bolyai, Geologie, XLIII/2, p.89-100, 5 figs., 4 pls., Cluj-Napoca.
CAROZZI A. (1953) – Découverte du genre Salpingoporella dans le Jurassique supérieur marin du Grand-Saléve (Haute-Savoie). Archives des Sciences S.P.H.N. Genève, 6/6, p.382-386, 1 fig., Genáve
CHAROLLAIS J., BRÖNNIMANN P., ZANINETTI (1966) – Troisième note sur les foraminifères du Crétacé inférieur de la région genevoise. Remarques stratigraphiques et description de Pseudotextulariella salevensis, n.sp.; Haplophragmoides joukowskyi, n.sp.; Citaella? Favrei n.sp. Archives des Sciences S.P.H.N. Genève, 19/1, p.23-48, 6 figs., 5 pls., Genève
CHIOCCHINI M., FARINACCI A., MANCINELLI A., MOLINARI V., POTETTI M. (1994) – Biostratigrafia a foraminiferi, dasicladali e calpionelle delle succesioni carbonatiche mesozoiche dell’Appenino centrale (Italia). Studi Geologici Camerti, vol. Spec. “Biostratigrafia dell’Italia centrale”, p.9-128, 22 figs., 42 pls., Camerino
CHIOCHINNI MP., MANCINELLI A. (1978) - Ricerche geologiche sul Mesozoico del Gran Sasso d'Italia (Abruzzo) III. Corelazioni microbiostratigrafiche tra facies di margine della piattaforma carbonatica e facies pelagiche del Giurasico e Cretacico inferiore. Studi geol. camerti, IV, p.19-36, 1 fig., 3 tabl., 11 pls. Camerino.
CHIOCCHINI M., MANCINELLI A. & MARCUCCI C. (1988) - Distribution of benthic foraminifera and algae in the Latium-Abruzzi carbonate platform facies (Central Italy) during Upper Malm-Neocomian. Revue de Paléobiologie, Vol. Spec. 2 (Benthos ’86), p.219-227, 3 figs., 2 pls., Genáve.
CHRISCEV CH., BAKALOVA D. (1974) - Distribution of the algae in the Emen Limestone Formation (the Lovech Urgonian group) (In Bulgarian, with English summary). Bulg. Acad. Sci., Bull. Geol. Inst., ser. Stratigraphy & Lithology, XXIII, p.65-89, Sofia.
CLAPS M., PARENTE M., NERI C., BOSELLINI A. (1996) – Facies and cycles of the S. Giovanni Rotondo Limestone (Lower Cretacious, Gargano Promontory, Southern Italy): the Borgo Celano Section. Annali dell’Università di Ferrara, vol.7, suppl., p.5-35, 17 figs., 6 pls., Ferrara
COCIUBA I. (1999) – Studiul stratigrafic al depozitelor mezozoice din sud-vestul Pădurii Craiului. Unpublished PhD Thesis, 235 p., 28 figs., 56 pls., Cluj-Napoca
CONRAD M.A. (1970) - Barremian and Lower Aptian Dasycladaceae in the area surronding Geneva (Switzerland). Geologica Romana, IX, p.63-100, 12 figs., 11
pls., Roma. CONRAD M.A. (1982) - Comments on Korkyrella and other
new Dasycladaceae. Rev. Paléobiol., 1/1, p.1-3, 1 pl., Genève.
CONRAD M.A. , PEYBERNES B. (1976) - Hauterivian-Albian Dasycladaceae from the Urgonian limestones in the French and spanish Eastern Pyrenées. Geologica Romana, XV, p.175-197, 16 figs., Roma.
CONRAD M.A., PEYBERNES B., RADOICIČ R. (1977) – Salpingoporella urladanasi, n.sp., une dasycladale du Crétacé inférieur d’Espagne et de Yougoslavie. Géologie Méditerranéenne, IV/2, p.73-82, 6 figs., 1 pl., Marseille.
CONRAD M.A. , PRATURLON A., RADOICIČ R. (1974) - The genus ActinoporellaG�mbel in Alth 1882, Dasycladales, green algae. A revision. Geologica Romana, XIII, p.1-15, 12 figs., Roma.
CONRAD M.A. & RADOICIČ R. (1978) – Salpingoporella katzeri n. sp., une dasycladale (algue calcaire) nouvelle su Berriasien et du Valanginien de la région méditerranéenne. Geol. Vjesnik, 30/1, p.69-72, 1 fig., 3 pls., Zagreb.
DRAGASTAN O. (1967) - Alge calcaroase în Jurasicul superior şi Cretacicul inferior din MunŢii Apuseni. Studii şi Cercet. Geol., Geofiz., Geogr., Geologie, 12/2, p.441-454, 8 pls., Bucureşti.
DRAGASTAN O. (1975) - Upper Jurassic and Lower Cretaceous microfacies from the Bicaz Valley basin (East Carpathians). Mem. Inst. Geol. Geophys., XXI, 87 p., 13 figs., 1 tabl., 95 pls., Bucureşti.
DRAGASTAN O. (1978) - Microfaciès de la série calcaire crétacée inférieure d'Aliman (Dobrogea de sud). D.S. Inst. Geol. Geofiz., LXIV/4 (1976-1977), p. 107-136, 2 figs., 4 tabl., 8 pls., Bucureşti.
DRAGASTAN O. (1980) - Alge calcaroase din Mezozoicul şi TerŢiarul României. Ed. Acad. RSR, 169 p., 115 figs., 80 pls., Bucureşti.
DRAGASTAN O. (1999) – Jurassic-Cretaceous calcareous algae of the Transylvanides, Inner dacides and Moesian platform (Romania). Revista Española de Micro-paleontologia, 31/2, p.185-218, 16 figs., 2 tabl., 6 pls., Madrid
DRAGASTAN O., COMAN M., ŞTIUCĂ E. (1988) - Bauxite-bearing formations and facies in the Pădurea Craiului and Bihor Mountains (Northern Apuseni). Rev. Roum. Géol., Géofiz. Géogr., Géologie, 32, p.67-81, 5 figs., Bucureşti.
DRAGASTAN O., ISTOCESCU D., DIACONU M. (1966) - Etude du niveau à Charophytes d'âge Crétacé inférieure des Monts Pădurea Craiului (Roumanie). Rev. Micropaléontol., 9/1, p.23-28, 2 pls., Paris.
DRAGASTAN O., MARINESCU M., GHEORGHE D., ŢINTEANU C. (1989) – Upper bauxite sensu D. Patrulius and some new algae of Pădurea Craiului Mts. (Northern Apuseni). Rev. Roum. Géol., Géophys., Géogr., Géologie, 33, p.55-67, 5 figs., 2 pls., Bucureşti
DRAGASTAN O., PURECEL R., BRUSTUR T. (1986) - The Upper Jurassic and Lower Cretaceous formations from the Bihor Mts. - central southern sector (Northern Apuseni). Analele Univ. Bucureşti, Geologie, XXXV, P.57-70, 9 figs., Bucureşti.
ENDO R. (1961) - Calcareous algae from the Jurassic Torinosu Limestone of Japan. The Sci. Rep. Saitama Univ., ser. B - Commemor. vol. R. Endo, p.53-75, 17 pls. Urawa.
FARINACCI A., RADOICIČ R. (1991) - Late Jurassic-Early Cretaceous dasycladales (green algae) from the western Pontides, Turkey. In: Farinacci A., Ager D.V. & Nicosia U. (eds.), “Geology and Paleontology of Western Pontides, Turkey”. Geologica Romana, XXVII, p.135-165, 1 fig., 12 pls., Roma.
FOURCADE E., JEREZ L., RODRIGUEZ T., JAFFREZO M. (1972) - El Jurasico terminal y el Cretacico inferior de la Sierra de la Muela (Provincia de Murcia). Consideraciones sobre las biozonas con foraminiferos del Albense-Aptense del sureste de Espana. Rev. Espan. Micropaleontol., Nr. Extraord. XXX Aniv. E.N. Adaro, p.215-248, 5 figs., 1 tabl., 9 pls., Madrid.
GRANIER B. (1987) - Revision de Likanella campanensis Azema er Jaffrezo, 1972, algue dasycladacée du Crétacé inférieur du sud-est de l'Espagne. Revue de Paléobiol., 6/2,

LOWER CRETACEOUS DASYCLAD ALGAE FROM THE PADUREA CRAIULUI MASSIF
63
p.207-212, 1 fig., 1 pl., Genève. GRANIER B. (1988) - Algues Chlorophyceae du Jurassique
terminal et du Crétacé inférieur en Alicante. Mediterranea, ser. estud. geol., 5 (1986), p.5-96, 12 figs., 4 tabl., 12 pls., Alicante.
GRANIER B. (1994) - The genus Actinoporella (G�mbel in Alth, 1881) and its representatives. A review. Beitrüge zur Palüontol., 19, p.113-127, 1 fig., 1 tabl., 4 pls., Wien.
GRANIER B. , BERTHOU P.Y. (1994) - Description de Milanovicella momciliana n. gen., n. sp., algue dasycladale du Portlandien de l'Algarve central (Portugal), et validation de quelques taxons affins. Rev. Micropaléontol., 37/2, p.113-121, 3 figs., 1 pl., Paris.
GRANIER B. , DELOFFRE R. (1993) - Inventaire critique des algues dasycladales fossiles. IIe partie - les algues dasycladales du Jurassique et du Crétacé. Rev. Paléobiol., 12/1, p.19-65, Genève.
GRANIER B., MASSE J.P., BERTHOU P.Y. (1994) - Heteroporella lepina Praturlon, 1967, revisited (followed by taxonomic notes on so-colled "Heteroporella" species). Beitrüge zur Palüontol., 19, p.129-141, 2 figs., 1 tabl., 2 pls., Wien.
GRANIER B. , TRABOLD G. (1995) - Occurence of genus Falsolikanella Granier during Hauterivian-early Aptian interval. 6th International Symposium on Fossil Algae and Carbonate Platforms. Abstracts, p. 13, Ankara.
IANOVICI V., BORCOS M., BLEAHU M., PATRULIUS D., LUPU M., DIMITRESCU R., SAVU H. (1976) - Geologia MunŢilor Apuseni. Ed. Acad. RSR, 631 p., 176 figs., 42 tabl., Bucureşti.K
JAFFREZO M. (1973) - Les algues calcaires du Jurassique supérieur et du Crétacé inférieur des Corbières. Rev. Micropaléontool., 16/2, p. 75-88, 1 fig., 1 tabl., 3 pls., Paris.
JAFFREZO M. (1980) - Les formations carbonatées des Corbières (France) du Dogger à l'Aptien: micropaléontologie stratigraphique, biozonation, paléoécologie. Extension des résultats à la Mésogée. Thèse Doct. d'Etat, Univ. P. er M. Curie, 614 p., 167 figs., 33 pls., Paris.
JAFFREZO M. , KOTETICHVILI E., TSIREKIDZE L. (1982) - Algues dasycladales des facies urgoniens de la RSS de Georgie (Caucase). Geobios, 15/5, p. 765-773, 1 fig., 1 tabl., 1 pl., Lyon.
JAFFREZO M., POISSON A., AKBULUT A. (1980) - Les algues du Crétacé inférieur des séries de type Bey Daglari 5Taurides Occidentales, Turquie). Bull. Min. Research Explor. Inst. Turkey, 91 (1978), p. 76-88, 2 figs., 1 tabl., 6 pls., Ankara.
JOHNSON J.H. (1954) - Cretaceous dasycladaceae from Gilespie Country, Texas. J. Paleontol., 28/6, p.787-790, 1 pl. (93), Washington.
JOHNSON J.H. (1969) - A review of the Lower Cretaceous Algae. Prof. Contrib. Colorado School Mines, 6, 110 p., 68 pls., Golden.
KUSS J., CONRAD M.A. (1991) - Calcareous algae from Cretaceous carbonates of Egypt, Sinai and Southern Jordan J. Paleontol., 65/5, p. 869-882, 6 figs., 1 tabl., Washington.
LEVY J. (1966) - Neomizzia (Dasycladacée) nouveau genre du Lias du Maroc. Rev. Micropaléontol., 9/&, p.37-39, 1 pl., Paris.
LUPERTO SINNI E., KOTETICHVILI E., TSIREKIDZE L . (1993) - New data on algae in the Urgonian limestones of Nakherala Hill (Caucasus, Georgia, CSI). In: Barattolo F. et al. (eds) - Studies on fossil benthic algae. Boll. Soc. Paleontol. Ital., Spec. vol. 1, p.287-293, 2 figs., 2 pls., Modena.
LUPERTO SINNI E., MASSE J.P. (1984) - Données nouvelles sur la micropaléontologie et la stratigraphie de la partie basale du "Calcare di Bari" (Crétacé inférieur) dans la région des Murges (Italie méridionale). Riv. Ital. Paleontol. Strat., 90/3, p.331-374, p.33-41, 6 figs., 9 pls. (33-41), Milano.
LUPERTO SINNI E., MASSE J.P. (1986) - Données nouvelles sur la stratigraphie des calcaires de plate-forme du Crétacé inférieur du Gargano (Italie méridionale). Riv. Ital. Paleontol. Strat., 92/1, p.33-66, 5 figs., 8 pls., Milano.
LUPERTO SINNI E., MASSE J.P. (1993) - The Early Cretaceous Dasycladales from the Apulia region (Southern Italy): biostratigraphic distribution and paleobiogeographic significance. In: Barattolo F. et al. (eds.) - Studies on fossil benthic algae, Bull. Soc. Paleontol. Ital., Spec. vol. 1, p.295-309, 4 figs., 3 pls., Modena.
MANCINELLI A (1992) - Distribuzione delle dasicladali (Algae verdi) nei sedimenti del Cretacico inferiore in facies di Piattaforma carbonatica dell'Appennino centro-meridionale (Lazio e Abruzzo). Studi Geologici Camerti, XII, p.7-29, 3 figs., 7 pls., Camerino.
MASSE J. P. (1976) - Les calcaires urgoniens de Provence. Valanginien-Aptien inférieur. Stratigraphie, paléontologie, les paléoenvironnements et leur évolution. Thèse Doct., 445 p., 124 figs., 11 tabl., 60 pls., Marseille.
MASSE J.P. (1993) - Early Cretaceous Dasycladales biostratigraphy from Provence and adjacent regions (South of France, Switzerland, Spain). A reference for Mesogean correlations. In: Barattolo et al. (eds.) - Studies on fossil benthic algae, Boll. Soc. Paleontol. Ital., Spec. vol. 1, p.311-324, 3 figs., 1 tabl., 2 pls., Modena.
MASSE J.P., LUPERTO SINNI E. (1989) - Cylindroporella lyrata, nouvelle espèce de dasycladale de l'Aptien inférieur du domain périméditerranéen nord-occidental. Rev. Micropaléontol., 32/1, p.30-38, 2 figs., 2 pls., Paris.
MASSE J.P. & SENTENAC F. (1987) – Variations bathimetriques et instabilité du trefonds: l’exemple des séries carbonatées à séquences loferitiques du Crétacé inférieur du Frioul (Italie septentrionale). Signification evenimentielle dans le cadre géodinamique de la Méditerranée occidentale. Mémoires géologiques de l’üniversité de Dijon, 11, p.157-166, 7 figs., Dijon.
PARENTE M. (1997) – Dasycladales from the Upper Maastrichtian of Salento Peninsula (Puglia, Southern Italy). Facies, 36, p.91-122, 8 figs, 3 tbl., 11 pls., Erlangen
PEYBERNES B. (1976) - Le Jurassique et le Crétacé inférieur des Pyrénées franco- espagnoles entre la Garonne et la Méditerranée. Thèse Doct. Sci. nat., 459 p., 149 figs., 42 pls., Toulouse.
PEYBERNES B. , CONRAD M.A. (1979) - Les algues du Crétacé inférieur de Hongrie. Bull. Cent. Rech. Explor.-Prod. Elf-Aquitaine, 3/2, p.743-752, 3 figs., 2 pls., Pau.
PEYBERNES B. , CONRQD M.A., CUGNY P. (1979) - Contribution à l'étude biostratigraphique, micropaléonto-logique et paléoécologique des calcaires urgoniens du Barrémo-Bédoulien bulgare (Prébalkan et Plate-fome Moesienne). Rev. Micropaléontol., 21/4, p.181-199, 6 figs., 3 pls., Paris.
RADOICIČ R. (1968) - Likanella? danilovae spec. nov. et quelques autres Dasycladacées crétacées inférieures des Dinarides externes. Vesnik zav. geol. geofiz. istr., 26 (A), p. 177-194, 2 figs., 16 pls., Beograd.
RADOICIČ R. (1969) - A new Lower cretaceous Dasycladacea, Clypeina pejovici, and note on some clypeinae. Gelogica Romana, VIII, p.71-84, 15 figs., Roma.
RADOICIČ R. (1970) - Algae in the Jurassic and the Cretaceous of South Hertzegovina. Geol. Glasnik, XV, p.99-107, 15 pls., Sarajevo.
RADOICIČ R. (1975) - Linoporella buseri sp. nov. from the Liassic of the Julian Alps (a preliminary report). Bull. Sci. (A), 20/9-10, p.277-278, Beograd.
REY J., BILOTTE M., PEYBERNES B. (1977) - Analyse biostratigraphique et paléontologique de l'Albien marin d'Estramadura (Portugal). Geobios, 10/3, p.369-393, 3 figs., 3 pls., Lyon.
SARTONI S. & CRESCENTI U. (1962) – Ricerche biostratigraphiche nel Mesozoico dell’Appennino Meridionale. Giornale di Geologia, sr.2, XXIX (1960-1961), p.161-304, 1 tabl., 52 pls., Bologna.
SCHINDLER U., CONRAD M.A. (1994) - The Lower Cretaceous dasycladales from the Northwestern Friuli platform and their distribution in chronostratigraphic and cyclostratigraphic units. Rev. Paléobiol., 13/1, p. 59-96, 10 figs., 5 tabl., 6 pls., Genève.
SCHLAGINTWEIT F. (1991) - Allochtone Urgonkalke im Mittlern Abschnitt der N�rdlichen Kalkalpen: Fazies,

I.I. BUCUR
64
Palüontologie und Palüogeographie. M�nch. Geowiss. Abhandlungen, A (Geol.-Pal.), 20, p.1-120, 37 figs., 6 tabl., 19 pls., M�nchen.
SCHUDACK M.E. (1993) – Die Charophyten in Oberjura und Unterkreide Westeuropas mit einer phylogenetischen Analyse der Gesamtgruppe. Berliner Geowiss. Abhandlungen, 8, 200 p., 70 figs., 20 pls., Berlin.
SENOWBARI-DARYAN B., BUCUR I.I., ABATE B. (1994) - Upper Jurassic calcareous algae from the Madonie Mountains, Sicily. Beitrüge zur Palüontol., 19, p.227-259, 2 figs., 1 tabl., 11 pls., Wien.
SOKAC B. (1987) - On some controversial Dasyclad genera and species and their stratigraphic position in the Lower Cretaceous deposits of the Dinarids. Geol. vjesnik, 40, p.9-38, 1 fig., 12 pls., Zagreb.
SOKAC B. (1996) – Taxonomic review of some Barremian and Aptian calcareous algae (Dasycladales) from the Dinaric and Adriatic Karst regions of Croatia. Geologica Croatica, 49/1, p.1-79, 3 figs., 9 tabl., 22 pls., Zagreb
SOKAC B. , VELIC I. (1978) - Biostratigrafska istrazivanja donje krede vajskih Dinarida (I). Neokom zapadne Istre. Geol. vjesnik, 30/1, p. 243-250, 8 pls., Zagreb.
SOKAC B., VELIC I. (1982) - Gyroporella lukicae n. sp. (Dasycladaceae) from the Lower Aptian of the surrounding of Jaice. Geol. vjesnik, 35, p. 37-41, 2 pls., Zagreb.
SOTAK J. (1987) - On distribution of dasycladacean algae in the Jurassic and Lower Cretaceous shallow-water limestones from the products of the Silesian Cordillera (Outer Western Carpathians). Miscellanea micropaleonto-logica, II/1, p.215-249, 1 fig., 11 pls., Hodonin.
SOTAK J., HOUSA V, MISIK M., SYKORA M. (1983) - West carpathian algae of the genus Triploporella Steinmann, 1880 - applications to Barattolo's classification. Geol. Zbornik - Geol. Carpathica, 39/3, p.323-352, 7 figs., 8 pls., Bratislava.
SOTAK J., MISIK M. (1993) - Jurassic and Lower Cretaceous dasycladalean algae from Western Carpathians. In: Barattolo et al. (eds.) - Studies on fossil benthic algae, Bull. Soc. Paleontol. Ital., Spec. vol. 1, p. 383-404, 2 figs., 12 pls., Modena.
TRAGELEHN H., BUCUR I.I., SYLVESTER Z. (1995) - On a new dasyclad species from the Paleocene of Austria and Romania. 6th International Symposium on Fossil Algae and Carbonate Platforms. Abstracts, p.28, Ankara.
VELIC I., SOKAC B. (1978) - Biostratigrafska analiza jure i donje krede sire okolice Ogulina (sredsnja Hrvatska). Geol. vjesnik, 30/1, p.309-337, 1 fig., 12 pls., Zagreb.

LOWER CRETACEOUS DASYCLAD ALGAE FROM THE PADUREA CRAIULUI MASSIF
65
PLATES
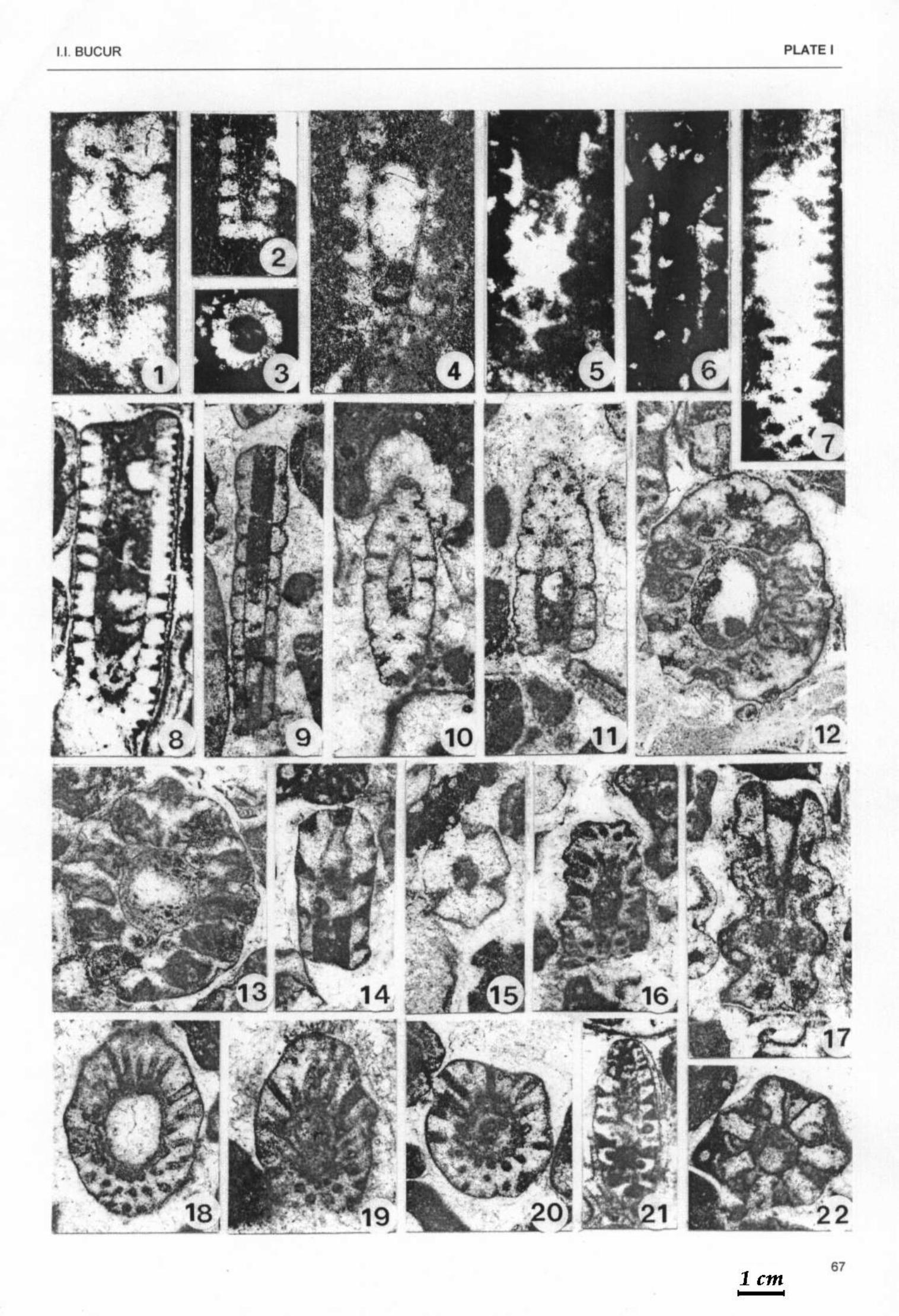
Plate I Figs.1-3 - Salpingoporella annulata CAROZZI. 1 - longutudinal-tangential section, sample 3688, x 85; 2 –
longitudinal-oblique section, sample 3732, x 40; 3 - transverse section, sample 3726, x 45. Figs. 4-6 - Salpingoporella katzeri CONRAD & RADOICIČ. 4 - longitudinal-oblique section, sample 3733, x 68; 5 -
oblique section, sample 3688, x 85; 6 - longitudinal section, sample 3726, x 40. Figs. 7, 8 - Salpingoporella melitae RADOICIČ. 7 - oblique-tangential section, sample 364 Beftia, x 45; 8 -
longitudinal-oblique section, sample 171c Vârciorog, x 45. Figs. 9 -11 - Salpingoporella muehlbergii (LORENZ). 9 - longitudinal section, sample 3779, x 33; 10, 11 - oblique
sections, sample 3782; 10 x 40; 11 x 50. Figs. 12, 13 - Salpingoporella genevensis (CONRAD). Transverse-oblique sections, sample 3800, x 33. Figs. 14, 15 - Salpingoporella sp. 14 - longitudinal-oblique section, sample 3781, x 40; 15 - transverse section,
sample 3783, x 35. Figs. 16, 21 - Salpingoporella cf. piriniae CARRAS & RADOICIČ. Longitudinal-oblique sections; 16 – sample 3781,
x40; 21 - sample 171a Vârciorog, x 45. Figs. 17, 22 - Salpingoporella cf. biokovensis SOKAC & VELIC. 17 - oblique section, sample 3783, x 45; 22 -
transverse section, sample 3780, x 50. Figs. 18-20 - Salpingoporella exilis (DRAGASTAN). Transverse-oblique sections. 18 - sample 3781, x 40; 19 -
sample 3783, x 50; 20 - sample 3780, x 45. Plate II Figs. 1-10 - Salpingoporella patruliusi BUCUR. 1 – longitudinal-oblique section, sample 3871, x 25; 2, 3 -
longitudinal-tangential sections (2, sample 3777, x 15; 3, sample 3782, x 25); 4-8 - transverse and transverse-oblique sections (4, sample 3779, x 30; 5, sample 3783, x 33; 6, sample 3769, x 40; 7, sample 3781, x 25; 8, sample 3781, x 30); 9, 10 - longitudinal-oblique sections (9, sample 3773, x 30; 10, sample 3781, x 20).
Fig. 11 - Bakalovaella elitzae (BAKALOVA). Oblique section, sample 182 b Vârciorog, x 45. Figs.12, 20 - Cylindroporella? elliptica BAKALOVA. Oblique sections. 12 - sample 144; 20 - sample 3164, x 45. Figs. 13, 16 - Cylindroporella cf. barnesii JOHNSON. 13 - oblique section, sample 3781, x 50; 16 - longitudinal-
tangential section, sample 3782, x 30. Figs. 14, 19 - Cylindroporella cf. lyrata MASSE & LUPERTO SINNI. 14 - transverse-oblique section; 19 - transverse
section, sample 3185A, x 40. Figs. 15, 18, 21 - Cylindroporella cf. pedunculata (JAFFREZO, POISSON & AKBULUT). 15, 18 - oblique sections
(15, sample 3757, x 40; 18, sample 3783, x 30); 21 - oblique-tangential section, sample 3780, x 50. Fig. 17 - Terquemella sp., sample 274 Aconi, x 50. Plate III Figs. 1- 6 – Anisoporella? cretacea (DRAGASTAN). 1 - longitudinal-oblique section, sample 3079, x 15; 2, 3 -
oblique-tangential sections (2, sample 57 Hiller, x 25; 3, sample 3079, x 40; 4, 6 - fragments in longitudinal section (4, sample 3079, x 45; 6, sample 3079, x 40); 5 - fragment in tangential section, sample 3079, x 45.
Figs. 7, 11, 12, 18 - Neomizzia dacica n. sp. Longitudinal and longitudinal oblique sections, sample 184 Aconi, x 45. Figs. 8, 19 - Falsolikanella? silvaeregis (BUCUR). 8 - longitudinal section, sample 7 Aconi, x 30; 9 - transverse-
oblique section, sample 184 Aconi, x 45. Fig.9 - Acroporella radoicicae (PRATURLON). Longitudinal-oblique section, sample 7 Aconi, x 45. Fig. 10 - Neomeris cretacea STEINMANN. Transverse section, sample 3163, x 25. Fig. 13 - Neomeris sp. Transverse-oblique section, sample 3781, x 20. Figs. 14, 15, 20 - Actinoporella sp. 14, 15 - oblique sections; 20 - transverse section, sample 355, x 45. Fig. 16 - Clypeina? maslovi PRATURLON. Transverse-oblique section, sample 7 Aconi, x 40. Figs. 17, 21 - Clypeina marteli EMBERGER. 17 - oblique section; 2& - longitudinal-tangential section, sample 3267,
x 30.

I.I. BUCUR
66
Plate IV Figs. 1-6 - Pseudoactinoporella fragilis CONRAD. 1 - Transverse-oblique section through the expanding part of the
thallus, sample 206/93 Cociuba, x 15; 2 - longitudinal oblique section; specimen from the Reşiţa-Moldova Nouă zone showing the enlaeged upper part of the thallus, sample 3955, x 12; 3, 4 - enlargements of the lower part of the specimen in Fig.2 showing the small ampullae (arrows), x 60; 5 - enlargement of the specimen in Fig. 6 showing the thin peduncle of the fertil ampulae (arrow), x 60; 6 - longitudinal-oblique section of a speciman from the Reşiţa-Moldova Nouă zone showing the bigining of the enlarged upper part of the thallus. Sample 3879, x 18.
Fig. 7 - Similiclypeina paucicalcarea (CONRAD). Oblique section, sample 291 Aconi, x 30. Figs.9-14, 16, 17 - Similiclypeina conradi BUCUR. 9, 12 - Fragments in longitudinal section (9, sample 3781, x 30;
12, sample 3782, x 30); 10 - tangential section, sample 3780, x 30; 11, 17 - oblique-tangential sections (11, sample 37??, x ?; 17, sample 3783, x 30); 13, 14, 16 - transverse sections (13, sample 3781, x 30; 14, sample 3780, x 20; 16, sample 3757, x 20).
Figs. 15, 19 - Similiclypeina cf. somalica (CONRAD, PEYBERNES & MASSE). Longitudinal-oblique sections. 15 - sample 3758, x 40; 19 - sample 3757, x 35.
Fig. 18 - Disocladella? sp. - transverse-oblique section, sample 355 Aconi, x 40 Plate V Figs. 1-11, 13, 14 - Falsolikanella danilovae (RADOICIČ). 1, 4 - Transverse-oblique sections (1, sample 3757, x 20;
4, sample 3783, x 20); 2, 5, 6 - transverse sections (2, sample 3781, x 20; 5, sample 3773, x 15; 6, sample 3783, x 20); 3, 7, 9 - logitudinal-oblique sections (3, sample 3757, x 15; 7, sample 3757, x 30; 9, sample 3782, x 20); 8, 13 - longitudinal-tangential sections (8, sample 3757, x 20; 13, sample 3781, x20); 10, 11, 14 - fragments in longitudinal section (10, sample 3782, x 25; 11, sample 184 Aconi, x 45; 14, sample 3757, x 13).
Fig. 12 - Praturlonella nerae (DRAGASTAN, BUCUR & DEMETER). Longitudinal section, sample 3732, x 55. Fig. 15 - Milanovicella? sp. Oblique section, sample 3781, x 35. Fig. 16 - Milanovicella? sp. Longitudinal section, sample 3780, x 30. Fig. 17 - Milanovicella? sp. Longitudinal-tangential section, sample 3782, x 40. Fig. 18 - Milanovicella pejovicae (RADOICIČ). Transverse and longitudinal-tangential sections, sample 3800, x 25. Plate VI Figs. 1-6 – Saklpingoporella annulata CAROZZI. 1 – Microfacies with Salpingoporella annulata, sample 3688, x 16; 2
– longitudinal-tangential section (enlargement of the specimen in fig.1), sample 3688, x 60; 3 – specimen illustrated by Jaffrezo (1973, pl.1, fig.11), x 45; 4 – specimen illustrated by Sartoni & Crescenti (1962, Pl.XLIV, fig.4), x 40; 5 – transverse sections, sample 3728, x 26; 6 – specimens illustrated by Carozzi (1953, figs.3-6), x 45.
Fig. 7 – Atopocara trivolvis PECK. Specimen illustrated by Dragastan et al. (1966, Pl.1, fig.1), x 60. Figs.8-10, 13 – Salpingoporella katzeri CONRAD & RADOICIČ. 8 – oblique section, sample 5744, x 58; 9 – oblique
section, sample 3726, x 58; 10 – transverse section, sample 5744, x 58. Figs.11, 12, 15, 16 – Clypeina parasolkani FARINACCI & RADOICIČ. 11 – Transverse-oblique section, sample
5744, x 58; 12 – transverse-oblique section, sample 3726, x 55; 15 – oblique section, sample 3726, x 58; 16 – transverse-oblique section, sample 3733, x 55.
Fig.14 – Salpingoporella cf. Hispanica CONRAD & GRABNER. Sample 5744, x 45. Figs. 17, 18 – Montsalevia / Vercorsella sp. (Pseudotextulariella n. sp. In BUCUR, 1988, Pl.I, figs.7-10). Figs.19-22, 26-28 – Montsalevia salevensis CHAROLLAIS, BROENNIMANN & ZANINETTI. 19 – longitudinal-
tangential section, sample 3689, x 100; 20 – transverse-oblique section, sample 6689, x 100; 21, 22, 26-28 – illustrations in Charollais et al., 1966, text-fig. pag.31 (figs.26, 27 in this paper) and pl.1, figs 1, 2 and 4 (figs. 28, 21 and 22 respectively in this paper), all figs. X 100.
Figs.23, 24, 29-31 – Haplophragmoides joukowskyi CHAROLLAIS, BROENNIMANN & ZANINETTI. 23 – Axial section, sample 3689, x 100; 24 – equatorial section, sample A12-Hiller, x 100; 29-31 – illustrations in Charollais et al., 1966, pl.2, figs. 1, 5 and 7 (figs. 29, 30 and 31 respectively in this paper), all figs. X 100.
Fig.25 – Meandrospira favrei (CHAROLLAIS, BEOENNIMANN & ZANINETTI), sample 3732, x 100.