maintenance of resting tension in the american eel (anguilla rostrata l.) heart is dependent upon...
TRANSCRIPT

JOURNAL OF EXPERIMENTAL ZOOLOGY 286:707–717 (2000)
© 2000 WILEY-LISS, INC.
Maintenance of Resting Tension in the American Eel(Anguilla rostrata L.) Heart Is Dependent UponExogenous Fuel and the Sarcoplasmic Reticulum
JOHN R. BAILEY, TARA BARTER, AND WILLIAM R. DRIEDZIC*Department of Biology, Mount Allison University, Sackville, New Brunswick,Canada E4L 1G7
ABSTRACT The relationship between extracellular glucose and management of cell Ca2+ inthe heart of the American eel (Anguilla rostrata) was indirectly assessed by monitoring the perfor-mance of isolated ventricular strips at 20°C. Twitch force increased in ventricular strips underspecific conditions of 30 bpm pacing and an extracellular Ca2+ challenge from 1.5 to 9.5 mM. Theresponse was independent of any exogenous metabolic fuel in the medium. Resting tension wasmaintained when glucose was available, but in the absence of a metabolic fuel, resting tensionincreased in response to the increase in extracellular Ca2+ level. When ventricular strips weretreated with iodoacetate to inhibit glycolysis, a Ca2+ challenge resulted in a decrease in twitchforce in association with an approximately equivalent increase in resting tension even in the pres-ence of exogenous glucose. However, when pyruvate (5 mM) was substituted as a metabolic fuel,twitch force increased as a function of extracellular Ca2+, and resting tension was maintained inthe presence of iodoacetate. Therefore, there is a need for an extracellular fuel but not a specificmetabolic requirement for glucose to maintain the performance characteristics, which are presum-ably related to the management of intracellular Ca2+ levels. Ventricular strips were treated withryanodine to inhibit Ca2+ release and uptake by the sarcoplasmic reticulum (SR). Ryanodine treat-ment impaired postrest potentiation at high extracellular Ca2+ levels. In the presence of ryanodine,the protective effect of glucose on the increase in resting tension in the face of an extracellularCa2+ challenge was eliminated. Considered together, the results reveal that the heart of the Americaneel has a requirement for an extracellular fuel to manage intracellular Ca2+ at high Ca2+ loads,and that the SR plays a role in the beat-to-beat regulation of Ca2+ at a frequency of 30 bpm, highCa2+ load, and 20°C. J. Exp. Zool. 286:707–717, 2000. © 2000 Wiley-Liss, Inc.
Grant sponsors: Natural Sciences and Engineering Research Coun-cil of Canada and the New Brunswick Heart and Stroke Foundation.
*Correspondence to: William R. Driedzic, Ocean Sciences Cen-tre, Memorial University of Newfoundland, St. John’s, NF, CanadaA1C 5S7.
Received 21 December 1998; Accepted 29 October 1999
Heart performance is dependent upon Ca2+ toactivate the contractile proteins and metabolic fu-els to provide energy to support ATP demand. Inthe teleost heart, contraction is generally consid-ered to be activated by Ca2+ entering across thesarcolemma (Tibbits et al., ’91), but at high tem-peratures, at least in the heart of rainbow troutand tuna, the sarcoplasmic reticulum (SR) maycontribute Ca2+ to the excitation-contraction pro-cess as well (Hove-Madsen, ’92; Shiels and Farrell,’97). Relaxation involves trans-sarcolemmal Ca2+
efflux and uptake by the SR in some species, un-der some conditions (Tibbets et al., ’91). Extra-cellular glucose, fatty acids, and stored glycogenare the primary metabolic fuels which supply theenergy necessary to rephosphorylate ADP. Fattyacids as metabolic fuels are generally more im-portant for the teleost heart at low, as opposed tohigh, temperatures (Driedzic, ’92). Thus, by theprocess of elimination, carbohydrate fuels aremore critical at higher temperatures.
There are a number of studies which suggestthat there is an interplay between glucose andCa2+ management that goes beyond their indepen-dent contributions to contraction. The companionpaper to this study shows that without glucose inthe bathing medium, ventricular strips fromAmerican eel heart do not relax under conditionsof increasing Ca2+ concentration (Bailey et al., ’99).In a marine teleost, Zoarces viviparous, ventricu-lar strips from hypoxia-adapted individuals dis-play a glucose-dependent, Ca2+ enhancement ofcontractile force under anoxia (Driedzic et al., ’85).In rat skeletal muscle, the inhibition of Ca2+ re-lease from the SR blocks enhanced glucose trans-port associated with hyperglycemia (Nolte et al.,

708 J.R. BAILEY ET AL.
’95). In rat heart, an intact glycolysis, even whenpyruvate is available, is required to decrease phar-macologically induced increases in intracellularCa2+ and subsequent hypercontracture (Nakamuraet al., ’93; Aasum et al., ’98). Under oxygen-limit-ing conditions, Ca2+ uptake by rat heart SR is de-pendent upon glycolytically generated ATP, as anumber of glycolytic enzymes are associated withthe sarcoplasmic reticular membrane (Xu et al., ’95).The above findings motivated us to conduct a se-ries of experiments to assess the interplay betweenextracellular Ca2+ and glucose on the contractilityof the American eel heart.
Twitch-force development and resting tensionwere monitored in ventricular strips under a va-riety of conditions of extracellular Ca2+ load, fuelavailability, and inhibition of sarcoplasmic reticu-lar function. The most salient finding is that inthe American eel heart an extracellular fuel, butnot necessarily glucose, is critical to relaxationunder some conditions. A functional SR contrib-utes to the presumptive decrease in cytosolic Ca2+
during diastole.
MATERIALS AND METHODSAnimals
American eel (Anguilla rostrata Lesueur) werepurchased from South Shore Trading Co., PortElgin, NB. Animals were transported to MountAllison and kept in flow-through aquaria at 13–15°Cuntil use. In all experiments, eels were removedfrom the holding tank, rendered torpid in ice wa-ter, and doubly pithed. Hearts were quickly excisedand placed in a beaker of cold Ringer’s solution.
MediaThe bathing medium was a basic Ringer’s for
freshwater fishes (Hoar and Hickman, ’67), contain-ing 111 mM NaCl, 2.0 mM KCl, 1.5 mM CaCl2, 1.0mM MgSO4, and 5.0 mM NaH2PO4, modified by ad-dition of extra bicarbonate (10 mM) to aid in long-term buffering to pH 7.8 at 20°C. The medium wasgassed with 0.5% CO2 : balance O2 for the durationof the experiment. Increased Ca2+ in the bathingmedium was achieved by the addition of aliquots ofa CaCl2 solution, giving a concentration increase of1 mM with each addition. Glucose (5 mM) wasadded as an exogenous substrate in some experi-ments, and in other cases pyruvate (5 mM) wasused. Iodoacetate (1 mM) was used as a glycolyticinhibitor in some experiments, and in othersryanodine (12.5 µM) dissolved in either DMSO orabsolute ethanol was used to inhibit the contribu-tion of SR Ca2+ movement. Low levels of ryanodine
lock the Ca2+ release channel in the open position(Coronado et al., ’94). The experimental tempera-ture was 20°C throughout.
Ventricular strip preparationsThe ventricle was dissected free of the atrium
and bulbus and bisected. Each section of ventriclewas rinsed in cold bathing medium to wash outblood. Two or more strips were cut from eachheart. Each strip was clamped in a bath contain-ing 20 mL of the medium. The ventricular stripswere placed between platinum wire electrodes andtied to a Harvard Apparatus (Saint-Laurent, Que-bec, Canada) isometric force transducer (model 60-2994), which was interfaced to a Biotronix BL-882strip chart recorder. Ventricular strips were stimu-lated to contract via field stimulation with a GrassMedical Instruments (Quincy, MA) model S9square-wave generator connected to the elec-trodes. Suprathreshold voltage, determined at thestart of each experiment, was used throughout.
Experimental protocolsCa2+ titration at variable frequency
This experiment assessed the impact of extra-cellular Ca2+ level at variable levels of Ca2+ traf-ficking. Three experimental frequencies, 12, 30,and 48 beats/min (bpm), were used as a means ofplacing different demands on the Ca2+ regulatorysystems. In each experiment, four ventricularstrips were prepared and equilibrated for 15 minat 30 bpm. Two strips had exogenous glucose inthe media. The other two had no exogenous fuelsource. Following the equilibration period, the fre-quency was set to 12, 30, or 48 bpm and pacingwas continued for 3 min. Ca2+ concentration wasincreased by 1 mM, and this step was repeated at3-min intervals until a final concentration of 9.5mM Ca2+ was obtained. Data from ventricularstrips under the same conditions were averaged.
Glycolytic requirementsThese experiments evaluated the necessity of
an intact glycolysis for contractile performance.Three ventricular strips were set up and allowedto equilibrate for 15 min at 30 bpm. One striphad glucose plus iodoacetate in the bath; one stripreceived iodoacetate but no glucose; and the thirdstrip received glucose but no iodoacetate. Follow-ing the equilibration period, a Ca2+ titration asdescribed above was performed. The second partof this study used pyruvate as the exogenous fuelsource rather than glucose; otherwise the proto-col remained the same.

RESTING TENSION IN THE EEL HEART 709
Frequency challenge with andwithout glucose
The purpose of this experiment was to assessthe role of extracellular glucose over a range offrequencies and, as such, different levels of Ca2+
trafficking. Four ventricular strips were preparedand equilibrated for 15 min at 30 bpm. Two ven-tricular strips had 5 mM glucose in the medium,and the other two ventricular strips had no exog-enous glucose present in the medium. Ca2+ con-centration was maintained at 1.5 mM. After theinitial equilibration period, the contraction fre-quency was set at 12 bpm. After 3 min the con-traction frequency was increased by 6 bpm andthe strips were paced at the higher frequency foranother 3 min. This step was repeated at 3-minintervals until a frequency of 60 bpm was reached.Data from ventricular strips under the same con-ditions were averaged.
Postrest potentiation with and withoutryanodine
The objective of these experiments and those de-scribed in the next section was to assess the role ofthe SR in Ca2+ management. The potentiation oftwitch force following a rest period is considered tobe indicative of Ca2+ storage and subsequent releaseduring the resumed contractions.
In the first study, two ventricular strips wereset up and allowed to equilibrate for 15 min at 30bpm with glucose in the medium, and at a Ca2+
concentration of 1.5 mM. Following the equilibra-tion period, stimulation was discontinued for ei-ther 0.5, 1, 5, or 10 min in randomized order.Contraction peak height was recorded for the firstthree contractions following the resumption ofstimulation. The strips were allowed to contractfor 5 min at 30 bpm between experimental peri-ods. Data from ventricular strips under the sameconditions were averaged.
In a second experiment, four ventricular stripswere set up and allowed to equilibrate for 15 minat 30 bpm with glucose present, and at a Ca2+
concentration of 4.5 mM. Two strips were treatedwith ryanodine dissolved in absolute ethanol (10µL); one strip was treated with an equivalent vol-ume of absolute ethanol; and one strip was un-treated. Postrest experiments were carried out asdescribed above. In this case, the data from theryanodine-treated strips were averaged.
Ca2+ titration with and without ryanodinePaired ventricular strips were equilibrated for
15 min at 30 bpm in basic medium without glu-
cose. One strip was treated with ryanodine dis-solved in DMSO (10 µL), and one strip receivedryanodine and glucose. The strips were left for 30min to equilibrate. A Ca2+ titration was done from1.5 to 9.5 mM, as described above.
In the second part of this experiment, pairedventricular strips were set up and equilibrated for15 min at 30 bpm in basic medium plus glucose.After this equilibration period, ryanodine, dis-solved in absolute ethanol (10 µL), was added toone bath, and a similar volume of absolute etha-nol was added to the second bath. The ventricu-lar strips were allowed to reequilibrate, as before.Following the second equilibration period, a Ca2+
titration from 1.5 to 9.5 mM was carried out, asdescribed above.
Data analysisThe twitch force of contraction was calculated
from strip chart recordings and expressed as apercentage of an initial twitch force measured at1.5 mM Ca2+ concentration. Resting tension wasexpressed as a percentage of the initial twitchforce of contraction. Postrest twitch force was cal-culated from the average peak height of the firstthree contractions following the resumption ofstimulation. There was little variation in the peakheight of the first three contractions. Postrest po-tentiation is expressed as a percentage of the con-traction immediately preceeding the cessation ofstimulation. Differences between means were ana-lyzed using a nonparametric Mann-Whitney “U”test (paired or nonpaired, as appropriate). Mul-tiple comparisons between groups and withingroups were carried out using a nonparametricKruskal-Wallis test. P < 0.05 was considered tobe significant.
RESULTSCa2+ titration at variable frequency
The presence or absence of glucose in the me-dium had no effect on twitch force generated byisolated ventricular strips with increasing Ca2+
concentration at any contraction frequency. Ven-tricular strips stimulated to contract at either 12bpm or 30 bpm showed a similar response in thatwhen extracellular Ca2+ concentration was in-creased there was a significant correlation withincreasing force of contraction (Fig. 1A,B). How-ever, strips stimulated to contract at a frequencyof 48 bpm did not display the increase in twitchforce in response to the increasing Ca2+ concen-tration seen at the other two frequencies (Fig. 1C).
There was no change in resting tension in re-

710 J.R. BAILEY ET AL.
sponse to an increase in Ca2+ level by ventricularstrips stimulated to contract at 12 bpm with eitherglucose present or glucose absent in the medium(data not shown). Ventricular strips stimulated tocontract at 30 bpm maintained resting tension ifglucose was available in the medium, but showedan increase in resting tension if glucose was ab-sent (Fig. 2A). In the absence of glucose, there wasa significant increase in resting tension when Ca2+
was increased from 1.5 to even 2.5 mM. Restingtension continued to increase to a level of about 30%of initial twitch force, at a Ca2+ level of 4.5 mM.Ventricular strips stimulated to contract at 48 bpm
Fig. 1. Force generation in ventricular strips from eelheart subjected to a Ca2+ challenge. All values are means ±SEM and N = 6 in all cases. Square symbols represent ven-tricular strips that had glucose in the medium. Circles repre-sent strips that did not receive an exogenous fuel. A: Pacedat 12 bpm. B: Paced at 30 bpm. C: Paced at 48 bpm.
Fig. 2. Resting tension in ventricular strips from eel heartsubjected to a Ca2+ challenge. All values are means ± SEMand N = 6 in all cases. Square symbols represent ventricularstrips that had glucose in the medium. Circles represent stripsthat did not receive an exogenous fuel. A: Paced at 30 bpm;asterisks indicate that resting tension is significantly differentbetween preparations with and without glucose in the mediumat Ca2+ levels of 2.5 mM and above. B: Paced at 48 bpm.
RESTING TENSION IN THE EEL HEART 711
showed a positive correlation between the restingtension and increasing Ca2+ concentration with orwithout glucose in the medium (Fig. 2B). The rest-ing tension reached about 30% of the initial twitchforce, at high Ca2+ levels in both cases.
Glycolytic requirementWhen ventricular strips, paced at 30 bpm, were
treated with iodoacetate and subjected to a Ca2+
challenge, twitch force failed after the 3.5 mM Ca2+
concentration such that by the 9.5 mM Ca2+ con-centration twitch force was approximately 75% ofthe initial force (Fig. 3A). The presence of glucosein the medium did not protect twitch force againstthe iodoacetate effect. Resting tension showed asignificant increase in ventricular strips treatedwith iodoacetate. Again, extracellular glucose didnot protect against this increase (Fig. 3B). The
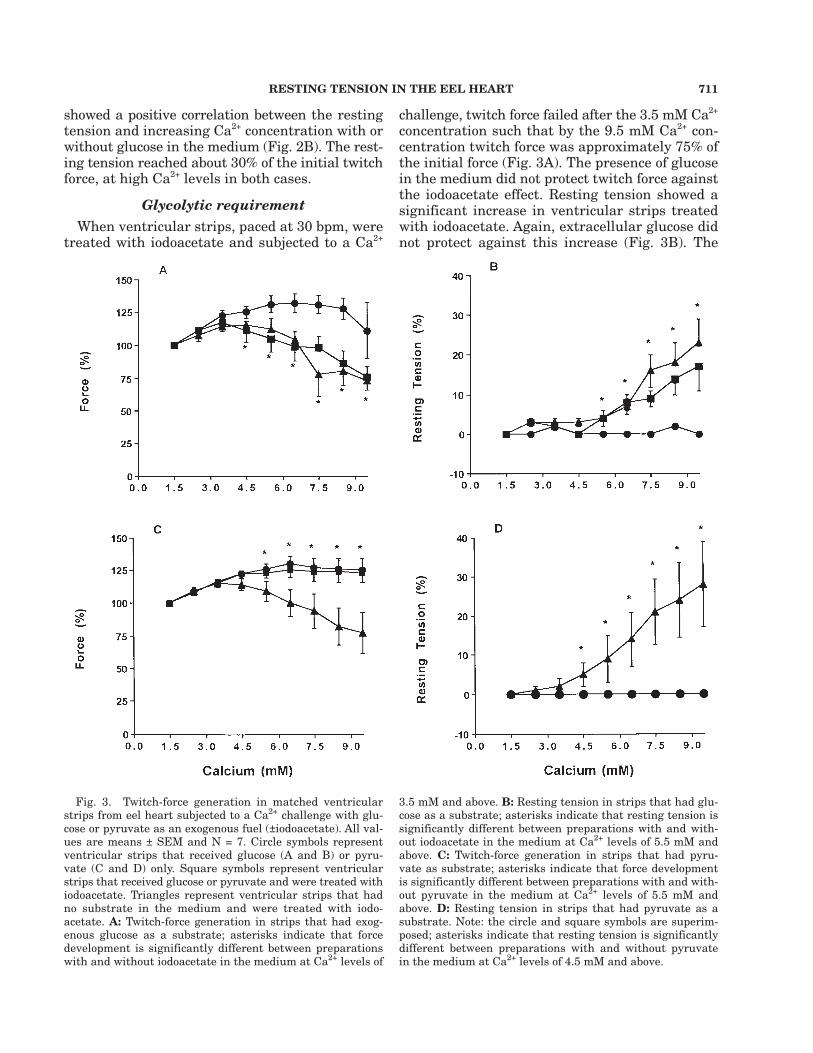
Fig. 3. Twitch-force generation in matched ventricularstrips from eel heart subjected to a Ca2+ challenge with glu-cose or pyruvate as an exogenous fuel (±iodoacetate). All val-ues are means ± SEM and N = 7. Circle symbols representventricular strips that received glucose (A and B) or pyru-vate (C and D) only. Square symbols represent ventricularstrips that received glucose or pyruvate and were treated withiodoacetate. Triangles represent ventricular strips that hadno substrate in the medium and were treated with iodo-acetate. A: Twitch-force generation in strips that had exog-enous glucose as a substrate; asterisks indicate that forcedevelopment is significantly different between preparationswith and without iodoacetate in the medium at Ca2+ levels of
3.5 mM and above. B: Resting tension in strips that had glu-cose as a substrate; asterisks indicate that resting tension issignificantly different between preparations with and with-out iodoacetate in the medium at Ca2+ levels of 5.5 mM andabove. C: Twitch-force generation in strips that had pyru-vate as substrate; asterisks indicate that force developmentis significantly different between preparations with and with-out pyruvate in the medium at Ca2+ levels of 5.5 mM andabove. D: Resting tension in strips that had pyruvate as asubstrate. Note: the circle and square symbols are superim-posed; asterisks indicate that resting tension is significantlydifferent between preparations with and without pyruvatein the medium at Ca2+ levels of 4.5 mM and above.

712 J.R. BAILEY ET AL.
increases in resting tension were similar in mag-nitude to the decreases in twitch tension and, assuch, peak tension remained about constant.
A separate set of preparations confirmed thatventricular strips exposed to iodoacetate losttwitch force at high Ca2+ levels in association withan increase in resting tension (Fig. 3C,D), butwhen pyruvate was present as a substrate therewas no loss of twitch force development as a re-sult of iodoacetate treatment. Also, ventricularstrips receiving pyruvate maintained resting ten-sion even with iodoacetate in the medium.
Frequency challenge with andwithout glucose
Ventricular strips were subjected to a frequencychallenge from 12 to 60 bpm. Twitch force wasmaintained between 12 and 36 bpm. Thereafter,twitch force had a negative correlation with fre-quency in ventricular strips, with or without glu-cose present in the medium (Fig. 4A). Increasesin contraction frequency resulted in an increasein resting tension whether or not glucose waspresent in the medium (Fig. 4B).
Postrest potentiation with andwithout ryanodine
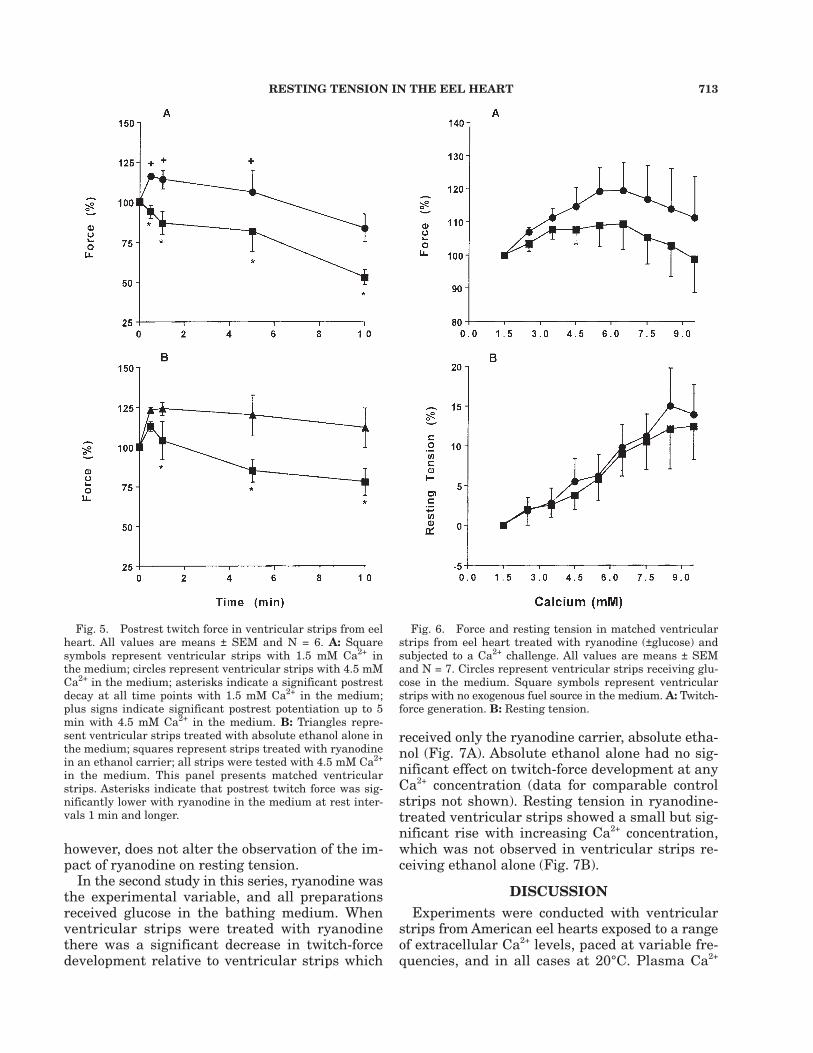
In these experiments, all ventricular strips re-ceived glucose. Postrest potentiation was assessedat extracellular Ca2+ levels of 1.5 and 4.5 mM.Figure 5A presents data from experiments con-ducted with different animals for the two Ca2+
levels. At 1.5 mM Ca2+, ventricular strips showedpostrest decay for all rest times, but when theCa2+ concentration in the bathing medium wasraised to 4.5 mM, ventricular strips showedpostrest potentiation at 0.5, 1.0, and 5.0 min ofrest, but not at 10 min of rest. In all cases,postrest potentiation was greater at Ca2+ concen-tration of 4.5 than 1.5 mM.
Postrest potentiation was assessed at an extra-cellular Ca2+ level of 4.5 mM and with ryanodinein the medium. Figure 5B presents data frompaired preparations. The ventricular strips treatedwith absolute ethanol, the ryanodine carrier,showed postrest potentiation. However, ryanodinetreatment turned the postrest potentiation intopostrest decay after 5 and 10 min of rest (Fig.5B). In all cases, postrest potentiation was greaterwithout than with ryanodine in the medium.
Ca2+ titration with ryanodineThe final experiments determined the effects of
ryanodine under a Ca2+ challenge. In the first
study, extracellular glucose was the experimentalvariable. Paired ventricular strips with and with-out glucose in the medium were treated withryanodine. There was no significant difference intwitch force between ventricular strips receivingglucose plus ryanodine and ventricular strips re-ceiving only ryanodine (Fig. 6A). There was a posi-tive correlation between the resting tension andincreasing Ca2+ concentration in ventricular stripstreated with ryanodine, with or without glucosein the medium (Fig. 6B). That is, in the presenceof ryanodine the protective effect of glucose onresting tension is removed. DMSO alone has a mi-nor negative impact on twitch-force developmentat high Ca2+ levels (data not shown). This factor,
Fig. 4. Force and resting tension in matched ventricularstrips from eel heart subjected to a frequency challenge. Allvalues are means ± SEM and N = 6. Square symbols repre-sent strips with glucose in the medium. Circles represent ven-tricular strips with no exogenous fuel the medium. A:Twitch-force generation. B: Resting tension.
RESTING TENSION IN THE EEL HEART 713
received only the ryanodine carrier, absolute etha-nol (Fig. 7A). Absolute ethanol alone had no sig-nificant effect on twitch-force development at anyCa2+ concentration (data for comparable controlstrips not shown). Resting tension in ryanodine-treated ventricular strips showed a small but sig-nificant rise with increasing Ca2+ concentration,which was not observed in ventricular strips re-ceiving ethanol alone (Fig. 7B).
DISCUSSIONExperiments were conducted with ventricular
strips from American eel hearts exposed to a rangeof extracellular Ca2+ levels, paced at variable fre-quencies, and in all cases at 20°C. Plasma Ca2+
Fig. 5. Postrest twitch force in ventricular strips from eelheart. All values are means ± SEM and N = 6. A: Squaresymbols represent ventricular strips with 1.5 mM Ca2+ inthe medium; circles represent ventricular strips with 4.5 mMCa2+ in the medium; asterisks indicate a significant postrestdecay at all time points with 1.5 mM Ca2+ in the medium;plus signs indicate significant postrest potentiation up to 5min with 4.5 mM Ca2+ in the medium. B: Triangles repre-sent ventricular strips treated with absolute ethanol alone inthe medium; squares represent strips treated with ryanodinein an ethanol carrier; all strips were tested with 4.5 mM Ca2+
in the medium. This panel presents matched ventricularstrips. Asterisks indicate that postrest twitch force was sig-nificantly lower with ryanodine in the medium at rest inter-vals 1 min and longer.
however, does not alter the observation of the im-pact of ryanodine on resting tension.
In the second study in this series, ryanodine wasthe experimental variable, and all preparationsreceived glucose in the bathing medium. Whenventricular strips were treated with ryanodinethere was a significant decrease in twitch-forcedevelopment relative to ventricular strips which
Fig. 6. Force and resting tension in matched ventricularstrips from eel heart treated with ryanodine (±glucose) andsubjected to a Ca2+ challenge. All values are means ± SEMand N = 7. Circles represent ventricular strips receiving glu-cose in the medium. Square symbols represent ventricularstrips with no exogenous fuel source in the medium. A: Twitch-force generation. B: Resting tension.

714 J.R. BAILEY ET AL.
concentration in the American eel is about 3.5 mMand increases to levels approaching 5 mM follow-ing air exposure (Hyde and Perry, ’87). Free Ca2+
levels will be approximately 60% of these valuesdue to binding to anions and proteins (Andreasen,’85). Given the absence of protein in the bathingmedium used in the current studies, and that bi-carbonate binds only about 10% of the Ca2+ inplasma, the total Ca2+ levels in the range of 1.5–4.5 mM would be in the physiological window ofCa2+ activity. Resting hearts rates for A. anguilla
have been reported as 25 bpm at 16°C (Peyraud-Waitzenegger et al., ’80) and 29 bpm at 12°C(Janvier, ’97). Adrenergic stimulation producedrates as high as 45 bpm (Peyraud-Waitzeneggeret al., ’80), and prostaglandin E2 reduced rates toas low as 13 bpm (Janvier, ’97). Resting heartrates in A. australis at 16–20°C is about 50 bpm,but can reach values as high as 65 bpm in someindividuals following pharmacological intervention(Hipkins, ’85). It is likely that the frequencies usedhere range from the low to high end of physiologi-cal rates for A. rostrata. Finally, 20°C is a typicalsummer temperature for A. rostrata living infreshwater.
A metabolic fuel is required for Ca2+
reduction under some conditionsIn the initial experiments, ventricular strips
were paced at 12, 30, or 48 bpm and subjected toincreases in extracellular Ca2+. At 12 and 30 bpm,increases in extracellular Ca2+ resulted in in-creases in twitch-force development, as has beenshown for ventricular strips from many fish spe-cies paced at 12 (e.g., Driedzic and Gesser, ’85;Bailey and Driedzic, ’90; Bailey et al., ’91) or 30bpm (Rantin et al., ’98). Twitch-force developmentis a function of intracellular Ca2+ activity and thetime that Ca2+ has to interact with the contrac-tile proteins (Yue, ’87). In the current studies, in-creased force development is considered, to a largeextent, to be due to an increase in Ca2+ influxacross the sarcolemma through L channels as afunction of excitation (Tibbits et al., ’91; Driedzicand Gesser, ’94). The increase in twitch force oc-curs with or without exogenous glucose, thusonboard fuels, presumably glycogen or fatty ac-ids, are able to supply the energy necessary tomaintain ATP levels. The heart of the Americaneel has an enzyme profile consistent with the mo-bilization of either of these fuels (Bailey et al.,’91; West et al., ’99). At 48 bpm however, increasesin extracellular Ca2+ level did not alter twitch-forcedevelopment with or without glucose in the me-dium. At this frequency it may be that the con-tractile fibrils are fully activated due to a highlevel of Ca2+ influx even at 1.5 mM extracellularCa2+. However, other explanations, such as the in-ability to supply ATP at an adequate sustainedrate, cannot be excluded.
Restoration of resting tension following a con-traction is dependent upon a decrease in Ca2+ ei-ther across the sarcolemmal membrane via theNa-Ca exchanger or uptake by the SR in somespecies (Tibbits et al., ’91). Resting tension was
Fig. 7. Force and resting tension in matched ventricularstrips from eel heart treated receiving glucose (±ryanodine)and subjected to a Ca2+ challenge. All values are means ±SEM, N = 8. Squares represent ventricular strips receivingglucose plus ethanol in the medium. Triangles represent ven-tricular strips receiving glucose plus ryanodine in an ethanolcarrier. A: Twitch-force generation; asterisks indicate thatforce development is significantly different between prepara-tions with and without ryanodine in the medium. B: Restingtension; asterisks indicate that resting tension is significantlydifferent between preparations with and without ryanodinein the medium.

RESTING TENSION IN THE EEL HEART 715
maintained at a pace rate of 12 bpm in the faceof increasing extracellular Ca2+, with or withoutglucose in the medium. That is, intracellular Ca2+
activity was fully decreased during the resting pe-riod, and onboard energy stores were mobilizedat an adequate rate to meet the ATP demand. At48 bpm, resting tension increased regardless ofthe presence or absence of glucose, implying thatthe capacity to decrease intracellular Ca2+ in theface of increasing Ca2+ influx was reached. Themost interesting condition was the intermediatelevel of Ca2+ trafficking and energy demand. At30 bpm, ventricular strips challenged with extra-cellular Ca2+ were able to maintain resting ten-sion, but only if glucose was present in themedium. Resting tension increased significantlyover the physiological range of Ca2+ concentration(i.e., 1.5–4.5 mM). This finding confirms the re-sult reported in the companion paper to this study(Bailey et al., ’99), with experiments conducted at24 bpm and 15°C.
Therefore, there is a window within the rangeof physiological Ca2+ concentration, frequency ofcontraction, and body temperature in which ex-tracellular glucose or at least an extracellularmetabolic fuel is required to maintain resting ten-sion in the American eel heart, most likely throughbeat-to-beat decreases in intracellular Ca2+ activ-ity. However, twitch force is not influenced by thepresence or absence of glucose. This finding is con-sistent with a study involving ventricular stripsfrom rat heart under anoxic conditions when glu-cose availability had no impact on the loss oftwitch force, but defended against an increase inresting tension (Anderson and Morris, ’78).
The next experiments assessed if there was aspecific requirement for glucose. Ventricular stripstreated with the glycolytic inhibitor iodoacetateand with or without a metabolic fuel in the me-dium showed an increase in resting tension anda decrease in twitch force development when chal-lenged with increases in extracellular Ca2+. Themagnitude of the increases in resting tension weresimilar to the decreases in twitch force; as such,peak force was maintained. Iodoacetate treatmentimpairs the metabolism of both extracellular glu-cose and stored glycogen since it acts at the levelof glyceraldehyde 3-phosphate dehydrogenase.Therefore, onboard fatty acids are not adequateto support increases in twitch-force development,a question which was unresolved by the first se-ries of experiments. When pyruvate was includedas a metabolic fuel, twitch force increased andresting tension was maintained in the face of in-
creases in extracellular Ca2+, even in the presenceof iodoacetate. That is, under these conditions,the myocytes were able to decrease intracellularCa2+ activity. Pyruvate is metabolized via the cit-ric acid cycle, which in turn leads to ATP produc-tion through oxidative phosphorylation. Theimportant point is that although there needs tobe an external energy source to support Ca2+ traf-ficking, glucose is not an obligatory requirementfor the process provided that aerobic metabolismis intact.
SR contributes to the maintenanceof resting tension
There is mounting evidence that at relativelyhigh temperatures the SR plays a role in beat-to-beat Ca2+ management of the rainbow trout andtuna heart (Hove-Madsen, ’92; Keen et al., ’92;Keen et al., ’94; Shiels and Farrell, ’97). Currentthinking based on the mammalian heart is thatat low temperature the SR Ca2+ release channelremains open in response to cytosolic Ca2+, thuseliminating the role of the organelle in Ca2+ man-agement. But at higher temperatures there is agreater probability that the channel will be closedand thus sensitive to opening via Ca2+ signalling(Tibbits, ’96).
Two means of obtaining an indirect assessmentof a functional SR are force frequency challengesand postrest potentiation studies. Ventricular strippreparations from fish hearts in which the SR isconsidered to be relatively unimportant show asharp decrease in twitch force as frequency is in-creased above 12 bpm. Preparations in which theSR is important for beat-to-beat Ca2+ managementusually display a positive followed by a negativestaircase (Driedzic and Gesser, ’85, ’88; Keen etal., ’92). It should be noted that rainbow trout maybe an exception to this general paradigm (Hove-Madsen, ’92; Shiels and Farrell, ’97). Ventricularstrips from the American eel heart maintainedtwitch-force development over the range of 12–36bpm, after which twitch force decreased in asso-ciation with a comparable increase in resting ten-sion. An increase in twitch force following a periodof noncontraction is considered to be indicative ofthe filling of Ca2+ stores during the rest period.Ventricular strips exhibited only postrest decayat low Ca2+ levels. However, when extracellularCa2+ was increased to 4.5 mM, the first beats fol-lowing a rest period were elevated. Further stud-ies utilized ryanodine to impair the SR. Ryanodineat the level used here should inhibit Ca2+ release(Coronado et al., ’94). As the SR cannot be a lim-

716 J.R. BAILEY ET AL.
itless sink for Ca2+, uptake should then be com-promised as well. Ryanodine treatment reducedor eliminated postrest potentiation at high levelsof extracellular Ca2+, a result similar to that ob-tained with rainbow trout (El-Sayed and Gesser,’89; Hove-Madsen, ’92). These experiments pro-vide indirect evidence that is consistent with afunctional SR in the beat-to-beat regulation of Ca2+
trafficking in the American eel heart at 20°C.In the final experiments, the impact of ryano-
dine on the response of the ventricular strips toincreases in extracellular Ca2+ was assessed. Asextracellular Ca2+ was increased, the blockage ofsarcoplasmic reticular function invariably resultedin an increase in resting tension. The protectiveeffect of glucose on the maintenance of resting ten-sion was eliminated. The impact of ryanodine ontwitch tension is less clear, as even control prepa-rations showed a decline in twitch force at highCa2+ levels in some cases, perhaps due to thelength of these studies. Also, decreases in twitchforce were usually associated with increases inresting tension, thus resulting in the maintenanceof peak force development.
CONCLUSIONSThe experiments presented here were restricted
to hearts from American eels. This species wasselected as a model system to capitalize on a sub-stantive background literature. Investigations arecurrently under way to assess if the findings areapplicable to other fish species. In the heart ofthe American eel there is a window within thephysiological range of contraction frequency andextracellular Ca2+ load where: (1) either an extra-cellular fuel source or onboard glycogen is requiredto support twitch force (i.e., onboard fatty acidsare not sufficient), and (2) an extracellular fuel isrequired to maintain resting tension (i.e., onboardglycogen is not sufficient). It appears that the en-ergy requirements associated with the processesinvolved in the maintenance of resting tension arecompartmentalized and dependent on externalfuel delivery. Metabolic compartmentation occursin other tissues. In smooth muscle, for instance,ATP requirement for the contractile fibrils is sup-ported by oxidative metabolism, and a separateenzyme cascade utilizing glycogen but not glucoseplays a role in this compartment (Hardin andPaul, ’95). In rat heart, glycolysis from glycogenand rates of oxidation of glycogen are differen-tially controlled, again implying metabolic com-partmentation (Allard, ’97). At 20°C, the Americaneel heart has a functional SR that plays a role in
the maintenance of resting tension, presumablyby decreasing cytoplasmic Ca2+ on a beat-to-beatbasis. ATP equivalents from onboard glycogen maynot be available to the Ca2+-ATPase of the SR. TheAmerican eel heart differs from the rat heart(Nakamura et al., ’93; Xu et al., ’95; Assum et al.,’98) in that there does not appear to be an obliga-tory requirement for extracellular glucose for thecontrol of intracellular Ca2+, as evidenced by con-tractile parameters, nor do our findings supportthe contention that Ca2+ uptake by the SR is gly-colytic dependent (Xu et al., ’95).
LITERATURE CITEDAasum E, Lathrop DA, Henden T, Sundset R, Larsen TS.
1998. The role of glycolysis in myocardial calcium control.J Mol Cell Cardiol 30:1703–1712.
Allard MF. 1997. Energy metabolism of the hypertrophiedheart. J Mol Cell Cardiol 29:A262.
Anderson GL, Morris RO. 1978. Role of glycolysis in the re-laxation process in mammalian cardiac muscle: compari-son of the influence of glucose and 2-deoxyglucose onmaintenance of resting tension. Life Sci 23:23–32.
Andreasen P. 1985. Free and total calcium concentrations inthe blood of the rainbow trout, Salmo gairdneri, duringstress conditions. J Exp Biol 118:111–120.
Bailey JR, Driedzic WR. 1990. Enhanced maximum frequencyand force development of fish hearts following temperatureacclimation. J Exp Biol 149:239–254.
Bailey JR, Sephton D, Driedzic WR. 1991. Impact of an acutetemperature change on performance and metabolism of pick-erel (Esox niger) and eel (Anguilla rostrata) hearts. PhysiolZool 64:697–716.
Bailey JR, Rodnick KJ, MacDougall R, Clowe S, Driedzic WR.2000. Anoxic performance of the American eel (Anguillarostrata L.) heart requires extracellular glucose. J Exp Zool286:699–706.
Coronado R, Morrisette J, Sukhareva M, Vaughan DM. 1994.Structure and function of ryanodine receptors. Am J Physiol266(Cell Physiol 35):C1485–C1504.
Driedzic WR. 1992. Cardiac energy metabolism. In: Hoar WS,Randall DJ, Farrell AP, editors. Fish physiology. Vol 12. NewYork: Academic Press. p 219–266.
Driedzic WR, Gesser H. 1985. Ca2+ protection from the nega-tive inotropic effect of contraction frequency on teleosthearts. J Comp Physiol B 156:135–142.
Driedzic WR, Gesser H. 1988. Differences in force-frequencyrelationships and calcium dependency between elasmo-branch and teleost hearts. J Exp Biol 140:227–241.
Driedzic WR, Gesser H. 1994. Energy metabolism and con-tractility in ectothermic vertebrate hearts: hypoxia, acido-sis, and low temperature. Physiol Rev 74:221–258.
Driedzic WR, Gesser H, Johansen K. 1985. Effects of hypoxicadaptation on myocardial performance and metabolism ofZoarces viviparous. Can J Zool 63:821–823.
El-Sayed MF, Gesser H. 1989. Sarcoplasmic reticulum, po-tassium, and cardiac force in rainbow trout and plaice. AmJ Physiol 257(Reg Integ Comp Physiol 26):R599–R604.
Hardin CD, Paul RJ. 1995. Metabolism and energetics of vas-cular smooth muscle. In: Sperelakis N, editor. Physiologyand pathophysiology of the heart. 3rd ed. New York: KluwerAcademic Publishers. p 1069–1086.

RESTING TENSION IN THE EEL HEART 717
Hipkins SF. 1985. Adrenergic responses of the cardiovascu-lar system of the eel, Anguilla australis. In vivo. J ExpZool 235:7–20.
Hoar WS, Hickman CP Jr. 1967. A laboratory companion forgeneral and comparative physiology. Englewood Cliffs, NJ:Prentice-Hall.
Hove-Madsen L. 1992. The influence of temperature onryanodine sensitivity and the force-frequency relationshipin the myocardium of rainbow trout. J Exp Biol 167:47–60.
Hyde DA, Perry SF. 1987. Acid-base and ionic regulation inthe American eel (Anguilla rostrata) during and after pro-longed aerial exposure: branchial and renal adjustments. JExp Biol 133:429–447.
Janvier J-J. 1997. Cardiovascular and ventilatory effects ofprostaglandin E2 in the European eel Anguilla anguilla. JComp Physiol B 167:517–526.
Keen JE, Farrell AP, Tibbits GF, Brill RW. 1992. Cardiacphysiology in tunas: II. Effect of ryanodine, calcium, andadrenaline on force frequency relationships in atrial stripsfrom skipjack tuna, Katsuwonus pelamis. Can J Zool70:1211–1217.
Keen JE, Vianzon D-M, Farrell AP, Tibbits GF. 1994. Effectof temperature and temperature acclimation on theryanodine sensitivity of the trout myocardium. J CompPhysiol B 164:438–443.
Nakamura K, Kusuoka H, Ambrosio G, Becker LC. 1993. Gly-colysis is necessary to preserve myocardial Ca2+ homeosta-sis during β-adrenergic stimulation. Am J Physiol 264(HeartCirc Physiol 33):H670–H678.
Nolte LA, Rincon J, Odegaard Wahlstrom E, Craig BW,Zierath JR, Wallberg-Henriksson H. 1995. Hyperglycemiaactivates glucose transport in rat skeletal muscle via a Ca2+-dependent mechanism. Diabetes 44:1345–1348.
Peyraud-Waitzenegger M, Barthelemy L, Peyraud C. 1980.Cardiovascular and ventilatory effects of catecholamines inunrestrained eels (Anguilla anguilla L.). J Comp Physiol B138:367–375.
Rantin FT, Gesser H, Kalinin AL, Guerra CDR, De FreitasJC, Driedzic WR. 1998. Heart performance, Ca2+ regula-tion and energy metabolism at high temperatures inBathygobius soporator, a tropical marine teleost. J ThermBiol 23:31–39.
Shiels HA, Farrell AP. 1997. The effect of temperature andadrenaline on the relative importance of the sarcoplasmicreticulum in contributing Ca2+ to force development in iso-lated ventricle trabeculae from rainbow trout. J Exp Zool200:1607–1621.
Tibbits GF. 1996. Towards a molecular explanation of the highperformance of the tuna heart. Comp Biochem Physiol113A:77–82.
Tibbits GF, Hove-Madsen L, Bers DM. 1991. Ca2+ trans-port and the regulation of cardiac contractility in teleo-sts: a comparison with higher vertebrates. Can J Zool69:2014–2019.
West JL, Bailey JR, Almeida-Val VMF, Val AL, Sidell BD,Driedzic WR. 1999. Activity levels of enzymes of energymetabolism in heart and red muscle are higher in northtemperate than in Amazonian teleosts. Can J Zool 77:690–696.
Xu KY, Zweier JL, Becker LC. 1995. Functional coupling be-tween glycolysis and sarcoplasmic reticulum Ca2+ transport.Circ Res 77:88–97.
Yue DT. 1987. Intracellular [Ca2+] related to rate of force de-velopment in twitch contraction of heart. Am J Physiol252(Heart Circ Physiol 21):H760–H770.