major histocompatibility complex class ii matching is necessary … · 2002-10-09 · laboratory...
TRANSCRIPT

The Role of Autoimmunity in Islet Allograft DestructionMajor Histocompatibility Complex Class II Matching IsNecessary for Autoimmune Destruction of Allogeneic IsletTransplants After T-Cell Costimulatory BlockadeLeila Makhlouf,
1Koji Kishimoto,
1Rex N. Smith,
2Reza Abdi,
1Maria Koulmanda,
3Henry J. Winn,
3
Hugh Auchincloss, Jr.,3
and Mohamed H. Sayegh1,4
Although it has often been assumed that transplanted
allogeneic islets can be destroyed by recurrent auto-
immunity in recipients with type 1 diabetes, definitive
evidence is lacking and the settings in which this may
occur have not been defined. To address these issues,
we compared the survival of islet transplants (subject
to tissue-specific autoimmunity) with cardiac trans-
plants (not subject to tissue-specific autoimmunity)
from various major histocompatibility complex (MHC)-
matched and -mismatched donors transplanted into
autoimmune NOD recipients. We found that when recip-
ients were treated with combined B7 and CD154 T-cell
costimulatory blockade, hearts survived best with bet-
ter MHC matching, whereas islets survived worst when
the donor and recipient shared MHC class II antigens. In
the absence of full or MHC class II matching, there was
no difference in the survival of islet and cardiac allo-
grafts. We also found that the tendency of NOD mice to
resist tolerance induction by costimulation blockade is
mediated by both CD4� and CD8� T-cells, not directly
linked to the presence of autoimmunity, and conferred
by non-MHC background genes. These findings have
clinical importance because they suggest that under
some circumstances, avoiding MHC class II sharing may
provide better islet allograft survival in recipients
with autoimmune diabetes, since mismatched allogeneic
islets may be resistant to recurrent autoimmunity. Our
results may have implications for the design of future
clinical trials in islet transplantation. Diabetes 51:
3202–3210, 2002
Islet transplants for patients with type 1 diabetespotentially face two distinct types of immune de-struction: one generated by the allogeneic responseto foreign tissues and the other generated by the
recurrence of the tissue-specific autoimmune process thatcaused the disease in the first place. Indeed, previousreports showed that human islets from genetically identi-cal twins (1) or cadaver donors (2) were subject torecurrent autoimmunity. Several other findings have sug-gested that recurrent autoimmunity might be responsiblefor destruction of xenogeneic (3) and allogeneic (3–6)islets. In interesting studies by Woehrle et al. (7) andMarkmann et al. (8) in BB rats, precultured major histo-compatibility complex (MHC)-mismatched islets were notsubject to recurrent autoimmunity, and the survival wassignificantly better for MHC-mismatched grafts than forMHC-matched grafts. Furthermore, until recently, humanislet transplants have generally been notably unsuccessfulcompared with other types of transplants in patients withtype 1 diabetes (9). In addition, several investigators havefound that strategies that induce long-term survival ofallogeneic islets in ordinary mice have been less success-ful when used in NOD mice that have developed sponta-neous autoimmune diabetes (5,10,11). In these cases, theassumption has been that the existence of the autoimmuneprocess in NOD mice was responsible for the failure of thetolerance-induction strategies.
On the other hand, other evidence has not supported theidea that recurrent autoimmunity plays a role in thedestruction of islets transplanted from genetically dispar-ate donors (12). First, although the results of clinical islettransplantation have often been poor, survival rates forwhole-organ pancreas transplants are similar to those ofkidneys (13), and even islet transplantation has been verysuccessful recently when performed with newer immuno-suppressive protocols (14). Second, it has been shown thatxenogeneic islets survive for longer periods than synge-neic islets in diabetic NOD mice when they have beentreated with a variety of immunomodulating therapies(15). Finally, Markees et al. (16) recently reported that theresistance of NOD mice to tolerance induction applies totissues other than islets, suggesting that this resistancemay not necessarily reflect recurrent tissue-specific auto-
From the 1Laboratory of Immunogenetics and Transplantation, Brigham andWomen’s Hospital, Harvard Medical School, Boston, Massachusetts; the2Department of Pathology, Massachusetts General Hospital, Harvard MedicalSchool, Boston, Massachusetts; the 3Department of Surgery, MassachusettsGeneral Hospital, Harvard Medical School, Boston, Massachusetts; and the4Nephrology Division, Children’s Hospital, Harvard Medical School, Boston,Massachusetts.
Address correspondence and reprint requests to Mohamed H. Sayegh, MD,Laboratory Immunogenetics and Transplantation, Renal Division, Departmentof Medicine, Brigham and Women’s Hospital, Boston, MA 02115. E-mail:[email protected].
Received for publication 21 March 2002 and accepted in revised form 8August 2002.
MHC, major histocompatibility complex; MST, mean survival time.
3202 DIABETES, VOL. 51, NOVEMBER 2002
immunity. Thus, it is not clear at this point what rolerecurrent autoimmunity plays in islet transplant destruc-tion and, if it does, under what circumstances. This is ofobvious clinical relevance to islet transplantation in hu-mans with type 1 diabetes who are potentially subject torecurrent autoimmunity in the transplanted islets.
To address these questions, we conducted a series oftransplant experiments in NOD mice, one of the bestavailable animal models for the study of type 1 diabetes(17), using two types of tissues: hearts, which would onlybe subject to alloreactive destruction, and islets, whichwould potentially be subject to recurrent autoimmunity inaddition to alloreactivity. We also used donors that ex-pressed a range of genetic disparities to determine whatdegree of antigen matching might allow for the expressionof recurrent autoimmunity. Our results indicate that whenB7 and CD154 T-cell costimulatory blockade is used toprolong islet transplant survival, recurrent autoimmunityis only evident when the MHC class II antigens arematched between the donor and recipient. Our data alsoshow that the resistance of NOD mice to tolerance bycostimulation blockade is mediated by both CD4� andCD8� T-cells, not absolutely linked to the presence of anautoimmune response, and conferred by non-MHC back-ground genes.
RESEARCH DESIGN AND METHODS
Mice. Diabetic female NOD/Lt (H-2 g7) mice (referred to as NOD) were usedas recipients. BALB/c (H-2 days), C57BL/6 (H-2b), NOD/LtSz-Prkdcscid/Prkdcscid (referred to as NOD-scid), B10.BR (H-2k), and B6.NOD (H-2 g7)mice, aged 8–12 weeks, were obtained from The Jackson Laboratory (BarHarbor, ME) and used as donors. BIOZZI ABH/RijHsdH (IA-g7) mice, referredto as BIOZZI mice, were purchased from Harlan (England, U.K.), and B10.HTG(H-2 g) were a kind gift of Dr. Hugh McDevitt (Stanford University).Murine model for islet transplantation. Once NOD mice spontaneouslydeveloped diabetes, 700 islets were transplanted under the renal capsule. Innonautoimmune recipients, diabetes was chemically induced with streptozo-tocin (225 mg/kg i.p.) and 400 islets were transplanted under the renal capsule.
Diabetes was defined as blood glucose �250 mg/dl on at least 2 consecu-tive days. Reversal of diabetes was defined as blood glucose �200 mg/dl for atleast 2 consecutive days. Graft rejection was defined as blood glucose �250mg/dl for at least 2 consecutive days and confirmed histologically. Islet graftfunction was assessed by blood glucose measurements (ACCU-Check advan-tage; Boehringer Mannheim) twice a week.
All experiments were performed in compliance with institutional guide-lines regarding animal care.Isolation of pancreatic islets. Pancreatic islets were isolated using colla-genase digestion followed by histopaque 1077 (Sigma 1077; Sigma, St. Louis,MO) density gradient separation and then handpicking as described elsewhere(18).Cardiac transplants. Cardiac transplants were performed in the same mousestrain combinations as islets. Cardiac graft survival was assessed by graftpalpation (19). Rejection was confirmed histologically.Histology. Grafts (three to six per group) were harvested soon after rejectionand fixed in 2% buffered formalin. Paraffin-embedded sections (4 �m) werestained with hematoxylin and eosin. Immunohistochemical staining wasperformed on both paraffin-embedded tissue sections using guinea pig anti-insulin (1/10 dilution; Dako) or rabbit anti-glucagon (1/10 dilution; Dako).Second antibodies were goat anti–guinea pig or goat anti-rabbit at 1/200(Vector Laboratories). Slides were blocked using an avidin-biotin blocking kit(Vector Laboratories, Burlingame, CA) and normal rabbit sera at 1/10 beforestaining with primary antibodies. After staining, slides were developed withavidin-conjugated horseradish peroxidase using the vectastain ABC Kit (Vec-tor Laboratories). Biotinylated isotype-matched antibodies were used asnegative controls for all staining experiments. Photographs were taken with aRT Color Spot camera (Diagnostic Instruments) and a Zeiss Axioskopmicroscope. All magnifications are 400�. Composite pictures were createdwith Adobe PhotoShop.Agents, antibodies, and in vivo T-cell depletion. MR1 is a hamstermonoclonal antibody that is specific for murine CD154 (the hybridoma is akind gift from Dr. Randy Noelle (Dartmouth College of Medicine, Lebanon,NH), and the antibody was manufactured commercially by Bioexpress, West
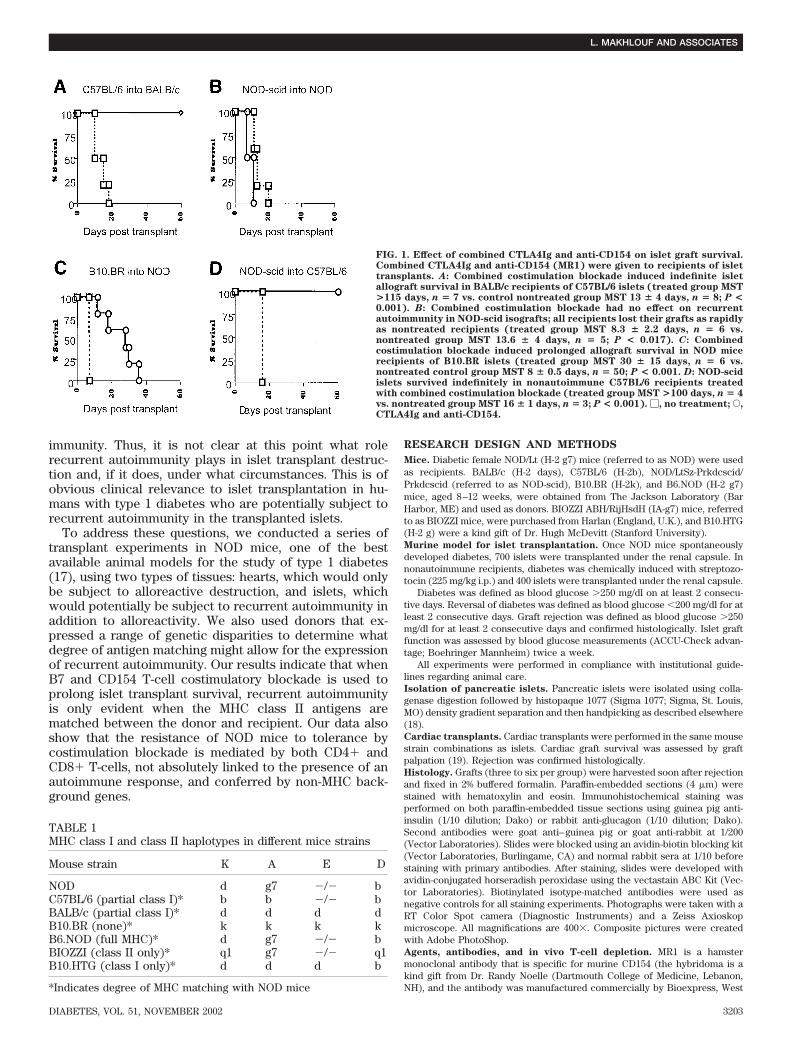
FIG. 1. Effect of combined CTLA4Ig and anti-CD154 on islet graft survival.Combined CTLA4Ig and anti-CD154 (MR1) were given to recipients of islettransplants. A: Combined costimulation blockade induced indefinite isletallograft survival in BALB/c recipients of C57BL/6 islets (treated group MST>115 days, n � 7 vs. control nontreated group MST 13 � 4 days, n � 8; P <0.001). B: Combined costimulation blockade had no effect on recurrentautoimmunity in NOD-scid isografts; all recipients lost their grafts as rapidlyas nontreated recipients (treated group MST 8.3 � 2.2 days, n � 6 vs.nontreated group MST 13.6 � 4 days, n � 5; P < 0.017). C: Combinedcostimulation blockade induced prolonged allograft survival in NOD micerecipients of B10.BR islets (treated group MST 30 � 15 days, n � 6 vs.nontreated control group MST 8 � 0.5 days, n � 50; P < 0.001. D: NOD-scidislets survived indefinitely in nonautoimmune C57BL/6 recipients treatedwith combined costimulation blockade (treated group MST >100 days, n � 4vs. nontreated group MST 16 � 1 days, n � 3; P < 0.001). �, no treatment; E,CTLA4Ig and anti-CD154.
TABLE 1MHC class I and class II haplotypes in different mice strains
Mouse strain K A E D
NOD d g7 �/� bC57BL/6 (partial class I)* b b �/� bBALB/c (partial class I)* d d d dB10.BR (none)* k k k kB6.NOD (full MHC)* d g7 �/� bBIOZZI (class II only)* q1 g7 �/� q1B10.HTG (class I only)* d d d b
*Indicates degree of MHC matching with NOD mice
L. MAKHLOUF AND ASSOCIATES
DIABETES, VOL. 51, NOVEMBER 2002 3203
Lebanon, NH). CTLA4Ig is a kind gift of Dr. Robert Peach (Bristol-MeyersSquibb, Princeton, NJ). The agents were administered intraperitoneally (250�g) at days 0, 2, 4, and 6 posttransplantation, as previously reported (20).Anti-CD8 ascites (116-13.1; anti-Lyt2.1 IgG2a) and anti-CD4 ascites (GK1.5;anti-L3T4 rat IgG2b) were obtained from the American Type Culture Collec-tion (Rockville, MD). Anti-CD8– and anti-CD4–treated mice received 0.2 mli.p. of unpurified ascites of the appropriate antibody (roughly equivalent to 200mg of purified antibody) on day �1 and day 0 and then twice a week aftertransplantation until rejection or day 60 posttransplantation in animals withsurviving grafts (21). This resulted in �98% depletion of CD4� or CD8�T-cells throughout the follow-up period.Statistics. Survival data were plotted by using the Kaplan-Meier method.Significance of difference between groups was tested by comparing groupmeans and medians (mean survival time [MST]) by either the two-tailed t test
or the Wilcoxon’s signed-rank test, as appropriate. Statistical significance wasdefined as a P value �0.05.
RESULTS
The effect of costimulatory blockade on islet trans-
plant survival in normal and NOD mice. We performedour studies using a regimen of T-cell costimulatory block-ade that has been found to be very effective in promotinglong-term allograft survival in several transplant models,including the stringent skin allograft model (20). Com-bined treatment with CTLA4Ig and anti-CD154 mAbcaused long-term islet allograft survival when C57BL/6
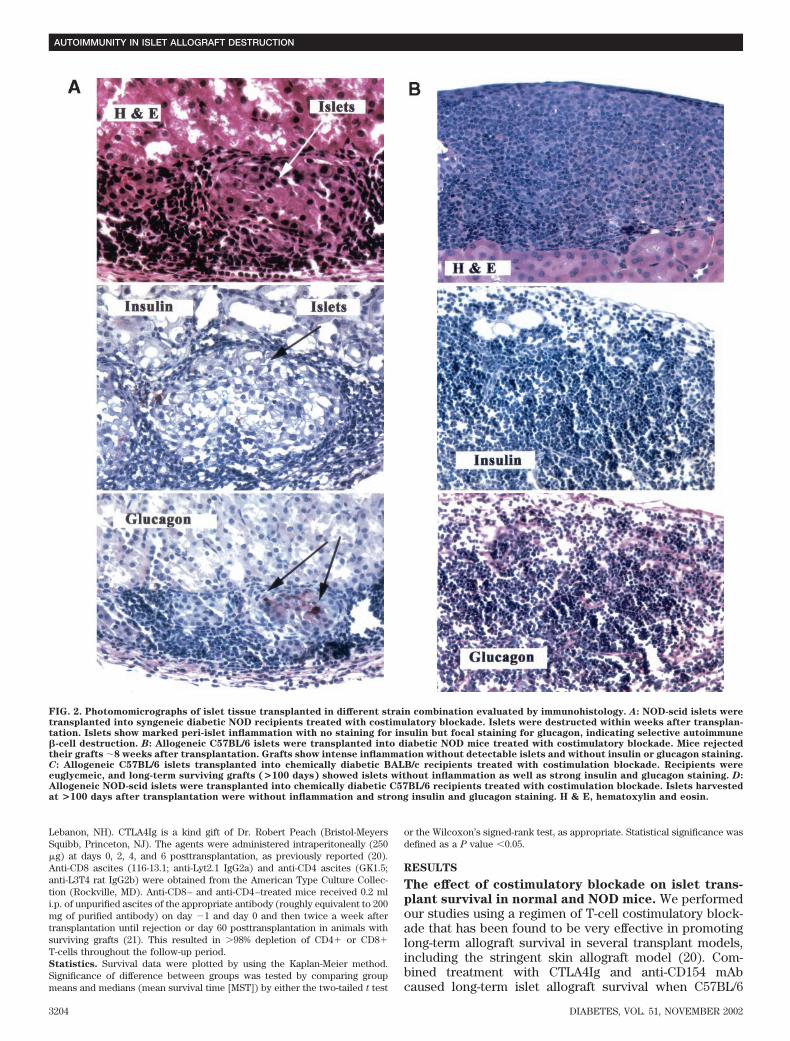
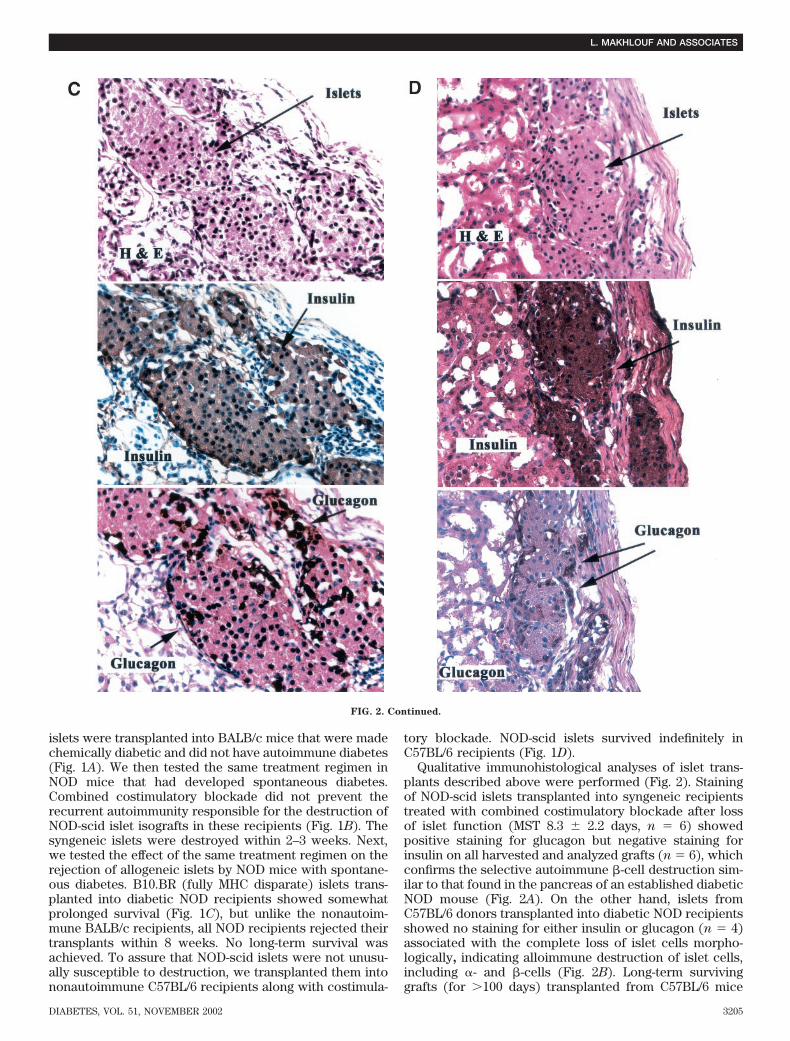
FIG. 2. Photomomicrographs of islet tissue transplanted in different strain combination evaluated by immunohistology. A: NOD-scid islets weretransplanted into syngeneic diabetic NOD recipients treated with costimulatory blockade. Islets were destructed within weeks after transplan-tation. Islets show marked peri-islet inflammation with no staining for insulin but focal staining for glucagon, indicating selective autoimmune�-cell destruction. B: Allogeneic C57BL/6 islets were transplanted into diabetic NOD mice treated with costimulatory blockade. Mice rejectedtheir grafts �8 weeks after transplantation. Grafts show intense inflammation without detectable islets and without insulin or glucagon staining.C: Allogeneic C57BL/6 islets transplanted into chemically diabetic BALB/c recipients treated with costimulation blockade. Recipients wereeuglycmeic, and long-term surviving grafts (>100 days) showed islets without inflammation as well as strong insulin and glucagon staining. D:Allogeneic NOD-scid islets were transplanted into chemically diabetic C57BL/6 recipients treated with costimulation blockade. Islets harvestedat >100 days after transplantation were without inflammation and strong insulin and glucagon staining. H & E, hematoxylin and eosin.
AUTOIMMUNITY IN ISLET ALLOGRAFT DESTRUCTION
3204 DIABETES, VOL. 51, NOVEMBER 2002
islets were transplanted into BALB/c mice that were madechemically diabetic and did not have autoimmune diabetes(Fig. 1A). We then tested the same treatment regimen inNOD mice that had developed spontaneous diabetes.Combined costimulatory blockade did not prevent therecurrent autoimmunity responsible for the destruction ofNOD-scid islet isografts in these recipients (Fig. 1B). Thesyngeneic islets were destroyed within 2–3 weeks. Next,we tested the effect of the same treatment regimen on therejection of allogeneic islets by NOD mice with spontane-ous diabetes. B10.BR (fully MHC disparate) islets trans-planted into diabetic NOD recipients showed somewhatprolonged survival (Fig. 1C), but unlike the nonautoim-mune BALB/c recipients, all NOD recipients rejected theirtransplants within 8 weeks. No long-term survival wasachieved. To assure that NOD-scid islets were not unusu-ally susceptible to destruction, we transplanted them intononautoimmune C57BL/6 recipients along with costimula-
tory blockade. NOD-scid islets survived indefinitely inC57BL/6 recipients (Fig. 1D).
Qualitative immunohistological analyses of islet trans-plants described above were performed (Fig. 2). Stainingof NOD-scid islets transplanted into syngeneic recipientstreated with combined costimulatory blockade after lossof islet function (MST 8.3 � 2.2 days, n 6) showedpositive staining for glucagon but negative staining forinsulin on all harvested and analyzed grafts (n 6), whichconfirms the selective autoimmune -cell destruction sim-ilar to that found in the pancreas of an established diabeticNOD mouse (Fig. 2A). On the other hand, islets fromC57BL/6 donors transplanted into diabetic NOD recipientsshowed no staining for either insulin or glucagon (n 4)associated with the complete loss of islet cells morpho-logically, indicating alloimmune destruction of islet cells,including �- and -cells (Fig. 2B). Long-term survivinggrafts (for �100 days) transplanted from C57BL/6 mice
FIG. 2. Continued.
L. MAKHLOUF AND ASSOCIATES
DIABETES, VOL. 51, NOVEMBER 2002 3205
AUTOIMMUNITY IN ISLET ALLOGRAFT DESTRUCTION
3206 DIABETES, VOL. 51, NOVEMBER 2002

into BALB/c recipients showed positive staining for bothglucagon and insulin (n 3) (Fig. 2C). Also, NOD-scidislets transplanted into C57BL/6 recipients treated withcostimulation blockade survived indefinitely, as well asstained positive for both glucagon and insulin (n 3)(Fig. 2D). Absence of cellular infiltrate was observedin all long-term surviving islets. The pattern of insulinand glucagon staining found in the islets in Fig. 2C andD is similar to that found in the islets of a normalpancreas.
These results confirm the general findings reported byothers: that immunomodulatory therapies that achievelong-term survival for allogeneic transplants are not nec-essarily effective in preventing autoimmune destruction ofsyngeneic islets in NOD mice, and that these therapies aregenerally less effective for allogeneic islet transplants inrecipients with spontaneous diabetes. However, it was notclear from these data whether the destruction of theallogeneic islets in the autoimmune recipients was due toa recurrence of autoimmunity.Is tissue-specific autoimmunity responsible for the
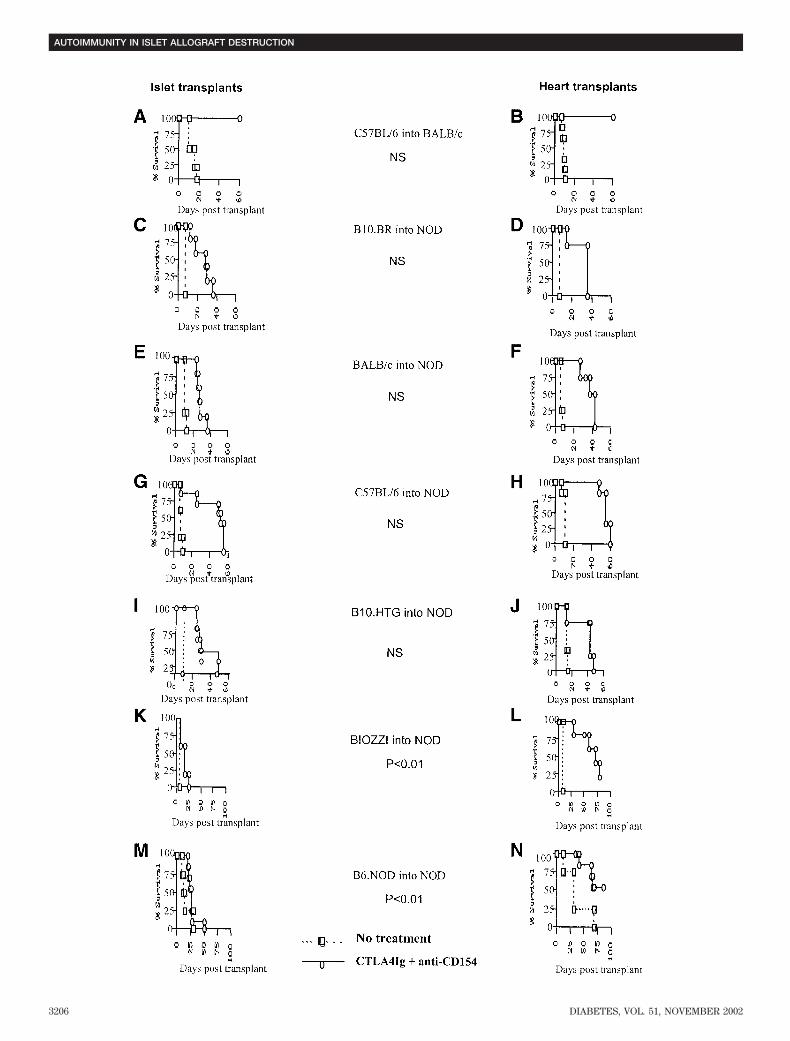
destruction of islet allografts?. To examine the role ofautoimmunity in the destruction of islets by NOD mice, wetransplanted either islet or cardiac allografts into NODmice that were treated with combined T-cell costimulatoryblockade as above. First, unlike islets (Fig. 1B), NOD-scidcardiac allografts were not rejected by NOD mice (graftsurvival �150 days, n 4). Therefore, we concluded thatcardiac grafts were not subject to tissue-specific autoim-munity. We then performed islet or cardiac transplantsusing a number of donor and recipient combinations thatwere designed to match some, or all, of the transplantationantigens of the donor and recipient (Table 1), assumingthat different mouse strain islets have similar inherentsensitivity to immune injury in the NOD mouse recipient.Figure 3 shows the results of these experiments. Thesurvival of cardiac allografts was nearly identical to thesurvival of allogeneic islets when MHC antigen sharingwas not present (Fig. 3C and D). It was also not statisti-cally significantly different when there was either partial orcomplete MHC class I antigen matching (compare Fig. 3E
with J). On the other hand, islet allografts were rejectedsignificantly quicker than heart grafts by NOD recipientswhen there was either complete or MHC class II antigenmatching. In these cases, there was significant prolonga-tion of, and sometimes indefinite, cardiac allograft survival
(Fig. 3L and N) but early destruction of islet allografts(Fig. 3K and M). The results of these experiments suggestthat matching for MHC class II antigens allows recurrentautoimmunity to play a role in islet allograft destruction.They provide no evidence to suggest that class I– orcompletely MHC–mismatched allografts are subject toautoimmune destruction.
It is interesting to note that despite the apparent role ofautoimmunity in the destruction of the MHC class II–matched islet allografts, the results of the immunohisto-logical analysis of these transplants showed that glucagon-staining cells, as well as insulin-staining cells, weredestroyed when class II–matched allografts were rejected(not shown). Thus, it appeared that while an autoimmuneresponse might be responsible for initiating and/or con-tributing to the early destruction of class II–matchedallografts, the final effector mechanisms also targeted theallogeneic cells. This is not surprising, since the majorityof cardiac allografts that are not subject to autoimmunedestruction were also ultimately rejected, albeit at asignificantly delayed time as compared with islets.Why are NOD mice resistant to tolerance induction?
The difficulty in achieving long-term survival of allogeneictransplants in NOD mice has been observed by others (16).We further examined the mechanisms of this resistance totolerance in several ways. First, we depleted either CD4�or CD8� T-cells vigorously in NOD recipients and thentransplanted either allogeneic or syngeneic islets. ChronicCD4� or CD8� T-cell depletion led to long-term survivalof the syngeneic islets (subject to autoimmune destructiononly), but allogeneic islets were still rejected within 3months (Fig. 4A and B). These data provide furthersupport to the notion that the difficulty in achievinglong-term allograft survival in NOD mice does not dependon the presence of autoimmunity.
CD8� T-cells have been shown to be resistant tocombined costimulation blockade in some stringent trans-plant models (22,23). However, the addition of CD8�T-cell depletion to our regimen of costimulatory blockadedid not further prolong the survival of allogeneic isletscompared with combined costimulatory blockade alone(Fig. 4B). Thus, the resistance of NOD mice to long-termallograft survival is not due to costimulation blockade–resistant alloreactive CD8� T-cells.
Finally, we used the same regimen of costimulatoryblockade in recipient B6.NOD (NOD MHC genes on the
FIG. 3. Survival of cardiac and islet allografts in NOD recipients. A: C57BL/6 islets transplanted into BALB/c recipients treated with combinedcostimulation blockade survived indefinitely (MST >60 days, n � 7; nontreated group MST 13 � 4 days; P < 0.005). B: C57BL/6 heartstransplanted into BALB/c recipients treated with combined costimulation blockade survived indefinitely (MST >60 days, n � 8; nontreated groupMST 8.6 � 1 days; P < 0.005). C: B10.BR islets transplanted into NOD recipients treated with combined costimulation blockade rejected at MST30 � 15 days, n � 6. MST of nontreated group: 8 � 0.5 days, n � 3; P < 0.005. D: B10.BR hearts transplanted into NOD mice treated with combinedcostimulation blockade rejected at MST 30 � 11 days, n � 4. Survival of B10.BR hearts versus islets: P � 0.9. MST of nontreated group: 7 � 0 days,n � 4; P < 0.005. E: BALB/c islets transplanted into NOD mice treated with costimulation blockade rejected at MST 26.7 � 10 days, n � 6. MSTof nontreated group: 9.8 � 1.5 days, n � 3; P < 0.005. F: BALB/c hearts transplanted into NOD recipients treated with combined costimulationblockade rejected at MST 39 � 9 days, n � 4. Survival of BALB/c hearts versus islets: P � 0.09. MST of nontreated group: 6.8 � 0.5 days, n � 3;P < 0.001. G: C57BL/6 islets transplanted into NOD mice treated with combined costimulation blockade rejected at MST 49 � 12 days, n � 6. MSTof nontreated group: 7 � 1 days, P < 0.005. H: C57BL/6 hearts transplanted into NOD mice treated with combined costimulation blockade rejectedat MST 56 � 4 days, n � 6. Survival of C57BL/6 hearts versus islets: P � 0.18. MST of nontreated group: 7 � 1 days, n � 5; P < 0.005. I: B10.HTGislets transplanted into NOD mice treated with combined costimulation blockade rejected at MST 32. �11.75 days, n � 6. MST of nontreatedgroup: 11 � 3 days, n � 4; P < 0.005. J: B10.HTG hearts transplanted into NOD mice treated with combined costimulation blockade rejected atMST 36.5 � 15.8 days, n � 4. Survival of B10.HTG hearts versus islets: P � 0.62. MST of nontreated group: 14.75 � 2.2 days, n � 4; P < 0.005.K: BIOZZI islets transplanted into NOD mice treated with combined costimulation blockade rejected at MST 19.2 � 4 days, n � 5. MST ofnontreated group: 7.5 � 0.5 days, n � 3; P < 0.005. L: BIOZZI hearts transplanted into NOD mice treated with combined costimulation blockaderejected at MST 58.5 � 20 days, n � 4. MST of nontreated group: 9 � 2 days, n � 2; P � 0.002. Survival of BIOZZI hearts versus islets: P � 0.004.M: B6.NOD islets transplanted into NOD mice treated with combined costimulation blockade rejected at MST 27.7 � 10.3 days, n � 7. MST ofnontreated group: 16.25 � 9.4 days, n � 4; P � 0.1. N: B6.NOD hearts transplanted into NOD mice treated with combined costimulation blockaderejected at MST 66 � 12.3 days, n � 7. MST nontreated group: 36 � 23 days, n � 4; P � 0.009. Survival of B6.NOD hearts versus islets: P � 0.0038.
L. MAKHLOUF AND ASSOCIATES
DIABETES, VOL. 51, NOVEMBER 2002 3207
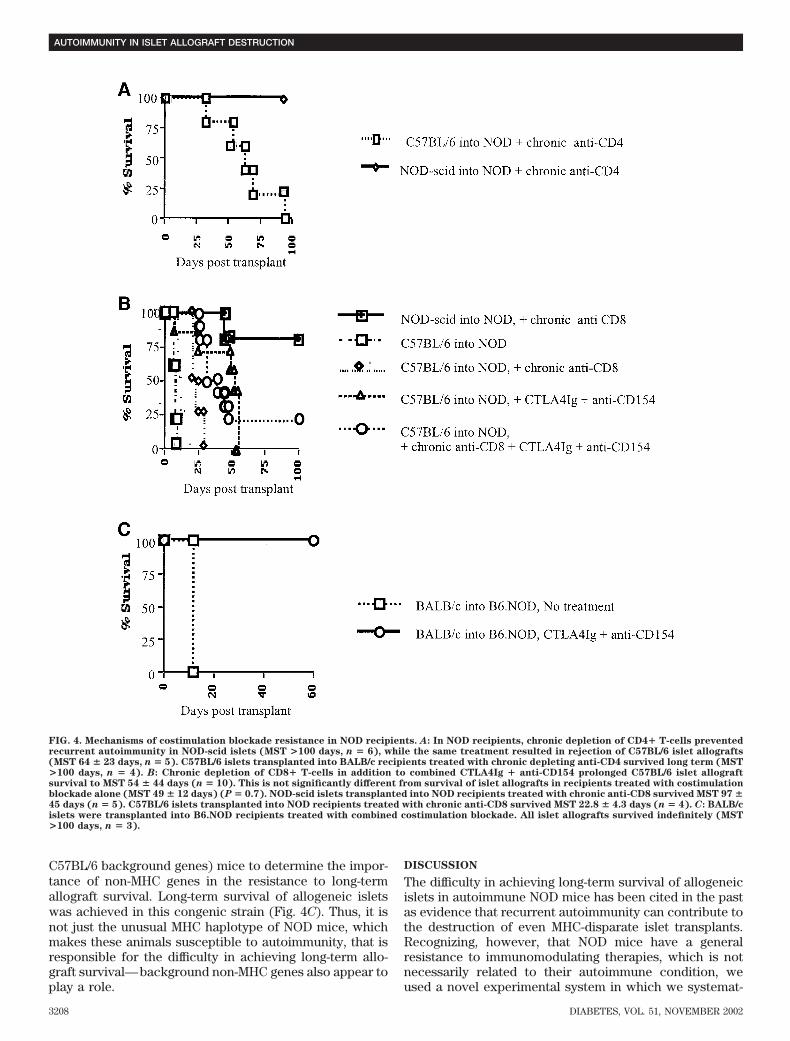
C57BL/6 background genes) mice to determine the impor-tance of non-MHC genes in the resistance to long-termallograft survival. Long-term survival of allogeneic isletswas achieved in this congenic strain (Fig. 4C). Thus, it isnot just the unusual MHC haplotype of NOD mice, whichmakes these animals susceptible to autoimmunity, that isresponsible for the difficulty in achieving long-term allo-graft survival—background non-MHC genes also appear toplay a role.
DISCUSSION
The difficulty in achieving long-term survival of allogeneicislets in autoimmune NOD mice has been cited in the pastas evidence that recurrent autoimmunity can contribute tothe destruction of even MHC-disparate islet transplants.Recognizing, however, that NOD mice have a generalresistance to immunomodulating therapies, which is notnecessarily related to their autoimmune condition, weused a novel experimental system in which we systemat-
FIG. 4. Mechanisms of costimulation blockade resistance in NOD recipients. A: In NOD recipients, chronic depletion of CD4� T-cells preventedrecurrent autoimmunity in NOD-scid islets (MST >100 days, n � 6), while the same treatment resulted in rejection of C57BL/6 islet allografts(MST 64 � 23 days, n � 5). C57BL/6 islets transplanted into BALB/c recipients treated with chronic depleting anti-CD4 survived long term (MST>100 days, n � 4). B: Chronic depletion of CD8� T-cells in addition to combined CTLA4Ig � anti-CD154 prolonged C57BL/6 islet allograftsurvival to MST 54 � 44 days (n � 10). This is not significantly different from survival of islet allografts in recipients treated with costimulationblockade alone (MST 49 � 12 days) (P � 0.7). NOD-scid islets transplanted into NOD recipients treated with chronic anti-CD8 survived MST 97 �45 days (n � 5). C57BL/6 islets transplanted into NOD recipients treated with chronic anti-CD8 survived MST 22.8 � 4.3 days (n � 4). C: BALB/cislets were transplanted into B6.NOD recipients treated with combined costimulation blockade. All islet allografts survived indefinitely (MST>100 days, n � 3).
AUTOIMMUNITY IN ISLET ALLOGRAFT DESTRUCTION
3208 DIABETES, VOL. 51, NOVEMBER 2002

ically compared the survival in NOD recipients of allo-grafts that might be subject to recurrent autoimmunity(islets) with that of grafts that would not be (hearts). Ourfindings indicate that when using B7 plus CD154 T-cellcostimulatory blockade for therapy, recurrent autoimmu-nity appears to contribute to the destruction of allogeneicislets only when there is MHC class II antigen matchingbetween the donor and recipient.
It is of course possible that recurrent autoimmunity iscontributing to the destruction of the class II–mismatchedislet allografts or the autoimmune process happens tocoincide in timing with the alloreactive process thatcauses cardiac graft rejection. There is no formal way ofexcluding this possibility as long as islet destructionoccurs. However, the rapid destruction of MHC classII–matched islets, like that of the NOD-scid islets, stronglysuggests that recurrent autoimmunity would be evidentmuch sooner than the late destruction that is seen whenusing class II-mismatched donors. In addition, the nearlyidentical timing of the islet rejection with that of thecardiac grafts, when there is no MHC class II antigenmatching, suggests that there is no need to invoke atissue-specific autoimmune component as an explanationfor the destruction of MHC-disparate islet allografts. It ispossible that autoreactive T-cells contribute to the recog-nition of mismatched donor strain islets; however, thefindings in Fig. 4, which show that vigorous CD4� orCD8� depletion completely prevented recurrent auto-immunity in NOD-scid islets transplanted into NODrecipients but was not as effective in promoting long-term survival of MHC-mismatched islet allografts inNOD recipients, provide strong support to our conclu-sion that it is predominantly the alloimmune responserather than the autoimmune response that is causingislets to be destroyed in NOD recipients of mismatchedislets.
Strictly speaking, our conclusions apply primarily to thesetting in which costimulatory blockade is used for treat-ment. It is possible that this particular immunomodulatingregimen eliminates some, but not all, of the components ofautoimmunity, leaving other components that can causedestruction of syngeneic and MHC class II–matched allo-geneic islets. This is unlikely since both CTLA4Ig (24) andMR1 (25) are relatively ineffective in reversing establishedautoimmunity. Still, other treatments might not eliminate acomponent of the autoimmune response that could causedestruction of MHC-disparate allografts. Thus, it is impor-tant to be cautious about generalizing from our experi-ments to conclude that recurrent autoimmunity willonly be a problem when there is MHC class II matchingwith all immunomodulatory strategies or in all clinicalsettings. It is possible that our observations do apply toother immunodulatory strategies. Indeed, there is onesupportive study in BB (autoimmune-prone) rats byMarkmann et al. (8) showing that precultured (an im-munomodulatory strategy that has been shown to pro-long islet graft survival) MHC-matched islet allograftsdo worse than precultured MHC-mismatched islets.While these studies did not compare islet and heart graftsurvival and did not dissect class I versus class IImatching effects, the data are consistent with our ob-servations in NOD recipients.
It is interesting to note that although it appears thatautoimmunity played a role in both cases, the rejection ofB6.NOD islets did not take place quite as quickly as thedestruction of NOD-scid islets in our experiments. Onepossible explanation for this observation is that some ofthe non-MHC genes responsible for the development ofdiabetes may do so by causing the islets in NOD mice to bemore susceptible to immune destruction. In addition, thedestruction of more than just the -cells in the classII–matched allogeneic islet transplants indicates that morethan just recurrent autoimmunity was involved. Thus, theprocess of allogeneic islet destruction, even when autoim-munity is involved, is not identical to the process ofautoimmune destruction of syngeneic islets. Apparently,the recurrence of the autoimmune response helps togenerate an allogeneic response that would otherwise bedelayed by costimulatory blockade. One implication ofthis finding is that it is not possible to determine theabsence of an autoimmune component in islet rejection bythe presence of generalized islet destruction.
The difficulty in defining the contribution of recurrentautoimmunity to the destruction of allogeneic islets stemsfrom the general resistance of NOD mice to immunomodu-lating therapies for all types of allografts (16). It is notclear, however, whether the features of the immune sys-tem of these mice that cause them to develop spontaneousautoimmunity are necessarily responsible for this generalresistance. Our data showing long-term survival of alloge-neic islet grafts in B6.NOD (Fig. 4C) indicate that theresistance to tolerance in NOD mice is not directly linkedto the presence of their autoimmune disease and is con-ferred by non-MHC genes. However, it is not clear whetherhumans with type 1 diabetes will show this generalresistance. Interestingly, the results of clinical kidneytransplantation using standard immunosuppressive drugshave been at least as good for patients with type 1 diabe-tes as for patients without autoimmune diabetes (26). Itremains to be determined, however, how diabetic patientsreceiving islet transplantation will respond to varioustypes of selective immunomodulating therapies in thefuture.
It is difficult to draw definitive conclusions about theclinical implications of our findings for patients undergo-ing islet transplantation. If we consider the NOD mouse asone of the best nonhuman models to study autoimmunediabetes (17), then on the one hand, our data suggest thatwhen using certain types of immunosuppression, islettransplants from MHC class II–mismatched donors may beresistant to recurrent autoimmunity. On the other hand,they suggest that unlike most solid organ transplants innonautoimmune recipients, class II–matched islet graftsfor patients with diabetes may be at a disadvantagebecause they will be susceptible to recurrent autoimmu-nity. Therefore, the possibility that MHC class II matchingmay lead to poorer clinical outcomes should be carefullyexamined in all future trials of islet transplantation. Fi-nally, our observations may have important implicationsfor the design of future immunomodulatory/tolerogenictherapies in islet transplantation based on the degree ofMHC sharing between donor and recipients with autoim-mune diabetes.
L. MAKHLOUF AND ASSOCIATES
DIABETES, VOL. 51, NOVEMBER 2002 3209

ACKNOWLEDGMENTS
This study was supported by the Juvenile Diabetes Re-search Foundation International (JDRFI) Center for IsletTransplantation at Harvard Medical School.
We thank Karla Stenger and Susan Shea for the invalu-able technical assistance. We also thank the JDRFI Centerfor Islet Transplantation core for islet isolation for prepa-ration of islets and the NOD mouse core for providingNOD mice.
REFERENCES
1. Sibley RK, Sutherland DE, Goetz F, Michael AF: Recurrent diabetesmellitus in the pancreas iso- and allograft: a light and electron microscopicand immunohistochemical analysis of four cases. Lab Invest 53:132–144,1985
2. Tyden G, Reinholt FP, Sundkvist G, Bolinder J: Recurrence of autoimmunediabetes mellitus in recipients of cadaveric pancreatic grafts. N Engl J Med
335:860–863, 19963. Guo Z, Wu T, Kirchhof N, Mital D, Williams JW, Azuma M, Sutherland DE,
Hering BJ: Immunotherapy with nondepleting anti-CD4 monoclonal anti-bodies but not CD28 antagonists protects islet graft in spontaneouslydiabetic NOD mice from autoimmune destruction and allogeneic andxenogeneic graft rejection. Transplantation 71:1656–1665, 2001
4. Stegall MD, Lafferty KJ, Kam I, Gill RG: Evidence of recurrent autoimmu-nity in human allogeneic islet transplantation. Transplantation 61:1272–1274, 1996
5. Stegall MD, Loberman Z, Ostrowska A, Coulombe M, Gill RG: Autoimmunedestruction of islet grafts in the NOD mouse is resistant to 15-deoxysper-gualin but sensitive to anti-CD4 antibody. J Surg Res 64:156–160, 1996
6. Bartlett ST, Schweitzer EJ, Kuo PC, Johnson LB, Delatorre A, Hadley GA:Prevention of autoimmune islet allograft destruction by engraftment ofdonor T cells. Transplantation 63:299–303, 1997
7. Woehrle M, Markmann JF, Silvers WK, Barker CF, Naji A: Transplantationof cultured pancreatic islets to BB rats. Surgery 100:334–341, 1986
8. Markmann JF, Posselt AM, Bassiri H, Brayman KL, Woehrle M, Hickey WF,Silvers WK, Barker CF, Naji A: Major-histocompatibility-complex re-stricted and nonrestricted autoimmune effector mechanisms in BB rats.Transplantation 52:662–667, 1991
9. Brendel MD, Hering BJ, Schultz AO, Bretzel RG: International IsletTransplant Registry. ITR Newsletter 9:1–20, 2001
10. Wang Y, Pontesilli O, Gill RG, La Rosa FG, Lafferty KJ: The role of CD4�and CD8� T cells in the destruction of islet grafts by spontaneouslydiabetic mice. Proc Natl Acad Sci U S A 88:527–531, 1991
11. Molano RD, Berney T, Li H, Cattan P, Pileggi A, Vizzardelli C, Kenyon NS,Ricordi C, Burkly LC, Inverardi L: Prolonged islet graft survival in NODmice by blockade of the CD40-CD154 pathway of T cell costimulation.Diabetes 50:270–276, 2001
12. Drachenberg C, Papadimitriou J, Weir M, Klassen D, Hoehn-Saric E,Bartlett S: Histologic findings in islets of whole pancreas allografts: lack of
evidence for recurrent cell mediated diabetes mellitus. Transplantation
62:1770–1772, 199613. Sollinger HW, Odorico JS, Knechtle SJ, D’Alessandro AM, Kalayoglu M,
Pirsch JD: Experience with 500 simultaneous pancreas-kidney transplants.Ann Surg 228:284–296, 1998
14. Shapiro A, Lakey J, Ryan E, et al: Islet transplantation in seven patientswith type 1 diabetes mellitus using a glucocorticoid-free immunosuppres-sive regimen. N Engl J Med 343:230–238, 2000
15. Mandel TE, Kovarik J, Koulmanda M, H.M. G: Cellular rejection of fetalpancreas grafts: differences between allo- and xenograft rejection. Xeno-
transplantation 4:2–10, 199716. Markees TG, Serreze DV, Phillips NE, Sorli CH, Gordon EJ, Shultz LD,
Noelle RJ, Woda BA, Greiner DL, Mordes JP, Rossini AA: NOD mice havea generalized defect in their response to transplantation tolerance induc-tion. Diabetes 48:967–974, 1999
17. Atkinson M, Leiter E: The NOD model for type 1 diabetes: as good as itgets. Nat Med 5:601–604, 1999
18. Lacey P, Kostianovsky M: Method for isolation of intact islets of Langer-hans from the rat pancreas. Diabetes 16:35–39, 1967
19. Kishimoto K, Dong VM, Issazadeh S, Fedoseyeva EV, Waaga AM, YamadaA, Sho M, Benichou G, Auchincloss H Jr, Grusby MJ, Khoury SJ, SayeghMH: The role of CD154-CD40 versus CD28–B7 costimulatory pathways inregulating allogeneic Th1 and Th2 responses in vivo. J Clin Invest
106:63–72, 200020. Larsen CP, Elwood ET, Alexander DZ, Ritchie SC, Hendrix R, Tucker-
Burden C, Cho HR, Aruffo A, Hollenbaugh D, Linsley PS, Winn KJ, PearsonTC: Long-term acceptance of skin and cardiac allografts after blockingCD40 and CD28 pathways. Nature 381:434–438, 1996
21. Yamada A, Kishimoto K, Dong VM, Sho M, Salama AD, Anosova NG,Benichou G, Mandelbrot DA, Sharpe AH, Turka LA, Auchincloss H Jr,Sayegh MH: CD28-independent costimulation of T cells in alloimmuneresponses. J Immunol 167:140–146, 2001
22. Trambley J, Bingaman AW, Lin A, Elwood ET, Waitze SY, Ha J, DurhamMM, Corbascio M, Cowan SR, Pearson TC, Larsen CP: AsialoGM1(�)CD8(�) T cells play a critical role in costimulation blockade-resistant allograft rejection. J Clin Invest 104:1715–1722, 1999
23. Williams MA, Trambley J, Ha J, Adams AB, Durham MM, Rees P, CowanSR, Pearson TC, Larsen CP: Genetic characterization of strain differencesin the ability to mediate CD40/CD28-independent rejection of skin allo-grafts. J Immunol 165:6849–6857, 2000
24. Lenschow DJ, Ho SC, Sattar H, Rhee L, Gray G, Nabavi N, Herold KC,Bluestone JA: Differential effects of anti-B7–1 and anti-B7–2 monoclonalantibody treatment on the development of diabetes in the nonobesediabetic mouse. J Exp Med 181:1145–1155, 1995
25. Balasa B, Krahl T, Patstone G, Lee J, Tisch R, McDevitt HO, Sarvetnick N:CD40 ligand-CD40 interactions are necessary for the initiation of insulitisand diabetes in nonobese diabetic mice. J Immunol 159:4620–4627, 1997
26. Kronson JW, Gillingham KJ, Sutherland DE, Matas AJ: Renal transplanta-tion for type II diabetic patients compared with type I diabetic patients andpatients over 50 years old: a single-center experience. Clin Transplant
14:226–234, 2000
AUTOIMMUNITY IN ISLET ALLOGRAFT DESTRUCTION
3210 DIABETES, VOL. 51, NOVEMBER 2002