malaya of universitystudentsrepo.um.edu.my/7753/9/pui_ling.pdf · 2019. 9. 23. · rhodophyta) from...
TRANSCRIPT

TAXONOMY AND MOLECULAR PHYLOGENY OF Halymenia SPECIES (HALYMENIACEAE, RHODOPHYTA)
FROM SOUTHEAST ASIA
TAN PUI LING
FACULTY OF SCIENCE
UNIVERSITY OF MALAYA KUALA LUMPUR
2017
Univers
ity of
Mala
ya

TAXONOMY AND MOLECULAR PHYLOGENY OF
Halymenia SPECIES (HALYMENIACEAE,
RHODOPHYTA) FROM SOUTHEAST ASIA
TAN PUI LING
THESIS SUBMITTED IN FULFILMENT OF THE
REQUIREMENTS FOR THE DEGREE OF DOCTOR OF
PHILOSOPHY
INSTITUTE OF BIOLOGICAL SCIENCES
FACULTY OF SCIENCE
UNIVERSITY OF MALAYA
KUALA LUMPUR
2017
Univers
ity of
Mala
ya

ii
UNIVERSITY OF MALAYA
ORIGINAL LITERARY WORK DECLARATION
Name of Candidate: TAN PUI LING (I.C/Passport No:
Matric No: SHC120092
Name of Degree: DOCTOR OF PHILOSOPHY
Title of Project Paper/Research Report/Dissertation/Thesis (“this Work”):
TAXONOMY AND MOLECULAR PHYLOGENY OF Halymenia SPECIES
(HALYMENIACEAE, RHODOPHYTA) FROM SOUTHEAST ASIA
Field of Study: Algae Biotechnology
I do solemnly and sincerely declare that:
(1) I am the sole author/writer of this Work;
(2) This Work is original;
(3) Any use of any work in which copyright exists was done by way of fair
dealing and for permitted purposes and any excerpt or extract from, or
reference to or reproduction of any copyright work has been disclosed
expressly and sufficiently and the title of the Work and its authorship have
been acknowledged in this Work;
(4) I do not have any actual knowledge nor do I ought reasonably to know that
the making of this work constitutes an infringement of any copyright work;
(5) I hereby assign all and every rights in the copyright to this Work to the
University of Malaya (“UM”), who henceforth shall be owner of the
copyright in this Work and that any reproduction or use in any form or by any
means whatsoever is prohibited without the written consent of UM having
been first had and obtained;
(6) I am fully aware that if in the course of making this Work I have infringed
any copyright whether intentionally or otherwise, I may be subject to legal
action or any other action as may be determined by UM.
Candidate’s Signature Date:
Subscribed and solemnly declared before,
Witness’s Signature Date:
Name:
Designation:
Univers
ity of
Mala
ya

iii
ABSTRACT
Halymenia is a red algal genus classified in the family Halymeniaceae of which
many of the species are poorly known. Despite the abundance of Halymenia species in
the tropical and subtropical waters, there are very few studies from Southeast Asia.
Traditionally, the identification of Halymenia is largely based on morphological
observation in particular the vegetative features. However, these features are not
sufficiently distinctive and may overlap with other taxa due to convergent evolution.
The lack of distinct morphological characters has led to a need for molecular approach
to address the taxonomic confusion in these red algae. Hence, both molecular analyses
and morphological examination were undertaken on specimen from Malaysia, Thailand,
Indonesia and the Philippines to enhance our understanding of the taxonomy and
phylogeny of Halymenia in Southeast Asia. The rbcL, COI-5P, UPA and LSU (28S
rDNA) markers were used to resolve the taxonomic position of Halymenia species.
Combination of the following main diagnostic vegetative characters is crucial for
species identification: habit, branching pattern, order of branching, presence or absence
of surface proliferations or spines, blade margins, blade thickness, cortex thickness,
shape and size of outer cortical cells, shape and size of inner cortical cells and presence
or absence of a stipe. The molecular analyses showed that the genus Halymenia is
polyphyletic and seven distinct species of Halymenia were present in our collections.
Among the seven Halymenia, four were previously described (H. durvillei, H. tondoana,
H. cf. dilatata, H. maculata), two were new species described from the current study (H.
malaysiana, H. johorensis) and one putative new species to be described (Halymenia sp.
A). Phylogenetic analyses indicated that both rbcL and COI-5P are suitable markers to
elucidate taxonomic position, resolve intraspecific genetic variation of Halymenia and
as potential DNA barcodes for Halymenia. In contrast, both UPA and LSU (28S rDNA)
Univers
ity of
Mala
ya

iv
are not suitable markers for molecular phylogenetics and DNA barcoding studies in
Halymenia.
Univers
ity of
Mala
ya

v
ABSTRAK
Halymenia merupakan genus alga merah yang dikelaskan dalam family
Halymeniaceae yang mana banyak spesiesnya kurang dikenali. Walaupun terdapat
banyak spesies Halymenia di perairan tropika dan subtropika, kajian alga ini dari Asia
Tenggara agak terhad. Secara tradisinya, kebanyakan identifikasi Halymenia adalah
berdasarkan pemerhatian morfologi terutamanya ciri-ciri vegetatif. Walau
bagaimanapun, ciri-ciri ini tidak cukup berbeza dan mungkin bertindih dengan taksa
lain disebabkan oleh konvergen evolusi. Kekurangan ciri-ciri morfologi yang berbeza
telah mendorong kepada penggunaan pendekatan molekular untuk menangani
kekeliruan taksonomi dalam alga merah ini. Justeru itu, analisis molekular dan
pemeriksaan morfologi telah dijalankan ke atas specimen dari Malaysia, Thailand,
Indonesia dan Filipina untuk meningkatkan pemahaman terhadap taksonomi dan
filogeni bagi Halymenia di Asia Tenggara. Empat marker molekular [rbcL, COI-5P,
UPA dan LSU (28S rDNA)] telah digunakan untuk menyelesaikan kedudukan
taksonomi spesies Halymenia. Gabungan daripada karakter vegetatif diagnostik utama
berikut adalah penting untuk mengenal pasti spesies: perincian thallus, corak cabangan,
susunan cabangan, kehadiran atau ketiadaan percambahan atau spina di permukaan
thallus, margin thallus, ketebalan thallus, ketebalan korteks, bentuk dan saiz sel dalam
korteks luaran, bentuk dan saiz sel dalam korteks dalaman, kehadiran atau ketiadaan
tangkai. Analisis molekular menunjukkan bahawa genus Halymenia adalah polyphyletic
dan terdapat tujuh spesies Halymenia di dalam koleksi kami. Antara tujuh spesies
Halymenia tersebut, empat daripadanya telah dihuraikan sebelum ini (H. durvillei, H.
tondoana, H. cf. dilatata, H. maculata), dua spesies baru yang dihuraikan dalam kajian
ini (H. malaysiana, H. johorensis) dan satu berkemungkinan merupakan spesies baru
yang perlu dihuraikan (Halymenia sp. A). Analisis filogenetik menunjukkan bahawa
kedua-dua rbcL dan COI-5P adalah marker molekular yang sesuai untuk menjelaskan
Univers
ity of
Mala
ya

vi
kedudukan taksonomi Halymenia, mengungkaikan variasi genetik Halymenia dan
berpotensi sebagai marker barkod DNA untuk Halymenia. Sebaliknya, kedua-dua UPA
dan LSU (28S rDNA) adalah marker molekular yang tidak sesuai untuk molekular
filogenetik dan kajian barkoding DNA Halymenia.
Univers
ity of
Mala
ya

vii
ACKNOWLEDGEMENTS
First and foremost, I would like to express my sincere gratitude to my supervisors,
Assoc. Prof. Dr. Lim Phaik Eem and Prof. Dr. Phang Siew Moi for their guidance,
insight and support throughout my study. Their utmost advices and efforts have
improved my knowledge in particular molecular taxonomy and bioinformatics thus
made the accomplishment of this research possible. I am grateful to Dr. Stefano
Draisma for providing specimens of Halymenia species for my research and Prof. Lin
Showe Mei for her guidance in preparing sections and examining morphological
structures of Halymenia. Their dedications have greatly boosted my interest in this
particular field.
Sincere thanks to Dr. Tan Ji for his guidance in molecular analysis since
undergraduate studies. I am extremely grateful to Dr. Ng Poh Kheng and Dr. Poong Sze
Wan for their utmost assistance and insightful suggestions throughout this whole
research project. Special appreciations go to Dr. Yow Yoon Yen and Dr. Ng Poh Kheng
who have been great companions in the sampling collection trips. I would also like to
thank my wonderful lab members including Miss Jeannette Lai, Miss Fiona Keng, Dr.
Yeong Hui Yin, Dr. Victoria Ng Fong Lee, Dr. Sim Mei Chea, Miss Ou Mei Cing, Mr.
Lee Kok Keong, Mr. James Lim Yong Kian, Mr. Tan Cheng Yau, Mr. Tan Yong Hao
and Miss Yong Wai Kuan for their support.
This study was funded by Ministry of Science, Technology and Environment E-
science Fund (04-01-03-SF0672) and University of Malaya Postgraduate Research Fund
(PG081-2013A). I thank University of Malaya for offering me Fellowship Scheme.
Last but not least, I want to express my appreciation for my family for their
understanding, moral support and encouragement. Again I would like to thank everyone
who helped me in completing this project.
Univers
ity of
Mala
ya

viii
TABLE OF CONTENTS
Abstract ............................................................................................................................ iii
Abstrak .............................................................................................................................. v
Acknowledgements ......................................................................................................... vii
Table of Contents ........................................................................................................... viii
List of Figures ................................................................................................................. xii
List of Tables ................................................................................................................. xiv
List of Symbols and Abbreviations ................................................................................. xv
List of Appendices ........................................................................................................ xvii
CHAPTER 1: INTRODUCTION .................................................................................. 1
1.1 Importance of taxonomy studies .............................................................................. 1
1.2 Algal taxonomy ....................................................................................................... 2
1.3 Research Question ................................................................................................... 4
1.4 Research Objectives................................................................................................. 4
1.5 Research Hypotheses ............................................................................................... 5
CHAPTER 2: LITERATURE REVIEW ...................................................................... 7
2.1 Introduction to algae ................................................................................................ 7
2.2 Red algae ................................................................................................................. 8
2.3 Halymeniaceae ....................................................................................................... 11
2.3.1 Aeodes J. Agardh ...................................................................................... 13
2.3.2 Thamnoclonium Kützing .......................................................................... 14
2.3.3 Grateloupia C. Agardh ............................................................................. 15
2.3.4 Cryptonemia J. Agardh ............................................................................. 16
Univers
ity of
Mala
ya

ix
2.4 Halymenia C. Agardh ............................................................................................ 18
2.4.1 Importance and economic potential of Halymenia ................................... 23
2.5 Genetic diversity of seaweeds ............................................................................... 25
2.6 Molecular phylogenetic methods ........................................................................... 26
2.7 Molecular approaches for taxonomic inference .................................................... 32
2.7.1 Random Amplified Polymorphic DNA (RAPD) ..................................... 33
2.7.2 Restriction Fragment Length Polymorphism (RFLP) .............................. 34
2.7.3 Amplified Fragment Length Polymorphism (AFLP) ............................... 34
2.7.4 Nucleic acid sequencing ........................................................................... 35
2.8 DNA barcoding ...................................................................................................... 36
2.9 Molecular marker for phylogenetic inference ....................................................... 38
2.9.1 Nuclear markers ....................................................................................... 38
2.9.2 Plastid markers ......................................................................................... 40
2.9.3 Mitochondrial markers ............................................................................. 41
2.10 Molecular studies in Halymenia ............................................................................ 43
CHAPTER 3: MATERIALS AND METHODS ........................................................ 47
3.1 Sample collection and processing .......................................................................... 47
3.2 Morphological and anatomical studies .................................................................. 50
3.3 Molecular analyses ................................................................................................ 50
3.3.1 DNA extraction ........................................................................................ 50
3.3.2 Spectrophotometric determination of DNA concentration and purity ..... 51
3.3.3 Polymerase chain reaction (PCR) amplification ...................................... 51
3.3.3.1 rbcL……. .................................................................................. 52
3.3.3.2 COI-5P ...................................................................................... 53
3.3.3.3 UPA….. ..................................................................................... 54
Univers
ity of
Mala
ya

x
3.3.3.4 LSU (28S rDNA) ...................................................................... 55
3.3.4 Determination of the amplification yield and quality by gel
electrophoresis, DNA purification and gene sequencing ........................ 55
3.3.5 Sequence and phylogenetic analyses ........................................................ 56
3.3.6 Haplotype network analyses ..................................................................... 59
CHAPTER 4: RESULTS .............................................................................................. 60
4.1 Morphological and anatomical observations ......................................................... 60
4.1.1 Halymenia malaysiana P.-L. Tan, P.-E. Lim, S.-M. Lin & S.-M. Phang. 60
4.1.2 Halymenia maculata J. Agardh ................................................................ 65
4.1.3 Halymenia cf. dilatata Zanardini ............................................................ 68
4.1.4 Halymenia johorensis P.-L. Tan, P.-E. Lim, S.-M. Lin & S.-M. Phang...70
4.1.5 Halymenia tondoana O. DeClerck & J.J. Hernández-Kantun.................. 74
4.1.6 Halymenia durvillei Bory de Saint-Vincent ............................................. 76
4.1.7 Halymenia sp. A ....................................................................................... 79
4.2 Molecular analyses ................................................................................................ 82
4.2.1 DNA extraction ........................................................................................ 82
4.2.2 PCR amplification .................................................................................... 82
4.2.3 Sequence analyses .................................................................................... 84
4.2.3.1 rbcL………. .............................................................................. 84
4.2.3.2 COI-5P ...................................................................................... 85
4.2.3.3 UPA…. ...................................................................................... 86
4.2.3.4 LSU (28S rDNA) ...................................................................... 87
4.2.4 Phylogenetic analyses ............................................................................... 88
4.2.4.1 rbcL…… ................................................................................... 88
4.2.4.2 COI-5P ...................................................................................... 91
Univers
ity of
Mala
ya

xi
4.2.4.3 UPA…. ...................................................................................... 93
4.2.4.4 LSU (28S rDNA) ...................................................................... 95
4.2.5 Intraspecific genetic diversity of Halymenia malaysiana ........................ 97
4.2.5.1 Haplotype network analysis of Halymenia malaysiana for
rbcL marker .............................................................................. 97
4.2.5.2 Haplotype network analysis of Halymenia malaysiana for
COI-5P marker .......................................................................... 99
CHAPTER 5: DISCUSSION ..................................................................................... 102
5.1 Morphological and anatomical studies ................................................................ 102
5.1.1 Morphological and anatomical distinction among Halymenia species .. 104
5.2 Molecular analyses .............................................................................................. 114
5.2.1 DNA extraction ...................................................................................... 114
5.2.2 PCR amplification .................................................................................. 115
5.2.3 Sequence analyses and molecular phylogenies ...................................... 116
5.2.4 Markers performance and potential DNA barcodes ............................... 119
5.2.5 Genetic diversity of Halymenia malaysiana .......................................... 122
CHAPTER 6: CONCLUSION ................................................................................... 126
6.1 General conclusion and appraisal of this study ................................................... 126
6.2 Future studies on Halymenia ............................................................................... 129
References ..................................................................................................................... 131
List of Publications and Papers Presented .................................................................... 160
Appendices………………………………………………………………………… .. 161
Univers
ity of
Mala
ya

xii
LIST OF FIGURES
Figure 1.1: Flow chart summarizing the research approach of this study.
6
Figure 2.1: Taxonomic classification of Halymeniaceae according to Saunders
and Karft (1996).
12
Figure 3.1: Map showing the collection sites (arrowed and circled) of the
samples in this study.
48
Figure 3.2: Map of the collection sites in Malaysia.
48
Figure 3.3: Map of the collection sites in the Philippines.
49
Figure 4.1: Thallus habit of Halymenia malaysiana.
62
Figure 4.2: Vegetative structures of Halymenia malaysiana. 63
Figure 4.3: Tetrasporangial and cystocarp morphology of Halymenia
malaysiana.
64
Figure 4.4 : Thallus habit of Halymenia maculata.
66
Figure 4.5: Vegetative structures, cystocarp and tetrasporangial morphology
of Halymenia maculata.
67
Figure 4.6: Thallus habit and anatomy of Halymenia cf. dilatata. 69
Figure 4.7: Habit and vegetative morphology of Halymenia johorensis. 72
Figure 4.8: Cystocarp and tetrasporangial morphology of Halymenia
johorensis.
73
Figure 4.9: Thallus habit and anatomy of Halymenia tondoana. 75
Figure 4.10: Thallus habit of Halymenia durvillei. 78
Figure 4.11: Vegetative, tetrasporangial and cystocarp morphology of
Halymenia durvillei.
79
Figure 4.12: Thallus habit and anatomy of Halymenia sp. A 81
Figure 4.13: Electrophoretogram showing amplicons of the plastid rbcL
(~1,400bp) and mitochondrial COI-5P (~600bp) genetic markers.
83
Figure 4.14: Electrophoretogram showing amplicons of the plastid UPA
(~400bp) and nuclear LSU (28S rDNA) (~600bp) genetic markers.
84
Figure 4.15: ML phylogeny inferred based on the rbcL sequences.
90
Figure 4.16: ML phylogeny inferred based on the COI-5P sequences. 92
Univers
ity of
Mala
ya

xiii
Figure 4.17: ML phylogeny inferred based on the UPA sequences.
94
Figure 4.18: ML phylogeny inferred based on the partial LSU (28S rDNA)
sequences.
96
Figure 4.19: Statistical parsimony network for rbcL haplotypes of Halymenia
malaysiana.
98
Figure 4.20: Statistical parsimony networks for COI-5P haplotypes of
Halymenia malaysiana.
100
Univers
ity of
Mala
ya

xiv
LIST OF TABLES
Table 2.1: Summary of several methods of phylogenetic analyses (Adapted from
Soltis and Soltis, 2003; Yang and Rannala, 2012 with modification).
29
Table 3.1: Primers used for amplification of rbcL.
53
Table 3.2: Primers used for amplification of COI-5P.
54
Table 3.3: Primers used for amplification of UPA.
54
Table 3.4: Primers used for amplification of LSU (28S rDNA).
55
Table 3.5: Model and parameters selected by Kakusan3 for ML analysis of
rbcL, COI-5P, UPA and LSU (28S rDNA) datasets.
58
Table 4.1: Percentage of pairwise distance between rbcL sequences of seven
Halymenia species examined in this study, excluding gaps and
ambiguities.
85
Table 4.2: Percentage of pairwise distance between COI-5P sequences of six
Halymenia species examined in this study, excluding gaps and
ambiguities.
86
Table 4.3: Percentage of pairwise distance between UPA sequences of six
Halymenia species examined in this study, excluding gaps and
ambiguities.
87
Table 4.4: Percentage of pairwise distance between LSU (28S rDNA) sequences
of six Halymenia species examined in this study, excluding gaps and
ambiguities.
88
Table 4.5: A summary of the rbcL haplotype diversity of Halymenia malaysiana
with number of individuals (N) and number of haplotypes (Nh) from
each location.
99
Table 4.6: A summary of the COI-5P haplotype diversity of Halymenia
malaysiana with number of individuals (N) and number of haplotypes
(Nh) from each location.
101
Table 5.1: Comparison of morphological and anatomical characters among
closely related foliose Halymenia species in this study.
107
Table 5.2: Comparison of morphological and anatomical characters among
closely related branched Halymenia species in this study.
109
Univers
ity of
Mala
ya

xv
LIST OF SYMBOLS AND ABBREVIATIONS
A : Adenine
AFLP : Amplified fragment length polymorphism
AICc : Corrected Akaike Information Criterion
BI : Bayesian Inference
BICc : Corrected Bayesian Information Criterion
BP : Bootstrap Percentage
bp : base pair
C : Cytosine
CI : Consistency Index
cm : centimeter
cox1 or COI : cytochrome c oxidase subunit 1
COI-5P : 5’ end of the cytochrome c oxidase subunit 1 gene
cox2 : cytochrome c oxidase subunit 2
cox2-3 spacer : spacer region between cytochrome c oxidase subunit 2 and 3
cox3 : cytochrome c oxidase subunit 3
dATP : Deoxyadenosine triphosphate
dCTP : Deoxycytidine triphosphate
dGTP : Deoxyguanosine triphosphate
DNA : Deoxyribonucleic acid
dNTP : Deoxyribonucleotide triphosphate
G : Guanine
HCl : Hydrochloric acid
ITS : Internal transcribed spacer
kb : kilobase
LSU : Large subunit of ribosomal DNA (28S rDNA)
m : meter
MCMC : Markov chain Monte Carlo
ML : Maximum likelihood
mM : milimolar
MP : Maximum parsimony
N : Number of individuals
NJ : Neighbour joining
Univers
ity of
Mala
ya

xvi
Nh : Number of haplotypes
ng : nanogram
OD : Optical density
PAUP : Phylogenetic analysis using parsimony
PCR : Polymerase chain reaction
pmol : Picomole
PP : Posterior probabilities
RAPD : Random amplified polymorphic DNA
rbcL : ribulose-1, 5-bisphosphate carboxylase/oxygenase large subunit
rbcS : ribulose-1, 5-bisphosphate carboxylase/oxygenase small subunit
rDNA : Ribosomal deoxyribonucleic acid
RFLP : Restriction fragment length polymorphism
RI : Retention index
RNA : Ribonucleic acid
RNase : Ribonuclease
rRNA : Ribosomal ribonucleic acid
RuBisCO : Ribulose-1, 5-bisphosphate carboxylase/oxygenase
SSU : Small subunit of ribosomal DNA (18S rDNA)
T : Thymine
U : unit
UPA : Universal plastid amplicon
UV : Ultraviolet
μL : microlitre
μm : micrometer
°C : degree Celcius
Univers
ity of
Mala
ya

xvii
LIST OF APPENDICES
Appendix A: List of specimens examined in this study with information on
herbarium number, locality, collector, date of collection and field
number.
162
Appendix B: List of published sequences used for rbcL analyses with collection
details and GenBank accession numbers.
172
Appendix C: List of published COI-5P, UPA and LSU (28S rDNA) sequences
with collection details and GenBank accession numbers for
analyses.
174
Appendix D: Uncorrected pairwise distance matrix of the rbcL sequences.
175
Appendix E: Uncorrected pairwise distance matrix of the COI-5P sequences.
183
Appendix F: Uncorrected pairwise distance matrix of the UPA sequences.
188
Appendix G: Uncorrected pairwise distance matrix of the LSU (28S rDNA)
sequences
190
Univers
ity of
Mala
ya

1
CHAPTER 1: INTRODUCTION
1.1 Importance of taxonomy studies
Taxonomy is the science that deals with identification, description, naming and
classification of living organisms (Lincoln et al., 1998; Wägele, 2005). It is fundamental
to the inventory of life on earth and understanding the variety of life forms (Lincoln et
al., 1998; Wägele, 2005). Without taxonomy, nobody would be certain of the identity of
organisms they were interested in, or whether they belonged to the same or different
species as the organisms studied by others (Nature, 2002). According to Narendran
(2000), it is absolutely necessary to recognize the correct name of the organism before
initiating any kind of studies. This is because the correct scientific name of the organism
acts as a functional label, using which various pieces of information concerning that
organism, including all the past work done on it, can be retrieved and stored ensuing
ease of reference (Narendran, 2000).
Taxonomy provides basic understanding about biodiversity that is a prerequisite for
all other biological research including medicine, bioprospecting, fisheries, quarantine,
defense, etc. (Narendran, 2000). It also plays a significant role in conservation by
documenting, describing, and cataloguing all the living things. Taxonomic information
is essential to understand the pattern of biodiversity which is useful in determining
biodiversity hotspots (regions with exceptionally high species richness) and
subsequently extra conservation resources are focused on those areas (Myers et al.,
2000). We cannot certainly expect to conserve organisms that we cannot identify, and
cannot develop the species conservation plans if we cannot recognize and describe the
interacting components of natural ecosystems (Rojas, 1992; Samper, 2004). Thus,
Univers
ity of
Mala
ya

2
effective control and management measures can only be executed when invasive species
are accurately and promptly identified. As revealed by Guerra-García et al. (2008), it is
estimated that about 90% of the world species are still unknown and most of the extinct
species still undescribed. Obviously, effective and prompt conservation measures must
be taken to halt this decline (Guerra-García et al., 2008).
1.2 Algal taxonomy
The exercise of discovering and documenting biodiversity has been given an
increased sense of urgency as the anthropogenic impacts are perilously altering the biota
of the Earth (Cardinale et al., 2012). Studies by De Clerck et al. (2013) have shown that
unlike the well-studied groups such as birds, mammals and higher plants which have a
decrease in the description rates as fewer species remained to be described (Costello and
Wilson, 2011, Joppa et al. 2011), there is no evidence for a decrease in the description
rates of algal species. Additionally, there is a gradual overall increase in the description
rates of algal species over time (De Clerck et al., 2013). Thus, the algae are a group of
organisms worth for study since many species have not yet been identified and the
precise number of species remains elusive (Robba et al., 2006).
Algal taxonomy studies have been the focus of research, particularly on the
economically important species (e.g. Kappaphycus, Eucheuma, Gracilaria) which have
great potential for the commercialisation of seaweed industries, in addition to
physiological aspects related to mass cultivation and the production of useful products
(Chan et al., 2006). In order to fully utilize the commercially important seaweeds, it is
important to understand their biochemical composition, ecology and more importantly
their taxonomic status. Therefore, algal taxonomy studies lies mainly in correct
identification for cultivation, exploitation and conservation purposes. However, the
Univers
ity of
Mala
ya

3
identification of algae, particularly the Rhodophyta, can be extremely difficult based on
morphological criteria alone due to their simple morphology and anatomy, rampant
phenotypic plasticity, convergence and alternation of heteromorphic generations
(Saunders, 2005). Therefore, molecular tools have been used to evaluate the limits of
morpho-species and to delineate boundaries between species (Manhart and McCourt,
1992; John and Maggs, 1997).
The ordinal classification of the Florideophyceae which based largely on the
characters of female reproductive anatomy before and after fertilization by Kylin (1956)
gave significant contribution to red algal systematics. The ultrastructure studies of pit
connections also leading to the refinement of the Kylinian ordinal classification.
However, molecular approaches to systematics provided significant insights into the
evolution of red algae and led to the proposal of several new orders. The application of
molecular techniques for use in algal taxonomy has also greatly improved our
understanding of species and their relationships. There are two approaches extensively
used by phycologists to assess algal species level diversity and discover new species:
(1) DNA taxonomy in which species are delineated based on sequence data using
evolutionary species concepts (Vogler and Monaghan, 2007) and (2) DNA barcoding
which identifies specimens based on sequence similarity against a database of a priori
defined species (Hebert et al., 2003). Phylogenies offer new ways to estimate
biodiversity, to assess conservation priorities, and to evaluate the evolutionary history in
any set of species (Mace et al., 2003). Nevertheless, molecular phylogenies are not
completely congruent with morphological taxonomy (Fama et al., 2002) and might
detect cryptic species in “species” complexes that were previously identified solely by
morphology (Zuccarello and West, 2003; Lewis and Flechtner, 2004). Consequently,
the combination of both molecular and morphological techniques is a promising
Univers
ity of
Mala
ya

4
approach for delineating species boundaries (Nam et al., 2000; Yoshida et al., 2000; de
Senerpont Domis et al., 2003; Kawai and Sasaki, 2004).
In the context of Halymenia, the taxonomy studies of this genus in Southeast Asia
remain scarce. The identification of Halymenia is problematic if based solely on
morphological characteristics due to its immense morphological plasticity and few
distinctive morphological features (Tan et al., 2015; 2017). This impels the use of
molecular techniques in the identification of Halymenia species. More studies should be
performed to better understand the biodiversity, genetic diversity and phylogeny of this
red seaweed because (1) Halymenia is rich in carrageenan and can be a potential source
for carrageenan and food production (Freile-Pelegrin et al., 2011; Kho et al., 2016); (2)
Southeast Asia is well known to be a biodiversity hotspot, with many organisms yet to
be identified (Sodhi et al., 2004). We believe that there are many yet to be discovered
Halymenia species in Southeast Asia albeit our attempts.
1.3 Research Question
How much biodiversity of Halymenia in Malaysia, Thailand, Indonesia and the
Philippines?
1.4 Research Objectives
The purpose of this study is to undertake both morphological examination and
molecular analyses to understand the species diversity of Halymenia in Malaysia,
Thailand, Indonesia and the Philippines, and to elucidate the relationships between these
species.
Univers
ity of
Mala
ya

5
The objectives of this study are:
1. To collect, describe and document the diversity of Halymenia from various localities
in Malaysia, Thailand, Indonesia and the Philippines based on morphological and
anatomical features
2. To elucidate the phylogenetic relationship between Halymenia species using
molecular approaches based on the DNA sequences of selected genetic markers from
different genomes
3. To assess the utility of the genetic markers for molecular phylogenetics studies and
their potential as DNA barcode for Halymenia
1.5 Research Hypotheses
a) H0: All morphological features were equally reliable as diagnostic characters
H1: Not all morphological features were equally reliable as diagnostic characters
b) H0: Identification based on molecular phylogenies were coherent with morphological
characters
H1: Identification based on molecular phylogenies were not coherent with
morphological characters
c) H0: Phylogenies of different molecular genetic markers were congruent and have
similar levels of resolution
H1: Phylogenies of different molecular genetic markers were not congruent and do
not have similar levels of resolution
Univers
ity of
Mala
ya

6
A flow chart summarizing the research approach of this study is presented in Figure
1.1.
Figure 1.1: Flow chart summarizing the research approach of this study.
Sample collection from various
localities in Malaysia, Thailand, Indonesia
and the Philippines
Morphological studies
Gross morphology
Anatomy
Derivation of conclusion
Samples preserved in
formalin seawater or
herbarium
Silica-gel preserved
samples or herbarium
Molecular studies
DNA extraction
Polymerase Chain
Reaction (PCR)
PCR purification
DNA sequencing
Phylogenetic analyses
Univers
ity of
Mala
ya

7
CHAPTER 2: LITERATURE REVIEW
2.1 Introduction to algae
Algae are photosynthetic organisms mainly living in aquatic habitat but excluding
seagrasses (aquatic angiosperm). They have a tremendously confusing array of cell
cycles, cell morphologies and live in a multitude of habitats (Bhattacharya and Medlin,
1998). They exhibit a broad range of morphological diversity, ranging from the
unicellular microscopic phytoplankton (e.g. Chlorella) to the macroscopic marine algae
(e.g. huge kelps over 50 meters long).
The unicellular and multicellular forms of algae are known as microalgae and
macroalgae respectively. Microalgae are generally photosynthetic and heterotrophic
organism with the potential for cultivation as energy crops. They can be cultivated
under certain conditions to give rise to various commercial byproducts such as oils, fats,
sugars and functional bioactive compounds. On the other hand, macroalgae, which are
mainly found in the Divisions Chlorophyta (green algae), Phaeophyta (brown algae) and
Rhodophyta (red algae), are commonly called seaweeds owing to their size,
multicellular construction and attachment to form substrata (Dawes, 1998).
As reported by Dhargalkar and Kavlekar (2004), the criteria used to distinguish the
different algal group are based on the photosynthetic pigments, storage food products,
cell wall component and fine structure of the cell and flagella. The green algae
(Chlorophyta) possess photosynthetic pigments such as chlorophyll a and b, giving
them a bright green colour, as well as the accessory pigments beta-carotene and
xanthophylls. The cell walls of green algae are generally composed of cellulose, with
some incorporation of calcium carbonate in some species. They stored their food in the
Univers
ity of
Mala
ya

8
form of starch in chloroplast (Leliaert et al., 2012). Likewise, the brown algae
(Phaeophyta) possess large quantities of brown coloured pigment fucoxanthin which
masks the colour of other pigments such as beta-carotene, xanthophylls, chlorophyll a
and c. The cell walls of brown algae are made up of cellulose and polysaccharides
known as alginic acid. Laminarin, mannitol are the food reserve of the brown algae
(Dhargalkar and Kavlekar, 2004). On the other hand, the red algae (Rhodophyta)
possess photosynthetic pigments chlorophyll a and the accessory pigments such as α
and β carotenes, xanthophylls zeaxanthin, lutein, r-phycocyanin, r-phycoerythrin, c-
phycocyanin and allophycocyanin. The cell walls of red algae has a firm inner layer
containing cellulose and a mucilaginous or gelatinous outer layer composed of
sulphated carbohydrates such as agar, carrageenan and porphyran. They stored their
food as floridean starch in the cystoplasm (Maggs et al., 2007).
2.2 Red algae
The red algae (Rhodophyta) are an ancient photosynthetic eukaryotic lineage,
predominating along the coastal and continental shelf areas of tropical, temperate and
cold-water regions (Lüning, 1990). They are comprised of about 6000 species and about
680 genera ranging from unicellular to complex multicellular taxa that found mainly in
the marine environment (Woelkerling, 1990; Yoon et al., 2010). They play essential
roles as primary producers, habitat formers for benthic communities and provide
nurseries for fisheries (Mann, 1973).
Despite the red algae have evolved a diverse range of modifications in cellular
organization and general morphology (Pueschel, 1990), they are distinguishable
amongst eukaryotic lineages by a combination of biochemical and ultrastructural
features (Maggs et al., 2007). The most noticeable feature of the red algae is the absence
Univers
ity of
Mala
ya

9
of flagella, basal bodies and centrioles in all life stages (Pueschel, 1990; De Clerck et al.,
2012). The chlorophyll a is the only chlorophyll in the red algae (van den Hoek et al.,
1995). They also possess α and β carotenes, xanthophylls zeaxanthin and lutein, and
phycobiliproteins such as r-phycocyanin, r-phycoerythrin, c-phycocyanin and
allophycocyanin as the accessory photosynthetic pigments (Dawes, 1998). Despite not
all Rhodophyta appears red, the red colour of these algae results from the predominantly
phycoerythrin pigments which absorb blue-green light and reflect red light (Boney and
Corner, 1960). The lack of external endoplasmic reticulum within chloroplast and the
presence of unstacked thylakoids with stalked phycobilisomes in the red algal plastids
are also significant ultrastructural features that distinguished them from other eukaryotic
lineages (Woelkerling, 1990; Maggs et al., 2007). The red algae are also characterized
by the presence of floridean starch as storage product in the cystoplasm, whereas the
green algae and plants store starch in the chloroplasts (Maggs et al., 2007). The red algal
cell wall has a firm inner layer containing cellulose and a mucilaginous or gelatinous
outer layer composed of sulphated carbohydrates such as agar, carrageenan and
porphyran. Possession of pit plugs is also a unique and distinctive feature of
Rhodophyta. The cytokinesis in red algae is incomplete and resulted in a small pore left
in the middle of the newly formed partition then the pit plug formed by deposition of
cytoplasmic substance in the wall of the gap connected to the cells (Pueschel and Cole,
1982; Maggs et al., 2007).
Rhodophyta was traditionally divided into two distinct classes, Bangiophyceae and
Florideophyceae, based on morphological, anatomical, and life-history differences of
the red algae (Dixon, 1973; van den Hoek et al., 1995; Müller et al., 2001). The smaller
class Bangiophyceae encompasses the most primitive red algal forms with relatively
simple morphologies (Müller et al., 2001). Little is known about the life histories of the
bangiophytes which seem to be diverse (Brodie and Irvine, 2003). Meanwhile, the more
Univers
ity of
Mala
ya

10
complex Florideophyceae has much diverse morphological structures and an intricate
triphasic life history (Verbruggen et al., 2010). Instead of diploid sporophyte, the
immediate product of post-fertilization unique to Florideophyceae is a hemiparasitic
diploid tissue termed gonimoblast surrounded by female nutritive tissues, which known
as cystocarp (Maggs et al., 2007). In order to compensate for the lack of motile sperm in
the red algae (Searles, 1980), plenty genetically identical diploid spores that give rise to
sporophytes are released.
The ultrastructure studies of pit connections gave significant contribution to red algal
systematics. A number of molecular phylogenetic studies based on different markers
were performed and provided significant insights into the evolution and relationships of
red algae particularly for the refinement at ordinal level (Freshwater et al., 1994; Ragan
et al., 1994; Saunders and Hommersand, 2004; Yoon et al., 2006). A new taxonomic
scheme was then proposed by Saunders and Hommersand (2004) based on previous
molecular phylogenies and ultrastructural characters including the Golgi-endoplasmic
reticulum (ER) association. A new phylum Cyanidiophyta with a single class
Cyanidiophyceae under the new subkingdom Rhodoplantae was proposed in addition to
the phylum Rhodophyta (Saunders and Hommersand, 2004). Additionally, three
subphyla were established for Rhodophyta: (1) Rhodellophytina with a single class
Rhodellophyceae of which composed of unicells or pseudofilaments with the cells
arranged in a row surrounded by the common gelatinous envelope; they have no sexual
reproduction; (2) Metarhodophytina with a single class Compsopogonophyceae of
which composed of filamentous or pseudoparenchymatous members which have a
biphasic life cycle; and (3) Eurhodophytina which contains the classes Bangiophyceae
and Florideophyceae, is defined by the occurrence of pit plugs in at least one of the
phases of the life history (Saunders and Hommersand, 2004). Subsequently, Yoon et al.
(2006) proposed a different classification system where Rhodophyta is divided into two
Univers
ity of
Mala
ya

11
subphylums- Cyanidiophytina and Rhodophytina. Cyanidophytina with one class,
namely Cyanidophyceae, while the Rhodophytina with six classes, namely (1)
Bangiophyceae, (2) Compsopogonophyceae, (3) Florideophyceae, (4)
Porphyridiophyceae, (5) Rhodellophyceae, and (6) Stylonematophyceae. To date,
taxonomic position of Rhodophyta is still in a state of flux due to the limited studies
above ordinal level.
2.3 Halymeniaceae
The Halymeniaceae is one of the taxonomically challenging families, in which the
diagnostic features especially cryptic or uncertain (Gargiulo et al., 2013). The
Halymeniaceae was previously placed under the large order of Cryptonemiales (Kylin,
1956). Subsequently, Saunders and Kraft (1996) proposed that two families, the
Halymeniaceae and Sebdeniaceae should be placed under the new, smaller order
Halymeniales based on molecular data, along with a review of relevant literature
depicting vegetative and reproductive features of the studied taxa. The taxonomic
classifications of this family are shown in Figure 2.1.
The Halymeniaceae is the most diverse family in the order Halymeniales, consists of
31 genera and approximately 317 species (Guiry and Guiry, 2017). It is characterized by
its multiaxial thallus structure with a “medulla of slender to robust, sparse to dense,
filaments and a cortex of ovoid cells in anticlinal filaments or pseudoparenchymatous,
medulla with or without stellate or refractive ganglioid cells” (Womersley and Lewis,
1994) and sexual reproduction, involving carpogonial branches and auxiliary cells borne
in separate filamentous ampullae (Chiang, 1970; Hommersand and Fredericq, 1990).
Members of this family have a triphasic life history with isomorphic gametophytes and
tetrasporophytes (Womersley and Lewis, 1994; Norris, 2014). Cruciately divided
Univers
ity of
Mala
ya

12
Classification:
Empire Eukaryota
Kingdom Plantae
Subkingdom Biliphyta
Phylum Rhodophyta
Subphylum Eurhodophytina
Class Florideophyceae
Subclass Rhodymeniophycidae
Order Halymeniales
Family Halymeniaceae
Figure 2.1: Taxonomic classification of Halymeniaceae according to Saunders and
Kraft (1996).
tetrasporangia either scattered over the thallus surface, grouped in sori or borne in
modified areas of tissue (nemathecia), while spermatangia are superficial on the thallus,
cut off from terminal cortical cells (Norris, 2014). Sexual thalli are monoecious or
dioecious. Connecting filaments develop from the fertilized carpogonium, contact and
diploidize the auxiliary cell, which then develops to the carposporophyte. Cystocarps
are embedded in the thallus and in most genera are surrounded by sparse to conspicuous
involucres originated from the ampullary filaments or also including medullary
filaments (Womersley and Lewis, 1994; Norris, 2014).
Chiang (1970) proposed that the shape and the structure of the auxiliary cell
ampullae could be useful to define some genera within the red algal family
Halymeniaceae. Five types of auxiliary cell ampullae have been proposed: Aeodes,
Cryptonemia, Halymenia, Grateloupia and Thamnoclonium (Chiang, 1970). In addition,
Kawaguchi et al. (2004) suggested that the structure of carpogonial-branch ampullae
Univers
ity of
Mala
ya

13
may also have taxonomic value similar to that of auxiliary cell ampullae. Even though
reproductive anatomy and postfertilization development have been used for separating
many genara of red algae (Kraft, 1977; Gargiulo et al., 1986; Hommersand et al., 1999),
reproductive uniformity within halymeniacean genera has been claimed and supported
by several authors (Kylin, 1956; Balakrishnan, 1961; Kawabata, 1963). Moreover,
postfertilization development is not well documented in most of the members of the
Halymeniaceae (Balakrishnan, 1960; Kraft, 1977; Gargiulo et al., 2013). Therefore,
vegetative features were emphasized rather than reproductive characters in genus-level
taxonomy (Kylin, 1956; Guiry and Irvine, 1974; Kraft, 1977). It is clear that separation
of many of the genera in this family requires further study and species concepts within
these genera are in need of review (De Smedt et al., 2001). Four genera with different
types of auxiliary cell ampullae as proposed by Chiang (1970): Aeodes, Thamnoclonium,
Grateloupia and Cryptonemia were selected and discussed as follows.
2.3.1 Aeodes J. Agardh
Aeodes J. Agardh is one of the red algal genera in the family Halymeniaceae with
four taxonomically accepted species (Guiry and Guiry, 2017). It is mostly distributed in
New Zealand, South Africa, Mediterranean Sea (Guiry and Guiry, 2017). Aeodes, based
on the generitype, Aeodes nitidissima J. Agardh, is characterized by foliose, lobed or
divided thallus, spreading laterally from the holdfast with very short stipe, a medulla
with few slender rhizoids and a relatively thick but loose involucre (Womersley and
Lewis, 1994). It is most closely related to Pachymenia J. Agardh which differs in the
above features such as the characteristics of stipe and medulla.
Cruciately divided tetrasporangia scattered, attached to mid cells of the cortex while
spermatangia developed from the surface cortical cells (Womersley and Lewis, 1994).
Univers
ity of
Mala
ya

14
The carposporophyte is surrounded by a prominent involucre developed from the
ampullary filaments (Womersley and Lewis, 1994). According to Chiang (1970), the
Aeodes-type auxiliary cell ampulla is very bushy, with up to four (rarely five) orders of
ampullar filaments and is cup-shaped in outline. Carpogonial branches are two-celled
and the carpogonial branch ampullae in Aeodes are the most complex ampullae which
branched to the third or fifth orders (Kawaguchi et al., 2004).
2.3.2 Thamnoclonium Kützing
Thamnoclonium Kützing is one of the red algal genera in the family Halymeniaceae
with only two taxonomically accepted species (Guiry and Guiry, 2017), including
Thamnoclonium dichotomum (J.Agardh) J.Agardh and Thamnoclonium lemannianum
Harvey. Thamnoclonium was founded by Kützing (1843) based on the generitype,
Thamnoclonium hirsutum Kützing collected in Western Australia. Thamnoclonium
hirsutum is now regarded as a synonym of Thamnoclonium dichotomum. This genus is
characterized by terete to compressed thalli with irregularly to subdichotmously
branches, covered throughout with short, irregularly branches excrescences, coated with
a thin layer of sponge, a thick secondary cortex with numerous growth rings developing
below and reproductive structures borne in special small fertile leaflets clustered at the
apices and upper margins (Womersley and Lewis, 1994).
Cruciately divided tetrasporangia in nemathecia on fertile leaflets, cut off from
subsurface cells while spermatangia cut off from outer cortical cells (Womersley and
Lewis, 1994). The carposporophyte is enclosed by a prominent involucre developed
from branched ampullary filaments (Womersley and Lewis, 1994). According to Chiang
(1970), the Thamnoclonium-type auxiliary cell ampulla is comprised of a single primary
ampullar filament and three or five 2- to 5-celled secondary ampullar filaments and is
Univers
ity of
Mala
ya

15
irregular in outline. Carpogonial branches are two-celled and the carpogonial branch
ampullae in Thamnoclonium are the simplest ampullae which branched only to the
second orders (Kawaguchi et al., 2004).
2.3.3 Grateloupia C. Agardh
Grateloupia C. Agardh is the largest red algal genus in the family Halymeniaceae,
comprising of 96 taxonomically accepted species (Guiry and Guiry, 2017). It is widely
distributed in warm temperate to tropical waters throughout the world (Lin et al., 2008;
Guiry and Guiry, 2017). Grateloupia, based on the generitype, Grateloupia filicina (J.
V. Lamouroux) C. Agardh, is characterized by terete to bladelike thalli that range from
lubricous to cartilaginous in texture, the presence of irregularly oriented filaments in the
medulla and a two-celled carpogonial branch borne in an ampulla composed of two
orders of branches (Womersley and Lewis, 1994; Lin et al., 2008). This genus includes
taxa with diverse range of habits, ranging from finely pinnate (e.g. G. filicina), foliose
(eg. G. turuturu Yamada) to subdichotomous blades (eg. G. dichotoma J. Agardh)
(Mateo-Cid et al., 2005).
Cruciately divided tetrasporangia embedded in the outer cortex, scattered over the
blade surface while spermatangia are borne superficially in whitish sori or scattered
over the blade surface (Norris, 2014). The carposporophyte is surrounded by a moderate
involucre derived from the ampullary filaments as well as the medullary filaments
(Womersley and Lewis, 1994; Norris, 2014). The carpogonial branch ampullae in
Grateloupia are the simplest ampullae which branched only to the second orders
(Kawaguchi et al., 2004). According to Chiang (1970), the Grateloupia-type auxiliary
cell ampulla is simple with a single primary ampullar filament and two or three 7- to 13-
celled secondary ampullar filaments and the mature ampulla is conical in outline.
Univers
ity of
Mala
ya

16
Following, Lin et al. (2008) reported two different patterns of the development of the
auxiliary-cell ampullae: (1) G. taiwanensis-type composed of three orders of
unbranched filaments that branch after diploidization of the auxiliary cell, and (2) G.
orientalis-type composed of two orders of unbranched filaments that do not branch after
diploidization of the auxiliary cell.
According to Womersley and Lewis (1994), Grateloupia and Halymenia differs in
the following aspects: (1) a lax medulla with irregularly oriented filaments in the former
and anticlinal filaments in the latter; and (2) the auxiliary cell ampullae are simple,
conical with the filaments converging above in the former and the open, spreading one
in the latter. Species identification in Grateloupia is difficult due to its high
morphological plasticity which variable in overall habit, texture, cortex structure, and
the location of reproductive structures (De Clerck et al., 2005; Wilkes et al., 2005; Yang
et al., 2013b). Although many taxa are still in need of review, recent studies combining
both molecular and morphological analyses have contributed to clearer species
circumscriptions especially for the morphologically similar species (Wang et al., 2000;
Kawaguchi et al., 2001; Gavio and Fredericq, 2002; Yang et al., 2013b).
2.3.4 Cryptonemia J. Agardh
Cryptonemia J. Agardh is a red algal genus comprising of 45 taxonomically accepted
species (Guiry and Guiry, 2017). It is mostly distributed in warm temperate to tropical
waters (Womersley and Lewis, 1994; Guiry and Guiry, 2017). Cryptonemia was
established by J. Agardh (1842) based on the generitype, Cryptonemia lactuta J.Agardh.
Cryptonemia lactuta is now regarded as a synonym of Cryptonemia lomation (Bertoloni)
J.Agardh.
Univers
ity of
Mala
ya

17
Members of this genus are primarily characterized by the well-developed stipe and /
or midrib, the presence of periclinal filaments and highly refractive cells in the medulla,
a relatively thin cortex and bushy ampullar filaments with up to four orders (Abbott,
1967; Chiang, 1970, Womersley and Lewis, 1994; Kim et al., 2012). Cruciately divided
tetrasporangia embedded in the cortex, scattered over the thallus surface while
spermatangia are superficial over the thallus (Norris, 2014). The carposporophyte is
surrounded by a slight involucre originated from elongation of the ampullary filaments
(Womersley and Lewis, 1994; Norris, 2014). According to Chiang (1970), the
Cryptonemia-type auxiliary cell ampulla is very similar to the Aeodes-type with bushy
ampullar filaments branched up to four orders but the outline of the Cryptonemia-type is
conical instead of cup-shaped in the Aeodes-type. Carpogonial branches are two-celled
and the carpogonial branch ampullae in Cryptonemia are reported to be branched to the
third and rarely fourth orders (Kawaguchi et al., 2004).
The distinction between Halymenia and Cryptonemia is difficult. According to
Abbott (1967), Cryptonemia can be differentiated from Halymenia by having inner
cortex of unmodified cells (Halymenia with elongate or stellate inner cortical cells) and
medulla with predominantly periclinal filaments in contrast to the predominantly
anticlinal filaments in Halymenia. The majority of Cryptonemia species usually have
cartilaginous, branched, perennial stalks and midribbed blades (Womersley and Lewis,
1994; Guimarães and Fujii, 1998). However, some species of Halymenia such as H.
stipitata I. A. Abbott has well-developed stipe and H. vinacea M.Howe & W.R.Taylor
has short midrib too (Guimarães and Fujii, 1998; Kawaguchi et al., 2002). Although
Cryptonemia and Halymenia are grouped under different types of auxiliary cell
ampullae based on Chiang’s generic concept, the reliability of these features has been
doubted by different authors who have found intermediate forms (e.g. H. assymetrica
Gargiulo, de Masi & Tripodi by Gargiulo et al., 1986; H. maculata J. Agardh by
Univers
ity of
Mala
ya

18
Kawaguchi et al., 2002). As shown by D’Archino et al. (2014), although not all the
issues in the genera Cryptonemia and Halymenia have been solved, molecular analysis
has contributed to the clarification of generic boundaries in the family Halymeniaceae.
Continued molecular analyses in concert with detailed anatomical studies will help to
clarify the taxonomy of this family and may also reveal the anatomical characters that
can be used to identify the groups (D’Archino et al., 2014). A molecular analysis by
Kim et al. (2012) showed that C. rotunda (Okamura) Kawaguchi is distantly related to
other members of the genus, thus this genus is in need of revision.
2.4 Halymenia C. Agardh
The marine red algal genus Halymenia C. Agardh is one of several species-rich red
algal genera in the family Halymeniaceae and includes 79 taxonomically accepted
species (Guiry and Guiry, 2017). It is mostly distributed in tropical and subtropical
regions (Gargiulo et al., 1986; Hernández- Kantun et al., 2009; Tan et al., 2015).
Halymenia was established by C. Agardh (1817) based on the generitype, Halymenia
floresii (Clemente) C. Agardh collected from Cádiz, Spain.
The genus is mainly characterized by gelatinous thalli, presence of anticlinal
filaments and refractive ganglionic cells in the medulla, stellate cells in the inner cortex,
and auxiliary cell ampullae with branched secondary filaments (Balakrishnan, 1961;
Abbott, 1967; Chiang, 1970; De Smedt et al., 2001). In Halymenia, the medulla is lax in
young parts with mainly anticlinal filaments and becoming denser and irregular in older
parts (Balakrishnan, 1961; Abbott, 1967; Womersley and Lewis, 1994). Cruciately
divided tetrasporangia embedded in the outer cortex, scattered over the blade surface
while spermatangia are borne in whitish sori at the cortical layer surface (Norris, 2014).
The carposporophyte is enclosed by a slight involucre derived from elongation and
Univers
ity of
Mala
ya

19
expansion of the ampullary filaments (Womersley and Lewis, 1994; Norris, 2014). The
carpogonial branches are two-celled and the carpogonial branch ampullae in Halymenia
are reported to be branched to the third and rarely fourth orders (Kawaguchi et al.,
2004). Chiang (1970) used the architecture of auxiliary cell ampullae as a primary
feature to group species at the generic level in the Halymeniaceae. According to
Chiang’s generic concept, simple or once or more branched secondary ampullar
filaments may emerge from long and slender primary ampullary filaments in the
Halymenia-type auxiliary cell ampullae. The auxiliary cell ampulla of Halymenia is
flattish, expanded when mature, and is intermediate between the Grateloupia type and
the Cryptonemia-type of ampulla based on its shape and the degree of branching
(Chiang, 1970). For instance, Hernández-Kantún et al. (2009) confirmed the assignment
of specimens from the Gulf of California (Halymenia actinophysa M. Howe) to the
genus Halymenia through the combination of female reproductive structures and tertiary
branching of auxiliary cell ampullae.
According to Abbott (1967), a vegetative feature- the anticlinally oriented filaments
has been considered more diagnostic than reproductive characters that seem to overlap
considerably among genera of the family (Kraft, 1977; Maggs and Guiry, 1982).
However, anticlinal medullary filaments are not exclusive to Halymenia and can be
found in other genera such as Cryptonemia and Kallymenia J. Agardh (Abbott, 1967;
Guimarães and Fujii, 1998). Additionally, stellate cells and refractive ganglionic cells
also present in the genera Weeksia Setchell and Kallymenia which are placed under
order Gigartinales (Abbott, 1967). Therefore, Halymenia should not be characterized by
a single feature. This has been supported by Kawaguchi and Lewmanomont (1999)
which stated that “no single feature most distinctively characterizes Halymenia”. A
combination of features is important in the characterization of Halymenia.
Univers
ity of
Mala
ya

20
To date, seven species of Halymenia have been reported from Malaysia, including H.
floresii (Clemente) C. Agardh, H. durvillei Bory de Saint-Vincent, H. dilatata Zanardini,
H. maculata J. Agardh, H. formosa Harvey ex Kützingand and two recently described
species from the current study- H. malaysiana P-L Tan, P-E Lim, S-M Lin & S-M
Phang and H. johorensis P-L Tan, P-E Lim, S-M Lin & S-M Phang (Kawaguchi et al.,
2002; Tan et al., 2015; Phang et al., 2016; Tan et al., 2017). In Thailand, H. durvillei, H.
dilatata and H. maculata are the only three Halymenia species have been recorded
(Lewmanomont and Kawaguchi, 2002; Tsutsui et al., 2012). On the other hand, a total
of 14 taxa and 22 taxa of Halymenia (including synonym) have been recorded in the
Philippines and Indonesia respectively (Silva et al., 1987; Verheij and Prud'homme van
Reine, 1993; Kraft et al., 1999; De Smedt et al. 2001; Atmadja and Prud'homme van
Reine, 2012). Most of the records here were from checklists and without detailed
morphological description. Thus, many taxa remain poorly known due to the scarce
information available.
Southeast Asia is well known to be a biodiversity hotspot, with many organisms yet
to be identified (Sodhi et al., 2004). Yet, there are relatively few studies of Halymenia in
this region. Several attempts have been made to study species of Halymenia in
Southeast Asia based solely on morphological characters. Kawaguchi and
Lewmanomont (1999) made a detailed morphological study of Halymenia dilatata
Zanardini by comparing the vegetative and reproductive features of the material from
Vietnam and Japan with Indian material, and by studying the pattern of spore
development to establish a better classification system for the western Pacific species.
The results showed that vegetative and reproductive features of of H. dilatata were in
accordance with the original and complementary descriptions by Zanardini (1851, 1858).
The carpospores development of H. dilatata was also similar to H. floresii from the
Mediterranean Sea (van den Hoek and Cortel-Breeman, 1970) and H. latifolia P.
Univers
ity of
Mala
ya

21
Crouan & H. Crouan ex Kützing from Ireland (Maggs and Guiry, 1982). According to
De Smedt et al. (2001), Halymenia specimens from the Philippines were examined by
studying their vegetative and reproductive morphology and four species were
recognized: H. durvillei, H. dilatata, H. maculata, and H. porphyraeformis Parkinson.
De Smedt et al. (2001) also reduced all varieties and formas within H. durvillei as
proposed by Weber-van Bosse (1921) to synonyms of H. durvillei since the minor
differences in gross thallus morphology and branching pattern observed were not
sufficient to warrant recognition at the species level. In the following year,
Lewmanomont and Kawaguchi (2002) compared the morphological and anatomical
structures of both H. dilatata and H. maculata from Thailand. These two species can be
distinguished from each other based on the texture of fresh plants, the margins, the
thickness of thallus and cortex, the number of cell layers in the cortex and the shape of
the cells in the outermost cortex layer. Kawaguchi et al. (2002b) studied the
morphology of a foliose red alga from Vietnam and revealed that it belongs in H.
maculata and is distinct from H. stipitata. The presence of three species of Halymenia,
H. durvillei, H. dilatata and H. maculata in Malaysia was confirmed by Kawaguchi et al.
(2002a) based on their gross morphology and anatomical features. This was also the
first time of describing the reproductive anatomy of H. durvillei including the
Halymenia-type auxiliary cell ampullae in detail. In 2004, Kawaguchi also verified the
presence of H. floresii in Malaysia by comparing the Malaysian material with the
lectotype and other authentic material of H. floresii.
Collins and Howe (1916) and Taylor (1960) had earlier described Halymenia species
separation based on the fronds dimension, branching pattern, thickness, degree of
cystocarp protrusion and presence or absence of ganglioid cells. According to Abbott
(1967), species delineation in Halymenia is based on habit, color, number of cortical
cell layers and quantity of medullary filaments. In addition, Gargiulo et al. (1986)
Univers
ity of
Mala
ya

22
recognized a new species Halymenia asymmetrica Gargiulo, de Masi & Tripodi in the
Mediterranean Sea by comparing following characters with other known species: (1)
habit, (2) branch pattern, (3) the presence or absence of marginal proliferations or
papillae on thallus surface, (4) dimensions of the thallus, with particular regard to blade
width and (5) presence or absence of secretory cells. Five diagnostic features have been
used by Hernández-Kantún et al. (2012) to identify four Halymenia species, including
order of branching, spines on the thallus surface, shape of the cells in the inner cortex,
thickness of cortex and stipe size. A number of morphological studies in Halymenia
have highlighted several features useful in delineating species. These include habit,
thallus size, blade margin, order of branching, presence or absence of a midrib in the
basal region, presence or absence of a stipe, presence or absence of marginal
proliferations, presence or absence of papillae or spines on thallus surface, blade
thickness, cortex thickness, shape of inner cortical cells, inner cortical cell size, and
presence or absence of refractive ganglionic cells (Gargiulo et al., 1986; Guimarães and
Fujii, 1998; De Smedt et al., 2001; Ballantine and Ruiz, 2004; Hernández-Kantún et al.,
2012; Tan et al., 2015; Azevedo et al., 2016a; 2016b; Tan et al., 2017). For example,
Guimarães and Fujii (1998) differentiated Halymenia brasiliana S.M.P.B.Guimarães &
M.T.Fujii from other Brazilian Halymenia species by its absence of a rib at the base of
the thallus and its absence of refractive ganglionic cells in the medulla. Furthermore,
De Smedt et al. (2001) initiated the use of stipe anatomy in Halymenia and proposed
that it may be useful in distinguishing species of Halymenia. In contrast, Guimarães
and Fujii (1998) indicated that the degree of cystocarp protrusion, colour, the diameter
and the number of medullary filaments are highly variable features, and thus not useful
in delineating Halymenia species. Additionally, the taxonomic significance of surface
maculation as a specific feature in Halymenia is controversial and needs to be verified
(Kawaguchi, 2002).
Univers
ity of
Mala
ya

23
Traditionally, the identification of Halymenia is based solely on morphological
characteristics, which is problematic due to its immense morphological plasticity and
few distinctive morphological features (Tan et al. 2015; 2017). In addition, comparative
studies of Halymenia species are disconcerted by variations in features used for species
delimitation (Hernández-Kantún et al., 2009). The lack of distinct morphological
characters has led to a need for molecular approach to address the taxonomic confusion
in these red algae. Recent molecular studies in concert with morphological examination
have led to the taxonomic revision of existing taxa and the discovery of new species
(Hernández-Kantún et al., 2012; Tan et al., 2015; Azevedo et al., 2016a; 2016b, Tan et
al., 2017).
2.4.1 Importance and economic potential of Halymenia
Carrageenans are sulphated cell wall polysaccharides found in Rhodophyta. They
have been greatly used in the food, cosmetics and pharmaceutical industries due to their
gelling, thickening and stabilizing properties (McHugh, 2003; Pereira et al., 2007). They
are useful for stabilizing and texturing products in the food industry. Additionally, their
strong antitumoral, immunomodulatory, anticoagulant, and antiviral activities (Campo
et al., 2009) make them useful in pharmaceutical and medical applications as excipients
and for controlled release of pharmaceutical compounds (Kranz et al., 2009). Nowadays,
carrageenan supplies have been mainly focused on Kappaphycus Doty and Eucheuma J.
Agardh (McHugh, 2003). However, the search for new or additional raw material
sources has been given an increased sense of urgency as worldwide demand and
development of new applications for carrageenan are increasing (Freile-Pelegrin et al.,
2011).
Univers
ity of
Mala
ya

24
As revealed by Kho et al. (2014), Halymenia is a promising carrageenan source. This
was supported by the studies of Kho et al. (2016) which shown that H. durvillei can be a
potential source for carrageenan production owing to its highest carrageenan yield
compared to another two Halymenia species (H. dilatata and H. maculata). In addition,
many studies have indicated the high carrageenan content in species of Halymenia
include Halymenia venusta Bøergesen (Semesi and Mshigeni, 1977; Parekh et al., 1987),
Halymenia porphyroides Børgesen (Parekh et al., 1989), Halymenia ceylanica Harvey
ex Kützing (Lai et al., 1994) and H. durvillei (Fenoradosoa et al., 2009). Thus, these
indicated that Halymenia is a potential source for carrageenan production which
generates lucrative returns to the industry and economy.
Besides, Halymenia is also a potential food source for human and animal. For
instance, Halymenia floresii is an edible species consumed in some Asian markets
(Godínez-Ortega et al., 2007). Garcia et al. (2016) highlighted the nutritional potential
of H. floresii as food, either as fresh produce or as a processed food ingredient. Three
species of Halymenia (H. durvillei, H. maculata and H. dilatata) have also the potential
to be used as raw material or ingredients in human diet and animal feed as reported by
Kho et al. (2016). Halymenia is also desired for its pigments. H. durvillei is a source of
the red pigment R-phycoerythrin which used as a food and cosmetic colorant, a
therapeutic agent owing to its immunomodulating and anti-cancer activity, and a
fluorescent agent (Bermejo Román et al., 2002; Spolaore, 2006). Halymenia floresii has
also been proved to be a good source for the extraction and preparation of R-
phycoerythrins (Malairaj et al., 2016). The lutein content of H. floresii may be of
particular interest for the market of edible seaweeds (Godínez-Ortega et al., 2007).
Apart from these, Halymenia can be used as a biofilter in an integrated aquaculture
system. Halymenia microcarpa (Montagne) P. C Silva was employed and proven to be
Univers
ity of
Mala
ya

25
a useful biofilter for the nutrient removal in a lobster-seaweed integrated aquarium
system (Chen and Chen, 1996).
2.5 Genetic diversity of seaweeds
Genetic diversity is the genetic variability within a populations or a species. It can be
referred to any variation either in its most primary level of nucleotides, genes,
chromosomes, or whole genomes of an individual (Wright, 1920; Fisher, 1930). Genetic
diversity assessment is important for a better understanding of the nature of forces
acting on genetic variation, pattern, and level of genetic variation, evolutionary history
and adaptation of an organism (Yow et al., 2011). According to Hughes et al. (2008),
genetic diversity within a population also has ecological effects on productivity, growth
and sustainability, as well as inter-specific interactions within communities and
ecosystem level processes.
Genetic variation holds the key to the ability of populations and species to persist
over evolutionary time through changing environments (Freeman and Herron, 1998). In
general, individuals in small populations are less able to adapt themselves to diverse
environmental conditions as they are probably to be homogenous in terms of genetic,
anatomy and physiology (Bagley et al., 2002). In contrast, larger populations are more
likely to have greater allele diversity and also the greater capacity for evolutionary
adaptation to survive in changing environments. As reported by Frankham et al. (2004),
loss of genetic diversity may diminish evolutionary potential and reproductive fitness of
a population to endurance in stressful environments.
Natural evolutionary forces for example mutation, natural selection, migration and
genetic drift may induce changes in the allele frequencies of populations (Valero et al.,
Univers
ity of
Mala
ya

26
2001). In addition, anthropogenic activities such as fisheries and aquaculture, global
climate change, land-use change and water pollution have threatened biodiversity as
well as genetic diversity in marine organisms in particular the seaweeds. Development
of islands and coastal areas into resorts, increase marine traffic which add oil to the
waters and untreated discharges from industries are also some of the human activities
which cause losses in seaweed resources (Phang et al., 2006). Introduction of non-
indigenous species associated with shipping vectors (eg. ballastwater and fouling of
vessel hulls), aquaculture and the aquarium trade have also impacted diversity of the
seaweed genetic resources and marine ecosystem (Schaffelke et al., 2006). A number of
studies have shown the dispersal of seaweed species across their native ranges owing to
anthropogenic activities (Rueness, 1989; Curiel et al., 1998; Fletcher and Farell, 1999;
Rueness and Rueness, 2000; Boudouresque and Verlaque, 2002; Smith et al., 2002;
Hwang et al., 2004).
Assessment of genetic diversity of seaweed with molecular tools has been
accelerated with advanced in DNA based molecular marker technologies. According to
Féral (2002), the utilization of genetic markers for genetic variation studies provide
valuable information for gene flow, population structure, phylogenetic relationships,
biogeographic studies, and parentage and relatedness analysis.
2.6 Molecular phylogenetic methods
Phylogenetic studies have vast applications in diverse fields, including ecology,
molecular biology, and physiology (Doyle et al., 2003). The fundamental importance of
phylogenetic studies is to provide insights into organismal relationships and evolution.
Phylogenetic trees outlining the evolutionary history of species can be derived from
nucleic acid or protein sequences from those species. Phylogenetic trees facilitate the
Univers
ity of
Mala
ya

27
interpretation of the evolution of diverse characters (molecular, physiological and
genetic). For instance, they are useful in the investigations of biosynthetic and
developmental pathways, natural products chemistry, origins and migrations of
evolutionary lineages, and conservation (Daly et al., 2001).
Phylogenetic inference can be defined as the process of determining the estimated
evolutionary history by analysis of a given data set (Swofford, 1996). The phylogenetic
inference methods are either distance based or character based. The distance based
methods involve the calculation of pairwise distance of all studied sequences and then
the resulting distance matrix is used for tree reconstruction (Yang and Rannala, 2012).
The most widely used distance-based method is neighbour joining (NJ). While the
character based method derives trees that optimize the distribution of the actual data
pattern for each character (Roy et al., 2014). The most commonly used character based
methods include Maximum Parsimony (MP), Maximum Likelihood (ML) and Bayesian
Inference (BI) methods. These methods simultaneously compare all sequences in the
alignment, considering one character (a site in the alignment) at a time to calculate a
score for each tree. The ‘tree score’ is the minimum number of changes for maximum
parsimony, the log-likelihood value for maximum likelihood and the posterior
probability for Bayesian inference (Yang and Rannala, 2012). Generally, the distance
based methods tend to be much faster than character based methods but they typically
yield little information beyond the basic tree structure (Dowell, 2008).
On the other hand, the phylogenetic inference methods can also be divided to model-
based or non-model based depend on whether the trees are inferred with or without
using evolutionary model. Model-based methods incorporate branch-length information,
explicit models of character evolution, and quantifying uncertainty in ancestral-state
estimates therefore they are generally preferable to non-model based methods as they
Univers
ity of
Mala
ya

28
are less likely to provide misleading results. NJ, ML and BI are model-based whereas
MP does not have an explicit model and its assumptions are implicit (Yang and Rannala,
2012). Additionally, an optimality criterion which is the score used to assess the value
of a particular tree, needs to be defined before using an algorithm to compute the value
of this function for various trees and search for the best tree that maximizes or
minimizes the criterion. For instance, the tree with the smallest sum of branch lengths is
the preferred tree in NJ. Likewise, the most optimal tree in ML is the one with the
highest likelihood score while the tree that minimizes the number of character-state
changes is the preferred MP tree.
Four criteria have been used to judge phylogenetic inference methods: (1)
consistency, (2) efficiency, (3) robustness and (4) computational speed (Yang and
Rannala, 2012). Soltis and Soltis (2003; Table II, page 1797) and Yang and Rannala
(2012; Table 2, page 309) summarised some of the strengths and weaknesses of
phylogenetic inference methods and is reproduced here with modification as Table 2.1.
Univers
ity of
Mala
ya
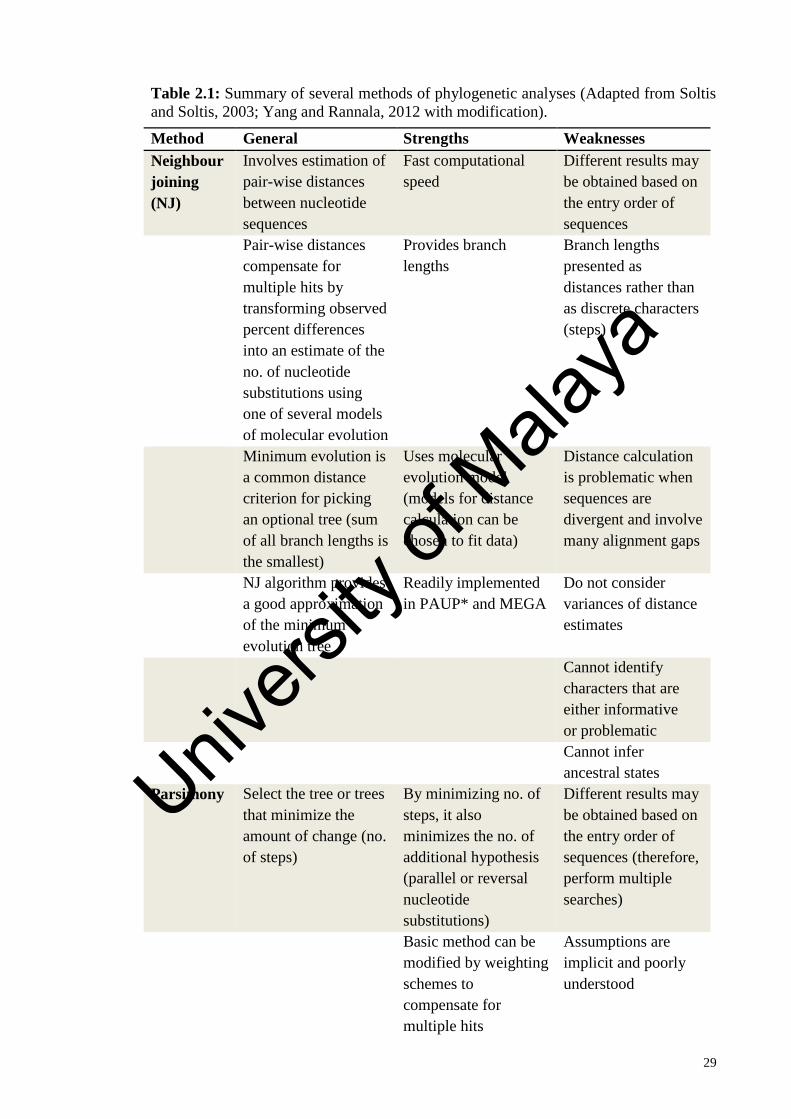
29
Table 2.1: Summary of several methods of phylogenetic analyses (Adapted from Soltis
and Soltis, 2003; Yang and Rannala, 2012 with modification).
Method General Strengths Weaknesses
Neighbour
joining
(NJ)
Involves estimation of
pair-wise distances
between nucleotide
sequences
Fast computational
speed
Different results may
be obtained based on
the entry order of
sequences
Pair-wise distances
compensate for
multiple hits by
transforming observed
percent differences
into an estimate of the
no. of nucleotide
substitutions using
one of several models
of molecular evolution
Provides branch
lengths
Branch lengths
presented as
distances rather than
as discrete characters
(steps)
Minimum evolution is
a common distance
criterion for picking
an optional tree (sum
of all branch lengths is
the smallest)
Uses molecular
evolution model
(models for distance
calculation can be
chosen to fit data)
Distance calculation
is problematic when
sequences are
divergent and involve
many alignment gaps
NJ algorithm provides
a good approximation
of the minimum
evolution tree
Readily implemented
in PAUP* and MEGA
Do not consider
variances of distance
estimates
Cannot identify
characters that are
either informative
or problematic
Cannot infer
ancestral states
Parsimony Select the tree or trees
that minimize the
amount of change (no.
of steps)
By minimizing no. of
steps, it also
minimizes the no. of
additional hypothesis
(parallel or reversal
nucleotide
substitutions)
Different results may
be obtained based on
the entry order of
sequences (therefore,
perform multiple
searches)
Basic method can be
modified by weighting
schemes to
compensate for
multiple hits
Assumptions are
implicit and poorly
understood
Univers
ity of
Mala
ya

30
Table 2.1, continued
Method General Strengths Weaknesses
Parsimony
Searches identify
numerous equally
parsimonious
(shortest) trees; treats
multiple hits as an
inevitable source of
false similarity
(homoplasy)
Relatively slow
(compared with NJ)
with large data sets
Readily implemented
in PAUP*
Lack of a model
makes it nearly
impossible to
incorporate our
knowledge of
sequence evolution
Can identify
individual characters
that are
informative or
problematic
Branch lengths are
substantially
underestimated when
substitution rates are
high
Can infer ancestral
states
Highly unequal rates
of base substitution
may cause
difficulties (e.g. long
branch attraction
Maximum
likelihood
(ML)
Involves estimating
the likelihood of
observing a set of
aligned sequences
given a model of
nucleotide substitution
and a tree
A statistical test (the
likelihood ratio test)
can be used to
evaluate properties of
trees
Computationally very
intensive (much
slower than other
methods
Flexible, models that
can incorporate
parameters of base
frequencies,
substitution rates, and
variation in
substitution rates
and, therefore, are
“general”
Practical with only
small nos. (fewer
than 50) of sequences
Univers
ity of
Mala
ya

31
Table 2.1, continued
Method General Strengths Weaknesses
Maximum
likelihood
(ML)
Uses all of the data
(invariable sites and
unique mutations are
still informative,
unlike parsimony
analysis
Easily implemented in
PAUP*
Nucleotide
substitution models
are used directly
in the estimation
process, rather than
indirectly (as in
parsimony)
Bayesian Uses a likelihood
function and an
efficient search
strategy
Based on the
likelihood function,
from which it inherits
many of its favorable
statistical properties
Markov chain Monte
Carlo (MCMC)
involves heavy
computation
Based on a quality
called the posterior
probability of a tree
Uses models as in ML In large data sets,
MCMC convergence
and mixing problems
can be hard to
identify or rectify
Researcher may
specify belief in a
prior hypothesis prior
to analysis
Prior probability
allows the
incorporation of
information or expert
knowledge
Uninformative prior
probabilities may be
difficult to specify.
Multidimensional
priors may have
undue influence on
the posterior without
the investigator’s
knowledge
Posterior probability
for trees and clades
have ease
interpretations
Posterior
probabilities
(measure of internal
support) can be
overestimates
Can be used to
analyze relatively
large data sets
Model selection
involves challenging
computation
Univers
ity of
Mala
ya

32
2.7 Molecular approaches for taxonomic inference
The adoption of molecular techniques since the 1980s has improved the
understanding of many species and overcame the problem in species identification in
red algal taxonomy (Robba et al., 2006; Maggs et al., 2007). Due to the limitations of
morphological features in identification, molecular approaches in species identification
have come to rely heavily by algal systematists for close to two decades (Harper and
Saunders, 2001). The application of molecular data such as DNA, RNA and protein
allows investigation of the phylogenetic relationship and evolutionary patterns of
diversity between organisms. Some of the molecular techniques employed in the
taxonomic inference and genetic diversity studies of red algae, include random
amplified polymorphic DNA (RADP), restriction fragment length polymorphism
(RFLP), amplified fragment length polymorphism (AFLP) and nucleic acid sequencing.
The development of polymerase chain reaction (PCR) in the late 1980s has
facilitated the development of molecular technologies and revolutionized many aspects
of current research. It is widely used in applications such as gene expression analysis,
sequencing, cloning and mutagenesis. PCR is used to amplify selected sections of DNA
using DNA polymerase to synthesize new strand of DNA complementary to the offered
template strand by adding a nucleotide only onto a preexisting 3'-OH group. At the end
of the PCR reaction, the specific sequence will be amplified in numbers of billions.
Generally, there are three steps involved in PCR, namely denaturation, annealing and
extension. During denaturation, double stranded DNA (template DNA) is denatured by
heat, resulting in two single stranded, complementary DNA. During the annealing step,
each of the two DNA strands act as the templates for the synthesis of new strands of
DNA at a temperature optimal for primers to bind to specific site of the template DNA.
Subsequently, DNA polymerases synthesize new DNA strands by adding nucleotides
Univers
ity of
Mala
ya

33
complementary to that of the template DNA in a 5' to 3' direction during extension step.
These denaturation, annealing and elongation steps (one cycle) are repeated for 25-40
times so as to achieve large quantities of the DNA of interest.
PCR method has had a significant impact on molecular taxonomy because of its
simplicity and rapidness. It has great sensitivity and is capable of amplifying sequences
from minute amounts of DNA. Owing to the huge advantages it offers, PCR has become
an indispensable technique with a wide range of applications, particularly in forensic
science, diagnosis, genetic linkage analysis and molecular paleontology studies.
2.7.1 Random Amplified Polymorphic DNA (RAPD)
Random Amplified Polymorphic DNA (RAPD) is a PCR-based technique which
involves the use of short random primers (8-12 nucleotides) resulting in the
amplification of many discrete DNA that are separated by gel electrophoresis (Williams
et al., 1990; Kumari and Thakur, 2014). The significant advantage of RAPD is it does
not require prior knowledge of the target genome (Bardakci, 2001). Additionally, RAPD
provides a rapid and efficient screen for DNA sequence based polymorphism at a very
high number of loci (Kumari and Thakur, 2014). Moreover, only small amount of DNA
is needed and no isotope labelling is required for sample detection (Williams et al.,
1990). This technique has been used in the taxonomy and classification of seaweeds at
the genus and species level. For instance, RAPD was used for differentiating
morphologically similar algal taxa (Lim et al., 2001), analyzing genetic diversity and
determining intraspecific relationships (Patwary et al., 1993; van Oppen et al., 1996;
Alberto et al., 1997; Wright et al., 2000) and assessing patterns of geographic dispersal
of populations (Alberto et al., 1999).
Univers
ity of
Mala
ya

34
2.7.2 Restriction Fragment Length Polymorphism (RFLP)
Restriction fragment length polymorphism (RFLP) is a technique that exploits
variations in homologous DNA sequences to genetically differentiate between
individuals. In RFLP, the DNA is digested by specific restriction enzymes. The
resulting restriction fragments are separated according to their lengths by gel
electrophoresis and then transferred to a membrane via the Southern blot procedure.
Hybridization of the membrane to a labeled DNA probe then determines the length of
the fragments which are complementary to the probe (Singh, 2012). The length of a
detected fragment varies between individuals. Therefore, RFLP is useful in
investigating genetic diversity within and between species (Pierce et al., 2000). This
technique has been widely used in the studies of phylogenetic relationships of algae. For
example, it was used to differentiate morphologically similar taxa and identify
geographically distant conspecific taxa (Bird and Rice, 1990; Candia et al., 1999;
Guillemin et al., 2008). However, RFLP techniques require lengthy technical procedure,
laborious and time-consuming.
2.7.3 Amplified Fragment Length Polymorphism (AFLP)
Amplified Fragment Length Polymorphism (AFLP) is a technique based on the
selective PCR amplification of restriction fragments from a total digest of genomic
DNA (Vos et al., 1995). In AFLP, the genomic DNA is digested with restriction
enzymes followed by the ligation of adaptors to the restriction fragments. Next,
selective amplification of a subset of the restriction fragments can be implemented by
using primers containing defined common sequences with one to three arbitrary
nucleotides. The amplified restriction fragments are separated by gel electrophoresis
then detected by using radioactively or fluorescent labeling of PCR primer. AFLP is a
Univers
ity of
Mala
ya

35
method which combines the advantages of both RAPD and RFLP. It is a highly
sensitive method for detecting polymorphisms in DNA, requires no prior sequence
information, and has high reproducibility (Féral, 2002; Meudt and Clarke, 2007; Yi et
al., 2010). This technique has been used in species identification and genetic diversity
studies of algae (Zhang et al., 2009; Pang et al., 2010; Shan et al., 2011; Li et al., 2013).
2.7.4 Nucleic acid sequencing
Nucleic acid sequencing is also one of the important techniques in the field of
molecular biology because the precise order of the nucleotide bases in a molecule of
DNA can be determined (Brown, 2001). The advent of DNA sequencing has
significantly advanced biological research and discovery. Since the sequence of the
nucleotides in a segment of DNA carries the genetic information, the nucleotide
sequence themselves can serve as valuable molecular tools in phylogenetic and
population genetic studies. Analyzing DNA sequences is useful in identifying
taxonomic groups, reconstructing phylogenies as well as evaluating the evolution of
genomes.
Nucleic acid sequencing offers several advantages of being rapid, time-saving, less
laborious and able to provide more informative data with large numbers of
independently evolving characters which can be used to reconstruct phylogenies if
compared to the much smaller number of biochemical, morphology and other characters.
Various nucleic acid regions have been extensively used in algal taxonomic studies, for
example rbcL (Freshwater and Rueness, 1994; Nam et al., 2000; McIvor et al., 2001;
Wang et al., 2001; Gavio and Fredericq, 2002; Kawaguchi et al., 2002; De Clerk et al.,
2005; Russell et al., 2009; Yang et al., 2013b; Kim et al., 2014), cox1 (Geraldino et al.,
2006; Sherwood, 2008; Yang et al., 2008; Yow et al., 2011; Tan et al., 2012; Lim et al.,
Univers
ity of
Mala
ya

36
2013; Yow et al., 2013), cox3 (Coyer et al., 2004; Uwai et al., 2006b; Kim et al., 2014),
cox2-3 spacer (Zuccarello et al., 1999; Zuccarello and West, 2002; Zuccarello et al.,
2006; Andreakis et al., 2007; Vis et al., 2008; Teasdale & Klein, 2010; Tan et al., 2012;
Lim et al., 2013; Tan et al., 2013), RuBisCO spacer (Destombe and Douglas, 1991;
Goff et al., 1994, Tan et al., 2013) and ITS rDNA (Bellorin et al., 2002; Broom et al.,
2002; Marston and Villard-Bohnsack, 2002; Cho et al., 2007; Hu et al., 2009; Moniz
and Kaczmarska, 2010; Russell et al., 2009; Kim et al., 2014). It is also enable
comparison of results from one laboratory to one another which is impossible for
molecular techniques such as RAPD, RFLP and AFLP owing to the lack of
methodology standardization. Therefore, the phylogenetic relationship of any species
can be studied by comparing the respective sequence data from GenBank with data
from any current study.
2.8 DNA barcoding
DNA barcoding is referring to DNA sequence analysis of a short, diagnostic segment
(specific region of a genome) to identify species through reference to DNA sequence
libraries or databases (Hebert et al., 2003). It is a useful tool, especially when coupled
with traditional taxonomic tools and is fundamental in revealing hidden diversity
(Hebert et al., 2004a). Hebert (2005) also denoted that, DNA barcoding will hasten the
pace of species discovery by allowing taxonomists to speedily sort specimens and by
highlighting divergent taxa that may represent new species. There are a lot of benefits
derived from barcoding such as enabling species identification from any stages of life or
fragment, facilitating species discoveries based on cluster analyses of gene sequences
and providing insight into the diversity of life (Savolainen et al., 2005). Hence, DNA
Univers
ity of
Mala
ya

37
barcoding is potential to be used as a practical tool in identifying all the eukaryotic
organisms on earth.
As revealed by Saunders and Kucera (2010), DNA barcoding eliminates the reliance
on morphological characters which mostly used for species discrimination. This is
especially important for algal taxonomic studies since algae are well known for their
simple morphologies and phenotypic plasticity. Additionally, different molecular
markers have been used for algal species identification and there is a lack of
standardized marker for rapid specimen identification. The use of DNA barcode which
act as standard marker will facilitate universal comparisons (Saunders, 2005). Thus,
Saunders commenced the use of DNA barcode for red algal species identification in
2005.
The 5’ end of the cytochrome c oxidase subunit 1 gene (COI-5P) was first proposed
by Herbert et al. (2003) as DNA barcode. It has been successfully used across diverse
animal taxa (Herbert et al., 2004a; b), as well as red (Saunders, 2005; Robba et al., 2006)
and brown algae (Kucera and Saunders, 2008). In general, plastid and mitochondrial-
encoded molecular markers were frequently proposed as good DNA barcodes for red
algae (Saunders, 2005; Robba et al., 2006; Saunders, 2008; 2009; Le Gall and Saunders,
2010; Rueness, 2010; Zuccarello and West, 2011). Previous studies have employed
DNA barcoding for uncovering cryptic red algal species and inferring phylogenetic
affinities of red algae (Clarkston and Saunders, 2010; Clayden and Saunders, 2010; Le
Gall and Saunders, 2010; Saunders and McDonald, 2010; Milstein and Saunders, 2012;
Hind and Saunders, 2013; Saunders and McDevit, 2013). DNA barcoding has also been
utilized in biodiversity surveys and biomonitoring efforts (Conklin et al., 2009; Rueness,
2010; Sherwood et al., 2010a; Carlile and Sherwood, 2013).
Univers
ity of
Mala
ya

38
2.9 Molecular marker for phylogenetic inference
The use of DNA sequences as a tool in numerous fields has been widely discussed in
the past years. It is important to select an appropriate marker before implementing a
DNA taxonomy or DNA barcoding system (Sonnenberg et al., 2007). The selection of
marker is greatly influenced by the taxonomic question to be addressed. According to
Patwardhan et al. (2014), an ideal marker should possess characteristics as follows: (1) a
single-copy gene may be more useful than multiple-copy gene; (2) prior to phylogenetic
analysis, the alignment of marker gene sequences should be easy; (3) the substitution
rate should be optimum so as to provide enough information sites; (4) primers should be
available to selectively amplify the marker gene; and (5) too much of base variation
among the taxa may not reflect the true ancestry thus is not preferable. Some of the
widely used markers in phylogenetic studies are described below in detail.
2.9.1 Nuclear markers
Nuclear encoded ribosomal DNA (rDNA) has been widely used in the taxonomy of
algae. It is present as head-to-tail tandem repeats in eukaryotes and encoded for rRNA
gene. The rDNA contains a transcribed region that gives rise to a pre-rRNA transcript
and an intergenic region which known as the non-transcribed spacer (NTS) (Leitch et al.,
2013). The pre-rRNA transcript includes the small subunit (SSU, 18S rRNA), the first
internal transcribed spacer (ITS1), the 5.8S rRNA, the second internal transcribed
spacer (ITS2) and the large subunit (LSU, 28S rRNA) (Leitch et al., 2013).
Nuclear encoded ribosomal DNA (rDNA) is good for studying phylogenetic
relationship because it is universal and is composed of highly conserved as well as
variable domains (Patwardhan et al., 2014). In addition, it is evolved more slowly than
Univers
ity of
Mala
ya

39
protein encoding genes rendering them valuable for phylogenetic inference studies at
the higher taxonomic levels. For instance, the SSU (18S rDNA) has been found not able
to differentiate closely related species of Gracilaria but useful in the study of the
generic relationship within the Gracilariaceae (Bellorin et al., 2002). The SSU
sequences have also proven useful for familial and ordinal level taxonomy for red algae
(Bird et al., 1990; Ragan et al., 1994; Saunders and Kraft, 1996).
The LSU (28S rDNA) has proved to be useful in the studies of higher-level
relationships of red algae as reported by Freshwater et al. (1999). It also provided higher
number of phylogenetically informative sites and more divergent than SSU thus may
resolve phylogenetic problems where SSU (18S rDNA) sequences are uninformative
(Freshwater et al., 1999). The utility of partial LSU (28S rDNA) sequence as DNA
barcode was assessed by Sherwood et al. (2010a; 2010b) in Hawaiian Rhodophyta
Biodiversity Survey and confirmed not useful for rapid identification in future red algal
biodiversity surveys due to its lack of a reliable species-level resolution.
The non-coding internal transcribed spacers (ITS) have been adopted in studies of
species identification, genetic diversity and biogeographical in red algae (Broom et al.,
2002; Marston and Villalard-Bohnsack, 2002; Hu et al., 2009; Russell et al., 2009; Kim
et al., 2014). In general, ITS regions are useful for molecular investigations of closely
related taxa but inappropriate for higher lever phylogenetic comparison. This is because
ITS regions are highly conserved intraspecifically, but variable between different
species (Bruns et al., 1991; Hillis and Dixon, 1991). Additionally, ITS regions evolve at
a more rapid rate than other conserved region of rDNA (Baldwin et al., 1995; Harper
and Saunders, 2001). According to Hu et al. (2009), ITS marker is also a potential DNA
barcode for the species identification of Rhodophyta. Even so, this marker has several
drawbacks. Extensive sequence variation, length polymorphism of ITS could be
Univers
ity of
Mala
ya

40
accounted for PCR amplification difficulty (Bown et al., 2003). Furthermore, the
common occurrence of mononucleotide runs and heterogeneity in the multiple copies of
the ITS within an individual made it difficult to acquire clean sequence from both
strands across the entire ITS region (Saunders, 2005). Excessive indels in ITS alignment
can also resulted in difficulty in sequence comparison between taxa especially distantly
related species. The 5.8S rRNA gene was included in phylogenetic studies of various
red algal lineages (Goff et al., 1994; Patwary et al., 1998; Bellorin et al., 2002;
Guillemin et al., 2008; Hu et al., 2009; Hind and Saunders, 2013).
Elongation factor 2 (EF2), which is a nuclear protein coding gene, was also useful
for assessing the relationships between eukaryotic lineages (Hashimo et al., 1997). For
example, the sister relationship between green plants and red algae was supported by
using EF2 (Moreira et al., 2000). In addition to resolving supraordinal relationships,
EF2 has been shown to be useful in improving the phylogenetic inference within
Florideophyceae by resolving some novel ordinal affinities within the Nemaliophycidae
and Rhodymeniophycidae (Le Gall and Saunders, 2007).
2.9.2 Plastid markers
Ribulose-l, 5-bisphosphate carboxylase/oxygenase (RuBisCO) is the enzyme that
facilitates the primary carbon dioxide fixation step in the photosynthesis (Freshwater
and Rueness, 1994). The RuBisCO cistron consists of a large subunit (rbcL) and a small
subunit (rbcS) separated by an intervening spacer and is plastid-encoded (Valentin and
Zetsche, 1989). The rbcL marker has been proved to be very powerful for taxonomic
research of red algae because it has good resolution at the species level as well as higher
taxonomic levels in the red algae (Hommersand and Fredericq, 2003). The rbcL has
been widely used in Halymeniaceae to clarify taxonomic status (Wang et al., 2001;
Univers
ity of
Mala
ya

41
Kawaguchi et al., 2002; De Clerk et al., 2005; Russell et al., 2009), to clarify taxonomic
distribution (Kim et al., 2014; Lee et al., 2016) and to delimit species (Kawaguchi et al.,
2013; Tan et al., 2015; Azevedo et al., 2016a). Its large size which provides many
characters for phylogenetic analysis, availability of conserved primers that permit rapid
amplification and sequencing are some of the advantageous of the utility of rbcL
(Freshwater and Rueness, 1994). Moreover, the absence of insertion or deletion
mutations in rbcL eliminates the problems of alignment. The large number of published
sequences available in GenBank also made it a useful marker for species identification
(Costa et al., 2012). On the other hand, the RuBisCO spacer had been used to resolve
the phylogenetic relationships between different red algal genera but insufficient for
differentiating closely related species (Destombe and Douglas, 1991; Goff et al., 1994).
Universal plastid amplicon (UPA), which is in domain V of the 23S plastid rRNA
gene, has been proposed as a DNA barcode for algae lineages (Presting, 2006;
Sherwood and Presting, 2007). Primer universality and the ease of data acquisition with
a single primer set uphold its potential as a DNA barcode for red algae (Sherwood and
Presting, 2007; Sherwood et al., 2010a; 2010b). Several studies have shown that UPA
can discriminate among species within a single genus (Sherwood et al., 2008; Clarkston
and Saunders, 2010). However, this marker is not suitable for species identification in
some groups (e.g. plants by Newmaster et al., 2008; Bangiales by Kucera and Saunders,
2012).
2.9.3 Mitochondrial markers
Mitochondrial markers have been proved useful in addressing the phylogenetic
issues and population genetic within animals owing to their haploid nature, rapid
evolution, uniparental inheritance and lack of recombination (Avise, 1994; Zuccarello et
Univers
ity of
Mala
ya

42
al., 1999). The success of the utility of mitochondrial marker cytochrome c oxidase
subunit 1 (cox1 or COI) in animals has led to the assessment of this marker for
application in DNA barcoding in red algae (Saunders, 2005).
The mitochondrial cytochrome c oxidase subunit 1 (COI) gene encoded for an
enzyme involved in the last step of electron transport chain. COI gene has been used for
phylogenetic inference in red algae (Geraldino et al., 2006; Yang et al., 2008; Yow et al.,
2011), whereas the 5’ end of the cytochrome c oxidase subunit 1 gene (COI-5P) has
been recommended for DNA barcoding studies in red algae (Saunders, 2005; Robba et
al., 2006). A number of studies have shown the utility of COI-5P for the intra- and
interspecific studies, barcoding studies as well as the genetic diversity and
phylogeographical studies for the rhodophytes (Saunders, 2009; Clarkston and Saunders,
2010; Clayden and Saunders, 2010; Le Gall and Saunders, 2010; Saunders and
McDonald, 2010; Gulbransen et al., 2012; Milstein and Saunders, 2012; Carlile and
Sherwood, 2013; Hind and Saunders, 2013; Saunders and McDevit, 2013; Xie et al.,
2015).
The cox2-3 spacer is the first mitochondrial marker developed for red algal
intraspecific studies. It is an intergenic spacer, which is also a non-coding region,
separating the conserved cytochrome oxidase subunit 2 (cox2) and 3 (cox3) genes
(Zuccarello et al., 1999). Although the size of this marker is small (around 350 bp), it
demonstrates sufficient DNA sequence variability for population and phylogeographic
studies in red algae (Zuccarello et al., 1999). This marker has proven useful in intra- and
interspecific studies (Zuccarello et al., 1999; 2006; Tan et al., 2012), as well as the
genetic diversity and phylogeographical studies for the Rhodophytes (Zuccarello et al.,
2002; Andreakis et al., 2007; Vis et al., 2008). According to Zuccarello and West
(2011), the mitochondrial cox2-3 spacer demonstrated greater intraspecific variation
Univers
ity of
Mala
ya

43
than the plastid RuBisCO spacer, implying that the mitochondrion was evolving at a
rate that could make it more useful in phylogenetic and population studies. The cox2-3
spacer has also been proposed as the potential DNA barcode for coralline red algae
(Hernández-Kantún et al., 2014), as well as Kappaphycus and Eucheuma (Tan et al.,
2012, 2013).
Another mitochondrial marker, cytochrome c oxidase subunit 2 (cox2), was proposed
by Tan et al. (2012) as a promising maker for the DNA barcoding of Kappaphycus and
Eucheuma. However, its use for species identification has yet to be verified in other red
algae (Tan et al., 2012). On the other hand, Kim et al. (2014) indicated that cytochrome
c oxidase subunit 3 (cox3) gene can be a suitable tool for evaluating the phylogeography
and population structure of red algae.
2.10 Molecular studies in Halymenia
In spite of the large number of species discovered in Halymenia, many species
remain poorly studied and in need of review (De Smedt et al., 2001). The identification
of Halymenia based solely on morphological characters is difficult owing to its
extensive morphological plasticity and few distinctive morphological features (Tan et al.
2015; 2017). For example, Schneider et al. (2010) showed that two morphologically
similar species- H. pseudofloresii Collins & M. Howe and H. floresii were distinct
based on molecular evidence. Thus, the doubt has been cast on species recognition
based solely on morphology and the use of molecular identification is recommended for
species assignment (Hernández-Kantún et al., 2012). The adoption of molecular
techniques overcomes the problem related to morphology-based taxonomy and proved
useful in elucidating species boundaries and relationship and evolutionary history
Univers
ity of
Mala
ya

44
(Kawaguchi et al., 2006; Schneider et al., 2010; Hernández-Kantún et al., 2012; Tan et
al., 2015; Azevedo et al., 2016a; 2016b; Tan et al., 2017).
The molecular studies of Halymenia are very limited when compared to other genus
in Halymeniaceae, for instance Grateloupia. The genetic record of Halymenia in
GenBank is scanty with most of the taxonomically accepted species have been studied
based only on morphological characters. A total of seven studies attempt the use of both
molecular analyses and morphological examination to clarify the taxonomic position of
Halymenia. The first molecular study on Halymenia was carried out by Kawaguchi et al.
(2006). The rbcL marker was used in this study to clarify the range of morphological
variations of H. durvillei and its taxonomic relationships with the allied taxa from Indo-
Pacific. The result showed that regardless of their wide external variations, all the
branched samples were H. durvillei since they were enclosed in a monophyletic clade
with H. durvillei from Malaysia. The rbcL phylogeny also supported the identification
of two foliose plants by morphological observation: H. maculata and H. dilatata.
Subsequently, COI-5P and UPA were used as markers to evaluate intraspecific
divergence of H. pseudofloresii collections from Bermuda (Schneider et al., 2010).
Molecular evidence showed that all five collections with wide range of morphological
variation belong to a single species H. pseudofloresii since their UPA sequences were
identical while COI sequences were identical or varying at a single site. COI-5P was
also used to evaluate interspecific divergence between H. floresii and H. pseudofloresii
collections from Bermuda. The genetic variation between H. floresii and H.
pseudofloresii collections (approximately 7%) was sufficiently high to warrant
recognition as distinct species. In the same studies, LSU (28S rDNA) and the protein-
coding elongation factor 2 (EF2) of H. pseudofloresii were compared with other
members of the Halymeniales (Schneider et al., 2010). In both LSU (28S rDNA) and
EF2 ML trees, H. pseudofloresii grouped closely with the type, H. floresii which was
Univers
ity of
Mala
ya

45
consistent with anatomical assignment of this species to the genus Halymenia. The
results also demonstrated the polyphyly of Halymenia that needs further taxonomic
study.
In addition, Hernández-Kantún et al. (2012) employed the rbcL marker to delineate
branched Halymenia species in the Indo-Pacific and this have led to the recognition and
description of two new species, H. hawaiiana Hernández-Kantún & A. R .Sherwood
and H. tondoana O. DeClerck & Hernández-Kantún. Hernández-Kantún et al. (2012)
also highlighted the need to re-examine H. durvillei and H. floresii in the Indo-Pacific
based on molecular analysis. The rbcL marker was also utilized by Tan et al. (current
study) to study foliose Halymenia species which led to the discovery of new species H.
malaysiana in 2015 and H. johorensis in 2017. Besides, Azevedo et al. (2016a) assessed
the diversity of branched Halymenia species in Brazil by utilizing both rbcL and COI-
5P markers. Both rbcL and COI-5P phylogenies demonstrated that samples previously
assigned to H. floresii and H. pseudofloresii indeed correspond to three molecularly
divergent but morphologically similar lineages in which proposed as new species (H.
ignifera C. A. A. Azevedo, Cassano & M. C. Oliveira, H. pinnatifida C. A. A. Azevedo,
Cassano & M. C. Oliveira and H. silviae C. A. A. Azevedo, Cassano & M. C. Oliveira).
Azevedo et al. (2016a) also emphasized that without the use of molecular tools, the
morphological variations observed in Brazilian samples could be misinterpreted as
morphological plasticity. In the same year, phylogenetic relationships and species
diversity within the genus Halymenia in Brazil was also investigated by Azevedo et al.
(2016b). Three molecular markers included rbcL, COI-5P and UPA were adopted in
this study and revealed the presence of five Halymenia species in Brazil with the
description of a new species (H. cearensis C. A. A. Azevedo, Cassano & M. C.
Oliveira). This study also indicated the existence of at least two new genera classified to
Univers
ity of
Mala
ya

46
the Halymeniaceae and emphasized the importance of molecular tools in better
understanding of the diversity of Brazilian marine flora.
In general, the rbcL marker is the most widely used marker in the genus Halymenia.
It is mainly used to clarify taxonomic position of unknown species and to elucidate
phylogenetic relationships between species of Halymenia. Besides, the COI-5P marker
is the supplementary marker which useful in studying intra- and interspecific
relationships and revealing cryptic species.
Univers
ity of
Mala
ya

47
CHAPTER 3: MATERIALS AND METHODS
3.1 Sample collection and processing
Samples of Halymenia were collected from various localities in Malaysia, Thailand,
Indonesia and the Philippines (Figure 3.1) intertidally or subtidally by snorkelling or
scuba diving. A majority of Malaysian specimens were collected from several islands
around Peninsular Malaysia (Figure 3.2): Pulau Merambong, Johor; Pulau Babi Besar,
Johor; Pulau Tinggi, Johor and Pulau Besar, Malacca. Sample collection in Malaysia
was also made in Sabah, East Malaysia at four sites – Pulau Labuan, Pulau Mata Pahi,
Pulau Karindingan and Tun Mustapha Park (TMP) (Figure 3.2). The Philippines
specimens were sampled from Busuanga Island, Uson Island, Grande Island,
Paglugaban Island, Siargao Island, Camiguin Island and Subic Bay (Figure 3.3), while
the Thailand specimens were collected from populations in Koh Dok Mai (Figure 3.1).
Sample collection was also conducted in several localities in Indonesia: Pulau Weh,
Aceh; Amed, Bali; Ekas, Pulau Lombok; Ternate Island; South Gam, Raja Ampat and
Pulau Rote (Figure 3.1). Specimens are normally collected from specimens attached to
bedrock, shell, stone and coral rubble or as drift materials.
Univers
ity of
Mala
ya
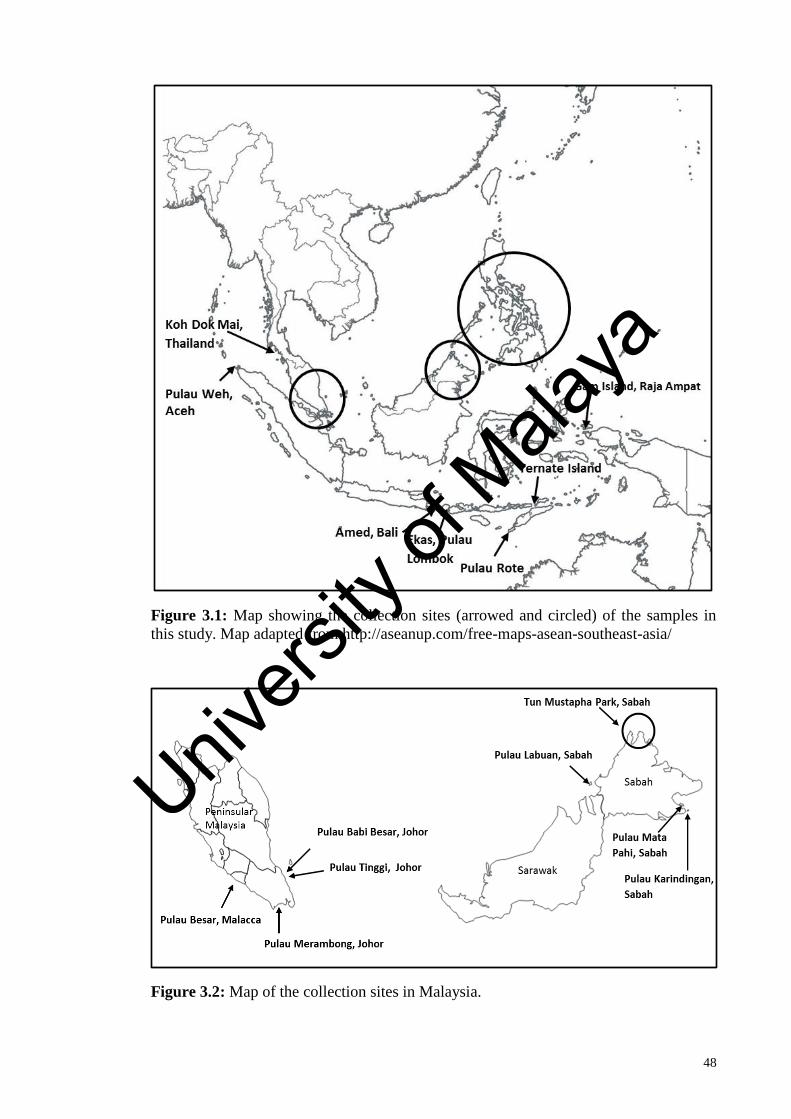
48
Figure 3.1: Map showing the collection sites (arrowed and circled) of the samples in
this study. Map adapted from http://aseanup.com/free-maps-asean-southeast-asia/
Figure 3.2: Map of the collection sites in Malaysia.
Univers
ity of
Mala
ya

49
Figure 3.3: Map of the collection sites in the Philippines. Map adapted from
http://www.d-maps.com/carte.php?num_car=587&lang=en
The collected specimens were cleaned with seawater to remove epiphytes, mud or
dirt. Specimens were pressed on herbarium sheets or preserved in 3-5% of formalin
seawater for morphological and anatomical study; whereas, a subsample of each
specimen was blotted-dry prior to dessicated in silica gel for molecular studies. Voucher
specimens are deposited in the University of Malaya Seaweeds and Seagrasses
Herbarium (KLU) and Leiden Herbarium (L). List of specimens used in this study is
provided in Appendix A.
Univers
ity of
Mala
ya

50
3.2 Morphological and anatomical studies
For morphological examination, the gross morphology of samples include thalli
colour, thalli size, blade shape, branching pattern (if applicable), order of branching (if
applicable), blade margin, presence or absence of stipe, presence or absence of spines or
proliferation on blade surfaces were observed and documented. Anatomical studies
were conducted by cutting thin sections using a razor blade. Sections were then stained
with 1% aniline blue acidified with diluted 1% HCl and subsequently mounted in 25- 30%
Karo syrup (Englewood Cliffs, NJ, USA) or were treated with Wittmann’s aceto-iron-
haematoxylin-chloral hydrate (Wittmann, 1965) and mounted in 50% Hoyer’s mounting
medium (Lin et al., 2008). The anatomical characters include shape and size of outer
cortical cells, shape and size of inner cortical cells, cortex thickness, presence or
absence of refractive ganglionic cells, size of cystocarps and tetrasporangia of stained
sections were then observed under an Olympus BX51 microscope (Olympus, Japan) and
the photomicrographs were taken with a DP72 digital camera.
3.3 Molecular analyses
3.3.1 DNA extraction
Genomic DNA of species of Halymenia were extracted from approximately 3 - 5 mg
silica gel-dried or herbarium specimens using the i-genomic Plant DNA Extraction Mini
Kit (iNtRON Biotechnology Inc., South Korea) according to the manufacturer’s
protocol with minor modifications. The seaweed materials were pulverized in liquid
nitrogen using a micropestle. This was followed by chemical lysis with the addition of
lysis buffer, RNase and Proteinase K to degrade proteins and RNA. The mixture was
vortexed vigorously before subjected to sonication for 10 minutes. After incubation at
Univers
ity of
Mala
ya

51
65°C for 45-60 minutes, the mixture was incubated on ice for 20 minutes to precipitate
proteins and polysaccharides. Subsequently, the glutinous mixture was centrifuged for
5 minutes and the clear lysate was transferred to a new tube to which binding buffer
with ethanol was added. The mixture was transferred to a spin column and followed by
centrifugation for binding of the DNA on the membrane. Contaminants such as
polysaccharides and proteins were further removed by washing buffer, and the DNA
bound to the membrane was eluted in two fractions of 50 μL elution buffer. The eluted
samples were later kept at -20˚C for long term storage.
3.3.2 Spectrophotometric determination of DNA concentration and purity
The concentration and purity of the isolated DNA were estimated using a
BioPhotometer (Eppendorf, Germany). For concentration measurement, the absorbance
reading of 1 at 260 nm corresponds to 50 μg/ ml for double-stranded DNA. Meanwhile,
the ratio of the absorbance at 260 nm and 280 nm (A260/A280) implies the purity of DNA.
A good quality DNA has an A260/A280 ratio of 1.8- 2.0, a lower ratio indicates DNA is
contaminated by protein, while a higher ratio suggests that DNA is contaminated by
RNA.
3.3.3 Polymerase chain reaction (PCR) amplification
The DNA was subjected to PCR amplification for different molecular markers,
including the mitochondrial COI-5P, nuclear LSU (28S rDNA), plastid rbcL and UPA.
The PCR amplification was carried out by Eppendorf EP Gradient S (Hamburg,
Germany) thermal cycler or Labnet MultiGene™ Gradient Thermal Cycler (Labnet,
USA) using the i-Taq™ Plus DNA Polymerase Kit (iNtRON Biotechnology, Korea).
Univers
ity of
Mala
ya

52
Each reaction contained 1-3 μL (25-50 ng) DNA template, 2 μL 10x i-Taq plus reaction
buffer, 2 μL dNTP mixture (consisting of 2.5 mM each of dNTP), 1.5 μL of each
forward and reverse primer (10 pmol/ μL), 0.25 μL i-Taq plus DNA polymerase
(iNtRON Biotechnology, Korea) and ultrapure water was added to make up the final
volume of 20 μL. A negative control with the same constituents but without template
DNA was prepared in each set of PCR reactions to detect contamination and monitor for
false positives. Concentration of DNA and annealing temperature were slightly
modified to achieve high yield product.
3.3.3.1 rbcL
Primers used for amplification of the rbcL gene were listed in Table 3.1. The almost
complete length of the rbcL gene was obtained using the primer pairs of F7/R1381 or
F7/RrbcS start. When this failed, two smaller overlapping fragments of rbcL were
amplified where the primer pairs F7/R753 or F57/R753 for amplification of the 5’ end
and primer pairs F577/R1150, F577/R1381 or F577/RrbcS start for amplification of the
3’end. When the attempt was unsuccessful, nested PCR were carried out to amplify two
smaller overlapping fragments of rbcL by using the diluted amplicons from the first
PCR run using the primer pairs F7/RrbcS start as the template DNA. The amplification
parameter of the rbcL gene is as follows: an initial denaturation at 94oC for 4 minutes,
followed by 35 cycles of denaturation at 93oC for 1 minute, annealing at 50
oC for 1
minute, extension at 72oC for 1.5 minutes; with a final extension at 72
oC for 7 minutes.
Annealing temperature was modified in the range of 48ºC to 52ºC to optimize
amplifications.
Univers
ity of
Mala
ya

53
Table 3.1: Primers used for amplification of rbcL.
Primer Primer Sequence (5’ → 3’) References
F7 AACTCTGTAGAACGNACAAG Gavio and Fredericq, 2002
F57 GTAATTCCATATGCTAATGAAGG Freshwater and Rueness, 1994
F577 GTATATGAAGGTCTAAAAGGTGG Freshwater and Rueness, 1994
R753 GCTCTTTCATACATATCTTCC Freshwater and Rueness, 1994
R1150 GCATTTGTCCGCAGTGAATACC Freshwater and Rueness, 1994
R1381 ATCTTTCCATAGATCTAAAGC Freshwater and Rueness, 1994
RrbcS start TGTGTTGCGCCGCCCTGTGTTAGTCTCAC Freshwater and Rueness, 1994
3.3.3.2 COI-5P
Primers used for amplification of the 5’ end of the COI gene were listed in Table 3.2.
Two degenerated primers were designed based on the sequences available in GenBank
using the Primer3 software (Untergasser et al., 2012). Amplifications of the COI-5P
region were accomplished using the primer pairs GHa1F/GHa1R, M13LF3/M13Ri or
COI1F/COI1R. When this failed, nested PCR were carried out using the primer pairs
M13LF3/M13Ri or COI1F/COI1R where the diluted amplicons from the first PCR run
using the primer pairs GHa1F/GHa1R as the template DNA. The amplification
parameter of the COI-5P region is as follows: an initial denaturation at 94oC for 4
minutes, followed by 35 cycles of denaturation at 93oC for 1 minute, annealing at 50
oC
for 1 minute, extension at 72oC for 1 minute; with a final extension at 72
oC for 7
minutes. Annealing temperature was modified in the range of 46ºC to 50ºC to optimize
amplifications.
Univers
ity of
Mala
ya

54
Table 3.2: Primers used for amplification of COI-5P.
Primer Primer Sequence (5’ → 3’) References
GHa1F TCAACAAATCATAAAGATATYGG Saunders, 2008
GHa1R CTTCWGGATGRCCAAAAAATCA Clarkston and
Saunders, 2010
M13LF3 TGTAAAACGACGGCCAGTACHAAYCAYAARGATATHGG Saunders and
Moore, 2013
M13Ri CAGGAAACAGCTATGACGGRTGICCRAARAAYCARAA Saunders and
Moore, 2013
COI1F GGAACACTTTAYTTAATTTTTGG This study
COI1R TGRTATARAATTGGATCWCC This study
3.3.3.3 UPA
UPA region was amplified using the primer pair p23SrV-f1/p23SrV-r1 as in
Sherwood and Presting (2007) (Table 3.3). The amplification parameter of the UPA is
as follows: an initial denaturation at 94oC for 3 minutes, followed by 40 cycles of
denaturation at 94oC for 30 seconds, annealing at 55
oC for 30 seconds, extension at
72oC for 50 seconds; with a final extension at 72
oC for 10 minutes. Annealing
temperature was modified in the range of 50ºC to 55ºC to optimize amplifications.
Table 3.3: Primers used for amplification of UPA
Primer Primer Sequence (5’ → 3’) References
p23SrV-f1 GGACAGAAAGACCCTATGAA Sherwood and Presting, 2007
p23SrV-r1 TCAGCCTGTTATCCCTAGAG Sherwood and Presting, 2007
Univers
ity of
Mala
ya

55
3.3.3.4 LSU (28S rDNA)
The central portion of the LSU (28S rDNA) was amplified using the primer pair
nu28SF/nu28SR as in Sherwood et al. (2010b) (Table 3.4). The amplification parameter
of the LSU is as follows: an initial denaturation at 94oC for 3 minutes, followed by 40
cycles of denaturation at 94oC for 30 seconds, annealing at 55
oC for 30 seconds,
extension at 72oC for 50 seconds; with a final extension at 72
oC for 10 minutes.
Annealing temperature was modified in the range of 50ºC to 55ºC to optimize
amplifications.
Table 3.4: Primers used for amplification of LSU (28S rDNA).
Primer Primer Sequence (5’ → 3’) References
nu28SF GGAATCCGCYAAGGAGTGTG Sherwood et al., (2010b)
nu28SR TGCCGACTTCCCTTACCTGC Sherwood et al., (2010b)
3.3.4 Determination of the amplification yield and quality by gel
electrophoresis, DNA purification and gene sequencing
The yield, size and quality of PCR amplicons were examined by electrophoresing
through a 1% agarose gel pre-stained with SYBR Safe DNA gel stain (Invitrogen, NY,
USA). A 1kb DNA ladder (Bioron, Germany) was used as a reference to estimate the
yield and the size of amplicons. Gel electrophoresis was carried out at 100 volts for 30
minutes, and then the gel was viewed under UV transilluminator using AlphaImager
2200 gel documentation system (Alpha Innotech, USA). After the electrophoretogram
was analyzed, desired products with a single band were directly purified while PCR
products with multiple bands underwent gel purification using LaboPass Gel and PCR
Clean-up Kit (Cosmo Genetech, Seoul, Korea) according to the manufacturer’s
protocols. The purified PCR products were then sent for automated DNA sequencing
Univers
ity of
Mala
ya

56
service (Genomics BioScience and Technology Co., Ltd., Taiwan) and were sequenced
on an Applied Biosystems 3730xl DNA Analyzer (Applied Biosystems, Foster City, CA,
USA) using the same primers used for PCR amplification and the BigDye Terminator
v3.1 Cycle Sequencing Kit (Applied Biosystems, USA).
3.3.5 Sequence and phylogenetic analyses
ChromasPro v1.5 (Technelysium Pty. Ltd., Australia) was employed to assemble the
forward and reverse sequence reads for each sample into contigs. Contigs were then
checked for nucleotide ambiguities and edited by examining electropherograms.
Published sequences of species of Halymeniaceae were downloaded from GenBank.
Consensus sequences and the published sequences were compiled and subsequently
aligned in ClustalX v. 2.0.8 (Larkin et al., 2007) to generate multiple sequence
alignments. The alignments were manually revised and trimmed in Bioedit v. 7.0.9.0
(Hall, 1999) by truncated sequences with longer lengths. In order to assess the level of
nucleotide variation between samples, MEGA6 (Tamura et al., 2013) was used to
compute uncorrected (p) pairwise genetic distances.
Two phylogenetic analyses ML and BI were implemented in this study. The
published sequences used for phylogenetic analyses are listed in Appendix B and C.
Each analysis dataset included the newly determined sequences for Halymenia and
reference sequences of Halymeniaceae downloaded from GenBank. Kakusan v.3
(Tanabe, 2007) was employed to search for the most appropriate model of sequence
evolution for ML and BI analyses with parameters inferred from the Akaike Information
Criterion (AIC) and the Bayesian Information Criterion (BIC) respectively. The models
and parameters selected for ML analyses are shown in Table 3.5. The best fit models
Univers
ity of
Mala
ya

57
for the BIC are as follow: GTR model for the rbcL dataset, HKY85 model for COI-5P,
HKY85+G for UPA and JC69+G for the LSU (28S rDNA) dataset.
ML analyses were performed in Treefinder v. October 2008 (Jobb et al., 2004) where
bootstrap support was estimated using 1000 bootstrap resamplings to evaluate
robustness. On the other hand, BI analyses were inferred in MrBayes v. 3.1.2 (Ronquist
and Huelsenbeck, 2003) where two parallel run of four chains of Markov Chain Monte
Carlo (MCMC) were conducted with one tree sampled every 100 generations for two
million generations. The two MCMC runs had reach convergence if the average
standard deviation of split frequencies less than 0.01. Convergence was evaluated using
Tracer v1.5 (Rambaut and Drummond, 2009b) by looking at the log likelihood plot.
Stationary was reached at generation 18,000. Therefore, the first 20,000 generations
were discarded as burn-in and the remaining generations were used to build the
consensus tree and determine the posterior probabilities.
All phylogenetic trees were visualized and processed using Figtree v1.3.1 (Rambaut
and Drummond, 2009a). All trees were rooted with appropriate outgroups and arranged
with decreasing node orders. Nodes with a bootstrap support lower than 70% and a
Bayesian posterior probability lower than 0.90 were weakly supported and considered
unresolved (Hillis and Bull, 1993; Huelsenbeck and Ronquist, 2001). Thamnoclonium
served as outgroups for the rbcL dataset, Grateloupia and Prionitis were the designated
outgroups for the COI-5P dataset, while Grateloupia selected as outgroups for both
UPA and LSU (28S rDNA) datasets.
Univers
ity of
Mala
ya

58
Table 3.5: Model and parameters selected by Kakusan3 for ML analysis of rbcL, COI-5P, UPA and LSU (28S rDNA) datasets.
Dataset Model
selected
-ln
likelihood
Nucleotide frequencies Substitution model rate matrix Gamma
distribution
shape
A C G T TC TA TG CA CG AG parameter
(alpha)
rbcL J2+G 7027.31 0.3132 0.1664 0.2037 0.3167 0.5118 0.0272 0.0504 0.2723 0.0504 0.3329 0.1814
COI-5P HKY+G 3447.21 0.2833 0.1534 0.1571 0.4062 0.4660 0.0170 0.0170 0.0170 0.0170 0.4660 0.1639
UPA HKY+G 1038.00 0.25 0.25 0.25 0.25 0.4523 0.2385 0.2385 0.2385 0.2385 0.4523 0.1450
LSU TVM+G 1108.32 0.2353 0.2249 0.3119 0.2280 0.2118 0.4128 0.0747 0.0765 0.0123 0.2118 0.1278
Univers
ity of
Mala
ya

59
3.3.6 Haplotype network analyses
In order to infer the genetic relationships among the haplotypes of H. malaysiana,
statistical parsimony implemented in TCS v1.21 (Clement et al., 2000) were employed
to construct haplotype networks for rbcL and COI-5P marker. This program collapsed
sequences with zero pairwise absolute distance into haplotype, treated gap as missing
data and connected the haplotypes with smaller differences until all haplotypes were
included in a single network based on parsimony limit (maximum number of
differences among haplotypes as a result of single substitutions with a 95% statistical
confidence).
Univers
ity of
Mala
ya

60
CHAPTER 4: RESULTS
4.1 Morphological and anatomical observations
Upon collection from field, the gross morphology of each specimen include thalli
colour, thalli size, blade shape, branching pattern (if applicable), order of branching (if
applicable), blade margin, presence or absence of stipe, presence or absence of spines or
proliferation on blade surfaces were observed and described. Anatomical characters
include shape and size of outer cortical cells, shape and size of inner cortical cells,
cortex thickness, presence or absence of refractive ganglionic cells, size of cystocarps
and tetrasporangia were then studied by examining the sections under microscope. The
morphological descriptions of each species were given below:
4.1.1 Halymenia malaysiana P.-L. Tan, P.-E. Lim, S.-M. Lin & S.-M. Phang,
Figures 4.1-4.3
Thalli are foliose (Figure 4.1), composed of mostly a single or, in some cases, few
blades, 6-25 cm long by 9-33 cm wide, arising from a small discoid holdfast without a
stipe, abruptly expanding into broad blades. Thalli ranging from pink to pinkish brown
with a supple cartilaginous and gelatinous (slimy) texture. Blades are oblong (Figures
4.1a-b) or suborbicular (Figure 4.1c) with a smooth surface, rarely with orbicular
proliferations (Figure 4.1d). Blade margins are mostly sinusoidally undulated (Figures
4.1a, 4.1c), sometimes minutely dentate (Figure 4.1b) or deeply cleft (Figure 4.1d).
The internal anatomy of blades is slightly different between young and old blades,
ranging from 150 µm to 700 µm in thickness (Figure 4.2) while the pigmented cortex is
Univers
ity of
Mala
ya

61
20-70 µm thick. Young blades are 150-200 µm in thickness, composed of 1-2 layers of
elongated outer cortical cells (8-13 µm long by 5-8 µm wide) and 2-3 layers of rounded
to ellipsoidal inner cortical cells (5-13 µm in diameter) (Figures 4.2a, 4.2c) with laxly
arranged medullary filaments (Figure 4.2b). Older blades are thickened, 240-700 µm in
thickness, consisting of elongated outer cortical cells and rounded inner cortex with
densely arranged medullary filaments (Figure 4.2d). Only few innermost cortical cells
become stellate, and refractive ganglionic cells are also present in the medulla, 40-90
µm in diameter, irregular in shape with 4-6 arms.
Spermatangial gametophytes were not found and only tetrasporic and female
structures were examined. Female gametophytes and tetrasporophytes are isomorphic.
Tetrasporangia sori are scattered over the thallus of fertile blades. Tetrasporangial
initials are first cut off from subcortical cells, then elongated (Figure 4.3a, arrows) and
divide into cruciately arranged tetraspores (Figures 4.3a-b). Mature tetrasporangia are
subspherical to oblong, 18-30 µm long by 15-18 µm wide (Figure 4.3b). Early post-
fertilization events were not observed. At one young stage of carposporophyte
development, ampullar filaments loosely surround the developing gonimolobes and
gonimoblast initial cell (Figure 4.3c, arrowhead), and the auxiliary cell (Figure 4.3c,
arrow) remains visible. Cystocarps are scattered over fertile blades and deeply
embedded in the layer between the inner cortex and medulla. Mature carposporophytes
are spherical, 90-120 µm in diameter, bearing several gonimolobes. Most cells of the
gonimolobes differentiate into ellipsoidal or ovoid carposporangia, 5-8 µm wide by 12-
20 µm long, released through an ostiole (Figures 4.3d-e).
Univers
ity of
Mala
ya

62
Figure 4.1: Thallus habit of Halymenia malaysiana. (a) A female thallus with oblong
blade (PSM12834). (b) A female thallus with minutely dentate margins (arrow)
(PSM12835). (c) A tetrasporic thallus with orbicular blade (PSM12850). (d) A sterile
thallus with orbicular proliferations on the surface (arrows) (PSM12838). [scale bars: a,
c, d = 5 cm; b = 1 cm]
Univers
ity of
Mala
ya

63
Figure 4.2: Vegetative structures of Halymenia malaysiana (PSM12835). (a) Cross-
section through the upper part of a young blade showing cortex and medulla. (b) Cross-
section through the median part of a young blade showing dense cortex and laxly
arranged medullary filaments. (c) Close up of Figure 4.2b showing details of cortical
and medullary filaments. (d) Cross-section through the basal part of an old blade
showing thickened cortex and densely arranged medullary filaments. [scale bars: a = 25
μm; c, d = 50 μm; b = 75 μm]
Univers
ity of
Mala
ya

64
Figure 4.3: Tetrasporangial (PSM12845) and cystocarp (PSM12834) morphology of
Halymenia malaysiana. (a) Cross-section through a tetrasporangia-bearing blade
showing tetrasporangial initials (arrows) and dividing tetrasporangium (arrowhead) cut-
off laterally from the subcortical cells. (b) Close up of a mature, cruciately divided
tetrasporangium (arrow). (c) Cross-section through a young cystocarp showing young
gonimolobe, gonimoblast initial (arrowhead), and basal fusion cell (arrow). (d) Cross-
section through a mature cystocarp. (e) Close up of mature cystocarp showing
carposporophyte borne on a basal fusion cell (arrow). [scale bars: a = 40 μm; b, d, e =
20 μm; c = 50 μm]
Univers
ity of
Mala
ya

65
4.1.2 Halymenia maculata J. Agardh, Figures 4.4-4.5
Thalli (Figure 4.4) are composed of single or few thick blades, 8-10 cm long by 9-15
cm wide, arising from a stiff stipe. Blades are circular to irregularly lobed and dark pink
to reddish brown in colour (Figure 4.4). Thallus texture is cartilaginous and gelatinous
(slimy). Blade surfaces are mottled with dark red, circular spots, some bearing
proliferations or small bladelets (Figures 4.4a-d). Margins of blades are irregularly to
regularly lobed (jigsaw-like) or denticulate (Figures 4.4a-b). In addition, some
specimens have surface protuberances on the thalli (Figure 4.4d).
The internal anatomy of blades is slightly different between young and old blades,
ranging from 170-480 µm in thickness (Figures 4.5a-b). Young blades are 170-220 µm
in thickness, composed of 1-2 layers of elongated outer cortical cells (8-23 µm long by
2-7 µm wide) and 3-4 layers of rounded to stellate inner cortical cells (4-16 µm in
diameter) with laxly arranged medullary filaments (Figure 4.5a). Older blades are
thickened, 250-480 µm in thickness, consisting of elongated outer cortical cells and
stellate inner cortex with densely arranged medullary filaments (Figure 4.5b). Refractive
ganglionic cells are also present in the medulla.
Spermatangial gametophytes were not found and only tetrasporic and female
structures were examined. Female gametophytes and tetrasporophytes are isomorphic.
Cystocarps are scattered over fertile blades and deeply embedded in the layer between
the inner cortex and medulla. Mature carposporophytes are spherical to pyriform, 80-
130 µm in diameter. Carposporangia are spherical to ovoid, 8-11 µm in diameter,
released through an ostiole (Figure 4.5c). Tetrasporangial sori are scattered over the
thallus of fertile blades. Tetrasporangial initials are first cut off from subcortical cells,
then elongated (Figure 4.5d) and divide into cruciately arranged tetraspores (Figure
Univers
ity of
Mala
ya

66
4.5e). Mature tetrasporangia are subspherical, 14-18 µm long by 16-20 µm wide (Figure
4.5e).
Figure 4.4: Thallus habit of Halymenia maculata. (a) A tetrasporic thallus showing
jigsaw-like blade margin and mottled surface (PSM12852). (b) A female thallus
showing jigsaw-like blade margin and mottled surface (PSM12833). (c) Part of a female
thallus showing surface proliferations (arrows) (PSM12833). (d) Part of a female thallus
showing surface protuberances (arrows) and bladelet (arrowhead) (PSM12833). [scale
bars: a,b = 2 cm; c, d = 1 cm]
Univers
ity of
Mala
ya

67
Figure 4.5: Vegetative structures (PSM12833), cystocarp (PSM12833) and
tetrasporangial (PSM12852) morphology of Halymenia maculata. (a) Cross-section
through the upper part of a young blade showing cortex and laxly arranged medullary
filaments. (b) Cross-section through the basal part of an old blade showing thickened
cortex and densely arranged medullary filaments. (c) Cross-section through a mature
cystocarp. (d) Cross-section through a tetrasporangia-bearing blade showing dividing
tetrasporangium (arrow) cut-off laterally from the subcortical cells. (e) Close up of a
mature, cruciately divided tetrasporangium (arrow). [scale bars: a, c = 50 μm; b, d = 30
μm; e = 20 μm]
Univers
ity of
Mala
ya

68
4.1.3 Halymenia cf. dilatata Zanardini, Figure 4.6
Thallus is foliose, reddish with darker spots, cartilaginous and gelatinous in texture.
Thallus (Figure 4.6a) is 7 cm high by 9 cm wide, arising from a short stipe (3 mm long,
2 mm wide) attached to a discoid holdfast. Blades are irregularly shaped (Figure 4.6a)
with smooth surface. Blade margins (Figure 4.6a) are minutely dentate.
Thallus blades (Figures 4.6b-c) are 180-200 µm thick in the apical region and 300-
320 µm thick in basal region. The pigmented cortex is 35-60 µm thick, with two to three
layers of subspherical to elongated outer cortical cells (7-12 µm long by 3-4 µm wide)
and two to four layers of irregularly stellate, tangentially flattened inner cortical cells (6-
18 µm in diameter). The medulla lax with anticlinal filaments in the apical region
(Figure 4.6b) and densely filled with both anticlinal and intertwined filaments (Figure
4.6c) in the basal region. Refractive ganglionic cells are present in the medulla but not
abundant, mostly with 6 arms.
Only female gametophyte was examined. Cystocarps are scattered over fertile blades
and deeply embedded in the layer between the inner cortex and medulla. Mature
carposporophytes are spherical to pyriform, 80-90 µm in diameter. Carposporangia are
spherical to ovoid, 10-17 µm in diameter (Figure 4.6d).
Univers
ity of
Mala
ya

69
Figure 4.6: Thallus habit and anatomy of Halymenia cf. dilatata (PSM12857). (a) A
female thallus. (b) Cross-section through the apical region of a blade showing cortex
and laxly arranged medullary filaments. (c) Cross-section through the basal region of a
blade showing thickened cortex and densely arranged medullary filaments. (d) Cross-
section through a mature cystocarp showing carposporophyte borne on a basal fusion
cell (arrow). [scale bars: a = 2 cm, b = 60 μm; c = 25 μm; d = 40 μm]
Univers
ity of
Mala
ya

70
4.1.4 Halymenia johorensis P.-L. Tan, P.-E. Lim, S.-M. Lin & S.-M. Phang,
Figures 4.7-4.8
Thalli are foliose, cartilaginous and gelatinous in texture, pink to pinkish red with
faded yellowish green in the middle regions of plants (Figures 4.7a-c). Thalli are 22-40
cm long by 10-30 cm wide, arising from a narrow-cuneate stipe (1-4 mm long, 2-3 mm
wide) attached to a discoid holdfast, expanding broadly into flat blades. Blades are
elliptical (Figure 4.7a), oblong (Figure 4.7b) or irregularly shaped (Figure 4.7c), incised
with some perforations. Blade surfaces are smooth or rugose with proliferations, while
rugose mainly found in the older plants (Figure 4.7c). Blade margins (Figures 4.7a-c)
are mostly irregularly dentate and cleft, sometimes smooth.
Thallus blades (Figures 4.7d–g) are 120-400 µm thick. The pigmented cortex is 15-
45 µm thick, with one to two layers of isodiametric outer cortical cells and one to four
layers of rounded to stellate inner cortical cells. The arrangement of cortex and medulla
in different aged blades differ slightly. Young blades are 120-200 µm thick, consisting
of one layer of isodiametric outer cortical cells (3-6 µm long by 2-4 µm wide) and one
to two layers of rounded to stellate inner cortical cells (4-12 µm in diameter) with laxly
arranged medullary filaments (Figure 4.7d). Older blades are 220-400 µm thick,
composed of one to two layers of isodiametric outer cortical cells and three to four
layers of rounded to stellate inner cortical cells with densely arranged medullary
filaments (Figure 4.7f). The medulla is primarily composed of anticlinal filaments in
young blades (Figures 4.7d-e) and both anticlinal and intertwined filaments (Figure 4.7f)
in older blades. A few refractive ganglionic cells are present in the medulla, these
irregular in shape with three to five arms (Figure 4.7g).
Male gametophytes were not found, and only female gametophytes and
tetrasporophytes were examined. Female gametophytes and tetrasporophytes are
Univers
ity of
Mala
ya

71
isomorphic. Carpogonial branches and early post-fertilization stages were not found.
The auxiliary cell ampulla is formed in the inner cortex, composed of a prominent
auxiliary cell and several orders of branched, filamentous cell rows (Figure 4.8a).
Following presumed diploidization, the auxiliary cell cuts off a gonimoblast initial
terminally from a cup-shaped depression (Figure 4.8b). The gonimoblast initial divides
to produce a primary gonimolobe. Basal ampullar cells become enlarged and the upper
ampullar cells are elongated (Figure 4.8b), while the inner cortical cells in vicinity of
the developing gonimoblasts produce secondary medullary filaments (Figure 4.8b). As
development of the gonimoblast continues, the auxiliary cell fuses with its adjacent
ampullar cells to form a fusion cell (Figures 4.8c-d). As cystocarp development
continues, more secondary medullary filaments are produced from the cell layer
between inner cortical cells and medulla (Figures 4.8c-d). Both elongated ampullar
filaments and secondary medullary filaments comprise the involucre that surrounds the
gonimolobe (Figures 4.8c–d). Mature carposporophytes (Figure 4.8e) are spherical to
pyriform, 80-250 µm in diameter, scattered over both surfaces of the fertile blades
except in the basal region. Cystocarps are embedded in the medulla but cause
protuberances on both sides of the thalli surfaces as small hemispherical blisters.
Carposporangia are spherical to ovoid, 6-12 µm in diameter, released through an ostiole
(Figure 4.8e). Mature tetrasporangia (Figure 4.8f, arrow) are cruciately divided,
subspherical to oblong, 14–20 µm long by 10–14 µm wide, scattered over the fertile
blades.
Univers
ity of
Mala
ya

72
Figure 4.7: Habit and vegetative morphology of Halymenia johorensis. (a) Cystocarpic
thallus (PSM12870). (b) Tetrasporic thallus (PSM10663). (c) Cystocarpic thallus
(PSM12873). (d) Cross section through upper part of a young blade showing thin cortex
and prominent anticlinal medullary filaments (PSM12870). (e) Cross section through
middle part of a young blade showing densely arranged cortical cells and laxly
arranged, filamentous medullary cells (PSM12870). (f) Cross section through middle
part of an old blade showing thickened cortex and densely arranged, filamentous
medullary cells (PSM12873). (g) Cross section through a young blade showing
refractive ganglionic cell (arrow) (PSM12874). [scale bars: a=7 cm; b=5 cm; c=2 cm; d,
f, g=50 µm; e=25 µm]
Univers
ity of
Mala
ya

73
Figure 4.8: Cystocarp (PSM12870) and tetrasporangial (PSM10663) morphology of
Halymenia johorensis. (a) Cross section through a young female blade showing
auxiliary cell ampulla, enlarged auxiliary cell (aux) and ampullar filaments
(arrowheads). (b) Young cystocarp showing auxiliary cell (aux) cutting off a
gonimoblast intial (gi), which produced a primary gonimolobe (g). Note that basal
ampullar cells (arrowheads) slightly enlarged, upper ampullar cells are elongated, and
the inner cortical cells in the vicinity of the gonimolobe have begun to send out
secondary medullary filaments (arrows). (c) Another young cystocarp showing a basal
fusion cell (fc), gonimoblast initial (gi) and gonimolobe enveloping by elongated
Univers
ity of
Mala
ya

74
ampullar filaments and secondary medullary filaments (arrows). (d) Immature cystocarp
showing fusion cell (fc), gonimoblast initial (gi), gonimoblasts enveloping by elongated
ampullar filaments and secondary medullary filaments (arrows). Note that the remnant
of ampullar filaments (arrowheads) do not divide any further. (e) Mature cystocarp
(arrow) showing gonimoblast initial (gi), basal fusion cell (fc) and gonimoblasts.
Arrowhead indicates ostiole. Note that most cells of gonimoblasts differentiate into
carposporangia. (f) Mature, cruciately divided tetrasporangium (arrow) immersed in the
cortex. [scale bars: a–e=25 μm; f= 15 μm]
4.1.5 Halymenia tondoana O. DeClerck & J.J. Hernández-Kantun, Figure 4.9
Thalli are pink to pinkish green, cartilaginous and gelatinous in texture. Thalli are 3-
6 cm long by 5–8 cm wide with unbranched cartilaginous stipe (7-10 mm long, 2-3 mm
in diameter) (Figure 4.9a). Blades resembling dichotomous bifurcations, branched up to
7 orders, branching distally from main axis. Marginal branchlets are abundant
throughout the thalli in the distal region. Blade surfaces are smooth with maculae
without spine. Margins of blades are lacerate and with proliferations.
Thallus blades are 150-400 µm thick. The pigmented cortex is 35-60 µm thick, with
one to two layers of elongated outer cortical cells and two to four layers of spherical
inner cortical cells. The arrangement of cortex and medulla in different aged blades
differ slightly. Young blades are 150-200 µm thick, consisting of one layer of elongated
outer cortical cells (5-8 µm long by 2-3 µm wide) and two to three layers of spherical
inner cortical cells (8-16 µm in diameter) with laxly arranged medullary filaments
(Figure 4.9b). Older blades are 300-400 µm thick, composed of two layers of elongated
outer cortical cells and three to four layers of spherical inner cortical cells with densely
arranged medullary filaments (Figure 4.9c). Refractive ganglionic cells are present in
the medulla, these irregular in shape with five to six arms.
Spermatangial gametophytes and tetrasporophytes were not found and only female
gametophytes were examined. Cystocarps are scattered over fertile blades and deeply
Univers
ity of
Mala
ya

75
embedded in the layer between the inner cortex and medulla. Mature carposporophytes
are pyriform, 100-150 µm in diameter. Carposporangia are spherical to ovoid, 10-15 µm
in diameter (Figure 4.9d).
Figure 4.9: Thallus habit and anatomy of Halymenia tondoana (PSM12955). (a) A
female thallus. (b) Cross-section through the apical region of a blade showing cortex
and laxly arranged medullary filaments. (c) Cross-section through the basal region of a
blade showing thickened cortex and densely arranged medullary filaments. (d) Cross-
section through a mature cystocarp showing carposporophyte borne on a basal fusion
cell (arrow). [scale bars: a = 2 cm; b, c = 30 μm; d = 60 μm]
Univers
ity of
Mala
ya

76
4.1.6 Halymenia durvillei Bory de Saint-Vincent, Figures 4.10-4.11
Thalli are red to reddish brown, cartilaginous and gelatinous in texture. Thalli
(Figures 4.10a-d) are up to 40 cm long with unbranched stipe (5-15 mm long, 1-6 mm
in diameter) bearing one or sometimes a few primary axes. Blades subdichotomously,
trichotomously or irregularly branched up to 7 orders (Figure 4.10a), axes either flat or
contorted and up to 4 cm wide, gradually tapering to apices. In some specimens,
secondary axes are arising as proliferations on the surface of the primary axes (Figure
4.10b). Blade surfaces are smooth in young plants (Figure 4.10c) and covered with
spiny proliferations in older plants (Figure 4.10b). Some specimens do not have surface
spine even in older thalli (Figures 4.10a, d). Margins of blades are dentate or laciniate,
sometime smooth with proliferations. Proliferations and spine-like marginal branchlets
are abundant throughout thalli in proximal and distal regions in some specimens
(Figures 4.10a-b) or abundant in the distal region but rare in the proximal region in
others (Figure 4.10d).
Thallus blades (Figures 4.11a-b) are 200-1300 µm thick. The pigmented cortex is 45-
150 µm thick, with one to four layers of subspherical to elongated outer cortical cells
and three to six layers of rounded to stellate inner cortical cells. The arrangement of
cortex and medulla in different aged blades differ slightly. Young blades are 200-600
µm thick, consisting of one to two layers of subspherical to elongated outer cortical
cells (8-13 µm long by 2-4 µm wide) and two to three layers of rounded to stellate inner
cortical cells (5-24 µm in diameter) with laxly arranged medullary filaments (Figure
4.11a). Older blades are 800-1300 µm thick, composed of elongated outer cortical cells
and rounded to stellate inner cortical cells with densely arranged medullary filaments
(Figure 4.11b). The medulla is primarily composed of longitudinal filaments in young
blades (Figure 4.11a) and both anticlinal and intertwined filaments (Figure 4.11b) in
Univers
ity of
Mala
ya

77
older blades. Refractive ganglionic cells are present in the medulla, these irregular in
shape with six to nine arms.
Spermatangial gametophytes were not found and only tetrasporophytes and female
gametophytes were examined. Tetrasporophytes and female gametophytes are
isomorphic. Tetrasporangial sori are scattered over the thallus of fertile blades.
Tetrasporangial initials are first cut off from subcortical cells, then elongated and divide
into cruciately arranged tetraspores (Figure 4.11c). Mature tetrasporangia (Figure 4.11c,
arrows) are cruciately divided, subspherical to oblong, 15-27 µm long by 10-15 µm
wide. Cystocarps are scattered over fertile blades and deeply embedded in the layer
between the inner cortex and medulla. Mature carposporophytes are spherical to
pyriform, 100-130 µm in diameter. Carposporangia are spherical to ovoid, 10-12 µm in
diameter, released through an ostiole (Figure 4.11d).
Univers
ity of
Mala
ya

78
Figure 4.10: Thallus habit of Halymenia durvillei. (a) Thallus which branched up to
seventh orders (PSM12836). (b) Thallus with surface spines, spine-like marginal
branchlets and several secondary axes are arising as proliferations on the surface of the
primary axes (PSM12999). (c) Young thallus with smooth surface (PSM12947). (d)
Thallus with proliferations and spine-like marginal branchlets which common in the
distal region but rare in the proximal region (PSM12942). [scale bars: a = 5 cm; b = 4
cm; c, d = 2.5 cm]
Univers
ity of
Mala
ya

79
Figure 4.11: Vegetative (PSM12942), tetrasporangial (PSM12939) and cystocarp
(PSM13011) morphology of Halymenia durvillei. (a) Cross-section through the median
part of a young blade showing cortex and laxly arranged medullary filaments. (b) Cross-
section through the basal part of an old blade showing thickened cortex and densely
arranged medullary filaments. (c) Cross-section through a tetrasporangia-bearing blade
showing dividing tetrasporangium (arrowhead) cut-off laterally from the subcortical
cells and cruciately divided tetrasporangia (arrows). (d) Cross-section through a mature
cystocarp showing carposporophyte borne on a basal fusion cell (arrow). Arrowhead
indicates ostiole. [scale bars: a, c = 20 μm; b, d = 40 μm]
4.1.7 Halymenia sp. A, Figure 4.12
Thalli are cartilaginous and gelatinous in texture. Thalli are 16-26.5 cm long by 11–
41 cm wide with stipe (4 mm long, 3 mm in diameter). Blades irregularly branched up
to 4 orders, branches and branchlets abruptly constricted at bases, gradually tapering to
apices (Figures 4.12a-b). Blade surfaces are smooth without spine or proliferations.
Univers
ity of
Mala
ya

80
Margins of blades are smooth with proliferations. Proliferations and spine-like marginal
branchlets are abundant throughout thalli in the distal region but rare in the proximal
region.
The internal anatomy of blades is slightly different between young and old blades,
ranging from 130 µm to 600 µm in thickness (Figures 4.12c-d) while the pigmented
cortex is 25-65 µm thick. Young blades are 130-300 µm in thickness, composed of 1-2
layers of subspherical to elongated outer cortical cells (5-9 µm long by 2-3 µm wide)
and 2-3 layers of rounded to stellate inner cortical cells (5-15 µm in diameter) with laxly
arranged medullary filaments (Figure 4.12c). Older blades are thickened, 400-600 µm in
thickness, consisting of subspherical to elongated outer cortical cells and rounded to
stellate inner cortex with densely arranged medullary filaments (Figure 4.12d). The
medulla is primarily composed of longitudinal filaments in young blades (Figure 4.12c)
and both anticlinal and intertwined filaments (Figure 4.12d) in older blades. Refractive
ganglionic cells are present in the medulla.
Spermatangial gametophytes and tetrasporophytes were not found and only female
gametophytes were examined. Cystocarps are scattered over fertile blades and deeply
embedded in the layer between the inner cortex and medulla. Mature carposporophytes
are spherical to pyriform, 85-100 µm in diameter. Carposporangia are spherical to ovoid,
8-10 µm in diameter (Figure 4.12e).
Univ
ersity
of M
alaya

81
Figure 4.12: Thallus habit and anatomy of Halymenia sp. A. (a) Cystocarpic thallus
(PSM12887). (b) Young cystocarpic thallus (PSM12888). (c) Cross-section through the
median part of a young blade showing cortex and laxly arranged medullary filaments.
(PSM12887). (d) Cross-section through the basal region of a blade showing thickened
cortex and densely arranged medullary filaments. (PSM12887) (e) Cross-section
through a mature cystocarp (PSM12887). [scale bars: a = 5 cm; b = 3 cm; c, d = 20 μm;
e = 30 μm]
Univers
ity of
Mala
ya

82
4.2 Molecular analyses
4.2.1 DNA extraction
The genomic DNA of species of Halymenia were extracted using the commercially
available i-genomic Plant DNA Extraction Mini Kit, (iNtRON Biotechnology Inc.,
South Korea) following the manufacturer’s protocol. A few minor modifications were
made by subjecting the mixture to sonication for 10 minutes before incubation,
extending the incubation time at 65°C and on ice. As measured by spectrophotometric,
the concentration of DNA extracted from herbarium was in the range of 10-25 ngμL-1
and the OD readings of A260/A280 were in the range of 1.40 to 1.80. While the
concentration of DNA extracted from silica gel-dried specimen was in the range of 30-
60 ngμL-1
and the OD readings of A260/A280 were in the range of 1.60 to 1.98. Only
good quality isolated DNA which have A260/A280 of 1.8 to 2.0 were subjected to PCR
amplification.
4.2.2 PCR amplification
The plastid rbcL, the mitochondrial COI-5P, the plastid UPA and the nuclear LSU
(28S rDNA) were subjected to PCR amplification. Most samples were successfully
producing amplicons over one PCR run. In contrast, a subsequent nested PCR was
performed to produce two smaller overlapping fragments of the rbcL and COI-5P,
which can thus be combined to form a contig representing the desired length of the
genetic marker.
Satisfactory amplifications of all genetic markers were attained with the optimum
annealing temperature of 50ºC. The amplicon sizes of the rbcL, COI-5P, UPA and LSU
(28S rDNA) were approximately 1400, 600, 400 and 600 base pairs (bp) respectively as
Univers
ity of
Mala
ya

83
shown in Figure 4.13 and Figure 4.14. Most of the species of Halymenia investigated in
this study were successfully producing amplicons for all genetic markers except H. cf.
dilatata which only the rbcL amplicon attained. The band of the highest intensity on the
gel mostly is the target band. In some cases, non-specific amplifications (more than one
band) were observed. This can be solved by identifying the desire band against the
ladder and subjected to gel purification the target band or adjusting the annealing
temperature for PCR amplification to get specific amplification.
Figure 4.13: Electrophoretogram showing amplicons of the plastid rbcL (~1,400bp)
and mitochondrial COI-5P (~600bp) genetic markers. Each number represents sample
as follow: 1= H. durvillei, 2= H. tondoana, 3= H. malaysiana, 4= H. maculata, 5= H.
johorensis, 6= Halymenia sp. A and 7= H. cf. dilatata. 1kb and –ve are 1 kilobase
DNA ladder and negative controls respectively.
Univers
ity of
Mala
ya

84
Figure 4.14: Electrophoretogram showing amplicons of the plastid UPA (~400bp) and
nuclear LSU (28S rDNA) (~600bp) genetic markers. Each number represents sample as
follow: 1= H. durvillei, 2= H. tondoana, 3= H. malaysiana, 4= H. maculata, 5= H.
johorensis and 6= Halymenia sp A. 1kb and –ve are 1 kilobase DNA ladder and
negative controls respectively.
4.2.3 Sequence analyses
4.2.3.1 rbcL
The rbcL gene for all samples was successfully amplified and sequenced. The length
of the rbcL sequences for Halymenia in this study ranged from 1250 to 1460 bp
depending on the primer sets used for sequencing. An unambiguous alignment was
generated since there is no insertion or deletion mutation in the rbcL sequences. The
length of the rbcL alignment used in this study was standardized and trimmed to 1250
bp.
There were no intraspecific divergences within samples of H. johorensis and
Halymenia sp. A respectively. The range of intraspecific rbcL sequence variation (Table
4.1) between H. durvillei populations, 0-2.56%, was considerably higher than other
Univers
ity of
Mala
ya

85
populations such as H. tondoana, 0.08-0.16%; H. maculata, 0.08-0.48%; H. malaysiana,
0-0.58%; H. johorensis, 0% and Halymenia sp. A, 0%. The interspecific pairwise
distance between these seven Halymenia species ranged from 1.22-1.46% between H.
maculata and H. cf. dilatata to 6.93-7.42% between H. maculata and H. malaysiana.
Halymenia tondoana and Halymenia sp. A were closely related as shown by their
genetic variation of 2.24-2.34%.
Table 4.1: Percentage of pairwise distance between rbcL sequences of seven
Halymenia species examined in this study, excluding gaps and ambiguities.
(1)
(2) (3) (4) (5) (6) (7)
(1) H. durvillei
0-2.56
(2) H. tondoana
5.04-6.00
0.08-0.16
(3) H. maculata
6.28-7.40 6.16-6.37 0.00-0.48
(4) H. malaysiana
5.48-6.70 6.66-7.11
6.93-7.42
0.00-0.58
(5) Halymenia sp. A
5.60-6.46 2.24-2.34 6.32-6.59
6.98-7.30 0.00
(6) H. johorensis
5.52-6.37
5.60-5.72 6.75-6.91 6.42-6.78 6.00 0.00
(7) H. cf. dilatata
6-7.03 5.89-5.93 1.22-1.46
6.45-6.77 6.13 6.45 -
4.2.3.2 COI-5P
The COI-5P region for all species of Halymenia examined in this study was
successfully amplified and sequenced except for H. cf. dilatata. The length of the COI-
5P sequences generated for Halymenia in this study ranged from 635 to 680 bp
depending on the primer sets used for sequencing. There was no alignment problem for
this dataset with no indels observed. The final alignment of the COI-5P sequences used
in this study was trimmed to 632 bp.
Univers
ity of
Mala
ya

86
There was no sequence variation between each sample of H. johorensis and
Halymenia sp. A respectively (Table 4.2). The range of intraspecific COI-5P genetic
variation between H. durvillei populations, 0-5.70 %, was remarkably higher than other
populations such as H. tondoana, 0.32%; H. maculata, 0.33%; H. malaysiana, 0-1.58%;
H. johorensis, 0% and Halymenia sp. A, 0%. H. tondoana was more closely related to
Halymenia sp. A with a divergence of 4.11-4.27%, while H. durvillei being most
divergent from H. maculata with a variation of 10.43-12.46%.
Table 4.2: Percentage of pairwise distance between COI-5P sequences of six
Halymenia species examined in this study, excluding gaps and ambiguities.
(1)
(2) (3) (4) (5) (6)
(1) H. durvillei
0.00-5.70
(2) H. tondoana
7.75-9.49 0.32
(3) H. maculata
10.43-12.46 10.22-10.91 0.33
(4) H. malaysiana
7.75-9.29 9.97-10.92 10.70-11.40 0.00-1.58
(5) Halymenia sp. A
7.21-8.54 4.11-4.27 10.54-11.07 9.81-10.13 0.00
(6) H. johorensis
6.49-8.07 7.91-8.07 9.27-9.59 7.44-8.07 8.07 0.00
4.2.3.3 UPA
The UPA region for all species of Halymenia examined in this study was
successfully amplified and sequenced with the primer pair p23SrV-f1/p23SrV-r1 except
for H. cf. dilatata. The length of the UPA sequences generated for the examined taxa
ranged from 410 to 420 bp. There was no alignment problem for this dataset since no
indels discovered. The final alignment of the UPA sequences generated in this study
was trimmed to 370 bp.
Univers
ity of
Mala
ya

87
The range of intraspecific UPA sequence divergence (Table 4.3) between H. durvillei
populations, 0-2.70%, was considerably higher than other populations such as H.
tondoana, 0%; H. maculata, 0.27-0.54%; H. malaysiana, 0.27-1.08%; H. johorensis, 0%
and Halymenia sp. A, 0%. The interspecific divergences between these six Halymenia
species ranged from 1.08% between H. tondoana and Halymenia sp. A to 4.59-5.14%
between H. maculata and H johorensis.
Table 4.3: Percentage of pairwise distance between UPA sequences of six Halymenia
species examined in this study, excluding gaps and ambiguities.
(1)
(2) (3) (4) (5) (6)
(1) H. durvillei
0.00-2.70
(2) H. tondoana
1.35-2.70 0.00
(3) H. maculata
2.70-4.59 3.24-3.78 0.27-0.54
(4) H. malaysiana
2.43-5.14 3.51-4.32 3.78-5.14 0.27-1.08
(5) Halymenia sp. A
1.35-2.70 1.08 3.24-3.78 3.51-4.32 0.00
(6) H. johorensis
2.97-3.78 3.24 4.59-5.14 3.78-4.59 3.24 0.00
4.2.3.4 LSU (28S rDNA)
The central portion of the LSU (28S rDNA) for all species of Halymenia examined
in this study was successfully amplified, sequenced with the primer pair
nu28SF/nu28SR except for H. cf. dilatata. The length of the LSU (28S rDNA)
sequences generated for the examined taxa ranged from 645 to 670 bp. The final
alignment of the LSU (28S rDNA) sequences generated in this study was limited to 643
sites, including a few introduced gaps.
Univers
ity of
Mala
ya

88
There were no intraspecific divergences within samples of each Halymenia species
except for H. durvillei with a variation of 0-0.16%. The interspecific divergences
between these six Halymenia species (Table 4.4) ranged from 0-0.16% between H.
tondoana and H. durvillei to 0.49-0.65% between H. durvillei and H. maculata.
Although there appeared to be no genetic distance between a few H. durvillei samples
(SGAD1405001, SGAD1209005, SGAD1012346, SGAD1211004 and SGAD1209204)
and also between these H. durvillei samples and H. tondoana, these entities actually
differed to each other by 2-3 bp indels which the gaps introduced were not taken into
consideration for the calculation of the sequence divergence.
Table 4.4: Percentage of pairwise distance between LSU (28S rDNA) sequences of
six Halymenia species examined in this study, excluding gaps and ambiguities.
(1) (2) (3) (4) (5) (6)
(1) H. durvillei
0.00-0.16
(2) H. tondoana
0.00-0.16 0.00
(3) H. maculata
0.49-0.65 0.49 0.00
(4) H. malaysiana
0.16 0.16 0.65 0.00
(5) Halymenia sp. A
0.16-0.32 0.16 0.64 0.32 0.00
(6) H. johorensis
0.32-0.49 0.32 0.81 0.49 0.49 0.00
4.2.4 Phylogenetic analyses
4.2.4.1 rbcL
A total of 128 taxa with 1250 bp in length were selected for rbcL phylogenetic
analyses. The identical sequences were collapsed to haplotypes. Two species of
Thamnoclonium Kützing (Halymeniaceae) were chosen as outgroups for this dataset.
Univers
ity of
Mala
ya

89
The topologies of rbcL phylogeny were inferred using both ML and BI phylogenetic
analyses and only the ML tree was presented (Figure 4.15).
The ML phylogenetic tree based on the rbcL sequences (Figure 4.15) showed that
the genera Halymenia, Gelinaria Sonder and Epiphloea J. Agardh formed an
assemblages with moderately bootstrap support (ML= 81%) but high posterior
probability (BI= 1.00). Seven species of Halymenia examined in this study were
distributed in three clades. Clade 1 composed mainly of branched Halymenia (H.
durvillei, H. harveyana, H. hawaiiana, H. tondoana, Halymenia sp. A, H. pinnatifida)
and a foliose Halymenia species (H. silviae) but their relationships were not supported
(ML<50%, BI<0.50). Halymenia durvillei was monophyletic and resolved with strong
bootstrap support (ML= 100%, BI= 1.00). There were three main subclades for H.
durvillei, dividing the specimens from (1) Malaysia, Indonesia, and the Philippines
(ML= 100%, BI= 1.00), (2) Sri Lanka, Indonesia, South Africa and Tanzania (ML=
100%, BI= 1.00) and (3) Japan, Malaysia, Indonesia and the Philippines (no support).
Halymenia tondoana formed a highly supported monophyletic clade (ML= 89%, BI=
1.00) which was also a sister to Halymenia sp. A (ML= 100%, BI= 1.00).
On the other hand, foliose Halymenia, two genera Gelinaria and Epiphloea were
resided in Clade 2 (ML= 51%, BI= 0.94). Halymenia malaysiana specimens were all
clustered together with strong support (ML= 96%, BI= 1.00) and occurred as a sister
group to H. dilatata from Japan with maximum support (ML= 100%, BI= 1.00).
Halymenia maculata formed a fully supported sister relationship (ML= 100%, BI= 1.00)
with H. cf. dilatata which itself was strongly supported (ML= 96%, BI= 1.00). Clade 3
consisted of H. johorensis which was sister to H. plana, this relationship was highly
supported (ML= 98%, BI= 1.00).
Univers
ity of
Mala
ya

90
Figure 4.15: ML phylogeny inferred based on the rbcL sequences. Numbers above each
branch indicate bootstrap percentage (BP) for ML and posterior probability (PP) for BI.
Branches without value indicate no support (BP <50% or PP < 0.50) or that internode
did not occur in the ML or BI tree. Specimens of this study are highlighted in bold.
Scale bar = 0.02 substitution per site. Asterisk (*) denoted H. malaysiana samples (S4,
S5, SGAD0712156, SGAD1007001, SGAD1205198, SGAD1205185, SGAD1205213,
J1, J2, J3, J4, J5, J6, J7, J9, J10, J12, J13, J14, J16, J20, J22, J24, J25, J26, J30, J31,
J33)
Univers
ity of
Mala
ya

91
4.2.4.2 COI-5P
The COI-5P sequence alignment for phylogenetic reconstruction included 99
sequences which the identical sequences were collapsed to haplotypes with a total
length of 632 bp. Grateloupia elliptica and Prionitis filiformis were selected as
outgroups for phylogenetic analyses.
The phylogenetic tree in Figure 4.16 showed that the species of Halymenia formed a
monophyletic assemblage with full support (ML= 100%, BI= 1.00). Six species of
Halymenia examined in this study were distributed in three clades. Clade 1 was
unsupported (ML<50%, BI= 0.55) and comprised of H. malaysiana, H. maculata, H.
johorensis and H. californica. Halymenia malaysiana and H. maculata were each
resolved with maximum support (ML= 100%, BI= 1.00). Halymenia maculata was also
resolved as a sister to H. johorensis, albeit with a long terminal branching and without
any support.
The monophyly of Clade 2, which composed solely of H. durvillei, was not
supported (ML<50%, BI= 0.60). Two major unsupported subclades were observed
within H. durvillei clade. In Clade 3, the monotypic H. tondoana was highly supported
(ML= 92%, BI= 0.99) and grouped with its sister taxon, Halymenia sp. A with high
support (ML= 88%, BI= 1.00).
Univers
ity of
Mala
ya

92
Figure 4.16: ML phylogeny inferred based on the COI-5P sequences. Numbers above
each branch indicate bootstrap percentage (BP) for ML and posterior probability (PP)
for BI. Branches without value indicate no support (BP <50% or PP < 0.50) or that
internode did not occur in the ML or BI tree. Specimens of this study are highlighted in
bold. Scale bar = 0.04 substitution per site. Asterisk (*) denoted H. malaysiana samples
(SGAD1007001, S4, S5, SGAD1205198, SGAD1205185, SGAD1205213, J1, J2, J3,
J4, J5, J6, J7, J9, J10, J12, J13, J14, J16, J20, J22, J24, J25, J26, J30, J31, J33)
Univers
ity of
Mala
ya

93
4.2.4.3 UPA
The UPA dataset involved 67 sequences of 370 bp in length. The identical sequences
were reduced to haplotypes. Two species of Grateloupia were chosen as outgroups for
phylogenetic analyses.
The phylogenetic tree depicted as Figure 4.17 showed that the species of Halymenia
formed a monophyletic cluster with maximum support (ML= 100%, BI= 1.00). Six
species of Halymenia were resolved in two main clades except for H. johorensis.
Halymenia johorensis was on its own and inferred as a basal to the rest of Halymenia
species including in this study.
In Clade 1, two major unsupported subclades were observed. The first subclade was
resolved as a polytomy, consisting of H. durvillei, H. formosa, H. cearensis and H.
pseudofloresii. Meanwhile, the second subclade was also a polytomy, where Halymenia
sp. A grouped with its sister taxon, H. silviae and formed an unsupported relationship
with H. pinnatifida and H. tondoana. In Clade 2, a polytomy of three lineages was
obtained. The first lineage was made up of H. malaysiana specimens, resolved with full
support (ML= 100%, BI= 1.00). Meanwhile, the second lineage comprised of H.
maculata specimens that was resolved with maximum support (ML= 100%, BI= 1.00)
and the third lineage, H. stipitata from Hawaii.
Univers
ity of
Mala
ya

94
Figure 4.17: ML phylogeny inferred based on the UPA sequences. Numbers above
each branch indicate bootstrap percentage (BP) for ML and posterior probability (PP)
for BI. Branches without value indicate no support (BP <50% or PP < 0.50) or that
internode did not occur in the ML or BI tree. Specimens of this study are highlighted in
bold. Scale bar = 0.008 substitution per site.
Univers
ity of
Mala
ya

95
4.2.4.4 LSU (28S rDNA)
The phylogenetic matrix for partial LSU (28S rDNA) involved 29 taxa with 643 sites,
including a few introduced gaps. The identical sequences were collapsed to haplotypes.
Grateloupia lanceolata and Grateloupia turuturu were used as outgroups for this
dataset.
As shown in the ML phylogeny (Figure 4.18), the genus Halymenia was resolved as
monophyletic with maximum support (ML= 100%, BI= 1.00). However, the
phylogenetic relationship within species of Halymenia was unresolved. Both partial
LSU (28S rDNA) sequences of H. tondoana from the Philippines were identical to the
H. formosa GenBank sequences (HQ422446). A few H. durvillei samples formed a
subclade with weak support (ML= 58%, BI<0.50). H. durvillei, H. malaysiana, H.
formosa, H. pseudofloresii, H. tondoana and Halymenia sp. A formed an unsupported
assemblages and were sister to a clade comprised of H. plana, H. maculata and H.
johorensis (ML= 80%, BI= 0.66). Halymenia maculata from Malaysia were not
grouped with H. maculata from Australia.
Univers
ity of
Mala
ya

96
Figure 4.18: ML phylogeny inferred based on the partial LSU (28S rDNA) sequences.
Numbers above each branch indicate bootstrap percentage (BP) for ML and posterior
probability (PP) for BI. Branches without value indicate no support (BP <50% or PP <
0.50) or that internode did not occur in the ML or BI tree. Specimens of this study are
highlighted in bold. Scale bar = 0.006 substitution per site.
Univers
ity of
Mala
ya

97
4.2.5 Intraspecific genetic diversity of Halymenia malaysiana
The usefulness of the rbcL and the COI-5P as potential markers to infer intraspecific
genetic variation of the Halymenia malaysiana from Southeast Asia were assessed. 42
specimens of H. malaysiana were collected from five localities in Malaysia: Pulau
Merambong (Johor), Pulau Tinggi (Johor), Pulau Karindingan (Sabah), Tun Mustapha
Park (Sabah) and Pulau Besar (Malacca); four localities in the Philippines: Siargao
Island, Grande Island, Subic Bay and Busuanga Island; and one locality in Indonesia:
Gam Island.
4.2.5.1 Haplotype network analysis of Halymenia malaysiana for rbcL marker
A total of 42 specimens with 1241 characters of the rbcL gene were employed for
haplotype analysis. Eight rbcL haplotypes were recognized in H. malaysiana (Figure
4.19) with nucleotide divergences 0-0.50% (0-7 bp). Three haplotypes were observed in
Tun Mustapha Park, two haplotypes were observed in Gam Island and Busuanga Island
while only one haplotype was found in other seven localities. Haplotype R1 was the
most frequent, detected in 27 of the 42 individuals of H. malaysiana sequenced. All H.
malaysiana populations in Peninsular Malaysia (Pulau Besar, Pulau Tinggi and Pulau
Merambong) were represented by haplotype R1, whereas three haplotypes represented
the populations in East Malaysia where haplotypes R1, R2 and R5 were identified from
Tun Mustapha Park (TMP) and one haplotype R5 from Pulau Karindingan (Figure 4.8).
Haplotypes R2, R3 and R4 were each exclusive to the population in Tun Mustapha Park,
Siargao Island and Gam Island. Haplotype R2 differed from haplotypes R1 and R4 by
two nucleotides each but only one nucleotide differences from R3. Halymenia
malaysiana populations in the Philippines were characterized by haplotypes R3, R6, R7
and R8. Haplotype R6 was comprised of samples from Grande Island and Subic Bay
Univers
ity of
Mala
ya

98
while two haplotypes (R7 and R8) were detected from Busuanga Island. Samples from
Gam Island, Indonesia were made up of haplotype R1 and R4. Haplotype R5 connected
to haplotype R1 by three nucleotides, R6 and R7 each by one nucleotide, and R8 by two
nucleotides.
Figure 4.19: Statistical parsimony network for rbcL haplotypes of Halymenia
malaysiana. Each circle represents a rbcL haplotype. The size of circles corresponds to
the haplotype frequency. Each line between haplotypes, bars and/ or branch points
indicates one mutation step. Missing haplotype indicated by small crosses. Haplotypes
shaded according to the geographic origin.
Univers
ity of
Mala
ya

99
Table 4.5: A summary of the rbcL haplotype diversity of Halymenia malaysiana with
number of individuals (N) and number of haplotypes (Nh) from each location.
Populations N Nh Haplotype
Peninsular Malaysia
Pulau Merambong 23 1 R1
Pulau Tinggi 3 1 R1
Pulau Besar 1 1 R1
East Malaysia
Tun Mustapha Park 3 3 R1, R2, R5
Pulau Karindingan 1 1 R5
Philippines
Siargao Island 2 1 R3
Subic Bay 1 1 R6
Grande Island 1 1 R6
Busuanga Island 5 2 R7,R8
Indonesia
Gam Island 2 2 R1, R4
Total 42 8 R1-R8
4.2.5.2 Haplotype network analysis of Halymenia malaysiana for COI-5P marker
A total of 41 specimens with 610 characters of the COI-5P region were employed for
haplotype analysis. Nine COI-5P haplotypes were recognized in H. malaysiana (Figure
4.20) with nucleotide divergences of 0-1.64% (0-10 bp). Three haplotypes were
observed in Tun Mustapha Park, two haplotypes were observed in Siargao Island and
only one haplotype was found in other eight localities. Haplotype C1 was the most
frequent (27/41 individuals), found in the populations in Peninsular Malaysia (Pulau
Merambong, Pulau Tinggi and Pulau Besar). On the other hand, populations in East
Malaysia were represented by four haplotypes where three haplotypes (C2, C4 and C7)
were identified from Tun Mustapha Park (TMP) and one haplotype C6 exclusive to
Pulau Karindingan. Haplotype C4 connected to haplotypes C2 and C3 by three
nucleotides, C1 and C6 each by two nucleotides, and C5 by one nucleotide. The sample
from Gam Island, Indonesia corresponded to haplotype C7. Halymenia malaysiana
Univers
ity of
Mala
ya

100
populations in the Philippines were characterized by haplotypes C3, C5, C8 and C9.
Haplotype C5 was exclusive to the populations in Busuanga Island while haplotype C3
was identified from Grande Island and Subic Bay. Haplotype C8 differed from
haplotype C9 by four nucleotides, both were exclusive to Siargao Island.
Figure 4.20: Statistical parsimony network for COI-5P haplotypes of Halymenia
malaysiana. Each circle represents a COI-5P haplotype. The size of circles corresponds
to the haplotype frequency. Each line between haplotypes, bars and/ or branch points
indicates one mutation step. Missing haplotype indicated by small crosses. Haplotypes
shaded according to the geographic origin.
Univers
ity of
Mala
ya
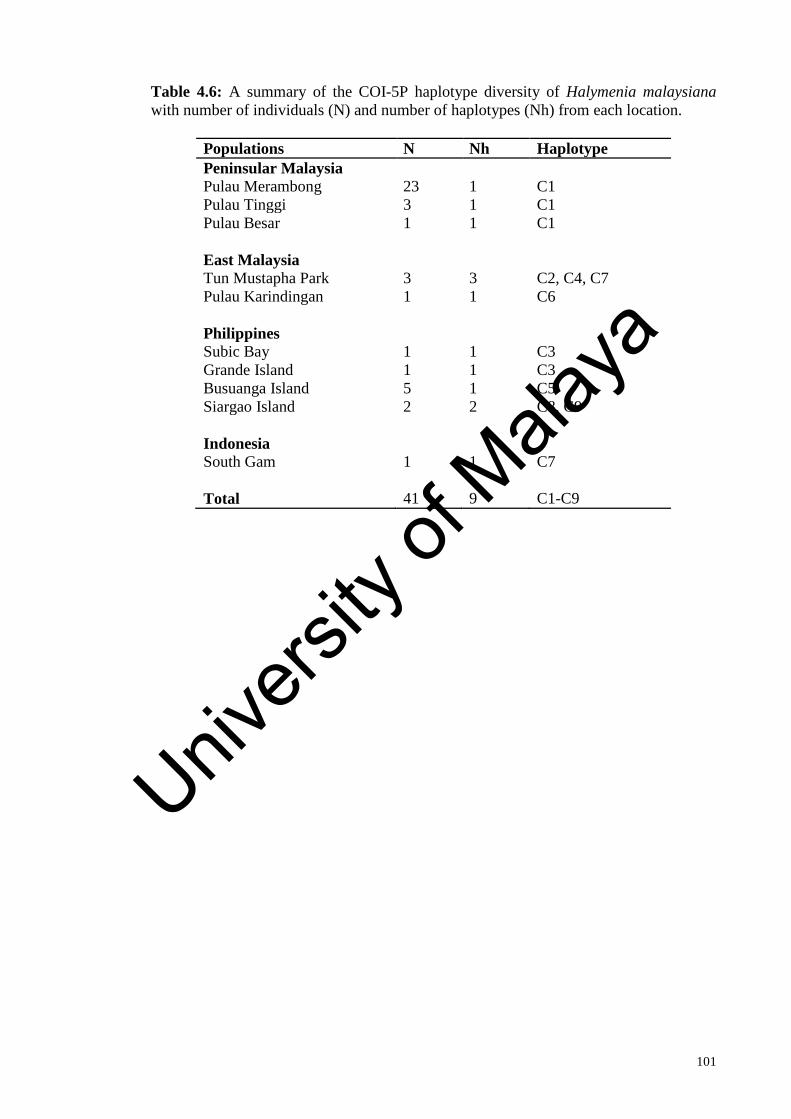
101
Table 4.6: A summary of the COI-5P haplotype diversity of Halymenia malaysiana
with number of individuals (N) and number of haplotypes (Nh) from each location.
Populations N Nh Haplotype
Peninsular Malaysia
Pulau Merambong 23 1 C1
Pulau Tinggi 3 1 C1
Pulau Besar 1 1 C1
East Malaysia
Tun Mustapha Park 3 3 C2, C4, C7
Pulau Karindingan 1 1 C6
Philippines
Subic Bay 1 1 C3
Grande Island 1 1 C3
Busuanga Island 5 1 C5
Siargao Island 2 2 C8, C9
Indonesia
South Gam 1 1 C7
Total 41 9 C1-C9
Univers
ity of
Mala
ya

102
CHAPTER 5: DISCUSSION
5.1 Morphological and anatomical studies
Generic concepts of Halymenia are based largely on the structure of auxiliary cell
ampullae (Chiang, 1970) and the presence of anticlinal medullary filaments (Abbott,
1967). Chiang (1970) proposed five types of auxiliary cell ampullae structure in order to
define the genera of the Halymeniaceae. Among the five types of auxiliary cell
ampullae, Halymenia-type auxiliary cell ampulla is flattish, expanded when mature and
generally regarded as having a primary ampullar filament bearing several branched
secondary filaments (Chiang, 1970). However, the development of the auxiliary cell
ampullae in many species of Halymenia is poorly known and calls for further study. In
addition, it is practically difficult to interpret the degree of branching of the auxiliary
ampullary structure and the extent of development of the pericarpial filaments (Maggs
and Guiry, 1982). According to Guimarães and Fujii (1998), the auxiliary cell ampullae
of the Brazilian species do not always conform with the scheme proposed by Chiang
(1970) for the genus. For instance, intermediate forms between the Halymenia-type and
Cryptonemia-type have been found by different authors (e.g. H. assymetrica Gargiulo,
de Masi & Tripodi by Gargiulo et al., 1986; H. maculata J. Agardh by Kawaguchi et al.,
2002). Moreover, collection of fertile specimens is often by chance, not all reproductive
stages were found in the collections. Some of the reproductive stages such as
carpogonial branches are rarely seen and difficult to interpret. Therefore, we agreed that
vegetative features should be emphasized rather than reproductive characters in genus-
level taxonomy (Kylin, 1956; Guiry and Irvine, 1974; Kraft, 1977).
Univers
ity of
Mala
ya

103
Presence of anticlinal filament is the most reliable taxonomic character of Halymenia
although it is not exclusive to Halymenia and can be found in other genera such as
Cryptonemia and Kallymenia (Abbott, 1967; Guimarães and Fujii, 1998). The presence
of stellate cells in the inner cortex and refractive ganglionic cells in the medulla are also
useful characteristic in recognition of Halymenia (Womersley and Lewis, 1994; De
Smedt et al., 2001; Guimarães and Fujii, 1998). However, stellate cells and refractive
ganglionic cells also present in the genera Weeksia and Kallymenia which are placed
under order Gigartinales (Abbott, 1967). Thus, a combination of features such as
presence of anticlinal filaments and refractive ganglionic cells in the medulla, stellate
cells in the inner cortex is important in the characterization of Halymenia.
The species identification of Halymenia is difficult due to its immense morphological
plasticity and few distinctive morphological features. Additionally, comparative studies
of Halymenia species are disconcerted by variations in features used for species
delimitation and insufficient information of taxonomically accepted species. A number
of morphological and anatomical features have been used as taxonomic characters for
species delineation of Halymenia but not all of them are equally reliable as diagnostic
characters. For instance, colour is not a good character for species identification of
Halymenia since the thalli colour become darker when dried and differentiated from the
colour observed in the field. We agree with Guimarães and Fujii (1998) who stated that
the degree of cystocarp protrusion, colour, the diameter and the number of medullary
filaments are highly variable features, and thus not useful in delineating Halymenia
species.
Our result agreed with previous studies that vegetative features are more diagnostic
than reproductive characters for species discrimination within Halymenia (Abbott, 1967;
Guimarães and Fujii, 1998; Hernández-Kantún et al., 2012). The present study showed
Univers
ity of
Mala
ya

104
that the combination of the following vegetative features is crucial for species
delineation in Halymenia: habit, thallus size, blade margins, order of branching,
branching pattern, presence or absence of a stipe, presence or absence of marginal
proliferations, presence or absence of surface proliferations or spines, blade thickness,
cortex thickness, shape of inner cortical cells, inner cortical cells size, and presence or
absence of refractive ganglionic cells. These vegetative features have been recognized
as taxonomically informative characters for species discrimination of Halymenia by
prior studies (Gargiulo et al., 1986; Guimarães and Fujii, 1998; De Smedt et al., 2001;
Ballantine and Ruiz, 2004; Hernández-Kantún et al., 2012; Tan et al., 2015; Azevedo et
al., 2016a; 2016b; Tan et al., 2017). The result of present study also suggested that the
size and shape of outer cortical cells may serve as taxonomic characters for
differentiating a few Halymenia species.
5.1.1 Morphological and anatomical distinction among Halymenia species
The specimens examined in this study were clearly assignable to the genus
Halymenia since these taxa agreed well with anatomical characters circumscribed for
Halymenia, including the presence of stellate cells in the inner cortex, anticlinal
filaments and refractive ganglionic cells in the medulla, and a medulla lax in young
parts and becoming denser in older parts (Balakrishnan, 1961; Abbott, 1967;
Womersley and Lewis, 1994).
Morphological evidence corroborated with our molecular findings in which seven
distinct species of Halymenia were present in our collections. Among the seven
Halymenia, four were previously described (H. durvillei, H. tondoana, H. cf. dilatata, H.
maculata), two were new species described from the current study (H. malaysiana, H.
johorensis) and one putative new species to be described (Halymenia sp. A). The
Univers
ity of
Mala
ya

105
highest number of Halymenia being examined is Halymenia malaysiana with 33
specimens, H. durvillei (32), H. johorensis (9), H. maculata (6), H. tondoana (3), H. cf.
dilatata (1), and the unidentified Halymenia sp. A (2). A few minor morphological
variations were observed between H. durvillei specimens. For instance, secondary axes
are arising as proliferations on the surface of the primary axes in some specimens and
surface spines present in some specimens but absent in other specimens. These
morphological variations were also reported by previous studies (De Smedt et al., 2001;
Hernández-Kantún et al., 2012). These results may suggest that H. durvillei consists of a
few cryptic species. Although molecular results in present study have supported the
existence of at least three species in H. durvillei complex, detailed data on morphology
and distribution were not yet available (Figures 4.10-4.11). Further examination of more
samples from different geographical regions and type specimen based on both
morphological and molecular analyses is needed to verify this finding. The
morphological and anatomical features of a fertile specimen collected from the
Phillippines conformed to the descriptions of H. dilatata in the literature (Hernández-
Kantún et al., 2012). However, its identity cannot be ensured since a detailed
examination on H. dilatata was not possible as the type specimen was unavailable for
the present study. Therefore, it was indicated here as H. cf. dilatata.
Tetrasporophytes and female gametophytes were found in all species examined in
this study except H. cf. dilatata and Halymenia sp. A in which only female
gametophytes were observed. Spermatangial gametophytes were not found and thus
remain to be described. Collection of fertile specimens is often by chance, thus not all
reproductive stages were found in collection. The developmental stages of the
carposporophyte were studied in detail in H. johorensis but not found in other species
examined in this study. The carpogonial branches, auxiliary cell ampullae and cystocarp
Univers
ity of
Mala
ya

106
development in many species of Halymenia are still poorly known and calls for further
study.
Among the seven Halymenia species studied in this study, three were branched
Halymenia (H. durvillei, H. tondoana, Halymenia sp. A) and four were foliose
Halymenia (H. cf. dilatata, H. maculata, H. malaysiana and H. johorensis). The
identification of branched and foliose Halymenia is fairly easy since they can
differentiated by observing their gross morphology. Thalli of branched Halymenia
branched up to several orders with different branching pattern and with abundant
marginal branchlets while thalli of foliose Halymenia are arising from a small discoid
holdfast with or without a stipe, abruptly expanding into broad blades. Branched
Halymenia in this study can be distinguished from one another based on five main
criteria, namely branching pattern, order of branching, cortex thickness, shape of inner
cortical cells and presence or absence of surface proliferations or spines. On the other
hand, foliose-type Halymenia are differentiated based on blade shape, blade margins,
presence or absence of surface proliferations or spines, cortex thickness, size and shape
of outer cortical cells and presence or absence of a stipe. All species of Halymenia in
this study can also be differentiated with following criteria: habit, thallus size, presence
or absence of marginal proliferations, blade thickness, size and shape of outer cortical
cells, size and shape of inner cortical cells, and presence or absence of refractive
ganglionic cells. Comparison of morphological and anatomical characters among
closely related branched and foliose Halymenia species in this study were summarized
in Table 5.1 and Table 5.2.
Morphological observations and molecular data both confirmed the presence of two
new species recognized from this study (H. malaysiana, H. johorensis) and one putative
new species, namely Halymenia sp. A. H. malaysiana, can be distinguished from other
Univers
ity of
Mala
ya

10
7
Table 5.1: Comparison of morphological and anatomical characters among closely related foliose Halymenia species in this study.
Characters H. malaysiana H. dilatata H. maculata H. porphyraeformis H. plana H. johorensis
Thallus size 6–25 cm high by 9–
33 cm wide
Up to 80 cm long and
50 cm wide
Up to 21 cm long and
38 cm wide
Up to 80 cm long, 50 cm
wide
10–30 cm high by 2–10
cm wide
22–40 cm high by 10–30
cm wide
Blade shape Oblong or
suborbicular
Irregular in outline Circular to irregularly
lobed
Irregularly circular to
elliptical
Irregularly divided into
broad lobed
Elliptical, oblong or
irregularly shaped
Blade margins Sinusoidal undulated,
sometimes minutely
dentate or deeply
cleft
Smooth or minutely
dentate
Irregularly to regularly
lobed (like jig-saw
pieces) or denticulate
Sinusoidal undulated Smooth to slightly
irregular
Irregularly dentate, cleft,
sometimes smooth
Blade surface Smooth, rarely with
orbicular
proliferations
Smooth, with small
orbicular proliferations
Rough, mottled, often
with proliferations,
spines or small
bladelets
Smooth, rarely with
orbicular proliferation
Smooth, mottled with
darker red, irregularly
shaped patches
Smooth or rugose with
perforations and
proliferations
Stipe Absent Present Present Present Present Present
Blade thickness 150–700 µm 285–400 µm 270–450 µm 65–410 µm 250–600 µm 120–400 µm
Cortex layers 6–7 layers 4–7 layers 5–7 layers 4–6 layers 3–6 layers 2–6 layers
Cortex thickness 20–70 µm
35–50 µm 50–100 µm 15–85 µm ND 15–45 µm
Shape of outer
cortical cells
Elongated Subspherical Elongated Subspherical to
elongated
Isodiametric Isodiametric
Outer cortical
cells size
8–13 µm long, 5–8
µm wide
6.5–11 µm long 7.5–35 µm long 6–10.5 µm across 3–7 µm across 3–6 µm long, 2–4 µm
wbide
Univers
ity of
Mala
ya

10
8
Table 5.1, continued
Characters H. malaysiana H. dilatata H. maculata H. porphyraeformis H. plana H. johorensis
Shape of inner
cortical cells
Rounded to
ellipsoidal
Irregularly stellate and
somewhat tangentially
flattened
Rounded to stellate Irregularly stellate Mostly not stellate Rounded to stellate
Inner cortical
cells size
5–13 µm in diameter 9.5–15 µm in diameter 11–33 µm in diameter 9.5–17.5 µm in diameter 15–22 µm across 4–12 µm in diameter
Refractive
ganglionic cells
(numbers of arm)
Present (4–6 arms) Common but not
abundant (mostly with
6 arms)
Present (5–9 arms) Present (5–8 arms) Prominent (ND) Few but present (3–5
arms)
References Tan et al., 2015; this
study
De Smedt et al., 2001 De Smedt et al. , 2001 De Smedt et al., 2001 Womersley and Lewis,
1994
Tan et al., 2017; this study
ND indicated non-available data
Univers
ity of
Mala
ya

10
9
Table 5.2: Comparison of morphological and anatomical characters among closely related branched Halymenia species in this study.
Characters H. durvillei H. tondoana H. hawaiiana Halymenia sp. A
Thallus size Up to 42 cm high Up to 22 cm high by 1.6–7cm
wide
10–30 cm high 16–26.5 cm long by 11–41 cm
wide
Branching pattern Subdichotomously,
trichotomously or irregularly
branched
Resembling dichotomous
bifurcations
Irregularly branched Irregularly branched, branches
and branchlets abruptly
constricted at bases
Order of branching Up to 7 Up to 7 Up to 3 Up to 4
Blade margins Rarely smooth, generally
dentate or laciniate
Lacerate, with proliferations Dentate and lacerate (not
laciniate)
Smooth with proliferations
Blade surface Smooth in young plants,
covered with spiny
proliferations in older thalli
Smooth with spots as granules
or macula but rare spiny
proliferations
Smooth with abundant spines
and macula
Smooth without spines
Stipe Present Present Present Present
Blade thickness 370–1625 µm 300–700 µm 250–800 µm 130–600 µm
Cortex layers 4–8 layers 4–5 layers 3–7 layers 4–6 layers
Cortex thickness 50–150 µm 25–60 µm 40–90 µm 25–65 µm
Shape of outer
cortical cells
Subspherical to elongated Elongated Elongated Subspherical to elongated
Univers
ity of
Mala
ya

11
0
Table 5.2, continued
Characters H. durvillei H. tondoana H. hawaiiana Halymenia sp. A
Outer cortical cells
size
8–19 µm long 5–7.5 µm long, 2.5–5 µm wide 5–15 µm long, 5–7.5 µm wide 5–9 µm long, 2–3 µm wide
Shape of inner
cortical cells
Rounded to stellate or
elongated parallel
Spherical Elongated parallel Rounded to stellate
Inner cortical cells
size
11–50 µm in diameter 7.5–20 µm in diameter 15–37.5 µm in diameter 5–15 µm in diameter
Refractive
ganglionic cells
(numbers of arm)
Abundant (6–9 arms) Common (5–6 arms) Common (4–5 arms) Present (-)
References De Smedt et al., 2001;
Kawaguchi et al., 2006
Hernández-Kantún et al., 2012 Hernández-Kantún et al., 2012 This study
Univers
ity of
Mala
ya

111
similar foliose Halymenia species in Southeast Asia, such as H. dilatata, H. maculata,
H. porphyraeformis and H. johorensis, by a combination of morphological and
anatomical characters (see Table 5.1). Morphologically, H. malaysiana resembles H.
dilatata and H. porphyraeformis, which display a smooth thallus surface and rarely
possess orbicular proliferations. However, H. malaysiana can be separated from H.
porphyraeformis and H. dilatata by its relatively small size (6-25 cm high by 9-33 cm
wide; H. dilatata and H. porphyraeformis: up to 80 cm long and 50 cm wide), relatively
thicker thallus (150-700 μm; H. dilatata: 285-400 µm; H. porphyraeformis: 65-410 µm),
oblong or suborbicular blade, and the lack of a stipe. Although H. malaysiana is similar
to H. maculata in terms of thallus size, the latter can be distinguished from the former
by having circular to irregularly lobed blades with jigsaw-like margin, rough mottled
surface with proliferations, spines or small bladelets, relatively thinner thallus but
thicker cortex (H. maculata: thallus 270-450 μm, cortex 50-100 µm; H. malaysiana:
thallus 150-700 μm, cortex 20-70 µm), longer elongated outer cortical cells (H.
maculata: 7.5-35 µm long; H. malaysiana: 8-13 µm long) and the presence of a firm
stipe. Halymenia johorensis is similar to H. malaysiana in thallus size (in regard to
blade width) and size of inner cortical cells, however these lineages differ by (1)
relatively thicker thallus in H. malaysiana (150-700 µm) than in H. johorensis (120-400
µm), (2) relatively thicker cortex in H. malaysiana (20-70 µm) than in H. johorensis
(15-45 µm), (3) elongated outer cortical cells in H. malaysiana (8-13 µm long, 5-8 µm
wide) in comparison to isodiametric outer cortical cells in H. johorensis (3-6 µm long,
2-4 µm wide) and (4) stipe absent in H. malaysiana but present in H. johorensis.
Halymenia malaysiana lacks a stipe, which is prominent in all other foliose Halymenia
species, including H. porphyraeformis, H. maculata, H. dilatata and H. johorensis.
Halymenia porphyraeformis has an unbranched stipe and H. maculata has a firm stipe
(De Smedt et al., 2001), H. johorensis has a narrow-cuneate stipe (Tan et al., 2017)
Univers
ity of
Mala
ya

112
while H. dilatata is characterized by the presence of a short but thick stipe (Zanardini,
1851; Zanardini, 1858). De Smedt et al. (2001) proposed the use of the stipe anatomy in
the genus Halymenia and suggested it might be a useful character in species
discrimination.
On the other hand, H. johorensis is similar to H. plana in having narrow-cuneate
stipe and one to two layers of isodiametric outer cortical cells. However, H. johorensis
can be differentiated from H. plana in having a relatively larger but thinner thallus (H.
johorensis: 22-40 cm high by 10-30 cm wide, 120-400 µm thick; H. plana: 10-30 cm
high by 2-10 cm wide, 250-600 µm thick), irregularly dentate and cleft margins, and
smaller inner cortical cells (H. johorensis: 4-12 µm in diameter; H. plana: 15-22 µm in
diameter). Furthermore, the blades of H. plana are irregularly divided into broad lobes
with surfaces mottled with dark red irregularly shaped patches. In contrast, the blades of
H. johorensis are elliptical, oblong or irregularly shaped with perforations and
proliferations. In addition, inner cortical cells of H. plana are mostly not stellate and the
species has prominent refractive ganglionic cells (Womersley and Lewis 1994: Figure
57B); whereas, H. johorensis has rounded to stellate inner cortical cells and few
refractive ganglionic cells.
Although H. johorensis resembles H. dilatata and H. porphyraeformis, it is
distinguished from the two by its relatively smaller thallus (H. johorensis: 22-40 cm
high by 10-30 cm wide; H. dilatata and H. porphyraeformis: up to 80 cm long and 50
cm wide), perforated blade, smaller isodiametric outer cortical cells (H. johorensis: 3-6
µm long; H. dilatata: 6.5-11 µm long, H. porphyraeformis: 6–10.5 µm long) and
smaller rounded to stellate inner cortical cells (H. johorensis: 4-12 µm in diameter; H.
dilatata: 9-15 µm in diameter; H. porphyraeformis: 9.5–17.5 µm in diameter).
Halymenia porphyraeformis is also differed from H. johorensis in displaying sinusoidal
Univers
ity of
Mala
ya

113
undulated margins and a thicker cortex (15–85 µm; 15–45 µm in H. johorensis).
Another foliose species, H. maculata, is readily distinguished from H. johorensis by its
circular to irregularly lobed margins, rough surface with proliferations, spines or small
bladelets, thicker cortex (50-100 µm; 15-45 µm in H. johorensis), longer elongated
outer cortical cells (7.5-35 µm; 3-6 µm in H. johorensis), and larger inner cortical cells
(11-33 µm in diameter; H. johorensis: 4-12 µm in diameter) despite their similarity in
terms of blade thickness and inner cortical cell shape.
Halymenia sp. A is similar to H. durvillei in having elongated outer cortical cells and
rounded to stellate inner cortical cells (see Table 5.2), however both lineages can be
differentiated by examining branching pattern and order of branching. Halymenia sp. A
irregularly branched up to 4 orders in which its branch and branchlets abruptly
constricted at bases; whereas, H. durvillei subdichotomously, trichotomously or
irregularly branched up to 7 orders in which its branch and branchlets with no
constricted at bases. Both lineages also differ by (1) relatively thinner thallus in
Halymenia sp. A (130-600 µm) than in H. durvillei (370-1625 µm), relatively thinner
cortex in Halymenia sp. A (25-65 µm) than in H. durvillei (50-150 µm), smaller outer
cortical cells in Halymenia sp. A (5-9 µm long) than in H. durvillei (8-19 µm long) and
relatively smaller inner cortical cells in Halymenia sp. A (5-15 µm in diameter) than in
H. durvillei (11-50 µm in diameter).
Although Halymenia sp. A resembles H. tondoana and H. hawaiiana, it is
distinguished from the two by its branching pattern and order of branching (Halymenia
sp. A: irregularly branched up to 4 orders in which its branch and branchlets abruptly
constricted at bases; H. tondoana: branching resembling dichotomous bifurcations and
branched up to 7 orders; H. hawaiiana: irregularly branched up to 3 orders).
Furtherrmore, Halymenia sp. A and H. hawaiiana differ by (1) spine absent on blade
Univers
ity of
Mala
ya

114
surface of Halymenia sp. A but present and abundant in H. hawaiiana, (2) relatively
thinner cortex in Halymenia sp. A (25-65 µm) than in H. hawaiiana (40-90 µm), (3)
smaller outer cortical cells in Halymenia sp. A (5-9 µm long, 2-3 µm wide) than H.
hawaiiana (5-15 µm long, 5-7.5 µm wide) and (4) smaller rounded to stellate inner
cortical cells in Halymenia sp. A (5-15 µm in diameter) in comparison to elongated
parallel inner cortical cells in H. hawaiiana (15-37.5 µm in diameter).
Despite above studies on Halymenia species, we believe that the diversity of
Halymenia species in Southeast Asia has not been fully explored. Additional sampling
from different localities, especially in the Coral Triangle which is well known as the
most biodiverse marine habitat, with detailed morphological and molecular
investigation is required to study the species richness of Halymenia in this region.
5.2 Molecular analyses
5.2.1 DNA extraction
Good quality DNA is indispensable for PCR amplification to gain reproducible
results. The choice of starting material used for DNA extraction is crucial in order to
obtain a good quality DNA. Silica gel-dried specimens are preferable starting material
for DNA extraction compared to herbarium specimens. Staats et al. (2011) reported that
DNA retrieval from aged herbarium specimens may be hindered due to natural DNA
degradation over time. Our studies concurred with the findings of Saunders (1993) that
dried specimens of red algae produced greater results for molecular analyses.
The amount of starting material used for DNA isolation is also one of the important
criteria in getting high quality DNA. Halymenia is rich in carrageenan (Freile-Pelegrin
et al., 2011; Kho et al., 2016). Isolation of DNA from Halymenia species is difficult due
Univers
ity of
Mala
ya

115
to the carrageenan gelation. In order to decrease the amount of gel formation during
isolation, only little starting material was used. In addition, constant vortexing during
incubation interval at 65˚C will promote homogenization and decrease gel formation.
Furthermore, longer durations for incubation on ice will reduce the co-isolation of
polysaccharides (Hu et al., 2004). In this study, the extraction of DNA using
commercially available kit proved beneficial since only small amount of starting
material was needed to gain high quality DNA.
5.2.2 PCR amplification
Based on the result of present study, the concentration of DNA isolated from
herbarium specimen was lower than the DNA isolated from silica gel-dried specimen.
Therefore, higher volume of DNA template from herbarium specimen was needed in
each PCR reaction. Most samples were successfully producing amplicons over one PCR
run. In contrast, a subsequent nested PCR was performed using primers with binding
sites internal to the primers used for the first PCR run to produce two smaller
overlapping fragments. This method worked well for the amplification of rbcL and
COI-5P sequences with extreme low concentration of DNA extracts. According to
Ekman (1999), nested PCR increased the yield and specificity of the amplification of
target DNA albeit time-consuming.
Present study preliminary test has found that COI-5P primers developed specifically
for red algae in Saunders (2005) did not work for specimens of Halymenia. This was
supported by Saunders (2008) which mentioned that red algal primers (Saunders, 2005)
scarcely work in some groups, for instance Halymeniales. In order to solve poor
amplification using abovementioned red algal primers and overcome contamination
problems, a lot of primer pairs were designed, tested and different combination of
Univers
ity of
Mala
ya

116
primers were used for DNA barcoding studies of different red algal species (e.g. Dilsea-
Neodilsea complex and Weeksia by Saunders, 2008; Gracilaria by Saunders, 2009;
Euthora by Clarkston and Saunders, 2010). The lack of universal amplification primers
is the major drawback of COI-5P (Clarkston and Saunders, 2010). In this study, two
degenerated primers (COI1F and COI1R) were designed based on the sequences
available in GenBank. The results showed that COI-5P sequences were easily amplified
and sequenced for most of the samples examined in this study using our newly designed
COI-5P primers. Both newly designed primers were also useful in sequencing COI-5P
sequences of red algal species in Halymeniaceae such as Cryptonemia and Grateloupia
(results not shown). Likewise, other COI-5P primers listed in Table 3.2 worked well for
Halymenia too.
5.2.3 Sequence analyses and molecular phylogenies
Both phylogenetic analyses ML and BI resulted in trees with near identical topology
for all well-supported nodes for the four genetic markers. Therefore, only ML trees
were presented (Figures 4.15-4.18). The rbcL phylogeny (Figure 4.15) agreed with
previous studies that demonstrated the polyphyly of Halymenia (Hernández-Kantún et
al., 2012; Tan et al., 2015; Azevedo et al., 2016a; 2016b; Tan et al., 2017), however a
taxonomic reassessment of the genera Gelinaria and Epiphloea is deferred until more
authentic materials are available for analysis. The rbcL results also indicated the
presence of seven distinct species of Halymenia in our collections. Among the seven
Halymenia, four were previously described (H. durvillei, H. tondoana, H. cf. dilatata, H.
maculata), two were new species described from the current study (H. malaysiana, H.
johorensis) and one putative new species to be described (Halymenia sp. A). Our result
corroborated with the findings by Hernández-Kantún et al. (2012) in which H. durvillei
Univers
ity of
Mala
ya

117
represent a complex of at least three distinct species. This was further supported by the
higher intraspecific rbcL sequence variation (0-2.56%) between H. durvillei populations
compared to other populations such as H. tondoana, 0.08-0.16%; H. maculata, 0.08-
0.48%; H. malaysiana, 0-0.58%; H. johorensis, 0% and Halymenia sp. A (see Table
4.1). However, we agreed with Hernández-Kantún et al. (2012) that referred these taxa
as the H. durvillei complex until more information is available and more detailed
reassessments of the species are possible using a broad range of samples from different
localities and type specimen.
Both rbcL analysis and pairwise distance (Figure 4.15; Table 4.1) clearly showed
that Halymenia sp. A are genetically distinct from other described Halymenia species.
The genetic variation between Halymenia sp. A and H. tondoana (2.24-2.34%) was
sufficiently high to warrant recognition as distinct species since the values were well
within the range of 0.7–10.6 % rbcL interspecific divergences observed in
Halymeniaceae (Wang et al., 2001; Gavio and Fredericq, 2002; Hernández-Kantún et al.,
2012; Tan et al., 2015; Azevedo et al., 2016a; 2016b). We have collected a fertile
individual that morphologically resembled with H. dilatata. However, its identity
cannot be ensured since no genetic records of authentic material are available within
GenBank for comparison. Therefore, it was indicated here as H. cf. dilatata. The record
of H. dilatata from Japan as shown by Wang et al. (2000) and its final disposition
require a further study as no detailed morphological description of this record has been
presented.
COI-5P, UPA and LSU (28S rDNA) sequences could not be successfully amplified
from the H. cf. dilatata sample identified in the present study so its phylogenetic
position cannot be established until sequence become available. Missing H. cf. dilatata
sequences in the COI-5P, UPA and LSU (28S rDNA) datasets did not affect the overall
Univers
ity of
Mala
ya

118
phylogeny. The phylogenetic reconstruction based on COI-5P and UPA markers
(Figures 4.16-4.17) showed that six distinct species of Halymenia were present in our
collections. Among the six Halymenia, three were previously described species (H.
durvillei, H. tondoana, H. maculata), two were new species described from the current
study (H. malaysiana, H. johorensis) and one unknown species (Halymenia sp. A).
UPA marker had lower resolution than COI-5P marker since the taxonomic status of
some branched Halymenia (H. durvillei and H. formosa) were not well resolved in UPA
phylogeny. The higher intraspecific divergence within H. durvillei (COI-5P: 0-5.70%;
UPA: 0-2.70%) compared to values formerly reported for Halymeniaceae (COI-5P: 0-
1.60%; UPA: 0-0.90%) (Yang and Kim, 2014; Manghisi et al., 2015) seemed to warrant
its recognition as a cryptic species complex. This assumption was also supported by the
findings of Saunders (2008) that COI-5P values higher than ca. 4% correspond to
distinct species. The interspecific divergence of Halymenia species (COI: 4.11-12.46%;
UPA: 1.08-5.14%) were comparable to values provided in previous studies (COI: 3.7-
14.0%; UPA: 0.6-4.6%) (Yang and Kim, 2014; Manghisi et al., 2015). The taxonomic
position and relationship of most species of Halymenia were not resolved in LSU (28S
rDNA) phylogeny (Figure 4.18), which can be attributed to the lack of available
GenBank sequences, in addition to the resolution limit of this marker. The drawback of
the LSU (28S rDNA) in phylogenetic reconstruction is noted and will not be discussed
in this context.
In general, the rbcL marker gave better resolution and clade support than other
markers. This is likely due to its large size which provides more characters for
phylogenetic analysis and greater number of available rbcL GenBank sequences
compared to other markers. Meanwhile, the other three markers gave lower resolution in
the order of COI-5P > UPA > LSU (28S rDNA) which is likely due to the scarcity of
GenBank sequences for taxonomically accepted species of Halymenia and the
Univers
ity of
Mala
ya

119
resolution limit of each marker. The pairwise distance of rbcL, COI-5P and UPA
markers (Table 4.1-4.3) were sufficiently high to discriminate species. Despite these
studies having improved our understanding of the phylogenetic relationship between
these red algae, many previously described Halymenia taxa are yet to be sequenced. In
order to better understand the species relationships, an effort to sequence the type
specimens is crucial.
5.2.4 Markers performance and potential DNA barcodes
In this study, four genetic markers were selected: the plastid rbcL and UPA, the
mitochondrial COI-5P and the nuclear partial LSU (28S rDNA). The aim of the marker
selection was to utilize DNA from different origin (i.e. plastid, mitochondrial and
nuclear) to study species diversity of Halymenia and elucidate the relationships between
Halymenia species in Malaysia, Thailand, Indonesia and the Philippines.
Accurate species identification is the utmost element in both molecular phylogenetics
and DNA barcoding. DNA barcoding has been proved useful for detection of cryptic
species, assignment of unknown samples to well-characterized taxa and delimitation of
species complexes (Hebert et al., 2003; Hebert et al., 2004; Newmaster and Ragupathy,
2009). An ideal DNA barcode should fulfil the criteria as follows: universality, short
length, ease of amplification and good species-resolving power (Hollingsworth et al.,
2009; Kucera and Saunders, 2012). The utility of each marker for molecular
phylogenetics and its potential as DNA barcode for Halymenia were assessed in this
study.
As mentioned before, the relationship of most species of Halymenia were not
resolved in LSU (28S rDNA) phylogeny. The same goes for Ceramium species in
Univers
ity of
Mala
ya

120
which LSU (28S rDNA) marker could not discriminate at the species level (Du et al.,
2014). Sherwood et al. (2010a) also demonstrated similar finding in which LSU (28S
rDNA) marker was not recommended for future red algal biodiversity surveys since it
could not resolved some members of the genera in Nemaliales. Among the four genetic
markers utilized in this study, LSU (28S rDNA) was the most conserved, followed by
UPA, rbcL and COI-5P. Despite partial LSU (28S rDNA) sequences were easily
amplified, this marker was too conserved for reliable species recognition. Since LSU
(28S rDNA) was not a reliable marker for both molecular systematics and DNA
barcoding studies in Halymenia, we did not pursue it further in this study.
The UPA marker has been proved useful for species level resolution (Sherwood and
Presting, 2007; Conklin et al., 2009; Clarkston and Saunders, 2010). It has also been
proposed as DNA barcoding marker for red algal species due to its primer universality
and the ease of data acquisition (Sherwood et al., 2010a; 2010b; Costa et al., 2012).
However, UPA was not sufficiently variable to differentiate species especially closely
related species (Sherwood et al., 2010b; Kucera and Saunders, 2012). This finding
concurred with our results in which UPA could not resolved some closely related
branched Halymenia, which most likely due to its low genetic variability (Kucera and
Saunders, 2012). As revealed by Ekrem et al. (2007), accuracy of species identification
is only noticeable and assessable in extensive datasets with reliable barcode libraries.
Thus, the limited number of reference UPA sequences for Halymeniaceae available in
GenBank might also impeded its application for species identification and DNA
barcoding. Our preliminary investigation did not support UPA as a suitable
phylogenetic marker or DNA barcode for Halymenia. Further studies are needed to
assess the feasibility of UPA marker for species delineation of Halymenia or as DNA
barcode when more GenBank reference sequences become available.
Univers
ity of
Mala
ya

121
The robustness of rbcL in resolving the phylogenetic relationships within
Halymeniaceae was consistent with previous studies (Wang et al., 2001; Kawaguchi et
al., 2004; Hernández-Kantún et al. 2012; Azevedo et al. 2016a; 2016b). The rbcL has
also been proposed as DNA barcode for land plants (Chase et al., 2005), green
macroalgae (Caulerpa by Kazi et al., 2013); green microalgae (Hadi et al., 2016). In this
study, the rbcL was shown to be less conserved (compared to 28S LSU and UPA) with
high species-resolving power which make it useful for species level phylogenetic
elucidation and thus recommended as a potential DNA barcode for Halymenia.
Additionally, the abundant and readily available rbcL sequences within the GenBank
(compared to COI-5P, UPA and 28S LSU sequences) uphold its potential as DNA
barcode.
On the other hand, the COI marker is widely recognized as the DNA barcode for red
algae (Robba et al., 2006; Saunders, 2008; Saunders, 2009; Clarkston and Saunders,
2010; Le Gall and Saunders, 2010). In this study, COI-5P was the most variable marker
among the four markers tested due to its high mutation rate. Its high genetic variability
(especially variable in the third codon position), make it useful in distinguishing
between even closely related species (Herbert et al., 2003). Our results agreed with the
findings of Azevedo et al. (2016a) on the use of COI-5P as a DNA barcode for
Halymenia.
Although COI-5P is recommended as a potential DNA barcode for Halymenia, its
relatively higher intraspecific variation and the limited number of reference COI-5P
sequences for Halymeniaceae available in GenBank may reduce the accuracy for
species identification. Likewise, the reduced genetic variation of rbcL may imply its
incapability to detect cryptic species and thus underestimate species richness of seaweed
(Hollingsworth et al., 2011; van Velzen et al., 2012). Therefore, two or more molecular
Univers
ity of
Mala
ya

122
markers are indeed essential to confirm the results. It is expected that the combined use
of rbcL and COI-5P would be better for species identification, discovery of cryptic
species, and phylogenetic reconstruction of Halymenia (Kucera and Saunders, 2012;
Yang and Kim, 2014).
In summary, we have illustrated that both rbcL and COI-5P markers are effective for
species identification in the genus Halymenia and also potential DNA barcodes for
Halymenia. On the contrary, both UPA and LSU (28S rDNA) markers are not suitable
markers for molecular phylogenetics and DNA barcoding studies in Halymenia.
5.2.5 Genetic diversity of Halymenia malaysiana
Little is known about the genetic diversity and phylogeographic structure of marine
seaweed in Southeast Asia despite their high biodiversity in the region. Several genetic
diversity studies of red algae have been undertaken (e.g. Gracilaria changii by Yow et
al., 2011 and Yow et al., 2013; Kappaphycus and Eucheuma by Lim et al., 2013;
Gracilaria salicornia and Gracilaria babae by Ng et al., 2015; Kappaphycus by
Dumilag et al., 2016; Pyropia acanthophora by Dumilag and Aguinaldo, 2017). Even
though molecular analyses have been employed to elucidate the taxonomic position of
Halymenia (Kawaguchi et al., 2006; Hernández-Kantún et al., 2012; Tan et al., 2015;
Azevedo et al., 2016a; 2016b; Tan et al., 2017), no study on intraspecific genetic
diversity and phylogeography of Halymenia has been implemented. This study has been
the first attempt in assessing the intraspecific genetic variation and phylogeographic
distribution of Halymenia focusing on H. malaysiana from three countries of Southeast
Asia, namely Malaysia, Indonesia and the Philippines. Halymenia malaysiana was
chosen for genetic diversity study due to its greater number of specimens available.
Both UPA and LSU (28S rDNA) markers are excluded in haplotype network analyses
Univers
ity of
Mala
ya

123
since they are not suitable for molecular phylogenetics of Halymenia as shown in results.
Only rbcL and COI-5P genetic markers were selected for haplotype network analyses.
The usefulness of plastid marker, rbcL (McIvor et al., 2001; Yang et al., 2008;
Destombe et al., 2010; Dumilag and Aguinaldo, 2017) and mitochondrial marker, COI-
5P (Yang et al., 2013a; Xie et al., 2015; Dumilag and Aguinaldo, 2017) in the studies of
genetic diversity on seaweeds have been addressed. In present study, the feasibility of
the rbcL and COI-5P as potential markers to infer intraspecific genetic variation of the
Halymenia malaysiana was assessed. Our results corroborated with the findings of
aforementioned studies that both rbcL and COI-5P are suitable markers for unravelling
intraspecies relationship and revealing patterns of genetic diversity in H. malaysiana.
However, COI-5P was more variable than rbcL, giving higher genetic variation (nine
COI-5P haplotypes compared to eight rbcL haplotypes; intraspecific pairwise
divergence of 0-1.64%, 0-10 bp for the COI-5P compared to 0-0.50%, 0-7 bp for the
rbcL).
All populations of H. malaysiana in Peninsular Malaysia (Pulau Besar, Pulau Tinggi
and Pulau Merambong) recovered the identical rbcL and COI-5P haplotypes (R1-C1),
showing no genetic divergence. Haplotypes R1 was the most common rbcL haplotypes
and accounted for 64% of the 42 samples examined. Meanwhile, haplotype C1 was the
most common COI-5P haplotypes and accounted for 66% of the 41 samples examined.
Diversity was highest in the Tun Mustapha Park in East Malaysia since three rbcL
haplotypes (R1, R2, and R5) and three COI-5P haplotypes (C2, C4 and C7) were
detected in the populations. These observations suggest that the populations of H.
malaysiana may have originated in Tun Mustapha Park. However, the origin of H.
malaysiana can only be ascertained with extensive sampling from a wider geographic
Univers
ity of
Mala
ya

124
range, especially in the Coral Triangle and also additional taxon sampling in each
locality.
An individual from Tun Mustapha Park, East Malaysia and another individual from
Gam Island, Indonesia were found to have rbcL haplotype R1 which prevailing in
Peninsular Malaysia. This genetic homogeneity could be due to the complex
oceanographic currents within this region. Current circulation has been proven as one of
the important characters in influencing the community structure of seaweeds (Uwai et
al., 2006a; Cheang et al., 2010; Buchanan and Zuccarello, 2012). The Southeast Asian
waters are mainly affected by monsoon seasons which will influence sea surface
currents circulation. During the Southwest Monsoon, the surface currents flow
northward in a clockwise direction, facilitating the dispersal of floating thalli or
fragments from Peninsular Malaysia to East Malaysia. In addition, genetic connectivity
between populations from Sabah, East Malaysia and Peninsular Malaysia is readily
since Borneo Island was part of the Sunda Shelf which connected to the Malay
Peninsula as a single landmass during the Pleistocene (Voris, 2000). Likewise, during
the Northeast Monsoon, the surface currents flow southward in an anticlockwise
direction (Wyrtki, 1961). Such surface current circulation patterns coupled with
directional current flows (e.g. Indonesian Thoughflow), could have promoted genetic
connectivity in Peninsular Malaysia and Indonesia.
On the other hand, the genetic connectivity that occurred among the different
lineages may also be a result of ship traffic (Broom et al., 2002; Xie et al., 2015). The
low level of genetic divergence observed in the haplotypes from Peninsular Malaysia
(R1-C1) and East Malaysia (R5-C4 and R2-C7) is likely since Borneo Island was
separated from the mainland peninsula by the South China Sea following the
submergence of Sundaland during the late Pleistocene (Voris, 2000).
Univers
ity of
Mala
ya

125
The results showed that the populations of H. malaysiana in the Philippines were
distinct from Malaysia and Indonesia. All four rbcL haplotypes (R3, R6, R7, R8) and
four COI-5P haplotypes (C3, C5, C8, C9) detected were endemic to the Philippines,
thus implying limited gene flow. This could be a result of the large geographical
separation and influenced by physical oceanographic processes. Genetic differentiation
was also evident in the Philippines populations of H. malaysiana where almost every
locality composed of unique haplotype except for Subic Bay and Grande Island shared
identical haplotypes (R6-C3) due to their relatively close proximity. Similar genetic
differentiation was also reported in red algae (Dumilag et al., 2017), where most of the
Philippine Pyropia acanthophora populations appeared specific to particular site. Deep
open ocean channels and variable currents among islands (Baums et al., 2005) are likely
to cause diversification in this region. Further studies involved more sampling sites in
this region may provide more evidence to elucidate the geographic pattern of the
Philippines populations of H. malaysiana.
The small sample size in this study did not allow us to infer the ancestral haplotypes
and elucidate phylogeographic pattern of H. malaysiana in detail. Additional sampling
from different geographic region is crucial as this will improve the distribution pattern
of the species (Zuccarello et al., 2006). The findings in the present study augmented our
understanding of the genetic diversity of H. malaysiana and offered a preliminary
outline of H. malaysiana distributions in Southeast Asia for future studies. This
information is also important for conservation and provides further insights into origin
and evolutionary relationships of H. malaysiana from Malaysia, Indonesia and the
Philippines.
Univers
ity of
Mala
ya

126
CHAPTER 6: CONCLUSION
6.1 General conclusion and appraisal of this study
Present study represents the first attempt to study the taxonomy of Halymenia
species from four countries of Southeast Asia, namely Malaysia, Thailand, Indonesia
and the Philippines, combining morphological features as well as molecular analyses.
This is also the first study in accessing the utility of UPA and LSU (28S rDNA) markers
in species delineation of Halymenia. Even though it is rather preliminary, it showed the
presence of seven species of Halymenia including H. durvillei, H. maculata, H.
tondoana, H. cf. dilatata with the description of two new species, H. malaysiana and H.
johorensis, and a potential new species, Halymenia sp. A. The findings in this study
augmented our understanding of species diversity of Halymenia in this warm water
region. It also provided new genetic records for species that had not previously been
analysed in a genetic context, for instance H. malaysiana and H. johorensis. The
following summarizes the significant results throughout the course of the study:
1) The genus Halymenia was shown to be polyphyletic, thus requiring more
detailed studies.
2) Both newly designed COI-5P primers (COI1F and COI1R) were useful in
sequencing COI-5P sequences of Halymenia and some red algal species in
Halymeniaceae.
3) The nuclear LSU (28S rDNA) genetic marker was shown to be the most
conserved in terms of genetic variations. Despite it was easy to acquire sequence data
for LSU (28S rDNA) marker, this marker was too conserved for reliable species
Univers
ity of
Mala
ya

127
recognition. It is not a reliable marker for both molecular phylogenetics and DNA
barcoding studies in Halymenia.
4) The plastid UPA genetic marker was not sufficiently variable to resolve some
closely related branched Halymenia, thus it is not a suitable phylogenetic marker or
DNA barcode for Halymenia. Further studies are needed to assess the feasibility of
UPA marker for species delineation of Halymenia or as DNA barcode when more
GenBank reference sequences become available.
5) The plastid-encoded rbcL gene was shown to be the best in terms of
phylogenetic resolution, less conserved (compared to 28S LSU and UPA) and has
abundant readily available rbcL sequences within the GenBank (compared to COI-5P,
UPA and 28S LSU sequences), thus a good marker for the systematics of Halymenia
and recommended as a potential DNA barcode for Halymenia.
6) The COI-5P genetic marker was shown to be the most variable in terms of
genetic variation, useful in distinguishing between even closely related species, thus a
promising DNA barcode for Halymenia.
7) The use of the molecular analyses in combination with morphological studies is
crucial for species identification.
8) Molecular results have supported the existence of at least three species in H.
durvillei complex but detailed data on morphology and distribution were not yet
available. Further examination of more samples from different geographical regions and
type specimen based on both morphological and molecular analyses is needed to verify
this finding.
9) Combination of the following vegetative features is crucial for species
delineation in Halymenia: habit, thallus size, blade margins, order of branching,
Univers
ity of
Mala
ya

128
branching pattern, presence or absence of a stipe, presence or absence of marginal
proliferations, presence or absence of surface proliferations or spines, blade thickness,
cortex thickness, shape and size of outer cortical cells, shape and size of inner cortical
cells, and presence or absence of refractive ganglionic cells.
10) Both morphological evidence and molecular findings led to the description of
two new species of Halymenia, namely H. malaysiana (Tan et al., 2015) and H.
johorensis (Tan et al., 2017); a potential new species of Halymenia, Halymenia sp. A.
This revealed that species of Halymenia in Southeast Asia is far richer than previously
recognized.
11) Both rbcL and COI-5P genetic markers were shown to be suitable for resolving
intraspecific genetic variation and are reliable markers for the study of genetic diversity
and phylogeographic distribution in Halymenia species i.e. H. malaysiana.
12) A statistical parsimony network of H. malaysiana yielded eight rbcL haplotypes
(R1-R8) and nine COI-5P haplotypes (C1-C9) which were closely related. COI-5P was
more variable than rbcL, giving higher genetic variation (nine COI-5P haplotypes
compared to eight rbcL haplotypes; intraspecific pairwise divergence of 0-1.64%, 0-10
bp for the COI-5P compared to 0-0.50%, 0-7 bp for the rbcL).
13) Haplotype R1 and C1 were the most common rbcL and COI-5P haplotypes of H.
malaysiana found in Peninsular Malaysia. The populations of H. malaysiana may have
originated in Tun Mustapha Park as contributing the highest intra-population diversity.
However, the origin of H. malaysiana can only be ascertained with extensive sampling
from a wider geographic range, especially in the Coral Triangle and also additional
taxon sampling in each locality.
Univers
ity of
Mala
ya

129
14) Current circulation, Pleistocene events, anthropogenic activities and natural
distributions have been postulated to affect the dispersal, in turn the genetic population
structure of H. malaysiana.
First and third alternative hypotheses and second null hypothesis are accepted
whereby a) not all morphological features were equally reliable as diagnostic characters,
b) identification based on molecular phylogenies were coherent with morphological
characters and c) phylogenies of different molecular genetic markers were not
congruent and do not have similar levels of resolution.
6.2 Future studies on Halymenia
We believe that the species diversity of Halymenia in this region is still
underestimated. Additional sampling from different geographic regions with detailed
morphological and molecular investigation is required to provide a greater
understanding of the phylogenetic relationships and species richness in this genus. The
putative new species discovered in this study will be described formally when more
specimens become available.
The sampling, morphological and genetic data of all taxonomically accepted species
will greatly assist in the systematics of Halymenia. Sequencing of type specimens in
particular H. dilatata, H. durvillei, Gelinaria ulvoidea and Epiphloea bullosa is crucial
to clarify their ambiguous taxonomic position. If there is lack of authentic materials,
sampling in type locality with the designation of neotype may be a feasible solution.
Further taxonomic study on the branched Halymenia is imperative in particular to reveal
the cryptic species found in H. durvillei complex.
Univers
ity of
Mala
ya

130
The development of the auxiliary cell ampullae in many species of Halymenia is
poorly known. Hence, subsequent taxonomic research should be more focused on the
study of female reproductive structures such as the developmental stages of the
carposporophyte when well-preserved specimens are available.
Univers
ity of
Mala
ya

131
REFERENCES
Abbott, I. A. (1967). Studies in some foliose red algae of the Pacific Coast I.
Cryptonemiaceae. Journal of Phycology, 3(3), 139-149.
Agardh, C. A. (1817). Synopsis algarum Scandinaviae: adjecta dispositione universali
algarum. Lundae: Ex officina Berlingiana.
Agardh, J. G. (1842). Algae maris Mediterranei et Adriatici, observationes in diagnosin
specierum et dispositionem generum. Paris: Apud Fortin, Masson et Cie.
Alberto, F., Santos, R., & Leitão, J. M. (1997). DNA extraction and RAPD markers to
assess the genetic similarity among Gelidium sesquipedale (Rhodophyta)
populations. Journal of Phycology, 33(4), 706-710.
Alberto, F., Santos, R., & Leitão, J. M. (1999). Assessing patterns of geographic
dispersal of Gelidium sesquipedale (Rhodophyta) through RAPD differentiation
of populations. Marine Ecology Progress Series, 191, 101-108.
Andreakis, N., Procaccini, G., Maggs, C., & Kooistra, W. H. C. F. (2007).
Phylogeography of the invasive seaweed Asparagopsis (Bonnemaisoniales,
Rhodophyta) reveals cryptic diversity. Molecular Ecology, 16(11), 2285-2299.
Atmadja, W. S., & Prud’homme van Reine, W. F. (2012). Checklist of the seaweed
species biodiversity of Indonesia with their distribution and classification:
Rhodophyceae (pp. 1-72). Jakarta: Coral Reef Information and Training Centre.
Coral Reef Rehabilitation and Management Programme. Indonesian Institute of
Sciences (LIPI).
Avise, J. C. (1994). Molecular markers, natural history and evolution. New York:
Chapman & Hall.
Azevedo, C. A. A., Cassano, V., & Oliveira, M. C. (2016a). Diversity of branched
Halymenia (Halymeniales, Rhodophyta) species on the Brazilian coast:
molecular and morphological analyses reveal three new species. Phycologia,
55(4), 431-444.
Azevedo, C. A. A., Cassano, V., & Oliveira, M. C. (2016b). Phylogenetic relationships
among Halymenia (Halymeniaceae, Rhodophyta) species on the Brazilian coast
with description of Halymenia cearensis sp. nov. Phytotaxa, 280(3), 241-258.
Univers
ity of
Mala
ya

132
Bagley, M J., Franson, S. E., Christ, S. A., Waits, E. R., & Toth, G. P. (2002). Genetic
diversity as an indicator of ecosystem condition and sustainability: utility for
regional assessments of stream condition in the eastern United States.
Cincinnati, Ohio: U.S. Environmental Protection Agency.
Balakrishnan, M. S. (1960). Reproduction in some Indian red algae and their taxonomy.
In P. Kachroo (Ed.), Proceedings of the Symposium on Algology (pp. 85-98).
New Delhi: Indian Council of Agricultural Research.
Balakhrishnan, M. S. (1961). Studies on Indian Cryptonemiales—III. Halymenia C. A.
Ag. Journal of the Madras University, Section B, 31(2), 183-217.
Baldwin, B. G., Sanderson, M. J., Porter, J. M., Wojciechowski, M. F., Campbell, C. S.,
& Donoghue, M. J. (1995). The ITS region of nuclear ribosomal DNA: a
valuable source of evidence on angiosperm phylogeny. Annals of the Missouri
Botanical Garden, 82, 247-277.
Ballantine, D. L., & Ruiz, H. (2004). Halymenia mirabilis sp. nov. (Cryptonemiales,
Rhodophyta), a spectacular alga from Grenada, Caribbean Sea. Phycologia,
43(1), 75-78.
Bardakci, F. (2001). Random amplified polymorphic DNA (RAPD) markers. Turkish
Journal of Biology, 25(2), 185-196.
Baums, I. B., Miller, M. W., & Hellberg, M. E. (2005). Regionally isolated populations
of an imperiled Caribbean coral, Acropora palmata. Molecular Ecology, 14(5),
1377-1390.
Bellorin, A. M., Oliveira, M. C., & Oliveira, E. C. (2002). Phylogeny and systematics of
the marine algal family Gracilariaceae (Gracilariales, Rhodophyta) based on
SSU rDNA and ITS sequences of Atlantic and Pacific species. Journal of
Phycology, 38(3), 551-563.
Bermejo Román, R., Alvarez-Pez, J. M., Acién Fernández, F. G., & Molina Grima, E.
(2002). Recovery of pure B-phycoerythrin from the microalga Porphyridium
cruentum. Journal of Biotechnology, 93(1), 73-85.
Bhattacharya, D., & Medlin, L. (1998). Algal phylogeny and the origin of land plants.
Plant Physiology, 116(1), 9-15.
Univers
ity of
Mala
ya

133
Bird, C. J., & Rice, E. L. (1990). Recent approaches to the taxonomy of the
Gracilariaceae (Gracilariales, Rhodophyta) and the Gracilaria verrucosa
problem. Hydrobiologia, 204(1), 111-118.
Boney, A. D., & Corner, E. D. S. (1960). A possible function of phycoerythrin in
intertidal red algae. Nature, 188(4755), 1042-1043.
Boudouresque, C. F., & Verlaque, M. (2002). Biological pollution in the Mediterranean
Sea: invasive versus introduced macrophytes. Marine Pollution Bulletin, 44(1),
32-38.
Bown, P., Plumb, J., Sánchez-Baracaldo, P., Hayes, P. K., & Brodie, J. (2003).
Sequence heterogeneity of green (Chlorophyta) endophytic algae associated with
a population of Chondrus crispus (Gigartinaceae, Rhodophyta). European
Journal of Phycology, 38(2), 153-163.
Brodie, J. A., & Irvine, L. M. (2003). Seaweeds of the British Isles Volume 1
Rhodophyta part 3B. Bangiophycidae. London: Intercept, Andover.
Broom, J. E., Nelson, W. A., Yarish, C., Jones, W. A., Aguilar Rosas, R., & Aguilar
Rosas, L. E. (2002). A reassessment of the taxonomic status of Porphyra
suborbiculata, Porphyra carolinensis and Porphyra lilliputiana (Bangiales,
Rhodophyta) based on molecular and morphological data. European Journal of
Phycology, 37(2), 227-235.
Brown, T. A. (2016). Gene cloning and DNA analysis: an introduction (6th ed.). United
Kingdom: John Wiley & Sons.
Bruns, T. D., White, T. J., & Taylor, J. W. (1991). Fungal molecular systematics.
Annual Review of Ecology and Systematics, 22(1), 525-564.
Buchanan, J., & Zuccarello, G. C. (2012). Decoupling of short- and long- distance
dispersal pathways in the endemic New Zealand seaweed Carpophyllum
maschalocarpum (Phaeophyceae, Fucales). Journal of Phycology, 48(3), 518-
529.
Campo, V. L., Kawano, D. F., da Silva, D. B., & Carvalho, I. (2009). Carrageenans:
Biological properties, chemical modifications and structural analysis–A review.
Carbohydrate Polymers, 77(2), 167-180.
Univers
ity of
Mala
ya

134
Candia, A., González, M. A., Montoya, R., Gómez, P., & Nelson, W. (1999).
Comparison of ITS RFLP patterns of Gracilaria (Rhodophyceae, Gracilariales)
populations from Chile and New Zealand and an examination of interfertility of
Chilean morphotypes. Journal of Applied Phycology, 11(2), 185-193.
Cardinale, B. J., Duffy, J. E., Gonzalez, A., Hooper, D. U., Perrings, C., Venail, P., . . .
Wardle, D. A. (2012). Biodiversity loss and its impact on humanity. Nature,
486(7401), 59-67.
Carlile, A. L., & Sherwood, A. R. (2013). Phylogenetic affinities and distribution of the
Hawaiian freshwater red algae (Rhodophyta). Phycologia, 52(3), 309-319.
Chan, C. X., Ho, C. L., & Phang, S. M. (2006). Trends in seaweed research. Trends in
Plant Science, 11(4), 165-166.
Chase, M. W., Salamin, N., Wilkinson, M., Dunwell, J. M., Kesanakurthi, R. P., Haidar,
N., & Savolainen, V. (2005). Land plants and DNA barcodes: short-term and
long-term goals. Philosophical Transactions of the Royal Society B: Biological
Sciences, 360(1462), 1889-1895.
Cheang, C. C., Chu, K. H., Fujita, D., Yoshida, G., Hiraoka, M., Critchley, A., . . . Ang,
P. O. (2010). Low genetic variability of Sargassum muticum (Phaeophyceae)
revealed by a global analysis of native and introduced populations. Journal of
Phycology, 46(6), 1063-1074.
Chen, C. S., & Chen, C. Y. (1996). Using red alga Halymema microcarpa as a biofilter
in an integrated aquarium system. Acta Oceanographica Taiwanica, 35, 357-367.
Chiang, Y. M. (1970). Morphological studies of red algae of the family
Cryptonemiaceae. Berkeley: University of California Press.
Cho, G. Y., Kogame, K., Kawai, H., & Boo, S. M. (2007). Genetic diversity of
Scytosiphon lomentaria (Scytosiphonaceae, Phaeophyceae) from the Pacific and
Europe based on RuBisCO large subunit and spacer, and ITS nrDNA sequences.
Phycologia, 46(6), 657-665.
Clarkston, B. E., & Saunders, G. W. (2010). A comparison of two DNA barcode
markers for species discrimination in the red algal family Kallymeniaceae
(Gigartinales, Florideophyceae), with a description of Euthora timburtonii sp.
nov. Botany, 88(2), 119-131.
Univers
ity of
Mala
ya

135
Clayden, S. L., & Saunders, G. W. (2010). Recognition of Rubrointrusa membranacea
gen. et comb. nov., Rhodonematella subimmersa gen. et comb. nov.(with a
reinterpretation of the life history) and the Meiodiscaceae fam. nov. within the
Palmariales (Rhodophyta). Phycologia, 49(3), 283-300.
Clement, M., Posada, D., & Crandall, K. A. (2000). TCS: a computer program to
estimate gene genealogies. Molecular Ecology, 9(10), 1657-1659.
Collins, F. S., & Howe, M. A. (1916). Notes on species of Halymenia. Bulletin of the
Torrey Botanical Club, 43(4), 169-182.
Conklin, K. Y., Kurihara, A., & Sherwood, A. R. (2009). A molecular method for
identification of the morphologically plastic invasive algal genera Eucheuma
and Kappaphycus (Rhodophyta, Gigartinales) in Hawaii. Journal of Applied
Phycology, 21(6), 691-699.
Costa, F. O., & Antunes, P. M. (2012). The contribution of the Barcode of Life initiative
to the discovery and monitoring of Biodiversity. In A. Mendonca, A. Cunha & R.
Chakrabarti (Eds.), Natural Resources, Sustainability and Humanity (pp. 37-68).
Netherlands: Springer.
Costello, M. J., & Wilson, S. P. (2011). Predicting the number of known and unknown
species in European seas using rates of description. Global Ecology and
Biogeography, 20(2), 319-330.
Coyer, J. A., Hoarau, G., Stam, W. T., & Olsen, J. L. (2004). Geographically specific
heteroplasmy of mitochondrial DNA in the seaweed, Fucus serratus
(Heterokontophyta: Phaeophyceae, Fucales). Molecular Ecology, 13(5), 1323-
1326.
Curiel, D., Bellemo, G., Marzocchi, M., Scattolin, M., & Parisi, G. (1998). Distribution
of introduced Japanese macroalgae Undaria pinnatifida, Sargassum muticum
(Phaeophyta) and Antithamnion pectinatum (Rhodophyta) in the Lagoon of
Venice. Hydrobiologia, 385(1), 17-22.
D’Archino, R., Nelson, W. A., & Zuccarello, G. C. (2014). Amalthea and Galene, two
new genera of Halymeniaceae (Rhodophyta) from New Zealand. Botanica
Marina, 57(3), 185-201.
Dawes, C. J. (1998). Marine Botany (2nd ed.). New York: John Wiley & Sons Inc.
Univers
ity of
Mala
ya

136
Daly, D. C., Cameron, K. M., & Stevenson, D. W. (2001). Plant systematics in the age
of genomics. Plant Physiology, 127(4), 1328-1333. De Clerck, O., Bogaert, K. A., & Leliaert, F. (2012). Diversity and evolution of algae:
primary endosymbiosis. Advances in Botanical Research, 64, 55-86.
De Clerck, O., Gavio, B., Fredericq, S., Barbara, I., & Coppejans, E. (2005).
Systematics of Grateloupia filicina (Halymeniaceae, Rhodophyta), based on
rbcL sequence analyses and morphological evidence, including the
reinstatement of G. minima and the description of G. capensis sp. nov. Journal
of Phycology, 41(2), 391-410.
De Clerck, O., Guiry, M. D., Leliaert, F., Samyn, Y., & Verbruggen, H. (2013). Algal
taxonomy: a road to nowhere? Journal of Phycology, 49(2), 215-225.
de Senerpont Domis, L. N., Famà, P., Bartlett, A. J., Prud'homme van reine, W. F.,
Espinosa, C. A., & Trono Jr., G. C. (2003). Defining taxon boundaries in
members of the morphologically and genetically plastic genes Caulerpa
(Caulerpales, Chlorophyta). Journal of Phycology, 39(6), 1019-1037.
De Smedt, G., De Clerck, O., Leliaert, F., Coppejans, E., & Liao, L. (2001).
Morphology and systematics of the genus Halymenia C. Agardh (Halymeniales,
Rhodophyta) in the Philippines. Nova Hedwigia, 73(3-4), 293-322.
Destombe, C., & Douglas, S. E. (1991). RuBisCO spacer sequence divergence in the
rhodophyte alga Gracilaria verrucosa and closely related species. Current
Genetics, 19(5), 395-398.
Destombe, C., Valero, M., & Guillemin, M. L. (2010). Delineation of two sibling red
algal species, Gracilaria gracilis and Gracilaria dura (Gracilariales,
Rhodophyta), using multiple DNA markers: resurrection of the species G. dura
previously described in the Northern Atlantic 200 years ago. Journal of
Phycology, 46(4), 720-727.
Dhargalkar, V. K., & Kavlekar, D. P. (2004). Seaweeds- a field manual. Dona Paula,
Goa: National Institute of Oceanography.
Dixon, P. S. (1973). Biology of the Rhodophyta. Edinburgh: Oliver and Boyd.
Dowell, K. (2008). Molecular Phylogenetics: An introduction to computational methods
and tools for analyzing evolutionary relationships. Math, 500, 18.
Univers
ity of
Mala
ya

137
Doyle, J. J., & Luckow, M. A. (2003). The rest of the iceberg. Legume diversity and
evolution in a phylogenetic context. Plant Physiology, 131(3), 900-910.
Du, G., Wu, F., Guo, H., Xue, H., & Mao, Y. (2015). DNA barcode assessment of
Ceramiales (Rhodophyta) in the intertidal zone of the northwestern Yellow Sea.
Chinese Journal of Oceanology and Limnology, 33(3), 685.
Dumilag, R. V., & Aguinaldo, Z.-Z. A. (2017). Genetic differentiation and distribution
of Pyropia acanthophora (Bangiales, Rhodophyta) in the Philippines. European
Journal of Phycology, 52(1), 104-115.
Dumilag, R. V., Orosco, F. L., & Lluisma, A. O. (2016). Genetic diversity of
Kappaphycus species (Gigartinales, Rhodophyta) in the Philippines. Systematics
and Biodiversity, 14(5), 441-451.
Ekman, S. (1999). PCR optimization and troubleshooting, with special reference to the
amplification of ribosomal DNA in lichenized fungi. Lichenologist, 31(5), 517-
531.
Ekrem, T., Willassen, E., & Stur, E. (2007). A comprehensive DNA sequence library is
essential for identification with DNA barcodes. Molecular Phylogenetics and
Evolution, 43(2), 530-542.
Famà, P., Jousson, O., Zaninetti, L., Meinesz, A., Dini, F., Di Giuseppe, G., . . .
Pawlowski, J. (2002). Genetic polymorphism in Caulerpa taxifolia
(Ulvophyceae) chloroplast DNA revealed by a PCR‐based assay of the invasive
Mediterranean strain. Journal of Evolutionary Biology, 15(4), 618-624.
Fenoradosoa, T. A., Delattre, C., Laroche, C., Wadouachi, A., Dulong, V., Picton,
L., . . . Michaud, P. (2009). Highly sulphated galactan from Halymenia durvillei
(Halymeniales, Rhodophyta), a red seaweed of Madagascar marine coasts.
International Journal of Biological Macromolecules, 45(2), 140-145
Féral, J.-P. (2002). How useful are the genetic markers in attempts to understand and
manage marine biodiversity? Journal of Experimental Marine Biology and
Ecology, 268(2), 121-145.
Figueroa, F. L., Korbee, N., Clerck, O. d., Bárbara, I., & Gall, E. A. (2007).
Characterization of Grateloupia lanceola (Halymeniales, Rhodophyta), an
obscure foliose Grateloupia from the Iberian Peninsula, based on morphology,
comparative sequence analysis and mycosporine-like amino acid composition.
European Journal of Phycology, 42(3), 231-242.
Univers
ity of
Mala
ya

138
Fisher, R. A. (1930). The genetical theory of natural selection. Oxford: The Clarendon
Press.
Fletcher, R. L., & Farrell, P. (1999). Introduced brown algae in the North East Atlantic,
with particular respect to Undaria pinnatifida (Harvey) Suringar. Helgoländer
Meeresuntersuchungen, 52, 259-275.
Frankham, R., Ballou, J. D., & Briscoe, D. A. (2004). A primer of conservation genetics.
Cambridge, UK: Cambridge University Press.
Freeman, S., & Herron, J. C. (1998). Evolutionary analysis. Upper Saddle River, New
Jersey: Prentice-Hall.
Freile-Pelegrin, Y., Azamar, J. A, & Robledo, D. (2011). Preliminary characterization
of carrageenan from the red seaweed Halymenia floresii. Journal of Aquatic
Food Product Technology, 20(1), 73-83.
Freshwater, D. W., Fredericq, S., & Bailey, J. C. (1999). Characteristics and utility of
nuclear-encoded large-subunit ribosomal gene sequences in phylogenetic studies
of red algae. Phycological Research, 47(1), 33-38.
Freshwater, D. W., Fredericq, S., Butler, B. S., Hommersand, M. H., & Chase, M. W.
(1994). A gene phylogeny of the red algae (Rhodophyta) based on plastid rbcL.
Proceedings of the National Academy of Sciences, 91(15), 7281-7285.
Freshwater, D. W., & Rueness, J. (1994). Phylogenetic relationships of some European
Gelidium (Gelidiales, Rhodophyta) species, based on rbcL nucleotide sequence
analysis. Phycologia, 33(3), 187-194.
Garcia, J. S., Palacios, V., & Roldán, A. (2016). Nutritional potential of four seaweed
species collected in the Barbate Estuary (Gulf of Cadiz, Spain). Journal of
Nutrition & Food Sciences, 6(3), 505.
Gargiulo, G. M., De Masi, F., & Tripodi, G. (1986). Structure and reproduction of
Halymenia asymmetrica sp. nov.(Rhodophyta) from the Mediterranean Sea.
Phycologia, 25(2), 144-151.
Gargiulo, G. M., Morabito, M., & Manghisi, A. (2013). A re-assessment of reproductive
anatomy and postfertilization development in the systematics of Grateloupia
(Halymeniales, Rhodophyta). Cryptogamie, Algologie, 34(1), 3-35.
Univers
ity of
Mala
ya

139
Gavio, B., & Fredericq, S. (2002). Grateloupia turuturu (Halymeniaceae, Rhodophyta)
is the correct name of the non-native species in the Atlantic known as
Grateloupia doryphora. European Journal of Phycology, 37(3), 349-359.
Geraldino, P. J. L., Yang, E.-C., & Bu, S.-M. (2006). Morphology and molecular
phylogeny of Hypnea flexicaulis (Gigartinales, Rhodophyta) from Korea. Algae,
21(4), 417-423.
Godínez-Ortega, J. L., Snoeijs, P., Robledo, D., Freile-Pelegrín, Y., & Pedersén, M.
(2008). Growth and pigment composition in the red alga Halymenia floresii
cultured under different light qualities. Journal of Applied Phycology, 20(3),
253-260.
Goff, L. J., Moon, D. A., & Coleman, A. W. (1994). Molecular delineation of species
and species relationships in the red algal agarophytes Gracilariopsis and
Gracilaria (Gracilariales). Journal of Phycology, 30(3), 521-537.
Guerra-García, J. M., Espinosa Torre, F., & García Gómez, J. C. (2008). Trends in
taxonomy today: an overview about the main topics in taxonomy. Zoológica
Baetica, 19, 15-49.
Guillemin, M.-L., Akki, S. A., Givernaud, T., Mouradi, A., Valero, M., & Destombe, C.
(2008). Molecular characterisation and development of rapid molecular methods
to identify species of Gracilariaceae from the Atlantic coast of Morocco. Aquatic
Botany, 89(3), 324-330.
Guimarães, S. M. P. B., & Fujii, M. T. (1998). Two species of foliose Halymenia
(Halymeniaceae, Rhodophyta) from Brazil. Botanica Marina, 41(1-6), 495-504.
Guiry, M. D., & Irvine, L. M. (1974). A species of Cryptonemia new to Europe. British
Phycological Journal, 9(3), 225-237.
Guiry, M. D., & Guiry, G. M. (2017). AlgaeBase. World-wide electronic publication,
National University of Ireland, Galway. Retrieved from http://www.algaebase.org.
Gulbransen, D. J., McGlathery, K. J., Marklund, M., Norris, J. N., & Gurgel, C. F. D.
(2012). Gracilaria vermiculophylla (Rhodophyta, Gracilariales) in the Virginia
coastal bays, USA: cox1 analysis reveals high genetic richness of an introduced
macroalga. Journal of Phycology, 48(5), 1278-1283.
Univers
ity of
Mala
ya

140
Hadi, S. I. I. A., Santana, H., Brunale, P. P. M., Gomes, T. G., Oliveira, M. D.,
Matthiensen, A., . . . Brasil, B. S. A. F. (2016). DNA barcoding green
microalgae isolated from neotropical inland waters. PLoS One, 11(2), e0149284.
Hall, T. A. (1999). BioEdit: a user-friendly biological sequence alignment editor and
analysis program for Windows 95/98/NT. Nucleic Acids Symposium Series, 41,
95-98.
Harper, J. T., & Saunders, G. W. (2001). The application of sequences of the ribosomal
cistron to the systematics and classification of the florideophyte red algae
(Florideophyceae, Rhodophyta). Cahiers de Biologie Marine, 42(1-2), 25-38.
Hashimo, T., Nakamura, Y., Kamaishi, T., & Hasegawa, M. (1997). Early evolution of
eukaryotes inferred from protein phylogenies of translation elongation factors 1α
and 2. Archiv für Protistenkunde, 148(3), 287-295.
Hebert, P. D. N., Cywinska, A., Ball, S. L., & De Waard, J. R. (2003). Biological
identifications through DNA barcodes. Proceedings of the Royal Society B:
Biological Sciences, 270(1512), 313-321.
Hebert, P. D. N., & Gregory, T. R. (2005). The promise of DNA barcoding for
taxonomy. Systematic Biology, 54(5), 852-859.
Hebert, P. D. N., Penton, E. H., Burns, J. M., Janzen, D. H., & Hallwachs, W. (2004a).
Ten species in one: DNA barcoding reveals cryptic species in the neotropical
skipper butterfly Astraptes fulgerator. Proceedings of the National Academy of
Sciences of the United States of America, 101(41), 14812-14817.
Hebert, P. D. N., Stoeckle, M. Y., Zemlak, T. S., & Francis, C. M. (2004b).
Identification of birds through DNA barcodes. PLoS Biology, 2(10), e312.
Hernández-Kantún, J. J., Riosmena-Rodriguez, R., Adey, W. H., & Rindi, F. (2014).
Analysis of the cox2-3 spacer region for population diversity and taxonomic
implications in rhodolith-forming species (Rhodophyta: Corallinales). Phytotaxa,
190(1), 331-354.
Hernández-Kantun, J. J., Riosmena-Rodriguez, R., & León-Cisneros, K. (2009).
Morphology and anatomy of Halymenia actinophysa (Halymeniales,
Rhodophyta) from the southwestern Gulf of California, Mexico. Botanica
Marina, 52(3), 248-255.
Univers
ity of
Mala
ya

141
Hernández-Kantún, J. J., Sherwood, A. R., Riosmena-Rodriguez, R., Huisman, J. M., &
De Clerck, O. (2012). Branched Halymenia species (Halymeniaceae,
Rhodophyta) in the Indo-Pacific region, including descriptions of Halymenia
hawaiiana sp. nov. and H. tondoana sp. nov. European Journal of Phycology,
47(4), 421-432.
Hillis, D. M., & Bull, J. J. (1993). An empirical test of bootstrapping as a method for
assessing confidence in phylogenetic analysis. Systematic Biology, 42(2), 182-
192.
Hillis, D. M., & Dixon, M. T. (1991). Ribosomal DNA: molecular evolution and
phylogenetic inference. The Quarterly Review of Biology, 66(4), 411-453.
Hind, K. R., & Saunders, G. W. (2013). Molecular markers from three organellar
genomes unravel complex taxonomic relationships within the coralline algal
genus Chiharaea (Corallinales, Rhodophyta). Molecular Phylogenetics and
Evolution, 67(2), 529-540.
Hollingsworth, P. M., Forrest, L. L., Spouge, J. L., Hajibabaei, M., Ratnasingham, S.,
van der Bank, M., . . . Little, D. P. (2009). A DNA barcode for land plants.
Proceedings of the National Academy of Sciences, 106(31), 12794-12797.
Hollingsworth, P. M., Graham, S. W., & Little, D. P. (2011). Choosing and using a
plant DNA barcode. PLoS One, 6(5), e19254.
Hommersand, M. H., & Fredericq, S. (1990). Sexual reproduction and cystocarp
development. In K. M. Cole & R. G. Sheath (Eds.), Biology of the red algae (pp.
305-345). New York: Cambridge University Press.
Hommersand, M. H., & Fredericq, S. (2003). Biogeography of the marine red algae of
the South African West Coast: a molecular approach. In A. R. O. Chapman, R. J.
Anderson, V. J. Vreeland & I. R. Davison (Eds.), Proceedings of the 17th
International Seaweed Symposium (pp. 325-336). Cape Town: Oxford
University Press.
Hommersand, M. H., Fredericq, S., Wilson Freshwater, D., & Hughey, J. (1999).
Recent developments in the systematics of the Gigartinaceae (Gigartinales,
Rhodophyta) based on rbcL sequence analysis and morphological evidence.
Phycological Research, 47(3), 139-151.
Hu, Z., Zeng, X., Wang, A., Shi, C., & Duan, D. (2004). An efficient method for DNA
isolation from red algae. Journal of Applied Phycology, 16(3), 161-166.
Univers
ity of
Mala
ya

142
Hu, Z. M., Guiry, M. D., & Duan, D. L. (2009). Using the ribosomal internal
transcribed spacer (ITS) as a complement marker for species identification of
red macroalgae. Hydrobiologia, 635(1), 279.
Huelsenbeck, J. P., & Ronquist, F. (2001). MRBAYES: Bayesian inference of
phylogenetic trees. Bioinformatics, 17(8), 754-755.
Hughes, A. R., Inouye, B. D., Johnson, M. T., Underwood, N., & Vellend, M. (2008).
Ecological consequences of genetic diversity. Ecology Letters, 11(6), 609-623.
Hwang, I. K., Kim, H. S., & Lee, W. J. (2004). Evidence for taxonomic status of
Pachydictyon coriaceum (Holmes) Okamura (Dictyotales, Phaeophyceae) based
on morphology and plastid protein coding rbcL, psaA, and psbA gene sequences.
Algae, 19(3), 175-190.
Jobb, G., Haeseler, A., & Strimmer, K. (2004). TREEFINDER: a powerful graphical
analysis environment for molecular phylogenetics. BMC Evolutionary Biology,
4(1), 18-26.
John, D. M., & Maggs, C. A. (1997). Species problems in eukaryotic algae: a modern
perspective. In M. F. Claridge, H. A. Dawah & M. R. Wilson (Eds.), The Units
of Biodiversity: Species in Practice (Vol. 54, pp. 83-107). London: Chapman
and Hall.
Joppa, L. N., Roberts, D. L., & Pimm, S. L. (2011). The population ecology and social
behavior of taxonomists. Trends in Ecology and Evolution, 26(11), 551-553.
Kawabata, S. (1962). A contribution to the systematic study of Grateloupiaceae from
Japan (2). Journal of Hokkaido Gakugei University, 13(2), 190-210.
Kawaguchi, S. (2004). Morphological observations of the type and some authentic
material of Halymenia floresia (Clemente y Rubio) C. Agardh, with notes on
previous reports of this alga. In I. A. Abbott & K. J. McDermid (Eds.),
Taxonomy of economic seaweeds with reference to the Pacific and other
locations (Vol. 9, pp. 143-156). Honolulu, Hawaii: Hawaii Sea Grant College
Program.
Kawaguchi, S., Kato, A., Masuda, M., Kogame, K., & Phang, S. M. (2002a).
Taxonomic notes on marine algae from Malaysia. VII. Five species of
Rhodophyceae, with the description of Lomentaria gracillima sp. nov. Botanica
Marina, 45(6), 536-547.
Univers
ity of
Mala
ya

143
Kawaguchi, S., & Lewmanomont, K. (1999). Morphology and culture study of a red
alga, Halymenia dilatata Zanardini, from Vietnam and Japan. In I. A. Abbott
(Ed.), Taxonomy of economic seaweeds with reference to some Pacific species
(Vol. 7, pp. 147-161). La Jolla, California: California Sea Grant College
Program.
Kawaguchi, S., Lewmanomont, K., & McDermid, K. J. (2002b). Morphology of
Halymenia maculata J. Agardh from Vietnam. In I. A. Abbott & K. J. Mcdermid
(Eds.), Taxonomy of economic seaweeds with reference to some Pacific species
(Vol. 8, pp. 259-266). La Jolla, California: California Sea Grant College
Program.
Kawaguchi, S., Shimada, S., Wang, H. W., & Masuda, M. (2004). The new genus
Yonagunia Kawaguchi et Masuda (Halymeniaceae, Rhodophyta), based on Y.
tenuifolia Kawaguchi et Masuda sp. nov. from Southern Japan and including Y.
formosana (Okamura) Kawaguchi et Masuda comb. nov. from Southeast Asia
Journal of Phycology, 40(1), 180-192.
Kawaguchi, S., Shimada, S., Wang, H. W., Faye, E. J., & Masuda, M. (2003). Polyopes
tosaensis Kawaguchi & Masuda, sp. nov. (Halymeniaceae, Rhodophyta) from
Japan. European Journal of Phycology, 38(4), 315-324.
Kawaguchi, S., Shimida, S., Abe, T., & Terada, R. (2006). Morphological and
molecular phylogenetic studies of a red alga, Halymenia durvillei,
(Halymeniaceae, Halymeniales) from Indo-Pacific. Coastal Marine Science,
30(1), 201-208.
Kawaguchi, S., Wang, H. W., Horiguchi, T., Sartoni, G., & Masuda, M. (2001). A
comparative study of the red alga Grateloupia filicina (Halymeniaceae) from the
northwestern Pacific and Mediterranean with the description of Grateloupia
asiatica, sp. nov. Journal of Phycology, 37(3), 433-442.
Kawai, H., & Sasaki, H. (2004). Morphology, life history, and molecular phylogeny of
Stschapovia flagellaris (Tilopteridales, Phaephyceae) and the erection of the
Stschapoviaceae fam. nov. Journal of Phycology, 40(6), 1156-1169.
Kazi, M. A., Reddy, C. R. K., & Jha, B. (2013). Molecular phylogeny and barcoding of
Caulerpa (Bryopsidales) based on the tufA, rbcL, 18S rDNA and ITS rDNA
genes. PLoS One, 8(12), e82438.
Kho, F. (2014). Biochemical composition of three red algae (Halymenia durvillaea
Bory de Saint-Vincent, Halymenia dilatata J. Agardh and Halymenia maculata
Zanardini) in Barangay Tubigan, Initao, Misamis Oriental. (Unpublished
Undergraduate's thesis). Mindanao State University, Philippines.
Univers
ity of
Mala
ya

144
Kho, F. B., Orbita, M. L. S., Manting, M. M. E., & Orbita, R. R. (2016). Carrageenan
and biochemical composition of three species of Halymenia (Halymenia
durvillaea Bory de Saint-Vincent, Halymenia maculata J. Agardh and
Halymenia dilatata Zanardini) in Initao, Misamis Oriental, Mindanao,
Philippines. International Journal of Biosciences, 8(5), 182-189.
Kim, S. Y., Manghisi, A., Morabito, M., Yang, E. C., Yoon, H. S., Miller, K. A., & Boo,
S. M. (2014). Genetic diversity and haplotype distribution of Pachymeniopsis
gargiuli sp. nov. and P. lanceolata (Halymeniales, Rhodophyta) in Korea, with
notes on their non-native distributions. Journal of Phycology, 50(5), 885-896.
Kim, S. Y., Seo, T. H., Park, J. K., Boo, G. H., Kim, K. M., & Boo, S. M. (2012).
Cryptonemia rotunda (Halymeniales) and Schizymenia apoda
(Nemastomatales), two new records of red algae from Korea. Algae, 27(1), 1-8.
Kraft, G. T., Liao, L. M., Millar, A. J. K., Coppejans, E. G. G., Hommersand, M. H., &
Freshwater, D. W. (1999). Marine benthic red algae (Rhodophyta) from Bulusan,
Sorsogon Province, Southern Luzon, Philippines. The Philippine Scientist, 36, 1-
50.
Kraft, G. T. (1977). The morphology of Grateloupia intestinalis from New Zealand,
with some thoughts on generic criteria within the family Cryptonemiaceae
(Rhodophyta). Phycologia, 16(1), 43-51.
Kraft, G. T., Liao, L. M., Millar, A. J. K., Coppejans, E. G. G., Hommersand, M. H., &
Freshwater, D. W. (1999). Marine benthic red algae (Rhodophyta) from
Bulusan, Sorsogon Province, Southern Luzon, Philippines. The Philippine
Scientist, 36, 1-50.
Kranz, H., Jürgens, K., Pinier, M., & Siepmann, J. (2009). Drug release from MCC-and
carrageenan-based pellets: experiment and theory. European Journal of
Pharmaceutics and Biopharmaceutics, 73(2), 302-309.
Kucera, H., & Saunders, G. W. (2008). Assigning morphological variants of Fucus
(Fucales, Phaeophyceae) in Canadian waters to recognized species using DNA
barcoding. Botany, 86(9), 1065-1079.
Kucera, H., & Saunders, G. W. (2012). A survey of Bangiales (Rhodophyta) based on
multiple molecular markers reveals cryptic diversity. Journal of Phycology,
48(4), 869-882.
Kumari, N., & Thakur, S. K. (2014). Randomly amplified polymorphic DNA-a brief
review. American Journal of Animal and Veterinary Sciences, 9(1), 6-13.
Univers
ity of
Mala
ya

145
Kützing, F. T. (1843). Phycologia generalis oder Anatomie, Physiologie und
Systemkunde der Tange. Mit 80 farbig gedruckten Tafeln, gezeichnet und gravirt
vom Verfasser. Leipzig: F.A. Brockhaus.
Kylin, H. (1956). Die Gattungen der Rhodophyceen. Lund: C. W. K. Gleerup.
Lai, M. F., Li, C. F., & Li, C. Y. (1994). Characterization and thermal behavior of six
sulphated polysaccharides from seaweeds. Food Hydrocolloids, 8(3-4), 215-232.
Larkin, M. A., Blackshields, G., Brown, N. P., Chenna, R., McGettigan, P. A.,
McWilliam, H., . . . Higgins, D. G. (2007). Clustal W and Clustal X version 2.0.
Bioinformatics, 23(21), 2947-2948.
Le Gall, L., & Saunders, G. W. (2007). A nuclear phylogeny of the Florideophyceae
(Rhodophyta) inferred from combined EF2, small subunit and large subunit
ribosomal DNA: establishing the new red algal subclass Corallinophycidae.
Molecular Phylogenetics and Evolution, 43(3), 1118-1130.
Le Gall, L., & Saunders, G. W. (2010). DNA barcoding is a powerful tool to uncover
algal diversity: a case study of the Phyllophoraceae (Gigartinales, Rhodophyta)
in the Canadian flora. Journal of Phycology, 46(2), 374-389.
Lee, H. W., Yang, M. Y., & Kim, M. S. (2016). Verifying a new distribution of the
genus Amalthea (Halymeniales, Rhodophyta) with description of A. rubida sp.
nov. from Korea. Algae, 31(4), 341-349.
Leitch, I. J., Greilhuber, J., Dolezel, J., & Wendel, J. (2013). Plant Genome Diversity
Volume 2: Physical Structure, Behaviour and Evolution of Plant Genomes.
Vienna: Springer-Verlag Wien.
Lewis, L. A., & Flechtner, V. R. (2004). Cryptic species of Scenedesmus (Chlorophyta)
from desert soil communities of Western North America. Journal of Phycology,
40(6), 1127-1137.
Leliaert, F., Smith, D. R., Moreau, H., Herron, M. D., Verbruggen, H., Delwiche, C. F.,
& De Clerck, O. (2012). Phylogeny and molecular evolution of the green algae.
Critical Reviews in Plant Sciences, 31(1), 1-46.
Univers
ity of
Mala
ya

146
Lewmanomont, K., & Kawaguchi, S. (2002). Foliose Halymenia (Halymeniaceae,
Cryptonemiales, Rhodophyta) from Thailand. In I. A. Abbott & K. J. McDermid
(Eds.), Taxonomy of economic seaweeds with reference to some Pacific species
(Vol. 8, pp. 267-277). La Jolla, California: California Sea Grant College
Program.
Li, S., Qiao, K., Shan, T., Pang, S., & Hou, H. (2013). Genetic diversity and
relationships of the brown alga Undaria pinnatifida cultivated along the Dalian
Coast as revealed by amplified fragment length polymorphism markers. Journal
of Applied Phycology, 25(4), 1255-1263.
Lim, P. E., Tan, J., Phang, S. M., Nikmatullah, A., Hong, D. D., Sunarpi, H., & Hurtado,
A. Q. (2013). Genetic diversity of Kappaphycus Doty and Eucheuma J. Agardh
(Solieriaceae, Rhodophyta) in Southeast Asia. Journal of Applied Phycology,
26(2), 1253-1272.
Lim, P. E., Thong, K. L., & Phang, S. M. (2001). Molecular differentiation of two
morphological variants of Gracilaria salicornia. Journal of Applied Phycology,
13(4), 335-342.
Lin, S. M., Liang, H. Y., & Hommersand, M. H. (2008). Two types of auxiliary cell
ampullae in Grateloupia (Halymeniaceae, Rhodophyta), including G.
taiwanensis sp. nov. and G. orientalis sp. nov. from Taiwan based on rbcL gene
sequences analysis and cystocarp development. Journal of Phycology, 44(1),
196-214.
Lincoln, R., Boxshall, G., & Clark, P. (1998). A dictionary of ecology, evolution and
systematics. Cambridge, UK: Cambridge University Press.
Lüning, K. (1990). Seaweeds: their environment, biogeography, and ecophysiology.
New York: John Wiley & Sons.
Mace, G. M., Gittleman, J. L., & Purvis, A. (2003). Preserving the tree of life. Science,
300(5626), 1707-1709.
Maggs, C. A., & Guiry, M. D. (1982). Morphology, phenology and photoperiodism in
Halymenia latifolia Kütz.(Rhodophyta) from Ireland. Botanica Marina, 25(12),
589-600.
Maggs, C. A., Verbruggen, H., & De Clerck, O. (2007). Molecular systematics of red
algae: building future structures on firm foundations. In J. Brodie & J. Lewis
(Eds.), Unravelling the algae: the past, present, and future of algal systematics
(Vol. 75, pp. 103-121). London: CRC Press.
Univers
ity of
Mala
ya

147
Malairaj, S., Muthu, S., Gopal, V. B., Perumal, P., & Ramasamy, R. (2016). Qualitative
and quantitative determination of R-phycoerythrin from Halymenia floresia
(Clemente) C. Agardh by polyacrylamide gel using electrophoretic elution
technique. Journal of Chromatography A, 1454, 120-126.
Manghisi, A., Morabito, M., Boo, G. H., Boo, S. M., Bonillo, C., De Clerck, O., & Le
Gall, L. (2015). Two novel species of Yonagunia (Halymeniales, Rhodophyta)
were uncovered in the South of Madagascar during the Atimo-Vatae Expedition.
Cryptogamie, Algologie, 36(2), 199-217.
Manhart, J. R., & McCourt, R. M. (1992). Molecular data and species concepts in the
algae. Journal of Phycology, 28(6), 730-737.
Mann, K. H. (1973). Seaweeds: their productivity and strategy for growth. Science,
182(4116), 975-981.
Marston, M., & Villalard-Bohnsack, M. (2002). Genetic variability and potential
sources of Grateloupia doryphora (Halymeniaceae, Rhodophyta), an invasive
species in Rhode Island waters (USA). Journal of Phycology, 38(4), 649-658.
Mateo-Cid, L. E., Mendoza-González, A. C., Gavio, B., & Fredericq, S. (2005).
Grateloupia huertana sp. nov.(Halymeniaceae, Rhodophyta), a peculiar new
prostrate species from tropical Pacific Mexico. Phycologia, 44(1), 4-16.
McHugh, D. J. (2003). Carrageenan. In A guide to the seaweed industry (pp. 61-72).
Retrieved from http://www.fao.org/3/a-y4765e.pdf
McIvor, L., Maggs, C. A., Provan, J., & Stanhope, M. J. (2001). RbcL sequences reveal
multiple cryptic introductions of the Japanese red alga Polysiphonia harveyi.
Molecular Ecology, 10(4), 911-919.
Meudt, H. M., & Clarke, A. C. (2007). Almost forgotten or latest practice? AFLP
applications, analyses and advances. Trends in Plant Science, 12(3), 106-117.
Milstein, D., & Saunders, G. W. (2012). DNA barcoding of Canadian Ahnfeltiales
(Rhodophyta) reveals a new species–Ahnfeltia borealis sp. nov. Phycologia,
51(3), 247-259.
Moniz, M. B. J., & Kaczmarska, I. (2010). Barcoding of diatoms: nuclear encoded ITS
revisited. Protist, 161(1), 7-34.
Univers
ity of
Mala
ya

148
Moreira, D., Le Guyader, H., & Philippe, H. (2000). The origin of red algae and the
evolution of chloroplasts. Nature, 405(6782), 69-72.
Müller, K. M., Oliveira, M. C., Sheath, R. G., & Bhattacharya, D. (2001). Ribosomal
DNA phylogeny of the Bangiophycidae (Rhodophyta) and the origin of
secondary plastids. American Journal of Botany, 88(8), 1390-1400.
Myers, N., Mittermeier, R. A., Mittermeier, C. G., Da Fonseca, G. A. B., & Kent, J.
(2000). Biodiversity hotspots for conservation priorities. Nature, 403(6772),
853-858.
Nam, K. W., Maggs, C. A., McIvor, L., & Stanhope, M. J. (2000). Taxonomy and
phylogeny of Osmundea (Rhodomelaceae, Rhodophyta) in Atlantic Europe.
Journal of Phycology, 36(4), 759-772.
Narendran, T. C. (2000). The importance of systematics. Resonance, 5(6), 60-68.
Nature (2002). Genomics and taxonomy for all. Nature, 417(6889), 573.
Newmaster, S. G., Fazekas, A. J., Steeves, R. A. D., & Janovec, J. (2008). Testing
candidate plant barcode regions in the Myristicaceae. Molecular Ecology
Resources, 8(3), 480-490.
Newmaster, S. G., & Ragupathy, S. (2009). Testing plant barcoding in a sister species
complex of pantropical Acacia (Mimosoideae, Fabaceae). Molecular Ecology
Resources, 9(s1), 172-180.
Ng, P. K., Lim, P. E., & Phang, S. M. (2015). Small-scale genetic structure of
Gracilaria salicornia and its red algal parasite, G. babae (Gracilariaceae,
Rhodophyta), in Malaysia. Botanica Marina, 58(3), 175-187.
Norris, J. N. (2014). Marine algae of the northern Gulf of California II: Rhodophyta.
Smithsonian Contributions to Botany, 96, 1-555.
Pang, Q., Sui, Z., Kang, K. H., Kong, F., & Zhang, X. (2010). Application of SSR and
AFLP to the analysis of genetic diversity in Gracilariopsis lemaneiformis
(Rhodophyta). Journal of Applied Phycology, 22(5), 607-612.
Parekh, R. G., Doshi, Y. A., & Chauhan, V. D. (1989). Polysaccharides from marine red
algae Acanthophora spicifera, Grateloupia indica and Halymenia porphyroides.
Indian Journal of Marine Sciences, 18, 139-140.
Univers
ity of
Mala
ya

149
Parekh, R. G., Doshi, Y. A., Rao, A. V., & Chauhan, V. D. (1987). Studies on a
phycocolloid from red alga Halymenia venusta Boergesen. Indian Journal of
Marine Sciences, 16, 274-276.
Patwardhan, A., Ray, S., & Roy, A. (2014). Molecular markers in phylogenetic studies-
a review. Journal of Phylogenetics & Evolutionary Biology, 2, 131.
Patwary, M. U., MacKay, R. M., & Meer, J. P. (1993). Revealing genetic markers in
Gelidium vagum (Rhodophyta) through the random amplified polymorphic
DNA (RAPD) technique. Journal of Phycology, 29(2), 216-222.
Patwary, M. U., Sensen, C. W., MacKay, R. M., & van der Meer, J. P. (1998).
Nucleotide sequences of small-subunit and internal transcribed spacer regions of
nuclear rRNA genes support the autonomy of some genera of the Gelidiales
(Rhodophyta). Journal of Phycology, 34(2), 299-305.
Pereira, L., van de Velde, F., & Mesquita, J. F. (2007). Cytochemical studies on
underutilized carrageenophytes (Gigartinales, Rhodophyta). International
Journal of Biology and Biomedical Engineering, 1(1), 1-5.
Phang, S. M. (2006). Seaweed resources in Malaysia: current status and future prospects.
Aquatic Ecosystem Health & Management Society, 9(2), 185-202
Phang, S. M., Yeong, H. Y., Ganzon-Fortes, E. T., Lewmanomont, K., Prathep, A., Hau,
L. N., . . . Tan, K. S. (2016). Marine algae of the South China Sea bordered by
Indonesia, Malaysia, Philippines, Singapore, Thailand and Vietnam. Raffles
Bulletin of Zoology Supplement, 34, 13-59.
Pierce, J. C., Tan, G., & Gaffney, P. M. (2000). Delaware Bay and Chesapeake Bay
populations of the horseshoe crab Limulus polyphemus are genetically distinct.
Estuaries, 23(5), 690-698.
Presting, G. G. (2006). Identification of conserved regions in the plastid genome:
implications for DNA barcoding and biological function. Botany, 84(9), 1434-
1443.
Pueschel, C. M. (1990). Cell structure. In K. M. Cole & R. G. Sheath (Eds.), Biology of
the red algae (pp. 7-41). New York: Cambridge University Press.
Pueschel, C. M., & Cole, K. M. (1982). Rhodophycean pit plugs: an ultrastructural
survey with taxonomic implications. American Journal of Botany, 69(5), 703-
720.
Univers
ity of
Mala
ya

150
Ragan, M. A., Bird, C. J., Rice, E. L., Gutell, R. R., Murphy, C. A., & Singh, R. K.
(1994). A molecular phylogeny of the marine red algae (Rhodophyta) based on
the nuclear small-subunit rRNA gene. Proceedings of the National Academy of
Sciences, 91(15), 7276-7280.
Rambaut, A., & Drummond, A. J. (2009a). FigTree (Version 1.3.1). Retrieved from
http://tree.bio.ed.ac.uk /software/figtree
Rambaut, A., & Drummond, A. J. (2009b). Tracer (Version 1.5). Retrieved from
http://beast.bio.ed.ac.uk/Tracer
Robba, L., Russell, S. J., Barker, G. L., & Brodie, J. (2006). Assessing the use of the
mitochondrial cox1 marker for use in DNA barcoding of red algae (Rhodophyta).
American Journal of Botany, 93(8), 1101-1108.
Rojas, M. (1992). The species problem and conservation: what are we protecting?
Conservation Biology, 6(2), 170-178.
Ronquist, F., & Huelsenbeck, J. P. (2003). MrBayes 3: Bayesian phylogenetic inference
under mixed models. Bioinformatics, 19(12), 1572-1574.
Roy, S. S., Dasgupta, R., & Bagchi, A. (2014). A review on phylogenetic analysis: a
journey through modern era. Computational Molecular Bioscience, 4(3), 39-45.
Rueness, J. (1989). Sargassum muticum and other introduced Japanese macroalgae:
biological pollution of European coasts. Marine Pollution Bulletin, 20(4), 173-
176.
Rueness, J. (2010). DNA barcoding of select freshwater and marine red algae
(Rhodophyta). Cryptogamie, Algologie, 31(4), 377.
Rueness, J., & Rueness, E. K. (2000). Caulacanthus ustulatus (Gigartinales,
Rhodophyta) from Brittany (France) is an introduction from the Pacific Ocean.
Cryptogamie, Algologie, 21(4), 355-363.
Russell, L. K., Hurd, C. L., Nelson, W. A., & Broom, J. E. (2009). An examination of
Pachymenia and Aeodes (Halymeniaceae, Rhodophyta) in New Zealand and the
transfer of two species of Aeodes in South Africa to Pachymenia. Journal of
Phycology, 45(6), 1389-1399.
Univers
ity of
Mala
ya

151
Samper, C. (2004). Taxonomy and environmental policy. Philosophical Transactions of
the Royal Society of London B: Biological Sciences, 359(1444), 721-728.
Saunders, G. W. (1993). Gel purification of red algal genomic DNA: an inexpensive
and rapid method for the isolation of polymerase chain reaction- friendly DNA.
Journal of Phycology, 29(2), 251-254.
Saunders, G. W. (2005). Applying DNA barcoding to red macroalgae: a preliminary
appraisal holds promise for future applications. Philosophical Transactions of
the Royal Society B: Biological Sciences, 360(1462), 1879-1888.
Saunders, G.W. (2008). A DNA barcode examination of the red algal family
Dumontiaceae in Canadian waters reveals substantial cryptic species diversity. 1.
The foliose Dilsea-Neodilsea complex and Weeksia. Botany, 86(7), 773-789.
Saunders, G. W. (2009). Routine DNA barcoding of Canadian Gracilariales
(Rhodophyta) reveals the invasive species Gracilaria vermiculophylla in British
Columbia. Molecular Ecology Resources, 9(s1), 140-150.
Saunders, G. W. (2014). Long distance kelp rafting impacts seaweed biogeography in
the Northeast Pacific: the kelp conveyor hypothesis. Journal of Phycology, 50(6),
968-974.
Saunders, G. W., & Hommersand, M. H. (2004). Assessing red algal supraordinal
diversity and taxonomy in the context of contemporary systematic data.
American Journal of Botany, 91(10), 1494-1507.
Saunders, G. W., & Kraft, G. T. (1996). Small-subunit rRNA gene sequences from
representatives of selected families of the Gigartinales and Rhodymeniales
(Rhodophyta). 2. Recognition of the Halymeniales ord. nov. Canadian Journal
of Botany, 74(5), 694-707.
Saunders, G. W., & Kucera, H. (2010). An evaluation of rbcL, tufA, UPA, LSU and ITS
as DNA barcode markers for the marine green macroalgae. Cryptogamie,
Algologie, 31(4), 487-528.
Saunders, G. W., & McDevit, D. C. (2013). DNA barcoding unmasks overlooked
diversity improving knowledge on the composition and origins of the Churchill
algal flora. BMC Ecology, 13(1), 9.
Univers
ity of
Mala
ya

152
Saunders, G. W., & McDonald, B. (2010). DNA barcoding reveals multiple overlooked
Australian species of the red algal order Rhodymeniales (Florideophyceae), with
resurrection of Halopeltis J. Agardh and description of Pseudohalopeltis gen.
nov. Botany, 88(7), 639-667.
Saunders, G. W., & Moore, T. E. (2013). Refinements for the amplification and
sequencing of red algal DNA barcode and RedToL phylogenetic markers: a
summary of current primers, profiles and strategies. Algae, 28(1), 31-43.
Savolainen, V., Cowan, R. S., Vogler, A. P., Roderick, G. K., & Lane, R. (2005).
Towards writing the encyclopaedia of life: an introduction to DNA barcoding.
Philosophical Transactions of the Royal Society B: Biological Sciences,
360(1462), 1805-1811.
Schaffelke, B., Smith, J. E., & Hewitt, C. L. (2006). Introduced macroalgae- a growing
concern. Journal of Applied Phycology, 18, 529-541.
Schneider, C. W., Lane, C. E., & Saunders, G. W. (2010). Notes on the marine algae of
the Bermudas. 11. More additions to the benthic flora and a phylogenetic
assessment of Halymenia pseudofloresii (Halymeniales, Rhodophyta) from its
type locality. Phycologia, 49(2), 154-168.
Searles, R. B. (1980). The strategy of the red algal life history. The American Naturalist,
115(1), 113-120.
Semesi, A. K., & Mshigeni, K. E. (1977). Contributions on the content and nature of the
phycocolloid from Halymenia venusta Boergesen (Rhodophyta,
Cryptonemiales). Botanica Marina, 20(4), 233-238.
Shan, T. F., Pang, S. J., Zhang, Y. R., Yakovleva, I. M., & Skriptsova, A. V. (2011). An
AFLP-based survey of genetic diversity and relationships of major farmed
cultivars and geographically isolated wild populations of Saccharina japonica
(Phaeophyta) along the northwest coasts of the Pacific. Journal of Applied
Phycology, 23(1), 35-45.
Sherwood, A. R. (2008). Phylogeography of Asparagopsis taxiformis
(Bonnemaisoniales, Rhodophyta) in the Hawaiian Islands: two mtDNA markers
support three separate introductions. Phycologia, 47(1), 79-88.
Sherwood, A. R., Kurihara, A., Conklin, K. Y., Sauvage, T., & Presting, G. G. (2010a).
The Hawaiian Rhodophyta Biodiversity Survey (2006-2010): a summary of
principal findings. BMC Plant Biology, 10, 258.
Univers
ity of
Mala
ya

153
Sherwood, A. R., & Presting, G. G. (2007). Universal primers amplify a 23S rDNA
plastid marker in eukaryotic algae and cyanobacteria. Journal of Phycology,
43(3), 605-608.
Sherwood, A. R., Sauvage, T., Kurihara, A., Conklin, K. Y., & Presting, G. G. (2010b).
A comparative analysis of COI, LSU and UPA marker data for the Hawaiian
florideophyte Rhodophyta: implications for DNA barcoding of red algae.
Cryptogamie, Algologie, 31(4), 451-465.
Sherwood, A. R., Vis, M. L., Entwisle, T. J., Necchi, O. Jr., & Presting, G. G. (2008).
Contrasting intra versus interspecies DNA sequence variation for representatives
of the Batrachospermales (Rhodophyta): insights from a DNA barcoding
approach. Phycological Research, 56(4), 269-279.
Silva, P. C., Meñez, E. G., & Moe, R. L. (1987). Catalog of the benthic marine algae of
the Philippines. Smithsonian Contributions to Marine Sciences, 27, 1-179.
Singh, A. K. (2012). Molecular taxonomy: use of modern methods in the identification
of a species. Indian Journal of Life Science, 2(1), 143-147.
Smith, J. E., Hunter, C. L., & Smith, C. M. (2002). Distribution and reproductive
characteristics of nonindigenous and invasive marine algae in the Hawaiian
Islands. Pacific Science, 56(3), 299-315.
Sodhi, N. S., Koh, L. P., Brook, B. W., & Ng, P. K. L. (2004). Southeast Asian
biodiversity: an impending disaster. Trends in Ecology & Evolution, 19(12),
654-660.
Soltis, D. E., & Soltis, P. S. (2003). The role of phylogenetics in comparative genetics.
Plant Physiology, 132(4), 1790-1800.
Spolaore, P., Joannis-Cassan, C., Duran, E., & Isambert, A. (2006). Commercial
applications of microalgae. Journal of Bioscience and Bioengineering, 101(2),
87-96.
Staats, M., Cuenca, A., Richardson, J. E., Vrielink-van Ginkel, R., Petersen, G., Seberg,
O., & Bakker, F. T. (2011). DNA damage in plant herbarium tissue. PLoS One,
6(12), e28448.
Swofford, D. L. (2003). PAUP*. Phylogenetic analysis using parsimony (* and other
methods) (Version 4). Sunderland, Massachusetts: Sinauer Associates.
Univers
ity of
Mala
ya

154
Swofford, D. L., Olsen, G. J., Waddell, P. J., Hillis, D. M., & Hillis, D. M. (1996).
Phylogenetic Inference. In D. M. Hiillis, D. Moritz & B. K. Mable (Eds.),
Molecular Systematics (pp. 407-514). Sunderland, Massachusetts: Sinauer
Associates.
Tamura, K., Stecher, G., Peterson, D., Filipski, A., & Kumar, S. (2013). MEGA6:
Molecular Evolutionary Genetics Analysis Version 6.0. Molecular Biology and
Evolution, 30(12), 2725-2729.
Tan, J., Lim, P. E., & Phang, S. M. (2013). Phylogenetic relationship of Kappaphycus
Doty and Eucheuma J. Agardh (Solieriaceae, Rhodophyta) in Malaysia. Journal
of Applied Phycology, 25(1), 13-29
Tan, J., Lim, P. E., Phang, S. M., Hong, D. D., Sunarpi, H., & Hurtado, A. Q. (2012).
Assessment of four molecular markers as potential DNA barcodes for red algae
Kappaphycus Doty and Eucheuma J. Agardh (Solieriaceae, Rhodophyta). PLoS
One, 7(12), e52905.
Tan, P. L., Lim, P. E., Lin, S. M., Phang, S. M., Draisma, S. G. A., & Liao, L. M.
(2015). Foliose Halymenia species (Halymeniaceae, Rhodophyta) from
Southeast Asia, including a new species, Halymenia malaysiana sp. nov.
Botanica Marina, 58(3), 203-217.
Tan, P. L., Lim, P. E., Lin, S. M., & Phang, S. M. (2017). Halymenia johorensis sp. nov.
(Halymeniaceae, Rhodophyta), a new foliose red algal species from Malaysia.
Journal of Applied Phycology. doi:10.1007/s10811-017-1104-8
Tanabe, A. S. (2007). Kakusan: a computer program to automate the selection of a
nucleotide substitution model and the configuration of a mixed model on
multilocus data. Molecular Ecology Notes, 7(6), 962-964.
Taylor, W. R. (1960). Marine algae of the eastern tropical and subtropical coasts of the
Americas. Ann Arbor: University of Michigan Press.
Teasdale, B. W., & Klein, A. S. (2010). Genetic variation and biogeographical
boundaries within the red alga Porphyra umbilicalis (Bangiales, Rhodophyta).
Botanica Marina, 53(5), 417-431.
Tsutsui, I., Hamano, K., Aue-Umneoy, D., Songphatkaew, J., Srisapoome, P.,
Ruangsomboon, S., . . . Maeno, Y. (2012). Common underwater plants in
coastal areas of Thailand. Tsukuba: Japan International Research Center for
Agricultural Sciences.
Univers
ity of
Mala
ya

155
Untergasser, A., Cutcutache, I., Koressaar, T., Ye, J., Faircloth, B. C., Remm, M., &
Rozen, S. G. (2012). Primer3- new capabilities and interfaces. Nucleic Acids
Research, 40(15), e115.
Uwai, S., Nelson, W., Neill, K., Wang, W. D., Aguilar-Rosas, L. E., Boo, S. M., . . .
Kawai, H. (2006a). Genetic diversity in Undaria pinnatifida (Laminariales,
Phaeophyceae) deduced from mitochondria genes-origins and succession of
introduced populations. Phycologia, 45(6), 687-695.
Uwai, S., Yotsukura, N., Serisawa, Y., Muraoka, D., Hiraoka, M., & Kogame, K.
(2006b). Intraspecific genetic diversity of Undaria pinnatifida in Japan, based
on the mitochondrial cox3 gene and the ITS1 of nrDNA. Hydrobiologia, 553(1),
345-356.
Valentin, K., & Zetsche, K. (1989). The genes of both subunits of ribulose-1, 5-
bisphosphate carboxylase constitute an operon on the plastome of a red alga.
Current Genetics, 16(3), 203-209.
Valero, M., Engel, C., Billot, C., Kloareg, B., & Destombe, C. (2001). Concept and
issues of population genetics in seaweeds. Cahiers de Biologie Marine, 42(1-2),
53-62.
van den Hoek, C., & Cortel-Breeman, A. (1970). Life-history studies on Rhodophyceae
II. Halymenia floresia (Clem.) Ag. Acta Botanica Neerlandica, 19(3), 341-362.
van den Hoek, C., Mann, D. G., & Jahns, H. M. (1995). Algae: an introduction to
phycology. New York: Cambridge University Press.
van Oppen, M. J. H., Klerk, H., Olsen, J. L., & Stam, W. T. (1996). Hidden diversity in
marine algae: some examples of genetic variation below the species level.
Journal of the Marine Biological Association of the United Kingdom, 76(1),
239-242.
van Velzen, R., Weitschek, E., Felici, G., & Bakker, F. T. (2012). DNA barcoding of
recently diverged species: relative performance of matching methods. PLoS One,
7(1), e30490.
Verheij, E., & Prud’Homme van Reine, W. F. (1993). Seaweeds of the Spermonde
Archipelago, SW Sulawesi, Indonesia. Blumea, 37(2), 385-510.
Univers
ity of
Mala
ya

156
Verbruggen, H., Maggs, C. A., Saunders, G. W., Le Gall, L., Yoon, H. S., & De Clerck,
O. (2010). Data mining approach identifies research priorities and data
requirements for resolving the red algal tree of life. BMC Evolutionary Biology,
10(1), 16.
Vis, M. L., Hodge, J. C., & Necchi, O. Jr. (2008). Phylogeography of Batrachospermum
macrosporum (Batrachospermales, Rhodophyta) from North and South America.
Journal of Phycology, 44(4), 882-888.
Vogler, A. P., & Monaghan, M. T. (2007). Recent advances in DNA taxonomy. Journal
of Zoological Systematics and Evolutionary Research, 45(1), 1-10.
Voris, H. K. (2000). Maps of Pleistocene sea levels in Southeast Asia: shorelines, river
systems and time durations. Journal of Biogeography, 27(5), 1153-1167.
Vos, P., Hogers, R., Bleeker, M., Reijans, M., Van de Lee, T., Hornes, M., . . . Kuiper,
M. (1995). AFLP: a new technique for DNA fingerprinting. Nucleic Acids
Research, 23(21), 4407-4414.
Wägele, J. W. (2005). Foundations of phylogenetic systematics. München, Germany:
Verlag Dr. Friedrich Pfeil
Wang, H. W., Kawaguchi, S., Horiguchi, T., & Masuda, M. (2000). Reinstatement of
Grateloupia catenata (Rhodophyta, Halymeniaceae) on the basis of morphology
and rbcL sequences. Phycologia, 39(3), 228-237.
Wang, H. W., Kawaguchi, S., Horiguchi, T., & Masuda, M. (2001). A morphological
and molecular assessment of the genus Prionitis J. Agardh (Halymeniaceae,
Rhodophyta). Phycological Research, 49(3), 251-261.
Weber-Van Bosse, A. (1921). Liste des algues du Siboga. II. Rhodophyceae. Premiere
partie. Protoflorideae, Nemalionales, Cryptonemiales. Siboga-Expeditie
Monograph, 59, 187-310.
Wilkes, R. J., McIvor, L. M., & Guiry, M. D. (2005). Using rbcL sequence data to
reassess the taxonomic position of some Grateloupia and Dermocorynus species
(Halymeniaceae, Rhodophyta) from the north-eastern Atlantic. European
Journal of Phycology, 40(1), 53-60.
Williams, J. G., Kubelik, A. R., Livak, K. J., Rafalski, J. A., & Tingey, S. V. (1990).
DNA polymorphisms amplified by arbitrary primers are useful as genetic
markers. Nucleic Acids Research, 18(22), 6531-6535.
Univers
ity of
Mala
ya

157
Wittmann, W. (1965). Aceto-iron-haematoxylin-chloral hydrate for chromosome
staining. Stain Technology, 40(3), 161-164.
Woelkerling, W. J. (1990). An introduction. In K. M. Cole & R. G. Sheath (Eds.),
Biology of the red algae (pp. 1-6). New York: Cambridge University Press.
Womersley, H. B. S., & Lewis, J. A. (1994). Family Halymeniaceae Bory 1828: 158. In
H. B. S. Womersley (Ed.), The Marine Benthic Flora of Southern Australia.
Part IIIA. (pp. 167-218). Canberra: Australian Biological Resources Study.
Wright, J. T., Zuccarello, G. C., & Steinberg, P. D. (2000). Genetic structure of the
subtidal red alga Delisea pulchra. Marine Biology, 136(3), 439-448.
Wright, S. (1920). The relative importance of heredity and environment in determining
the piebald pattern of guinea-pigs. Proceedings of the National Academy of
Sciences of the United States of America, 6(6), 320-332.
Wyrtki, K. (1961). Physical oceanography of the Southeast Asian waters. In Scientific
results of marine investigations of the South China Sea and the Gulf of Thailand
1959-1961 (Vol. 2, pp. 195). La Jolla, California: Scripps Institution of
Oceanography.
Xie, Z. Y., Lin, S. M., Liu, L. C., Ang, P. O., & Shyu, J. F. (2015). Genetic diversity
and taxonomy of foliose Bangiales (Rhodophyta) from Taiwan based on rbcL
and cox1 sequences. Botanica Marina, 58(3), 189-202.
Yang, E. C., Kim, M. S., Geraldino, P. J. L., Sahoo, D., Shin, J.-A., & Boo, S. M.
(2008). Mitochondrial cox1 and plastid rbcL genes of Gracilaria
vermiculophylla (Gracilariaceae, Rhodophyta). Journal of Applied Phycology,
20(2), 161-168.
Yang, M. Y., Geraldino, P. J. L., & Kim, M. S. (2013a). DNA barcode assessment of
Gracilaria salicornia (Gracilariaceae, Rhodophyta) from Southeast Asia.
Botanical Studies, 54(1), 27.
Yang, M. Y., Han, E. G., & Kim, M. S. (2013b). Molecular identification of
Grateloupia elliptica and G. lanceolata (Rhodophyta) inferred from plastid rbcL
and mitochondrial COI genes sequence data. Genes & Genomics, 35(2), 239-246.
Univers
ity of
Mala
ya

158
Yang, M. Y., & Kim, M. S. (2014). Taxonomy of Grateloupia (Halymeniales,
Rhodophyta) by DNA barcode marker analysis and a description of
Pachymeniopsis volvita sp. nov. Journal of Applied Phycology, 27(3), 1373-
1384.
Yang, Z., & Rannala, B. (2012). Molecular phylogenetics: principles and practice.
Nature Reviews Genetics, 13(5), 303-314.
Yi, H., Sui, Z., & Bao, Z. (2010). Preliminary analysis of population genetic diversity of
cultivated Laminaria japonica sporophyte via AFLP technique. Journal of
Ocean University of China, 9(1), 53-58.
Yoon, H. S., Müller, K. M., Sheath, R. G., Ott, F. D., & Bhattacharya, D. (2006).
Defining the major lineages of red algae (Rhodophyta). Journal of Phycology,
42(2), 482-492.
Yoon, H. S., Zuccarello, G. C., & Bhattacharya, D. (2010). Evolutionary history and
taxonomy of red algae. In J. Seckbach & D. J. Chapman (Eds.), Red algae in the
genomic age (Vol. 13, pp. 25-42). Netherlands: Springer Science.
Yoshida, T., Stiger, V., & Horiguchi, T. (2000). Sargassum boreale sp. nov.(Fucales,
Phaeophyceae) from Hokkaido, Japan. Phycological Research, 48(3), 125-131.
Yow, Y. Y., Lim, P. E., & Phang, S. M. (2011). Genetic diversity of Gracilaria changii
(Gracilariaceae, Rhodophyta) from west coast, Peninsular Malaysia based on
mitochondrial cox1 gene analysis. Journal of Applied Phycology, 23(2), 219-226.
Yow, Y. Y., Lim, P. E., & Phang, S. M. (2013). Assessing the use of mitochondrial
cox1 gene and cox2-3 spacer for genetic diversity study of Malaysian Gracilaria
changii (Gracilariaceae, Rhodophyta) from Peninsular Malaysia. Journal of
Applied Phycology, 25(3), 831-838.
Zanardini, G. (1851). Algae novae vel minus cognitae in mari Rubro a Portiero
collectae. Flora, 34, 33-38.
Zanardini, G. (1858). Plantarum in mari Rubro hucusque collectarum enumeratio
(juvante A. Figari). Memorie delle Reale istituto Veneto di scienze, lettere ed
arti, 7, 209-309.
Univers
ity of
Mala
ya

159
Zhang, D., Sui, Z. H., Yi, H., Tong, Z., & Bao, Z. M. (2009). The genetic diversity
analysis of the sporophyte s of Undaria pinnatifida (Phaeophyceae) for Qingdao
population by AFLP markers. Periodical of Ocean University of China, 39(4),
664-668.
Zuccarello, G. C., Buchanan, J., & West, J. A. (2006). Increased sampling for inferring
phylogeographic patterns in Bostrychia radicans/ B. moritziana
(Rhodomelaceae, Rhodophyta) in the eastern USA. Journal of Phycology, 42(6),
1349-1352.
Zuccarello, G. C., Sandercock, B., & West, J. (2002). Diversity within red algal species:
variation in world-wide samples of Spyridia filamentosa (Ceramiaceae) and
Murrayella periclados (Rhodomelaceae) using DNA markers and breeding
studies. European Journal of Phycology, 37(3), 403-417.
Zuccarello, G. C., & West, J. A. (2002). Phylogeography of the Bostrychia calliptera–
B. pinnata complex (Rhodomelaceae, Rhodophyta) and divergence rates based
on nuclear, mitochondrial and plastid DNA markers. Phycologia, 41(1), 49-60.
Zuccarello, G. C., & West, J. A. (2003). Multiple cryptic species: molecular diversity
and reproductive isolation in the Bostrychia radicans/B. moritziana complex
(Rhodomelaceae, Rhodophyta) with focus on North American isolates. Journal
of Phycology, 39(5), 948-959.
Zuccarello, G. C., & West, J. A. (2011). Insights into evolution and speciation in the red
alga Bostrychia: 15 years of research. Algae, 26(1), 21-32.
Zuccarello, G. C., West, J. A., Kamiya, M., & King, R. J. (1999). A rapid method to
score plastid haplotypes in red seaweeds and its use in determining parental
inheritance of plastids in the red alga Bostrychia (Ceramiales). Hydrobiologia,
401, 207-214.
Univers
ity of
Mala
ya

160
LIST OF PUBLICATIONS AND PAPERS PRESENTED
Publications
1. Tan, P.-L., Lim, P.-E., Lin, S.-M., Phang, S.-M., Draisma, S. G. A., & Liao, L. M.
(2015). Foliose Halymenia species (Halymeniaceae, Rhodophyta) from Southeast
Asia, including a new species, Halymenia malaysiana sp. nov. Botanica Marina,
58(3), 203-217.
2. Tan, P.-L., Lim, P.-E., Lin, S.-M., & Phang, S.-M. (2017). Halymenia johorensis sp.
nov. (Halymeniaceae, Rhodophyta), a new foliose red algal species from Malaysia.
Journal of Applied Phycology. doi:10.1007/s10811-017-1104-8
Conference and seminar presentations
1. Phylogeny Study of Halymenia from Parts of South East Asian Countries at the 3rd
World Conference on Marine Diversity, 12-16 October 2014, Qingdao, China
2. Phylogenetic relationships of Halymenia (Halymeniaceae, Rhodophyta) from
Southeast Asia based on rbcL sequences at the 19th
Biological Sciences Graduate
Congress (BSGC), 12-13 December 2014, National University of Singapore,
Singapore.
3. Introduction to foliose Halymenia (Halymeniaceae, Rhodophyta) from Southeast
Asia including the description of one new species, Halymenia malaysiana for the
Institute of Biological Sciences Postgraduate Seminar, University of Malaya, on the
22 April 2015.
Posters presented
1. Molecular delineation of species in Halymenia (Halymeniaceae, Rhodophyta) from
Southeast Asia at the 9th
Asia-Pacific Conference on Algal Biotechnology (APCAB),
15-18 November 2016, Bangkok, Thailand.
Univers
ity of
Mala
ya

Botanica Marina 2015; 58(3): 203–217
*Corresponding authors: Phaik-Eem Lim, Instititute of Ocean and Earth Sciences, University of Malaya, 50603 Kuala Lumpur, Malaysia; and Institute of Biological Sciences, University of Malaya, 50603 Kuala Lumpur, Malaysia, e-mail: [email protected]; and Showe-Mei Lin, Institute of Marine Biology, National Taiwan Ocean University, Keelung 20224, Taiwan, R.O.C., e-mail: [email protected] Tan and Siew-Moi Phang: Instititute of Ocean and Earth Sciences, University of Malaya, 50603 Kuala Lumpur, Malaysia; and Institute of Biological Sciences, University of Malaya, 50603 Kuala Lumpur, MalaysiaStefano G.A. Draisma: Instititute of Ocean and Earth Sciences, University of Malaya, 50603 Kuala Lumpur, Malaysia; and Faculty of Science, Department of Biology, Prince of Songkla University, Hatyai, Songkhla, 90112 ThailandLawrence M. Liao: Graduate School of Biosphere Science, Hiroshima University, 1-4-4 Kagamiyama, Higashi-Hiroshima 739-8528, Japan
Pui-Ling Tan, Phaik-Eem Lim*, Showe-Mei Lin*, Siew-Moi Phang, Stefano G.A. Draisma and Lawrence M. Liao
Foliose Halymenia species (Halymeniaceae, Rhodophyta) from Southeast Asia, including a new species, Halymenia malaysiana sp. nov.
Abstract: Despite the large number of species discov-ered in Halymenia, many remain poorly known due to the scarce information available. In order to facilitate species discrimination of foliose Halymenia species in Southeast Asia, molecular analysis and morphologi-cal studies were made on Halymenia collections from Malaysia, the Philippines, and Indonesia. The rbcL phy-logenetic analyses showed that there are at least six taxa of foliose Halymenia occurring in Southeast Asia. Among the six taxa, a new species, Halymenia malaysiana P.-L. Tan, P.-E. Lim, S.-M. Lin et S.-M. Phang, is proposed based on both rbcL sequence analyses and morphologi-cal observations. Halymenia malaysiana is characterized by thalli possessing oblong or suborbiculate blades with a supple cartilaginous structure and gelatinous (slimy) texture, arising from a small discoid holdfast without a stipe, abruptly expanding into a broad blade and having a smooth surface with sinusoidally undulated margins. The phylogenetic analyses also revealed that Halymenia is a polyphyletic genus, which requires further taxonomic studies.
Keywords: Halymenia malaysiana sp. nov.; red algae; Southeast Asia; taxonomy.
DOI 10.1515/bot-2015-0004Received 7 January, 2015; accepted 24 April, 2015
IntroductionThe red algal genus Halymenia C. Agardh, comprising 69 currently accepted species, is one of the largest genera in terms of species within the family Halymeniaceae (De Smedt et al. 2001, Guiry and Guiry 2014). It is mostly dis-tributed in tropical and subtropical regions (Gargiulo et al. 1986, Kawaguchi and Lewmanomont 1999, Hernán-dez-Kantun et al. 2009). The genus is mainly character-ized by gelatinous thalli, presence of anticlinal filaments and refractive ganglionic cells in the medulla, stellate cells in the inner cortex, and auxiliary cell ampullae with branched secondary filaments (Balakrishnan 1961, Abbott 1967, Chiang 1970, De Smedt et al. 2001).
Halymenia was established by C. Agardh (1817) and the generitype is Halymenia floresii (Clemente) C. Agardh collected from Cádiz, Spain. Chiang (1970) used the archi-tecture of auxiliary cell ampullae as a primary feature to group species at the generic level in the Halymeniaceae. According to Chiang’s generic concept, simple or once or more branched secondary ampullar filaments may emerge from long and slender primary ampullary filaments in Halymenia-type auxiliary cell ampullae. The auxiliary cell ampulla of Halymenia is flattish, expanded when mature, and is intermediate between the Grateloupia-type and the Cryptonemia-type of ampulla based on its shape and the degree of branching (Chiang 1970). Several attempts have been made to study foliose Halymenia species in Southeast Asia. Kawaguchi and Lewmanomont (1999) made a detailed morphological study of Halyme-nia dilatata Zanardini by comparing the vegetative and reproductive features of the material from Vietnam and
Brought to you by | National Taiwan Ocean UniversityAuthenticated | [email protected]
Download Date | 6/23/15 2:16 AM
Univers
ity of
Mala
ya

9TH ASIA PACIFIC CONFERENCE ON ALGAL BIOTECHNOLOGY- BANGKOK
Halymenia johorensis sp. nov. (Halymeniaceae, Rhodophyta),a new foliose red algal species from Malaysia
Pui-Ling Tan1,2& Phaik -Eem Lim1
& Showe-Mei Lin3& Siew-Moi Phang1,2
Received: 5 December 2016 /Revised and accepted: 19 February 2017# Springer Science+Business Media Dordrecht 2017
Abstract A new Halymenia species, Halymenia johorensissp. nov., from southern Peninsular Malaysia is proposed basedon plastid-encoded large subunit of ribulose-1,5-bisphosphatecarboxylase/oxygenase (rbcL) gene analyses and detailedmorphological observations. The new species is characterizedby having (1) elliptical, oblong, or irregularly shaped blades,incised with some perforations, arising from a narrow-cuneatestipe attached to a discoid holdfast; (2) blades with a cartilag-inous and gelatinous texture, a smooth to rugose surface, andirregularly dentate and cleft margins; and (3) isodiametric out-er cortical cells and rounded to stellate inner cortical cells.RbcL sequence analyses have shown H. johorensis to be ge-netically distinct from other Halymenia species. AlthoughH. johorensis is sister to Halymenia plana, these two speciescan be distinguished both molecularly and morphologically.Further studies are necessary to investigate the phylogeneticrelationships and species diversity in this genus.
Keywords Halymenia johorensis . Malaysia . New species .
rbcL . Rhodophyta . Taxonomy
Introduction
The marine red algal genus Halymenia C. Agardh is one ofseveral species-rich red algal genera in the familyHalymeniaceae and includes 79 taxonomically accepted spe-cies (Guiry and Guiry 2016), widely distributed in tropical andsubtropical regions (Gargiulo et al. 1986; Hernández-Kantúnet al. 2012; Tan et al. 2015). Agardh (1817) established theHalymenia based on the generitype, Halymenia floresii(Clemente) C. Agardh, from Cádiz, Spain.
Generic concepts of Halymenia are based largely on thestructure of auxiliary cell ampullae, as proposed by Chiang(1970), and the presence of anticlinal medullary filaments(Abbott 1967). However, several authors believe that vegeta-tive features are more diagnostic than reproductive characters,which overlap greatly among genera of Halymeniaceae(Abbott 1967; Guimarães and Fujii 1998; Hernández-Kantún et al. 2012). Vegetative features used to delineate spe-cies of Halymenia include habit, thallus size, blade margin,order of branching, presence or absence of a midrib in thebasal region, presence or absence of a stipe, presence or ab-sence of marginal proliferations, presence or absence of papil-lae or spines on thallus surface, blade thickness, cortex thick-ness, shape of inner cortical cells, inner cortical cell size, andpresence or absence of refractive ganglionic cells (Guimarãesand Fujii 1998; De Smedt et al. 2001; Ballantine and Ruiz2004; Hernández-Kantún et al. 2012; Tan et al. 2015;Azevedo et al. 2016).
To date, six species of Halymenia have been reported fromMalaysia, including three foliose species, H. dilatataZanardini, H. maculata J. Agardh, and H. malaysiana P-LTan, P-E Lim, S-M Lin & S-M Phang (Kawaguchi et al.2002; Tan et al. 2015; Phang et al. 2016). Kawaguchi et al.(2002) confirmed the existence of two foliose species(H. dilatata and H. maculata) and one branched species
* Phaik -Eem [email protected]
1 Institute of Ocean and Earth Sciences, University of Malaya,50603 Kuala Lumpur, Malaysia
2 Institute of Biological Sciences, University of Malaya, 50603 KualaLumpur, Malaysia
3 Institute of Marine Biology, National Taiwan Ocean University,Keelung 20224, Taiwan, Republic of China
J Appl PhycolDOI 10.1007/s10811-017-1104-8
Univers
ity of
Mala
ya

16
1
Appendix A: List of specimens examined in this study with information on herbarium number, locality, collector, date of collection and field number.
Taxa Herbarium
number
Locality and collector Date of collection Field number DNA sequences and GenBank accession
numbers where applicable rbcL/COI-
5P/UPA/LSU (28S rDNA)
Halymenia cf. dilatata
Zanardini
PSM12857 Camiguin Island, Philippines
S. Draisma
4 April 2014 SGAD1404050 + (KP202340) / - / -
Halymenia durvillei
Bory de Saint-
Vincent
PSM12899 Pulau Labuan, Sabah, Malaysia
S. Draisma
28 June 2012 1206070 + / + / - / -
H. durvillei PSM12986 Pulau Merambong, Johor,
Malaysia
PLT
17 December 2014 J15 + / + / + / -
H. durvillei PSM12989 Pulau Merambong, Johor,
Malaysia
PLT
17 December 2014 J19 + / + / + / -
H. durvillei PSM12903 Tun Mustapha Park (TMP),
Sabah, Malaysia
S. Draisma
17 September 2012 SGAD1209202 + / + / + / -
H. durvillei PSM12905 Tun Mustapha Park (TMP),
Sabah, Malaysia
S. Draisma
17 September 2012 SGAD1209204 + / + / + / +
H. durvillei PSM12901 Tun Mustapha Park (TMP),
Sabah, Malaysia
S. Draisma
7 September 2012 SGAD1209005 + / + / + / +
H. durvillei PSM12894 Pulau Babi Besar, Johor,
Malaysia
S. Draisma
30 May 2012 SGAD1205234 + / + / + / +
H. durvillei PSM12929 Busuanga Island, Calamian
Islands, Philippines
S. Draisma
1 March 2014 SGAD1403004 + / + / - / -
Univers
ity of
Mala
ya

16
2
Appendix A, continued
Taxa Herbarium
number
Locality and collector Date of collection Field number DNA sequences and GenBank accession
numbers where applicable rbcL/COI-
5P/UPA/LSU (28S rDNA)
H. durvillei PSM12939 Busuanga Island, Calamian
Islands, Philippines
S. Draisma
4 March 2014 SGAD1403091 + / + / - / -
H. durvillei PSM12942 Uson Island, Calamian Islands,
Philippines
S. Draisma
6 March 2014 SGAD1403116 + / + / - / -
H. durvillei PSM12925 Paglugaban Island, Palawan,
Philippines
S. Draisma
24 January 2014 SGAD1401050 + / + / + / -
H. durvillei PSM12884 Pulau Mata Pahi, Semporna,
Sabah, Malaysia
S. Draisma
17 December 2010 SGAD1012344 + / + / + / +
H. durvillei PSM12885 Pulau Mata Pahi, Semporna,
Sabah, Malaysia
S. Draisma
17 December 2010 SGAD1012346 + / + / + / +
H. durvillei PSM12836 Pulau Karindingan, Sabah,
Malaysia
PEL
23 June 2010 S6 + / - / + / -
H. durvillei PSM12837 Pulau Karindingan, Sabah,
Malaysia
PEL
23 June 2010 S7 + / + / - / -
H. durvillei PSM12968 Pulau Weh, Aceh, Indonesia
S. Draisma
29 April 2014 SGAD1405001 + / + / + / +
H. durvillei PSM12970 Pulau Weh, Aceh, Indonesia
S. Draisma
6 May 2014 SGAD1405014 + / + / + / -
H. durvillei SGAD0712160* Gam Island, Raja Ampat,
Indonesia
S. Draisma
25 November 2007 SGAD0712160 + / + / + / - Univ
ersity
of M
alaya

16
3
Appendix A, continued
Taxa Herbarium
number
Locality and collector Date of collection Field number DNA sequences and GenBank accession
numbers where applicable rbcL/COI-
5P/UPA/LSU (28S rDNA)
H. durvillei SGAD0712161* Gam Island, Raja Ampat,
Indonesia
S. Draisma
25 November 2007 SGAD0712161 + / + / + / -
H. durvillei SGAD0712476* Gam Island, Raja Ampat,
Indonesia
S. Draisma
4 December 2007 SGAD0712476 + / + / + / +
H. durvillei PSM12913 Ternate Island, Pantar Strait,
East Nusa Tenggara, Indonesia
S. Draisma
4 April 2013 SGAD1304049 + / + / - / -
H. durvillei PSM12954 Siargao Island, Philippines
S. Draisma
14 April 2014 SGAD1404200 + / + / - / -
H. durvillei PSM12911 Pulau Rote, East Nusa
Tenggara, Indonesia
S. Draisma
15 November 2012 SGAD1211004 + / + / + / +
H. durvillei PSM13015 Padang Bai, Bali, Indonesia
S. Draisma
24 June 2015 SGAD1506097 + / + / - / -
H. durvillei PSM13011 Amed, Bali, Indonesia
S. Draisma
20 June 2015 SGAD1506071 + / + / - / -
H. durvillei PSM12999 Ekas, Pulau Lombok, Indonesia
PEL
15 May 2015 C1 + / + / + / -
H. durvillei PSM13000 Ekas, Pulau Lombok, Indonesia
PEL
15 May 2015 C2 + / + / + / -
H. durvillei PSM13001 Ekas, Pulau Lombok, Indonesia
PEL
15 May 2015 C3 + / + / + / -
H. durvillei PSM13002 Ekas, Pulau Lombok, Indonesia
PEL
15 May 2015 C4 + / + / + / -
Univers
ity of
Mala
ya

16
4
Appendix A, continued
Taxa Herbarium
number
Locality and collector Date of collection Field number DNA sequences and GenBank accession
numbers where applicable rbcL/COI-
5P/UPA/LSU (28S rDNA)
H. durvillei PSM12947 Camiguin Island, Philippines
S. Draisma
4 April 2014 SGAD1404077 + / + / + / -
H. durvillei PSM12948 Camiguin Island, Philippines
S. Draisma
4 April 2014 SGAD1404081 + / + / + / +
H. durvillei PSM12951 Camiguin Island, Philippines
S. Draisma
6 April 20114 SGAD1404128 + / + / - / -
Halymenia johorensis P.-L. Tan, P.-E. Lim,
S.-M. Lin & S.-M.
Phang sp. nov.
PSM12870 Merambong Shoal, Johor,
Malaysia
PEL
23 August 2009 S8 + (KX958402) / + / + / +
H. johorensis PSM12871 Merambong Shoal, Johor,
Malaysia
PEL
23 August 2009 S1 + (KX958403) / + / + / +
H. johorensis PSM12875 Pulau Merambong, Johor,
Malaysia
PEL
23 August 2009 S2 + / + / + / +
H. johorensis PSM12872 Merambong Shoal, Johor,
Malaysia
PEL
23 August 2009 S12 + / + / - / -
H. johorensis PSM12873 Merambong Shoal, Johor,
Malaysia
PEL
23 August 2009 S13 + / + / + / -
H. johorensis PSM12874 Pulau Merambong, Johor,
Malaysia
PEL
23 August 2009 S14 + / - / - / -
H. johorensis PSM10663 Merambong Shoal, Johor,
Malaysia
PEL
10 March 2009 S11 + / + / + / - Univ
ersity
of M
alaya

16
5
Appendix A, continued
Taxa Herbarium
number
Locality and collector Date of collection Field number DNA sequences and GenBank accession
numbers where applicable rbcL/COI-
5P/UPA/LSU (28S rDNA)
H. johorensis PSM12832 Pulau Merambong, Johor,
Malaysia
PEL
28 February 2009 S15 + / + / - / -
H. johorensis PSM12882 Merambong Shoal, Johor,
Malaysia
PEL
13 July 2014 B1 + (KX958404) / + / + / +
Halymenia maculata J.
Agardh
PSM12833 Pulau Merambong, Johor,
Malaysia
PEL
24 August 2009 S3 + (KP202334) / - / + / +
H. maculata PSM12840 Tanjong Tigasamil, Tun
Mustapha Park (TMP), Sabah,
Malaysia
S. Draisma
16 September 2012 SGAD1209183 + (KP202335) / + / + / +
H. maculata PSM12841 Tun Mustapha Park (TMP),
Sabah, Malaysia
S. Draisma
17 September 2012 SGAD1209209 + (KP202336) / + / + / -
H. maculata PSM120906 Tun Mustapha Park (TMP),
Sabah, Malaysia
S. Draisma
17 September 2012 SGAD1209208 + / - / + / -
H. maculata PSM12902 Tun Mustapha Park (TMP),
Sabah, Malaysia
Bert Hoeksema
16 September 2012 SGAD1209190 + / - / + / -
H. maculata
PSM12897 Pulau Labuan, Sabah, Malaysia
S. Draisma
28 June 2012 SGAD1206066 + / - / - / -
Halymenia malaysiana
P.-L. Tan, P.-E. Lim,
S.-M. Lin et S.-M.
Phang sp. nov.
PSM12834 Pulau Merambong, Johor,
Malaysia
PEL
24 August 2009 S4 + (KP202322) / + / + / - Univers
ity of
Mala
ya

16
6
Appendix A, continued
Taxa Herbarium
number
Locality and collector Date of collection Field number DNA sequences and GenBank accession
numbers where applicable rbcL/COI-
5P/UPA/LSU (28S rDNA)
H. malaysiana PSM12835 Pulau Merambong, Johor,
Malaysia
PEL
24 August 2009 S5 + (KP202323) / + / + / -
H. malaysiana PSM12838 Pulau Besar, Malacca, Malaysia
S. Draisma
12 July 2010 SGAD1007001 + (KP202324) / + / + / -
H. malaysiana PSM12850 Pulau Karindingan, Sabah,
Malaysia
PEL
23 June 2010 S10 + (KP202325) / + / - / -
H. malaysiana PSM12845 Siargao Island, Philippines
S. Draisma
10 April 2014 SGAD1404132 + (KP202326) / + / - / -
H. malaysiana PSM12846 Siargao Island, Philippines
S. Draisma
12 April 2014 SGAD1404182 + (KP202327) / + / - / -
H. malaysiana PSM12847 Grande Island, Subic Bay,
Luzon, Philippines
S. Draisma
29 July 2014 SGAD1406043 + (KP202328) / + / - / -
H. malaysiana PSM12848 Subic Bay, Luzon, Philippines
S. Draisma
29 July 2014 SGAD1406045 + (KP202329) / + / - / -
H. malaysiana PSM12851 Busuanga Island, Calamian
Islands, Philippines
S. Draisma
1 March 2014 SGAD1403003 + (KP202330) / + / - / -
H. malaysiana PSM12853 Busuanga Island, Calamian
Islands, Philippines
S. Draisma
4 March 2014 SGAD1403055 + (KP202331) / + / - / -
H. malaysiana PSM12854 Busuanga Island, Calamian
Islands, Philippines
S. Draisma
4 March 2014 SGAD1403087 + (KP202332) / + / - / - Univers
ity of
Mala
ya

16
7
Appendix A, continued
Taxa Herbarium
number
Locality and collector Date of collection Field number DNA sequences and GenBank accession
numbers where applicable rbcL/COI-
5P/UPA/LSU (28S rDNA)
H. malaysiana SGAD0712156* Gam Island, Raja Ampat,
Indonesia
S. Draisma
25 November 2007 SGAD0712156 + (KP202333) / - / - / -
H. malaysiana SGAD0712471* Gam Island, Raja Ampat,
Indonesia
S. Draisma
3 December 2007 SGAD0712471 + / + / - / -
H. malaysiana PSM12890
Tanjung Gua Hujan, Pulau
Tinggi, Johor, Malaysia
S. Draisma
28 May 2012 SGAD1205185 + / + / + / -
H. malaysiana PSM12891 Tanjung Gua Hujan, Pulau
Tinggi, Johor, Malaysia
S. Draisma
28 May 2012 SGAD1205198 + / + / + / -
H. malaysiana PSM12893 Ibol, Pulau Tinggi, Johor
Malaysia
S. Draisma
29 May 2012 SGAD1205213 + / + / + / -
H. malaysiana PSM12900 Tun Mustapha Park (TMP),
Sabah, Malaysia
S. Draisma
7 September 2012 SGAD1209001 + / + / + / +
H. malaysiana PSM12904 Tun Mustapha Park (TMP),
Sabah, Malaysia
S. Draisma
17 September 2012 SGAD1209203 + / + / + / +
H. malaysiana PSM12909 Tun Mustapha Park (TMP),
Sabah, Malaysia
S. Draisma
23 September 2012 SGAD1209396 + / + / + / -
H. malaysiana PSM12927 Busuanga Island, Calamian
Islands, Philippines
S. Draisma
1 March 2014 SGAD1403001 + / + / - / -
Univers
ity of
Mala
ya

16
8
Appendix A, continued
Taxa Herbarium
number
Locality and collector Date of collection Field number DNA sequences and GenBank accession
numbers where applicable rbcL/COI-
5P/UPA/LSU (28S rDNA)
H. malaysiana PSM12935 Busuanga Island, Calamian
Islands, Philippines
S. Draisma
4 March 2014 SGAD1403056 + / + / - / -
H. malaysiana PSM12972 Pulau Merambong, Johor,
Malaysia
PLT
17 December 2014 J1 + / + / + / -
H. malaysiana PSM12973 Pulau Merambong, Johor,
Malaysia
PLT
17 December 2014 J2 + / + / + / -
H. malaysiana PSM12974 Pulau Merambong, Johor,
Malaysia
PLT
17 December 2014 J3 + / + / + / -
H. malaysiana PSM12975 Pulau Merambong, Johor,
Malaysia
PLT
17 December 2014 J4 + / + / + / -
H. malaysiana PSM12976 Pulau Merambong, Johor,
Malaysia
PLT
17 December 2014 J5 + / + / + / -
H. malaysiana PSM12977 Pulau Merambong, Johor,
Malaysia
PLT
17 December 2014 J6 + / + / + / -
H. malaysiana PSM12978 Pulau Merambong, Johor,
Malaysia
PLT
17 December 2014 J7 + / + / + / -
H. malaysiana PSM12980 Pulau Merambong, Johor,
Malaysia
PLT
17 December 2014 J9 + / + / + / - Univers
ity of
Mala
ya

16
9
Appendix A, continued
Taxa Herbarium
number
Locality and collector 17 December 2014 Field number DNA sequences and GenBank accession
numbers where applicable rbcL/COI-
5P/UPA/LSU (28 S rDNA)
H. malaysiana PSM12981 Pulau Merambong, Johor,
Malaysia
PLT
17 December 2014 J10 + / + / + / -
H. malaysiana PSM12983 Pulau Merambong, Johor,
Malaysia
PLT
17 December 2014 J12 + / + / + / -
H. malaysiana PSM12984 Pulau Merambong, Johor,
Malaysia
PLT
17 December 2014 J13 + / + / + / -
H. malaysiana PSM12985 Pulau Merambong, Johor,
Malaysia
PLT
17 December 2014 J14 + / + / + / -
H. malaysiana PSM12987 Pulau Merambong, Johor,
Malaysia
PLT
17 December 2014 J16 + / + / + / -
H. malaysiana PSM12990 Pulau Merambong, Johor,
Malaysia
PLT
17 December 2014 J20 + / + / + / -
H. malaysiana PSM12992 Pulau Merambong, Johor,
Malaysia
PLT
17 December 2014 J22 + / + / + / -
H. malaysiana PSM12993 Pulau Merambong, Johor,
Malaysia
PLT
17 December 2014 J24 + / + / + / -
H. malaysiana PSM12994 Pulau Merambong, Johor,
Malaysia
PLT
17 December 2014 J25 + / + / + / - Univers
ity of
Mala
ya

17
0
Appendix A, continued
Taxa Herbarium
number
Locality and collector 17 December 2014 Field number DNA sequences and GenBank accession
numbers where applicable rbcL/COI-
5P/UPA/LSU (28S rDNA)
H. malaysiana PSM12995 Pulau Merambong, Johor,
Malaysia
PLT
17 December 2014 J26 + / + / + / -
H. malaysiana PSM12996 Pulau Merambong, Johor,
Malaysia
PLT
17 December 2014 J30 + / + / + / -
H. malaysiana PSM12997 Pulau Merambong, Johor,
Malaysia
PLT
17 December 2014 J31 + / + / + / -
H. malaysiana PSM12998 Pulau Merambong, Johor,
Malaysia
PLT
17 December 2014 J33 + / + / + / -
Halymenia sp. A PSM12887 Koh Dok Mai, Thailand
S. Draisma
11 August 2012 SGAD1208001 + / + / + / +
Halymenia sp. A PSM12888 Koh Dok Mai, Thailand
S. Draisma
11 August 2012 SGAD1208002 + / + / + / +
Halymenia tondoana
O. DeClerck et J.J.
Hernández-Kantun
PSM12955 Siargao Island, Philippines
S. Draisma
15 April 2014 1404246 + / + / + / +
H. tondoana PSM12952 Siargao Island, Philippines
S. Draisma 10 April 2014 1404143 + / + / - / -
H. tondoana PSM12956 Siargao Island, Philippines
S. Draisma
15 April 2014 1404247 + / + / + / +
GenBank accession numbers were provided for taxa of which DNA sequences were published in Tan et al., 2014. Abbreviations for collector are: S. Draisma – Stefano G.A.
Draisma; PEL – Phaik Eem Lim; PLT – Pui Ling Tan. PSM indicate reference code of herbarium at the University of Malaya Seaweeds and Seagrasses Herbarium (KLU). The
herbaria deposited in Leiden Herbarium are denoted with ‘*’. Sequences obtained and included in phylogenetic analyses are denoted with ‘+’. Unavailable sequences or irrelevant
data are denoted with ‘- .́
Univers
ity of
Mala
ya

17
1
Appendix B: List of published sequences used for rbcL analyses with collection details and GenBank accession numbers.
Species Collection information GenBank accession
number
Carpopeltis phyllophora (J.D. Hooker et Harvey) F. Schmitz Black Island, Esperance Bay, Western Australia FN908147
Codiophyllum natalense J.E. Gray Protea Banks, Northern Pinnacle, Kwazulu-Natal,
South Africa
FN908160
Cryptonemia lomation (Bertoloni) J. Agardh Calvi, Cap de la Revellata FN908155
Cryptonemia luxurians (C. Agardh) J. Agardh Nagaiso, Namikata, Ehime Prefecture, Japan AB061374
Epiphloea bullosa (Harvey) De Toni Julien Bay, Escape Island, Western Australia FN908149
Galene meridionalis D'Archino et Zuccarello Port Pegasus, Stewart Island, New Zealand KJ606650
Galene profundae D'Archino et Zuccarello Hou Hou Point, Marlborough Sounds, South
Island, New Zealand
KJ606645
Gelinaria ulvoidea Sonder Julien Bay, Escape Island, Western Australia FN908154
Halymenia cearensis C.A.A.Azevedo, Cassano &
M.C.Oliveira
Ceara, Brazil KX586174
Halymenia dilatata Zanardini Debana, Hachijo Island, Tokyo, Japan AB038604
Halymenia durvillei Bory de Saint-Vincent Pulau Rebak Besar, Langkawi, Kedah, Malaysia AB038603
H. durvillei Beruwela, Sri Lanka AY772020
Halymenia floresii (Clemente) C. Agardh Illes Formigues, Palamos, Girona, Spain AY772019
H. floresii Armacao de Pere, Portugal KU382051
Halymenia harveyana J. Agardh Jurien Bay, Escape Island, Western Australia JQ976885
Univers
ity of
Mala
ya

17
2
Appendix B, continued
Species Collection information GenBank accession
number
H. harveyana Quarter Mile Reef, Sodwana Bay, South Africa JQ976886
Halymenia hawaiiana J.J. Hernández-Kantun et A.R.
Sherwood
Pupukea, Oahu, Hawaii, USA AM422899
Halymenia ignifera C.A.A.Azevedo, Cassano et
M.C.Oliveira
Rio do Fogo Beach, Rio do Fogo, RN, Brazil KT807829
Halymenia maculata J. Agardh Pulau Satang Besar, Santubong, Kuching,
Sarawak, Malaysia
AB061397
Halymenia pinnatifida C.A.A.Azevedo, Cassano et
M.C.Oliveira
Rio do Fogo Beach, Rio do Fogo, RN, Brazil KT807828
Halymenia plana Zanardini Reef west of Verona Sands boat launch, in mouth
of Huon River, Tasmania, Australia
KU726713
Halymenia silviae C.A.A.Azevedo, Cassano et M.C.Oliveira Rio do Fogo Beach, Rio do Fogo, RN, Brazil KT807827
Halymenia stipitata I.A. Abbott Kaalawai, Oahu, Hawaii JQ976891
Halymenia tondoana O. DeClerck et J.J. Hernández-Kantun Dancalan, N of Bulusan, SE Luzon, Philippines JQ976888
Spongophloea tissotii (Weber-van Bosse) Huisman, De
Clerck, Prud'homme van Reine et Borowitzka
Monkey Mia, Shark Bay, Western Australia FN908162
Thamnoclonium dichotomum (J.Agardh) J.Agardh North Twin Peaks Island, Esperance Bay, Western
Australia
FN908151
Thamnoclonium lemannianum Harvey Cottesloe, Western Australia FN908161
Univers
ity of
Mala
ya

17
3
Appendix C: List of published COI-5P, UPA and LSU (28S rDNA) sequences with collection details and GenBank accession numbers for analyses.
Species Collection information GenBank accession number
COI-5P UPA LSU
Grateloupia angusta (Okamura) S.Kawaguchi &
H.W.Wang
Jeju, South Korea - KJ648575 -
Grateloupia elliptica Holmes Jeju, South Korea JX475023 - -
Grateloupia lanceolata (Okamura) S.Kawaguchi Spain - - AM420418
Grateloupia subpectinata Holmes Jeju, South Korea - KF543073 -
Grateloupia turuturu Yamada Japan - - AM420447
Halymenia californica G.M.Smith & Hollenberg California, Lover's Point, Pacific Grove, USA KM254858 - -
Halymenia cearensis C.A.A.Azevedo, Cassano &
M.C.Oliveira
Ceara, Brazil KX586151 - -
H. cearensis Ceara, Brazil KX586166 KX586140 -
Halymenia floresii (Clemente) C.Agardh Armacao de Pere, Portugal GQ862071 - GQ471912
Halymenia formosa Harvey ex Kützing Oahu, Hawaii, USA - HQ421565 HQ422446
Halymenia ignifera C.A.A.Azevedo, Cassano &
M.C.Oliveira
Rio Grande do Norte, Brazil KT807817 KX586118 -
H. ignifera Espirito Santo, Brazil KT807819 KX586122 -
Halymenia maculata J. Agardh Lord Howe Island, New South Wales, Australia - - GQ471913
Halymenia pinnatifida C.A.A.Azevedo, Cassano
& M.C.Oliveira
Rio Grande do Norte, Brazil KT807811 KT586108 -
Halymenia plana Zanardini Point Lonsdale Lighthouse Reef, Victoria,
Australia
- - GQ471914 Univ
ersity
of M
alaya

17
4
Appendix D: Uncorrected pairwise distance matrix of the rbcL sequences.
S1 KU726713 0712160 1304049 C4 1404200 1211004 1404077 1404128 AB038603 JX000238 JX000244 J19 1209005 S1 N KU726713 0.02480 0712160 0.05680 0.05680 1304049 0.05765 0.05765 0.00080 C4 0.05520 0.05520 0.00160 0.00240 1404200 0.05840 0.05840 0.00320 0.00400 0.00320 1211004 0.05680 0.05680 0.00160 0.00240 0.00160 0.00320 1404077 0.06080 0.05920 0.01040 0.01121 0.01040 0.01200 0.01040 1404128 0.06160 0.05840 0.01120 0.01201 0.01120 0.01280 0.01120 0.00080 AB038603 0.05680 0.05680 0.01120 0.01201 0.01120 0.01280 0.01120 0.01360 0.01440 JX000238 0.05871 0.05479 0.00587 0.00686 0.00587 0.00783 0.00489 0.00978 0.00978 0.00978 JX000244 0.06164 0.05577 0.01272 0.01371 0.01272 0.01468 0.01174 0.02055 0.02055 0.01957 0.01468 J19 0.05873 0.05546 0.01631 0.01714 0.01631 0.01794 0.01631 0.02202 0.02284 0.02121 0.01566 0.00098 1209005 0.06058 0.05735 0.01616 0.01698 0.01616 0.01777 0.01616 0.02181 0.02262 0.02100 0.01566 0.00098 0.00163 J15 0.06080 0.05760 0.01520 0.01601 0.01520 0.01680 0.01520 0.02080 0.02160 0.02080 0.01468 0.00000 0.00082 0.00081 1206070 0.06175 0.05854 0.01604 0.01685 0.01604 0.01764 0.01604 0.02005 0.02085 0.02005 0.01566 0.00098 0.00163 0.00162 S7 0.06109 0.05949 0.01608 0.01689 0.01608 0.01768 0.01608 0.02010 0.02090 0.01929 0.01370 0.00489 0.00489 0.00485 AY772020 0.06480 0.06320 0.01840 0.01761 0.01840 0.01840 0.01840 0.02400 0.02480 0.02160 0.01566 0.01957 0.02284 0.02262 JQ976882 0.06240 0.06240 0.01840 0.01761 0.01840 0.01840 0.01840 0.02240 0.02320 0.02000 0.01566 0.02153 0.02284 0.02262 JQ976884 0.06019 0.05728 0.01650 0.01555 0.01650 0.01650 0.01553 0.02233 0.02233 0.01942 0.01576 0.02167 0.02233 0.02233 JQ976886 0.04960 0.04480 0.02720 0.02802 0.02720 0.02880 0.02720 0.03120 0.03200 0.02880 0.02348 0.02935 0.03100 0.03312 JQ976885 0.05040 0.04560 0.02640 0.02722 0.02640 0.02800 0.02640 0.03040 0.03120 0.02800 0.02153 0.02740 0.03018 0.03231 AM422899 0.05600 0.05120 0.03600 0.03683 0.03600 0.03760 0.03600 0.04000 0.04080 0.03600 0.03327 0.03816 0.03670 0.03796 1403091 0.06160 0.06000 0.01760 0.01841 0.01760 0.01920 0.01760 0.02160 0.02240 0.02160 0.01468 0.00587 0.00571 0.00565 1405001 0.06374 0.06209 0.01904 0.01821 0.01904 0.01904 0.01904 0.02483 0.02566 0.02070 0.01531 0.01939 0.02280 0.02258 GQ995519 0.05600 0.05120 0.03600 0.03683 0.03600 0.03760 0.03600 0.04000 0.04080 0.03600 0.03327 0.03816 0.03670 0.03796 1404143 0.05600 0.05360 0.05040 0.05124 0.05040 0.05200 0.05040 0.05360 0.05440 0.04880 0.04599 0.05186 0.05302 0.05493 1404246 0.05721 0.05479 0.05077 0.05161 0.05077 0.05238 0.05077 0.05399 0.05479 0.04996 0.04697 0.05284 0.05383 0.05574
Univers
ity of
Mala
ya

17
5
Appendix D, continued
S1 KU726713 0712160 1304049 C4 1404200 1211004 1404077 1404128 AB038603 JX000238 JX000244 J19 1209005 1404247 0.05604 0.05364 0.05044 0.05124 0.05044 0.05204 0.05044 0.05364 0.05444 0.04884 0.04701 0.05289 0.05306 0.05497 JQ976888 0.05583 0.05337 0.04926 0.05008 0.04926 0.05090 0.04926 0.05255 0.05337 0.04844 0.04706 0.05294 0.05337 0.05419 1208001 0.06000 0.05840 0.05600 0.05685 0.05600 0.05760 0.05600 0.05920 0.06000 0.05600 0.05382 0.05969 0.05873 0.06058 KT807828 0.05444 0.05685 0.05124 0.05204 0.05124 0.05284 0.05124 0.05364 0.05444 0.05444 0.05191 0.05485 0.05143 0.05335 KT807827 0.04800 0.05200 0.04560 0.04644 0.04560 0.04720 0.04400 0.04800 0.04880 0.04560 0.04501 0.04990 0.04649 0.04847 1406043 0.06581 0.06501 0.05859 0.05939 0.05859 0.06019 0.05859 0.05778 0.05859 0.05778 0.05796 0.05894 0.05646 0.05916 S10 0.06501 0.06421 0.05778 0.05859 0.05778 0.05939 0.05778 0.05698 0.05778 0.05698 0.05697 0.05796 0.05565 0.05835 1403055 0.06421 0.06340 0.05698 0.05778 0.05698 0.05859 0.05698 0.05618 0.05698 0.05618 0.05599 0.05697 0.05483 0.05754 0712471 0.06782 0.06700 0.05873 0.05955 0.05873 0.06038 0.05873 0.05790 0.05873 0.05873 0.05964 0.06163 0.05955 0.06038 1209396 0.06640 0.06560 0.05920 0.06005 0.05920 0.06080 0.05920 0.05840 0.05920 0.05840 0.05773 0.05871 0.05710 0.05977 1404132 0.06581 0.06501 0.05859 0.05939 0.05859 0.06019 0.05859 0.05778 0.05859 0.05778 0.05697 0.05796 0.05646 0.05916 J1 0.06720 0.06640 0.06000 0.06085 0.06000 0.06160 0.06000 0.05920 0.06000 0.05920 0.05871 0.05969 0.05791 0.06058 1403056 0.06480 0.06400 0.05760 0.05845 0.05760 0.05920 0.05760 0.05680 0.05760 0.05680 0.05675 0.05773 0.05546 0.05816 AB038604 0.06560 0.06320 0.06080 0.06165 0.06080 0.06240 0.06080 0.06000 0.06080 0.06160 0.05871 0.05871 0.05873 0.06058 1209183 0.06748 0.06829 0.06341 0.06428 0.06341 0.06504 0.06341 0.06585 0.06667 0.06585 0.06556 0.06556 0.06362 0.06341 1209190 0.06913 0.06994 0.06511 0.06597 0.06511 0.06672 0.06511 0.06752 0.06833 0.06752 0.06556 0.06556 0.06444 0.06462 1206066 0.06800 0.06880 0.06400 0.06485 0.06400 0.06560 0.06400 0.06640 0.06720 0.06560 0.06360 0.06360 0.06281 0.06300 AB061397 0.07360 0.07360 0.06480 0.06565 0.06480 0.06640 0.06480 0.06720 0.06800 0.06640 0.06458 0.06458 0.06362 0.06300 1404050 0.06452 0.06532 0.06290 0.06371 0.06290 0.06290 0.06290 0.06532 0.06613 0.06290 0.06225 0.06225 0.06003 0.06026 FN908149 0.07040 0.06480 0.06160 0.06245 0.06160 0.06320 0.06160 0.06400 0.06480 0.06240 0.05675 0.05773 0.06117 0.06220 JQ976891 0.07760 0.07200 0.07920 0.08006 0.07920 0.08080 0.07920 0.08160 0.08240 0.07920 0.07926 0.08219 0.07993 0.08078 FN908154 0.06000 0.05520 0.05840 0.05925 0.05680 0.06000 0.05840 0.06080 0.06160 0.06000 0.05871 0.06067 0.06036 0.06220 AY772019 0.04800 0.04560 0.04720 0.04804 0.04560 0.04880 0.04720 0.04800 0.04880 0.04880 0.04697 0.05186 0.04812 0.05008 KU382051 0.05200 0.04960 0.04960 0.05044 0.04960 0.05120 0.04960 0.05040 0.05120 0.05120 0.04795 0.05284 0.05057 0.05250 KX586174 0.05642 0.05315 0.05070 0.05155 0.05070 0.05233 0.05070 0.05315 0.05233 0.05233 0.05186 0.05675 0.05478 0.05560 KT807829 0.05120 0.05360 0.04960 0.05044 0.04960 0.05120 0.04960 0.05200 0.05280 0.05120 0.04892 0.05088 0.05057 0.05089 KJ606645 0.05887 0.05315 0.06132 0.06056 0.06132 0.06296 0.06132 0.06296 0.06378 0.06541 0.06556 0.06849 0.06460 0.06541 KJ606650 0.05887 0.05478 0.06623 0.06547 0.06623 0.06787 0.06623 0.06787 0.06868 0.06868 0.07045 0.07143 0.06787 0.06868
Univers
ity of
Mala
ya

17
6
Appendix D, continued
S1 KU726713 0712160 1304049 C4 1404200 1211004 1404077 1404128 AB038603 JX000238 JX000244 J19 1209005 FN908151 0.06520 0.06112 0.06601 0.06525 0.06601 0.06764 0.06601 0.06683 0.06764 0.06438 0.06067 0.06556 0.06688 0.06520 FN908161 0.07680 0.07280 0.07440 0.07366 0.07440 0.07600 0.07440 0.07600 0.07680 0.07840 0.08023 0.08611 0.07667 0.07680 FN908147 0.07699 0.07050 0.07536 0.07461 0.07536 0.07861 0.07699 0.08185 0.08104 0.07861 0.08023 0.07828 0.07504 0.07699 FN908155 0.06400 0.05680 0.05920 0.06005 0.05920 0.06080 0.05920 0.06000 0.06080 0.06080 0.05577 0.05871 0.05954 0.06400 AB061374 0.06160 0.05920 0.05200 0.05284 0.05200 0.05360 0.05200 0.05280 0.05360 0.05040 0.04990 0.05088 0.05220 0.06160 FN908160 0.06000 0.05600 0.05760 0.05845 0.05600 0.05920 0.05760 0.05840 0.05760 0.05760 0.05773 0.06262 0.05954 0.06000 FN908162 0.06710 0.07201 0.06301 0.06383 0.06301 0.06465 0.06137 0.06710 0.06792 0.06465 0.06968 0.06771 0.06056 0.06710
Univers
ity of
Mala
ya

17
7
Appendix D, continued
J15 1206070 S7 AY772020 JQ976882 JQ976884 JQ976886 JQ976885 AM422899 1403091 1405001 GQ995519 14004143 J15 N 1206070 0.00080 S7 0.00402 0.00482 AY772020 0.02160 0.02245 0.02251 JQ976882 0.02160 0.02245 0.02251 0.00480 JQ976884 0.02136 0.02233 0.02233 0.00583 0.00194 JQ976886 0.03200 0.03288 0.03296 0.03120 0.02960 0.02816 JQ976885 0.03120 0.03208 0.03215 0.03040 0.02880 0.02621 0.00400 AM422899 0.03840 0.03929 0.03859 0.03760 0.03680 0.03592 0.02960 0.03040 1403091 0.00560 0.00561 0.00080 0.02400 0.02400 0.02330 0.03440 0.03360 0.04080 1405001 0.02235 0.02324 0.02246 0.00083 0.00579 0.00607 0.03146 0.03063 0.03725 0.02483 AM422899 0.03840 0.03929 0.03859 0.03760 0.03680 0.03592 0.02960 0.03040 0.00000 0.04080 0.03725 1404143 0.05520 0.05613 0.05547 0.05920 0.05680 0.05243 0.04720 0.04800 0.04480 0.05760 0.05877 0.04480 1404246 0.05560 0.05641 0.05641 0.05963 0.05721 0.05340 0.04754 0.04835 0.04593 0.05721 0.06005 0.04593 0.00081 1404247 0.05524 0.05618 0.05551 0.05925 0.05685 0.05345 0.04724 0.04804 0.04484 0.05765 0.05877 0.04484 0.00160 JQ976888 0.05419 0.05501 0.05501 0.05829 0.05583 0.05350 0.04680 0.04762 0.04516 0.05583 0.05772 0.04516 0.00657 1208001 0.06080 0.06175 0.06109 0.06480 0.06240 0.05922 0.05040 0.05120 0.05280 0.06320 0.06457 0.05280 0.02240 KT807828 0.05364 0.05457 0.05310 0.05765 0.05685 0.05539 0.04884 0.04964 0.04804 0.05524 0.05712 0.04804 0.04964 KT807827 0.04960 0.05052 0.04904 0.05360 0.05120 0.04951 0.04160 0.04240 0.04080 0.05120 0.05215 0.04080 0.04720 1406043 0.05939 0.06034 0.05887 0.06501 0.06260 0.06140 0.06019 0.06260 0.05859 0.06100 0.06540 0.05859 0.06661 S10 0.05859 0.05953 0.05806 0.06421 0.06180 0.06043 0.05939 0.06180 0.05778 0.06019 0.06457 0.05778 0.06742 1403055 0.05778 0.05873 0.05726 0.06501 0.06260 0.06140 0.05859 0.06100 0.05698 0.05939 0.06540 0.05698 0.06661 0712471 0.06038 0.06121 0.06038 0.06534 0.06286 0.06410 0.06038 0.06286 0.06038 0.06121 0.06593 0.06038 0.06948 1209396 0.06000 0.06095 0.05949 0.06560 0.06320 0.06214 0.06080 0.06320 0.05920 0.06160 0.06623 0.05920 0.06880 1404132 0.05939 0.06034 0.05887 0.06501 0.06260 0.06140 0.06019 0.06260 0.05859 0.06100 0.06540 0.05859 0.06982 J1 0.06080 0.06175 0.06029 0.06640 0.06240 0.06117 0.06160 0.06400 0.05840 0.06240 0.06705 0.05840 0.06960 1403056 0.05840 0.05934 0.05788 0.06560 0.06320 0.06214 0.05920 0.06160 0.05760 0.06000 0.06623 0.05760 0.06720 AB038604 0.06080 0.06175 0.06109 0.06800 0.06560 0.06311 0.06000 0.06240 0.05680 0.06320 0.06871 0.05680 0.06640
Univers
ity of
Mala
ya

17
8
Appendix D, continued
J15 1206070 S7 AY772020 JQ976882 JQ976884 JQ976886 JQ976885 AM422899 1403091 1405001 GQ995519 14004143 1209183 0.06341 0.06423 0.06341 0.07073 0.06992 0.06990 0.06260 0.06504 0.06585 0.06423 0.07239 0.06585 0.06260 1209190 0.06913 0.06994 0.06511 0.06597 0.06511 0.06672 0.06511 0.06752 0.06833 0.06752 0.06556 0.06556 0.06444 1206066 0.06800 0.06880 0.06400 0.06485 0.06400 0.06560 0.06400 0.06640 0.06720 0.06560 0.06360 0.06360 0.06281 AB061397 0.07360 0.07360 0.06480 0.06565 0.06480 0.06640 0.06480 0.06720 0.06800 0.06640 0.06458 0.06458 0.06362 1404050 0.06452 0.06532 0.06290 0.06371 0.06290 0.06290 0.06290 0.06532 0.06613 0.06290 0.06225 0.06225 0.06003 FN908149 0.07040 0.06480 0.06160 0.06245 0.06160 0.06320 0.06160 0.06400 0.06480 0.06240 0.05675 0.05773 0.06117 JQ976891 0.07760 0.07200 0.07920 0.08006 0.07920 0.08080 0.07920 0.08160 0.08240 0.07920 0.07926 0.08219 0.07993 FN908154 0.06000 0.05520 0.05840 0.05925 0.05680 0.06000 0.05840 0.06080 0.06160 0.06000 0.05871 0.06067 0.06036 AY772019 0.04800 0.04560 0.04720 0.04804 0.04560 0.04880 0.04720 0.04800 0.04880 0.04880 0.04697 0.05186 0.04812 KU382051 0.05200 0.04960 0.04960 0.05044 0.04960 0.05120 0.04960 0.05040 0.05120 0.05120 0.04795 0.05284 0.05057 KX586174 0.05642 0.05315 0.05070 0.05155 0.05070 0.05233 0.05070 0.05315 0.05233 0.05233 0.05186 0.05675 0.05478 KT807829 0.05120 0.05360 0.04960 0.05044 0.04960 0.05120 0.04960 0.05200 0.05280 0.05120 0.04892 0.05088 0.05057 KJ606645 0.05887 0.05315 0.06132 0.06056 0.06132 0.06296 0.06132 0.06296 0.06378 0.06541 0.06556 0.06849 0.06460 KJ606650 0.05887 0.05478 0.06623 0.06547 0.06623 0.06787 0.06623 0.06787 0.06868 0.06868 0.07045 0.07143 0.06787 FN908151 0.06520 0.06112 0.06601 0.06525 0.06601 0.06764 0.06601 0.06683 0.06764 0.06438 0.06067 0.06556 0.06688 FN908161 0.07680 0.07280 0.07440 0.07366 0.07440 0.07600 0.07440 0.07600 0.07680 0.07840 0.08023 0.08611 0.07667 FN908147 0.07699 0.07050 0.07536 0.07461 0.07536 0.07861 0.07699 0.08185 0.08104 0.07861 0.08023 0.07828 0.07504 FN908155 0.06400 0.05680 0.05920 0.06005 0.05920 0.06080 0.05920 0.06000 0.06080 0.06080 0.05577 0.05871 0.05954 AB061374 0.06160 0.05920 0.05200 0.05284 0.05200 0.05360 0.05200 0.05280 0.05360 0.05040 0.04990 0.05088 0.05220 FN908160 0.06000 0.05600 0.05760 0.05845 0.05600 0.05920 0.05760 0.05840 0.05760 0.05760 0.05773 0.06262 0.05954 FN908162 0.06710 0.07201 0.06301 0.06383 0.06301 0.06465 0.06137 0.06710 0.06792 0.06465 0.06968 0.06771 0.06056
Univers
ity of
Mala
ya

17
9
Appendix D, continued
1404246 1404247 JQ976888 1208001 KT807828 KT807827 1406043 S10 1403005 0712471 1209396 1404132 J1 1403056 1404246 N 1404247 0.00081 JQ976888 0.00739 0.00657 1208001 0.02337 0.02242 0.02627 KT807828 0.05081 0.04964 0.05090 0.05604 KT807827 0.04835 0.04724 0.04762 0.05200 0.02802 1406043 0.06791 0.06661 0.06661 0.06982 0.06501 0.06180 S10 0.06871 0.06742 0.06743 0.07063 0.06421 0.06260 0.00080 1403055 0.06791 0.06661 0.06661 0.06982 0.06340 0.06180 0.00161 0.00080 0712471 0.07031 0.06948 0.07060 0.07279 0.06617 0.06286 0.00496 0.00414 0.00496 1209396 0.07010 0.06886 0.06897 0.07200 0.06405 0.06240 0.00321 0.00241 0.00321 0.00165 1404132 0.07114 0.06982 0.06990 0.07303 0.06501 0.06340 0.00401 0.00321 0.00401 0.00248 0.00080 J1 0.07091 0.06966 0.06979 0.07280 0.06485 0.06320 0.00321 0.00241 0.00321 0.00331 0.00160 0.00241 1403056 0.06849 0.06725 0.06732 0.07040 0.06405 0.06240 0.00241 0.00161 0.00080 0.00579 0.00400 0.00482 0.00400 AB038604 0.06769 0.06645 0.06732 0.06960 0.06325 0.06320 0.01445 0.01364 0.01445 0.01572 0.01440 0.01525 0.01440 0.01520 1209183 0.06341 0.06265 0.06240 0.06423 0.06672 0.06423 0.07096 0.07015 0.06933 0.07279 0.07154 0.07259 0.07236 0.06992 1209190 0.06366 0.06356 0.06322 0.06592 0.06919 0.06592 0.07258 0.07177 0.07097 0.07361 0.07315 0.07419 0.07395 0.07154 1206066 0.06205 0.06165 0.06158 0.06320 0.06725 0.06560 0.07063 0.06982 0.06902 0.07196 0.07120 0.07223 0.07200 0.06960 AB061397 0.06285 0.06325 0.06240 0.06400 0.07126 0.06960 0.07624 0.07544 0.07464 0.07610 0.07680 0.07785 0.07760 0.07520 1404050 0.05930 0.05887 0.05868 0.06129 0.06935 0.06290 0.06613 0.06532 0.06452 0.06782 0.06694 0.06774 0.06774 0.06532 FN908149 0.06608 0.06485 0.06486 0.07120 0.07046 0.06320 0.06581 0.06661 0.06581 0.06948 0.06800 0.06902 0.06880 0.06640 JQ976891 0.07977 0.07926 0.07964 0.08400 0.07926 0.07840 0.07705 0.07624 0.07544 0.08023 0.07760 0.07865 0.07840 0.07600 FN908154 0.06205 0.06085 0.06076 0.06640 0.05845 0.05920 0.06501 0.06421 0.06340 0.06534 0.06400 0.06501 0.06480 0.06400 AY772019 0.04835 0.04724 0.05008 0.05200 0.05204 0.04640 0.05377 0.05297 0.05217 0.05624 0.05440 0.05538 0.05520 0.05280 KU382051 0.05077 0.04964 0.05255 0.05440 0.05444 0.04720 0.05778 0.05698 0.05618 0.06038 0.05840 0.05939 0.05920 0.05680 KX586174 0.05151 0.05074 0.05090 0.05805 0.05728 0.05070 0.06727 0.06809 0.06727 0.06959 0.06787 0.06727 0.06868 0.06787 KT807829 0.05963 0.05845 0.05993 0.06240 0.05444 0.05040 0.06180 0.06100 0.06019 0.06369 0.06240 0.06180 0.06320 0.06080 KJ606645 0.06541 0.06465 0.06486 0.06705 0.06628 0.06214 0.07055 0.06973 0.06891 0.07291 0.07114 0.07219 0.07032 0.06950
Univers
ity of
Mala
ya

18
0
Appendix D, continued
1404246 1404247 JQ976888 1208001 KT807828 KT807827 1406043 S10 1403005 0712471 1209396 1404132 J1 1403056 KJ606650 0.06868 0.06792 0.06814 0.07195 0.06628 0.06214 0.07383 0.07465 0.07383 0.07788 0.07604 0.07547 0.07522 0.07441 FN908151 0.07498 0.07579 0.07504 0.07553 0.07661 0.06852 0.06927 0.07686 0.07604 0.07498 0.07579 0.07504 0.07553 0.07661 FN908161 0.08480 0.08622 0.08487 0.08539 0.08400 0.08086 0.07840 0.08347 0.08427 0.08480 0.08622 0.08487 0.08539 0.08400 FN908147 0.08185 0.08266 0.08191 0.08210 0.08509 0.08678 0.07942 0.08780 0.08699 0.08185 0.08266 0.08191 0.08210 0.08509 FN908155 0.06160 0.06285 0.06165 0.06322 0.06720 0.06245 0.05840 0.07865 0.07785 0.06160 0.06285 0.06165 0.06322 0.06720 AB061374 0.06000 0.06124 0.06085 0.06240 0.06560 0.06405 0.05760 0.07303 0.07223 0.06000 0.06124 0.06085 0.06240 0.06560 FN908160 0.06800 0.06930 0.06805 0.06979 0.06720 0.06085 0.05840 0.06982 0.06902 0.06800 0.06930 0.06805 0.06979 0.06720 FN908162 0.07938 0.08020 0.07938 0.07970 0.08183 0.07119 0.06792 0.07535 0.07453 0.07938 0.08020 0.07938 0.07970 0.08183
Appendix D, continued
KT807829 KJ606645 KJ606650 FN908151 FN908161 FN908147 FN908155 AB061374 FN908160 FN908162 KT807829 N KJ606645 0.05805 KJ606650 0.06214 0.01472 FN908151 0.07009 0.06132 0.06378 FN908161 0.08000 0.06868 0.07032 0.07009 FN908147 0.07536 0.07195 0.07441 0.08068 0.08509 FN908155 0.05920 0.05070 0.05233 0.06275 0.07120 0.07699 AB061374 0.05280 0.05478 0.05478 0.06194 0.07520 0.07455 0.02400 FN908160 0.05520 0.04988 0.05642 0.06194 0.06480 0.07212 0.04400 0.04480 FN908162 0.06792 0.05984 0.06475 0.07038 0.07692 0.07856 0.05728 0.05646 0.04173 N
Univers
ity of
Mala
ya

18
1
Appendix D, continued
AB038604 1209183 1209190 1206066 AB061397 1404050 FN908149 JQ976891 FN908154 AY772019 KU382051 KX586174 AB038604 N 1209183 0.06423 1209190 0.06511 0.00244 1206066 0.06320 0.00244 0.00482 AB061397 0.06880 0.00650 0.01206 0.00720 1404050 0.05887 0.01230 0.01459 0.01290 0.02016 FN908149 0.06400 0.04309 0.04502 0.04320 0.04880 0.04032 JQ976891 0.07520 0.06179 0.06270 0.05920 0.06640 0.05887 0.06240 FN908154 0.06320 0.06016 0.06350 0.05840 0.06400 0.05806 0.06560 0.06880 AY772019 0.05360 0.06423 0.06511 0.06560 0.06960 0.06290 0.06160 0.07760 0.05360 KU382051 0.05760 0.06829 0.06913 0.06960 0.07360 0.06694 0.06560 0.08160 0.05920 0.00560 KX586174 0.06623 0.06787 0.06868 0.06868 0.07114 0.06430 0.06705 0.07686 0.06296 0.04334 0.04742 KT807829 0.06160 0.06585 0.06672 0.06480 0.06880 0.06048 0.06160 0.07120 0.05520 0.04240 0.04480 0.04988 KJ606645 0.07114 0.07114 0.06950 0.07114 0.07359 0.06843 0.07359 0.08095 0.06214 0.05805 0.05887 0.06378 KJ606650 0.07604 0.07686 0.07686 0.07686 0.07931 0.07502 0.07441 0.08422 0.06950 0.06460 0.06378 0.06705 FN908151 0.07905 0.07905 0.08150 0.07661 0.07905 0.07560 0.07905 0.09046 0.07498 0.06275 0.06194 0.07441 FN908161 0.08160 0.08862 0.09084 0.08800 0.09040 0.08468 0.08480 0.09440 0.08240 0.07120 0.07200 0.08258 FN908147 0.08914 0.08130 0.08347 0.08023 0.08185 0.07598 0.08995 0.09643 0.08266 0.07455 0.07699 0.07686 FN908155 0.07520 0.07398 0.07556 0.07120 0.07520 0.06855 0.06560 0.08480 0.06640 0.06000 0.06080 0.06051 AB061374 0.07120 0.07154 0.07315 0.06880 0.07280 0.06613 0.06480 0.08480 0.06320 0.05760 0.05840 0.06051 FN908160 0.07120 0.07154 0.07315 0.07040 0.07440 0.06774 0.07120 0.08480 0.06640 0.05840 0.06240 0.06132 FN908162 0.07774 0.07365 0.07529 0.07283 0.07529 0.07078 0.07938 0.09165 0.07692 0.07038 0.07283 0.07459
Univ
ersity
of M
alaya

18
2
Appendix E: Uncorrected pairwise distance matrix of the COI-5P sequences.
1209183 1209209 1404246 1404247 1404143 1208002 KT807809 KT807814 KT807811 J15 1209204 1209202 1209005 1206070 1209183 N 1209209 0.00334 1404246 0.10909 0.10383 1404247 0.10909 0.10224 0.00316 1404143 0.10744 0.10224 0.00316 0.00316 1208002 0.11074 0.10543 0.04272 0.04272 0.04114 KT807809 0.12310 0.12266 0.08432 0.08432 0.08263 0.09106 KT807814 0.12648 0.12606 0.08769 0.08769 0.08600 0.09444 0.00337 KT807811 0.13752 0.13356 0.10866 0.10866 0.10696 0.09508 0.11036 0.11375 J15 0.10465 0.10433 0.08307 0.08147 0.08147 0.08466 0.09322 0.09661 0.09898 1209204 0.10579 0.10543 0.08386 0.08386 0.08228 0.08228 0.09106 0.09444 0.09847 0.00160 1209202 0.10744 0.10703 0.08544 0.08544 0.08386 0.08386 0.09275 0.09612 0.10017 0.00319 0.00158 1209005 0.10744 0.10703 0.08228 0.08228 0.08070 0.08386 0.08938 0.09275 0.10017 0.00319 0.00158 0.00316 1206070 0.10579 0.10543 0.08703 0.08703 0.08544 0.08228 0.08769 0.09106 0.09168 0.00799 0.00633 0.00791 0.00791 S7 0.10909 0.10863 0.09494 0.09494 0.09335 0.08386 0.08938 0.09275 0.09677 0.01597 0.01424 0.01582 0.01582 0.01741 C4 0.11371 0.11396 0.08668 0.08507 0.08507 0.07384 0.08703 0.09044 0.09949 0.04984 0.04815 0.04976 0.04976 0.05136 1506071 0.11333 0.11236 0.08494 0.08333 0.08333 0.07212 0.08844 0.09184 0.09915 0.04808 0.04647 0.04808 0.04808 0.04968 1211004 0.11240 0.11182 0.07911 0.07911 0.07753 0.07278 0.07926 0.08263 0.10187 0.04952 0.04747 0.04905 0.04589 0.05063 0712476 0.11405 0.11342 0.08386 0.08386 0.08228 0.07753 0.08769 0.09106 0.10526 0.05591 0.05380 0.05538 0.05222 0.05696 1404200 0.11901 0.11821 0.08861 0.08861 0.08703 0.07911 0.09106 0.09106 0.10526 0.05591 0.05380 0.05538 0.05222 0.05696 1404049 0.11500 0.11396 0.08654 0.08494 0.08494 0.07692 0.08844 0.09184 0.10256 0.05288 0.05128 0.05288 0.05288 0.05449 1405001 0.12397 0.12460 0.09335 0.09335 0.09177 0.08544 0.09275 0.09612 0.09677 0.04952 0.04747 0.04589 0.04905 0.05063 1404081 0.10744 0.10703 0.07911 0.07911 0.07753 0.07595 0.08432 0.08769 0.09168 0.04633 0.04430 0.04589 0.04589 0.04747 KT807819 0.10793 0.10733 0.09275 0.09275 0.09106 0.09949 0.10118 0.10455 0.10357 0.07966 0.08094 0.08263 0.07926 0.08094 KT807817 0.10793 0.10733 0.09275 0.09275 0.09106 0.09949 0.10118 0.10455 0.10696 0.07966 0.08094 0.08263 0.07926 0.08432 KX586166 0.10793 0.10733 0.08600 0.08600 0.08432 0.08600 0.08600 0.08938 0.10526 0.07119 0.06914 0.07083 0.06745 0.06914 KX586151 0.10624 0.10562 0.08432 0.08432 0.08263 0.08432 0.08432 0.08769 0.10357 0.06949 0.06745 0.06914 0.06577 0.06745 GQ862073 0.08430 0.08466 0.07595 0.07595 0.07437 0.08386 0.08769 0.09106 0.09677 0.05272 0.05380 0.05538 0.05222 0.05380
Univers
ity of
Mala
ya

18
3
Appendix E, continued
1209183 1209209 1404246 1404247 1404143 1208002 KT807809 KT807814 KT807811 J15 1209204 1209202 1209005 1206070 GQ862071 0.12066 0.11821 0.10443 0.10443 0.10285 0.10285 0.10961 0.11298 0.11205 0.09105 0.08861 0.08703 0.08703 0.08861 S13 0.09587 0.09265 0.08070 0.08070 0.07911 0.08070 0.08938 0.09275 0.09508 0.07508 0.07278 0.07437 0.07437 0.06962 1403055 0.10744 0.10703 0.10601 0.10601 0.10443 0.09968 0.11804 0.12142 0.10866 0.08307 0.08070 0.08228 0.08228 0.07753 1209203 0.10909 0.10863 0.10443 0.10443 0.10285 0.09810 0.11636 0.11973 0.10696 0.08466 0.08228 0.08386 0.08386 0.07911 1406043 0.11405 0.11342 0.10918 0.10918 0.10759 0.09968 0.11804 0.12142 0.10526 0.08946 0.08703 0.08861 0.08861 0.08386 S10 0.11240 0.11182 0.10759 0.10759 0.10601 0.10127 0.11973 0.12310 0.10696 0.08786 0.08544 0.08703 0.08703 0.08228 1209001 0.11405 0.11342 0.10285 0.10285 0.10127 0.09968 0.12142 0.12479 0.10526 0.08626 0.08386 0.08544 0.08544 0.08386 S5 0.10909 0.10863 0.10443 0.10443 0.10285 0.09810 0.11636 0.11973 0.10357 0.08466 0.08228 0.08386 0.08386 0.07911 1404182 0.10744 0.10703 0.10285 0.10285 0.10127 0.09968 0.11298 0.11636 0.10696 0.08626 0.08386 0.08544 0.08228 0.08070 1404132 0.10909 0.10863 0.10127 0.10127 0.09968 0.09810 0.11467 0.11804 0.10696 0.08626 0.08386 0.08544 0.08228 0.08070 1209396 0.10909 0.10863 0.10127 0.10127 0.09968 0.09810 0.11130 0.11467 0.10526 0.08466 0.08228 0.08386 0.08070 0.07911 KM254858 0.10744 0.10703 0.09652 0.09652 0.09652 0.09810 0.10287 0.10624 0.11885 0.09904 0.09652 0.09810 0.09494 0.09652 JX475023 0.12414 0.12191 0.10379 0.10214 0.10379 0.12521 0.12676 0.13028 0.13404 0.10430 0.10544 0.10379 0.10379 0.10379 KM254920 0.13884 0.13578 0.11076 0.11076 0.11076 0.10759 0.13153 0.13491 0.12903 0.10863 0.10918 0.11076 0.10759 0.10918
Univers
ity of
Mala
ya

18
4
Appendix E, continued
S7 C4 1506071 1211004 0712476 1404200 1404049 1405001 1404081 KT807819 KT807817 KX586166 KX586151 GQ862073 1209183 0.10909 0.10863 0.09494 0.09494 0.09335 0.08386 0.08938 0.09275 0.09677 0.01597 0.01424 0.01582 0.01582 0.01741 1209209 0.11371 0.11396 0.08668 0.08507 0.08507 0.07384 0.08703 0.09044 0.09949 0.04984 0.04815 0.04976 0.04976 0.05136 1404246 0.11333 0.11236 0.08494 0.08333 0.08333 0.07212 0.08844 0.09184 0.09915 0.04808 0.04647 0.04808 0.04808 0.04968 1404247 0.11240 0.11182 0.07911 0.07911 0.07753 0.07278 0.07926 0.08263 0.10187 0.04952 0.04747 0.04905 0.04589 0.05063 1404143 0.11405 0.11342 0.08386 0.08386 0.08228 0.07753 0.08769 0.09106 0.10526 0.05591 0.05380 0.05538 0.05222 0.05696 1208002 0.11901 0.11821 0.08861 0.08861 0.08703 0.07911 0.09106 0.09106 0.10526 0.05591 0.05380 0.05538 0.05222 0.05696 KT807809 0.11500 0.11396 0.08654 0.08494 0.08494 0.07692 0.08844 0.09184 0.10256 0.05288 0.05128 0.05288 0.05288 0.05449 KT807814 0.12397 0.12460 0.09335 0.09335 0.09177 0.08544 0.09275 0.09612 0.09677 0.04952 0.04747 0.04589 0.04905 0.05063 KT807811 0.10744 0.10703 0.07911 0.07911 0.07753 0.07595 0.08432 0.08769 0.09168 0.04633 0.04430 0.04589 0.04589 0.04747 J15 0.10793 0.10733 0.09275 0.09275 0.09106 0.09949 0.10118 0.10455 0.10357 0.07966 0.08094 0.08263 0.07926 0.08094 1209204 0.10793 0.10733 0.09275 0.09275 0.09106 0.09949 0.10118 0.10455 0.10696 0.07966 0.08094 0.08263 0.07926 0.08432 1209202 0.10793 0.10733 0.08600 0.08600 0.08432 0.08600 0.08600 0.08938 0.10526 0.07119 0.06914 0.07083 0.06745 0.06914 1209005 0.10624 0.10562 0.08432 0.08432 0.08263 0.08432 0.08432 0.08769 0.10357 0.06949 0.06745 0.06914 0.06577 0.06745 1206070 0.08430 0.08466 0.07595 0.07595 0.07437 0.08386 0.08769 0.09106 0.09677 0.05272 0.05380 0.05538 0.05222 0.05380 S7 0.10909 0.10863 0.09494 0.09494 0.09335 0.08386 0.08938 0.09275 0.09677 0.01597 0.01424 0.01582 0.01582 0.01741 C4 0.11371 0.11396 0.08668 0.08507 0.08507 0.07384 0.08703 0.09044 0.09949 0.04984 0.04815 0.04976 0.04976 0.05136 1506071 0.11333 0.11236 0.08494 0.08333 0.08333 0.07212 0.08844 0.09184 0.09915 0.04808 0.04647 0.04808 0.04808 0.04968 1211004 0.11240 0.11182 0.07911 0.07911 0.07753 0.07278 0.07926 0.08263 0.10187 0.04952 0.04747 0.04905 0.04589 0.05063 0712476 0.11405 0.11342 0.08386 0.08386 0.08228 0.07753 0.08769 0.09106 0.10526 0.05591 0.05380 0.05538 0.05222 0.05696 1404200 0.11901 0.11821 0.08861 0.08861 0.08703 0.07911 0.09106 0.09106 0.10526 0.05591 0.05380 0.05538 0.05222 0.05696 1404049 0.11500 0.11396 0.08654 0.08494 0.08494 0.07692 0.08844 0.09184 0.10256 0.05288 0.05128 0.05288 0.05288 0.05449 1405001 0.12397 0.12460 0.09335 0.09335 0.09177 0.08544 0.09275 0.09612 0.09677 0.04952 0.04747 0.04589 0.04905 0.05063 1404081 0.10744 0.10703 0.07911 0.07911 0.07753 0.07595 0.08432 0.08769 0.09168 0.04633 0.04430 0.04589 0.04589 0.04747 KT807819 0.10793 0.10733 0.09275 0.09275 0.09106 0.09949 0.10118 0.10455 0.10357 0.07966 0.08094 0.08263 0.07926 0.08094 KT807817 0.10793 0.10733 0.09275 0.09275 0.09106 0.09949 0.10118 0.10455 0.10696 0.07966 0.08094 0.08263 0.07926 0.08432 KX586166 0.10793 0.10733 0.08600 0.08600 0.08432 0.08600 0.08600 0.08938 0.10526 0.07119 0.06914 0.07083 0.06745 0.06914 KX586151 0.10624 0.10562 0.08432 0.08432 0.08263 0.08432 0.08432 0.08769 0.10357 0.06949 0.06745 0.06914 0.06577 0.06745 GQ862073 0.08430 0.08466 0.07595 0.07595 0.07437 0.08386 0.08769 0.09106 0.09677 0.05272 0.05380 0.05538 0.05222 0.05380
Univers
ity of
Mala
ya

18
5
Appendix E, continued
S7 C4 1506071 1211004 0712476 1404200 1404049 1405001 1404081 KT807819 KT807817 KX586166 KX586151 GQ862073 GQ862071 0.08703 0.08668 0.08814 0.08228 0.09177 0.09177 0.09295 0.08861 0.07437 0.08769 0.08769 0.09949 0.09781 0.07278
S13 0.07437 0.08026 0.07853 0.07753 0.08070 0.08228 0.07853 0.06962 0.06487 0.07757 0.08094 0.07926 0.07757 0.06329
1403055 0.08386 0.08668 0.08654 0.08544 0.08861 0.09177 0.08974 0.08544 0.07911 0.08769 0.09106 0.08600 0.08432 0.08070
1209203 0.08544 0.08507 0.08494 0.08386 0.08703 0.09019 0.08814 0.08386 0.07753 0.08600 0.08938 0.08432 0.08263 0.07911
1406043 0.09019 0.08668 0.08654 0.08861 0.09177 0.09494 0.09295 0.08861 0.08228 0.09106 0.09444 0.08938 0.08769 0.08386
S10 0.08861 0.08507 0.08494 0.08703 0.09019 0.09335 0.09135 0.08703 0.08070 0.08938 0.09275 0.08769 0.08600 0.08228
1209001 0.09019 0.08347 0.08333 0.08544 0.08861 0.09177 0.08974 0.08861 0.07911 0.09106 0.09444 0.08938 0.08769 0.08386
S5 0.08544 0.08186 0.08173 0.08386 0.08703 0.09019 0.08814 0.08386 0.07753 0.08600 0.08938 0.08432 0.08263 0.07911
1404182 0.08703 0.08347 0.08333 0.08228 0.08861 0.09177 0.09295 0.08544 0.08228 0.08263 0.08600 0.08432 0.08263 0.07753
1404132 0.08703 0.08668 0.08654 0.08544 0.08544 0.09177 0.09295 0.08544 0.08228 0.08938 0.09275 0.08432 0.08263 0.07753
1209396 0.08544 0.08186 0.08173 0.08070 0.08386 0.08703 0.08814 0.08386 0.08070 0.08432 0.08769 0.08263 0.08094 0.07595
KM254858 0.09968 0.10754 0.10897 0.10127 0.10127 0.10285 0.09936 0.10601 0.09019 0.09275 0.09275 0.09949 0.09781 0.08544
JX475023 0.11532 0.11901 0.11755 0.11862 0.12356 0.12356 0.12417 0.12356 0.11532 0.11972 0.12148 0.11268 0.11092 0.10049
KM254920 0.11076 0.11557 0.11699 0.11392 0.11551 0.11551 0.11699 0.11392 0.11392 0.12310 0.12142 0.11130 0.10961 0.10601
Univers
ity of
Mala
ya

18
6
Appendix E, continued
GQ862071 S13 1403055 1209203 1406043 S10 1209001 S5 1404182 1404132 1209396 KM254858 JX475023 KM254920 GQ862071 N S13 0.08386 1403055 0.10759 0.07753 1209203 0.10601 0.07595 0.00158 1406043 0.11076 0.07753 0.00633 0.00475 S10 0.10918 0.07911 0.00475 0.00316 0.00475 1209001 0.10759 0.08070 0.00633 0.00475 0.00633 0.00475 S5 0.10601 0.07595 0.00475 0.00316 0.00475 0.00316 0.00475 1404182 0.10443 0.07437 0.01266 0.01108 0.01266 0.01108 0.01266 0.01108 1404132 0.10759 0.07753 0.01582 0.01424 0.01582 0.01424 0.01582 0.01424 0.00633 1209396 0.10285 0.07595 0.01108 0.00949 0.01108 0.00949 0.01108 0.00949 0.00475 0.00791 KM254858 0.11867 0.08544 0.09810 0.09652 0.09810 0.09968 0.10127 0.09652 0.09494 0.09494 0.09652 JX475023 0.12850 0.11203 0.13509 0.13344 0.13839 0.13674 0.13344 0.13344 0.12850 0.13015 0.13015 0.12521 KM254920 0.12658 0.11709 0.13608 0.13449 0.13608 0.13449 0.13924 0.13449 0.13291 0.13291 0.13133 0.11392 0.09226 N
Univers
ity of
Mala
ya

18
7
Appendix F: Uncorrected pairwise distance matrix of the UPA sequences
1205213 S4 1209203 1209208 1209209 1209183 KX586118 KX586122 1208002 KX586103 1404247 KX586108 GQ471923 1205213 N S4 0.00811 1209203 0.00270 0.01081 1209208 0.04324 0.05135 0.04324 1209209 0.04054 0.04865 0.04054 0.00270 1209183 0.03784 0.04595 0.04054 0.00541 0.00270 KX586118 0.02703 0.03514 0.02973 0.02973 0.02703 0.02432 KX586122 0.02432 0.03243 0.02703 0.02973 0.02703 0.02432 0.00270 1208002 0.03514 0.04324 0.03784 0.03784 0.03514 0.03243 0.01351 0.01622 KX586103 0.03784 0.04595 0.04054 0.04054 0.03784 0.03514 0.01081 0.01351 0.01351 1404247 0.03514 0.04324 0.03784 0.03784 0.03514 0.03243 0.01351 0.01622 0.01081 0.01892 KX586108 0.03514 0.04324 0.03784 0.03784 0.03514 0.03243 0.00811 0.01081 0.01081 0.01351 0.01081 GQ471923 0.03243 0.04054 0.03514 0.03514 0.03243 0.02973 0.01622 0.01892 0.01892 0.02703 0.01892 0.01892 KX586140 0.03784 0.04595 0.04054 0.04054 0.03784 0.03514 0.02162 0.02432 0.01892 0.02703 0.01892 0.01351 0.01081 1209202 0.03784 0.04595 0.04054 0.04054 0.03784 0.03514 0.01351 0.01622 0.02162 0.02432 0.02162 0.01081 0.02162 1012344 0.03243 0.04054 0.03514 0.03784 0.03514 0.03243 0.01081 0.01351 0.01892 0.02162 0.01892 0.01351 0.01892 HQ421565 0.03784 0.04595 0.04054 0.04054 0.03784 0.03514 0.01351 0.01622 0.01622 0.02432 0.02162 0.01622 0.02162 1211004 0.02973 0.03784 0.03243 0.03243 0.02973 0.02703 0.01081 0.01351 0.01351 0.01622 0.01351 0.01351 0.01351 0712476 0.02703 0.03514 0.02973 0.03514 0.03243 0.02973 0.01351 0.01622 0.01622 0.01892 0.01622 0.01622 0.01622 1404077 0.04324 0.05135 0.04595 0.04595 0.04324 0.04054 0.02432 0.02703 0.02703 0.02432 0.02703 0.02703 0.02703 1405001 0.02432 0.03243 0.02703 0.03784 0.03514 0.03243 0.01622 0.01892 0.01892 0.02703 0.01892 0.01892 0.01892 HQ421574 0.02162 0.02973 0.02432 0.02432 0.02162 0.01892 0.00541 0.00811 0.01351 0.01622 0.01351 0.01351 0.01081 S1 0.03784 0.04595 0.04054 0.05135 0.04865 0.04595 0.02703 0.02973 0.03243 0.03243 0.03243 0.03243 0.02973 KJ648575 0.06486 0.07297 0.06757 0.07297 0.07027 0.06757 0.06216 0.06486 0.07027 0.07297 0.07027 0.07027 0.05946 KJ543073 0.06216 0.07027 0.06486 0.07297 0.07027 0.06757 0.06216 0.05946 0.07027 0.06757 0.07027 0.07027 0.06216
Univers
ity of
Mala
ya

18
8
Appendix F, continued
KX586140 1209202 1012344 HQ421565 1211004 0712476 1404077 1405001 HQ421574 S1 KJ648575 KJ543073 KX586140 N 1209202 0.02162 1012344 0.02432 0.00811 HQ421565 0.02703 0.01622 0.01351 1211004 0.01892 0.01351 0.01081 0.01351 0712476 0.02162 0.01622 0.01351 0.01622 0.00270 1404077 0.03243 0.02703 0.02432 0.02703 0.01351 0.01622 1405001 0.02432 0.01892 0.01351 0.01892 0.01081 0.01351 0.02432 HQ421574 0.01622 0.01622 0.01351 0.01622 0.00811 0.01081 0.02162 0.01351 S1 0.03514 0.03784 0.03514 0.03784 0.02973 0.03243 0.03784 0.03514 0.02703 KJ648575 0.06486 0.07027 0.06757 0.07027 0.06216 0.06486 0.07027 0.06486 0.05946 0.06216 KJ543073 0.06757 0.07568 0.07297 0.07568 0.06757 0.07027 0.07027 0.07027 0.06216 0.05676 0.02432
Univers
ity of
Mala
ya

18
9
Appendix G: Uncorrected pairwise distance matrix of the LSU (28S rDNA) sequences
S2 GQ471913 GQ471914 GQ471915 1404246 1209001 1211004 1209204 1208001 1405001 1209005 KJ594951 1209183 S2 N GQ471913 0.00162 GQ471914 0.00000 0.00162 GQ471915 0.00324 0.00485 0.00324 1404246 0.00485 0.00647 0.00485 0.00162 1209001 0.00487 0.00649 0.00487 0.00162 0.00325 1211004 0.00485 0.00647 0.00485 0.00162 0.00324 0.00162 1209204 0.00485 0.00646 0.00485 0.00000 0.00162 0.00162 0.00161 1208001 0.00485 0.00647 0.00485 0.00162 0.00324 0.00324 0.00323 0.00161 1405001 0.00324 0.00485 0.00324 0.00000 0.00162 0.00162 0.00162 0.00000 0.00162 1209005 0.00324 0.00485 0.00324 0.00000 0.00162 0.00162 0.00161 0.00000 0.00161 0.00000 KJ594951 0.00485 0.00647 0.00485 0.00162 0.00324 0.00325 0.00324 0.00162 0.00324 0.00162 0.00162 1209183 0.00808 0.00969 0.00808 0.00485 0.00647 0.00647 0.00645 0.00644 0.00645 0.00485 0.00484 0.00324 HQ422446 0.00334 0.00501 0.00334 0.00000 0.00000 0.00168 0.00167 0.00000 0.00167 0.00000 0.00000 0.00167 0.00501 AM420447 0.03859 0.04027 0.03859 0.04034 0.04034 0.04209 0.04195 0.04188 0.04195 0.04027 0.04027 0.03866 0.04181 AM42041 0.04027 0.04195 0.04027 0.04202 0.04202 0.04384 0.04370 0.04362 0.04370 0.04202 0.04202 0.04034 0.04355
Appendix G, continued
HQ422446 AM420447 AM420418 HQ422446 N AM420447 0.04089 AM420418 0.04259 0.00491 N
Univers
ity of
Mala
ya