margaret t.r-- inside the closed world of the brain.pdf
DESCRIPTION
Inside the Closed World of the BrainTRANSCRIPT




INSIDETHECLOSEDWORLDOFTHEBRAINHOWBRAINCELLSCONNECT,SHAREANDDISENGAGE—ANDWHYTHISHOLDSTHEKEYTOALZHEIMER’SDISEASE
MARGARETT.REECEPHDREECEBIOMEDICALCONSULTINGLLC
MANLIUS,NEWYORK

TextCopyright©2015byMargaretT.Reece
Imageslicensedfromwww.shutterstock.comincludefigures1-2,1-7,2-1,2-2,2-6,3-1,4-4,4-5,4-9,6-2,6-3,6-4,6-5,7-13 and 7-14. Images in the public domain in theUnited States are indicated in the figure legends. The remainingfiguresarelicensedundervariouscreativecommonslicensesatWikiMedia.
All rights reserved.No part of this publicationmay be reproduced, distributed or transmitted in any formor by anymeans, including photocopying, recording, or other electronic or mechanical methods, without the prior writtenpermission of the publisher, except in the case of brief quotations embodied in critical reviews and certain othernoncommercial uses permitted by copyright law. For permission requests, write to the publisher, at “Attention:PermissionsCoordinator,”attheaddressbelow.
MargaretT.ReecePhD/ReeceBiochemicalConsultingLLC
8195CazenoviaRoad
Manlius,NewYork13104
www.medicalsciencenavigator.com
BookLayout©2013BookDesignTemplates.com;CoverImage©Viktoriya,Shutterstock.com
OrderingInformation:
Quantity sales. Special discounts are available on quantity purchases by corporations, associations, and others. Fordetails,contactthe“SpecialSalesDepartment”attheaddressabove.
InsidetheClosedWorldoftheBrain/MargaretT.ReecePhD—1sted.
ISBN978-0-9963513-1-7(ebook)

ForallwhoarededicatedtoeradicationofthelonggoodbyethatisAlzheimer’sdisease.

PrefaceMOSTEVERYONEHASHEARDofAlzheimer’sdisease, but fewknowmuch
about it. Because I teach human physiology, friends, acquaintances, students, familymembersandstrangersfrequentlyaskmequestionsaboutit.WhatisAlzheimer’sdisease?How is Alzheimer’s disease different than just getting old? Can I avoid Alzheimer’sdiseasebykeepingmycholesterol levelundercontrol?Comingupwithaclearaccurateanswertotheseandsimilarquestionsovercoffeeorlunchisachallenge.First,wordsthatdescribehowabrainroutinelyworksrequireexplanation.Second,somemythsaboutthehuman brain must be dispelled. Third, the phases of Alzheimer’s disease prior to theappearance of symptoms need to be described. My goal with this book is to providereaders with state-of-the-art knowledge of how brain cells normally work together andwheretheymaygoastraytoestablishAlzheimer’sdisease.Thereisconsiderablereasontobelieve that ongoing research effortswill produceways toprevent, or sufficiently slow,Alzheimer’s disease so that people in the future can live a normal lifespan withoutexperiencingthisformofdementia.
MargaretT.Reece,PhD

IntroductionTHETEMPTATIONTOREADchapter8,“WhenItAllGoesWrong—Alzheimer’s
Dementia” first is understandable. For readerswith a background in neuroscience, thatapproach should not be a problem. Others will find reference throughout chapter 8 toearlier chapters with needed background material. Chapters 1-7 are organized toprogressively build a basic vocabulary for newcomers to the science.Medical studentswill find numerous facts on every page that are extracted from actual Step 1 examquestions.
Chapter1 presents tactics for quickly learning the necessarywords. The secondchapterprovidesanexplanationofthegeneralorganizationofthehumanbrainbothatthevisualandmicroscopiclevel.Thenextchapterdescribesthebrain’selaboratesystemforqualitycontrolofthefluidssurroundingitscells.Twochaptersaredevotedtoneurons,thesuperstars of the brain cell community. The first discusses where neurons get theirelectricityandthesecondexplainshowneuronscommunicatewitheachother.Inchapter6thebrain’sother,non-neuron,cellsareintroduced,andtheirpartnershipwithneuronsisexplained. Inchapter7, the consensuswithinpsychology andneuroscience is presentedconcerningcriticalelementsofmemoryformationandlanguageacquisition.
GlossaryandFurtherReadingsectionsareincludedattheend.FurtherReadingisapartiallistoftheoriginalpapersconsultedincreatingthisbook.

CONTENTS
Tips&TricksforLearningScientificLanguage
LanguageandSound
ScientificVocabulary
StrategiesandTactics
NamingBrainElements
UsefulTools
SummaryChapter1
HowtheHumanBrainIsOrganized
TheVisibleBrain
BrainSubdivisions
GrayMatterandWhiteMatter
InsidetheBrain
SummaryChapter2
QualityControlofBrain’sExtracellularFluids
FluidSurroundingCells
CerebrospinalFluid
CerebralBloodSupply
SummaryChapter3
Neurons—HowTheyMakeElectricity
NeuronCompartments
Brain’sElectricity
NeuronsatRest
Voltage-sensitiveIonChannels
AxonSignaling
AxonHousekeeping
SummaryChapter4

NeuronSynapses—ExcitatoryandInhibitory
BrainSynapses
PresynapticCompartment
PostsynapticCompartment
ExcitatoryandInhibitoryNeurons
OtherNeurotransmitters
SUMMARYCHAPTER5
IntroductiontotheGliaandMicroglia—MeettheStageCrew
StemCells
AdultGliaandMicroglia
FourPartSynapses
FunctionalPartnerships
MetabolismintheBrain
RepairofBrainDamage
InflammationandInfection
SummaryChapter6
Brain’sInfrastructureforMemoryandLanguage
InformationFlow
MappingtheBrain’sNeurons
LinkingAnatomytoPurpose
HumanMemory
AnatomicStructureofMemory
LearningLanguage
SummaryChapter7
WhenItAllGoesWrong—Alzheimer’sDementia
Alzheimer’sBrain
Alzheimer’sTherapies
Pre-symptomaticAlzheimer’s

Amyloid-βandTauPhysiology
NeuronDamageandLoss
ActivationofGliaandMicroglia
Alzheimer’s-likeBrainwithoutDementia
NewAvenuesforProgress
SummaryChapter8
FurtherReading
Glossary

“Somethingsneedtobebelievedtobeseen.”STEVEJOBS

[1]
Tips&TricksforLearningScientificLanguageTHESTRANGEWORDSUSEDinanatomyandphysiologymakeitdifficult to
followdiscussionsofthescience.Becausescientificlanguageisanobstacleformany,thisbook begins by describing the secret to understanding thewords needed to learn aboutwhathappensinsidethehumanbrain.
Human anatomic names were assigned when scholars wrote and lectured inClassical Latin. Classical Latin was the universal language of large segments of thewesternscientificworldfromthetimeoftheRomanEmpire(Figure1-1)throughthe17thcentury. The good news is Latin can be translated intomodern languages. Psychologyresearchdiscoveredwordsarelearnedfastbythehumanbrainwhentheyareassociatedwith something familiar. Thus, assigning meaning to the Latin names makes them fareasiertoremember.
Figure:1-1:RangeofLatinlanguageusein60ADshowningreen.Illustration:©HannesKarnoefel
LANGUAGEANDSOUNDInfants and young children acquire their primary language through their brain’s
instinctiveinterpretationofauditoryinput.Byjusthearingthesubsetofsoundsusedinthelanguagespokennearthemtheycansortthesoundsintotheirproperorderandmapthemtoimportance.Mostbrainstructuresdedicatedtoprocessingofauditorysignalsaresuperbatdiscerningpitchofthehumanvoiceandassigningimplicationtotonesandinflection.
Infantscandistinguishallofthesoundsofalloftheworld’slanguagesuntilaboutagesixmonths.Betweensixmonthsandayear,brainpathwaysdevotedtolanguagebeginto form in support of the soundsmost often heard. Learning to recognize and speak alanguageisinstinctiveforinfants.

Adultstryingtolearnasecondlanguagefindreadingandwritinganewlanguageisnotenoughtodevelopfluency.Listeningtolanguagespokeninthecorrectmanneroveranextendedperiodof time isneeded.Auditory input is required tobuildnew languagepathwaysinthebraintoparallelthoseofthenativelanguagelearnedininfancy.
Likewise,justreadingscientificterminologydoeslittletoestablishitinmemory.FewpeoplespeakClassicalLatinanymore,soasubstituteauditorystrategyisneededtohelpthemindmapthesoundsofscientificnamestotheirmeaning.
SCIENTIFICVOCABULARYToday much of the world’s population is at least familiar with the English
language.SomeargueEnglishshouldbetheprimarylanguageusedtoteachscience.And,English in its various forms is, for themost part, derived fromLatin. Latin andGreekscientificwordspresentagreaterchallengeforthosewhosenativelanguageisnotderivedfromLatin.
Translation of compound scientific words is not always direct. The simpledescriptivenature isoftenhiddenbecauseof thepatched togetherarrangementofmanyideas.The solution is tobreak the longwords intoparts and toassignmeaning toeachpart.Thenthepartsmustberearrangedintoasensibleorder,andwordorderisnotalwaysthesamefromlanguagetolanguage.Forexample,inLatinadjectivesfollownounsunlikeEnglishwhereadjectivesprecedenouns.
Becausepeoplebecomesouncomfortablewiththesoundofscientificwords,theyalsofailtospeakandwritethemwithprecision.Scientificterminologyisoftencomposedof made-up words, which seem almost like brief descriptive pseudo-sentences. If thecompoundwordsarenotspokenwithprecision,thevariouspartsmaybecomemixedinahaphazard sequence producing nonsense descriptions. To keep the parts of compoundscientificnamesinproperorder,speakingandlisteningmustbeincludedinthelearningprocess.
STRATEGIESANDTACTICSRecent studies at colleges experimented with approaches to help students learn
scientific and medical terminology. Design of the education experiments relied uponconclusions of investigators who study the brain’s process for learning language.Educatorsfoundreadinganewanddifficultwordoutloudthreetofivetimeseachdayforseveral days improved students’ ability to remember theword, to spell it and to betterabsorb printed material using the word. Adding auditory input to reading of scientificwordswasmoreeffectiveincreatingwordmemorythanreadingalone.
Theremainingsectionsof thischapterdiscusssomebasic terminologyneededtodescribehowthebrainworks.Thisvocabularywillbeusedoftenintherestofthebook.ImportantwordswillbepresentedinitalicsandthemeaningoftheoriginalLatinorGreekwordwillbeunderlined.
There areonline tools available for learninghow topronounce anatomicnames.Thetoolsprovideanacceptablepronunciationinmanynativelanguages.Anexampleofthese tools can be found by opening a computer or tablet device to the internet atwww.translate.google.com.

AtGoogle translate, start bypickingEnglish above theboxon the left and type‘neuron’intothebox.Next,tohearthewordneuroninasecondlanguage,pickthesecondlanguageabovetheboxontheright.Neuronwillbetranslatedintotheselectedlanguage.BelowtheboxontheleftwhenEnglishisthechosenlanguagetherewillbeadefinitionofwhatthewordmeans.
Beloweachboxisasmallmicrophoneicon.Clickoneachicontolistentoneuronpronouncedintheselectedlanguage.Thewordneuron,eventhoughspelledthesameinseveral languages, may be pronounced in various ways because the alphabet ispronouncedinavarietyofwaysfromlanguagetolanguage.
Practice pronouncing the word neuron after the computer speaks it in eachlanguage.Repeatthisprocessthreetofivetimesforbothformsoftheword.Therepetitionwillmapthesequenceofthesoundstomemory.KeepGoogletranslateopen,andasnewscientificwordsappearcontinuetopracticelisteningtothemandsayingthemoutloud.
NAMINGBRAINELEMENTSNaming the cells of the brain offers a good place to begin learning how the
anatomic labeling system works. For studying the brain, the scientific names neuron,nervecellandnerveareessential.Nerveisoftenusedasifitmeansthesameasneuronornerve cell. But, that is not correct. Both neuron and nerve cell refer to an individualelectricalcellofthebrainorspinalcord.
Incontrast,anerve isacable-likebundle.Thebundle includes just thepartofaneuroncalledanaxon.ThewordaxoncomesfromtheGreekwordforaxis,astraightline.Many neurons contribute their elongated axons to a nerve.Each axon in a nerve is thelengthyextensionofasingleneuron(Figure1-2andFigure1-3).
Nervesareenclosedbyatoughsheathoftissue.Thewordneuro,fromtheGreeklanguage,meanssineworstring.Nervesinfactlooklikewhitestringwhenseeninlivingtissue.Theindividualcellsofthenervoussystem,neurons,werenotobservedbyscholarsuntil longafternervesweredescribed (Figure1-3).Some,but not all neurons, are longandstringylikenerves.Neuronsassumemanydifferentshapes.
Practice reading and saying neuron, nerve and axon usingwww.translate.google.com.
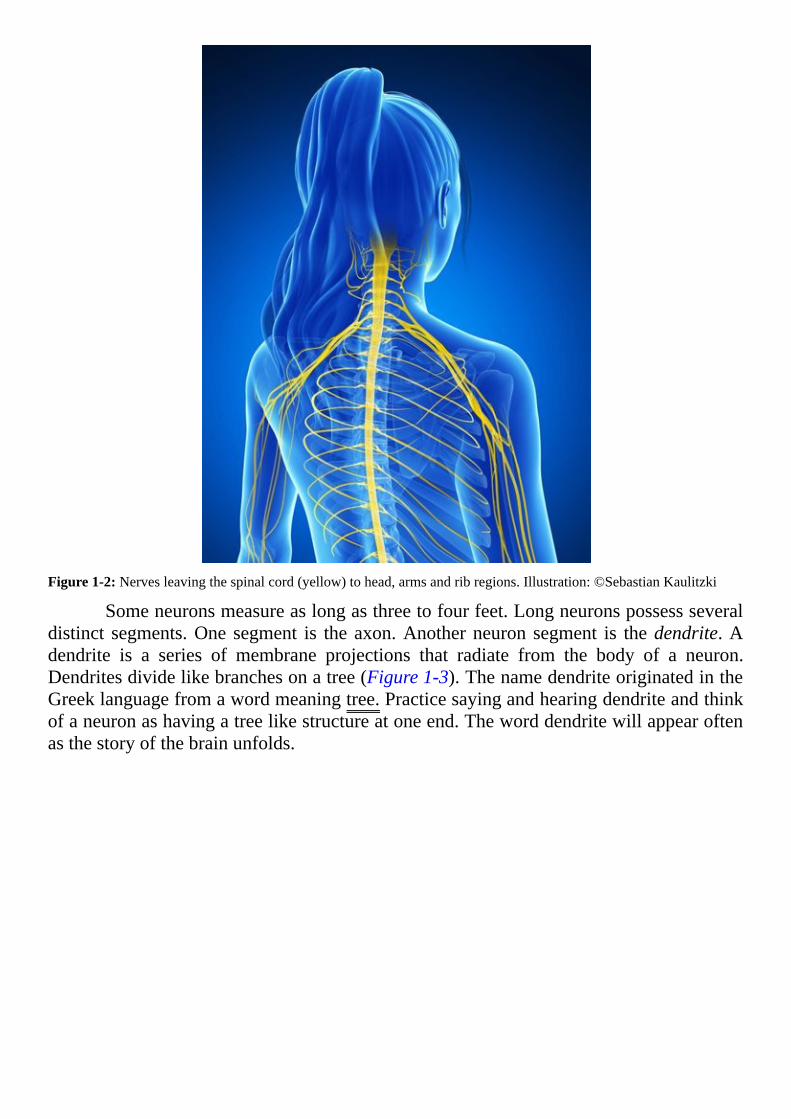
Figure1-2:Nervesleavingthespinalcord(yellow)tohead,armsandribregions.Illustration:©SebastianKaulitzki
Someneuronsmeasureaslongasthreetofourfeet.Longneuronspossessseveraldistinct segments.One segment is the axon.Anotherneuron segment is thedendrite.Adendrite is a series of membrane projections that radiate from the body of a neuron.Dendritesdividelikebranchesonatree(Figure1-3).ThenamedendriteoriginatedintheGreeklanguagefromawordmeaningtree.Practicesayingandhearingdendriteandthinkofaneuronashavingatreelikestructureatoneend.Theworddendritewillappearoftenasthestoryofthebrainunfolds.

Figure 1-3: Brain neuronswith different shapes.Drawing: SantiagoRamón yCajal about 1900, thiswork is in thepublicdomain
Dendritesdisplaysmallmembraneprotrusionscalleddendriticspines(Figure1-4).Heredendrite is changed to thedescriptive form,dendritic.Spine is a derivative of theLatin word spina meaning thorn-like structures on a stem. Each dendrite may displayseveral thousand dendritic spines. Dendritic spines change their shape over time inresponse to their local environment. Their properties draw considerable attention inmodernneurosciencestudies.

Figure1-4:Closeupofadendriteofabrainneurondisplayingdendriticspines.Photomicrograph:©CopperKettle
Each neuron includes a nucleus, an area within its body to house its geneticinformation. As sometimes happens in anatomy, the word nucleus has two differentmeanings in the central nervous system, which is the brain and spinal cord. Whendescribing the location of the genetic information in a neuron, nucleusmeans the samecompartmentfoundinothercellsforhousinggeneticinformation.
But, in the brain and spinal cordnucleus alsomeans a collection of neuron cellbodies.Brainareasmarkedbyagroupofneuroncellbodiesfine-tuneparticularfunctionaloperationslikefingerstypingonakeyboard.NucleusoriginatesfromtheLatinwordforkernelornut,whichisasuitabledescriptionof theappearanceof theclustersofneuroncell bodies in the brain and of the subdivision of all cellswhere genetic information isstored.
Theprocessofneuroplasticityisarathernewconceptinscienceofthebrainthatdates back only to the 1980s. It refers to the brain’s ability to rearrange its neurondendrites and dendritic spines in response to sensory stimulation like sound and light.Neuroplasticity happens while saying, hearing and reading new scientific words. Theregions of the human brain dedicated to learning new ideas are particularly busymodifyingthewayneuronsconnect.
Before1980,scientistsbelievedallneuronconnectionsinthebrainsofmammalsand birds remained permanently in place after puberty. The earliest accounts ofneuroplasticitydescribed seasonal changes inbrainneuronconnections in songbirds. Itwasnotuntilafter2000thatneuroplasticitywasconfirmedinhumanbrain.Contemporarystudiesreporthumanbrainneuronsadapttotheirenvironmentthroughoutlife.
The word neuroplasticity is a combination of two words, neuro and plasticity.Plasticity originates from two similar words one Greek, the other Latin describing theprocesstomold.Therefore,thecompoundwordneuroplasticitymeanstomoldormodify

howneuronsconnect.
Axon terminals exist at the far end of the neuron’s axon (Figure 1-5). Axonterminalspossessspecialcharacteristicsallowingthemtocommunicatewithanothercell.Where the terminal end of a neuron contacts another cell, a structure forms named asynapse.SynapsederivesfromaGreekworddescribingapointofcontact.
Figure1-5:Generalstructureofaneuron.Illustration:©NickGorton
At a synapse an axon terminal releases a chemical substance named aneurotransmitter (Figure 1-6). Again scientists combined two words to create a newdescriptive label. Theword transmitter stems from aLatinwordmeaning tosend. Thecombinationwordreferstoachemicalaneuronreleasesasasignal.
A subdivision of neuroplasticity is synaptic plasticity. Synaptic plasticity isremolding of anatomic structures where axon terminals make contact, synapses. Itincludes changes in the type and amount of neurotransmitter released by the axonterminal.Italsoincorporatesanymodificationofthereceivingcell’sabilitytorespondtoneurotransmitter.

Figure1-6:Asimplified illustrationofanaxon terminalsynapseonadendriticspine.Thebeigespheres in theaxonterminal represent neurotransmitter. A small space exists between the two structures throughwhich neurotransmittertravels.Illustration:©CurtisNeveu
Another recently recognized process for the adult brain is neurogenesis. In thiscase, two words combine to describe one process. The first part of the word, neuro,describes an electrical cell of the brain or spinal cord. The second part of the word,genesis, refers tobeingborn.Combining the twoparts intoneurogenesiscreatesawordinferring the bringing of neurons into existence. The word genesis originates from theGreek word for birth. Genesis is a word used often in physiology. For example,osteogenesisisbirthofnewbone.OsteoisfromtheGreekwordosteonmeaningbone.
Young neurons develop from stem cells known as neuroblasts. Again scientistscreated a description from two words. The suffix blast appears again and again inphysiologywithvariousprefixes.Blastisdefinedasanimmatureembryonicstageinthedevelopmentofacelltomaturity.BlastcomesfromtheGreekwordforbud.Neuroblastsare, therefore,stemcellscommittedtobecomingneuronsbyproceedingthroughseveralintermediateformslikeflowerbuds.Intheadultbrain,neuralstemcellsrepresentoneoftheintermediateformsofneuroblastsontheirwaytobecomingneuronsandglia.
Neuronscomprise10%ofthepopulationofcellsinthebrain.Theremaining90%of cells in brain tissue are named glia and microglia. The word glia is symbolic ofscientists’lackofunderstandingofthesecellsuntilrecentyears.GliaoriginatesfromtheGreekwordforglue.Dictionariesstillmistakenlydescribegliaasanetworkofbranchedcellsandfibersgluingtogetherthetissueofthebrainandspinalcord.
Brain gliawas at first divided into two classes,microglia andmacroglia, baseduponthephysicalsizeofthecells.Thatis,smallglialcells,microgliaandlargeglialcells,macroglia (Figure 1-7). Later, it was learned that size was not the distinguishing

characteristic.Newerstudiesdiscoveredmicrogliaisnotpartoftheglia,becauseitdoesnot originate in the embryo from neuroblasts like glia, but rather from the embryo’sprimitiveyolksaccells.
Figure 1-7: Illustration depicting four of the five types of cells in brain tissue, astrocyte, microglia, neuron andoligodendrocyte.Afifthcelltypeependymalcellisnotincludedinthisillustration.Illustration:©AlilaMedicalMedia
Microgliamigratesoveralongdistanceintheembryotojointheneuroblastsinthedeveloping brain. Microglia is related tomacrophages of the body’s immune system.Phage comes from the Greek word phagos meaning to eat. Both microglia andmacrophageseatcelldebrisindamagedtissue.Twocelltypesincludedinglia,astrocytesand oligodendrocytes (Figure 1-8), develop from the same embryonic stem cell asneurons.

Figure1-8:Neurons,oligodendrocytesandastrocytesalldescend from the sameneuroblasts, stemcells thatdevelopinto brain cells. Cell lineage but not size relationships are depicted. Illustration: This work is in the public domaincourtesyofNationalInstitutesofHealth,UnitedStates
The suffix cytes is used often in physiology combined with other descriptivewords.Cytesmeans cell, and it comes from aGreekword describing a hollow vessel.Cellsseenwiththefirstmicroscopesappearedtobehollowemptyvessels.
The prefix added to cyte always describes some characteristic of the cell indiscussion.Astrocytes appear as star shapedcells.TheGreekwordastronmeans a star.Therearetwoprefixesbeforecyteinthecaseofoligodendrocytes.OligoisaGreekwordforlittle.Dendromeanstree.Putting it together,oligodendrocytes translates tocells thatarelittleandbranchedliketrees.
Thebrain’sependymalcellsarealsoderivedfromneuroblasts.Theependymaisalayerofcubeshapedcellscoveringthesurfaceofthebrain’sfourinteriorchambers, theventricles, and the central canal of the spinal cord.Ventricle comes from a Latinwordmeaningbellyorcavity.EpendymaisderivedfromaGreekwordforacovering.
Ependymal cells secrete cerebrospinal fluidwhich cushions the brainwithin theskull.Thenameofthisfluiddescribesitslocation.CerebraisaLatinwordforbrainandspinain thiscaserefers to thehard,pointedbackbonewhichencloses thecordofaxonsleavingandenteringthebaseofthebrain.
USEFULTOOLSFormostanatomicterms,theLatinandGreekrootwordscanbefoundwithalittle
research.Often textbooks include a glossary containing someof them.Also,Wikipediaoffers a helpful list of Latin and Greek root words athttp://en.wikipedia.org/wiki/List_of_Greek_and_Latin_roots_in_English.

AnotherhelpfulreferenceisTheAmericanHeritageCollegeDictionary,publishedbyHoughtonMifflinCompany.Inthedictionary,thedefinitionofeachwordisfollowedby theLatinorGreek sourceword and itsmeaning.Discovering the senseof scientificwordsandhearingthemspokenoverandoveriscritical tothebrain’sabilitytoretrievethemfrommemorywhenneeded.
The scientificwords described here are found throughout this book and inmosttalksdescribingneuroscience.ItmayseemthischaptertakesalongtimetocompleteasyouworkwithGoogletranslate.Donotworryaboutit.Learningthisvocabularynowwillsaveagreatdealoftimelater.
SUMMARYCHAPTER1Humanbrain’sabilitytolearnanewlanguageisinfluencedbythelanguageitlearnedfirst
Humanbeingsrememberbetternewwordstheyhearthannewwordstheyread
Adding auditory input to reading of scientific words is an effective tool forcreatingwordmemory
HumananatomicstructureswerefirstnamedbyteacherswhospokeClassicalLatin
Memory of words forms quicker when meaning of new words is tied tosomethingalreadyknown
The American Heritage College Dictionary provides Latin and Greek rootwordswithitsdefinitionofEnglishwords
The Google translate website is a useful tool for practicing the language ofbrainscience

[2]
HowtheHumanBrainIsOrganizedBRAINSTRUCTUREISDESCRIBEDinthreeways.First,visualobservationof
thewhole brain establishes the overall layout of larger structures. Second,microscopicvisualization of fixed, sliced and stained brain tissue displays its cell structure. Third,videosof livingbrainobtainedwith computerizedmicroscopesdemonstratemobility ofresidentneurons.
Neuron signaling practices of the human brain aremore complex than those ofother species.Yet, the gross organization of brain tissue is similar betweenmammalianspecies.And,agreatdealofwhatisknownaboutthehumanbrain’soperationalsystemscomesfromobservationsofrats,miceandnon-humanprimates.
THEVISIBLEBRAINThehumanbrainisasoftfragileorganprotectedfrominjurybythehardbonycase
of the head.Because of its soft character, the brainwas considered an irrelevant organuntil the late 1800s. Today scientists recognize the brain as the physical location ofconsciousness.
Theexpressiongrossanatomy refers to theexternalfeaturesofadissectedtissueororgan.Itincludeseverythingapersonseeswhenviewingabodypartwithoutthehelpofamicroscope.Itmayalsoincludethetextureofthetissue.Forexample,doesthetissuefeelfirmorspongy?
Thecowbrain(Figure2-1),likethehumanbrainhasacerebellumandarightandleft hemisphere. The hemispheres connect to each other by a bridge of neuron axonsnamed the corpus callosum. Corpus callosum comes from two Latin words, corpusreferring to a body of tissue and callosum indicating its hard texture, much like theconsistencyofacallus.
Thecorpuscallosumappearsasabroadwhitebandoftissuecomposedofaxonsoftheneuronsresidinginthebrainhemispheres.Theaxonsofthecorpuscallosumconnectcorresponding parts of the hemispheres, and permit the right hemisphere and the lefthemispheretocoordinatetheiractivity.Theponsisastructurethatattachesthecerebellumtotherestofthebrain.

Figure2-1:Grossanatomyofadissectedcowbrain.Thisfigureshowstheplacementoflargebrainformations.Labeledareascorrespondtosimilarstructuresfoundinhumanbrains.Photo:©decade3d
Incontrast to thesoftconsistencyofdissectedcowbrainshowninFigure2-1,abrainpreservedwithchemicalsfeelslikerubber.Figure2-2isapictureofahumanbraintreated with formalin to preserve it from decay. The increased mechanical strength ofpreservedbrainallows the tissue tobe sliced into thin sequential tissue sections.Tissuesectionsmay be stainedwith various dyes to observe their cellular organizationwith amicroscope.
Notice the deep folds in the surface of the human hemispheres (Figure 2-2).Increased depth of the surface folds permits greater expansion of the volume of thehemisphereswithoutrequiringthehumanskulltoenlarge.Similarfoldsofthecowbrain(Figure2-1)areshallowincomparison.
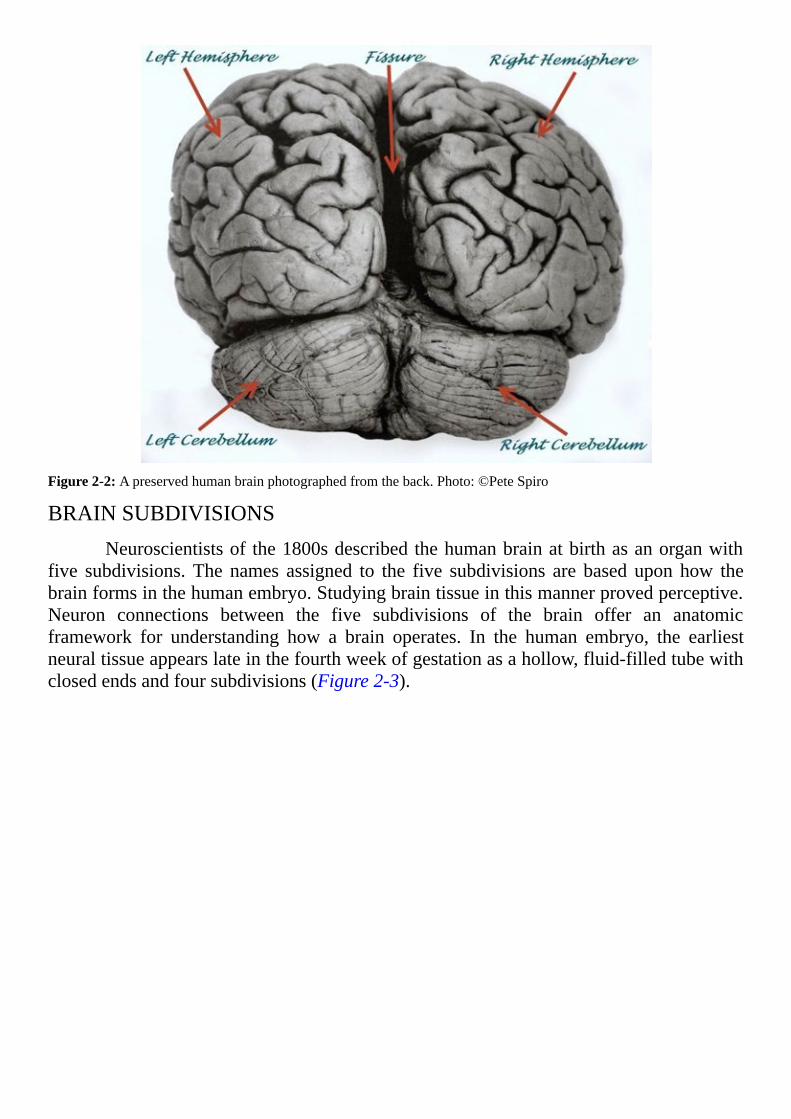
Figure2-2:Apreservedhumanbrainphotographedfromtheback.Photo:©PeteSpiro
BRAINSUBDIVISIONSNeuroscientistsof the1800sdescribedthehumanbrainatbirthasanorganwith
five subdivisions.The names assigned to the five subdivisions are based upon how thebrainformsinthehumanembryo.Studyingbraintissueinthismannerprovedperceptive.Neuron connections between the five subdivisions of the brain offer an anatomicframework for understanding how a brain operates. In the human embryo, the earliestneuraltissueappearslateinthefourthweekofgestationasahollow,fluid-filledtubewithclosedendsandfoursubdivisions(Figure2-3).

Figure 2-3: This diagram shows the position of the neural tube at about four weeks gestation in a human embryo.Illustration:©Kurzon
ThewhitedotshownintheblueprosencephalonofFigure2-3develops into theopticnerve,retinaandirisoftheeyes.Eyesoccupyauniquesettingbeingtheonlypartsofthebrainwithoutabonycover.
Forallmammalsastheembryomatures,theprosencephalondividesintotwoareasnamed the telencephalon and diencephalon. The embryonic subdivision identified asmesencephalon continues tomaintain its original name even after birth.The embryonicsubdivision in Figure 2-3 labeled rhombencephalon, also matures into two brainsubdivisionsnamedthemetencephalonandthemyelencephalon.Thespinalcordmaturestobecometheadultspinalcord.
Thus,thenamesofthefivesubdivisionsofthehumanbrainatbirthare:
Telencephalon
Diencephalon
Mesencephalon
Metencephalon
Myelencephalon
Remembering thenamesof the five anatomical subdivisionsof thebrain canbeapproached in the same way as the scientific terms in Chapter 1, “Tips & Tricks forLearningScientificLanguage.”Noticethenamesofthebrainsubdivisionallcontainthesuffix cephalon.Cephalon originates from a Greek word meaning head. Therefore, ananatomicnameincludingcephalonindicatesapartofaperson’shead.
Knowing the names of the brain subdivisions is useful, because neuroscientistsrefer to themoften.For example, knowledgeof the positionof the brain’s subdivisions

wouldbeneededtodescribethepathtothespinalcordofthebrain’sneuronsdevotedtoinitiatingmusclemovement.
TELENCEPHALONThe prefix of each subdivision name is descriptive of its gross anatomy. In the
humanbrainthe telencephalon is therightandlefthemispheres,seenwhenlookingatawhole brain as in Figure 2-1 and Figure 2-2. In Greek telos meant far end. Duringembryologicdevelopmentthispartofthebrainmaturesatthefarendoftheneuraltube.
Across species the telencephalon is thebrain subdivisionmost recently evolved.The telencephalon is required for rational thought,making decisions and implementingchoices.The outermost layer of the telencephalon is named thecerebralcortex.CortexderivesfromaLatinworkmeaningbark,asintreebark.Thecerebralcortexconsistsoflargeneuronsinlayers(Figure2-4).
Figure2-4:ThepositionofcerebralcortexneuronsdrawnbySantiagoRamónyCajal.Neuroncellbodiesarearrangedin horizontal layers labeled A-F. Axons and dendrites form a network for optimum interaction between neurons.Drawing:Thisworkisinthepublicdomain.
The neurons of the frontal cerebral cortex, the brain region under a person’sforehead, serve as decisionmakers.Axons of cortical neurons are sometimes long, andmany of them connect with multiple areas of the brain and spinal cord. For example,axons of cerebral cortical neurons devoted to control of body movement may extendseveral feetbeforeconnecting to spinal cordneurons,which in turn send their axons tomusclestocausecontraction.
METENCEPHALON
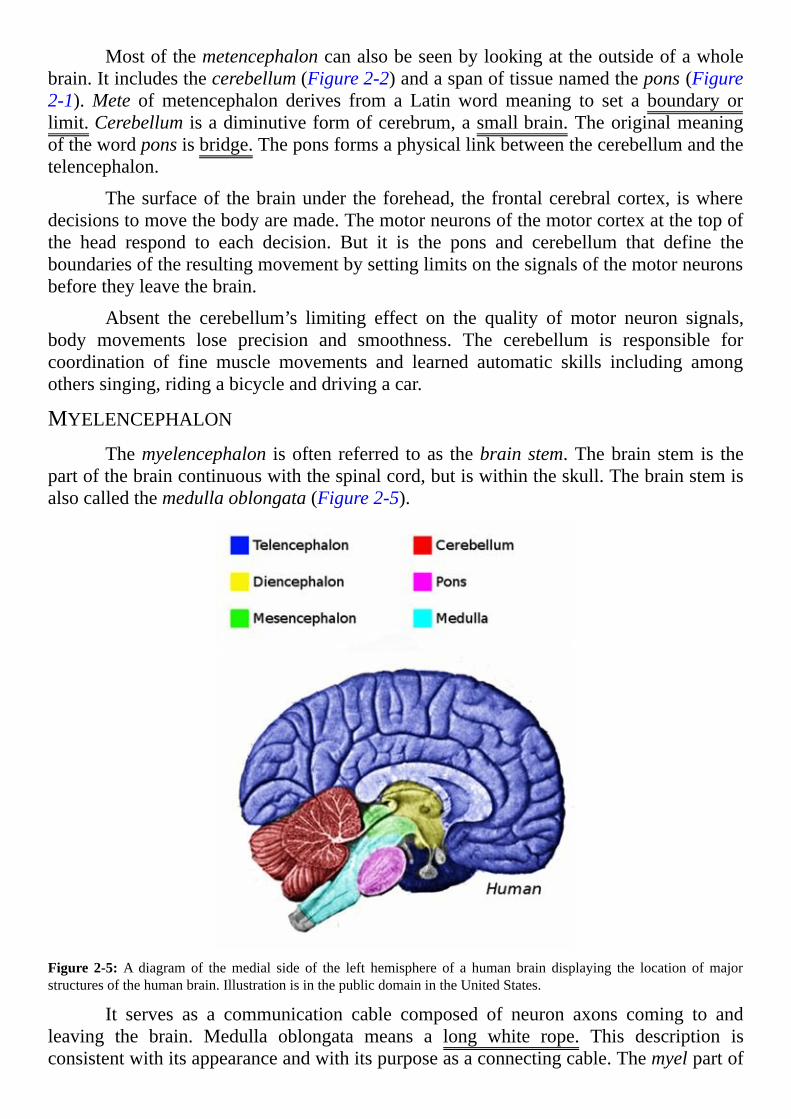
Mostofthemetencephaloncanalsobeseenbylookingattheoutsideofawholebrain.Itincludesthecerebellum(Figure2-2)andaspanoftissuenamedthepons(Figure2-1).Mete ofmetencephalon derives from a Latinwordmeaning to set a boundary orlimit.Cerebellum isadiminutiveformofcerebrum,asmallbrain.Theoriginalmeaningofthewordponsisbridge.Theponsformsaphysicallinkbetweenthecerebellumandthetelencephalon.
Thesurfaceof thebrainunder theforehead, thefrontalcerebralcortex, iswheredecisionstomovethebodyaremade.Themotorneuronsofthemotorcortexatthetopofthe head respond to each decision. But it is the pons and cerebellum that define theboundariesoftheresultingmovementbysettinglimitsonthesignalsofthemotorneuronsbeforetheyleavethebrain.
Absent the cerebellum’s limiting effect on the quality of motor neuron signals,body movements lose precision and smoothness. The cerebellum is responsible forcoordination of fine muscle movements and learned automatic skills including amongotherssinging,ridingabicycleanddrivingacar.
MYELENCEPHALON
Themyelencephalon isoftenreferred toas thebrainstem.Thebrain stem is thepartofthebraincontinuouswiththespinalcord,butiswithintheskull.Thebrainstemisalsocalledthemedullaoblongata(Figure2-5).
Figure 2-5:A diagram of themedial side of the left hemisphere of a human brain displaying the location ofmajorstructuresofthehumanbrain.IllustrationisinthepublicdomainintheUnitedStates.
It serves as a communication cable composed of neuron axons coming to andleaving the brain. Medulla oblongata means a long white rope. This description isconsistentwithitsappearanceandwithitspurposeasaconnectingcable.Themyelpartof

thenamemyelencephalonreferstothefattymaterialwrappedaroundneuronaxonsnamedmyelin.
DIENCEPHALONThe diencephalon of the human brain cannot be seen without dissection. It lies
immediatelybelowtherightandlefthemispheresandthecorpuscallosum(Figure2-5).Thediencephalonincludestwoparts.Inthiscasetheprefixdidescribesadivisionofthebraincontainingtwoclustersofneuroncellbodies,thethalamusandhypothalamus.
Theoriginalmeaningofthalamusisanteroomorentrance.Thisisanappropriatedescriptivename for thisdivision,becausealmostall informationcoming into thebrainmustbeprocessedby the thalamusbefore reachingother regions.The thalamusplaysacentral role in managing information arriving from the eyes, ears and other sensoryorgans.
Thehypothalamus sitsbelow the thalamus.Theprefixhypo isGreekandmeansunderorbeneath.Thehypothalamuscontrolsbodytemperature,hunger,thirstandreleaseofhormonesfromthebody’smasterendocrinegland,thepituitary.Thepituitarysitsinapocket in the bone of the skull below the hypothalamus and is connected to thehypothalamusbythepituitarystalk,asmalltubeofneuraltissue.Pituitaryisanothernamebaseduponamistakenscientificbelief.ItcomesfromtheLatinformucus,becauseitwasthoughttobethesourceofmucusinthenoseandsinuses.
MESENCEPHALON
Themesencephalonormidbrain is locateddeep in thecenterof thehumanbrain(Figure2-5).MesisavariationoftheGreekwordmesosormiddle.Thisisanolddivisionintermsofthebrain’sevolution.Allanimalspossessthisbraindivisionthatco-ordinatescomplex reflex reactions.Themesencephalonworkswith the brain stem to initiate andperform the vital unconscious processes of the body like regulation of breathing. Themesencephalonisdetachedfromintellectualreasoning.

Figure2-6:MagneticResonanceImage(MRI)ofalivinghumanbraindisplayingthemedialsideofthelefthemisphere.MRIImage:©Cessna152
TrytoidentifythestructureslabeledinFigure2-5intheimageofalivinghumanbrain shown in Figure 2-6. Where are the cerebellum and the pons in Figure 2-6?ComparingFigure2-5andFigure2-6estimatethelocationofthemidbrain.
InFigure2-6,thecorpuscallosumisthewhite,curvedstructureinthemiddleoftheimage.Themedullaoblongataisimmediatelyabovethespinalcord.Thespinalcordisat the bottomof the image between vertebrae and is outside the skull.The scalp is thewhiteouter lineover theskull.Thedarkbandbeneath the scalp isboneof theskull.Ahole in the bottom of the skull permits neuron axons to leave and enter the brain. Theopeningisnamedtheforamenmagnumandliterallymeansalargehole.
GRAYMATTERANDWHITEMATTER
Figure2-7:Cutsurfaceofafixedhumanbrainshowinggraymatterandwhitematter.Photo:©JohnA.Beal

When formalin fixed brain is sliced open, part of the interior appearswhite andpart is a light gray color. (Figure2-7). Late in the 19th centurywhen dyes specific forneuronsbecameavailable for the first time, itwasdiscoveredgraymatter is clustersofneuron cell bodies.Whitematter iswhite because it contains a largenumberof neuronaxonscoveredwithwhitemyelin.
INSIDETHEBRAIN
DEADBRAINMICROSCOPY
Microanatomyreferstoanatomicfeaturesofatissuedetectablebythehumaneyeonlyaftermagnification.Themicroscopewas invented in the late1500s.Cells in livingtissuewere first described byRobertHooke in 1665.Yet, as late as themid-nineteenthcenturysomescientistsstillbelievedthebrainanexceptiontotherulethatalllivingtissueis made up of cells. Cells could not be seen in brain tissue because the fatty myelininterferedwithdyesnecessarytoseetheoutlineofthecells.
The first tissue-specific stain for neuron cell bodies was discovered in 1884 byFranz Nissl. Near the same time Carl Weigert developed a dye absorbed by the fattymyelinmaterial of the brain and not by other brain tissue components. Comparison ofthese two staining methods confirmed gray brain matter contains large collections ofneuron cell bodies, and white matter is white because of the large number of axonscoveredwithmyelin.
Inthelate1800sandduringtheinitialyearsofthe20thcentury,SantiagoRamónyCajalmadehisfirstrevealingdrawingsofneuronsinbraintissue(Figure2-8).Thesilverstainingprocessheusedwasdevelopedin1873byCamilloGolgi.Golgi’sstaindisplaysonlyasmallpercentageoftheneuronsinatissueslice.Thisisfortunatebecauseastainthatmarksalloftheneuronsinthetissuewouldobscuretheshapeofindividualneurons.Nowover150differenttypesofbrainneuronsaredistinguishablebasedupontheshapeoftheirdendritesalone.Themanyuniquedendritepatternsmaketheneuronthemostdiversecelltypeinthebody.

Figure2-8:Adrawingofneuronsinthechickcerebellum.Noticethevarietyintheshapeandsizeoftheneuroncellbodies(darkroundandovalstructures),axonsanddendrites.Drawing:SantiagoRamónyCajal1905.Thisworkisinthepublicdomain.
During the 20th century, many brain specific stains were developed formicroscopic evaluation of fixed tissue slices. Modern methods permit a more detailedanalysis of the cells in various brain regions. Thin pieces of tissue evaluated withcontemporarytechniquespresentadifferentviewofthebrainthanobservedbySantiagoRamónyCajal.
Modern staining protocols produce images where the number and size of thebrain’s neurons, microglia and glia become visible. In photos taken through amicroscope’s magnifying lens, neurons exhibit larger cell bodies than other brain cells(Figure2-9).
Figure2-9:AhighmagnificationphotomicrographofaHPS(hematoxylinphloxinesaffron)stainedbrainbiopsy.Thispieceofbraintissueismostlygraymatterwithasmallamountofwhitematterinthelowerleftquarteroftheimage.Photomicrograph:©Nephron

Ittakesalittlepracticetoseethedifferencebetweengraymatterandwhitematterin stained brain sections.Areas dominated by large neuron cell bodies are graymatter.Areas where neurons are few in number are white matter. Glia is found dispersedthroughoutbothgraymatterandwhitematter.
Notice inFigure2-9 themany large neurons present in the section labeled graymatter.Thecellpointedoutasaglialcellisasmalleranddarkerstainingbodythantheneurons.Theglialcellmaybeanastrocytebecausemanyastrocytessurroundtheneuroncellbodiesofthegraymatter.Thesmallestdarkstainingcellsencircledbyawhitehaloareoligodendrocytes.Thehalo iswhere fat of theirmyelinmembranewas removedbychemicalsusedinthestainingprocedure.Thepartofthephotomicrographlabeledwhitematterisprimarilyaxonsoftheneuronslocatedinthegraymattermixedwithglia.
LIVEBRAINMICROSCOPY
Amethodofmicroscopydevelopedsince1990allowsscientiststostudyalivingbrain. This procedure employs instruments known as optical imaging systems. Opticalimaging systems provide the spatial resolution necessary to reveal individual neurondetailsliketheshapeofdendriticspines.Thisisaninvasiveprocedurerestrictedtouseinanimalstudies.Itrequireseitherthinningoftheboneoftheskull,orskullremovalovertheareaofinterest.Studiesconductedinmice,rats,catsandnon-humanprimatesprovidemostofthisdata.
When using rats, a permanent window can be implanted where the skull isremoved and imaging living neurons can be performed for a year ormore in the sameanimal.Miniportabledevicesallowimagingofneuronstooccurwhileratsexploretheirenvironment.
Atpresentthefinestopticalbrainimagespenetratetoadepthofabout1millimeterofthebrainsurface.Forthistypeofoptical imagingofneuronstobeuseful inhumans,thedepthoflightpenetrationmustbeimproved,andawaytoavoidanopenskullmustbefound. Improvement of optical imaging technology is a goal of the present worldwideemphasisonbrainresearch.
In reality thepicture detectedbyoptical imaging systems, confocalmicroscopesand2photonimagingmicroscopes,cannotbeseendirectlybyhumaneyes.Thesearenotmicroscopes in the same sense Robert Hooke’s instrument is a microscope. Lightinformationfrommodernimagingsystemsissenttoacomputerandthecomputerformsapicturethatthehumaneyerecognizes(Figure2-10).

Figure 2-10: A pyramidal neuron expressing Green Fluorescent Protein (GFP) in a mouse visual cortex.Photomicrograph:©Nrets
Light captured from brain tissue by optical imaging systems is produced byfluorescentmolecules.A focused laser beam is used to increase the energy level of thefluorescentmolecules in the tissuewithfastpulsesof infrared light.Betweenpulses thefluorescentmolecules return to their normal energy state. In the process of returning totheir baseline energy level, each fluorescent molecule emits a photon of light at aparticularwavelengthinthevisiblelightrange.
The amount of light produced by fluorescent molecules is so low it must beenhancedaspartofthedetectionprocess.Thus,acomputerisrequiredtocompileintoavisualimagethelightemitted,thelightscatterinformationandthepositionofthefocusedlaserbeaminthetissue.
For brain cells to possess fluorescentmolecules, the animalmustmake thembyusingitsowncellmachineryforsynthesizingmolecules.Animalsmodifiedtodothisarenamedtransgenicanimals.Acommonwaytoproducetransgenicanimalsistoinjectthegenes required for synthesis of the fluorescentmolecule into the nucleus of a fertilizedegg.
Correcttimingofthestepsoftheprocedureisessential.Foraforeigngenetobeincorporated into an animal’s geneticmaterial, the alien genemust be addedbefore thechromosomesof thespermandoocytemerge (Figure2-11). Inmostexperiments,up to40% of the mice born from such embryos will express the foreign gene and makefluorescentprotein.Expressionoftheforeigngenecanberestrictedtoaparticulartissueintheanimalbyincludingapartthatrespondstomoleculesuniquetothetissue.

Figure 2-11: Diagram of sperm injection into an oocyte to create a fertilized egg. Themicro-manipulator holds theoocytewhilethemicro-injectorontherightplacesasinglespermintheoocyte.Illustration:©KDS444
Computerized microscopic techniques not only permit observation of brainanatomyingreaterdetailbutalsodisplaytheanimatednatureofbraintissue.Videosmadeby Michael J. Schell http://youtu.be/Mhcaz6_fzZA and http://youtu.be/Cjjdky96ubcdemonstratethemotionofdendriticspinesinrealtime.
SUMMARYCHAPTER2The organization of the major subdivisions of the brain is similar in mostmammalianspecies
Thehumanbraindevelopsinto5anatomicsubdivisionsbybirth
Eachbrainsubdivisionincludesadistinctivefeaturereflectedinitsname
Residentneuronsconnectbrainregionstoeachotherwiththeiraxons
Thegraymatterofformalinfixedbraintissuecontainsneuroncellbodies
Thewhitematterofformalinfixedbrainisneuronaxons
Neurons distribute into anatomic categories based upon the shape of theirdendrite
Over 150 different types of neuron categories exist based on the shape ofdendrites
Optical imaging systems for living animals deliver the spatial resolution

necessary to reveal individual neurons, their extensions and over time theirmobility

[3]
QualityControlofBrain’sExtracellularFluidsTHEHUMANBRAINISISOLATEDfromtherestofthebodyinmultipleways.
Itscellsmanagetheirbusinesslikemembersofanindependentsociety.Thebraindependsupontherestofthebodyonlyforanadequatesupplyofoxygenandglucoseandasmall,select group of nutrients and growth factors. It connects to the outsideworld primarilythroughitsownneuron-basedsensorysystems.
Physiologicmechanismsadapttothebrain’suniquecircumstances.Onevariationofnormalphysiologyisrevealedintheunusualcharacteristicsofthebrain’smaintenanceprogramforitsfluidcompartments.Threefluidcompartmentssupportbraincellactivities.They are the intracellular fluid named cytoplasm and extracellular fluids known asinterstitial fluid that surrounds blood vessels, neurons, glia and microglia and thecerebrospinalfluidthatcushionsthebrainwithintheskull.
Whileexchangeofmoleculesbetweenfluidcompartmentsisadynamicprocessinallbodytissues,braintissueexhibitsanunusualandelegantformofmolecularexchangebetweenits fluidcompartments.Thischapterfocusesuponqualitycontrolof thebrain’sextracellular fluids.The followingchaptersexplainhowdynamicsbetweenextracellularand intracellular fluid compartments support, and are vital to, the electrical signalingsystemsinthebrain.
FLUIDSURROUNDINGCELLS
VIRCHOW-ROBINSPACEInterstitial fluid surroundingbrain cells andbloodvessels consistsofwaterwith
dissolved sugars, salts, fatty acids, amino acids, hormones, neurotransmitters and thewater solublewaste products generated by cell activity. Largerwater-solublemoleculesincludingproteinsareabsentinnormalcircumstances.
Unlikeelsewhereinthebody,interstitialfluidaroundneuronsandgliaisseparatedfrominterstitialfluidaroundthebrain’sarteriesandarteriolesbypiamater,anultra-thinmembrane. The name pia mater comes fromMedieval Latin and translates into tendermother. The space created by the presence of the pia mater around arterial vessels isknownastheVirchow-Robinspace.
Virchow-Robin is a composite of the names of the two investigators whodemonstrated existence of this fluid space in the brain, RudolphVirchow andCharles-Philippe Robin. No comparable pia mater sheath is present around the veins. TheVirchow-Robinspaceendswherebloodcapillariesbegin.Thereitiseliminatedbyfusionofthecapillaryendothelialcellswiththemembraneofastrocyteglialcells.
Virchow-Robin space isolates interstitial fluid from all proteins and other largemolecules leaked from arterial blood vessels. Virchow-Robin space also helps to clearwaste-containing interstitial fluid surrounding neurons and glia. Waste-containing

interstitialfluidflowsthroughthepiamaterintotheVirchow-Robinspaceandthendrainsintothelymphaticsystemoftheheadandneck(Figure3-1)forreturntotheheart.
Figure3-1:Anatomicmodeloftheveinsandlymphaticvesselsoftheheadandneck.Photo:©Tinydevil
Lymphocytes, white blood cells of the immune system, escaping from bloodvessels become trapped in the Virchow-Robin space and are returned to the blood. Inhealthybrain,lymphocytesandothercellsoftheperipheralimmunesystemareexcludedfrom the interstitial fluidaroundneuronsandglia.Onlywhen thebrain’sown immune-likecells,themicroglia,becomeoverpoweredbyinfectionortraumadoimmunesystemlymphocytesenterintointerstitialfluidsurroundingneurons.
BLOODBRAINBARRIERBlood in vascular vessels, arteries and veins, is often included in the normal
descriptionof thebody’sextracellularfluidcompartments.Elsewherein thebody,directexchangeofwaterandwater-solublemoleculesbetweenbloodandtheinterstitialfluidisunconstrained at capillaries. In brain, however, membranes of the capillaries and postcapillarysmallveinslimitpassageoflowmolecularweightmaterialandwater.
The quantity of low molecular weight substances, hormones, amino acids,neurotransmitters and othermetabolites oscillates in blood under normal circumstances.Fluctuationsinthequantityofthesemetabolitesintheinterstitialfluidofthebrainwouldcauseunacceptabledisruptionofneuronfunction.About98%ofblood’ssmallmoleculesdonotenterthebrainthroughitscapillarysystem.
The limitationon releaseof lowmolecularweightmaterial fromblood increasesosmotic pressure in brain capillaries. Brain capillary osmotic pressure, a force pullingwaterintocapillariescreatedbythehighnumberofmoleculesunabletoleavethebloodis

great.Fartoomuchwaterwouldberemovedfromthebrain’s interstitialfluidwithoutareduction in permeability of brain capillaries for water. Entry of water and moleculesnecessaryforbrainwell-beingusesadifferentpaththatisdescribedbelow.
FIGURE3-2:Diagramshowingtightjunctionsatthebloodbrainbarrierincludingsomeofthemainproteins.Blut=blood,Gehirn=brain,Illustration:©ArminKübelbeck
Abloodbrainbarrier is createdby tight lacing together of capillary endothelialcells by protein formationswith the descriptive namesadherens junctions (Figure 3-2).Thejunctionscausemoleculesinbloodtobetransportedthroughthecapillaryendothelialcells.
The blood capillary membrane is composed of a cell type named endothelial.Endothelialcellmembraneshaveproteinsthattransferspecificmoleculesfromonesidetothe other. About 10-15% of the proteins in the brain’s capillary system transportmolecules. Glucose, the brain’s main energy source, is transported into the brain by aproteinnamedtheglucosetransportertype1(GLUT1)ofcapillaryendothelialcells.
GLUT1doesnot requireenergyor insulin toperformits transferofglucose intothebrain.Itfacilitatesdiffusionofglucosefromthebloodtothebrain’sinterstitialfluidina passive fashion. Glucose is almost always about twice as high in blood as in braininterstitial fluid. Itshighbloodconcentration favorsglucosemovement intobrain tissuethroughchannelsintheGLUT1proteinoptimizedforthestructureofglucose.
Insulinmadeinthepancreasalsoenterssomebrainregionsbyusinganendothelialcell transporter protein. In the brain pancreatic insulin regulates cell growth. Pancreaticinsulin is reported to support survivalofneuronswithoutaffecting theiruseofglucose.Theneuronsofthecerebralcortex,hypothalamusandcerebellumareparticularlysensitiveto insulin. Insulin supports brain uptake of the amino acids needed to synthesize the

neurotransmittersnorepinephrine,dopamineandserotonin.
Gasexchangeisnotaffectedbythebrain’scapillarybarrier.Becausebothoxygenand carbondioxide dissolve in the fattymatrix of cellmembranes, they passwith easethroughthebrain’scapillaries.Othersmallfattymoleculesalsopassthroughthecapillarymembraneofthebloodbrainbarrier.However,manyofthemaretransferredbackintothebloodbymembraneproteins.
Anadditionallayerofphysicalandoperationalsupportforthebloodbrainbarriercomesfrommembraneprojectionsofanotherbraincellcalledanastrocyte(Figure3-3).The astrocyte’s foot-like structures make direct contact with brain capillaries. Recentstudiesproduceddatasuggestingastrocytes, inadditiontoprovidingmechanicalsupportfor the blood brain barrier, also participate in secretion of chemical factors to blockimmunecellsfromenteringthebrain.
Figure3-3:Anartist’srenditionofabloodbrainbarriercapillarywithattachedastrocytes.Illustration:©BenBrahimMohammed
Thebloodbrainbarrier’smostimportantpurposeispreventionofentryofbacteria,virus,immunecellsandlargeproteinslikeantibodies.Immunesystemcellsenterthebrainonlywhenitsbloodvesselssufferdamage.Thepresenceofimmunecellsintissueleadstoinflammation.Typicalinflammationisnotwelltoleratedbybraintissuebecauseitmakescapillariesleakypermittingpassageofwaterandotherbloodbornemolecules.
THEMENINGES
Themeningesarelayersofmembranescoveringtheentiresurfaceofthebrainandspinalcord.TheirnameisderivedfromtheGreekwordmeninxmeaningmembrane.Themeninges providemechanical protection for brain tissue, furnish a pathway for flowofcerebrospinal fluid to cushion the brain and spinal cord and supply support for bloodvessels entering and leaving the brain. Formation of cerebrospinal fluid and its passagethroughthebrainiscoveredinthefollowingsection.
Three layersofmeningealmembranes,duramater, arachnoidmembraneandpiamatercoverthebrain(Figure3-4).ThenameduramaterisfromGreek,andittranslatestotoughmother.Duramaterisathickfibroustissue.Therearetwolayersofduramater,a

layerneartheboneoftheskullcontinuouswiththebone’sownmembranouswrapandaninnerlayerclosertothebrain.Thetwolayersofduramaterencloseandsupportthelargevenouschannelsandsinusesthatreturnbloodtotheheart.
Duramater also forms a sac around the next layer ofmembrane, thearachnoidmembrane. The arachnoid membrane is thin transparent fibrous tissue. It acquired itsnamebecauseofthespiderwebappearanceofitsdelicatefibersthatconnectittothelayerofpiamaterunderneath.TheGreekwordforspiderisarachneandthesuffixoidmeansintheimageof.
Figure3-4:Thisdiagram represents a section across the topof thehuman skull. It describes the arrangementof themeningealmembranescoveringthebrain.Theorangearea,thesubarachnoidcavityfillswithcerebrospinalfluid.Thisfigureisbasedonplate769fromGray’sAnatomy.Illustration:©OpenStaxCollege
Betweenthearachnoidmembraneandthenextlayerofmeninges,thepiamater,isawebbedspacefilledwithcerebrospinalfluid,thesubarachnoidcavity.Thesub-arachnoidcavityreceivescerebrospinalfluidflowingoutof thefourthventricle.Thesubarachnoidcavityencirclestheentirebrainandspinalcordprovidingapadofprotection.Allbloodvessels entering the brain, branches of the internal carotids and vertebral arteries, passthrough the subarachnoid cavity. Cranial nerves exiting the bottom of the brain alsopassagethroughthesubarachnoidcavity.
Piamateristheinnermostmembraneanditadherestothecerebralcortexrunningdown into the surface fissures.At the surface fissures,piamater is a thin fibrous tissuethat is impermeable to fluid. It forms a sheer translucent envelope spanning almost theentirebrain.Piamaterisanchoredtothesurfaceofthebrainbymembraneextensionsofastrocyteslikethosefoundreinforcingthebloodbrainbarrier(Figure3-3).
Pia mater also forms a sheath around the cerebral arteries passing through thesubarachnoidcavity.ThepiamaterarterialsheathinthesubarachnoidcavityiscontinuouswiththepiamateroftheVirchow-Robinspacedescribedabove.Thepiamatersheathofthe Virchow-Robin space is permeable to fluid, but the pia mater sheath of thesubarachnoidcavityiswatertightsupplyingabarrierbetweencerebrospinalfluidexitingintothevenoussinusandincomingbloodvessels.
CEREBROSPINALFLUID
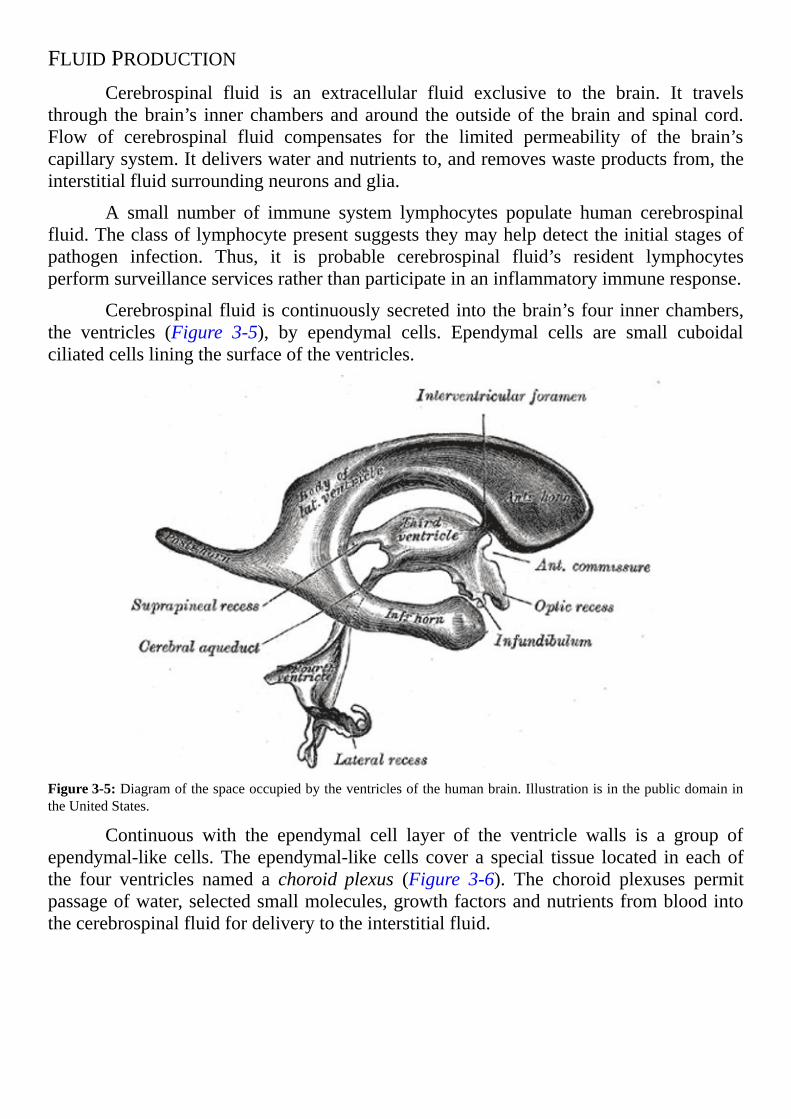
FLUIDPRODUCTIONCerebrospinal fluid is an extracellular fluid exclusive to the brain. It travels
through thebrain’s inner chambersandaround theoutsideof thebrainand spinal cord.Flow of cerebrospinal fluid compensates for the limited permeability of the brain’scapillarysystem.Itdeliverswaterandnutrientsto,andremoveswasteproductsfrom,theinterstitialfluidsurroundingneuronsandglia.
A small number of immune system lymphocytes populate human cerebrospinalfluid.Theclassoflymphocytepresentsuggeststheymayhelpdetecttheinitialstagesofpathogen infection. Thus, it is probable cerebrospinal fluid’s resident lymphocytesperformsurveillanceservicesratherthanparticipateinaninflammatoryimmuneresponse.
Cerebrospinalfluid iscontinuouslysecreted into thebrain’sfour innerchambers,the ventricles (Figure 3-5), by ependymal cells. Ependymal cells are small cuboidalciliatedcellsliningthesurfaceoftheventricles.
Figure3-5:Diagramofthespaceoccupiedbytheventriclesofthehumanbrain.IllustrationisinthepublicdomainintheUnitedStates.
Continuous with the ependymal cell layer of the ventricle walls is a group ofependymal-likecells.Theependymal-likecells cover a special tissue located in eachofthe four ventricles named a choroid plexus (Figure 3-6). The choroid plexuses permitpassageofwater,selectedsmallmolecules,growthfactorsandnutrientsfrombloodintothecerebrospinalfluidfordeliverytotheinterstitialfluid.

Figure3-6:Thislateralventricleofadissectedhumanbrainshowsachoroidplexus.Photo:©Anatomist90
The interior of a choroid plexus is a convoluted vascular network of looseconnective tissue and large capillaries. These capillaries possess a structure similar tosmall perforated veins.Most blood-borne smallmolecules pass through and around theendothelial cells of these capillaries bringingwaterwith them into the fluidwithin thechoroidplexus.The ependymal-like cells covering the choroidplexusdisplay extensivefoldson their side facing thebloodvessels.Creases in themembrane expand the cell’sabilitytoabsorbfluidreleasedfromthesieve-likecapillaries.
Fluid within the choroid plexus must be processed through the ependymal-likecells to become cerebrospinal fluid.Tight junctions tie together ependymal-like cells ofthe choroid plexus preventing fluids from going around them. Like other cells lininghollow spaces, the characteristics of their cell membrane on the side facing the bloodcapillaries,theirbasalmembrane,differfromcharacteristicsoftheirmembranefacingtheopenspaceoftheventricle.Thepartoftheirmembranefacingtheopenchamberiscalledtheirapicalmembrane.
Proteins in the apical membrane transfer ions from the ependymal-like cell’scytoplasm into the cerebrospinal fluid. The ions create an osmotic pressure in thecerebrospinal fluid. When sufficient osmotic pressure is generated by the ion transferprocess,waterispulledoutofthecellsintothecerebrospinalfluidincreasingitsvolume.Nutrientsandgrowthfactorspickedupbythebasalmembranepassagethroughthecellsbeforefollowingwaterintothecerebrospinalfluid.Ependymalcellsalsomanufactureandsecreteawidevarietyofbiologicsubstancestosupportbrainhealth.
In addition to secreting cerebrospinal fluid, the choroid plexus is active in theclearanceofdrugsandpollutantsfromthebrain.Inareverseprocess,underappropriatecircumstance, the ependymal-like cells reabsorb cerebrospinal fluid. Reabsorbedcerebrospinalfluidmovesbackintothechoroidplexuscapillaries.

The ependymal cells lining thewall of the ventricles also absorbs cerebrospinalfluid, but for a different purpose than the choroid plexus. Along the surface of theventricles absorption of cerebrospinal fluid is amechanism for exchanging nutrients incerebrospinalfluidforcellularwasteproductsininterstitialfluid.
Inhumansabout500millilitersofcerebrospinal fluid isproducedeachday.Thevolumeoftheentiresystemofchannelsforcerebrospinalfluidisonly150–270milliliters.Therefore,cerebrospinalfluidisreplacedabout2–4timesperday.Pressuregeneratedbythecontinuousproductionofcerebrospinalfluidcausesit toflowthroughtheventricles,thespinalcordandmembranessurroundingthebrainandspinalcord.
PATHTHROUGHTHECENTRALNERVOUSSYSTEMToillustrate thepath takenbycerebrospinal fluid through,aroundandoutof the
brain, the anatomy of the systemmust be reviewed.The central portion of the brain isoccupiedbylateralspacesinthehemispheresnamedthelateralventriclesandtwootheropenareasnamedthethirdventricleandthefourthventricle(Figure3-5).Anopenarea,thecentralcanal,alsorunsthelengthofthespinalcord.
Cerebrospinal fluid formed in the two lateral ventricles passes through theinterventricular foramen, a small open area connecting the ventricles, into the thirdventricle.ReferbacktoFigure3-5toseethelocationoftheopenspacesincludedinthispath.
The third ventricle sits between the thalamus of the right hemisphere and thethalamusofthelefthemisphere.Therecerebrospinalfluidofthelateralventriclesmixeswith cerebrospinal fluid formed in the third ventricle. From the third ventricle thecombinedcerebrospinalfluidofthreeventriclesmovesthroughthecerebralaqueductintothefourthventricle.
Thefourthventricle liesbetweentherightandleftponsandcerebellum.Mergedcerebrospinal fluid of the four ventricles exits the fourth ventricle and enters into thecentralcanalofthespinalcordandintosubarachnoidcavityofthemeninges.
Cerebrospinalfluidflowfromtheventriclesreachingthetopoftheheadwithinthesubarachnoid cavity passes through structures in the arachnoid membrane namedarachnoidgranulations.Itflowsthroughthearachnoidgranulationsandmixeswithbloodinthevenoussuperiorsagittalsinus(Figure3-4).
Thearachnoidgranulationsactasonewayvalvesforflowofcerebrospinal fluidoutofthebrain.Pressureinthecerebrospinalfluidisusuallyhigherthanpressureinthevenoussinus.But,evenwhenthepressuredifferenceisreversednobackflowofvenousbloodoccursthrougharachnoidgranulationvalvesintothesubarachnoidcavitybecauseofthestructureofthevalves.
CEREBRALBLOODSUPPLY
ARTERIALAlthoughthebloodbrainbarrierlimitsflowofmanymaterialsfrombloodintothe
brain, a steady cerebral circulation to deliver glucose, oxygen and a small set of other

moleculesiscriticalforbraintissuesurvival.Neuronsrequireaconstantsourceofglucoseforenergyproduction.Structures insideneuronsnamedmitochondria synthesizemobilehigh-energymoleculesofadenosinetriphosphate(ATP)fromglucose.TheydependuponasteadysupplyofoxygentosupporttheirproductionofATP.Interruptionofoxygenflowto brain tissue, and therefore synthesis of ATP, for as little as four minutes can causepermanentneurondamage.
Twomajorsetsofarteriesontherightandleftsideofthebodyprovidebloodthatis rich in oxygen and glucose to the brain. They are the internal branch of the carotidarteriesoftheneckandthevertebralarteries(Figure3-7).
The external branch of carotid arteries supplies blood to the face. The internalcarotidarteriesperfusethefrontandmiddleportionsofthebrainandthevertebralarteriesperfuseitsbackportion.Thebrain’sarteriesspreadoveritssurfacewithinthemeningesinthesubarachnoidspacebeforetheypenetratedeepintobraintissue.
The internal carotid arteries run deep in neck tissue to the right and left of thetrachea.Theyentertheskullthroughthecarotidcanalsoftheskull’stemporalbone.Theretheybranchintotheanteriorcerebralarteriesandthemiddlecerebralarteries.

Figure3-7:Arteriestothebrainshownontherightsideofthehead.ThisillustrationisareproductionofalithographplatefromGray’sAnatomy,publishedin1918.ThisworkisinthepublicdomainintheUnitedStates.
The vertebral arteries are smaller than the internal carotid arteries. The pair ofvertebralarteriesbranchfromthelargearteriessupplyingtheshoulders,lateralchestandarms.TheyrunthroughthelateralholesinthetransverseprocessofthecervicalvertebraeC6 toC1 (Figure3-7). They then travel across the cervical vertebraeC1 and enter thebrain through the foramenmagnum in the base of the skull.Within the skull they fusetogethertomakethebasilararterythatsuppliesbloodtothemidbrain.Thebasilararterybranchesfurthertotraversetheposteriorpartofthebrain.
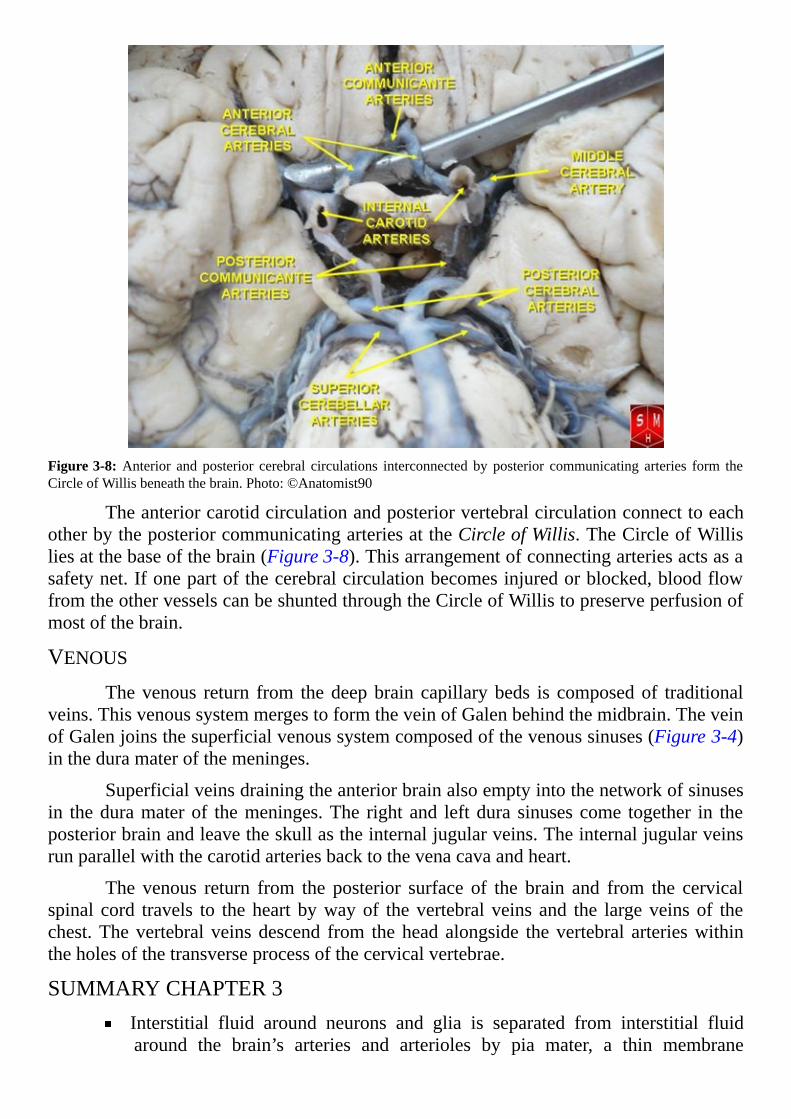
Figure3-8:Anterior and posterior cerebral circulations interconnected by posterior communicating arteries form theCircleofWillisbeneaththebrain.Photo:©Anatomist90
TheanteriorcarotidcirculationandposteriorvertebralcirculationconnecttoeachotherbytheposteriorcommunicatingarteriesattheCircleofWillis.TheCircleofWillisliesatthebaseofthebrain(Figure3-8).Thisarrangementofconnectingarteriesactsasasafetynet.Ifonepartofthecerebralcirculationbecomesinjuredorblocked,bloodflowfromtheothervesselscanbeshuntedthroughtheCircleofWillistopreserveperfusionofmostofthebrain.
VENOUSThevenous return from thedeepbrain capillarybeds is composedof traditional
veins.ThisvenoussystemmergestoformtheveinofGalenbehindthemidbrain.TheveinofGalenjoinsthesuperficialvenoussystemcomposedofthevenoussinuses(Figure3-4)intheduramaterofthemeninges.
Superficialveinsdrainingtheanteriorbrainalsoemptyintothenetworkofsinusesin theduramaterof themeninges.The rightand leftdura sinusescome together in theposteriorbrainandleavetheskullastheinternaljugularveins.Theinternaljugularveinsrunparallelwiththecarotidarteriesbacktothevenacavaandheart.
The venous return from the posterior surface of the brain and from the cervicalspinal cord travels to theheart bywayof thevertebral veins and the largeveinsof thechest.Thevertebral veinsdescend from thehead alongside thevertebral arterieswithintheholesofthetransverseprocessofthecervicalvertebrae.
SUMMARYCHAPTER3Interstitial fluid around neurons and glia is separated from interstitial fluidaround the brain’s arteries and arterioles by pia mater, a thin membrane

creatingtheVirchow-Robinspace
Virchow-Robinspaceassistsinblockingaccesstothebrainoflymphocytesandproteinsofthebody’simmunesystem
The blood brain barrier prevents direct exchange of water and most smallmoleculesbetweenbraincapillariesandinterstitialfluid
The blood brain barrier is created by tight connections between capillaryendothelialcellsandissupportedbyastrocytemembranousextensions
Themeningesarelayersofmembranescoveringtheentiresurfaceofthebrainandspinalcord
Cerebrospinalfluidissecretedbyependymal-likecellsofthechoroidplexuseslocatedineachofthebrain’sfourventricles
Cerebrospinalfluiddeliversnutrients,growthfactorsandwaterfrombloodtotheinterstitialfluidofthebrainandremovescellularwastematerial
Cerebrospinalfluidflowsfromthelateralventriclesintothethirdventricleandthenintothefourthventricle
Cerebrospinalfluidflowsfromthefourthventricleintothecentralcanalofthespinalcordandintothesubarachnoidcavityofthemeninges
In the subarachnoid cavity, cerebrospinal fluid flows to the top of the headwhereitpassesthroughone-wayvalvesintothelargevenoussinuses
Acontinuousflowofbloodtothebrainfordeliveryofglucoseandoxygenisessential
Majorarteriessupplyingoxygenatedbloodtothebrainaretheinternalbranchofthecarotidarteriesandthevertebralarterieslocatedoneachsideofthebody
Theinternalcarotidsperfusetheanteriorandmiddleportionsofthebrainandthevertebralarteriesperfuseitsposteriorportion
Veinsdrainingbloodfromtheanteriorbrainemptyintothenetworkofsinusesintheduramaterofthemeninges
Thedurasinusesmergeintheposteriorbrainandleavetheskullastheinternaljugularveins
Thevenousreturnfromtheposteriorsurfaceofthebrainandfromthecervicalspinalcordtravelstotheheartbywayofthevertebralveinsandthelargeveinsofthechest

[4]
Neurons—HowTheyMakeElectricityTHE LARGER, VISIBLE ASPECTS of brain anatomy and physiology are
described in previous chapters.Here the narrative shifts to the realmof the less visibleaspectsofthebrain,itselectricalcurrents.
Neurons,theprincipalelectricalcellsandtheunmistakablesuperstarsofthebraincell community, occupy the central hub of almost all efforts to clarify how the brainworks. These remarkable cells form the physical substance of themind and a person’ssenseofself.
Brain’sformofelectricityisprobablythemostdifficultconceptforpeoplenewtoscience to get their mind around. It is not the same as the electricity encountered ineverydaylife,anditishumantodoubtideasthatareunexpectedandwithoutprecedent.
Yet, theseeminglybizarreexplanationofneuronelectricitypresentedhere is thenet result of over 40 years of scientific experimentation. And, the report is not yetcomplete. Neuroscientists continue to fine-tune their understanding of how neuronscommunicate.
Becauseneuroncommunicationiscomplex,thischapterreviewsonlytheelectricalpropertiesofneuronaxons.Thenextchapterconsiderssynapses,theinterfaceneuronsusetocommunicatewitheachotherandtheeffectofsynapseactivityondendritesandeventswithinthebodyofaneuron.
NEURONCOMPARTMENTS
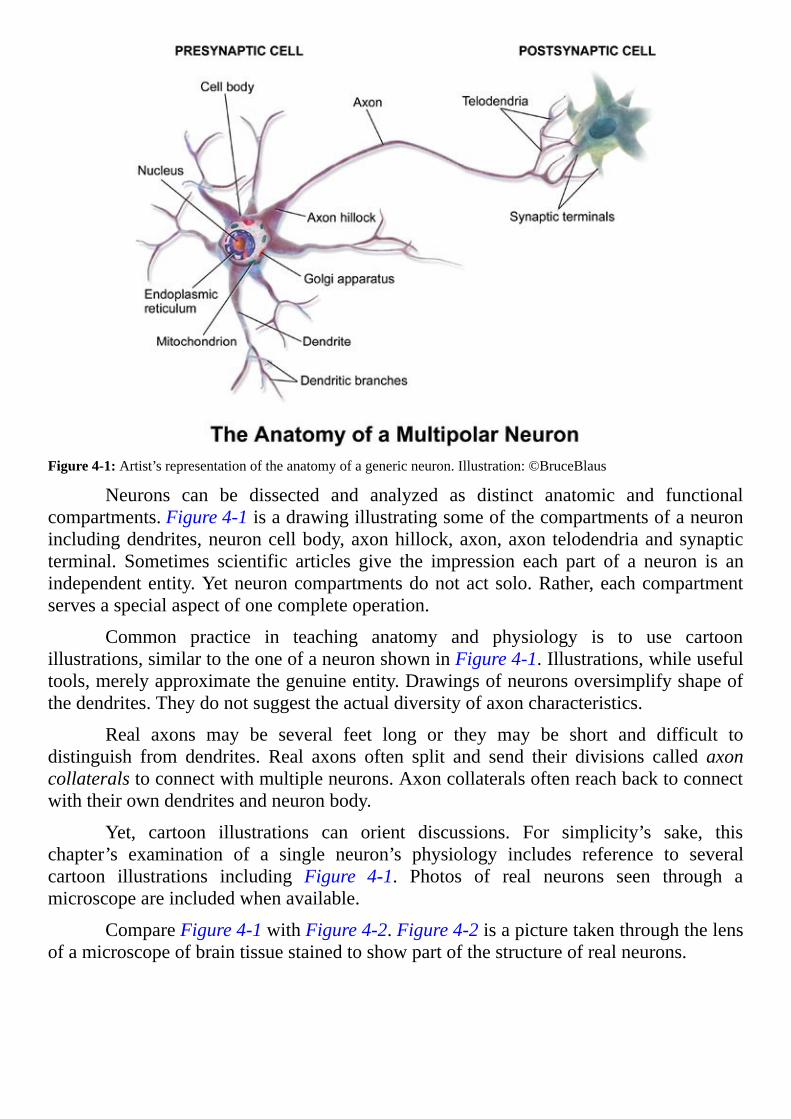
Figure4-1:Artist’srepresentationoftheanatomyofagenericneuron.Illustration:©BruceBlaus
Neurons can be dissected and analyzed as distinct anatomic and functionalcompartments.Figure4-1isadrawingillustratingsomeofthecompartmentsofaneuronincludingdendrites,neuroncellbody,axonhillock,axon,axon telodendriaandsynapticterminal. Sometimes scientific articles give the impression each part of a neuron is anindependententity.Yetneuroncompartmentsdonotactsolo.Rather,eachcompartmentservesaspecialaspectofonecompleteoperation.
Common practice in teaching anatomy and physiology is to use cartoonillustrations,similartotheoneofaneuronshowninFigure4-1.Illustrations,whileusefultools,merelyapproximatethegenuineentity.Drawingsofneuronsoversimplifyshapeofthedendrites.Theydonotsuggesttheactualdiversityofaxoncharacteristics.
Real axons may be several feet long or they may be short and difficult todistinguish from dendrites. Real axons often split and send their divisions called axoncollateralstoconnectwithmultipleneurons.Axoncollateralsoftenreachbacktoconnectwiththeirowndendritesandneuronbody.
Yet, cartoon illustrations can orient discussions. For simplicity’s sake, thischapter’s examination of a single neuron’s physiology includes reference to severalcartoon illustrations including Figure 4-1. Photos of real neurons seen through amicroscopeareincludedwhenavailable.
CompareFigure4-1withFigure4-2.Figure4-2isapicturetakenthroughthelensofamicroscopeofbraintissuestainedtoshowpartofthestructureofrealneurons.

Figure 4-2: Image of neurons in the human hippocampus stained with the Golgi method. 40X magnification.Photomicrograph:©MethoxyRoxy
Notice how the dendritic extensions of the neuron bodies pack tight togetherwithin the interstitial fluid. Dark round and oval spots in the right half of the imagedelineate neuron bodies. Dendrites appear as long hair-like structures attached to theneuronbodies.
Thehumanbrain tissuesectionshowninFigure4-2wasstainedusing theGolgisilverstainmethod.Golgisilverstaininglabels1%-3%ofneuronspresentinanysection,butevensothespaceappearscrowded.Inreality,35-100timesmoreneuronswereinthistissuesectionthanabsorbedthesilver.
Thegeometryoftheinterstitialfluid-filledspacearoundbrainneuronsmodeledbyengineers isdescribedasanetworkofporesand tunnels less than100nanometers(10-7meter)across.Withanestimated86billionneuronsinthehumanbrainandtentimesthatmany glia plus 15-25 square meters of blood and lymph vessels, space available tointerstitial fluid tunnels is limited. Tight packing creates a setting where chemicalcommunicationbetweenneuronsandtheinterstitialfluidoccursacrossspacesjustafewnanometerswide.Ananometerisonepartofameterthatisdividedintoonebillionequalparts.
BRAIN’SELECTRICITY
HOWNEURONSCONNECT

Figure4-3:Howneuronsconnectwitheachother.Illustration:ThisworkisinthepublicdomaincourtesyoftheUnitedStatesNationalInstitutesofHealth.
Deciding where to start when describing how neurons communicate with eachother is a challenge, because it is a circular narrative.The receiving endof oneneuronrespondstoinputfromthemessagingendofanotherneuron.Then,inturn,thereceivingneuronsendsamessageabout thesignal it receivedas input to thereceivingendof thenextneuron(Figure4-3).Thedilemma is to decidewhere in the neuron’s structure thestoryofitselectricalmembraneshouldbegin.
AtthetopofFigure4-3,noticewheretheaxonterminalsofaneuronoutsidetheframeof thepicturemakeconnectionson thedendritesand thebodyofa largeneuron.Electric signals shown as jagged arrows proceed along the surface of the receivingneuron’sbodyanddown its axon.The axon terminalsof the largeneuron then connectwiththebodyanddendritesofthenextneuroninsequence.
Theinsertinthelowerrightcorneroftheillustrationisamagnificationoftheaxonterminal’s connection, a synapse. Notice the enlarged section presents the neurons asseparatecells.Anarrowspaceexistsbetweentheaxonterminalandthenextneuron.Thisconfiguration of neurons connecting in sequence is the basic component of the brain’sneuronnetworks.

ELECTRICALCURRENTINTHEBRAINThischapterdescribescharacteristicsofneuronaxons (Figure4-1).Onoccasion
axonsarecomparedtoelectricalwires,becausetheaxon’sjobistotransmitanelectricalsignal from the body of the neuron to the axon terminal. However, the electrical wiremetaphoristoosimpleandmaybeconfusingratherthanhelpful.Inreality,whileelectriccurrents spread the length of axons, the process is complex and unlike the flow ofelectronsthroughacopperwire.
Electrical current in the brain consists of a stream of atoms called ions. Ionspossesseitherapositiveoranegativecharge.Thequalityandquantityofanion’schargeis governed by the atom’s lack of a match between its number of protons, positiveparticles,andelectrons,negativeparticles.
Whentheconcentrationofaparticularionisnotthesameintheinterstitialfluidasin the neuron’s cytoplasm, the ion can use open passages in the neuron’smembrane tomove toward the fluid compartmentwhere its concentration is least.This is a chemicalprocessknownasdiffusion.Diffusion iswhenmolecules relocate bymoving through asolution from the place where their concentration is high to another place where theirconcentrationislower.
Theimportantionsforneuronsignalingincludesodiumions(Na+),potassiumions(K+),chlorideions(Cl-)andcalciumions(Ca++).Theconcentrationdifferencesoftheseions between neuron cytoplasm and interstitial fluid is sufficient to draw them throughopen tunnel-shaped proteins described as ion channels in the neuron membrane. Ionchannelproteinsaccommodatepassageofspecificions.Mostionchannelsopenandcloseinresponsetoparticularsignals.Intheabsenceofspecificsignals,membraneionchannelsremainclosed,ionscannotdiffuseacrossthemembraneandthereisnoelectricalcurrent.
Thepreviouschapterdescribedwholebrainmechanismsforkeepingthequalityofbrain’sinterstitialfluidsteady.Inadditiontostrategiesforinterstitialfluidqualitycontrol,all cells including neurons use ion exchange pumps to protect their cytoplasm’s ionconcentration(Figure4-4).

Figure4-4:Energyrequiringpumpsmaintaintheioniccompositionofneuroncytoplasm.Illustration:©AlilaMedicalImages
The pumps aremembrane proteins that compensate for ions relocating betweencytoplasm and interstitial fluidwhen ion channels are open. Pumps,moving ions fromwheretheirconcentrationislowtowheretheirconcentrationishigh,requireenergyfromthemolecular bonds of adenosine triphosphate,ATP.A great deal of the brain’s energysupplyisusedtopowerneuronmembraneionpumps.
DIRECTIONOFNEURONIONFLOW
WhenK+ channels open,K+ diffusesOUT of a neuron’s cytoplasm.WhenNa+
channels open, Na+ diffuses INTO a neuron’s cytoplasm. Diffusion caused by ionconcentrationdifferencesonoppositesidesofamembraneisdescribedinphysiologyasions moving down their concentration gradient. And, these two ions move down theirconcentration gradient across neuronmembranes creating an opposite flow of positive,chemical-basedelectricalcurrent.
The opposite flow of Na+ and K+ across open channels in neuron membraneappearsonthesurfacetobeasimpleconcept.Buttheoutcomeofthissimpleprocessisquitepowerfulinthenervoussystem.Itprovidesthefoundationuponwhichinformationiscarriedbyneuronalcircuits.
Thecytoplasmofneurons,andofmostcells,possessesahighconcentrationofK+
and of large, negative charged soluble proteins. Nomembrane channels exist for largemoleculessosolubleproteinsremaininthecytoplasm.Theybalancethepositivechargeof the high amount of K+ within cytoplasm and maintain the cytoplasm’s electricalneutrality.

Incontrast toK+, bothNa+ andCl- are kept low in cytoplasmof adult neurons.Ca++ is highwithin neurons but stays confined in compartmentswithin the cytoplasm.Ca++isnotfreetodiffusewithincytoplasmexceptinresponsetospecifictriggers.Unlikecytoplasm,interstitialfluidsurroundingadultneuronsislowinK+buthighinCa++,Na+
andCl-.Whenopenchannelsareavailableallthreeions,Ca++,Na+andCl-movedowntheirconcentrationgradientintoneuroncytoplasmcreatingelectricalcurrent.
Whenaneuronisatrest,mostionchannelsintheaxonmembraneremainclosedandlittleioniccurrentcanflow.Avarietyofcircumstancescausemembraneionchannelsof different cell types to open. Each ion channel responds to its own specific openingtrigger. Ion channels in a neuron’s axon membrane for Na+ and K+ open whencharacteristics of the membrane surrounding them alters in the manner discussed infollowingsections.
NEURONSATREST
TRANSMEMBRANEPOTENTIALSAll cellmembranes have an electrical potential on their inside and their outside
surface.Anelectricalpotentialisanenergycreatedbythepresenceofelectricalcharges.When cells are inactive, ions of the cytoplasm and ions of the interstitial fluid createelectricalenergyofdissimilarmagnitudeonthetwosidesofthemembrane.Acomparisonoftheelectricalpotentialoftheinteriorsideofacell’smembranetotheelectricalpotentialof the exterior side produces a measurable inequality described as a transmembranepotential.
Conveniently, the definition of voltage is a difference in electrical potentialbetweentwopoints.Whenamembrane’spotentialisdifferentonitstwosides,thereisavoltageacrossthemembrane.Thisallowstheelectricalforcedifferencebetweenthetwosides of a cell’s membrane to be expressed as a numerical value. Transmembranepotentialsofneuronmembranesquantifyinmillivolts,onethousandthsofavolt.
Voltage is always a relative term.By convention in physiology the quantity andquality,positiveornegative,of transmembranevoltage isalwaysstatedas theelectricalpotentialoftheinsidemembranesurfacerelativetotheelectricalpotentialoftheoutsidemembrane surface. Therefore, a transmembrane potential of -70 millivolts means theinsidesurfaceofthemembranepossessesanelectricalpotential70millivoltslessthantheelectricalpotentialoftheoutsidesurface.
Thetransmembranerestingpotentialisthemembranepotentialofneuronsthatarenottransmittingsignals.Thetransmembranerestingpotentialofneuronsmostoftenfallsintherangeof-60millivoltsto-90millivolts.
ORIGINOFTRANSMEMBRANEPOTENTIALSTransmembrane potentials exist because a small number of each cell’s ion
channelsforNa+andK+alwaysremainopenallowingaslowconstantdiffusionoftheseions. Ion channels remaining permanently open are referred to as leaky channels orpassivechannels.

Passiveionchannelscreateatransmembranepotential.Theydifferfromthelargenumber of Na+ and K+ voltage-sensitive channels used by neurons to send a signal.Opening and closing of voltage-sensitive neuron channels is triggered by transientfluctuationsofthetransmembranepotentialandwillbedescribedinthenextsection.
In neuron membrane, passive channels for K+ outnumber, by far, the passivechannels for Na+. Because K+ is at a far higher concentration inside the neuron thanoutside,itdiffusesoutoftheneuronthroughitspassivechannels.IncomingNa+throughthefewerpassiveNa+channelscannotmakeupforthepositivechargelostwiththeexitofK+,andtheinsideoftheneuronmembranebecomesrelativelynegative.Themagnitudeofthe negativity of the electrical field on the inside surface of a neuron membrane isdeterminedbytheamountofK+leavingthecell.
Once outside of the neuron, positive K+ is drawn to the outer surface of themembranebytheexcessofnegativechargeleftattheinsidesurface.WhiletheexitofK+
from the neuron through passive channels is driven forward by its high cytoplasmicconcentration,otherforcesstopitsexodus.IntimethebuildupofnegativechargeontheinsideofthemembraneholdstheremainingK+back,andthepositivechargebuiltupontheoutsideofthemembranebytheearlierexitofK+repelsit.BythetimeforcesonK+
balance,anegativetransmembranerestingpotentialisestablished.
Little exit of K+ is needed to create a transmembrane potential. A theoreticaltransmembrane potential can be calculated for cells with passive K+ channels but nopassiveNa+channels.Thistheoreticaltransmembranepotentialisnamedtheequilibriumpotential for potassium. For neurons, because of the number of passive K+ channelspresent,theequilibriumpotentialforpotassiumisabout-92millivolts.
The theoretical transmembrane potential when a cell membrane possesses onlypassiveNa+channelsistheequilibriumpotentialforsodium.Forneuronstheequilibriumpotentialforsodiumisabout+50millivolts.Theinsideofthemembraneachievesgreaterpositivitythantheoutsideofthemembraneby50millivolts.
Actual neuron transmembrane resting potentials measure slightly more positivethan the -92 millivolts equilibrium potential for potassium. This is because of thecontributionofthesmallnumberofpassiveNa+channelsinneuronmembranes.
The final component in the establishment of a stable transmembrane restingpotential is thepresenceofprotein ionexchangepumps illustratedabove inFigure4-4.Without these energy requiring pumps the passive channels would dissipate ionconcentrationgradientsbetweenthecytoplasmandinterstitialfluid.SeveralkindsofionexchangepumpsexistinthemembranesofbraincellsinadditiontothoseforNa+andK+.
Ionexchangepumpsandionchannelsrepresentdifferentclassesofcellmembraneproteinswithseparatespheresofoperation.Theyshouldnotbeconfusedwitheachother.Ion exchange pumps move ions from one fluid compartment to another against theirconcentration gradient. Ion channels establish open passages in themembrane allowingionstodiffusefromthefluidcompartmentwheretheirconcentrationishightothefluidcompartmentwheretheirconcentrationislow.

VOLTAGE-SENSITIVEIONCHANNELS
IONCHANNELSTRUCTURECharacteristics of proteinmoleculesmake themexcellent ion channel structures.
Proteinsconsistofassembliesof20uniquemolecularunitsnamedaminoacids.Individualamino acids are fat-soluble or water-soluble. Some are large molecules and some aresmall.Someaminoacidscarryacharge,andsomeremainneutral.
Tobuildaprotein,aminoacidsconnecttogetherinanarrangementdescribedasapeptidebond.Theaminoacids linkoneafter theother toformachainnamedapeptidebecause of nature of the bonds. Some protein chains become long, greater than 2000aminoacids,andsomeremainshort,10to200aminoacids.
Proteinchainsfoldoveronthemselvesbecauseofthedifferentsizeandchargeofindividualaminoacids.Eachaminoacidsearchesforspaceandacomfortableelectricalfieldenvironment.Theportionswithpositivechargerepeleachother.Theportionswithnegativechargerepeleachother,andtheoppositechargedportionsgather together.Oneconfiguration,favoredbysomesectionsofproteinsastheyfold,isaspiralshapenamedanalphahelix,aright-handedcoil(Figure4-5).
Figure 4-5: Section of a protein showing open-loop peptide backbone structures on both endswith an alpha helicalpeptidebackbonestructurebetween.The linedrawingsattached to thebackbonestructures representaminoacid sidechains.Aminoacidsidechainsdistinguishoneaminoacidfromanother.Illustration:©molekuul.be
If thesequenceofaminoacidsinahelixisarrangedwhereall theneutralaminoacid side chains occur on one side of the helix and the charged amino acid side chainsoccupytheoppositeside,thehelixisabletoinsertitselfinacell’slipidmembranewiththe neutral amino acid side chains becomingmembrane anchors. The part of the helixawayfromthemembraneisabletoretainelectricalpropertiesattractivetowaterandions.Proteinsectionspassingthroughcellmembranesalwayshaveanalphahelicalform.
Another feature of proteins, which makes them exceptional molecules forconstructingionchannels,istheirabilitytoadjusttheirshapewhenothermoleculesentertheirelectricalfield.Whenthetransmembranerestingpotentialofaneurondeviatesfrom

itsnormalvalue,voltage-sensitiveproteinchannelsrespondbychangingtheirshape.Theresultingchangeinthepositionoftheaminoacidsidechainsopensatunnelthroughthemembrane.
Voltage-sensitivechannelproteinsforNa+andforK+inneuronaxonmembranesappearsimilartoeachotherinstructure.Thebasicorganizationofavoltage-sensitiveionchannelconsistsoffourdomainseachwithsixtransmembranealphahelices.
Thevoltage-sensitiveK+ionchannel(Figure4-6)isacompositeoffourseparateproteins, each with a configuration of a single domain with six transmembrane alphahelices. Incontrast the fourdomainarrangement is shownspreadout asa singleaminoacidchainforthevoltage-sensitiveNa+channelinFigure4-7.
EXCLUSIVITYOFK+CHANNELSMembrane ion channels allow exclusive passage of particular ions. How ion
channel selectivity is accomplished remained amystery until recent data uncovered theoriginoftheuniquenessoftheionchannelsforK+andNa+.ThesedatasolvethepuzzleofwhyNa+,asmallerion,cannotpassthroughionchannelssimilarinstructuredesignedforthelargerK+.TheK+channeldiagramedinFigure4-6presentsanexcellentexampleofhowaproteinchannelisorganizedtofavoraparticularion.
Figure4-6:Viewofpotassium(K+)inthefilterofaK+channel.Thisviewisfrominsidethecell.K+isthepurpledotat thecenterof the structure.The reddots representoxygenatoms;graydotsdenotecarbonatoms;bluedots signifyhydrogenatoms.Noticethesymmetricalplacementoffouridenticalsubunits.Illustration:BaseduponPublicDataBank1BL8isreleasedtopublicdomainbyBensaccountatWikimediaproject.
When ions likeNa+ andK+ disperse in water they become surrounded by, andbound to, water molecules by a type of weak chemical bond described as a hydrogen

bond. In order for ions to pass through theK+ channel’s filter, the sphere of hydrogenbondedwatermoleculessurroundingK+mustbestrippedaway.Strippingoffmoleculesofwaterrequiresahydratediontoencounterwithintheionchannelotheratomscapableofdisplacingthewatermolecules.
Structuralanalysisofvoltage-sensitiveK+channels,similartothosefoundinthehumanbrain,revealedapassagewithanarrowfilterareacreatedbycarbonyloxygeninfour critical positions.Carbonyl oxygen is an oxygen atomwith two strong bonds to acarbonatom.Carbonyloxygenisacomponentofthepeptidebondlinkingaminoacidstoeachother.
In this particular channel, the four carbonyl oxygen components localize to theexact positions necessary to drive water away from around the K+. Yet, because theoxygenatomofcarbonyloxygenisbondedstronglytocarbon,itsbindingtoK+isweakenoughtoallowK+tocontinuethroughthechannel.
TheprecisemovementofwaterandK+throughthevoltage-sensitiveK+channelisstillunderactive investigation.Acontemporary theoryhypothesizesonewatermoleculefrom each hydration sphere also passes through the channel’s filter. The sequence ofmolecularmovement through the selectivity filter is thought tobealternatingpotassiumionandwatermolecule,K+-H2O-K+-H2O-K+-H2O.
ThesameprocessdoesnotworkforNa+attheK+channel,becauseNa+issmallerthanK+. The four carbonyl oxygen components forming the selectivity filter of theK+
channel are spaced too far apart to rid the smaller Na+ of its hydrogen bonded watermolecules.Withitswaterhydrationsphereleftintact,Na+istoolargetopassthroughtheK+channel.
SELECTIVENESSOFNA+CHANNELS
Voltage-sensitiveNa+channelslocatedinneuronaxonsconsistofonelargealphaproteinassociatedwithtwoorthreesmallerbetaproteins.ThealphaproteincontainsthechannelwithaselectivefilterforNa+.Thebetaproteinsalterthecellularlocationofthealphachannelandalteritsvoltagesensitivity.
Figure4-7showsthealphasubunitofaNa+channelspreadoutinamembranetoillustrateitindetail.

Figure4-7:Sectionsof thealphasubunitofavoltage-sensitiveNa+channelspreadout inamembrane(whiteband).Notice the fourdomains, I, II, IIIandIV,eachwith6 transmembranealphahelices through themembrane.G=siteswheresugarscanbeattached,P=aminoacidswherephosphategroupscanbeattached,Slabelsthelocationoftheionchannelfiltercomponents,I=aminoacidsaffectedwhenthechannelclosesafterbeingopen.Illustration:©Cthuljew
In the axon membrane, the four domains labeled I-IV form a circular clusterresembling the four subunitsof theK+ channel shown inFigure4-6.When thedomainscluster,theproteinsectionsillustratedasloopsoutsidethecellcloseovertheareaslabeledSinthediagram.TheseloopscreatetheNa+channelfilter.Theremainderofthechannelinthemembraneisformedbythefoursetsofhelicesnumbered5and6intheillustration.Theareabetweenhelices5and6becomestheopenchannel linkingthe interstitialfluidwiththeneuron’scytoplasm.
Eventhoughtheoverallorganizationofthealphasubunitofthevoltage-sensitiveNa+channelissimilartothevoltage-sensitiveK+channel,thecontourofitsionselectivityfilterisquitedifferent.TheentranceoftheNa+channelfilterislargerthantheentranceoftheK+channelfilter.TheNa+channelfilterisformedbyfournegativechargedglutamateaminoacids.TheglutamatesidechainsstripmostbutnotallofthewatermoleculesfromNa+.Small,partlyhydratedNa+moves throughits ionchannelfilterwith theremainingwatermoleculesformingabridgetothefilterwalls.HydratedK+istoolargeforthesamebridgingeffectwiththefilterwalltooccur.
AXONSIGNALING
ARRANGEMENTOFVOLTAGE-SENSITIVECHANNELS
Thepositionofthevoltage-sensitiveNa+andK+channelsalongthelengthoftheaxoniscritical tofidelityofelectricalconductionalongneuronaxons.Thewell-orderedclustersofvoltage-sensitiveNa+andK+channels,andtheirsequentialpatternofopeningand closing, transmit chemical electricity from the axon’s initiation site near the axonhillockoftheneuronbodytotheterminalendoftheaxon.
TopaintaclearpictureofhowclustersofNa+andK+voltage-sensitivechannelsdistribute along axons another type of brain cell, the oligodendrocytes, must beintroduced. Oligodendrocytes possess a cell body central to many long membraneextensions (Figure 4-8). Oligodendrocytes use their slender extensions to hang on toaxons.

Figure4-8:AmouseoligodendrocytelabeledwithGFP(GreenFluorescentProtein).Theimagewasrecordedusingafluorescentmicroscopewitha63xobjective.Photomicrograph:©JurjenBroeke
Where oligodendrocyte extensions contact neuron axons they convert to flatsheets.Theflattenedsheetofmembranewindsaroundneuronaxonscreatingacoverwithmanylayers.Thelayeredportionoftheoligodendrocytemembranecompactsandpushesoutmostofitsowncytoplasmshrinkingthewraptotightlayersofmembrane.
Themembraneofoligodendrocyteextensionscontainsahighlevelofavarietyoffat molecules. The entire group of fat molecules is described collectively as myelin.Myelinactsasanaxon’selectrical insulation.Myelinalsoprotects fragileaxonsagainstinflammatoryandoxidativeinjury.Asingleoligodendrocytemayencloseasingleaxonoritmayreachoutandenclosetheaxonsofmanyneuronsatonce.
For simplicity, illustrations showing myelination of neurons leave outoligodendrocyte cell bodies and picture just the covers made by individual membraneextensionsasshowninorangeinFigure4-9.Thelayerofmyelinisnotcontinuousalongtheaxon.Betweenthemyelinlayersbarepatchesofaxonoccur.Thebarepatches,knownasNodesofRanvier,arenamedforthemanwhodiscoveredthem,FrenchpathologistandanatomistLouisAntoineRanvier.

Figure 4-9: Drawing of a neuron showingmyelin sheaths colored orange. The oligodendrocytes cell bodies are notpresent.Illustration:©martan
Dense clusters of voltage-sensitiveNa+ channels occur at theNodes ofRanvier.The assembly ofNa+ channels at the Nodes of Ranvier includes secretion of adhesiveproteins by the oligodendrocytes. Clusters of the channels form around the adhesiveproteins. Fibrous protein components within the axon, collectively described as itscytoskeleton,alsoplayaroleinanchoringNa+channelsinplace.
In contrast to the position of the voltage-sensitive Na+ channels on the axon,voltage-sensitiveK+ channels gather together under the edges of themyelinwrap. TheedgesoftheoligodendrocytewrapmergewiththeaxonmembranetoformathinpartitionseparatingNa+andK+channelclusters.AxonalK+ channels residenear,but inside, thepartitionsunderthemyelin.
Axonmyelination is an ongoing process throughout brain development and intoadulthood.Thefrontal lobeof thecerebralcortex, theareaassociatedwithlearningnewconceptsandmemory,isthelastbrainregiontoaddmyelintoitsaxons.Itisalsotheareainadultswiththemaximumamountofaxonmyelination.
Thedensityofaxonswithmyelinintheadulthumanbrainvariestoagreatextentamong regions. Many axons in the brain lack oligodendrocytes. Axons withoutmyelination also display voltage-sensitive Na+ and voltage-sensitive K+ channels inalternatingclustersspreadalongtheaxonalmembranebutthespeedofsignaltransmissionis comparatively slow. Axons with myelin conduct signals about 50 times faster thanaxonswithoutmyelin.

Oligodendrocytesmayinfluencethedesignandfunctionofbraincircuitry.Patchesof myelin on living axons appear in an irregular pattern, unlike myelin illustrated inFigure 4-9. Some neurons in the cerebral cortex maintain extensive stretches of axonwithoutmyelin,followedbystretcheswithmyelin.Manytheoriesexistaboutwhythisis.OnespeculationislargerNodesofRanvierallowneighboringneuronstosynapseonthebarepartsofanaxon.Anotherguessismyelinsheathsproducechemicalstoblockaxoncollateral formation. Axons split and send collaterals off to synapse onmultiple targetcells,butaxoncollateralsonlyformwhereanaxonisbare.Bothsynapsesonaxonsandaxoncollateralsparticipateascentralelementsofcomplexbraincircuits.
TRANSIENTMEMBRANEVOLTAGEPATTERNS
ThesignaltravelingalonganaxoniselectricalbecauseionsflowingthroughNa+
andK+ voltage-sensitivemembrane channels carry a charge.Moving a signal along anaxon requires setting in motion a pattern of voltage transients, short-lived and rapidfluctuations in the transmembrane potential called action potentials (Figure 4-10).Consecutive action potentials progress the length of an axon producing a signal thatmaintainsitsstrengthasittravelsfromitsinitiationsitetotheaxon’sterminalend.
Voltage-sensitiveNa+andK+channel filters remainclosed to flowof ionsat theaxon’snegativetransmembranerestingpotential.Whenatransmembranepotentialmovesaway from its negative resting value to a sufficiently less negative voltage, a thresholdpotential,voltage-sensitiveionchannelfiltersopenforashortperiodoftime.
Apositivedriftofthetransmembranerestingpotentialattheinitialsegmentoftheaxon is the result of events within the neuron body at the axon hillock (Figure 4-9).Diffusion of Na+ from the axon hillock depolarizes the initial segment of an axonmembranetoitsthresholdpotential.
Depolarizemeanstobringelectricalpotentialsontwosidesofamembranecloserto being equal. The threshold potential causes the first cluster of the axon’s voltage-sensitiveNa+channelstoopen(Figure4-10).

Figure 4-10: Transmembrane voltage transients during an action potential. Notice the time between the membranereaching threshold and thepeakchange in transmembranevoltage is less thanone thousandthof a second (ms).Themagnitude of the threshold potential needed for initiation of an action potential varies between neurons. Illustration:©Synaptidude
The axon hillock is also able to originate action potentials, but its thresholdpotentialismorepositivethantheaxon’sthresholdpotential.Membranedepolarizationatthe axon hillock is a result of action potential–like activity named dendritic spikesspreadingoverthemembraneoftheneuronbodyfromthedendrites.Dendriticspikes,theresult of signaling at neuron synapses, are explained further in Chapter 5, “NeuronSynapses—ExcitatoryandInhibitory.”
The electrical field adjustment associated with depolarization of thetransmembrane potential changes the alignment of amino acid side chains in the Na+
channelprotein.RepositioningofaminoacidsidechainsopenstheNa+channel’sionfilterand permits passage of Na+ across the membrane into the axon’s initial segment. Na+diffusingthroughopenchannelfilters,inturn,movesthetransmembranepotentialnearertozeromillivolts.ArepresentativeactionpotentialisdiagramedinFigure4-10.
SPREADOFACTIONPOTENTIALSThe transmembrane threshold potential necessary to open an axon’s voltage-
sensitiveNa+channels isreachedbeforethethresholdpotentialrequiredforopeningtheK+channels.Thatis,amembranedepolarizationofgreatermagnitudeisrequiredtoopenvoltage-sensitiveK+ channels. Because of this difference in response to changes in thetransmembranepotential,axonNa+channelsalwaysopenbeforeK+channels.
Open voltage-sensitiveNa+ channels allowNa+ to diffuse into the axon until itapproaches the equilibrium potential for sodium—the potential where the positiverepellingenvironmentwithintheaxoncytoplasmovercomestheforceoftheconcentrationgradientdrivingNa+intothecell.Thepeaktransmembranepotentialofanactionpotentialisabit less than theactualNa+ equilibriumpotential for tworeasons.First thevoltage-

sensitive K+ channels begin to open before the potential peaks allowing K+ to carrypositivechargeoutoftheaxon.
Second,theentireclusterofNa+channelsisnotopenatthesametime.Beforethepeak of the action potential, part of the group ofNa+ channels begins a gradual close.ReferbacktoFigure4-7andnoticetheNa+channelfilterislocatedatadifferentplaceonthe alpha channel protein than the inactivation sites that cause it to close. Thisarrangement of the Na+ channel’s voltage-sensitive amino acids makes opening andclosingtheNa+channelseparateprocesses.Oncethechannelfiltercloses,aperiodoftimemustpassbeforetheproteinrearrangesitselfintoapositionwherethefilterareacanagainrespondtoatransmembranepotential.
AsNa+enterstheaxon,itdiffusesawayfromitschannel.Itspreadsovertheinsideofthemembraneinalldirections.SomeoftheNa+travelstowardtheaxonterminalunderthemyelinwrapinthedirectionofthenextNodeofRanvier.WhensufficientNa+reachesthenextNodeofRanvier,itdepolarizesthatsegmentoftheaxontothresholdandelicitsanother action potential. This process repeats itself from node to node until an actionpotentialisproducedattheaxonterminal.
SomeoftheNa+enteringtheaxonalsodiffusesbacktowardtheneuron’sbody.Atthe axonhillock, backdiffusionofNa+ brings the transmembranepotential of the axonhillocktothresholdproducinganactionpotentialthatspreadsalongthemembraneoftheneuronbody.Thebody’sactionpotentialtravelstowardthedendrites.Howfarthisactionpotential moves over the membrane surface and into the dendrites depends upon themembrane’spatternofvoltage-sensitive ionchannels.Thedistributionof thesechannelsvaries from neuron to neuron. The flow of membrane action potentials from the axonhillock toward the dendrites informs the rest of the neuron that an output signal wasgenerated.
Thesequenceofionchanneleventsalonganaxoninsuresaxonalactionpotentialsmove along in one direction, from the initial segment to the axon terminal. Any Na+diffusing in the direction of ion channels at the previous node is unable to bring thetransmembrane potential at the previous node to threshold. This is because the lateropeningK+channelsremainopenthereforK+todiffuseoutoftheaxoncarryingpositivechargewithit.
Not all the ion channels in a cluster open and close in unison. There is a timeduringeachactionpotentialwhenmostoftheNa+andK+channelsareopen.Duringthislullbeforechannelsclose,theaxoncannotproduceanotheractionpotential.Thispauseisnamedtheaxon’sabsoluterefractoryperiod.Anaxon’sabsoluterefractoryperiodlimitsthe transmission frequency of action potentials and helps to force action potentials toprogress toward the axon terminal. An absolute refractory period lasts about onethousandthofasecond.
Someaxonsalsoexhibitarelativerefractoryperiodwhenmost,butnotall,ofthevoltage-sensitive Na+ and K+ channels again respond to depolarization of thetransmembrane potential. During the relative refractory period a greater than normalmembrane depolarization is required to elicit an action potential. This is because the

transmembrane potential immediately following an action potential is closer to thepotassium equilibrium potential than to the normal transmembrane resting potential.Relativerefractoryperiodslastabout2thousandthsofasecond.
AXONHOUSEKEEPINGComponentswithin neuron cytoplasm also transfer along axons.Axon terminals
dependuponthemachineryoftheneuronbodyforsynthesisofstructuralelements.Foranaction potential to create the desired effect at the axon’s terminal end, the terminal endmust possess an adequate stock of proteins, mitochondria, vesicles, nutrients, energymoleculesandothercriticalitems.Everythingrequiredforwell-beingoftheaxonterminalmustbetransportedfromtheneuronbodythroughtheaxon.
Afeedbackmessagingsystem isalsonecessary to regulateappropriate timingoftheflowofmoleculesanditemsfromtheneuronbodytotheaxonterminal.Thismeanssubstancesmust be transported in both directions. Two terms describe the direction offlowofbiologicmaterialinaxons.Itemstransportedanterogrademovetowardtheaxonterminal.Itemstransportedretrogradetraveltowardtheneuronbodyfromthedirectionoftheaxonterminal.
In neurons, as in other cells, shuttlingof cytoplasmic components fromplace toplaceisacontinuousprocessrequiringenergy.Re-arrangementofbiologicmoleculesandstructuresisaccomplishedbymotorproteinsbindingcargoandconveyingitalongfibroustracksdescribedascytoskeleton.
Axonscontainanextensivecytoskeletonalongtheirentireinnerlength.Itprovidesa scaffold for cargo delivery to and from the neuron body. Things travelling inmembranous sacks move with speed along the axon’s cytoskeleton. Enclosed materialincludesamongotherthings,aminoacids,variousproteins,lipidsandsugars.
Motorproteinsbindtospecificmembraneproteinsonthesackseitherdirectlyorwith the help of cytoplasmic linker proteins. Cargo not enclosed inmembranous sacksmoves sluggishly through the length of the axon. The mechanism for movement ofunpackageditemsalongthecytoskeletonalsoincludesmotorproteins,buttheprocessislesscontinuous.Slowmovingitemsprogresspartwayalongtheaxonandthenstopforaperiodof timeand thenmoveonagain.The intermittentnatureof thisprocess suggeststheremaybemultiplemonitoringpointsalongtheway.
SUMMARYCHAPTER4Ions,chargedatoms,carrythechemical-basedelectricalcurrentsofthebrain
Thedifferenceintheconcentrationofionsbetweenneuroncytoplasmandbraininterstitialfluidpermitionstodiffusethroughopenmembranechannels
Sodium ion (Na+), potassium ion (K+), chloride ion (Cl-) and calcium ion(Ca++)createtheelectricalcurrentsinbrain
Neuronmembranesdisplayadifferentelectricalpotentialonthesidebathedbyinterstitialfluidthanonthesidebathedbycytoplasm
Transmembrane voltage,which ismeasured inmillivolts, is always stated as

insideelectricalpotentialrelativetooutsideelectricalpotential
Atransmembranepotentialof-70millivoltsmeanstheinsidesurfaceofthecellmembrane possesses an electrical field force 70 millivolts less than theelectricalfieldforceoftheoutsidesurface
Voltage-sensitive ion channels of the axon membrane occur as large proteincomplexes
Voltage-sensitive channelsopenwhen the transmembranepotential near themreachesathresholdvalue
The transient fluctuation of the transmembrane potential of neuron axons,causedbytheopeningandclosingofvoltage-sensitiveNa+andK+channels,isknownasanactionpotential
Action potentials travel along an axon from the neuron’s body to the axonterminal
Materialsmovebetween theaxon’s terminal and theneuron’sbodyalong theaxon’scytoskeleton

[5]
NeuronSynapses—ExcitatoryandInhibitoryARRIVALOFACTIONPOTENTIALS at axon terminals sets inmotion events
supporting communication between neurons. Transfer of information from neuron toneuron, or from neuron to non-neuron cell, is in most cases a molecular rather thanelectricalphenomenon.
SantiagoRamónyCajal shared the1906NobelPrizewithCamilloGolgi.Theyreceived the prize because of their extensive body of work describing the structure ofbrain neurons. SantiagoRamón yCajal proposed neuronsmay be separate entities thatcommunicatewith each other across narrow spaces of about 20 nm (20 x 10-9meter).Neuroscientists did not embrace this idea at first. However, Dr. Ramón y Cajal wascorrect. The structures at the places where neurons connect with each other are nowcollectively assigned the name synapse (Figure 5-1). Synapses and their surroundingstructuresattractconsiderableongoingattention.
Figure5-1:Somelocationsofneuronsynapsesfoundinthebody.Illustration:©BruceBlas
BRAINSYNAPSESBrain synapses occur at neuron dendrites, at the neuron cell body and at axons.
But,neurons throughout thebody,peripheralneurons,alsoformsynapsesonothercellslikeskeletalmuscleandbloodcapillarysmoothmuscle.
The complex placement of synaptic structures in the brain permits fine-tunedregulatorycontrolofneuroninputandoutputandcreatescomplexneuronalcircuits thatactasunits.
Themost common type of synapse in the brain is onewhere a smallmolecularweight chemical called a neurotransmitter is released from the axon terminal.Neurotransmitter is stored in the axon terminal in vesicles. The section of the axonterminal membrane dedicated to neurotransmitter release is called the presynapticcompartment(Figure5-2)ofthesynapse.
Neurotransmitterreleasedfromthepresynapticcompartmentdiffusesintoagapofabout 20 nanometers between the presynaptic compartment and a modified section ofneuroncellmembraneacrossthegapnamedthepostsynapticdensity(Figure5-1).

The gap itself is called a synaptic cleft. When the postsynaptic cell is a brainneuron,thepostsynapticdensitymaybeonadendrite,onadendriticspine,ontheneuronbody,ontheaxonoronanaxon’sterminal.
Figure5-2:Illustrationofsomeofthekeyelementsfoundatneurontoneuronsynapses.Illustration:©CurtisNeveu
PRESYNAPTICCOMPARTMENT
NEUROTRANSMITTERRELEASE
Neurotransmitter is stored by the presynaptic neuron in vesicles inside axonterminals(Figure5-2).Whenactionpotentialsarriveattheaxonterminals,themembranedepolarizationtheyinitiatecausesvoltage-sensitivecalciumion(Ca++)channelstoopen.Ca++ diffuses through the open channels into the presynaptic compartment from theinterstitial fluid. Ca++ entering the neuron is called free Ca++ to distinguish it fromcytoplasmicCa++boundtocellularproteins.
FreeCa++isathousandtimeshigherintheinterstitialfluidsurroundingadultbrainneurons than in neuron cytoplasm.The introduction of a high level of freeCa++ to thecytoplasm when voltage-sensitive Ca++ channels open sets off a series of molecularevents.These events transport neurotransmitter filledvesicles out of a reservepool anddeliverthemtothemembraneofthepresynapticcompartment.Uponarrival,thevesicle’smembranemergeswiththemembraneofthepresynapticcompartment,opensandreleasesitsneurotransmitterintothesynapticcleft.
Theaxonterminal’svoltage-sensitiveCa++channelscontainoneoffivedifferent

combinations of four protein subunits. All voltage-sensitive Ca++ channel subunitcombinations include an alpha subunit with the ion channel. The Ca++ alpha subunitdisplays an organizational pattern similar to the axon’s Na+ channel alpha subunitdescribedinthepreviouschapter.Thealphasubunitofthevoltage-sensitiveCa++channelcontainsafilterspecificforCa++.
The alpha subunits of voltage-sensitive Ca++ channels vary in their amino acidcomposition depending upon their location in the body and their precise physiologicfunction. Three variations of the Ca++ alpha subunit reside in the brain to serviceneurotransmitter release. Three associate proteins help anchor it in the axon terminalmembraneandmodulateitsfilterproperties,affectingitsactivationthresholdandrateofinactivation. Associate proteins also control transfer of the Ca++ alpha subunit proteinfromitsplaceofsynthesisintheneurontotheaxonterminalmembrane.
RETRIEVALOFNEUROTRANSMITTER
Neurotransmitter wears out its welcome in a synaptic cleft in a short period oftime. Action potentials travel to an axon terminal at a fast pace, often just a fewthousandthsofasecondapart.Ifneurotransmitterreleasedbyeachsignalisnotremovedrapidly,thesynapticcleftbecomessaturatedmakingfurthersignalsineffective.
Most neurons depend upon neurotransmitter reuptake pumps called transporterproteinstoremoveitfromthesynapticcleft,andtherebyenditseffectivenessatthenextneuron. Neurotransmitter transporter proteins represent one of the multiple classes ofproteins found in themembrane of the presynaptic compartment. Inmany areas of thebrain, neurotransmitter in the synaptic cleft is also transported into astrocyte glial cellssurroundingneuronsynapses.Overtwentyrecognizedproteinscomprisethecollectionofbrain neurotransmitter transporters. Each neurotransmitter uses a special subset of thetransporterproteins.
Transporter proteins take advantage of ion concentration differences on the twosidesofsynapticmembranestosupplytherequiredenergyforpumpingneurotransmitteroutof the synapticcleft.These transportersalsogoby thegenericnameofsymporters.Symportersmove twoormoredifferentmolecules,or ions, in thesamedirectionacrosscellmembraneswithat leastoneof themdiffusingdown its concentrationgradient andonebeingmovedagainstitsconcentrationgradient.Atthesynapse,themoleculemovingagainstitsconcentrationgradientistheneurotransmitter.Infact,thebrainusesmuchofitsenergypumpingsymporterionsbackoutofthecell.
MostneurotransmittersymportersdependuponNa+andCl-.InterstitialfluidinthesynapticcleftishigherinNa+andCl-thantheneuroncytoplasm.Thetransporterprotein’sshapeallowsNa+andCl- todiffuseacrosstheneuronmembraneintothecytoplasmandcarryneurotransmitterwiththem.Ionconcentrationgradientscanbethoughtofasindirectsourcesofenergy,becauseenergyconsumingpumpsarerequiredtomaintainthem.
An exception to reuptake of neurotransmitter from the synaptic cleft bytransporters occurs when an axon terminal releases the neurotransmitter namedacetylcholine. Acetylcholine is inactivated in the synaptic cleft rather than transported

back into the presynaptic terminal. There the enzyme acetylcholinesterase splits it intocholineandacetate.
Acetylcholinesterase is located on the interstitial fluid side of the postsynapticmembrane. Choline is then transported back into the presynaptic compartment forsynthesisonceagainintoacetylcholine.Thismayseemlikeacomplicatedapproach.But,theresultisarapidrateofremovalofacetylcholinefromthesynapticcleft.Theachievedratewouldbeunattainableusingatransporter.
POSTSYNAPTICCOMPARTMENT
POSTSYNAPTICDENSITYSynapsesonneurondendritescontinuetobethebrainsynapsesmostoftenstudied.
The postsynaptic compartments of neuron dendrites occur on small projections of thedendrite’smembraneandonthedendrite’smainshaft.
Thesmallprojectionsarenameddendriticspinesbecauseoftheirshape.Neuronswith extensive, branched dendritesmay contain up to thousands of dendritic spines perbranch(Figure5-3).Thegreater thebranchingof itsdendrites, the larger thevolumeofincomingsignalsaneuronaccommodates.
Dendrites experience simultaneous neurotransmitter input from multiplepresynaptic neurons. The response of the dendrite depends upon its complement ofproteins capable of interacting with neurotransmitters. The location and distribution ofvarioustypesofneurotransmitter-responseproteins influence thenetsignalpassedon tothe neuron cell body. Postsynaptic proteins that respond to neurotransmitter are calledreceptors.
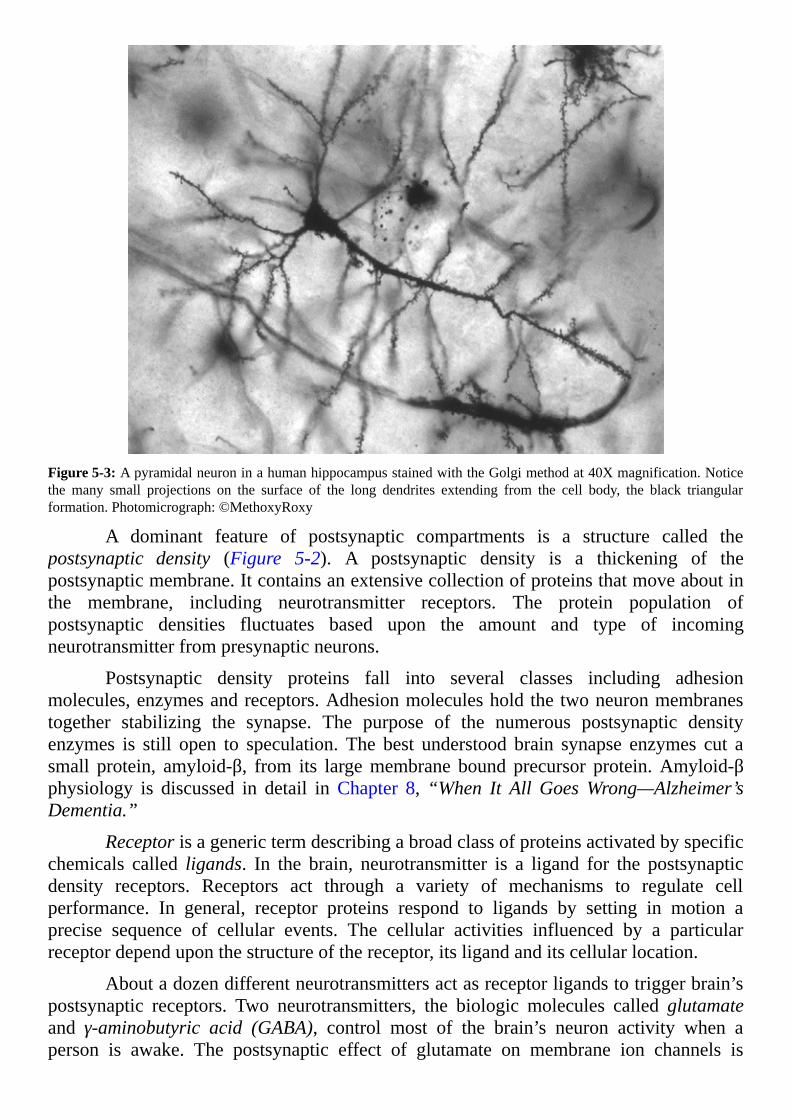
Figure5-3:ApyramidalneuroninahumanhippocampusstainedwiththeGolgimethodat40Xmagnification.Noticethe many small projections on the surface of the long dendrites extending from the cell body, the black triangularformation.Photomicrograph:©MethoxyRoxy
A dominant feature of postsynaptic compartments is a structure called thepostsynaptic density (Figure 5-2). A postsynaptic density is a thickening of thepostsynapticmembrane.Itcontainsanextensivecollectionofproteinsthatmoveaboutinthe membrane, including neurotransmitter receptors. The protein population ofpostsynaptic densities fluctuates based upon the amount and type of incomingneurotransmitterfrompresynapticneurons.
Postsynaptic density proteins fall into several classes including adhesionmolecules,enzymesandreceptors.Adhesionmoleculesholdthetwoneuronmembranestogether stabilizing the synapse. The purpose of the numerous postsynaptic densityenzymes is still open to speculation. The best understood brain synapse enzymes cut asmall protein, amyloid-β, from its largemembrane bound precursor protein.Amyloid-βphysiology is discussed in detail in Chapter 8,“When It All GoesWrong—Alzheimer’sDementia.”
Receptorisagenerictermdescribingabroadclassofproteinsactivatedbyspecificchemicals called ligands. In the brain, neurotransmitter is a ligand for the postsynapticdensity receptors. Receptors act through a variety of mechanisms to regulate cellperformance. In general, receptor proteins respond to ligands by setting in motion aprecise sequence of cellular events. The cellular activities influenced by a particularreceptordependuponthestructureofthereceptor,itsligandanditscellularlocation.
Aboutadozendifferentneurotransmittersactasreceptorligandstotriggerbrain’spostsynaptic receptors. Two neurotransmitters, the biologic molecules called glutamateand γ-aminobutyric acid (GABA), control most of the brain’s neuron activity when aperson is awake. The postsynaptic effect of glutamate on membrane ion channels is

estimated to consume about one third of the entire energy used by the brain.Most ofbrain’s energy consumption is byATP-requiring ion exchange pumps thatmaintain thetransmembranepotential.
Ligands glutamate and GABA trigger different dendrite receptors and opendifferent ion channels.Openglutamate receptor channels permit entryofNa+ andCa++
intodendriticspines.OpenGABAreceptorchannelspermitCl-toenter,andK+toleave,thedendrite’s shaft.EntryofNa+andCa++ throughglutamate receptors depolarizes theneuronmembranemaking the transmembrane potentialmore positive. Entry ofCl- andexit of K+ through GABA receptors generates an extremely negative, hyperpolarizedtransmembranepotential.
When the transmembrane potential is hyperpolarized, the Na+/Ca++ receptorchannelscannotopen,eveninthepresenceofglutamate.Thus,GABAreceptoractivationblocks the purpose of glutamate receptors. Glutamate is called an excitatoryneurotransmitterbecauseitcausesactionpotential-likedendriticspikes.GABAiscalledan inhibitory neurotransmitter because it blocks the ability of glutamate to initiatedendriticspikes.
NEUROPLASTICITYThe process by which the size and shape of dendritic spines alter over time, a
progressioncalledneuroplasticity,isoneofthemostexcitingareasofcontemporarybrainresearch.Dynamicdendriticspinesprobablyformtheanatomicinfrastructureofmemory.
Neuroscientistsbelieve thatneuronswithagreatnumberof largespineson theirdendrites are most likely to participate in formation of complex memories. Dendriticspinesexperiencingabundant input frompresynapticneuronspersist andenlarge.Thosereceivingfewsignalsshrinktoastubbyshapeandsometimesdisappearcompletelyfromthedendrite.SpineshapesusuallyobservedareshowninFigure5-4.

Figure5-4:Illustrationdepictingmainshapesfordendriticspines.Thisillustrationisreleasedtopublicdomainbytheartist
Dendritic spines operate as discrete neuron compartments. They display a highdegreeofmolecularorganization.Postsynapticglutamatereceptoractivationproducestwogeneralcategoriesofactionwithindendriticspines,anelectricalactionandabiochemicalaction
OpeningofglutamatereceptorionchannelspermitsentryintothedendriticspineofbothNa+andCa++.EnteringNa+flowsalongthemembraneofthedendriticspineintothedendrite.Na+ depolarizes the dendrite’smembrane toward the threshold needed forinitiationofdendriticspikes,whicharesimilartoactionpotentials,toalerttheneuronofincomingsignals.
In contrast, free Ca++ within the dendritic spine sets in motion a series ofintracellular biochemical reactions that ultimately create new molecules to enlarge thedendriticspineandincreaseitsnumberofneurotransmitterreceptors.
ELECTRICALOUTPUTOFDENDRITICSPINES
Na+ current, originating frommany dendritic spines is summed in the dendrite.The summedNa+ initiates a dendritic spike.Although dendritic spikes resemble actionpotentialsof theneuron’saxon, the locationof thevoltage-sensitive ionchannelsof thedendrite lack the rigid organization of an axon’s voltage-sensitive ion channels.Information about organization of axon voltage-sensitive ion channels is presented in

Chapter4,“Neurons—HowTheyMakeElectricity.”
Actionpotential transmembrane transientsof axonsareachievedusingonlyNa+
andK+voltage-sensitivechannels.Incontrast,dendriticspikesrequireNa+,Ca++andK+
voltage-sensitive channels.DendriticNa+,K+ andCa++ voltage-sensitive channels openwhenthetransmembranepotentialofthedendritereachesanappropriatethresholdvalue.Voltage-sensitiveK+ channels always open last. The threshold potential for a dendriticspike is far higher than the threshold for an action potential at the axon. Therefore, agreaterincreaseinNa+inthecytoplasmisrequiredforinitiationofadendriticspike.
Dendriticspikesmove toward theneuroncellbodyslower thanactionpotentialsmovealonganaxon.Therateofdendriticspikeprogressdependsuponthenumberandpattern of voltage-sensitive channels present and on the degree of branching of thedendritebeforethesignalarrivesatthecellbody.Nearthecellbodydendriticspikesarestrongerthandendriticspikesoriginatingatdistalpointsonthedendrite.Dendriticspikesflowovertheneuroncellbodymembraneandmergewiththecellbodyactionpotentialsinitiated by backflow of Na+ from action potentials of the axon. The net effect of theelectricalpotential along thecellbodymembranekeeps themetabolicmachineryof theneuronawareofbothincomingandoutgoingsignals.
BIOCHEMICALRESPONSETONEUROTRANSMITTER
Newbiochemicalmolecules created in response to the entry of freeCa++ into adendritic spine, transfer through the spine neck and the dendrite to the nucleus in theneuroncellbody.Abiochemicalconnectionbetweendendriticspinesandtheneuroncellnucleus is necessary to maintain a large population of proteins in the postsynapticcompartment.Dendriticspinesdependupontheneuronnucleustoenhancetheirsupplyofreceptors and structural proteins when there is an increase in the number of incomingneurotransmittersignals.
Biochemical molecules synthesized in dendritic spines and later found in theneuronnucleus include certain kinases and transcription factors.Kinase enzymes attachphosphate groups to other proteins. Protein transcription factors,which bind to nuclearDNA,initiatecopyingoftheDNAgeneticcodeintomessengerRNA(mRNA).CellsusemRNAasatemplateforconstructionofnewproteins.
SpecifictranscriptionfactorsinitiatereadingofthecodeofindividualgenesoftheDNA.Kinasesfromthedendriticspinesconverttranscriptionfactorsalreadyintheneuronnucleusfromaninactivetoanactiveform.Atleast300genesarethoughttoberegulatedbythebiochemicalmoleculesoriginatingindendriticspines.
ItappearsthemRNAcreatedinthenucleusinresponsetosynapticactivityatthedendriticspines,andthemolecularequipmentneededforquicksynthesisofproteinfromthatmRNA,transporttodendriticspinesingranularvesicles.ThelocationintheneuronwherepackagingofthemRNAoccursisnotknown,noristhenatureofthevehicleusedto deliver the granules to the dendritic spines identified. However, granular vesiclescontainingmRNAdotravelthroughneuroncytoplasmtodendriticspines.Thesegranularvesiclesremaininalatentprotectedstateuntilneeded.

With increased neurotransmitter activity, mRNA released from the granulescommencesnewproteinsynthesiscausingthedendriticspinestoenlarge.Wheninputofneurotransmitter to thepostsynapticcompartmentslows,freeCa++ inthedendriticspinedecreases and the process of supplying newmRNA/ribosomegranules from the neuronnucleusalsoslowsandthedendriticspineshrinks.
EXCITATORYANDINHIBITORYNEURONS
EXCITATORYGLUTAMATESYNAPSESNeurons releasing the neurotransmitter glutamate are excitatory neurons.
Glutamate synapses display a greater complexity than the generic synapse presentedearlier (Figure5-2).This isnotsurprisingbecauseexcitatoryglutamateneuronssupportmultifacetedaspectsofmemoryformationandmemoryrecall.Glutamateneuronsynapsesarelostearlyasdementiaprogresses.
Glutamatesynapsesinclude,inadditiontotwoneurons,cellsthatarenotneurons.Other brain cells forming part of glutamate synapses are glial astrocytes and surveyingmicroglia.Glialastrocytessurroundneuroncellbodiesandparticipateinthetimelyuptakeofneurotransmitterfromsynapticclefts.Surveyingmicrogliamonitors thewell-beingofsynapseactivitymanytimesperhour.Chapter6,“IntroductiontotheGliaandMicroglia—MeettheStageCrew”reviewstheroleofthegliaandmicrogliaatsynapses.
There are multiple forms of the glutamate receptor in the brain. Subtypes ofglutamate receptors that open ion channels may be separated from each other by theirability to bind pharmaceutical drugs. An added feature at glutamate synapses is thepresenceofanothervarietyofglutamatereceptorthatdoesnotopenionchannels.Insteadthese receptors respond toglutamateby activating internalneuronprocesses.Glutamatereceptors forming ion channels localize to the postsynapticmembrane in, and near, thepostsynaptic density.The glutamate receptorswithout an ion channel are found both inpre-andpostsynapticmembranes.
Glutamatereceptors,whichdonotformionchannels,bindglutamateonaportionofthereceptorextendingoutsidetheneuronthatcausesachangeinthepositionofapartof the receptor located inside the neuron. Repositioning of the cytoplasmic part of thisglutamate receptor allows it to bind to cytoplasmic regulatory molecules. The boundregulatory molecules set in motion synthesis of new molecules in the cytoplasm that,depending upon neuron circumstances,may enhance or reduce the effectiveness of thesynapse.
Glutamate’s ion channel receptors arenamedAMPAandNMDAafter thedrugsspecified as alpha amino-3-hydroxyl-5-methyl-4-isoxazole-proprionate andN-methyl-D-aspartate,respectively.AMPAandNMDAglutamatereceptorsworkasateamresponsiblefor glutamate’s immediate excitatory action at the postsynaptic density. The AMPAreceptorrespondsfirst.
Mobile AMPA receptor proteins rapidly cycle into and out of the postsynapticdensityfromadjacentareasofmembrane.AMPAreceptorsincludefourdifferentproteinsubunits named GluA1, GluA2, GluA3 and GluA4. Four of these subunits in variouscombination,mostoftentwoGluA1withtwoGluA2,clustertogetherinthepostsynaptic

membranecreatinganionchannelatthecenteroftheircomplex(Figure5-5).
Figure5-5:TheAMPAreceptorforglutamateboundtoaglutamateagonistshowingtheaminoterminal,ligandbindingdomain and transmembrane domain. This reconstruction is based upon file 3KG2 in the protein data bank.©CurtisNeveu
AMPAandNMDAreceptorsbothformnon-selectivecationchannels.Cationsareions carrying a positive charge.Non-selective cation channel filters allow simultaneouspassageofNa+,Ca++ and someK+. Na+ andCa++ enter dendritic spines through thesechannelsandK+flowsoutofthedendriticspineaseachionmovesdownitsconcentrationgradient. The net result of the larger influx of Na+ compared to efflux of K+ isdepolarizationofthetransmembranepotential.
TheAMPAandNMDAformsoftheglutamateionchannelreceptorsexistasapairon glutamate responsive dendritic spines. The AMPA receptor is the first to open inresponse to glutamate. The NMDA receptor’s ion filter is blocked by magnesium ion(Mg++)whenthemembranepotentialisatitsnormalnegativerestingvalue.
NMDA receptor relies upon theAMPA receptor to initiate depolarization of themembrane in response to glutamate. Glutamate binding to the AMPA receptor rapidlycausesmembranedepolarizationforcingMg++awayfromtheNMDAfilter.GlutamateisthenabletoopentheNMDAfilterandexpandcationflowintothedendriticspine.
TheNMDA receptor like theAMPA receptor is a four subunit protein complexcomposedoftwoGluN1andtwoGluN2subunits.Differentgenescodefourisoformsof

theGluN2subunit.Thisdiversityallows thehumanbrain todisplaymanyvariationsofglutamateionchannelreceptors.
InadditiontotheprerequisiteofmembranedepolarizationbytheAMPAreceptorbefore theNMDA ion channel can open,NMDA receptor requires binding of a secondmolecule.Thesecondmoleculemaybeeitheroftheaminoacidsglycineorserine(Figure5-6).Bothoftheseaminoacidsareprobablycontributedbytheastrocyteglialcells thatsurroundbrain synapses.There are alsoother areason theNMDA receptorwheredrugmoleculescanbind,change its shapeandmodify itsability to functioncalledallostericsites.
Figure 5-6: Illustration of two of the four subunits of awholeNMDA receptor showing the relative location of theglutamateandtheglycinebindingsites.Glutamateandglycinedonotoccupythesamesubunit.FoursubunitNMDAreceptorsrequirethebindingoftwomoleculesofglutamateandtwoofglycineorserineforactivation.©CurtisNeveu
INHIBITORYGABASYNAPSESGABA is synthesized by enzymes in the brain from glutamate. Neurons using
GABAasaneurotransmitterformsynapsesontheneckofthedendriticspines(Figure5-4)oronthedendriteitselfbetweendendriticspines.GABAneuronsalsoformsynapsesonthemembraneofneuronpresynapticaxonterminals,onneuroncellbodiesandsometimesat sections of axonswithoutmyelin cover.Reuptake ofGABA from a synaptic cleft isaccomplishedbythepresynapticaxonterminalwithsomeassistancefromastrocyteglialcellsinthearea.
TherearetwotypesofGABAreceptorsdesignatedGABAtypeAandGABAtypeB.GABAbindingtoaGABAtypeAreceptoropensanionchannel that isselectiveforchlorideion,Cl-.BecauseCl- ishigher in the interstitial fluid thaninneuroncytoplasm,Cl-diffusesthroughGABAionchannelreceptorsintothepostsynapticcompartment.The

additional negative charge brought into the neuron by Cl- causes the transmembranepotentialtofallbelowitsnormalrestingvalue.
NegativehyperpolarizationofaneurontransmembranepotentialmakesitdifficultforNa+enteringthroughglutamateionchannelreceptorstomakethemembranepotentialpositive enough to initiate dendritic spikes.Neurons usingGABAas a neurotransmitteracquiretheirgenerictitle,inhibitoryneurons,becausetheyblocktheabilityofglutamatetoperformitsnormalexcitatorypurpose,initiationofdendriticspikes.
In contrast to GABA type A receptors, GABA type B receptors traverse themembrane without forming an open channel. When GABA binds to the extracellularportionofaGABAtypeBreceptor,theportionoftheproteininsidetheneuronrepositionsitself to better facilitate a series of cytoplasmic events. Those cytoplasmic events overtimecausesomeoftheneuron’sK+channelstoopenandtoremainopenforaperiodoftime.
TheexitofK+ fromtheneuronhyperpolarizesthetransmembranepotentialevenfurtherthantheentranceofCl-alone.ThecombinedinfluenceofthetwotypesofGABAreceptorsatdendriteseliminates theeffectofopenglutamate ionchannels leading toanabsence of dendritic spikes and lack of new structural proteins and receptors for thepostsynapticcompartment.
TheGABA typeA receptor, schematically diagramed inFigure 5-7, consists offive subunits arranged around a central channel for Cl-. Each subunit contains fourtransmembrane domains. Opening of the Cl- channel requires binding of two GABAmolecules.Thesizeof thefilter in thechannelcanbemodifiedbythepresenceofdrugmoleculesbindingatvariousallostericsites.
Figure5-7:SchematicdrawingoftheGABAtypeAreceptorproteinillustratingthefivecombinedsubunitsformingtheCl-channel.ThepositionsofthetwoGABAbindingsitesandthedrugbindingsiteforbenzodiazepineareillustrated.Thisillustrationisinthepublicdomain.
Many pharmaceutical drugs bind toGABA typeA receptors and influence theiroperation.DrugmoleculesbindatallostericsitesseparatefromGABA’sbindingsite.Thephysiologiceffectsofthedrugsincludesedation,amnesiaandanticonvulsantactivity.

OTHERNEUROTRANSMITTERSAll brain synapses present a variation of the basic theme introduced with
discussionofglutamateandGABAsynapses.Toomanyneurotransmitter receptor typesandsynapseconfigurationsexistinthehumanbraintodiscussthedetailsofeachofthemhere. However, like glutamate and GABA, the other neurotransmitters are ligands forreceptorsthatsetinmotioncomplexbiochemicalpathwayswithinneurons.
SUMMARYCHAPTER5Synaptic transmission is used by neurons to communicate with each otheracrossnarrowspacescallsynapticclefts
Brainsynapsesoccuratneurondendrites,atcellbodies,ataxonsandataxonterminals
Action potentials arriving at the axon terminal cause voltage-sensitive Ca++
channelstoopenallowingfreeCa++toentertheaxonterminalcytoplasm
CytoplasmicfreeCa++intheaxonterminalsetsinmotionprocessesleadingtothereleaseofasmallchemicalcalledaneurotransmitter
Neurotransmitter activates receptors of the postsynaptic compartment thatresideinthepostsynapticdensity
Neurotransmitterisremovedfromthesynapticcleftbyreuptakepumpsofthepresynapticcompartmentandbyastrocyteglialcells
Synapsesatneurondendritesoccureitheronthemainshaftofthedendriteoronprojectionsofthedendrite’smembranecalleddendriticspines
Neurons using glutamate or GABA as a neurotransmitter control the largestportionofbrainactivitywhenapersonisawake
TherearetwoclassesofglutamateandGABAreceptorsinthebrain,oneclassquicklyopensionchannels in thepostsynapticdensity, theotherclassslowlychangescellularprocesseswithinneurons
Glutamatereceptorsofthepostsynapticdensitydepolarizethetransmembranepotentialinthepresenceofglutamate
GABAreceptorsonthedendritehyperpolarizethetransmembranepotentialandblockthedepolarizingeffectsofglutamate
Na+currentsenteringthedendritethroughglutamatereceptorstriggeranactionpotential-likeresponseatthedendrite’smembranecalledadendriticspike
Ca++ currents entering dendritic spines set in motion synthesis of additionalproteinsneededforfurtherdevelopmentofthesynapticconnection
GABAreceptorshyperpolarizethedendritebyopeningchannelsforCl-andK+
Excitatoryneuronsreleasetheneurotransmitterglutamate
InhibitoryneuronsreleasetheneurotransmitterGABA

[6]
IntroductiontotheGliaandMicroglia—MeettheStageCrew
ONLY 10% OF BRAIN CELLS are neurons. The remaining 90% of cellsparticipateaspartnersoftheneuronsinmanagementofbrainfunction.Thegeneralnameforthenon-neuroncellsinbraintissueisglia.ThelabelgliaoriginatesfromawordintheGreek language that means glue. The first brain anatomists thought glia suppliedscaffoldingforneurons.Thefragileappearanceofneuronssuggestedaneedforsometypeofphysicalsupport.Hencethemisnomerimplyinggliaisgluetocementneuronstogether.
Thereisalsoagroupofsmallcellsinthebrainoncethoughttobepartoftheglia.Becauseof theirsize theywerenamedmicroglia.However,observationsinrecentyearsestablished brain microglia is not part of the glia population. The studies showedmicrogliaoriginatesfromadifferentstemcellpopulationthantheglia.
STEMCELLS
RADIALCELLSIn embryos radial cells produce embryonic neurons. Radial cells (Figure 6-1),
named for their radiating appearance, originate from the cells forming the neural plateduringtheinitialphasesofembryonicdevelopment.Radialcellscanalsomatureintoglia.Maturegliaincludestwodistinctcellpopulations,astrocytesandoligodendrocytes.

Figure6-1:Radialglia,alsoknownasBergmannglia,ofthecerebellumofamouseaged7days.ThisworkisinthepublicdomainintheUnitedStatesbecauseitwasproducedbyanofficeroremployeeoftheUnitedStatesGovernmentaspartoftheperson’sofficialduties.
Asradialstemcellsbegintomature,aportionoftheirpopulationstopsanddoesnot continue on to fullmaturity. These partially differentiated cells become the brain’sneuralstemcells.Theneuralstemcellpopulationpersistsinjuvenileandadultbrainandprovidesasourceofreplacementneuronsandoligodendrocytes.
Neuralstemcellscapableofbecomingnewneuronsexistintheadultbrainunderthelayerofependymalcellsliningthelateralventriclesandinazoneofthehippocampusregion of the cerebral cortex. The hippocampus, discussed in the next chapter, isrecognizedforitsprominentroleinmemoryformationandmemoryrecall.
ASTROCYTESInmanyregionsof thebrainandspinalcord,astrocytegliadevelopsfromradial
cells after the original neuronsmature. Few astrocytes display measurable rates of celldivision inhumanbrain tissue.Replacementofastrocytes seldomoccurs. It appears themajorityoftheoriginalastrocytessurvivemostofalifetime.
OLIGODENDROCYTESIn theembryo, radialcellsdestined tobecomeoligodendrocytes, sometimesalso
referredtoasoligodendrocyteprogenitorcells(OPCs)firstappearintheventralforebrain.OPCs travel longdistances from the forebrain to reach theirultimatedestinations in thedevelopingbrain.
Manymonitoringmolecules support the progress of oligodendrocytesmigrationthroughbraintissuetotheneuronstheywillprotect.Regulatormoleculesguidecontact-basedmigrationofOPCsoverextracellularmatrices,axon tractsandastrocytesurfaces.Expansion and retractionof slendermembrane extensionsofmaturingoligodendrocytes

aidtheirmigrationandenhancetheirabilitytomakecontactwithneuronaxons.
AportionoftheOPCsdistributingthroughbraindoesnotimmediatelymatureintooligodendrocytes.Theprogenitorcellsremaindormantinthetissueaspartoftheneuralstem cell population until encouraged by chemical signals in the brain to resumedevelopment.
MICROGLIA
Microglia originates from the extra-embryonic yolk sac during initial blood cellformation in early embryogenesis. Microglia expresses some but not all of the sameantigens as macrophages formed during blood formation by bone. However, microgliaseedsthecentralnervoussystembeforebloodformationbeginsinbone.
Microglia is present at all stages of brain development. In healthy brain,maintenance and local expansion of the microglial population is dependent upon self-renewal of resident cells. All precursor cells for microglial expansion in adult brainarrivedtherepriortobirth.
Thediscovery thatmicroglia,maturebraincells, retains theability to self-renewindefinitely in the same fashionas adult stemcellshas caused scientists to review theirdescriptionofstemcells.Bydefinitionstemscellsareimmaturecellsthatcanproceedintwodirections.Theycanmatureintospecializedcells,ortheycandividetoproducemorestemcells.
Dogmauntilrecentlystatedthatmaturecellscannotself-renew,thatisreproducethemselves indefinitely like stem cells. This property of brainmacrophage-like cells toself-renewisalsoobservedinthemacrophagepopulationofcertainothertissues.
ADULTGLIAANDMICROGLIA
ASTROCYTESAstrocytegliacomprisesthemostabundantcellpopulationintheadultbrainand
spinalcord.Astrocytesoutnumberneuronsbyoverfivefold.Noregionofthebrainlacksastrocytes(Figure6-2).

Figure 6-2: Protoplasmic astrocytes in the human cerebral cortex stained with Cajal’s gold sublime method.Photomicrograph:©JoseLuisCalvo
Astrocytes tile the brain in an ordered non-overlapping arrangement. Individualastrocytes manage distinct domains. Only the most distal tips of membrane extensionsradiatingfromindividualastrocytecellbodiestouchoneanother.Eachastrocyteexhibits5-10mainradiatingextensionsthatgiverisetonumerousfinerbranches.
Astrocytes divide into twomain subtypes,protoplasmic and fibrous, based upontheirmorphologyandanatomiclocationinthebrain.Protoplasmicastrocytesspreadinthegray matter near neuron bodies. Fibrous astrocytes (Figure 6-3) position themselvesthroughoutallwhitematterwheretheymakecontactwithaxonsatNodesofRanvier.

Figure6-3:FibrousastrocytesofbraintissueshowingmanylongthinmembraneextensionsstainedbytheGolgisilverchromatemethod.Photomicrograph:©JoseLuisCalvo
Bothtypesmakeextensivecontactwithbloodvesselsandregulatebloodflowbysecretingmoleculestoeitherdilateorconstrictthebraincapillaries(Figure6-4).
Figure 6-4: Astrocyte membrane extensions with end feet to the walls of a blood vessel stained with Cajal’s goldsublimemethod.Photomicrograph:©JoseLuisCalvo
OLIGODENDROCYTESOligodendrocyteslocatedattheaxonsofneuronsproducethewhitecolormarking
neuron tracts in thebrain.Whitematterof the adult brain is crowdedwithbothmature
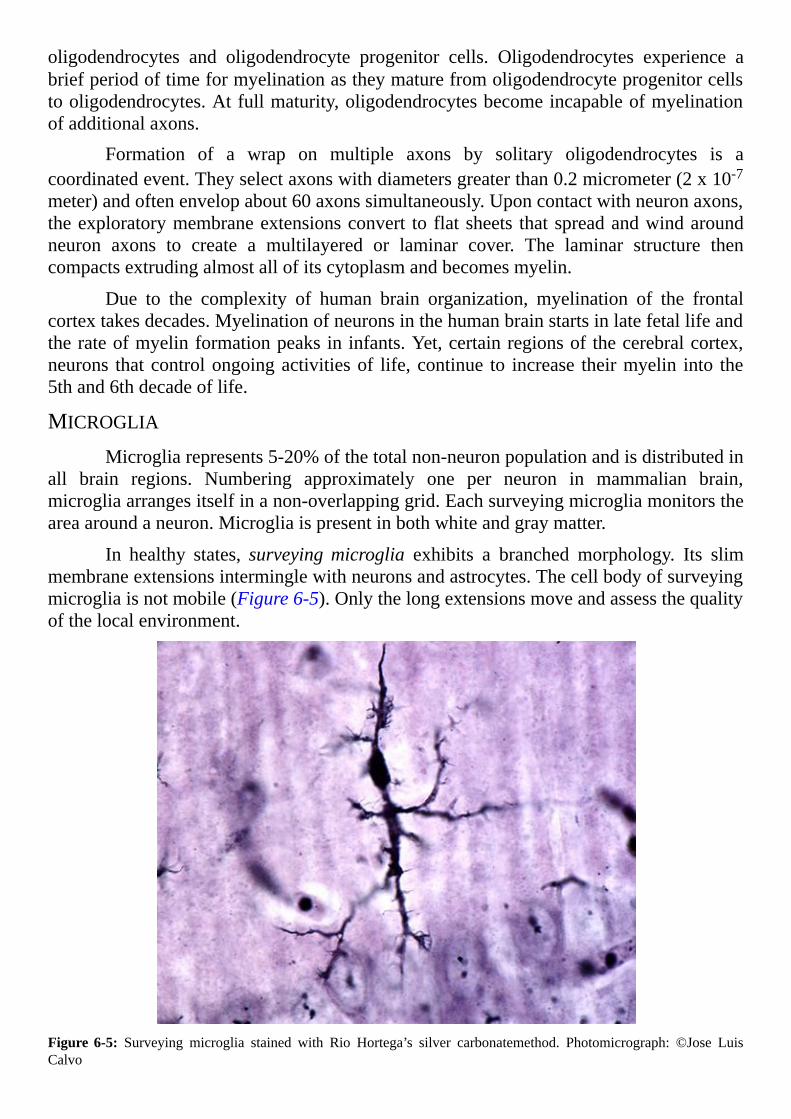
oligodendrocytes and oligodendrocyte progenitor cells. Oligodendrocytes experience abriefperiodoftimeformyelinationastheymaturefromoligodendrocyteprogenitorcellstooligodendrocytes.Atfullmaturity,oligodendrocytesbecomeincapableofmyelinationofadditionalaxons.
Formation of a wrap on multiple axons by solitary oligodendrocytes is acoordinatedevent.Theyselectaxonswithdiametersgreaterthan0.2micrometer(2x10-7meter)andoftenenvelopabout60axonssimultaneously.Uponcontactwithneuronaxons,theexploratorymembraneextensionsconvert to flatsheets thatspreadandwindaroundneuron axons to create a multilayered or laminar cover. The laminar structure thencompactsextrudingalmostallofitscytoplasmandbecomesmyelin.
Due to the complexity of human brain organization, myelination of the frontalcortextakesdecades.Myelinationofneuronsinthehumanbrainstartsinlatefetallifeandtherateofmyelinformationpeaksininfants.Yet,certainregionsof thecerebralcortex,neurons thatcontrolongoingactivitiesof life,continue to increase theirmyelin into the5thand6thdecadeoflife.
MICROGLIA
Microgliarepresents5-20%ofthetotalnon-neuronpopulationandisdistributedinall brain regions. Numbering approximately one per neuron in mammalian brain,microgliaarrangesitselfinanon-overlappinggrid.Eachsurveyingmicrogliamonitorstheareaaroundaneuron.Microgliaispresentinbothwhiteandgraymatter.
In healthy states, surveyingmicroglia exhibits a branched morphology. Its slimmembraneextensionsinterminglewithneuronsandastrocytes.Thecellbodyofsurveyingmicrogliaisnotmobile(Figure6-5).Onlythelongextensionsmoveandassessthequalityofthelocalenvironment.
Figure 6-5: Surveying microglia stained with Rio Hortega’s silver carbonatemethod. Photomicrograph: ©Jose LuisCalvo

FOURPARTSYNAPSESThepreviouschapterreviewedthetwopartsofaneuronsynapse,thepresynaptic
compartment and the postsynaptic density. In real brains, however, the synapse whereneurons come together is a four part structure. The four-part synapse includes thepresynapticandpostsynapticneurons,anastrocytenetworkandsurveyingmicroglia.Thecrowded collection of several cell types aroundneuron synapses presents a diversity ofcommunication opportunities for maintenance of healthy brain function and fornetworkingamongcells.
ASTROCYTEWEBS
Astrocytes, connected by gap junctions where the tips of their membraneextensions come in contact, create cellular webs. Gap junctions, membrane structuressharedbytwocellswithacentralopenchannel,jointhecytoplasmofmultipleastrocytes.The gap junctions allow ions, nutrients and othermolecules to passwithout restrictionbetweenastrocytes.Thispermitsmultipleastrocytestobehavelikeasinglecell.
The larger astrocyte webs reachmillimeter (1 x 10-3 meter) size in some brainregionsandincludehundredstothousandsofastrocytes.Movementofsubstancesthroughastrocyte networks is modified by activity in neighboring neurons. Neuron signalinginitiates changes in astrocyte cytoplasmic concentrations of calcium (Ca++) and sodium(Na+),powerfulionsfortriggeringcytoplasmicresponses.
Astrocyte networks organize themselves into separate anatomic compartments.Astrocyte networks near the neurons committed to a unique functional activity mayexperience an influential relationship exclusive to that set of neurons. Alternately,astrocytenetworksmayactasbarrierstoisolateneuronsystemsandrestrictinformationtransferbetweenneuroncircuits.Neuroncircuitsincludemultipleneuronsperformingasasingle unit. Astrocyte barriers may provide effective segregation of neuron activity invariouspartsofthebrain.
Boundaries formed by both protoplasmic and fibrous astrocytes take part inguiding the migration of oligodendrocyte progenitor cells. And, both subtypes ofastrocytesprovidephysicalandfunctionalsupportatbraincapillaries.Astrocytesmaintainintegrity of the blood brain barrier and regulate blood flow to match it to neuronrequirement for glucose and oxygen. Protoplasmic astrocytes link neuron bodies anddendriteswithbloodvessels.Fibrousastrocyteslinkaxonswithbloodvessels.
SURVEYINGMICROGLIA
Surveying microglia, sometimes called resting microglia, possesses a tinystationary cell body (Figure 6-5). The membrane extensions emanating from the bodyelongate and retract as they explore the brain tissue environment. Surveyingmicrogliascans its localareawithprecisionandmakesphysicalcontactwith synapses tomonitorsynapticactivity.
Surveyingmicrogliamakesbrief,about5min,contactswithneuronsynapsesatafrequencyofaboutonceperhour.Thetimingofthecontactsdependsonthefrequencyofneurotransmitter release by the presynaptic compartment. Surveying microglia contacts

synapseslessoftenwhenneuronactivitylessens.
Surveyingmicrogliamakes contactwithneurons at synapsesondendritic spinesbutnotwiththeshaftsofdendrites.Itscontactatsynapsesmaybemediatedbyavarietyof chemo-attractant and repellant signaling molecules. Proposed signaling moleculesinclude glutamate,ATP,molecules named chemokines, brain-derived growth factor andioncurrents.
FUNCTIONALPARTNERSHIPS
ASTROCYTESANDNEURONSThe extent of astrocyte engulfment of neuron synapses and Nodes of Ranvier
variesbybrainregion.However,allsynapsesinthebrainmaintainsomeinteractionwithastrocytes.
Inareasofthebrainlikethehippocampusandcerebralcortex,themanybranchesof a single protoplasmic astrocytemay contact several hundred dendrites frommultipleneuronsandenvelopupto100,000ormoresynapses.Thisaffordsindividualastrocytesaswellasastrocytenetworksextensiveneuronmonitoringpotential.
A primary function of fibrous astrocytes is to monitor and maintain steady ionconcentrations in the interstitial fluid around axons. Their extensive contactwith bloodcapillariespermitsthemtocorrectanydeviationfromtheacceptablerangeofinterstitialfluidionconcentration,whichisessentialtoactionpotentialpropagation.
Astrocytes, like postsynaptic compartments of neurons express neurotransmitterreceptors on their distal branches. The neurotransmitter receptors expressed match theneurotransmitterreceptorsoftheneuronsynapsestheycontact.
Yet, the timing of ion currents at the astrocyte bodies indicates neurotransmitterreceptors on their branches do not respond to normal levels of neurotransmitter.Understressful conditions, astrocytes possess the potential to release the neurotransmitter toactivatetheirownreceptors.Inhealthybrain,however,thisisnotthoughttooccur.
Neurons, which are active when the brain is awake, release a neurotransmitternamedglutamate.Glutamateisremovedfromsynapticcleftsbyresidentastrocytes.Thecellmembraneof astrocytesnearaglutamate synapseexpresses transporterproteins forglutamate.
The drain of glutamate fromneurons is compensated for by flowof amoleculenamedglutaminefromastrocytestoneurons.Aboutonethirdoftheglutamatetakenupbyastrocytes is converted to glutamine and returned to the neuron. Within the neuronglutamine is converted back to glutamate and repackaged into vesicles for synapticrelease.
Astrocytes supplement glutamine made from neuron glutamate with glutaminesynthesizeddenovofromglucose.Theremainingtwothirdsoftheglutamatetakenupbyastrocytes replenish components of the astrocytes mitochondrial ATP-producing cycle.Neuronslacktheenzymesnecessarytomakeglutamatefromglucose.
Onetheoryisthisarrangementprovidesanenergyadvantagetoneurons.Itpermits

ALL the glucose entering neurons to bemovedwith high efficiency throughglycolysisandALLtheresultingpyruvatetobetransferredintotheirmitochondriatoproducehighenergymoleculesofATP.Ifneuronsmadeglutamate,partoftheirpyruvatefromglucosemetabolism would be drawn off for glutamate synthesis and would result in a loweroverallATPproduction.
OLIGODENDROCYTESANDNEURONAXONSThe thickness of an axon’s myelin sheath varies. Variation in myelin thickness
along with differences in the length of individual Nodes of Ranvier may provideadditionalopportunitiesforneuronstoadjustthespeedoftheiractionpotentials.Becauseonlylargeraxonsattractoligodendrocytes,itisassumedelectricalactivityoftheaxonisanessentialsignalformyelination.Insupportofthisidea,whenintricatemotorskillsarelearned, axon myelination increases in synchrony with increased motor cortex neuronactivity.
Neuronaxontransportsystemsandoverallneuronviabilityalsoappeartodependuponsupportivesignalsfromoligodendrocytes.Oligodendrocytesinsurethewell-beingofaxons by releasing growth stimulatingmolecules including glial cell neurotropic factor,brain-derived neurotropic factor and insulin-like growth factor. Axons with damagedmyelin sheaths display alteration in the stability of their cytoskeleton which leads toadversechangesinthetransportratesofmaterialbetweentheneuronbodyandtheaxonterminal.
Oligodendrocytesmay play a larger role in establishing electrical circuits in thebrain than is appreciated. Proteins in the oligodendrocytes’ membrane inhibit axoncollateral formation. Observed pattern variation in the length of the Nodes of Ranvier,therefore,may affect a neuron’s opportunities for using axon collaterals in its circuitrydesign.Inaddition,synapsesonaxonsoccuronlywhereanaxonisbareattheNodesofRanvier.
MICROGLIAANDNEURONSMicrogliaconnectswithastrocytesandneuronsatsynapses.Thereisaligandfora
brain receptor on the membrane of neurons, astrocytes and oligodendrocytes namedCD200.ThereceptorforCD200isexclusivetomicroglia.TheCD200moleculesonthesurface of neighboring cells keepmicroglia in its surveying state. Surveyingmicrogliaalsoexpressesreceptorsformostbrainneurotransmittersthatallowittomonitoractivityatallbrainsynapses.
The behavior of surveying microglia is regulated and guided by activity ofindividualneurons.NeuronswithhighlevelsofactivityreleaseATP.ATPinthesynapseinducesmicroglia to polarize its extensions toward these dynamic synapses. Surveyingmicroglia removes dendritic spines when neuron activity slows. Pruning of synapsesreceiving little neurotransmitter input is required to maintain the dynamic character ofneuron circuits. In the developing visual system, live images observed with confocalmicroscopy followed by electron microscopy confirm surveying microglia removessynapticelements.
METABOLISMINTHEBRAIN

GLUCOSEFORENERGYThebrainreliesonbloodglucoseasitsprimarysourceofenergy.Glucosemoves
into the brain through capillary endothelial cells with the help of transporter proteins.Astrocyteswhoseextensionsreinforcethebloodbrainbarrierarewellpositionedtotakeup entering glucose. Neurons also absorb incoming glucose. To assist neurons in thisendeavor,astrocytessecretemoleculesthatdilatebloodvessels.Astrocytesadjustmentofbloodflowmatchesactivityatsynapsestoavailabilityofglucose(Figure6-4).
Glucosecanbestockpiledinsomecellsintheformofastoragemoleculenamedglycogen. Astrocytes serve as the principle storage site for glycogen in the brain.Glycogen containing astrocytes exist in areas where synapses pack close together.Astrocyteglycogenstockssupportnormalneuronactivityintimesoflowbloodglucose.Exceptionally active neurons create areaswith lowblood glucose.When blood glucosebecomesinadequate,glycogenisconvertedfirsttoglucoseandthenmetabolizedtolactatebyastrocytesfordeliverytoneurons.TheneuronsconvertlactatetopyruvatetofueltheirproductionofATP.
Whenbrainactivity is in itsnormal range, theastrocytestockpileofglycogen isgraduallybrokendownandrestored.Theroutineuptakeofglutamateneurotransmitterbyastrocytestriggersconversionofglycogentoglucose.Inadditiontousingglucosetomakeglutamine to send to the neurons, astrocytes make ATP to support their own Na+ ionpumps.Whenastrocytesremoveglutamatefromasynapticcleftusingtransporters,Na+isbrought into the cell along with glutamate. Ion pumps must then restore astrocytecytoplasmtoitsnormallowlevelofNa+.
Brain cells, other than neurons and astrocytes, also use glucose for energy.Oligodendrocytes during the myelination procedure consume substantial amounts ofglucose, oxygen andATP. Surveyingmicroglia is not thought to require a high rate ofglucose utilization, butactivatedmicroglia, needed to repair brain damage or stave offinfection,increasesitsglucoseuptake.
CHOLESTEROLFORMEMBRANESTRUCTUREMany different kinds of lipids, fat molecules, are used to form brain cell
membranes.Somelipidscrossintothebrainfromblood,otherscannotnavigatethroughthebloodbrainbarrier.Cholesterolisamongthelipidsthatcannotcrossthebloodbrainbarrier.Thereisnomixingofbraincholesterolwithcholesterolinblood.Allcholesterolusedbybraincellsismadeinthebrain.
Thebraincontainsmorecholesterol thananyotherorgan.Twenty to twenty-fivepercentofaperson’scholesterolissynthesizedinthebrainandresidesthere.Cholesterolisamajorcomponentofbraincellmembranes(Figure6-6).

Figure6-6:Illustrationofcholesterol’sroleinlipidmembraneformation.Arrowspointtocholesterol(yellow)insertedbetweenstructurallipidsinacellmembrane.Illustration:©WarX,releasedtothepublicdomain
Cholesterol is critical to synapse formation, dendrite formation, axon elongationand is a major component of myelin. Brain cholesterol is also vital to efficientneurotransmitter release from axon terminals. It is an important component of vesiclesusedfortransportofmaterialswithinneuronsalongcytoskeletalcomponents.Cholesterolmakes regions of membranes less fluid and more resistant to the passage of chargedmolecules. It presence at synapses supports the functionof ion channels andmembranereceptors.
Astrocytesandoligodendrocytesproduce2-3timesmorecholesterolthanneurons.Astrocytesprovide cholesterol for growthof neurons and to support synapse structures.Theconcentrationof cholesterol inbrain is kept stable. Its half-life is reported tobe aslongas5years.Incontrast,inbloodplasmathehalf-lifeofcholesterolisafewhours.
Brain cells take up cholesterol enclosed in apolipoprotein E (ApoE) with amembranereceptor.ApoE,producedbyastrocytesandmicroglia,isthemajortransporterof cholesterol within the brain. Astrocytes secrete small ApoE particles composed ofphospholipidandcholesterolthatperformlikehigh-densitylipoprotein(HDL)circulatingoutside the brain. The larger triglyceride-containing low-density lipoprotein (LDL)particlesproducedbyliverappearabsentinthecentralnervoussystem.
Elimination of cholesterol from the brain requires its conversion to 24S-hydroxycholesterol. This enzymatic change occurs mainly in neurons. The conversionproductcrossesthebloodbrainbarrierandexitsneuronsandthebrainwithoutrestriction.
REPAIROFBRAINDAMAGE
REACTIVEASTROCYTESNeurons respond to areas of injury in the brain by adjusting their circuits in
undamaged sections of the brain to by-pass failure of the damaged tissue. Isolation ofinjured tissue followed by tissue repair or elimination of damaged tissue is left toastrocytes,oligodendrocytesandmicroglia.
Astrocytes respond to all types of central nervous system injury. Astrocytes

reacting to tissue rupture remove excess glutamate. This is protective because surplusglutamate in brain fluids is toxic to neurons. Astrocytes shield cells in damaged braintissue from oxidative stressors by producing the anti-oxidant glutathione. Glutathioneneutralizes highly destructive oxygen-containing molecules released by the injury.Astrocytes also facilitate repair of damage to the blood brain barrier and reduce excessfluidaccumulation,edema.
Aresponseknownasreactiveastrogliosisisahallmarkofstructuraldamageinthebrain. Reactive astrogliosis is a graded response. Small injuries produce a mild tomoderateformofastrogliosis.Largerinjuriesproduceseverediffuseastrogliosiswithorwithoutglialscarformation.
Duringmildtomoderateastrogliosissomeastrocytesdisplayenlargementoftheirbody and extensions. There is no extensive reorganization of the tissue structure. Thisformofastrogliosis isassociatedwithmild,non-concussion traumaandviral infections.Withresolutionoftheinjury,astrocytesreturntotheirnormalstate.
Incontrast,severediffuseastrogliosisisfoundsurroundinglargefocallesionsandinfections and in those brain areas experiencing neuron deterioration. The volume ofastrocytebodiesandmembraneextensionsenlarges.Astrocyteextensionsspreadbeyondtheir normal domain and overlap those of their neighbors. This type of astrogliosisproduceslastingreorganizationofthetissue,butitdoesnotproducescartissue(Figure6-7).
Figure 6-7: High magnification photomicrograph showing reactive astrocytes restructuring tissue in this image ofischemicbrain.Photomicrograph:©Nephron
Astrocyte response to highly destructive insults to the brain including tissuepenetrationbyaforeignobject,blockageofbloodflowandtorntissueincludesallofthefeaturesofseverediffuseastrogliosisplusformationofdense,compactglialscars.Scarsform along the edges of tissue destruction. Astrocytes interact with oligodendrocytes,

microglia and cells of the meninges to deposit dense collagen. The deposited materialcontainsmolecularsignalstoinhibitneuronregenerationwithinthedamagedarea.Inthisformofastrogliosis,structuralchangesinthebraintissuelastlongaftertheinjuredsectionisenclosed.
AMOEBOIDMICROGLIA
Surveying microglia responds to mechanical brain injury by changing to itsmacrophage-like form.Tornbrain tissue releasesATP fromneurons andother damagedcells. Surveying microglia travels toward released ATP and then makes an executivedecision whether neurons may be salvaged or not. If the neurons cannot be repaired,microgliaassumesitsmacrophage-likeform,releaseslethalcompoundstofinishthekillandremovesdebrisbyengulfingit.
For salvageable neurons, microglia releases factors to assist in their repair andrecruits oligodendrocyte progenitor cells. The restoration of myelin to damaged axonscannot be performed by existing mature oligodendrocytes. Instead oligodendrocyteprogenitor cells, distributed throughout the brain, must be persuaded to mature. Thetransitionofoligodendrocyteprogenitorcellsintothematurationsequenceappearstobeinitiatedbyfactorssecretedbyactivatedastrocytes andmicroglia thatdraw them to thedamagedsite.
INFLAMMATIONANDINFECTIONSurveyingmicrogliaisthefirstrespondertoinfectioninthebrain.Whenmicroglia
detectspathogenswithitsmembranereceptors,whichresemblethereceptorsonimmunecells outside the brain, it becomes amoeboid and it multiplies. Then, likemacrophage,microglia engulfs and destroys the pathogen. Pathogen-activated microglia releasesnitrousoxide,chargedoxygenmolecules,pro-inflammatorycytokines,proteindestroyingenzymesandtoxiclevelsofglutamate.Activatedmicrogliaalsointerfereswithastrocyteuptakeofglutamate,furtherincreasingdestructiveglutamateinthetissue.
Inflammation in response to viral and bacterial infection in the central nervoussystem is different than classical inflammation elsewhere in thebody. It doesnot resultfrom the entry of immune cells from blood. It does not produce heat or edema in thetissue. Rather, activatedmicroglia and reactive astrocytes producemolecules similar tothosereleasedbyimmunecellsoutsidethebraintodestroythepathogen(Figure6-8).
Astrocytes make and secrete molecules with either pro- or anti- inflammatorypotentialdependinguponthebrain’ssituation.Inresponsetopathogentheysecretepro-inflammatorymoleculestoassistmicrogliaindestructionofthepathogen.Astrocytescanbeinfectedbyvirusesandactasreservoirforseveralviralinfections.Thepreciseroleofastrocytesinviralinfectionremainsunclear.

Figure 6-8: Photomicrograph of brain tissue stained with hematoxylin and eosin showing viral encephalitisinflammationonrightandnormalbraintissueonleft.Noticethelargeincreaseindarkstainingcellnucleiontheright.Photomicrograph:©vetpathologist
Virus also infects and replicates in microglia. Virus-infected microglia andastrocytes producemolecules toxic to neurons. Death of neurons during viral infectionappearstoresultfromanerroneousexchangeofsignalmoleculesbetweenastrocytesandmicrogliaingrayandwhitematter.
The description, inflammation, is expanded in the central nervous system toinclude diseases not related to infection. In the brain, inflammation produced bydegenerativediseasesisnotpartofahealingmethodbutratherachronicmalfunctionofanormalprotectivemechanism.Therewillbeadditionaldiscussionoftheinflammationofdegenerativebraindisordersinthefinalchapterofthisbook,Chapter8,“WhenItAllGoesWrong—Alzheimer’sDementia.”
SUMMARYCHAPTER6Radial cells of the embryo mature into neurons, oligodendrocytes andastrocytes
The portion of radial cells that fails to completelymature during embryonicdevelopmentpersistsinadultbrainasneuralstemcells
Neuralstemcellsdevelopintoreplacementneuronsandnewoligodendrocytes
Astrocytespresentatbirthsurvivemostofalifetime
Neural stemcells capable ofmaturing into oligodendrocytes exist throughouttheadultbrain
Microgliadoesnotoriginate from radial stemcells but rathermigrates to theneural plate from the extra-embryonic yolk sac prior to blood formation inbone
Astrocytestilethebraininanorderedarrangementwithnoregionofthebrainleftuncovered
Astrocytes, connected at the tips of their long extensions by gap junctions,createlargecellularwebs

Due to the complexity of the human brain, oligodendrocytes myelination ofneuronaxonscontinuesintothe5thand6thdecadeoflife
Eachsurveyingmicrogliamonitorstheareaaroundoneneuron
Fibrous astrocytes monitor and maintain steady ion concentrations in theinterstitialfluidaroundaxons
Protoplasmic astrocytes remove glutamate from neuron synapses and returnglutaminetotheneuronforconversionbacktoglutamate
Oligodendrocytes myelination of neuron axons insulates, speeds actionpotentials, provides growth factors and affects synapses on axons and axoncollateralformation
Microgliaexpresses receptors formostneurotransmitterspermittingmicrogliatomonitorneurotransmitterreleaseatallbrainsynapses
Braincellsdependonglucosefortheirprimaryenergysource
Thestorageformofglucose,glycogen,isonlyfoundinastrocytes
Allcholesterolinbrainismadeinthebrain
Brain cholesterol is critical to synapse formation, dendrite formation, axonelongationandisamajorcomponentofmyelin
Theresponseofastrocytestobraininjuryisnamedreactiveastrogliosis
Activatedmicrogliainitsmacrophage-likeformeliminatesdamagedneurons
Thefirstrespondertopathogeninthebrainismicroglia
Astrocytes secrete pro-inflammatory molecules to assist microglia indestructionofthepathogen
Inflammationinresponsetoviralandbacterialinfectioninthecentralnervoussystemisproducedbypro-inflammatorymoleculessecretedbyastrocytesandmicroglia

[7]
Brain’sInfrastructureforMemoryandLanguagePSYCHOLOGY STUDIES DISCOVERED many clues about how human
memoryworksoverthepast40years.Yet,mechanismsessentialforconsciousnessandasense of self remain unclear. When reading this chapter, keep in mind much of thepresentationisstilltheory.Onewaytothinkaboutdescriptionsofhowabrainworksistovisualize theparticulars as a fuzzy snapshotwith too fewpixels.Contemporary sciencerecognizes some fundamental truths about how the brainworks, but a complete pictureremainselusive.
Neuroscientists agree on the general details of how neurons communicate witheach other.But, few undisputed facts support themany theories of how neurons createsomethingasintangibleasthoughts.Bothgeneticsandaperson’senvironmentcontributetoself-awareness,howeverlittleisknownabouthowthebrain’sinfrastructuresupportsanindividual’sperceptionoftheworld.
Thedogmathattheadulthumanbraincannotreplaceitsneuronswasreversedin1998byevidenceofneurogenesisinthehippocampusofacancerpatient.Incorporationofemergingknowledgeofneurogenesisandplasticityofneuronconnectionsintotheoriesofhuman awareness requires reinterpretation of older data. Also, the part played by thebrain’sglialcellsinneuronsignalingisbeginningtobeappreciated.Humanmemorymaynotbetheexclusivedomainofneuronsworkinginisolation.
Modern speculation occasionally proposes brains operate like computers. Yet,contemporary studies of the central nervous system imply brains work different thantypical computers.Most computers calculate in a linear sequence solvingproblemsonestep at a time. Brains solve multiple problems and engage in complex environmentalanalysisall at the same time.Theydo thiswith littleerror inawatery structure tensofmillionsoftimesslowerthanstate-of-the-artdigitalcomputers.
Inbrainmodelsofthelastcentury,neuronswereimaginedtosignalinasequential‘on’and‘off’manner.However,newermodelsofneuronsignaling,whicharebaseduponemerging evidence from physiology and psychology, include interconnected processingunitsoperatinginparalleltogenerategradedandvariablesignals.Withparalleloperatingmodels,asmallchangeinoneunitadjustsalotofinformationinthesystematthesametime. Designers of new computers are working to copy the brain’s parallel operatingorganizationintotheirsystems.
Thischapterpresentssomeofthemajorhypothesesbuiltaroundexistingevidenceof the brain’s mechanism for forming memories. Most of the neuroscience discussedcomes from investigations conducted in the last 15 years andwas obtainedwith newernon-invasive measuring devices developed for human medical testing. Languageacquisition,oneformofmemory,ispresentedasanexampleofthecomplexityofhumanmemory systems. Present understanding of language acquisition permits relevant, ifincomplete,discussionoftheneuroncircuitsthatprocesssensoryinformationtocreatea

formofmemoryuniquetohumans.
This chapter first considers neuron coding of information, anatomic patterns ofneuron connections andnon-invasive tools used to link activity of humanbrain circuitswithspecifictasks.Thendataobtainedwiththesetoolswillbeusedtooutlinerecentideasaboutmemoryformationandlanguageacquisition.
INFORMATIONFLOW
NEURONCODESTheoriginalhypothesis stating that signalingbetweenbrainneurons iscodedby
stringsof ‘on’and ‘off’ inputemergedduringstudiesof sensory input to thebrain.Thepresenceofexternalstimuliincludinglight,sound,taste,smellandtouchcauseneuronsofsensory organs to fire sequences of action potential in various temporal patterns.Manystudiesconclude thepatternofactionpotentialsentering thebrainfromasensoryorganencodes information about the sensory stimulus. An extension of those conclusionsassumesneuronsignalingwithinthebrainiscodedinasimilarway.
Analysis of sensory action potential patterns uses statistical methods andprobability theory.An intense debate continues aboutwhether sensory neurons use ratecodingortemporalcoding.Ratecodingofactionpotentialpatternsassumesinformationaboutthesensorystimulusiscontainedinthefiringrateoftheneuron.Temporalcodingassumes precise timing of each action potential in the train contains the information.Acquiring data that separates these theories is a challenge because individual neuronsdisplaybackgroundfiringactivityequivalenttonoise.
Incontrast,newertheoriesofinformationprocessingwithinthebrainhypothesizeaction potential coding is carried out by populations of neuronsworking together. Thismodelclaimsgroupsofneurons,neuroncircuits,performbetter than individualneuronsbecausenoise in thegroupgetsaveragedout.This is frequently referred toasensemblecoding.
Otherhypothesesclaimbalancebetweentheaverageamountsofdepolarizingandhyperpolarizingsynapticcurrents isabasicformofbrain informationprocessing.Thesehypothesesareconsistentwiththerhythmsinbrainelectricalactivityobservedwithscalpelectrodes. If the balance model is correct, any shift in the equilibrium toward netexcitationornet inhibitionmayallowmultiple sensory inputs tobeclarifiedasoneall-inclusivemessageatcerebralcorticalneurons.Possibly,analignmentofbrainelectricalrhythmswithensemblecodingcontributestoquickdistributionofinformationthroughoutthebrain.
RHYTHMICBRAINWAVES
Brainneuronsarebelievednever tooperate in isolation.Rather, theyperformasmembersoforganizedgroups labeledneuroncircuits.Eachneuron circuit is thought toprocessspecifickindsofinformation.
Theflowofactionpotentials,movingfromneurontoneuronbecauseofsynapticactivity,isoftencomparedtoelectricalcurrentinhouseholdcircuits.Electricalengineersdescribeacircuitasanetworkconsistingofaclosedloopthatprovidesareturnpathfor

anelectricalcurrent,oftenelectronsflowingthroughawire.
The following discussion of brain activity will use the term ‘circuit’ tomean aneuron path within the brain where ion flow creates a pattern of action potentials anddendriticspikes.Unlikehouseholdcircuitsforelectricity,braincircuitsdonotformclosedloopswhereactionpotentialstravelbacktotheirpointsofinitiation.
Neurons come in many sizes and shapes. Neuron circuits incorporate the largeneurons described in Chapter 4, “Neurons—How They Make Electricity” plus manysmaller neurons identified as interneurons. Interneuron axons and dendrites often lookalikebecauseoftheirsmallsize.Anothernameforinterneuronsisassociationneuronsorrelayneurons.Interneuronscontributegreatlytotheobservedelectricaloscillationsknownasbrainwaves.
Interneurons influence the output of neuron circuits because they regulate theactivity of the larger neurons. They fine-tune the characteristics of a brain’s ongoingelectrical activity. Most interneurons release a neurotransmitter that hyperpolarizes thepresynapticand/orpostsynaptic transmembranepotentialatneuronsynapses.Membranehyperpolarization by inhibitory interneurons slows the flow of action potentials, andthereforecurrent,throughaneuroncircuit.
The total ongoing neuron activity in the brain sums into rhythmic patterns.Sometimes the pattern is the result of synchrony in action potentials arriving at thesynapses of a group of neurons. When action potentials of large groups of neuronssynchronizetheycauselargeamplitudefluctuationsofthelocalfieldpotential.Thelocalfield potential is created by opening of membrane ion channels at synapses and theresulting net flow of ions into neurons from the interstitial fluid and out of neuroncytoplasmintotheinterstitialfluid.
Fluctuationsof local fieldpotentialsalsoappearbecauseofconnectionsbetweendistant brain areas. Some brain areas form feedback circuits by linking together in acircularpattern.Timingofactionpotentialrepetitionsinvariouspartsofafeedbackloop’scircuitscausesoscillationsinregionalfieldpotentials.Electrodesplacedonthesurfaceoftheheadmeasureshiftsinlocalfieldpotentials.
Voltagemeasurements,thedifferencesinthefieldpotentialbetweenpairsofscalpelectrodes, produce an oscillatory recording over time named an electroencephalogram(EEG). An EEG measures the synchronous activity of millions of neurons at a time(Figure7-1).

Figure7-1:Simulationofbrainactivity showing10Hzoscillations.Neuronspiking,actionpotentialsarrivingat thepresynaptic compartment, is simulated by a rate-modulated Poisson process (upper panel). Local field potential issimulatedasthesumionflowcausedbytheneuronspiking,representingthemeanactivityofalargenumberofneurons(lowerpanel).Illustration:©TjeerdB
MAPPINGTHEBRAIN’SNEURONS
HUMANCONNECTOME
The termconnectome is anotherof science’smade-upnames.The ‘connect’partalludes tomapping all the neuron connections throughout the brain. The ‘-ome’ part isused to suggest a comparison to the genome project thatmapped all of an individual’sgenes.Someliketocomparethebrainconnectometoawiringdiagram.Onamicroscaleitdescribesthecharacteristicsofneuronconnectionssurroundingasynapse.Onalargerscale the brain connectome is a description of neuron connectivity between distantsectionsofthebrain.
Defining the entire connectome of the human brain is not feasible at present.However, data collected from tiny volumes of brain tissue using automated electronmicroscopy are confirming the complexity of neuron connections.Brain tissue is slicedinto thin serial sections before processing for electron microscopy. The microscopysections, reconstructed after analysis, produce a three dimensionalmodel of neuronconnectionsinthetissue.
Automated electron microscopy data confirm cells around brain synapses packtight together. Yet, the presence of synapses cannot be inferred by where axons anddendrites intersect. How individual variation will affect the outcome of ongoingconnectomeprojectsremainsuncertain.But,automatedelectronmicroscopybrainmaps,atrillion times finer than thoseobtainablewithexternal imaging techniques,addessentialinformation.

DIAGRAMSOFTHECEREBRALCORTEXThe first brain region mapped by anatomists of the nineteenth century is the
cerebral cortex. Based upon expansion of brain regions inmammalian species throughevolution,themostrecentlydevelopedpartofthecerebralcortexisnamedtheneo-cortex.Theancientpartofthecerebralcortexisthehippocampus(Figure7-2).
Figure7-2:Brainmapofamacaquemonkeydisplayingthedarkouterbandofneo-cortex.Theoldestphylogeneticareaofthecortex,thehippocampus,isthecircledregionintheredboxatthebottom.Illustration:©brainmaps.org
Inhumansasinmonkeys,thehippocampusliesundertheneo-cortexinthemedialtemporal lobe. It contains five regions, dentate gyrus, CA1, CA2, CA3 and CA4. Thehippocampus codes memory into neuron signaling patterns. It consolidates short-termmemoryintolong-termmemoryandguidesaperson’snavigationofspace.
Anatomistsofthelate19thcenturywereintriguedbythelayeredarrangementofcortexneuronsdrawnbySantiagoRamónyCajalandothers(Figure7-3).

Figure7-3:Theexterior surfacesof the cortical sections are locatedat the topof the illustration.Left:Nissl stainedneuronbodies of the visual cortex of an adult human.Middle:Nissl-stainedmotor cortex of an adult human.Right:Golgi stained cortex of a 1½month old infant displaying neuron bodies, dendrites and axons of a random1-3%ofneuronspresent.Noticehow thepatternofneuronbodies,dark spots, varies.Drawings:BySantiagoRamónyCajalfoundinthebook“ComparativeStudyoftheSensoryAreasoftheHumanCortex.”ThisworkisinthepublicdomainintheUnitedStates.
The large neurons of the neo-cortex occupy six layers from exterior surface tointerior surface. Neurons from various layers connect to form small microcircuits incorticalcolumns.Thehumancerebralcortexisonly2to4millimetersthick.Inspiteofitssize this small band of tissue plays a critical role in language, memory, attention,awareness,thoughtandconsciousness.
Anatomists noticed neuron bodies and neuron axons display diverse patterns ofarrangement invarious regionsof theneo-cortex (Figure7-3).Theconfigurations showmarkeddifferencesinneuronorganizationanddefiniteborderswhereonepatternchangesto another design. Each area defined by one of the patterns, when stimulated withelectricalcurrent,producesadistinctresponse.
TheGermananatomist,KorbinianBrodmann,wasthefirsttomapneuronpatternsoftheentireneo-cortex.HeusedNisslstaintolocateneuronbodies.Brodmannnumberedhisareas1through52.Becauseofhisinterestinevolutionoftheneo-cortex,hestudiedtheneo-cortexofhumansinpostmortemspecimensandcomparedpatternshefoundwiththoseofeightotherspecies.Hediscoveredsomebrainareas,Brodmannareas12,13,14-16and48-51inlowermammals,areabsentinhumans.

Many of Brodmann’s areas were renamed when evidence of their purposeemerged.Forexample,Brodmannarea17isnowrecognizedastheprimaryvisualcortexandBrodmannarea4istheprimarymotorcortex.ModerntechniquessubdividemanyofBrodmann’s areas based upon smaller neuron groups with distinctive responses toelectricalstimulation.Althoughrefinedandrenamedforoveracentury,Brodmann’sareasremainthebestknownandmostoftencitedwhendiscussingthecerebralcortex(Figure7-4).
Figure7-4:NumbersrefertoareaswhereBrodmanndetectedapatternofneuronbodieswithcharacteristicsdifferentthanthoseofthesurroundingarea.ColoredareascorrelateBrodmannareaswithlaterdatarevealingtheresponsibilityofparticularneurongroups.Illustration:©OpenStaxCollege
Underthecerebralcortexdeepinwhitematter,neuronbodiescollectintogroups.Eachgroupiscalledanucleus.Inpreservedbrain,nucleiappearasislandsofgraymatterwithin white matter. Neurons included in each brain nucleus form connections withneuronsinothernucleiandwithneuronsinthecerebralcortex.
Largerbrainnucleipossessdescriptivenamesbecausetheirroleinbrainactivityisextensivelydescribed.Modernmethodsroutinelyaddadditionalsmallnucleitothehumanbrain map. The purpose of most of the multitude of small nuclei once too little to benoticedisstillbeinginvestigated.
NEURONAXONTRACTSA traditionalmethod formapping neuron connections between brain nuclei and
cortical areas relies upon injection of a labeling agent, often a dye. This technique isdescribedas tract tracing.Tract tracing is considered thegold standard for charting thepathofneuronsthroughbraintissue.
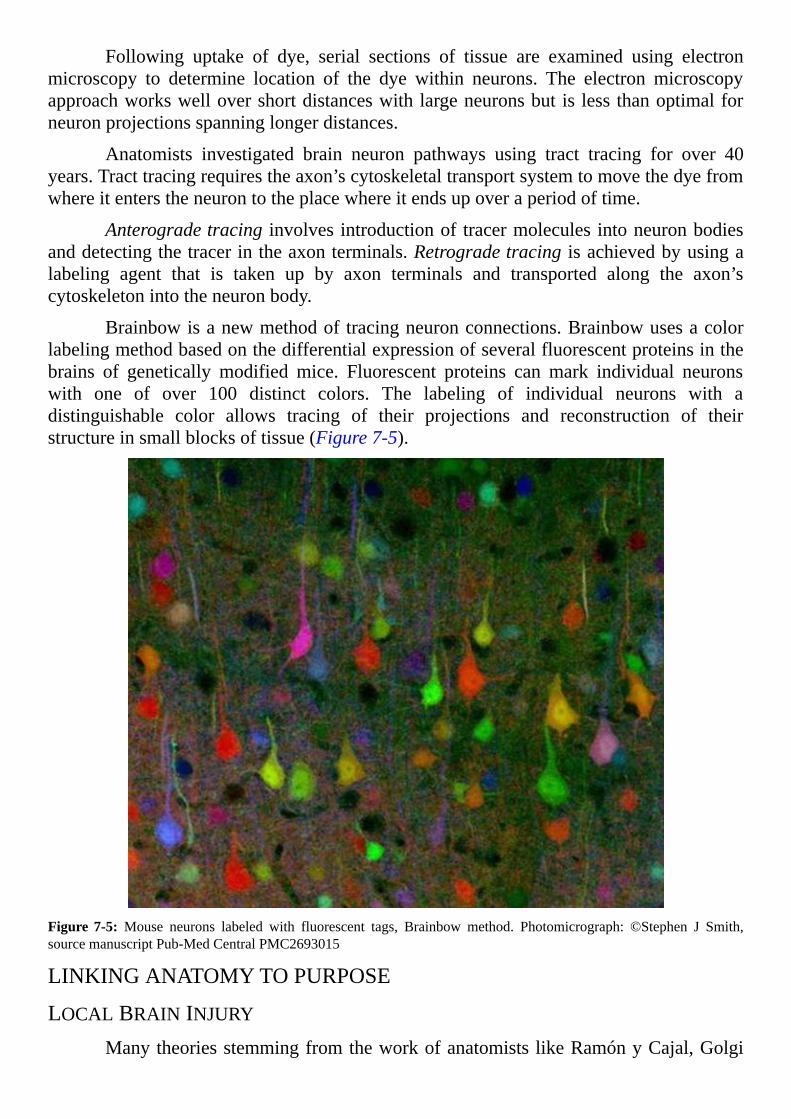
Following uptake of dye, serial sections of tissue are examined using electronmicroscopy to determine location of the dye within neurons. The electron microscopyapproachworkswellovershortdistanceswithlargeneuronsbut is lessthanoptimalforneuronprojectionsspanninglongerdistances.
Anatomists investigated brain neuron pathways using tract tracing for over 40years.Tracttracingrequirestheaxon’scytoskeletaltransportsystemtomovethedyefromwhereitenterstheneurontotheplacewhereitendsupoveraperiodoftime.
Anterogradetracing involves introductionof tracermolecules intoneuronbodiesanddetectingthetracerintheaxonterminals.Retrogradetracing isachievedbyusingalabeling agent that is taken up by axon terminals and transported along the axon’scytoskeletonintotheneuronbody.
Brainbowisanewmethodoftracingneuronconnections.Brainbowusesacolorlabelingmethodbasedonthedifferentialexpressionofseveralfluorescentproteinsinthebrains of genetically modified mice. Fluorescent proteins can mark individual neuronswith one of over 100 distinct colors. The labeling of individual neurons with adistinguishable color allows tracing of their projections and reconstruction of theirstructureinsmallblocksoftissue(Figure7-5).
Figure 7-5: Mouse neurons labeled with fluorescent tags, Brainbow method. Photomicrograph: ©Stephen J Smith,sourcemanuscriptPub-MedCentralPMC2693015
LINKINGANATOMYTOPURPOSE
LOCALBRAININJURYManytheoriesstemmingfromtheworkofanatomistslikeRamónyCajal,Golgi

andBrodmann speculate that variation in neuron structure, neuron location in the brainandneuronconnectivityregulatedistinctcharacteristicsofbrainperformance.
Confirmation that intellectual pursuits align with discrete regions of the humanbrainemergedfromexaminationofpatientsintheclinicwhosufferedlocalbraindamage.For example, damage to specific brain areas is able to cause a general loss ofunderstanding, deficits in the ability to use language or a lack of ability to control ofemotion. Injury to the hippocampus affects the formation and duration ofmemories oflife’sexperiencescausingvariousdegreesofamnesia.
Whilepatientswithbraininjuriescontributeagreatdealtounderstandingofbrainmechanisms, it should not be assumed all aspects of amissing capability aremanagedsolely by the areawith obvious damage.Normal activity requires dynamic interactionsbetween several brain regions. A multistep process can be interrupted by removing orrestricting a single step. Information carried by neuron circuits is splintered intomanysmallpartsasitprogressesthroughthebrain.Multiplebitsofinformationaboutasingleeventdistributeacrossmultiplebrainregions.Whythiscomplexsystemisanadvantageisnotknown.
EVENT-RELATEDPOTENTIALSModernhumanbrainstudiesapplynon-invasivetechniquestoexploreinformation
handling by neuron circuits. Four non-invasive neuroimaging techniques dominate thisresearch.First,thereisEvent-RelatedPotentials(ERPs),partofanelectroencephalogram(EEG).WithERPs theelectricalactivitymeasuredbyscalpelectrodes is time locked topresentation of a sensory stimulus, a picture or spoken word. ERPs are exceptionallysensitivetovoltagechangesduetoneuronactivityinthecerebralcortex.Thismethodisoftenusedtostudytheregionsofthecerebralcortexdedicatedtolearninglanguage.
MAGNETICRESONANCEIMAGING
Asecondformofbrainimagingusedforhumanstudiesisavariationofmagneticresonance imaging (MRI)namedfunctionalMRI(fMRI).AbasicMRIscanproducesastatic anatomical picture of large brain structures in virtual slices of living tissue. Thebasic MRI serves to identify large brain structures associated with neuron activitymeasured by fMRI. fMRI is often employed for research andMRI is used for routineclinicalevaluationofbrainpathology.
Resonance in physics is the tendency of a system to oscillate with greateramplitudeatcertainfrequencies.MostmedicalapplicationsofMRIuseastrongmagnettosetupanefficientresonantfrequency.Astheresonanceofmagneticallyexcitedhydrogenatomsinthebraindecays,aradiofrequencysignalisemitted.
Dependingupontimingoftheresonanceinducedbythemagnet,andoftheradiofrequencysignalmeasurements,imagesareclassifiedasT1andT2.T1andT2acquiredimages look different. Tissue areas with high water content, a lot of hydrogen atoms,appearblackinT1images(Figure7-10)andwhiteinT2images(Figure7-6).Comparetheareaoftheventriclesfilledwithcerebrospinalfluidinthetwofigures.
InT1imageswhitematterislightgray,graymatterisgrayandcerebrospinalfluidappearsblack.T2imagesshowcerebrospinalfluidandedemainthetissueaswhiteareas
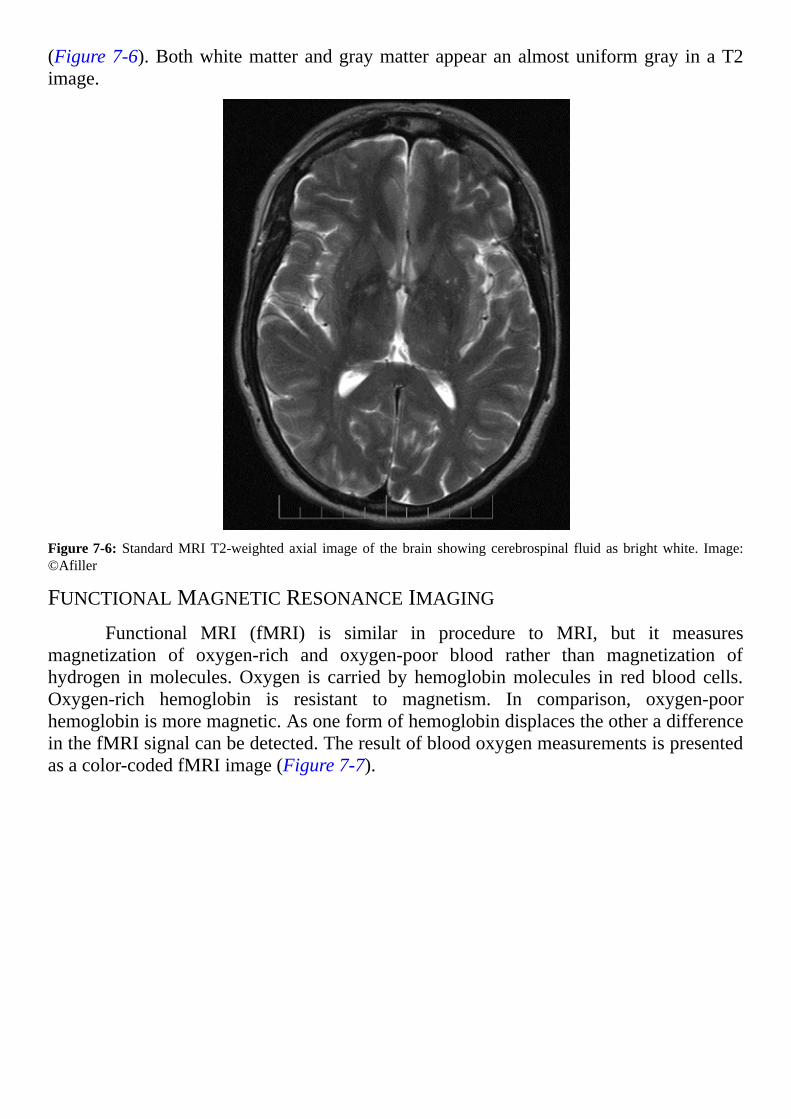
(Figure7-6).Bothwhitematterandgraymatterappearanalmostuniformgray inaT2image.
Figure7-6:StandardMRIT2-weightedaxial imageof thebrain showingcerebrospinal fluidasbrightwhite. Image:©Afiller
FUNCTIONALMAGNETICRESONANCEIMAGING
Functional MRI (fMRI) is similar in procedure to MRI, but it measuresmagnetization of oxygen-rich and oxygen-poor blood rather than magnetization ofhydrogen inmolecules.Oxygen is carried by hemoglobinmolecules in red blood cells.Oxygen-rich hemoglobin is resistant to magnetism. In comparison, oxygen-poorhemoglobinismoremagnetic.AsoneformofhemoglobindisplacestheotheradifferenceinthefMRIsignalcanbedetected.Theresultofbloodoxygenmeasurementsispresentedasacolor-codedfMRIimage(Figure7-7).

Figure 7-7: An fMRI image with yellow/orange areas showing brain areas with increased blood flow. Image:©OpenStaxCollege
Neuron activity requires a large amount of energy. Creation of energy richmoleculesfromglucoseusesalotofoxygenextractedfromthehemoglobinofredbloodcells.Astrocytesdeterminetherateoflocalbrainbloodflow.Whenneuronsincreasetheirsignalingactivity,surroundingastrocytessignalforincreasedlocalbloodflowtoinsureanadequateoxygensupply.Oxygenrichblooddisplacesoxygen-depletedblood inabout2seconds.
The slow dynamics of the blood flow response, seconds, compared to neuronresponse in milliseconds means a signal detected by fMRI is a summation of neuronactivityinsmalltissueareas.Thepurposeofthiskindofdataistocorrelatebrainneuronactivitywith a task performed by the tested subject.Multiple brain areas in use duringrecognitionofobjects, emotional statesor listening to speechcanbeevaluated for theirdegreeofassociationwithfMRI.fMRIproducesdatasetsfromupto100,000locationsatthe same time.Thedataprovide three-dimensionalmapswithhigh spatial resolutionofareasworkingtogetheracrossthewholebrain.
MAGNETOENCEPHALOGRAPHY
A third type of non-invasive imaging used in brain research ismagnetoencephalography (MEG). Breaking this long scientific term into its root partsproduces the approximate translation ‘mappingmagnetic fields in the head’. TheMEGsignalmeasuresneuroncurrentrather than theoxygenationstateofhemoglobin inbrainbloodvessels.MEGrecordsmagneticfieldsproducedbycurrentsflowingthroughbraincircuitsusingsophisticatedsensorsnamedmagnetometers.Itallowspreciselocalizationofthe neuron currents responsible for the magnetic fields. MEG resolves events with aprecisionof10milliseconds,or less.Magnetometers reside in ahelmetplacedover thesubject’shead(Figure7-8).

Figure7-8:PersonundergoingaMEG.PictureisfromUnitedStatesNationalInstituteofMentalHealth.ThisimageisinthepublicdomainintheUnitedStates.
Sometimes MEG is combined with fMRI when mapping a response to anexperimentalcondition.Thedegreeofagreementbetweendatacollectedwith these twotechniques varies depending upon the complexity of the neuron circuitry in brain areasrespondingtothestimulus.
FUNCTIONALNEARINFRAREDSPECTROSCOPYThefourthmethod,functionalnearinfraredspectroscopy(fNIRS),takesadvantage
of the transparency of skin, bone and brain tissue to light in the near infra-red lightspectrum, 700-900 nanometers. Most of the thermal radiation emitted by objects nearambient temperature is in the infra-red spectrum. This light energy is invisible to theunaided human eye. Data obtained with fNIRS are similar to data produced by fMRIbecausetheyalsomeasurechangesinbloodoxygenationoftissue.
AnadvantagefNIRSisitrequiresasimplehelmetdeviceandisthereforefarlessexpensive thanfMRI.Adisadvantageof fNIRSisnear infrared lightcannot reachbrainareas deeper than about 4 centimeters. Its use in humans is limited to studies of thecerebral cortical neurons. Another disadvantage of fNIRS is its lack of anatomicalreferencepointsfortheobservedchangesinbloodoxygenation.
HUMANMEMORY

CATEGORIESOFMEMORY
Memorymolds consciousness,making eachmind unique. Patientswho sufferedbrain damage to their hippocampus provided the first definitive knowledge that humanmemory is stored in, and recalled from separate brain regions. These patients lost theirability to form and recall particular types of memory, while retaining other types ofmemory.Individuallossesdependeduponthepreciselocationoftheinjury.
Modern non-invasive brain imaging studies are producing volumes of datadefining neuron circuits dedicated to storage and recall of human memory. There aremultiple forms of humanmemory and different neuron pathways support each type. Inbrief,themajordivisionsofmemoryarelong-termmemoryandshort-termmemory.
Most systems for cataloging the defining characteristics of human memoryoriginated in the field of psychology. Psychologists report long-term memory can bedividedintotwoseparatecategories,explicit/declarativememoryand implicit/proceduralmemory.Short-termmemory,alsoknownasworkingmemory,ismorecomplexthanlong-termmemory.
EXPLICIT/DECLARATIVEMEMORY
Explicit memory includes memory available at will. Explicit memories fit intothree categories: episodic memory, semantic memory and autobiographical memory.Episodicmemoryrecallsspecificevents,situationsandpersonalexperience.Forexample,anepisodicmemorymaybeofaperson,ofaperson’snameortheplaceandtimespentwithaparticularperson.
Incontrast,semanticmemoryislimitedtofactslikethenumberofhoursinadayorthemeaningofaword.Autobiographicalmemoryissimilartoepisodicmemorybutisrestrictedtothoseeventsinaperson’sownlifehistory.Memoryofeventscorrespondingtoaperson’scollegegraduationorgettingmarriedisautobiographicalmemory.
IMPLICIT/PROCEDURALMEMORY
Implicit memory, unlike explicit memory, is below the level of consciousawareness. Implicit memory is an unintentional memory formed as an experience isrepeated.Implicitmemoryguidesperformanceofapracticedactivity.It istheautomaticformofmemoryusedwhenreadingthisbook.Attentiontothedetailsofhowtoreadisnot required because an implicit memory of reading is already present. In anotherexample, implicit memory recognizes that music belongs to particular categories likeclassical,country,bluesorrock.
WORKING/SHORT-TERMMEMORY
Workingmemory isnotpartof long-termmemory.Ratherworkingmemory isadynamicformofmemorycombiningmanythingsfrompastlearningtogetherwithpresentexperiences.Itdrawsonpreviousexperiencestomanagethepresent.
Workingmemoryalso includesresistingdistractionwhenprocessing informationforstorageaslong-termmemory.Forexample,whenstudentshearinstructionsforhowtoproceedwithanexam,workingmemorystoresdetailsof theexamprocess in long-term
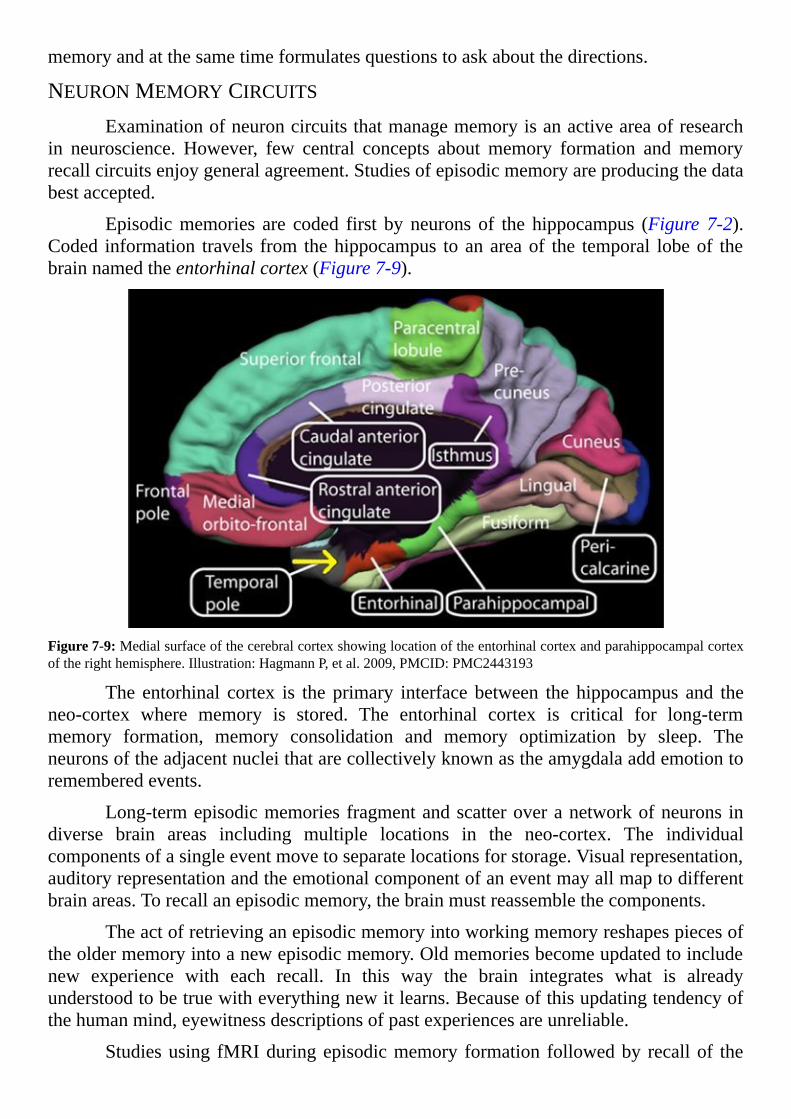
memoryandatthesametimeformulatesquestionstoaskaboutthedirections.
NEURONMEMORYCIRCUITSExaminationofneuroncircuitsthatmanagememoryisanactiveareaofresearch
in neuroscience. However, few central concepts about memory formation andmemoryrecallcircuitsenjoygeneralagreement.Studiesofepisodicmemoryareproducingthedatabestaccepted.
Episodicmemories are coded first by neurons of the hippocampus (Figure 7-2).Coded information travels from thehippocampus toanareaof the temporal lobeof thebrainnamedtheentorhinalcortex(Figure7-9).
Figure7-9:Medialsurfaceofthecerebralcortexshowinglocationoftheentorhinalcortexandparahippocampalcortexoftherighthemisphere.Illustration:HagmannP,etal.2009,PMCID:PMC2443193
The entorhinal cortex is the primary interface between the hippocampus and theneo-cortex where memory is stored. The entorhinal cortex is critical for long-termmemory formation, memory consolidation and memory optimization by sleep. Theneuronsoftheadjacentnucleithatarecollectivelyknownastheamygdalaaddemotiontorememberedevents.
Long-termepisodicmemories fragmentandscatteroveranetworkofneurons indiverse brain areas including multiple locations in the neo-cortex. The individualcomponentsofasingleeventmovetoseparatelocationsforstorage.Visualrepresentation,auditoryrepresentationandtheemotionalcomponentofaneventmayallmaptodifferentbrainareas.Torecallanepisodicmemory,thebrainmustreassemblethecomponents.
Theactofretrievinganepisodicmemoryintoworkingmemoryreshapespiecesoftheoldermemoryintoanewepisodicmemory.Oldmemoriesbecomeupdatedtoincludenew experience with each recall. In this way the brain integrates what is alreadyunderstoodtobetruewitheverythingnewitlearns.Becauseofthisupdatingtendencyofthehumanmind,eyewitnessdescriptionsofpastexperiencesareunreliable.
Studiesusing fMRIduringepisodicmemory formation followedby recallof the

memory discovered remembering reactivates the same neuron circuitry as the originalexperience.Thereisgeneralagreement,at leastforepisodicmemory, thatrecallfollowsthesameneuronpaththroughthebrainastheactualexperience,butinreverse.However,the number of details remembered about a previous event depends upon the amount ofattentionfocusedupontheeventwhenitoccurred.
Unlike episodicmemory, themain neuron circuit controlling implicit/proceduralmemory is a loopconnectingvariouspartsof theneo-cortex, thecerebellum, the spinalcord,thethalamusandthebasalganglia(Figure7-10).
Figure7-10:Traversesectionofthestriatum,thelargestofthebasalganglianuclei,labeledinredonT1MRIimage.MRI:©LindsayHanford,thisimageisreleasedtothepublicdomain.
Body movement is initiated by the neurons of the motor cortex in response toexecutive decisions made in the frontal cerebral cortex. The cerebellum fine-tunes theoutput of themotor neurons on theirway to the spinal cord.The neurons of themotorcortexsynapseon themotorneurons in thespinalcordsendingasignal to themuscles.Sensoryreceptorsdetectingpositionofthebodysendinformationaboutmusclepositionsbacktothespinalcord.Thespinalcordforwardspositioninformationtothebrain.
Allfeedbackfromoutsidethebrainabouttaskimplementationentersthebrainbywayofthespinalneurontractstothethalamus.Thethalamusrelaysfeedbackinformationfromthemusclestothecerebralcortex,cerebellumandbasalganglia.
The basal ganglia, a group of brain nuclei deep beneath the cortex, smooth outvoluntarymovementbycoordinatingthemovementwithconsciousmotivation.Thebasalgangliaaccomplish this through theirconnections to the thalamus, themotorneuronsofthe cortex, the brainstem and other brain nuclei. The unconscious memory of manyiterations of an activity becomes an implicit/procedural memory that is thought to bestoredasaneurontemplatewithinthecerebellum.
ANATOMICSTRUCTUREOFMEMORY

NEURONCONNECTIONSMemoryformationinthehumanbraininvolvesmorethanneuroncurrentmoving
through stationary neuron circuits. Dynamic anatomic reorganization of neuronconnectionsduringmemoryformationisessential.Neuroncircuitreorganizationincludesrestructuringofneuronensembleunits,remodelingofneuronsynapsesandreplacementofsomeneurons.Theseeventsareenhancedbysleep.
There is evidence for activity-dependent recruitment of neurons from reserveneuronpools toensemblecircuits.Reserveneuronpoolsexist insignalingensemblesofvertebratebrainsfromfrogstoprimates.Thereservepoolneuronsshareinputandoutputcircuitswiththecoreensemblebutexpressdifferentneurotransmittersandexertdifferenteffects on common targets. Incoming stimuli affect both the core and reserve neuronpools. The flexible composition of ensembles allows various combinations ofneurotransmitteroutputinresponsetoincomingsignals.
Neuronensemblespossessanadditionalmechanismforrespondingtovariationinmultiple sensory inputs of diverse intensity. The presumption asserting a neuron’sneurotransmitter never changes is incorrect. For example, neuroscience once thoughtdopamineneuronsalwaysreleaseddopamineandnorepinephrineneuronsalwaysreleasednorepinephrine.Todaythereisevidencethatneuronscanrespondtofluctuatingincomingelectricalactivitybychangingtheneurotransmitter theyreleaseandtheneurotransmitterreceptorstheydisplay.
Whitematterreorganizationisalsopartofmemoryformation.Modificationofaxonsinacircuitincludealterationinthenumberofaxonsinamemorypathway,changesinthediameteroftheaxonsandvariationinthedegreeofdensepacking,axonbranchingandtheamountofaxonmyelination.Theabilityofneuronstoadjusttheirpropertiesduringmemorycreationprovidesadynamicrangeofoptionsforoptimizingmemoryconstruction.
PRESERVATIONOFDENDRITICSPINESMemory programing begins at neuron synapses. Over time synapse structures
enlargeorshrinkdependinguponthenumberofincomingactionpotentials.Thissuggeststhe amount of sensory input is correlated with the functional capacity of neuronensembles. The more a pathway is used, the better it works. The process that allowssynapses to become stronger or weaker is called neuroplasticity. A discussion of theplasticity of neuron synapses is presented in Chapter 5, “Neuron synapses—ExcitatoryandInhibitory.”
Studies during the 1970s predicted synaptic plasticity. In those studies, smallgroups of brain neurons were activated with an electrical current from tiny electrodes.Investigators reported a phenomenon they named long-term potentiation (LTP). Long-termpotentiationisdescribedasanincreaseintheabilityofbrainsynapsestorespondtoneurotransmitter after receiving a rapid burst of signaling activity induced by externalelectrodes.Theabove-normalsensitivityofsynapsestoneurotransmitterpersistedinthesestudiesforminutestomanymonths.
The biologic mechanism supporting LTP remained unclear for a long time.
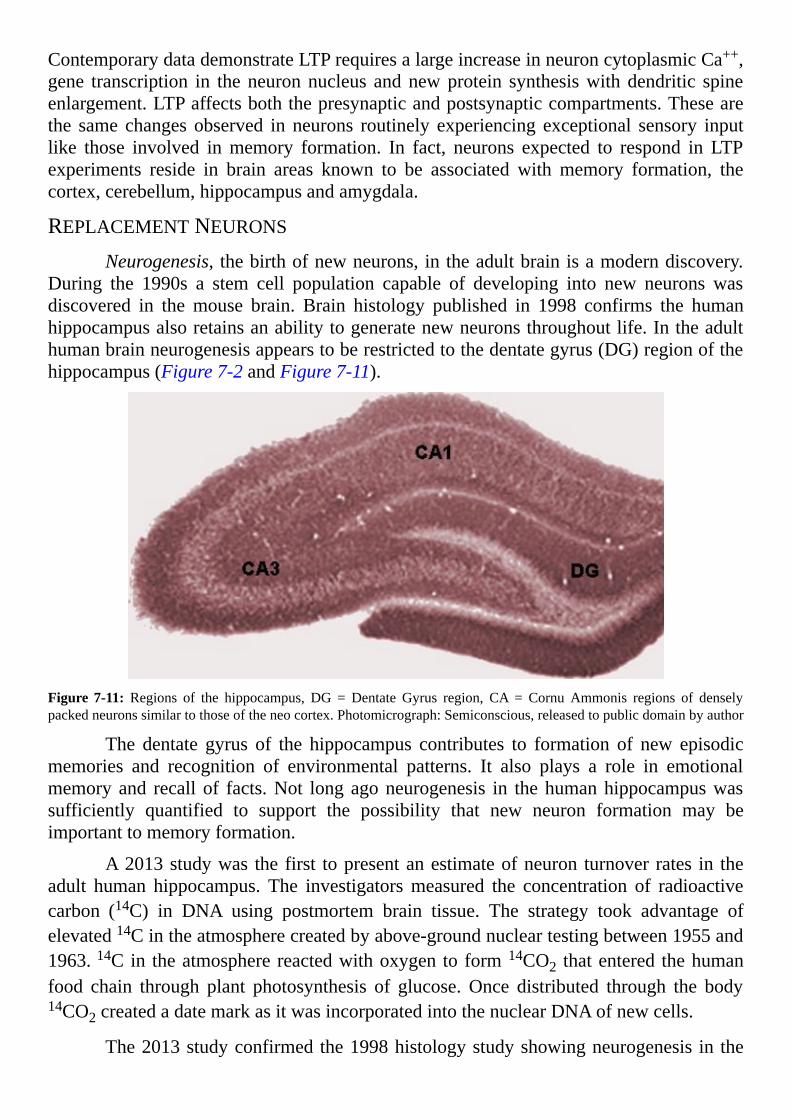
ContemporarydatademonstrateLTPrequiresalargeincreaseinneuroncytoplasmicCa++,gene transcription in theneuronnucleus andnewprotein synthesiswithdendritic spineenlargement.LTPaffectsboththepresynapticandpostsynapticcompartments.Thesearethe same changes observed in neurons routinely experiencing exceptional sensory inputlike those involved inmemory formation. In fact, neurons expected to respond in LTPexperiments reside in brain areas known to be associated with memory formation, thecortex,cerebellum,hippocampusandamygdala.
REPLACEMENTNEURONSNeurogenesis, thebirthofnewneurons,intheadultbrainisamoderndiscovery.
During the 1990s a stem cell population capable of developing into new neurons wasdiscovered in the mouse brain. Brain histology published in 1998 confirms the humanhippocampusalsoretainsanabilitytogeneratenewneuronsthroughoutlife.Intheadulthumanbrainneurogenesisappearstoberestrictedtothedentategyrus(DG)regionofthehippocampus(Figure7-2andFigure7-11).
Figure 7-11: Regions of the hippocampus, DG = Dentate Gyrus region, CA = Cornu Ammonis regions of denselypackedneuronssimilartothoseoftheneocortex.Photomicrograph:Semiconscious,releasedtopublicdomainbyauthor
The dentate gyrus of the hippocampus contributes to formation of new episodicmemories and recognition of environmental patterns. It also plays a role in emotionalmemoryand recall of facts.Not longagoneurogenesis in thehumanhippocampuswassufficiently quantified to support the possibility that new neuron formation may beimportanttomemoryformation.
A2013studywas the first topresentanestimateofneuron turnover rates in theadult human hippocampus. The investigatorsmeasured the concentration of radioactivecarbon (14C) in DNA using postmortem brain tissue. The strategy took advantage ofelevated14Cintheatmospherecreatedbyabove-groundnucleartestingbetween1955and1963.14Cin theatmospherereactedwithoxygen to form14CO2 thatentered thehumanfood chain through plant photosynthesis of glucose.Once distributed through the body14CO2createdadatemarkasitwasincorporatedintothenuclearDNAofnewcells.
The2013studyconfirmedthe1998histologystudyshowingneurogenesis in the

human hippocampus is restricted to the dentate gyrus. Because 14C is radioactive andemits a lowenergybeta particlewith a half-life of 5,700±40years, these investigatorswereabletodeterminetherateofneuronturnoverinthedentategyrus.Theyconcludedthe vast majority of human dentate gyrus neurons are replaced over a lifetime. Theirmathematicalmodelspredictamedianturnoverrateof1.75%peryear,correspondingtoabout 700 new neurons per day in each hippocampus, right and left hemisphere.Neurogenesiscontinueswithoutdeclineintoatleastthe5thdecadeoflifebasedupontheoldestindividualsinthestudy.
Thehalf-lifeofreplacementneuronsintheadulthumandentategyrusisabout7.1years.Thisis10timesshorterthanthehalf-lifeofnon-renewingneuronsintheremainderof the hippocampus. Studies in rodents, where dentate gyrus neurons also turnover,indicate adult-born hippocampus neurons demonstrate enhanced synaptic plasticity and,therefore,agreaterinfluenceonhippocampusmemoryformation.
A twopartmodelproposespattern separation,which insures similar experiencespersistasdistinctmemories, requiresreplacementhippocampusneuronsforcoding.Thesecondcomponentofthismodelmaintainsolderneuronsofthehippocampus,outsidethedentate gyrus, are devoted to pattern completion. Pattern completion is essential forreconstructionofastoredmemorybasedonpartialclues.
SLEEPREINFORCEMENTOFNEURONCONNECTIONSManyobservationalstudiesofhumanlearningconcludethereisapositiveeffectof
sleep on the consolidation of recent events into long-termmemory.However, empiricaldataaredifficulttocollectandproposedneuronmechanismsremaincontroversial.Studiesinvestigating neuron synapse adjustments when learning new skills report sleepdeprivationinhibitsrevisionofdendriticspinesynapses.
Imaging studies of the hippocampus report neurons there continue to be activeduring all phases of sleep. Neuron ensemble signaling patterns suggestive of episodicmemoryformationarealsodetectedduringsleep.Formammals,EEGwaveformsduringsleepandwakefulnessarewellestablishedandcanbeassociatedwithperiodsofspecifictypesofmemoryformation.
BasedonEEGwaveforms,sleepisdividedintotwoforms,rapideyemovement(REM)andnon-REMsleep.TheAmericanAcademyofSleepMedicine furtherdividesnon-REMsleepintothreestagesN1,N2andN3.Sleepstagesfolloweachotherinasleepcycle.Sleepcycleslastabout75to100minutes(Figure7-12).
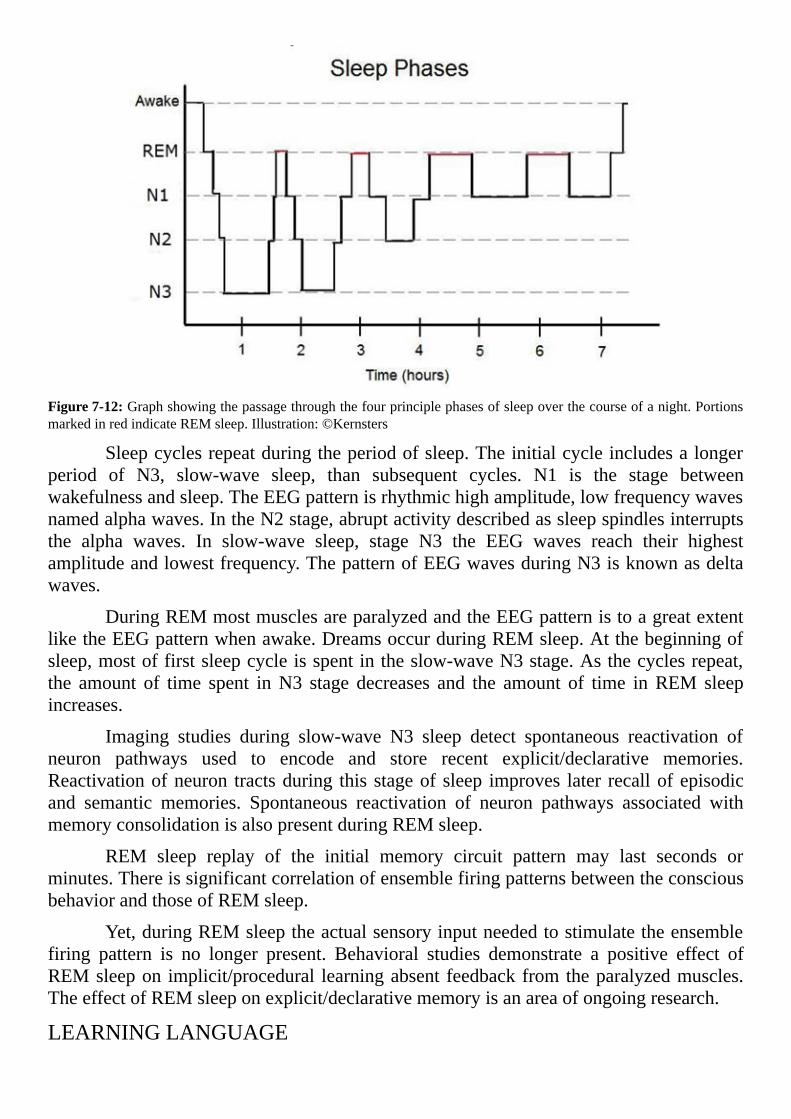
Figure7-12:Graphshowingthepassagethroughthefourprinciplephasesofsleepoverthecourseofanight.PortionsmarkedinredindicateREMsleep.Illustration:©Kernsters
Sleepcyclesrepeatduringtheperiodofsleep.Theinitialcycleincludesalongerperiod of N3, slow-wave sleep, than subsequent cycles. N1 is the stage betweenwakefulnessandsleep.TheEEGpatternisrhythmichighamplitude,lowfrequencywavesnamedalphawaves.IntheN2stage,abruptactivitydescribedassleepspindlesinterruptsthe alpha waves. In slow-wave sleep, stage N3 the EEG waves reach their highestamplitudeandlowestfrequency.ThepatternofEEGwavesduringN3isknownasdeltawaves.
DuringREMmostmusclesareparalyzedandtheEEGpatternistoagreatextentliketheEEGpatternwhenawake.DreamsoccurduringREMsleep.Atthebeginningofsleep,mostoffirstsleepcycleisspentintheslow-waveN3stage.Asthecyclesrepeat,the amount of time spent inN3 stage decreases and the amount of time inREM sleepincreases.
Imaging studies during slow-wave N3 sleep detect spontaneous reactivation ofneuron pathways used to encode and store recent explicit/declarative memories.Reactivationofneuron tractsduring this stageof sleep improves later recallofepisodicand semantic memories. Spontaneous reactivation of neuron pathways associated withmemoryconsolidationisalsopresentduringREMsleep.
REM sleep replay of the initial memory circuit pattern may last seconds orminutes.ThereissignificantcorrelationofensemblefiringpatternsbetweentheconsciousbehaviorandthoseofREMsleep.
Yet,duringREMsleeptheactualsensoryinputneededtostimulatetheensemblefiring pattern is no longer present. Behavioral studies demonstrate a positive effect ofREMsleepon implicit/procedural learningabsent feedbackfromtheparalyzedmuscles.TheeffectofREMsleeponexplicit/declarativememoryisanareaofongoingresearch.
LEARNINGLANGUAGE

NATIVELANGUAGEOnaverage,hippocampusmaturityisreachedataboutage3-4andcoincideswith
theagewhenaninfantacquirestheabilitytofitwordstogetherinanativelanguage.Thisisalsotheagewhenpeopledeveloptheearliestofthelong-termmemoriesrevisitedlaterinlife.Thisconvergenceofdevelopmentaleventsimplieslanguageisneededforepisodicmemoryformation.
The ability of humans to learn, communicate with and think in a languagefascinates both neuroscientists and psychologists. The advent of non-invasive brainimaging techniques expanded knowledge of particular brain areas and neuron circuitryessential for learning language.Language acquisition studies include infants and youngchildren,becausetheyaresuperiorlearnersoflanguagecomparedtoadults.
Remarkable data are accumulating describing establishment of a person’s nativelanguageand theneuroncircuits supporting language ingeneral.Exposureof infants tolanguagealone,likehearingarecordingofahumanvoice,appearsinadequate.Totriggerlearningfromexposuretolanguagetheremustbeasocialinteractionbetweentheinfantandanotherhumanbeing.Criticalaspectsofthesocialcomponentsremainunclear.
Theinfantbrainiswiredforreceivingauditoryinputpriortobirth.Dataindicateinfants recognize theirmother’s voice at birth. They begin learning the sounds of theircaregivers’ languageduring the first12monthsof life.Their implicitmemorycomes torecognizethatsoundsfallintopatternswithregularitiesintheverbalflow.Thepatternedsoundsbecomewordsoccupyinggrammaticalblueprints(Figure7-13).
Figure7-13:Youngmothertalkingwithherbaby.Photo:©Kletr
Vocabulary expands fast around18monthsof age.Fittingwords together into anative language begins between 18 and 36months. Speaking the native language with

fluencyistypicallymasteredbyage8.
Among the theories of why adults experience greater difficulty learning a newlanguageistheideaofinterference.Neuroncircuitssupportingpatternsofanestablishedlanguagemayhinderrecognitionofthenewlanguage’spatterns.Additionallanguagescanbe learned at any age but fluency of a native speaker is seldom attained if the newlanguageisacquiredafterpuberty.HOWSMALLCHILDRENARESTUDIED
Event-Related Potentials (ERPs) reveal language processing in little children(Figure7-14).ERPsmeasurethepartsofanelectroencephalogram(EEG)correspondingintimetoaspecificformofsensoryinput,inthiscasespeech.ERP/EEGisinexpensiveincomparison to other techniques. There is a good association between exposure to astimulus and measurement of the responding neuron activity. Being noiseless is anadditional advantage for auditory studies. Its major drawback is its sensitivity tomovement.
Figure7-14:Twoyearoldboywith24hourEEGelectrodesattached.Photo:©SteveBuckley
Magnetoencephalography(MEG)anothernoiselesstechniqueisusedforlanguageacquisition studies of children up to 2 years of age.MEG’s advantage is its ability todetectthepreciselocationoftheneuroncircuitsactivatedbyastimulus.Movementdoesnot affect MEG data because the helmet contains head tracking sensors. MEG allowstracingofneuronactivity inbothyoungchildren andadults. It canbeused to compareneuronactivity fundamental to learninganative languagewith circuits activatedduringadultlearningofasecondlanguage.Itsmajordisadvantageishighcost.
Near Infrared Spectroscopy (NIRS) also provides good spatial resolution of thebrainareaswithincreasedneuronactivitywhenaspecificsensorystimulusispresented.Thehelmet is smallandeasy to fit toan infantand theprocedure isnoiseless.BecauseNIRSissensitivetomovement,studiesareoftencompletedwhileaninfantisasleep.
Functional magnetic resonance imaging (fMRI) is used most often for adult

studies.Itsspatialresolutionofneuronresponseisexcellentbecauseitincludesanatomiclandmarks.However,fMRIissensitivetomovementandisnoisy.Itsnoiselevellimitsitsvalue in studies of small children because sound protection is necessary. fMRI is alsoexpensivetouseinresearchprotocols.
LANGUAGECIRCUITSA mere 600 consonants and 200 vowels are used to create all of the world’s
languages.TheEnglishlanguageusesasetofabout40non-identicalsoundsmadeupofconsonantsandvowels referred toasphonemes.Thedictionarydefinesphonemeas thesmallest speech unit in a language capable of conveying a distinction in meaning.Languagesvarytoagreatextentinthenumberofphonemestheyuse.
Babiesbegin learning languagebyselectingand figuringout themeaningof thesoundsinthespeechtheyhear.Ifaninfant’sparentsusetwodifferentnativelanguages,theinfantisabletolearntherulesforbothlanguagesat thesametime.It isimprobablethisoccursbecausetheinfantbrainstoresinformationaboutthestatisticalpatternsofthelanguages heard. Rather, multiple memory pathways induced by socialization play yetundocumentedrolesattachingmeaningtothesoundsoflanguage.
TheauditoryandthespeechcortexarelocatedatBrodmannareas22,41,42,44and45.Inhumansdamagetotheprimaryauditorycortexcausesonlyalossofawarenessofsound.Reflexivereactiontosoundremains.Theauditoryandspeechcorticalareasareonlysomepartsofthebraininvolvedinlearninglanguage.
Permanent loss of language due to brain injury requires failure of specificsubcorticalareasaswell.Thebasalganglia,andpathwaystoandfromthesebrainnuclei,playasignificantrole.Comprehendingthemeaningcommunicatedbytheorderofwordsrequiresactivityinthebasalgangliacircuitstotheprefrontalcortex.
Investigators puzzled for years over how the brain extracts and encodes speechfrom a noisy background. Linguists Roman Jakobson, Nikolai Trubetzkoy and SergejKarcevskijina1928paperclassifiedphonemes,soundsusedinspeech,intocategoriesbyhowairismovedthroughanoralcavitywhenthephonemeisspoken.Thethreecriteriausedwere1)where air is compressed in thepassageon itswayout, 2) vibrationornovibrationofthevocalcords3)howairisreleased.Theydemonstratedphonemescouldbeassignedtogroupsbaseduponthesevocalizationcharacteristics.
DataobtainedbyrecentstudiesofthelanguagecentersinthehumanbrainsupportJacobson’sgroupingofphonemes.StudiesrecordingdirectcorticalactivityinhumansaspartoftheirclinicalevaluationforepilepsysurgerydiscoveredthemajorityoftheEnglishspeech-activatedlocationsintheauditorycortexrespondtoaparticularphonemegroup.
Thisresultisincontrasttoanearlierexpectationthateachphonemewouldactivatea different location on the auditory cortex. Segregation of auditory cortex responseelementsbygroupsofphonemesbasedupon themechanicsof speaking implies speechandlanguagemaydeveloptogether.
Some investigatorshypothesize thedifficultyexperiencedbyadults in learninganewlanguageisduetocircuitryconstructedininfancytosupportphonemegroupsofthenative language. Overlap of phoneme groups between languages could trigger

inappropriateassociationsintheadultlearner.Itmayseemtotheadultlearnerthatcertainwordsrepresentsomethingdifferentthantheirtruesense.
Language ismanaged inadifferentway in thebrain’s leftandrighthemisphere.Thelefthemisphereistheprimarylanguagemanagerforthemajorityofpeople.Therighthemisphere is less involved, but communicationbetween the hemispheres is critical forcomprehensionofambiguouswordslike‘lead’,whichcaneithermean‘ametalsubstance’orto‘actasaguide’.
Resultsofrecentimagingstudiesdiscouragetheoriesofcompletedominancebyasingle hemisphere of any particular property of language.BothMEGand fMRI studiesreport the involvement of both hemispheres in many aspects of language processing.Dominance is nowdiscussed in terms of better performance of individual tasks by onehemisphereortheother.
Largestudiescontinueworldwidetoimprovetoolsavailableforinvestigationandmodelingofbraincircuits.Braincircuitsappearagooddealmorecomplexandmorefluidthan ever imagined. It is also becoming clear that neuron circuitry is influenced by thenon-neuron cells of the brain. Both astrocyte networks and patterns of myelination byoligodendrocytesimpacttheoutputofneuronensembles.
Neuron ensemble signaling pools and neurons capable of changing theirneurotransmitter in response to environmental fluctuation add an additional challenge.Uncoveringthemysteriesoftheclosedworldofthehumanbrainwillrequireco-operativeefforts by mathematicians, physicists, chemists, engineers and neuroscientists. There ismuchlefttodiscover,butnewstudieseachyearopenaclearerpictureofhowthehumanbrainworks.
SUMMARYCHAPTER7Evidence for how memory is formed is the closest science comes tounderstandingconsciousness,pleasure,anger,happiness,loveandhate
The 2-4 millimeter deep layer of neurons composing the human cortex iscritical to language, memory, attention, awareness, thought anddecisionmaking
Neuroninformationcodinginthebrainiscarriedoutbypopulationsofneuronsworkingtogetherinensembles
The connectome projects aremapping all neuron connections throughout thehumanbrain
EEGsmeasure largefluctuation in thebrainfieldpotentialscausedbyneuronensemblesfiringactionpotentialsinsynchrony
Four forms of non-invasive imaging for brain activity studies include Event-Related Potentials (ERP part of an EEG), functional magnetic resonanceimaging(fMRI),magnetoencephalography(MEG)andfunctionalnearinfraredspectroscopy(fNIRS)
Imagingstudiesdiscoveredsensoryinformationfromasingleeventisdividedintomanysmallpiecesandisdistributedwidelyacrossbrainregions

Storage and retrieval of episodicmemoriesby thebrainuse the sameneuroncircuits,butthedirectionofcurrentflowforretrievalisthereverseofthatforstorage
Memorydividesintotwotypes,consciousexplicit/declarativememoryoffactsandeventsandunconsciousimplicit/proceduralmemoryofapracticedskill
The neurons of the hippocampus codememory into signals that activate thenearbyentorhinal cortex,which in turnmovespiecesofmemory to theneo-cortexforstorage
A template for unconscious memory of a practiced activity is thought to bestoredwithinthecerebellum
Anessentialcomponentofmemory formation isanatomical reorganizationofneuronsynapsescalledneuroplasticity
Neurogenesis in anadulthumanbrain is confined to thedentategyrusof thehippocampus
Sleepproducesapositiveeffectonmemoryformation
Infantsandyoungchildrenaresuperiorlearnersoflanguagecomparedtoadults
Segregationofauditoryresponseelementsbyphonemegroupsalignedwiththemechanicsofspeakingimpliesspeechandlanguagemaydeveloptogether

[8]
WhenItAllGoesWrong—Alzheimer’sDementia
LIVING A LONG LIFE is highly desirable. Even though memory processingadjustsasthehumanbrainagescertainpeople, includingsomewholivepast100years,retain theirmental competence.Yet,manyadults fearonsetofdementia ingeneral, andAlzheimer’s disease in particular. According to the United States Centers for DiseaseControl and Prevention, Americans are two times more afraid of losing their mentalcapacitythantheirphysicalcapacitywhentheygrowolder.
But,what exactly isAlzheimer’s disease, and is it the same thing as dementia?Contemporary data indicate Alzheimer’s disease is a gradual change in the brain thatoccurs over decades before mental dysfunction is noticed. People who developAlzheimer’sdiseaselosetheirabilitytorememberandreasoninaprogressionthatisslow,frustratingandultimatelyfatal.
Graduallyneuronsdieintheregionsofthebrainneededformemoryofpeopleandevents.During thepre-symptomaticphaseofAlzheimer’sdisease, survivingcells in thebraincompensateforthelossofneuronsuntilfinallythenumberlostbecomestoogreat.The long pre-symptomatic phase ofAlzheimer’s diseasemay be viewed as good news,because it provides a lengthy period when prevention strategies can be designed andtested.
Abundantlossofneurons,andaresultingdecreaseinbrainvolumethatproceedssequentiallythroughspecificregionsassociatedwithmemoryandreasoning,differentiatesAlzheimer’s disease from other forms of dementia. An estimated 44 million peopleworldwidelivedwithdementiain2014.Thenumberisexpectedtoriseto70millionby2030.Alzheimer’sdiseaseisthemostcommontypeofdementiaanditaccountsfor60%to80%ofallcases.By2050theprojectedcostofcaringforindividualswithAlzheimer’sdiseasecouldreach$1.2trillionintheUnitedStatesalone.
This chapter examines the power of brain imaging techniques to unravel thecellular behaviors that produce Alzheimer’s disease. Advances in imaging technology,designedformedicaldiagnosis,openanewviewofthetransformationsoccurringinthehuman brain with age. New understanding of the physiology of the brain presentsunexploredavenuesfordiseaseprevention.
ALZHEIMER’SBRAINDifferentiation of Alzheimer’s disease from other forms of dementia is
accomplished by examination of brain tissue after death. Postmortem evaluation ofAlzheimer’sbrainrevealsshrinkageinthehippocampusandinareasofthetemporallobe,parietal lobe and frontal lobe of the brain (Figure 8-1). These brain areas managelanguage,memory,emotionandtheabilitytoreason.

Figure8-1:Lobesofthehumanbrainshowingmainsulciandboundaries.Illustration:©Sebastian023
TheclassicaltheoryofAlzheimer’sdiseaseassumesdepositsofamyloid-β,asmallbutaggregatedprotein,insomeunidentifiedmannerinitiateasequenceofeventsleadingtodeathofbrainneurons. In turn,neurondeathcausesshrinkageof theparticularbrainareas believed to govern memory and emotions. In support of this theory, brains ofdeceasedAlzheimer’s patients display large aggregates of amyloid-β in areas lacking anormalquantityofneurons.
Postmortem histology of Alzheimer’s brain detects signs of damage withinsurviving neurons and large amyloid-β deposits surrounding them. The most obviousinternalneurondamage is thepresenceofa fibrousdepositdescribedas tangled tau. Inhealthy brain, tau is a soluble protein within axons and dendrites and it would not beobservedbyroutinehistologictechniques.TheanatomyofaneuronisillustratedinFigure4-1inChapter4,“Neurons—HowTheyMakeElectricity.”
The intellectual impairmentobserved inAlzheimer’sdiseasecorrelateswellwiththe number of neurons displaying tau tangles and the decreased size of affected brainregions.Thepresenceof tangled tau in theneuronsofAlzheimer’sbrains isassumedtosignify a compromised cellular transport system. Soluble tau regulates assembly anddisassembly of train track-like structures that transfer material from one section of aneuron to another. The damaged neuron transport system probably contributes to theshrunkenappearanceofneuronsinAlzheimer’sbrain.
ALZHEIMER’STHERAPIESUntil a short time ago, it was hoped removal of amyloid-β plaque would halt
disease progression for patients assumed to be experiencing Alzheimer’s dementia.Clinical treatment trials involveddeliveryofantibodydrugstothebrain.Theantibodieswere expected to assist the brain’s innate immune system, the microglia, destroy theamyloid-βdeposits.
Unfortunately treating patients with mild to moderate dementia, who alsopresented a genetic predisposition to Alzheimer’s disease, with an antibody againstamyloid-βfailedtodelayprogressoftheirdisease.Theantibodyemployedintheclinical

trials passed through the blood brain barrier and cleared amyloid-β deposits, butparticipantsshowednoimprovementandtheirdiseaseprogressed.
Lack of positive results in clinical trials targeting amyloid-β plaque led someinvestigatorstoquestionwhetheritspresenceinAlzheimer’sisacauseorsymptomofthedisease.GBIResearch(GlobalBusiness IntelligenceResearch) reported inMarch,2015that583newdrugsareindevelopmentforAlzheimer’sdisease.Mostoftheseareinthediscoveryandpre-clinicaltestingphase.
New drug candidates favor different mechanisms of action than the earliertreatments,because therapiesforAlzheimer’sdiseasesince2006failedatarateof95%acrossall threephasesofclinical testing. Innovativenewdrugdesigns feature the latestdiscoveriesabouttheagingbrain.
PRE-SYMPTOMATICALZHEIMER’S
INVESTIGATIVETOOLSCombinations of non-invasive and invasive imaging techniques investigate
accumulation of amyloid-β and tau in the brains of people at high risk for developingAlzheimer’sdisease.EvidenceimpliesprogressionofAlzheimer’sdiseasebeginsyearstodecadesbeforesymptomsofintellectualfailuresurface.Sequential,progressivechangeinamyloid-βdepositsandtau’spresenceinbrainfluidproceedsforanaverageof10yearsbeforeevenminimalmentalimpairmentisexperienced.
Pre-symptomaticAlzheimer’sdisease is investigatedusingnon-invasivemethodsincluding electroencephalogram (EEG), magnetic resonance imaging (MRI), functionalmagnetic resonance imaging (fMRI) andmagnetoencephalography (MEG). Specifics ofthese methods are described in Chapter 7, “Brain’s Infrastructure for Memory andLanguage.”
Positronemissiontomography(PET),anotherimagingtechniqueforstudyingpre-symptomaticAlzheimer’sdisease, is invasivebecauseitrequiresinjectionofradioactivemolecules.Aradioactivetracermustbeincorporatedintoabiologicalmoleculecapableofcrossingthebloodbrainbarrier.Inthebrain,theradioactivemoleculeemitsasignalthatcan bemeasured after binding to a brain substance or after itsmodification by a brainenzyme.
The biologic tracer molecules chosen for PET studies of Alzheimer’s diseaseinclude fluorodeoxyglucose (FDG), an analogue of glucose, and Pittsburg compoundB(PiB),amoleculethatbindstoamyloid-βplaques.WithFDG,Alzheimer’sinvestigatorslook for brain areas with decreased utilization of glucose (Figure 8-2). Decreasedconsumption of glucose is interpreted as decreased neuron electrical signaling activity.PETdatacanbeoverlaidonMRIstructuraldataasinFigure8-2forapreciselocationofareaswithlowneuronactivity.

Figure8-2:Atransaxialsliceofthebrainofapatienttakenwithpositronemissiontomography(PET).Redareasshowahighamountof18F-FDGtraceraccumulationandblueareaslowtonotraceraccumulation.Image:©JensMaus
PET and fMRI remain the imaging techniques most often employed to studyAlzheimer’sdisease.InSeptember,2014asearchofPubMed,theUnitedStatesNationalLibraryofMedicineat theNationalCenter forBiological Information,producedfor thesearchterms“Alzheimer’s+fMRI+human”alistof5,165papers.Theearliestpaperonthe listwas published in 1983.A similar searchwith the terms “Alzheimer’s + PET +human” produced a list of 1,981 papers, and again the earliest paper on the list waspublishedin1983.
BothfMRIandPETuseindirectmeasurementsforevaluationofneuronsignalingactivity.fMRImeasuresflowofoxygenatedhemoglobinintosmallareasofbrainandPETmeasures glucose consumption. MEG, unlike fMRI and PET, measures actual neuronelectricalactivityand,thoughexpensive,maybeusedtoagreaterextentinfuturestudiesofAlzheimer’sdisease.APubMedsearchwiththeterms“Alzheimer’s+MEG+human”producedalistof111studiessince1996.
BIOMARKERSOFALZHEIMER’SDISEASEAbnormal levels of soluble tau and soluble amyloid-β in the cerebrospinal fluid
precedemildcognitivedeclinebyseveralyears.Tauincreasesandamyloid-βdecreasesinthecerebrospinalfluidduringthistime.Thedecreaseinamyloid-βincerebrospinalfluidisthoughttoreflectareductioninitsnormalrateofeliminationfromthebrain.
The increased presence of soluble tau in the fluids bathing neurons and incerebrospinalfluidpriortoonsetofAlzheimer’ssymptomssuggestsabnormalregulationof tau.Tau’snormalwork is thought tobeaccomplished insideneurons.Because tau isalsopresentinsidethenon-neuroncellsofthebrain,theglia,neuronscannotbeverifiedasthe sole source of elevated tau in brain fluids during the pre-symptomatic phase ofAlzheimer’sdisease.
Presenceoftauoutsideneuronsandthetangledformoftauproteinwithinneuronsarenot restricted toAlzheimer’sdisease.Anumberofotherneurodegenerativediseasesdisplay irregularities in the properties of tau protein within neurons and within glia.Neurodegenerativediseaseswithtauanomaliesareassignedthegeneralnametauopathies.They include among others frontotemporal dementia with Parkinsonismlinked to

chromosome17(FTDP-17T),gangliogliomaandleadencephlaopathy.
All tauopathies include neuron death and loss of the structures neurons use tocommunicatewitheachothernamedsynapses.Thenamesynapsederives fromaGreekterm,sunapsismeaningpointof contact.An indepthdescriptionof activities atneuronsynapsesispresentedinChapter5,“NeuronSynapses—ExcitatoryandInhibitory.”
PET imaging with Pittsburg compound B (PiB) estimates a brain’s insolubleamyloid-β load and indicates its distribution in various brain regions. PET PiB studiesdetectaccumulationofamyloid-βdepositsinthebrainformanyyearspriortoappearanceofmemoryloss in theportionof thepopulationathighriskfordevelopingAlzheimer’sdisease.
BelownormalconsumptionofglucoseinthebrainisanothercharacteristicofallAlzheimer’s patients. Diminished utilization of glucose in particular brain regions isconsidered an indicator of neuron loss. PET with 18F-FDG detects low glucosemetabolismintheposteriorcingulate,parietallobe,temporallobeandprefrontallobeofsomepeopleatriskfordevelopingAlzheimer’sdisease.
Peopleat riskbecauseofvariations in theirgenes,andwhodevelop thedisease,sometimesexhibitadeclineintheirbrain’sconsumptionofglucose20-30yearspriortosymptoms.Yet,noteverypersonwithpredisposinggenesdevelopsAlzheimer’sdisease.And,onlyabout50%ofpeoplewithAlzheimer’sdiseasehavegenesthatareidentifiedashighriskvariants.
Thepatternofactivitybetweenneuroncircuitsinvariousbrainregionsisstudiedusing fMRI and MEG. One theory based on fMRI and MEG data proposes multiplefunctional rearrangements of neuron circuit activitymay postpone cognitive difficultiesduring thepre-symptomaticphaseofAlzheimer’sdisease.Neuroncircuitry reroutingofinformationisestimatedtooccurasearlyas10yearsbeforesymptomsappear.
AMYLOID-βANDTAUPHYSIOLOGY
AMYLOID-βPHYSIOLOGYItseemsreasonabletoassumethebrainwouldnotpossessanelaborateprocessfor
creating amyloid-β if it served no purpose. Amyloid-β as a single peptide chain is aflexibleandunstructuredprotein.Thenormalconcentrationofsolubleamyloid-βproteininthehumanbrainandcerebrospinalfluidisabout10-9Molar,whichis4.5microgramsofamyloid-βperliteroffluid.
During sleep-wake cycles, amyloid-β production is enhanced during periods ofincreasedneuronactivityandisdecreasedwhenneuronactivityislow.Theoriesfromthisandotherdataproposeamyloid-βispartofanegativefeedbackloopcontrollingneuronsignaling.
Amyloid-βisasmallpeptidecutfromalargerprotein,amyloidprecursorprotein(APP) that isanchored inneuronmembranes.Amyloid-βexcisionand replenishmentofamyloid precursor protein in the membrane increase with rapid neuron signaling.Production continues until the concentration of amyloid-β becomes high enough at thesynapse to decrease the effectiveness of the neurotransmitter released during signaling

events(Figure8-3).
Figure8-3:Amyloidprecursorprotein(APP)moleculesinacellmembrane.ThesectionoftheAPPmoleculesshowningreenisamyloid-β.Illustration:NationalInstituteofAging,inthepublicdomainintheUnitedStates.
A complex bell-shaped relationship between synaptic amyloid-β and neuronactivity isproposed.Thebell-shapedmodelhypothesizes intermediateconcentrationsofamyloid-β,10-12Molarto10-9Molar,encouragereleaseofneurotransmitter. Incontrast,levelsbelow10-12Molarblockneurotransmitterrelease.Highlevelsofamyloid-β,above10-9Molar,alsoinhibitneuronsignaling,buttheyaccomplishitbyblockingtheeffectsofneurotransmitterratherthanaffectingitsreleasefromneurons.Similarbell-shapedactivityrelationshipsarecommoninbiologicsystems.
Less isknownabout thepurposeof the larger segmentof theamyloidprecursorprotein released by neuron enzymes during amyloid-β formation. Amyloid precursorproteinisanancientandconservedmoleculethroughoutmammalianevolution.Itoccurson the membrane of cells outside the brain as well as on neurons. Some of amyloidprecursorprotein’sstructuralcharacteristicsappearsimilar tothoseofgrowthpromotingmolecules like epidermal growth factor. Based upon its structure alone, the large piecereleasedwhenamyloid-βiscutoutmayactasagrowthfactorinthebrain.
AMYLOID-βPLAQUE
As the concentration of amyloid-β increases to 10-6Molar, it binds to itself andacquires a protein structure known as a beta sheet. The dramatic change in theconfiguration of amyloid-β from a floppy single strand peptide to a variety of multi-

stranded, rigid structures encourages a theorymaintaining each of amyloid-β’smultipleformsparticipatesinauniquebrainfunction.
Acontrastingtheoryproposesbetasheetamyloidstructuresmayprovideasimpleandefficientwaytoremoveexcessfunctionalamyloid-βbytransformingit intoashapenolongerusefulatsynapses.Thereductionofamyloid-βincerebrospinalfluidduringpre-symptomatic Alzheimer’s disease is interpreted failure of normal clearance of themoleculefrombrainratherthandecreasedproductionofthemoleculebyneurons.
The reasoning argues that amyloid-β would not accumulate as plaque if itsproduction decreased. And, a slower than normal clearance rate of amyloid-β wouldencourage formation of the beta sheet proteins which aggregate into plaque. Proposedmechanisms for reduction of amyloid-β in cerebrospinal fluid include changes in itsuptakebybraincellsoradecreaseinitstransportacrossthebloodbrainbarrierintothecerebralbloodvessels.
Figure8-4:Pinocytosisisaformofcellularuptake.Itusesareasofthecellmembranetograbsmallmoleculesfromthesurroundingfluid.Illustration:ThisworkisreleasedintothepublicdomainworldwidebyitscreatorJacekFH
A variety of cells in healthy brain remove soluble single-strand amyloid-β.Surveying microglia, the neuron monitoring system of the brain, internalizes solubleamyloid-β by pinocytosis (Figure 8-4) and delivers it to lysosomes for degradation.Soluble, single peptide amyloid-β is sensitive to themany enzymes that as amatter ofcoursedigestproteinsthatarenolongerneeded.
Astrocyte glial cells and the endothelial cells of blood capillaries use a differentmechanism than surveyingmicroglia for removal of soluble amyloid-β.Both astrocytesandcapillaryendothelialcellsparticipateinsafekeepingofthebloodbrainbarrierandintransfer of amyloid-β across the barrier. They use a membrane protein, LDL receptor-relatedprotein1(LRP1),totransportsingle-strandamyloid-βoutofthebrain.
LRP1 is abundant on the surface of the capillary endothelial cells of the bloodbrainbarrier.LRP1receptorisalsofoundonothercellsthroughoutthebrain,becauseitis

themajorvehicle foruptakeof cholesterol.Cholesterol is a critical componentofbrainmembranesingeneralandofneuronsynapsemembranesinparticular.
For fatty cholesterol to bind to LRP1 itmust be surrounded by awater-solubleprotein named apolipoprotein E (ApoE). In the brain, astrocytes make cholesterol andpackageitinApoEbeforesecretingitforusebyneuronsandotherbraincells.Incontrast,solubleamyloid-βdoesnotappeartobeincorporatedintoApoEpackagesbeforebindingtoLRP1.Rather,dataindicatesingle-strandamyloid-βbindsdirectlytoLRP1competingwithApoEforLRP1bindingsites.
Thecompetitionbetweenamyloid-βandApoEopensthepossibilitythatavariant,strongerbindingApoEmaypreventamyloid-βbindingtoLRP1.Peoplewhoinheritgenescoding particular variant forms of ApoE suffer a high risk of developing Alzheimer’sdisease.Possibly,specificvariantsofApoEpreventefficienteliminationofamyloid-βbyLRP1.
TAUPHYSIOLOGYUndernormalcircumstances, tau is a solubleproteinwithinneurons. Itprovides
for adequate spacing between track-like assemblies for moving material from place toplacewithin neurons. Tau’s efforts are controlled by an array of enzymes that add andremove phosphate groups at various positions.At least 30 sites on tau protein undergomodificationbyregulatoryenzymes.
Inadditiontoitswell-documentedroleinneuronaxons,tauisdetectedinthelongdendritesattheotherendofneurons.Itspresenceinaxons,dendrites,dendriticspinesandtheneuronbodysuggeststauisinvolvedintheongoingtransportofmaterialbetweenallpartsofneurons.
Neurons oftenmature into long cells where proteins andmoleculesmust travellong distances from their place of production to their destination. Feedback moleculesfromvariouspartsoftheneuronalsotravelalongtauregulatedassembliestosynchronizeenzymeactivityandgenetranscriptionintheneuronbody.
Tauprotein lacksanecessaryaminoacidsequencefor itsdirectpassage throughcellmembranes.Yetsoluble tau is found innormalcerebrospinal fluid.Howandwherealongtheneuroncellmembranetaumaymoveintotheneuron’sbathingfluid,andfromthereintothecerebrospinalfluid,isunknown.AdditionalinformationaboutthefluidsofthebrainispresentedinChapter3,“QualityControlofBrain’sExtracellularFluids.”
Tau without phosphate groups attached is a flexible soluble protein lackingstructuresimilartosingle-strandamyloid-β.Recentstudiesofthenon-phosphorylatedtaumoleculedetect regions thatassumeahelical formationwhennear the lipidsof thecellmembrane.Allproteinscapableofpassingthroughcellmembranesassumeahelicalformwithin themembrane.Discoveringwhether this property is an important factor in tau’sabilitytomoveoutofneuronsrequiresfurtherinvestigation.
TauobservedwithinneuronsinpostmortemAlzheimer’sbrainpossessesa largernumberofphosphategroupsthanrequiredforitsnormalactivities.Theextraphosphateschangetaufromaflexiblemoleculetoahelicalfilamentwithanegativeelectriccharge.Pairsofhelicaltaufilamentsaggregateintothetautanglesvisiblewithinneurons.

Extracellular, soluble tau is elevated in the neuron synapses duringAlzheimer’sdisease.SolubletauwithintheAlzheimer’ssynapsesacquiressomephosphategroupsbutnotsomanythatitprecipitatesoutofsolution.
NEURONDAMAGEANDLOSS
MEMORYCIRCUITSPostmortem Alzheimer’s brain shows a dramatic decrease in total volume in
additiontoamyloid-βdepositsandtautanglesinsideremainingneurons.Theentirelossinsize of the brain appears to be due to disappearance of neurons. The astrocyte cellpopulationappearstoremainstableandthemicrogliapopulationexpands.
Neuronlossisparticularlyevidentinthehippocampusandcerebralcortexofthetemporal,parietalandfrontallobes.Regionallossofvolumeissequentialandconsistentwithdiseaseprogression(Figure8-5).
Briefperiodsof extreme lossof recallof events, situationsandpersonal experiencesprovideaninitialwarningofAlzheimer’sdisease.Thepartofthebrainthatcodesmemoryofevents,episodicmemory,isthehippocampus.ThehippocampusanditsadjacentbrainareasareamongthefirsttosufferdamagebyAlzheimer’sdisease.
The hippocampus receives sensory input from auditory, visual and olfactory corticalneurons.Italsoreceivesinputfromtheneuronsofthefrontalcortex,thepartofthebraininchargeofmakingexecutivedecisions.Hippocampusneuronssendtheirmemorycodestoaportionofthetemporallobenamedtheentorhinalcortex.Fromtherepiecesofeachmemorymove to other parts of the cerebral cortex for storage until they needed in thefuture.Neuroninputfromtheneighboringamygdalatothehippocampusaddsemotionaltexturetomemories.

Figure8-5:Alzheimer’sdiseasebraincomparison.The top isan illustrationofnormalhumanbrainstructureandthebottomisanillustrationofabrainwithAlzheimer’sdisease.©Garrondo,,releasedtothepublicdomain
A largeMRI study evaluated volume and shape change in areas of the medialtemporal lobe of people at risk for developing Alzheimer’s disease. MRI scans wereacquired from1995 to2005.By the time the studypublished in2014participantswerefollowed for18years.Thegoalof the studywas todetermine the sequenceof affectedbrainareasduringpre-symptomaticAlzheimer’sdisease.

Figure8-6:AreasofthebrainthatfailasAlzheimer’sdiseaseprogresses.Illustration:©7mike5000
InthisMRIstudy,theentorhinalcortexshowssignificantvolumechangeassoonas8to10yearsbeforetheonsetofsymptoms.Atrophicchangesintheentorhinalcortexalign with the pre-symptomatic time points when amyloid-β first decreases incerebrospinalfluid.Thehippocampusshowsatrophy2to4yearspriortosymptoms.Andthe amygdala displays significant changes 3 years prior to mild cognitive impairment.Atrophic regions correspond to brain areas postmortem with shrunken neurons, heavydepositsoftautanglesandamyloid-βplaque(Figure8-6).
GLUCOSEMETABOLISM
Adecrease inglucosemetabolism revealedbyPETwith 18F-FDG, a radioactiveglucose analog, aligns with onset of brain atrophy detected by MRI. Neurons dependalmostexclusivelyuponglucoseforenergy,andtheirsignalingproceduresrequireabout20% of the body’s total energy supply. PET data combined with fMRI data showingreduced blood volume in the brain areas affected by Alzheimer’s disease suggest thedisease initiating factor may be a failure in the brain’s energy producing metabolicpathways.Complexmodelshavebeenproposedforhowneuronsandastrocytesmayshareenergyproduction.Thedetailsofthepathwaysarelessimportantforthisdiscussionthanthefactthatthemetabolicrelianceofneuronsandastrocytesuponeachotheriscomplex.
If the metabolism theory of Alzheimer’s disease is correct, then the metabolicdefectmaybeanastrocytedefect,oraneurondefectorboth.Glucosemetabolisminbrainrequires a partnership between neurons and astrocytes. It entails a dynamic sharing ofmolecularconversionpathwaysbetween the twocell types.Adetaileddiscussionof thenormal dynamic relationshipbetweenneurons and astrocytes for energy conservation is

presentedinChapter6,“IntroductiontotheGliaandMicroglia—MeettheStageCrew.”
ACTIVATIONOFGLIAANDMICROGLIA
REACTIVEASTROCYTESAstrocytes tile thebrain inacloseordered lattice.Basedupontheirmorphology,
three types of astrocytes reside in the brain. They are protoplasmic astrocytes, fibrousastrocytes and reactive astrocytes. Protoplasmic astrocytes surround neuron bodies.Fibrousastrocytes associatewithneuronaxons.Reactiveastrocytes disperse throughoutdamagedbrain.
Whenbraintissueexperiencesaninjury,protoplasmicandfibrousastrocytesreactbyshifting theircharacteristics to thoseofreactiveastrocytes.Theastrocyteresponse tobrain damage is termed reactive astrogliosis. Astrogliosis is a variable modification ofastrocytebehaviordependinguponthecontextofthedamage.Inthelesssevereformofastrogliosis, reactive astrocytes adjust and repair local brain structure without scarformation.Whendamageissevere,reactiveastrocytesproliferateandformscarstowalloffthedamage.
AlargenumberofreactiveastrocytesinhabitbrainswithAlzheimer’sdisease.Theshiftfromaninterspersedpopulationofsupportiveprotoplasmicandfibrousastrocytestoa population where reactive astrocytes predominate begins in the initial stages ofAlzheimer’s disease. With increasing duration of disease, the portion of the astrocytepopulationinthereactiveformincreases.
CarefulstudiesoftheastrocytepopulationofAlzheimer’sbrainssuggestthetotalpopulation of brain astrocytes does not change during disease progression. Rather thepercent of the population functioning as protoplasmic and fibrous astrocytes decreaseswhilethepercentfunctioningasreactiveastrocytesincreases.
ACTIVATEDMICROGLIA
Two functional forms ofmicroglia live in brain tissue, surveyingmicroglia andactivated microglia. During the switch from surveying form to the activated form,microglia changes its shape,proliferates andbegins to expressmarkers characteristicofthemacrophagesofthebody’simmunesystem.Theactivatedformofmicrogliaremovesneuronswithseveredamageandengulfsamyloid-βplaque.
Ofnote,inAlzheimer’sdiseasealldeadneuronsaremissingfromthebrain.Deadand dying neurons are not identified in the tissue. In healthy brain surveyingmicrogliamonitorsthewell-beingofneuronsynapses.Ifaneuronisdamagedsurveyingmicrogliaattemptstorepairitbysecretinggrowthfactors.However,ifaneuronisdamagedbeyondrepairsurveyingmicrogliatransformstoactivatedmicroglia,finishesthekillandremovesthedebris.
InAlzheimer’sdisease it isunknownwhethersurveyingmicroglia fails to repairdamagetoneuronscausedbysomeyetunknownfactor,orifsurveyingmicrogliaplaysaroleininitiatingneurondeathbeforebecomingactivatedanddisposingoftheremains.
Uptakeofsingle-strandamyloid-βbysurveyingmicrogliatoclearitfrombrainis

different than the response of activated microglia to amyloid-β plaque. Activatedmicrogliatreatsamyloid-βplaqueasaforeignintruder.Itattemptstosurroundanddestroythe plaque. This is the same form of microglia that removes dying neurons. Becauseactivated microglia responds to amyloid-β plaque as a foreign intruder, Alzheimer’sdisease isdescribedasan inflammatorydisease.This tends to causeconfusion,becausethe inflammation ofAlzheimer’s disease exhibits different characteristics than classicalinflammation.
Classical inflammation isaprocess involvingcellsof thebody’s immunesystemresponding to the presence of a foreign substance.The classical inflammatory responsedilates blood vessels and increases their permeability to water and cells. Water andimmune cells then gain access to the foreign object. Heat, swelling, redness and paindevelopintheaffectedtissue.Destructionorwallingoffoftheforeignobjectisfollowedbyhealinginthetissue.
Classicalinflammationdoesnotoccurinthebrainunlessthebloodbrainbarrierisbroken.ThebloodbrainbarrierofdeceasedAlzheimer’spatientsshowsnofunctionallossbeyond that normally expected because of age. Pathologists label Alzheimer’s brain“degenerative” rather than “inflammatory” on postmortem evaluation. The term‘inflammation’when characterizing a braindisplayingAlzheimer’s disease refers to thepresenceofalargequantityofreactiveastrocytesandactivatedmicroglia.
InAlzheimer’s brain activatedmicroglia surrounds dense amyloid-β plaques butappear unable to remove them. Like macrophages of classical inflammation, activatedmicroglia discharges cytokines, chemicals that attract other cells to the areas of plaqueinvasion.Thecytokinesreleasedbymicrogliadrawreactiveastrocytestotheplaque.
Reactive astrocytes internalize amyloid-β using their cell membrane proteins.Reactiveastrocytesalsoencircleamyloid-βplaquesinformationssimilartotheglialscarstheycreateafterbrain trauma.Glial scar-like formationmaycreateabarrier to separateamyloid-βplaquesfromhealthybraintissue.
ALZHEIMER’S-LIKEBRAINWITHOUTDEMENTIAAlzheimer’sdiseasewasoriginallydescribedasaformofdementiaaccompanied
by the presence in the brain of large amyloid-β plaques between neurons and tangledmaterial within neurons. A feature lacking in histologic investigations of Alzheimer’sdiseaseupuntilpublicationoftheNunstudyin1997wasacontrolgroupofbrainsfromoldpeoplewhoremainedmentallycompetentduringtheirlifetime.
Asimilarbutlargerstudy,theReligiousOrdersStudy,fundedbytheUnitedStatesNationalInstituteonAgingatRushUniversityMedicalCenterinChicagowillcontinuethroughJune,2016.By the timeofcompletion, the studywillhave22yearsof clinicaldataonmorethan1,000participantsandbraintissuefromover350people.Nuns,priestsandbrothersfrom31CatholicordersintheUnitedStatesareparticipating.
Asurprisingoutcomeof thecontrolledstudiesofageandAlzheimer’sdisease isthe discovery of participants with intact intellectual function whose brains displayAlzheimer’s-like amyloid-β deposits and tau tangles at postmortem examination. ThisphenomenonappearsintheresultsofboththeNunStudyandtheReligiousOrdersStudy.

Inthesestudies,12%to30%ofparticipantswithoutcognitiveimpairmentatdeathdisplayedsubstantialamountsofamyloid-βplaqueandtautangles in theirneurons.Yet,these individuals retained the neurons, synaptic elements, axon geometry and corticalthicknessneededfornormalmemoryfunction.BrainsoftheseindividualsarereferredtointheremainderofthischapterasAlzheimer’s-likebrains.
Observation ofAlzheimer’s-like brainwith normal cognitive function is helpinginvestigators re-evaluate factors involved in the Alzheimer’s disease progression. Themost dramatic non-neuron characteristic of Alzheimer’s-like brain is a lack of reactiveastrocytes. The large number of reactive astrocytes observed in the majority ofAlzheimer’sbrainsatpostmortemexaminationisabsentinAlzheimer’s-likebrains.
Other differences exist between Alzheimer’s brain and Alzheimer’s-like brain.Brains from Alzheimer’s patients contain more of the structured beta sheet forms ofamyloid-β and have amyloid-β plaques that are larger than those of Alzheimer’s-likebrains.Alzheimer’s-likebrainshave lesssoluble tau influidssurroundingsynapses thanAlzheimer’sdiseaseandothertauopathies.
Two neuron synaptic proteins, reduced about 50% in Alzheimer’s diseasecompared to normal brain, remain unchanged in Alzheimer’s-like brain. One of theseproteins is acommoncomponentof thevesicles that storeand releaseneurotransmitter,andtheother isacomponentof theregionof thesynapsethatcontainsneurotransmitterresponseelements.
Similarities between Alzheimer’s brains and Alzheimer’s-like brains includeextensivetautangleswithinneurons,asubstantialquantityofamyloid-βplaqueandequalamountsofsingle-strandflexibleamyloid-βwithinsynapses.
NEWAVENUESFORPROGRESSRevelationoftheNunStudyandtheReligiousOrdersStudythatbrainswithintact
reasoning ability sometimes display the classical markers of Alzheimer’s disease is animportant advance in knowledge about this illness. These brain control studies providecompellingevidencethatamyloid-βplaquearound,andtautangleswithin,neuronsarenotsufficienttocausethemassivelossofneuronsassociatedwithAlzheimer’sdementia.
Some theories suggest rogue reactive astrocytesmay kill rather than protect theneurons.Alternately,amereswitchofastrocytephenotypetothereactiveformwithoutanincrease in their total populationmay simply deprive neurons of themetabolic supporttheyrequireforsurvival.AnimportantdistinguishingfeatureofAlzheimer-likebrainwasa lack of reactive astrocytes. Because astrocytes normally become reactive only in thepresence trauma or a signal from activated microglia, it is important to discover theactivation signal(s) astrocytes receive from microglia with progression of Alzheimer’sdisease.
In Alzheimer’s-like brain astrocytes supportive of neuron well-being werepreserved.Microgliaevidentlydidnotsignalthoseastrocytestoparticipateindestructionofneurons,becausetheneuronpopulationsurvived.CoulditbethatsurveyingmicrogliaofAlzheimer’s-likebrainfailedtoreceivethe‘killme’signalfromneurons?Eventhoughmoreneeds tobe learnedabout indicatorsofneurondistressatsynapses,microgliamay

provideanexcellentdrugtargetrightnowtoslowprogressionofAlzheimer’sdisease.
Microglia initiated inflammation is caused by secretion of molecules similar tothoseusedbyperipheralmacrophage.Inflammationisusuallyatightlycontrolledprocesswhere pro-inflammatory and anti-inflammatorymolecules are secreted in a pattern thatdestroysaninvaderandthenhealsthetissue.Whenthesystembecomesunbalancedandpro-inflammatorymoleculespersistentlydominate,tissuedestructionoccurs.
Until very recently peripheral macrophage was thought to be different thanmicrogliabecausewornout cells are replacedbybonemarrowstemcells.Microglia incontrastseedsneural tissueduringembryonicdevelopmentandtheoriginalmaturecellsprovide expansions of the population for an entire lifetime. This odd capability ofmicroglia,amaturecell,torenewitselfforcedscientiststoreevaluatetheirexplanationofstemcells.Dogmawasthatonlystemcellsarecapableofself-renewal.However,itisnowknown that several tissues outside the brain also host a fully differentiatedmacrophagepopulationthatisabletoself-renewlikemicroglia.
Self-renewingmacrophageoftissuesoutsidethebrainmayprovideanaccessiblemodel for further investigation of microglia inflammatory response. Anti-inflammatorydrugs specifically targeted to microglia present practical advantages for treating anddelayingonsetofAlzheimer’sdisease.Alargedatabaseisalreadyavailableforextendeduseofvariousanti-inflammatorymedicationstoaidindesignoftheresearch.
Drugstargetingmicroglia-inducedinflammationareamongthosecurrentlyinpre-clinicalandearlyphaseclinicaltesting.ItisverylikelythatthekeytodelayingonsetandprogressionofAlzheimer’sdiseasewillbefoundinthewayneuronsandnon-neuronbraincellsconnect,shareanddisengage.Theoldbeliefthatapersononlyuses10%ofhis/herbraincellsbecauseamere10%ofbraincellsareneuronshasbeenprovenfalse.
SUMMARYCHAPTER8The classical theory of the last 30 years that proposes deposits of amyloid-βbegin the sequence of events leading to death of neurons associated withAlzheimer’sdiseaseisbeingreevaluated
The degree of intellectual impairment observed with Alzheimer’s diseasecorrelateswith thenumberofneuronswith tau tanglesanddecreasedsizeofaffectedbrainregions
Sequential,progressivechangeinamyloid-βdepositsandaccumulationoftautangles proceeds for an average of 10 years before mental impairment isexperiencedinpatientswithAlzheimer’sdisease
Accumulation of amyloid-β into dense plaque formations throughoutAlzheimer’sbrainisbelievedtobeanoutcomeofdecreasedclearanceof thenormalbrainmolecule
Abnormal tau in cerebrospinal fluid is not specific to Alzheimer’s diseasebecause an elevated quantity of tau is present in other forms ofneurodegenerativediseasesaswell
LossofneuronsinparticularbrainareasisasequentialprocessinAlzheimer’s

disease
ThefirstbrainareaaffectedbyAlzheimer’sdiseaseisaregioninthetemporallobes adjacent to the entorhinal cortex, followed in order by the entorhinalcortex, hippocampus, amygdala, areas of the temporal and parietal lobesassociatedwithlanguagefunctionandthentheprefrontalcortex
Earliest changes observed in the shape and volume of the entorhinal cortexappearassoonas8to10yearspriortomemorylosssymptoms
Thehippocampusshowsatrophy2to4yearsbeforesymptoms
In people at high risk for Alzheimer’s disease, a decrease in glucosemetabolismcanbedetecteddecadesbeforetheonsetofsymptoms
Brains of some healthy people retain intact neuron pathways yet display themarkersofAlzheimer’sdisease,amyloid-βdepositsandtautangles
Themost strikingdifferencebetweenAlzheimer’s-likebrain andAlzheimer’sdiseasebrain is theportionof the astrocytepopulation that is in the reactiveform
Most astrocytes in late stage Alzheimer’s disease are of the damage-controlreactivephenotyperatherthanthefunctional-supportprotoplasmicandfibrousphenotype
Thelargepopulationofreactiveastrocytes inAlzheimer’sdiseasemayplayaroleinkillingneurons
Alternatively, a switch in phenotype of astrocytes to their reactive formmaydepriveneuronsofthesupporttheyrequireatsynapsesforsurvival
Analysts report of the 583 drug candidates for Alzheimer’s disease underdevelopment in2015by thepharmaceutical industry, themajority are still inthediscoveryandpre-clinicaltestingphase
NewdrugcandidatesforAlzheimer’sdiseasefocusuponthelatestevidenceofhowneuronsandnon-neuronbraincellsinteractwitheachother

FurtherReadingTHEFOLLOWINGPAPERSAREAFEWofthereferencesconsultedinwriting
thisbook.Thelistisfarfromexhaustive,butitprovidesasampleofthecurrentscientificliterature discussed. Most of these papers are available for free athttp://www.ncbi.nlm.nih.gov/pubmed/. There is a search box at the top of the page atPubMedwhereyoumaytypeineitherthePubMedIdentificationnumber(PMID)orthePubMedCentralIdentificationnumber(PMCID)listedwitheachcitation.Onceyoureachthefreepaperitcanbedownloadedinvariousformats.
2104 Alzheimer’s Disease Facts & Figures,http://www.alz.org/downloads/facts_figures_2014.pdf,Alzheimer’sAssociationReport
BennettDAetal.,OverviewandFindingsfromtheReligiousOrdersStudy,2012,CurrAlzheimerRes,9:628-645,PMID:22471860PMCID:PMC3409291
BergelsonEandSwingleyD,Theacquisitionofabstractwordsbyyounginfants,2013,Cognition127:391-397,PMID:23542412,PMCID:PMC3633664
BirrenS.J.andMarderE.,PlasticityintheNeurotransmitterRepertoire,2013,Science340:436-437,PMID:23620040
Cakir T et al., Reconstruction and flux analysis of coupling between metabolicpathways of astrocytes and neurons: application to cerebral hypoxia, 2017, Theor BiolMedModel4:48Review,PMID:18070347,PMCID:PMC2246127
Catterall WA, Voltage-gated sodium channels at 60: structure, function andpathophysiology, 2012, J Physiol 590:2577–2589, PMID: 22473783, PMCID:PMC3424717
Chan MY et al., Decreased segregation of brain systems across the healthy adultlifespan, 2014, Proc Natl Acad Sci USA 111:E4997-5006, PMID: 25368199 PMCID:PMC4246293
Corkin S., Permanent Present Tense, 2013, Basic Books, New York. ISBN9780465031597
Dulcis D and Spitzer NC, Reserve pool neuron transmitter respecification: Novelneuroplasticity, 2012, Dev. Neurobiol. Vol. 72:1-15, PMID: 21595049, PMCID:PMC3192250
Eriksson PS et al., Neurogenesis in the adult human hippocampus, 1998, NatureMedicineVol.4:1313-1317,PMID9809557,FreeatNatureMedicine
FowlerPWet al.,Detailed examination of a single conduction event in a potassiumchannel,2013,JPhysChemLett4:3104–109,PMID:24143269,PMCID:PMC3797101
Greer PL and Greenberg ME, From synapse to nucleus: Calcium-dependent genetranscriptioninthecontrolofsynapsedevelopmentandfunction,2008,Neuron59:846–860,PMID:18817726,CellPressOpenAccess
Häusser M et al., Diversity and dynamics of dendritic signaling, 2000, Science290:739–744,PMID:11052929

HillmanEM,Opticalbrainimaginginvivo:techniquesandapplicationsfromanimalto man, 2007, J Biomedical Optics, 12:051402 Review, PMID 17994863, PMCIDPMC2435254
Kawakami R et al., Visualizing hippocampal neurons with in vivo two-photonmicroscopy using 1030 nm picosecond pulse laser, 2013, Scientific Reports 3: 1014,PMID:23350026,PMCID:PMC3553458
Kuhl PK, Early language learning and literacy: neuroscience implications foreducation, 2011, Mind Brain Educ, Vol. 5, 128-142, PMID: 21892359, PMCID:PMC3164118
Lazarczyk MJ et al., Preclinical Alzheimer disease: identification of cases at riskamong cognitively intact older individuals, 2012, BMC Medicine 10:127-139, PMID:23098093,PMCID:PMC3523068
Lloren-Martin’Met al.,Selectivealterationsofneuronsandcircuits related toearlymemory loss in Alzheimer’s disease, 2014, Frontiers in Neuroanatomy 8:38-49, PMID:24904307,PMCID:PMC4034155
McKenzie IA et al.,Motor skill learning requires active central myelination, 2014,Science346:318,PMID:25324381
MedinaMandAvilaJ,TheroleofextracellularTauinthespreadingofneurofibrillarypathology, 2014, Frontiers in Neuroscience, 8:113-119, PMID: 24795568, PMCID:PMC4005959
Merin-Serrais P et al., The influence of phospho-T on dendritic spines of corticalpyramidal neurons in patients with Alzheimer’s disease, 2013, Brain 136:1913-1928,PMID:23715095,PMCID:PMC3673457
Miller JF et al.,Neural activity in humanhippocampal formation reveals the spatialcontextofretrievedmemories,2013,ScienceVol.342,1111-1114,PMID:24288336Freeatwww.sciencemag.org
MillerG,Mysteriesofthebrain.Howarememoriesretrieved?2012,Science338:30-31,PMID:23042864
NeherJJetal.,Primaryphagocytosisofneuronsbyinflamedmicroglia:potentialrolesinneurodegeneration,2012,FrontiersinPharmacology3:1-9,PMID:22403545,PMCID:PMC3288722
PanzaFetal.,Istherestillanyhopeforamyloid-basesimmunotherapyforAlzheimer’sdisease?2014,CurrOpinPsychiatry,27:128-137,PMID:24445401
Perez-Nievas BG et al.,Dissecting phenotypic traits linked to human resilience toAlzheimer’s pathology. 2013, Brain 136:2510-2526, PMID: 23824488, PMCID:PMC3722351
RanasingheKGetal.,RegionalfunctionalconnectivitypredictsdistinctimpairmentsinAlzheimer’sdiseasespectrum,2014,NeuroImage:Clinical5:385-395,PMID:25180158,PMCID:PMC4145532
ReeceM,Physiology:Customdesignedchemistry,2012,ReeceBiomedicalConsulting

LLC,CreateSpaceISBN9781482326611
SallowaySetal.,Twophase3trialsofBapineuzumabinmild-to-moderateAlzheimer’sDisease, 2014, New England Journal of Medicine, 370:322-333, PMID: 24450891,PMCID:PMC4159618
Serrano-Pozo A et al., A phenotypic change but not proliferation underlies glialresponse in Alzheimer’s disease, 2013, American Journal of Pathology 182:2332-2344,PMID:23602650,PMCID:PMC3668030
SertbaᶊMetal.,Systemicanalysisof transcription-leveleffectsofneurodegenerativediseases onhumanbrainmetabolismbyanewly reconstructedbrain-specificmetabolicnetwork,2014,FEBSOpenBio4:542-553,PMID:25061554,PMCID:PMC4104795
Sieweke MH and Allen JE, Beyond stem cells: self-renewal of differentiatedmacrophages, 2013, Science, 342 342(6161):1242974. doi: 10.1126/science.1242974,PMID:24264994
SnowdonDA,AgingandAlzheimer’sdisease:LessonsfromtheNunStudy,1997,TheGerontologist, 37:150-156, http://gerontologist.oxfordjournals.org/content/37/2/150.long,PMID:9127971
SpaldingKLetal.,Dynamicsofhippocampalneurogenesisinadulthumans,2013Cell153:1219-1227,PMID23746839,FreeatCellPressOpenAccess
SwingleyD,Therootsofearlyvocabularyininfants’ learningfromspeech,CurrDirPsycholSci17:308-311,PMID:20523916,PMCID:PMC2879636
Tomassy GS et al.,How big is the myelinating orchestra? Cellular diversity withinoligodendrocyteslineage:factsandhypotheses,2014,FrontCellNeurosciVol8,Article201,PMID:25120430,PMCID:PMC4112809
Younes Let al., Inferring change point times ofmedial temporal lobemorphometricchangeinpreclinicalAlzheimer’sdisease,2014,NeuroImage:Clinical5:178-187,PMID:25101236,PMCID:PMC4110355
WongADetal.Theblood-brainbarrier:Anengineeringperspective,2013,FrontiersinNeuroengineering,6:1–22,PMID:24009582,PMCID:PMC3757302

GlossaryActionpotential–apatternofvoltagetransientsacrossanaxon’smembrane
Activatedmicroglia–aformofmicrogliathatdestroyspathogensandremovesdeadcells
Adherensjunction–acomplexofproteinsthattiethemembranesoftwocellstighttogether
Allostericbindingsite–apositiononaproteinwherethebindingofadrugmoleculeforcesachangeintheshapeoftheprotein
Alzheimer’sdisease–aprogressiveformofdementiacharacterizedbyabundantlossofneuronsinbrainareasassociatedwithmemoryformation
Amyloid-β – a small peptide with an undefined function produced on the outsidesurface of brain neurons that accumulates as large insoluble deposits in the brains ofAlzheimer’spatients
Anterograde–directionofmovementwithinneuronsofmaterialtravellingawayfromthecellbody
ApolipoproteinE–abrainproteinthatsurroundscholesterolasitistransportedfromoneareatoanother
Arachnoidmembrane – a membrane covering of the brain and spinal cord whosedelicatefibersgiveitaspiderweb-likeappearance
Astrocyte–astarshapedbraincellthatworksinpartnershipwithneurons
Autobiographical memory – a form of memory similar to episodic memory butlimitedtoeventsinaperson’sownlifehistory
Axon–a longprojectionofaneuron’scellmembrane thatconductschemicalbasedelectricity
Axoncollateral–abranchoffofanaxonthattravelsthroughthebraininadifferentdirectionthanthemainaxon
Axonhillock–theregionofaneuronbodythatconnectstotheaxon
Axoninitialsegment–thepartoftheaxonclosesttotheneuronbodythatcontainstheaxon’sfirstsetofvoltage-sensitivesodiumchannels
Axon terminal – the far end of the axon away from the neuron body whereneurotransmitterisstored
Bloodbrainbarrier–bloodcapillariesthatregulatetransferofmoleculesintoandoutofthebrain
Braininflammation–aconditioninthebrainproducedbyprolongedmalfunctionofanormalprotectivemechanismtodestroybacteriaandvirus
Brainstem–thepartofthebraincontinuouswiththespinalcord

BrodmannAreas–amapofcorticalneuronpatternsbaseduponmarkeddifferencesinneuronconfigurationandconnectivity
Carotidarteries–arteriesof theneck thatsupplyblood to thefaceand to thefrontandmiddleportionofthebrain
Cerebellum – structure at the back of the brain critical for refinement of motormovements
Cerebralcortex–thesixlayersoflarge,intricatelyconnectedneuronsthatcoverthebrainhemispheres
Cerebrospinalfluid– fluidcirculating throughandaround thebrainandspinalcordthatisproducedbythechoroidplexusesandtheependymalcellsliningthesurfaceoftheventricles
Choroidplexus – a tissue located in eachof thebrain’s fourventricles that extractsnutrients from specialized blood capillaries and secretes a solution called cerebrospinalfluid
Circle ofWillis – a safety net of blood vessels at the base of the brain where theanterior carotid and posterior vertebral circulations are connected by the posteriorcommunicatingarteries
Connectome–aproposedmapofalltheneuronconnectionsinthehumanbrain
Corpuscallosum–abridgeofneuronaxonsthatconnectcorrespondingareasofthebrain’srightandlefthemispheres
Cytoplasm–thefluidcompartmentwithincellsthatisoutsidethenucleus
Cytoskeleton–theinternalframeworkofacellcomposedprimarilyofactinfilamentsandmicrotubules
Dementia–deteriorationofaperson’sintellectualabilities
Dendrite–abranchedextensionofaneuronbodythatcollectsandconductsimpulsesfromadjacentneuronsinwardtowardthecellbody
Dendriticspikes–membranevoltagetransientsoriginatinginneurondendritessimilartoactionpotentials
Dendritic spines – thorn-like projections of dendritemembranewhere synapses arelocated
Diencephalon–thedivisionofthebrainthatincludesthethalamusandhypothalamus
Diffusion– inchemistry the relocationofmoleculeswithinasolutionawayfromanareawheretheyareinhighconcentration
Duramater – a double layer of thick fibrousmembrane between the brain and theboneoftheskull
Electroencephalogram,EEG– thedifferenceinfieldpotentialsdetectedbypairsofscalpelectrodes
Ensemblecoding–actionpotentialcodingofinformationcarriedoutbypopulations

ofneuronsworkingtogether
Entorhinalcortex–anareaof the temporal lobeof thebrainnear thehippocampusthat acts as an interfacebetween thehippocampusand theneo-cortexwherememory isstored
Ependymalcells–smallcuboidalciliatedcells lining thesurfaceofbrainventriclesthatsecreteandabsorbcerebrospinalfluid
Episodicmemory – thememory form that recalls specific events, people, situationsandpersonalexperiences
Equilibriumpotentialforpotassium–acalculatedtransmembranepotentialforcellswithpassiveK+channelsbutnopassiveNa+channels
Equilibriumpotential for sodium – a calculated transmembrane potential for cellswithpassiveNa+channelsbutnopassiveK+channels.
Event-related Potentials, ERPs – electrical activity measured by pairs of scalpelectrodesthatistimelockedtopresentationofasensorystimulussuchasapicture
Excitatory neuron – a neuron that releases a neurotransmitter that causes dendriticspikesinpostsynapticneurons
Explicit/declarativememory–memorythatcanberecalledatwill
Fibrousastrocytes – glial cells that position themselves throughout allwhitematterwheretheymakecontactwithaxonsatNodesofRanvier
Foramen magnum – a large hole in the base of the skull where the spinal cordconnectswiththebrainstem
FunctionalMagneticResonanceImaging,fMRI–avariationofMRIthatmeasuresflowofoxygenatedbloodintosmallvolumesofbraintissue
FunctionalNearInfraredSpectroscopy,fNIRS–animagingmethodthatmeasureschanges in brain blood flow by recording near infrared thermal radiation penetratingthroughbraintissue,boneandskin
Gapjunctions–membranestructuressharedbytwocellswithacentralopenchannelthatconnectstheircytoplasm
Glycogen–anstorageformofglucosewithincells
Graymatter–braintissuedominatedbylargeclustersofneuroncellbodies
Grossanatomy–theexternalfeaturesofadissectedtissueororgan
Hippocampus – the ancient part of the cerebral cortex that receivesmultiple inputsfromsensoryorgansandusesthatinformationtocodenewmemories
Implicit/proceduralmemory–memorybelowthelevelofconsciousawarenessthatformsasanexperienceisrepeated;recognitionofthecategorytowhichapieceofmusicbelongsisanexampleofimplicitmemory
Inhibitory neuron – a neuron that releases a neurotransmitter that blocks dendriticspikingatapostsynapticneuron

Interneuron–asmallinhibitoryneuronthatregulatestheactivityoflargerneurons
Interstitialfluid–theproteinfreefluidthatsurroundscells
Ion – an atom that lacks a match in its number of positive particles, protons, andnegativeparticles,electrons
Ion exchange pump – an energy consuming protein that moves ions across a cellmembraneagainsttheirconcentrationgradient
Ligand–asmallmoleculeswho’sbindingtocellproteinsregulatescellperformancethroughavarietyofmechanisms
Local field potential – an electrical potential in the brain created by opening ofmembrane ionchannelsatmultiple synapsesand the resultantmovementof ionswithinthetissue
Long term memory – includes both explicit/declarative and implicit/proceduralmemory
Longtermpotentiation – an increase in the ability of brain synapses to respond toneurotransmitter after receiving a rapid burst of signaling activity induced by externalelectrodes
Macrophage–acellofthebody’simmunesystemthatsurroundspathogensanddeadcellsandrecyclestheircomponents
Magnetic Resonance Imaging, MRI – a noninvasive procedure that produces ananatomicpictureoflargebrainsstructuresinvirtualslicesoflivingtissue
Magnetoencephalography,MEG – a noninvasive procedure that records magneticfieldsproducedbyelectricalcurrentsflowingthroughbraincircuits
Medullaoblongata–anothernamefortheaxonsmakingupthebrainstemthemeanslongwhiterope
Meninges–protectivemembranescoveringtheentirebrainandspinalcord
Mesencephalon–adivisionofthebraindeepinthecenteroftheorganalsocalledthemidbrainthatcoordinatescomplexreflexreactions
Metencephalon–thecerebellumandthepons
Microglia–nonneuroncellsofthebrainthatmonitorsneuronwell-beingandprovidethebrain’simmuneresponsetopathogens
Mitochondria–structuresinthecytoplasmofcellscontainingenzymesnecessaryforconversionoffoodtouseableenergy
Myelencephalon–anothernameforthebrainstem
Myelin–thewhitefattymaterialcoveringneuronaxons
Neo-cortex–throughevolutionthemostrecentlydevelopedpartofthecerebralcortex
Nerve–abundleofneuronaxons
Neural stem cell– partially differentiated brain stem cell capable of providing

replacementneuronsandoligodendrocytes
Neurogenesis–thebirthofnewneuronsfromneuralstemcells
Neuroglia–theoligodendrocytesandastrocytesofthebrain
Neuron–anindividualelectricalcellofthebrainorspinalcord
Neuroncircuit–anorganizedgroupofneuronsthatoperatetogetherasasingleunit
Neuroplasticity– thebrain’sability torearrangeitsdendritesanddendriticspines inresponsetosensorystimulationsuchassoundandlight
Neurotransmitter–achemicalreleasedbyanaxonterminaltosignaltoanothercell
NodesofRanvier–barepatchesofaxonbetweenmyelinlayers
Nucleus – in brain a cluster of neuron cells bodies in thewhitematter beneath thecerebralcortex;withincellsacompartmentthathousesgeneticmaterial
Oligodendrocytes–braincellsthatwrapneuronaxonswithmyelin
Passiveionchannel–acellmembraneporeforionsthatremainsconstantlyopen
Phonemes – the smallest units of speech in a language capable of conveying adistinctioninmeaning
Piamater – innermost of the membranes covering the brain adhering to the outersurfaceofthecerebralcortexandformingasheatharoundarteriesenteringthebrain
Pons–aspanofbraintissuethatconnectsthecerebellumtothecerebralhemispheres
Positronemissiontomography,PET–abrainimagingmethodthatrequiresinjectionof a radioactivemolecule that can cross the blood brain barrier and emit a signal afterbindingtoabrainsubstance
Postsynaptic density – a thickening of the neuron membrane that containsneurotransmitterresponseelements
Prosencephalon–theforebrainintheembryothatmaturesintotheretina,opticnerve,iris,cerebralhemispheres,thalamusandhypothalamus
Protoplasmic astrocytes – glial cells of the gray matter that surround neuron cellbodies
Radial cells – embryonic stem cells that produce neurons, astrocytes andoligodendrocytes
Reactive astrocytes – astrocytes that respond to damage in the brain by removingexcess toxic glutamate, producing anti-oxidants and walling off damaged tissue fromhealthytissuewithscarformations
Receptor–agenerictermforabroadclassofproteinsactivatedbyspecificchemicalscalledligandsthatregulatecellperformancethroughavarietyofmechanisms
Retrograde–directionofmovementwithinneuronsofmaterialtravellingtowardthecellbody
Rhombocephalon – in the embryo the last brain division which matures into the

cerebellum,ponsandbrainstem
Semanticmemory–memoryoffactslikenumberofhoursinadayandmeaningofaword
Shorttermmemory–anothernameforworkingmemorywhichisadynamicformofmemorycombiningmanythingsfrompastlearningtogetherwithpresentexperiences
Superiorsagittalsinus–anareabetweenthelayersofduramaterwherevenousbloodandcerebrospinalfluidpoolontheirwaybacktotheheart
Surveying microglia – the form of microglia that monitors neuron synapses andprovidesrepairtodamagedneurons
Symporter–aproteinpumpthatmovestwoormoredifferentmolecules,orions,inthe same direction across amembrane where at least onmolecule is moving down itsconcentrationgradientandoneisbeingmovedagainstitsconcentrationgradient
Synapse–structureattheplacewhereneuronscontacteachotherorcontactothercells
Synapticcleft–thegapataneuronsynapseofabout20nanometers
Tau–asolubleproteinwithinneuronsessentialforregulationofthemicrotubulesthatmovematerialfromplacetoplaceinthecell
Tau tangles – a form of tau that precipitates within neurons as pairs of helicalfilamentsandaggregatesintoastructurevisiblewithalightmicroscope
Telencephalon–intheembryothefarendoftheneuraltubethatmaturesintotherightandlefthemispheres
Thalamus–thepartofthebrainthatservesasanentrypointforinformationcomingtothebrainfromtherestofbody
Tract tracing –charting thepathofneuronaxons throughbrain tissuesby injectingdyeandobservingitsmovementwithinthecell
Transmembranepotential–thedifferenceinelectricalfieldpotentialcreatedbyionsandchargedproteinsontwosidesofacellmembrane
Ventricles–thefourhollowchambersinthecenterofthebrain
Vertebralarteries –branchesof the largearteries supplyingblood to the shoulders,lateralchestandarmsthatrunthroughthecervicalvertebraeandintotheheadwheretheyperfusethebackofthebrain
Virchow-RobinSpace–theareaaroundbrainarteriescreatedbythepiamatersheath
Voltage-sensitiveionchannel–achannelforionsthroughacellmembranethatopensandclosesinresponsetochangesinthetransmembranepotential
Whitematter–areasofthebrainwithfewneuroncellbodiesbutmanyneuronaxons
Workingmemory–adynamicformofmemorycombiningmanythingsfrompastlearningtogetherwithpresentexperiences

ABOUTTHEAUTHOR
Margaret Thompson Reece PhD, former Senior Scientist in academic medicine andChiefScientificOfficeratSerometrixLLC,headsReeceBiomedicalConsultingLLC.
Dr.Reecehelpsstudentswhostruggletofigureouthowtostudyhumananatomyandphysiology throughherwebsitehttp://www.medicalsciencenavigator.com/media-kit/, andher speaking and writing. Dr. Reece consults privately with a small selected group ofstudents.ShelivesinupstateNewYork.