mathematical modeling of opiate binding to mouse brain membrane
TRANSCRIPT

Bulletin ofMathematicalBiology Vol. 47, No. 4, pp. 503-512, 1985. Printed in Great Britain
0092-8240/85S3.00 + 0.00 Pergamon Press Ltd.
�9 1985 Society for Mathematical BioLogy
M A T H E M A T I C A L M O D E L I N G O F O P I A T E B I N D I N G T O M O U S E B R A I N M E M B R A N E
�9 HERBERT D. LANDAHL,* JAVIER GARZ~)N~ and NANCY M. LEE Department of Biochemistry and Biophysics* and Department of Pharmacology and Langley Porter Institute, University of California, San Francisco, CA 94143, U.S.A.
The heterogeneity of rat brain opiate receptors was examined by analyzing competi t ion data. The binding of three prototypical tri t iated opioid agonists, [3H]-dihydromorphine ([3HI-DHM), [3H]-D-alaLD-leuS-enkephalin ([3H]-DADLE), and [3H]-ethylketocycla- zocine ([3H]-EKC) was determined in the presence of varying concentrations of each of these unlabeled ligands, generating nine displacement curves. A computer program was then used to find the best fit of a model system to these data, assuming two, three or four independent binding sites. The best fit was a four-site model. One of these sites is specific for DHM; two are relatively selective for DHM and DADLE respectively, but also bind EKC. The remaining site binds only EKC with high affinity. These results, together with displacement data using naloxone, FK33824, and D-alaLmetS-enkephalinamide, are discussed in terms of current opiate receptor models.
Introduction. It is now widely accepted that multiple types of receptors exist for opioid ligands, yet the nature of these receptors remains mysterious in several ways. For one thing, though every class of receptors presumably interacts with a specific, endogenous ligand, the identity of this ligand is in many cases unknown. Indeed, the prototypical or 'pure' opioid correspond- ing to each receptor type is still controversial. These receptors were originally defined as those sites specifically or selectively interacting with one class of opioids. Thus/z sites are selective for morphine, 6 for the enkephalins, and ~: for ethylketocyclazocine (EKC). This definition is problematical, how- ever, in that all of these drugs may interact with other receptor types.
Conventionally, the Scatchard plot has been used to help define a popula- tion of receptors binding a particular kind of opiate. Scatchard plots of almost any opiate reveal a curvilinear pattern, however, suggesting that the ligand binds to several sites. It is also difficult to use this method to cover a wide range of drug concentrations, with data at very low and high concentrat ions-- that is, binding to highest and lowest affinity s i tes-- being subject to large experimental error. Error is also increased by the fact that both ordinates depend on the free drug concentration, which may be difficult to determine with precision.
] Current address: Cajal Institute (C.S.I.C.), Department of Neurophaxmacology, Valasquez 144, Madrid, 28006, Spain.
503
504 H.D. LANDAHL et al.
As an alternative approach to Scatchard plots, we have studied opiate receptor heterogeneity using a displacement technique. Here we report the results of these studies and some of the advantages this approach offers.
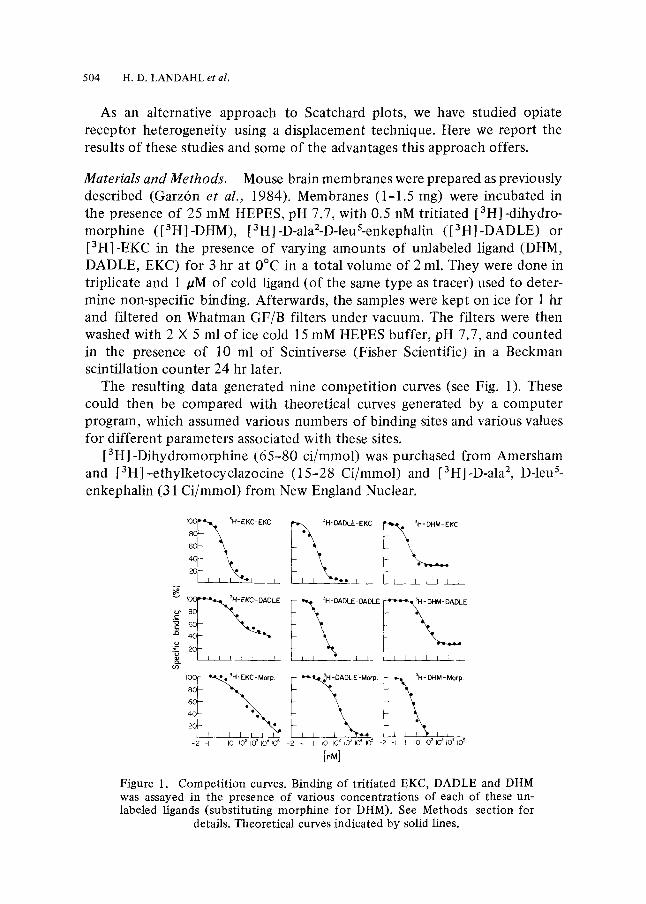
Materials and Methods. Mouse brain membranes were prepared as previously described (Garz6n et al., 1984). Membranes (1-1.5 rag) were incubated in the presence of 25 mM HEPES, pH 7.7, with 0.5 nM tritiated [3HI -dihydro- morphine ([3H]-DHM), [3H]-D-ala2-D-leuS-enkephalin ([3H]-DADLE) or [3H]-EKC in the presence of varying amounts of unlabeled ligand (DHM, DADLE, EKC) for 3 hr at 0~ in a total volume of 2 ml. They were done in triplicate and 1 /sM of cold ligand (of the same type as tracer) used to deter- mine non-specific binding. Afterwards, the samples were kept on ice for 1 hr and filtered on Whatman GF/B filters under vacuum. The filters were then washed with 2 • 5 ml of ice cold 15 mM HEPES buffer, pH 7.7, and counted in the presence of 10 ml of Scintiverse (Fisher Scientific) in a Beckman scintillation counter 24 hr later.
The resulting data generated nine competit ion curves (see Fig. 1). These could then be compared with theoretical curves generated by a computer program, which assumed various numbers of binding sites and various values for different parameters associated with these sites.
[3H]-Dihydromorphine (65-80 ci/mmol) was purchased from Amersham and [3H]-ethylketocyclazocine (15-28 Ci/mmol) and [3Hl-D-ala2, D-leu s- enkephalin (31 Ci/mmol) from New England Nuclear.
~ 3H'EKC-EKc ~ ~
E 80 7 m
4 0
2 0
I00 16080 ~ - Mot p.
2 0 I I ]
-2 -I I I0 JO ~ I0 ~ ~0" FO 5
~ KC
I I I I 1 ~ 5 _
-2 -I I I0 102103104 I0 ~ -2 -I I I0 ~02 I0 ~ tO 4105
[oM]
Figure 1. Competition curves. Binding of tritiated EKC, DADLE and DHM was assayed in the presence of various concentrations of each of these un- labeled ligands (substituting morphine for DHM). See Methods section for
details. Theoretical curves indicated by solid lines.

MATHEMATICAL MODELING OF OPIATE BINDING 505
Morphine and EKC were gifts from NIDA and [3H]-D-alaZ-D-leu s- enkephalin, D-ala2-metS-enkephalinamide (DAMA) and FK33824 were purchased from Peninsula Laboratories (San Carlos, California). Naloxone was from Endo Corporation.
Mathematical Model. For each site i, of total number Rot, the number of tracer molecules of concentration (T) bound to the receptor will be (TRi) = (T)(Ri)/Kir, where Ri is the total number of occupied receptors and KiT is the dissociation constant of receptor i for tracer T; likewise, the number of drug molecules of concentration D bound to the receptor will be (DRi) = (D)(Ri/KiD). The total number Roi is just the sum of the free R i plus (TRi) and (DRi) from which:
so that
Ri = Roi/(1 + T/KiT + D/KiD) (1)
(TRi) = RoiT/(KiT + T + DKiT/KiD). (2)
The measured value YTD is the ratio of the sum of the amounts of tracer bound to each receptor in the presence of tracer and drug (in amount D) divided by the total amount bound when the tracer has a concentration T, but D = 0. That is, the specific binding for each of the nine curves in Fig. 1 is given by:
YrD = y~ (TRi)/Z (TRi)D=O (3)
or substituting (2) into (3),
~- (Roi)/(KiT + T + DKir/Kio) YTO = (4)
ZRoi/(Kir + T)
If T and D refer to the same drug, T and D are concentrations of labeled and unlabeled drug, respectively, and KiT = KiD. In all experiments T = 0.5 nM. The quanti ty T is retained in equation (4) since a number of K values were found which were substantially less than T.
Results and Discussion. The best fit under these conditions for the data shown in Fig. 1 proved to be a model assuming four independent sites, two of which were DHM-specific sites (sites 3 and 4).
For reasons to be explained later, one can estimate from asymptotic values in the curves [3H]-DHM-EKC and [3H]-DHM-DADLE the number of one of the DHM-'specific' sites (site 4) to be one-third of the number of the other DHM-sensitive site (site 3). Inspection of the [3H]-DHM-morphine curve indicates that the K value (association constant) of DHM-'specific' site 4 must be close to that for DHM-specific site 3, because the Scatchard

506 H.D. LANDAHL et al.
plot is essentially linear. If we tentatively assume them to be equal, this leaves nine K values and two fractions, the ratio of the number of sites No. 2 to No. 1 and that of No. 3 to No. 1, to be determined from the nine curves. These calculations were carried out using a non-linear least-squares procedure to obtain the coefficient giving the least sum of the squares of differences between calculated and observed values for all 109 points of the nine curves of Fig. 1, the error estimates on the parameters being made from the covariance matrix using the linear expansion about the solution. Since the system is far from linear, the error ranges should be used as guide- lines only. All points were weighted equally. The program ensured that all coefficients remained positive.
In our example, if site 3 (m) refers to one DHM site and 4 (rn*) refers to the other DHM site, then KmM = Krn,M, approximately, while Km* E =
Km*D = oo (M referring to DHM, E to EKC and D to DADLE): the site 4 is referred to as 'specific' because the dissociation constants for all but DHM are effectively infinite (association being zero). In addition, Rom* = Rom/3. Since there is an arbitrary scale factor in the Roi, we can set Rol = 1, leaving Ro2 and Roa as unknowns, together with the nine unknown values KIM , K1E , K1D . . . . , K3D. The results presented in the accompanying figures show computed and observed values for each of the nine curves. The parameters are summarized in Table I. The curve [3H]-EKC-DADLE needs to be extended to higher concentrations of DADLE to determine the coefficients more fully.
A useful consequence of the model is to be able to give a better estimate as to what the tritiated drugs are binding to. Since they are at low concentra- tions, the amount of activity associated with each site is the ratio Roi/KiT. If
TABLE I Association Constant (nM)
Site Error* (S.D.)
1 2A 2B 3
EKC 1.68 -+ 0.23 2.31 -+0.79 t 3 . 2 0 + 0 . 4 5 / DADLE 4.02 + 0.50 56700-+ 14000 ~ 51.3-+ 7.0 I 0 ,042 DHM 253 + 41 19600 + 5700 t 4.66 -+ 0.43 Nx 19.6 -+ 5.7 0.10 -+ 0.03 ~ 1 0 6 5.63 -+ 0.76 0.048 FK 36.3 + 2.4 6000 -+4600 ~ 1 0 6 1.23 + 0.11 0,028 DAMA 0.17-+0.01 5 4 0 0 - + 3 0 0 0 220,000-+- 110,000 0 .37-+0.05 0,042 Roi 1.0 2.03 -+ 0.68 1.82 -+ 0.49
* Root mean square error for three curves using three tracers, EKC, DADLE and DHM for each drug.
Site 4 is not in the table, since only DHM binds to 4 and does with approximately equal affinity to 3.
~ Results cannot distinguish between 2A and 2B.

MATHEMATICAL MODELING OF OPIATE BINDING 507
Fi is the normalized value of this ratio, then for [3HI-DHM, F1 = 0.01, F 2 = 0.00, F 3 = 0.74, and F4 = 0.25, with the result that one has a nearly pure mixture of morphine (site 3) and morphine-'specific' (site 4) binding sites in the proport ion 3 : 1. For [3tt]-DADLE* the proport ions F/ are 0.87, 0.00, 0.13, 0.00, so that with [3H]-DADLE* one has a large pre- ponderance of No. 1 binding sites, with the remainder consisting of No. 3 binding sites. For [3H]-EKC the proport ions are 0.29, 0.43, 0.28, 0.00. From these results one may refer to site 1 as a DADLE-preferred site, 3 as a DHM-preferred site, and 4 as a DHM-specific site. These values are shown in Table II.
TABLE II
Apparent Occupancy (F) for Various Drugs
F1 F3 F4 F2A F2B
EKC 0.291 0.278 0.000 0.151 0.279 DADLE 0.875 0.125 0.000 0.000 0.000 DHM 0.009 0.743 0.248 0.000 0.000
Nx 0.007 0.043 0.000 0.950 0.000 FK 0.018 0.982 0.000 0.000 0.000 DAMA 0.540 0.459 0.001 0.000 0.000
Note that in the entire above analysis we have assumed that K4,M = K3,M. If one uses the appropriate values given K4,M in the table to compute the data for [3H]-DHM-morphine curves, but increases or decreases K4,M relative to K3,M one finds that a factor of three decreases the fit of the data very noticeably. From this we conclude that K4,M = K3,M, within a factor of three. While this may seem a rather wide range, we should remember that the K values vary from 0.1 to more than 106. In this connection, we note that because of the way the Ks enter into the equations, the quanti ty i -+ 0.5 is perhaps bet ter interpreted as being 1 within a factor of 1.5.
In addition to the above analysis, an a t tempt was made to fit the data to a model with two sites in which there is interaction between sites. The efforts were unsuccessful even though twelve parameters are involved, six K values and six interaction coefficients, these in addition to the morphine-'specific' site.
The effects of three more drugs, naloxone (Nx), FK33824, and D-ala 2- metS-enkephalinamide (DAMA), were also analyzed for the binding of the three tracers, generating nine more curves (Fig. 2). Using the K values for the tracers and the amounts Roi of receptors already estimated, the three curves for each new drug were analyzed to obtain three K values for each

508 H.D. LANDAHL et al.
I O O ~ g H - E KC - Nolo•
4C ~ e
2c i �9 E I C s
60 �9 �9
I ~ ) ~ 3H-EKC-DAMA
4c I 2C
I
] ] I I ] I I
~ 1 I I l I ~ _ - 2 -I I I0 I0 'z I0 3 10 4 I0 ~ -2 -[ I I0 f{3 2 I0 ~ PO 4 IO ~ -2 -I I I0 IO t I0 ~ I0 4 pO s
[riM]
Figure 2. Competi t ion curves. As in Fig. 1, except the unlabeled ligands used were Nx, FK 33824, and DAMA.
drug. None of these drugs had any detectable interaction with the DHM- 'specific' site. The proportions of m* in the experiments using these drugs, as estimated from the asymptotic values, were from about 0.2 to 0.3. There were a few cases in which there was duplication of experiments over a period of time with results that indicated significant variation in this fraction. Hence, rather than force them to be equal to the value of 0.25 (1 : 3) used for the previous nine curves, the individual asymptotic values were used. This had no appreciable effect on the parameter estimates but appreciably reduced the average error and reduced the size of the error estimates. Since [3H] -DHM is almost exclusively tagged to sites 3 and 4 but the drugs do not react with 4, K3x for each drug X can be estimated directly from the [aH]-DHM-X data. Since [3H]-DADLE is about 85% bound to site 1, and K3x is now known, one can obtain Klx almost independently of the K3x value from the [3H]-DADLE-X data. There remains to be estimated the K values for site 2 from the [3H]-EKC-X data.
However, there remained substantial discrepancies with the curves involv- ing [3H] -EKC which could be greatly reduce if site 2 is, in fact, two distinct sites which happen to have comparable K values with respect to EKC. Unfortunately, introducing this distinction into the model, of course, requires assuming a proportion for each. Trying proportions from 1/4 to 1/2 showed that at about 0.3-0.4, the fit was best. The data for each drug were then reanalyzed using 0.35, these results being shown in Fig. 2 and Tables I and II. An inspection of the figures shows that the computed curves give a satisfactory representation of the data with two possible exceptions.

MATHEMATICAL MODELING OF OPIATE BINDING 509
The data for [3H]-DADLE-Nx in Fig. 2 suggest that the theoretical curve should perhaps be steeper. This could occur if the site 1 were double, at least with respect to Nx, and if there were a strong cooperative interaction between the Nx molecules at the two sites. But we do not feel that the present data warrant introducing this complexity.
In the case of [3H]-EKC-DAMA, the computed curve (solid curve, Fig. 2 K2A = 5800, K2B = 220,000) is considerably below the data at low concen- trations. By arbitrarily increasing Ro2 by 1.5 times its error estimate while decreasing Ro3 by 1.5 times its error estimate one can reduce the average error from 0.11 to 0.06, the resulting curve falling between the two curves in Fig. 2. This could be justified on the grounds that there are variations in the proportions of the different sites as, for example, the variability in the proport ions of site 4, and because the experiments were necessarily carried out over an extended period of time. However, the fit is still not satisfactory and suggests that some interaction needs to be introduced. The discrepancy between theory and experiment suggest that DAMA, although binding substantially at low concentrations to site 3, does not displace the EKC tracer. Since this type of discrepancy is even more pro- nounced in the case of dynorphin and many of its fragments (unpublished results), it is justified to consider the possibility that site 3 is a pair of sites such that none of the above drugs can occupy them simultaneously, with the exception of EKC and DAMA. For the case in which EKC and DAMA can occupy either site 3' or 3" (with constants KyD and Ka"D), and if we neglect moieties occupied by two tracers, then YTD for EKC as tracer and DAMA (D) as competi tor will be given by (4) in which the third term (i = 3), corresponding to (TRi)/T is replaced by:
where
in which
(TRs)/T = Ro3G
K37, + TG + K3rD/K~o + K3rD2/(4PDD/K~D) (5)
G = 1 4-D/(PTDK3D)
K3D = KyD Ky'D/(Ka'D + Ky'D),
K3T = KyTK3, ,T / (Ky T + Ky'T)
PTD = (KyT + K3"T)(KyD + Ky'D)
(KyD Ky'T + KY'D K3'7)
P
PDO = POD (KyD + K3"D)2/KyD KY'D.

510 H.D. LANDAHL et al.
Here K3D, K3T, PDD and PTD are apparent coefficients corresponding to the intrinsic (primed) coefficients, /)TO measuring tracer-drug interaction and POD measuring drug-drug interaction, the Ks being the apparent single site values. If PTD = oo, the competi t ion between tracer and drug is complete. If Pro = 1, then T a n d D act independently at sites 3' and 3", while PTD < 1 corresponds to positive cooperativity; and similarly for POD. Note that two additional parameters have been introduced. If we apply the case with PDD = oo to the data for [3H]-EKC-DAMA interaction, we obtain a curve close to the dashed curve shown in Fig. 2, the error being 0.033, a sub- stantial improvement over 0.06 and 0.11, the values mentioned above. Since P2"o is approx. 1, the tracer and drug do not interfere with each other but only one of the pair of sites can be occupied by the drug at a given moment in this case. If we do not force PDO to be infinite, then we have the result shown by the dashed curve for [3H]-EKC-DAMA in Fig. 2 for which K2A = 5400 -+ 3000, K2B = 220,000 -+ 110,000, POT = 0.62 + 0.08 and PDD = 2.0 -+ 0.6 there being only moderate interaction, the coefficients being not too far from unity. We note in passing that if site 1 were a double site instead of 3, one would obtain similar results, though not as satisfactory.
For Nx and FK, the K value for either 2A or 2B tended to exceed 108, that is, there was no detectable association. In every case, a search for a solution was made again, starting with values 2A and 2B interchanged. In each case the fit was not as good as before, in some cases much worse. However, in view of the possible variations in relative amounts of different receptor binding sites as discussed above, not too much should be made for any one case, and these K values could possibly be interchanged.
The values of the dissociation constants in Table I are suggestive. For example, using EKC as the tracer, addition of large amounts of two drugs which have no effect on 2B will have no additive effect, and similarly for 2A. But addition of one drug in large concentration having an effect on 2A, and another having an effect on 2B, will result in complete loss of binding of [aH] -EKC according to the model. Data of this kind are needed for further characterization of the sites 2A and 2B.
Table lI shows also the proport ions of activity of each of the receptor sites for each of the three additional drugs. Assuming that a tracer label for each is possible, the values/;1, F3, �9 �9 are the proport ion of each drug at tracer concentration attached to sites 1, 3 . . . . Viewed in this way, the following conclusions are apparent: (1) FK prefers site No. 3; (2) DADLE is the drug most specific with respect to 1; (3) Nx appears to be highly specific for site 2A and thus useful in distinguishing 2A from 2B; (4) only EKC has an appreciable fraction associated with 2B; and (5) even though DHM can block [3H]-EKC associated with 2B, their fractions associated with 2B are at very low levels.

MATHEMATICAL MODELING OF OPIATE BINDING 511
Note that although DAMA has a higher affinity for site 1 than does DADLE, the ratio of K for this site to the site of the next highest affinity is about 10-fold (51/4.0) for DADLE, as compared to a value of a little more than 2 (0.37/0.17) for DAMA. Hence, DADLE shows the greatest selectivity for site No. 1.
Although DHM has less affinity to the 'p' site (No. 3) than three of the other four drugs, it is the most selective with respect to this site, the ratio being 54(253/4.66) compared to a value of 30(36/1.2) for FK, the only comparable value.
Several models of the opiate receptor have recently been proposed that explicitly take into account the existence of multiple binding sites. The traditional model assumed that each ligand bound to a distinct receptor molecule, though ligands specific for one type of receptor were believed able to bind, with somewhat lower affinity, to the other receptors (Lord et al., 1977; Chang and Cuatrecasas, 1979). Several years ago, however, we pro- posed a receptor model containing both p and 6 sites in a single molecular complex (Lee and Smith, 1980). A similar model was later proposed by Rothman and Westfall (Rothman and Westfall, 1982a, b), who reported evidence that the Is and 6 sites allosterically interact with one another, so that ligands specific for one site may act as non-competitive inhibitors of binding at the other site. In this study, it is clear that site 1 is equivalent to the 6 site and site 3, the ~ site in both these models. On the other hand, Pasternak (Pasternak, 1982)has suggested that both p and 6 ligands interact with a single high affinity site.
How the additional sites suggested by this study could fit into one of these schemes remains to be determined. It is possible that some or all of them exist in a complex with p and 6. This arrangement would be consistent with the non-competitive interaction observed between different ligands in this study. For example, if dynorphin binds directly to a ~: site in the brain, the existence of this site in close association with the other sites would enable a modulating action to be achieved through allosteric effects, as preliminary reports seem to indicate (Tulunay et al., 1981; Garz6n et al., 1984).
However, it is conceivable that non-competitive effects could occur between binding sites on distinct molecules. Furthermore, it seems unlikely that a single receptor complex would contain as many as five distinct types of sites. This conclusion also seems incompatible with available data on the brain regional distribution of different opiate binding sites (Goodman et al., 1980).
Currently, we are applying a similar approach to endogenous opioid peptides, such as dynorphin, B-endorphin and enkephalins and their frag- ments. The peptide dynophin is of particular interest because, as we have

512 H.D. LANDAHLetal.
shown previously (Friedman e t al., 1981), it modulates the action of the morphine. This modula tory action may result from dynorphin 's ability to
interact with several o f these receptors.
S u m m a r y . The use o f displacement curves allows us to approach the issue of multiple receptors in a way that avoids many o f the problems associated with Scatchard plots. We have been able to identify five different opiate binding sites, and characterize each according to the affinity and relative amount of binding it exhibits with respect to six different ligands. While we cannot, of course, rule out the possibility of there being still more sites, these five are necessary and adequate to explain the current data.
The work was supported in part by National Institute on Drug Abuse Grant DA 02643. J. G. was recipient of a MEC Fellowship and N. M. L. is recipient of a National Institute on Drug Abuse Research Scientist Career Award.
LITERATURE
Chang, K. J. and P. Cuatrecasas. 1979. "Multiple Opiate Receptors: Enkephalin and Morphine Bind to Receptors of Different Specificities." J. biol. Chem. 254, 2610.
Friedman, H. H., M. F. Jen, J. K. Chang, N. M. Lee and H. H. Loh. 1981. "Dynorphin: A Possible Modulatory Peptide on Morphine or ~-Endorphin Analgesia in Mouse." Europ. J. PharmacoL 69, 357.
Garz6n, J., P. Sfinchez-Blfizquez, J. Gerhart, H. H. Loh and N. M. Lee. 1984. "Dynor- phin 1-13: Interaction with Other Opiate Ligand Bindings in Vitro." Brain Res. 302, 392.
Goodman, R. R., S. H. Snyder, M. J. Kuhar and W. S. Young, III. 1980. "Differentiation of Delta and Mu Opiate Receptor Localization by Light Microscopic Autoradiography." Proc. natn. Acad. ScL U.S.A. 77, 6239-6243.
Lee, N. M. and A. P. Smith. 1980. "A Protein-Lipid Model of the Opiate Receptor." Life Sci. 26, 1454.
Lord, J. A. H., A. A. Waterfield, J. Hughes and H. W. Kosterlitz. 1977. "Endogenous Opioid Peptides: Multiple Agonists and Receptors." Nature 267, 495.
Pasternak, G. W. 1982. "High and Low Affinity Opioid Binding Sites: Relationship to Mu and Delta Sites." Life ScL 31, 1303.
Rothman, R. B. and T. C. Westfall. 1982a. "Morphine AUosteric Coupling Between Morphine and Enkephalin Receptors in Vitro." Mol. Pharmacol. 21, 548.
- - a n d - - . 1982b. "Morphine Allosterically Modulates the Binding of 3H-Leucine- enkephalin to a Particulate Fraction of Mouse Brain." Mol. PharmacoL 21,538.
Tulunay, F. C., M. F. Jen, J. K. Chang, H. H. Loh and N. M. Lee. 1981. "Possible Regula- tory Role of Dynorphin on Morphine- and Endorphin-induced Analgesia." J. Pharmac. exp. Ther. 219, 256.
RECEIVED 2-18-85