mathematicalmodellingofnitricoxideregulationofretepegforma ...jas/paperpdfs/nitricoxide1.pdf ·...
TRANSCRIPT

J. theor. Biol. (2002) 214, 1}16doi:10.1006/jtbi.2001.2400, available online at http://www.idealibrary.com on
Mathematical Modelling of Nitric Oxide Regulation of Rete Peg Formationin Psoriasis
NICHOLAS J. SAVILL*-, RICHARD WELLER? AND JONATHAN A. SHERRATT*
*Centre for ¹heoretical Modelling in Medicine, Department of Mathematics, Heriot}=att ;niversity,Edinburgh EH14 4AS, ;.K. and ?Department of Surgery, ;PMC, E 1557 Biomedical Science ¹ower,
200 ¸othrop Street, Pittsburgh, PA 15261, ;.S.A.
(Received on 9 November 2000, Accepted in revised form on 16 July 2001)
Recent experiments have shown that in patients with psoriasis, highly elevated levels of nitricoxide (NO) are released at the surface of psoriatic plaques. Nitric oxide is a central biologicalregulator of many aspects of physiology, and it is a natural possibility that the high nitric oxidelevels in psoriasis play a causal role in the onset of the disease. Here, we use mathematicalmodelling to investigate this possibility. We begin by discussing a simple model consisting ofa single equation for nitric oxide concentration, which enables nitric oxide secretion rates inthe basal epidermis to be calculated from the observed NO release rates at the skin surface.Using this key parameter value, we then develop an extended model that tests the hypothesisthat nitric oxide regulates the formation of the extended rete pegs seen in psoriatic plaques.This occurs via the peroxynitrite-dependent activation of the collagenase MMP}8, which isproduced by neutrophils present at high levels in psoriatic plaques. The plausibility of thehypothesis is demonstrated and speci"c testable quantitative predictions about the roles of thevarious cell types and signalling molecules are made.
( 2002 Academic Press
1. Introduction
Nitric oxide has long been known as an environ-mental pollutant. But in the late 1980s it was alsodiscovered to be a key physiological regulator,when a substance known previously as en-dothelium-derived relaxing factor was identi"edas nitric oxide (Ignarro et al., 1987). It was for thisand related work that Furchgott, Ignarro andMurad were awarded the 1998 Nobel prize forphysiology and medicine. Blood vessels dilate inresponse to nitric oxide, and blood #ow is regu-lated by the production of nitric oxide by theendothelial cells lining blood vessels. This dis-
-Author to whom correspondence should be addressed.E-mails: [email protected], [email protected], [email protected]
0022}5193/02/010001#16 $35.00/0
covery was rapidly followed by the "nding thatnitric oxide was also responsible for the killing ofbacterial cells by macrophages (Hibbs et al.,1989). In the last decade, there has been an ex-plosion of experimental research on nitric oxide:it was named &&molecule of the year'' in 1992 bythe journal Science (Culotta & Koshland Jr,1992), and is now known to regulate biologicalfunction in most tissues in the body [see Mon-cada (1999) for review]. One of the many medicalconditions in which nitric oxide may play a role ispsoriasis.
Psoriasis is one of the world's most commonskin diseases, a!ecting about 2% of the popula-tion; nevertheless, its biological cause is un-known. Normal skin consists of two layers: theepidermis, consisting of predominantly a single
( 2002 Academic Press

2 N. J. SAVILL E¹ A¸.
close-packed cell type called keratinocytes andbelow this the dermis, composed of a collagen-based matrix within which reside "broblast cells,blood vessels, nerve endings, and epidermal ap-pendages such as hair follicles and sweat glands.The boundary between the epidermis and dermishas characteristic spatial oscillations known asrete ridges. In psoriasis, the epidermis is signi"-cantly thickened and poorly di!erentiated, caus-ing #aking at the skin surface; it is also in"ltratedby lymphocytes (white blood cells). The interfacebetween the epidermis and dermis also changes,with very pronounced epidermal (rete) pegs pro-truding into the dermis [compare Fig. 1(a) and1(b)]. In the most common form of psoriasis(&&chronic plaque psoriasis''), these variouschanges occur in discrete plaques, separated byuna!ected skin. These features characterizingpsoriasis have all been known for many years,but the underlying molecular cause(s) of the dis-ease remain unknown.
A recent development in the understanding ofpsoriasis has been the discovery that psoriaticplaques (a.k.a. lesions) actively produce nitric ox-ide. Kolb-Bachofen et al. (1994) "rst showed thatepidermal keratinocytes in plaques produce theenzyme &&inducible nitric oxide synthase'', whichis part of a nitric oxide production pathway, andWeller et al. (1996a) showed that nitric oxidesynthesis is indeed increased in psoriasis. Healthyhuman skin does produce nitric oxide, at verylow levels, as a result of chemical reactions withinsweat (Weller et al., 1996b). However, productionis about ten times higher in non-lesional skin ofpsoriatics, and about ten times higher again inthe plaques themselves (Weller & Ormerod, 1997;Orem et al., 1997; Ormerod et al., 1998). Theseobservations are highly suggestive of a role fornitric oxide in the development of psoriasis, al-though no mechanism has yet been established.From a clinical viewpoint, this is an excitingpossibility, because nitric oxide levels can be al-tered relatively easily, by topical application ofnitric oxide donors or inhibitors. However, a de-tailed quantitative understanding of nitric oxidelevels is an essential precursor to this, since thee!ects of nitric oxide are highly concentration-dependent.
In this paper, we begin (Section 2) by usinga mathematical model to estimate nitric oxide
concentrations within psoriatic plaques, based onexperimental data on nitric oxide release rates atthe skin surface. We then (Section 3) use an en-larged model to show how nitric oxide produc-tion can give rise to the characteristic pattern ofpronounced rete pegs within the plaques. This isthe "rst quantitative testable hypothesis fora plausible mechanism of rete peg formation.Mathematical modelling of nitric oxide in bio-logy has a relatively long history. In 1994, Lan-caster published a very in#uential paper in whichhe used a reaction}di!usion model to study thespatiotemporal distribution of nitric oxidearound a single blood vessel; this has sub-sequently been re"ned and revised (Lancaster,1997; Butler et al., 1998; Vaughn et al., 1998).Modelling has also been used to quantify thee!ects of nitric oxide donors (Schmidt et al.,1997), to study the kinetics of nitric oxide'scytotoxic e!ects (Laurent et al., 1996), and toinvestigate the potential role of nitric oxide inwound healing pathologies (Cobbold & Sherratt,2000). By contrast, psoriasis has, to the best ofour knowledge, not previously been studied usingmathematical modelling.
2. Modelling Nitric Oxide Productionin Psoriatic Plaques
The high production rates of nitric oxide inpsoriatic plaques have been established bymeasuring release rates of nitric oxide from theskin surface. This involves a procedure, de-veloped by Weller et al. (1996a), in which a nitro-gen-"lled vessel is placed over the skin surface,and the amount of nitric oxide collected overa prescribed time period (about 20 min) is mea-sured by chemiluminescence. This procedureprovides a quantitative comparison betweennitric oxide production in di!erent skin regions,di!erent individuals, etc. However, it does notgive direct information about the key issue ofnitric oxide concentration levels within psoriaticplaques; our "rst modelling goal is to predictthese concentrations.
Within psoriatic plaques, nitric oxide is se-creted mainly by the basal layer of keratinocytesas observed by staining for iNOS (Kolb-Bachofen et al., 1994; Ormerod et al., 1998). Weassume that the nitric oxide production rate is

FIG. 2. Schematic illustration of the model set-up withboundary conditions and spatial dimensions. Basal cellsmake up roughly 30% of the cells in the rete pegs. In themodel, the width of the basal cell layer needs to be about12 lm in order to achieve this percentage coverage.
MATHEMATICAL MODELLING OF NITRIC OXIDE 3
uniform within the basal layer of the epidermis,and study the nitric oxide release rate at the skinsurface that results from this. The model consistsof a single partial di!erential equation for nitricoxide concentration n (r
1, t). We restrict the atten-
tion to a two-dimensional cross-section of theskin, illustrated schematically in Fig. 2, and as-sume uniformity in the third dimension; thus,r1"(x, y) denotes a spatial position in two dimen-
sions, and t denotes time. The de"nitions of themodel's variables and parameters are given inTable 1 and a discussion of the parameter valuesis given in Appendix A.
The nitric oxide molecule is small and is sol-uble in phospholipid membranes and thus willdi!use at approximately the same (high) rate indi!erent layers of the skin, unimpeded by cells,extracellular matrix components, etc. However,the reaction kinetics will be quite di!erent fordi!erent regions. In the suprabasal epidermis,nitric oxide decays at a constant rate j due toreactions with a range of free radicals (Lancaster,1994). In the basal layer there is, in addition tothis decay, production of nitric oxide at a rate a
1.
Finally, within the dermis, the intrinsic decayrate is augmented by active scavenging of nitricoxide by blood, due to the rapid binding of nitricoxide to haemoglobin (Gow & Stamler, 1998).This is a key part of the model because it intro-duces an important nonlinearity; nitric oxidecauses blood vessel dilation, which will in turnresult in a higher volume of blood in the plaque,and thus more rapid removal of nitric oxide. Wedenote by g (n) the increase in diameter of bloodvessels relative to that in the healthy skin. Experi-mental data (Ku, 1996) indicate that an appropri-ate functional form is g (n)"g
0n/(g
1#n) with
g1+20 nM.The discussion implies the following form for
our mathematical model:
LnLt
"Dn+ 2n#
G!jn in suprabasal epidermis,
!jn#a1
in basal layer,
!jn!d1n(1#g(n)) in dermis.
(1)
For the purposes of this section, we imposethe divisions between these regions, based onthe observed form of psoriatic plaques (Fig. 2).We consider the nitric oxide release rate at theskin surface over an area much larger than theintracapillary distance. Therefore, the spatial do-main should be considered as an average overseveral square millimetres of skin. In the nextsection of the paper, we will go on to discuss therole played by nitric oxide in regulating the shapeof the epidermal}dermal junction.
We solved eqn (1) on a rectangular domainwith x and y coordinates parallel and perpen-dicular to the skin surface, respectively (seeFig. 2). The boundaries at x"0 and ¸ representthe centres of a rete peg and a dermal papilla,respectively, and no #ux conditions are appliedon these boundaries. Beyond the skin surface(y"0) is air [or, in experimental measurementof nitric oxide release, a nitrogen-"lled vessel

TABLE 1Parameter de,nitions and values for the two models
Parameter Description Value
d1
NO scavenging by Hb 100 s~1 (10 s~1 in model 1)d2
ONOO~ reaction CO2
58 s~1
d3
ONOO~ scavenging by Hb 30 s~1
d4
ProMMP-8 decay 0.01 s~1
d5
MMP-8 decay 0.01 s~1
d6
Collagen degradation by "broblasts 100 s~1
k1
No reaction with O~2
19 nM~1 s~1
k2
ProMMP-8 activation by ONOO~ 10~5 nM~1 s~1
k3
Collagen degradation by MMP-8 5]10~6 nM~1 s~1
a1
NO production by basal cells 170 nMs~1
a2¸ ProMMP-8 production by neutrophils 170 nM s~1
a3
Collagen production by "broblasts 5.36]104 lg mg~1 s~1 cell~1 lm3
Dn
NO di!usion coe$cient in tissue 3300 lm2 s~1
Dp
ONOO~ di!usion coe$cient in tissue 2600 lm2 s~1
g0
Maximal change in capillary cross-sectional area 9g1
[NO] for half-maximal change in capillary 20 nMj NO removal in tissue 0.17 s~1
s O~2
concentration in tissue 0.01 nMF Fibroblast density in dermis 10~8 cell lm~3
cT
Threshold collagen density 100 lg mg~1
n [NO]p [ONOO~]m@ [proMMP-8]m [MMP-8]c Collagen density
Note: See Appendix A for details and references.
4 N. J. SAVILL E¹ A¸.
(Ormerod et al., 1998)]. The di!usion coe$cientof nitric oxide in these media is about 1000 timesgreater than in biological tissues, and thus anynitric oxide released will di!use away from theskin surface e!ectively instantaneously. Thus, theappropriate boundary condition at y"0 isn"0. The fourth boundary y"> is included forcomputational necessity; we use Ln/Ly"0 asa boundary condition, but this is not signi"cantprovided that > is su$ciently large.
Our aim is a quantitative solution of thismodel, and thus, good parameter estimates areessential; fortunately, there is a large literature onquantitative properties of nitric oxide on whichthese parameters can be based. The di!usioncoe$cient D
nof nitric oxide in tissue was mea-
sured as 3300 lm2 s~1 by Malinski et al. (1993),using microsensor measurements. The parameterj is the decay rate of nitric oxide in tissues dueto reactions with free radicals, which is estimatedas 0.17 s~1, corresponding to a half-life of 4 s
(Lancaster, 1994). The parameter d1
can be esti-mated from the rate constant of nitric oxide bind-ing to haemoglobin in #owing blood, which is5]104 M~1 s~1 (Liao et al., 1999; note that thecorresponding rate for free haemoglobin is about1000 times larger). Blood vessels constitute about10% of dermal papillae in a typical psoriaticplaque (Auer et al., 1994) so that a typical concen-tration of 2 mM ("14 g per 100 ml) haemoglo-bin in the blood (Thibodeau & Patton, 1997)gives an average concentration of 0.2 mM in thedermal papillae. Thus, d
1"(5]104)](0.2]
10~3) s~1"10 s~1. We base the value of g0
onestimates that blood #ow in the dermal papillaeof psoriatic plaques is between two and "ve timesgreater than in normal skin (Klemp & Staberg,1983; Krogstad et al., 1995). Assuming Poiseuille#ow in the vessels, this implies an increase inblood vessel area by factor of between 1.4 and 2.2(J2}J5). Thus, the average value of 1#g(n) inthe dermal papillae dermis should be about 1.8.
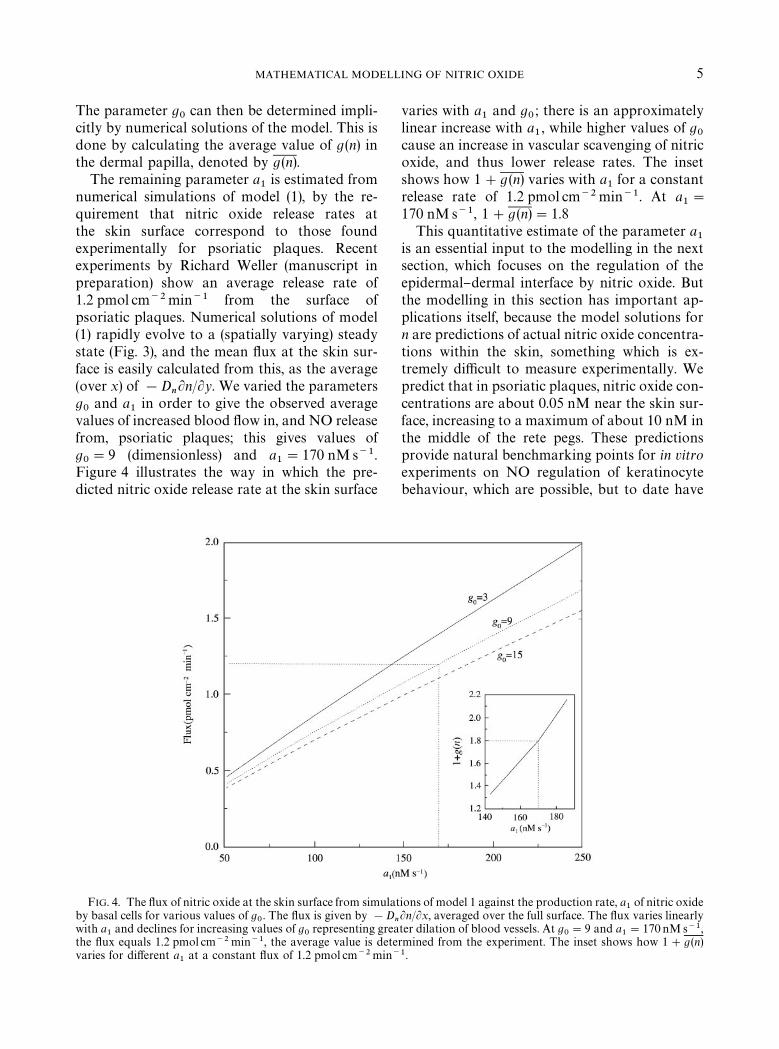
MATHEMATICAL MODELLING OF NITRIC OXIDE 5
The parameter g0
can then be determined impli-citly by numerical solutions of the model. This isdone by calculating the average value of g(n) inthe dermal papilla, denoted by g(n).
The remaining parameter a1
is estimated fromnumerical simulations of model (1), by the re-quirement that nitric oxide release rates atthe skin surface correspond to those foundexperimentally for psoriatic plaques. Recentexperiments by Richard Weller (manuscript inpreparation) show an average release rate of1.2 pmol cm~2min~1 from the surface ofpsoriatic plaques. Numerical solutions of model(1) rapidly evolve to a (spatially varying) steadystate (Fig. 3), and the mean #ux at the skin sur-face is easily calculated from this, as the average(over x) of !D
nLn/Ly. We varied the parameters
g0
and a1
in order to give the observed averagevalues of increased blood #ow in, and NO releasefrom, psoriatic plaques; this gives values ofg0"9 (dimensionless) and a
1"170 nMs~1.
Figure 4 illustrates the way in which the pre-dicted nitric oxide release rate at the skin surface
FIG. 4. The #ux of nitric oxide at the skin surface from simulaby basal cells for various values of g
0. The #ux is given by !D
with a1
and declines for increasing values of g0
representing grethe #ux equals 1.2 pmol cm~2 min~1, the average value is detvaries for di!erent a
1at a constant #ux of 1.2 pmol cm~2min
varies with a1
and g0; there is an approximately
linear increase with a1, while higher values of g
0cause an increase in vascular scavenging of nitricoxide, and thus lower release rates. The insetshows how 1#g (n) varies with a
1for a constant
release rate of 1.2 pmol cm~2min~1. At a1"
170 nMs~1, 1#g (n)"1.8This quantitative estimate of the parameter a
1is an essential input to the modelling in the nextsection, which focuses on the regulation of theepidermal}dermal interface by nitric oxide. Butthe modelling in this section has important ap-plications itself, because the model solutions forn are predictions of actual nitric oxide concentra-tions within the skin, something which is ex-tremely di$cult to measure experimentally. Wepredict that in psoriatic plaques, nitric oxide con-centrations are about 0.05 nM near the skin sur-face, increasing to a maximum of about 10 nM inthe middle of the rete pegs. These predictionsprovide natural benchmarking points for in vitroexperiments on NO regulation of keratinocytebehaviour, which are possible, but to date have
tions of model 1 against the production rate, a1of nitric oxide
nLn/Lx, averaged over the full surface. The #ux varies linearly
ater dilation of blood vessels. At g0"9 and a
1"170 nMs~1,
ermined from the experiment. The inset shows how 1#g(n)~1.

6 N. J. SAVILL E¹ A¸.
been expressed in terms of concentrations of NOdonors rather than nitric oxide itself (Krischelet al., 1998).
3. Regulation of Rete Pegs by Nitric Oxide
Rete pegs extend from 50 to 500 lm into thedermis, their lower tips reaching the horizontalvenous plexus (Bacharach-Buhles et al., 1994a).Each dermal papilla contains a single tortuouscapillary surrounded by ECM (Irwin et al., 1977).At some locations, particularly at the tip of thedermal papillae, the capillary directly abuts thebasal cell layer. The basal cells form an unbrokenlayer between the dermis and the other epidermalkeratinocytes. In vertical sections of biopsies, thedermal papillae appear to have widely varyingwidths and sometimes appear as isolated islandssurrounded by epidermis. This is an artifact of 2Dsectioning of 3D "nger-like structures, the islandsoccur because the axes of some papillae are ob-lique to the cut of the section. How these retepegs and dermal papillae are formed is stillunclear. Do the capillaries grow upwards intothe dermis or does the epidermis grow downand around the capillaries? These two mecha-nisms are not mutually exclusive.
Capillary growth or angiogenesis is known tooccur in psoriatic plaques; speci"cally, some en-dothelial cells in the vertical limbs of capillaryloops label positively with the nuclear prolifer-ation marker MAb MIB-1, while endothelial cellsin non-lesional skin do not (Creamer et al.,1997a). In addition, a plethora of angiogenic fac-tors have been discovered in plaques (Ettehadiet al., 1994; Elder et al., 1989; Nickolo! et al.,1994; Detmar et al., 1994; Creamer et al., 1997b;Bhushan et al., 1999). Even though angiogenesisdoes occur, there is no evidence that it leads tocapillary growth into the epidermis and there isno evidence of endothelial cell death in regressingplaques as one might expect (Braverman & Sib-ley, 1982). One possible e!ect of angiogenesismay be the observed capillary tortuosity (Bullet al., 1992). Therefore, the evidence for angiogen-esis is circumstantial at best.
Evidence in favour of epidermal growth intothe dermis is stronger. Lymph capillaries and thehorizontal subpapillary venous plexus are fre-quently observed to lie within some dermal papil-
lae (Bacharach-Buhles et al., 1994a, 1997). Thereis no evidence for or against lymphogenesis sothis observation is still inconclusive (Cli! et al.,1999). However, the second observation, if true,is a strong evidence for epidermal growth intothe dermis. Another unexplained observation isthe existence of ECM within dermal papillae: ifthe capillaries grow upwards into the dermis, howis the new ECM formed, or if the epidermis growsdownwards, why does some of the ECM remain?
In this paper, we adopt the hypothesis putforward by Bacharach-Buhles et al. (1994a) thatepidermis grows downwards and neglect any ef-fect of angiogenesis. A future paper will considerangiogenesis as the rete peg formation mecha-nism. A model of the formation of rete pegs needsto account for the following. (i) The loss of ECMbetween capillaries over several weeks thatcauses the sinking of the basal layer into thedermis. (ii) The cessation of ECM loss givinga layer of ECM roughly 10 lm thick between thebasal cells and the capillaries.
In order for the epidermis to grow between thecapillaries, the ECM must be degraded fasterthan it is reformed by "broblasts. Fibroblastsalso degrade ECM; however, it is unlikely thatthey cause increased degradation because thenormal turnover rate of ECM is on the order ofseveral years (Uitto, 1986) and plaques developwithin days. More likely candidates for theelevated degradation rates are the numerousneutrophils that in"ltrate psoriatic plaques.Neutrophils migrate from the capillaries throughthe dermis and into the epidermis degrading bothECM and basement membrane. In resolvingplaques, neutrophils disappear "rst, followed bythe loss of dermal papillae and tortuous capillar-ies, suggesting that the "broblasts reform theECM once the neutrophils have left the plaque.
Assuming neutrophils are the cause of ECMloss, why does some ECM remain in the dermalpapillae? If the epidermis moves downwards, thissuggests that the ECM density at the epider-mal}dermal junction is below a critical thresholdsuch that basal cells are able to move into thedegraded ECM. In a mature plaque, however, theECM density at the junction should be at a criti-cal threshold so that basal cells cannot movefurther into the ECM. Therefore, we need afactor (signalling molecule) that causes strong

MATHEMATICAL MODELLING OF NITRIC OXIDE 7
neutrophil-induced ECM-degradation when thebasal cells are far from the capillaries, butweakens degradation as the basal cells movecloser to the capillaries. In addition, there mustbe a threshold density of ECM below which basalcells can move into the ECM.
A strong candidate for this factor is nitric ox-ide. Because basal cells are a source of nitricoxide and capillaries are a sink, the closer thebasal cells are to the capillaries, the lower is thenitric oxide concentration within the dermis.Moreover, nitric oxide concentration determinesthe rate of neutrophil degradation of ECM: nitricoxide reacts rapidly with the free radical super-oxide (O~
2, a by-product of metabolism) to form
peroxynitrite (ONOO~). Peroxynitrite is astrong activator of neutrophil procollagenase(Okamoto et al., 1997; Maeda et al., 1998), theinactive form of collagenase secreted by neu-trophils (also called MMP-8). These reactionscan be represented as follows:
NO#O~2
k1&"ONOO~,
ONOO~#proMMP-8 k2&"MMP-8,
MMP-8#collagen k3&"MMP-8.
Peroxynitrite, like nitric oxide, is a small andhighly reactive free radical species, which there-fore has a high di!usion coe$cient and fast kinet-ics. MMP-8 is a 85 kDa molecule and is knownto degrade collagen only in the locality of thesecretory neutrophils due to its inhibition by tis-sue inhibitors of metalloproteinases (TIMPs).
The above reaction kinetics plus di!usion ofnitric oxide and peroxynitrite can be written asa system of di!erential equations:
Nitric oxide:
LnLt
"Dn+ 2n! k
1sn
hij
3%!#5*0/8*5) O~
2
# /(n)hij
130$6#5*0/!/$ 3%.07!-
(2)
Peroxynitrite:
LpLt
"Dp+ 2p# k
1sn
hij
3%!#5*0/8*5) O~
2
! d2p
hij
$%#!:
#
t(n, p,m@)hgigj
3%.07!-
(3)
Procollagenase:
dm@dt
"!k2pm@
hgigj!#5*7!5*0/
!d4m@
hij
$%#!:
# a2¸
hij
130$6#5*0/": /%65301)*-4
(4)
Collagenase:
dmdt
" k2pm@
hgigj!#5*7!5*0/
!d5m
hij
$%#!:
(5)
Collagen:
dcdt
" a3F
hij
4%#3%5*0/": &*"30"-!454
! k3mc
hij
"3%!,$08/": #0--!'%/!4%
! d6Fc
hij
"3%!,$08/": &*"30"-!454
(6)
where
/(n)"Ga1
in basal layer,
!d1n(1#g (n)) in capillaries,
0 otherwise,
and
t(n, p, m@)"G!k
2pm@ in ECM,
!d3p(1#g (n)) in capillaries,
0 otherwise.
The variables n, p, m@ and m are the molar concen-trations of nitric oxide, peroxynitrite, procol-lagenase and collagenase, respectively, and c isthe dry weight density of collagen. Nitric oxidedynamics are as in Section 2, except that theparameter j has been replaced by k
1s, where s is
the constant molar concentration of superoxide.As discussed above, the reaction of nitric oxidewith superoxide produces peroxynitrite, whichhas a constant background removal rate (d
2),
primarily by reaction with carbon dioxide, and isalso removed actively (modelled by the functiont). They key property of peroxynitrite is its ac-tivation of procollagenase (rate constant k
2),
which is produced at a constant rate a2
by neu-trophils (density ¸). Both pro and active forms ofcollagenase have intrinsic decay rates, denoted byd4
and d5, respectively; d
5incorporates the e!ects
of TIMPs. The "nal equation represents collagenproduction by "broblasts (density F) and removalboth by active collagenase and by "broblasts
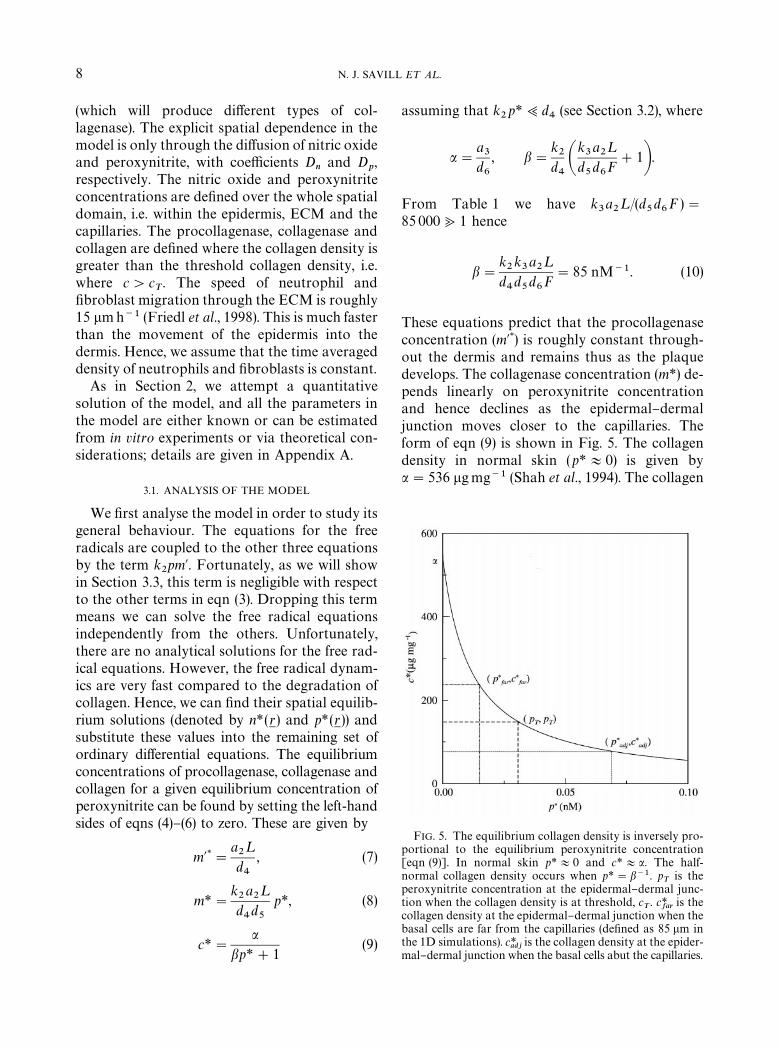
FIG. 5. The equilibrium collagen density is inversely pro-portional to the equilibrium peroxynitrite concentration[eqn (9)]. In normal skin p*+0 and c*+a. The half-normal collagen density occurs when p*"b~1. p
Tis the
peroxynitrite concentration at the epidermal}dermal junc-tion when the collagen density is at threshold, c
T. c*
faris the
collagen density at the epidermal}dermal junction when thebasal cells are far from the capillaries (de"ned as 85 lm inthe 1D simulations). c*
adjis the collagen density at the epider-
mal}dermal junction when the basal cells abut the capillaries.
8 N. J. SAVILL E¹ A¸.
(which will produce di!erent types of col-lagenase). The explicit spatial dependence in themodel is only through the di!usion of nitric oxideand peroxynitrite, with coe$cients D
nand D
p,
respectively. The nitric oxide and peroxynitriteconcentrations are de"ned over the whole spatialdomain, i.e. within the epidermis, ECM and thecapillaries. The procollagenase, collagenase andcollagen are de"ned where the collagen density isgreater than the threshold collagen density, i.e.where c'c
T. The speed of neutrophil and
"broblast migration through the ECM is roughly15 lmh~1 (Friedl et al., 1998). This is much fasterthan the movement of the epidermis into thedermis. Hence, we assume that the time averageddensity of neutrophils and "broblasts is constant.
As in Section 2, we attempt a quantitativesolution of the model, and all the parameters inthe model are either known or can be estimatedfrom in vitro experiments or via theoretical con-siderations; details are given in Appendix A.
3.1. ANALYSIS OF THE MODEL
We "rst analyse the model in order to study itsgeneral behaviour. The equations for the freeradicals are coupled to the other three equationsby the term k
2pm@. Fortunately, as we will show
in Section 3.3, this term is negligible with respectto the other terms in eqn (3). Dropping this termmeans we can solve the free radical equationsindependently from the others. Unfortunately,there are no analytical solutions for the free rad-ical equations. However, the free radical dynam-ics are very fast compared to the degradation ofcollagen. Hence, we can "nd their spatial equilib-rium solutions (denoted by n*(r
1) and p*(r
1)) and
substitute these values into the remaining set ofordinary di!erential equations. The equilibriumconcentrations of procollagenase, collagenase andcollagen for a given equilibrium concentration ofperoxynitrite can be found by setting the left-handsides of eqns (4)}(6) to zero. These are given by
m@*"a2¸
d4
, (7)
m*"k2a2¸
d4d5
p*, (8)
c*"a
bp*#1(9)
assuming that k2p*;d
4(see Section 3.2), where
a"a3
d6
, b"k2
d4Ak3a2¸
d5d6F#1B.
From Table 1 we have k3a2¸/(d
5d6F )"
85 000<1 hence
b"k2k3a2¸
d4d5d6F"85 nM~1. (10)
These equations predict that the procollagenaseconcentration (m@*) is roughly constant through-out the dermis and remains thus as the plaquedevelops. The collagenase concentration (m*) de-pends linearly on peroxynitrite concentrationand hence declines as the epidermal}dermaljunction moves closer to the capillaries. Theform of eqn (9) is shown in Fig. 5. The collagendensity in normal skin (p*+0) is given bya"536 lgmg~1 (Shah et al., 1994). The collagen

MATHEMATICAL MODELLING OF NITRIC OXIDE 9
density approaches zero as p*PR, and the half-normal density is given when p*"b~1"0.012 nM.
The key idea underlying our hypothesizedmechanism of rete peg formation is that collagendensity will be high near the capillary, but willdecrease rapidly away from the capillary, en-abling the epidermis to invade. We hypothesizethat this gradient in collagen occurs in responseto that in peroxynitrite, which will decrease sig-ni"cantly near a capillary, due to haemoglobinscavenging of nitric oxide and peroxynitrite.With this mechanism in mind, eqn (9) gives im-portant quantitative insights. For the model toreproduce rete pegs the peroxynitrite concentra-tion when the basal cells are far from the capillar-ies should be large enough to reduce the collagendensity at the epidermal}dermal junction to be-low the threshold collagen density (i.e. c*
far(c
Tand p*
far'p
T, see Fig. 5). As the junction moves
closer to the capillaries the peroxynitrite concen-tration decreases due to scavenging of peroxy-nitrite and nitric oxide by haemoglobin, andthe collagen density at the epidermal}dermaljunction rises. If the collagen density reaches thethreshold density before the basal cells reach thecapillaries, then rete peg formation stops and eachdermal papilla contains a single capillary sur-rounded by ECM (i.e. c*
adj'c
Tand p*
adj(p
T).
Two other cases are possible. (i) When c*far
'cT,
then no rete pegs will form and (ii) whenc*adj
(cT, then the basal cells will directly abut
the capillaries.From a therapeutic viewpoint the parameter
b is very interesting. A reduction in its valuemeans that more peroxynitrite is needed to givethe same reduction in collagen density. There-fore, any physiological change to any of the para-meters in eqn (10) that decrease b would havea negative e!ect on rete peg formation. Goodcandidates reduce the neutrophil density and in-crease the "broblast density, both of which canbe controlled. The other therapeutically access-ible variable is peroxynitrite concentration, vianitric oxide depletion by NO-inhibitors.
3.2. ONE-DIMENSIONAL SIMULATIONS
Detailed simulation of rete peg formation inthe model (2}6) requires a simulation in two
spatial dimensions; however, we have been un-able to solve the full model in 2D because ofcomputational constraints. Therefore, we beginby solving the full model in a one-dimensionalanalogue of rete peg formation, before solving(Section 3.3) a simpli"ed system of equations intwo space dimensions. Our one-dimensional for-mulation consists of a "nite domain of length100 lm, with a 5 lm region at the bottom bound-ary corresponding to a capillary. The basal cellsare modelled as a 12 lm wide movable region,initially placed at the top boundary. As the solu-tion evolves, we check the collagen density at theepidermal}dermal junction, and if this is belowthe critical threshold c
T, the basal cells are shifted
towards the capillary until the density at thejunction is at the threshold. Initially, all variablesare set to 0 except the collagen density which isset to a
3/d
6in the dermis, and there are zero #ux
end conditions on both boundaries.A typical 1D simulation over a period of 12
days is shown in Fig. 6. Time runs from left toright and space runs from top to bottom. Thenitric oxide and peroxynitrite concentrationsequilibrate within a few seconds due to their fastdi!usion and kinetics. The procollagenase andcollagenase equilibrate within several minutesand the collagen density equilibrates over severaldays. Figure 7 shows how the spatial distributionof the variables changes as the basal layer movescloser to the capillary. Each curve is a snapshot intime taken every 105 s. The arrows at the top ofeach graph show the position of the epider-mal}dermal junction at each snapshot. At thecapillary (x"95 lm), all variables except col-lagen change very little as the basal cells movecloser. The peroxynitrite concentration is alwaysappreciable leading to some degradation of col-lagen at the capillary.
The story is di!erent at the epidermal}dermaljunction. As the basal layer moves closer, bothnitric oxide and peroxynitrite concentrations falldue to their scavenging by haemoglobin. Theydecrease almost linearly with distance due totheir high di!usion coe$cients. The procol-lagenase concentration is high (at lM levels) dueto a large production by neutrophils and slowactivation by peroxynitrite [eqn (7)]. Its concen-tration changes very little because its activation isslower than its degradation (k
2p*;d
4). The
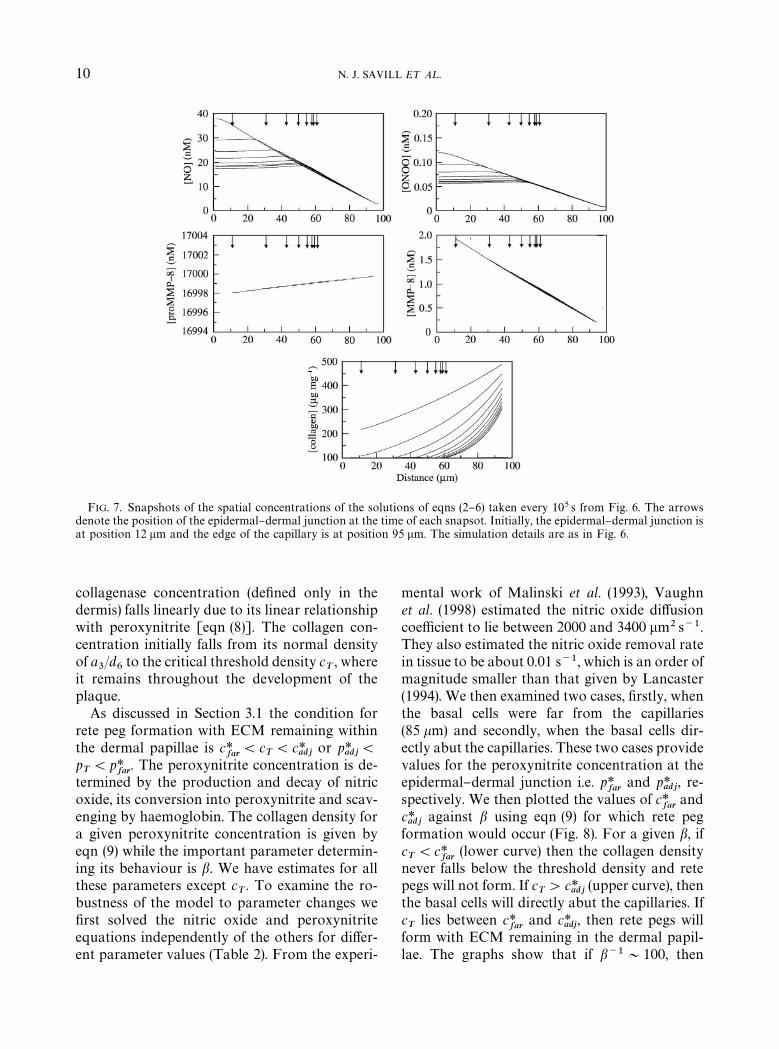
FIG. 7. Snapshots of the spatial concentrations of the solutions of eqns (2}6) taken every 105 s from Fig. 6. The arrowsdenote the position of the epidermal}dermal junction at the time of each snapsot. Initially, the epidermal}dermal junction isat position 12 lm and the edge of the capillary is at position 95 lm. The simulation details are as in Fig. 6.
10 N. J. SAVILL E¹ A¸.
collagenase concentration (de"ned only in thedermis) falls linearly due to its linear relationshipwith peroxynitrite [eqn (8)]. The collagen con-centration initially falls from its normal densityof a
3/d
6to the critical threshold density c
T, where
it remains throughout the development of theplaque.
As discussed in Section 3.1 the condition forrete peg formation with ECM remaining withinthe dermal papillae is c*
far(c
T(c*
adjor p*
adj(
pT(p*
far. The peroxynitrite concentration is de-
termined by the production and decay of nitricoxide, its conversion into peroxynitrite and scav-enging by haemoglobin. The collagen density fora given peroxynitrite concentration is given byeqn (9) while the important parameter determin-ing its behaviour is b. We have estimates for allthese parameters except c
T. To examine the ro-
bustness of the model to parameter changes we"rst solved the nitric oxide and peroxynitriteequations independently of the others for di!er-ent parameter values (Table 2). From the experi-
mental work of Malinski et al. (1993), Vaughnet al. (1998) estimated the nitric oxide di!usioncoe$cient to lie between 2000 and 3400 lm2 s~1.They also estimated the nitric oxide removal ratein tissue to be about 0.01 s~1, which is an order ofmagnitude smaller than that given by Lancaster(1994). We then examined two cases, "rstly, whenthe basal cells were far from the capillaries(85 km) and secondly, when the basal cells dir-ectly abut the capillaries. These two cases providevalues for the peroxynitrite concentration at theepidermal}dermal junction i.e. p*
farand p*
adj, re-
spectively. We then plotted the values of c*far
andc*adj
against b using eqn (9) for which rete pegformation would occur (Fig. 8). For a given b, ifcT(c*
far(lower curve) then the collagen density
never falls below the threshold density and retepegs will not form. If c
T'c*
adj(upper curve), then
the basal cells will directly abut the capillaries. IfcT
lies between c*far
and c*adj
, then rete pegs willform with ECM remaining in the dermal papil-lae. The graphs show that if b~1&100, then
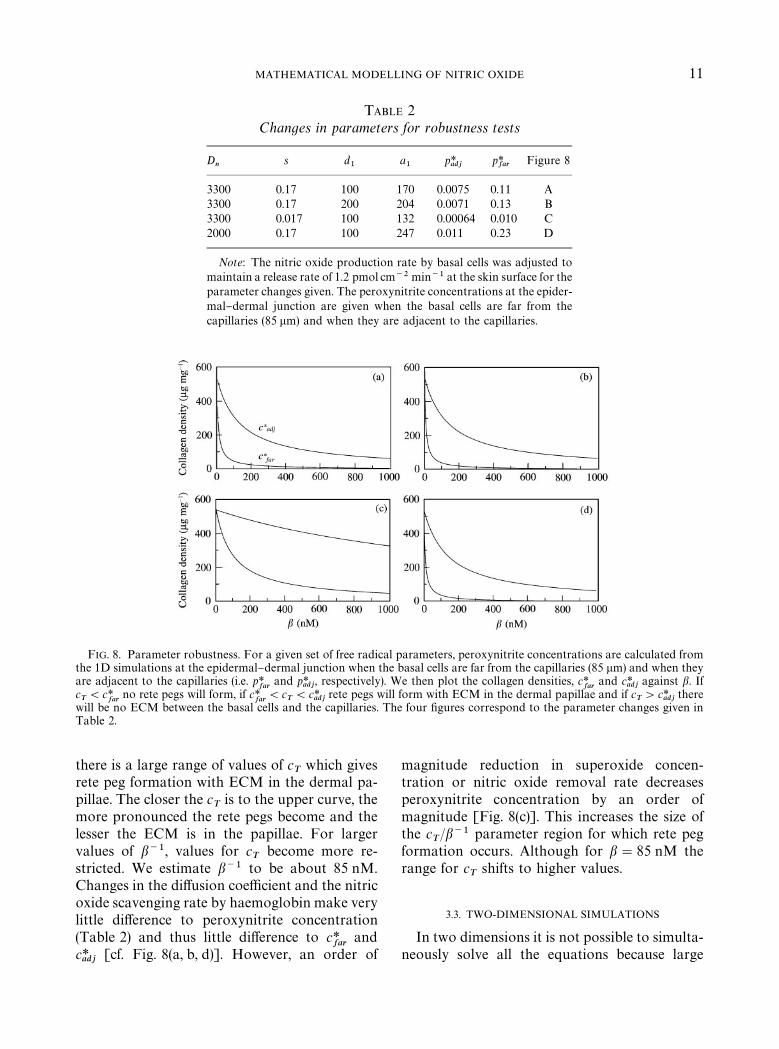
TABLE 2Changes in parameters for robustness tests
Dn
s d1
a1
p*adj
p*far
Figure 8
3300 0.17 100 170 0.0075 0.11 A3300 0.17 200 204 0.0071 0.13 B3300 0.017 100 132 0.00064 0.010 C2000 0.17 100 247 0.011 0.23 D
Note: The nitric oxide production rate by basal cells was adjusted tomaintain a release rate of 1.2 pmol cm~2 min~1 at the skin surface for theparameter changes given. The peroxynitrite concentrations at the epider-mal}dermal junction are given when the basal cells are far from thecapillaries (85 lm) and when they are adjacent to the capillaries.
FIG. 8. Parameter robustness. For a given set of free radical parameters, peroxynitrite concentrations are calculated fromthe 1D simulations at the epidermal}dermal junction when the basal cells are far from the capillaries (85 lm) and when theyare adjacent to the capillaries (i.e. p*
farand p*
adj, respectively). We then plot the collagen densities, c*
farand c*
adjagainst b. If
cT(c*
farno rete pegs will form, if c*
far(c
T(c*
adjrete pegs will form with ECM in the dermal papillae and if c
T'c*
adjthere
will be no ECM between the basal cells and the capillaries. The four "gures correspond to the parameter changes given inTable 2.
MATHEMATICAL MODELLING OF NITRIC OXIDE 11
there is a large range of values of cT
which givesrete peg formation with ECM in the dermal pa-pillae. The closer the c
Tis to the upper curve, the
more pronounced the rete pegs become and thelesser the ECM is in the papillae. For largervalues of b~1, values for c
Tbecome more re-
stricted. We estimate b~1 to be about 85 nM.Changes in the di!usion coe$cient and the nitricoxide scavenging rate by haemoglobin make verylittle di!erence to peroxynitrite concentration(Table 2) and thus little di!erence to c*
farand
c*adj
[cf. Fig. 8(a, b, d)]. However, an order of
magnitude reduction in superoxide concen-tration or nitric oxide removal rate decreasesperoxynitrite concentration by an order ofmagnitude [Fig. 8(c)]. This increases the size ofthe c
T/b~1 parameter region for which rete peg
formation occurs. Although for b"85 nM therange for c
Tshifts to higher values.
3.3. TWO-DIMENSIONAL SIMULATIONS
In two dimensions it is not possible to simulta-neously solve all the equations because large

12 N. J. SAVILL E¹ A¸.
di!erences in time-scales between nitric oxideand collagen dynamics cause prohibitively longsimulation times. However, it is possible to de-couple the equations and hence simulate themindependently as discussed in Section 3.2. The1D simulations show that the term k
2pm@ is of the
order 10~2, whereas the term k1sn is of the order
1 and the terms d2p and d
3p (1#g(n)) are of the
order 10 (units of nMs~1). Therefore, the termk2pm@ can reasonably be neglected, and the equa-
tions for nitric oxide and peroxynitrite then be-come independent of the other three. Moreover,the equations for procollagenase and collagenaseare independent of the equation for collagen.Hence, we can simulate the fast equilibration ofnitric oxide and peroxynitrite, then analyticallycalculate the equilibrium concentrations ofprocollagenase and collagenase by setting theLHS of their equations to 0. Finally, we can thensimulate the degradation of collagen over time,moving the epidermal}dermal junction so thatthere is the collagen density at the critical thre-shold value. With the new con"guration of basalcells, the nitric oxide and peroxynitrite equilib-rium concentrations can be recalculated and theprocesses repeated until no further movement ofthe basal cells occurs.
Figure 9 shows one such simulation. The basalcells are modelled as a 12 lm wide movable re-gion, initially placed 100 lm below the skin sur-face. The capillaries are 10 lm wide, and we havearbitrarily chosen their con"guration to demon-strate the robustness of the pattern formingmechanism. The boundary conditions are as inSection 2. Figure 9(a) shows the initial con"-guration of capillaries and basal layer with equi-librium concentrations of nitric oxide andperoxynitrite. The density of collagen at the epi-dermal}dermal junction has just dropped belowthe critical threshold and a new con"guration ofthe basal layer is just about to be calculated.Figure 9(b) shows the "nal con"guration of basalcells, where no more movement is possible. This"nal pattern closely resembles the con"gurationsseen in psoriatic plaques [see Fig. 1(b)].
4. Conclusion
In this paper, we have presented two di!erentmodels related to nitric oxide activity in psoriasis.
The "rst model assumes a "xed skin geometry,and predicts nitric oxide density within, and re-lease from, the skin, as a result of production inthe basal epidermis. Using data on nitric oxiderelease rates from psoriatic plaques, this modelpredicts an NO production rate by basal epider-mal cells of about 170 nM s~1. Within the tissue,NO concentrations reach about 10 nM and atthe skin surface about 0.05 nM. We found noexperimental reports on NO production ratesfrom basal epidermal cells nor any data onNO concentrations within psoriatic plaques.Vaughn et al. (1998) have estimated from thework of Malinski et al. (1993) an NO productionrate by endothelial cells about an order of a mag-nitude larger than our estimate for basal epider-mal cells.
Building on this, the second model considersthe regulation, by nitric oxide, of the interfacebetween the epidermis and dermis. This modelpredicts that the NO production in the basalepidermis can cause the formation of the pro-nounced rete pegs seen in psoriatic plaques. Wehypothesize that the basic mechanism for this isthe rapid conversion of NO to peroxynitrite,which in turn regulates the activation of procol-lagenase secreted by neutrophils. As the collagendegrades the basal cells and the epidermis movecloser to the capillaries. At a critical collagendensity, basal cells can no longer move into theECM thus halting their movement and creatingthe rete pegs and dermal papillae observed in thepsoriatic plaques. From the estimated parametervalues from the literature we have shown thatthis is a plausible model that is robust to changesin parameter values. A critical parameter inthe model is b which relates collagen density inthe ECM to the peroxynitrite concentration.Because b depends on many of the model's para-meters we can quantitatively predict howchanges in these parameters a!ect rete pegformation.
From in vitro experimental work on collagendegradation (Okamoto et al., 1997) we predicta procollagenase activation rate by peroxynitriteof 10~5 nM~1 s~1 and a collagen degradationrate by collagen of 5]10~6nM~1 s~1. The depthof the rete pegs depends on how a collagen den-sity at the epidermal}dermal junction varies withperoxynitrite concentration (Fig. 5) and on the

MATHEMATICAL MODELLING OF NITRIC OXIDE 13
collagen density threshold below which theepidermis can invade the dermis.
We have focussed on the role of nitric oxide incontrolling the formation of highly pronouncedrete pegs in psoriasis. However, nitric oxide isalso a potential regulator of a number of otheraspects of cell behaviour, which may play a rolein psoriasis, in particular the growth and di!eren-tiation of keratinocytes (Krischel et al., 1998) andthe endothelial cell growth and migration (Zicheet al., 1994). Detailed studies of the e!ects of theseactivities of nitric oxide in psoriasis are essentialbefore a full assessment can be made of the over-all role of nitric oxide in the onset of the disease.This is a natural area for future mathematicalmodelling.
From a therapeutic viewpoint, the suggestionthat nitric oxide may play a causal role inpsoriasis is an exciting one, because nitric oxidelevels can be altered by topical application ofcreams containing either NO-donors or NO-inhibitors. In fact, a study has recently beenpublished in which an NO-inhibitor was appliedtopically in psoriasis: NO release was reduced,but with no e!ect on clinical outcome (Ormerodet al., 2000). However, because the e!ects of nitricoxide are so concentration-dependent, it is quitepossible that di!erent treatment regimes may besuccessful. Our research forms a part of a grow-ing body of work providing, for the "rst time,clues to the underlying molecular causes ofpsoriasis, and this has the potential to lead toa new generation of anti-psoriasis therapies.
NJS and JAS are supported in part by SHEFCResearch and Development grant 107 &&Centre forTheoretical Modelling in Medicine''. JAS is sup-pported in part by an EPSRC Advanced ResearchFellowship.
REFERENCES
AUER, T., BACHARACH-BUHLES, M., EL GAMMAL, S.,STUG CKER, M., PANZ, B., POPP, C., HOFFMANN, K.,HAPPE, M. & ALTMEYER, P. (1994). The hyperperfusion ofthe psoriatic plaques correlates histologically with dilationof vessels. Acta Derm. <enereol. S186, 30}32.
BACHARACH-BUHLES, M., EL GAMMAL, S., PANZ, B. &ALTMEYER, P. (1994a). The pseudo-elongation of capillar-ies in psoriatic plaques. Acta. Derm. <enereol. (Stockh) 186,133}137.
BACHARACH-BUHLES, M., EL GAMMAL, S., PANZ, B. &ALTMEYER, P. (1994b). The topography of epithelium andvascular plexuses in psoriasis. In: Electron Microscopy in
Dermatology*Basic and Clinical Research (Ishibashi, Y.Nakagawa, H. & Suzuki, H. eds), pp. 345}351. Amster-dam: Elsevier Science.
BACHARACH-BUHLES, M., EL GAMMAL, S., PANZ, B. &ALTMEYER, P. (1997). In psoriasis the epidermis, includingthe subepidermal vascular plexus, grows downwards intothe dermis. Br. J. Dermatol. 136, 97}101.
BHUSHAN, M., MCLAUGHLIN, B., WEISS, J. B. & GRIFFITHS,C. E. M. (1999). Levels of endothelial cell stimulatingangiogenesis factor and vascular endothelial growthfactor are elevated in psoriasis. Br. J. Dermatol. 141,1054}1060.
BRAVERMAN, I. M. & SIBLEY, J. (1982). Role of microcircula-tion in the treatment and pathogenesis of psoriasis.J. Invest. Dermatol. 78, 12}17.
BULL, R. H., BATES, D. O. & MORTIMER, P. S. (1992).Intravital video-capillaroscopy for the study of the micro-circulation in psoriasis. Br. J. Dermatol. 126, 436}445.
BUTLER, A. R., MEGSON, I. L. & WRIGHT, P. (1998). Di!u-sion of nitric oxide and scavenging by blood in the vascula-ture. BBA-Gen. Subjects 1425, 168}176.
CLIFF, S., BEDLOW, A. J., STANTON, A. W. B. & MORTIMER,P. S. (1999). An in vivo study of the microlymphatics inpsoriasis using #uorescence microlymphography. Br.J. Dermatol. 140, 61}66.
COBBOLD, C. A. & SHERRATT, J. A. (2000). Mathematicalmodelling of nitric oxide activity in wound healing canexplain keloid and hypertrophic scarring. J. theor. Biol.204, 257}288.
CREAMER, D., ALLEN, M. H., SOUSA, A., POSTON, R.& BARKER, J. N. W. N. (1997a). Localization of endothelialproliferation and microvascular expansion in active plaquepsoriasis. Br. J. Dermatol. 136, 859}865.
CREAMER, D., JAGGER, R., ALLEN, M., BICKNELL, R.& BARKER, J. (1997b). Overexpression of the angiogenicfactor platelet-derived endothelial cell growth factor/thymidine phosphorylase in psoriatic plaques. Br. J.Dermatol. 137, 851}855.
CULOTTA, E. & KOSHLAND JR, D. E. (1992). NO news isgood news. Science 258, 1862}1865.
DENICOLA, A., SOUZA, J. M. & RADI, R. (1998). Di!usion ofperoxynitrite across erythrocyte membranes. Proc. NatlAcad. Sci. ;.S.A. 95, 3566}3571.
DETMAR, M., BROWN, L. F., CLAFFEY, K. P., YEE, K. T.,KOCHER, O., JACKMAN, R. W., BERSE, B. & DVORAK, H. F.(1994). Overexpression of vascular permeability factor/vascular endothelial growth factor and its receptors inpsoriasis. J. Exp. Med. 180, 1141}1146.
ELDER, J. T., FISHER, G. J., LINDQUIST, P. B., BENNETT, G. L.,PITTELKOW, M. R., COFFEY, R. J., ELLINGSWORTH, L.,DERYNCK, R. & VOORHEES, J. J. (1989). Overexpression oftransforming growth factor alpha in psoriatic epidermis.Science 243, 811}814.
ETTEHADI, P., GREAVES, M. W., WALLACH, D., ADERKA, D.& CAMP, R. D. R. (1994). Elevated tumour necrosis factor-alpha (TNF-a) biological activity in psoriatic skin lesions.Clin. Exp. Immunol. 96, 146}151.
FRIEDL, P., ZAG NKER, K. S. & BROCKER, E. B. (1998). Cellmigration strategies in 3D extracellular matrix: di!erencesin morphology, cell matrix interactions, and integrin func-tion. Microsc. Res. ¹ech. 43, 369}378.
GOW, A. J. & STAMLER, J. S. (1998). Reactions between nitricoxide and haemoglobin under physiological conditions.Nature 391, 169}173.

14 N. J. SAVILL E¹ A¸.
HAKIM, T. S., SUGIMORI, K., CAMPORESI, E. M. &ANDERSON, G. (1996). Half-life of nitric oxide in aqueoussolutions with and without haemoglobin. Physiol. Meas.17, 267}277.
HIBBS, J. B., TAINTOR, R. R., VAVRIN, Z. & RACHLIN, E. M.(1989). Nitric oxide: a cytostatic activated macrophagee!ector molecule. Biochem. Biophys. Res. Commun. 157,87}94.
IGNARRO, L. J., BYRNS, R. E., BUGA, G. M. & WOOD, K. S.(1987). Endothelium-derived relaxing factor from pulmon-ary artery and vein possesses the pharmacologic andchemical properties identical to those of nitric oxide rad-ical. Circ. Res. 61, 866}879.
IRWIN, M., BRAVERMAN, M. D. & AGNES YEN, B. S. (1977).Ultrastructure of the capillary loops in the dermal papillaeof psoriasis. J. Invest. Dermatol. 68, 53}60.
KISSNER, R., NAUSER, T., BUGNON, P., LYE, P. G. &KOPPENOL, W. H. (1997). Formation and properties ofperoxynitrite as studied by laser #ash photolysis, high-pressure stopped-#ow technique, and pulse radiolysis.Chem. Res. ¹oxicol. 10, 1285}1292.
KLEMP, P. & STABERG, B. (1983). Cutaneous blood #ow inpsoriasis. J. Invest. Dermatol. 81, 503}506.
KOLB-BACHOFEN, V., FEHSEL, K., MICHEL, G. & RUZICKA, T.(1994). Epidermal keratinocyte expression of inducible ni-tric oxide synthase in skin lesions of psoriasis vulgaris.¸ancet 344, 139.
KRISCHEL, V., BRUCH-GERHARZ, D., SUSCHEK, C.,KRONCKE, K. D., RUZICKA, T. & KOLB-BACHOFEN, V.(1998). Biphasic e!ect of exogenous nitric oxide on prolif-eration and di!erentiation in skin derived keratinocytesbut not "broblasts. J. Inves. Dermatol. 111, 286}291.
KROGSTAD, A. L., SWANBECK, G. & WALLIN, B. G. (1995).Axon-re#ex-mediated vasodilation in the psoriatic plaque?J. Invest. Dermatol. 104, 872}876.
KU, D. D. (1996). Nitric oxide- and nitric oxide-donorinduced relaxation. Meth. Enzymol. 269, 107}119.
LANCASTER, J. R. (1994). Simulation of the di!usion andreaction of endogenously produced nitric oxide. Proc. NatlAcad. Sci. ;.S.A. 91, 8137}8141.
LANCASTER, J. R. (1997). A tutorial on the di!usibility andreactivity of free nitric oxide. Nitric Oxide: Biology andChemistry 1, 18}39.
LAURENT, M., LEPOIVRE, M. & TENU, J. P. (1996). Kineticmodelling of the nitric oxide gradient generated in vitro byadherent cells expressing inducible nitric oxide synthase.Biochem. J. 314, 109}113.
LIAO, J. C., HEIN, T. W., VAUGHN, M. W., HUANG, K. T.& KOU, L. (1999). Intravascular #ow decreases erythrocyteconsumption of nitric oxide. Proc. Natl Acad. Sci. ;.S.A.96, 8757}8761.
LIU, X., MILLER, M. J. S., JOSHI, M. S., SADOWSKA-KROWICKA, H., CLARK, D. A. & LANCASTER JR, J. R.(1998). Di!usion-limited reaction of free nitric oxide witherythrocytes. J. Biol. Chem. 273, 18 709}18 713.
LYMAR, S. V. & HURST, J. K. (1995). Rapid reaction betweenperoxonitrite ion and carbon-dioxide*implications forbiological-activity. J. Am. Chem. Soc. 117, 8867}8868.
MAEDA, H., OKAMOTO, T. & AKAIKE, T. (1998). Humanmatrix metalloprotease activation by insults of bacterialinfection involving proteases and free radicals. Biol. Chem.379, 193}200.
MALINSKI, T., TAHA, Z., GRUNFELD, S., PATTON, S.,KAPTURCZAK, M. & TOMBOULIAN, P. (1993). Di!usion of
nitric oxide in the aorta wall monitored in situ by por-phyrinic microsensors. Biochem. Biophys. Res. Commun.193, 1076}1082.
MONCADA, S. (1999). Nitric oxide: discovery and impact onclinical medicine. J. Roy. Soc. Med. 92, 164}169.
NICKOLOFF, B. J., MITRA, R. S., VARANI, J., DIXIT, V. M.& POLVERINI, P. J. (1994). Aberrant production of inter-leukin-8 and thrombospondin-1 by psoriatic keratinocytesmediates angiogenesis. Am. J. Pathol. 144, 820}828.
OKAMOTO, T., AKAIKE, T., NAGANO, T., MIYAJIMA, S.,SUGA, M., ANDO, M., ICHIMORI, K. & MAEDA, H. (1997).Activation of human neutrophil procollagenase by nitro-gen dioxide and peroxynitrite: a novel mechanism forprocollagenase activation involving nitric oxide. Arch.Biochem. Biophys. 342, 261}274.
OREM, A., ALIYAZICIOGLU, R., KIRAN, E., VANIZOR, B.,CIMNOCODEIT, G. & DEGER, O. (1997). The relationshipbetween nitric oxide production and activity of thedisease in patients with psoriasis. Arch. Dermatol. 133,1606}1607.
ORMEROD, A. D., COPELAND, P. & SHAH, S. A. A. (2000).Treatment of psoriasis with topical N-G-monomethyl-L-arginine, an inhibitor of nitric oxide synthesis. Br.J. Dermatol. 142, 985}990.
ORMEROD, A. D., WELLER, R., COPELAND, P., BENJAMIN, N.,RALSTON, S. H., GRABOWSKI, P. & HERRIOT, R. (1998).Detection of nitric oxide and nitric oxide synthases inpsoriasis. Arch. Dermatol. Res. 290, 3}8.
SCHMIDT, K., DESCH, W., KLATT, P., KUKOVETZ, W. R.& MAYER, B. (1997). Release of nitric oxide from donorswith known half-life: a mathematical model for calculatingnitric oxide concentrations in aerobic solutions. Naunyn-Schmiedebergs Arch. Pharmacol. 355, 457}462.
SHAH, M., FOREMAN, D. M. & FERGUSON, M. W. J. (1994).Neutralising antibody to TGF-b
1,2reduces cutaneous
scarring in adult rodents. J. Cell Sci. 107, 1137}1157.THIBODEAU, G. A. & PATTON, K. T. (1997). ¹he HumanBody in Health and Disease. St. Louis: Mosby YearBook, Inc.
UITTO, J. (1986). Interstitial collagens. In: Biologyof the Integument 2: <ertebrates (Bereiter-Hahn, J.Matoltsy, A. G. & Richards, K. S. eds). pp. 800}809. Berlin:Springer-Verlag.
VAUGHN, M. W., KUO, L. & LIAO, J. C. (1998). Estimationof nitric oxide production and reaction rates in tissueby use of a mathematical model. Am. J. Physiol. 43,H2163}H2176.
WELLER, R. & ORMEROD, A. (1997). Increased expression ofnitric oxide synthase. Br. J. Dermatol. 136, 132}148.
WELLER, R., ORMEROD, A. & BENJAMIN, N. (1996a). Nitricoxide generation measured directly from psoriatic plaquesby chemiluminescence. Br. J. Dermatol. 134, 569.
WELLER, R., PATTULLO, S., SMITH, L., GOLDEN, M.,ORMEROD, A. & BENJAMIN, N. (1996b). Nitric oxide isgenerated on the skin surface by reduction of sweat nitrate.J. Invest. Dermatol. 107, 327}331.
WHEATER, P. R., BURKITT, H. G., STEVENS, A. & LOWE, J. S.(1985). Basic Histopathology:A Colour Atlas and ¹ext.London: Churchill Livingstone.
ZICHE, M., MORBIDELLI, L., MASINI, E., AMERINI, S.,GRANGE, R. H. J., MAGGI, C. A., GEPPETTI, P. & LEDDA, F.(1994). Nitric-oxide mediates angiogenesis in-vivo and en-dothelial-cell growth and migration in-vitro promoted bysubstance-P. J. Clin. Invest. 94, 2036}2044.

MATHEMATICAL MODELLING OF NITRIC OXIDE 15
APPENDIX A
Dn, D
p: The di!usion coe$cient for nitric oxide
in intact tissue is roughly 3300 lm2 s~1 (Malinskiet al., 1993). Vaughn et al. (1998) give estimatesfor the di!usion coe$cient between 2000 and3400 lm2 s~1 from the data of Malinski et al.(1993). There is no experimental value for thedi!usion coe$cient of peroxynitrite. However, ithas a similar permeability across phospholipidmembranes as nitric oxide (Denicola et al., 1998).Hence, its di!usion coe$cient will be roughlysimilar to that of nitric oxide with some compen-sation for its larger mass. This compensationfactor is (15/31)1@3, giving a di!usion coe$cient ofroughly 2600 lm2 s~1.
j, k1, s: The rate constant for the reaction of
nitric oxide with superoxide is 19 nM~1 s~1(Kissner et al., 1997). The background removalrate of nitric oxide in tissue is roughly 0.17 s~1(Lancaster, 1994). Vaughn et al. (1998) give esti-mates for the removal rate of between 0.006 and0.01 s~1 from the data of Malinski et al. (1993).We do not have any estimate for the superoxideconcentration in tissue. The reaction betweenNO and superoxide is very fast compared withother NO reactions. Thus, if we assume that this isthe predominant reaction, the superoxide concen-tration should be &0.01 nM for a removal rate of0.17 s~1. Smaller superoxide concentrations willgive correspondingly smaller peroxynitrite con-centrations. Note that in model 1, j"k
1s.
d1: Liu et al. (1998) found a decay rate of
385 s~1 for nitric oxide in blood. Independently,Hakim et al. (1996) found a maximum rateconstant for nitric oxide and red blood cellhaemoglobin of 2]105 M~1 s~1. Taking a hae-moglobin concentration in blood as 2 mM givesa maximum decay rate of 400 s~1. However, Liaoet al. (1999) showed that the rate constant in#owing blood is 5]104 M~1 s~1, which is thevalue we use here, hence d
1"100 s~1.
d2: In tissue, the dominant reaction of per-
oxynitrite is with carbon dioxide. This occurs ata rate of 5.8]104 M~1 s~1 (Lymar & Hurst,1995). The concentration of carbon dioxide isapproximately 1 mM giving an approximatedecay rate of peroxynitrite in tissue of 58 s~1.
d3: Denicola et al. (1998) measured the reac-
tion rate for peroxynitrite and haemoglobin as
1.04]104 M~1 s~1 at 253C. This gives a decayrate of 20 s~1; we take the value at body temper-ature to be d
3"30 s~1.
d4: There is no experimental data for the re-
moval rate of procollagenase, although we couldexpect it to be roughly on the order of a fewminutes. We chose its value to be 0.01 s~1.
d5, k
2, k
3: Okamoto et al. (1997) performed in
vitro studies on the peroxynitrite activation ofneutrophil procollagenase. From their experi-mental set-up we constructed the followingmodel:
pR "!k2pm@!dp#y(t), (A.1)
mR @"!k2pm@, (A.2)
mR "k2pm@!d
5m, (A.3)
cR"!k3mc, (A.4)
where
y(t)"3ad (t!nq) for n"0,2,R, (A.5)
where d (t) is the delta function. The initial condi-tions are p(0)"0, m@ (0)"0.6 lM, m (0)"0 andc(0)"1.8 lM and the known parameter valuesare d"0.7 s~1, a"0.4}1200 lM and q"300 s.By solving the model and "tting the solution totheir data we estimated the following parametervalues: k
2"10~5 nM~1 s~1 and k
3/d
5"5]
10~4 nM~1. Assuming an inhibition rate of col-lagenase by TIMPs on the order of a few minutes,we set d
5"0.01 s~1 and hence k
3"5]
10~6 nM~1 s~1.a3, d
6, F: The cell density of "broblasts is
about 10~8 cells lm~3 (Cobbold & Sherratt,2000). The collagen density in normal skin is536 lgmg~1 dry weight (Shah et al., 1994). Fromour model this means that a
3/d
6"536. The time
taken for rete pegs to return to normal frompsoriatic skin is of the order of weeks. This im-plies a regrowth time for collagen of about 106 s.From eqn (6) the regrowth time is of the order536/(a
3F). Hence, we take a
3"5.36]104
lg mg~1 lm3 s~1 cell~1 and d6"100 s~1.
a2¸: To estimate the term a
2¸ we note that
the rate of degradation of collagen is given by the

16 N. J. SAVILL E¹ A¸.
factor k3m in eqn (6). The time-scale for this is on
the order of a few days, therefore k3m&
10~5 s~1. At equilibrium, m*"k2
a2¸p*/
(d4d5) by assuming that k
2p*;d
4. The value of
p* is taken from the epidermal}dermal junction
when the basal cells are far from the capillaries.From the 1D simulations this value is 0.12before the basal cells have moved towardsthe capillary. This gives a value of roughly170 nMs~1 for a
2¸.

FIG. 1. (a) Normal skin showing the lightly stained de-rmis, the medium stained epidermis and the darkly staineddead outer layer of cells. (b) Skin from a psoriatic plaquewith elongated narrow rete pegs (R). Between the rete pegs,the epidermis is thinned over oedematous and prominentdermal papillae (DP) in which dilated capillaries are prom-inent (c). The epidermis contains small aggregations of neu-trophils forming microabscesses (M). There is a variablechronic in#ammatory in"ltrate in the upper dermis (D).Kindly reproduced from Wheater et al. (1985).
FIG. 3. Example of the steady-state solution of nitricoxide (model 1). The skin surface is at the top. The solutionis calculated using the alternating-direction-implicit methodwith dt"2]10~3 s and dx"dy"1lm. Equilibrium isreached within a few seconds. The colour-bar gives the nitricoxide concentration in nM.
doi:10.1006/jtbi.2001.2400N. J. SAVILL E¹ A¸.
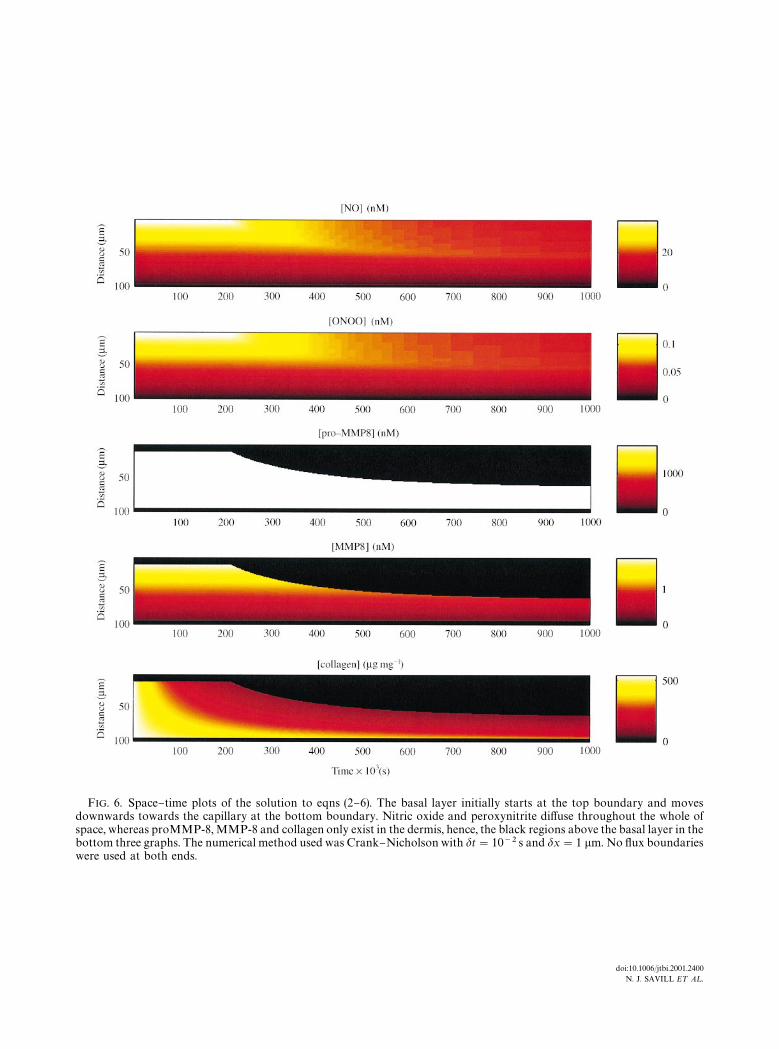
FIG. 6. Space}time plots of the solution to eqns (2}6). The basal layer initially starts at the top boundary and movesdownwards towards the capillary at the bottom boundary. Nitric oxide and peroxynitrite di!use throughout the whole ofspace, whereas proMMP-8, MMP-8 and collagen only exist in the dermis, hence, the black regions above the basal layer in thebottom three graphs. The numerical method used was Crank}Nicholson with dt"10~2 s and dx"1 lm. No #ux boundarieswere used at both ends.
doi:10.1006/jtbi.2001.2400N. J. SAVILL E¹ A¸.

FIG. 9. The two-dimensional solution of eqns (2}6). The position of the basal layer is represented as the white band and thecapillaries are in black in the bottom two plots. (a) The initial con"guration of basal layer and capillaries with a newcon"guration of the basal layer is about to be computed because collagen density has fallen below the critical threshold. (b)The "nal con"guration with no more movement of the basal layer is possible. The numerical method for the solution of nitricoxide and peroxynitrite is ADI with dt"5]10~3 s and dx"dy"1 lm. Once these solutions have equilibrated theproMMP!8 and MMP!8, spatial concentrations are calculated from eqns (7) and (8). The collagen density is then solvedover time using a "rst-order Runge}Kutta algorithm with dt"500 s and a spatial resolution of 1 lm2. Movies of thissimulation can be downloaded from http://www.ma.hw.ac.uk/&njs/psoriasis.html.
doi:10.1006/jtbi.2001.2400N. J. SAVILL E¹ A¸.