mechanism of plasma iron incorporation into hepatic ferritin*
TRANSCRIPT

THE JOURNAL OF BIOLOGICAL CHEMISTRY Vol. 235, No. 3, March 1960
Printed in U.S.A.
Mechanism of Plasma Iron Incorporation into Hepatic Ferritin*
ABRAHAM MAZUR, SAUL GREEN, AND A. CARLETON
From the Department of Medicine, Cornell University Medical College and The New York Hospital, New York, New York
(Received for publication, September 22, 1959)
Except for recent studies concerned with the release of iron from hepatic ferritin (1, 2), little experimental evidence is avail- able bearing on the identity of the biochemical mechanisms in- volved in the transport of iron in the animal organism. Nishida and Labbe (3, 4) have demonstrated the presence of an enzyme in liver which is required for the incorporation of iron into heme, and Jandl et al. (5) have shown a need for oxidative metabolism in the reticulocyte for the incorporation of plasma-bound iron into hemoglobin. Saltman et al. (6) found that oxidative metab- olism of the liver cell was not concerned with the incorporation of iron into that organ. However, in the latter studies, iron in the form of ferric ammonium citrate was used at rather high concentrations instead of the physiological form of plasma-bound iron which is present in much lower concentrations in the cir- culation. From results of in vitro studies, Loewus and Fineberg (7) have suggested the specific need for ascorbic acid during the incorporation of iron, as ferric ammonium citrate, into ferritin.
Results of the present study demonstrate that the release of iron from its linkage to the plasma iron-binding protein, trans- ferrin, and its subsequent incorporation into hepatic ferritin, is dependent on oxidative metabolism of the liver cell, specifically for the continued synthesis of ATP. In addition to ATP, ascorbic acid is also required for the incorporation reaction. A mechanism is suggested to explain the roles of ATP and ascorbic acid in this reaction.
EXPERIMENTAL
Radioactive iron bound to the iron-binding protein of plasma or serum was prepared in two ways. Fe++59 citrate was incu- bated with rat plasma at room temperature for 30 minutes before its use. This form of iron will be called “plasma Fe59.” Radio- active iron salts were injected intraperitoneally into a rat and blood withdrawn from the abdominal aorta after 30 minutes. The serum separated from thii blood was used for most experi- ments in order to make quite certain that we were dealing with iron bound firmly to plasma transferrin, as it exists in the nor- mal circulation. This type of iron will be called “serum Fe5g.” In all cases the quantity of chemical iron, in the form of radio- active iron, was too small to influence the normal iron-binding capacity of the serum.
Female rats of the Carworth-Nelson strain, weighing 150 to 200 g were used throughout. Ferritin was isolated from rat liver slices after homogenization of 1.0 g wet weight aliquots of liver with 7 ml of cold Ringer-phosphate solution, heating of the
* Supported by a Grant No. A-1655 from the National Institutes of Health, United States Public Health Service.
homogenate, with careful stirring, to 70”, and clarification of the cooled extract by centrifugation. The insoluble residue was ex- tracted once with 4 ml of the salt solution and the pooled extracts mixed with excess rabbit antihorse spleen ferritin serum. The mixture was incubated for one hour at 37” or until flocculation of the antigen-antibody complex occurred. If time was not imme- diately available, the mixture was allowed to stand in the refriger- ator overnight. The ferritin-antibody precipitate was recovered by centrifugation and washed 3 times with 10 ml of saline. It was then counted in a well-scintillation detector and analyzed for total chemical iron. In a separate experiment it was found that recrystallized rat liver ferritin could be quantitatively pre- cipitated from solution by addition of suitable quantities of antihorse ferritin serum. Since the preparation of crystalline rat liver ferritin in quantities adequate for antiserum production is extremely costly, rabbit antiserum to crystalline horse spleen ferritin (8) was used throughout.
Radioactive iron was obtained from the Abbott Laboratories, Chicago, nucleotide phosphates from Sigma Chemical Company, St. Louis, and Schwarz Laboratories, 1Mt. Vernon. DEAE cellu- lose, type 20, was obtained from Brown Company, Berlin, New Hampshire. Purified ascorbic acid oxidase was a gift from Dr. C. A. Dawson and Miss Jean Dayan of Columbia University.
Distribution of Plasma FeSg in Rat-Rats were injected intra- venously with Fe59-labeled rat plasma in an attempt to deter- mine the quantity of plasma-bound iron which is directed to the liver under normal circumstances. Previous determinations of the rate of incorporation into liver or liver ferritin of plasma iron have been reported using the guinea pig (9), mouse (lo), and rat (11). In the latter experiments the iron was injected intraperi- toneally or intravenously and, where rats were used, the animals were fed or injected with abnormally large quantities of chemical iron in order to stimulate ferritin synthesis. In the present ex- periments, insignificant quantities of chemical iron were injected. Four to six animals were killed at varying time intervals and pooled specimens of washed red cells, plasma, perfused liver and liver ferritin analyzed for radioactivity. The results in Table I show that the liver had taken up a maximal quantity of circulat- ing Fe59 after two days, amounting to 20’% of the injected dose. One-half of the Fe% in the liver was present as ferritin. From values obtained for the 13th day, it may be concluded that the remainder of injected Fe5g, some SO%, had been taken up by the bone marrow for incorporation into red cells. The specific ac- tivity of ferritin iron remained fairly constant from the 2nd to 4th day, but dropped to lower values by the 13th day. These results demonstrate that the liver and its ferritin account for a substantial proportion of the iron which is removed normally
595
by guest on Novem
ber 27, 2018http://w
ww
.jbc.org/D
ownloaded from

596 Plasma Iron Incorporation into Ferritin Vol. 235, No. 3
TABLE I
Distribution of plasma-Fes9 in rat Each rat was injected with a pre-incubated mixture of rat
plasma and Fe69, as ferrous citrate, equivalent to less than 0.2 rg of chemical iron, containing 100,000 c.p.m. Fe69.
centrifugation of the cooled mixture until the solution was 0.5 saturated with the salt. The insoluble material was separated by centrifugation, dissolved in water, and reprecipitated at 0.5 satu- ration with ammonium sulfate. Thii process was repeated. The final precipitate was dissolved in a small quantity of water and dialyzed free of ammonium sulfate against 0.05 M phosphate buffer, pH 7.4, containing 0.15 mole per liter of NaCl. The resulting solution was then subjected to differential high speed centrifugation yielding four fractions. In Table II these are called top, intermediater, intermediates and bottom. Aliquots of each of these fractions were mixed with an excess of antiferritin serum and the washed specific precipitates analyzed for radio- activity, total iron, and total nitrogen. Table II lists the results expressed in two ways: as specific activity of ferritin iron (counts per minute per Fug of ferritin iron) and as counts per minute per pg of total nitrogen in the antigen-antibody precipitate. The latter is an index of the degree of association of radioactive iron with ferritin protein since the quantity of total nitrogen in the ferritin-antibody precipitate varies directly with the quantity of ferritin protein in that precipitate (8). The data obtained from this experiment, although they demonstrate a higher rate of incorporation of plasma iron into the top “apoferritin-rich” frac- tion than into the bottom “iron-rich” fraction, do indicate that, even as early as 1 hour after injection of Fe59, this iron has be- come associated to an appreciable extent with the highly aggre- gated iron-rich bottom fraction. By the 8th day the bottom fraction contained more than 5 times the quantity of Fesg per pg of protein N than that of the top fraction. The data, ex- pressed as specific activity of ferritin iron, illustrate the move- ment of radioactive iron with time from the top to the bottom fraction until by the 15th day all fractions have almost identical specific activities. These results suggest that the sites for iron incorporation are probably present in ferritin as well as in apofer- ritin molecules.
Incorporation of Plasma Iron into Liver Slices and Ferritin- Jandl et al. (5) have demonstrated that the incorporation of plasma-bound iron into reticulocytes is dependent on oxidative metabolism and that the rate for reticulocytes is much greater than that which occurs with liver slices. They also demonstrated that the state of saturation of the plasma iron-binding protein determines the availability of this iron for transfer to the liver; the more highly saturated the plasma globulin, the greater the extent of iron transfer. These results suggest that iron added to plasma iron-binding protein to the limits of its binding capac- ity is not held as tightly by this protein as is iron associated with an unsaturated iron-binding plasma protein, and therefore be- haves more like ionic iron. Saltman et al. (6) studied the trans- fer of iron into rat liver slices using ionic iron in relatively large quantities (90 ,ug per 100 mg wet weight of liver) and observed a relatively large uptake of iron into liver slices incubated in a nitrogen atmosphere, concluding that oxidative energy was not required for the transfer.
Since our experiments in viva had shown a relatively slow nor- mal rate of transport of plasma iron to the liver as compared with bone marrow, experiments were performed with liver slices incubated in a Krebs-bicarbonate medium containing Fe5g bound to normal rat plasma. In addition to measurement of radio- activity associated with the well washed slices, ferritin was iso- lated from an aliquot of such slices and assayed for Fess and total chemical iron. Table III shows clearly a marked inhibition of plasma iron incorporation into ferritin of liver slices when these
- 1 Injected Fe”8 in Specific activity
of iron in
Time
2 hr 5 hr
24 hr 2 days 3 days 4 days
13 days
Liver ferritin Yti:ll Liver
globint ferritin :ed cells* Plasma* Liver
% % 3.6 43.4 4.6 13.4
34.4 1.4 48.4 1.4 64.0 1.5 73.6 2.4 74.4 6.1
_-
-
% % c.#.m./pg Fe 8.9 3.9 0.6 13.3
14.3 5.8 0.7 16.8 15.0 6.4 5.4 22.3 19.3 9.5 8.3 26.2 15.2 7.0 11.1 21.9 15.2 7.5 12.1 22.4 12.4 7.5 12.6 15.9
F
_- .-
L
* Values for total red cells and plasma are based on the assump- tion of a blood volume equal to 8% of body weight.
t Values calculated from concentrations of erythrocyte iron and hemoglobin.
TABLE II
Incorporation of plasma Fe69 into ferritin fractions Each rat was injected with plasma-bound Fe69 as for Table I.
I- -
T Tiie Intermediatel Bottom
c.p.m./ c.p.m./ /a Fe Pg. N
1.5 1.8 6.7 4.3
10.8 9.3 8.6 6.4 6.4 5.1
TOP
c.pm/ pg Fe
3.0 14.0 14.0
8.1 6.6
:4&y
2.6 5.6 8.0 3.7 3.3
‘;“,.$ ‘
7.4 31.2 14.8
6.4 6.0
-
‘ 3 2.9 6.1 3.7 1.2 1.2
y;/
4.5 25.3 14.5
7.4 6.1
:;“y
2.8 9.0 4.9 1.2 2.4
-
1 hr 4 hr
24 hr 8 days
15 days 2.-
from the plasma. They also show a continuous movement of plasma iron to and from hepatic ferritin in the normal animal.
Incorporation of Plasma Iron into Fe&in Fractions in Vivo- Fineberg and Greenberg (12) have shown that C14-glycine in- corporated into hepatic ferritin at the earliest time interval after its injection, appears to the greatest extent in that portion of the ferritin molecular mixture which is rich in apoferritin and low in iron content. Rothen (13) had shown that a solution of crystalline ferritin could be separated by high speed centrifuga- tion into fractions with varying ratios of iron : nitrogen; the upper layer of the solution contained molecules with the slowest sedi- mentation rate which were essentially free of iron. The remain- ing solution contained molecules in an aggregated state with increasing iron : nitrogen ratios, as their relative sedimentation rates increased. Differential high speed centrifugation in the Model L Spinco enables one to separate such fractions (14). In the experiments of Fineberg and Greenberg (12), this tech- nique was utilized to demonstrate that apoferritin was formed first and that iron was incorporated later to form ferritin. In the present study a similar technique was used to separate four arbitrary fractions of rat ferritin at various time intervals after the intravenous injection of plasma Fe5g. Livers from 6 to 10 rats, for each time interval, were pooled, homogenized with Ringer-phosphate solution and the homogenate heated to 70”. Ammonium sulfate was added to the clear extract obtained after
by guest on Novem
ber 27, 2018http://w
ww
.jbc.org/D
ownloaded from

March 1960 A. Maxur, X. Green, and A. Carleton 597
are incubated in an atmosphere of nitrogen. In contrast to thii effect, anaerobiosis did not inhibit the uptake of Fesg by the total liver; indeed, there was some increase above that obtained with slices incubated aerobically. Analyses for chemical iron of ferritin isolated from slices incubated anaerobically demonstrated a progressive loss of ferritin iron as compared with that from aerobic slices. These results are in confirmation of our earlier studies (1, 2) of the mechanism of stimulation of ferritin iron release by liver slices at low oxygen tensions. Despite the loss of ferritin iron due to anaerobic conditions shown in Table III, calculation of specific activities of ferritin iron isolated from these slices shows a marked inhibition of plasma iron incorporation into ferritin in the absence of oxygen.
Table IV shows the stimulation of plasma iron incorporation into rat liver slices produced by addition to the medium of a variety of substrates for oxidative enzymes. The values reported are compared with those obtained for slices incubated in a sub- strate-free medium, whose total liver Fesg content and ferritin iron specific activities were adjusted to 1.0. All substrates were present in the form of neutralized solutions at a final concentra- tion of 0.01 M and aerobic incubation terminated at the end of one hour. Acids of the Krebs cycle or compounds which can be converted to such acids were found to be stimulatory. Citric acid was most effective, whereas glucose or /3-hydroxybutyric acid were without effect. These results confirm the need for oxidative metabolism during the incorporation of plasma iron into liver ferritin and are quite unlike those obtained by Saltman et al. (6) who used ionic iron and found that compounds such as citric acid were inhibitory to the incorporation of iron into the liver presumably by forming an undissociable complex with ionic iron and making such iron unavailable for incorporation into the liver cell.
Table IV also lists the effect of addition of various inhibitors to the medium, all at a final concentration of 5 X 10e3 M. In- hibition by these compounds strongly suggests the need for par- ticipation of intact metabolizing cells during the incorporation of iron bound to the iron-binding protein of plasma into ferritin.
In order to resolve several of the differences in results obtained with plasma-bound iron as compared with that reported for ionic iron, liver slices were incubated with radioactive iron bound to plasma and in separate flasks with an equivalent amount of radio- active iron in ionic form. Table V shows clearly that ionic iron is incorporated into the liver cell as well as into ferritin to an extent 7 times as great as an equivalent amount of plasma-bound iron. However, whereas anaerobic conditions do not decrease incorporation of either form of iron into the total liver cell, inhi- bition of incorporation into ferritin takes place only if plasma- bound iron is used. Further differences in incorporation into ferritin between ionic and plasma iron are noted when inhibitors are added to the aerobic incubation mixtures. Whereas iodo- acetamide, a general sulfhydryl alkylating agent, inhibits the incorporation of either form of iron to the same extent, arsenite is more inhibitory when plasma iron is used, and dinitrocresol inhibits only the incorporation of plasma iron into ferritin of liver slices.
These results suggest that some aspect of oxidative metabolism, possibly oxidative phosphorylation, because of the striking differ- ence in effects of dinitrocresol, is involved in the removal of iron from its normal linkage to the plasma iron-binding protein. The results also suggest that oxidative metabolism is not involved
TABLE III Incorporation of plasma Fe69 by liver slices
Each incubation flask contained 2.0 g of pooled rat liver slices, 10 ml of Krebs-Ringer-bicarbonate and 0.5 ml of diluted rat plasma tagged with 100,000 c.p.m. of Fe69. Incubation was car- ried out at 37” in an atmosphere of 95% oxygen-5% carbon dioxide, or 95yo nitrogen-5yo carbon dioxide. The slices were washed three times with 50-ml portions of 0.9% sodium chloride solution before removal of aliquots of 0.5 g forcountingand 1.0 g forferritin isolation.
Time
min
30 60 90
120
Radioactivity of
Liver Ferritin
c.p.?n./g liver
602 211 766 288 862 473
1050 550
Nitrogen
Ferritin iron
M 88 80 82 83
Radioactivity of Ferni;in
Liver Ferritin
c.#.m./g zioer
705 32 if 962 34 53
1131 29 42 1231 38 31
TABLE IV
E$ect of substrates and inhibitors on plasma Fe69 incorporation into liver slices
Conditions the same as in Table III, using oxygen-carbon diox- ide. Control flasks contained no added substrate, and the Fe69- content of the total liver as well as the specific activity of the fer- ritin obtained from this control liver are adjusted to values of 1.0. Substrates were present at a final concentration of 0.01 M, whereas inhibitors were present at a final concentration of 5 X 10-s M.
Radioactivity in Substrate -
Liver
Control.. . 1.0 Citrate. . . . . 2.1 Oxalacetate.. . . 1.8 Malate.. . . . 1.5 Fumarate....... 1.5 Lu-Ketoglutarate. 1.1 Succinate . . . 1.0 Pyruvate 1.0 Lactate......... 1.0 Aspartate. 1.0 Glutamate. 1 .O Glucose......... 1.0
fl-Hydroxybutyr- ate.. . . . . . . . . . 1.1
Ferritin
1.0 4.6 2.7 2.7 2.4 1.8 1.8 1.8 1.7 1.6 1.2 1.0
Inhibitor
Control. . Iodoacetamidc N-Ethyl male-
imide . . . . . Cyanide...... Arsenite.. Dinitrocresol Dinitrophenol Azide . .
T Radioactivity in
-_
I.
-
Liver
1.0 1.2
1.0 1.1 1.0 0.9 0.8 1.2
Ferritin
1.0 0.1
0.1 0.2 0.2 0.3 0.5 0.5
once the iron is liberated from such a protein bond, in which state it passes into the liver cell unimpeded.
Incorporation of Plasma Iron into Ferritin of Homogenates- Rat liver homogenates, prepared by use of a solution of 0.15 M KC1 containing 0.64 mmole per liter of KHCO,, were incubated with plasma-bound Fesg in an atmosphere of oxygen and the effect on iron incorporation into its ferritin noted when various compounds were added. Table VI demonstrates the stimulatory effect of nicotinamide, DPN and TPN. These results confirm the need of oxidative enzymes for the incorporation reaction. Nicotinamide probably protects DPN and TPN from destruction by intracellular enzymes (15, 16). The greatest enhancement
by guest on Novem
ber 27, 2018http://w
ww
.jbc.org/D
ownloaded from

TABLE V Comparison of incorporation of ionic iron with plasma iron into
liver slices Each flask contained 2.0 g of liver slices from a common pool
together with 10 ml Krebs-Ringer-bicarbonate medium and lOO,- 000 c.p.m. of Fe59 and were incubated for one hour at 37”. The values listed in parentheses for the control flasks are c.p.m. per g of liver, adjusted to 1.0 for comparison with the experimental flasks.
I Ferric59 citrate T Plasma-irons9 Additions
Liver Ferritin Liver Ferritin
Control. . 1.0 (7020) 1.0 (2874) 1.0 (1070) 1.0 (355) Nitrogen 1.1 1.2 1.4 0.2 Iodoacetamide. 1.0 0.2 1.1 0.2 Arsenite. 1.6 0.6 1.0 0.3 Dinitrocresol 1.3 1.1 1.0 0.3
TABLE VI Incorporation of serum Fe59 into ferritin of liver homogenates
Each flask contained the equivalent of 1.0 g wet weight of rat liver in the form of a homogenate prepared with0.15 M KCl-0.0006 M KHCO, and serum-bound Fe59 (50,000 c.p.m.). Flasks were in- cubated at 37” for 30 minutes in an atmosphere of 100% oxygen. Specific activites of ferritin iron of homogenates in the experi- mental flasks were compared with that isolated from the control flask which was adjusted to 1.0.
Additions Relative specific ac- tivity of ferritin Fe
598 Plasma Iron Incorporation into Ferritin Vol. 235, No. 3
the incorporation of serum Fe59 into ferritin of a supernatant solution prepared from a rat liver homogenate as well as the effect of added ATP. The supernatant, without additional ATP had a low level of activity, and this activity fell off after 60 min- utes. Addition of 10 hmoles ATP increased the reaction velocity but straight line kinetics held only for 90 minutes. The presence of 20 pmoles ATP yielded the greatest activity which was main- tained essentially unaltered for 2 hours. These results indicate that ATP is destroyed during the incubation procedure, but it is not possible from these results to determine whether this altera- tion of ATP is due to the iron incorporation reaction or to en- zymes present in the supernatant solution.
Participation of Ascorbic Acid during Incorporation of Plasma Iron into Hepatic Fe&tin-The effect of adding ATP to the su- pernatant solution prepared from a rat liver homogenate was
TABLE VII Distribution of serum Fe59 incorporating activity in
rat liver fractions
Rat liver was homogenized with 0.25 M sucrose and subjected to differential centrifugation, yielding the following fractions: nuclei, mitochondria, microsomes, and supernatant solution. Each flask contained the equivalent of 0.1 g of the appropriate fraction, recrystallized horse-spleen ferritin (100 pg of total iron), serum-bound Fe@ (50,000 c.p.m.), 2.5 ml 0.154 M KC1-0.0006 M KHCOs, 2.5 ml 0.2 M Tris buffer, pH 7.4, and 0.154 M KC1 to make 10.0 ml. Incubation was continued for 30 minutes at 37” in an atmosphere of 100% oxygen before ferritin isolation.
Fractions Ferritin iron specific activity
c.p.m./pg iron None........................................ Nicotinamide (0.1 M). Nicotinamide (0.1 M) + DPN (3.4 pmoles). Nicotinamide (0.1 M) + TPN (2.6 pmoles) Nicotinamide (0.1 M) + F- (0.1 M) Nicotinamide (0.1 M) + F- (0.1 M) + ATP (5
pmoles) . . Nicotinamide (0.1 M) + 1;” (0.1 M) + ATP (10
rmoles).................................... Nicotinamide (0.1 M) + F- (0.1 M) + ATP (20
rmoles).....................................
1.0 1.7 2.0 4.0 9.2
10.1
12.2
14.9
All. 11.1 Nuclei........................................ 0.7 Mitochondria 5.5 Microsomes................................... 0.0 Supernatant solution.. 11.9 Supernatant solution + Nuclei.. 12.4 Supernatant solution + Mitochondria. 15.5 Supernatant solution + Microsomes.. 10.2
I I I I I I I I 100 -
of plasma iron incorporation into ferritin occurred after the addi- tion of F- together with nicotinamide; an increase of g-fold above that obtained with the unfortified homogenate. Since F- is known to protect ATP from hydrolysis by ATPase in homoge- nates (17), the effect of additional ATP was studied. The results demonstrated the effectiveness of added ATP and suggest that the homogenate was not saturated with ATP insofar as its action on the iron incorporation reaction is concerned.
Fractionation of a rat liver homogenate prepared with 0.25 M sucrose by the method of Schneider (18) led to the finding (Table VII) that most of the system responsible for the incorporation of plasma iron is present in the clear supernatant solution. For this experiment an endogenous source of ferritin, horse-spleen ferritin, was used as the acceptor. It was incubated with the appropriate fraction, the latter in quantities equivalent to 0.1 g of original wet weight of liver. Mitochondria possess some ac- tivity which may be due to the presence of ATP or ATP-generat- ing systems in such particles.
60 90 120 Time (minutes)
FIG. 1. Incorporation of serum-bound Fe59 into ferritin of a clear extract prepared from a rat liver homogenate. Each reac- tions flask contained 50,000 c.p.m. Fe5Q, a quantity of extract equivalent to 1.0 g wet weight of rat liver which contained 100 pg of ferritin total iron, 3.5 ml of 0.2 M Tris buffer, pH 7.4, and 0.154 M KC1 to make 10 ml. Flasks were incubated in air at 37” with _ . _. Fig. 1 shows the time-course of the reaction which results in gentle shaking.
by guest on Novem
ber 27, 2018http://w
ww
.jbc.org/D
ownloaded from

March 1960 A. Mazur, S. Green, and A. Carleton 599
next investigated. As in the previous experiment, the source of ferritin was recrystallized horse-spleen ferritin. The results in Table VIII indicate that a factor, in addition to ATP, is present in the supernatant solution, and that this factor is concerned with the iron incorporation reaction. This factor is fairly stable to heating to 100” for one minute, but is almost completely removed by overnight dialysis against saline. Because of the reported involvement of ascorbic acid in the incorporation reaction (7), the identity of ascorbic acid with the factor present in the super- natant solution was investigated. The heated solution was mixed with ascorbic acid oxidase, at pH 5.6, for 15 minutes at room temperature and its activity tested in the presence as well as in the absence of added ATP after adjusting the solution to pH 7.4. Ascorbic acid oxidase completely destroyed the iron incorporation activity present in the supernatant solution, con- firming the presence of ascorbic acid in this solution and demon- strating the need for both ATP as well as ascorbic acid during the incorporation of serum iron into ferritin.
Serum Iron Incorporation into Various Fe&tins by ATP and Ascorbic Acid-Table IX lists the results of an experiment de- signed to determine the optimal ratio of ATP:ascorbic acid re- quired for the incorporation reaction. For this purpose rat fer- ritin was prepared in a partially purified state from the clear extract which results after centrifugation of a rat liver homoge- nate at 8500 x g for 1 hour. Ferritin was precipitated at 0.5 saturation with ammonium sulfate, the precipitate dissolved in water and dialyzed free of salts. An optimal molar ratio of ATP:ascorbic acid of 2 : 1 was obtained.
Table X lists the comparative activities of a variety of ferritins, tested as acceptors for serum-bound Fess in the presence of opti- mal quantities of ATP and ascorbic acid. Recrystallized horse- spleen ferritin was prepared as previously reported (19). Crys- talline rat ferritin was prepared from a large pool of rat livers by the same technique as that used for horse-spleen ferritin. Frac- tion A represents the clear extract obtained after centrifugation of a rat liver homogenate at 8500 x g for one hour, followed by exhaustive dialysis. Fraction B was prepared by precipitation of the ferritin from fraction A at 0.5 saturation with ammonium sulfate, solution of the precipitate in water, dialysis to remove salts, adjustment to pH 4.6, removal of an insoluble precipitate and adjustment of the resulting solution to pH 7.4. Fraction B was heated to 70” and the coagulated protein removed, yielding fraction C. Fraction C was centrifuged in the Model L Spinco for one hour at 80,000 X g and the top one-third of the solution removed and discarded. An equivalent quantity of saline was added, the solutions mixed and centrifuged again. This process was repeated for a total of four times, yielding fraction D. Purili- cation is indicated by a rise in the ratio of ferritin iron to total nitrogen and demonstrates that much of the nonferritin contami- nating protein is of lower density than the bulk of ferritin. Final purification was achieved by separating fraction D on a DEAE cellulose exchange column using increasing concentrations of phosphate buffer, pH 7.7, for gradient elution. Ferritin was strongly adsorbed to the cellulose at a concentration of buffer of 0.005 M but was eluted at 0.05 M buffer. This fraction, called E in Table X, was about 30yo pure when compared with crys- talline rat ferritin. It may be seen that continued purification of crude rat liver ferritin resulted in an increase in activity as an acceptor for serum-bound iron. These results cast doubt on the presence in liver of factors other than ATP and ascorbic acid required for the incorporation reaction. The fact that fraction
TABLE VIII Properties of factor, in addition to ATP, required for serum
Fe69 incorporation into ferritin
Each flask contained horse-spleen ferritin with KCl-KHCOZ solution and quantities of supernatant, prepared by the cen- trifugating of a rat liver homogenate at 9000 X g for 1 hour, equivalent to 0.1 g wet weight of liver. For treatment with ascorbic acid oxidase the pH of the supernatant was adjusted to pH 5.6,116 units of enzyme added, and the mixture incubated for 15 minutes at room temperature. It was then adjusted to pH 7.4 and tested as for Table VII.
Specific activity of ferritin iron Treatment of supernatant
Without ATP With ATP
c.p.m./pg iron Untreated............................. 6.8 1 19.3 Heated to lOO”......................... 6.8 11.2 Dialyzed.............................. 3.2 3.3 Heated supernatant + ascorbic acid
oxidase.............................. 0.0 0.0
TABLE IX
Optimal ratio of ATP:ascorbic acid for serum Fe59 incorporation into rat ferritin
Source of ferritin was rat liver whose preparation is described in text. Serum Fe69 (50,000 c.p.m.) was added to each flask which was incubated for 1 hour at 37”. Concentration of ATP and ascorbic acid is expressed as pmoles.
Additions Ferritin iron speciiic activity
C.Q.?n.l pg iron
None ......................................... 0.7 20ATP ....................................... 3.4 20ATP + lascorbate ........................ 33.8 20 ATP + 5 ascorbate ........................ 50.2 20ATP + loascorbate ....................... 54.0 20 ATP + 20 ascorbate ....................... 52.0
TABLE X
Serum Fe69 incorporation into various ferritins by ATP and ascorbic acid
Each flask contained a quantity of ferritin equivalent to 100 pg of total iron together with Tris buffer and serum-bound Fe69 as in Table IX. Ascorbic acid (10 pmoles) and ATP (20 pmoles) were added before incubation for 30 minutes in an atmosphere of nitrogen. Ferritin was isolated immunochemically.
Source of ferritin Ferritin Fe I I Specific Total N activity
ferritin iron
Horse spleen (recryst. 4 times). . Rat liver (recryst. 1 time). . Ratliver;A............................. Rat liver; B.. . Rat liver; C............................ Ratliver;D............................. Rat liver; D + Cd++ (2 X N+M) . Rat liver; D + p-chloromercuribenzoate
mg ironjmg N c.p.m./pg iron 1.87 20.8 1.22 13.2 0.006 8.4 0.016 18.8 0.043 17.9 0.112 18.5 0.112 9.8
(2 x 10-M) 0.112 5.0 Ratliver;E............................ 0.433 25.0
by guest on Novem
ber 27, 2018http://w
ww
.jbc.org/D
ownloaded from
600 Plasma Iron Incorporation into Ferritin Vol. 235, No. 3
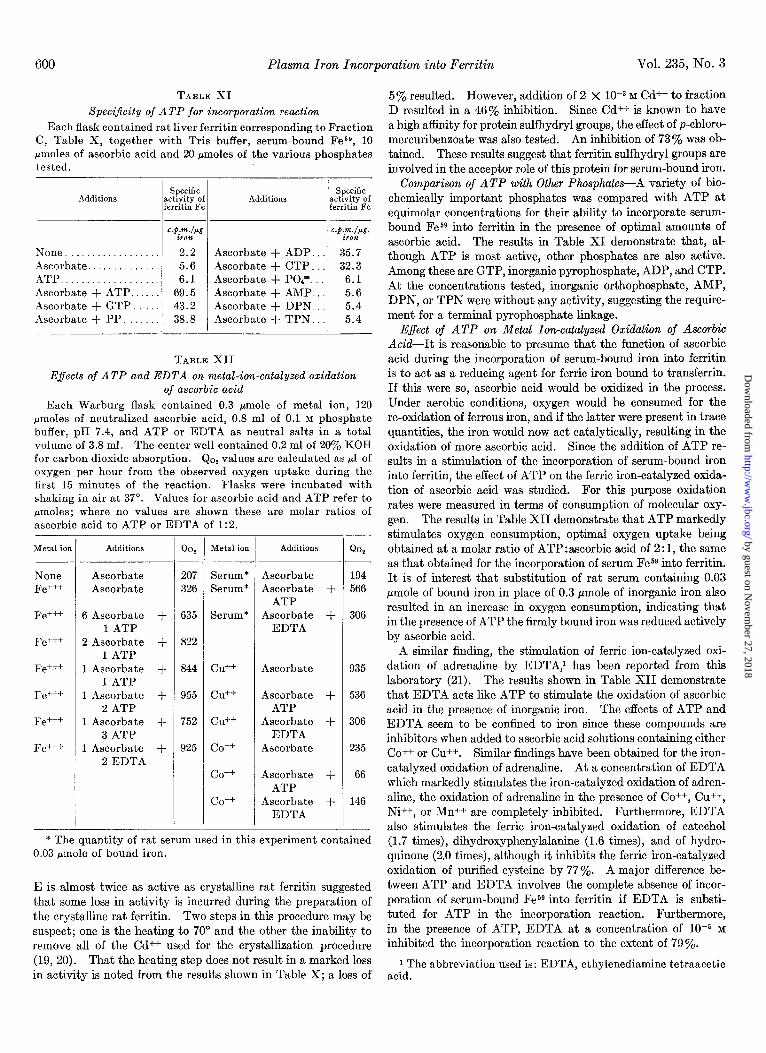
TABLE XI
Speci$city of ATP for incorporation reaction
Each flask contained rat liver ferritin corresponding to Fraction C, Table X, together with Tris buffer, serum-bound Fes9, 10
pmoles of ascorbic acid and 20 pmoles of the various phosphates tested.
Additions
None. Ascorbate. ATP Ascorbate + ATP. Ascorbate + GTP. Ascorbate + PP.
Specific ctivity of xritin Fe
Additions
:.p.m./pg .P.dP&-. iron iron 2.2 Ascorbate + ADP. 35.7 5.6 Ascorbate + CTP. 32.3 6.1 Ascorbate + POP.. 6.1
69.5 Ascorbate + AMP.. 5.6 43.2 Ascorbate + DPN. 5.4 38.8 Ascorbate + TPN. 5.4
TABLE XII
Specific ctivity of xritin Fe
E.fects of ATP and EDTA on metal-ion-catalyzed oxidation of ascorbic acid
Each Warburg flask contained 0.3 pmole of metal ion, 120 pmoles of neutralized ascorbic acid, 0.8 ml of 0.1 M phosphate buffer, pH 7.4, and ATP or EDTA as neutral salts in a total volume of 3.8 ml. The center well contained 0.2 ml of 20% KOH for carbon dioxide absorption. Qop values are calculated as ~1 of oxygen per hour from the observed oxygen uptake during the first 15 minutes of the reaction. Flasks were incubated with shaking in air at 37”. Values for ascorbic acid and ATP refer to pmoles; where no values are shown these are molar ratios of ascorbic acid to ATP or EDTA of 1:2.
Metal ion Additions
Ascorbate Ascorbate
None Serum* 194 Fe+++ Serum* 566
Fe+++ Serum* 306
Fe+++
Fe+++ Cd+ 935
Fe+++ Cd-’ 536
Fe+++ cu++ 306
Fe+++ co++ 235
co++ 66
co++ 146
* The quantity of rat serum used in this experiment contained 0.03 &mole of bound iron.
6 Ascorbate + 1 ATP
2 Ascorbate + 1 ATP
1 Ascorbate + 1 ATP
1 Ascorbate + 2 ATP
1 Ascorbate + 3 ATP
1 Ascorbate + 2 EDTA
- Qo,
207 326
635
822
844
955
752
925
Ascorbate Ascorbate +
ATP Ascorbate +
EDTA
Ascorbate
Ascorbate + ATP
Ascorbate + EDTA
Ascorbate
Ascorbate + ATP
Ascorbate + EDTA
Metal ion Additions Qo,
E is almost twice as active as crystalline rat ferritin suggested that some loss in activity is incurred during the preparation of the crystalline rat ferritin. Two steps in this procedure may be suspect; one is the heating to 70” and the other the inability to remove all of the Cd++ used for the crystallization procedure (19, 20). That the heating step does not result in a marked loss in activity is noted from the results shown in Table X; a loss of
5% resulted. However, addition of 2 x 1O-3 M Cd++ to fraction D resulted in a 46% inhibition. Since Cd++ is known to have a high afhnity for protein sulfhydryl groups, the effect of p-chloro- mercuribenzoate was also tested. An inhibition of 730/, was ob- tained. These results suggest that ferritin sulfhydryl groups are involved in the acceptor role of this protein for serum-bound iron.
Comparison of ATP with Other Phosphates-A variety of bio- chemically important phosphates was compared with ATP at equimolar concentrations for their ability to incorporate serum- bound Fesg into ferritin in the presence of optimal amounts of ascorbic acid. The results in Table XI demonstrate that, al- though ATP is most active, other phosphates are also active. Among these are GTP, inorganic pyrophosphate, ADP, and CTP. At the concentrations tested, inorganic orthophosphate, AMP, DPN, or TPN were without any activity, suggesting the require- ment for a terminal pyrophosphate linkage.
E$ect of ATP on Metal Ion-catalyzed Oxidation of Ascorbic Acid-It is reasonable to presume that the function of ascorbic acid during the incorporation of serum-bound iron into ferritin is to act as a reducing agent for ferric iron bound to transferrin. If this were so, ascorbic acid would be oxidized in the process. Under aerobic conditions, oxygen would be consumed for the re-oxidation of ferrous iron, and if the latter were present in trace quantities, the iron would now act catalytically, resulting in the oxidation of more ascorbic acid. Since the addition of ATP re- sults in a stimulation of the incorporation of serum-bound iron into ferritin, the effect of ATP on the ferric iron-catalyzed oxida- tion of ascorbic acid was studied. For this purpose oxidation rates were measured in terms of consumption of molecular oxy- gen. The results in Table XII demonstrate that ATP markedly stimulates oxygen consumption, optimal oxygen uptake being obtained at a molar ratio of ATP:ascorbic acid of 2:1, the same as that obtained for the incorporation of serum Fe59 into ferritin. It is of interest that substitution of rat serum containing 0.03 pmole of bound iron in place of 0.3 pmole of inorganic iron also resulted in an increase in oxygen consumption, indicating that in the presence of ATP the firmly bound iron was reduced actively by ascorbic acid.
A similar finding, the stimulation of ferric ion-catalyzed oxi- dation of adrenaline by EDTA,’ has been reported from this laboratory (21). The results shown in Table XII demonstrate that EDTA acts like ATP to stimulate the oxidation of ascorbic acid in the presence of inorganic iron. The effects of ATP and EDTA seem to be confined to iron since these compounds are inhibitors when added to ascorbic acid solutions containing either CO++ or Cu++. Similar findings have been obtained for the iron- catalyzed oxidation of adrenaline. At a concentration of EDTA which markedly stimulates the iron-catalyzed oxidation of adren- aline, the oxidation of adrenaline in the presence of CO++, Cu++, Ni++, or Mn++ are completely inhibited. Furthermore, EDTA also stimulates the ferric iron-catalyzed oxidation of catechol (1.7 times), dihydroxyphenylalanine (1.6 times), and of hydro- quinone (2.0 times), although it inhibits the ferric iron-catalyzed oxidation of purified cysteine by 77%. A major difference be- tween ATP and EDTA involves the complete absence of incor- poration of serum-bound Fe59 into ferritin if EDTA is substi- tuted for ATP in the incorporation reaction. Furthermore, in the presence of ATP, EDTA at a concentration of 1O-5 M inhibited the incorporation reaction to the extent of 79%.
i The abbreviation used is: EDTA, ethylenediamine tetraacetic acid.
by guest on Novem
ber 27, 2018http://w
ww
.jbc.org/D
ownloaded from

March 1960 A. Maxur, S. Green, and A. Carleton 601
DISCUSSION
The importance of iron for hemoglobin synthesis by bone mar- row has directed attention away from the movement of plasma iron to other tissues. In the rat, 20 % of the iron which normally leaves the plasma enters the liver cell where one-half of this iron is incorporated into ferritin. The rapidity of ferritin protein synthesis (22) and of incorporation of iron into various ferritin protein molecules as illustrated in Table II, suggests that this protein, although it may function as a storage depot for iron in an emergency, is normally in a constant state of renewal and that its iron passes into and out of its molecules constantly, to and from the circulating plasma. This gives rise to an iron cycle: plasma -+ hepatic ferritin + plasma, comparable to, although quantitatively smaller than, the iron cycle involving hemoglobin synthesis: plasma -+ marrow + red cell + senescent red cell -+ plasma.
Values for the rate of incorporation of plasma iron into various ferritin fractions suggest that iron is incorporated by attachment to sites on the protein, of which more are available in those mole- cules which contain little iron (apoferritin) and which have been most recently synthesized (12). Inhibition of incorporation of plasma-bound iron into ferritin by Cd++ or p-chloromercuriben- zoate suggests that such sites involve sulfhydryl groups. Ana- lytical data reported from this laboratory (23) have shown that the apoferritin-like fraction of ferritin contains more SH groups than do the others although SH groups are present even in those ferritin molecules containing the largest amount of iron. These results agree with our present findings which show a substantial incorporation of plasma iron into all fractions as early as one hour after the iron has been injected intravenously into the ani- mals.
The marked differences observed in experiments with liver slices between ferric citrate and plasma-bound iron emphasize the necessity for use of the physiological form of iron in experi- ments in vitro. When ionic iron is injected into the intact animal the iron is rapidly bound by the plasma iron-binding globulin, transferrin, although, even in this type of experiment, one must avoid the administration of quantities of iron which will saturate circulating transferrin. From the results of Jandl et al. (5) it is unlikely that transferrin saturated with iron binds all of its iron as avidly as is the case for iron normally present in the circulating plasma. Although greater incorporation of iron into the liver slice and its ferritin takes place with use of ferric citrate, this process is entirely independent of the living cell and may be con- sidered an artifact. The relatively slow incorporation of plasma- bound iron, however, is in accord with the rate of uptake of plasma iron by the liver in the intact animal. In the intact animal iron in the plasma is linked with transferrin with a bind- ing constant of approximately 10’ (24). This binding serves as a brake against the rapid movement of iron to the tissues. Al- though the use of ferric citrate together with liver slices leads to incorporation of iron into ferritin, it is uncertain whether iron bound to the protein in this way is identical with that which exists in native ferritin.
Regulation of iron transport from circulating plasma to hepatic ferritin is determined by the state of oxidative metabolism of the liver cell, e.g. oxygen tension in the blood flowing through the liver. Aerobic metabolism in the liver is needed for the synthe- sis of ATP which, together with ascorbic acid, serve to transport plasma-bound iron to ferritin. The fact that liver slices bind plasma iron regardless of oxygen tension or the presence of in-
(l/2 Ascorbic Acid) 0
PH -- .- (# 6) C-N<> ,%,
(,,)_\,!JY& ‘p=o -- P I
I \ (Transferrin) Q
\ P=O \
etc-(Ri hosed FIG. 2. Complex postulated for ATP, ascorbic acid, and serum-
bound iron.
hibitors of oxidative enzymes, suggests that the binding site on the liver cell wall is not dependent on energy sources (ATP) or that such a site has a very low rate of turnover. It is likely, however, that ATP and ascorbic acid are present in the aerobic cell at or near this binding site in order to remove the iron from its linkage with transferrin. It is postulated that this linkage is broken and transferrin, minus its iron, returned to the circulation, on the basis of the great differences in turnover of plasma iron as compared with that of plasma globulins (25, 26), as well as by the fact that alteration of the level of transferrin in the plasma does not affect the rate of movement of iron from the plasma (27). It is apparent that the removal of iron from its linkage to transferrin requires the presence of a compound with strong binding properties for iron. ATP is eminently suitable for this purpose since, as Neuberg and Mandl (28) have shown, ATP is capable of preventing the precipitation from solution of ferrous or ferric sulfides.
The role of ascorbic acid in the reaction which leads to the transport of plasma iron to ferritin presumably involves its action as a reducing agent, converting transferrin iron from the ferric to the ferrous state. This conversion takes place to a limited extent at pH 7.4, if at all, unless ATP is present. It may there- fore be presumed that a complex is formed among ATP, ascorbic acid, and the iron attached to transferrin. Our data reveal that such a complex would consist of 2 moles of ATP for each mole of ascorbic acid. Formulation of such a complex is shown in Fig. 2 and is based on the Mg++-ATP complex suggested by Szent-Gyorgyi (30), as well as on the results of our experiments with phosphate derivatives related to ATP. Whereas Mg++ would be expected to form a quadridentate complex, Fe+++ is shown in the form of a hexadentate structure involving 2 oxygen atoms of the terminal pyrophosphate linkages of ATP, the N atom attached to carbon atom 6 and N 7 of the adenine rings, one of the oxygen atoms of ascorbic acid which is capable of electron transfer and, finally, the unknown linkage of the iron atom with transferrin. Transfer of an electron from ascorbic acid to the iron atom would result in its reduction to the ferrous state, presumed weakening of its linkage to transferrin and liber- ation of an Fe++-ATP complex. How the iron from such a complex is transferred to ferritin is not known although the participation of SH groups of the protein is evident. One
by guest on Novem
ber 27, 2018http://w
ww
.jbc.org/D
ownloaded from

602 Plasma Iron Incorporation into Few&n Vol. 235, No. 3
possibility involves the splitting of ATP at its terminal phosphate bond due to its linkage with iron. Such a linkage, as Boyer and Harrison have pointed out (30) would favor an increase in posi- tive charge on the phosphorus atom and lead to an increased ease of attack by nucleophilic reactants (OH-). Indeed, Bamann et al. (31) have demonstrated the phosphatase-like activity of ionic iron leading to a splitting of the terminal phosphate of ATP, although the reaction is slow. In this regard, it is of great inter- est that native ferritin does contain phosphorus (20) and the possibility exists that this phosphorus arises from ATP which carries iron to ferritin for its incorporation. The use of ATP whose terminal P is labeled will be helpful in elucidating this point.
Model experiments involving the measurement of oxidation rates of ascorbic acid by Fe+++ in the presence of ATP or EDTA emphasize the activation effect of formation of covalent com- plexes of metal ions. ATP accelerates the oxidation rate of ascorbic acid in the presence of small amounts of Fe+++ or serum- bound iron, and EDTA which forms a complex with Fe+++ ex- tremely similar to that postulated for ATP, also stimulates the Fe+++-catalyzed oxidation of ascorbic acid. The effects of ATP and EDTA are specific for Fe+++; although Cu++ or Co++ cata- lyze the oxidation of ascorbic acid, the reaction is not stimulated by addition of either complexing agent. Preliminary studies suggest that ascorbic acid is not entirely specific since the Fe+++- catalyzed oxidation of adrenaline is stimulated by both ATP and EDTA and that of catechol, dihydroxyphenylalanine or hydroquinone is stimulated by EDTA, whereas the Fe+++-cata- lyzed oxidation of cysteine is inhibited by EDTA. There ap- pears to be a requirement for an acidic OH group, probably for the purpose of bonding via one of the coordinate valences of Fe+++. The results cited for cysteine are in disagreement with that reported by Pfleger and Rummel (32). We have found it necessary to purify cysteine with extreme care using the tech- nique of Warburg and Sakuma (33) in order to remove extraneous metal ions which interfere with the EDTA effect. The fact that both ATP and EDTA stimulate the Fe+++-catalyzed oxidation of ascorbic acid whereas only ATP accelerates or makes possible the incorporation of serum-bound iron into ferritin suggests once again that the Fe++-ATP complex is probably split during or immediately before the attachment of the iron to the ferritin molecule, since such a splitting of the Fe++-EDTA complex is not possible.
As a result of the present findings, an interesting relationship appears between the biochemical mechanisms involved in iron release and iron incorporation involving ferritin. The release of iron from hepatic ferritin to the plasma is favored by decreased oxygen supply to the liver cell causing an accelerated breakdown of nucleotides including ATP, yielding elevated cellular levels of hypoxanthine and xanthine, and a consequent increase in oxida- tion of these substrates by xanthine oxidase, a process which involves ferritin iron reduction and release (1, 2). As is now evident, the reverse reaction, the transfer and incorporation of plasma-bound iron into hepatic ferritin is dependent on energy- yielding reactions for the continued synthesis of ATP, which, together with ascorbic acid, releases iron from its linkage to the plasma iron-binding protein and serves to incorporate it into ferritin.
It is quite likely that the mechanism for incorporation of plasma iron into hepatic ferritin which has been described also functions with respect to the insertion of iron into ferritin in
other tissues, such as marrow, spleen, intestinal mucosa and placenta, since ATP is common to all tissues. If intercellular rather than plasma ascorbic acid is needed for this reaction it is of interest that the vitamin is found to the greatest extent in the liver cell supernatant fraction (34)) and that oral administra- tion of ascorbic acid is extremely effective in increasing the ex- tent of oral iron absorption (35) during which time increasedquan- tities of ferritin appear in the mucosal cells (36). The effect of ascorbic acid deficiency in the guinea pig on the incorporation of plasma iron into ferritin is now under study.
SUMMARY
Fesg bound to plasma or serum has been used to study the mechanism of incorporation of iron into hepatic ferritin. Of the iron which leaves the plasma of a normal rat, 20% is incorporated into the liver; one-half of this iron is present as ferritin. Distribution of this iron among the ferritin molecules of varying total iron content indicates that plasma-bound iron is at first incorporated to a somewhat greater extent into low den- sity iron-poor molecules which with time are converted to high density ferritin molecules of high iron content.
Incorporation of plasma-bound iron into ferritin of liver slices or homogenates is dependent on energy produced during oxida- tive metabolic reactions, specifically for the synthesis of adeno- sine triphosphate (ATP). In addition to ATP, ascorbic acid is required for the incorporation reaction. Using pure compounds, neither ATP nor ascorbic acid alone is capable of transferring plasma-bound Fess to ferritin. However, when added together, ATP and ascorbic acid perform this transfer reaction. Evidence is presented which suggests the presence of ferritin SH groups as part of the acceptor site.
The mechanism whereby ATP and ascorbic acid mediate the transfer of plasma-bound iron to ferritin has been studied. The formation of a complex involving 2 moles of ATP, 1 mole of ascorbic acid, and the iron-transferrin protein of plasma is sug- gested. ATP markedly stimulates the oxidation of ascorbic acid in the presence of ionic iron or iron bound to rat serum. It is suggested that this reaction serves to reduce the plasma-bound ferric iron to the ferrous state, thus releasing it from its bond to the protein and making it available for incorporation into ferritin. Model experiments using ATP and ethylenediamine tetraacetic acid point to the specificity of Fe+++ in this reaction, although compounds other than ascorbic acid which contain an acidic OH group may participate.
The physiological significance of the dependence of plasma iron incorporation into ferritin on aerobic metabolism in the liver is discussed.
REFERENCES 1. GREEN, S., AND MAZUR, A., J. Biol. Chem., 227, 653 (1957). 2. MAZUR, A., GREEN, S., SAHA, A., AND CARLETON, A., J.
Clin. Invest., 37, 1809 (1958). 3. NISHIDA, G., AND LABBE, R. F., Biochim. et Biophys. Acta, 31,
519 (1959). 4. LABBE, R. F., Biochim. et Biophys. Acta, 31, 589 (1959). 5. JANDL, J. H., INMAN, J. K., SIMMONS, R. L., AND ALLEN, D.
W., J. C&n. Invest., 38, 161 (1959). 6. SALTMAN, P. FISKIN, R. D., BELLINGER, S. B., AND ALEX, T.,
J. Biol. Chem., 220, 751 (1956). 7. LOEWUS, M. W., AND FINEBERG, R. A., Biochim. et Biophys.
Acta, 26, 441 (1957). 8. MAZUR, A., AND SHORR, E., J. Biol. Chem., 182, 607 (1950). 9. THEORELL, H., BEZNAH, M., BONNICHSEN, R., PAUL, K. G.,
AND AKESON, A., Acta Chem. Stand., 6,445 (1951).
by guest on Novem
ber 27, 2018http://w
ww
.jbc.org/D
ownloaded from

March 1960 A. Mum-, S. Green, and A. Carleton 603
10. HAMPTON, J. K., JR., AND KAHN, J. B., JR., Am. J. Physiol., 174, 226 (1953).
11. LOFTFIELD; R. h., AND BONNICHSEN, R., Acta Chem. Stand., 10, 1547 (1956).
12. FINEBERG, R. A., AND GREENBERG, D. M., J. Biol. Chem., 214, 107 (1955).
13. ROTHEN, A., J. Biol. Chem., 163, 679 (1944). 14. MAZUR, A., LITT, I., AND SHORR, E., J. Biol. Chem., 187, 473
(1950). 15. MANN, P. J. G., AND QUASTEL, J. II., Biochem. J., 36,
502 (1941). 16. HANDLER, P., AND KLEIN, J. R., J. Biol. Chem., 143,49 (1942). 17. POTTER, V. R., J. Biol. Chem., 169, 17 (1948). 18. SCHNEIDER, W. C., J. Biol. Chem., 176, 259 (1948). 19. MAZUR, A., AND SHORR, E., J. Biol. Chem., 176, 771 (1948). 20. GRANICH, S., Chem. Revs., 38, 379 (1946). 21. GREEN, S., MAZUR, A., AND SHORR, E., J. Biol. Chem., 220,237
(1956). 22. LOFTFIELD, R. B., AND EIGNER, E. A., J. Biol. Chem., 231,
925 (1958). 23. MAZUR, A., BAEZ, S., AND SHORR, E., J. Biol. Chem.. 213, 147
(1955). . I
24. COHEN, E., SURGENOR, D., AND HUNTER, M., in J. T. EDSALL (Editor), Enzymes and enzyme systems: their state in nature,
Harvard University Press, Cambridge, Massachusetts, 1951, D. 107.
25. CARTWRIGHT, G. E., AND WINTROBE, M. M., J. Clin. Invest., 28, 86 (1949).
26. GITLIN, D., JANEWAY, C. A., AND FARR, L. E., J. C.&in. Invest., 36, 44 (1956).
27. LAURELL, C. B., Pharmacol. Revs., 4, 371 (1952). 28. NEUBERG, C., AND MANDL, I., Arch. Biochem., 23, 499 (1949). 29. SZENT-GYORGI, A., in 0. H., GAEBLER (Editor), Enzymes:
units of biological structure and function, Academic Press, Inc., New York, 1956, p. 395.
30. BOYER, P. D., AND HARRISON, W. H., in W. D., MCELROY AND B. GLASS (Editors), The mechanism of enzyme action, The Johns Hopkins Press, Baltimore, 1954, p. 666.
31. BAMANN, E., FISCHLER, F., AND TRAPYANN, H., Biochem. Z., 326, 413 (1954).
32. PFLEGER, K., AND RUMMEL, W., Naturwissenschaften, 4, 88 (1957).
33. WARBURG, O., AND SAKUMA, S., Arch. ges. Physiol. Pjluger’s, 200, 203 (1923).
34. GERO, E., AND Roux, J. M., Compt. rend. sot. biol., 152, 329 (1958).
35. MOORE, C. V., AND DUBACH, R., Science., 116, 527 (1952). 36. GRANICK, S., J. Biol. Chem., 164, 737 (1946).
by guest on Novem
ber 27, 2018http://w
ww
.jbc.org/D
ownloaded from

Abraham Mazur, Saul Green and A. CarletonMechanism of Plasma Iron Incorporation into Hepatic Ferritin
1960, 235:595-603.J. Biol. Chem.
http://www.jbc.org/content/235/3/595.citation
Access the most updated version of this article at
Alerts:
When a correction for this article is posted•
When this article is cited•
to choose from all of JBC's e-mail alertsClick here
http://www.jbc.org/content/235/3/595.citation.full.html#ref-list-1
This article cites 0 references, 0 of which can be accessed free at
by guest on Novem
ber 27, 2018http://w
ww
.jbc.org/D
ownloaded from