mechanisms of heterogeneous distribution of monoclonal...
TRANSCRIPT

(CANCER RESEARCH 48, 7022-7032, December 15, 1988]
Mechanisms of Heterogeneous Distribution of Monoclonal Antibodies and OtherMacromolecules in Tumors: Significance of Elevated Interstitial Pressure1
Rakesh K. Jain2 and Laurence T. Baxter3
Department of Chemical Engineering, Carnegie Mellon University, Pittsburgh, Pennsylvania 15213-3890
ABSTRACT
The therapeutic efficacy of monoclonal antibodies (MAbs), bound toradionuclides, chemotherapeutic agents, toxins, growth factors, or effector antibodies, depends upon their ability to reach their target in vivo inadequate quantities. Despite the high vascular permeability and interstitial transport coefficients in tumor tissue compared to several normaltissues, MAbs and their fragments do not distribute homogeneously in atumor. Heterogeneity of tumor-associated antigen expression alone cannot explain this maldistribution of MAbs in tumors. We propose that inaddition to the heterogeneous blood perfusion, hindered diffusion in theinterstitium, and extravascular binding of MAbs, elevated interstitialpressure is responsible for the poor penetration of MAbs into tumors.Elevated interstitial pressure principally reduces the driving force forextravasation of fluid and macromolecules in tumors, and also leads toan experimentally verifiable, radially outward convection which opposesthe inward diffusion. We present here mathematical models for transportof fluid and macromolecules in a tumor. To illustrate the significance ofelevated interstitial pressure, these models are used to describe theinterstitial pressure, interstitial fluid velocity, and concentration of non-binding macromolecules as a function of radial position in a uniformlyperfused tumor.
The key result of these models is that the filtration of fluid from bloodvessels in a uniformly perfused tumor is (a) spatially heterogeneous, (A)a result of elevated interstitial pressure, and (c) sufficient to explain theheterogeneous distribution of macromolecules in tumors. Nonuniformblood flow, and extravascular binding would enhance this heterogeneityin the solute distribution considerably. The results of the models alsoagree with the following experimental data: (a) tumor interstitial pressureis low in the periphery and it increases toward the center of the tumor;(A) the radially outward fluid velocity at the tumor periphery predictedby the model is of the same order of magnitude as measured in tissue-isolated tumors; and (c) immediately after bolus injection, the concentration of macromolecules is higher in the periphery than in the center;however, at later time periods the peripheral concentration is lower thanin the center. These results have significant implications not only forMAbs and their fragments, but for other biologically useful macromolecules (e.g., cytokines) produced by genetic engineering for cancer diagnosis and treatment.
INTRODUCTION
Substantial evidence in the literature indicates that MAbs4
and their fragments do not distribute homogeneously and inadequate quantities in tumors. There is a preferential distribution of antibodies in areas closest to blood vessels, and thereare many regions with zero or negligible antibody (1, 2). Thisis surprising since tumors have high vascular permeability andhigh interstitial transport coefficients compared to several normal tissues (3, 4). What factors are responsible for this heterogeneity?
Received 4/8/88: revised 6/14/88. 8/8/88; accepted 9/21/88.The costs of publication of this article were defrayed in part by the payment
of page charges. This article must therefore be hereby marked advertisement inaccordance with 18 U.S.C. Section 1734 solely to indicate this fact.
1Supported by the National Cancer Institute (CA-36902). This work waspresented at the Radiation Research Society Annual Meeting in Philadelphia.April 16-21,1988, and at the Annual Meetings of the Microcirculation Societyand the BiomédicalEngineering Society in Las Vegas, April 30 to May 5, 1988.
2To whom requests for reprints should be addressed.1Recipient of NSF Predoctoral Fellowship.' The abbreviation used is: MAb. monoclonal antibody.
Several investigators have attributed the maldistribution ofantibodies to the well-known heterogeneity of tumor-associatedantigen expression (5, 6). The fact that antibodies and theirfragments do not penetrate and bind significantly to the antigen-rich regions of a tumor (1) suggests that there may be nonim-munological reasons for the inability of MAbs to reach allregions of a tumor.
It is well established that tumor blood supply is highly heterogeneous (7, 8). Therefore, immediately after injection, onewould expect the blood-borne MAbs to accumulate primarilyin the richly perfused regions. Once extravasated, the interstitialmatrix would hinder the diffusion of macromolecules (9-11).The binding of MAbs to the tumor-associated antigens in theextravascular space would further reduce the amount of mobileantibody molecules available for diffusion away from a vessel(12, 13). In addition to the heterogeneous blood supply, hindered interstitial diffusion, and rapid extravascular binding, arethere other mechanisms responsible for maldistribution ofMAbs in tumors?
Recently, we proposed another mechanism for this heterogeneity: high interstitial pressure in tumors (3, 4). The elevatedinterstitial pressure primarily reduces the driving force forextravasation of fluid and macromolecules inside tumors, andhence leads to nonuniform filtration of fluid and macromolecules from blood vessels. In addition, existence of pressuregradients from the tumor center to its periphery leads to anexperimentally verifiable, radially outward interstitial convection in tumors. Butler et al. (14) calculated the net fluid lossfrom the tumor periphery (Qip) from the arteriovenous difference in blood hematocrit of tissue-isolated tumors. In fourdifferent mammary carcinomas (2-5 g), average Q¡Fwas foundto be 0.14-0.22 ml/h/g-tissue, which is significantly greaterthan the lymph drainage in most normal tissues (~0.002-0.07ml/h/g). Similar values of Q¡Fhave been found recently inWalker 256 carcinoma in rats (15) and in human tumor xeno-grafts in nude rats (16). The radially outward fluid velocity atthe tumor periphery, UK,can be calculated by dividing the fluidloss by tumor surface area:
UR= pVQ,f!A = pRQir/3 (A)
where, p is the tumor density (g/ml); and A, A, and V are,respectively, the tumor radius, surface area and volume (cm,cm2, cm') (17). For a tumor of 1-cm radius (~4 g) the value of
URis -0.1-0.2 Mm/s! A macromolecule must have an inwarddiffusional velocity greater than the outward convective velocityto effectively penetrate into a tumor.
Despite the overwhelming evidence and importance of interstitial convection in tumors, there are no direct measurementsof the magnitude and direction of convective velocity of interstitial fluid or antibodies within a tumor. In addition, there areno mathematical models in the literature which account for theelevated and nonuniform interstitial pressure in tumors in vivo(18). Dedrick and «workers (19, 20) have developed physiologically based pharmacokinetic models for the uptake of meth-otrexate in a canine lymphosarcoma and in murine Lewis lungcarcinoma. Jain et al. (21) have also developed a similar model
7022
on June 25, 2018. © 1988 American Association for Cancer Research. cancerres.aacrjournals.org Downloaded from

HETEROGENEOUS DISTRIBUTION OF ANTIBODIES
for the transport of methotrexate in Walker 256 carcinoma andhepatoma 5123 in rats. These compartmental models do notprovide the spatial distribution of molecules in tumors. Jainand Wei (22) developed a distributed parameter model todescribe the radial distribution of methotrexate and sulfur-mustard in solid tumors. Again their model is valid only forsmall molecules (M, < 1000) for which convection is negligiblecompared to diffusion.
We report here two interrelated, physiologically based, distributed parameter models: (a) a fluid transport model to describe the interstitial pressure (/>,-)and interstitial fluid velocity(M,-)as a function of radial position in a tumor; and (/>)a solute
transport model to describe the interstitial concentration (C¡)of macromolecules as a function of radial position and timepostinjection. To elucidate the role of elevated /»,-,we considerhere the most simple case of a spherical tumor with uniformblood perfusion and no binding between antibody and antigen.As explained later (see "Discussion") nonuniform blood supply
and binding would further increase the heterogeneity of antibody distribution in a tumor. Although the solute transportmodel is applicable for any macromolecule, we show heresimulations for IgG (M, ~ 150,000) and two of its fragments,F(ab')2, (M, ~ 100,000) and Fab (M, ~ 50,000). Two modes of
injection are analyzed: continuous infusion which leads to aconstant plasma concentration ((',,"), and a bolus injection
which leads to a Cf which decays exponentially with time.Extensive simulations and sensitivity analysis are carried out toidentify key parameters responsible for heterogeneous distributions of MAbs and other macromolecules in tumors.
of a vessel into the interstitial space, and most of it is reabsorbed at thevenous end. The net fluid loss from a vessel, J, (ml/s) is given byStarling's Law (4):
Jv —¿�LpS\p, —¿�p¡—¿�aT(tr, —¿�7T,)] (B)
where Lp is the hydraulic conductivity of the microvascular wall (cm/mmHg-s); S is the exchange area of the vessel (cm2); pv and p¡are the
vascular and interstitial pressures (mmHg); irvand JT,are the osmoticpressures of the plasma and interstitial fluid (mmHg); and ar is theaverage osmotic reflection coefficient for the plasma proteins, definedas the fraction of solute filtered through a membrane under conditionsof zero concentration difference and high filtration rate.
In normal tissue this residual fluid, J,. is reabsorbed by the lymphatics. In tumors, due to the absence of anatomically well-defined, functioning lymphatics (23), the net fluid leaked from a vessel can only goin two directions: towards the center of the tumor or towards theperiphery. The inward flow would lead to an increase in the interstitialpressure until p¡becomes equal to an effective pressure, pf\- p, -aT(*, —¿�TÕ)],at which point the fluid leakage would stop (J, = 0). The
outwardly flowing fluid would ooze into the surrounding normal tissuewhere it may be reabsorbed by the normal, functioning lymphatics. Thevelocity of this fluid in the interstitium, «„can be related to theinterstitial pressure gradient, dp¡/dr,by Darcy's Law for flow through
a porous medium (24):
i = -Kdp¡/dr (Q
where K is the hydraulic conductivity of the interstitium (cm2/mmHg-s), and r is the radial position in the tumor (cm). Balancing thefluid filtered from the vessels with the fluid moving towards the tumorperiphery yields the following conservation of mass equation:
DEVELOPMENT OF MODELS
Shown in Fig. 1 is the schematic of a spherical tumor embedded innormal tissue. Consider a vessel in the tumor's periphery; according toStarling's hypothesis, some of the plasma filters from the arterial end
Fig. 1. Schematic of tumor cross-section showing a blood vessel in the periphery and another in the interior of the tumor. Note that the interstitial pressure ishigher in the center than in the periphery of the tumor. This pressure gradientleads to a radially outward convection ( -»-)which opposes the radially inwarddimisión ( ^». Most importantly, the lower pressure in the periphery leads to ahigher filtration of fluid and macromolecules (—>)from the blood vessels in theperiphery.
drJ,V
(D)
Substituting Equations B and C into D leads to:
The boundary conditions are: (a) no fluid flux or pressure gradientat center of the tumor, and (b) continuous pressure and flow rate acrossthe interface between the tumor and the surrounding normal tissue.These equations were solved analytically to obtain profiles for thenormalized pressure [p = (p¡- p*>)/(p, - p,)} and normalized velocity[n,(r)/«A](25).
The extravasation of solute molecules, J, (g/s), from an exchangevessel occurs by convection and diffusion. Convection is proportionalto the rate of fluid leakage, J„from the vessel, and diffusion isproportional to the difference between the plasma concentration, Cp,and the interstitial concentration, C¡(4):
J, = J,(\- a)Cp + PS(CP - C,)Pe,l(ep'- -
[convection] [diffusion](F)
where P is the vascular permeability coefficient (cm/s); Pev is thetransvascular Peclet number, the ratio of convection to diffusion[Jv(\ —¿�a)IPS]; and, a is the osmotic reflection coefficient for the M Ab.
For a macromolecule, convection is the dominant mode of transport innormal tissues. It should also be noted that pressure and concentrationdistributions around individual blood vessels are not considered in thesemodels, but the fluxes given by Equations B and F are used as distributed source terms, rather than local sources due to individual vessels.
Once a molecule has extravasated its movement in the interstitiumalso occurs by convection and diffusion. Convection is proportional tothe interstitial fluid velocity, u¡,and diffusion is proportional to theconcentration gradient in the interstitium, dd/dr. Mathematically the
7023
on June 25, 2018. © 1988 American Association for Cancer Research. cancerres.aacrjournals.org Downloaded from

HETEROGENEOUS DISTRIBUTION OF ANTIBODIES
Table 1 Baseline parameter values used in simulations
ParameterLf(cm/mmHg-s)A'
(cm:/mmHg-s)S/V(cm-')P.
(mmHg)-,(rumili;!T,
(mmHg)f"rTfO(KT'cmVs)/».„(Ifl-'cm/s)r,(h)TissueNormalTumorNormalTumorNormalTumorBothBothNormalTumorBothNormalTumorBothTumor*Tumor*PlasmaBaseline
value0.36x10-'°-*2.8
x 10-'*•'8.53xIO'54.13xIO"'70°20015.6'2V10150.95.
0.90.0.50^0.91'0.82*1.01.3,
2.0,4.4/-l'J5.73.17.3,1497-1103.8,6.1. 3.07-*Ref.Table
2' in Reference4Fig.
6' in Reference24Table
6' in Reference4Table
6Cin Reference84442744249,
101027
" Value representative of hindlimb.4 Determined from whole organ studies.' Refers to table/figure in the article cited.'' Assumed 7.8 times that for normal tissue, based on the ratio of tumor to
normal vascular permeability as measured by Gerlowski and Jain (10).' Assumed equal in both tissue types.f Values for IgG, F(ab'):, and Fab, respectively." Value for albumin in subcutaneous tissue (for normal tissue) and in mesentery
(for tumor).* When convection dominates the interstitial transport in the tumor periphery
(uiR/D » 1). the solution for C, in the tumor is not sensitive to the values of Pand D chosen for the surrounding tissue.
' These are effective values which include both convettive and diffusive com
ponents. The permeability coefficients for the fragments were calculated by scalingthe value for IgG by the interstitial diffusion coefficient in normal tissue. Theyagree in magnitude with the values assumed in Ref. 27.1The diffusion coefficients for IgG and fragments in tumor tissue were takenas a factor of 2.5 lower than for dextran (9).'
* The half-lives are 72. 4.2. and 2.0 h for IgG, F(ab')2, and Fab, respectively.
interstitial flux /, (g/cm2-s) is given by (24):
/,=Tfltid - Dd C,/dr
[convection] [diffusion] (G)
where D is the interstitial diffusion coefficient and r/is the retardationfactor (the ratio of the solute velocity to the fluid velocity). Due to thehindrance provided by the interstitial matrix, the value of D in a tissueis less than its value in water (3). Balancing the solute leaving the vessel(Equation F) with the solute moving in the interstitium (Equation G)leads to the following convection-diffusion equation:
ao ,D±2dC¡ l d'7^r[ (H)
This equation was solved using the Galerkin finite-element method(26) subject to the following boundary conditions: the concentrationand flux are continuous across the interface between the tumor andsurrounding normal tissues (25), and there is no interstitial solute fluxin the center. The solution provided C, as a function of radial position,r, and time, t, postinjection.
The key assumptions incorporated in these two models are: (a) thetumor is spherical and radially symmetric; (¿>)the physiological parameters: P, D, S, V, Lf, p„Try,JT„and a, are independent of radial positionwithin the tumor, but p¡,«,,C¡,J,. and J, are spatially dependent; (c)the tumor growth rate is slow compared to transport rates, thereforeall transport coefficients are independent of time; (d) the concentrationof macromolecules in tumor vessels, C„is independent of radial position in the tissue, due to the slow rate of extravasation; (e) the tumorradius is large compared to ¡ntercapillarydistances; (/) there are noanatomically well-defined functional lymphatics in the tumor; and (g)the macromolecule does not bind to any site in the tissue.
All parameters used in these models have been recently compiled byJain (3, 4. 8) for various normal and neoplastic tissues. Values used inthe current work are listed in Table 1 along with their sources. Sensi-
' Clauss and Jain, unpublished results.
tivity analysis is carried out by varying each of the parameters tosimulate the physiological variability and to identify key parametersresponsible for heterogeneous transport.
RESULTS
Interstitial Pressure. Shown in Fig. 2 are interstitial pressureprofiles in tissue-isolated tumors. The shape of the profiles isessentially the same for tumors surrounded by normal tissue,except that the interstitial pressure becomes equal to p. a shortdistance (<20% of tumor radius) into the surrounding tissue(25). To generalize the results, the radial position has beennormalized with respect to the tumor radius (f = r/R), and theinterstitial pressure has been normalized with respect to theeffective vascular pressure (p = [p¡—¿�p*]/{p, —¿�p«]).As a result
of these normalizations the pressure profiles can be describedin terms of a single parameter, a|= R[(Lf/K)(S/Y)]t/:i\, which
includes the ratio of vascular to interstitial hydraulic conductivities.
Note that similar to the experimental data reported in theliterature, the calculated pressure is highest in the center of atumor and it decreases to the normal value at the tumor's edge
(3). In addition, with increasing a, pressure increases in thecenter until it reaches p... the interstitial pressure required tostop transcapillary filtration, and the profile becomes steepernear the periphery. An increase in a may result from: (a) anincrease in the tumor radius, R: (b) an increase in the exchangesurface area per unit volume, S/V; and (c) an increase in theratio of the hydraulic conductivity of the vessel wall to that ofthe interstitial space, LP/K. If a2 is less than 100, then the
pressure in the center of the tumor becomes a function of a,and hence the tumor radius. Indeed, several investigators sincethe work of Young et al. (29) have shown that tumor pressureincreases with growth initially and then levels off (3). Thesecond and third approaches to increasing a suggest that if fluidleakage from vessels is faster than its removal from the interstitium the tumor pressure will increase. While plausible thereis no conclusive data in the literature to support these approaches. They, however, support the observation of an increasein tumor blood flow following fractionated irradiation (4). It ispossible that fractionated radiation may reduce the cancer celldensity without destroying the tumor vasculature. The reduction in tumor cell density may lead to an increase in the tissuehydraulic conductivity. A', and hence lower the interstitial pres-
0.0
0.0 02 0.4 0.6 08
NORMALIZED RADIUS (r/R)Fig. 2. Normalized interstitial pressure \p - [p, - p.]/[p, - p~\\ as a func
tion of normalized radial position (r R) in a tumor for different values of <>(--R[(¿,//0(5/f)]"2, see text). For tumor parameters given in Table 1, a2 = 1356.Center: r/R = 0, edge: r/R = 1. The pressure profile shown is for an isolatedtumor. The shape of the profile is essentially the same for a tumor surrounded bynormal tissue, and the interstitial pressure becomes equal to p* a short distance(<20% of the tumor radius) into the surrounding tissue.
7024
on June 25, 2018. © 1988 American Association for Cancer Research. cancerres.aacrjournals.org Downloaded from

HETEROGENEOUS DISTRIBUTION OF ANTIBODIES
sure, which in turn may restart blood flow in previously collapsed vessels (8). If this hypothesis is indeed true, then fractionated radiation would be useful in lowering tumor pressureand increasing MAb uptake.
The most important conclusion to be drawn from thesecalculations (Fig. 2) is that as p¡tends to pe in the center of thetumor, the extravasation of fluid and macromolecules decreasesdrastically according to Equations B and F. However, at theperiphery of the tumor, p¡is near normal tissue pressure, andthus extravasation is highest there.
Interstitial Fluid Velocity. Shown in Fig. 3 are interstitialfluid velocity profiles in tumors. To generalize the results, theradial position and velocity have been normalized with respectto the tumor radius (r = r/R), and the velocity at the periphery[u,-(r)/Uff], respectively. Similar to the pressure profiles, thevelocity profiles can be characterized by the parameter a.
As required by Darcy's Law (Equation C) flat pressure pro
files lead to low velocities in the center of a tumor, and steeppressure profiles in the periphery lead to large velocities there.Also, as a increases the velocity profiles become steeper in theperiphery. Since «,is nearly zero in the center of the tumor,interstitial convection is only important in the periphery.
Using the parameter values given in Table 1, the total fluidloss predicted by this model (= 4ir/Ã2M/¡)is 0.77 ml/h for a 4.2-
g tumor, which agrees well with the mean value of 0.76 ml/hmeasured by Butler et al. (14). A more precise estimate of thefluid loss would require incorporation of heterogeneous perfusion and more accurate values of physiological parameters.
Concentration Profiles. Shown in Figs. 4 through 9 are theinterstitial concentration profiles of IgG, F(ab')2, and Fab in
tumors at various times post-injection. Figures 4-6 are for acontinuous infusion which leads to a constant plasma concentration (Cp = C„°),and Figs. 7-9 are for a bolus injection in
which the plasma concentration decreases exponentially withtime (Cp = Cp0e~"rp). The radial position is normalized with
respect to the tumor radius (r = r/R, =0 at the center, =1 atthe edge), and C, is normalized with respect to Cf°.These
figures illustrate the importance of heterogeneous filtrationfrom blood vessels, which is a function of the interstitial pressure, vascular permeability and molecular weight. An importantparameter in these figures is P/Ptn, which characterizes thefraction of the transcapillary solute flux which is due to diffusion alone for the effective permeability coefficients reported inthe literature. Note that the values of Ptn reported in Table 1include both convective and diffusive components. Although
ohlN
K
NORMALIZED RADIUS (r/R)
Fig. 3. Normalized interstitial velocity [u¡(r)/Uj,]as a function of normalizedradial position (r/R) in a tumor for different values of «|=R\(Lf/K)(S/V)\lli, seetext|. Center: r/R = 0, edge: r/R = 1. The velocity profile shown is for an isolatedtumor, and is similar to that seen for a tumor surrounded by normal tissue. Fora2 = 1356, UK= 0.17 >im/s in an isolated tumor, and 0.016 nm/s in a tumor
surrounded by normal tissue.
o aO
O a.O
oo.O
72.0
720
720
0.2 0.4 0.6 0.8 IO
r/R
Fig. 4. Interstitial concentration (C,) profiles of IgG in a tumor with a radiusof 1 cm, during 72 h of continuous infusion (constant C, = C,°):A, P/P,n = 0: B,P/P,n = 1%; C P/P,a = 10%. The concentration in the interior of the tumorincreases as the diffusive vascular permeability (P) increases. Center: r/R = 0,edge:r/A= 1.
the value of the diffusive component of the transcapillary fluxhas been estimated to be 0-70% for albumin, recent studiessuggest the likely range to be 0-10% (28, 30). The fraction ofthe flux due to diffusion is expected to be even smaller for IgGwhich is a larger molecule than albumin. As a result, in ourcalculations we have used diffusive permeability, P, which is 0,1, and 10% of the effective permeability coefficient, Pcn, foundby Gerlowski and Jain (10). Note that this does not imply that
7025
on June 25, 2018. © 1988 American Association for Cancer Research. cancerres.aacrjournals.org Downloaded from
HETEROGENEOUS DISTRIBUTION OF ANTIBODIES
IO-I
o aO
00 02 04 06 08
720
B
OdO
08
06
04-
0.2-
0.0
\:v -72.0
^24.0
00 02 04 06 08 IO
r/R
00.O
10-
08-
06-
0.4-
0.2-
00
I;X24.0
00 02 04 06 08-Ki.o
1.0
r/R
0.0 0.2 0.4 0.6 0.8
480
480
lO-i
08-
OQ. 0.6HO
'-72.0 ö04
0.2-
0.0
48.0
80
2.0
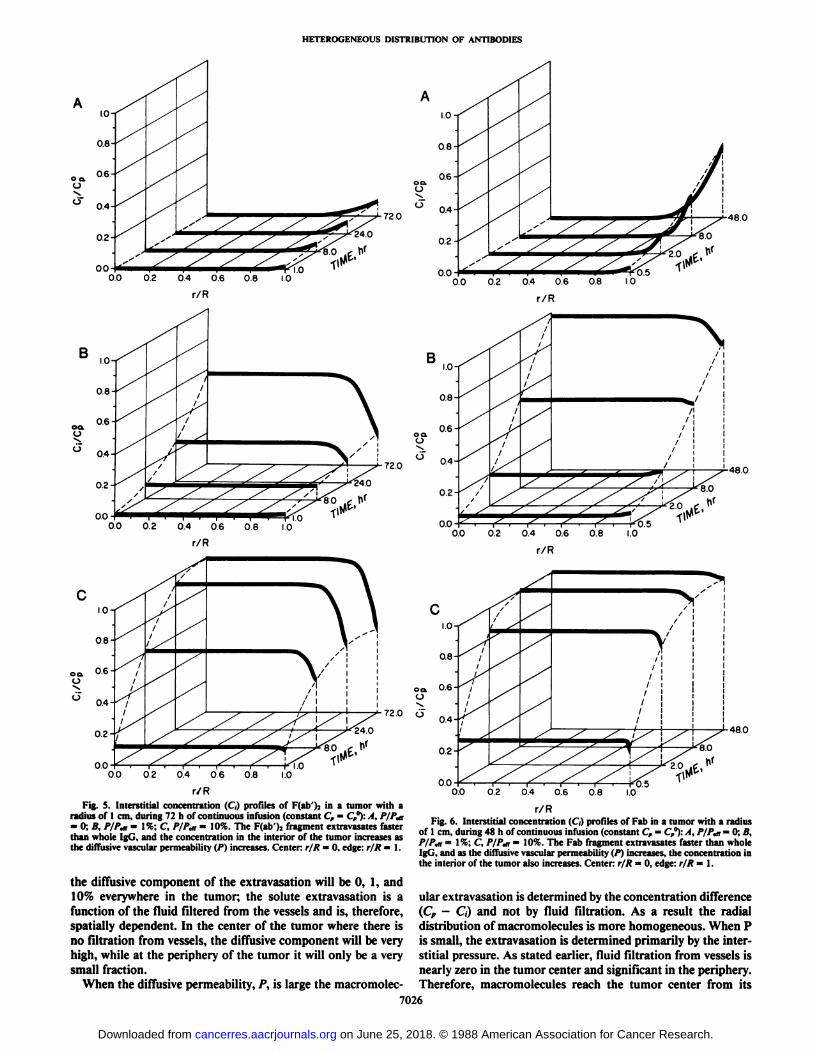
00 02 0.4 06 0.8 1.0Fig. S. Interstitial concentration (C) profiles of F(ab')j in a tumor with a
radius of 1 cm. during 72 h of continuous infusion (constant C, = C,°):/4, P/P,n= 0; A, /V/%iT= 1%; C, /V/V = 10%. The F(ab')2 fragment extravasates faster
than whole IgG, and the concentration in the interior of the tumor increases as(he diffusive vascular permeability (P) increases. Center: r/R = 0. edge: r/R = 1.
r/RFig. 6. Interstitial concentration (C) profiles of Fab in a tumor with a radius
of 1 cm, during 48 h of continuous infusion (constant C, —¿�C,°):A, P/Ptn= 0; B,P/P,n = I %: C P/P,n = 10%. The Fab fragment extravasates faster than wholeIgG, and as the diffusive vascular permeability (P) increases, the concentration inthe interior of the tumor also increases. Center: r/R = 0, edge: r/R = 1.
the diffusive component of the extravasation will be 0, 1, and10% everywhere in the tumor; the solute extravasation is a ular extravasation is determined by the concentration differencefunction of the fluid filtered from the vessels and is, therefore, (C, - C,) and not by fluid filtration. As a result the radial
spatially dependent. In the center of the tumor where there is distribution of macromolecules is more homogeneous. When Pno filtration from vessels, the diffusive component will be very is small, the extravasation is determined primarily by the inter-high, while at the periphery of the tumor it will only be a very stitial pressure. As stated earlier, fluid filtration from vessels issmall fraction. nearly zero in the tumor center and significant in the periphery.
When the diffusive permeability, P, is large the macromolec- Therefore, macromolecules reach the tumor center from its
7026
on June 25, 2018. © 1988 American Association for Cancer Research. cancerres.aacrjournals.org Downloaded from
HETEROGENEOUS DISTRIBUTION OF ANTIBODIES
00 02 0.4 06 0.8 1.0
72 0
720
J-72.0
0.0 0.2 0.4 0.6 0.8 1.0
r/R
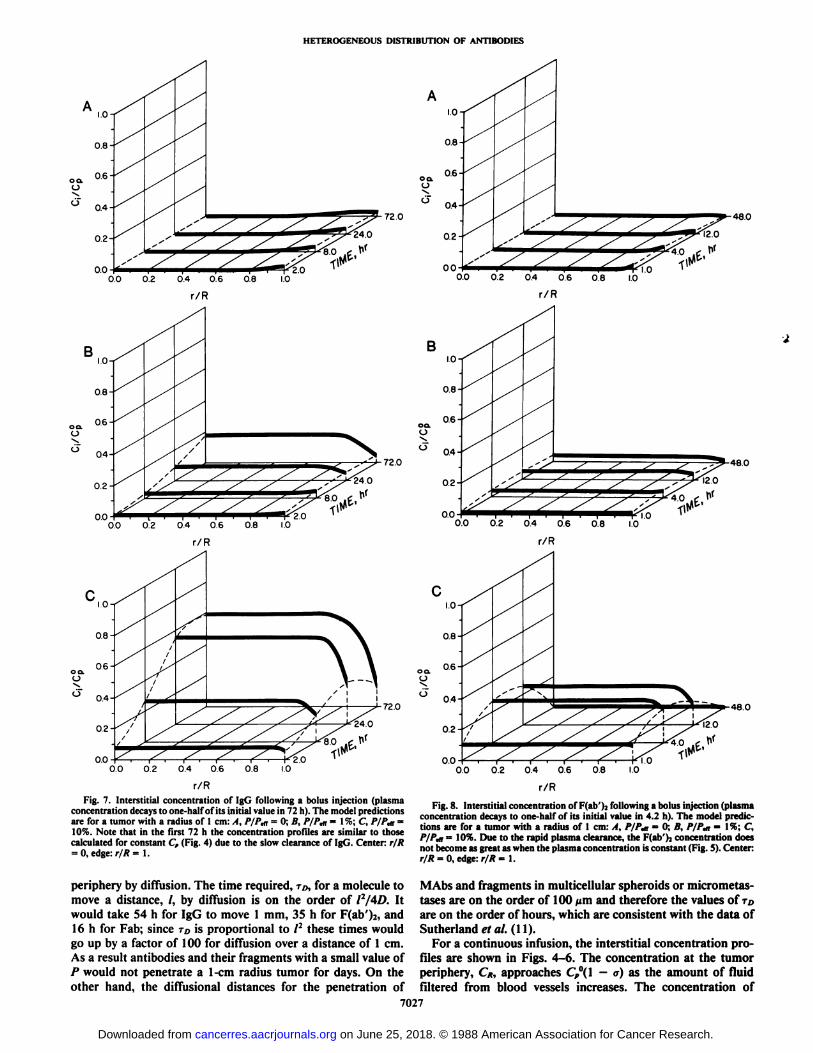
Fig. 7. Interstitial concentration of IgG following a bolus injection (plasmaconcentration decays to one-half of its initial value in 72 h). The model predictionsare for a tumor with a radius of 1 cm: A, P/P,n = 0; B, PIP,«= \%; C, P/P,n =10%. Note that in the first 72 h the concentration profiles are similar to thosecalculated for constant Cp (Fig. 4) due to the slow clearance of IgG. Center: r/R= 0. edge: r/R = 1.
periphery by diffusion. The time required, TD,for a molecule tomove a distance, /, by diffusion is on the order of 12/4D. Itwould take 54 h for IgG to move 1 mm, 35 h for F(ab')2, and16 h for Fab; since TDis proportional to /2 these times would
go up by a factor of 100 for diffusion over a distance of 1 cm.As a result antibodies and their fragments with a small value ofP would not penetrate a 1-cm radius tumor for days. On theother hand, the diffusional distances for the penetration of
0.0 02 0.4 06 08
0.0 0.2 0.4 0.6 08
0.0 0.2 0.4 0.6 08 10
480
480
480
Fig. 8. Interstitial concentration of Hali' ).•following a bolus injection (plasma
concentration decays to one-half of its initial value in 4.2 h). The model predictions are for a tumor with a radius of 1 cm: A, PIP«,= 0; B, PIP«,= \%; C,P/P,n = 10%. Due to the rapid plasma clearance, the F(ab')2 concentration does
not become as great as when the plasma concentration is constant (Fig. 5). Center:r/R = 0, edge: r/R = 1.
MAbs and fragments in multicellular spheroids or micrometas-tases are on the order of 100 urn and therefore the values of TDare on the order of hours, which are consistent with the data ofSutherland et al. (11).
For a continuous infusion, the interstitial concentration profiles are shown in Figs. 4-6. The concentration at the tumorperiphery, C/¡,approaches Cf°(\—¿�a) as the amount of fluid
filtered from blood vessels increases. The concentration of7027
on June 25, 2018. © 1988 American Association for Cancer Research. cancerres.aacrjournals.org Downloaded from
HETEROGENEOUS DISTRIBUTION OF ANTIBODIES
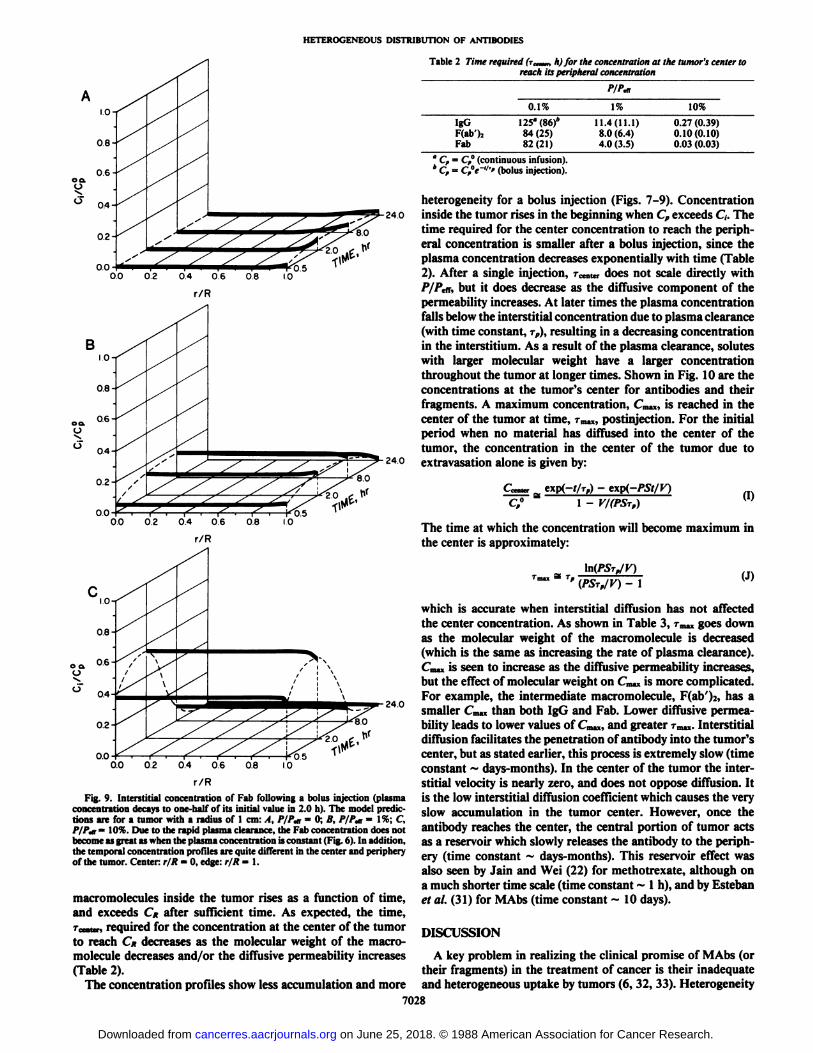
Table 2 Time required (r„,„„h) for the concentration at the tumor's center to
reach its peripheral concentration
P/P,a
240
240
0.0 02 0.4 0.6 0.8 10
r/R
Fig. 9. Interstitial concentration of Fab following a bolus injection (plasmaconcentration decays to one-half of its initial value in 2.0 h). The model predictions are for a tumor with a radius of 1 cm: A, P/Pat = 0; B, P/P«,= 1%; C,/'/',„= 10%. Due to the rapid plasma clearance, the Fab concentration does notbecome as great as when the plasma concentration is constant (Fig. 6). In addition,the temporal concentration profiles are quite different in the center and peripheryof the tumor. Center: r/R = 0. edge: r/R = 1.
macromolecules inside the tumor rises as a function of time,and exceeds CK after sufficient time. As expected, the time,renter, required for the concentration at the center of the tumorto reach CK decreases as the molecular weight of the macro-molecule decreases and/or the diffusive permeability increases(Table 2).
The concentration profiles show less accumulation and more
IgGF(ab')2
Fab0.1%125"
(86)*
84 (25)82(21)1%11.4(11.1)
8.0 (6.4)4.0 (3.5)10%0.27
(0.39)0.10(0.10)0.03 (0.03)
°Cf = C,°(continuous infusion).4 Cf = Cft~"'' (bolus injection).
heterogeneity for a bolus injection (Figs. 7-9). Concentration24o inside the tumor rises in the beginning when Cp exceeds C¡.The
time required for the center concentration to reach the peripheral concentration is smaller after a bolus injection, since theplasma concentration decreases exponentially with time (Table2). After a single injection, r,.,.„„.,does not scale directly withP/P,a, but it does decrease as the diffusive component of thepermeability increases. At later times the plasma concentrationfalls below the interstitial concentration due to plasma clearance(with time constant, TP),resulting in a decreasing concentrationin the interstitium. As a result of the plasma clearance, soluteswith larger molecular weight have a larger concentrationthroughout the tumor at longer times. Shown in Fig. 10 are theconcentrations at the tumor's center for antibodies and their
fragments. A maximum concentration, Cmax,is reached in thecenter of the tumor at time, Tma«,postinjection. For the initialperiod when no material has diffused into the center of thetumor, the concentration in the center of the tumor due toextravasation alone is given by:
_. exp(-t/T,) - e\p(-PSt/Y)1 - V/(PSrp) (I)
The time at which the concentration will become maximum inthe center is approximately:
\n(PS-r„/V)'" (PSrp/V) - l (J)
which is accurate when interstitial diffusion has not affectedthe center concentration. As shown in Table 3, Tmiugoes downas the molecular weight of the macromolecule is decreased(which is the same as increasing the rate of plasma clearance).Cm.xis seen to increase as the diffusive permeability increases,but the effect of molecular weight on Cm.»is more complicated.For example, the intermediate macromolecule, F(ab')2, has a
smaller Cm,x than both IgG and Fab. Lower diffusive permeability leads to lower values of Cm„,and greater Tma)l.Interstitialdiffusion facilitates the penetration of antibody into the tumor's
center, but as stated earlier, this process is extremely slow (timeconstant ~ days-months). In the center of the tumor the interstitial velocity is nearly zero, and does not oppose diffusion. Itis the low interstitial diffusion coefficient which causes the veryslow accumulation in the tumor center. However, once theantibody reaches the center, the central portion of tumor actsas a reservoir which slowly releases the antibody to the periphery (time constant ~ days-months). This reservoir effect wasalso seen by Jain and Wei (22) for methotrexate, although ona much shorter time scale (time constant ~ 1 h), and by Estebanet al. (31) for MAbs (time constant ~ 10 days).
DISCUSSION
A key problem in realizing the clinical promise of MAbs (ortheir fragments) in the treatment of cancer is their inadequateand heterogeneous uptake by tumors (6, 32, 33). Heterogeneity
7028
on June 25, 2018. © 1988 American Association for Cancer Research. cancerres.aacrjournals.org Downloaded from

HETEROGENEOUS DISTRIBUTION OF ANTIBODIES
1.00-
Fig. 10. Interstitial concentration in thecenter of the tumor as a function of time. .I.IgG with continuous infusion; B, IgG afterbolus injection (plasma clearance half-life of72 h); C, F(ab')2 with continuous infusion; D,F(ab')2 after bolus injection (plasma clearancehalf-life of 4.2 h); /., Fab with continuousinfusion; F, Fab after bolus injection (plasmaclearance half-life of 2.0 h). It is seen that thedecaying plasma concentration has a more significant role for the F(ab')2 and Fab fragments.
It is also seen that as the diffusive vascularpermeability (P) increases the center concentration increases more rapidly, but the macro-molecule is also cleared faster out of the tumor.
0.00
1.00
20 40 60 80 100
TIME (hours) TIME(hours)
J.OO-
0.00
20
TIME (hours) TIME (hours)
Table 3 Time required (T^¿,h) to reach maximum concentration (C^J in thetumor's center after bolus injection
P/P«,IgGF(ab')2
Fab0.1I'm.,3403010.7%cm„/c,°0.037
0.00730.029Tm«154
175.01%cwc,°0.23
0.0610.19TO«46
7.01.610%(WC/0.64
0.320.59
of antigen expression alone cannot explain the inability ofMAbs to penetrate the center of a tumor. We propose that themaldistribution of MAbs in tumors results from (a) heterogeneous perfusion; (b) elevated interstitial pressure; (c) slow interstitial diffusion; and (d) rapid and heterogeneous extravas-cular binding. To quantify these mechanisms we have developedtwo interrelated mathematical models: a fluid transport modeland a solute transport model, which provide profiles of pressure,velocity and concentration of macromolecules in the tumorinterstitium. To examine the role of elevated interstitial pressure alone, we have applied these models to a homogeneously
perfused tumor injected with a non-binding macromolecule.The main conclusion of the fluid transport model is that /?,
at the edge of the tumor is essentially equal to the normal tissuepressure and it increases in a monotonie fashion as one movestowards the center of a tumor (Fig. 2). For the physiologicalparameters listed in Table 1, the maximum value of p¡in atumor is equal to the effective vascular pressure, p, (=11.5 mmHg) (Fig. 2). These results are in qualitative agreement with theavailable interstitial pressure data (3). Since the driving forcefor fluid filtration is the difference between p, and p¡(EquationB), the net rate of filtration of fluid from vessels is maximumat the tumor periphery and zero in the center. This nonuniform-ity in fluid filtration from vessels has a profound effect on theextravasation of macromolecules for which convection is thedominant mode of transcapillary exchange. In the extreme caseof a macromolecule with zero diffusive permeability, the mac-romolecular efflux from vessels will parallel the fluid efflux(Equation F), i.e., zero in the center and maximum at theperiphery. In such a case the only way a macromolecule canreach the tumor's center from its periphery is by interstitial
7029
on June 25, 2018. © 1988 American Association for Cancer Research. cancerres.aacrjournals.org Downloaded from

HETEROGENEOUS DISTRIBUTION OF ANTIBODIES
diffusion. As shown earlier, this process is extremely slow formacromolecules. This shows again the importance of heterogeneous filtration; if the molecules could get out of the vascu-lature throughout the tumor then they would only have to travelthe intercapillary distance which is on the order of 100 /¿m(which would take 0.54 h for IgG, 0.35 h for F(ab')2, and 0.16
h for Fab) instead of up to 1 cm to achieve a uniform concentration throughout the tumor. A similar problem is encounteredwith transport from the peritoneal cavity to the surroundingtissues (34). Only in micrometastases would diffusion be rapidenough to diminish heterogeneity.
There are a number of factors which determine the centralversus peripheral accumulation of macromolecules within atumor. For a bolus injection, there are temporal phases ofaccumulation which depend on whether Cp is high or lowcompared to C¡,i.e., whether the macromolecule is extravasat-ing from the blood vessels or being cleared from the tissue. Atearly times peripheral accumulation of macromolecules in atumor is favored by large interstitial convection, high centralinterstitial pressure, and a large vascular to interstitial hydraulicconductivity ratio. A more uniform or central accumulation ofmacromolecule is favored by a large interstitial diffusion coefficient, or a large diffusive vascular permeability.
However at longer time periods, when the MAbs are beingwashed out of the tissue, many of the same factors that favoredperipheral accumulation will now begin to favor central accumulation. It should be noted that this is not due to an increasein the central concentration, but rather a rapid decrease in theperipheral concentration. This reservoir effect is due to the factthat fluid with a low MAb concentration is filtered out of thevessels, and driven by convection into the surrounding normaltissue (or a body cavity) where functioning lymphatics (andblood vessels to some extent) remove the macromolecules.These competing mechanisms determine whether central orperipheral accumulation occurs in a given tumor.
Limitations of Current Models. While the fluid and solutetransport models presented here are simple they provide considerable insight into the mechanisms of heterogeneous distribution of macromolecules in tumors. Due to the assumption ofa homogeneous tumor with no binding the models underestimate the heterogeneity in the distribution of antibodies; whereasthe assumption of no anatomically well-defined functioninglymphatics in the tumor and an abrupt change in the physiological parameters for the surrounding normal tissue overestimatethe gradients at the tumor periphery. In what follows we willdiscuss these assumptions and their implications.
Although the interstitial pressure profiles and interstitial fluidvelocity at the tumor's edge, UK,predicted by the fluid transport
model are in qualitative agreement with the available data, thepredicted pressure gradients in the tumor periphery are extremely steep. This steepness is a result of not incorporatingany functioning lymphatics in the tumor in the model presentedhere. Although the tumors do not themselves produce lymphatics, it is likely that during invasion into a normal tissue, atumor incorporates the functioning lymphatics of the host tissuein its periphery. If present, these lymphatics would facilitatethe removal of interstitial fluid from the outer rim of a tumorand would make the pressure profile less steep. This wouldreduce the value of UK.
The second limitation of the current analysis is the assumption of a homogeneous tumor. In reality a tumor may havemultiple nodes and each node may contain several zones: ne-crotic, seminecrotic, viable, and advancing (8). This heterogeneity would make the physiological parameters incorporated in
these models spatially dependent. In particular, the pressuresp„7r„,and T, may not be constant. If so, the pressure profilewould be affected, and a less steep profile might result. But,most importantly, lack of functional blood vessels in the ne-crotic zones of a tumor would preclude delivery of blood-borneantibodies and lead to a more heterogeneous distribution ofantibodies than shown by the current analysis.
The pressure and concentration distributions around individual blood vessels are not considered in these models, but thefluxes from blood vessels are used as distributed source terms,rather than local sources. This may lead to an overestimationof the fluid filtration because capillary-capillary interactionwould lead to pressure gradients around the blood vessels whichwould oppose filtration (35). The concentration profiles aroundindividual vessels is expected to be unimportant when theinterstitial concentration is much smaller than the plasma concentration. It is also unimportant when the transcapillary soluteflux is primarily due to convection, such as at the periphery ofthe tumor.
Finally, the assumption of no binding leads to a C¡/CP°ratioless than or equal to one. If the antibody binds to a tumor-associated antigen, the concentration profile would be evenmore heterogeneous than when binding is ignored. This is dueto the fact that more MAb is available for binding at theperiphery (i.e., in the richly perfused region with low interstitialpressure) and less inside (i.e., in the poorly perfused region withhigh interstitial pressure). In addition, if the solute is bound atthe periphery of the tumor, it is not free to be washed out bythe large convective flux present at the tumor edge. Significantbinding in the low pressure regions of a tumor may lead to veryhigh MAb concentration averaged over the entire tumor(d/Cf° > 1) while the local concentration of MAb may beextremely small inside a tumor (C,/Q0 «sc1). From a therapeutic
point of view, the quantity of interest is the local concentrationof MAb and not the average tissue to plasma ratio (Uj/Cp°)as
commonly reported.The physiological models presented here are currently being
extended to include lymphatic drainage in the tumor periphery,heterogeneous perfusion, and extravascular binding in order toevaluate the relative contribution of each of these mechanismsin determining the heterogeneous distribution of antibodies (25,36).
Implications for Cancer Diagnosis and Therapy. Given theresults of these models and their limitations, how can thepenetration of nonbinding macromolecules be increased in auniformly perfused tumor? The most obvious approach is toincrease the fluid filtration from vessels inside the tumor. Thiswould require increasing the effective vascular pressure and/orlowering the tumor interstitial pressure. The latter may beachieved by: (a) increasing the hydraulic conductivity, K, of thetumor interstitium; (b) lowering the hydraulic conductivity, Lp,of the tumor vessels; (c) decreasing the exchange surface areaper unit volume (S/V) of tumor vessels; or (d) decreasing thetumor radius. Of all these possibilities, lowering K seems mostfeasible. A' is a function of the cell density and glycosaminogly-
can content (24). Any physical (e.g., radiation) or chemical (e.g.,proteases) agent which can reduce the cell density or glycosa-minoglycan content without destroying the vasculature maylead to higher filtration. Filtration may also be increased usingvasoactive drugs which increase the microvascular pressure, pv.Unfortunately, the increase in pr is short-lived.
An alternate approach to increasing the penetration of MAbsis to increase the diffusive component of the vascular permeability. Since the diffusive transcapillary flux depends on
7030
on June 25, 2018. © 1988 American Association for Cancer Research. cancerres.aacrjournals.org Downloaded from

HETEROGENEOUS DISTRIBUTION OF ANTIBODIES
(Cp —¿�Ci) and not on the fluid filtration, C, will continue toincrease and approach Cf. The concentration at the tumor'sedge approaches (1 —¿�a)Cp when there is a significant fluid flux
coming out of the vasculature at the periphery. The rate atwhich d will approach Cf inside the tumor will depend uponthe magnitude of the diffusive component of the transcapillaryflux. This is borne out clearly in Figs. 4-6 (for continuousinfusion). Increasing P/Ptn from 0.1 to 10% reduces the timerequired for the center concentration to equal the peripheralconcentration by two orders of magnitude (Table 2). Loweringthe molecular weight of macromolecules also has the sameeffect when Cp is constant.
In the case of a bolus injection the advantage of betterpenetration which the MAb fragments have is offset by theirrapid plasma clearance. This point is borne out in Figs. 7-9.Increasing P/Pen also leads to a faster washout of nonbindingantibodies or their fragments from a homogeneously perfusedtumor. These conclusions are in general agreement with thepublished data on nonspecific antibodies (37).
The significant aspects of this work for clinical use of macromolecules may be summarized as follows: (a) the majortherapeutic deficiency of inadequate and nonuniform tumoruptake of monoclonal antibodies may be explained by transportconsiderations alone without invoking antigenic heterogeneity;(b) a number of situations do exist where achieving uniformdistribution is less of a problem, e.g., in very small tumors(micrometastases), due to short diffusion lengths, or in cancerdetection, where uniform distribution is not required; (c) whenthe limiting factors for transport are known, specific measuresmay be taken to circumvent difficulties, e.g., the use of vascularpermeability modulating agents when microvascular permeability is too low (38), or radiation therapy when the low hydraulic conductivity is dominant; (d) there are large effects ofthe molecular weight on the total accumulation and the distribution within the tumor, which are different for bolus versuscontinuous injection (Figs. 4-9); and (e) from a therapeuticpoint of view, the quantity of interest is the local concentrationof MAb and not the average tissue to plasma ratio as commonlyreported. These results have significant implications not onlyfor MAbs and their fragments, but for other biologically usefulmacromolecules (e.g., cytokines).
ACKNOWLEDGMENTS
We would like to thank Drs. R. L. Dedrick, P. M. Cullino, and B.Rippe for their helpful comments on this manuscript.
APPENDIX
Cidc„
DI,J„J,
K
L,
P
P,if
Interstitial concentration, g/mlAverage interstitial concentration of whole tumor, g/mlPlasma concentration, g/mlPlasma concentration at t = 0, g/mlInterstitial concentration at outer edge of tumor, r = R, g/mlMaximum interstitial concentration at center of tumor, ;•= <),
g/mlInterstitial diffusion coefficient for tumor, cur"'sInterstitial solute flux, g/cm2-s
Transcapillary solute flux (g/s) and fluid flux (ml/s), respectivelyHydraulic conductivity of tumor interstitium (Darcy's Lawconstant), cm2/mmHg-sHydraulic conductivity coefficient of vascular wall, cm/mmHg-sDiffusivevascular permeability coefficient, cm/sEffectivevascular permeability coefficient, cm/s
Pe, =p¡ =p, =p, =/>. -p -QIF =r/ =
r =r =R =S/V =Õ =«, =UK =a =
iti, it, —¿�
a =<ST =
Peclet number for vascular wall |=7»(1 - a)/PS\Interstitial fluid pressure, mmHgEffectivevascular pressure [—p,—¿�OT(TT,—¿�ir¡)],mmHgMean vascular pressure in exchange vessels, mmHgInterstitial fluid pressure in surrounding normal tissue, mmHgDimensionless interstitial pressure [=(p¡—¿�p*)/(p, —¿�p,)]Rate of fluid loss from tumor, ml/h/g-tissueRetardation factor, the ratio of the solute velocity to theinterstitial fluid velocityDistance from center of tumor, cmDimensionless radial position (—r/R)Outer radius of tumor, cmSurface area per unit volume for transcapillary exchange, cm"1
time, hRadial interstitial fluid velocity, cm/sValue of Uiat outer edge, r = R, cm/sDimensionless ratio of hydraulic conductivities (=R[(LP/K)
Colloid osmotic pressure (mmHg) of interstitial fluid, andplasma respectivelyOsmotic reflection coefficient for MAb. F(ab'>2.or Fab
Averageosmotic reflection coefficient for plasma proteinsTime constant for interstitial diffusion, hTime constant for plasma clearance, hTime required for center concentration to equal C«Time to reach maximum central concentration (Cm.,)
REFERENCES
1. Jones, P. k.. Gallagher, B. M., and Sands, H. Autoradiographic analysis ofmonoclonal antibody distribution in human colon and breast tumor xeno-grafts. Cancer Imm mini. Immunother.. 22: 139-143, 1986.
2. Sands, H., Jones, P. L., Shah, S. A.. PÃame,D.. Vessella. R. L.. and Gallagher,B. M. Correlation of vascular permeability and blood flow with monoclonalantibody uptake by human clouser and renal cell xenografts. Cancer Res.,48: 188-193, 1988.
3. Jain, R. K. Transport of molecules in the tumor interstitium: a review. CancerRes., 47: 3039-3051, 1987.
4. Jain, R. K. Transport of molecules across tumor vasculature. Cancer Metastasis Rev.. 6: 559-594, 1987.
5. Epenetos, A. A., Snook, D., Durbin, H., Johnson, P. M., and Taylor-Papadimitriou, J. Limitations of radiolabeled monoclonal antibodies forlocalization of human neoplasms. Cancer Res., 46: 3183-3191, 1986.
6. Dykes, P. W.. Bradwell, A. R., Chapman, C. E., and Vaughan, A. T. M.Radioimmunotherapy in cancer: clinical studies and limiting factors. CancerTreat. Rev., 14: 87-106, 1987.
7. Jain, R. K., and Ward-Hartley, K. Tumor blood flow—characterization,modifications, and role in hyperthermia. IEEE Transact, sonics ultrasonics,SU-3I: 504-526, 1984.
8. Jain, R. K. Determination of tumor blood flow: a review. Cancer Res., 48:2641-2658, 1988.
9. Nugent, L. J.. and Jain, R. K. Extravascular diffusion in normal and neo-plastic tissues. Cancer Res., 44: 238-244, 1984.
10. Gerlowski, L. E., and Jain, R. K. Microvascular permeability of normal andneoplastic tissues. Microvasc. Res., 31: 288-305, 1986.
11. Sutherland, R., Buchegger, F., Schreyer, M., Vacca, A., and Mach, J. P.Penetration and binding of radiolabelled anti-carcinoembryonic antigenmonoclonal antibodies and their antigen binding fragments in human colonmulticellular tumor spheroids. Cancer Res., 47: 1627-1633, 1987.
12. Astarita, G. Mass Transfer with Chemical Reactions. Amsterdam: Elsevier,1967.
13. Weinstein, J. N., Eger, R. R., Covell, D. G., Black, C. D. V., Mulshine, J.,Carrasquillo, J. A., Larson, S. M.. and Keenan, A. M. The pharmacology ofmonoclonal antibodies. Ann. NY Acad. Sci., 507; 199-210, 1987.
14. Butler, T. P., Grantham, F. H., and Cullino, P. M. Bulk transfer of fluid inthe interstitial compartment of mammary tumors. Cancer Res., 35: 3084-3088, 1975.
15. Sevick, E. M., and Jain, R. K. Blood flow and efferent blood pH of tissue-isolated Walker 256 carcinoma during hyperglycemia. Cancer Res., 48:1201 -1207, 1988.
16. Vaupel, P.. Fortmeyer, H. P., Runkel. S., and Kallinowski, F. Blood flow,oxygen consumption, and tissue oxygénationin human breast cancer xenografts in nude rats. Cancer Res., 47: 3496-3503, 1987.
17. Jain. R. K. Transvascular and interstitial transport in tumors. In: S. Chien(ed.). Vascular Endothelium in Health and Disease, pp. 215-220. Amsterdam: Elsevier. 1988.
18. Nakagawa, H., Groothius, D. R., Owens. E. S.. Fenstermacher, J. D., Patlak,C. S., and Blasberg, R. G. Dexamethasone effects on ['"Ijalbumin distribution in experimental RG-2 gliomas and adjacent brain. J. Cerebral BloodFlow and Metabolism, 7: 687-701, 1987.
7031
on June 25, 2018. © 1988 American Association for Cancer Research. cancerres.aacrjournals.org Downloaded from

HETEROGENEOUS DISTRIBUTION OF ANTIBODIES
19. Lutz. R. J., Dedrick, R. L., Straw, J. A., Hart, M. M., Klubes, P.. andZaharko. D. S. The kinetics of methotrexate distribution in spontaneouscanine lymphosarcoma. J. Pharmacokinet. Biopharm., 3: 77-97, 1975.
20. Yang, K. H., Fung, W. P., Lutz, R. J., Dedrick. R. L., and Zaharko, D. S.In vivo methotrexate transport in murine Lewis lung tumor. J. Pharm. Sci.,6«:941-945. 1979.
21. Jain. R. K., Wei. J., and Cullino. P. M. Pharmacokinetics of methotrexatein solid tumors. J. Pharmacokinet. Biopharm.. 7: 181-194, 1979.
22. Jain, R. K., and Wei, J. Dynamics of drug transport in solid tumors:distributed parameter model. J. Bioengineering. I: 313-330, 1977.
23. Cullino. P. M. Extracellular compartments of solid tumors, ¡n:F. F. Becker(ed.). Cancer, pp. 327-354. New York: Plenum Publishing Corporation,1975.
24. Swabb, E. A., Wei, J.. and Cullino, P. M. Diffusion and convection in normaland neoplastic tissues. Cancer Res., 34: 2814-2822, 1974.
25. Baxter, L. T.. and Jain, R. K. Transport of fluid and macromolecules intumors. I. Role of interstitial pressure and convection. Microvasc. Res., inpress.
26. Becker, E. B., Carey, G. F., and Oden, J. T. Finite Elements: An Introduction,Volume 1. Eaglewood Cliffs, NJ: Prentice-Hall, Inc., 1981.
27. Covell, p. G.. Barbet, J.. Holton, O. D., Black, C. D. V., Parker, R. J., andWeinstein, J. N. Pharmacokinetics of monoclonal immunoglobulin Gl,F(ab')2 and Fab' in mice. Cancer Res., 46: 3969-3978, 1986.
28. Curry, F. E. Mechanics and thermodynamics of transcapillary exchange. In:E. M. Renkin and C. C. Michel (eds.). Handbook of Physiology, Section 2.The Cardiovascular System IV, pp. 309-374. Bethesda, MD: AmericanPhysiological Society, 1984.
29. Young, J. S., Lumsden, C. E., and Stalker, A. L. The significance of the"tissue pressure" of normal testicular and of neoplastic (Brown-Pearce car
cinoma) tissue in the rabbit. J. Pathol. Bacteriol., 62: 313-333, 1950.30. Rippe. B., and Haraldsson, B. Fluid and protein fluxes across small and large
pores in the microvasculalure. Application of two-pore equations. ActaPhysiol. Scand., 131: 411-428, 1987.
31. Esteban, J. M.. Schlom, J., Mornex, F., and Colcher. D. Radioimmunother-apy of athymic mice bearing human colon carcinomas with monoclonalantibody B72.3: histologie and autoradiographic study of effects on tumorsand normal organs. Eur. J. Cancer, 23: 643-655. 1987.
32. Sands. H.. and Jones, P. L. Physiology of monoclonal antibody accretion bytumors. In: D. M. Goldenberg (ed.). Cancer Imaging with RadioactiveAntibodies. Boston: Martinus Nijhoff, in press.
33. Dedrick, R. L., and Flessner, M. F. Pharmacokinetic considerations onmonoclonal antibodies. In: M. Mitchell (ed.). Immunity to Cancer II. Proc.of 2nd conference on Immunity to Cancer. Williamsburg, VA. in press.
34. Flessner, M. F., Fenstermacher, J. D., Blasberg, R. G., and Dedrick, R. L.Peritoneal absorption of macromolecules studied by quantitative autoradi-ography. Am. J. Physiol., 248: H26-H32, 1985.
35. Fleischman, G. J., Secomb, T. W., and Gross, J. F. Effect of extravascularpressure gradients on capillary fluid exchange. Mathematical Biosci., fil:145-164, 1986.
36. Jain, R. K. Physiological barriers to delivery of monoclonal antibodies andother macromolecules in tumors. Cancer Res. (suppl.), in press.
37. Blasberg, R. G., Nakagawa. H., Bourdon, M. A., Groothius, D. R., Patlak,C. S., and Bigner, D. D. Regional localization of a glioma-associated antigendefined by monoclonal antibody 8IC6 in vivo: kinetics and implications fordiagnosis and therapy. Cancer Res., 47: 4432-4443, 1987.
38. Baxter. L. T., Jain, R. K., and Svensjö. E. Vascular permeability andinterstitial diffusion of macromolecules in the hamster cheek pouch: effectsof vasoactive drugs. Microvasc. Res., 34: 336-348, 1987.
7032
on June 25, 2018. © 1988 American Association for Cancer Research. cancerres.aacrjournals.org Downloaded from

1988;48:7022-7032. Cancer Res Rakesh K. Jain and Laurence T. Baxter of Elevated Interstitial PressureAntibodies and Other Macromolecules in Tumors: Significance Mechanisms of Heterogeneous Distribution of Monoclonal
Updated version
http://cancerres.aacrjournals.org/content/48/24_Part_1/7022
Access the most recent version of this article at:
E-mail alerts related to this article or journal.Sign up to receive free email-alerts
Subscriptions
Reprints and
To order reprints of this article or to subscribe to the journal, contact the AACR Publications
Permissions
Rightslink site. Click on "Request Permissions" which will take you to the Copyright Clearance Center's (CCC)
.http://cancerres.aacrjournals.org/content/48/24_Part_1/7022To request permission to re-use all or part of this article, use this link
on June 25, 2018. © 1988 American Association for Cancer Research. cancerres.aacrjournals.org Downloaded from