mejoramiento de medio de cultivo para …
TRANSCRIPT

MEJORAMIENTO DE MEDIO DE CULTIVO PARA FERMENTACIONES DE
Bacillus thuringiensis serovar. israelensis
Autor: Rodrigo F. González B.
Firma:
Asesor: Paula Atehortúa Osorio Co-asesor: Astrid Altamar
Firma: Firma:
Asesor: José G. Vélez Cuartas Asesor: Sergio Orduz Peralta
Firma: Firma:
Universidad de los Andes
Facultad de Ingeniería
Departamento de Ingeniería Química
Bogotá D. C., Cundinamarca
Corporación para Investigaciones Biológicas
Unidad de Biotecnología y Control Biológico
Medellín, Antioquia

IQ-2003-2-31
1
TABLA DE CONTENIDO
Pág.
1. INTRODUCCIÓN . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .4
2. RESUMEN . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 7
3. MARCO TEÓRICO . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 9
3.1 Bacillus thuringiensis . . . . . . . . . . . . . . . . . . . . . . . . . . . . 9
3.1.1 Morfología . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 9
3.1.2 Biología de Bacillus thuringiensis . . . . . . . . . . . . . . . . .10
3.1.2.1 Germinación y fase vegetativa. . . . . . . . . . . . . . . . . . 10
3.1.2.2 Fase estacionaria: Esporulación y Formación del cristal
paraesporal: . . . . . . . . . . . . . . . . . . . . . . . . . . . . .11
3.1.3 Acción de las Toxinas . . . . . . . . . . . . . . . . . . . . . . . .12
3.1.4 Producción de Bacillus thuringiensis . . . . . . . . . . . . . . .13
3.1.4.1. Medio de Cultivo y Condiciones Ambientales . . . . . . . . . 13
3.1.4.2. Sistemas de producción de Bacillus thuringiensis . . . . . . 14
3.1.4.3. Productos comerciales a base de Bacillus thuringiensis . . . .15
3.2 REVISIÓN DE LA LITERATURA ACERCA DE MEDIOS DE CULTIVOS
PARA Bacillus thuringiensis . . . . . . . . . . . . . . . . . . . . . . . .17

IQ-2003-2-31
2
4. MATERIALES Y MÉTODOS . . . . . . . . . . . . . . . . . . . . . . . . 22
4.1 DISEÑO EXPERIMENTAL . . . . . . . . . . . . . . . . . . . . . . . . . . 22
4.2 OBTENCIÓN DE SOLUCIÓN DE ESPORAS DE Bacillus thuringiensis
serovar. israelensis. . . . . . . . . . . . . . . . . . . . . . . . . . . . . 23
4.3 PREPARACIÓN DEL PREINÓCULO E INÓCULO DE LA
FERMENTACIÓN . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 24
4.4 PROCESO DE FERMENTACIÓN EN EL REACTOR DE 20 LITROS . . 25
4.5 DETERMINACION DE CONCENTRACIÓN DE PROTEINAS . . . . . . .27
4.5.1 Purificación de proteínas . . . . . . . . . . . . . . . . . . . . . . 27
4.5.2 Determinación de Proteínas Solubles en Medio Alcalino por
medio del Método de Bradford . . . . . . . . . . . . . . . . . . .27
4.6 DETERMINACIÓN DE CONCENTRACIÓN DE BIOMASA . . . . . . . . 28
4.7 DETERMINACIÓN DE TOXICIDAD . . . . . . . . . . . . . . . . . . . . .29
4.7.1 Preparación del Ingrediente Activo por Liofilización . . . . . . 29
4.7.2 Bioensayo . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .29
4.7.3 Análisis Probit . . . . . . . . . . . . . . . . . . . . . . . . . . . . 30
5. RESULTADOS . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 31
5.1 COMPORTAMIENTO DE pH y OXÍGENO DISUELTO EN LAS
FERMENTACIONES . . . . . . . . . . . . . . . . . . . . . . . . . . . . .31
5.2 TIEMPO DE FERMENTACIÓN . . . . . . . . . . . . . . . . . . . . . . . 32
5.3 PRODUCCIÓN DE PROTEÍNA TÓXICA . . . . . . . . . . . . . . . . . . 34
5.4 PRODUCCIÓN DE BIOMASA . . . . . . . . . . . . . . . . . . . . . . . .35

IQ-2003-2-31
3
5.5 TOXICIDAD . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 37
5.6 COSTOS . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 38
6 ANÁLISIS . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .41
6.1 PRODUCCIÓN DE INGREDIENTE ACTIVO PARA BIOPESTICIDAS . .41
6.2 COSTOS DE PRODUCCIÓN . . . . . . . . . . . . . . . . . . . . . . . . 43
7. CONCLUSIONES . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .46
8. ANEXOS . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 47
ANEXO 1. Graficas de Comportamiento de pH y Oxígeno Disuelto . . . . 47
ANEXO 2. Análisis de Varianza del Tiempo de Fermentación . . . . . . . .52
ANEXO 3. Análisis de Varianza de Producción de Proteínas . . . . . . . . 53
ANEXO 4. Análisis de Varianza de Producción de Biomasa . . . . . . . . .54
ANEXO 5. Análisis de Varianza de la Toxicidad . . . . . . . . . . . . . . . . 55
9. BIBLIOGRAFÍA . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .56

IQ-2003-2-31
4
1. INTRODUCCIÓN
El Dengue ha sido un problema de salud pública en todas las zonas tropicales del
planeta. Esta es una patología de alto poder epidémico que en los últimos años se
ha venido acrecentando, principalmente en países en vía de desarrollo. En nuestro
país no ha sido la excepción debido a la alta dispersión del vector de la enfermedad.
En Colombia se han utilizado como métodos de control del vector Aedes aegypti
los pesticidas de origen químico, sin tener en cuenta las implicaciones que tienen
el uso de este tipo de productos, sin utilizar productos alternativos como los
biológicos.
A nivel mundial la utilización de biopesticidas en el control de insectos plaga y
vectores de enfermedades es ampliamente aceptado e implementado por los
países de la Comunidad Económica Europea, así como en Estados Unidos y
Canadá. Entre los pesticidas biológicos más utilizados se encuentran las
formulaciones basados en protoxinas de Bacillus thuringiensis (B. thuringiensis).
Los biopesticidas que contienen B. thuringiensis se han empleado ampliamente
para el control de plagas de insectos en agricultura y vectores de enfermedades

IQ-2003-2-31
5
en salud pública; además, son ambientalmente seguros y efectivos en una
variedad de situaciones (Siegel, 2001).
B. thuringiensis es el pesticida biológico más utilizado debido a su efectividad,
rapidez en el control del insecto plaga, estabilidad durante el almacenaje, toxicidad
específica contra el insecto plaga (Dípteros, Lepidópteros y Coleópteros entre
otros) y ausencia total de toxicidad contra invertebrados no blanco y vertebrados
(Siegel, 2001; Deacon, 1983). En la actualidad, la producción industrial de este
tipo de toxina ya está implementada en diferentes países principalmente de
Europa, Asia y Norte América; en Colombia no existe ninguna empresa que
produzca este tipo de biopesticidas, razón por la cual es necesaria su importación
a altos costos. Por lo anterior es indispensable el desarrollo de tecnología para la
producción de B. thuringiensis serovar. israelensis a nivel local para el control del
mosquito vector del Dengue.
La metodología más usada para su producción es el cultivo discontinuo por su fácil
implementación y por los bajos riesgos de contaminación que éste sistema de
producción implica. El estudio de la optimización en la producción de B.
thuringiensis está encaminado a incrementar los rendimientos de biomasa,
esporas, concentración y toxicidad del ingrediente activo, desarrollando medios de
cultivo económicos y optimizando las condiciones de operación para reducir
costos de producción (Avignone Rossa et al., 1992).

IQ-2003-2-31
6
La aplicación de biopesticidas basados en B. thuringiensis es limitada debido a su
alto costo de producción (Dulmage, 1981). Muchos de estos biopesticidas
comerciales son producidos en medios sintéticos que contienen derivados de
soya, pescado, glucosa, extracto de levadura y otros componentes que mejoren el
rendimiento. El costo de producción de B. thuringiensis depende de varios
factores, sin embargo la materia prima comprende entre un 30-40% de la
producción total (Ejiofor, 1991; Lisansky et al., 1993). Debido a esto es necesario
buscar el mejor medio para disminuir los costos de producción. Siendo uno de los
principales objetivos de estudio en la planta piloto de la Unidad de Biotecnología
y Control Biológico (B&CB) de la Corporación para Investigaciones Biológicas
(CIB) el desarrollo de medios de cultivo para B. thuringiensis económicos, de alto
rendimiento en la producción de cristales, que conserven los niveles de toxicidad
necesarios y que sean de fácil adquisición en la región.
Muchas variables interactúan entre ellas, afectando el proceso global de
fermentación. La relación entre la fuente de carbono y nitrógeno es una de estas
variables, al igual que las concentraciones iniciales de estas fuentes (Farrera et
al., 1998; Yang et al., 2000). De este modo, el objetivo general de este trabajo
consiste en evaluar el efecto de la relación de carbono:nitrógeno y las
concentraciones iniciales de la fuente de nitrógeno y de carbono sobre la
producción de proteínas y biomasa en fermentaciones discontinuas con B.
thuringiensis serovar. israelensis.

IQ-2003-2-31
7
2. RESUMEN
Este trabajo consistió en la evaluación del efecto de las concentraciones iniciales
de la fuente de nitrógeno y de carbono y energía sobre el proceso fermentativo de
B. thuringiensis serovar. israelensis. Se realizó un diseño factorial 22 con punto
medio, evaluando 3 relaciones diferentes de carbono:nitrógeno (3.3, 5.2 y 7) que
corresponden a 5 conjuntos de concentraciones iniciales de estas fuentes. En total
se realizaron 10 fermentaciones en un reactor de tanque agitado de 20 litros con
volumen efectivo de 11 litros en operación discontinua, y control automático de pH
entre 6.5 y 8.5, temperatura en 30°C y formación de espuma. Se cuantificaron las
principales variables de respuesta del proceso, como tiempo de fermentación,
concentración de biomasa, concentración de proteínas y toxicidad contra Aedes
aegypti. Se encontró que a medida que aumenta la relación C:N, el tiempo de
fermentación aumenta, de igual manera, cuando se aumenta la concentración de
las fuentes estudiadas manteniendo la relación C:N constante, es más significativo
el aumento del tiempo de proceso. En cuanto a la toxicidad, los resultados
muestran que ésta aumenta con el aumento de la relación C:N, y este mismo
comportamiento ocurre con la producción de proteínas. El aumento en la

IQ-2003-2-31
8
producción de biomasa se ve favorecido tanto con el aumento de la relación C:N,
como con el aumento de las concentraciones iniciales. Con un análisis de costos
se encontraron dos medios de cultivo en los cuales disminuyen el costo de
producción de biomasa y proteínas con respecto al costo con el medio de cultivo
actualmente utilizado en las fermentaciones en la Unidad de Biotecnología y
Control Biológico de la Corporación para Investigaciones Biológicas.
Adicionalmente con estos dos medios se obtuvo un alto rendimiento y su toxicidad
no fue estadísticamente diferente respecto a la obtenida con el medio base, lo que
muestra que se mantiene la calidad del producto.

IQ-2003-2-31
9
3. MARCO TEÓRICO
3.1 Bacillus thuringiensis
3.1.1 Morfología
Esta es una bacteria con un tamaño de entre 3 y 5 µm de largo y 1,0 a 1,2 µm de
ancho. Pertenece a la familia Bacillaceae y al genero Bacillus. Esta bacteria
pertenece a los bacilos gram-positivos, productores de esporas. Éste a diferencia
de otros bacilos produce cuerpos cristalinos adyacentes a la espora durante la
esporulación algunos de los cuales son tóxicos para ciertas especies de insectos
(Figura 1). Este cristal tiene diferentes formas y tamaños dependiendo de las
proteínas que lo compongan. Los cristales son bipiramidales, cúbicos,
romboidales, esféricos, rectangulares, triangulares o irregulares y su tamaño va
desde 350 nm hasta 2 µm de longitud (Caballero e Iriarte., 2001).

IQ-2003-2-31
10
Figura 1. B. thuringiensis esporulado y cristales tóxicos.
3.1.2 Biología de Bacillus thuringiensis
B. thuringiensis es un microorganismo anaerobio facultativo, quimiorganotrofo y
actividad catalasa positiva. Este puede fermentar glucosa, fructosa, maltosa,
trealosa y ribosa entre otras fuentes de carbono.
3.1.2.1 Germinación y fase vegetativa:
La primera etapa de B. thuringiensis es la germinación de la espora de resistencia,
proceso que tarda unos minutos. Aparece entonces la célula vegetativa que se
multiplica activamente en condiciones aeróbicas aunque es anaerobia facultativa
(Caballero e Iriarte., 2001).

IQ-2003-2-31
11
3.1.2.2 Fase estacionaria: Esporulación y Formación del cristal paraesporal:
La esporulación normalmente está inducida por el empobrecimiento del medio de
cultivo (Figura 2.) y coincide con el cambio de fase de crecimiento exponencial a
fase estacionaria. En la fase estacionaria tiene lugar simultáneamente la
formación de la endoespora de resistencia y la del cristal paraesporal. Para ello
es necesaria la presencia de algunos oligoelementos en el medio de cultivo, por lo
que se puede utilizar caldo nutritivo suplementado con sales de manganeso, o
bien medios pobres inductores de la esporulación (Stewart et al., 1981).
Figura 2. Esporulación y formación de cristal tóxico.
Simultáneamente a la formación de la espora, desde el momento del
englobamiento de la preespora hasta su maduración, tiene lugar la síntesis de
varios cristales paraesporales que pueden representar entre 20 y 30% del peso
seco del esporangio (Bulla et al., 1980).

IQ-2003-2-31
12
3.1.3 Acción de las Toxinas
B. thuringiensis es una de las bacterias que ha desarrollado factores de virulencia
que permiten infectar diferentes organismos, entre las que se encuentran las
fosfolipasas, proteasas, quitinasas, alfa exotoxinas o exotoxinas termolabiles, que
son probablemente lectinanas tipo C, las alfa exotoxinas, las cuales son toxinas
que funcionan como análogos de ATP (Snepf y Whiteley, 1985; Levinson 1990),
las proteínas VIP, que son proteínas insecticidas que se producen en la fase
vegetativa del crecimiento y las δ endotoxinas, que son proteínas que se
acumulan en la fase de esporulación formando cristales paraesporales y que
tienen muy diferentes especificidades. Se han descrito δ endotoxinas activas
frente a insectos, ácaros y otros invertebrados (Feitelson, 1993) .
Existen dos tipos de endotoxinas: las proteínas Cry y las proteínas Cyt. Hasta la
fecha se han clonado y secuenciado 166 genes Cry diferentes y 16 genes Cyt
diferentes. Desde el punto de vista del uso de las proteínas de B. thuringiensis
como materia activa de bioinsecticidas, las proteínas Cry son de un gran interés
por su elevada toxicidad y su relativa especificidad, ya que, en general, su
espectro de huéspedes lo constituyen un reducido número de especies de
insectos (Hofte y Whiteley, 1989). En cambio, las proteínas Cyt muestran una
mejor toxicidad y mayor inespecificidad, siendo incluso capaces de lisar in vitro un
amplio rango de tipos celulares, incluidas las células de mamíferos.

IQ-2003-2-31
13
Las proteínas son ingeridas por la larva del insecto blanco. Tras la ingestión, las
toxinas son disueltas debido al pH alcalino de los jugos del intestino de la larva.
Una vez las protoxinas han sido activadas, se unen específicamente a receptores
de membrana de las células epiteliales del intestino medio de los insectos,
produciendo un choque osmótico debido al intercambio de fluidos entre la luz
intestinal y la cavidad hemocélica. Este choque lleva al insecto a la muerte en
unas pocas horas (Knowles y Ellar, 1987).
3.1.4 Producción de Bacillus thuringiensis
Los primeros intentos en la utilización de B. thuringiensis para el control de
insectos plaga tuvieron lugar en la década de los años veinte para el control del
barrenador europeo del maíz, Ostrinia nubilalis. El primer producto basado en B.
thuringiensis fue comercializado en Francia en 1938 y recibió el nombre de
Sporeine. (Weiser, 1986) Desde esta fecha en adelante se ha trabajado en el
estudio del microorganismo, y la optimización en la producción de biopesticidas de
bajo costo.
3.1.4.1. Medio de Cultivo y Condiciones Ambientales
B. thuringiensis requiere para su crecimiento y para una buena producción de
proteínas tóxicas de una fuente de carbono, aminoácidos y algunas sales. La
elección de estos nutrientes dependen de dos factores fundamentales: la

IQ-2003-2-31
14
disponibilidad en la locación de producción y el costo de esta materia prima. La
fuente de carbono es uno de los factores más importantes en el crecimiento de la
bacteria, por lo que la elección de su concentración es importante para un buen
crecimiento. Otros factores medioambientales importantes para el crecimiento de
la bacteria y la síntesis de toxinas, son el pH, la temperatura y la concentración
oxígeno disuelto presente en el medio.
El medio de cultivo debe suministrar los requerimientos nutricionales del
microorganismo y, por lo tanto, proporcionar un ambiente adecuado para el
crecimiento, esporulación y síntesis de proteínas tóxicas, además debe ser de
bajo costo y encontrarse fácilmente en el lugar de producción (González et al.,
2001).
3.1.4.2. Sistemas de producción de B. thuringiensis
B. thuringiensis se ha producido utilizando tanto fermentaciones discontinuas o
tipo batch, como fermentaciones continuas y semicontinuas o fed-batch. De éstas,
la fermentación tipo batch ha sido la más ampliamente utilizada para la producción
industrial de B. thuringiensis (Rowe y Margaritis, 1987).
La fermentación discontinua consiste en la adición inicial de medio de cultivo para
que el microorganismo crezca de forma exponencial hasta que se agote el
sustrato limitante para el crecimiento. En esta fase hay una gran de manda de

IQ-2003-2-31
15
oxigeno y el pH tiende a disminuir debido a la acumulación de ácidos segregados
por la bacteria (Mingone y Avignone-Rossa, 1996).
La fermentación denominada fed-batch consiste en la adición de medio de cultivo
nuevo, sin la remoción de medio agotado. Hay dos tipos de estas fermentaciones,
la continua donde se introduce continuamente los nutrientes y la semicontinua en
donde se hace por pulsos en momentos estratégicos de la fermentación (Mignone
y Avignone-Rossa, 1993).
Los sistemas continuos consisten en mantener los flujos de salida y los de
entrada iguales, ya sea por adición o remoción, o por reflujos en el sistema como
pueden ser las células. Kang et al. (1993) desarrollaron un método de
fermentación continuo con retención celular completa donde se elimina el medio
agotado. Estos cultivos continuos son los menos utilizados debido a la disminución
de toxicidad del producto (González et al., 2001).
3.1.4.3. Productos comerciales a base de B. thuringiensis
El descubrimiento de B. thuringiensis como insecticida microbiano permitió que en
la década de 1930 se iniciara en Europa el uso de B. thuringiensis como agente de
control de insectos. Durante las dos siguientes décadas se realizaron pruebas de
campo contra varios insectos lepidópteros en Europa y Estados Unidos. El primer
producto comercial estaba disponible en Francia en 1938 (Aronson et al., 1986).

IQ-2003-2-31
16
Un desarrollo de productos comerciales más extenso se llevo acabo en la década
de 1950 en varios países como URSS, Checoslovaquia, Francia y Alemania; en
Estados Unidos también se inicio este interés en su uso (Benhard y Utz, 1993).
El contar con productos comerciales a partir de 1960 dio comienzo a un gran
periodo de pruebas de campo en el área agrícola y forestal.
Entre 1960 y 1970 se determinó el alto grado de especificidad entre las cepas;
sepas diferentes exhibían diferente espectro de actividad insecticida. Al parecer,
las especies de insectos difieren en su susceptibilidad a cepas de B. thuringiensis
particulares (Dulmage, 1970).
En los últimos 20 años el mercado de los bioinsecticidas ha sido dominado por
productos que contienen con ingrediente activo una mezcla de cristal y esporas de
la cepa HD1 para el control de plagas agrícolas y forestales, dando lugar a la
sustitución de algunos insecticidas convencionales. El descubrimiento de B.
thuringiensis serovar. israelensis, abrió nuevos mercados, siendo el primer
producto comercial de B. thuringiensis para el control de larvas de mosquito y
mosca negra, incrementándose así las ventas globales en 0.5% de los mercados
insecticidas (Goldberg y Margalit, 1977).
B. thuringiensis es el agente de control biológico de más popularidad, con ventas
en 1995 de noventa millones de dólares, con 67 productos registrados y mas de
450 usos y formulaciones. El surgimiento de B. thuringiensis como bioinsecticidas

IQ-2003-2-31
17
necesitó varias disciplinas aparte de la biología molecular, tales como el
aislamiento de cepas, tecnologías de producción, técnicas de aplicación en
campo, formulación etc., encaminadas a controlar un número importante de plagas
del orden lepidóptero. Dicho esquema ha sido utilizado durante varios años como
un modelo de producción industrial. Posteriormente, la identificación de cepas de
B. thuringiensis con actividad hacia otros organismos plaga como dípteros,
coleópteros, nematodos, ácaros y platelmintos expandió el uso de bioinsecticidas
a base de B. thuringiensis hacia otros mercados (Cerón, 2001).
3.2 REVISION DE LA LITERATURA ACERCA DE MEDIOS DE CULTIVOS
El estudio en planta piloto para la producción de B. thuringiensis, necesita un
especial énfasis en la preparación de medio de cultivo para la obtención de
beneficios económicos a escala industrial (Margarit et al., 1982).
Arcas et al. (1984) estudiaron varios medios de cultivo en proceso discontinuo y
encontraron que la adición de varios micronutrientes como Mg, Mn y Ca aumentan
la producción de biomasa. También encontraron que el uso de nitrógeno orgánico
era necesario para una buena productividad, y que la combinación de extracto de
levadura con sulfato de amonio como fuentes de nitrógeno y la glucosa como
fuente de carbono eran las más apropiadas para el crecimiento del
microorganismo y la producción de biomasa.

IQ-2003-2-31
18
Pearson et al. (1988) observaron que la cantidad de carbohidratos y proteínas en
el medio de cultivo tienen una alta relación en la esporulación y producción de
bioinsecticidas basados en B. thuringiensis.
Skidar et al. (1991) estudiaron el efecto de diferentes sales, encontrando las
concentraciones requeridas de Ca, Mg, Mn y Cu para fermentaciones de B.
thuringiensis. Además encontraron que las concentraciones óptimas de nutrientes
para la esporulación no eran las mismas que para la síntesis de proteínas tóxicas.
Avignone Rossa et al. (1993) realizaron un estudio en donde se encontró que los
cultivos discontinuos de B. thuringiensis producían un ingrediente activo más
tóxico que otros procesos de fermentación. También observaron que la limitación
de fuente de carbono o nitrógeno en la fermentación lleva a producir una toxicidad
inferior, comparada a la máxima obtenible.
Asano et al. (1994) estudiaron la toxicidad de B. thuringiensis contra cuatro
especies diferentes de insectos (Plutella xylostella, Adoxophyes sp., Bómbix mori y
Spodoptera litura). Evaluando el cultivo completo final (CCF), el sedimentado de la
centrifugación del CCF y el sobrenadante de dicho centrifugado, encontrando que
el producto sedimentado era el de mayor toxicidad.

IQ-2003-2-31
19
Liu y Bajpai (1995) desarrollaron un medio de cultivo modificado, la partir del
medio propuesto por Arcas (1984) en el que mejoraron el rendimiento en
fermentaciones fed-batch.
Morris et al. (1996) encontraron que utilizando un medio con extracto de semillas
de algodón y glucosa como fuentes de nitrógeno y carbohidratos respectivamente,
se mejoraba la potencia del complejo espora cristal del producto de la
fermentación.
Farrera et al. (1998) encontraron que la relación entre el carbono y el nitrógeno y
la concentración inicial de sólidos en el medio tienen una repercusión importante
en la producción de espora y de proteínas tóxicas de B. thuringiensis.
Liu B. y Tzeng (1998, 2000) realizaron un modelo para predecir la producción de
esporas, con base en componentes del medio de cultivo, encontrando unas
concentraciones óptimas de tapioca, derivados de pescado y sulfato de amonio.
Estos mismos autores también encontraron que la fuente de carbono afecta el
tiempo de iniciación de la esporulación.
Tyagi et al. (2001, 2002) Encontraron que el lodo de aguas residuales municipales
hidrolizado es un buen medio de cultivo para producir una alta entomotoxicidad en
el producto final. Además encontraron que el valor óptimo en la relación C:N esta
entre 7.9 y 9.9 para lodos combinados de aguas residuales.

IQ-2003-2-31
20
Yang y Wang (2000) estudiaron el efecto de las fuentes de nitrógeno y carbono en
la fase de esporulación y encontraron que un exceso de fuente de carbono
beneficia la expresión de endoproteínas a diferencia de la fuente de nitrógeno que
es perjudicial para esta expresión. También sugieren que el agotamiento del
nitrógeno en el medio es el activador para que se de inicio a la fase de
esporulación en B. thuringiensis.
Wu et al. (2001) propusieron un método de selección de medio de cultivo a partir
de la motilidad del microorganismo, obteniendo mejores resultados en la selección
de medios que los métodos convencionales.
Poopathi et al. (2002) estudiaron medios de cultivo de bajo costo para la
producción de larvicidas de mosquitos, encontrando un medio de cultivo basado
en papa como el más económico para su utilización a nivel industrial.
Içgen et al. (2002) encontraron que los diferentes compuestos de nitrógeno
inorgánico agregados al medio regulan diferencialmente la formación de las
diferentes proteínas tóxicas Cry.
Finalmente Ozkan et al. (2002) estudiaron el efecto de diferentes fuentes de
carbono y nitrógeno en la síntesis de proteínas Cry 11Aa y Cry 4Ba, hallando que
la glucosa, el almidón y las melazas eran supresoras para la producción de

IQ-2003-2-31
21
proteínas Cry 4Ba. También encontraron que el Mn es el factor más crítico para la
síntesis de estas proteínas.

IQ-2003-2-31
22
4. MATERIALES Y MÉTODOS
4.1 DISEÑO EXPERIMENTAL
Se planteó un diseño experimental factorial de 22 con punto medio, de dos
repeticiones, para las concentraciones iniciales de fuente de nitrógeno y de
carbono descritas en la Figura 4; donde se buscaba observar el efecto de la
relación entre las fuentes de carbono y nitrógeno (C:N), y el efecto del cambio de
concentraciones iniciales de estas fuentes en relaciones de C:N constantes, sobre
el rendimiento y producción de proteínas tóxicas de B. thuringiensis.
Se realizaron dos fermentaciones para cada condición, y para cada una de éstas
se evaluaron la concentración de proteína tóxica soluble en medio alcalino, la
concentración de biomasa, la toxicidad sobre Aedes aegypti, el tiempo de
fermentación y el costo de operación de fermentación.

IQ-2003-2-31
23
CIB 3x C:N = 7
F. de Carbono = 50,4 g/l F. de Nitrógeno = 24 g/l
CIB 5x C:N = 7
F. de Carbono = 84 g/l F. de Nitrógeno = 40 g/l
CIB 4x C:N = 5,2
F. de Carbono = 49,5 g/l F. de Nitrógeno = 32 g/l
CIB 3x C:N = 3,3
F. de Carbono = 24 g/l F. de Nitrógeno = 24 g/l
CIB 5x C:N = 3,3
F. de Carbono = 40 g/l F. de Nitrógeno = 40 g/l
Figura 4. Diseño factorial de 22 con punto medio.
4.2 OBTENCIÓN DE SOLUCIÓN DE ESPORAS DE Bacillus thuringiensis
serovar. Israelensis.
Se utilizó la cepa IPS 82 de B. thuringiensis serovar israelensis, perteneciente a la
colección de microorganismos de la unidad de Biotecnología y control Biológico de
la Corporación para Investigaciones Biológicas (CIB), debido a su alto potencial
tóxico contra insectos del orden Díptera.
Se inoculó una colonia de B. thuringiensis de una caja de petri con medio Luria
Bertani (LB) en 5 ml de medio de cultivo CIB1 (Vallejo et al., 1999) y se incubó a
30 ºC y 300 rpm durante 8 horas. Con este cultivo se inoculó 25 ml de medio CIB1
contenido en un erlenmeyer de 125 ml y se incubó a 30 ºC y 300 rpm durante 13

IQ-2003-2-31
24
horas. 5 ml de este cultivo se inocularon en un erlenmeyer de 500 ml con 90 ml de
medio CIB1 que fueron incubados a 30 ºC y 300 rpm durante 48 horas. Después
de verificada la esporulación y la no presencia de contaminantes se centrifugó el
cultivo a 4 ºC y 8000 rpm durante 15 minutos para descartar el sobrenadante y
suspender el sedimentado en 50 ml de NaCl 1 M y se incubó a 37 ºC y 125 rpm
durante 30 minutos. Se centrífugo nuevamente y se realizaron dos lavados más en
agua estéril a las mismas condiciones de centrifugación mencionada. Finalmente
se resuspendió el sedimentado en una solucion estéril solucion buffer salina de
fosfatos (PBS) y glicerol al 20% v/v y se realizaron alícuotas de 5 ml en tubos tapa
rosca de 15 ml. Los tubos fueron calentados en baño de maría a 80 ºC durante 10
minutos y seguidamente se almacenaron a –20 ºC hasta su uso.
4.3 PREPARACIÓN DEL PREINÓCULO E INÓCULO DE LA FERMENTACIÓN
Se inoculó 5 ml de la solución de esporas previamente preparada un erlenmeyer
de 500 ml con 100 ml de medio CIB1 y se incubó a 30 ºC y 250 rpm durante 13
horas. Luego se inoculó este cultivo en dos erlenmeyers de 2000 ml con 500 ml de
medio CIB1 y se incubo a 30 ºC y 250 rpm durante las siguientes 5 horas.

IQ-2003-2-31
25
4.4 PROCESO DE FERMENTACIÓN EN EL REACTOR DE 20 LITROS
Se utilizó el reactor de tanque agitado con volumen nominal de 20 litros de la
planta piloto de fermentaciones de la Unidad de Biotecnología y Control Biológico
de la CIB (Figura 3). Este reactor dispone de sensores de pH, concentración
oxígeno disuelto, temperatura y nivel de espuma. Además cuenta con un sistema
de control de temperatura, pH y espuma. Al reactor se le agrega el medio de
cultivo a evaluar y se inocula el cultivo de un litro incubado 5 horas. El medio de
cultivo y los reactivos de control de pH, ácido nítrico 3N e hidróxido de potasio 3N
y el antiespumante son esterilizados a 120 ºC previamente a la inoculación.
Las condiciones de operación del fermentador, fueron las óptimas que se han
encontrado para este proceso en otros estudios (Gonzalez et al., 2001). La
temperatura se mantuvo en 30 ºC, el pH se mantuvo entre 6.5 – 8.5. El flujo de
aire fue de 2 vvm equivalente a 22 litros de aire por minuto una agitación de 400
rpm. El volumen de uso fue de 11 litros y se mantuvo a una presión de 6 psig.
Se realizaron muestreos periódicamente para observar el estado de la
fermentación; revisar que no hubiera contaminación y determinar la finalización de
la fermentación, que se tomó cuando el cultivo tuviera una lisis celular (después de
esporulada la bacteria) mayor o igual al 90%. Finalmente se tomó una muestra del
cultivo completo final (CCF) ajustado a un pH de 4.1, para hacerse el análisis

IQ-2003-2-31
26
correspondiente a producción de proteínas y de biomasa, a toxicidad y
concentración remanente de fuente de carbono.
Figura 3. P&ID del Fermentador de 20 litros de la Planta Piloto de la CIB.

IQ-2003-2-31
27
4.5 DETERMINACIÓN DE CONCENTRACIÓN DE PROTEÍNAS
4.5.1 Purificación de proteínas
Las proteínas de cada muestra se purificaron centrifugando 1 ml de ésta a 4 ºC y
10000 rpm durante 10 minutos, se resuspendió el sedimentado en NaCl 1M y se
incubó a 37 ºC y 180 rpm durante una hora. Después se le realizó dos lavados con
agua estéril centrifugando a las mismas condiciones y el sedimentado fue
resuspendido en NaOH 100 mM e incubado a 37 ºC y 180 rpm durante 30
minutos para solubilizar la proteína. Finalmente sé centrífugo a 4 ºC y 10000 rpm
durante 10 minutos y se guardó a –20 ºC hasta calcular la concentración de
proteínas por el método de Bradford.
4.5.2 Determinación de Proteínas Solubles en Medio Alcalino por el Método
de Bradford
Se determinó cuantitativamente el contenido de proteínas solubles en medio
alcalino de los cultivos completos finales de fermentaciones realizadas en este
trabajo por el método de Bradford donde la unión del colorante de Coomassie a
las proteínas produce un cambio de color en respuesta a varias concentraciones
de proteína, cambio que es cuantificado por espectrofotometría.

IQ-2003-2-31
28
Se preparó una curva de patrón mediante la dilución en agua estéril del estándar
proteico BSA (Suero de Albúmina Bovina) cuyas concentraciones estaban entre
0.2 mg/ml y 1.2 mg/ml de proteína. Con la proteína solubilizada de cada muestra
también se realizaron diluciones para que su concentración estuviera entre los
límites de la curva patrón. En un vial de 1.5 ml se agregó 100µl de la dilución a
evaluar y de la curva patrón y se le adicionó 320 µl de agua estéril y 80 µl de
reactivo de Bradford. De cada una de estas soluciones se agregó 200 µl en cada
pozo del plato de lectura al que se le midió la absorbancia lector de ELISA a 595
nm. Finalmente con la regresión lineal de la curva patrón se calculó la
concentración de proteína en las muestras.
4.6 DETERMINACIÓN DE CONCENTRACIÓN DE BIOMASA
Se determinó la concentración de biomasa de las fermentaciones mediante el
método de peso seco. Se tomó 5 ml del CCF de cada fermentación al cual se le
agregó 35 ml de agua destilada y se centrífugo a 4 ºC y 8000 rpm durante 10
minutos. Se lavó el sedimentado con 30 ml de agua destilada y se centrífugo
nuevamente a las mismas condiciones. El sedimentado obtenido se resuspendió
en 20 ml de agua y se depositó en copas de aluminio previamente pesadas en un
horno a 100 ºC. Finalmente, se pesaron las copas hasta obtener peso constante.

IQ-2003-2-31
29
4.7 DETERMINACIÓN DE TOXICIDAD
4.7.1 Preparación del Ingrediente Activo por Liofilización
Se centrífugó el CCF de las fermentaciones a 4ºC y 8000 rpm durante 15 minutos.
Después se resuspendió el sedimentado en un volumen igual a 10 ml por cada 50
ml del volumen inicial de CCF centrifugado de NaCl 1M y se incubó a 37ºC y 125
rpm durante 30 minutos. Completado nuevamente el volumen inicial con agua
destilada se centrífuga a 4ºC y 8000 rpm durante 15 minutos. Se lava el
sedimentado dos veces mas con agua destilada a las mismas condiciones. Se
resuspende el sedimentado en agua estéril con volumen de 1/20 del volumen
inicial CCF y se congela a –30ºC durante 4 horas. Después se liofilizó la muestra
congelada a –50ºC y 0.05 milibares.
4.7.2 Bioensayo
El efecto de B. thuringiensis sobre los insectos susceptibles sólo puede ser
medido cuantitativamente mediante bioensayo, que se puede definir como
“cualquier método que mida alguna propiedad de un factor, en términos de
respuesta biológica” (Ibarra y del Rincón, 2001). Para el caso de B. thuringiensis
serovar. israelensis que tiene una alta toxicidad frente al orden Díptera, se utilizó
se como insecto blanco Aedes aegypti con estadío larval acuático de tercer instar

IQ-2003-2-31
30
para evaluar la concentración letal media (CL50), concentración a la cual un
insecticida mata el 50 % de la población.
Se realizaron varias diluciones de ingrediente activo en PBS, las cuales un
pequeño volumen de estas se añadieron a vasos de plástico de 260 ml con 100 ml
de agua y 10 larvas de tercer instar A. aegypti. Se incubó los vasos con diferentes
concentraciones de ingrediente activo a 30 ºC durante 24 horas. Finalmente se
registró el número de larvas muertas para cada dosis. También se realizó un
control negativo al que no se le agrego nada para observar la mortalidad natural
de las larvas.
4.7.3 Análisis Probit
Para calcular CL50 se utilizó el programa estadístico probit para lo que se requiere
el número total de larvas muertas en cada dosis, el número total de larvas
adicionadas por dosis y la concentración de ingrediente activo en cada dosis
obtenido en los bioensayos. Este análisis consiste en la obtención de unidades de
probabilidad (unidades probit) que corresponden al área bajo una distribución
normal. Con la ayuda de la transformada probit es fácil calcular la CL50 debido a
que se puede realizar una recta de los porcentajes de mortalidad contra el
logaritmo de las concentraciones (Ibarra y del Rincón, 2001).

IQ-2003-2-31
31
5. RESULTADOS
5.1 COMPORTAMIENTO DE pH y OXÍGENO DISUELTO EN LAS
FERMENTACIONES
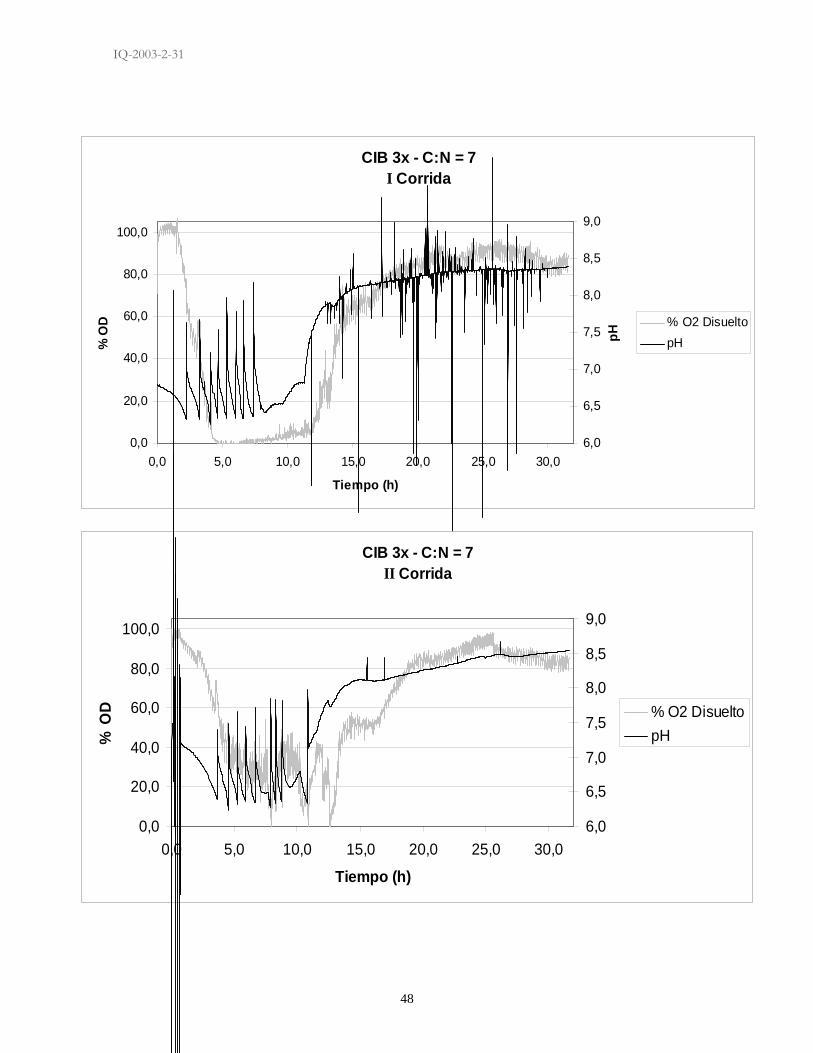
El comportamiento tanto del pH como de la concentración de oxígeno disuelto en
el medio de las fermentaciones evaluadas es muy similar. Se puede observar que
al inicio de todas las fermentaciones el pH tiende a disminuir al igual que la
concentración de oxígeno disuelto y que a partir del momento en que la
concentración de oxígeno disuelto empieza a incrementar el pH del medio se
vuelve cada vez más básico (Anexo 1). El comportamiento es igual para todas las
fermentaciones cambiando únicamente los tiempos a las que estas respuestas se
dan. En el medio más rico tanto en fuente de carbono y nitrógeno el tiempo donde
el oxígeno disuelto estuvo por debajo del 20% duró mucho más que este tiempo
en el medio de cultivo más pobre en nutrientes. De igual manera mientras más rico
fuera el medio de cultivo los controles de pH representados en las gráficas de pH
en el tiempo (Anexo 1) como picos, fueron más numerosos que en los medios de
cultivo con concentraciones bajas de las fuentes evaluadas.

IQ-2003-2-31
32
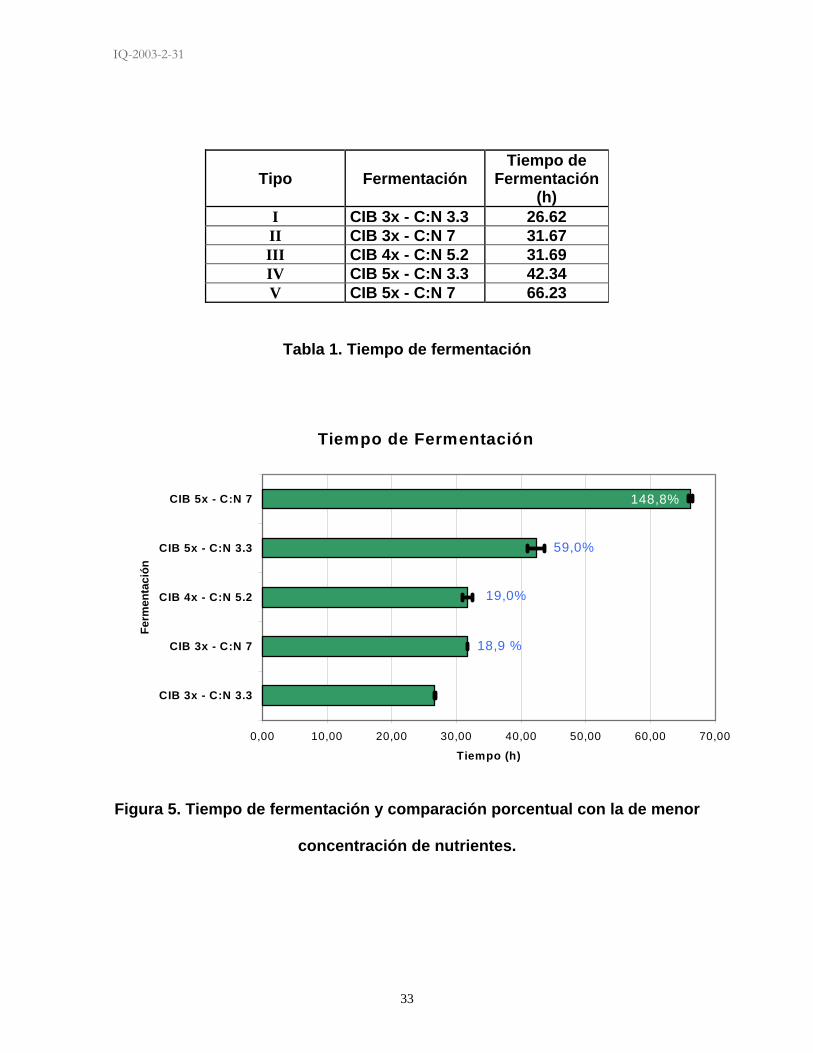
5.2 TIEMPO DE FERMENTACIÓN
El tiempo de fermentación varió a medida que las concentraciones de fuente de
carbono y nitrógeno fueron manipuladas. Las fermentaciones tipo I (Tabla 1) en
donde la concentración de nitrógeno y carbono eran las más bajas duraron
aproximadamente 26.62 horas en alcanzar lisis celular (liberación de cristales y
esporas al medio) de más del 90%, criterio con el que se decretó la finalización de
todas las fermentaciones. En la fermentación tipo II, el tiempo aumentó cinco
horas aproximadamente, teniendo una duración en promedio de 31.67 horas. En
la fermentación tipo IV el tiempo de fermentación tuvo un aumento considerable
con respecto a la tipo I, esta fermentación duró 42.34 horas. La fermentación más
rica en ambas fuentes (fermentación tipo V) tuvo un tiempo de fermentación muy
superior a las cuatro restantes, su tiempo fue de 66.32 horas. Esta fermentación
tuvo muchas dificultades al momento de determinar su finalización debido a la
desincronización de las fases de crecimiento del microorganismo. Finalmente el
punto central del diseño experimental, es decir la fermentación tipo III tuvo un
tiempo promedio fue de 31.69 horas.
En la Figura 5 se observa como fue el comportamiento del tiempo a las diferentes
concentraciones de fuentes de nutrientes evaluadas, y se observa el porcentaje de
aumento de tiempo con respecto a la fermentación de concentraciones más bajas.
IQ-2003-2-31
33
Tipo Fermentación Tiempo de
Fermentación (h)
I CIB 3x - C:N 3.3 26.62 II CIB 3x - C:N 7 31.67 III CIB 4x - C:N 5.2 31.69 IV CIB 5x - C:N 3.3 42.34 V CIB 5x - C:N 7 66.23
Tabla 1. Tiempo de fermentación
Tiempo de Fermentación
0,00 10,00 20,00 30,00 40,00 50,00 60,00 70,00
CIB 3x - C:N 3.3
CIB 3x - C:N 7
CIB 4x - C:N 5.2
CIB 5x - C:N 3.3
CIB 5x - C:N 7
Ferm
enta
ción
Tiempo (h)
148,8%
18,9 %
19,0%
59,0%
Figura 5. Tiempo de fermentación y comparación porcentual con la de menor
concentración de nutrientes.

IQ-2003-2-31
34
5.3 PRODUCCIÓN DE PROTEÍNA TOXICA
La producción de proteína fue favorable en todas las fermentaciones evaluadas en
comparación a la fermentación tipo I (Tabla 2). Esta fermentación produjo en
promedio 4.13 g/l. En la fermentación tipo II en la que se aumenta la fuente
carbono para ajustar la relación C:N a 7, se ve un aumento significativo en la
concentración de proteínas encontradas en el CCF, esta fermentación produjo una
concentración de proteína de 5.37 g/l, valor más alto que el obtenido en la
fermentación tipo IV que tuvo una concentración de 5.01 g/l. La fermentación tipo
V, la más rica en concentración de nutrientes fue la fermentación que mayor
producción de proteínas obtuvo, su producción fue de 8.22 g/l en promedio. La
fermentación que tuvo también una alta producción de proteínas fue el punto
medio (fermentación tipo III) que obtuvo una concentración de 7.06 g/l, casi 3 g/l
más que la fermentación base.
Tipo Fermentación Concentración de Proteína (g/l)
I CIB 3x - C:N 3.3 4.13 II CIB 3x - C:N 7 5.37 III CIB 4x - C:N 5.2 7.06 IV CIB 5x - C:N 3.3 5.01 V CIB 5x - C:N 7 8.22
Tabla 2. Tiempo de fermentación

IQ-2003-2-31
35
En la Figura 6 se observa como fue el comportamiento de la producción de
proteínas en las diferentes fermentaciones probadas y el porcentaje de aumento
en la producción con respecto a la fermentación de concentraciones más bajas.
Concentración de Proteína
3,00
4,00
5,00
6,00
7,00
8,00
9,00
10,00
CIB 3x - C:N 3.3 CIB 3x - C:N 7 CIB 4x - C:N 5.2 CIB 5x - C:N 3.3 CIB 5x - C:N 7
Fermentación
Con
cent
raci
ón d
e Pr
oteí
na
98,8%
29,9%
70,7%
21,1%
Figura 6. Producción de proteína y comparación porcentual con la de menor
concentración de nutrientes.
5.4 PRODUCCIÓN DE BIOMASA
La producción de biomasa, que comprende tanto las esporas como los cristales
proteícos tuvo un comportamiento muy parecido al de la producción de proteínas
pero sin tanta diferencia respecto a la fermentación tipo I (Tabla 3). La

IQ-2003-2-31
36
fermentación tipo I tuvo una producción de biomasa de 6.39 g/l. Con el aumento
de fuente de carbono, la fermentación tipo II incrementó la producción, esta
fermentación produjo 8.35 g/l. La fermentación tipo IV tuvo un aumento en la
producción, más no fue tan significativo como el resto de combinaciones
evaluadas, la producción de esta fue de 7.14 g/l. Como en el caso de producción
de proteínas las fermentaciones de mayor producción de biomasa fueron la del
punto medio (fermentación tipo III) con una producción de 8.71 g/l, y la de mayor
concentración de nutrientes (fermentación tipo V) la cual obtuvo una alta
producción de biomasa de 11.03 g/l.
Tipo Fermentación Concentración
de Biomasa (g/l)
I CIB 3x - C:N 3.3 6.39 II CIB 3x - C:N 7 8.35 III CIB 4x - C:N 5.2 8.71 IV CIB 5x - C:N 3.3 7.14 V CIB 5x - C:N 7 11.03
Tabla 3. Producción de biomasa
La Figura 7 muestra el comportamiento de la producción de biomasa en las
diferentes fermentaciones utilizadas y el porcentaje de aumento en la producción
con respecto a la fermentación de concentraciones más bajas.

IQ-2003-2-31
37
Concentración de Biomasa
4.00
5.00
6.00
7.00
8.00
9.00
10.00
11.00
12.00
13.00
14.00
CIB 3x - C:N 3.3 CIB 3x - C:N 7 CIB 4x - C:N 5.2 CIB 5x - C:N 3.3 CIB 5x - C:N 7
Fermentación
Con
cent
raci
ón d
e Pr
oteí
na
72,6% 30,7%
36,3%
11,7%
Figura 7. Producción de biomasa y comparación porcentual con la de
menor concentración de nutrientes.
5.5 TOXICIDAD
La toxicidad medida como concentración letal 50, fue muy similar en todas las
fermentaciones, menos en la fermentación tipo V donde las concentraciones de
nutrientes eran las mayores. En esta fermentación se ve un detrimento de la
toxicidad del ingrediente activo de la fermentación al aumentar la concentración
necesaria para matar la mitad de la población (Figura 8).

IQ-2003-2-31
38
Tipo Fermentación Toxicidad
CL50 (ng/ml)
I CIB 3x - C:N 3.3 10.06 II CIB 3x - C:N 7 11.17 III CIB 4x - C:N 5.2 9.79 IV CIB 5x - C:N 3.3 8.38 V CIB 5x - C:N 7 15.51
Tabla 4. Concentración letal media.
Toxicidad
0,00
2,00
4,00
6,00
8,00
10,00
12,00
14,00
16,00
18,00
20,00
CIB 3x - C:N 3.3 CIB 3x - C:N 7 CIB 4x - C:N 5.2 CIB 5x - C:N 3.3 CIB 5x - C:N 7
Fermentación
CL
50 (n
g/m
l)
Figura 8. Concentración letal media.
5.6 COSTOS
Los costos de cada una de las cinco fermentaciones evaluadas se obtuvieron
teniendo en cuenta los costos directos y los gastos operativos. A partir de las

IQ-2003-2-31
39
diferentes concentraciones de medio de cultivo se obtuvo el costo de materia
prima, incluyendo el costo de los preinóculos y el costo de los reactivos de control
de pH y espuma. En cuanto los gastos de operación, a partir de el tiempo de
fermentación se estimó el costo de los servicios industriales requeridos en cada
una de las fermentaciones, tales como vapor, agua, energía eléctrica,
alcantarillado, y el costo de mano de obra tanto del ingeniero y el asistente de
laboratorio.
Tipo Fermentación Costo
Fermentación ($)
I CIB 3x - C:N 3.3 $ 98,344,67 II CIB 3x - C:N 7 $ 114,908,56 III CIB 4x - C:N 5.2 $ 116,934,64 IV CIB 5x - C:N 3.3 $ 149,534,38 V CIB 5x - C:N 7 $ 222,728,93
Tabla 5. Costo Fermentación.
En la Figura 9 se muestra el costo de cada una de las diferentes fermentaciónes y
el cambio porcentual del costo contra la fermentación tipo I.

IQ-2003-2-31
40
Costos
$ 0,00
$ 50,000,00
$ 100,000,00
$ 150,000,00
$ 200,000,00
$ 250,000,00
CIB 3x - C:N 3.3 CIB 3x - C:N 7 CIB 4x - C:N 5.2 CIB 5x - C:N 3.3 CIB 5x - C:N 7Fermentación
Cos
tos
x Fe
rmen
taci
ón
26,5%
16,8% 18,9%
52,0%
Figura 9. Costo Fermentación.

IQ-2003-2-31
41
6 ANÁLISIS
6.1 PRODUCCIÓN DE INGREDIENTE ACTIVO PARA BIOPESTICIDAS
El comportamiento del pH en el tiempo donde se observa una caída de éste en las
primeras horas de fermentación se debe a que B. thuringiensis en su fase de
crecimiento exponencial produce ácidos. Durante la fase de esporulación y lisis
son segregados metabolitos alcalinos, lo cual hace que aumente el pH del medio
de cultivo. Entonces, a partir de los picos del límite inferior de control observados
en las gráficas de pH (Anexo 1), se puede observar el tiempo aproximado de
crecimiento de la bacteria. Este tiempo de la fase vegetativa también se puede
detectar a partir del comportamiento del oxígeno disuelto, puesto que la mayor
demanda de oxígeno durante la fermentación es en esta fase.
El efecto en el tiempo de fermentación respecto al cambio de relación entre la
fuente de carbono y nitrógeno muestra un aumento en este tiempo. Esto se debe a
que la fuente de carbono es el nutriente más importante en la fase de crecimiento,
la cual al ser aumentada, hace que exista un mayor crecimiento. En cuanto al

IQ-2003-2-31
42
aumento de concentración inicial de estas fuentes, el aumento en el tiempo de
fermentación es todavía más considerable. Se observa entonces que la
concentración inicial de la fuente de nitrógeno del medio tiene un efecto importante
en el tiempo total de fermentación. Por medio de un análisis de varianza se
observa que las fermentaciones tipo II y tipo III (Tabla 1) no presentan una
diferencia estadísticamente significativa (Anexo 2).
El efecto de la relación C:N en la producción de proteínas tóxicas y biomasa es
muy similar. La producción aumenta a medida que el medio de cultivo es más rico
en fuente de carbono. Este aumento es mucho más considerable cuando las
concentraciones iniciales de las fuentes son mayores. Cuando aumenta la
concentración inicial de las fuentes estudiadas y se mantiene la relación C:N, la
producción de proteínas y de biomasa también se ve favorecida, sin embargo este
aumento de productividad es más significativo siempre y cuando se tenga una
adecuada relación C:N. Haciendo un análisis de varianza en la producción de
proteínas (Anexo 3) se observa que entre las fermentaciones tipo II y tipo VI
(Tabla 2) no hay diferencias estadísticamente significativas, mientras que si hay
diferencias frente al punto medio que es uno de los de mayor productividad en
este aspecto. En cuanto a producción de biomasa entre las fermentación tipo II y
tipo III (Tabla 3) no existen diferencias estadísticamente significativas (Anexo 4).
La concentración letal media utilizada como medida de la toxicidad de las
fermentaciones evaluadas se realizo con el fin de comparar la calidad del producto

IQ-2003-2-31
43
final de cada fermentación. La calidad de cuatro de las cinco diferentes
fermentaciones evaluadas presentaron una calidad similar al obtener un CL50
promedio de alrededor de 10 ng/ml. Esto se evidencia con el análisis de varianza
(Anexo 5) donde se ve que no hay diferencias significativas en la toxicidad de las
fermentaciones, excepto en la fermentación tipo V (tabla 4) la más rica en
nutrientes, la cual tuvo una toxicidad un poco más baja a las demás, sin embargo
se encuentra en el mismo orden de magnitud.
6.2 COSTOS DE PRODUCCIÓN
El costo de las fermentaciones aumentó con respecto a la fermentación base
(fermentación tipo I) debido al incremento en la cantidad de materia prima y al
aumento en el tiempo de fermentación. El costo de la materia prima frente al costo
de operación es casi despreciable, debido a que la mano de obra es el factor más
importante en las fermentaciones evaluadas en la planta piloto. Se observó que
las fermentaciones tipo II y tipo III (Tabla 5) tuvieron un incremento muy bajo con
respecto a la fermentación tipo I, esto debido a que el tiempo de fermentación de
estas fue muy similar. A diferencia de éstas, las fermentaciones tipo IV y tipo V
tuvieron un costo muy superior a las anteriores debido al largo tiempo de
fermentación.

IQ-2003-2-31
44
Con la productividad y el costo de cada una de las fermentaciones se calculó el
valor de producción por gramo de proteína y biomasa (Tabla 6).
Tipo Fermentación Costo
Proteína ($/g)
Costo Biomasa
($/g) I CIB 3x - C:N 3.3 $ 2,162,63 $ 1,398,58 II CIB 3x - C:N 7 $ 1,944,63 $ 1,250,55 III CIB 4x - C:N 5.2 $ 1,506,49 $ 1,220,13 IV CIB 5x - C:N 3.3 $ 2,715,37 $ 1,903,48 V CIB 5x - C:N 7 $ 2,464,20 $ 1,835,02
Tabla 6. Costo de Producción.
En la Figura 11 se ve la comparación en porcentaje del costo de producción con
respecto a la fermentación base. Aquí se ve claramente que en dos de las
fermentaciones evaluadas (tipo II y tipo III) se logró producir ingrediente activo
para biopesticidas a un menor costo con respecto a las fermentaciones realizadas
con el medio de cultivo actualmente utilizado en la planta piloto de la CIB.

IQ-2003-2-31
45
Costos de Produccion por Gramo de Producto
$ 0,00
$ 500,00
$ 1,000,00
$ 1,500,00
$ 2,000,00
$ 2,500,00
$ 3,000,00
CIB 3x - C:N 3.3 CIB 3x - C:N 7 CIB 4x - C:N 5.2 CIB 5x - C:N 3.3 CIB 5x - C:N 7
Fermentación
Cos
to x
Gra
mo
($/g
)
Costo Proteína ($/g) Costo Biomasa ($/g)
31,2%-10,1%
-10,6%-30,3%
-12,76%
25,6%
36,1%
13,94%
Figura 11. Costo de Producción.

IQ-2003-2-31
46
7. CONCLUSIONES
Se obtuvieron dos medios de cultivo en los que el costo de producción tanto de
biomasa como de proteínas tóxicas disminuyó con respecto al medio de cultivo de
referencia (CIB 3x, C:N 3.3), y además la toxicidad de esos medios no presentó
diferencias estadísticamente significativas frente a la del medio base. El mejor
costo de producción tanto de proteínas como de biomasa se obtuvo con un medio
de cultivo con concentraciones iniciales de fuente de nitrógeno y carbono de 32 y
49 g/l respectivamente y una relación C:N 5.2.
Por otra parte se observó que el aumento de la relación entre las fuentes de
carbono y de nitrógeno favorece la producción de proteínas tóxicas y biomasa.
También se observó que al aumentar las concentraciones iniciales de carbono y
nitrógeno hay un aumento en la producción de proteínas y biomasa, este aumento
es más significativo desde que exista una buena relación de concentraciones de
estas fuentes. Finalmente con los resultados obtenidos se puede decir que la
concentración inicial de fuente de nitrógeno parece ser más determinante en el
tiempo total de fermentación que la fuente de carbono.

IQ-2003-2-31
47
CIB 3x - C:N = 3,3 I Corrida
0,0
20,0
40,0
60,0
80,0
100,0
0,0 5,0 10,0 15,0 20,0 25,0
Tiempo (h)
% O
D
6,0
6,5
7,0
7,5
8,0
8,5
9,0
pH
% O2 DisueltopH
8. ANEXOS
ANEXO 1. Graficas de Comportamiento de pH y Oxígeno Disuelto
CIB 3x - C:N = 3,3 II Corrida
0,0
20,0
40,0
60,0
80,0
100,0
0,0 5,0 10,0 15,0 20,0 25,0
Tiempo (h)
% O
D
6,0
6,5
7,0
7,5
8,0
8,5
9,0
pH
% O2 DisueltopH
IQ-2003-2-31
48
CIB 3x - C:N = 7 I Corrida
0,0
20,0
40,0
60,0
80,0
100,0
0,0 5,0 10,0 15,0 20,0 25,0 30,0
Tiempo (h)
% O
D
6,0
6,5
7,0
7,5
8,0
8,5
9,0
pH
% O2 DisueltopH
CIB 3x - C:N = 7 II Corrida
0,0
20,0
40,0
60,0
80,0
100,0
0,0 5,0 10,0 15,0 20,0 25,0 30,0
Tiempo (h)
% O
D
6,0
6,5
7,0
7,5
8,0
8,5
9,0
% O2 DisueltopH
IQ-2003-2-31
49
CIB 4x - C:N = 5.2I Corrida
0,0
20,0
40,0
60,0
80,0
100,0
0,0 5,0 10,0 15,0 20,0 25,0 30,0
Tiempo (h)
% O
D
6,0
6,5
7,0
7,5
8,0
8,5
9,0
% O2 DisueltopH
CIB 4x - C:N = 5.2II Corrida
0,0
20,0
40,0
60,0
80,0
100,0
0,0 5,0 10,0 15,0 20,0 25,0 30,0
Tiempo (h)
% O
D
6,0
6,5
7,0
7,5
8,0
8,5
9,0
% O2 DisueltopH

IQ-2003-2-31
50
CIB 5x - C:N = 3,3 I Corrida
0,0
20,0
40,0
60,0
80,0
100,0
0,0 5,0 10,0 15,0 20,0 25,0 30,0
Tiempo (h)
% O
D
6,0
6,5
7,0
7,5
8,0
8,5
9,0
% O2 DisueltopH
CIB 5x - C:N = 3,3 II Corrida
0,0
20,0
40,0
60,0
80,0
100,0
0,0 5,0 10,0 15,0 20,0 25,0 30,0
Tiempo (h)
% O
D
6,0
6,5
7,0
7,5
8,0
8,5
9,0
% O2 DisueltopH


IQ-2003-2-31
52
ANEXO 2. Análisis de Varianza del Tiempo de Fermentación
Gráfica 1. ANOVA
Duncan test; variable Tiempo (h) (Tiempo.sta) Probabilities for Post Hoc Tests
Error: Between MS = .48188, df = 5.0000
CIB 3x - C:N 3.3 CIB 3x - C:N 7 CIB 5x - C:N 3.3 CIB 5x - C:N 7 CIB 4x - C:N 5.2CIB 3x - C:N 3.3 0,000978 0,000098 0,000068 0,000977
CIB 3x - C:N 7 0,000978 0,000152 0,000098 0,972530CIB 5x - C:N 3.3 0,000098 0,000152 0,000246 0,000262
CIB 5x - C:N 7 0,000068 0,000098 0,000246 0,000143CIB 4x - C:N 5.2 0,000977 0,972530 0,000262 0,000143
Tabla 1. Test de Duncan
Caracteristica; LS MeansCurrent effect: F(4, 5)=1048.6, p=.00000
Effective hypothesis decompositionVertical bars denote 0.95 confidence intervals
CIB 3x - C:N 3.3CIB 3x - C:N 7
CIB 5x - C:N 3.3CIB 5x - C:N 7
CIB 4x - C:N 5.2
Caracteristica
202530354045505560657075
Tiem
po (h
)

IQ-2003-2-31
53
ANEXO 3. Análisis de Varianza de Producción de Proteínas
Gráfica 1. ANOVA
Duncan test; variable Proteina (g/l) (Proteina.sta) Probabilities for Post Hoc Tests
Error: Between MS = .63602, df = 151.00
CIB 3x - C:N 3.3 CIB 3x - C:N 7 CIB 5x - C:N 3.3 CIB 5x - C:N 7 CIB 4x - C:N 5.2CIB 3x - C:N 3.3 0,000011 0,000065 0,000004 0,000003
CIB 3x - C:N 7 0,000011 0,035948 0,000011 0,000009CIB 5x - C:N 3.3 0,000065 0,035948 0,000003 0,000011
CIB 5x - C:N 7 0,000004 0,000011 0,000003 0,000009CIB 4x - C:N 5.2 0,000003 0,000009 0,000011 0,000009
Tabla 1. Test de Duncan
Caracteristica; LS MeansCurrent effect: F(4, 151)=136.75, p=0.0000
Effective hypothesis decompositionVertical bars denote 0.95 confidence intervals

IQ-2003-2-31
54
8.4. ANEXO 4. Análisis de Varianza de Producción de Biomasa
Gráfica 1. ANOVA
Duncan test; variable Biomasa (g/l) (Biomasa.sta)
Probabilities for Post Hoc Tests
Error: Between MS = 2.0218, df = 109.00 CIB 3x - C:N 3.3 CIB 3x - C:N 7 CIB 5x - C:N 3.3 CIB 5x - C:N 7 CIB 4x - C:N 5.2
CIB 3x - C:N 3.3 0,000071 0,080107 0,000029 0,000046CIB 3x - C:N 7 0,000071 0,005224 0,000057 0,398863
CIB 5x - C:N 3.3 0,080107 0,005224 0,000046 0,000535CIB 5x - C:N 7 0,000029 0,000057 0,000046 0,000105
CIB 4x - C:N 5.2 0,000046 0,398863 0,000535 0,000105
Tabla 1. Test de Duncan Tabla 1. Test de Duncan
Caracteristica; LS MeansCurrent effect: F(4, 109)=37.469, p=0.0000
Effective hypothesis decompositionVertical bars denote 0.95 confidence intervals
CIB 3x - C:N 3.3CIB 3x - C:N 7
CIB 5x - C:N 3.3CIB 5x - C:N 7
CIB 4x - C:N 5.2
Caracteristica
5
6
7
8
9
10
11
12
13
Bio
mas
a (g
/l)

IQ-2003-2-31
55
8.5. ANEXO 5. Análisis de Varianza de la Toxicidad
Gráfica 1. ANOVA
Duncan test; variable CL50 (ng/ml) (Toxicidad.sta)
Probabilities for Post Hoc Tests
Error: Between MS = 2.1553, df = 5.0000 CIB 3x - C:N 3.3 CIB 3x - C:N 7 CIB 5x - C:N 3.3 CIB 5x - C:N 7 CIB 4x - C:N 5.2
CIB 3x - C:N 3.3 0,483929 0,314362 0,015713 0,859446CIB 3x - C:N 7 0,483929 0,126385 0,031855 0,399980
CIB 5x - C:N 3.3 0,314362 0,126385 0,005979 0,381110CIB 5x - C:N 7 0,015713 0,031855 0,005979 0,013821
CIB 4x - C:N 5.2 0,859446 0,399980 0,381110 0,013821
Tabla 1. Test de Duncan
Caracteristica; LS MeansCurrent effect: F(4, 5)=6.8677, p=.02898
Effective hypothesis decompositionVertical bars denote 0.95 confidence intervals
CIB 3x - C:N 3.3CIB 3x - C:N 7
CIB 5x - C:N 3.3CIB 5x - C:N 7
CIB 4x - C:N 5.2
Caracteristica
4
6
8
10
12
14
16
18
20
CL 5
0 (ng/
ml)

IQ-2003-2-31
56
9. BIBLIOGRAFÍA Abdel-Hameed, A. 2001. Stirred tank culture of Bacillus thuringiensis H-14 for the production of the mosquitocidal δ-endotoxin; mathematical modeling and scaling-up studies. World Journal of Microbiology and Biotechnology. 17: 857-861. Arcas, J., O. Yantormo, E. Arrarás y R. Ertola. 1984. A new medium for growth and delta-endotoxin production by Bacillus thuringiensis var. kurstaki. Biotech. letters. 6, (8): 495-500. Aronson, A., W. Beckman y P. Dunn. Bacillus thuringiensis and related insect. Pathogens. Microbiol. Rev. 50; 1-14 Asano, S., H. Hori y y. Cui. 1994. A unique insecticidal activity in Bacillus thuringiensis growth medium. Appl. Entomol. Zool. 29: 39-45. Avignone Rossa, C. y C. Mignone. 1993. δ Endotoxin activity and spore production in batch and fed-batch culture of Bacillus thuringiensis. Biotech. letters. Vol. 15, 3: 295-300. Avignone Rossa, C., J. Arcas y C. Mignone. 1992. Bacillus thuringiensis growth, sporulation and δ-endotoxin production in oxygen limited and non-limited cultures. World Journal of Microbiology and Biotechnology. 8: 301-304. Benhard, K. y R. Utz. 1993. Production of Bacillus thuringiensis insecticides for experimental and commercial uses. Pp 255-267. En Practice Entwistle, P.F., J.S. Cory, M.J. Bailey y S. Goggs, (ed.), Bacillus thuringiensis An environmental biopesticide. Theory and Wiley, Chichester, UK. Bulla, L.A., Jr., D.B. Bechtel, J.K. Kromer, Y.I. Shethna, A.I. Aronson y P.C. Fitzjames. 1980. Ultrastructure, physiology, and biochemistry of Bacillus thuringiensis. CRC Crit. Rev. Microbiol. 8: 147-204. Caballero, P. e Iriarte, J. 2001. Biología y ecología de Bacillus thuringiensis. Capítulo 1: 15-44. En: Caballero, P. Y Ferré, J., (ed.), Bioinsecticidas: fundamentos y aplicaciones de Bacillus thuringiensis en el control integrado de plagas. PHYTOMA, España. Cerón, J.A. 2001. Productos comerciales nativos y recombinantes a base de Bacillus thuringiensis. p 153-168 en: Caballero, P. Y Ferré, J., (ed.), Bioinsecticidas: fundamentos y aplicaciones de Bacillus thuringiensis en el control integrado de plagas. PHYTOMA, España.

IQ-2003-2-31
57
Dulmage, H.T. 1970. Insecticidal activity of HD1, a new isolate Bacillus thuringiensis var. alesti. J. Invert. Pathol. 15: 232-239. Farrera, R.R., F. Pérez-Guevara y M. de la Torre. 1998. Carbon:nitrogen ratio interacts with initial concentration of total solids on insecticidal crystal protein and spore production in Bacillus thuringiensis. Appl. Microbiol. Biotechnol. 49: 758-765. Feitelson, J.S. 1993. The Bacillus thuringiensis family tree. p 63-71 En: Kim, L., (ed.), Advanced Engineered pesticides. Marcel Dekker Inc. New York. Goldberg, L.J. y J. Margarit. 1977. A bacterial spore demonstrating rapid larvicidal activity against Anopheles segentii, Uranotacnia unguiculata, Culex univitattus, Aedes aegypti and Culex pipiens. Mosq. News. 37 355-358 González, A., A. Restrepo y S. Orduz. 2001. Producción de Bacillus thuringiensis. p 109-132 en: Caballero, P. Y J. Ferré, (ed.), Bioinsecticidas: Fundamentos y aplicaciones de Bacillus thuringiensis en el control integrado de plagas. PHYTOMA, España. Hofte, H. y H.R. Witeley. 1989. Insecticidalcrystal proteins of Bacillus thuringiensis. Microbiol. Rev. 53: 242-255. Honée, G. Y B. Visser. 1993. The mode of action of Bacillus thuringiensis crystal proteins. Entomol. Exp. Appl. 69: 145-155. Ibarra, J.E. y M.C. Del Rincón. 2001. Cuantificación toxicológica de Bacillus thuringiensis. Capítulo 6: 15-44. En: Caballero, P. Y Ferré, J., (ed.), Bioinsecticidas: fundamentos y aplicaciones de Bacillus thuringiensis en el control integrado de plagas. PHYTOMA, España. Icgen, Y., B. Icgen y G. Ozcengiz. 2002. Regulation of crystal protein biosinthesis by Bacillus thuringiensis: I. Effects of mineral elements and pH. Res. Microbiol. 153 : 599-604. Icgen, Y., B. Icgen y G. Ozcengiz. 2002. Regulation of crystal protein biosinthesis by Bacillus thuringiensis: I. Effects of carbon and nitrogen sources. Res. Microbiol. 153 : 605-609. Karim, M.I., R.J. Lucas, K.J. Osburne y P.L. Rogers.. 1993. The effect of oxygen on the sporulation and toxicity of Bacillus sphaericus 2362. Biotech. letters. Vol. 15, 1: 47-50. Khowles, B.H. y J.A.T. Dow. 1993. The crystal delta-endotoxins of Bacillus thuringiensis models for their mechanisms of action of the insects gut. BioEssays 15: 469-476.

IQ-2003-2-31
58
Levinson, B.L. 1990. High performance liquid chromatography analisis of tow β-exotoxons produced by some Bacillus thuringiensis strains, p 115-136 en: Hickle, L.A. y W.L. Fitch, (ed.), Analytical chemistry of Bacillus thuringiensis. American Chemical Society , Washington. Liu, B.L. y Y.M. Tzeng. 1998. Optimization of growth medium for the production of spores from Bacillus thuringiensis using response surface methodology. Bioprocess Engineering 18: 413-418. Liu, B.L. y Y.M. Tzeng. 2000. Characterization study of the sporulation kinetics of Bacillus thuringiensis using response surface methodology. Biothechnology and Bioengineering 68: 11-17. Liu, W.M. y R.K. Bajpai. 1995. A modified growth medium for Bacillus thuringiensis. Biotech. Prog. 11: 589-591. Margalit, J. y H.Oren. 1982. Experience in Production of Bacillus thuringiensis var. israelensis H – 14 in Israel. Special Programme for Research and Training in Tropical Diseases. Morris, O.N., V. Converse, P. Kanagaratnam y J.S. Davies. 1996. Effect of cultural conditions on spore-crystal yield and toxicity of Bacillus thuringiensis subsp. Azawai (HD133). Journal of Invertebrate Pathology. 67: 129-136. Ozcan, M., F.B. Dilek, U. Yetis y G. Ozcengiz. 2003. Nutritional and cultural parameters influencing antidipterian delta-endotoxin production. Res. Microbiol. 154 : 49-53. Pearson, D. y O.P. Ward. 1988. Effect of culture conditions on growth and sporulation of Bacillus thuringiensis supsp. Israelensis and development of media for production of protein crystal endotoxin. Biotech. letters. Vol. 10, 7: 451-456. Poopathi, S., K. Anup, L. Kabilan y V. Sekar. 2002. Development of low-cost media for thr culture of mosquito larvicides, Bacillus spaericus and Bacillus thuringiensis serovar. israelensis. World Journal of Microbiology and Biotechnology. 18: 209-216. Porcar, M. y P. Caballero. 2001. Biología y ecología de Bacillus thuringiensis. p 15-44 en: Caballero, P. Y J. Ferré, (ed.), Bioinsecticidas: Fundamentos y aplicaciones de Bacillus thuringiensis en el control integrado de plagas. PHYTOMA, España. Sacci, F.V., P. Parenti, G.M. Hanozet, B. Giordona, P. Lulhy y M.G. Wolfersberger. 1986. Bacillus thuringiensis toxin inhibits K+ gradient-dependent

IQ-2003-2-31
59
aminoacid transport across the brush border membrane of Pieris brassicae midgut cells. FEBS Lrtt. 204: 213-118. Shnepf, H.E. y H.R Whiteley 1985. Protein toxin of Bacilli? p 209-216 en: Hoch, J.A. y P. Setlow, (ed.), Milecular biology of microbial differentiation. American Society of Microbiology. Sikdar, D.P., M.K. Majumdar y S.K- Majumdar. 1991. Effect of minerals on tha production of the delta endotoxin by Bacillus thuringiensis supsp. Israelensis. Biotech. letters.13 (7) 511-514. Stewart, G.S.A., K. Johnstone, E. Hagelberg y D.J. 1981. Commitment of bacterial spores to germinate. Biochem. J. 198: 101-106. Tirado, M. de L., R.D. Tyagi y J.R. Valero. 2001. Wastewater treatment sludge as raw material for the production of Bacillus thuringiensis Based pesticides. Wat. Res. 35 (16): 3807-3816. Vidyarthi, A.S., R.D. Tyagi, J.R. Valero y R.Y. Surampalli. 2002. Wastewater treatment sludge as raw material for the production of Bacillus thuringiensis Based pesticides. Wat. Res. 36: 4850-4860. Wu, W.T., S. Chen y J.Y. Hong. 2001. A droplet medium selection for cultivation of Bacillus thuringiensis. Biotech. Prog. 17: 778-780. Yang, X.M. y S.S. Wang. 2000. Phase specific optimization of multiple endotoxin protein production with genetically engineered Bacillus thuringiensis. Biothecnil. Appl. Biochem. 31: 71-76.