melanie l. lews department of sciences · abstract in many parts of the world atrazine is the most...
TRANSCRIPT

MELANIE L. L E W S DEPARTMENT OF PLANT SCIENCES
Submitted in partial fuifilment of the requirements for the degree of
Master of Science
Faculty of Graduate Studies The University of Western Ontario
London, ON August 1998
0 Melanie L. Lewis t 998

National Library Bibliothèque nationale du Canada
Acquisitions and Acquisitions et Bibliographie Services seMces bibliographiques
395 mio on Street 395. nie Wellington OttawaON K I A W OttawaON KIAON4 Canada Canada
The author has granted a non- exclusive licence ailowhg the National Library of Canada to reproduce, loan, distriibute or sell copies of this thesis in microform, paper or electronic formats.
The author retains ownershrp of the copyright in this thesis. Neither the thesis nor substantial extracts £iom it may be printed or otherwise reproduced without the author's permission.
L'auteur a accordé une Licence non exclusive permettant à la Bibliothèque nationale du Canada de reproduire, prêter, distribuer ou vendre des copies de cette thèse sous la forme de microfiche/film, de reproduction sur papier ou sur format électronique.
L'auteur conserve la propriété du droit d'auteur qui protège cette thèse. Ni la thèse ni des extraits substantiels de celle-ci ne doivent être imprimés ou autrement reproduits sans son autorisation.

ABSTRACT
In many parts of the world atrazine is the most cornmonly used herbicide for control
of broad leaf weeds in a vanety of crops. Its intensive use and persistence in soil has led to
concerns regardinj gound and surface water contamination. Consequently there is
considerable interest in developing agricultural management practices that would minimize
the potential for ground and surface water pollution by atrazine and its metabolites. One
possibility is to use soil bactena that degrade atrazine to non-toxic metabolites. Atrazine-
degrading bacteria have recently been isolated From agncultural fields in Ontario and
Québec.
In the present study, rep-PCR using the consensus pnmers BOXAlR and
ERIC 1 WERIC2, plasmid profiles and biochemical reactions were used to characterize 1 2
gram-negative atrazine degraders isolated from a single site near Ottawa, Ontario. Results
indicated that there was very limited diversity among these isolates.
A unique atrazine-degrading gram-positive bactenum from Sr. Hyacinthe. Québec.
designated C 1 87, also was characterized. 16s rDNA sequencing indicated that C 1 87 was
a new species in the genus Nocardioides. Nocardioides sp. strain C 187 used atrazine as a
sole source ofnitrogen and as a carbon source. Although it could d e p d e atrazine it couid
not mineralize it to CO1 and ammonia. High pressure liquid chromatography revealed that
atrazine was degraded to the non-phytotoxic metabolite hydroxyatrazine under aerobic and
oxygen-free conditions. Hydroxyatrazine was also degraded under these conditions. This
suggested that the first two steps in the degradation pathway were catalyzed by hydrolases,
as has been described for Psezrdomonar sp. strain ADP (De Souza et al.. 1996 and Boundy-
... 111

Mills et al.. 1997). The hydrolases (atrazine chlorohydrolase and hydroxyatrazine
ethylaminohydrolase) synthesized by Psewdomonas sp. strain ADP are cncoded by the genes
atzA and utzB. Dot-blot hydridization with DIG-labelled probes prepared From at1.4 and ut=B
indicated no sequence similarity with purified jenomic DN.4 fiorn .Vocat-diozcles sp. strain
C 157. The gram-positive bactenum. Rhodococnrs corallinzrs YRRL B- l 5 U J R . degades
dethylaminoatrazine via the enzyme s-triazine hydrolase. which is rncoded by r ~ 4 (Shao et
al., 1995). Dot-blot hydndization usinp a DIG-labelled probe prepared from trz.4 indicated
no sequence homology with .Vocordiozdes sp. strain C 1%'. These results sugsest that the
hydrolases responsible for atrazine and hydroxyatrazine degradation by .Vucardiordes sp.
strain C 1 87 are not encoded by genes homologous to ar;-l. acB or rr-4 .
Cell free extracrs prepared bom strain C 187 cells gown with atrazine and without
a m i n e were analysed for their ability to degrade atrazine. For both treatrnents. no lag phase
was present, atrazine was degraded ro hydroxyatrazine at the same rate. and hydroxyanazine
was no t metabolized. These results suggest that anazine dec hloi-ination is constitutive.
The results of this study indicate that atrazine-degrading bactena isolared on a plot
scale are not diverse. This is the first report of a .Vocardioides species degrading atrazine.
Atrazine degradation by :Vocardzoides sp. C 1 87 is constitutive and the aromatic nucleus of
atrazine is not rnetabolized. While ,Vocardioides sp. strain CI87 degrades a m i n e
hydrolytically it does not possess sequences homologous to utz-f , ucB or r r -4 .
Keywords: atrazine, rnineralizaton, bacteria, degrading, hybridization, diversity.

.4CLiYOWLEDGEMENTS
A number of people both in the lab and outside of the lab whose contributions and
effons are highly appreciated were instrumental towards the completion of this project.
First and forernost. I would like to thank my supervisors Dr. Edward Topp and
Dr. Diane Cuppels. for their academic guidance throughout the past two years. 1 thank
my advisory cornmittee members, Dr. Andre Lachance and Dr. Susanne Kohalmi for
their advice and invaluable suggestions.
1 am extrernely gratefùl to Sandra LMillar, Henry Bork, Teresa Ainsworth, and Lou
Ann Verellen for their technical advice and assistance.
1 would like to thank M. De Souza, Dr. L. Wackett and Dr. M. Sadowsky at the
University of Minnesota, for providing me with Prer~domonar sp. strain ADP. piMD4
and pATZB-2. Thanks to Dr. D. Cuppels for providing me with bacterial strains that
were essential to many of my experiments.
Finally, I would like to thank Jeff Ivey for his endless help with the mathematical
and statistical analysis required for this thesis and for his emotional support as i
completed this project.
This project was financially supported by The University of Western Ontario and
Novartis Crop Protection.

TABLE OF CONTENTS
TITLE PAGE ......................................................................................................
CERTIFICATE OF EXAIMINATION .......................................................-.........
...................................................................................................... ABSTELKT..
ACKNOWL EDGEMENTS.. ...............................................................................
TABLE OF CONTENTS ..................,...........................................................
LIST OF FIGURES ............................................................................................
............................................................................................. LIST OF TABLES
............................................................................................ AB B REVIATIONS
Chapter 1 - INTRODUCTION ......................................................................-.
Chapter 2 - LITERATURE REVIEW ............................................................
........................................................................................ 2.1 Atrazine
2.2 Atrazine-degradation. ....................................................................
2.3 Pseudomonas sp. strain ADP.. ........................................................
2.4 Rhodococczis corallintrs NRRL B- 1 5 U R .....................................
Chapter 3 - MATERIALS AND METHODS ......................~...........-...-....-...-.
3.1 Bactenal strains and plasmids ..........................................-.~~--..--..-..
3.2 Media and growth conditions ..................................................--.....-
3.3 Characterization of bacteria ....................................................--......
3 -4 Analytical techniques.. .................................................-..........---......
vi
Page
1
* .
I l
. * .
111
v
vi
xiv

. . Atrazine minerahzatron ........................ .. ............................
High pressure liquid chromatogaphy (HPLC) analysis ......
Preparation of C 187 ce11 free extract (CFE) .......................
Detemination of rate of atrazine-degradation by cell free
.............................................................................. extract
Preparation of C 187 whole cells and determination
of the rate of atrazine-degradation .....................................
.......................................................... . 3 5 DNA manipulation techniques
................................................. 3 3 . 1 isoIation of plasrnid DN A.
3.5.2 Isolation of total genomic DNA .........................................
3 . 5 . 3 Preparation of DIG-labelled DNA ......................................
3.5.4 Recovery of DIG-labelled DNA
k a p e n t s from agarose .................... ............ ......................
- - 3 . 3 Dot blot preparation ..........................................................
33 .6 Hybridization and detection of DIG-labelled DNA
.............................................................................. probes
.......................................................................................... 3.6 tep-PCR
...................................................................... 3.7 16s rDNA sequencing
3.7.1 Preparation of PCR product for
....................................................... 16s rDNA sequencing
............................................. 3 J .2 16s rDNA sequence analysis

Chapter 4 - RESL'LTS .......................................................................................
4.1 Determination of diversity among atrazine-degrading
bacteria isolated from agicultural soils in Ottawa Ontario ................
4.1. I Assessrnent of biochemical diversity ...................................
. . 4.12 .Assessrnent of genetic diversity ..........................................
4.2 Characterization of C 187-an atrazine degrader from
St. Hyacinthe, Quibec. Canada .......................................................
4.3.1 Characterization of C 187 .............................................. ..-.
4.7.2 Determination of sequence sirnilarity with
Pseitdornonas sp. strain .4DP genes encoding atrazine
hydroiase and hydroxyatrazine ethylaminohydrolase and
with the Rhodococ~zî~ cora1liw.s 'IRRL B- l 5 W R
gene sncoding s-triazine chlorohydrolase ............ ............-.
4-23 'ulineralization of atrazine by C 1 57 ...... . . .. .. .... .-. - ... -. . - .-. . . -. . -
4.2.4 Degradation of atrazine to hydroxyatrazine by ce11
kee extract .... .... .. .. . ... .... . ... ... . ...... ..-. . . . . .. . . .. ... .. .-.... . .. .. . . . . . ...
4-23 DegradationofatrazinebywholrcellsofC1S7 ..................
Chapter 5 - DISCUSSION ........................................................... .....-... .-... .......-
5.1 Diversity and plasmid distribution in amine-degading
. . bactena isolated on a plot scale .........................................................
5.2 Characterization of a unique gram-positive bacteriun
from St. Hyacinthe, Québec .............................................................
. . . V l l l

. 5.2.1 IdentificationofC1S/ ..................... .. ............................. 7 5
5.2.2 Genetic and biochemical analysis of the initial steps
in the atrazine-degrading pathlva y. ..................................... 79
5 2.3 Regulation of atrazine degradation .............. ... .............. SZ
......................................................................................... REFERENCES
CLWCL!LLM VITI2 .............................................................................

LIST OF FIGURES
FIGURE DESCRIPTION PAGE
1. Chernical structure and properties of
- atrazlne.. ............................. ... .. 3
3 Structural and functionaI models of Photosystern iI 7 -, ..........
3. Proposed degadation pathways of atrazine
. . by sorl rnicroorganisms ..................................................... 13
4. Parhway for atrazine catabolisrn to cyanuric
acid in Psedomonas sp. strain A û P ............................... 15
5. Restriction rnap of Pset~rionionus sp. strain
ADP genomic DNA fragment cloned in
plasmid pMD 1. which contains the uc;l
and urrB senes .................... ... ....................................
Proposed pathway for the microbial degradation
of atrazine by Rhodococcrti cordlin~ls
NRRL B - l 5 W R ...........................................................
Dendrograms of the Ottawa amine-degrading
bacteria based on the biochemical responses to the
............................................. API 20E and ,srPI NFT test
Genomic fingerprints of anazine-degrading
bacteria isolated fiom Ottawa, Ontario, Canada
generated fiom ERTC-PCR. ............................................
X

Genomic fingerprints of atrazine-degading
bacteria isolated from Ottawa. Ontario. Canada
yrnerated Frorn BOX-PCR ............. ... ..........................
Plasmid profiles of selected atrazine-degading
bacteria isolated fiom Ottalva Ontario. Canada.. ............
'lucleoride sequence of the 16s rDNA jene
fiom Cl87 ......................................................................
Phylogene~ic tree showing the positions of C 1 S7.
rnembers of the genus iVocartiioides
and other related species .............. .... ..........................
Dot blot hybridization of purified DNA from C 187 with
DIG-labeiled nrr.4 and tznB probes ............... ..... ..........
Dot blot hybridization of purified DNA tiom C 157 with
.............................................. DIG-labelled tc.4 probe ....
Tram formation of atrazine to hydroxy atrazine
by CFE from C 187 when cells were g o w n
with atrazine and without atrazine, as demonstrated
by reverse phase HPLC analysis .......... ... .....................
Dissipation of anazine and formation of
hydroxyatrazine by CFE from C 187 .................................
Degradation of atrazine by Cl 87 whote cells as
demonstrated by HPLC analysis .......................................

Dissipation of atrazine. formation of hydroxyatrazine
and subsequent degradation of hydroxyatrazine by
C 187 whole cells ..............---...................-..-....--... . .-. . .. -. .-.

LIST OF T-ABLES
DESCRIPTION
.\nazine-degrading bactena used in this study ............................... 23
Plasmids and additionai bacteria used in this study ......................... 24
Primers used in this study 34 ............................................................
Biochemical responses of atrazine-degading bacteria isolated
tiom Ottawa. Ontario to .QI IOE biochemicai tesrs ...................... 43
Biochernicai responses of atrazine-degrading bacteria isolated
from Ottawa. Ontario to API >TT biochemical tests .............. .... ... W

ai
h p
* 4 i i S
bp
BCtP
CFE
cm
Cm
CFU
CC
Ci
Da
DIG
ABBREVTATIONS
- Percentage
- Degrees Centigrade
- Microgram (s)
- Microliter (s)
- Micrometer (s)
- Micromolar (s)
- Micromole (s)
- Absorbante
- Active Ingredient
- Ampicillin
Atrazine iMineral Salts
Basepair (s)
5-Bromo-4-Chloro-3- tndoly 1 Phosphate
Cell Free Extract
Centimeter (s)
Chlorarnphenicol
Colony Forming C'nit (s)
Cubic Centimeter (s)
Curie (s)
Dalton (s)
Digoxigenin
xiv

CSPD
JNTP
DN A
dLTP
DPM
ERIC
EDTA
EMBL
0 3
ha
HPLC
h
Km
kb
kg
LSC
L
LlMP
LB
- Disodium 3-(CMethoxyspiro { 1.2-Dioxetane-3.2'4 5-Chloro Tricyclo[3.3.1 .'.-IDecan! -4-yl) Phenyl Phosphate
- Deoxynucleoside Triphosphate
- Deosyribonucleic Acid
- Deosyuradine Triphosphate
- Disintegration Per Minute
- Enterobactenai Reperitive Intergenic Consensus
- Ethy lenediamine Tetraacetic Acid Disodium Salt
- European Molecular Biology Laboratory
- Hectare
- High Pressure Liquid Chromatogaphy
- Hour ( s )
- Kanamycin
- Kilobase (s)
- Kilogram (s)
- Liquid Scintilation Counter
- Liter ( s )
- Low Melting Point
- Luria-Bertani
- Milliarnpere (s)
- .Millicune (s)

min
$1
P P ~
PPm
Pen
pmol
nm
N
Nm
PV-
PCR
REP
- MilIigrarn (s)
- Milliliter (s)
- >Iillimeter (s)
- Millirnolar
- Minute (s)
- Mohr
- Mole(s)
- Nanometer (s)
- Nanomole (s)
- Nitroblue Tetrazolium
- Nutrient Broth-Yeast Extract
- Parts Per Billion
- Parts Pet MiIIion
- Penicillin
- Picornole (s)
- Nanometer (s)
- Normal
- Neomycin
- Pathovar
- Polymerase Chain Reaction
- Repetitive Extragenic Palindromic
- Repetitive Elernent Polymerase Chain Reaction
xvi

SSC
SDS
std, dev.
Sm
Su
Tc
Tra
Tn
TB E
Tris
TSA
G v
v
Vkm
v/v
w/v
- Resistant
- Revolutions Per Minute (s)
- Ribonucleic Acid
- Ribosomai Deoxyribonucieic Acid
- Ribosomal Ribonucleic Acid
- Sensitive
- Sodium Chloride Sodium Citrate
- Sodium Dodecyl Sulphate
- Standard Deviation
- Streptornycin
- Suppressor
- Tetracycline
- Transfer ( Conjugal)
Transposon
Tris Boric Acid EDTA Disodiurn
Tr ima Base (Tris [Hydro'rymethyl] Aminomethane
Tryptone Soy Agar
Ultraviolet
voit (s)
Volts Per Centimeter (s)
Vehme Per Volume
Weight Per Volume
xvii

1
Chapter 1. Introduction
Atrazine is the most widely used s-triazine herbicide for the control of annual broad leaf
weeds. About 36 x 106 kkg of atrazine are applied yearly in the Cnited States on crops such
as corn and sorghum (Mulbry, 1994). Atrazine was first introduced into Canada in 1960.
Current application rates are 1 to 1.70 kg a i h a depending on weed pressure. soil propenies,
climate conditions. and the crop planted (Ministry of Agiculture, Food and Rural
,4ffairs. 1996). The intensive use of atrazine in both the United States and Canada has led
to contamination of soil and gound water (Goodnch et al., 199 1 ; Belluck et al., 199 1 ; Frank
and Skons, 1985). Since atrazine may be a hazard to human health (Bouwer. 1989),
attention must be given to its fate in soil and to the chemical and biological processes
involved in its degradation.
In soil. biodegadation is the major pathway of atrazine dissipation (Erickson and
Lee, 1989). AIthough chemical degradation piays an important role. the present study
concentrates on microbial degradation. Initially atrazine was considered to be
nonmetabolized by most soil bacteria. It was no[ until the 1980s that microbial atrazine
degradation was proposed to occur by Y-dealkylation. resulting in the accumulation of
aminotriazine compounds (Behki and Khan, 1986; Giardina et al.. 1950, 1952;
Giardi et al, 1985). In the last 5 years pure cultures of bacteria have been described that c m
catabolize (mineralize) atrazine to CO, and amrnonia (Behki and Khan, 1994; Assaf and
Turco, 1994; Mandelbaum et al., 1995; Moscinski et al., 1996). Such bactena rnay be useful
for the decontamination of atrazine spill and waste sites. Atrazine degradation c m be
initiated by removal of the isopropyl and ethyl side chains. However recent studies

(De Souza et al.. 1995) indicate that degradation can also be initiated by hydrolytic
dechlorination. yielding the non-phytotoxic metabolite hydroxyatrazine. Funher degradation
ofthe ethylarnino and isopropylarnino side chains yields the metabolite cyanunc acid. which
can be used by many soit bacteria as a sole source of nitrogen (Cook. 1987; Cook et al.,
1985; Cook and Hütter, 1984; Eaton and Kams. 1991; Jutzi et al., 1952). Three genes
cncoding enzymes for atrazine catabolism to cyanuric acid have been cloned and sequenced
€rom the ritrazine-degrader Pseziï/olonronas sp. strain ADP (De Souza et al., 1996; Boundy-
Mills et al.. 1997: Sadowsky et al.. 19%). These genes. designated u c A BC. were shown to
be present in atrazine-catabolizing pure cultures isolated from sites throughout the world
(De Souza et al., 199s).
Atrazine-degrading bactena have been isolated from Canadian (Behki et al.. 1993;
Behki and Khan, 1957; Shao and Behki, 1996), American (Assaf and Turco, 1994;
Mandelbaum et al.. 1991) and French (Bouquard et al., 1997; Topp et al.. 1998) soils
previously exposed to atrazine. Recently our lab has isolated two groups of atrazine-
degrading bacteria (gnm-positive and gram-negative) from diverse soils in the agicultural
regions surrounding Ottawa, Ontario, St. Hyacinthe, Quebec and Winchester, Ontario (Topp
et al.. 1997). The two main objectives of this thesis are to deterrnine the divenity of the
gm-negative atrazine-degrading bacteria from Ottawa and to characterize the initial steps
involved in atrazine-degradation by a gm-positive aû-azine-degader from St. Hyacinthe.


Figure I . Chernical struct~ire of the a-icuitural herbicide atrazine i 2-cliloro-4-
ethy lamino-6-isopropylamino-s-tnazine) ( Spencer. 1 087).


Figure 2: Structural (-A) and hnctional (B) models of Photosytem 11. rr. a tyrosine
residur acting as an zlectron donor to p 6 W . the arrows show thr: direction
of sicctron tram fer: OEC. oxygen-svohng cornpiex: p68li. PSI1 reaction
cenrre: phaeo. phaeophytin a: Q., and Q,. quinones: PQ. mobile plastoquinonc
pool. Figure modi tied kom Cobb ( 1 YI?).


s
and mobility in soi1 (Topp et al.. 1W5L Atrazine is classifïed as a moderately persistent
herbicicic nith a hrilf-Iik rringing tiom 15 co 20 days in estuanne sediments to sc\.eral
months. up ro a year. in soi1 (Jones cr al.. 1983. .*y atrazine rcmaining in thc soi1 may be
triinsportrd into groundwater. surface waters. streams and tile drainage via surtace ninoff and
~roundwatcr intsrtlow. Jones et al. ( 1W3) reported that approsimately O. 1"% to 3% of the - atruinc placed on agicultural ficlds is iost to the aquatic environment. In many cases the
lzvrls o t'atrazint detectrd in gound watzr. surface w t s r s and jtreams esceeds the masimilm
contaminant Ievei sct by the Cnitcd Statcs Environmental Protection Agncy of 2 pans per
billion ( ppb) ( Ktllo. 1 'IS9). -4trazinc hÿs also been detrcred in the atrnospliere and min watcr
(Nations and Halberg, 1992). The solubiiity of atrazine in wiiter at 25 'C is 33 mgL. Once
airuine and othcr pesricidrs reacli pundwater and aquifers t k y c m persist t'or many yean.
For those who use goundwatsr as the sole source al'drinkiny watcr this can prcseni major
hralth prohlcms such as cancer. nen-ous systrm disorders. birth drfects and male strnlity
(Bouwer. 1089). In lin attrmpt to detoxify contaminated aquifers expensive physical and
chsmical treatment plants have bccn devrloped to rcmovc the contaminants at the whole
h o u x point of zntry (Goodrich ct al.. 199 1 j. Although trcatrncnt plants arc necessa-
(because ofprevious conramination). prevcntativc methods arc prcfcrrcd sjnce tliey are less
expznsivc and long tem. Consequently. there is considerable inrerest and need for the
dcvelopmsnt oT agicultural mamyement pncticas that minimize the potential for atrazine
to enter goundwater and surface water. One such application being studied is the addition
of organic amendmrnts. such as manure. to the soi1 in an attempt to alter the rate ot'atrazine
degndation. Topp et al. (1996) and Entry and Emmingharn ( 1995) both found that the

9
addition of Jairy manure to tields cropped to corn increassd the rats ofatrazins degradation.
[n order ro devclop iurther practicrs. characterizarion of atrazine degradation in the soi1 is
essential.
2.2 Atrazine-deyradation.
In the soi1 citrazine can be degradcd non-biologically and biologically. Non-
biological degrridation can occor \via photodccornposition. volatilization. soi1 ~dsorption. and
b y two soi 1-associarcd chernical rwctions: hydrosy lation and deal kylation ( Jordan ct al..
1 970). Although these processes are occumng. biological cisgadat ion. speci fically microbial
degradation. is the major pathway for atnzine dissipation in the soi1 iErickson and Lee.
19S9) . While çarlicr studies only reponed atrazine degradarion by rnixed rnicrobial
consoniü. more recent repons have indicated that several pure microbial srrains can degrade
atrazine and cise i t as a source of nitroyen and'or carbon ro suppon yrouth. Tliese strains
include Rhorioc*occirs strain B-30 ( Behki and Khan. 1994). Rliodococczts sp. strain NIS6iZ1
( Nagy et al.. 1995). Rlrodococcw sp. strain T E 1 ( Sliao and Behki. 1993). .-lci~zerob~rcrer
~dco~ierictts ( Mirgain et ai.. 1993 ), a new bacterial species closely related to .-lgrohacrericmr
i-citiiohmtcr ( Radosevich et ai.. 1 3 V o r i i Giardina ct ni., 1980. 1982:
Giardi et al.. 1985). and several strains of Pseltdonloms including Pseltdomorzus sp. strain
ADP (Mandelbaurn et al.. 19953. It is believed that atrazine c m be dejraded in the presencc
or absence of oxygen. depending on the organisrn in question. For instance. De Souza et al.
( 1996) showed that Psezrdomonus sp. stnin A D P could degrade atrazine under both aerobic
and oxygen-limiting conditions, while Behki et al. ( 1993) demonstrated that strains of

I O
R/~othcoc.cus could only degrade atrazins under aerobic conditions.
TIic tirst merabolic srep in the biodegradation ofatrazine has been sugsested to be
ï-dealkylation. wirh remocd of the cthyl sidr chain prcceding rernoval of the isopropyl sidc
chain i Radosevich st al.. 1995 ). Howvcr. Behki and a i a n i Ic)S61. reporteci that sorne
species of Pse~idoi~zomrs dralkylate the isoproppl yroup tirst. in eirhrr case the
microoganisms are using the olkyl side choins 3s a carbon source. Man)- microorganisms
do not fiirther metabolize rhe cierilkylated met3bolites. Sincc .V-dcalkylation is not sufficicnt
to desrroy the phytotoxic p r 0 p e ~ k of atrazine and the rnctaboiitcs may have unknow-n
effects on mimals and othcr organisms ( Kaufinan and Blake. 1970: Kaufman and Kramey,
1970) funher degradation is desirable. Additional depda t ion of arrazine metabolites by
Jeamination. cicchlorination and'.or ring cleavage i mincralization) lias been ubsewed in some
species i Boundy-Mills et ai.. 1997). Howcvcr. dechlorination to producs hydrosyanalojs
is suficient to dcsiroy phytotosicity ( Mandelbaurn ct al.. 19%). While the mechanism for
chcmicril dechlorination by hydrolysis is well undcrstood. the mechanism and rolc of
rnicrobial dechlorination (by hydrolysis or osygenic dcchlonnation) is less clear (Behki and
Khan. 1986). Cntil recently ir was believcd that soi1 bacteria ciid not have the snzyrnatic
systems to hydrate the aromatic carbon-halogen bond. Howevrr. recent studies of atrazine
dejradation by .-lgrohocreritrrn J I Jri (Moscinski et al.. 1996). Ralsroniü SA9 1-3
(Radosevich et al.. 1995), Hhodococc~rs sp. (Shao and Behki. 1995; Shao ct al.. 1995).
Rliizohirmi sp. (Bouquard ct al.. 1997). and Pselrdonionrir sp. s tn in ADP (De Souza et al..
1996; .Llandelbaum et al.. 1993. 1995) have indicrited that microbial dehalogenation is
occurring and in Fact may be the îïrst step in atrazine biodegradation by Psercdonioncls sp.

I I
strain ADP.
Cornpicte mineralization of otrazine occurs by clravage of the s-triazine ring. Dircct
attack on atrazine to sleaw the s-triazins ring has not been reported to date. How-ever.
studies by Cook et ai. ( 19S5). Ericltson and Lee i 19891. Radosevich et al. (1995) and
\Iandeibaurn et al. ( 19931 have shown that some organisrns. including soms ritrazine-
degraders are catabolizing the metabolite cljanuric acid to carbon dioxide and arnrnonia.
Althoush the exact pathwa'; in mazinc-dcgraders is unclear- i t has been hypotheslzed that
cyanuric acid is clcanxi cnzymatica11y to produce biurct and carbon Jioxide. Biuret is thcn
coniw-ted to urea. n-hich is dcgraded to the final products of mineralization. ornrnonia and
carbon dioxidc ( Figure 3 ).
2.3 Pseudomonas sp. strain ADP.
F'seucfonto~~cls sp. strain ADP is a gram-negati r-è ritruine degrader that was isolatsd
(rom a herbicide spi11 site in Minnesota. CS.;\. r Mandelbaum et al.. 1995). Pserdoi~roiras
sp. stmin ADP is capable of mineraiking the r-tnazine ring of atrvine via the intermediates
hydrosyatrazine and c~~anuric acid. in doing so i t uses ritrazine as its sole nitrosen source.
Most of our current undsrstanding of the genes and enzymes involved in atrazine
degradation cornes tiom studies of Pserdonzonu.~ sp. strain ADP. The enzymes for atrazine
catabolism to cyanuric acid (by Pserrdonio~ius sp. snain ADP) are encodcd by the genes utz.4.
rlcB and rrïzC. which are located on a self-transmissible plasmid (De Souza et al.. 1998).
The tirsr gene in the degradation pathway. c ic .4 . catalyzes atrazinc dechlorination to the non-
phytotoxic metaboiite hydroxyauazine (De Souza et al.. 1995) (Figure 4). The urz.4 gene is

Figure 3. Proposrd degadation pathways of atrazine by soi 1 microor~anisms exc luding
P.seiidomorr~rs sp. strain ADP. 3. dechlonnation: b. .V-dcalkylation: c .
deamination; and ci, ring c l s w a g .

Atrazine -
Deethylatrazine H ydroxyatrazine Deisopropylatrazine
Deeth yl hydroxyatrazine Deethyldeisopropylatrazine De i~~pr~p!~ lhydr~~yat raz i f le
Deethyldeiso propyl hyd roxyatrazine (Ammeline)
Cyanuric acid
Biuret
U rea
COz+ NH,

Figure 4. Pathway for atrazine catabol km to cyanunc acid in Psezctlon~o~im sp. strain
ADP (De Souza ct al.. 1998). L i r r l . ~ i r A 3 . and 'icC encods the enzymes
a i ru ine chlorohydroiase. Iiydrosymxzinc cthylarninohydrolase and Li
isopropylarnmelide isopropylaminohydrolase respectively.
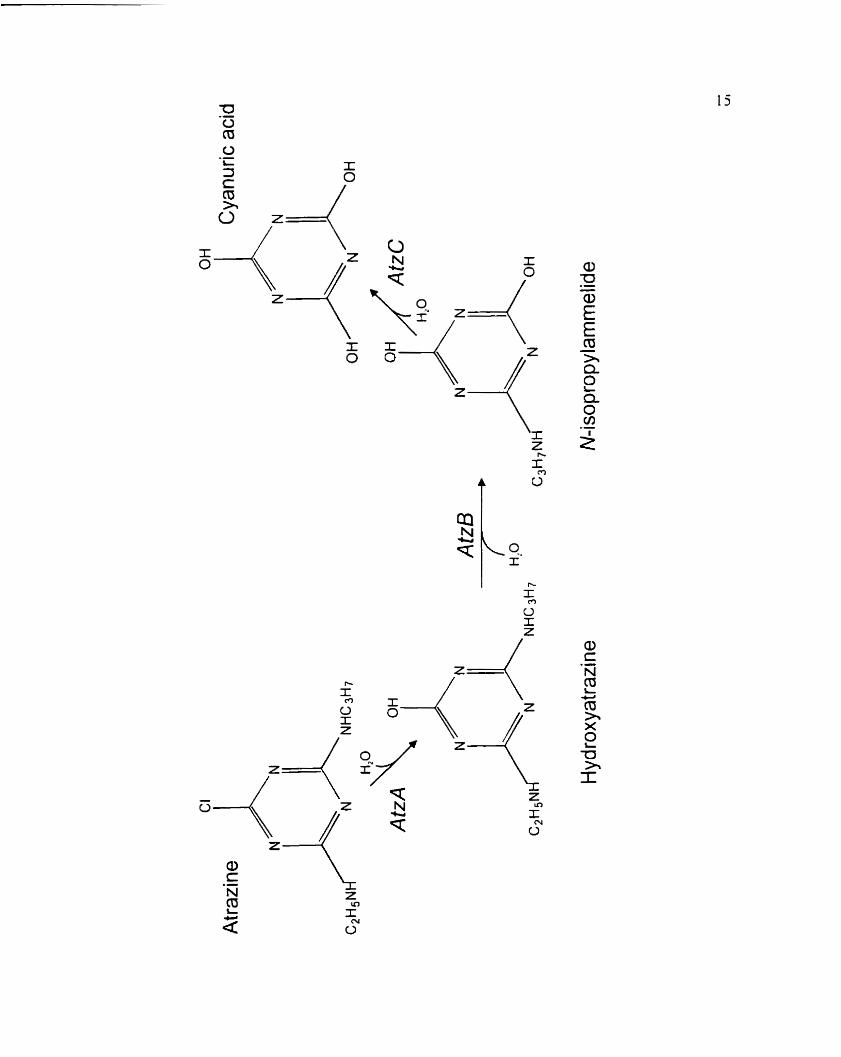

16
located on a 2 l .j kilobase ( k b ) EcoR l fragment. This fragment. ~vhich was cloned into
pLAFR3 and designatcd pMD 1. cncoded atrazine degradation activity in E. coli DHju
( Figure 5 ) ( De Souza et al.. 1995 ). Atrazinc degradation was dcmonstratcd by a zone of
cleariny on agar rncdium containing crystallinc atrÿzinc. .A Scnc con ferring the atrazine-
clraring plienotype \vas s~ibcloned as a 1.9 kb .-fiuI fragment in pXCYC 1 8-1. which was
dcsignatrd pMD4 (Dc Souza et al.. 19951. The same 2 1.5 kb EcoRl hyment also contained
thc second gene in the degradative pathway. utzB t Figure 5 ) . <icB is located 8 kb
downstrcam o f ~ r 1 z - f and encodes the enzyme hydroxyatrazinc cthylarninohydrolase. which
rrmsfomis hydroxyatnzins to .V-isopropylammelide tBoundy-Mills et al.. 1997) (Figure 4).
trcC: thc third gene in the dcgradativc pathway. encodcs the enzyme Y-isopropylamrnelide
isopropylaminoliydrolase ( AtzC) which transfoms X-isopropylarnmclide to cyanuric acid
and isopropylaminr (Figure 4). rrcC has been cloned and cxpressed in E, coli DH5u
(Sadowky et al.. 1998).
Rrcent studics by De Souza et al. ( 1998) h a ~ e shown that ti\-e geographically-distinct
atrazine-deyrading bacteria (.4lculigmes strain SG 1. .Agi-oh~icrer-hnr strain J 1 Ja, Strain 3Si38,
R~rlstorritr strain M9 1-3 and Clasihcrcter-) contained genes homologous to nc.4. -B. -C. This
suggests that atrazine-catabolic genes are conserved in diverse bacterial genera thereby
making Psercdomoriirs sp. strain .4DP an ideal mode1 system for the study of atrazine-

A, Restriction map of Psezldornonas sp. strain ADP smomic D.N-4
hagnent clonrd in plasmid pMD 1. which contains the uc.4 and L I ~ B jenes.
B. Rsstnction rnap of subclons pATZB-2. -4 4.0 Kb Chi frrigmcnr w s
subcloned into p .KYC 184 to produce pATZB-2. This figure is kom the
journal article published by Boundy-Mills st al. i 190-) and used n i<h their
permission.


t .4 Rhodococcus coralliri ris X RRL B- 1 5444R.
R . cor-crliirtrrs N R R L B- 15444R is a gram-positik-e coryneform bacterium that \vas
obtiiincd tiom soi Is tliat were exposrd to higli Isvels of s-triazine herbicidzs for '0 years.
R. ru)-dliiriis NRRL B- 1 5 - W R is capable of uti lizing deisopropy laminoatrazinc and
destliylaminoatrazine but not atrazins (or simazine) as a solc nitrogcn source for growth
(Cook and Hütter. 1986). Deisopropylaminoatrazine. also called deethylsirnazine is a
product of aerobic atrazine and simazine degradation. whereas deeth y laminoatrazine is a
product of atrazine degrcidation only. R. coruilorrrs NRRL B- I j - I U R has the ability to
dechlorinate md deaminate deisopropylaminoatrazine (and deethylaminoatrazinc), both
aerohically and anasrobicnlly. to the metabolites .V-ethy 1-ammclinc and :V-cthy 1-ammelide
respectivrly (Figure 6 ) (Cook aid Hütter. 1986). The enzyme for deamination lias not been
isolated ro h i e . Cook and Hüttcr (19S6) purifid 2 hydrolases (180 000 Da. and 450 000
Da.) lrom R. coi-riiltirru NRRL B - I 5 4 U R which wcrc capable of dechlorination of
deisopropylaminoatrazine (and deethylaminoatrazine). However. Mulbry ( 1994) in an
attrmpt to reproducs the exprriments was only able to purify onc hydrolase composed of
four identical subunits of 54 000 Da. The enzyme. s-triazine hydrolase is encoded by the
sene r i z 4 (Shao et al., 1995).
Although atnzine is not a substratr for s-triazine hydrolase. gene probes for the gene
sequence would be useful for isolatinj reiated atrazine degradation genes from other
microbial isolates. In addition. manipulation of the genes responsible for degradation may
yield enzpat ic activities that do attack atrazine.

Figure 6. Proposed pathway for the microbial degradation of deisopropy latrazine b y
Rlto<lococctcs cor-dliir iis NRRL B- l54WR ( Mulbry. 1 994). Dec hionnation
of deisopropylatrazine occurs via rhc snzymc s-tria~ine hydrolase and is
encoded by try.4.

Deisopropylatrazine

Chapter 3. Materials and methods
3.1 Bacterial strains and plasmids
Thc bacterial strains and plasmids usrd in this study arc listed in Tablcs 1 a and 1 b.
Thc iitrazine-degadiny bacteria from Ottawa were isolated from soils described by Topp et
ai. ( 1996). C 1 S7 was isolatcd from soi1 ncar St. Hyacinthe that u s cropped with mixed
ccrcais and ne\-rr trcated with litruine. Major soi1 properties are as follows: texture: loarn.
3S0,1 sand. 30" (1 silt. 32"0 clay. I .-ln.n organic rnatter. and pH 6.
3.2 Media and orowth conditions
AI1 atrazine deyraden were g r o m at W C on atrazine mineral salts (.;LLLS) ( 1 -6 L
K2HPOJ. 0.4 SL KH,PO,. 0.2 g L MgS0,-7H@. 0.1 NaCl. 0.03 CnCI-ZHZO and 1 mL!L
trrice elements [IO mg. L ZnS0,-7H,O. 3 mgL MnC1,-4H,O. 30 m g L HjBOI. 20 m y L
CoC1,-6H,O. 1 rngL CuC1,-2H:O. 2 rn*L - 'iiC1,-1H,O. and 3 m-L N~LVOO,-LH~O
(modified from Lapage and Mitchell. 1 W O ) ] ) medium (modi fied from Mandelbaum et al..
1993) supplemcnted u n i t 1 1 citrazine (500 m j in 2 mL methanoliL). sodium citrate dihydrate
( 1 g L). Lynch vitarnins ( 1 mL, L ) (Lynch et al., 1980) and FeS0,-6H20 (5 mg/L). Lynch
vitamins and FeS0,-6HI0 were filter stenlized (0.2 um acrodisc. Gelman Sciences) and
added to cooled, autociaved media. The degradation of atrazine by bacteria was indicated
by a zone of clearing surrounding the colonies. P. syriiigue pt . [onzc~to "as gro\vn on Kings
B medium (King et al.. 1954) at ZYC. E. coli strains were grown on Luria-Bertani (LB)
agar (Sambrook et al., 1989) at 30' C. When requircd. chloramphenicol (30 @mL) and
kanamycin (50 pJrnL) were added to the medium. Clmibacter nriclzigarie,isis subsp.

Table 1 a. Atrazine-degrading bacteria used in this study.
S train number Geographical origin Source rind/or re ferencc
C 147. C 1%. C 1%. C 1 OO. Ottawa, Ontario C163. C17-L. C175. C176. C177. C l X C179. Cl80
Pserulonzoti~~s sp. strain .AD P
St. Hyacinthe. Quebec
Winchester. Ontario
Minnesota. US..\.
E. Topp: Topp ct al.. 1996
E. Topp
E. Topp
blandeibaum et al.. 1995

Table 1 b. Plasmids and additional bacteria used in this study.
Strain! plasmid Relevant characteristics Rcferencs and or source
Bacteria
DC3000 spontaneous Ri P derivatiw of NCPPB 1 106.68-kb
E. coli
host E. cok 5 3 . 146-kb, Tra' . h p r Km' Penr Su
D. Cuppels. 1'186
BRL
J . Dick
E. Topp
Shao et al.. L995
host E. coli CVASO3, Selvaraj and lyer. 30-kb, Cmr Kmr, TnS donor 1983

Table Ib continueid. Plasmids and additional bacteria used in this study.
S train/ p lasrnid Rele~mt characteristics Rzference andor source
Plasmids
pRK70 1 3 host E. coli HB I O 1. -CS-kb. Km' Nm' Tr3'. mobilizins factor. hcipcr plasrnid
host E. coli 55-3. 93-kb. Km' Ampr Smr Su Cm'
host E. coli D H k . vcctor pXCYC 1 84. MI-4
host E. coli DHSu. vcctor p X Y C 1 84. 'nrB
Boiindy-kIilIs ct al.. 1 9'17

36
/triclrig~irre~rsls JDS3- I \vas grown on t y t o n c soy agar (TSX) ( DIFCO. Detroit. MI) at room
tcmperature. For long terni storagc bacteria were suspended in 30°G glycerol or nutrient
brotli-yeast -tract <NB)-) (Vidin-er. 19671 brotli containing 15% ylycerol and stored at
-70 - C.
3.3 Characterization of bacteria
The Gram stain reaction \vas iised to discriminate between gram-positive and
gram-nryatiw atrazinc dryraden. .Al1 atrazine-degadins strains wsre grown on XVlS agar
supplsmentrd with atrazinc for 6 days at W C . The sram-positive control. Cim:ihacier
nrichigmeiuis subsp. niichtgmlemïis JDS3-1. uas grown on TSA for 72 h at room
temperature. The gram-ncgativc control. Pserrdotuoii<is siet-i~tgtre pv. rorrimo DC3000 was
crown on hB\i agar m room temperature for 72 h. The cells were Gram-stained according Cr
to tlie nianuf~cturer's instructions (Signa Cliem. Co.. St. Louis. Mo.). Cslls which retaincd
the p n m q dyr (crystal violet) u w e considered ,mm-positivc. Cclls which lost the pnmary
dye during the decoloration step were considered gram-negative.
The potassium hydroxide (KOH) test (Suslow et al.. 1952) was used to confirm the
Gram-reaction. .A drop (50 uL) ot'jO/o (wi'v) aqueous potassium hydroxide was placcd on
a clean siide. Bacterial cells were transferred to the KOH with a stcrilc wooden toothpick-
The cells were mixed thoroughly on the slide. After sevcral seconds. the toothpick was
alternately raised and lowered in order to detect a stnnging effect. The KOH test was
considered positive if the viscosity increased and stringing occurrcd within 15 seconds. A
positive rcaction is typical of gram-negative bacteria.

-4trazinc-degadins bacteria were testrd for cytochrome c ovidase acti\:ity according
to the procedure describsd by Smibert and Kneg ( 198 1 ). -411 strains. including the negativs
control. E. coli. azre y0a .n on NB)- ayar for 72 h nt 30 'C. Psrtr~lonioi~~~s~liro,.esceiis. the
positive control. \vas yrown on hB1- aga . îbr -IS h at 25 'C. Bacterial colonics were smeared
tvith a stenle loop ont0 \Chatman 3 C N filter paper rnoistençd rvith I o , ) ( w L-) trtrmeth-1-p-
phenylenzdiarnine dihydrochlonde (Sigma). Dcvsloprnsnt of a purple color wirhin 30
seconds was scored as a positivc reaction.
Substrate utilirarion w s cletetmineci for stnins C 147. C 155. C 156. C 160. C 163.
C 174. C 1 77. C 175. C 179 and C 1 SO usiny thc .et X E and .-\PI ';FT stnps ris recommmded
by rhc manufacturer ( .AP I Laboratory Products. Ltd.. St. Laurent. QuGbec. Canada). Samples
n-erc incubated at 30 'C and rcad at 24. 48 and 72 II.
3.4 .Inalvtical techniques
3.4.1 Atrazine rnineralization
Steri le A M S supplemsntcd with uni fomily nnj-labrllsd [lJC]atrazine
(50000 disintegrations pet minute I D P W ~ L ) (specific xtivity 4.5 mCi. mmol. radioactive
puri- 95%. S i l a ) \vas cidded to sterils microtitre wells i 200 u L well) (96 WeIl Cc11 Culture
Clustcr. Costar Corporation). tndividual \vrlls were inoculatcd wirh one colony of C 1 S7 or
Pserrrloniotlus sp . srrain ADP (positive conrrol). The remainin~ wells were supplied with
stcrile distilled water to maintain humidity levels. The microtitre plate was incubated at
3O'C. in a Zipiock bag for 48 h. .A Falcon tube (Fisher Scientific Ltd.) containing 1 mL of
1 'I XaOH was addrd to the Ziplock bas as a I4COZ trap. Samples (50 uL or 100 u L ) were

2s
takcn after 24 h and 48 h. The total radioactivity remaining in the samples \vas quanti fied
by Liquid Scintillation Counting ( LSC) (Beckman LS 580 I . Beckman Instruments Inc..
[mine. CA) iising I O rnL of CnivsrSol cocktail KN. Costa Mesa. CA) and corrected for
quenching tvith an cxtemal standard.
3.4.2 High oressure liauid ch romoto~raohv (HPLC) analvsis
Atrazine disappcarance and metabolite formation \vas determined using HPLC
analysis (Topp et al.. 1993). Microtitre plate wells u-ere fiilcd with 200 PL of .LMS
containing 92-76 UM (20 mgL) atrazine and the wells were inoculated with C 157. C 147. or
Psezrdonzo~rtzs sp. strain ADP. The microtitre plate was incubated at 3O'C overnight. .\fier
incubation. 100 u L samples were addcd to an equal volume of methanol and centrifused at
15 400 .u g for 4 min at room temperature to remove cslls and othcr insoluble matenals-
HPLC analysis of the supcmatant was perfomed with n U-atcrs 5 1 O Liquid Chromarogaph
Systsm equipped \vit11 a Waters 490 programmable-wavelength detector. Atrazine and its
metabolites were resol\-cd by using an analsicd C,, reverse phase CVhatnan HPLC column
(4.6 mm x 250 mm. 10 um diameter. irrejular packing). The opcntin_o conditions were as
follows: injection volume. 40 PL: drtector waveizngth. 210 nm: mobile phase. 7096
CH ,OH. 30% 5 mM 'iaH,PO, (pH 9.0) or 30% CH,OH. 5096 10 mk1 ammonium acetate:
flow rate. 1 mLimin. Identification of cornpounds \vas detemined by cornparin: retention
times with analytical standards. Concentrations (mg/L andor uM) were determined by
comparing peak areas with peak areas of known concentrations of analytical standards.
Resultant chrornatop.rns were generated using the sohvare prograns Lotus 1-14 97 (Lotus

' 9
Devsloprnental Corporation. Cambridge. MX) and Origin (Version 4.1 O. Microcal Sofnvare.
Inc.. 'lorthampton. M A ) .
3.4.3 Preoaration of C 187 cell free extract (CFm
Mid esponential-phase crlls grown in sither NBY broth containing 5 9 L jlucose and
1 S j 3 2 UM (40 mg.! L) atrazine or ' IBY broth containing 5 5 L glucose were han-csted by
centn fugation ai 12 300 s y for 10 min at 4-C and ivashcd tlvice in I O mL of 10 mM sodium
phosphate buffer (pH 7 . 2 ) (Topp ct al.. 1993. The cclls were resuspended in fresh bufler
to an A,,,,, of 4.5 (4.5 s iO" CFC1mL,. Cnide ceIl extracts Lvere prepared bp sonicating the
cells for ;i total of 1 O min ( 2 min. biirsts. 1 min. rest intenals) using ;i Braun-Sonic 2000
ultrasonic Iiomogenizçr (intermediate 13T titanium probe tip [3.:SW]. low powcr position.
approsimately 60 Watts. B. Braun Instruments. Burlinyame. C.1.j. The supernatant s a s
clanfied by centrifugation at 12 300 s y for 10 min nt 4 'C. The cmdc cxtract was kept on
ice and used immediately. To determine the minimum sonication time required to obtain
maximum protein. cclls t 4.5 s 10" CFC:m L ) w r e sonicatcd for O. 2. 4. 6 , S. 10. 12. 14 and
1 6 min. After rach time inrenal 100 uL samples were removed and cinalysed for protein
concentration. A 7 mL simple of cmde cxtract \vas removed and autoclaved (5 min. 121 ' C l
.Auroclaved CFE (200 u L ) and non-autoclaved CFE (100 u L ) \vas stored at -10'C for protein
mal ysis. The remairtins CFE \vas used io determine the rate of atrazine degradation (Section
3 -3.4).
The protein content was assayed using the Bio-Rad Protein Micro hssay (Bio-Rad).
Dilutions of protcin standard (bovine semm albumin) containing 3 to 18 p g m L were

30
prepared. CFE set aside for protein analysis was diluted 10-fold (SO u L sarnple into -20 u L
srenle sodium phosphate buffer [pH '2. 10 mM]). .-\ 0.8 mL aliquot of standard. diluted
sample. or sterïle sodium phosphate butEr (blank) (pH 7.1. I O mu) w;is miscd with 0.2 mL
of Jye rengcnt concentratc (Bio-Rad) m d lightly vortexed. .-\lier 10 min the A;.,I
detrrminsd for a c h sample using ;i Beckman DU 6-10 spcctrophotometsr (Beckman
Instnirnenrs Inc. ). -4 standard ciin-e plotting .AI,,, and standard protein concentration (3 pg-
I 5 ,LE) was prepared rach time the assa' \vas performed. The protein concentration of each
sample was cietermined from the standard curve.
3.4.4 Determination of rate of atrazine deuradation hy ceil free estract
Cd1 free extract was prepared as descnbed in section 3.4.3. Cell iiee extract and 10
mbl sodi~im phosphate buffcr (pH 7.1) containing 92.76 UM (20 m g L ) strazine were
prehearcd to 30-C. One rnL ol'CFE \vas mised with an equal volumc oisodium phosphate
butTcr ( 1 0 mhl. pH 7.2) in a 3 - m L sçnirn bottls. The reaction mixtures wsre incubated at
30 -C with shaking ( 1 50 rpm ). 1 00-uL samplcs a w c removed at 0. 1 0. 30. 30. -10. 50. GO.
90. 120. 151). 11 0, 271) and 1440 min intemals. and mixed with 100 u L of 95'6 methanof.
The samples were ccntnfuged at 15 400 s g for 4 min at room temperaturc. Xtrazine
dissipation and metabolite formation werc rcsolved using re-wrse phase HPLC analysis as
described in section 3.6.3. Caiculations of rates and rate constants were bascd on the
assumption that atrazine-degradation followed f i n t order kinetics. Exponential c w e fitting
was used to determine first order rate constants. The derivative of the decreasing exponential
at T=O was also used to demonstrate the rate of atrazine-degradation. Each rate was

3 1
normalized based on the mount of prorein in the reaction. Intercxpenmenral \-ariabilitp was
determinrd iisiny .WOV.\ (Statistica. Version 4.5. Tulsa. OK). Significant differences
benvecn treatments i \v ith atrazins \.ersus wi thout atrazine) w r e detennined using the paircd
t-test (Statistica. Version 4.5. Tulsa. OKA.
3.4.5 Preoaration of Cl87 whole cells and determination of the rate of atrazine-
de~radation-
Cclls growiny cxponentially in NBY broth containing 5 JL glucose and I S5 ULM (40 m2L)
atrazine were hm*ested by centrifugation at 12 300 x y for 20 min at 4-C and washed twice
in 1 O mL of 1 O mM sodium phosphate hutrer ( pH '3. Tlic crlls were resuspended in fresh
bufier to an .i ,,*,, o f 4.5 (4.5 Y 10" CFC mL). Three millilitcr samples of the cells were
preheated to 3O'C anci placed into 4 25-rnL serurn bottles. The scntrn bottles w x e scaled
wich geey butyl nibber stoppers md aluminurn cnmp tops. In order to crcatc m osyyen -kc
environment ovygen \vas evacuated under vacuum from nvo o f the sealcd s e m bottles and
backfiilcd to iitmosplieric pressure with nitrojen gas iising a r-acuum desiccaror. Three
rnilliliters of osyyen-tiee sodium phosphate buffer ( 1 O mM. pH 7.2. preheated to 3O'C)
containing 97.76 uM (20 myL) atrazine \vas added to al1 4 serurn boitles using an osygen-
Cree 10 cc alerile syringe. The reaction mixtures were incubated at 3O'C with shaking ( 1 50
rpm). Samples ( 100 yL) were removed at 0. 10. 20. 30.40. 50. 60.90. 110. 150. 11 0. 270
aiid 1440 min. mixed with 100 u L of 959th methanol and centrifuged at 14 000 rpm for 4 min
at room temperature. Atrazine dissipation and metabolire formation were resolved using
reverse phase HPLC analysis of the supernatant as descnbed in section 3 - 6 2 Data analysis

33
was performed usiny the proccddures descnbed in section 3.4.4 sxccpt that rate constants and
rates ivere nomalized based on the CFL mL rather than thc protein mnccntration.
3.5 DNA mani~ulation techniaues
3.5. f Isolation of ~îasrnid DNA
Cncut plasmid DNA was isolated from strains C 1-17. Cl 55. C 156. C 160. C 1 b3.
C 177. CI 7s. C 1 3 . and CI SO ticcording to the alkaline Iysis rnethod of Kado und Liu ( 198 1 ).
Atrazine degraders wrre grown on NBY agar for 73. h at 30 ' C. E. c d i strains wçre grorvn
on NBY agar supplernented with 50 p y m L of kanamycin for -48 h at 30 'C. One bactenal
colon- was picked Iiorn the plate. suspended (with gentle tlicking) in 150-uL lysing solution
i3'k SDS in 50 m'l Tris. pH 12.0) and then incubated for 60 min at 60-C. Tu the ceil lysatc
2 volumes of iinbuffercd phenol:chioroform ( 1 : 1 ) wxs added. Ccil &bris was removsù by
centrifugation at 15 400 .; 2 for 4 min at room temperature. Plasmid D'IX was visualized
by Iiorizontal gzl clectrophoresis in { J . j 0 i ultra pure agarose (Life Tschnoio~ies. Inc..
Gaithersburg. MD) at 32 V'cm for 19.5 h using O.5X TBE buffer ( IOX: 0.S9 M Tris. 0.89 .M
borïc acid. 0.01 .LI EDTA disodium. pH 8.4). Gels werc stained in dilute ethidium bromide
( 1 .U u g m L ) for 2U min. destained in distilled water for 20 min. and photogaphed under LV
light with a Polaroid camera (Kodak film. type F665. Rochester. YY).
3.5.2 Isolation of total genornic DNA
Punfied total genornic DNA was isolated from C 157. C 193. Psetulomonus sp. strain
ADP. and DC3000. C 157. C 192. and Pseirdomonus sp. strain ADP were Town on Abils

33
plus atrazins piates at 30 -C for 6 days. DG000 was yrown on Kings B medium at room
temperature Ior '2 h. Cells were waashsd from thcir plates wirh 2 mL of sterile distilled water
and placed in s t ede i 5 m L microfugs tubes. The suspensions were L-onsxed vigorously.
The cclls wrrc pelleted by centrifugation for I O min at U O O s g and the supernatants
discardrd. The pellets were then suspcndcd in ISO u L of lysozyme incubation but'fsr
(20 mgmL lysozyme. 30 mM Tris-HCl (pH S). 2 rn.M EDTA. 1 -39;) Triton) and incubated
60 min at 37'C. .A 35 ,uL aliquot of Proteinase K and 100 u L ofbuffer A L (Qiamp Tissue
Kit. Qiayen. Chatswonh. c.41 ivas added to çach and mised by uxtexing. Sarnples were
incubated at 70 -C for 30 min and 9 Y C for an additional 30 min. Total senomic DXA was
extractcd iising the Qiamp Tissuc Kit following the manufacturer's instructions (procceding
from step 1). Absorbancc values (.Alb, and .Azso) were obtained iisinj a B e c h a n D L 640
spectrophotometer (Becliman Insrniments Inc.). The A,,,, A,,, ratio for al1 samples was
behveen 1 .S and 1 -9. The final DNA concsntntion was calculated from the absorbante value
at 760 nm iising the rquation 1 A?,, = 50 pg, mL (Sambrook et al.. 1989).
3.5.3 Preparation of DIG-labeiled DNA
DIG-labelled probes were generated with the PCR DIG Probe Sythesis Kit
(Boehnnger Mannheim Biochemicals. Lavai. Québec). DIG-labelled probes mere generated
by incorponting DIG- 1 1-dUTP dunng a PCR according to manufacturer instructions
(Boehringer Mannheim Biochemicals). Custom internai primers (Table 7) were designed
specifically for ur1.4 and atzB with the software prograrns OLLGO (Ver 4.0. National
Biosciences, Inc., Plymouth, MN) and D N A i h Y (Version 2.5. Lynnon BioSott, Vaudreuil.

Table 3. Primers used in this studv
Primer Primer sequence* Region Rekrcncc and or amplitisd source
Dl
6SjrZ
530f
1 1 OOr
926f
l@?r
ERIC 1 R
ERIC2
BOXA 1 R
1 O7SF 1
1 S2iRZ
2358F1
2435R1
16s rDXA
16s rDN.4
16s rD3i.A
16s rDN.4
16s rDIi-4
16s rD'i.4
ERIC
ERIC
BOXA
tf l1.4
L I C Z . ~
a d 3
a c B
Lans. 1991
Martin et al.. 1992
This study
This study
This stud';
This study

35
Que.. Canada) and primcrs were synthesized by Li fc Technologies Inc. Xfter ampli fication.
ri 4-UL iiliq~iot of the 50-uL reaction mixture was analyzed by horizontal gel elec trophoresis
using 1. joo a;aros<r in 1 S TBE bufkr 3t 100 V For 45 min. The gel was stained brietly in
cthidium brornide ( 2 m g LI. destained and visualizcd under short ~vavclcngth UV light. The
vield of DIG-[abelled probe sas estimated by a colorirnetric assay with nitroblue tetrazolium
( NBT) ( Boehringer blannhcim Biochrmicals) and 5-bromo-4-c hloro-3-indol y 1 phosp hatc
(BCLP) (Boehringer Mannheim Biochemicals) as substrates for alkaline phosphatase
conjugated to anti-DIG mtibody. Scrial dilutions of the DIG-labelled probe were compared
n-ith serial dilutions of controi DiG-labclled DNA of known concentration. The DIG-
Iabellcd probe was stored at -10'C.
3.54 Recover>* of DIG-labelled DY.4 fragments from agarose
DNA bands werc cscised from agarose gels according to the msthods described by
Kutlu and Cuppcls ( 1997). DIG-labcllcd DNA wÿs electrophoresed in 1 % low-rnelting point
(LMP) a p o s e in IS TBE buffer at 60 Vkm for 70 min at 1'C. The gel was stained for 15
min in rthidium bromidc (2.0 m y L ) and visualized under short wavelength LW light. The
band of interest was escised. weighed and incubatrd with 0.04 volumes of 25X asarase
buffer (750 mM Bis-Tris. 250 mM EDTA) for 15 min at 65'C. The molten agarose was
cooled to 45 T and digested with 2.1 Units of DNAase-free agarase ( 1 Unit per 100 mg of
aiprose) at 45'C overnight. Remaining oligosacchandes were removed by addin;
0.1 volumcs of 3 M NaOAc (pH 5.5) and incubatins the solution for 15 min on ice. The
oiigosaccharaides were removed by centrifugation (1 5 100 x g for 10 min) at 1'C. DNA was

recovered from the supematant by cold ( -NT) ethanol precipitation.
3.5.5 Dot blot ~ r e ~ a r a t i o n
For preparation of dot blots (Cuppels ct al.. !990) whole ceIl suspensions were
prepared by suspending colonies that were gown on .\.LIS supplemcnted with atrazine. LB
plus kanamycin (50 ug,mL) or XBY agar in 500-uL of s t ede distilled water to an A,,,,,, of
2.0 (2.0 u 10" CFC mL). For soms dot blots. puri tied genornic DN.4 was prepared (Section
3.5.2) and used nrher than ivhols cells. Aliquots of whole cells O pi) or purified gcnornic
DNA (200 ng) were pipetted onto uncharsed 52 mm diameter nylon membranes for colony
and plaque hybridization (Boehnnjer blannheim Biochemicals). The cclls were lysed and
the DNA cienatured by placing the membranes on Uhatman 3mrn papzr saturatcd with
denaturation solution (0.5 N NaOH. 1.5 hf XaCl) for 15 min. The membranes wcre then
neutnlized by placing the membranes on Alter paper saturatsd wirh nrutralization solution
( 1 .O hl Tris[pH SI. 1 .j 41 NaCl) for 15 min. Aftcr being incubated for 5 min on filter paper
moistzned with 2X SSC the membranes wçre air dricd and placed in a L1; cross-linker
(LVP. Inc.. Lpland. CA). The membranes were storcd dry in a desiccator untii required for
hybridization.
3.5.6 Hvbridization and detection of DIG-labelled DiVA probes
Target DNA immobilized on nylon membranes was hybridized with a DIG-labelled
DNA probe (Cuppels et al.. 1990; Sambrook et al.. 1999). The dot blots were incubated for
2 h at J Y C in a prehybridization buffer containing 5X SSC. 50% deionized formamide,

37
O. 1 " A N-[auroyl-sarcosine. 0.07''L SDS. and 5% blocking reayent (Boehringer Mannheim
Biochemicals). Thcy were then tnnslerred to new bags containing 10 ng'rnL of heat-
denatureci probe in 3 mL of prchybridization buffer and incubatcd ovemight at 42 ' C . After
hybridization thc membranes were washcd twice. 10 min per wash. in 2 X wash solution
( 2 N SSC. 0. SDS) at room ternpsrature followed by two washes. 1 5 min per wash. in
prewarmed O.5X wasli solution (O.5X SSC. 0.1 ?& SDS) at 68 'C. The membranes wcre
incubated in Genius buffer 1 ( 100 rnM Tris. 150 m M NaCl. pH 7.5) for I min and then in
Genius bufkr Z ( 1 %[wh-] blocking rcascnt in Genius bu ffer 1 ) for 2 h at roorn temperature
wi th gentlc shaking. A Aer a 30 min incubation in alkaline phosphatase-conjugated anti-D IG
anribody diluted 1 : 10000 in Genius buffer 2. the blot was washed twice. 15 min per wash.
with Grnius buffer 1 and then equilibratcd in Genius buffer 3 for 2 min. The probe-target
DN .A hybrids Lverc detected usin$ the cherniluminescent substrate disodium
3-(4-rncthosyspiro ; 1 .I-dioxetane-3.2'-(Schloro) tricycl0[3.3.1.' 'ldecan: -4-yl) phenyl
phosphate (CSPD) (Boehringer ~Mannheim BiochcmicaIs). The membranes were moistened
with 1 mL of CSPD (diluted 1 500 in Genius buffer 3), scaled in hybndization bags and
rxposcd to X-ray tilm (Kodak X-Omat AR. 14" x 17". Richardson .Y-Ray, London, ON) for
45 to 120 min. To stnp membranes OF DIG-labelled probe they were washed nvice. 10 min
per wash, in 0.2 N NaOHiO. 1 % SDS with shaking at 37°C. The membranes were then rinsed
in 2X SSC and stored dry.

3.6 r e ~ - P c R
.\trazine-degadins bacteria isolated from Ottawa (Table 1 a) wcrc fingerprinted using
rep-PCR and the consensus primcrs BOXAI R and Enterobacterial Repetitive Intcrgenic
Consensus ( ERIC) 1 R and ERIC? (modified From Pooler et al.. 1996). Wlole cells of each
isolate were g o w n on TSA for 4s h. Single colonics were removed from the plates and
suspcnded in 1 mL of stede distilled water. The ceIl concentration rvas adjusied to an A,
of2 ( 2 s 10" CFU.mL) and used as remplate for PCR. The oligonuclcotide pnmers (Table 2)
were syntliesizrd by BRL based on published sequences ( Hulton et al.. 1992: Martin et al.,
1 ) . ERIC- PCR reactions wcrc carricd out in a 25 uL volume containing 9.6 uL sten le
viater. 2.5 ,pL 1OX buffcr (Promesa. -Madison. WI). 3 u L MgCl: (Promega), 2.5 pL sterile
gelatin ( l rny'rnL). 0.2 g L dNTP (25 mM), 1 pL each of ERICIR and ERIC2 prirners
(50 PM) and 0.7 UL of T C I ~ polynierase (Promepa). BOXA- PCR rcactions were cmied out
in a 25 u L volume containins 10.6 u L stenle water. 2.5 uL 1OX bufler (Promega), 3 pL
MgCI? (Promcga). 2.5 pL s tede gelatin ( 1 mymL) . 0.1 uL dNTP (3 miCl). 1 pL of the
BOXA 1 R primer (50 uM), and 0.2 pL of Tclq polymerase (Promcga). Each reaction tube
was covered with one drop of stcnlc minera1 oil. The amplifications were perfomed with
a H ybaid OmniGene Thermocycler (Interscience Inc., Markham. ON.. Canada). For both
the ERIC primers and BOXA primer (Table 1) the cycles used were as follows: 1 cycle at
94-C for lmin, 30 cycles at 9J3C for i min. at 52'C for 1 min. and ar 72'C for 1 min. ARer
amplification 4 pL of the BO1U and ERIC PCR products were separated by horizontal gel
electrophoresis in 1.5% agarose (Nusieve 3: 1 ) in 1 X TBE buffer (lOX: 0.89 M Tris. 0.89 M
boric Acid. 0.02 M EDTA disodium, pH 8.4) at 100 V For 45 min. A 1 pL aliquot of 1-kb

39
DKA Laddcr ( Li tè Technologies Inc.) was addcd to the first and last lane of each gel. Gels
\vere stained in dilute cthidiiim bromide ( 1 p ~ m L ) for 20 min. destained in distilled water
tUr 20 min and photographed under L1; liyht \vit11 a Polaroid camerci (Kodak film. type
F665).
3.7 16s rD.i.4 seauencing
3.7.1 Preoaration of PCR oroduct for 16s rDN4 seauencin~,
Strain Cl S7 was groum on . h i l S plus atrazine agar for 6 days. Cells were washed
from the plarcs with 2 mL of sterilc distilled tvater and placed in a stenle disposable test tube.
The cell concentration \vas adjusted to an A,,,,, of 2 ( 2 x 10'' CFUimL) and used as templatc
Ior PCR. PCR was carried out in a 75 u L volume containing 17 ,uL sterile distillcd water,
7 .5 u L 1 O S buffer (Promeja). 4.5 pL 41gClI (25 mM) (Promega), 0.6 uL 25 mM dNTP.
3-75 uL of rach opposing primer ( 1 O PM) and 0.1 uL of T.«q polymcrasc (Promega). Each
reaction mixture was coverrd with I drop of minera1 oil. PCR-fragments were amplificd
using a Hybaid OmniGene Thermocycler (Interscience Inc.. Markham. 03.. Canada) using
the following program: L cycle at 94'C for 3 min. 30 cycles at 94'C for 1 min
(denaturation), at 502C for 1 min (annealing) and at 72°C for 3 min (elongation); 1 cycle at
72-C for 5 min (final extension): and stabilization at 3O'C for 1 min. Published primcrs
(Table 2) specific fior the 16s rDNA molecule were purchased fiom Life Technoiogies inc.
The primers were paired together as follows: 926f-1492r; tD 1 -68jrI; and 530f-l l OOr. Three
PCR products of approximately 500 bp in length were obtained. The PCR products were
concentrated to a volume of 2 1 p L using Microcon- 100 microconcentrators (Amicon Inc.,

40
Bevsrlp. MA) 3s recommendrd by the manufacturer. Final DNA concentrations were
calculated liorn the absorbance values ar 160 nrn using the equation drscribed in section
3 - 5 2 The PCR products t 30 n j u L ) and corresponding primers ( 1.2s pmol.',uL) were sent
to The John P. Robans Research Instirute DNX Sequencing Facilit- (Cni\.rnic) of Western
Ontano. London. ON.. Canada) for sequencing (AB[ Prism Mode1 3 77 scquencer. Pcrkin
Elmer Applied Biosystems. Foster City. CA). Threr sequences wsre generatcd.
3.7.1 16s rDN.4 sequence analpis
For each strmd. the 3 srquencrs wrre mer@ into one 1462 bp sequrnce usinj the
so fhvarc program DX.AlM.4N (Version 2.7; Lynnon BioSoti. Vaudreuil. Qué.. Canada). The
seqlience was edited manuail- by cornparisons of the two strands to clirninate discrepancies.
Ambiyuous bases were resolvsd by esamination oithc t.lectrophoregrarns. Selected 200 bp
sequenccs from the 1500 bp sequence were cornpared nith 16s rDNA sequences h m
Gcnbank md E'VIBL sequencc databases. The 16s rDNA sequrnce of C 187 \vas then aligned
with 16s rDNA sequences of al1 species of the jenus .Vocctniioides and other related species.
using the algonthm Clustal W included in D'IAiLIh'l. The following nucleotide sequcnces
were obtained from the Genbank and EMBL databases iinder the following accession
niirnbers: AFOOjO 17 (,,Locardioicles fiiliws). AF005007 (rVoccinlioides 1~trelt.s). hFOO5006
( A'ocurdioides jer~ser~ii), S53 2 1 1 (iVoc~irtlioides dbtis), AF0050 1 5 (iVocurclioiciesjlav~is),
AF005007 (~Voccrrdioicies luteus). AF00500S (rVocnrriioicies plunrarum). AF0050 18
(LVoccrrclioities rlzermoliltci~ttis), AF005009 (iVocurdioides simplex). AF005070
(iVoccu-tlioitles sp. ATCC 2941 9). AF005021 (rVoccrrdioides sp. N S P 4 1 ). X92364

4 1
( (;eo<len~zorop/lil~~s sp. ). X93 1 S 5 (.4 crirtopiitles hrmilietrsis 1. SS3 806 ( C~'11dotnottc~s
[ru-hanr 1. DS-974 (.VociirdÏoirles sp. i . C9USS tStrepron1~res ilze~-rnoci~rho~~~~loi~o~-c~~zs DSXl
-14395 1. L-10627 ( Fr-cïiikici sp. 1. CO I 298 ( .Vuc~u.dioides sp. OS-4). XS067 1 ( Rlio~iococn~s
- o i ~ i i Sirnilanty \ducs and c\.olutionary distances wsre calculated from thc alignrnent
i DN.OI.AW .A phylogenetic tree \vas constructed by neighbour-joining with 1000 bootstrap
iterations.

Chapter 4. Results
4 1 Determination of diversity amonp atrazine-de radina bacteria isolated from
aericu ltu ral soils in Ottawa.
4.1.1 .\ssessrnen t of biochernical diversim.
Szvcral atrazine-de-adiny bacteria werc isolatrd tiom ihe Central Espenmental
Farm in Ottawa (Table 1 a. ). On .\.LIS pIus atrazine medium they produccd small whire
colonies ivith a distinct zone of cleanng around each colony. The Gram stain ÿnd KOH tests
revealed that al1 the isoiates were sram-nrgativc. ;\II of the isolates were cytocliromc c
ovidase positive. The cytochrome c ouidase-positive control strain P. jlziot-mcrris and thc
ouidase-nçgativc control strain E. roli gave the rxpecred rcactions. HPLC andysis. to
confirm that a11 the isolates degaded atnzine. showed rhar one of the isolatrd strnins. C 154.
\vas unablc to degrade atrazine (data not shown). Since C 174 was unable to degrade atrazinr
i t Lvas nor considered hrther. The .VI ZOE md .VI NFT biochemical strips were used to
comparc di fferenccs in biochemical reactions berwsen oach isolatc (Table 3a. Table 3b ). The
. V I X E systrm tests for fermentarive bacterio and rhe .&PI ';FT system tests for non-
fèrmentative bacteria. Dendrogarns slicited by cluster anaiysis OC the biochemical responses
to API X E and AP t 'iFT by atrazine-degading isolates formcd a dense clustcr in wliich
C 155 acted as tin outlier (Figure 7). Examination of the raw data indicatcd that C 155 differed
from the otherç in its ability to reduce nitrate cornplercly to nitroyen sas. Based on the API
ZOE strip al1 the isolates reacted negatively to tlic hllowing biochemical tests: transformation
of lysine into cadaverine. transfomarion of ornithine into putrecine. utilization of citrate as



Figure 7. A. Dendrognm of the Ottaua atrazine-degrading bacteria based on the
biochcrnicai responses to the .\PI 20E test. B. Dendro gram of the Ottawa
atrazinr-degrading bactcria based on the biochemical responses to rhs .\PI
'IFT test. In each case the dendrogam was simplified from that created by
equally weighted average linkagc cluster analysis to show the number of
biochemical responscs which ivcre di f i rent betwecn each isolatr.

Number of differences
Num ber of differences

1 7
the sole sourcc of carbon. the production of hydrogen sulfidc tiorn thiosulfatc. metabolisrn
of tnprophan. and the producrion of acetoin from sodium pyw-atc. .\II of the isolates were
riiso linable to ferment the crirbohydrates mannitol. inositol. sorbito 1. rhrirnnosc, mxose.
tneiibiose and amygdalin into an acid. Based on the APT NFT bioctiernical tests 211 rhe
isolatcs were unable to assirnilate D-gluconic acid. capric acid. adipic acid. L-arabinose and
L-rnalic acid. Both thc .VI ZOE and . V I NFT biochemical strips showsd a negati~qe reaction
for rhe followin~ biochemical tests: liqustàction of sslatin by protcolytic enzymes.
Iiydrolysis of ONPG to iiirrophcnol and yalacrose bu p-galxtosidasc. tomarion of indok
tiom the substnte rnptophan. transfomation of arginine into ornithine. ÿmmonia and carbon
Jioxide. and krmentation of $ucosc. .As w c l l both biochemical strips showed that al1 ofthe
isolates u-erc able to reduce nitrate to nirrite. Csin; the . l P I 30E biochcmical snip C 175 and
C 155 ivtire cinabie to produce ammonia î?om irea a. However. the .\PI SFT biocliemical s~r ip
show-sd that al1 of the isolates were capable of urea hydrolysis.
4.1.2 Xssessment of oenetic diversi-.
DNA tingerpnnts to determine the gcnctic di\-rrsitp of the atrÿzine-drgrading
bacteria [rom Ottawa wsre yenerated using rcp-PCR. Rep-PCR iising BOXA and ERIC
consensus primers. generated cornplex tingerprint patterns consistins of 20 or more PCR
products that ranged in size Iiom 0.2 kb to 1.6 kb ( Figure S and 9). Fingerprint profiles for
both primer sets were very sirnilar. with minor differences limited to the prescncc or absence
of one or two hi$ molecular weight bands (Figure S. lanes S. 9. 10. and 12). Thcsc results
suggest limited genetic divrrsity among the isolates. E. colt D H j a (Figure 8 and 9. lans 13)

Figure S. Rcp-PCR generated DNA tïngerprints of atrazine-degrading bactcna from
Ottawa. The Iinyerprint patterns were derived from whole cells using the
ERIC primer set t ERICIR and ERIC?: Table 2 ) . PCR products were
slectrophoresed in a 1 .jOG agarose gel and stained with srhidium bromidc.
Limes: 1 and 15: 1 kb DiI-4 Ladder: lane 2. C 147: lane 3. C 155: lime 4.
C 156: lane 5. C 160: lane 6. C 163: lane 7. C 175: lane S. C 176. lane 9. C 1 77:
Iane 10. C 178: lane 1 1. C 179: lane 12. C I SO: lane 13. E cofi DH5u: and lane
14. water control.


Fiyure 9. Rcp-PCR gensrated D-iA tingerpnnts of atrazine-degrading bactsria (rom
Ottawa. Tlis finyerpnnt patterns werc den\-sd frorn whole cclls using the
BOXA primer ( BOXA I R; Table 3. PCR products lwre stectrophorcsed
in a 1 . j o 6 agarose gel and stained with cthidium bromidc. Lanes: 1 and
15: 1 kb DNA Ladder: Ime 2. C 147; lane 3. C 155: lane 4. C 156: lanc 5.
C 160: lane O. C 163: lane '. C I 75: lane S. C 176. lane '1. C 1 - 7 lanc 10.
C 1 7s: lane 1 I . î 1°C): lane 12. C I SO: lane 13. E. c.oir DHSu; and lane 14.
water conrrol.


52
\\.as used as an outlicr and gave a distinct banding pattern cornparcd to the other isolates.
To tiirtlier compare the Ottawa isolates plasmid protiles were yenented u s i n j the
alkaline Iysis nicthod of isolation describecl by Kado and Liu ( 198 1 ). The plrisrnid protiles
arc shonm i n Figure 1 O. The size and nuniber of plasmids varicd for cach isolate. howevcr.
C 147. C 1 -T and C 1 7 s had siniilar profiles as did C 1 55 and C 156. .A sinjle 97-kb plasmid
\vas cornmon to al1 isolates. The s i x of the cornmon plasmid was dctcrmincd by creating
a standard cL1n.e iising plasmids ofknown molrcular weight from vanous bactcria (Figure
1 O. lanes 1 to 3 ).
4 . Characterization of C187-an atrazine-de~rader from St. Hvacinthe. Ouébec,
Canada.
4.2.1 Characterization of C 187.
On AMS supplemcntcd with atrazine C 1 S7 produces small (0.5 to I mm) white
colonies with a clear zonc surrounding rach colon-. The presrncc of thcse halos \vas
attributed to the metabolism of atrazine to soluble products. The Gram stain and KOH test
rcvealed Cl87 to be gram-positive nnging from rods to cocci in shape. The cytochrome c
oxidase test indicated that C 157 was oxidase positive. The 16s rDKA gene of C 187. which
\vas 1462 bp long (Figure I I ) . was sequcnced and compared with seqiiences published in
Genbank and EMBL. The 16s rDNA sequrnce of C l87 was aligncd with the 16s rDNX
sequences of al1 known iVocrirdioides species and other related species. Similarity values
and cvolutionary distances were deterrnined and a phylogenetic tree (Figure 12) was
constmcted by the ncighbour-joining mcthod. C 187 foms a distinct lineage within the

Figure 1 O. Plasmid profiles of selectrd anazinr-drgading bacrerin isolated fiom Ottawa.
Lncut plasmids isolated by the alkaline !-sis rnrthod of Kado and Liu ( 19s 1
were subjcctrd to horizontal agarose gel elrctrophoresis in 0.jU;, ayarosc ycls
at 3 2 V cm for 1.1.5 In. Size and mobilitp of fi1-e uncut reference plasmids
(pR4O.A [146 kb]. pR1 [93 kb]. DC3OOO [6S kb]. pRK7013 [AS kb] and pGS9
[30 kb]) are indicatrd on the leA ( [anes 1 to 5 ). Lane 6. C 147: lane -. C 155:
lane S. C 1 j6; Iane 9. C 160: lane 1 O. C 163: Ianr 1 1. C 1 ": lane 11. C 1-S:
lanc 1 3. C 1 79: and lane 1-4. C 1 W .


Fisure 1 1. Nucleotidr sequence of the 16s rDNA gene from C 187. The cornpiete
nucleotide sequence ( 1462 bp) of both strands was arnplified using published
primen (Stahl and ;\mman. 1991 ). The PCR product was sequenced at the
John P. Robarts Research Institute DPiX Ssquencing Facility (LWO.
London, ON).

CCTCTTTCAG AGW.GMGC.2
AGAGGTATGC SIWATGCGC.1 SGTTCTCTGG
ZGGAGGAAGG
CGTCTCAGST
CACGAAAGTT TGTSGAGGGA
1 4 4 1 GAAYTCGTAA
GGCGGCGTGC CTTCGYGGGT G,;iGC.WTCTG Û A 4 Y C TAV--S-- ~ r - - r - du L uGFAAG ATSAGCTTGT CC-UGTAGCCS TGAGACACGG MTATTGGAC CGSGAGGGAT CAGGGACGAA CCC-GZC.nJ.CT TÛCGAGCGTT -AGULGGTC'ïG CCCTGAGZCI AGGGGAGXiC GATATCAGGA G C A T T X C T G CASGATTAGA GCGCTAGGTG GCT-WCYCAT A b v ~ i M.?GAC GGCGGAGCAT TTACCTAGGT 2AGGTLCGT.A GTCAAGCTCGT G A G C C C S C C GGGACTCATA TGGGGATGAC CTTCACGCAT ATCGCAAGAT CGGATTGGGG GCTAGT19ATC CCCGGGCCTT GGCW-CACCC GCCGTCÛAAT CAAGGTMGCC
TTAACACATG ACACYAGCÛY SCCTTC-zlCAT SCSI\&TAC SA TTP ,;LCGGCGGT TGÛTUGGGTA GCCTGAGAGG CCCAGACTCC .UTGGGCGAA GACYGCCTTC GCGCAAGTGA A C G T G C C A K CTCCGGPATT TCACGTCGGG GCTTCCSATA GG.WTTCCTG GGAACACCGG ACGCTGAGGA TACCCTGGTA TC-GGACTCAT 7,AAGCGCCCC TCA.UGGAF.7 GCGGATTGAT TTGACATATA AYGOTCCTAT GTLGTGAGAf CTLGTCTTAT AGAGACTGCC G T C M G T C A T GCTACPATGG GGAGCGAATC TCTGCAACTC GCAGATC.4GC GTACACACCG GAAGCCGGTG GTGGGGCGAG GT
cr.AGTCG?.GC CGAACG(JYT5 ZS.GGAT-?J-C'T LC?.CTTC%L-G GTiAG GAA*r ..CSGCC,CACC STGACCEGCC TACSGGAGGC AGCCTGATCE GGGSTGTWJ. CGGTACCTSC AGCCGCGGTA ASTGGGZG'ï>. AGTGAPtMCT ICC-GGCAGHCT GTGTAGCGGT TGGCG=GGL G C G M G T G T STCCACACCG TCCACGAGTT GCCTGGGGAG 3GF.LGSGGGC TCGATGCFAC GGGFAATCTS ACAGGSGGTG STTGGGTT.AA GTTGCC.4GC.A GGGGTCAACT CATGCCCCTT CLSÛTACAPA CCXLV4.A-GCC GACCCCATGA AACGCTGCGG CCCGTCACGT GCCCA?-CCCT C G A T T G G G K

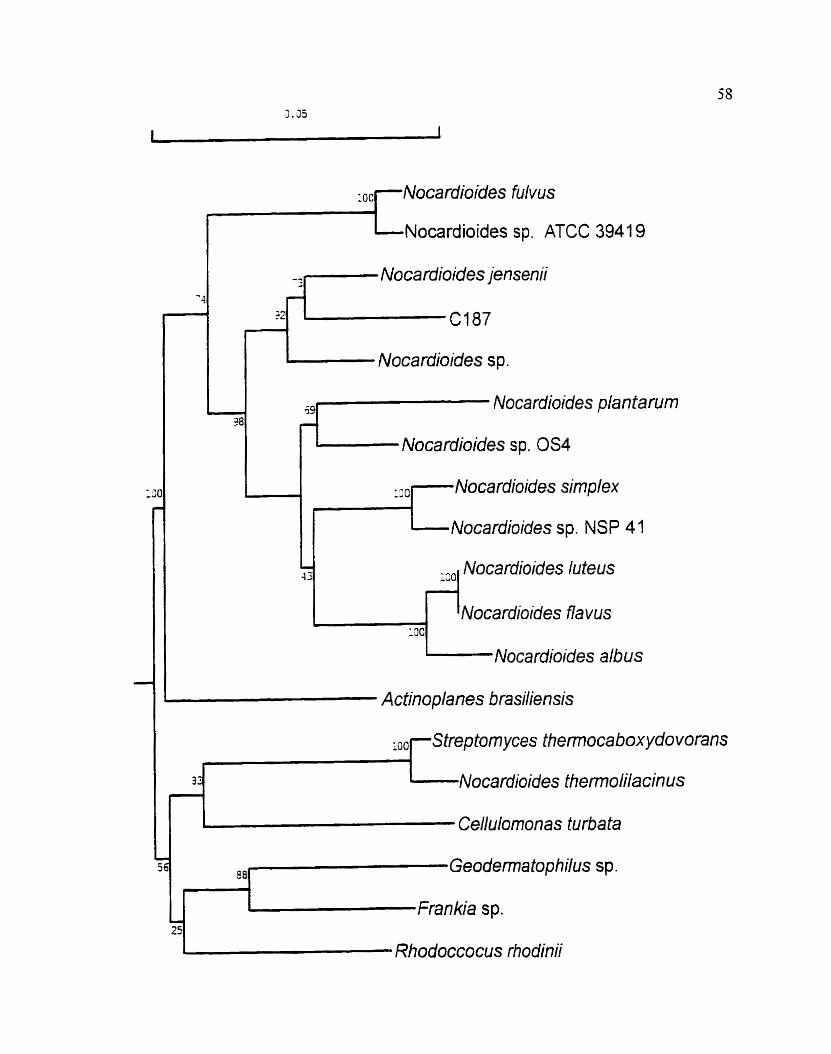
Figure 12. Phylogenetic tree showing the positions of C 157. members of the genus
:Vocardioides and other reiated species. The tree was generated using the
neighbour-joining method and was based on a cornpanson of the 16s rDNA
sequences. The numben at the nodes indicate the lsvels of bootstrap support
based on data for 1000 replicares. The bar scale indicatss 0.05 substitution
per nucleotide position.
9 Nocardioides sp. ATCC 3941 9
''1- Nocardioides jensenii
Nocardioides s p.
4 Nocardioides plantarum
Nocardioides sp. OS4
::cl- Noca rdioides simplex
r-'- Nocardioides sp. NSP 41
130 Nocardioides luteus
Nocardioides f h vus l m
Nocardioides albus
I Actinoplanes brasiliensis
1 q-Streptom yces thennoca box ydovorans r
32 I Noca rdioides themolila cin us r 1
Cellulornonas turba ta
7 os( Geodematophilus sp.
'1' Frankia sp.
Rhodoccocus rhodinii

senus .Vocrr~'ciioide.s and is most similar to .V. jerrsenii.
4.2.2 Determination of sequence similari with Psertdomonas sp. strain ADP ~ e n e s
encodin~ atrazine hvdrolase and hydrosvatrazine ethvlaminohvdroiase and with the
Rliodococcrrs coraffin ris 'VRRL B- 15444R oene encodin~ s-triazine ch lorohvdrolase.
Dot blot hybridization was used to determine if CI 87 had sequence sirnilarity with
the genes encoding atrazine cnlorohydrolase ( 4 ) and hydroxyatrazine
ethylarninohydrolasc (trtzl3) from the atrazine-degrader Pserrdornonus sp. strain ADP. PCR
using custom primers (Table 7 ) was used to ampli- and DIG-label sequences internai to the
NI-4 and utzB yenes fiom E. coli DHSu pMD4 m d E. coli DH5u pATZB-2 respectiveiy.
Amplification of ricB yielded a single product of approximately 3 0 bp in length (dara not
shown). .hplification of c r c 4 yielded 3 producrs of approximately 7500. 1700 and 900 bp
in length (dara not shown). The 900 bp product. which was the evpected product size based
on primer design. \vas purified from agarose sel and used the probe. The 900 bp DIG-
labelled product did not cross-hybridize with crcB. When the cntire PCR product mixture
(which included al1 three product sizes) tvas used as the ~rt--..l probe there was
cross-hybridization with r i t 3 . suggesting that the nvo additionai products ( 1700 bp and 2500
bp) were the result of non-specific primer binding during PCR. Both the DIG-labelled utzA
probe and the DIG-labelled mzB probe did not hybridize to purified genomic DNA from
Cl87 (Figure UA. l3B, lane 1 ) indicating that this strain had no sequence sirnilarity with
the two genes (crrz.4 and atzB) From Pseiidonionas sp. strain ADP.
Dot blot hybridization also was used to determine if CI 87 had sequence sirnilarity

F e 1 3 Dot biot hybridization of purified DNA From Cl87 with DIG-labeied ntz.4
(-4) and DIG-labeied atzB (B) probes from E. coli D H j a (pbfD4) and E. coli
D H j a (pATZB-2) respectively. 1. C 187: 2, Cl 93: 3. DC3000: 4. E. coli
DHja (piMD4); and 5. E. coli DHja pATZB-2.


62
with the gene rncoding s-mazine hydrolase from Rlrodococcrrs cot-dli)rzrs hiRRL B- I j J J J R .
A 600 bp BamHI fragment from the r r i A gene was DiG-labelled using rmdom pnming
(Zhu. unpublished). The rrs.4 probe did not hybridize to punfied genomic DNA from
C 187 (Figure 14. lane 1 ) indicating no sequence similarity with the rr-s.-l gene fiom
Rl~odococc~rs cordlinrrs 'IRRL B- 1 5 W R .
4.3.3 Mineralization of atrazine bv Cl 87.
To determine if C 157 was capable of metabolizing the aromatic nucleus of citrazine.
cells were incubatrd in .LMS medium containinj [C-ring "Cl atrazine. The remaining
radioactivity was quantified afier 14 and 4 3 h by LSC. The ndioüctivity decreased by 4%
afisr 48 h incubation for C 157. G% for the uninoculated control. and 94% for the positive
connol Pretîdotno>tus sp. strain ADP. Sratistical analysis using the t-test indicated that there
was no ~i~gnificant difference in the decline in radioactivity benvcen the uninoculated control
and C 187. There was a significant difference (p< 0.000 1. n= 4) in the mineralization
capabilities between C l S7 and Psezrdoino>rus sp. strain .-P. Thus C 1 57 is unable to
mineralize atrazine to CO1 and mmonia.
42.4 Degradation of atrazine to hvdroxvatrazine bv cell free estract.
In order to determine the regulation of amine-degading enzymes. CFE from C 157
gown in the presence of atrazine or in the absence of a m i n e was cvaluated for the ability
to degrade atrazine. CFE combined with PB (pH 7.1. 10 miM) containing atrazine was
analysed by HPLC at varying time intervals, and the resultant chromatograms were

Figure 14. Dot blot hybridization of purified DNA kom C 187 with DIG-labeled t r i -4
probe fiom Rhodacocczls cordlinr~ NRRL B- I j W R . 1. C 157; 2. DC3000;
and 3. R. cornllr'nrrs NRlU B- 1 5 W R .


65
comparecl with those of atrazine and hydrox yatrazine standards. Atrazine was no longer
detected afrer 24 11 of incubation. The disappearance of atrazine correlatcd with the
accumulation of a single metabolite with a rctcntion time of 4.6 min or 7.1 min depcnding
on the solvcnt system used (Figure 15). The rctention tirne of the metabolite corresponded
to thai of a hydroxyatrazine standard (4.4 min with 70041 methanol: 304% watrr. 7.15 min with
5O0h methanol: 50?" 1 O rnM ammonium acetate).
Figure i 6 shows the degradation of atrazine and the formation of hydrouyatrazine b y
CFE prepared tiom celis gown in NBY with (40 m g L) and without ütrazine. In both cases
(with and without atrazine) there was no lag precedins atrazine degradation. The rate
constant ( k ) of atrazine-degradation per mg protein for cells g o w n in atrazine was 5.5 u 1 0 '
min-' ( n = 4 standard deviatbn [std. dev.]= 2.4 x 10' min") and the rate (V,,- ,,,) was
30.6 nrnobmj protein - min ( n = 4 std. dev. = I 5.6 nmol/ms protein - min). For cslls grown
without atrazine k=5.7 'r min-' ( n = 4 std. dev.= 2.7 u 10.' min ' ) and V, , ,,,= 28.1
nrnoiim~ protein - min (n=4. std. dev.=14.9 nmol/mg protein - min). The variation
demonstrated by the larse standard deviations can be partially explained by intcrcxpcrimental
variability (.kVOV.4. p= 0.023). The assumption that degradation kinetics were tirst order
was substantiated by the linearity of the log rransformcd concentration versus time graph (6
> 0.93 for a11 graphs). The paired t-test showed no significant differencc bcnvcen the rate
of degradation for cells gown in atrazine versus those grown without atrazine. Thesc results
suggest that atrazine degradation by C 187 is expressed constitutively.

Figure 15. Transformation of a tmine to hydroxyatnzine by CFE from C 1 S: when cells
were g o u m with atrazine (B) and without asruine t C ) . as demonstrateci by
reverse phase HPLC. ( -4) Authentic standards of atrazine (46.38 uM) and
hydroxyatrazine (46.38 uM). Mobile phase. X O h CHIOH. ~ O ' ' O 5 mM
NaH2POI at a tlow rate of 1 mlmin .

d - 5 --
TU - - Hydroxyatrazine
J 1 6 3 'O
Retention Time (minutes)

Figure 16. Dissipation of atrazine (+) and formation of hydroxyatrazine ( W ) by CFE
from Cl57. ( A ) Activity of CFE when cells were groown in the presence of
atrazine. (B) Activity of CFE when cells were g o w n in the absence of
atrazine. Exponential cunre titting wri; used to determine first order rate
constants for atrazine degradation. Each rate constant was normalized based
on the arnount of protein per ml CFE. Error ban indicate the standard srror
among two replicates within one experiment.

Time (minutes)

4.2.5 Deyradation of atrazine bu whoie cells of Cl8%
Whols ccl ls gown in the presence of atrazinc w-erc ecaluared for rheir ribility to
degads atrazine under aerobic and oxyyen- ties conditions. .AM S containiny arrazinc \vas
inoculated with whole cclls and anaiysed by reverse phase HPLC at T=O and T=71 h.
Resultant chromatogams were cornparcd with thosr of atrazine and hydrosyatrazine
standards (Fiyurc 1 3 . Atrazine \vas no lonser dstected riftcr 23 h. .A metabolite ~v ich a
retention time of4.6 min was Jetscred rifter 24 hour. The retention time of the metabolite
corrcsponded ro that O t' a hyùrosyatrazinr standard 14.43 min ~ v i th 70"zh methanol: 30%
water 1.
To determine if the enzymes involved in atrazine-degradarion werc oxypen dependent
whole cclls of C 187 rnived with an q u a 1 volume of phosphate buffer conraining iitrazine
( 20 rns L) Lverc anaiysed for rheir abilit'. to ciegradc atritzins nhen incubated i\.ith 0s)-gcn
and without osygen. Figure 1 S shows the degradation of atrazine. accumulation of
Iiydrosyatrazinc and the subsequent degradation of hydroxyatrazine by tvhole cells for borh
incubations. The per cell rate (V, ,=,,,) of auazine degradation for cells incubated with oxygen
kvas 4.8 s 1W- nmol/CFC - h (n-6. srd. dcv.=j.l .; IO-'nmol.'CFC - hl (1.0 s I O - " 2 of
atrazine:CFC - h ) and the rate constant ( k ) per ce11 was 7 . 2 s 10"' h-i (n=6.
std. dev.=,.j x 1 9 " h-'1. In the absence of oxygen V,,=,, ,=l.4 s 10.' nmol!CFC - h (n=j .
std. dev.=l.O x 1WS nmol/CFC . 11) (3.0 x 10-" g oTatrazinuCFC . h ) and k=1.7 s 10'12 h- '
(n=5, std. dey. 4 . 7 s 1 O-" h+l). In al1 cases the 6 values for the 10% transfomed graphs were
yreater than 0.91. At 72 h hydroxyatrazine w u completely degraded under aerobic
conditions. Anaerobically, hydroxyatrazine was completely degadcd after 72 h (data not

Figure 17. Degradarion of atrazine by C 1 S7 whole c r l l s as drrnonstrated by HPLC
~inalysis. (-41 Authentic standards ot'arrazine 46-38 uM and hydroqatrazinc
40.3 S uM. ( B) C i 87 whole cells incubated with . A M containin3 atrazine and
sarnpled at T=O and T=14 hours. Mobile phase. 70% CH,OH. 30% 5 m.LI
NûHIP02 ai a tlow rate of 1 rnl'rnl.

Retention Tirne (minutes)

Figure I S. Dissipation of atrazine ( + 1. formation ofhydroxyatrazinc ( W ) and subsequent
degradation of hydroxyatrazine by Cl 87 whole cells. . Concentration
changes of anazine and hydroxyatrazine over 7 2 hrs whcn cells were
incubated aerobically . tB) Concentration changes of atrazine and
hydmxyatrazine over 73 hn when cells were incubated anarrobicallg. Inscn
gaphs show atrazine degradation and hydrouyatrazine formation in the iirst
hr for each incubation. Exponential curve fitting was used to detemine îïrst
ordcr rare constants for atrazins degradation. Error bars indjcate the standard
crror m o n g six replicates wirhin one sxperirnent.

Time (hrs)

75
iho~vn ). In both cases there ivas no accumulation of an additional rnctabolite. Statistical
anülysis using the t-test showcd a significant diffcrence between atrazine degradation
aerobically and anacrobically (p=0.000003. n=9. Cndcr maerobic conditions. atrazine was
degraded 3 tirnes s lowr . These results suggest that the enzymes reqiiired for atrazine and
hydroxyatrazine degradation are not dependent on mo lecular oxyjen.

Chapter S. Discussion
5.1 Diversitv and ~ ia smid distribution in rttrazine-de~radina bacteria isola ted on a l o t
scale.
Prokaryotic genomes contain srnall interspersed rcpeii tive sequences scparating
longer single-copy DNA scqurnces (Lupski and Weinstock. 1991). Thesr sequences are
non-encoding. between 1 5 and several hundrcd base pairs in length. and c m be ~ised as
oiigonucleoride binding sites for PCR rnediated genomic fingcrpnnting (Versalovic ct al..
9 1). -4lthough the sequcnces were idenrificd in a single species or yenus. several arc
conszrved and dispened in the genomes of rnmy diverse gram-negative and sram-posi tive
bacteria (\*crsaiovic et al.. 1994). Three such sequences are ERIC. repetitive extragenic
palindrornic (REP) and BOXA; REP and ERIC arc palindromic units. Genornic
fingrpnnting with BOXA. ERIC andior REP sequcnces is possible becausc individual
strains va- with respect to the number and distribution of these systcms in the cliromosome
( De Bruijn. 1992; Poolcr et a[., 1996; Judd et al.. 1993: Versalovic et al.. 199 1 ). In the
presenr study BOXA and ERIC consensus primers in combination with PCR were used to
determine the divenity of a group of gram-neyativc atrazine degraders isolated korn
a-gicultural soi1 near Ottawa. Xlthough there were occasional differences in the intensity and
prcsence of high molecular weight bands. the overall genomic fin~erprints of the soi1 isolates
suggests that the population of atnzine-degading bactcria is homogeneous. Some random
variation in the higher molecular weight region of the profiles is not uncxpected since
extension efficiency by Tkq polymerase decreases as the length of the target sequence
increases (Sambrook et al., 1989; Versalovic et al., 1994).

Bactena of diverse type and habitat are known to harbour plasmid DNA
(Stanisich. 1983. The çizc and number of plasmids a bacterium harbours can vary
tremendously ( Stanisich. 1985). .-\ varicty of plasmid isolation protocols haïe bccn
Jescribed (Kado and Liu. 198 1 ; Wheatcro fi and L~'i1liams. L 9s 1 ). d l of ichich exploit the
sniall çize (conipared to chromosornal DNA) and covalently clossd circle nature ofpiasmid
DNA i Sambrook et ai.. 1989). LVhile the plasmid profilcs of the airw.ine-degrading bacteria
were uniquc. one plasmid of iipprosirnatcly O X b \vas common to al1 isolatcs. Rccent
plasmid isolation studies b!. De Souza et al. ( 1998) on Pseridoi?rotit~s sp. strain .\DP
indicarcd that the genes for atrzine caiabolism to cyanunc acid: il[=-1. -B. and -C. are
locaiized on a 96-kb self-transmissible plvmid (pADP- I ). The rclationship benveen pADP-
1 and the 97-kb plasmid from the Ottawa isolates has yer ro b r detcrmincd. However.
knoiving rhat the Ottawa isolatcs haive sequencc similarity with ~ i r z . 4 anci -4tzB (data not
sliown) and tliat horizontal genc tnnsfcr (the transmission of DNA lragments between
different specics) (Hemck et al.. 1997) in narurally occumng microbial commitnities is nor
uncornmon (Fulthorpe and Wyndham. 1991: Hemck et al.. 1997: Lorcnz and
Wackemagcl. 1994). this 97-kb plasmid may in fact be the same one found in Psezr~lorm.mcrs
sp. strain ADP. As mentioncd above. the Ottawa isoiates yenerall y had di Kferent profiles.
Differcnccs between each isolate was not totally unespectcd since plsrnids Iiavs the ability
to be integnted into chromosornal DNA. c m be dispenscd by host cells under varying
growth conditions m d can underso horizontal tram fer (Madigan ct al.. 1997). Furthemore,
plasmid isolation techniques may result in plasmids of one isolate (and not another) being
nicked. resulting in different migration distances and hence the appearance of different

7s
plasmid profiles.
Substrate utilization and biochemicai reactions. using the .-\Pi systcms. were also
usrd to assess isolate Jiversity. Thsse spstcms which are standardizcd. miniatiinzed wrsions
of con\-entional biochemical procedures nllow for the quick identification and
characterization of E~treroh~icreriacme and other gam-negativç bacteria (bioblerieus Vitek
Inc.. 1095). .Amonj thc 1 1 strains. strain C 155 \vas the most uniquc. The major differencr
between Ci 55 and the othcrs was its ability to reduce nitrate ro nitrogen =as. Amongst the
remaining isolates thsrc wcre vcry f c ~ v differences in substra~s urilization and biochemical
- reactions as shown in Figure ,.
5.2 Characterization of a uniaue oram-oositive bacterium frorn St. Hyacinthe,
Québec-
5.1.1 Identification of Cl 87.
[dentification of C1S7 was bascd on 16s rDNA sequsncins. Traditionaliy the
classi fication a d identi ficntion of organisms has becn based on morphological.
developmental and nutntional characteristics ( Lane ct al.. I9S5: Moritz and Hillis. 1996).
However. these characteristics have the disadvantage in that they rire Iiighly intluenccd by
cnvironmental conditions and therefore directly affected bp cvolutionary convcrsence (when
organisms share a common phenotype because of common e v o l u r i o n ~ pressures) (Lane et
al.. 1985). For this rcason phylogenetic analysis based on macromolecules (,DNA. RNA.
protein and lipids) is more desirable. Of these macromolecules. ribosomai RNA (or DNA).
particularly the 10s rRNA coding region has been the rnost useful for establishing

79
relationships and identity between rnicroorganisms. since i t is universal. homologous. and
ücccssible (Hillis and Davis. 1986: Mindel1 and Honeycurt. 1990: Weller and Ward. 1989).
Cornpaison of the complete 16s rDKA sequencr tiom C 1 S7 ivith sequences published in
Gcnbank and EMBL indicatcd trhat C 1 S7 is a new species in the genus .Vocwdioides and is
most similar to .Y jemenii (AFQOjOO6). .LIernbers of the jenus .Loci~rtiioi&s are gram-
positive. pleomorphic. arrobic. mesophilic nocardiofom actinomycetss that r om well-
devcloped vegetative rnycçlia. Escept for the production of vegetativs myceliiim ( when
yrown on .\.LIS agar supplsmented with iitrazine). C1S7 hris a11 of ihese characteristics.
.Vocm-cltoitles sp. strain C l 87 may fom vegetative mycelium. However. the conditions
required for vrgeiative mycelium formation upere not defined in this study. .Vocrirciioides
species have been found to degrade a variety of organic pollutants including picnc acid
(2.4.6-trinitrophenol). pyridinc. dibenzothiophene and 2.4.5-tnchlorophenoxyacctic acid
iGolovleva et al.. lC19O: Rajan et al.. 1996: Yoon et al.. 1997: Sandhya et al.. 1995). While
other gram-positive bacteria including .locurciiu (Giardina et 31.. 1982) and Rlioducoccrrs
(Nagy et al.. 1995: Shao and Behki. 1996) san degrade ritruine (by .V-dealkylation), this is
the tint rcport of a .Voctrrdioirles species degrading atrazine.
5 2 . 2 Genetic and biochemical analvsis of the initial stem in the atrazine-degrading
pathwav.
Reverse phase HPLC analysis was used to follow a m i n e degradation by whole cells
oi'Xoccu-chdes sp. strain Cl 57 incubated aerobically and anacrobically. In reverse phase
HPLC. the stationary phase is non-polar and the mobile phase is polar. HPLC separates

50
molecules based on adsorption. ion-exchange and size esclusion. [ts superior resolution.
speed and hiyh sensitivip pcrmits the separarion. quantification and identification of srnall
qiiantities o t' metabolire. Howcvcr. HPLC doés not always provide conclusivc proof of
idcntity since thsrc is the possibility that some metabolites nmay have similar retznrion tirnes
whcn run in the sarne solvcnr system. Ldeally HPLC results would be coniïrmcd by linking
retention times with mass spectrometry (De Souza et ai.. S . Mass spectrometry
compares the complcte mass spectmm of a compound witli the mass spcctra of known
compounds (Eagle sr JI.. 1 0 9 1 I. Thc contirmation of identification (of metabolites) by mass
spectrometry was Sryond th<: scope of this study. Atrazine was dsgradsd by strnin C 187
under asrobic and anaerobic conditions. Howevçr. degradation was approsimately 3 times
hstcr aerobically. In both cascs whiic atrazinc was beziny drgraded hydrosyatrazine as
accurnulating. Once suazinc had dissipateLi .Vocu~-(fio~~irs sp. strain C 1 S- bcgan to degrade
hydrosyatrazins. Degradation of atrazinz and hydroxyritrazinc under osygrn-lirniting
conditions indicates that thc enzymes invoivcd are not mono or diosy,vsnases. which require
rnoircular oxygrn. but rathcr hydrolases. Most gram-positive bacteria that degradc ritrazine
are distinct in that degrridation is limited to .V-dealkylation and occurs under oxygen-rich
conditions by mcans o h PJ5,-like monooxygenase (Giardina ct al.. IW2; Nagy et al.. 1995;
Shao and Behki. 1996). However, RItodococcris cor-ullinrrs YRRL B-1SWR can
dechlorinate and deaminate dsisopropylarninoatrazine (but not atrazinc) anaerobically and
thcrefore presumably hydrolytically (Cook and Hütter. 1986). There has also been reports
o f gram-negative bacteria. including Psetcdomoiius sp. strain A D P (De Souza et al.. 1996).
degrading atrazine or its metabolites under oxygen-limirin; conditions

S I
(Radosetich ct ai.. 1995: Jessee et al.. 1983). Dc Souza et al. ( 1996) indicatcd that the
cnzymes Lvere hydro lases. The exacr rnechanism of the hydrolases synthesized b y strain
Cl 57 was not detsnnincd. Howsver. in the case of atrazine dechlorination. the enzyme
would catalyse the direct substitution ofchloride for hydroxide ( kom watcr). It is of interest
that the dec hlorination of pentachlorophenol to tcrrac hloro hydroquinone by
FZu-ohcicteri~rni sp. \vas originalIy thought to occur via a hydrolytic enzyme (Steisn and
Crawford. 1056) and was later shown to be catalysed by a monooxygrnasc (Sun and
Oner. 1991 ). . h y arnbiguity about atmzinc dechlorination by strain C 157 could be resolved
by enperimrnts using H,"O. howsver this mas not investigatrd in this study. If in fact the
cnzvrnc is a hydrolase tlien ['W]hydroxyatruine would bc yieided when whole ceils or CFE
of .Vocc~rdioides sp. strain C 157 are incubared with atrazine in H,"O. This result would ruls
out the possibility that the enzyme catal ysed cxchange aftcr an initial rnonooxygenasc
reaction that used traces of oxygen that remained in sealsd reaction vessels.
Nucltic acid hybridization offers a rapid approach to tht idtntificarion of
hornoloyous gene sequences (Cuppels et al.. 1990). Givcn that the appropriate DNA
scquence (probe) and hybridization conditions are chosen. thc sensitivity and specificity of
this mrthod is quite h i j h (Cuppels et al.. 1990). DG-labtllrd probes are preferred over
radio-labeilcd probes because thcy requirr shorter rxposure times and digosigenin poses no
health or environmental hazards. In addition. the short half-life of radioisotopes necessirates
kequent preparation of the probe. whereas DiG-labelled probes arc stable for up to one year
and can be reused several times (Boehnnger .LLannheirn Biochernicals. 19953. To determine
if .Vocnrd~ocrles sp. strain C 1 57 had seq uence similarity rvi th the Psetrdoinonas sp. strain

ADP ;enes sncoding atrazine chlorohydrolase
etliylarninohydrolase ( ~ l c B ) . dot blot hybridization
82
1 ) and Iiydrox yatrazine
usiny DIG-labeiled probes was
performed. ;\lthough .Vocui-dioides sp. s tnin CI S7 dechlorinates atrazine and degrades
hydroxyarrazine Iiydrolyticall y. its enzymes are not cncodcd by genes with Iiornology to K.-l
and KB. Thus. .Vbccir-diotdes sp. strriin C I S7 and Psetidolon~oizus sp. strain hDP may have
comp Iereiy different hydrolase genes.
To determine if .Vonri-diotdes sp. strain C 157 had scquence simiiarity with ri=.-l. the
gene cncoding s-{nazine Iiydrolase in Rliodococcz~s corolliittrs XRRL B - I 3 U R
( Shao et al.. 1095) dot blot hybridization us@ a DIG-labelled probe was performed. The
hydrolase responsibie for dechlorination of a m i n e is not cncoded by a gene with homolozy
to irz-1. susgesting that the Iipdrolase synthesized by strain Cl S Ï ( for dechlorination) is
cncoded by a diffcrcnt sene sequence.
5.2.3 Re~ulation of atrazine degradation.
Reverse phase HPLC was used to tollow atrazine degradation by strain C 1 S? CFE
in order to detennine enzyme regulation. CFE from .\ioccir-riioides sp. strain Cl S? cells
erown in NBY or NBY siipplernented with atrazine degraded atrazine at similar rates - (V,,.,,,=2S. 1 nmol/mghnin and V ,,.,$0.6 nrnolimgmin respectively ). In both cases
hydroxyatrazine was accurnulated: the concentration accumulated was equivalent to the
concentration of atrazine degradeci. Hydroxyatrazine was not degraded. Nearl y identical
rates dong with the absence of a lag phase indicates that the gene sncoding the hydrolase
requircd for dechlorination is continually transcribed. Thus. degradation of atrazine to

53
hydroxyatrazine by 3'occir-dioides sp. strain CI87 is constitutive. The accumulation of
hydroxyatrazine by CFE and not whole cells may suggest that the hydrolase required for
Iiydrosyatrazine degradation is an extracellular enzyme. however it is more likcly that the
protein responsible for hydroxyatrazine degradation was sensitive to the procedures
(sonication and centri fuytion) used to prepare the CFE.
Hydrosyatrazine is nonherbicidal and nontoxic (Gysin and Knuesli. I 9 G O ) and does
not leach t'rorn soi1 as reridily as atrazine (Erickson and Lee, 1989). Thus, hydrolytic
dechlorination is an idcal mechanisrn for degrading atrazine with the goal of bioremediation
of agricultural soils. In surnmary. this study presents the first report of a .Voc~it*dcoides
specirs capable of constitutive. hydrolytic degadation of atrazine and hydroxyatrazine. This
study Funher indicates that on a plot scale there is lirnited diversity among atrazine-degading
bactena.

84
References
Assaf N. 4.. and R. F. Turco. 1994. Accelerated biodegadation of atrazinr by a microbial
consortium is possible in culture and soil. Biodegadation 2: 29-35.
Brhki. R. M.. and S. H. Khan. 1956. Degradation of atrazinç hy Pîczidonioriu-:
.Uealkylation and dehalogenation of atrazine and its metabolites. I. Aync. Food. Chem.
3 4: 746-719.
Behki. R. M.. and S. H. Khan. 1987. Isolation anci charactenzation o t' an s - e t h y i - 5 s -
diprop y 1 thiocarbamats-dcsrading .di-tiwohacrer stnin and evidence for plasmid-associated
s-ethyl-Y-N-dipropy lthiocarbamate degradation. Appl. Environ. bLicrobio1. 53: 1088- 1093.
Behki. R. M.. and S. H. Khan. 1991. Degradation of atrazins. propazinc. and sirnazine by
Rlrodococcrcs strÿin B-3). J. Aync. Food Chsrn. 42: 1 3 7- 134 1.
Behki. R.. E. Topp. and B. A. Blachvell. L994. Ring hydroxylation of .L--rncthylcarbamate
insecticides by Rhodococc~is TE L . J. A - ~ C . Food Chem. 45: 13 75- 1 3 78
Beki . R.. E. Topp. W. Dick. and P. Germon. 1993. 'vktabolism of the herbicide atrazine
by Rhodococc~is strains. Appl. Environ. Microbiol. 59: 1955- 1959-

SS
Bzlluck. D.. S. L. Benjamin. and T. Dawson. 199 1. Grouridwater contamination by citmine
and its metabolites: nsk assessment. policy. and legal implication. p. 254-'-- . Ill L .
Somasiindaram. and J. R. Coats (rd.). Pesticide transfomation products: h te and
signitigance in the cnvironment. Amencan Chernical Society. Washington. D. C.
bioMerieiis Vitek. Inc. 1995. 3pi 30E Sysrem. biokleneus .\PI klaniial.
Boehringer blannlieirn Biochemicals. 9 The DIG system rissr's guide for tiltcr
hybridization.
Boundy-klills. K.L.. bl. L. De Souza. R. T. blandelbaurn. L. P. Wiickett. and M. I.
Sadowsky. 1997. The i tcB gene of Pseldonio~iris sp. srrain ADP encodes r l i r second
enzyme of a novel titrazinc dsjrndation pathway. Appl. Environ. Llicrohiol. 63: 9 16-923.
Bouquard. C.. J. Ouazzani. J. Prome. Y. Michei-Briand. and P. Plésiat. 1997.
Dechlorination of atrazine by a Rlrizohitm sp. isolate. Appl. Environ. Microbiol. 63: 362-
S M .
Boucvcr. H. 1989. Agiculture and proundwatcr quality. Civil Eng. 59: 60-63
Bukhari. A. 1.. J. -4. Shapiro. and S. L. Adhya. 1977. DNA insertion clcrncnts. plasmids,
and episomes. Cold Spring Harbor Laboratory, Cold Spring Harbor. N. Y.

Cobb. A. 1997. Herbicide and plant physioiogy. Chapman and Hall. New York. Y. Y.
Cook. .A. LI. lclS7. Biodegadation of s-triazine senobiotics. FEMES blicrobiol. Rrv. 16:
93-1 16.
Cook. -4. 41.. P. Beilstsin. H. Grossenbacher. and R. Hüttsr. 1985. Ring clcawgc and
drgradative pathway ol'cyanuric acid in bactena. J . Biochem. 131:35-30.
Cook. A. M.. and R. Hütter. 1984. Deethyisirnizine: bacterial dechlorination. deaminiarion.
and cornpletc degradation. J. .Agric. Food Chsm. 32: j S 1-587.
Cook. -4. M.. and R. Hüttsr. 1986. Ring dechlorination of derthylsimazins by hydrolascs
from Rlzodococcii~ cor-dliriits. FEMS Microbiol. Lrtters. 34: 33-33s .
Cuppels. D. A.. R. A. Moore. and V. L. Morris. 1990. Constniction m d use of a
nonradioactivc DNA hybridization probe for detection of Pseinioi?iorl~u syi-tti,ocle pv. iornuto
on tomato plants. Appl. Environ. Microbiol. 61 : 3530-3536.
De Bruijn. F.J. 1992. Use of repetitive extragenic palindrornic and enterobacterial repctitive
intergenic consensus sequences and the polymerase chain reaction ro tingerprint the genomes
of RlzCohitrm ~rreiiioti isolates and other soi1 bacteria. Xppl. Environ. -Microbiol. 58: 2 180-
11 57.

57
De Souza -M. L.. D. Newcombe. S. Alvey. D. E. Crowley, A. Hay. -M. J. Sadowsky, and L.
P. Wackett. 1998. Moiecular basis of a bacterial consortium: Intenpecies catabolism of
atrazine. Appl. Environ. Microbiol. 64: 178-1 84.
De Souza M. L.. M. J. Sadowsky. and L. W. Wackett. 1996. -4trazine chlorohydrolase tiom
Pseti~lornonns sp. strain ADP: gene sequencr. enzyme purification. and proiein
characterization. J. Bacteriol. 178: 489 J--NOO.
De So~iza. kt. L.. L. P. Wackett. K. L. Boundy-Mills. R. T. Manddbaurn. and M. J.
Sadowsky. 1995. Cloning, characterization. and expression of a gene region from
Psericlornorim sp. strain ADP invo lved in dec hlorination of ritrazine. Appl. Environ.
Microbiol. 61: 3373-3378.
De Souza. M. L.. L. P. Wackett. and 41. J. Sadowsky. 1998. The 1ic.4BC genes encodin~
atrizine catabolisrn are located on a sel f-transmissible plasmid in Pseudonionas sp. strain
ADP. Appl. Environ. Microbiol. 64: 323-2324 .
Eagle, D. J.. J. L. O. Jones, E. J. Jewell. and R. P. Paxton. 1991. Determination of pesticides
by gas chromatography and high-pressure Liquid chrornatography. Marcel Dekker, Inc. Y.
Y . . N. Y .

88
Eaton. R. W.. and J. S. K m s . 1991. Cloning and cornparison of the DNA encoding
amrnelide aminohydro lase and cyanunc acid arnidohydro lase from three s-triazine-degrading
bacterial strains. J. Bacteriol. 173: 1363- 1 366.
Entry. J. A.. and W. H. Emrningham. 1995. The influence ofdairy manure on atrazine and
2.4-dichloropheno?tyacetic acid mineralization in Pasture soils. C m . J. Soi1 Sci. 75: 379-333.
Erickson. E. L.. and K. H. Lee. 1989. Degradation of atrazine and related s-triazines. Crit.
Rev. Environ. Contam. 19: 1-13.
Figurski. D. H.. and D. R. Helinski. 1979. Replication of an orijin-containing derivative
of piasmid RKZ dependent on a plasrnid hc t ion provided in tram. Proc. 'latl. Acad. Sci.
76: 1648- 1651.
Frank. R.. and G. J. Skons. 1 983. Dissipation of a m i n e kom soil. Bull. Environ. Contarn.
Toxicol. 34: 541-548.
Fulthorpe, R. R., and R. C. Wyndham. 1991. Transfer and expression of the catabolic
plasmid pBRC6O in wild bacterial recipients in a freshwater ecosystem. Appl. Environ.
Microbiol. 57: 1546- 1553.

89
Giardi, M. T., M. C. Giardina and G. Filacchioni. 1985. Chernical and biological
degradation of prirnary metabolites of' anazine by a iVocczrdia strain. Agit. Biol. Chem. 49:
1551-1558.
Giardina M. C., -M. T. Giardi, and G. Filacchioni, 1980. 4-arnino-3-chloro- 1.3.5-triazine:
.A new metabolite of atrazine by a soi1 bacterium. .A,sric. Biol. Chem. 44: 2067-2072.
Giardina -M. C.. _M. T. Giardi. and G. Filacchioni. 1982. Atrazine metabolism by .Vocnrciia:
elucidation of initial pathway and synthesis of potential metabolites. Agric. Biol. Chem. 46:
1439-1445
Golovleva. L. A.. N. R. Pertsova. L. 1. Ec-tushenko. and B. P. Baskunov. 1990. Degradation
of 2.4, 5-tnchlorophenoxyacetic acid by .Voccirdiozdrs sirnplex culture. Biodegradation. 1 :
263-271.
Goodrich. J. A.. B.W. Lykins, Jr.. and R. M. Clark. 1991. Drinking water from
agriculturally contarninated groundwater. J. Environ. Qual. 20: 707-71 7.
Gysin. H.. and E. Knuesli. 1960. Chernistry and herbicidal properties of triazine derivatives.
Adv. Pest Control Res. 3289-358.

90
Herrick. J. B.. K. G. Stuart-Keil. W. C. Ghiorse, and E. L. ,Madsen. 1997. 'faturai horizontal
transfer of a naphthalene dioxygenase gene beiween bacteria native to a coai rar-
contarninated field site. .Appl. Environ. .Microbiol. 63: 2330-2337.
Hillis. D. M.. and S. K. Davis. Evolution ofribosomal DNA: fiey million years of recorded
h i s t o ~ in the frog senus Runa. 1986. Evolution. 40: 1275- 1288.
Hulton. C. S. J.. C. F. Higgins, and P. W. Sharp. ERIC sequences: a novrl family of
repetitive elements in the jenomes of Escherichia coli. Salmonella ryhimcu-irrrn and other
enterobacteria. 199 1. -Mol. Microbiol. 5: 525-334.
Jessee. J. A.. A. C. Hendnc ks, G. C. Allen. and J. L. Neal. 1 983. .'uiaero bic degradation of
cyanuric acid. cpsteine. and atrazine by a facultative bacterium. Appl. Environ. Microbiol.
45: 97- 102.
Jones. T. W.. W. .M. Kemp. J. C. Stevenson. and J.C. Means. 1982. Degradation of ntrazine
in estuarine watedsediment systems and soils. J. Environ. Qual. 1 1 : 633-638.
Jordan. L. S., W. J. Farmer, I. R. Goodin. and B. E. Day. 1970. Nonbiological detoxication
of the s-tnazine herbicides. Res. Rev. 32: 267-285.

9 1
Judd. A. K.. M. Schneider, M J. Sadowsky, and F. J. De Bruijn. 1993. Use ofrepetitive
sequences and the ploymerase chah reation technique to classifi genetically related
Brnrtvrhizobi~tm jnponicurn serocluster 123 strains. Appl. Environ. Microbiol. 59: 1702-
1708.
Jutzi. K.. -4. M. Cook. and R. Hütter. 1982. The degradative pathway of the s-triazine
melamine. Biochem. J. 208: 679-654
Kado. C. 1.. and S. T. Liu. 1981. Rapid procedure for detection and isolation of large and
small plasmids. J. Bactenol. 145: 1365- 1373.
Kaufman. D. D., and I. Blake. 1970. Degradation of atrazine by soi1 fungi. Soi1 Biol.
Biochem. 3: 73-50
Kaufman, D. D., and P. C. Keamey. 1970. 'ulicrobial degradation of s-triazine herbicides.
Residue Rev. 32: 235-265.
Kello, D. 1989. LW0 drinking water quality guidelines for selected herbicides. Food
Addit. Contam. 6(Suppl.): S79-S85.
King, E. O., M. K. Ward, and D. E. Raney. 1954. Two simple media for the demonstration
of pyocyanin and fluorescein. J. Lab. Clin. Med. 44: 301-307.

Kutlu, K., and D. A. Cuppels. 1997. Development of a diagnostic probe for Xantharnonads
causing bacterial spot of peppers and tomatoes. Appt. Environ. Microbio 1. 63 : 44624470.
Lane, D. J. 1991. 16S/23S rkVA sequencing. p. 133 In Stackebrandt, E. and M.
Goodfellow. (eds.). Nucleic Acid Techniques in Bacterial Systematics. John Wiley and Sons
Ltd. New York, hY.
Lane. D. J.. B. Pace, G. J. Olsen, D. A. Stahl. M. L. Sogin. and N. R. Pace. 1982. Rapid
determination o f 16s ribosomal E2NA sequences for phylogenetic analysis. Proc. Natl. Acad.
Sci. CSA. 82: 6955-6959.
Lapage. S. P., and T. G. Mitchell. 1970. Media for the maintenance and presewation of
bacteria p. 1-134. In J. R. Norris and D. W. Ribbons (ed.), Methods in Microbiology. Voi.
3A. &ademic, New York, NY.
Lorenz. M. G., and Wackemagel. 1994. Bacterial gene transfer in the environment.
Microbiol. Rev. 58: 563-602.
Lupski. J. R., and G. M. Weinstock. 1992. Short, interspersed repetitive DNA sequences
in prokaryotic genomes. .J. Bacteriol. 174: 4525-4529.

93
Lynch, M. J., A. E. Wopat. and M. L. O'Connor. 1980. Characterization of two new
facultative methanotrophs. Appl. Environ. Microbiol. 40: 400-407
Madigan. M. T.. 1. 41. Martinko. and J. Parker. 1997. Microbial jenetics. p. 329-33 1. IN
B io l o g of .lIicroorganisrns. Prentice-Hall. Lrpper Saddle River. NJ.
blandelbaurn. KT.. D. L. -\ilan and L. P. Wackett. 1995. lsoiation and characterization
of a Pseudonionas sp. that mineraiizrs the s-triazine herbicide atrazine. Appl. Environ.
Microbiol. 61: 145 1-1157.
Mmdelbaum. R. T.. L. P. Wackett. and D. L. Allan. 1993. blineralization of the s- tnazine
ring of atrazine by stable bacterial mixed cultures. Appl. Environ. Microbiol. 59: 1695-
1701.
Martin, B..O. Humbert, M. Carnÿra. E. Guenzi. J. Walker. T. Mitchell, P. Andrew. 41
Prudhomme, G. Alloing, R. Hakenbeck. D. A. SLorrison. G. J. Boulnois. and J-P. Clave-S.
L992. A highly conserved repeated DNA elernent located in the chromosome of
Sireprococciîs pnertmoniae. Xucleic Acids Res. 20: 3 479-3 483.
Mindell, D.P., and R. L. Honeycutt. 1990. Ribosornal RNA in vertebrates: evolution and
phylogenetic applications. Annu. Rev. Ecol. Syst. 21 : 54 1-66.

94
Mirgain, I., G. A. Green, and H. Monteil. 1993. Degradation of atrazine in iaboratory
microcosms: isolation and identification of the biodegrading bacteria. Environ. Toxicol.
Chem. 12: 1627-1634.
Moritz, C.. and D. M. Hiliis. 1996. MolecuIar Systematics: Context and
Controversies. p. 1-7. In D. M. Hillis. C. Moritz, and B. K. Mable (ed.). klolecular
Systematics Second Edition. Sinauer Associates. Inc., Sunderland. MA.
Moscinski. J. K.. K. K. Jayachandran, and T. B. Moorman. 1996. Mineralization of the
herbicide atrazine by rlgrobacreritm ~czdiobacrer. abstr. 4-4 1 4, p. 458. In Abstracts of the
96"' General Meeting of the b e r i c a n Society for microbiology. Washington. D. C.
blulbry, W. 1994. Purification and characterization of an inducible s-triazine hydrolase
from Rhodococcza cordinrw NRRL B- 1 5444R. Appl. Environ. LMicrobiol. 60: 6 13-6 1 8.
Nagy. I., F. Compemolle, K. Ghys, J. Vanderleyden and R. De Mot. 1995. X single
cytochrome P-450 system is involved in degradation of the herbicides
EPTC (s-ethyldipropylthiocarbamate) and atrazine by Rhodococctts sp. Strain M86/2 1.
Appl. Environ. Microbiol. 61 9056-2060.

95
Nagy, I., G. Schoofs. F. Compemolle. P. Proost. J. Vanderleyden. and R. De Mot. 1995.
Degradation of the thiocarbamate herbicide EPTC (s-ethyl dipropylcarbamothioate) and
biosafening by Rhodococcus sp. strain NI86i21 invole an inducible cytochrome P-450
system and aldehyde dehydrogenase. J. Bacteriol. 177: 676-687.
Nations. B. K.. and G. R. Halberg. 1992. Pesticides in Iowa Precipitation. I. Environ. Quai.
31 : 356-492.
Pooler, ,M. R., D. F. Ritchie, and J. S. Hartung. 1996. Genetic re[ationships amonp strains
of .lCnnrhomonasfragarine based on random arnpiified polymorphic DNA PCR, repetitive
extragenic palindromic PCR. and entcrobactetial repetitive intergenic consensus PCR data
and generation of multiplexed PCR pnmers useful for the identification of this
phytopathogen. Appl. Environ. Microbiol. 62: 3 12 1-3 127.
Radosevich, M., S. I. l'raina, Y.Hao, and 0. H. Tuovinen. 1995. Degradation and
mineralization of atrazine by a soi1 bacterial isolate. Appl. Environ. blicrobiol. 61 : 297-307.
Raja., J., K. Valli, R. E. Perkins, F. S. Sariaslani, S. M. Barns, A. L. Reysenbach, S. Rehrn,
M. Ehringer, and N. R. Pace. 1996. ~Mineralization of 2, 4, 6-trinitrophenol (picnc acid):
characterization and phylogenetic identification of microbial strains. J. Ind. Micro bio 1. 16:
3 19-324.

96
Sadowsky, M. J., Z. Tong, M. De Souza. and L. P. Wacken. 1998. AtzC is a new member
of the arnidohydrolase protein superfamiiy and is homologous to other atrazine-metabolizing
enzymes. J. Bacteriol. 180: 152- 158.
Sambrook. L, E. F. Fritsch, and T. Maniatis. 1989. Molecular Cloning: a laboratop manual.
2nd ed. Cold Spring Harbor Laboratory Press, Cold Spring Harbor. N. Y.
Sandhya. S., S. K. Prabu, and R. B. T. Sundari. 1995. blicrobial degradation o f
dibenzo thiop hene by :Vocarrlioicies. J'. Environ. Sci. Health. 30: 1995-2006.
Selvaraj. G.. and V. 'I. Iyer. 1983. Suicide plasmid vehicle for insertion mutagenesis in
Rhizobirrrn meliloti and related bacteria J. Bacteriol. 156: 1297- 1300.
Shao, 2. Q.. and R. Behki. 1995. Cloning of the genes for degradation of the herbicides
EPTC (s-ethy l diprop y lthiocarbamate) and atrazine ftom Rhodococars sp. S train TE 1 . App 1.
Environ. Microbiol. 61 : 206 1-1065.
Shao, Z. Q., and R. Behki. 1996. Characterization of the expression of the rhcB gene.
coding for a pesticide-depding cytochrome PA50 in Rhodococnrs strains. Appl. Environ.
~Microbiol. 62: 403-407.

Shao. Z.Q.. W. Seffens, W. Mulbry. and R. M. Behki. 1995. Cloning and expression of the
s-tnazine hydrolase gene ( i c A ) From Rhodococcus corczilinits and development of
Rhodococnis recombinant strains capable of dealkylating and dechlorinating the herbicide
atrazine. J. Bacteriol. 177: 5748-5755.
Shaw. J. E. (ed.). 1996. Guide to weed control. Publication 73. Ontario blinistry of
A-g-iculture, Food and Rural Affairs. Queen's Printer. Ottawa. ON.
Srnibert. R. .M.. and N. R. Krieg. 198 1. Chapter 20: General charactenzation. p. 01 Gerhardt.
P.. 41urray. R. G. E.. Costilow. R. Y.. 'lester. E. W.. Wood. W. -4.. Kneg, N. R.. and G. B.
Phillips. (ed. ), Manual of Methods for Generai Bactenology. Washington D.C.: ASM.
Spencer. E. Y. 1982. Guide to the chernicals used in crop protection. Canadian
Government Publishing Centre. Ottawa. Canada.
Stanisich, V. A. 1985. Identification and analysis ofplasmids at the genetic level. p. j ?tl
Sennett. P. M., and J. Grinsted. (eds.), Methods in Microbiology. Acadernic Press hc..
Orlando, Florida.
Steiert, J. G., and R. L. Crawford. 1986. Catabolism of pentachlorophenol by a
Flavobacreriurn sp. Biophys. Res. Commun. 141 : 825-830.

98
Suslow, T. V., M. N. Schroth, and .M. isaka. 1982. .Application of a rapid method for Gram
differentiation of plant pathogenic and saprophytic bactena without staining.
Phytopathology. 72: 9 17-9 18.
Topp, E., D.W. Gutman. B. Bourgoin, I. Millette, and D. Gamble. L995. Rapid
mineralization of the herbicide atrazine in alluvial sediments and enrichment cultures.
Environ. Toxical. Chem. 14: 733-747.
Topp, E., R. S. Hanson. D. B. Rjngelberg, D. C. Wlite, and R. Wheatcro fi. 19%. Isolation
and charac terization of an .V-methylcarbamate insecticide-degrading methy 10 trop hic
bacteriurn. Appl. Environ. Microbiol. 59: 3339-3349.
Topp, E., L. Tessier, and E. G. Gregorich. 1996. Dairy rnanure incorporation stimulates
rapid atrazine mineralization in an agicultural soil. Can. 1. Soi1 Sci. 76: 4-10.
Topp, E., L. Tessier, and M. Lewis. 1997. Characterization of atrazine-degradin bacteria
isolated fiom agncultural soil. abstr. Q-407, p. 523. Absûacts of the 97Ih Gcneral Meeting
of the Arnerican Society for Microbiology. Washington. D. C.
Topp. E., H. Zhu, M. Lewis, and D. Cuppels. 1998. Triazine herbicide degradation in soils.
abstr. SAZ, p. 62. Programme and abstracts of the 48th h u a 1 Canadian Society of
Microbiologists. Guelph, ON.

C.S. Environmental Protection Agency. 199 1. Rules and regulaticlns. Fed. Regist. 56:
Versalovic, J.. T. Koeuth. and I. R. Lupski. 199 1. Distribution of repetitive DNA sequences
in eubacteria and application to fingerprinting of bacterial genomes. Nucleîc Acids Res. 19:
6823-683 1.
Versalovic, J., M. Schneider. F. J. De Bmijn. and J. R. Lupski. 1994. Genomic
fingerprintinj of bacteria using reprtitive sequence-based polymerase chain reaction.
blethods Mol. Ce11 Biol. 5: 3 -40 .
Vidaver, A. K. 1967. Synthetic and cornplex media for the rapid detection of fluoresence
O t^ phyopathogenic pseudornonads: efiect of the carbon source. App 1. Micro biol. 15: 1 5 23 -
15%.
Weisburg, W. G., S. M. Barns, D. A. Pelletier and D. J. Lane. L99 1. 16s ribosomal
DNX amplification for phylogenetic study. J. Bacteriol. 173: 697-703-
Weller, R., and D. M. Ward. 1989. Selective recovery of 16s rRKA sequences fkom Natural
rnicrobial cornrnunities in the form of cDNA. Appl. Microbiol. 55: L 8 1 8- 1 822.

1 O0
Wheaicrofi, R., and P. A. Williams. 198 1. Rapid methods for the study of both stable and
unstabie plasmids in Pse~rdornonas. J. Gen. Microbiol. 1 24: 433-43 7.
Xun. L., and C. S. Orser. 1991. Purification and properties of pentachlorophenol
hydroxylase, a flavoprotein £Yom Flavobnc~eriurn sp. strain ATCC39723. J. Bacteriol. 1 73:
44-47-4453.
Yoon. J.. S. Rhee, J. Lee, Y . Park, and S. Lee. 1997. .Vocnrc/ioi<les pyriciino&ricrts sp.
Nov., a pyridine-degrading bacterium isolated korn the oxic zone of an oil shale column.
Int. J. Syst. Bacteriol. 47: 933-938.
IMAGE EVALUATION TEST TARGET (QA-3)
APPLIED - IMAGE. lnc = 1653 East Main Street - ,=- Rochester. NY 14609 USA -- --= Phone: 71614826300 -- --= Fax: 71 W2M-5989
O '993. A@& Image. Inc.. All Rights Raserved