mendel’s stem length gene (le) encodes a …€™s stem length gene (le) encodes a gibberellin...
TRANSCRIPT

The Plant Cell, Vol. 9, 1435-1443, August 1997 O 1997 American Society of Plant Physiologists
Mendel’s Stem Length Gene (Le) Encodes a Gibberellin 3P-Hydroxylase
Diane R. Lester,’ John J. ROSS, Peter J. Davies,2 and James 6. Reid Department of Plant Science, University of Tasmania, Hobart, Tasmania 7001, Australia
We describe the isolation of the Le gene of pea, which controls internode elongation and originally was described by Mendel. Heterologous screening of a pea cDNA library yielded a partia1 clone that was 61 ?40 identical to coding regions of the putative Arabidopsis gibberellin 30-hydroxylase gene, GA4. DNA gel blot analysis with this cDNA revealed a Hindlll restriction fragment length polymorphism between pea isolines differing at Mendel’s Le locus. Genomic clones of the GACrelated gene were isolated from the Le and /e isolines. Polymerase chain reaction combined with restriction fragment length polymorphism analysis were used to show that the gene mapped to the Le locus. A cDNA containing a complete open reading frame of the pea GACrelated gene was amplified by polymerase chain reaction from each iso- line. Recombinant expression in Escbericbia co/i demonstrated that the product of the Le cDNA was a gibberellin 30- hydroxylase that is able to convert GA,, to the bioactive GA,. Substantially reduced levels of gibberellin 3P-hydroxylase activity were measured, after expression of the /e cDNA, by using identical methods. This reduced activity was associ- ated with an alanine-to-threonine substitution in the predicted amino acid sequence of the enzyme near its proposed active site.
INTRODUCTION
Our understanding of the basis of heredity was derived orig- inally from seven genes of the garden pea. These genes control the traits followed by Gregor Mendel in breeding ex- periments from which he established the laws of inheritance (Mendel, 1866). The traits are seed shape (Rlr), cotyledon color (/ l i ) , seed and flower color (Ala), pod shape (Vlv), pod color (Gplgp), flower position (Falfa), and stem length (Lelle) (Mendel, 1866; White, 1917). Full recognition of Mendel’s work came belatedly and posthumously in 1900 (Bateson, 1901). Fortunately, however, the mutants with which he had worked were readily identified because they were well known and widely dispersed in Europe during the last cen- tury (White, 1917). To date, only one of Mendel’s genes has been cloned: gene R (Rugosus), controlling the round seed trait, encodes a starch branching enzyme (Bhattacharyya et al., 1990).
The stem length gene Le principally controls the length between nodes of the pea plant (White, 1917). Plants ho- mozygous for the recessive /e allele are dwarfed and clearly distinguishable from the wild type and heterozygous rela- tives: “Even in their quite young state they can be easily picked out by their compact growth” (Mendel, 1866).
’To whom correspondence should be addressed. E-mail d.lesterO utas.edu.au; fax 61 -3-6226-2698. *0n leave from the Section of Plant Biology, Cornell University, Ithaca, NY 14853.
A more detailed understanding of the Le gene eluded researchers until relatively recently, when the physiology of its action was determined. Evidence that it might act via the gibberellin (GA) pathway came with the measurement of substantially reduced endogenous GA,-like activity in /e plants (Potts et al., 1982). Subsequent findings that dwarf plants were unable to convert radiolabeled GA20 to GA, and that their phenotype could be restored to that of the wild type by the application of GA, indicated that Le controlled GA biosynthesis at the step of 3p-hydroxylation (Ingram et al., 1984). A direct relationship between GA 3p-hydroxy- lation and plant height emerged in pea (Ross et al., 1989) and in other species through the study of the analogous mutations dwarfl of maize, dy of rice, and ga4 of Arabidop- sis (Spray et al., 1984; Kobayashi et al., 1989; Talon et al.,
The simplest interpretation of the effect of /e is that the al- lele represents a mutation of a GA 3p-hydroxylase gene (Ross et al., 1989). Nonetheless, the possibility that Le is a regulatory gene cannot be excluded. The fortuitous isolation by transposon tagging of a putative GA 3 p-hydroxylase gene, GA4, from Arabidopsis (Chiang et al., 1995) provided a tool with which to investigate these theories. This study de- scribes the isolation and characterization of a GA Sp-hydrox- ylase gene from pea. It presents evidence that we have cloned a second of Mendel’s classic genes, Le, and ex- plores the molecular basis of the dwarf trait that he studied.
1990).

1436 The Plant Cell
RESULTS
Isolation of a GA4-Related Gene from Le and leLines of Pea
Low-stringency screening of a pea (cv Alaska; Le) Xgt11cDNA library with a portion of the GA4 cDNA produced apartial cDNA, pODSc, that showed homology with 2-oxoglu-tarate-dependent dioxygenases (2ODDs), the family of en-zymes to which the GA 3(3-hydroxylase belongs (Lange andGraebe, 1993).
The phenotypes of the Le (tall) and le (dwarf) pea isolinesare shown in Figure 1. DMA gel blot analysis with pOD3c gavea similar pattern of hybridization in each isoline for bothEcoRI and EcoRV digests, as shown in Figure 2. A restric-tion fragment length polymorphism (RFLP) was detected byHindlll between the two lines. No faintly hybridizing bandswere observed under low-stringency hybridization conditions.
The 4.25-kb EcoRI fragment that hybridized with pODScwas isolated from the le line via a subgenomic library andsequenced to give a map of the pea G/W-related gene andflanking regions, as shown in Figure 3. A putative open read-ing frame containing a single intron was identified: it is lo-cated centrally in the fragment. Identification of the mostlikely translation initiation codon, ATG, was based on the ex-
EcoRILe le
Hindlll EcoRVLe le Le le
Figure 1. Mendel's Stem Length Trait.
Lines 205f (Le/Le; tall) and 205 (te//e; dwarf) of pea show the differ-ence in stem length controlled by the le locus.
kb8.5-
4.8-
3.6-2.8-
1.9-
Figure 2. Genomic DMA Gel Blot Showing an RFLP in the Pea GA4-Related Gene.
Shown is the result of DMA gel blot analysis of 15 |xg of genomicDMA from isolines differing at the Le locus digested with EcoRI,Hindlll, and EcoRV, probed with the G/44-related cDNA of pea, andwashed at low stringency. Numbers at left indicate the length of thefragments in kilobases.
pected approximate length of the N-terminal region of theprotein (estimated from comparisons with the ArabidopsisGA4 cDNA sequence and other 2ODD cDNA sequences) aswell as the presence of a putative TATA box 80 bp up-stream. The exact position of the intron was determined bya comparison of the cDNA and genomic sequences and alsoby the presence of putative splice sites. It is 544 bp long,compared with 433 bp for the intron in the Arabidopsis GA4gene, and is located at a similar position in the coding region.
A 2.2-kb fragment encompassing the coding region, theTATA sequence, and some of the flanking sequence of theG/W-related gene from the Le line was isolated by poly-merase chain reaction (PCR) amplification (Figure 3). Bycomparing the open reading frame of the Le allele with Gen-Bank sequences, we determined that this sequence is mosthighly matched with the GA4 cDNA. The 1122-bp nucleotidesequence is 61 % identical to the GA4 cDNA, which trans-lates to an identity of 53% for the predicted 374-amino acidprotein. An alignment of the deduced amino acid sequencesof the two genes is shown in Figure 4.
A comparison of the alleles of the pea G/W-related genefrom the Le and te lines in the region of their overlap re-vealed two minor sequence differences. The possibility thatthese differences represented PCR artifacts in the Le allelewas minimized by performing duplicate reactions and usingan enzyme mix with proofreading ability. The intron in thegene from the le line contained an extra two nucleotidebases, AT, at position 1954 that were not present in the in-tron of the Le gene. Nucleotide A was present at position2654 in the te genomic fragment instead of a G residue atthe same position in the wild-type Le gene. This change waspredicted to replace the amino acid alanine at residue 229with threonine in the te gene product (Figure 4).

Linkage Analysis of the Le Locus
The 4.25-kb EcoRl genomic fragment carrying the pea GA4- related gene isolated from the /e line contained two Hindlll sites (Figure 3). Restriction mapping predicted that one of these, situated 900 bp upstream of the gene's coding regions, corresponded to the site that produced the RFLP seen in the DNA gel blot analysis. A 1.2-kb fragment spanning this site was amplified by PCR from each isoline and then di- gested with Hindlll. The PCR product of theLe plant remained intact after Hindlll digestion, whereas two bands of 1 .O and 0.2 kb resulted from the /e product, confirming the presence of a Hindlll polymorphism between alleles (Figure 3).
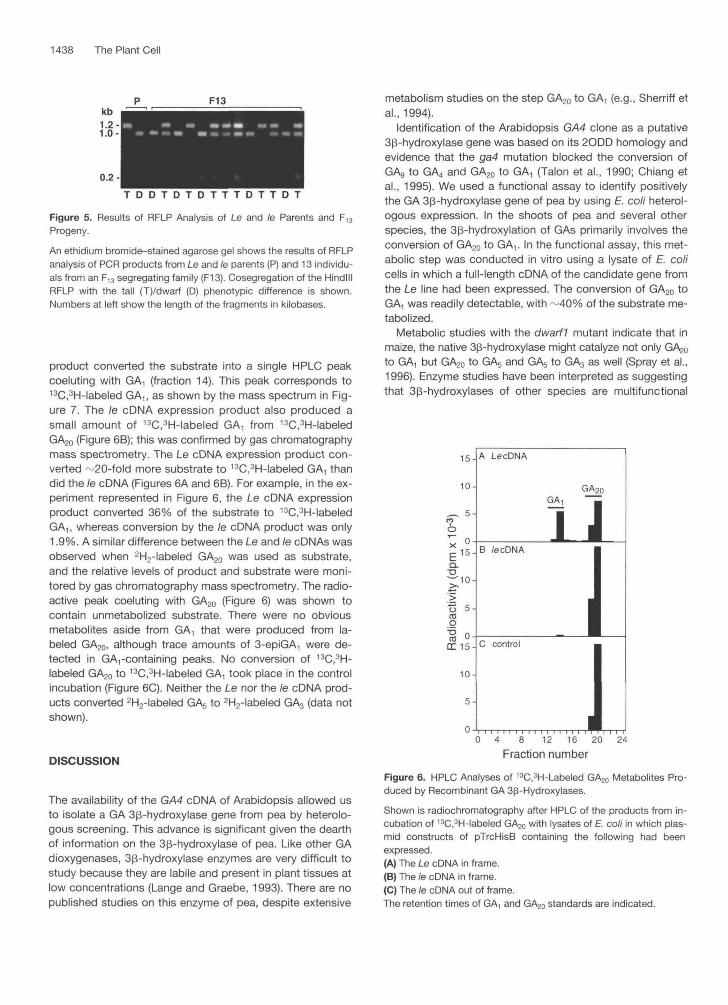
RFLP analysis of the aforementioned PCR products of 40 plants from two FI3 segregating families demonstrated co- segregation of the Hindlll polymorphism with the tall/dwarf phenotypic difference. All 12 dwarf plants in this population, like the dwarf parent, had the 1.0- and 0.2-kb bands only. Twenty-one tal1 plants had all three bands of 1.2, 1 .O, and 0.2 kb, whereas seven tal1 plants displayed the 1.2-kb band only (x2, 1:2:1 = 1.35; P > 0.5). Thus, the GA4-related gene of pea mapped to the Le locus. The results of the RFLP analysis of the Le and /e parents and 13 individuals from an FI3 segregating family are shown in Figure 5.
Functional Assay of Le and /e Recombinant Gene Products
The genomic sequences enabled us to design PCR primers predicted to amplify the complete open reading frame of the pea GA4-related gene from a template of cDNA. These primers amplified the expected 1 . l -kb product from both Le
Le --------- I I I
I I I I I EcoRl EcoRV
I Hindlll EcoRl Hindlll
/e I
--c c Od302 od306 -
1 kb
Figure 3. Map of the GA4-Related Pea Gene and Flanking Regions from Le and /e Genotypes.
The two exons of the gene are represented by the boxed regions. The locations of EcoRI, Hindlll, and EcoRV restriction sites are indi- cated for both fragments. The portion of the EcoRl fragment from the Le gene that was sequenced is indicated by unbroken lines. The restriction map of the Le gene outside of sequenced regions was deduced using the results of the genomic DNA gel blot analysis (Fig- ure 2). The region of both alleles containing the polymorphic Hindlll site that was amplified by PCR and subjected to RFLP analysis is defined by the positions of PCR primers od302 and od306. This GA4-related gene of pea was shown to encode a GA Sp-hydroxy- lase (see text) and has the GenBank accession number U93210.
Le Encodes a Gibberellin 3p-Hydroxylase 1437
33 35
68
69
C P H A K L I G P T K P P L Y R S V T W N E Y L G T K A K H F N K A L 348 S i V P U V S U V E S ~ l O ~ l K ~ H ~ T ~ 349
S S V R L C T P I N G L F O V N D S N K N S V O V G 374 Pea3Bh O M I i N H R E E . . . . . . . . . . . . . . . . . 358 Ara GA4
Figure 4. Alignment of Deduced Amino Acid Sequences of the Pea GA 3P-Hydroxylase Gene and the Arabidopsis GA4 cDNA.
The pea 36-hydroxylase (pea 3ph) sequence represents the Le (wild type) gene. The intron position (indicated by the arrow) was confirmed by the sequence of a cDNA with the GenBank accession number U85045. The GA4 sequence (Chiang et al., 1995) has the accession number L37126 in volume 1 O0 of GenBank. ldentical resi- dues of both sequences are boxed. Residues showing high conser- vation throughout plant 20DDs are marked with triangles. These were identified from a multiple alignment of flavonol synthase from petunia (GenBank accession number S67953), flavanone 3p-hydrox- ylase from maize (GenBank accession number U04434), GA 20-oxi- dase from Arabidopsis (EMBL accession number X53381), pea (GenBank accession number U58830), and spinach (GenBank ac- cession number U33330) and GA4 from Arabidopsis (GenBank ac- cession number L37126). The three residues that are invariant throughout plant 20DDs and related enzymes whose side chains act as iron ligands are marked with circles. The predicted protein of the /e GA 36-hydroxylase gene has a threonine instead of the alanine at position 229 (marked with an asterisk). Dots indicate spaces in- serted to conserve the alignment. Ara, Arabidopsis.
and /e lines. Both cDNAs were ligated into pTrcHisB and se- quenced to confirm sequence data obtained from the ge- nomic clones and also to confirm that their frame and orientation were correct for expression. Escherichia coli was transformed with these constructs and also, as a negative control, with pTrcHisB containing the /e cDNA in the oppo- site orientation.
The results obtained from incubation of i3C,3H-labeled GA,, with lysates of cells in which each plasmid construct had been expressed are shown in Figure 6. The Le cDNA
1438 The Plant Cell
F13
0.2-
T D D T D T D T T T D T T D T
Figure 5. Results of RFLP Analysis of Le and te Parents and F13Progeny.
An ethidium bromide-stained agarose gel shows the results of RFLPanalysis of PCR products from Le and te parents (P) and 13 individu-als from an F,3 segregating family (F13). Cosegregation of the HindlllRFLP with the tall (T)/dwarf (D) phenotypic difference is shown.Numbers at left show the length of the fragments in kilobases.
product converted the substrate into a single HPLC peakcoeluting with GA, (fraction 14). This peak corresponds to13C,3H-labeled GA,, as shown by the mass spectrum in Fig-ure 7. The le cDNA expression product also produced asmall amount of 13C,3H-labeled GA, from 13C,3H-labeledGA20 (Figure 6B); this was confirmed by gas chromatographymass spectrometry. The Le cDNA expression product con-verted ~20-fold more substrate to 13C,3H-labeled GA, thandid the te cDNA (Figures 6A and 6B). For example, in the ex-periment represented in Figure 6, the Le cDNA expressionproduct converted 36% of the substrate to 13C,3H-labeledGA,, whereas conversion by the te cDNA product was only1.9%. A similar difference between the Le and te cDNAs wasobserved when 2H2-labeled GA20 was used as substrate,and the relative levels of product and substrate were moni-tored by gas chromatography mass spectrometry. The radio-active peak coeluting with GA20 (Figure 6) was shown tocontain unmetabolized substrate. There were no obviousmetabolites aside from GA, that were produced from la-beled GA20, although trace amounts of 3-epiGA, were de-tected in GA,-containing peaks. No conversion of 13C,3H-labeled GA20 to 13C,3H-labeled GA, took place in the controlincubation (Figure 6C). Neither the Le nor the le cDNA prod-ucts converted 2H2-labeled GAs to 2H2-labeled GA3 (data notshown).
DISCUSSION
The availability of the GA4 cDNA of Arabidopsis allowed usto isolate a GA 3p-hydroxylase gene from pea by heterolo-gous screening. This advance is significant given the dearthof information on the 3p-hydroxylase of pea. Like other GAdioxygenases, 3p-hydroxylase enzymes are very difficult tostudy because they are labile and present in plant tissues atlow concentrations (Lange and Graebe, 1993). There are nopublished studies on this enzyme of pea, despite extensive
metabolism studies on the step GA20 to GA, (e.g., Sherriff etal., 1994).
Identification of the Arabidopsis GA4 clone as a putative3p-hydroxylase gene was based on its 2ODD homology andevidence that the ga4 mutation blocked the conversion ofGA9 to GA4 and GA20 to GA, (Talon et al., 1990; Chiang etal., 1995). We used a functional assay to identify positivelythe GA 3p-hydroxylase gene of pea by using E. coli heterol-ogous expression. In the shoots of pea and several otherspecies, the 3p-hydroxylation of GAs primarily involves theconversion of GA20 to GA,. In the functional assay, this met-abolic step was conducted in vitro using a lysate of E. colicells in which a full-length cDNA of the candidate gene fromthe Le line had been expressed. The conversion of GA20 toGA, was readily detectable, with ~40% of the substrate me-tabolized.
Metabolic studies with the dwarfl mutant indicate that inmaize, the native 3p-hydroxylase might catalyze not only GA^to GA, but GA20 to GA5 and GAg to GA3 as well (Spray et al.,1996). Enzyme studies have been interpreted as suggestingthat 3p-hydroxylases of other species are multifunctional
15-
10-
coo
5-
EQ."O
15-
5-(0g
rr 15
10-
5-
A LecDNA
GA20
GA,
B /ecDNA
C control
04 8 12 16 20 24
Fraction number
Figure 6. HPLC Analyses of 13C,3H-Labeled GA20 Metabolites Pro-duced by Recombinant GA 3(3-Hydroxylases.
Shown is radiochromatography after HPLC of the products from in-cubation of 13C,3H-labeled GA20 with lysates of E. coli in which plas-mid constructs of pTrcHisB containing the following had beenexpressed.(A) The Le cDNA in frame.(B) The te cDNA in frame.(C) The te cDNA out of frame.The retention times of GA, and GA20 standards are indicated.

Le Encodes a Gibberellin 3p-Hydroxylase 1439
' O 0 1
P 4 a
2 1 5 0
5071
I d l A
4 1 7 o 1, li I I '+ I' I I , " # '; IYL .1. h 200 2 5 0 3 0 0 350 400 440 500
MassKharge Ratio
Figure 7. Mass Spectrum of Derivatized 13C,3H-Labeled GA, Pro- duced by Recombinant GA 3p-Hydroxylase.
Shown is the mass spectrum of derivatized 13C,3H-labeled GA, from fraction 14 (see Figure 6) after HPLC analysis of products from the incubation of the Le cDNA expression cell lysate with 13C,3H-labeled GA20. The spectrum closely matches that of a GA1 standard; the Kovats retention indices were 2672 for the spectrum above and 2671 for the standard. (Only the 13C-ion shows in the spectrum be- cause of the low specific activity of the 3H.)
(Hedden and Kamiya, 1997). However, the pea recombinant 3p-hydroxylase did not convert GAZ0 to GA5 or GA5 to GA3 and showed no obvious signs of other activities, such as 2p- hydroxylation. The conversion of GA, to GA, does occur in pea shoots (Poole et al., 1995), but it seems that this step occurs either through the action of another enzyme or by nonenzymatic processes.
Consistent with the classification of the 3p-hydroxylases as 20DDs, the pea 3p-hydroxylase gene contains all of the domains characteristic of plant 20DDs (Prescott, 1993; de Carolis and de Luca, 1994; Figure 4). Beyond the 20DDs, these domains also classify the pea 3p-hydroxylase in a wider group of enzymes, which includes 1 -aminocyclopro- pane-1 -carboxylate oxidase and hyoscyamine-6s-dioxyge- nase from plants as well as isopenicillin N-synthase and deacetoxycephalosporin C synthase from fungi (de Carolis and de Luca, 1994; Roach et al., 1995).
Among all of the published sequences, coding regions of the pea GA 3p-hydroxylase gene display greatest identity with the Arabidopsis GA4 cDNA. The deduced amino acid sequences of the two genes are 53% identical. This leve1 of identity is considerably lower than the 74% average identity observed between species for a 20DD of the flavonoid bio- synthetic pathway, flavanone 3p-hydroxylase (Britsch et al., 1993). However, it is comparable with levels observed be- tween species for genes encoding GA 20-oxidases, which is the 20DD that catalyzes the previous step of the GA path- way (Hedden and Kamiya, 1997). In severa1 species, includ-
ing pea, 20-oxidase genes make up multigene families (Hedden and Kamiya, 1997). In contrast, the 3p-hydroxylase gene of pea appeared to be a single-copy gene. However, the existence of another relatively divergent 3p-hydroxylase gene cannot be excluded-the large genome size of pea (-20 times larger than Arabidopsis) might make such a gene undetectable by DNA gel blot analysis. Close similarities are evident between the structure of the pea 3p-hydroxylase gene and the Arabidopsis GA4 gene. Both contain single in- trons of roughly comparable size at the same position in their coding regions. One of two introns of the Arabidopsis GA 20-oxidase gene is also at this position (Xu et al., 1995).
Tight linkage between an RFLP and the tall/dwarf pheno- typic difference strongly implied that in cloning the GA 3p- hydroxylase gene, we had cloned Mendel's Le locus. This conclusion substantiates a body of evidence obtained from GA application and analytical and metabolic studies show- ing that Le controls GA 3p-hydroxylation (Ingram et al., 1984; Ross et al., 1992; Smith et al., 1992). It also disproves the theory that abnormal expression of an inhibitor of 3p- hydroxylase activity gives rise to the /e phenotype (Smith, 1 992).
In light of the RFLP result, we sought to demonstrate its corollary-the presence of a deleterious mutation in the le allele that could account for the dwarf phenotype. Another mutation at the Le locus, /ed (Ross and Reid, 1987), is rele- vant when considering the molecular basis of the /e muta- tion. The greater severity of the phenotype resulting from /ed indicates that /e is leaky, that is, some Le-like activity occurs in le plants.
In this study, all findings are consistent with the expecta- tion that at least a degree of 3p-hydroxylase activity is pre- served in le plants. The isolation of genomic clones from Le and le plants showed that there was no gross disturbance to the 3p-hydroxylase gene of dwarf plants. An extra 2 bp were present in the intron of the le 3p-hydroxylase gene, but this would not be expected to affect gene function. The se- quence of the le cDNA confirmed that a correctly spliced RNA transcript was present in /e plants. We found 3p- hydroxylase RNA to be undetectable in apical and expand- ing internode tissue by mRNA gel blot analysis, but quantita- tive PCR amplification of cDNA detected no marked difference in levels of 3p-hydroxylase RNA between Le and /e plants (data not shown). According to sequence data, the only difference between the Le and le predicted proteins was the replacement of alanine with threonine at position 229. Because this residue does not show high conservation throughout 20DDs, it cannot be immediately assigned im- portance in relation to function of the molecule, unlike the in- variant cysteine that is replaced by glycine in the ga4 mutant (Chiang et al., 1995). Although the substitution of threonine for alanine is not radical, it introduces a polar side chain in place of a nonpolar one.
Nevertheless, the functional assay revealed a stark differ- ence between the ability of the Le and /e cDNA products to convert GA,, to GA,. The amount of 3p-hydroxylase activity

1440 The Plant Cell
produced from E. coli expression of the le cDNA was -20- fold less than that produced by the Le cDNA under identical conditions. The finding of residual amounts of activity in the le gene product is consistent with the “leakiness” of the le mutation. The difference in activity between the Le and le re- combinant enzymes is comparable in magnitude to the im- pact of the le mutation on the metabolism of fed 13C,3H- labeled GA,, by intact plants (Ross et al., 1989; Sherriff et al., 1994). Endogenous GA1 levels show a smaller (10-fold) difference between Le and /e plants (Ross et al., 1992), which probably results from feedback regulation of the pre- ceding GA 20-oxidase and hence GA,, levels (Martin et al., 1996).
We found that although the recombinant enzymes were active in this system, they did not accumulate to levels de- tectable by protein staining of SDS-polyacrylamide gels. This was our experience previously with another pea 20DD of the GA pathway, the 20-oxidase, using the same expres- sion vector (Lester et al., 1996). It appears that in a soluble (and therefore presumably active) form, these enzymes are unstable at high concentrations in E. coli. Although it was not possible to check the concentrations of recombinant protein before the activity assay, we believe that compari- son of results between samples is meaningful because the E. coli expression system is highly controlled. Certainly, the magnitude of the difference in activity displayed between Le and le enzymes was closely reproducible. We were unable to check the molecular masses of Le and le recombinant 3P-hydroxylases as a consequence of their lack of accumu- lation. Therefore, all information regarding translation of the recombinant enzymes is based on predictions made from their DNA sequences.
The Le and /e recombinant proteins were predicted to be the same size, with only one difference in amino acid se- quence-the alanine-to-threonine substitution at residue 229. The markedly different levels of 3P-hydroxylase activity between the two proteins suggest that this substitution caused the reduction in activity of the le gene product. Thus, we examined more closely the possible role of Ala-229 in the function of the molecule. Although the altered residue it- self is not conserved throughout 20DDs, it is situated in a domain of the predicted protein where conserved residues are concentrated, as illustrated in Figure 4. Most notably, it is situated close to the submotif histidine-X-aspartic acid (His-X-Asp, where X is any amino acid) at positions 231 to 233, which is invariant in plant 20DDs, 1 -aminocyclopro- pane-1 -carboxylate oxidases, and other related enzymes (de Carolis and de Luca, 1994; Prescott and John, 1996). An exciting development in understanding plant 20DDs came recently with the proposal of a model for their action (Roach et al., 1995). According to this model, His-231 and Asp-233 of the pea 3P-hydroxylase are central to enzyme action, pro- viding two of the three amino acid side chains that coordi- nate the active site iron (Roach et al., 1995).
We have noted high consensus in regions flanking the His-X-Asp motif among sequences of the other 20DDs that
have been obtained from severa1 species, that is, flavanone 3P-hydroxylase and GA 20-oxidases (Britsch et al., 1993; Hedden and Kamiya, 1997). This consensus might reflect a role for these residues in enzyme function deriving from their proximity to the active site. The Arabidopsis GA4 sequence, like the wild-type pea 3P-hydroxylase, contains alanine at the position altered in the predicted le gene product. Possi- bly, the small nonpolar side chain of alanine is required in this position near the His-X-Asp motif for optimal3P-hydroxylase activity. However, more detailed analyses of protein struc- ture/function relationships in 20DDs are required to confirm that this relatively subtle alteration in the le 3P-hydroxylase impairs its activity and therefore that it represents the mo- lecular basis of the dwarf phenotype.
The subtlety of the le mutation contrasts with the nature of the mutation discovered in the first of Mendel’s genes to be cloned. The wrinkled seed trait arises from a mutation at the R locus in the form of an 800-bp transposon-like insertion (Bhattacharyya et al., 1990). The insertion is toward the 3‘ end of a starch branching enzyme gene and causes trunca- tion of the gene product (Bhattachatyya et al., 1990). The presence of this insertion was suspected from DNA gel blot analysis results and confirmed by the sequence of both al- leles (Bhattacharyya et al., 1990).
The 3P-hydroxylase gene is one of a number of structural genes controlling GA metabolism that has been isolated from pea recently (Lester et al., 1996; Martin et al., 1996; Ait- Ali et al., 1997). Because pea is well suited for hormonal studies, the availability of these clones is likely to accelerate progress toward an understanding.of GA metabolism and its regulation.
METHODS
Plant Material
The isolines 205+ (LelLe) and 205- (/e//e) of pea (Pisum sativum) were developed from the cross L2 x L77 at the University of Tasmania (Hobart, Tasmania, Australia). All Le and /e material used in this study was from these two lines. Two segregating F,, families from the L2 X
L77 cross were grown for restriction fragment length polymorphism (RFLP) analysis.
cDNA Library Screening
Two hundred and fifty thousand plaques of a Xgtl 1 cDNA library pre- pared from pea cultivar Alaska apical buds (RNA pooled from buds grown in 7 days of darkness and buds grown in 5 days of darkness and then 2 days of light [ClonTech, Palo Alto, CAI) were screened with a fragment spanning nucleotides 241 to 821 of the GA4 cDNA (Chiang et al., 1995). Filters were hybridized at 42°C in 2 x SSC (1 x SSC is 0.15 M NaCl and 0.015 M sodium citrate), 1% SDS, 20% for- mamide, and 10% dextran S04 and washed at 50°C in 2 x SSC and 0.1 % SDS. A single positive plaque was purified, and the insert was subcloned into the EcoRl site of pBluescript KS- (Stratagene, La

Le Encodes a Gibberellin 3p-Hydroxylase 1441
Jolla, CA). The DNA sequence of this clone, pOD3c, was determined from plasmid DNA by using dye-terminator cycle sequencing meth- ods (Applied Biosystems, Foster City, CA) and an automated se- quencer (model Prism 377; Applied Biosystems) at CSlRO Marine Laboratories (Hobart).
lsolation of Genomic Clones from Le and /e Lines
Genomic DNA was prepared by the method of Ellis (1994) from leaves of the /e line and digested completely with EcoRI. The di- gested DNA was size fractionated by agarose gel electrophoresis, and fragments in the range of 2.8 to 4.8 kb were purified and ligated into the EcoRl site of hgtl0. Phage were plated in Escherichia co/i KW251 and screened with inserl DNA of pODSc, which corresponds to nucleotides 180 to 1050 of the full-length Le cDNA (GenBank ac- cession number U85045). A single positive plaque was purified, and its insert was subcloned into the EcoRl site of pBluescript KS-. Se- quences of both strands of the insert were determined from plasmid DNA prepared using either Qiagen (Hilden, Germany) or Bresapure (Bresatec, Adelaide, Australia) plasmid purification kits, with se- quencing methods otherwise as described above. The sequence of the 4.25-kb /e fragment enabled us to design the primers OD3up and ODSdo, which encompassed the entire coding region of the GA4- related gene. These primers were used to amplify the corresponding region of the Le gene. Polymerase chain reaction (PCR) was per- formed with KlenTaq (ClonTech) in a volume of 20 pL containing 10 ng of genomic DNA and primers OD3up and OD3do at 0.4 yM, with the reaction conditions otherwise as recommended by ClonTech. Reactions were held at 94°C for 2 min and then cycled 35 times at 94°C for 20 sec, 62°C for 20 sec, and 72°C for 2 min. PCR products were ligated into pGEM-T (Promega), and plasmid DNA was then prepared for the sequencing of inserts. PCR amplification, cloning, and sequencing of the Le gene were performed in duplicate reac- tions. The sequence of the 2.2-kb Le genomic fragment was com- pared with the GenBank database using the BLAST program facility at the National Center for Biotechnology lnformation (Bethesda, MD).
DNA Gel Blot and RFLP Analyses
Genomic DNA was prepared from the Le and /e isolines and 40 plants of FI3 progeny (two families) of the Le x /e cross. DNA gel blot analysis was performed by standard methods (Southern, 1975), with hybridization conditions being the same as those used for cDNA li- brary screening, except that 50% instead of 20% formamide was used in the hybridization buffer. PCR was performed using the Klen- Taq enzyme with primers OD302 and OD306 at 0.4 pM in a volume of 20 pL containing 10 ng of genomic DNA and otherwise using re- action conditions as described previously. Reactions were held at 94°C for 2 min and then cycled 35 times at 94°C for 20 sec, 62°C for 20 sec, and 72°C for 2 min in a Perkin-Elmer thermal cycler. PCR products were diluted three times, digested with Hindlll, separated by electrophoresis in a 1 % agarose gel, and visualized by ethidium bromide staining.
Primer Sequences
The primer sequences are listed 5' to 3' below. The nucleotide posi- tions to which they correspond in the coding strand (cs) or noncoding strand (ncs) of the 4.25-kb /e genomic fragment (GenBank accession
number U9321 O) are given: OD302, 5 '-GTCGTAAAGAATGGT-
GTGCTTGTGGTTAACGTGC-3', 1468 to 1495 ncs; OD3nt, 5'-TGA- TGAAGATGGTTGATG-3', 271 to 300 CS; OD306,5'-GAAATCAGG-
CGAATTCACTATGCCTTCACTCTCCGMGCCTA-3', 1425 to 1452 CS; OD3ct, 5'-GACAGAATTCCTATTTAGCCCACTTGGACACTA-3', 3080 to 31 03 ncs; OD3up, 5 '-GTAAGCGCAACATTGTCGCCA- TCTTCTC-3', 1313 to 1340 cs; and OD3do, 5'-TAGATGCGCACG- TACGGTTGTTGCCAC-3', 3528 to 3554 ncs.
E. coli Expression
Sequences from genomic clones were used to design PCR primers OD3nt and OD3ct for amplification of a complete open reading frame from the cDNA template of the Le and /e lines. Total RNA was pre- pared from apical and expanding internode tissue of 3-week-old Le and /e plants (Ausubel et al., 1994), and mRNA was selected using an mRNA isolation kit (Boehringer Mannheim). First-strand cDNA was prepared from 200 ng of mRNA in a total volume of 25 pL, using the Superscript Preamplification System (Gibco BRL). PCR was per- formed as described previously, except that the primers used were OD3nt and OD3ct, the template was 0.5 pL of the cDNA reaction, and cycling was 35 times at 94°C for 20 sec, 58°C for 20 sec, and 72°C for 20 sec.
EcoRl sites included in these primers enabled ligation of the cDNAs into the EcoRl site of the expression vector pTrcHisB (Invitro- gen, La Jolla, CA). Constructs containing the Le and /e cDNAs in the correct orientation were selected. These were predicted to encode the 45-amino acid N-terminal fusion peptide of pTrcHisB in frame with a threonine residue followed by the 374-amino acid sequence encoded by the Le and /e GA4-related genes. A construct of pTrcHisB containing the /e cDNA in the opposite orientation was se- lected for use as a negative control.
Starter cultures of transformed E. coli TOPIO cells were grown overnight at 37"C, and 0.5 mL was used to inoculate 100 mL of 2 x YT broth (NaCI [ I g/L], yeast extract [ I g/L], and tryptone [I .6 g/L]) containing 1 O0 pg/mL ampicillin. Cultures were grown with shaking at 32°C until they reached an optical density of 0.3 at 600 nm. Isopro- pyl p-D-thiogalactopyranoside was added to 5 mM, and cultures were grown for an additional 75 min. Kept at 4"C, cells were har- vested by centrifugation, washed with 2 x YT and then 200 mM Tris- HCI, pH 7.8, and 20 mM DTT, and resuspended in 300 pL of 200 mM Tris-HCI and 20 mM DTT. Lysozyme (Sigma) was added to 2 mg/mL, and cells were incubated on ice for 1 O min and then lysed by two cy- cles of freezehhawing. DNase I (Progen Industries, Darra, Australia) was added to a concentration of 400 units per mL, and samples were left on ice for 10 min. Aliquots of 120 pL were removed for assay of 3p-hydroxylase activity. The E. coli expression procedure and the subsequent activity assays were performed on three separate occa- sions to ensure the reproducibility of findings.
GA 3P-Hydroxylase Enzyme Assay
Enzyme preparations were incubated at 30°C with GA substrates in 200 pL of 100 mM Tris, pH 7.8, containing 4 mM DTT, 4 mM ascor- bate, 4 mM 2-oxoglutarate, 0.5 mM FeSO,, and 1 mg/mL catalase. The ability of the Le and /e cDNA products to metabolize GA,, was determined by using two different substrates: 1 7-13C,3H,-labeled GAZ0 (55,000 dpm, 88% carbon-13, 30 mCi mmol-I), and 17,l 7-2Hz- GA,, (100 ng). The 13C,3H-labeled GAZ0 was prepurified by HPLC. 17,17-2H,-GA5 (100 ng) was also tested as a substrate. Reactions

1442 The Plant Cell
were stopped after 1 hr by adding 25 FL of glacial acetic acid, and samples were then purified using Sep-Pak (Waters Associates, Mil- ford, MA) C18 cartridges. Products from 13C,3H-labeled GAZ0 were analyzed by radiocounting after HPLC, as described in Ross et al. (1995), whereas those from 1 7,17-2H,-GA,, were monitored by gas chromatography mass spectrometry, as given in Lester et al. (1996). The HPLC program was 20 to 75% methanol (in 0.4% acetic acid) over 25 min (linear) at a flow rate of 2 mL/min. One-minute fractions were collected, and 50% aliquots were radiocounted (Hasan et al., 1994). Fractions containing radioactivity at the retention time of GAl were dried, methylated, and trimethylsilylated, as given in Hasan et al. (1 994). Gas chromatography m a s spectrometry was performed using a gas chromatograph (model 5890; Hewlett-Packard, Palo Alto, CA) coupled to a m a s spectrometer (Concept ISQ; Kratos, Manchester, UK) (Hasan et al., 1994). The column and gas chroma- tography program have been described previously (Ross et al., 1995). Full-scan mass spectra were acquired by scanning the range m/z 180 to m/z 550 at 0.8 sec per decade in the raw data mode and enhanced by averaging and background subtraction. Kovats reten- tion indices were determined using a mixture of n-alkanes (Sigma).
ACKNOWLEDGMENTS
We thank Drs. Howard Goodman and Hui-Hwa Chiang (Harvard Medical School and Department of Molecular Biology) for the GA4 clone, Dr. Noel Davies (Central Science Laboratory, University of Tasmania) for help with mass spectrometry, Alasdair MacKenzie- Hose and Christopher Williams for technical assistance, Dr. Christine Willis (University of Bristol) for 13C,3H-labeled GAZo, Lewis Mander (Australian National University) for deuterated GAs, and Drs. Peter Hedden and Andrew Phillips (Long Ashton Research Station) for ad- vice. This project was funded by the Australian Research Council.
Received March 27, 1997; accepted June 10,1997.
REFERENCES
Ait-Ali, T., Swain, S.M., Reid, J.B., Sun, T.-p., and Kamiya, Y. (1997). The Ls locus of pea encodes the gibberellin biosynthesis enzyme ent-kaurene synthase A. Plant J. li, 443-454.
Ausubel, F.M., Brent, R., Kingston, R.E., Moore, D.D., Seidman, J.G., Smith, J.A., and Struhl, K., eds (1994). PhenoVSDS method for plant RNA preparation. In Current Protocols in Molecular Biol- ogy, Vol. 1 (New York: Wiley Interscience), pp. 4.3.1-4.3.3.
Bateson, W. (1901). Experiments in plant hybridization by Gregor Mendel. J. R. Hortic. SOC. 24, 1-32.
Bhattacharyya, M.K., Smith, A.M., Ellis, T.H.N., Hedley, C., and Martin, C. (1990). The wrinkled-seed character of pea described by Mendel is caused by a transposon-like insertion in a gene encoding starch-branching enzyme. Cell 60, 11 5-1 22.
Britsch, L., Dedio, J., Saedler, H., and Forkmann, G. (1993). Molecular characterization of flavanone 3p-hydroxylases. Eur. J. Biochem. 217,745-754.
Chiang, H.-H., Hwang, I., and Goodman, H.M. (1995). lsolation of the Arabidopsis GA4 locus. Plant Cell 7, 195-201.
de Carolis, E., and de Luca, V. (1994). 2-Oxoglutarate-dependent dioxygenases and related enzymes: Biochemical characterization. Phytochemistry 36,1093-1 107.
Ellis, T.H.N. (1994). Approaches to the genetic mapping of pea. In Modern Methods of Plant Analysis, Vol. 16, Vegetables and Vege- table Products, H.-F. Linskens and J.F. Jackson, eds (Berlin: Springer-Verlag), pp. 11 7-160.
Hasan, O., Riddout, B.G., Ross, J.J., Davies, N.W., and Reid, J.B. (1994). ldentification and quantification of endogenous gibberel- lins in apical buds and cambial regions of Eucalyptus. Physiol. Plant. 90,475-480.
Hedden, P., and Kamiya, Y. (1 997). Gibberellin biosynthesis: Enzymes, genes and their regulation. Annu. Rev. Plant Physiol. Plant MOI. Biol. 48, 431-460.
Ingram, T.J., Reid, J.B., Murfet, I.C., Gaskin, P., Willis, C.L., and MacMillan, J. (1984). lnternode length in Pisum. The Le gene controls the 3p-hydroxylation of gibberellin A,, to gibberellin Al. Planta 160, 455-463.
Kobayashi, M., Sakurai, A,, Saka, H., and Takahashi, N. (1989). Quantitative analysis of endogenous gibberellins in normal and dwarf cultivars of rice. Plant Cell Physiol. 30,963-969.
Lange, T., and Graebe, J.E. (1993). Enzymes of gibberellin synthe- sis. In Methods in Plant Biochemistry, Vol. 9, P.J. Lea, ed (Lon- don: Academic Press), pp. 403-430.
Lester, D.R., Ross, J.J., Ait-Ali, T., Martin, D.N., and Reid, J.B. (1 996). A gibberellin 20-oxidase cDNA (accession no. U58830) from pea (Pisum safivum L.) seed (PGR96-050). Plant Physiol. 111,1353.
Martin, D.N., Proebsting, W.M., Parks, T.D., Dougherty, W.G., Lange, T., Lewis, M.J., Gaskin, P., and Hedden, P. (1996). Feed-back regulation of gibberellin biosynthesis and gene expression in Pisum safivum L.. Planta 200, 159-1 66.
Mendel, G. (1866). Versuche über Pflanzen-hybriden. Verh. Natforsch. Ver. Brünn 4,3-47. English translation from web site http://netspace. students.brown.edu/MendelWeb/Mendel.html.
Poole, A.T., Ross, J.J., Lawrence, N.L., and Reid, J.B. (1995). ldentification of gibberellin A4 in Pisum safivum L. and the effects of applied gibberellins A,, 4, A5 and A3 on the /e mutant. Plant Growth Regul. 16,257-262.
Potts, W.C., Reid, J.B., and Murfet, I.C. (1982). lnternode length in Pisum. I. The effect of the Le//e gene difference on endogenous gibberellin-like substances. Physiol. Plant. 55,323-328.
Prescott, A.G. (1993). A dilemma of dioxygenases (or where bio- chemistry and molecular biology fail to meet). J. Exp. Bot. 44, 849-861.
Prescott, A.G., and John, P. (1 996). Dioxygenases: Molecular structure and role in plant metabolism. Annu. Rev. Plant Physiol. Plant MOI. Biol. 47,245-271.
Roach, P.L., Clifton, I.J., Fülop, V., Harlos, K., Barton, G.J., Hajdu, J., Andersson, I., Schofield, C.J., and Baldwin, J.E. (1 995). Crystal structure of isopenicillin N synthase is the first from a new structural family of enzymes. Nature 375, 700-704.
Ross, J.J., and Reid, J.B. (1987). lnternode length in Pisum. A new allele at the /e locus. Ann. Bot. 59, 107-1 09.

Le Encodes a Gibberellin 3pHydroxylase 1443
Ross, J.J., Reid, J.B., Gaskin, P., and MacMillan, J. (1989). Inter- node length in Pisum. Estimation of GA, levels in genotypes Le, /e and /ed. Physiol. Plant. 76, 173-176.
Ross, J.J., Reid, J.B., and Dungey, H.S. (1992). Ontogenetic varia- tion in levels of gibberellin A, in Pisum. lmplications for the control of stem elongation. Planta 186, 166-1 71.
Ross, J.J., Reid, J.B., Swain, S.M., Hasan, O., Poole, A.T., Hedden, P., and Willis, C.L. (1995). Genetic regulation of gibberellin deac- tivation in Pisum. Plant J. 7, 513-523.
Sherriff, L.J., McKay, M., Ross, J.J., Reid, J.B., and Willis, C.L. (1 994). Decapitation reduces the metabolism of gibberellin A,, to A, in Pisum sativum L., decreasing the Lelle difference. Plant Physiol. 104, 277-280.
Smith, V.A. (1992). Gibberellin A, biosynthesis in Pisum sativum L. II. Biological and biochemical consequences of the /e mutation. Plant Physiol. 99, 372-377.
Smith, V.A., Knatt, C.J., Gaskin, P., and Reid, J.B. (1992). The dis- tribution of gibberellins in vegetative tissues of Pisum sativum L. I. Biological and biochemical consequences of the /e mutation. Plant Physiol. 99, 368-371.
Southern, E.M. (1975). Detection of specific sequences among DNA fragments separated by gel electrophoresis. J. MOI. Biol. 98,503-517.
Spray, C., Phinney, B.O., Gaskin, P., Gilmour, S.J., and MacMillan, J. (1984). lnternode length in Zea mays L. The dwarf-7 mutation controls the 3P-hydroxylation of gibberellin AZO to gibberellin A,. Planta 160,464-468.
Spray, C.R., Kobayashi, M., Suzuki, Y., Phinney, B.O., Gaskin, P., and MacMillan, J. (1996). The dwarf-7 (d7) mutant of Zea mays blocks three steps in the gibberellin-biosynthetic pathway. Proc. Natl. Acad. Sci. USA 93, 1051 5-1 051 8.
Talon, M., Koornneef, M., and Zeevaart, J.A.D. (1990). Endoge- nous gibberellins in Arabidopsis thaliana and possible steps blocked in the biosynthetic pathways of the semidwarf ga4 and ga5 mutants. Proc. Natl. Acad. Sci. USA 87, 7983-7987.
White, O.E. (1917). Studies of inheritance in Pisum 11. Proc. Am. Phi-
Xu, Y., Li, L., Wu, K., Peeters, A.J.M., Gage, D.A., and Zeevaark, J.A.D. (1995). The GA5 locus of Arabidopsis thaliana encodes a multifunctional gibberellin 30-oxidase: Molecular cloning and functional expression. Proc. Natl. Acad. Sci. USA 92, 6640-6644.
10s. SOC. 56,487-588.

DOI 10.1105/tpc.9.8.1435 1997;9;1435-1443Plant Cell
D R Lester, J J Ross, P J Davies and J B ReidMendel's stem length gene (Le) encodes a gibberellin 3 beta-hydroxylase.
This information is current as of April 9, 2019
Permissions 98X
https://www.copyright.com/ccc/openurl.do?sid=pd_hw1532298X&issn=1532298X&WT.mc_id=pd_hw15322
eTOCs http://www.plantcell.org/cgi/alerts/ctmain
Sign up for eTOCs at:
CiteTrack Alerts http://www.plantcell.org/cgi/alerts/ctmain
Sign up for CiteTrack Alerts at:
Subscription Information http://www.aspb.org/publications/subscriptions.cfm
is available at:Plant Physiology and The Plant CellSubscription Information for
ADVANCING THE SCIENCE OF PLANT BIOLOGY © American Society of Plant Biologists