mmm1p spans both the outer and inner mitochondrial membranes
TRANSCRIPT

Kondo-Okamoto et al.
- 1 -
Mmm1p Spans Both the Outer and Inner Mitochondrial Membranes, and
Contains Distinct Domains for Targeting and Foci Formation
Noriko Kondo-Okamoto, Janet M. Shaw*, and Koji Okamoto*
Department of Biology, University of Utah, Salt Lake City, UT 84112
*To whom correspondence should be addressed:
Department of Biology, University of Utah
257 South 1400 East, Salt Lake City, UT 84112
Tel.: (801) 585-6205 (to J.M.S.) / (801) 587-9209 (to K.O.)
Fax: (801) 581-2174
E-mail: [email protected] (to J.M.S.) / [email protected] (to K.O.)
Running Title: Mmm1p Is a Double Membrane-spanning Protein
Copyright 2003 by The American Society for Biochemistry and Molecular Biology, Inc.
JBC Papers in Press. Published on September 12, 2003 as Manuscript M308436200 by guest on M
arch 21, 2018http://w
ww
.jbc.org/D
ownloaded from

Kondo-Okamoto et al.
- 2 -
SUMMARY
In the yeast Saccharomyces cerevisiae, the integral membrane protein Mmm1p is required for
maintenance of mitochondrial morphology and retention of mitochondrial DNA (mtDNA).
Mmm1p localizes to discrete foci on mitochondria that are adjacent to mtDNA nucleoids in the
matrix, raising the possibility that this protein plays a direct role in organizing, replicating or
segregating mtDNA. Although Mmm1p has been shown to cross the outer membrane with its C-
terminus facing the cytoplasm, the location of the N-terminus has not been resolved. Here we
show that Mmm1p spans both the outer and inner mitochondrial membranes, exposing its N-
terminus to the matrix. Surprisingly, deletion of the N-terminal extension decreased steady-state
levels of the Mmm1 protein, but did not affect mitochondrial morphology or mtDNA
maintenance. Moreover, expression of Neurospora crassa MMM1, which naturally lacks a long
N-terminal extension, substituted for loss of Mmm1p in budding yeast. These results indicate
that the matrix-exposed portion of Mmm1p is not essential for mtDNA nucleoid maintenance.
Additional studies revealed that the transmembrane segment and C-terminal domain of Mmm1p
are required for foci formation and mitochondrial targeting, respectively. Our data suggest that
Mmm1p’s double membrane-spanning topology at the membrane contact site is critical for
formation of tubular mitochondria.
by guest on March 21, 2018
http://ww
w.jbc.org/
Dow
nloaded from

Kondo-Okamoto et al.
- 3 -
INTRODUCTION
Mitochondria are dynamic organelles that change size, shape and distribution in response to
changing cellular needs (1-4). Because mitochondria are essential and cannot be synthesized de
novo, most cell types employ cytoskeletal elements and molecular motors to partition the
organelles into newly formed daughter cells during division (5). Studies performed in a variety of
organisms including budding yeast reveal that maintenance of proper organelle morphology is
critical for mitochondrial function. In Saccharomyces cerevisiae, mitochondria form highly
branched tubular networks that continually divide and fuse (6-10). Mutations in a variety of
nuclear genes disrupt wildtype mitochondrial morphology, converting the tubular network into
small fragments, aggregates, large spheres, collapsed tubules, nets, or other aberrant shapes (11).
In some cases, these morphological changes lead to loss of mitochondrial DNA (mtDNA) and
defective respiratory activity (12-14). For example, mitochondria fragment and lose mtDNA in
cells lacking the large GTPase Fzo1p (15,16) or Ugo1p, (17) two integral outer membrane (OM)
proteins that mediate mitochondrial fusion. Similar mtDNA loss is observed in cells lacking the
intermembrane space (IMS) GTPase Mgm1p (18-21) required for mitochondrial fusion and inner
membrane (IM) remodeling, and the IM Rhomboid-type protease Mdm37p/Pcp1p/Rbd1p (11,22-
24) required for the biogenesis of Mgm1p. In the examples cited above, mtDNA loss is thought
to be a secondary consequence of changes in mitochondrial shape, since there is currently no
evidence that these proteins interact directly or indirectly with mtDNA nucleoids (mtDNA-
protein complexes). A second class of mutations that converts tubular mitochondria into large
spheres also leads to mtDNA loss. These mutations affect three integral OM proteins, called
Mdm10p (25), Mdm12p (26), and Mmm1p (27), which are thought to form a complex. Roles for
this complex in cytoskeletal attachment and mtDNA nucleoid maintenance have been proposed.
by guest on March 21, 2018
http://ww
w.jbc.org/
Dow
nloaded from

Kondo-Okamoto et al.
- 4 -
The function of the Mmm1 protein has been studied in some detail. S. cerevisiae Mmm1p is
predicted to have a single transmembrane (TM) segment of ~25 residues (amino acids 92-116)
(27). Cells lacking functional Mmm1p display defects in mitochondrial motility and inheritance,
and mitochondria isolated from these cells are defective in ATP-sensitive actin-binding activity
in vitro (28,29). Based on these observations, it was proposed that Mmm1p is required for
interactions of mitochondria with the actin cytoskeleton. When tagged at the C-terminus with the
green fluorescent protein (GFP), Mmm1p-GFP localizes as discrete foci on the surface of
mitochondria (30). Strikingly, these Mmm1p-GFP foci appear adjacent to mtDNA nucleoids in
the matrix (30). Additional biochemical data suggest that Mmm1p resides at contact sites
between the inner and outer mitochondrial membranes (30). These findings raise the possibility
that Mmm1p is connected either directly or indirectly to mtDNA nucleoids.
Studies of the filamentous fungus Neurospora crassa MMM1 (NcMMM1) suggest that the
cellular role of this protein varies in different cell types. In N. crassa, mitochondria form tubular
arrays that co-align with microtubules rather than actin filaments (31). Unlike S. cerevisiae, N.
crassa cannot survive in the absence of mitochondrial respiration, and loss of mtDNA results in
lethality. Although NcMMM1 is also required for maintenance of normal mitochondrial
morphology, it is dispensable for interactions with microtubules (32,33). Moreover, the Mmm1
protein is not essential for mtDNA stability in N. crassa, since loss-of-function mutations are not
lethal (32). Thus, while the role of Mmm1p in mitochondrial morphology maintenance has been
conserved during evolution, its roles in cytoskeletal association and/or mtDNA retention may
have been altered or lost.
The phenotypes observed in mmm1 mutants suggest that Mmm1p contains distinct domains
with different functions. A better understanding of Mmm1p structure, topology and domain
by guest on March 21, 2018
http://ww
w.jbc.org/
Dow
nloaded from

Kondo-Okamoto et al.
- 5 -
function may reveal how this protein carries out activities within the cell. S. cerevisiae Mmm1p
contains one predicted TM segment that spans the outer mitochondrial membrane leaving the C-
terminal domain in the cytoplasm (27). Although the N-terminus faces the interior of the
organelle, its exact location (IMS or matrix) has not been determined. In addition, the specific
functions of the Mmm1p N-terminal extension, TM segment and C-terminal domain have not
been explored. In this study, we determined the submitochondrial location of the Mmm1p N-
terminus, and identified domains important for mitochondrial targeting and foci formation. In
vivo site-specific cleavage and in vitro protease protection assays revealed that the N-terminus of
Mmm1p is exposed to the matrix, a topology that requires the protein to span both the outer and
inner mitochondrial membranes. Although deletion of the N-terminal extension resulted in
reduction of steady-state Mmm1p protein levels, it did not affect protein targeting to the
organelle, maintenance of mitochondrial morphology, or mtDNA retention. Moreover, the
mtDNA loss phenotype of the S. cerevisiae mmm1 null (mmm1 ) mutant could be rescued by
expressing NcMMM1, which naturally lacks a long N-terminal extension. These findings
indicate that the matrix-exposed, N-terminal extension of Mmm1p does not play a direct role in
mtDNA maintenance. We further show that the TM segment and C-terminal domain play
essential roles in Mmm1p foci formation and mitochondrial targeting, respectively. These
combined results suggest that Mmm1p’s double membrane-spanning topology at the membrane
contact site is crucial for establishment and maintenance of tubular mitochondrial morphology.
Implications of these findings with respect to mtDNA maintenance and cytoskeletal attachment
are discussed. Models for the double-membrane spanning topology of Mmm1p are also
presented.
by guest on March 21, 2018
http://ww
w.jbc.org/
Dow
nloaded from

Kondo-Okamoto et al.
- 6 -
EXPERIMENTAL PROCEDURES
Yeast Strains and Growth Conditions—All strains are derivatives of the FY10 strain (34).
Standard genetic methods were used to grow, transform and manipulate yeast (35) and bacterial
(36) strains. Strains used in this study include: JSY1825 (MAT , his3 200, leu2 1, lys2 -202,
trp1 63, ura3-52) and NOY84 (MAT , his3 200, leu2 1, lys2 -202, trp1 63, ura3-52,
mmm1 ::HIS3, [pRS416-MMM1-His]). Plasmids containing MMM1 constructs were introduced
by plasmid shuffle into NOY84. Yeast transformants were grown at 30°C in SC media or on
dropout plates containing one of the following carbon sources: 2% dextrose (SD), 2% glycerol
(SGly), 2% raffinose (SR), or 2% galactose plus 2% raffinose (SGalR).
Plasmid Construction—For pRS416-MMM1-His and pRS315-MMM1-His, a BamHI-EcoRV
fragment encoding the 0.52 kb MMM1 5’-UTR and Mmm1p (amino acids 1-426) plus 8xHis,
and an EcoRV-XhoI fragment encoding the 0.45 kb MMM1 3’-UTR were generated by PCR and
cloned into the BamHI-XhoI sites of the low-copy yeast vectors pRS416 and pRS315,
respectively (American Type Culture Collection). For pRS315-mmm1 (2-75)-His, pRS315-
mmm1 (2-90)-His, and pRS315-mmm1 (2-120)-His, the 1.1 kb BamHI-SalI fragment of
pRS315-MMM1-His was replaced by a PCR-generated BamHI-SpeI fragment encoding the 0.52
kb MMM1 5’-UTR plus ATG and a PCR-generated SpeI-SalI fragment encoding Mmm1p
(amino acids 76-190), (amino acids 91-190), and (amino acids 121-190), respectively. For
pRS315-MMM1-GFP, a BamHI-XmaI fragment encoding the 0.52 kb MMM1 5’-UTR and
Mmm1p (amino acids 1-426), XmaI-EcoRV fragment encoding HA plus GFP, and an EcoRV-
XhoI fragment encoding the 0.45 kb MMM1 3’-UTR were generated by PCR and cloned into the
BamHI-XhoI sites of pRS315. For pRS315-mmm1 (2-75)-GFP and pRS315-mmm1 (2-90)-
by guest on March 21, 2018
http://ww
w.jbc.org/
Dow
nloaded from

Kondo-Okamoto et al.
- 7 -
GFP, the 1.1 kb BamHI-SalI fragment of pRS315-MMM1-GFP was replaced by a 0.87 kb
BamHI-SalI fragment from pRS315-mmm1 (2-75)-His and a 0.82 kb BamHI-SalI fragment
from pRS315-mmm1 (2-90)-His, respectively. For pRS315-T20(TM)-mmm1(C)-His, a BamHI-
SpeI fragment encoding the 0.52 kb MMM1 5’-UTR plus ATG, a PCR-generated NheI-SpeI
fragment encoding Tom20p (amino acids 2-50), a SpeI-XhoI fragment encoding Mmm1p (amino
acids 121-426) plus 8xHis, and the 0.45 kb MMM1 3’-UTR were cloned into the BamHI-XhoI
sites of the pRS315 variant that lacks an SpeI site in the MCS. For pRS315-T20(TM)-mmm1(C)-
GFP, the 1.1 kb BamHI-SalI fragment of pRS315-MMM1-GFP was replaced by a 0.88 kb
BamHI-SalI fragment from pRS315-T20(TM)-mmm1(C)-His. For pRS315-T20(TM)-GFP, the
1.4 kb SpeI-XhoI fragment of pRS315-T20(TM)-mmm1(C)-His was replaced by a PCR-
generated SpeI-NcoI fragment encoding GFP (amino acids 2-56) and a 1.0 kb NcoI-XhoI
fragment from pRS315-MMM1-GFP. For pRS315-NcM(TM)-mmm1(C)-His, the 0.67 kb
BamHI-SpeI fragment of pRS315-T20(TM)-mmm1(C)-His was replaced by a PCR-generated
BamHI-NheI fragment encoding the 0.52 kb MMM1 5’-UTR plus ATG and N. crassa MMM1
(amino acids 2-39). For pRS315-NcM(TM)-mmm1(C)-GFP, the 1.1 kb BamHI-SalI fragment of
pRS315-MMM1-GFP was replaced by an 0.85 kb BamHI-SalI fragment from pRS315-
NcM(TM)-mmm1(C)-His. For pRS425-MMM1-GFP, a 3.0 kb BamHI-XhoI fragment from
pRS315-MMM1-GFP was cloned into the BamHI-XhoI sites of pRS425 (American Type Culture
Collection). For pRS425-mmm1(1-135)-GFP, pRS425-mmm1(1-180)-GFP, pRS425-mmm1(1-
275)-GFP, and pRS425-mmm1(1-365)-GFP, the 1.8 kb BamHI-XmaI fragment of pRS425-
MMM1-GFP was replaced by PCR-generated BamHI-XmaI fragments encoding the 0.52 kb
MMM1 5’-UTR plus Mmm1p (amino acids 1-135), (amino acids 1-180), (amino acids 1-275),
and (amino acids 1-365), respectively. For pRS425-mmm1(1-390)-GFP and pRS425-mmm1(1-
by guest on March 21, 2018
http://ww
w.jbc.org/
Dow
nloaded from

Kondo-Okamoto et al.
- 8 -
415)-GFP, the 0.71 kb SalI-XmaI fragment of pRS425-MMM1-GFP was replaced by a PCR-
generated SalI-XmaI fragments encoding Mmm1p (amino acids 191-390) and (amino acids 191-
415), respectively. For pRS425-mmm1 (2-120)-GFP, the 1.1 kb BamHI-SalI fragment of
pRS425-MMM1-GFP was replaced by a 0.74 kb BamHI-SalI fragment from pRS315-mmm1 (2-
120)-His. For pRS315-TCS-MMM1-His and pRS315-TCS-MMM1-GFP, the 0.58 kb SpeI-SalI
fragment of pRS315-MMM1-His and pRS315-MMM1-GFP, respectively, was replaced by a
PCR-generated SpeI-NcoI fragment encoding 3xMyc, an NcoI-BglII fragment encoding TCS
(TEV cleavage site: ENLYFQG) plus 3xHA, and a BglII-SalI fragment encoding Mmm1p
(amino acids 2-190). For pRS315-TCSmut-MMM1-His, the 0.12 kb NcoI-BglII fragment of
pRS315-TCS-MMM1-His was replaced by a PCR-generated NcoI-BglII fragment encoding the
mutated TCS (ANLDFRP) plus 3xHA. For pYX142-TCS-NcMMM1, an EcoRI-SpeI fragment
encoding 3xMyc-TCS-3xHA and a SpeI-EcoRV fragment encoding N. crassa MMM1 (amino
acids 2-415) were generated by PCR and cloned into the EcoRI-EcoRV sites of pYX142, a low-
copy yeast expression vector with the strong constitutive TPI1 promoter (Novagen). For
pYX142-TCS-MMM1-His, the 1.4 kb NcoI-EcoRV fragment of pYX142-TCS-NcMMM1 was
replaced by a 0.69 kb NcoI-SalI fragment from pRS315-TCS-MMM1-His, and a 0.73 kb SalI-
EcoRV fragment from pRS416-MMM1-His. For pYX142-Su9-TCS-GFP, a SpeI-BglII fragment
encoding 3xHA plus TCS was generated by PCR, and the SpeI site was converted to blunt ends
using Mung Bean Nuclease. The fragment was cloned into the BamHI-BglII sites of pYX142-
mtGFP (the BamHI site was converted to blunt ends using DNA polymerase I, Large Fragment)
(37). For pYX142-b2-TCS-GFP, the 0.21 kb EcoRI-BglII fragment of pYX142-mtGFP was
replaced by a PCR-generated EcoRI-NheI fragment encoding Cyb2p (amino acids 1-220) and a
SpeI-BglII fragment encoding 3xHA plus TCS. For pRS416GAL1-b2-TEV, a PCR-generated
by guest on March 21, 2018
http://ww
w.jbc.org/
Dow
nloaded from

Kondo-Okamoto et al.
- 9 -
NheI-BglII fragment encoding Cyb2p (amino acids 1-220) and a BamHI-XhoI fragment from
pWP1039 (provided by Will Prinz) encoding the TEV (tobacco etch virus) protease were cloned
into the XbaI-XhoI sites of pRS416GAL1 (American Type Culture Collection). All clones used
in this study were verified by DNA sequencing.
Site-specific TEV Cleavage Assay—Yeast cells harboring two plasmids encoding a TEV
protease and a substrate protein were pregrown to an OD600 of 1.5-3 in SD media. To induce TEV
protease expression, cells were washed three times with 1 ml of SGalR media, resuspended to an
OD600 of 0.4-1, and grown to an OD600 of 1-2 in SGalR media. 2 OD600 units of cells were
collected 3 h (for Su9-TCS-GFP), 6 h (for b2-TCS-GFP), or 16 h (for TCS-Mmm1p-His, TCSmut-
Mmm1p-His, and TCS-Mmm1p-GFP) after induction, resuspended in 1 ml of H2O, and lysed by
adding 160 µl of 1.85 M NaOH, 7.4% 2-mercaptoethanol on ice for 10 min. The protein extracts
were precipitated with 6% trichloroacetic acid on ice for 10 min. The precipitates were rinsed
with 1ml of 1M Tris base, resuspended in 60 µl of SDS gel-loading buffer, and boiled for 3 min.
15 µl of each sample was analyzed by SDS-PAGE and Western blotting.
Protease Protection Assay—Yeast cells expressing TCS-Mmm1p-His or TCS-Mmm1-GFP
were grown in SR media, and the mitochondria were isolated as previously described (38),
except that oxalyticase (Enzogenetics) was used for spheroplast formation (6 x 104 U in 30 ml
suspension containing 2 x 103 OD600 units of cells). Mitochondria (200 µg of protein) were
resuspended in 1 ml of HS buffer (20 mM HEPES-KOH, pH 7.4, 0.6 M sorbitol), H buffer (20
mM HEPES-KOH, pH 7.4), or HT buffer (20 mM HEPES-KOH, pH 7.4, 1% Triton X-100). If
indicated, the samples were treated with proteinase K at a final concentration of 200 µg/ml, and
by guest on March 21, 2018
http://ww
w.jbc.org/
Dow
nloaded from

Kondo-Okamoto et al.
- 10 -
kept on ice for 30 min. The reaction was stopped by the addition of 2 mM phenylmethylsulfonyl
fluoride (PMSF) on ice for 5 min. For the samples in HS and H buffers, centrifugation was
performed at 16,000g for 10 min at 4°C. The pellet was washed with 1 ml of HS and H buffers
containing 1 mM PMSF, collected by centrifugation at 16,000g for 5 min at 4°C, and
resuspended in SDS gel-loading buffer. The sample in HT buffer was directly added to SDS gel-
loading buffer. The samples corresponding to 30 µg of mitochondria were boiled for 3 min, and
analyzed by SDS-PAGE and Western blotting.
Immunoblot Analysis—Western blots were decorated with mouse monoclonal antibodies
generated against HA (1:1,000; University of Utah Core Facility), GFP (1:1000; Covance), or 3-
PGK (1:2,000; Molecular Probes), and rabbit antisera raised against the N-terminal domain of
Fzo1p (1:2,000; Shaw laboratory), Cyb2p (1:2,000; Carla Koehler), or Ssc1p (1:1,000; Elizabeth
Craig) in TBS buffer containing 5% non-fat dry milk for 1 h, and washed three times with TBS
buffer for 10 min. The blots were decorated with the goat anti-mouse or -rabbit HRP conjugate
(1:8,000; Sigma) in TBS buffer containing 5% non-fat dry milk for 1 h, and washed three times
with TBS buffer for 10 min. Proteins were detected using ECL Plus and Hyperfilm ECL
(Amersham). All steps were performed at room temperature.
Microscopy and Imaging—Cells were visualized by a Zeiss Axioplan 2 Imaging microscope
(Carl Zeiss) equipped with differential interference contrast (DIC) optics, epifluorescence
capabilities, and a Zeiss Plan-Apochromat 100x (N.A. 1.4) objective. Filter sets (excitation /
beamsplitter / emission) for GFP and RFP fluorescence were BP 470 ± 20 / FT 495 / LP 525 ±
25, and BP 546 ± 6 / FT 560 / BP 575-640, respectively. Images were captured using a Zeiss
by guest on March 21, 2018
http://ww
w.jbc.org/
Dow
nloaded from

Kondo-Okamoto et al.
- 11 -
AxioCam Mm monochrome digital camera and Zeiss AxioVision 3.1 software, and assembled
using Adobe Photoshop (Adobe Systems). Plasmids encoding organelle marker proteins, and a
staining probe used in this study were pVT100U-mtGFP (37), pRS424ADH-mtRFP (Shaw
laboratory), and MitoFluor Red 589 (Molecular Probes) for mitochondria, and pJK59 (Sec63p-
GFP) (39) for the endoplasmic reticulum.
RESULTS
The N-terminus of Mmm1p Is Exposed to the Mitochondrial Matrix—To investigate whether
the N-terminus of Mmm1p resides in the mitochondrial matrix or the IMS, we performed an in
vivo site-specific cleavage assay using Tobacco Etch Virus (TEV) protease (40) targeted either to
the matrix or the IMS. This protease recognizes the seven amino acid consensus sequence,
EXXYXQ(S/G). Cleavage occurs between the conserved glutamine and serine or glycine
residues (41). In vivo processing by TEV protease in budding yeast has been applied successfully
to examine localization and topology of peroxisomal and mitochondrial proteins tagged with
TEV cleavage sites (TCS) (42,43).
We constructed plasmids encoding TEV proteases fused to the C-termini of different
mitochondrial presequences including, Su9 (amino acids 1-69) from ATPase subunit 9 of N.
crassa, and b2 (amino acids 1-220) from cytochrome b2 of S. cerevisiae (Su9-TEV and b2-TEV,
respectively). Su9 and b2 are well-characterized presequences that specifically target various
passenger polypeptides, including TEV protease, to the matrix and the IMS, respectively
(44,37,43) (J. W. Thatcher and J. M. Shaw, unpublished data). To determine the location of the
N-terminus, Mmm1p-His was fused to the C-terminus of a wildtype TCS (TCS-Mmm1p-His) or
a mutated TCS (TCSmut-Mmm1p-His) sequence (Fig. 1A). In addition, GFP was fused to the C-
by guest on March 21, 2018
http://ww
w.jbc.org/
Dow
nloaded from

Kondo-Okamoto et al.
- 12 -
terminus of TCS-Mmm1p (TCS-Mmm1p-GFP) (Fig. 1A). Each construct also contained a 1x-3x
HA epitope tag, allowing recognition by immunoblot analysis. The TCS-Mmm1p-His, TCSmut-
Mmm1p-His, and TCS-Mmm1p-GFP constructs were fully functional and restored normal
mitochondrial networks and glycerol growth when expressed in mmm1 cells (data not shown).
In addition, TCS-Mmm1p-GFP formed mitochondrial foci (data not shown).
Processing of the TCS-tagged Mmm1p variants by TEV protease is predicted to generate
proteins reduced in size by ~5 kDa that can be detected with anti-HA antibodies. When TCS-
Mmm1p-His was coexpressed with Su9-TEV, low levels of the ~5 kDa shorter form were
detected (Fig. 1B, lane 2, arrow; for each substrate, Su9-TEV and b2-TEV were expressed under
identical conditions and for identical times). In contrast, very little of the ~5 kDa shorter form of
TCS-Mmm1p-His was produced by the coexpression of b2-TEV (Fig. 1B, lane 4). In time-course
experiments, the amount of cleavage product produced by expression of Su9-TEV protease was
consistently larger than that produced by b2-TEV expression (data not shown). The cleavage of
TCS-Mmm1p-His by Su9-TEV occurred within the TCS, since Su9-TEV did not generate the ~5
kDa shorter form of the TCSmut-Mmm1p-His substrate (Fig. 1B, lane 2). Moreover, specific Su9-
TEV dependent processing was observed in cells expressing TCS-Mmm1p-GFP (Fig. 1B, lane 2,
open arrowhead). Although the processing of TCS-Mmm1p-His and TCS-Mmm1p-GFP was not
complete, additional control experiments indicated that it was specific to Su9-TEV. In particular,
the substrates Su9-TCS-GFP and b2-TCS-GFP were processed only by Su9-TEV and b2-TEV,
respectively (Fig1B, lane2, closed arrowhead and 4, closed circle). The latter results demonstrate
that TEV proteases are active in the matrix and IMS, and that the processing occurs only when
both a TEV protease and a substrate are localized in the same mitochondrial subcompartment.
Together, these results suggest that the N-terminus of Mmm1p is exposed in the matrix.
by guest on March 21, 2018
http://ww
w.jbc.org/
Dow
nloaded from

Kondo-Okamoto et al.
- 13 -
To further examine the topology of Mmm1p, we carried out a conventional protease
protection assay in vitro. Mitochondria were isolated from mmm1 cells expressing TCS-
Mmm1p-His or TCS-Mmm1p-GFP, and incubated in the presence or absence of proteinase K
(PK) with no treatment (to keep the organelle intact), osmotic shock treatment (to expose the
IMS), or Triton X-100 treatment (to solubilize the organelle). As shown in Figure1C, lanes 1 and
5, anti-HA antibodies decorated the full-length ~80 kDa TCS-Mmm1p-His and ~107 kDa TCS-
Mmm1p-GFP polypeptides in mitochondrial samples that were not treated with protease. In
contrast, when intact mitochondria were exposed to PK (200 µg/ml), a ~40 kDa proteolytic
fragment was detected (Fig. 1C, lanes 2 and 6). The apparent molecular weight of the protected
polypeptide was larger than predicted for this substrate (a cleavage product consisting of 3xMyc-
TCS-3xHA plus the first 120 amino acid residues of Mmm1p should migrate at ~22 kDa). This
size increase was not due to incomplete proteolysis, as treatment with 1 mg/ml PK also generated
a ~40 kDa form (data not shown). During the course of this study, we noticed that all variants of
Mmm1p migrated slower (appeared larger) on SDS gels than predicted. The reason for this
altered migration is not known. Nevertheless, when osmotically shocked mitochondria
containing TCS-Mmm1p-His or TCS-Mmm1p-GFP were treated with PK, the ~40 kDa clipped
form was not degraded further, even though the OM was clearly ruptured, allowing the IMS
protein Cyb2 to be digested (Fig. 1C, lane 3 and 7). In contrast, the protected ~40 kDa form was
completely degraded and could not be detected in PK-treated samples containing Triton X-100
(Fig. 1C, lane 4 and 8). Under this condition, both mitochondrial membranes were lysed as
assessed by degradation of the matrix protein Ssc1. Since the HA tag recognized in these
experiments resides near the N-termini of the substrates, these results indicate that the N-termini
of TCS-Mmm1p-His and TCS-Mmm1p-GFP can only be digested when PK has access to
by guest on March 21, 2018
http://ww
w.jbc.org/
Dow
nloaded from

Kondo-Okamoto et al.
- 14 -
proteins in the matrix. Similar results were obtained when experiments were performed using a
different protease, trypsin (data not shown). Based on the combined results, we conclude that
Mmm1p has the topology of Nmatrix-Ccytoplasm, spanning the outer and inner mitochondrial
membranes.
The N-terminal Extension of Mmm1p Is Not Required for Maintenance of Mitochondrial
Morphology and MtDNA—To test whether the N-terminal extension of Mmm1p is critical for
mitochondrial morphology and/or mtDNA maintenance, we constructed plasmids encoding
Mmm1 mutant proteins that lack amino acid residues 2-90 and 2-120 (Mmm1∆(2-90)p-His and
Mmm1∆(2-120)p-His, respectively) (Fig. 2A). Mmm1∆(2-90)p-His lacks nearly the entire N-terminal
extension in front of the predicted TM segment (amino acids 92-116) (27). Both the N-terminal
extension and the TM segment are deleted in Mmm1∆(2-120)p-His. When expressed in mmm1
cells, Mmm1∆(2-90)p-His restored normal mitochondrial tubular networks that were
indistinguishable from those in mmm1 cells expressing full-length Mmm1p-His (Fig. 2B and
Table I). In contrast, mmm1 cells expressing Mmm1∆(2-120)p-His exhibited the large spherical
mitochondria typically seen in mmm1 cells containing an empty plasmid (Fig. 2B and Table I).
Although Mmm1p-His and Mmm1∆(2-90)p-His restored glycerol growth when shuffled into
mmm1 cells, mmm1 cells expressing Mmm1∆(2-120)p-His or containing an empty plasmid did
not grow on glycerol medium (Fig. 2C). No glycerol growth defect was observed in mmm1
cells expressing Mmm1∆(2-90)p-His at 37°C (data not shown). Consistent with this growth pattern,
the morphology and number of DAPI-stained mtDNA nucleoids in mmm1 cells expressing
Mmm1p-His or Mmm1∆(2-90)p-His were similar, while mmm1 cells expressing Mmm1∆(2-120)p-
His or containing an empty plasmid lacked such structures (data not shown). The failure of
by guest on March 21, 2018
http://ww
w.jbc.org/
Dow
nloaded from

Kondo-Okamoto et al.
- 15 -
Mmm1∆(2-120)p-His to complement mutant phenotypes in mmm1 cells is likely due to the
absence of the TM segment, which is required for mitochondrial membrane insertion (see Fig. 6).
Our data suggest that the N-terminal extension of Mmm1p is dispensable for the maintenance of
normal mitochondrial tubular networks and mtDNA.
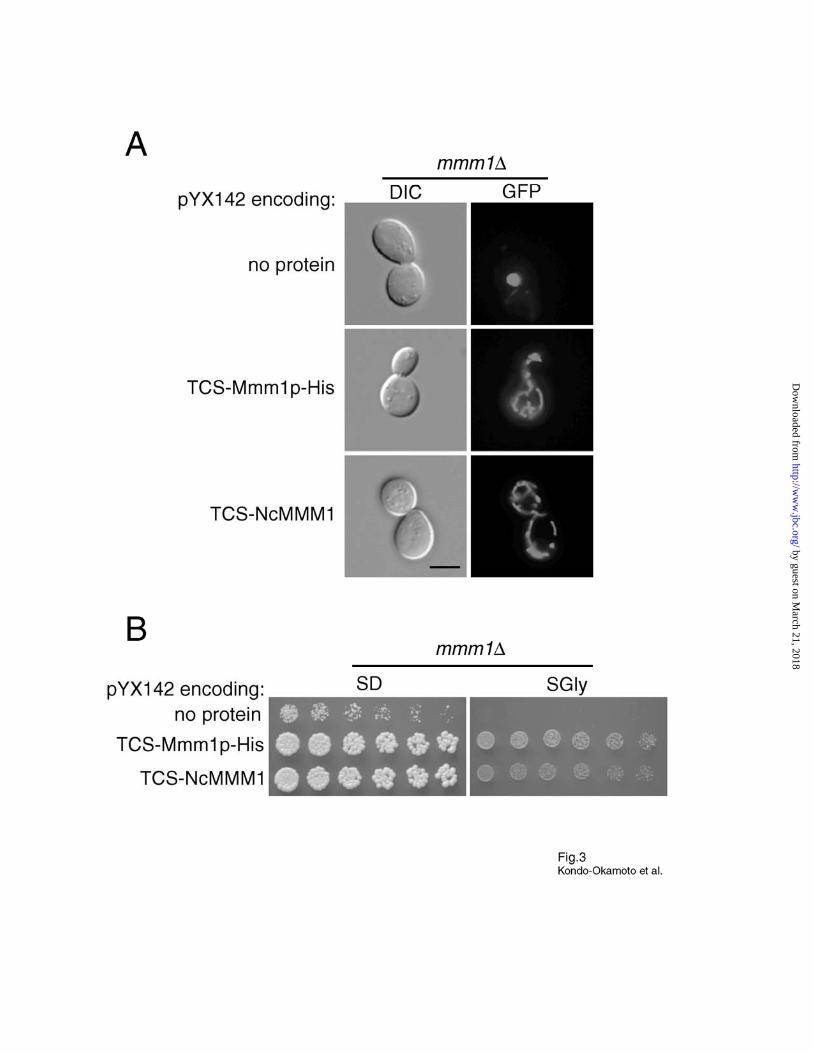
Yeast mmm1 Cells Expressing NcMMM1 Can Maintain Normal Mitochondrial Tubular
Networks and MtDNA—Although S. cerevisiae Mmm1p is fairly similar to the N. crassa
homologue NcMMM1 (30% identity and 54% similarity), there is a significant difference
between the two proteins at the N-termini. In particular, Mmm1p has ~90 amino acid residues N-
terminal to the predicted TM segment, while NcMMM1 has ~10 amino acid residues (32). The
fact that Mmm1∆(2-90)p-His is as functional as Mmm1p-His prompted us to test whether
NcMMM1 can substitute for the yeast homologue. We constructed a yeast expression vector
encoding NcMMM1 fused to the C-terminus of a TCS (TCS-NcMMM1). The same vector was
used for the expression of TCS-Mmm1p-His as a control. When expressed in mmm1 cells,
TCS-NcMMM1 restored normal mitochondrial tubular networks that were virtually identical to
those in mmm1 cells expressing TCS-Mmm1p-His (Fig. 3A and Table I). In addition, the
glycerol growth defect of mmm1 cells was suppressed by the expression of TCS-NcMMM1 or
TCS-Mmm1p-His at 30°C (Fig. 3B) and 37°C (data not shown). DAPI-staining confirmed that
normal mtDNA nucleoids were present in mmm1 cells expressing TCS-NcMMM1 or TCS-
Mmm1p-His (data not shown). In vivo TEV cleavage and in vitro protease protection assays
suggest that the N-terminus of NcMMM1 is also exposed to the matrix of yeast mitochondria (N.
Kondo-Okamoto, unpublished data). Together, these results demonstrate that, despite the lack of
an N-terminal extension, NcMMM1 can perform the function of Mmm1p in yeast.
by guest on March 21, 2018
http://ww
w.jbc.org/
Dow
nloaded from

Kondo-Okamoto et al.
- 16 -
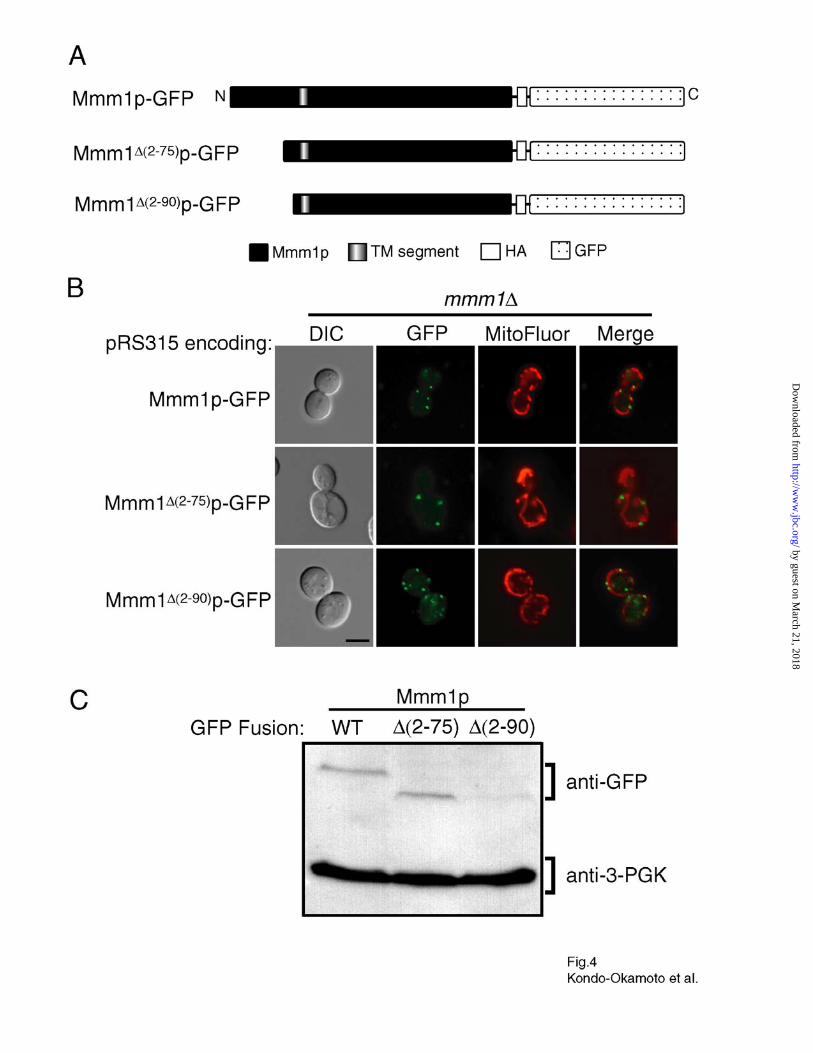
Deletion of the Mmm1p N-terminal Extension Does Not Affect Foci Formation but Reduces
the Steady-state Level of Mmm1p—To examine whether the N-terminal extension of Mmm1p is
required for foci formation, we constructed plasmids encoding Mmm1p-GFP mutant proteins
lacking the amino acid residues 2-75 and 2-90 (Mmm1∆(2-75)p-GFP and Mmm1∆(2-90)p-GFP,
respectively) (Fig. 4A). When expressed in mmm1 cells, both Mmm1∆(2-75)p-GFP and Mmm1∆(2-
90)p-GFP formed discrete foci on mitochondrial tubules identical to those observed in mmm1
cells expressing Mmm1p-GFP (Fig. 4B) (30). The number of cells exhibiting these Mmm1p foci,
the number of foci per cell, and the fluorescence intensity of the foci were similar for all three
constructs, Mmm1∆(2-75)p-GFP, Mmm1∆(2-90)p-GFP, and Mmm1p-GFP (data not shown).
Moreover, the morphology of restored mitochondrial networks was identical in mmm1 cells
expressing Mmm1∆(2-75)p-GFP and Mmm1∆(2-90)p-GFP, and Mmm1p-GFP (Fig. 4B) (30). These
observations suggest that the N-terminal extension of Mmm1p is dispensable for foci formation.
Surprisingly, we found that the steady-state level of Mmm1∆(2-90)p-GFP was four- to six-fold
lower than that of Mmm1p-GFP and Mmm1∆(2-75)p-GFP (Fig. 4C). This observation indicates that
amino acid residues 75-90 somehow affect expression or turnover of the Mmm1 mRNA or
protein. Since mutant fusion proteins lacking the N-terminal extension function as well as the
wildtype fusion protein and produce similar numbers of Mmm1p foci, these data suggest that the
steady-state level of Mmm1p expressed in wildtype cells is not limiting.
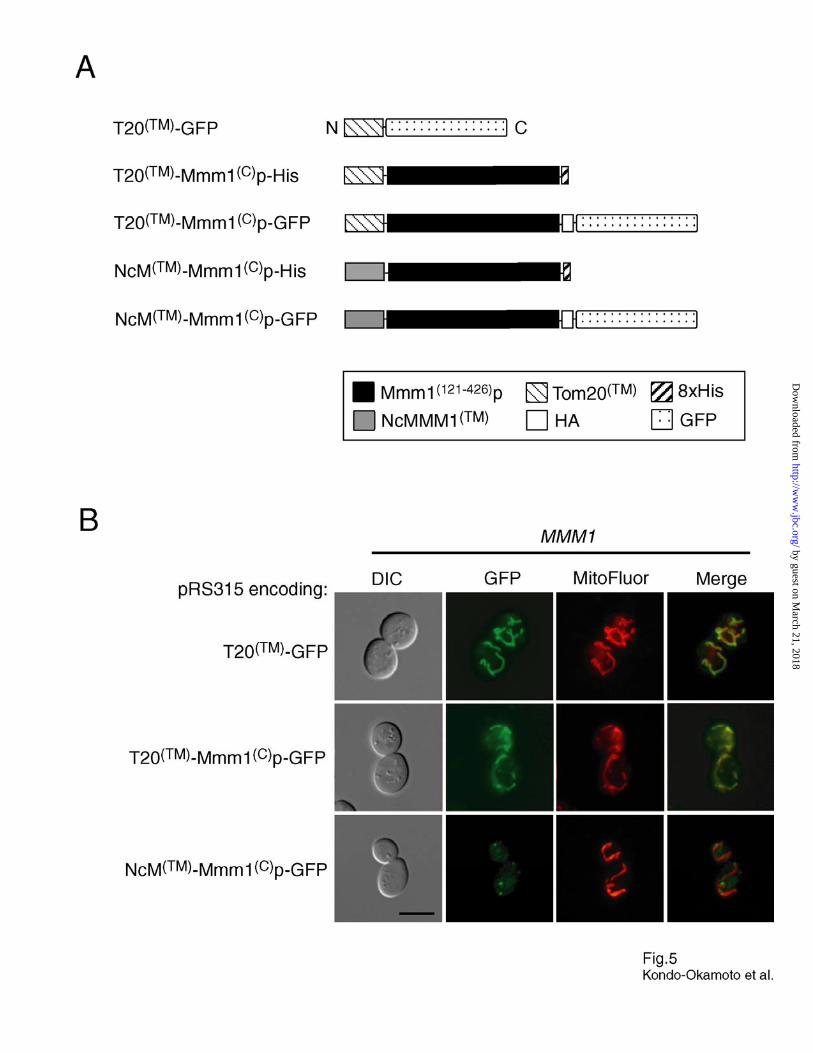
The TM Segment of Mmm1p Is Essential for Foci Formation—To investigate the role of the
Mmm1p TM segment in foci formation, we constructed plasmids encoding chimeric proteins
containing the TM segment of Tom20p or NcMMM1, and the large C-terminal domain of
by guest on March 21, 2018
http://ww
w.jbc.org/
Dow
nloaded from

Kondo-Okamoto et al.
- 17 -
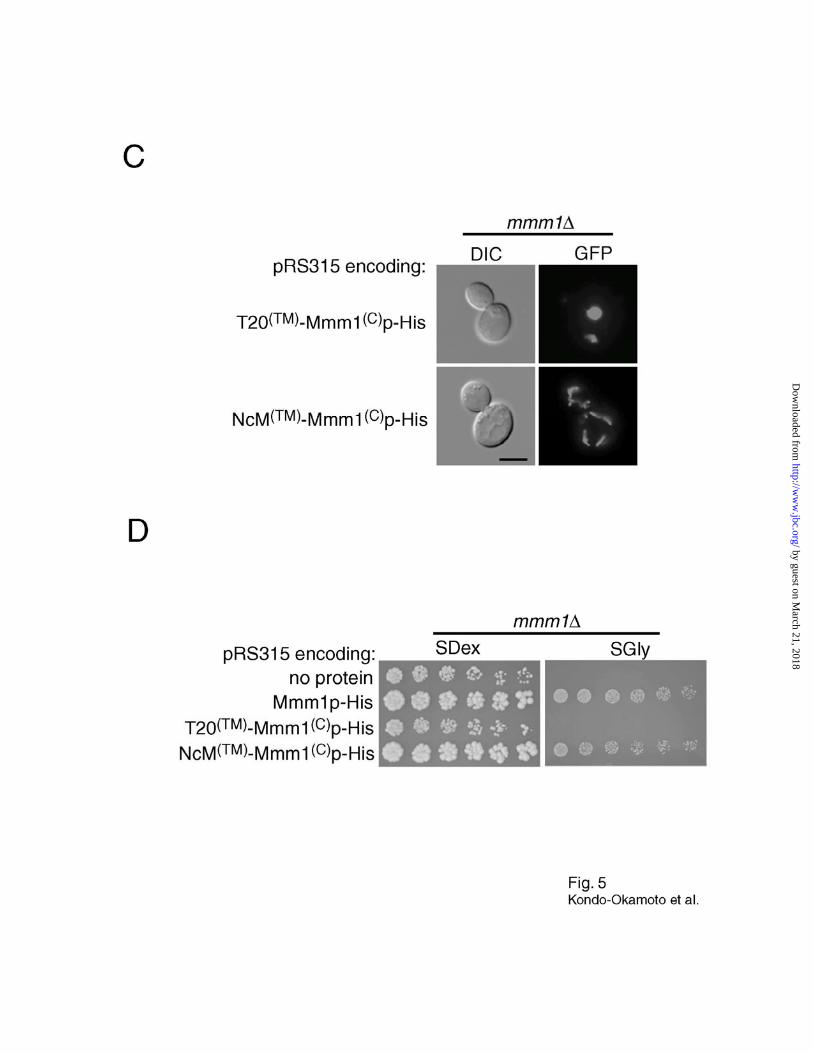
Mmm1p-His (T20(TM)-Mmm1(C)p-His and NcM(TM)-Mmm1(C)p-His, respectively) (Fig. 5A). For
localization studies, GFP was fused at the C-terminus of T20(TM)-Mmm1(C)p and NcM(TM)-
Mmm1(C)p (T20(TM)-Mmm1(C)p-GFP and NcM(TM)-Mmm1(C)p-GFP, respectively) (Fig. 5A).
Tom20p contains a single TM segment at the N-terminus embedded in the outer mitochondrial
membrane, and has the topology of Nin-Ccytoplasm (45). A previous study showed that the human
Tom20 TM segment plus some flanking residues fused at the N-terminus of GFP can be targeted
and inserted into the OM in humans (46).
When expressed in wildtype cells, GFP fused to the C-terminus of the Tom20p TM segment
(T20(TM)-GFP, Fig. 5A) colocalized with mitochondria and was evenly distributed throughout the
tubular network (Fig. 5B). Similarly, T20(TM)-Mmm1(C)p-GFP localized throughout the
mitochondrial network without forming visible foci (Fig. 5B). In contrast, NcM(TM)-Mmm1(C)p-
GFP formed foci that colocalized with mitochondrial tubules (Fig. 5B). These results indicate
that the TM segment of Mmm1p is essential for foci formation, and can be replaced with that of
NcMMM1. Interestingly, T20(TM)-Mmm1(C)p-His did not restore normal mitochondrial tubular
networks and glycerol growth in mmm1 cells, whereas NcM(TM)-Mmm1(C)p-His suppressed
these defects in the mutant (Fig. 5C, D and Table I). These data, together with the observation
that the N-terminus of NcMMM1 is exposed to the matrix of yeast mitochondria (N. Kondo-
Okamoto, unpublished data), suggest that the double membrane-spanning topology is crucial for
Mmm1p to localize as discrete foci where the protein performs its function.
The C-terminal Domain of Mmm1p Is Indispensable for Targeting to Mitochondria—To
define the region required for targeting Mmm1p to mitochondria, we expressed plasmids
encoding GFP fused to the C-terminus of various Mmm1p deletion mutants in wildtype cells
by guest on March 21, 2018
http://ww
w.jbc.org/
Dow
nloaded from

Kondo-Okamoto et al.
- 18 -
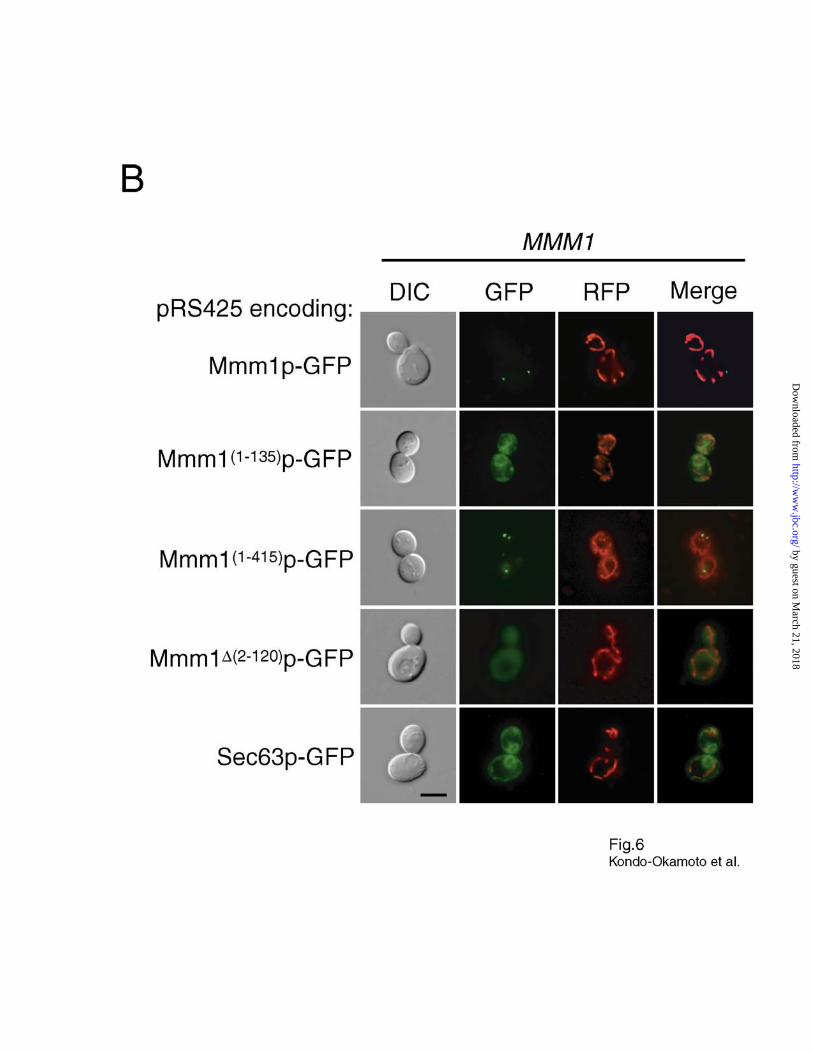
(Fig. 6A). Surprisingly, Mmm1(1-135)p-GFP was targeted to perinuclear structures similar to those
labeled with the endoplasmic reticulum (ER) marker Sec63p-GFP (39) (Fig. 6B). Since we
already showed that removal of the N-terminus does not interfere with Mmm1p targeting or foci
formation, this finding indicates that the TM segment of Mmm1p does not contain a complete
signal for targeting to mitochondria. Additional studies established that Mmm1(1-180)p-GFP and
Mmm1(1-275)p-GFP were also targeted to the ER, while Mmm1(1-365)p-GFP and Mmm1(1-390)p-GFP
were targeted to both the ER and cytoplasm (Fig. 6A). In contrast, Mmm1(1-415)p-GFP formed
discrete foci that were colocalized with the mitochondrial network (Fig. 6B), and restored normal
mitochondrial morphology and glycerol growth in mmm1 cells (data not shown). These results
indicate that the last 11 amino acids of Mmm1p are not required for mitochondrial targeting and
foci formation. Mmm1∆(2-120)p-GFP, a mutant lacking the TM segment, was targeted to the
cytoplasm (Fig. 6B). Collectively, these observations demonstrate that, together with the TM
segment, the C-terminal domain plays an important role in targeting Mmm1p to mitochondria.
DISCUSSION
Double Membrane-spanning Topology of Mmm1p—Previous studies suggested that: 1)
Mmm1p spans the OM with its C-terminus exposed to the cytoplasm (27), and 2) Mmm1p forms
discrete foci at membrane contact sites, adjacent to mtDNA nucleoids in the matrix (30).
Although the latter finding raised the possibility that the N-terminus of Mmm1p spans the IM,
the submitochondrial location of Mmm1p’s N-terminus has not been reported (30). In this study,
we used in vivo site-specific cleavage and in vitro protease protection assays to demonstrate that
the N-terminus of Mmm1p is exposed to the mitochondrial matrix. Together, these findings
establish that Mmm1p spans both mitochondrial membranes with the topology of Nmatrix-Ccytoplasm.
by guest on March 21, 2018
http://ww
w.jbc.org/
Dow
nloaded from

Kondo-Okamoto et al.
- 19 -
Tim23p is the only other protein reported to span both mitochondrial membranes in yeast
(47). This protein contains an N-terminal domain anchored in the OM, a middle (hydrophilic)
domain exposed to the IMS, and four, C-terminal TM helices embedded in the IM (47). Unlike
Tim23p, Mmm1p contains only one predicted TM segment and is unlikely to contain a stretch
exposed to the IMS. This interpretation is supported by our observation that the N-terminal
extension of Mmm1p was not degraded further by externally added protease even when the OM
was ruptured by osmotic shock (Fig. 1C).
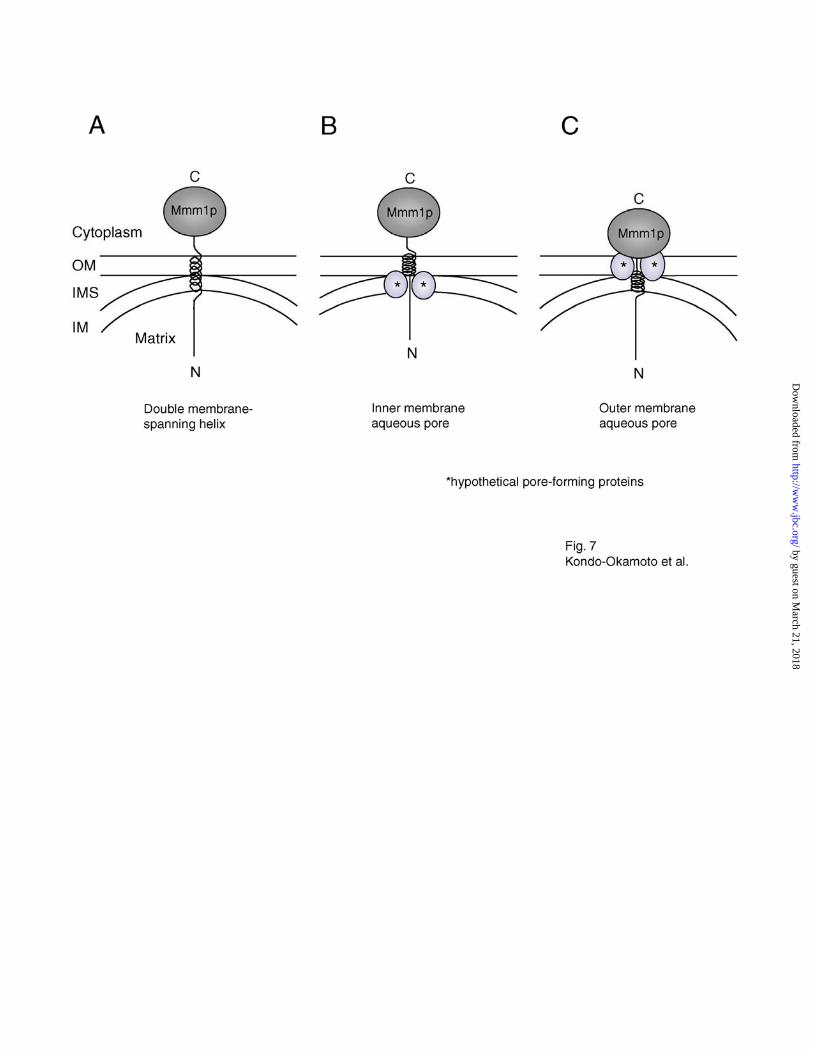
In the absence of a second TM segment, at least three different models could account for the
double membrane-spanning topology of Mmm1p. First, Mmm1p might span both mitochondrial
membranes once via its single TM segment, as depicted in Fig. 7A. This scenario is problematic,
if the TM helix of Mmm1p is only ~25 amino acid residues in length (amino acids 92-116), as
previously predicted (Kyte-Doolittle hydrophobicity prediction) (27). Unless this region adopts
an unconventional structure (e.g. a dramatically extended helix), additional amino acids flanking
the predicted TM helix would be required to span two lipid bilayers. In fact, other prediction
programs including SOSUI (amino acids 102-123), HMMTOP (amino acids 101-120) and
TMHMM (amino acids 100-122), predict slightly different TM helices within the Mmm1 protein
(ExPASy Molecular Biology Server). When the results of all four prediction programs are
combined, an Mmm1p TM segment of 32 residues (amino acids 92-123) results. If a TM
segment of this length exists in Mmm1p, it might be sufficient to span both mitochondrial
membranes. Second, the TM segment of Mmm1p could be embedded in the OM and the N-
terminal extension could span the IM through an aqueous pore formed by other members of a
protein complex (Fig. 7B). Third, the C-terminal flanking region of the Mmm1p TM segment
could span the OM through an aqueous pore formed by proteins in the OM (Fig. 7C). Further
by guest on March 21, 2018
http://ww
w.jbc.org/
Dow
nloaded from

Kondo-Okamoto et al.
- 20 -
studies are required to define the length, position, structure and environment of Mmm1p’s
double membrane-spanning domain.
A recent study indicates that two additional OM proteins, Mdm10p and Mdm12p, form
mitochondrial foci adjacent to mtDNA nucleoids (48). Like the mmm1 strain, cells lacking
Mdm10p and Mdm12p contain spherical mitochondria and lose mtDNA nucleoids (25,26). In
addition, all three proteins behave as if they form a complex in immune precipitation
experiments (48), and localization of Mmm1p to mitochondrial foci does not occur in the
absence of Mdm10p and Mdm12p (48) (N. Kondo-Okamoto, unpublished data). Based on these
observations, it is likely that Mdm10p and Mdm12p interact with Mmm1p, forming a complex at
the membrane contact site. The topology of Mdm10p and Mdm12p with respect to both
mitochondrial membranes, and the domains of those proteins required for interaction with
Mmm1p are important issues to be addressed in the future.
Role of the Mmm1p TM Segment in Foci Formation and Generation of Contact Sites—Our
finding that the double membrane-spanning topology of Mmm1p is critical for foci formation
(Fig. 5) supports the notion that Mmm1p functions at contact sites between the inner and outer
mitochondrial membranes. Although we cannot rule out the possibility that a subgroup of
Mmm1p molecules is distributed throughout mitochondria in a single membrane-spanning
fashion, the results presented here suggest that the membrane contact site is the place where
Mmm1p forms a functional complex. Despite three decades of research on mitochondrial
structure and protein import, proteins that direct the formation of these morphologically defined
membrane contact sites have proven difficult to identify (49). Here we demonstrate that the
double membrane-spanning protein Mmm1 serves as a marker of this intramitochondrial
by guest on March 21, 2018
http://ww
w.jbc.org/
Dow
nloaded from

Kondo-Okamoto et al.
- 21 -
structure. Is Mmm1p a structural determinant of the mitochondrial membrane contact site,
attaching the OM to the IM? Based on the observation that the IM is strikingly disorganized in
mmm1 cells, a role for Mmm1p in forming this structure has been proposed (30). However, the
disorganized IM observed in mmm1 mutants could also be a secondary consequence of
converting mitochondrial membranes from tubular networks to spherical compartments.
Moreover, it has not been firmly established whether the membrane contact sites are lost and/or
destabilized in mitochondria lacking Mmm1p. High-resolution electron microscopy and 3D
tomography of mmm1 mitochondria will help to address this issue.
Role of the Mmm1p N-terminal Extension in MtDNA Nucleoid Maintenance—The unique
Nmatrix-Ccytoplasm topology of Mmm1p raised the possibility that this is a bifunctional protein with
two topologically distinct domains, an N-terminal extension for mtDNA nucleoid maintenance
and a C-terminal domain for mitochondrial morphology maintenance. This hypothesis predicts
that deletion of the N-terminal extension will lead to mtDNA loss but will not affect
mitochondrial morphology. However, data presented here indicate that the N-terminal extension
of Mmm1p is not required for maintenance of normal tubular networks or mtDNA nucleoids
(Fig. 2). Thus, it is unlikely that Mmm1p directly anchors mtDNA nucleoids to the matrix side of
the IM. This interpretation is supported by our observation that, despite the lack of an N-terminal
extension, the N. crassa MMM1 homologue is fully functional in budding yeast (Fig. 3).
How is Mmm1p involved in maintenance of mtDNA? We cannot exclude the possibility that
Mmm1p functions indirectly to attach mtDNA nucleoids to the IM. For example, Mmm1p may
interact with one or more proteins whose matrix-exposed domains directly bind mtDNA
nucleoids. In the absence of functional Mmm1p, such protein(s) might not target, assemble or
by guest on March 21, 2018
http://ww
w.jbc.org/
Dow
nloaded from

Kondo-Okamoto et al.
- 22 -
function properly to anchor mtDNA nucleoids at the membrane contact site, ultimately resulting
in mtDNA loss. Alternatively, unknown factor(s) might anchor mtDNA nucleoids to the
membrane contact site independently of Mmm1p. In this scenario, mtDNA nucleoid aggregation
and the subsequent mtDNA loss phenotypes of mmm1 mutants (30) may be a secondary
consequence of mitochondrial morphology defects that grossly alter IM structure.
Role of Mmm1p in Cytoskeletal Attachment—Although a role for Mmm1p in cytoskeletal
attachment has been proposed, two lines of evidence suggest that this protein does not directly
mediate interactions between mitochondria and the cytoskeleton. First, in N. crassa,
microtubules are the cytoskeletal elements utilized for mitochondrial morphology, distribution
and inheritance (31). However, in vitro studies showed that NcMMM1-depleted mitochondria
still bind to microtubules (33), indicating that NcMMM1 is not essential for microtubule-
dependent mitochondrial behavior. Second, we show here that NcMMM1 can substitute for
Mmm1p in S. cerevisiae (Fig. 3). In contrast to N. crassa, S. cerevisiae utilizes the actin
cytoskeleton for mitochondrial attachment and movement (5,50). Thus, the primary function of
Mmm1p has apparently been conserved between S. cerevisiae and N. crassa, independent of the
cytoskeletal system used by the cell to regulate mitochondrial dynamics. Based on these results,
it seems unlikely that the Mmm1 protein acts as a direct molecular bridge between mitochondria
and cytoskeletal elements to maintain tubular networks.
How, then, can we reconcile the observation that S. cerevisiae mitochondria bind actin
filaments in an Mmm1p-dependent fashion in vitro (28)? One possibility is that S. cerevisiae and
N. crassa Mmm1 proteins bind molecular adaptors that bridge interactions between mitochondria
and actin filaments in both cell types. Loss of actin interactions would have a severe affect in
by guest on March 21, 2018
http://ww
w.jbc.org/
Dow
nloaded from

Kondo-Okamoto et al.
- 23 -
budding yeast, since actin filaments and cables are the major cytoskeletal system found in the
cytoplasm. In contrast, both actin and microtubules are found as prominent cytoplasmic
structures in N. crassa. Since N. crassa relies most heavily on microtubules for mitochondrial
dynamics and presumably has additional proteins that mediate microtubule attachment, loss of
Mmm1p actin-binding activity produces only subtle (if any) defects. Identifying the complete
set of cytoskeleton-binding and motor proteins that interact with mitochondria in both organisms
will help to clarify this discrepancy.
The Mmm1p Cytoplasmic Domain Is Required for Mitochondrial Targeting—To our surprise,
we found that Mmm1p mutant proteins lacking the cytoplasmic C-terminal domain were targeted
to the ER rather than mitochondria (Fig. 6). This result indicates that all or a part of the C-
terminal domain serves as a “subsignal” for mitochondrial targeting of Mmm1p. To date, only
one other protein is known to require cytoplasmic signals for mitochondrial targeting. An alpha
helix at the N-terminus of the single TM protein Bax was recently shown to contribute
significantly to mitochondrial targeting (51). These cytoplasmic “subsignals” may interact with
receptor proteins or specific recruiting factors that facilitate faithful targeting.
Why are Mmm1p mutants lacking the C-terminal domain targeted to the ER? This
phenomenon may be due to the hydrophobicity and/or length of the membrane-spanning domain.
For example, human Tom20 variants with increased hydrophobicity and length in the TM
segment are targeted to ER/Golgi compartments (46). Similarly, increasing the TM domain
hydrophobicity of another outer mitochondrial membrane protein, Fis1, also results in ER
targeting (52). In the absence of cytoplasmic C-terminal sequences, the excess hydrophobicity
and/or length of the Mmm1p TM segment may define a default signal for ER targeting.
by guest on March 21, 2018
http://ww
w.jbc.org/
Dow
nloaded from

Kondo-Okamoto et al.
- 24 -
What are the proteins that interact with the C-terminal domain of Mmm1p and recruit
Mmm1p to mitochondria? As described above, recent studies suggest that at least two other
proteins, Mdm10p and Mdm12p, are required to target Mmm1p to mitochondria (48) (N. Kondo-
Okamoto, unpublished observations). In addition to components of the OM protein import
machinery, other, as yet unidentified, proteins may play a role in targeting Mmm1p to
mitochondria. Studying how Mmm1p is targeted to mitochondria, inserted into the outer and
inner membranes, and assembled into a complex at the membrane contact site will help us to
understand the molecular function of Mmm1p and the mechanisms and pathways underlying
protein transport to a distinct intramitochondrial structure, the membrane contact site.
Acknowledgments—We thank Benedikt Westermann (University of Munich, Munich,
Germany) for plasmids pVT100U-mtGFP, pYX142-mtGFP, and the cDNA encoding NcMMM1,
Will Prinz (National Institute of Health, Bethesda, MD) for plasmids pJK59 and pWP1039, Carla
Koehler (University of California Los Angeles, Los Angeles, CA) for anti-Cyb2p antisera,
Elizabeth Craig (University of Wisconsin, Madison, WI) for anti-Ssc1p antisera, and members of
the Shaw laboratory for stimulating discussions and careful review of the manuscript. N.K.-O.
and K.O. are deeply grateful to Walter Neupert (University of Munich, Munich, Germany) for
his generous support during the initial stage of this study.
This work was supported by National Institutes of Health Grant GM-53466 awarded to J.M.S
and a grant from the United Mitochondrial Disease Foundation awarded to K.O. The University
of Utah DNA and Peptide Facility is supported in part by a grant from the National Cancer
Institute (CA42014).
by guest on March 21, 2018
http://ww
w.jbc.org/
Dow
nloaded from

Kondo-Okamoto et al.
- 25 -
REFERENCES
1. Hermann, G. J., and Shaw, J. M. (1998) Ann. Rev. Cell Dev. Biol. 14, 265-303
2. Yaffe, M. P. (1999) Science 283, 1493-1497
3. Griparic, L., and van der Bliek, A. M. (2001) Traffic 2, 235-244
4. Jensen, R. E., Aiken Hobbs, A. E., Cerveny, K. L., and Sesaki, H. (2001) Microsc. Res.
Tech. 51, 573-583
5. Boldogh, I. R., Yang, H.-C., and Pon, L. A. (2001) Traffic 2, 368-374
6. Nunnari, J., Marshall, W. F., Straight, A., Murray, A., Sedat, J. W., and Walter, P. (1997)
Mol. Biol. Cell 8, 1233-1242
7. Yoon, Y., and McNiven, M. A. (2001) Curr. Biol. 11, R67-R70
8. Shaw, J. M., and Nunnari, J. (2002) Trends Cell Biol. 12, 178-184
9. Westermann, B. (2002) EMBO Rep. 3, 527-531
10. Mozdy, A. D., and Shaw, J. M. (2003) Nat. Rev. Mol. Cell Biol. 4, 468-478
11. Dimmer, K. S., Fritz, S., Fuchs, F., Messerschmitt, M., Weinbach, N., Neupert, W., and
Westermann, B. (2002) Mol. Biol. Cell 13, 847-853
12. Berger, K. H., and Yaffe, M. P. (2000) Trends Microbiol. 8, 508-513
13. Contamine, V., and Picard, M. (2000) Microbiol. Mol. Biol. Rev. 64, 281-315
14. Scott, S. V., Cassidy-Stone, A., Meeusen, S. L., and Nunnari, J. (2003) Curr. Opin. Cell
Biol. 15, 482-488
15. Hermann, G. J., Thatcher, J. W., Mills, J. P., Hales, K. G., Fuller, M. T., Nunnari, J., and
Shaw, J. M. (1998) J. Cell Biol. 143, 359-373
16. Rapaport, D., Brunner, M., Neupert, W., and Westermann, B. (1998) J. Biol. Chem. 273,
20150-20155
by guest on March 21, 2018
http://ww
w.jbc.org/
Dow
nloaded from

Kondo-Okamoto et al.
- 26 -
17. Sesaki, H., and Jensen, R. E. (2001) J. Cell Biol. 152, 1123-1134
18. Jones, B. A., and Fangman, W. L. (1992) Genes Dev. 6, 380-389
19. Guan, K., Farh, L., Marshall, T. K., and Deschenes, R. J. (1993) Curr. Genet. 24, 141-148
20. Shepard, K. A., and Yaffe, M. P. (1999) J. Cell Biol. 144, 711-720
21. Wong, E. D., Wagner, J. A., Gorsich, S. W., McCaffery, J. M., Shaw, J. M., and Nunnari,
J. (2000) J. Cell Biol. 151, 341-352
22. Esser, K., Tursun, B., Ingenhoven, M., Michaelis, G., and Pratje, E. (2002) J. Mol. Biol.
323, 835-843
23. Herlan, M., Vogel, F., Bornhovd, C., Neupert, W., and Reichert, A. S. (2003) J. Biol.
Chem. 278, 27781-27788
24. McQuibban, G. A., Saurya, S., and Freeman, M. (2003) Nature 423, 537-541
25. Sogo, L. F., and Yaffe, M. P. (1994) J. Cell Biol. 126, 1361-1373
26. Berger, K. H., Sogo, L. F., and Yaffe, M. P. (1997) J. Cell Biol. 136, 545-553
27. Burgess, S. M., Delannoy, M., and Jensen, R. E. (1994) J. Cell Biol. 126, 1375-1391
28. Boldogh, I., Vojtov, N., Karmon, S., and Pon, L. A. (1998) J. Cell Biol. 141, 1371-1381
29. Yang, H.-C., Palazzo, A., Swayne, T. C., and Pon, L. A. (1999) Curr. Biol. 9, 1111-1114
30. Aiken Hobbs, A. E., Srinivasan, M., McCaffery, J. M., and Jensen, R. E. (2001) J. Cell
Biol. 152, 401-410
31. Steinberg, G., and Schliwa, M. (1993) J. Cell Sci. 106, 555-564
32. Prokisch, H., Neupert, W., and Westermann, B. (2000) Mol. Biol. Cell 11, 2961-2971
33. Fuchs, F., Prokisch, H., Neupert, W., and Westermann, B. (2002) J. Cell Sci. 115, 1931-
1937
34. Winston, F., Dollard, C., and Ricupero-Hovasse, S. L. (1995) Yeast 11, 53-55
by guest on March 21, 2018
http://ww
w.jbc.org/
Dow
nloaded from

Kondo-Okamoto et al.
- 27 -
35. Burke, D., Dawson, D., and Stearns, T. (2000) Methods in Yeast Genetics, 2000 Ed., Cold
Spring Harbor Laboratory, Cold Spring Harbor, NY
36. Sambrook, J., and Russell, D. W. (2001) Molecular Cloning, 3rd Ed., Cold Spring Harbor
Laboratory, Cold Spring Harbor, NY
37. Westermann, B., and Neupert, W. (2000) Yeast 16, 1421-1427
38. Diekert, K., de Kroon, A. I., Kispal, G., and Lill, R. (2001) Methods Cell Biol. 65, 37-51
39. Prinz, W. A., Grzyb, L., Veenhuis, M., Kahana, J. A., Silver, P. A., and Rapoport, T. A.
(2000) J. Cell Biol. 150, 461-474
40. Dougherty, W. G., and Semler, B. L. (1993) Microbiol. Rev. 57, 781-822
41. Carrington, J. C., and Dougherty, W. G. (1988) Proc. Natl. Acad. Sci. U. S. A. 85, 3391-
3395
42. Faber, K. N., Kram, A. M., Ehrmann, M., and Veenhuis, M. (2001) J. Biol. Chem. 276,
36501-36507
43. Wong, E. D., Wagner, J. A., Scott, S. V., Okreglak, V., Holewinske, T. J., Cassidy-Stone,
A., and Nunnari, J. (2003) J. Cell. Biol. 160, 303-311
44. Esaki, M., Kanamori, T., Nishikawa, S., and Endo, T. (1999) Proc. Natl. Acad. Sci. U. S.
A. 96, 11770-11775
45. Ramage, L., Junne, T., Hahne, K., Lithgow, T., and Schatz, G. (1993) EMBO J. 12, 4115-
4123
46. Kanaji, S., Iwahashi, J., Kida, Y., Sakaguchi, M., and Mihara, K. (2000) J. Cell Biol. 151,
277-288
47. Donzeau, M., Kaldi, K., Adam, A., Paschen, S., Wanner, G., Guiard, B., Bauer, M. F.,
Neupert, W., and Brunner, M. (2000) Cell 101, 401-412
by guest on March 21, 2018
http://ww
w.jbc.org/
Dow
nloaded from

Kondo-Okamoto et al.
- 28 -
48. Boldogh, I. R., Nowakowski, W. D., Yang, H.-C., Chung, H., Karmon, S., Royes, P., and
Pon, L. A. (2003) Mol. Biol. Cell in press
49. Reichert, A. S., and Neupert, W. (2002) Biochim. Biophys. Acta 1592, 41-49
50. Itoh, T., Watabe, A., Toh-e. A., and Matsui, Y. (2002) Mol. Cell. Biol. 22, 7744-7757
51. Cartron, P. F., Priault, M., Oliver, L., Meflah, K., Manon, S., and Vallette, F. M. (2003) J.
Biol. Chem. 278, 11633-11641
52. Beilharz, T., Egan, B., Silver, P. A., Hofmann, K., and Lithgow, T. (2003) J. Biol. Chem.
278, 8219-8223
by guest on March 21, 2018
http://ww
w.jbc.org/
Dow
nloaded from

Kondo-Okamoto et al.
- 29 -
FIGURE LEGENDS
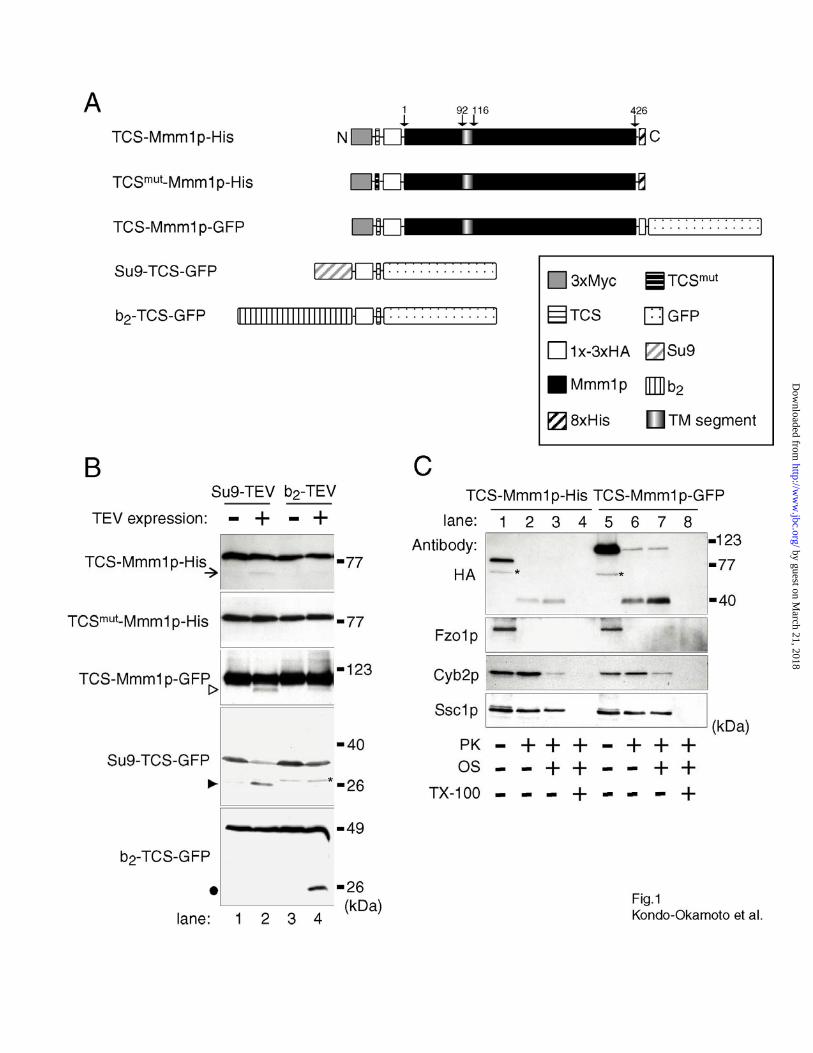
Fig. 1. The N-terminus of Mmm1p is exposed to the mitochondrial matrix. A, schematic
representation of proteins used for in vivo site-specific cleavage and in vitro protease protection
assays. Mmm1p is 426 amino acids in length. The TM segment designated in Burgess et al.
(1994) is shown (amino acids 92-116). B, in vivo site-specific cleavage in cells harboring
plasmids that encode a TEV protease (top) and a substrate protein (left). The expression of TEV
proteases was repressed by dextrose (lanes 1 and 3) or induced by galactose (lanes 2 and 4).
Protein extracts from 0.5 OD600 units of cells were analyzed by immunostaining with antibodies
specific for HA (for TCS-Mmm1p-His, TCSmut-Mmm1p-His and TCS-Mmm1p-GFP expressed
in mmm1 cells) and GFP (for Su9-TCS-GFP and b2-TCS-GFP expressed in wildtype cells). The
asterisk indicates a form of Su9-TCS-GFP produced by the mitochondrial processing peptidase.
TEV protease cleavage products for each substrate are marked to the left of each panel by an
arrow, open arrowhead, closed arrowhead or closed circle. C, in vitro proteinase K (PK)
treatment of mitochondria isolated from mmm1 cells expressing TCS-Mmm1p-His or TCS-
Mmm1p-GFP. Mitochondria (30 µg of protein) were mock-treated (lanes 1 and 5), treated with
200 µg/ml PK under osmotic conditions (lanes 2 and 6) or subjected to osmotic shock (OS) in
the absence (lanes 3 and 7) or presence (lanes 4 and 8) of Triton X-100 (TX-100). Samples were
analyzed by immunostaining with antibodies specific for HA (for TCS-Mmm1p-His and TCS-
Mmm1p-GFP), Fzo1p (for the OM protein), Cyb2p (for the IMS protein) and Ssc1p (for the
matrix protein). Asterisks indicate forms produced by non-specific degradation.
Fig. 2. The N-terminal extension of Mmm1p is not essential for mitochondrial
morphology or mtDNA maintenance. A, schematic representation of Mmm1p deletion mutant
by guest on March 21, 2018
http://ww
w.jbc.org/
Dow
nloaded from

Kondo-Okamoto et al.
- 30 -
proteins. B, mmm1 cells carrying plasmid pRS315 alone (no protein expressed) or encoding
Mmm1p-His, Mmm1∆(2-90)p-His, or Mmm1∆(2-120)p-His were transformed with pVT100U-mtGFP,
grown to log phase in SD media and observed by fluorescence microscopy to visualize
mitochondria. Bar, 5 µm. C, cells described in B (without pVT100U-mtGFP) were grown
overnight in SD media, spotted as two-fold serial dilutions, and incubated for three (SD plates)
or six (SGly plates) days.
Fig. 3. N. crassa MMM1 can substitute for Mmm1p in yeast. A, mmm1 cells carrying the
plasmid pYX142 alone (no protein expressed) or encoding TCS-Mmm1p-His, or TCS-
NcMMM1 were transformed with pVT100U-mtGFP, grown to log phase in SD media, and
observed by fluorescence microscopy to visualize mitochondria. Bar, 5 µm. B, cells described in
A (without pVT100U-mtGFP) were grown overnight in SD media, spotted as two-fold serial
dilutions, and incubated for three (SD plates) or six (SGly plates) days.
Fig. 4. Deletion of the Mmm1p N-terminal extension does not affect foci formation but
reduces steady-state protein levels. A, schematic representation of Mmm1p deletion variants
fused to the N-terminus of GFP. B, mmm1 cells carrying the plasmid pRS315 encoding
Mmm1p-GFP, Mmm1∆(2-75)p-GFP, or Mmm1∆(2-90)p-GFP were grown to log phase in SD media,
stained with MitoFluor Red 589, and observed by fluorescence microscopy. Bar, 5 µm. C,
protein extracts from 0.5 OD600 units of cells described in B were analyzed by immunostaining
with antibodies specific for GFP (for Mmm1p-GFP, Mmm1∆(2-75)p-GFP, and Mmm1∆(2-90)p-GFP)
and 3-PGK (as a cytoplasmic protein loading control).
by guest on March 21, 2018
http://ww
w.jbc.org/
Dow
nloaded from

Kondo-Okamoto et al.
- 31 -
Fig. 5. The double membrane-spanning topology of Mmm1p is crucial for foci
formation. A, schematic representation of chimeric fusion proteins. B, wildtype cells carrying
the plasmid pRS315 encoding T20(TM)-GFP, T20(TM)-Mmm1(C)p-GFP, or NcM(TM)-Mmm1(C)p-GFP
were grown to log phase in SD media, stained with MitoFluor Red 589, and observed by
fluorescence microscopy. Bar, 5 µm. C, mmm1 cells carrying the plasmid pRS315 encoding
T20(TM)-Mmm1(C)p-His or NcM(TM)-Mmm1(C)p-His were transformed with pVT100U-mtGFP,
grown to log phase in SD media, and observed by fluorescence microscopy to visualize
mitochondria. Bar, 5 µm. D, mmm1 cells carrying the plasmid pRS315 alone (no protein
expressed) or encoding Mmm1p-His, T20(TM)-Mmm1(C)p-His or NcM(TM)-Mmm1(C)p-His were
grown overnight in SD media, spotted as two-fold serial dilutions, and incubated for three days
(SD plates) or six days (SGly plates).
Fig. 6. The C-terminal domain of Mmm1p plays an essential role in mitochondrial
targeting. A, schematic representation of Mmm1p deletion variants fused to the N-terminus of
GFP. Wildtype cells containing pRS425 encoding each GFP fusion protein or pJK59 encoding
the ER marker Sec63p-GFP were transformed with pRS424ADH-mtRFP, grown to log phase in
SD media and observed by fluorescence microscopy. Localization of these GFP fusion proteins is
summarized at the right. B, representative images of wildtype cells expressing Mmm1p-GFP,
Mmm1(1-135)p-GFP, Mmm1(1-415)p-GFP, Mmm1∆(2-120)p-GFP, or Sec63p-GFP. Bar, 5 µm.
Fig. 7. Schematic models for the double membrane-spanning topology of Mmm1p.
Asterisks mark hypothetical pore-forming proteins in the outer or inner membrane. OM, outer
membrane; IMS, intermembrane space; IM, inner membrane. See text for details.
by guest on March 21, 2018
http://ww
w.jbc.org/
Dow
nloaded from

Kondo-Okamoto et al.
- 32 -
Table I. Mitochondrial Membrane Morphology in mmm1 Cells
Plasmid %Tubule %Intermediate* %Sphere
pRS315 5 30 65pRS315-MMM1-His 89 7 4pRS315-mmm1 (2-90)-His 94 4 2pRS315-mmm1 (2-120)-His 7 45 48
pRS315-T20(TM)-mmm1(C)-His 14 65 21pRS315-NcM(TM)-mmm1(C)-His 87 10 3
pYX142 12 41 47pYX142-TCS-MMM1-His 85 12 3pYX142-TCS-NcMMM1 96 2 2
Numbers are percent of cells grown to log phase in SD media at 30°C (n=200). Mitochondrial
morphology was visualized with matrix-targeted GFP, and scored in the three different categories
shown. *Intermediate denotes coexistence of tubules and spheres, or fragmented thick tubules.
by guest on March 21, 2018
http://ww
w.jbc.org/
Dow
nloaded from



Noriko Kondo-Okamoto, Janet M. Shaw and Koji Okamotodistinct domains for targeting and foci formation
Mmm1p spans both the outer and inner mitochondrial membranes, and contains
published online September 12, 2003J. Biol. Chem.
10.1074/jbc.M308436200Access the most updated version of this article at doi:
Alerts:
When a correction for this article is posted•
When this article is cited•
to choose from all of JBC's e-mail alertsClick here
by guest on March 21, 2018
http://ww
w.jbc.org/
Dow
nloaded from