modeling of calcium signaling pathways stefan schuster and beate knoke dept. of bioinformatics...
Post on 21-Dec-2015
223 views
TRANSCRIPT

Modeling of Calcium Signaling PathwaysStefan Schuster and Beate Knoke
Dept. of BioinformaticsFriedrich Schiller University Jena
Germany

1. Introduction
• Oscillations of intracellular calcium ions are important in signal transduction both in excitable and nonexcitable cells
• A change in agonist (hormone) level can lead to a switch between oscillatory regimes and stationary states digital signal
• Moreover, analogue signal encoded in frequency • Amplitude encoding and the importance of the
exact time pattern have been discussed; frequency encoding is main paradigm

Ca2+ oscillations in various types of nonexcitable cells
Astrocytes
Hepatocytes Oocytes
Pancreatic acinar cells

Vasopressin
Phenylephrine
Caffeine
UTP
Calmodulin
Calpain
PKC
…..
Effect 1
Effect 2
Effect 3
Bow-tie structure of signalling
How can one signal transmit several signals?
Ca2+ oscillation
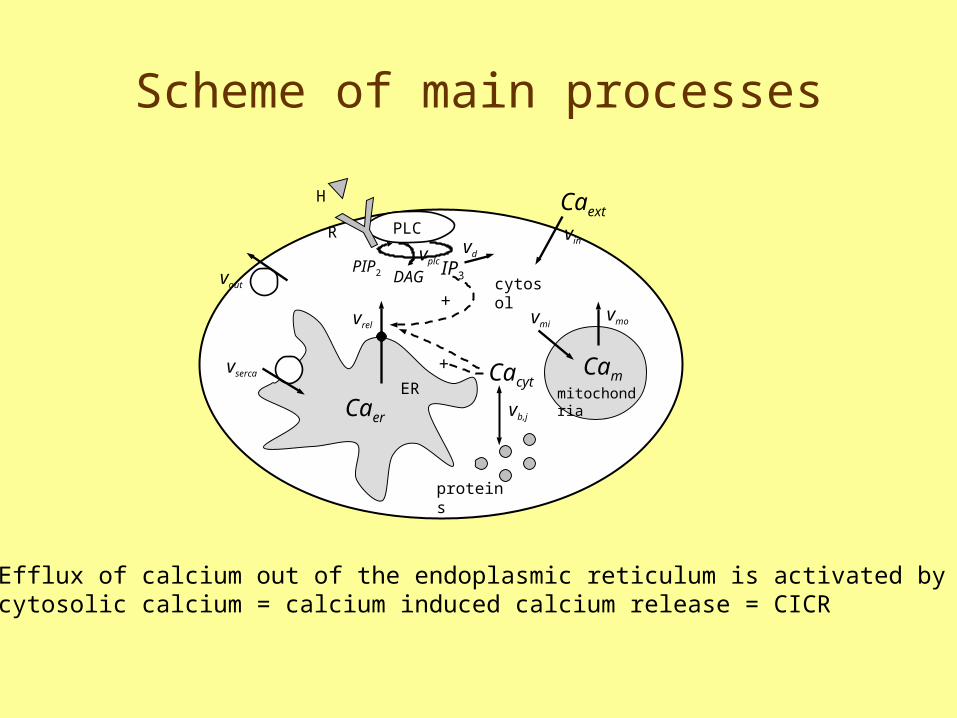
Scheme of main processes
PLCR
H
vout
vin
cytosol+
vrel
vserc
a
IP3
mitochondria
vmivmo
CamCacyt
vb,j
proteins
vplcvd
Caext
+
PIP2 DAG
ERCaer
Efflux of calcium out of the endoplasmic reticulum is activated by cytosolic calcium = calcium induced calcium release = CICR

Somogyi-Stucki model
• Is a minimalist model with only 2 independent variables: Ca2+ in cytosol (S1) and Ca2+ in endoplasmic reticulum (S2)
• All rate laws are linear except CICR
R. Somogyi and J.W. Stucki, J. Biol. Chem. 266 (1991) 11068
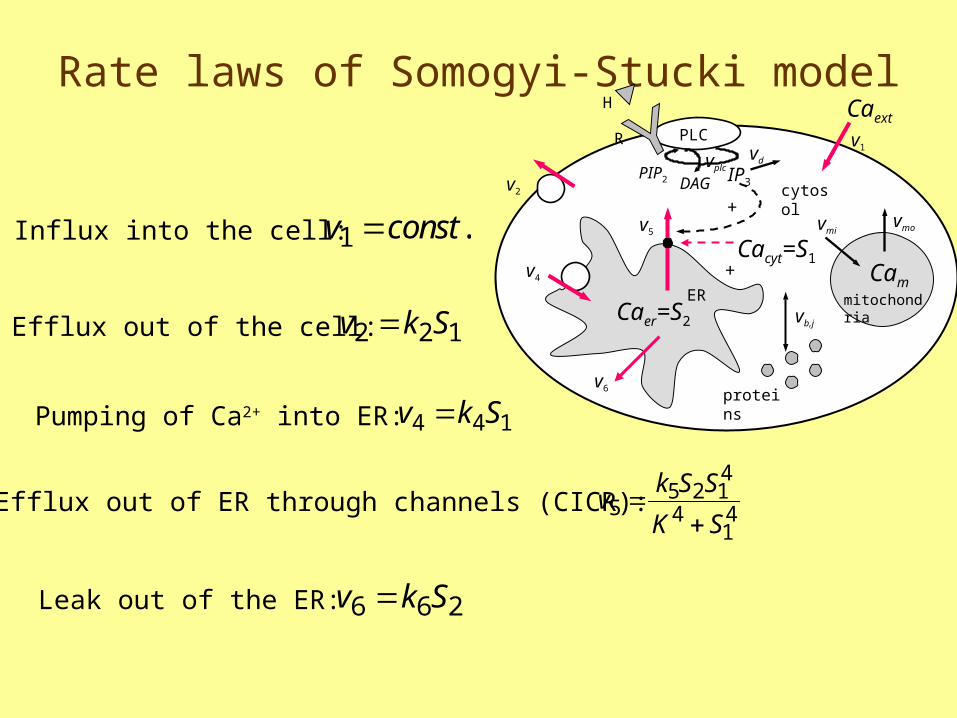
Rate laws of Somogyi-Stucki model
v const1 .
v k S2 2 1
144 Skv
Influx into the cell:
Efflux out of the cell:
Pumping of Ca2+ into ER:
Efflux out of ER through channels (CICR): vk S S
K S5
5 2 14
414
Leak out of the ER: v k S6 6 2
PLCR
H
v2
v1
cytosol+
v5
v4
IP3
mitochondria
vmivmo
Cam
Cacyt=S1
vb,j
proteins
vplcvd
Caext
+
PIP2 DAG
ERCaer=S2
v6

fast movement
slow movement
Relaxationoscillations!
Temporal behaviour

Many other models…
• by A. Goldbeter, G. Dupont, J. Keizer, Y.X. Li, T. Chay
etc.
• Reviewed, e.g., in Schuster, S., M. Marhl and T. Höfer.
Eur. J. Biochem. (2002) 269, 1333-1355 and Falcke, M.
Adv. Phys. (2004) 53, 255-440.
• Most models are based on calcium-induced calcium
release.

2. Bifurcation analysis of two models of calcium oscillations
• Biologically relevant bifurcation parameter in Somogyi-
Stucki model: rate constant of channel, k5 (CICR),
dependent on IP3
• Low k5 : steady state; medium k5: oscillations; high k5:
steady state.
• Transition points (bifurcations) between these regimes
can here be calculated analytically, be equating the trace
of the Jacobian matrix with zero.

Usual picture of Hopf bifurcations
Supercritical Hopf bifurcation Subcritical Hopf bifurcation
parameter
varia
ble
parameterva
riabl
e
stable limit cycle stable limit cycle
unstable limit cycle
Hysteresis!

Bifurcation diagram for calcium oscillations
Subcritical HB Supercritical HB
From: S. Schuster &M. Marhl, J. Biol. Syst.9 (2001) 291-314
oscillations

Schematic picture of bifurcation diagram
parameter
varia
ble
Bifurcation
Very steep increase in amplitude. This is likely to be physiologically advantageous because oscillations start with a distinct amplitude and, thus,misinterpretation of the oscillatory signal is avoided.
No hysteresis – signal is unique function of agonist level.

Global bifurcations
• Local bifurcations occur when the behaviour near a
steady state changes qualitatively
• Global bifurcations occur „out of the blue“, by a global
change
• Prominent example: homoclinic bifurcation

Homoclinic bifurcation
Saddle point Saddle point
Homoclinic orbit
Before bifurcationAt bifurcation
After bifurcation
Limit cycle
Saddle point
Necessary condition in 2D systems: at least 2 steady states(in Somogyi-Stucki model,only one steady state)
Unstable focus
S1
S2

Model including binding of Ca2+ to proteins and effect of ER transmembrane potential
PLCR
H
vout
vin
cytosol+
vrel
vserca
IP3
mitochondria
vmivmo
CamCacyt
vb,j
proteins
vplcvd
Caext
+
PIP2 DAG
ERCaer
Marhl, Schuster, Brumen, Heinrich, Biophys. Chem. 63 (1997) 221

PrCaPrleak ER,pump ER,ch ER,cyt
d
dJJJJJ
t
Ca
)(d
dleak ER,ch ER,pump ER,
ER
ERER JJJt
Ca
System equations
with
)(~2cyt
21
2cyt
ch ER,
CaCa ECaK
CagJ
cytpump ER,pump ER, CakJ )( cytERleak ER,leak ER, CaCakJ
PrcytPr CakJ PrCakJ CaPr
Nonlinear equation for transmembrane potential
2D model

…this gives rise to ahomoclinic bifurcation
varia
ble
Saddle point
oscillation
As the velocity of the trajectory tends to zero when it approachesthe saddle point, the oscillationperiod becomes arbirtrarily longnear the bifurcation.
parameter
Hopf bifn.
Schuster &Marhl, J. Biol. Syst.9 (2001) 291
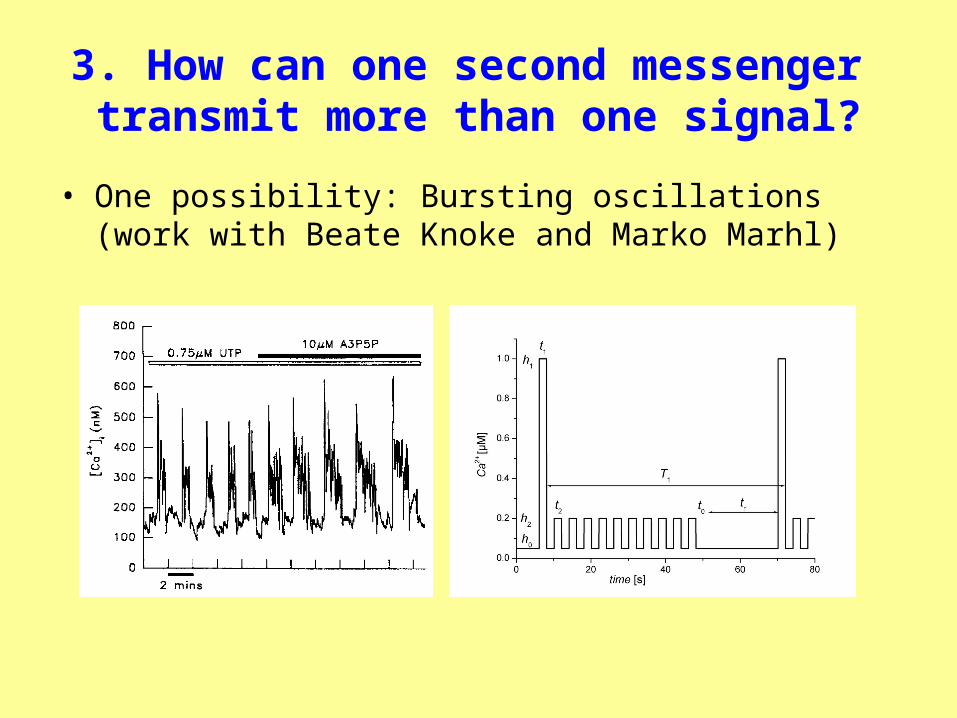
3. How can one second messenger transmit more than one signal?
• One possibility: Bursting oscillations (work with Beate Knoke and Marko Marhl)

Differential activation of two Ca2+ - binding proteins
41T
1 4 41
*Prot CaProt Ca
K Ca
42T
2 4 44
2I
*
( )* 1
Prot CaProt Ca
CaK Ca
K

Selective activation of protein 1
Prot1
Prot2

Selective activation of protein 2
Prot1
Prot2
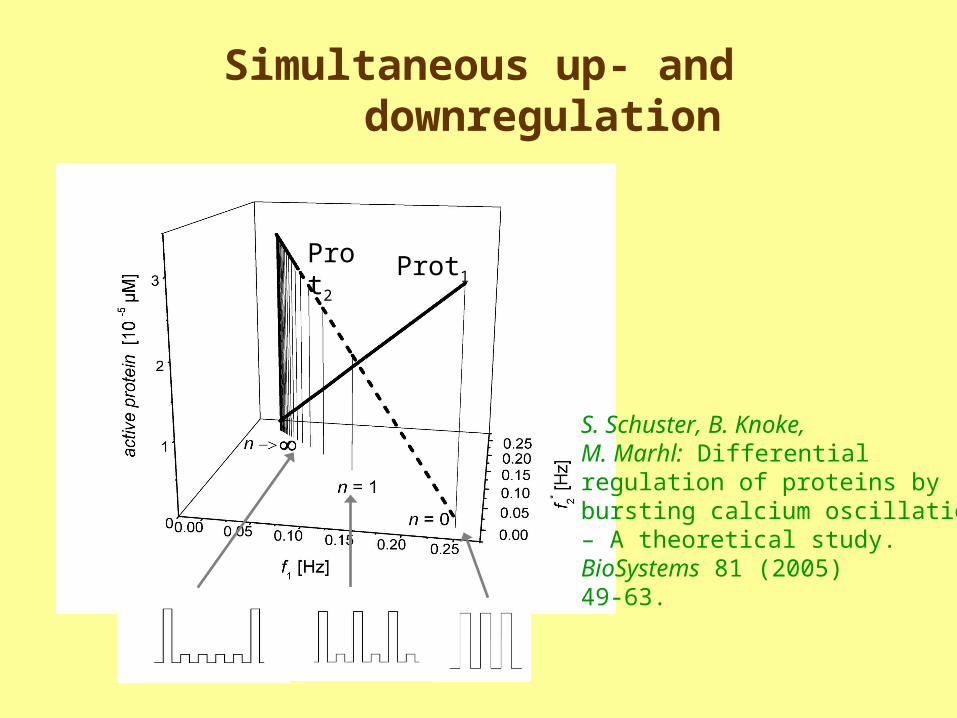
Simultaneous up- and downregulation
Prot1Prot2
S. Schuster, B. Knoke, M. Marhl: Differential regulation of proteins by bursting calcium oscillations – A theoretical study. BioSystems 81 (2005)49-63.
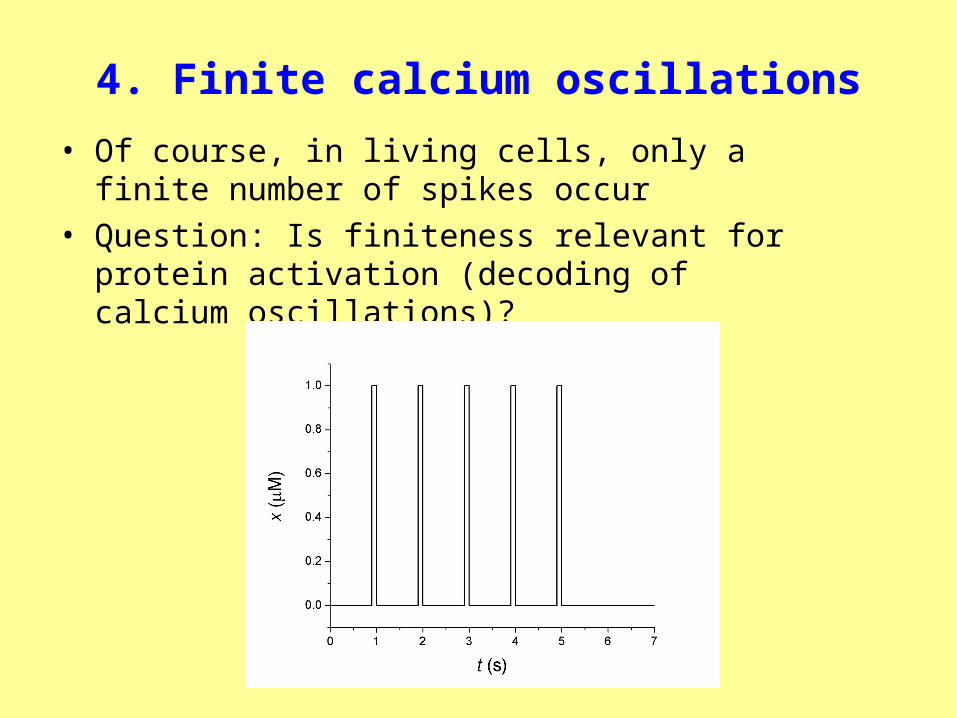
4. Finite calcium oscillations
• Of course, in living cells, only a finite number of spikes occur
• Question: Is finiteness relevant for protein activation (decoding of calcium oscillations)?

Intermediate velocity of binding is best
kon = 1 s-1M-4
kon = 15 s-1mM-4kon = 500 s-1mM-4
koff/kon = const. = 0.01 M4
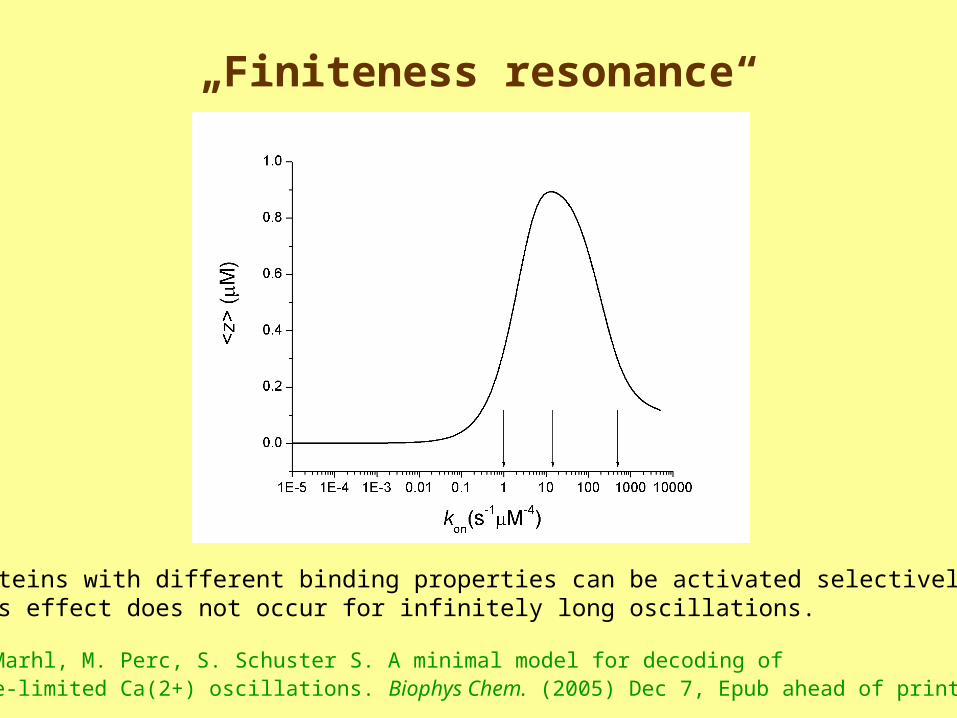
„Finiteness resonance“
Proteins with different binding properties can be activated selectively. This effect does not occur for infinitely long oscillations.
M. Marhl, M. Perc, S. Schuster S. A minimal model for decoding of time-limited Ca(2+) oscillations. Biophys Chem. (2005) Dec 7, Epub ahead of print

5. Discussion
• Relatively simple models (e.g. Somogyi-Stucki) can give rise to
complex bifurcation behaviour.
• Relaxation oscillators allow jump-like increase in amplitude at
bifurcations and do not show hysteresis.
• At global bifurcations, oscillations start with a finite (often large)
amplitude.
• Physiologically advantageous because misinterpretation of the
oscillatory signal is avoided in the presence of fluctuations.

Discussion (2)• Near homoclinic bifurcations, oscillation period can get
arbitrarily high.
• This may be relevant for frequency encoding. Frequency can be varied over a wide range.
• Bursting oscillations may be relevant for transmitting two signals simultaneously – experimental proof is desirable
• Thus, complex oscillations as found in, e.g. hepatocytes, may be of physiological importance
• Finite trains of calcium spikes show resonance in protein activation
• Thus, selective activation of proteins is enabled

Cooperations
• Marko Marhl (University of Maribor, Slovenia)
• Thomas Höfer (Humboldt University, Berlin, Germany)
• Exchange with Slovenia supported by Research Ministries of both countries.