modularizacin de la funcin inmunolgica en la …148.206.53.84/tesiuami/uami13151.pdf · el sistema...
TRANSCRIPT



NOMBRE: Pérez Hernández José de Jesús MATRÍCULA: 99334642 TELÉFONO: 015959281079 LICENCIATURA: Biología Experimental DIVISIÓN: Ciencias Biológicas y de la Salud UNIDAD UNIVERSITARIA: Iztapalapa TRIMESTRE LECTIVO: 05-P TÍTULO DEL PROYECTO DE INVESTIGACIÓN DEL QUE DEPENDE EL PDI: Compartamentalización de la respuesta inmunológica en el SNC TITULO DEL PDI: Modularización de la función inmunológica en la corteza somatosensorial primaria del ratón adulto NOMBRES DE LOS ASESORES EXTERNOS: Dra. Anhaí Chavarria Krauser NOMBRES DE LOS ASESORES INTERNOS: Dra. Elisa Vega Avila

MODULARIZACIÓN DE LA FUNCIÓN INMUNOLÓGICA EN LA CORTEZA
SOMATOSENSORIAL PRIMARIA DEL RATÓN ADULTO
Antecedentes y Marco Teórico
El sistema nervioso central (SNC) está constituido por unidades modulares
denominadas columnas que agrupan células nerviosas y conexiones que procesan un tipo de
información semejante (Mountcastle, 1979; Leise, 1990). Por ejemplo, en la corteza visual
de los primates y felinos, existen columnas de células que procesan preferencialmente
información proveniente del ojo izquierdo o del ojo derecho (columnas de dominancia
ocular; Leise, 1990). La evidencia anatómica hasta ahora acumulada, junto con diversos
estudios neurofisiológicos, sugiere que las columnas corticales participan en el
procesamiento de información de tipo neural (Mountcastle; 1979, Livingstone et al; 1984,
Leise, 1990). Por otro lado, observaciones previas realizadas en el laboratorio sugieren que
la organización de la respuesta inmunológica difiere entre las distintas regiones del sistema
nervioso. Por ejemplo, cisticercos ubicados en el parénquima cerebral o en los surcos, se
asocian con niveles bajos de citocinas y anticuerpos específicos en el líquido
cefaloraquídeo. En contraste, los parásitos localizados en el espacio subaracnoideo de la
base o intraventriculares, inducen una respuesta más pronunciada con altos niveles de
citocinas y de anticuerpos específicos (Chavarria et al., 2005). Así mismo, los niveles de
expresión de citocinas en diversas regiones parenquimatosas del cerebro de mono difieren
entre sí en respuesta a la infección por Plasmodium coatneyi (Tongren et al., 2000). Esta
heterogeneidad no solamente se observa consecutiva a la infección directa del SNC, sino
también en las respuestas inmunológicas del mismo a infecciones periféricas. Para ilustrar
este punto cabe mencionar que la infección experimental con tripanosomas en la rata
conduce a una producción más elevada de citocinas en los plexos coroideos con relación al
parénquima cerebral (Sharafeldin et al, 1999). De esta forma, estas observaciones apoyan
la existencia de compartimentos inmunológicos en el SNC, cuya identidad estructural aún
desconocemos.
¿Cuales son las bases estructurales y fisiológicas de la compartamentalización de la
respuestas inmunológica en el SNC? Hasta donde sabemos aún no existe una respuesta a

esta pregunta. Es posible, sin embargo, que existan módulos definidos por el tipo, la
distribución y la densidad de células presentadoras de antígeno (Matyszak et al; 1996,
McMenamin et al; 2003, Guillemin et al; 2004, Galea et al; 2005). En apoyo a esta
posibilidad, se ha observado que los plexos coroideos y las meninges muestran
principalmente macrófagos y células dendríticas, mientras que en el parénquima cerebral la
microglia y los macrófagos perivasculares son los tipos celulares predominantes (Matyszak
et al; 1996, McMenamin et al; 2003, Guillemin et al; 2004, Galea et al; 2005). También se
ha observado que la expresión del MHCII, una molécula necesaria para la presentación de
antígenos, varía en diferentes regiones del SNC, siendo muy baja en la corteza cerebral y
alta en la médula espinal (Vass et al; 1990).
Para evaluar la hipótesis referida utilizamos el modelo de la corteza somatosensorial
primaria del ratón adulto. Esta corteza contiene un mapa corporal constituido por módulos
columnares denominados barriles (White et al; 1993). Cada uno de los barriles representa
grupos de mecanoreceptores localizados en las vibrisas faciales del ratón. Debido a su
definición anatómica, podemos identificarlos con mucha precisión (Woolsey et al; 1970;
Killackey et al; 1979). Esto facilitó la evaluación de la relación entre la modularidad neural
con la inmunológica.
Hipótesis
Existe una relación estructural y funcional entre los módulos neurales y los módulos
inmunológicos.
Objetivo General
Evaluar la relación anatomo-funcional entre los módulos neurales y los módulos
inmunológicos.
Objetivos Específicos
1. Evaluar la distribución y densidad de células microgliales, macrófagos
perivasculares y astrocitos en el interior de los barriles (modular) y en el tejido
circundante (no modular) en ratones adultos, utilizando técnicas de
inmunohistoquímica y análisis digital de imágenes.

2. Evaluar la distribución y densidad de células microgliales, macrófagos
perivasculares y astrocitos en el interior de los barriles (modular) y en el tejido
circundante (no modular) en ratones adultos, después de realizada una lesión
circunscrita, utilizando técnicas de inmunohistoquímica y análisis digital de
imágenes.
3. Evaluar la presencia de citocinas en el interior de los barriles (modular) y en el
tejido circundante (no modular) en ratones adultos, utilizando técnicas de
inmunohistoquímica y análisis digital de imágenes.
4. Evaluar la presencia de citocinas en el interior de los barriles (modular) y en el
tejido circundante (no modular) en ratones adultos, después de realizada una lesión
circunscrita, utilizando técnicas de inmunohistoquímica y análisis digital de
imágenes.
Metodología
Animales de experimentación
Se usaron ratones machos adultos de la cepa CD1 de 60 a 80 días de edad
(n=4/grupo experimental), los cuales fueron mantenidos en un ciclo luz:obscuridad 14:10,
con agua y comida ad libitum. Todos los procedimientos experimentales fueron aprobados
por los comités de ética locales.
Lesiones circunscritas de la zona modular (zona de los barriles) y la zona no modular
de la corteza somatosensorial
Para realización de las lesiones, los ratones adultos fueron anestesiados con
ketamina y xilacina (90µg/10µg/g de peso). Los animales se colocaron en un aparato
estereotáxico, después se expuso el cráneo y se les realizó un pequeño agujero de 0.5mm
con un dremmel. Posteriormente se realizó una lesión circunscrita en la zona modular (zona
de los barriles) de la corteza somatosensorial (coordenadas: anteroposterior: 1.5mm;
profundidad: 0.5mm; lateral: 2.3mm relativo a bregma) o en la zona no modular
(coordenadas: anteroposterior: 1.8mm; profundidad: 0.5mm; lateral: 2.3mm relativo a
bregma) utilizando una aguja estéril de 30 G 1/2. Transcurridos dos minutos, se retiró la
aguja, y los animales fueron suturados y colocados en cajas aisladas hasta su recuperación.

Los animales se sacrificaron (ver abajo) a los dos días de realizada la lesión. Se eligió este
periodo debido a que la respuesta inflamatoria en el SNC es de inicio rápido y a las 24-48
horas hay reactividad de la glia. Antes de su sacrificio, los animales fueron anestesiados
con pentobarbital (20 mg/kg de peso corporal), perfundidos con solución salina y
posteriormente con paraformaldehído al 4%. Los animales fueron decapitados, se
obtuvieron los cerebros, se disectó la corteza cerebral lesionada y la contralateral para ser
postfijadas entre dos laminillas y procesadas por métodos de inmunohistoquímica para
determinar la respuesta inmunológica circundante a la lesión. Se realizaron las tinciones de
inmunohistoquímica para determinar la respuesta de astrocitos (antígeno GFAP), de
macrófagos y microglia (antígenos MHCII y F4/80) y la expresión de citocinas
inflamatorias (IFNγ, TNFα) y anti-inflamatorias (IL4, IL10) en el tejido lesionado y
circundante.
Inmunohistoquímica
Ratones normales, lesionados en la zona modular y en la zona no modular se
sacrificaron a los 2 días posteriores a la lesión. Cumplido este periodo, los animales fueron
anestesiados con pentobarbital sódico (20 mg/kg de peso corporal), prefundidos,
decapitados y sus cerebros cuidadosamente removidos para evitar dañar la corteza cerebral.
Se separaron las cortezas de cada cerebro y se colocaron sobre portaobjetos recubiertos con
cinta de teflón para extender las cortezas y facilitar la visualización del mapa de los
barriles. Las muestras se congelaron por inmersión en 2-metilbutano pre-enfriado con hielo
seco. Después de ser congeladas, las muestras se almacenaron a -74° C hasta efectuar los
experimentos morfológicos. Para ello, se realizaron cortes tangenciales seriados (35µm de
espesor) de las cortezas cerebrales a una temperatura de -20° C utilizando un criostato. Los
cortes fueron recuperados y colocados en pozos con amortiguador de fosfatos (PB; 0.1M,
pH 7.4). En seguida se inactivó la actividad de peroxidasa endógena en el tejido con 3%
H202 en PB durante 15 minutos y se lavaron 3 veces por 5 minutos con PB. Los cortes
fueron incubados con una solución de bloqueo que contiene albúmina sérica bovina (ABS;
3%) y tritón X-100 (0.3%) disueltos en PB, en agitación y a temperatura ambiente por 90
minutos. Se agregó el anticuerpo primario rata anti-TNFα de ratón, rata anti-F4/80 de
ratón, rata anti-IL4 de ratón, rata anti-IL10 de ratón (todos de Serotec; a una dilución 1:50),

o rata anti-MHCII de ratón, rata anti-IFNγ de ratón y conejo anti-GFAP (todos de
Chemicon; a una dilución 1:500 excepto anti-GFAP 1:300), diluidos en la solución de
bloqueo. El anticuerpo primario se incubó toda la noche a 4° C en agitación. Como control
de los experimentos se omitió el anticuerpo primario y se incubó sólo con suero de bloqueo.
Al día siguiente los cortes fueron lavados 3 veces por 15 minutos en agitación con una
solución de lavado que contenía Tritón X-100 (0.3%) en PBS. Posteriormente, los cortes se
incubaron por 1 hora 30 minutos a temperatura ambiente en agitación con los anticuerpos
secundarios biotinilados dirigidos contra IgGs de rata (Chemicon, a una dilución 1:500) o
contra IgGs de conejo, todos de Vector, a una dilución 1:500) en correspondencia con los
anticuerpos primarios utilizados. Pasado ese tiempo, se retiró el exceso de anticuerpos y los
cortes se lavaron tres veces con la solución de lavado. Los cortes se incubaron con el
complejo avidina-peroxidasa (Elite AB Kit, Vector Laboratories, Burlingame, CA, USA)
por 1 hora 30 minutos a temperatura ambiente en agitación, se lavaron tres veces con PB, y
la actividad de peroxidasa se reveló con un kit de 3,3- diaminobencidina y peróxido de
hidrógeno de acuerdo al protocolo recomendado por el proveedor (Vector Laboratories).
Los cortes se tiñeron para visualizar los barriles de la corteza somatosensorial con violeta
de cresilo por 25 minutos, se lavó, con agua destilada para posteriormente ser deshidratados
en soluciones crecientes de etanol. Al final del procedimiento, los cortes se montaron sobre
laminillas cubiertas de gelatina y se dejaron secar hasta el día siguiente en ambiente libre de
polvo, se montaron en Cytoseal y se observaron en un microscopio de campo claro o
epifluorescencia (Optiphot-2, Nikon, Tokio, Japan o Axioscop 40, Carl Zeiss, Gottingen,
Germany). Las fotografías fueron tomadas con una cámara digital (Coolpix4300, Nikon o
AxioCam MRc, Carl Zeiss) y convertidas a escala de grises con el programa Adobe
Photoshop®. Estas fotografías fueron utilizadas para mapear la distribución de astrocitos y
células presentadoras de antígenos dentro y fuera de los barriles. Para ello, se estimó su
densidad. Los datos obtenidos fueron contrastados estadísticamente utilizando un t de
Student fijando la significancia estadística a una p<0.05.

Resultados
Distribución y densidad de células microgliales, macrófagos perivasculares y astrocitos
en la corteza somatosensorial primaria de ratones adultos normales
El análisis inmunohistoquímico efectuado en la corteza somatosensorial primaria de
ratones machos CD1 en condiciones fisiológicas, reveló la ausencia de astrocitos (Fig. 1
A), así como del Complejo Mayor de Histocompatibilidad clase II (MHCII de sus siglas en
inglés), una molécula necesaria para la presentación antigénica (Fig. 1 B). Por otra parte,
encontramos una menor densidad de células microgliales y macrófagos perivasculares con
respecto al resto de la corteza cerebral (Fig. 1 C). En la Fig. 1 D se observan algunos
macrófagos perivasculares (ver la cabeza de flecha) y un mayor número de células
microgliales en el septo o borde (ver flechas) con respecto al interior de los barriles.
Presencia de citocinas en la corteza somatosensorial primaria de ratones adultos
normales
En cuanto a la respuesta inmunológica, nosotros estudiamos dos citocinas pro-
inflamatorias (INFγ y TNFα) y dos anti-inflamatorias (IL4 e IL10). Las citocinas pro-
inflamatorias y la IL4 no fueron detectadas en la zona modular de los barriles (Fig. 2 A, B y
C). Sin embargo, llama la atención la presencia de IL10 en el interior de los barriles y no
así en el borde o septo de los mismos (Fig. 2 D). La imagen muestra que esta citocina se
encuentra principalmente en las terminaciones nerviosas de las neuronas (Fig. 2 D, ver
flechas).
Distribución y densidad de células microgliales, macrófagos perivasculares y astrocitos
en la corteza somatosensorial de ratones adultos después de realizada una lesión
circunscrita
Después de realizar una lesión circunscrita de tipo penetrante observamos una
respuesta glial pronunciada tanto en las lesiones realizadas en la zona modular como en
aquellas fuera de la zona modular (Fig. 3 A-F). Es notable la activación de la microglia y
de los macrófagos perivasculares ya que expresan MHCII (Fig. 3 C y D), la cual en
condiciones fisiológicas no está presente (ver Fig. 1 B). Los astrocitos muestran poca

reactividad como se puede observar en la Fig. 3 E y F. Llama la atención que en las
lesiones realizadas dentro de la zona modular o en los barriles, la respuesta glial difiere en
su distribución con respecto a las lesiones realizadas fuera de los barriles. Podemos
observar que solamente los barriles cercanos a la lesión muestran reactividad celular (Fig. 3
A y C). Por otro lado, la respuesta de macrófagos y células microgliales es más intensa en
el septo que en el interior de los barriles (Fig. 3 A y C).
Presencia de citocinas en la corteza somatosensorial de ratones adultos después de
realizada una lesión circunscrita
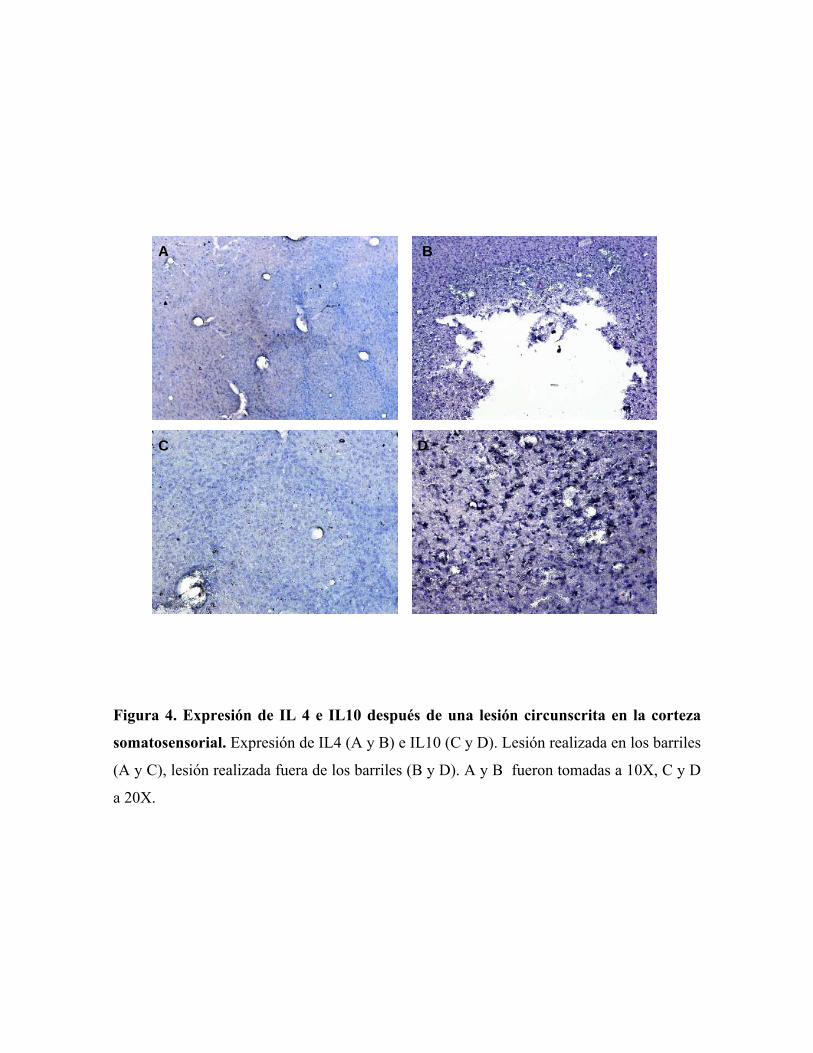
Con respecto a la expresión de las citocinas anti-inflamatorias, observamos un
aumento en la expresión de IL 4 después de realizada la lesión tanto en la zona modular
como no modular (Fig. 4 A y B). Sin embargo, se puede ver que la distribución de IL4 en la
zona modular es más pronunciada en el septo de los barriles con respecto al interior de los
mismos (Fig. 4 A). Por otro lado se observa que la expresión de IL10 es únicamente
detectable en el borde de la lesión fuera de la zona modular (Fig. 4 D) mientras que en los
barriles está ausente (Fig. 4 C).
En contraste con la baja expresión de IL 4 e IL 10, INFγ se encuentra expresado en
gran cantidad tanto en las lesiones dentro de la zona modular como fuera (Fig. 5 A y B).
TNFα muestra un perfil similar aunque menos pronunciado (Fig. 5 C y D). Nuevamente se
puede observar que en las lesiones realizadas dentro de la zona modular la expresión de
ambas citocinas pro-inflamatorias es mayor en el septo de los barriles con respecto al
interior de los mismos (Fig. 5 A y C).
Discusión
Los datos obtenidos en este trabajo apoyan la existencia de módulos inmunológicos
y que éstos están relacionados estructural y funcionalmente con los módulos neurales. En
este proyecto nos enfocamos a estudiar la respuesta inmunológica en una zona modular de
la corteza somatosensorial correspondiente al mapa corporal de las vibrisas faciales,
denominada barriles.
Nuestras observaciones muestran que la corteza somatosensorial tiene una
organización inmunológica particular en condiciones fisiológicas. En primera instancia

notamos que la distribución de células gliales es diferente en el área de los barriles con
respecto a otras zonas de la corteza cerebral, mostrando una menor densidad de microglia y
macrófagos perivasculares en los barriles, y que su distribución es preferentemente en los
septos y escasa en el interior de los mismos. Estos tipos celulares participan tanto en
fenómenos de protección contra patógenos, de inmuno-regulación y de reparación de
tejidos fagocitando restos celulares, como en procesos regenerativos (Chen L et. al. 2002).
Sin embargo, la microglia y los macrófagos son los principales participantes en eventos
inflamatorios que pueden llevar a daño tisular (Chavarria et al., 2004), por lo que su menor
densidad en el área de los barriles pudiera conferir cierta “protección” ante un estímulo
inflamatorio. Por otro lado, la ausencia de la expresión del MHCII en la corteza cerebral
también pudiera ser relevante al contribuir al ambiente inmuno-privilegiado del SNC.
Otro hallazgo relevante es la presencia de IL10 en condiciones fisiológicas
únicamente en el interior de los barriles en las terminaciones nerviosas. La IL10 es una
citocina con propiedades anti-inflamatorias o inmuno-reguladoras. Adicionalmente se le ha
atribuido el papel de neuroprotectora, ya que previene la muerte celular al bloquear la
caspasa 3 (Beachis et al., 2001). La presencia de la IL10 en el interior de los barriles
pudiera conferirle características inmunológicas diferentes que al septo de los mismos.
Las diferencias que hallamos en la corteza somatosensorial con respecto al resto de
la corteza cerebral apoyan la existencia de módulos inmunológicos, estas observaciones se
ven reforzadas por los datos obtenidos al lesionar fuera y dentro de la zona modular.
Observamos que la respuesta de las células gliales difiere, cuando la lesión es realizada
fuera del área modular la distribución glial es alrededor del borde de la lesión y muestra una
respuesta intensa que disminuye en zonas alejadas de la lesión. Esta distribución contrasta
con la que hallamos al lesionar en la zona modular, ya que la respuesta glial es únicamente
en los barriles pegados a la lesión y es más intensa en los septos con respecto al interior de
los barriles. Se sabe que los astrocitos tienen un papel importante en la modulación de la
respuesta inmunológica en el SNC (Aschner, 1998), sin embargo, no observamos una
respuesta de astrocitos en las lesiones tanto en la zona modular como en la zona no
modular; esto pudiera deberse a que los astrocitos requieren más tiempo para su activación
que las células microgliales (Cornet. et.al, 2000 ) y que los animales de experimentación
fueron sacrificados a los dos días después de realizada la lesión. La respuesta celular más

marcada que encontramos es principalmente de la microglia y los macrófagos. La microglia
muestra un perfil de activación no sólo por morfología sino también al encontrar la
expresión de MHCII tanto en el borde de la lesión como en zonas alejadas. En el caso de
las lesiones de la zona modular encontramos la expresión del MHCII en concordancia con
la distribución de las células microgliales y macrófagos ya mencionada. Estas
características descritas pudieran conferirle a la zona modular una respuesta más delimitada
y controlada con respecto al resto de la corteza cerebral.
La presencia de citocinas anti-inflamatorias fue muy limitada tanto en lesiones en el
área modular como fuera. Cabe destacar que la presencia de IL10 en el interior de los
barriles, que se observó en condiciones fisiológicas, desaparece totalmente después de la
lesión, es probable que esta citocina haya sido utilizada para regular el proceso
inflamatorio, y de esta manera delimitar el daño así como prevenir la muerte neuronal
(Beachis et al., 2001). Mientras que en las lesiones realizadas fuera de los barriles
observamos la presencia de IL10 e IL4 en células con morfología de macrófagos.
Previamente se ha reportado que los macrófagos al fagocitar mielina producen moléculas
anti-inflamatorias (Bavone et al., 2006). Dado que en el borde de la lesión hay muerte
celular incluso de oligodendrocitos, es posible que los macrófagos activados puedan
fagocitar restos de mielina, dando como resultado la producción de IL10 e IL4. Este
fenómeno no ocurre con la misma intensidad en el área de los barriles, lo que siguiere que
el mecanismo de inmuno-regulación pudiera ser diferente.
En contraste con la respuesta anti-inflamatoria, la respuesta de citocinas pro-
inflamatorias es considerablemente más intensa, especialmente la de INFγ. La presencia de
esta citocina es muy marcada en el borde de las lesiones y su periferia de las zonas no
modulares y contrasta con la intensidad y distribución de IFNγ en los barriles, donde se
expresa nuevamente más en los septos con respecto al interior de los barriles.
Adicionalmente se observa que el área de expresión de INFγ es menor en los cerebros
lesionados que en el área modular. INFγ puede ser neurotóxico (Lambertsen et al., 2004),
pudiera ser que la baja expresión de INFγ en la zona modular limite la extensión del daño.
Dado que observamos que la expresión de IFNγ es menor en el interior de los barriles con
respecto a los septos, es posible que la IL10 haya regulado o incluso suprimido en esta área
la respuesta inflamatoria de IFNγ. Así mismo, llama la atención la expresión de IFNγ en la

microglia activada y en macrófagos de las zonas no modulares. Por otro lado, la expresión
de TNFα en las lesiones tanto en la zona modular como no modular es menor con respecto
a INFγ. Llama la atención que muestra una distribución similar a la respuesta de IFNγ, es
decir, en la zona modular es más pronunciada en los septos que en el interior de los barriles.
TNFα es una molécula pleiotrópica que tiene efectos tanto pro-inflamatorios como efecto
neuroprotector al inducir neuroplasticidad cuando está presente en cantidades pequeñas
(William C et al., 2000). Es posible que la baja expresión de esta citocina en el área
modular pudiera estar relacionada más con el efecto neuroprotector que inflamatorio.
Conclusión
Los datos obtenidos en este proyecto apoyan la existencia de módulos
inmunológicos en el SNC y que estos compartimentos están relacionados estructural y
funcionalmente con los módulos neurales. Los módulos inmunológicos pueden conferirle a
los módulos neurales características inmunológicas diferentes que pudieran estar
relacionados con eventos de mayor o menor inflamación y por lo tanto con mayor o menor
daño tisular y la pérdida o conservación de la función. Nuestras observaciones abren nuevas
perspectivas en el estudio y la comprensión de la fisiología y patología del SNC.

A B
C D
Fig. 1 Distribución de las células gliales en la zona modular de los barriles en ratones
normales. Expresión de GFAP para ver astrocitos (A), del MHCII (B) y de F4/80 (C, D)
para ver macrófagos (ver la cabeza de flecha) y microglia (ver las flechas). A, B y D fueron
tomadas a 20X, C a 4X.

B
A
C D
Fig. 2 Presencia de IL 10 en el interior de barriles. Expresión de INFγ (A), TNFα (B),
IL4 (C) e IL10 (D). Las flechas muestran terminaciones nerviosas dentro de los barriles. A,
B y C fueron tomadas a 20X, D a 40X.

A B
C D
E F
Figura 3. Distribución de células gliales después de una lesión circunscrita en la
corteza somatosensorial. Expresión del marcador F4/80 de microglia y macrófagos (A y
B), del MHCII (C y D) y de GFAP en astrocitos (E y F). Lesión realizada en los barriles
(A, C y E), lesión realizada fuera de los barriles (B, D y F). Las fotos fueron tomadas a
10X( A, B, C, D y F) y 4X( E )
A B
C D
Figura 4. Expresión de IL 4 e IL10 después de una lesión circunscrita en la corteza
somatosensorial. Expresión de IL4 (A y B) e IL10 (C y D). Lesión realizada en los barriles
(A y C), lesión realizada fuera de los barriles (B y D). A y B fueron tomadas a 10X, C y D
a 20X.

A B
C D
Figura 5. Expresión de INFγ y TNFα después de una lesión circunscrita en la corteza
somatosensorial. Expresión de INFγ (A y B) y TNFα (C y D). Lesión realizada en los
barriles (A y C), lesión realizada fuera de los barriles (B y D). Las fotos fueron tomadas a
4X.

Referencias
Bachis A, Colangelo AM, Vicini S, Doe PP, De Bernardi MA, Brooker G, Mocchetti I.
Interleukin-10 prevents glutamate-mediated cerebellar granule cell death by blocking
caspase-3-like activity.J Neurosci. 2001 May 1;21(9):3104-12.
Boven LA, Van Meurs M, Van Zwam M, Wierenga-Wolf A, Hintzen RQ, Boot RG, Aerts
JM, Amor S, Nieuwenhuis EE, Laman JD. Myelin-laden macrophages are anti-
inflammatory, consistent with foam cells in multiple sclerosis. Brain. 2006 Feb;129(Pt
2):517-26.
Chavarria-Krauser A, Schurr U. Is damage in central nervous system due to inflammation?
Autoimmun Rev. 2004 Jun;3(4):251-60.
Chavarria A, Fleury A, Garcia E, Marquez C, Fragoso G, Sciutto E. Relationship between
the clinical heterogeneity of neurocysticercosis and the immune-inflammatory profiles. Clin
Immunol. 2005;116(3):271-8.
Chen L, Yang P, Kijlstra A. Distribution, markers, and functions of retinal microglia.
Ocul Immunol Inflamm. 2002 Mar;10(1):27-39.
Anne Cornet , Estelle Bettelli , Mohamed Oukka , Christophe Cambouris , Virginia
Avellana-Adalid , Kostas Kosmatopoulos , Roland S. Liblau. Role of astrocytes in antigen
presentation and naive T-cell activation. Journal of Neuroimmunology 106: 69–77.
Eskandari F, Sternberg EM. Neural-immune interactions in health and disease. Ann N Y
Acad Sci. 2002;966:20-7.
Galea I, Palin K, Newman TA, Van Rooijen N, Perry VH, Boche1 D. Mannose Receptor
Expresión Specifically Reveals Perivascular Macrophages in Normal, Injured, and
Diseased Mouse Brain. Glia 2005; 49:375–384.
Guillemin GJ, Brew BJ. Microglia, macrophages, perivascular macrophages, and pericytes:
a review of function and identification. J. Leukoc. Biol. 75: 388–397; 2004.
Leise E. Modular construction of the nervous system: a basic principle of design for
invertebrates and vertebrates. Brain Res Reviews 1990, 15: 1-23.
Lambertsen KL, Gregersen R, Meldgaard M, Clausen BH, Heibol EK, Ladeby R, Knudsen
J, Frandsen A, Owens T, Finsen B. A role for interferon-gamma in focal cerebral ischemia
in mice.J Neuropathol Exp Neurol. 2004 Sep;63(9):942-55.

Livingstone MS, HubelDH. Anatomy and Physiologyof a color system in the primate
visual cortex. J Neurosci 1984, 4: 2830- 2835.
Killackey HP, Belford GR. The formation of afferent patterns in the somatosensory cortex
of the neonatal rat. J Comp Neurol. 1979;183(2):285-303.
McMenamin PG, Wealthall RJ, Deverall M, Cooper SJ, Griffin B. Macrophages and
dendritic cells in the rat meninges and choroid plexus: three-dimensional localisation by
environmental scanning electron microscopy and confocal microscopy. Cell Tissue Res.
2003;313(3):259-69.
Matyszak MK, Perry VH. The potential role of dendritic cells in immune-mediated
inflammatory diseases in the central nervous system. Neuroscience 1996. 74: 599–608.
Meredith EJ, Chamba A, Holder MJ, Barnes NM, Gordon J. Close encounters of the
monoamine kind: immune cells betray their nervous disposition. Immunology. 2005;
115(3):289-95.
Michel Aschner. Inmune and inflammatory responses in the CNS; modulation by
astrocytes. Toxicology Letters 102-103 (1998) 283-287
Mountcastle V. An organizing principle for cerebral function: the unit module and
distributed system. In Smitt FO, Worden FG (Editors), The neurosciences forth study
program, MIT, Cambridge, MA, 1979: 21-42.
Sharafeldin A, Hamadien M, Diab A, Li HL, Shi FD, Bakhiet M. Cytokine Profiles in the
Central Nervous System and the Spleen During the Early Course of Experimental African
Trypanosomiasis. Scand. J. Immunol. 1999; 50: 256–261,
Vass K, Lassmann H. Intrathecal application of interferon gamma. Progressive appearance
of MHC antigens within the rat nervous system. Am J Pathol. 1990;137(4):789-800.
Vegeto E, Ciana P, Maggi A. Estrogen and inflammation: hormone generous action spreads
to the brain. Mol Psychiatry. 2002;7(3):236-8.
Weigent DA, Blalock JE. Associations between the neuroendocrine and immune systems. J
Leukoc Biol. 1995;58(2):137-50.
White E, Peters A. Cortical modules in the Posteromedial barrel subfield (Sml) of the
mouse. J Comp Neurology 1993, 334: 86-96.
William C. Covey, Tracey A. Ignatowski , Paul R. Knight, Robert N. Spengler. Brain-
derived TNFa: involvement in neuroplastic changes implicated in the

conscious perception of persistent pain. Brain Research 859 2000 113–12.
Woolsey TA, Van der Loos H. The structural organization of layer IV in the somatosensory
region (SI) of mouse cerebral cortex. The description of a cortical field composed of
discrete cytoarchitectonic units. Brain Res. 1970;17(2):205-42.