molecular basis of abnormal conduction in … · molecular basis of abnormal conduction in mice...
TRANSCRIPT

MOLECULAR BASIS OF ABNORMAL CONDUCTION IN MICE OVER-EXPRESSING
ENDOTHELIN-1
by
Erin Elizabeth Mueller
A thesis submitted in conformity with the requirements for the degree of PhD
Graduate Department of Laboratory Medicine and Pathobiology
Copyright by Erin Elizabeth Mueller (2011)

ii
MOLECULAR BASIS OF ABNORMAL CONDUCTION IN MICE
OVER-EXPRESSING ENDOTHELIN-1
Erin Elizabeth Mueller
Doctor of Philosophy, 2011
Department of Laboratory Medicine & Pathobiology, University of Toronto
ABSTRACT
Binary transgenic (BT) mice with doxycycline (DOX)-suppressible cardiac-specific over-
expression of endothelin-1 (ET-1) exhibit progressive heart failure, QRS prolongation, and
death following DOX withdrawal. However, the molecular basis and reversibility of the
electrophysiological abnormalities in this model were not known. Here we assess the
mechanisms underlying ET-1-mediated electrical remodelling, and its role in heart failure.
Prior attempts to prevent this model of ET-1 induced cardiomyopathy with ET receptor
antagonism were not beneficial. We now propose to evaluate the effectiveness of blocking
the synthesis of ET-1 with CGS 26303, a dual inhibitor of endothelin converting enzyme
(ECE) and neutral endopeptidase.
BT vs. littermate control mice were withdrawn from DOX and serially studied with
ultrasound biomicroscopy, octapolar catheters, multi-electrode epicardial mapping,
histopathology, Western blot, immunohistochemistry and qRT-PCR. Prolonged ventricular
activation and depressed rate of ventricular activation were detected as early as 4 wks after
transgene activation, when structure and function of the heart remained unaffected. By 8 wks
of ET-1 over-expression, biventricular systolic and diastolic dysfunction, myocardial fibrosis,
cardiomyocyte hypertrophy, prolonged ventricular activation and repolarization, depressed

iii
rate of ventricular activation, and abnormal atrioventricular nodal function were observed.
Within 4 wks of ET-1 induction, reduction were observed in connexin-43 mRNA, protein,
and phosphorylation, Nav1.5 mRNA and protein, Na+ conductance, K+ channel interacting
protein-2 mRNA and Kv4.2 mRNA. Chromatin immunoprecipitation revealed that nuclear
factor κB preferentially binds to Cx43 and Nav1.5 promoters. Importantly, the associated
electrophysiological abnormalities at this time point were reversible upon suppression of
ET-1 over-expression and completely prevented the development of structural and functional
remodelling. Treatment with CGS-26303 (5 mg/kg/day) failed to improve survival, or
hemodynamic and contractile decline.
ET-1-mediated ventricular conduction delays correlates with gap junction and ion channel
remodelling, and precedes heart failure. The sequence and reversibility of this phenotype
suggest that a primary abnormality in electrical remodelling may contribute to the
pathogenesis of heart failure. CGS 26303 failed to prevent this cardiomyopathic phenotype.
These data suggest that chronically high levels of bigET-1, as seen in heart failure, may
induce increased ECE activity and/or non-ECE ET-1 synthesis, thus circumventing the
efficacy of ECE blockade in this model.

iv
ACKNOWLEDGEMENTS
Firstly, I would like to thank my supervisors, Mansoor Husain and Duncan Stewart
for providing insight, focus, motivation, contined support, and positive reinforcement
throughout my PhD. Secondly, I would like to thank the Department of Laboratory Medicine
and Pathobiology and CLAMPS for providing a great learning environment. I would also like
to thank my committee members, Peter Backx and Kumaraswamy Nanthakumar for serving
as mentors, and guiding my research progress. Thank you for taking an active and
enthusiastic interest in my project and for providing invaluable insights, helpful discussions,
and electrophysiological expertise. In particular, thank you to Stéphane Massé from
Nanthakumar’s lab, for his continued assistance with electrophysiological experiments.
Additionally, I would like to thank my family and friends, particularly my husband,
for their continued support, encouragement, and patience throughout my graduate studies.
Thank you to Peter Sabatini, Karolina Kolodziejska, Sonya Hui, Shivalika Handa, Jae Choi,
Kiwon Ban, Dorota Dajnowiec, and Dan Trcka for your support, camaraderie, and all the
wonderful memories over the last 7 years.
I would like to thank all past and present members of the Husain lab, in particular, the
surgical skills of Abdul Momen and Golam Kabir, mouse colony management and
genotyping support of Haiyan Xiao and Changsen Wang, the cell culture and qRT-PCR
expertise of Karolina Kolodziejska, general lab advice from Talat Afroze, primer design
assistance and office antics of Omar El-Mounayri, the guidance and reliable advice of Hassan
Zaidi, and the superb everyday support and administrative skills of Tracey Richards.

v
I would also like to thank my MSc supervisor, Susan Howlett for instilling me with a
love of science. And finally, I would like to thank the Ontario Graduate Scholarship in
Science and Technology for funding throughout my PhD program.

vi
TABLE OF CONTENTS
ABSTRACT ............................................................................................................................ II
ACKNOWLEDGEMENTS ................................................................................................. IV
TABLE OF CONTENTS ..................................................................................................... VI
LIST OF TABLES ................................................................................................................ XI
LIST OF FIGURES ............................................................................................................ XII
LIST OF APPENDICES ................................................................................................... XIV
LIST OF ABBREVIATIONS ............................................................................................. XV
CHAPTER 1. LITERATURE REVIEW .............................................................................. 1
1.1.1 Definition ........................................................................................................................ 2
1.1.2 Etiology and prevalence ................................................................................................ 2
1.1.3 Symptoms and classifications........................................................................................ 3
1.1.4 Pathophysiology ............................................................................................................. 4 1.1.4.1 Neurohormonal activation ........................................................................................ 5 1.1.4.1 LV remodelling ......................................................................................................... 7
1.1.5 Treatments ...................................................................................................................... 9
1.2 ENDOTHELIN ............................................................................................................... 10
1.2.1 Distribution, regulation and synthesis of ET-1 ......................................................... 10
1.2.2 Clearance of ET-1 ........................................................................................................ 11
1.2.3 ECE ............................................................................................................................... 13
1.2.4 ET Receptors and signal transduction ....................................................................... 13
1.2.5 Transgenic mouse models: genetic manipulation of ET-1 system ........................... 16
1.2.6. Role of ET-1 in inflammation, hypertrophy, and fibrosis ....................................... 19

vii
1.2.7 Pathophysiology of ET-1 in HF .................................................................................. 23
1.3 ELECTRICAL REMODELLING ................................................................................ 25
1.3.1 Excitation in the healthy heart .................................................................................... 25 1.3.1.1 Ionic basis of cardiac action potential ..................................................................... 26 1.3.1.2 Electrophysiological mapping ................................................................................ 26 1.3.1.3 Excitation-contraction coupling and Ca2+ cycling .................................................. 28 1.3.1.4 Ca2+ handling proteins and Ca2+ current ................................................................. 28 1.3.1.5 Sodium current ........................................................................................................ 32 1.3.1.6 Transient outward K+ current (Ito) .......................................................................... 35 1.3.1.7 Delayed rectifier K+ current (IK) ............................................................................. 36 1.3.1.8 Inward rectifier K+ current (IK1) .............................................................................. 36 1.3.1.9 Gap junctions .......................................................................................................... 37
1.3.2 Electrical remodelling and HF .................................................................................... 40 1.3.2.1 Ca+ channel remodelling ......................................................................................... 41 1.3.2.2 Na+ channel remodelling ......................................................................................... 45 1.3.2.3 K+ channel remodelling .......................................................................................... 45 1.3.2.4 Gap junction remodelling ....................................................................................... 46
1.3.3 Electrical remodelling and ET-1................................................................................. 46 1.3.3.1 Regulation of Ca2+ handling by ET-1 .................................................................... 46 1.3.3.2 Regulation of cardiac repolarization by ET-1 ......................................................... 48 1.3.3.3 Regulation of cardiac conduction by ET-1 ............................................................. 48
1.4 RATIONALE, HYPOTHESIS, OBJECTIVES ........................................................... 49
1.4.1 Rationale ....................................................................................................................... 49
1.4.2 General hypothesis ....................................................................................................... 50
1.4.3 Hypotheses .................................................................................................................... 50
1.4.4 Objectives...................................................................................................................... 50
CHAPTER 2. PHARMACOLOGICAL FAILURE OF LONG-TERM DUAL ECE-NEP INHIBITION WITH CGS-26303 IN AN ET-1 MODEL OF CARDIOMYOPATHY......................................................................................................... 52
2.1 INTRODUCTION........................................................................................................... 53
2.2 MATERIALS & METHODS......................................................................................... 55
2.2.1 Experimental animals .................................................................................................. 55

viii
2.2.2 Drug administration .................................................................................................... 55
2.2.3 Invasive LV hemodynamics ........................................................................................ 57
2.2.4 ET-1 / BigET-1 ELISA ................................................................................................ 57
2.2.5 ANP ELISA .................................................................................................................. 58
2.2.6 Histopathology.............................................................................................................. 58
2.2.7 ECE activity .................................................................................................................. 59
2.3 RESULTS ........................................................................................................................ 59
2.3.1 Short term treatment with CGS-26303 inhibits ECE and NEP activity................. 59
2.3.2 Long term treatment with CGS-26303 fails to prevent cardiomyopathic phenotype................................................................................................................................................. 61
2.4 DISCUSSION .................................................................................................................. 66
CHAPTER 3. ET-1 INDUCED ELECTRICAL REMODELLING PRECEDES LV DYSFUNCTION IN ET-1 INDUCED CARDIOMYOPATHY ....................................... 70
3.1 INTRODUCTION........................................................................................................... 71
3.2 MATERIALS & METHODS......................................................................................... 72
3.2.1 Experimental animal ................................................................................................... 72
3.2.2 Surface ECG and intracardiac electrophysiological evaluation .............................. 72
3.2.3 Epicardial mapping ..................................................................................................... 74
3.2.4 Invasive LV hemodynamics ........................................................................................ 76
3.2.5 Ultrasound biomicroscopy .......................................................................................... 78
3.2.6 Histopathology.............................................................................................................. 78
3.2.7 Statistical analysis ........................................................................................................ 78
3.3 RESULTS ........................................................................................................................ 79
3.3.1 Electrical defects in mice with cardiac-specific ET-1 over-expression ................... 79
3.3.2 Electrical remodelling is triggered as early as 4 wks after ET-1 over-expression . 79

ix
3.3.3 HF develops by 8 weeks after ET-1 over-expression ................................................ 80
3.3.4 Inhibiting ET-1 expression at the onset of electrical remodelling prevents progression to HF .................................................................................................................. 84
3.4 DISCUSSION .................................................................................................................. 86
CHAPTER 4. REDUCED CONNEXIN-43 AND SODIUM CHANNEL NAV1.5 IS ASSOCIATED WITH ET-1 INDUCED ELECTRICAL UNCOUPLING ..................... 89
4.1 INTRODUCTION........................................................................................................... 90
4.2 MATERIALS & METHODS......................................................................................... 91
4.2.1 Experimental animals .................................................................................................. 91
4.2.2 RNA isolation & quantitative real-time RT-PCR analysis ...................................... 91
4.2.3 Western blotting ........................................................................................................... 91
4.2.4 Immunohistochemistry ................................................................................................ 95
4.2.5 HL-1 cell culture .......................................................................................................... 96
4.2.6 Optical mapping ........................................................................................................... 96
4.2.7 Isolation of NMVM ...................................................................................................... 97
4.2.8 Promoter analysis......................................................................................................... 97
4.2.9 ChIP .............................................................................................................................. 99
4.2.10 Isolation of adult mouse ventricular myocytes ...................................................... 100
4.2.11 Patch clamp recordings ........................................................................................... 100
4.2.12 Statistical analysis .................................................................................................... 101
4.3 RESULTS ...................................................................................................................... 101
4.3.1 ET-1 mediated electrical remodelling correlates with reduced Cx43, p-Cx43, Cx40, Nav1.5, and Na+ channel conductance ............................................................................... 101
4.3.2 In vitro validation of ET-1 induced electrical remodelling .................................... 108
4.3.3 ET-1 induced reductions in Cx43 and Nav1.5 may be induced by NFκB ............. 108

x
4.4 DISCUSSION ................................................................................................................ 113
CHAPTER 5. SUMMARY AND FUTURE DIRECTIONS ........................................... 120
5.1 SUMMARY AND CONCLUSIONS ........................................................................... 121
5.2 FUTURE DIRECTIONS .............................................................................................. 122
5.2.1 ET-1 and atrial electrical remodelling ..................................................................... 122
5.2.2 ET-1 and K+/Ca2+ channel remodelling ................................................................... 123
5.2.3 Role of NFκB p50 in ET-1 induced gap junction/ion channel remodelling .......... 124
5.2.4 Polymorphisms in ET-1 signaling components ....................................................... 124
REFERENCES .................................................................................................................... 125
APPENDICES ..................................................................................................................... 148

xi
LIST OF TABLES
Table 1.1. Remodelling of ion channels, connexins, and Ca2+ handling proteins in the failing ventricle................................................................................................................................... 43 Table 3.1. Temporal progression and prevention of electrical remodelling in mice over-expressing ET-1 during pacing ............................................................................................... 81 Table 3.2. Progression and prevention of cardiac structural and functional abnormalities as evaluated by invasive hemodynamics ..................................................................................... 82 Table 3.3. Progression and prevention of cardiac dysfunction as evaluated by ultrasound biomicroscopy ......................................................................................................................... 83 Table 4.1. Real-time PCR Primer Sequences ......................................................................... 94 Table 4.2. Real-time PCR Primer Sequences-promoters for ChIP for NFkB binding sites ... 98 Table 4.3. Progression and prevention of molecular remodelling as evaluated by LV mRNA expression levels ................................................................................................................... 102

xii
LIST OF FIGURES
Figure 1.1 Production and degradation of ET-1 ..................................................................... 12 Figure 1.2. ET-1 mediated signaling via Gαq/s/i ....................................................................... 14 Figure 1.3. DOX-off system of cardiac over-expression of ET-1 .......................................... 18 Figure 1.4. Multifaceted nature of ET-1 signaling ................................................................. 20 Figure 1.5. Illustration of NFκB activation ............................................................................. 22 Figure 1.6. Schematic of the cardiac conduction system, the ionic currents contributing to the cardiac action potential, and the surface electrocardiogram ................................................... 27 Figure 1.7. Ca2+ cycling in healthy cardiac myocytes ........................................................... 29 Figure 1.8. Regional connexin expression in the heart ........................................................... 38 Figure 1.9. Electrical remodelling of cardiac myocytes in the failing heart ........................... 42 Figure 1.10. ET-1 induced electrical remodelling in cardiac myocytes ................................. 47 Figure 2.1. Schematic of ET-1 synthesis, degradation, and treatment with CGS 26303 ........ 54 Figure 2.2. Schematic of experimental design ........................................................................ 56 Figure 2.3. Ex vivo validation of CGS 26303 ........................................................................ 60 Figure 2.4. Short term treatment with CGS-26303 inhibited ECE and NEP activity ............. 62 Figure 2.5. Long term treatment with CGS-26303 fails to preserve hemodynamic and LV contractile integrity in BT mice .............................................................................................. 63 Figure 2.6. Long term treatment with CGS-26303 fails to inhibit ECE and NEP activity ..... 64 Figure 2.7. Therapy with CGS-26303 does not improve survival in BT mice ....................... 65 Figure 2.8. Model depicting short vs. long term treatment with dual ECE-NEP inhibitor .... 69 Figure 3.1. Schematic of experimental design ........................................................................ 73 Figure 3.2. Electrophysiological evaluation using intracardiac mapping reveals electrical defects in mice over-expressing ET-1 after 8-10 weeks of transgene induction .................... 75 Figure 3.3. Temporal progression and prevention of electrical remodeling in mice over-expressing ET-1 ...................................................................................................................... 77 Figure 3.4. Myocardial fibrosis appears after 8 wks of ET-1 over-expression ....................... 85 Figure 3.5. Model showing effects of 4 vs. 8 wks of ET-1 induction .................................... 87 Figure 4.1. Schematic of experimental designs ...................................................................... 92 Figure 4.2. Reduced LV Cx43 and Nav1.5 expression in mice as early as 4 wks after ET-1 induction ............................................................................................................................... 103 Figure 4.3. Prolonged ET-1 over-expression leads to progressive loss and lateralization of Cx43 ...................................................................................................................................... 105 Figure 4.4. Prolonged ET-1 over-expression lead to progressive loss and lateralization of p-Cx43 ...................................................................................................................................... 106 Figure 4.5. Na+ channel conductance reduced in LV myocytes isolated from mice 4wks post DOX withdrawal ................................................................................................................... 107 Figure 4.6. ET-1 treatment had no effect on Cx43 mRNA, protein, or p-Cx43/ Total Cx43 protein expression in HL-1 cells ........................................................................................... 109 Figure 4.7. ET-1 treatment had no effect on conduction velocity in HL-1 cells .................. 110 Figure 4.8. Four weeks of ET-1 transgene induction had no affect levels of left atrial ....... 111 Cx43 or Nav1.5 mRNA expression ...................................................................................... 111 Figure 4.9. ET-1 reduces Cx43 levels in NMVM ................................................................. 112 Figure 4.10. NFκB p50 preferentially binds to sites within the Cx43 and Nav1.5 promoters in the LV of ET-1 over-expressing mice .................................................................................. 114

xiii
Figure 4.11. Model illustrating mechanim(s) of ET-1 induced reductions of Cx43 and Nav1.5 after 4 wks of ET-1 over-expression ..................................................................................... 116

xiv
LIST OF APPENDICES
Appendix 1. Myocardial ET- levels after short vs. long term CGS 26303 treatment ........... 148 Appendix 2. Reduced molecular determinants of Ito (Kv4.2 and KChIP2 mRNA expression) after 4 wks of ET-1 over-expression ..................................................................................... 149 Appendix 3. Cx43 core promoter sequence .......................................................................... 150 Appendix 4. Nav1.5 core promoter sequences ...................................................................... 151 Appendix 5. NFκB p50 preferentially binds to sites within the NFκBIB promoter in the LV of ET-1 over-expressing mice ............................................................................................... 152 Appendix 6. NFκB p50 preferentially binds to a site within the Tbx2 promoter in the LV of ET-1 over-expressing mice ................................................................................................... 153 Appendix 7. NFκB p50 does not preferentially bind to a site within the Tbx3 promoter in the LV of ET-1 over-expressing mice ........................................................................................ 154 Appendix 8. NFκB p50 does not preferentially binds to sites within the Nkx2.5 promoter in the LV of ET-1 over-expressing mice .................................................................................. 155

xv
LIST OF ABBREVIATIONS
ACE angiotensin converting enzyme
AH atrial-His
Ang II angiotensin II
ANP atrial natriuretic protein
AP action potential
APD action potential duration
AV2:1 2:1 AV block cycle lengths
AVERP AV effective refractory periods
AVWENK AV Wenckebach
AVN atrioventricular node
bigET-1 big Endothelin-1
BNP brain natriuretic peptide
BT binary transgenic
Cav1.2 α1C-subunit of the L-type Ca2+ channel
CN calcineurin
CV conduction velocity
Cx connexin
DBP diastolic blood pressure
DOX doxycycline
dP/dt maximum positive and negative rate of LV pressure change
-dV/dt maximum negative rate of voltage change
E/A ratio of peak early to atrial diastolic inflow velocities
ECE endothelin converting enzyme
ET-1 endothelin-1
FS fractional shortening
hET-1 human ET-1
HF heart failure
HR heart rate
HV His-ventricular

xvi
HW/BW heart weight body weight ratio
ICa-L L-type Ca2+ current
ICa-T T-type Ca2+ current
IKr rapid delayed rectifier K+ current
IKs slow delayed rectifier K+ current
IK1 inward rectifier K+ current
Itos slow recovering transient outward K+ current
Itof fast recovering transient outward K+ current
IκB inhibitor of κB
IL-6 interleukin-6
IP3R inositol triphosphate receptor
KChIP2 K+ channel interacting protein 2
LQT long QT syndrome
LV left ventricular
LVSP left ventricular systolic pressure
MHC myosin heavy chain
MiRP minK-related peptides
MMP matrix metalloproteinase
Nav1.5 α-subunit of the cardiac Na+ channel
NBT non-binary transgenic
NCX Na+/Ca2+ exchanger
NE Norepinephrine
NEP neutral endopeptidase
NFκB nuclear factor κB
NHE Na+/H+ exchanger
NMVM neonatal mouse ventricular myocytes
NRVM neonatal rat ventricular myocytes
p-Cx43 phosphorylated Cx43
PKC protein kinase C
RAAS renin angiotensin aldosterone system
RV right ventricular

xvii
RyR ryanodine receptor
PLB phospholamban
SAN sinoatrial node
SBP arterial systolic blood pressure
SERCA2a sarcoplasmic reticulum Ca2+-ATPase
SR sarcoplasmic reticulum
TNFα tumor necrosis factor α
tTA tetracycyline-transactivator
VT ventricular tachycardia
VF ventricular fibrillation

1
CHAPTER 1. LITERATURE REVIEW

2
1.1 HEART FAILURE
1.1.1 Definition
Heart failure (HF) is a syndrome where the heart does not pump adequately to meet
the metabolic demands of the body. HF is typically a chronic condition that progresses slowly
over time. However, rapid-onset (acute) HF may develop suddenly from a
structural/functional insult or as a consequence of abrupt worsening of chronic HF. HF can
result from systolic or diastolic dysfunction of either or both sides of the heart. Common risk
factors for the development of HF include hypertension, myocardial infarction, coronary
artery disease, damaged heart valves, kidney conditions, congenital heart defects, diabetes,
arrhythmia, viral infection, anaemia, hyperthyroidism, age, obesity, a sedentary lifestyle, a
family history of cardiomyopathy, stress, sleep apnea, smoking, and alcohol/drug abuse.
1.1.2 Etiology and prevalence
Cardiovascular disease is the leading cause of death in Canada, responsible for 30% of
all deaths1 and affecting 1.3 million Canadians (5% of population)2. In Canada, heart disease
and strokes claim a life every 7 minutes and costs the economy more than $22.2 billion every
year2. In particular, HF currently affects ~500,000 Canadians with an additional 50,000 new
cases diagnosed annually3, with a 50% five-year survival rate4.
Coronary artery disease and hypertension are the main causes of HF. Valve disease,
cardiomyopathy, as well as infiltrative, infective, genetic, endocrine, and nutritional
conditions can also trigger HF. Valve disease can result from congenital defects, infections,
and coronary artery disease. Hypertrophic/dilated/restrictive/idiopathic cardiomyopathy can
be induced by toxins (alcohol, cocaine, chemotherapy drugs), tachyarrythmias, infections and

3
inflammation (Chagas disease, myocarditis, pericarditis, endocarditis, sepsis), and genetics
(mutations in genes encoding sarcomeric/cytoskeletal/Ca2+ handling proteins). Infiltrative
disorders ensue from amyloidosis, sarcoidosis, hemochromatosis, and connective tissue
disease. Endocrine disorders are attributed with diabetes, hypo- and hyper-thyroidism, and
Cushing’s syndrome. Nutritional conditions result from cachexia, obesity, and insufficient
thiamine, selenium, and carnitine. Conditions that prompt high cardiac output such as anemia,
arteriovenous fistula, thyroid disease, beriberi, and pregnancy can also cause HF.
1.1.3 Symptoms and classifications
Common HF symptoms include shortness of breath, fatigue, fluid retention, dizziness,
rapid or irregular heart beat as a result of reduced perfusion of various organs and
compensatory mechanisms. Left sided HF primarily affects the pulmonary system, where fluid
accumulation in the lungs causes shortness of breath (particularly when in a horizontal
position), wheezing, and coughing. Right sided HF can occur alone or as a consequence of left
sided HF; it affects systemic fluid imbalance as fluid accumulates in the veins, resulting in
peripheral edema, ascites, hepatomegaly, nausea, and weight gain.
There are presently two commonly recognized HF classification systems. The New
York Heart Association classification scheme for HF is based on the ease of executing
physical activity. Class I HF patients do not develop symptoms from physical activity. Class II
HF patients have no symptoms at rest, although regular physical activity is mildly limited by
shortness of breath, fatigue, and palpitations. Class III HF patients also have no symptoms at
rest, however, ordinary exercise is moderately limited by shortness of breath, fatigue, and
palpitations. Class IV HF patients exhibit symptoms at rest and with any physical activity.

4
The American College of Cardiology and American Heart Association established a
second classification scheme for HF which integrates the progression and development of the
disease. The first two stages (Stage A, B) encompass those patients at risk for developing HF,
and the last two stages (Stage C, D) include patients that exhibit moderate/severe HF
symptoms1. Stage A designated patients lack both symptoms of HF and structural heart
disease, but are at a high risk for developing HF as a consequence of coronary artery disease,
hypertension, or diabetes. Patients in Stage B have no signs or symptoms of HF, despite
structural heart disease as a result of a previous myocardial infarction, left ventricular
hypertrophy, or asymptomatic valve disease. Stage C patients have structural heart disease
with symptoms of HF such as fatigue, shortness of breath, and exercise intolerance. Stage D
patients have severe end-stage HF; these patients are resistant to current pharmacological
interventions and cannot perform day-to-day activities, display severe fatigue and are
vulnerable to repeated and/or prolonged hospitalizations.
1.1.4 Pathophysiology
HF has been described as a cardiorenal model, as severe renal Na+ and water retention
is caused by cardiac dysfunction. A hemodynamic model has also been proposed to account
for the changes in blood pressure/flow in the vasculature and myocardium that occur in HF.
An alternative neurohormonal model has been described to reflect the involvement of the
neurological and hormonal systems in the progression of HF. A biomolecular model of HF
combines the neurohormonal models with the key molecular and cellular changes that mediate
left ventricular (LV) remodelling in the progression of HF.

5
1.1.4.1 Neurohormonal activation
Initially, HF is characterized by impaired cardiac output due to acute or chronic
myocardial insult. Several compensatory mechanisms such as the sympathetic nervous
system, the renin angiotensin aldosterone system (RAAS), and cytokine systems are then
activated, which temporarily restore cardiovascular function by increasing chronotropy,
inotropy, systemic vascular resistance, and Na+ and water retention. Chronic neurohormonal
and cytokine activation eventually become maladaptive, resulting in LV remodelling and
cardiac decompensation (HF progression).
In HF, reduced cardiac output triggers activation of the sympathetic nervous system as
inhibitory input from baroreceptors decreases and excitatory input increases. Increased
circulating levels of norepinephrine (NE) ensue in HF and act on β1- and α1-adrenergic
receptors to elicit increased systemic vascular resistance, release of renin, Na+ retention,
chronotropy, inotropy, and lusitropy. NE also stimulates production of the vasoconstrictor
endothelin-1 (ET-1), aldosterone that promotes additional Na+ and water retention, and
arginine vasopressin that further increases water retention and systemic vascular resistance.
The role of ET-1 in HF will be discussed in section 1.2.7. These adaptive responses help
maintain short-term cardiac output, however, at the expense of increased myocardial energy
requirement, enhanced arrhythmia susceptibility, and ultimately worsened HF.
Renal renin release is activated by reduced renal perfusion, reduced renal Na+ delivery,
and the sympathetic nervous system. Initially, renin cleaves four amino acids from the
precursor peptide angiotensinogen in the circulation yielding angiotensin I. Two amino acids
are then cleaved from angiotensin I by the membrane-bound angiotensin converting enzyme
(ACE) to yield the biologically active angiotensin II (Ang II). Alternatively, Ang II can also

6
be produced from ACE -independent mechanisms, including other proteases such as mast cell
chymase. In HF, ACE mRNA, protein, and activity are increased. Ang II exerts its effects
through binding to G protein coupled receptors, angiotensin type 1 and type 2 receptors.
Binding to angiotensin type 1 receptors mediates vasoconstriction, cell growth, aldosterone
production, and NE release, while binding to angiotensin type 2 receptors stimulates
antagonizing effects. Ang II increases systemic vascular resistance, stimulates release of
aldosterone from the adrenal cortex, arginine vasopressin from the posterior pituitary, NE
release, and cardiac/vascular hypertrophy.
To counteract the deleterious vasoconstrictive effects of chronically elevated levels of
NE, arginine vasopressin, ET-1, Ang II, and aldosterone, several antagonizing vasodilatory
neurohormonal systems are activated in HF. These counter neurohormonal systems include
the natriuretic peptides, bradykinin, adrenomedullin, and vasodilating prostraglandins. Atrial
natriuretic peptides (ANP) and brain natriuretic peptides (BNP) are stimulated by atrial and
ventricular stretch, respectively, in addition to Ang II and ET-1. ANP/BNP are synthesized as
prepro-ANP/BNP and cleaved by corin/furin to yield inactive N-terminal fragments NT-
ANP/NT-BNP and biologically active ANP/BNP. Both natriuretic peptides are degraded by
neutral endopeptidase (NEP) or are cleared by the natriuretic peptide receptor C. The half-life
for ANP is 3 min, while the half-life for BNP is 20 min. These peptides bind to natriuretic
peptide- A and -B receptors to exert natriuresis, vasodilation, inhibition of renin and
aldosterone, and inhibition of cardiac hypertrophy. Evaluating plasma levels of BNP and NT-
BNP has proven beneficial for the diagnosis and prognosis of HF.

7
1.1.4.1 LV remodelling
In HF, LV remodelling is characterized by changes in myocyte biology, myocyte loss,
non-myocyte gain, and loss of structural integrity. Alterations in myocyte biology include
hypertrophy, excitation-contraction coupling, ion channels, electrical coupling, contractile
proteins, cytoskeletal proteins and beta-adrenergic desensitization. Hypertrophy is an adaptive
response to hemodynamic overload. Pressure overload results in concentric hypertrophy,
where the addition of sarcomeres in parallel results in myocyte widening and increased LV
wall thickness. Volume overload causes eccentric hypertrophy, where sarcomeres added in
series results in myocyte lengthening and LV dilation. Myocyte hypertrophy leads to enlarged
mitochondria and nuclei, and progressive loss and disruption of myofibrils. Abnormal
contractile and regulatory proteins consist of reversion to fetal troponin-T and myosin heavy
chain (MHC) isoform (loss of α-MHC, gain of β-MHC), and loss of myofilaments.
Cytoskeletal protein changes include down-regulation of titin and up-regulation of desmin,
vinculin, and dystrophin. Finally, excessive β-adrenergic signalling is blunted by receptor
mediated internalization and degradation. Changes in excitation-contraction coupling, Ca2+
handling, ion channels, and electrical coupling in HF will be discussed in section 1.3.2.
Myocyte loss in HF results from increased apoptosis, necrosis, and autophagy. Gain of
non-myocytes such as fibroblasts and mast cells results in myocardial fibrosis and
extracellular matrix degradation. Upon mechanical or neurohormonal stimulation (Ang II,
aldosterone, ET-1), cardiac fibroblasts produce collagen I/III/IV, laminin, and fibronection to
repair extracellular matrix. Marked collagen synthesis in HF exemplified by increased
collagens I/III/IV/VI, fibronectin, laminin, and vimentin results in perivascular, interstitial, or
replacement/scarring fibrosis. Progressive myocardial fibrosis in addition to reduced collagen

8
cross-linking and linkage with individual myocytes leads to myocardial stiffness, LV dilation
and dysfunction, and increased propensity for lethal cardiac arrhythmias. Mast cells modify
the collagen matrix by releasing the proteases tryptase and chymase, thus mediating the
degradation of the extracellular matrix by activating the matrix metalloproteinases (MMP 1-
3,9). Mast cells can also produce cytokines IL-1/4/5/10, tumor necrosis factor-alpha (TNF-α),
leukotrienes, and nitric oxide2. In HF, increased myocardial mast cell density correlates with
increased chymase activity, MMP-2 activation, collagen degradation, and LV dilation3.
The balance between pro- and anti-inflammatory cytokines is unstable in HF,
favouring pro-inflammatory cytokines such as TNF-α and interleukin-6 (IL-6)4. Cytokines are
produced in the myocardium by myocytes and a variety of other cell types to initiate repair in
response to injury. Chronic release of inflammatory cytokines contributes to progressive LV
remodelling by provoking myocyte hypertrophy, fetal gene re-programming, stimulation of
fibroblasts, increased MMP secretion from fibroblasts, increased collagen matrix degradation,
myocardial fibrosis, and myocyte loss5. Substantial cross-talk exists between inflammatory
cytokines and the RAAS pathway, as Ang II mediates increased TNF-α mediated by nuclear
factor κB (NFκB), while inflammatory cytokines activate ACE and chymase to augment
RAAS signalling. The chronic activation of these maladaptive processes in LV remodelling
leads to increased energy requirements, hypertrophy, dilation, fibrosis, and myocyte loss, and
serves to fuel further LV remodelling and amplification of these maladaptive signalling
systems.

9
1.1.5 Treatments
The treatment strategy is based on the severity of HF symptoms; treatments include
lifestyle changes, medications to antagonize symptoms, device therapies, and surgery.
Therapy for patients in Stage A and B focuses on lifestyle changes as well as medications or
device therapies to treat the underlying condition to prevent the development of HF. Lifestyle
changes include achieving and maintaining a healthy weight through diet and regular exercise,
low sodium diet, 2L fluid restriction, blood pressure monitoring, medication compliance,
smoking cessation, and moderate alcohol consumption. Device therapies include implantation
of a pacemaker for cardiac resynchronization (biventricular pacing) or a cardioverter
defibrillator to combat lethal arrhythmias. A variety of pharmacological agents have been
developed in order to prevent the progression and to reverse HF. These medications include
ACE inhibitors, Ang II receptor blockers, beta blockers, digoxin, diuretics, aldosterone
antagonists, vasodilators, anticoagulants, and antiplatelet agents. The treatment approach for
patients with Stage C HF includes lifestyle changes, a cocktail of conventional
pharmacological agents, and implantation of pacemaker/ defibrillator for biventricular pacing
and to ensure sustained normal cardiac rhythm. For patients with end-stage HF where standard
medications have failed (Stage D), the treatment strategies include chronic infusion of
vasodilators and inotropes, permanent mechanical circulatory support via implantation of a
left ventricular assist device, repairing the underlying cause of HF such as a coronary bypass
or a valve replacement, cardiac transplantation, and experimental surgical/medicinal
approaches. Advances in blocking the hyper-activated ET-1 system in HF is examined in
section 1.2.7, while possible targets for gene therapy treatment in HF will be discussed in
section 1.3.2.1.

10
1.2 ENDOTHELIN
1.2.1 Distribution, regulation and synthesis of ET-1
ETs are 21 amino acid vasoconstrictive peptides involved in the regulation of vascular
tone and the pathophysiology of cardiovascular disease. Three ET peptides have been
identified on three separate genes, ET-1, ET-2, and ET-3. ET-1 is the main isoform with
cardiovascular actions; it is produced not only by endothelial cells, but also by vascular
smooth muscle cells, cardiomyocytes, fibroblasts, macrophages, and leukocytes6-8.
ET-2 is found in the ovary and intestinal epithelial cells, while ET-3 is found in
endothelial cells and intestinal epithelial cells. ET-1 also plays a role in neural, pulmonary,
reproductive, and renal physiology. As such, hyper-ET-1 signaling has been implicated in the
pathophysiology of HF, atherosclerosis, pulmonary hypertension, asthma, cancer, diabetes,
glaucoma, pain, sexual dysfunction, fibrosis, renal failure, inflammation, and cerebral
vasospasm. Polymorphisms in the ET-1 promoter region are associated with cardiac
hypertrophy and asthma9, 10.
ET-1 expression is primarily regulated at the transcriptional levels by various stimuli
that act on elements in the regulatory region of the ET-1 gene. ET-1 production is promoted
by Ang II, vasopressin, epinephrine, thrombin, low-shear stress, hypoxia, inflammatory
cytokines, and insulin. The production of ET-1 is inhibited by nitric oxide, bradykinin, high-
shear stress, heparin, prostaglandins, natriuretic peptides, estrogen, and progesterone.
Differential and tissue specific ET-1 expression is also regulated by mRNA instability,
epigenetics, and microRNAs. The half-life for ET-1 mRNA is 15 min11, this may result from
suicide motifs present at the 3’ region that affect mRNA stability by enhancing proteasomal

11
degradation12. The first intron of the ET-1 gene is subject to methylation and gene silencing
in mouse dermal fibroblasts13. Aldosterone regulates ET-1 transcription via histone
modification in rat inner medullary collecting duct cells14. Recently, microRNAs (miR-199,
and miR-155) have been shown to negatively regulate ET-1 expression in rat liver sinusoidal
epithelial cells and human microvascular epithelial cells15.
ET-1 is synthesized as preproET-1 and undergoes a series of proteolytic cleavage
reactions to yield the active ET-11-21 peptide. It is initially synthesized as the precursor
preproET-1(1-212), released into the cytoplasm as proET-1, and then cleaved by a furin-like
endopeptidase to yield bigET-1(1-38). The final step involves the cleavage of bigET-1 at Trp21
to the mature ET-1 peptide by an endothelin-converting enzyme (ECE). Additionally, bigET-1
can also be cleaved by chymase to yield a 31 amino acid peptide, which can then be
subsequently cleaved to the active ET-1 by a neutral endopeptidase (NEP) and/or ECE (Fig.
1.1)16, 17.
1.2.2 Clearance of ET-1
The half-life of ET-1 in plasma is less than 2 min due to efficient extraction in the
kidney and lungs. ET-1 is subject to either receptor- or enzyme-mediated degradation. ET-1 is
principally cleared by ETB receptor mediated internalization and degradation, and secondarily
by enzymatic metabolism by NEP. Although the ET-1 is degraded rapidly, its biological
effects last much longer due to the near irreversible binding nature of ET-1 with its receptors.

prepro ET-1
Big ET-11-38
ET-11-21
ET-11-31
Furin
Chymase
ECE
cleared / metabolized
NEP, ETBR
ETAR ETBR
+-
NO, ANP, CNPprostacyclin, heparin
Ang II, vasopressin, cytokines, thrombin, shearing forces, ROS
α1-PDX, CMK, furin prodomain
TY-51469, NK3201, BCEAB, TEI-F00806
BQ123, Darusentan, YM-598, ABT-627 BQ788, A-192621
CGS 26303, CGS 34043,Phosphoramidon, SLV-306
Bosentan, Tezosentan, Enrasentan, LU-420627
CGS 26303, CGS 35066, PD 069185, SM-19712
Figure 1.1. Production and degradation of ET-1. Prepro-ET-1 is regulated at the transcription level and is reduced to BigET-11-38 by a furin-like enzyme. The majority of BigET-11-38 is then reduced to mature ET-11-21 by ECE, or to ET-11-31 by a chymase and subsequently cleaved by a metalloprotease to yield active ET-11-21. ET-1 can then exert its biological effects through binding to G protein coupled receptors (ETAR, ETBR). The production of ET-1 can be inhibited at several steps of the signaling cascade. The use of ET receptor antagonism and ECE inhibition have been evaluated in conditions with chronically high levels of ET-1.
12

13
1.2.3 ECE
ECE is a membrane-bound zinc metalloprotease. Zinc metalloproteases are responsible
for processing and metabolizing peptide hormones, immunoregulatory proteins, and
neuropeptides. Although there are 3 ECE isoforms (ECE-1-3), ECE-1 is the main functional
ECE. ECE-1 has a widespread distribution as it is highly expressed in the cardiovascular,
endocrine, and reproduction systems. There are four ECE-1 splice variants (ECE1a-d) and
differ in sub-cellular localization. ECE-1 is not only responsible final processing of bigET-1
to mature ET-1, but can also hydrolyze bradykinin, substance P, Ang II, and insulin. NEP and
ACE are also zinc metalloprotease. NEP is 37% homologous with ECE and metabolizes
natriuretic peptides, bradykinin, and ET-1. ACE is responsible for the catalysis of angiotensin
I to Ang II.
1.2.4 ET Receptors and signal transduction
ET-1 acts locally in an autocrine and paracrine fashion through binding to the
G-protein coupled receptors ETA and ETB. ETA and ETB receptors differ in their affinity for
ETs, their distribution, and their association with various G-protein α subunits (Gαs, Gαi/o,
Gαq/11, Gα12/13) and thus signal transduction pathways (Fig. 1.2). ET receptors have a very high
affinity for ET-1, resulting in a nearly irreversible coupling. Low circulating levels of ET-1
combined with high tissue ET-1 levels are attributed to strong ET-1/ET receptor binding
kinetics.
ETA receptors bind to ETs with different affinities, ET-1>ET-2>ET-3; they are
distributed widely throughout the cardiovascular system, pulmonary system, central nervous
system, sensory nervous system, immune system, gastrointestinal system, kidney, prostate,

ET A
ET-1
Gαs
ET A
/B
ET-1
Gαq
ETB
ET-1
GαiAC
cAMP
PKA
RAP
MEK
PLC
DAG
PKC
Raf
SOS
ERK
IP3
↑Ca2+
CaM
CNAkt
PI3K
IKKs
NFAT NFκB
SHC GRB2
Pyk2
Ras
c-Src
↑Ca2+
14

Figure 1.2. ET-1 mediated signaling via Gαq/s/i. Gαq activation triggers PLC to produce intracellular messengers IP3 and DAG. IP3 elicits the release of SR Ca2+ stores via the IP3
receptor. The rise in intracellular Ca2+ activates CaM mediated activation of CN, enabling the nuclear translocation of NFAT to induce changes in gene transcription. DAG stimulates PKC to activate Raf, which in turn leads to the initiation of ERKs via MEKs, which regulates gene expression by activating transcription factors. Gαq mediated activation of Pyk2 also leads to initiation of the ERK/MEK cascade by activating the SHC/GRB2/SOS complex to stimulate Ras, which leads to the subsequent activation of Raf-MEK-ERK. Ras mediates activation of Akt via PI3K, enabling the phosphorylation of IKKs and subsequent nuclear translocation of NFκB to trigger changes in gene transcription. PKC also mediates activation of NFκB via inhibition of IKKs. Gαs activates AC to produce cAMP, enabling activation of PKA and Rap. Rap activates the Raf-MEK-ERK pathway. PKA acts on a variety of proteins to stimulate the release of Ca2+ from the SR and the influx of Ca2+ across the plasma membrane, among others. Gαi signalling inhibits AC, thus blunting cAMP levels and PKA-mediated signalling. Gαi also activates c-Src, which triggers the MEK pathway via activation of the SHC/GRB2/SOS complex. PLC= phospholipase C, IP3=inositol trisphosphate, DAG = diacylglycerol, CaM = calmodulin, CN = calcineurin, NFAT = nuclear factor of activated T-cells, PKC = protein kinase C, MEK = mitogen-activated protein kinase kinases, ERK = extracellular signal-regulated kinases, PYK2 =proline-rich tyrosine kinase-2, GRB2 = growth factor receptor-bound protein-2, SOS = son of sevenless, Ras = rat sarcoma, PI3K =phosphoinositide 3-kinase, IKK = inhibitor of κB kinase, NFκB = nuclear factor-κB, AC = adenylyl cyclase, cAMP = cyclic adenosine 3,5-monophosphate, PKA = protein kinase A, RAP = ras-related protein, SR = sarcoplasmic reticulum
15

16
ovary, and pancreas. ETA receptors are up-regulated by hypoxia, cyclosporine, epidermal
growth factor, basic fibroblast growth factor, cAMP, and estrogen, and are down-regulated by
ET-1, Ang II, platelet-derived growth factor, and transforming growth factor. ETA receptors
are coupled to Gαs, Gαq/11, and Gα12, and generally induce vasoconstriction, mitogenesis,
angiogenesis, matrix formation, inflammation, apoptosis, and electrical remodeling18-20.
Selective ETA receptor antagonists have been discovered: ZD4054, atrasentan, darusentan,
macitentan, ambrisentan, and sitaxsentan.
ETB receptors bind to all ETs with equal affinity and are distributed less extensively;
they are found in the cardiovascular system, the pulmonary system, neurons, bone, pancreas,
and kidney. ETB receptors are up-regulated by C-type natriuretic peptide and Ang II, and
down-regulated by cAMP and catecholamines. ETB receptors are coupled to Gαi/o, Gαq/11, Gα13
and generally mediate vasodilation, natriuresis, ET-1 clearance, vasoconstriction, and anti-
apoptosis21-23. Agonists (sarafotoxin 6c and IRL1620) and selective antagonists (BQ788,
A192621, RES7011, and IRL2500) have been discovered for the ETB receptor. Several non-
selective ET-1 receptor antagonists have also been developed: bosentan, tezosentan,
enrasentan, and LU-420627.
1.2.5 Transgenic mouse models: genetic manipulation of ET-1 system
The ET-1 system is necessary for normal embryonic development, more specifically in
the development of tissues derived from embryonic neural crest. ET-1/ETA receptor activity is
essential to normal cranial and cardiovascular development resulting from impaired neural
crest cell communication. ET-1 knockout mice die at birth due to craniofacial abnormalities
resulting from respiration failure24. They also display cardiovascular, thyroid, and thymus

17
malformations25, 26. Mice with cardiac specific deletion of ET-1 have a reduced hypertrophic
response and with age develop dilated cardiomyopathy, increased fibrosis and apoptosis, and
impaired NFκB activation27, 28. ETA receptor-deficient mice develop craniofacial and
cardiovascular malformations similar to the ET-/- phenotype29.
Over-expression of human ET-1 led to high transgene levels in the brain, lung, and
kidney; these mice developed renal cysts, fibrosis, glomerulosclerosis and pulmonary fibrosis
and inflammation30, 31, while over-expression of human ET-2 led to glomerulosclerosis32.
Endothelium specific over-expression of ET-1 led to elevated tissue and plasma levels of ET-1,
vascular remodelling, and endothelial dysfunction33. Cardiac specific over-expression of
human ET-1 led to an inflammatory cardiomyopathy characterized by increased expression of
inflammatory cytokines, NFκB nuclear translocation, LV dilation and contractile dysfunction
and death (Fig. 1.3)34.
ET-3/ETB receptor activity is essential to normal epidermal melanocyte and enteric
neuron development. Disruptions in either ET-3 or ETB receptor genes result in aganglionic
megacolon and pigmentary disorders35, while ET-3 over-expression results in
hyperpigmentation. Endothelial specific deletion of ETB receptors results in endothelial
dysfunction, impaired nitric oxide release, elevated plasma ET-1 levels, and resistance to
high-salt-diet-induced hypertension36.
ECE-1 knockout mice exhibit similar development defects as those seen in ET-1 and
ETAR knockout mice, as well as those seen in ET-3 or ETB receptor knockout mice. The
majority of ECE-1 knockout mice die in utero due to severe cardiac abnormalities. The
surviving mice exhibit craniofacial and cardiac abnormalities similar to those observed in both
ET-1 and ETA receptor knockout mice37. ECE-1-/- mice also lack epidermal melanocytes

Yang et al, Circulation, 2004
ppET-1 pBi β-gal
+ DOX
αMHC promoter tTA
Figure 1.3. DOX-off system of cardiac over-expression of ET-1. Mice harboring the tetracycyline transactivatior (tTA) under the control of the α-myosin heavy chain promoter were crossed with a line harboring a human ET-1 transgene under control of a tTA-responsive promoter, and thus using a DOX-off system of conditional cardiac-specific over-expression of human ET-1.
18

19
and enteric neurons, a phenotype that parallels that seen in ET-3 or ETB receptor knockout
mice. Mice lacking ECE-1 can produce substantial levels of ET-1. ECE-2 knockout mice
develop normally, however double ECE-1-/-/ECE-2-/- mice exhibit a more severe ECE-1-/-
embryonic phenotype and persistent mature ET-1 levels38. Thus, it is apparent that other non-
ECE proteases can also generate mature ET-1 from bigET-1. Chymase, NEP, and MMP-2 are
likely candidates, as they can also metabolize bigET-1. Tissue specific distribution of ECE,
and consequently ET-1 production are important in normal development, as developmental
defects persist in these mice despite retaining the ability to synthesize ET-1.
1.2.6. Role of ET-1 in inflammation, hypertrophy, and fibrosis
In the heart, ET-1 contributes to inflammation, hypertrophy, fibrosis, and electrical
remodelling (Fig. 1.4). ET-1 acts as a pro-inflammatory cytokine by priming neutrophils and
stimulating the release elastase from neutrophils and histamine from mast cells39-41. ET-1
stimulates NFκB dependent IL-6 release from vascular smooth muscle cells and interleukins
and adhesion molecules from leukocytes42, 43. ET-1 also induces monocytes to produce
inflammatory cytokines and chemokines; CD40 production is mediated by NFκB, while
macrophage inflammatory protein-1β is dependent on hypoxia-inducible factor-1α, AP-1 and
NFκB44-46.
NFκB is a transcription factor that responds to a number of extracellular stimuli (pro-
inflammatory cytokines, pro-apoptotic/necrotic, viral/bacterial antigens). NFκB functions as a
homo- or hetero-dimer of structurally similar subunits: p50, p52, p65/RelA, RelB, and c-rel.
The N-terminus of all 5 NFκB subunits contains a conserved DNA binding domain, a
dimerization domain, and a nuclear localization signal. RelA, RelB, and c-rel have

Vascular Smooth Muscle Cell
Fibroblast
Macrophage
Endothelial Cell
↑ Collagen↑ Fibrosis
↑ Adhesion↑ Migration↑ Cytokines
Vasodilation↑ ET-1 Clearance
VasoconstrictionProliferation
MigrationETA
ETA
ETA
ETB
ETB
ETB
ETB
Cardiomyocyte
ETA ETB
HypertrophyElectrical Remodelling
ET-1
Figure 1.4. Multifaceted nature of ET-1 signaling. Depending on the target cell type, ET-1 stimulates vasoconstriction, vasodilation, inflammation, fibrosis, hypertrophy, and electrical remodelling.
20

21
muscle cells and epithelial cells through ETA receptor mediated mechanisms47, 48. In
fibroblasts, ET-1 induces chemotaxis, proliferation, collagen production, and inhibition of
MMP expression through both ETA and ETB receptor dependant mechanisms49-51, as well as
transactivation domains in their C-terminus. The other NFκB proteins, p50 and p52, are
synthesized as precursors p105 and p100, respectively. These precursors contain ankyrin
repeats at their C-terminus that enable them to act as inhibitors of κB (IκB) proteins. Cleavage
and proteasomal degradation of the p105 and p100 yield mature p50 and p52. Unlike the
other NFκB proteins, p50 and p52 have transrepression domains in their C-terminus. However,
they can also mediate transactivation by dimerization with RelA, RelB or c-rel. Inactive
NFκB is sequestered in the cytoplasm by its interaction with an IκB protein. The IκB family
consists of IκBα, IκBβ, IκBγ, IκBε, Bcl-3, p105 and p100. All IκB proteins contain ankyrin
repeats that enable them to interact with NFκB dimers. This interaction inhibits the nuclear
translocation of NFκB by masking their nuclear localization signal. A variety of external
stimuli cause phosphorylation of two residues of the IκB by IκB kinase (IKK).
Phosphorylation enables dissociation and subsequent proteasomal degradation of IκB, thus
relieving the inhibition of NFκB. This enables NFκB translocation to the nucleus where it
binds to specific DNA binding sites in order to transcriptionally repress or activate genes
involved in cell proliferation, survival, differentiation, immunity and inflammation (Fig 1.5).
Heightened NFκB activation is associated with cancer, autoimmune-, neurodegenerative-, and
cardiovascular-diseases such as atherosclerosis, hypertrophy, and HF52-54. ET-1 can activate
NFκB by PKC, PI3K/Akt, ERK1/2, and p38 MAPK dependent mechanisms (Fig. 1.2). Lack
of NFκB p50 has been shown to improve survival and LV remodeling in a model of TNFα-
induced cardiomyopathy and after myocardial infarction55, 56.

Figure 1.5. Illustration of NFκB activation. In the cytoplasm, NFκB is kept in the inactive state whilst bound to the inhibitory protein IκBα. When activated by various extracellular stimuli, such as ET-1, IKK phosphorylates IκBα, leading to the dissociation and subsequent proteasomal degradation of IκBα, thereby leaving NFκB free to translocate to the nucleus to transcriptionally repress or activate target genes. NFκBi= inactive, NFκBa= active
ETA
ET-1
IKK
p65
p50
IκBα
p65
p50
IκBαPP
p65
p50
IκBαPP proteasome
degradation
p65
p50
transcriptional repression /activation
NFκBi
NFκBa
22

23
As a pro-fibrotic factor, ET-1 regulates the expression and degradation of several
components of the extracellular matrix. ET-1 induces remodelling of the extracellular matrix
by stimulating the synthesis and release of collagens and fibronectin from vascular smooth
stimulates the differentiation of fibroblasts to contractile myofibroblasts through the ETA
receptor mediated PI3K/Akt pathway57. ET-1 also induces the differentiation of epithelial
cells to fibrotic mesenchymal cells58.
ET-1 acts as a pro-hypertrophic factor by inducing hypertrophic gene reprogramming
via activation of the transcription factors NFκB, NFAT, and zinc finger protein 260. ET-1
induces cardiomyocyte hypertrophy by triggering a rise in intracellular Ca2+ through
activation of NHE and reverse mode NCX, elevated intracellular Ca2+ enables activation of
calcineurin and subsequent dephosphorylation and nuclear translocation of NFAT59. ET-1 also
triggers hypertrophy via the calcineurin/NFAT pathway by inducing nuclear Ca2+ release by
perinuclear IP3R60. ET-1 has been shown to stimulate cardiomyocyte hypertrophy through
activation of ERK, activation of NFκB by p38 MAPK, and activation of the zinc finger
protein 260 by PKC.61-63 The role of ET-1 in electrical remodelling will be discussed in
section 1.3.3.
1.2.7 Pathophysiology of ET-1 in HF
ET-1, bigET-1, ECE-1, ETA receptors are elevated in human and animal models of
HF64-67. In addition, ETA and ETB receptor binding is increased, and ETB receptors are
downregulated in HF68, 69. Levels of plasma ET-1 and bigET-1 also correlate with disease
severity and survival in HF due to increased ET-1 production in the circulation and
myocardium66, 70. Blocking ET-1 signaling for the treatment of HF is of great interest, as

24
ET-1 signaling is hyper-activated in HF and contributes to increased systemic vascular
resistance, cardiac hypertrophy and fibrosis, inflammation and arrhythmia predisposition.
Inhibiting ET-1 signaling via receptor antagonism has not proven beneficial in the
treatment of clinical HF. Initially, ET-1 receptor blockade looked like a promising avenue for
the treatment of HF as many experimental models of HF showed improved survival,
hemodynamics and ventricular remodelling with both non-selective ET-1 receptor blockade
and ETA receptor blockade71-73. Also, preliminary small-scale clinical trials with acute doses
of bosentan, darusentan, BQ-123, and tezosentan showed short-term hemodynamic benefit in
patients with HF74-77. However, the subsequent RITZ (Randomized Intravenous TeZosentan)
trial showed no therapeutic benefit on mortality or HF progression, and was associated with
adverse side effects such as elevated levels of liver transaminases. The VERITAS-1 and -2
(value of endothelin receptor inhibition with tezosentan in acute HF study) trials were
discontinued after a year due to lack of improvement78. Four large-scale clinical trials did not
show improvement in HF status, mortality, or hospitalization. The REACH-1 (randomized
endothelin antagonism in chronic HF) study was discontinued early due to worsened HF and
development of adverse side effects such as high levels of liver transaminases, anaemia, and
fluid retention with treatment of high-dose bosentan79. Next, ENABLE-1 and -2 (endothelin
antagonist bosentan for lowering cardiac events in HF) studies showed treatment with low-
dose bosentan lead to early worsening of HF, abnormal liver function, fluid retention, and no
survival benefit80. Treatment with enrasentan in the ENCOR (enrasentan cooperisedative
randomized evaluation) study resulted in higher rates of HF and mortality. Lastly, the EARTH
(endothelin A receptor antagonist trial in HF) trial showed no benefit to survival or HF

25
progression with darusentan treatment81. The disappointing outcome of ET-1 receptor
antagonism for the treatment of HF may reflect a problem with dosage, receptor selectivity of
the drug, timing of treatment, or adverse effects due to combination therapy with existing
treatments. However, ET-1 receptor blockade has been effective in the treatment of clinical
pulmonary arterial hypertension.
Blocking ET-1 synthesis with the use of ECE inhibitors has also been evaluated in
order to blunt ET-1 over-production in various disease states. Many ECE inhibitors are also
dual ECE/NEP or triple ECE/NEP/ACE inhibitors. Using non selective ECE inhibitors would
simultaneously block the synthesis of potent vasoconstrictors ET-1 and Ang II, while
promoting the beneficial vasodilator actions of natriuretic peptides and bradykinin by
inhibiting their degradation. In experimental models of HF, dual ECE/NEP inhibitors have
been beneficial in increasing cardiac output, diuresis, and reducing cardiac/vascular
remodelling and secretion of neurohormones82-86. The use of the dual ECE/NEP inhibitor SLV
306 is currently being tested in a phase II clinical trial for the treatment of HF.
1.3 ELECTRICAL REMODELLING
1.3.1 Excitation in the healthy heart
Cardiac excitation and conduction depend on the flux of ions across the cell membrane,
efficient Ca2+ cycling, myocyte architecture, and electrical coupling between myocytes. Here,
we describe the ion channels, Ca2+ handling proteins, and gap junctions that facilitate cardiac
impulse generation and propagation.

26
1.3.1.1 Ionic basis of cardiac action potential
Electrical impulses in the heart are initiated by the pacemaker cells of the sinoatrial
node (SAN), propagate through the right and left atria, across the atrioventricular node (AVN)
to the His-bundle, and finally, travel through the Purkinje fibers (Fig. 1.6A). Coordinated
propagation of the electric impulse is coupled to the rhythmic contraction of the heart. The
cardiac action potential (AP) is shaped by the intricate balance between inward depolarizing
currents and outward repolarizing currents across the cell membrane (Fig. 1.6B). The shape of
the AP varies with cell type and is composed of 4 phases. At rest, the transmembrane potential
is -80mV. During phase 0, the cell is rapidly depolarized by inward Na+ current (INa). Here,
the amount of INa is proportional to the maximum rate of voltage change (dV/dt).
Depolarization of the membrane then triggers activation of Ca2+ and K+ currents. The brief
repolarization in phase 1 is caused by the activation of the transient outward K+ current (Ito).
In atrial myocytes, the ultrarapid delayed rectifier K+ current also plays a role in phase 1. Next,
the plateau phase is maintained by inward L-type Ca2+ current (ICa-L) and outward K+ currents.
Phase 3 repolarization is influenced by the progressive activation of the rapid delayed rectifier
(IKr) and the slow delayed rectifier K+ current (IKs). The final repolarizing phase is generated
by the inward rectifier K+ current (IK1).
1.3.1.2 Electrophysiological mapping
Various electrophysiological techniques have been developed to characterize cardiac
electrical activity at the organism, tissue, cell, and channel levels. The electrocardiogram
(ECG) is used to map the electrical activity through all regions of the heart at the organism
level (Fig. 1.6C). Intracardiac mapping is used to locate regions of electrical disturbance and

SA node
AV node
His Bundle
Bundle branches
Purkinje fibers
P
R
Q
S
T
QRS interval QT
interval
PR interval
0 4
12
3
INa
IK1
Ito ICa-L
IKr, IKs
A
B
C
Figure 1.6. Schematic of the cardiac conduction system, the ionic currents contributing to the cardiac action potential, and the surface electrocardiogram. (A) Spread of excitation throughout the heart. (B) Ionic currents underlying the four phases of a typical ventricular action potential. (C) Various parameters of the surface electrocardiogram; PR = time from earliest-atrial to -ventricular activation, QRS = ventricular depolarization time, QT = ventricular depolarization and repolarization time.
27

28
identify conduction anomalies between the atria and His-bundle, or between the His-bundle
and the ventricles. Optical mapping with voltage-sensitive fluorescent dies and epicardial
mapping with microelectrodes can be used to study the propagation of electrical activity
throughout the tissue or cardiomyocyte mono-layer. Patch-clamp can be used to study the AP
waveform, whole-cell currents, and individual ion channel recordings.
1.3.1.3 Excitation-contraction coupling and Ca2+ cycling
Cardiac contraction is activated by a rise in intracellular free Ca2+. To work
effectively, Ca2+ homeostasis in cardiac myocytes is maintained on a beat to beat basis (Fig.
1.7). Excitation-contraction coupling is the process that links depolarization of the cell
membrane to contraction of the myofilaments. During an AP, depolarization of the cell
membrane causes an influx of Ca2+ into the cytoplasm via voltage-gated Ca2+ channels in the
plasma membrane. This local rise of Ca2+ triggers the release of Ca2+ stored in the
sarcoplasmic reticulum (SR) into the cytoplasm via the ryanodine receptor (RyR2) to activate
the contractile machinery. The amplitude of contraction is dependent on the level of Ca2+ that
is in the vicinity of the myofilaments. Relaxation occurs due to repolarization of the plasma
membrane and a rapid reduction in intracellular Ca2+ levels. This decline in intracellular Ca2+
is mediated principally by the sequestration of Ca2+ into the SR via the SR Ca2+-ATPase
(SERCA2a) and the efflux of Ca2+ from the cell by the Na+/Ca2+ exchanger (NCX1).
1.3.1.4 Ca2+ handling proteins and Ca2+ current
The SR regulates efficient contraction and relaxation of the contractile machinery on a
beat to beat basis by controlling the release and sequestration of Ca2+. The two types of Ca2+

Na+
Ca2+
NCX
Ca2+
Myofilaments
Ca2+
Ca2+
SR
SERCA
RyR
Ca2+
Ca2+
Figure 1.7. Ca2+ cycling in healthy cardiac myocytes. Ca2+ handling is tightly controlled on a beat to beat basis; it provides the link provides between excitation of the cell membrane and contraction of the myofilaments. Depolarization of the cell membrane triggers an influx of Ca2+ into cell via the L-Type Ca2+ channel. This local rise in Ca2+ causes the release of SR Ca2+ stores into the cytoplasm via the RyR to activate the contractile machinery. Relaxation occurs when Ca2+ exits the cytoplasm principally by the re-uptake of Ca2+ into the SR by SERCA, or is extruded via NCX. GJ = gap junction
K+
Na+
GJ
Na+
NHX
H+
IP3R
ATPase
K+
PLB
29

30
release channels on the SR are the RyR2 and the inositol triphosphate receptor (IP3R). IP3R2
(gene ITPR2) are expressed at low abundance in the heart and mediate IP3 induced release of
Ca2+; their role in excitation-contraction coupling is not known. RyR2 (gene RYR2) is the
dominant Ca2+ release channel in the heart and plays key role in excitation-contraction
coupling87, 88. These channels are tetrameric proteins, with Ca2+ activation and inactivation
sites, as well as several regulatory sites. RyR2 are found in close proximity to the L-type Ca2+
channels in the cell membrane for efficient excitation-contraction coupling. They are found in
a macromolecular complex with kinases, kinase anchoring proteins, phosphatases, and FK-
506 binding proteins, which regulate channel activity89. FK-506 binding proteins function to
stabilize RyR, and inhibit diastolic Ca2+ release from the SR90. Mutations in RyR2 have been
found in patients with catecholaminergic polymorphic ventricular tachycardia and more
recently, long QT syndrome (LQT)91.
During diastole, Ca2+ is primarily sequestered back into the SR through SERCA2a, or
is extruded from the cell through the NCX1. NCX1 (gene SLC8A1) normally operates in
forward mode, exchanging one intracellular Ca2+ ion for three extracellular Na+ ions, resulting
in a net inward current. However, NCX1 activity is reversible and its direction depends on the
electrochemical ion gradient. Reverse mode supports Ca2+ entry and Na+ extrusion when high
intracellular Na+ levels are generated from rapid inward INa during AP depolarization, or by
the Na+/H+ exchanger (NHE1). NCX1 is localized to T-tubules in a macromolecular complex;
it is regulated by Ca2+, Na+, pH, phosphorylation, phosphatidylinositol bisphosphate and
several interacting proteins such as phospholemman, CN, and 14-3-3 proteins92. SERCA2a
(gene ATP2A2) is responsible for reducing intracellular Ca2+ during relaxation by restoring the
SR stores for subsequent Ca2+ release during the next beat. SERCA activity is regulated by

31
phospholamban (PLB; gene PLN). PLB inhibits SERCA2a activity in its unphosphorylated
state. Phosphorylation induces a conformational change in PLB and relieves its inhibition of
SERCA, and thereby increases the Ca2+ pump rate and the reuptake of Ca2+ into the SR93. The
inhibition of SERCA is restored upon dephosphorylation of PLB by SR associated
phosphatases94. PLB mutations have been found in patients with familial dilated
cardiomyopathy95-97.
In the heart, there are two types of voltage-gated Ca2+ channels, T-type and L-type
Ca2+ channels. T-type Ca2+ channels exhibit a small conductance and activate at low voltages.
ICa-T is carried by the α subunits Cav3.1 (gene CACNA1G) and Cav3.2 (gene CACNA1H).
Their role in excitation-contraction coupling is not clear, as they are expressed in the working
myocardium during development, but are restricted to the cardiac conduction system in
adulthood and are re-expressed during cardiac pathological conditions98, 99.
The L-type Ca2+ channel is the main Ca2+ channel in working cardiac myocytes, ICa-L
provides the main influx of Ca2+ which triggers excitation-contraction coupling. L-type Ca2+
channels exhibit a large conductance and activate at high voltages. Cav1.2 (gene CACNA1C) is
the dominant α pore forming subunit in the heart and is composed of 4 homologous domains
of 6 transmembrane α helices that contain the voltage/Ca2+ sensor, gating mechanism, and
regulatory sites. Cav1.2 co-assembles with a β subunit and an α2δ subunit. Cavβ subunits
(Cavβ1-4, gene CACNB1-4) and α2δ subunits (α2δ1-4, gene CACNA2D1-4) regulate Cav1.2
expression and function. Channel inactivation is voltage and Ca2+ dependent, as calmodulin
enhances inactivation of ICa-L by sensing the local accumulation of Ca2+. L-type Ca2+ channels
are modulated by kinases, phosphatases, sorcin, and cytoskeletal ahnak protein and are found
in T-Tubules adjacent to RyR2 in the SR. Recently, the K+ channel interacting protein 2

32
(KChIP2) has been shown to interact with the N-termini of Cav1.2 and positively regulate ICa-
L100. Several L-type Ca2+ channel channelopathies have recently been discovered. Loss of
function mutations in CACNA1C and CACNB2b are linked to sudden cardiac death and
Brugada syndrome101, 102. Gain of function mutations in CACNA1C are associated with
Timothy syndrome (a multisystem disorder exhibiting congenital heart disease), LQT, and
lethal arrhythmias103.
1.3.1.5 Sodium current
INa generates the depolarizing current for phase 0 of the cardiac AP and contributes to
its propagation throughout the myocardium. Nine functional α-subunit of voltage-gated Nav
channels (Nav1.1-1.9) have been discovered, which differ in channel kinetics and response to
neurotoxins. The majority of neuronal Nav channels (Nav1.1-1.3, 1.6-1.7) and skeletal Nav
channels (Nav1.4) are sensitive to the neurotoxin tetrodotoxin, while the α-subunit of the
cardiac voltage gated Na+ channel Nav1.5 (gene SCN5A) and neuronal Nav1.8 are mostly
resistant. Although Nav1.5 is the main Nav channel in the heart, several other skeletal (Nav1.4)
and neuronal Nav isoforms (Nav1.1-1.3, 1.6-1.8) have also been found in the heart. More
specifically, Nav1.1 has been found in the SAN, His bundle, Purkinje fibers PF, and ventricles;
Nav1.2 in the His bundle, Purkinje fibers, and ventricles; Nav1.3 in SAN, AVN, and
ventricles; Nav1.4 mRNA in SAN, AVN, atrium, and ventricles; Nav1.6 in SAN and
ventricles; Nav1.7 mRNA in AVN, and and finally Nav1.8 in atrium104, 105. Various studies
have shown that tetrodotoxin-sensitive Nav channels account for 5-10% of INa in ventricular
myocytes106, and Nav1.1 and Nav1.3 have been linked with SAN automaticity107. Also, a

33
Nav1.8 variant has been linked with prolonged cardiac conduction (P wave/ PR/QRS intervals)
and increased susceptibility to heart block and ventricular arrhythmias108.
Nav1.5 is a 220 kDa protein comprised of 4 homologous domains each containing 6
transmembrane segments, 3 intracellular linking domains, and cytoplasmic N- and C-termini.
β-subunits (β1-β4; gene SCN1B-SCN4B) are 30-35 kDa proteins with 1 transmembrane,
spanning domains that interact with Nav1.5 to regulate channel function, density, and location.
Several proteins have been shown to interact with and regulate the expression and function of
Nav1.5, including ankyrin-G, α1-syntrophin, multicopy suppressor of gsp1, Nedd-like
enzymes, calmodulin kinase II, protein tyrosine phosphatase H1, 14-3-3η, caveolin-3,
fibroblast growth factor homologous factor 1B, calmodulin, glycerol-3-phosphate
dehydrogenase like protein, telethonin, and plakophilin-2. Recently, NFκB p50 has been
linked with transcriptional repression of SCN5A109.
Nav1.5 is essential to ventricular development, as well as initiation and propagation of
the cardiac electrical impulse. Mice lacking Nav1.5 develop defects in ventricular
morphogenesis and die in utero by E11.5110. Heterozygous SCN5A mice exhibit delayed atrial,
AVN, and ventricular conduction, reduced Na+ channel conductance, and increased
susceptibility to ventricular arrhythmias110. Older SCN5A +/- mice develop extensive
ventricular conduction delays, fibrosis, and disturbed expression of connexin 43 (Cx43)111.
Mutations in SCN5A have been discovered in patients with various cardiac disorders.
Gain of function mutations generally increase late INa and are linked with congenital LQT3112.
Loss of function mutations decrease INa and are linked with Brugada syndrome, cardiac
conduction disease, Lenegre disease, congenital AV block, sudden infant death syndrome, and

34
sick sinus syndrome113-118. SCN5A variants have also been linked with atrial fibrillation and
dilated cardiomyopathy119, 120.
Several mutations in proteins that regulate the function of Nav1.5 have also been
discovered in patients with arrhythmias. Mutations in caveolin-3 (gene CAV3) have been
linked with congenital LQT9 and sudden infant death syndrome121, 122, while mutations in α1-
syntrophin (gene SNTA1) are associated with congenital LQT12123, 124. Mutations in all four of
the Nav1.5 accessory β-subunits have been identified; SCN1B with Brugada syndrome type 5,
cardiac conduction disease, and atrial fibrillation125, 126; SCN2B with atrial fibrillation126;
SCN3B with Brugada syndrome type 7127; and SCN4B with congenital LQT10128.
While the above studies have linked abnormalities in Na+ channels to rhythm disorders,
Na+ channels blockers have also been used to treat various arrhythmias. Na+ channel blockers
are classified as class I antiarrhythmic agents (class Ia, Ib, Ic), with each sub-class differing in
degree of blockade, and effects on phase 0 depolarization and APD. As changes in phase 0
depolarization do not always affect APD, shortened APD may reflect inhibition of Na+
currents operational in later stages of repolarization. Class Ia agents are moderate Na+ channel
blockers used to treat atrial fibrillation/flutter, supraventricular tachycardia (SVT), and
ventricular tachycardia (VT). These agents moderately reduce phase 0 depolarization and
prolong APD. Class Ib agents are weak Na+ channel blockers used to treat VT; they
marginally affect phase 0 depolarization and shorten APD. While class Ic agents are robust
Na+ channel blockers used to treat life-threatening SVT and VT, they markedly depress phase
0 depolarization and have no appreciable effect on APD.

35
1.3.1.6 Transient outward K+ current (Ito)
Ito is a rapidly activating and inactivating voltage-gated K+ current that contributes to
the early phase of repolarization in working myocytes. It is comprised of two distinct
components, a slow recovering component (Ito,s) carried by the Kv1.4 (gene KCNA4) α subunit,
and a fast recovering component (Ito,f) carried Kv4.2 (gene KCND2) and Kv4.3 (gene KCND3)
α subunits. In mice, Kv4.2 is critical for functional Ito in the working myocardium129. Classical
voltage-gated K+ channels are composed of a tetramer of Kvα subunits each contains 6
transmembrane segments (S1-S6), a voltage sensor (S4), a pore region (selectivity filter)
between S5 and S6, and cytoplasmic N- and C-termini. The pore region confers K+ selectivity
and the N-terminus contains the tetramerization domain for Kvα subunit assembly. Kvα
subunits associate with several accessory proteins: Kv channel interacting proteins (KChIPs),
MinK-related peptides (MiRP), and Kvβ subunits. KChIP tetramers associate with the N-
termini of Kv4α subunits and affect channel expression and kinetics. MinK is a single
transmembrane spanning accessory subunit that regulates Kvα channel trafficking and kinetics.
Kvβ tetramers interact with the N-termini of Kvα subunits and regulate channel trafficking and
kinetics.
KChIP2, MiRP1, MiRP2, Kvβ and diaminopeptidyl transferase-like protein-6 regulate
Kv4α channels. KChIP2 (gene Kcnip2) is a Ca2+ binding protein that is essential to murine
Itof130. Recent in vitro studies suggest that INa and Ito are structurally and functionally coupled
via accessory subunits such as KChIP2131. Several protein kinases such as PKA, PKC, ERK,
and calmodulin kinase II modulate Kv1.4 and Kv4α subunits. Phosphorylation by p90
ribosomal S6 kinase, c-Src, and G-protein-coupled kinase-2 has been shown to modulate Itof
and membrane trafficking132-134. Cytoskeleton proteins filamin and α-actinin, scaffolding

36
proteins post-synaptic density protein and synapse-associated protein 97, and syntaxin 1A
associate with and regulate Ito channels135. To date, a gain of function mutation in MiRP2
(gene KCNE3) has been linked with Brugada syndrome136.
1.3.1.7 Delayed rectifier K+ current (IK)
IK plays a role during late repolarization of the AP and maintaining the AP duration
(APD). It is composed of a slowly activating component (IKs) carried by the α protein
KvLQT1 (gene KCNQ1) and a rapidly activating component (IKr) carried by the α protein
HERG (gene KCNH2). KvLQT1 functionally form tetramers and associates with the β-subunit
MinK (gene KCNE1) in a 4:2 ratio, while HERG co-assembles with the β-subunit MiRP1
(gene KCNE2). Mutations in KCNE1 are linked with LQT5 and familial atrial fibrillation137,
138, and mutations in KCNE2 also correlate with atrial fibrillation139. IKs is modulated by PKA,
CaM, Nedd4-2, and PIP2. IKr is regulated by the membrane protein KCR1 and rho-GTPase
activating protein 6. KvLQT1 and HERG channels are functionally and structurally linked140.
Several loss of function mutations in KCNQ1 and KCNH2 are associated with LQT1 and
LQT2 respectively141-143. In contrast, gain of function mutations in KCNQ1 are linked with
atrial fibrillation and short QT syndrome type 2144, 145, while KCNH2 mutations are associated
with short QT syndrome type 1146.
1.3.1.8 Inward rectifier K+ current (IK1)
IK1 is carried mainly by Kir2.1 (gene KCNJ2) and is important during the final phase
of AP repolarization as well as for stabilizing the resting membrane potential. IK1 is
characterized by a smaller outward K+ current at depolarized membrane potentials (0 mV)

37
and by a larger outward K+ current at more negative membrane potentials approaching -96
mV (equilibrium potential for K+) due to a voltage-dependent block of outward current by
intracellular polyamines and Mg2+. A functional IK1 channel is composed of a tetramer of
Kir2.1 channels, each comprised of a pore region and 2 transmembrane spanning domains.
Kir2.1 channels are regulated by PIP2. Several Kir2.1 channelopathies have been discovered.
Gain of function mutations are linked with familial atrial fibrillation and short QT syndrome
type 3147, 148, whereas loss of function mutations are associated with catecholaminergic
polymorphic ventricular tachycardia 3 and LQT7-related Andersen-Tawil syndrome 1149, 150.
1.3.1.9 Gap junctions
Gap junctions mediate electrical and metabolic coupling between cells. In the heart,
these channels enable propagation of the action potential and the diffusion of ions, second
messengers and small metabolites less than 1000 Da. Gap junction channels are formed by the
docking of two hemichannels found on adjacent cells. Hemichannels are formed from the
oligomerization of 6 connexin proteins. Each connexin is composed of 4 transmembrane α-
helices, 2 extracellular loops, a cytoplasmic loop, and cytoplasmic N- and C- termini. Channel
function is regulated by Ca2+, pH, voltage, and phosphorylation. The C-terminal domain
contains numerous regulatory sites for phosphorylation and protein-protein interactions.
The mammalian heart contains Cx30.2 (gene GJD3), Cx40 (gene GJA5), Cx43 (gene
GJA1), and Cx45 (gene GJA7) (Fig 1.8). Cx43 is the principal Cx in the heart; it is expressed
in all working cardiomyocytes, bundle branches and Purkinje fibers (Fig 1.8A). Cx40 is
located in atrial myocytes, His bundle, bundle branches, and Purkinje fibers (Fig 1.8B),
whereas Cx45 and Cx30.2 are only found in the specialized cells of the conduction system

Cx43
Cx40
Cx45/Cx30.2
A
B
C
Figure 1.8. Regional connexin expression in the heart. (A) Cx43 is expressed in the atria, ventricles, and His-Purkinje system. (B) Cx40 is mainly expressed in the atria. (C) Cx45 and Cx30.2 are expressed throughout the specialized tissues of the conduction system.
38

39
(Fig 1.8C). As the half-life for Cx43 is less than 5 hrs, the life cycle of Cx43 is tightly
controlled by several kinases151. Cx43 is subject to both lysosomal and proteasomal
degradation152. Cx43 phosphorylation regulates Cx43 trafficking to the cell membrane,
channel assembly, gating, and turnover. PKC-induced phosphorylation of p-Ser368 reduces
unitary channel conductance153, while Src-induced phosphorylation of Tyr265/247 induces
reduced gap junction communication154 , CK1-induced phosphorylation of Ser325/328/320
regulates gap junction assembly 155, PKA-induced phosphorylation of Ser364/365 increases gap
junction assembly by inducing Cx43 trafficking 156, 157, and MAPK-induced phosphorylation
of Ser255/279/282 reduces gap junction communication, likely via reduced open channel time158.
Cx43 is normally found at intercalated disks in a multi-protein complex, interacting with
various structural proteins such as zona occcludens-1, cadherins, catenins, microtubules, and
tubulins159. Interaction with zona occcludens-1at Pro375/377 regulates gap junction size and
localization at intercalated disks160, while binding of a Cx43 interacting protein of 85kDa
(CIP85) to residues 253-256 induces Cx43 turnover via the lysosomal pathway161. The T-box
transcription factor Tbx2 and Tbx3, and Nkx2.5 have been shown to negatively regulate Cx43
transcription162, 163.
Cx43 is a high conductance channel essential to normal cardiac development and
propagation of the electrical impulse in the working ventricular myocardium. Mice lacking
Cx43 develop ventricular conduction delays and arrhythmias and die perinatally due to
cardiac malformations of the RV outflow tract164, 165. Neonatal mouse ventricular myocytes
(NMVM) from Cx43-/- mice exhibit 25 fold slower ventricular conduction166. Heterozygous
Cx43 mice express 50% less Cx43 protein and exhibit slowed ventricular conduction and
increased susceptibility for spontaneous arrhythmias167, 168. Cardiac specific deletion of Cx43

40
results in delayed ventricular conduction and sudden cardiac death within 2 months169.
Inducing deletion of Cx43 post-natally results in slowed ventricular conduction and death
during bradyarrhythmia170. Chimeric Cx43 mice display heterogeneous Cx43 expression
pattern in the heart, delayed conduction, and reduced contractility171.
Cx40 is a high conductance channel required for impulse propagation in the atrial and
conduction system. During development, Cx40 is expressed strongly in the entire working
myocardium and is restricted to the atria and conduction system in adulthood. Studies on mice
lacking Cx40 have generally revealed an increased susceptibility to atrial arrhythmias, and
delayed conduction at all levels of the conduction system (SAN, intra-atrial, AVN, right
bundle branch, ventricular)172-175.
Cx45 and Cx30.2 are low conductance channels involved in propagation of the
electrical impulse in the specialized cells of the conduction system. Cx30.2 plays a role in
delaying the cardiac electrical impulse in the AVN to ensure coordinated activation of the
ventricles as mice lacking Cx30.2 have increased AVN conductance176. Cx45 is essential
during early cardiogenesis. During development, Cx45 is expressed in all regions of the heart,
however, it is down-regulated and restricted to cells of the conduction system in adulthood.
Mice lacking Cx45 develop AVN block, several cardiac and vascular defects, severe apoptosis
and die of HF in utero around E10177, 178. Mice with cardiac restricted deletion of Cx45
develop a similar phenotype as Cx45-/- mice and also die in utero around E10179.
1.3.2 Electrical remodelling and HF
Impaired cardiac conduction, membrane repolarization, automaticity, intracellular Ca2+
handling, and myocardial architecture contribute to electrical remodelling in HF. More

41
specifically, remodelling of ion channels, connexins, and proteins involved in intracellular
calcium cycling contribute to increased APD, heterogeneity of APD, prolonged QT, and
susceptibility to lethal arrhythmias in HF (Fig. 1.9; Table 1.1).
1.3.2.1 Ca+ channel remodelling
Excitation-contraction coupling and intracellular Ca2+ handling are severely
compromised in HF due to altered ICa-L, ICa-T, SERCA2a, PLB, NCX1, RyR2, and IP3R
function (Table 1.1). The Ca2+ transient amplitude and rate of decay are diminished in HF due
to abnormal SERCA2a activity. SERCA2a expression and function are reduced in HF, leading
to defective sequestration of Ca2+ into the SR180-182. Reduced PLB expression and function, as
well as increased PLB dephosphorylation in HF, also contribute to impaired SERCA2a
activity183-185. Interestingly, mutations in PLB causing dysfunctional Ca2+ sequestration have
been identified in patients with HF96. Increased forward mode NCX1 may act to compensate
for reduced SERCA2a, as NCX1 expression and activity is increased in HF, and results in a
net increase in inward current during the plateau phase of the AP186, 187. An increase in late
depolarizing current would impair repolarization and lead to AP prolongation and/or early
after depolarizations. The trigger and gain of excitation-contraction coupling are reduced in
HF due to remodelled ICa-L and RyR2 function. RyR2 expression and function are reduced and
RyR2 phosphorylation is increased182, 188. Hyperphosphorylation of RyR2 results in defective
Ca2+ release from the SR, increased diastolic Ca2+ leak, and dissociation of FK-506 binding
proteins. Diastolic Ca2+ leak could generate Ca2+ waves and cause delayed after
depolarizations. IP3R are also increased in HF and may contribute to further altered Ca2+
release from the SR189. Phosphorylation of L-type Ca2+ channel is increased, while its
expression, ICa-L and T-tubule density are reduced in HF190-192. Reduced ICa-L inactivation may

Na+
NCX
Ca2+
Ca2+
Ca2+
SR
↓SERCA
↓RyR
Ca2+
Ca2+Ca2+
Figure 1.9. Electrical remodelling of cardiac myocytes in the failing heart. Electrical coupling between cells is severely impaired in HF as the expression and phosphorylation of Cx43 is reduced, and Cx43 is extensively misslocalized. Meanwhile, several K+ currents (IK1, IK, Ito, INa-K), INa, and Ca2+ currents/channels (ICa-L, RyR, SERCA) are reduced, while other currents/channels (ICa-T, INHX, INCX, IP3R) are increased. Contraction of the myofilaments is diminished as impaired Ca2+ cycling leads to reduced [Ca2+]i available to activate myocardial contraction.
K+
Na+
↓Cx43
↓Cx43~P
Na+
NHE
H+
↑IP3R
ATPase
K+
Cx43 misslocalization
↓INa ↓ICa-L, ↑ICa-T↓INa-K ↑INCX↑INHE ↓IK1, IK, Ito
↓[Ca2+]i
PLB
P
P
P
P
42

43
Table 1.1. Remodelling of ion channels, connexins, and Ca2+ handling proteins in the failing ventricle
Protein Gene Changes in Heart Failure REF Current INa Nav1.5 SCN5A ↓F, ↓M, ↓P, ↑NaL 193-195 ICa-L Cav1.2 CACNA1C ↓F, ↓P, ↑~P h,e 190-192 ICa-T Cav3.1/ Cav3.2 CACNA1G/H ↑F e 196 Ito,f Kv4.2/Kv4.3 KCND2/3 ↓F, ↓M, ↓P h,e 197-199 Ito,s Kv1.4 KCNA4 ↓F, ↓M e 197, 200 IKr HERG KCNH2 ↔F h,e 201, 202 IKs KvLQT1 KCNQ1 ↓F h,e 202, 203 IK1 Kir2.1 KCNJ2 ↓F h,e 201, 202 Connexins Cx43 GJA1 ↓M, ↓P ↓~P, ↑ lateralization h,e 204-206 Ca2+ handling proteins
NCX SLC8A1 ↑F, ↑M, ↑P h,e 186, 187 SERCA2a ATP2A2 ↓F, ↓M, ↓P h,e 180-182 RyR2 RYR2 ↓F, ↓M, ↓P, ↑~P h,e 182, 188 IP3R2 ITPR2 ↑M, ↑ P h 189 PLB PLN ↓F, ↓M ↓~P, h,e 183-185
F=function, M=mRNA expression, P=protein expression, ~P=phosphorylation, h= human HF, e=experimental HF

44
lead to persistent late ICa-L and thus increased APD as a result of abnormal dephosphorylation
and open channel probability. Also, ICa-L displays a blunted response to continued β-
adrenergic stimulation in HF. Prolonged APD may also predispose to reactivation of ICa-L and
consequently early after depolarizations. Of note, mice with cardiac specific over-expression
of Cav1.2 develop HF207. ICa-T is re-expressed in the failing heart and could contribute to
increased spontaneous depolarisations and altered excitation contraction coupling196. The
balance between protein kinase and phosphatase activity is altered in HF and impairs
excitation contraction coupling and Ca2+ homeostasis.
As several cardiomyopathies have been linked with mutations in genes encoding
sarcomeric, cytoskeletal, and Ca2+ handling proteins, efforts aimed at re-establishing the wild-
type proteins are therapeutically attractive. Gene therapy has emerged as a potential
therapeutic tool to treat HF. SERCA2a is a likely candidate to improve cardiac contractility
and Ca2+ handling in HF, as restoring SERCA2a activity by increasing SERCA2a levels
and/or relieving the inhibition of SERCA2a mediated by PLB has proven beneficial in
experimental models of HF. 208-211 Recently, the efficacy and safety of myocardial gene
targeted increase of SERCA2a (MYDICAR) in patients with phase III or IV HF was evaluated
in the phase II clinical trial CUPID (Ca2+ up-regulation by percutaneous administration of
gene therapy in cardiac disease). MYDICAR treatment has successfully reduced and delayed
LV assist device implantation or cardiac transplantation, and improved exercise tolerance, HF
symptoms, echocardiographic measures, and NT-proBNP levels212.

45
1.3.2.2 Na+ channel remodelling
HF is associated with reduced INa, Nav1.5 mRNA and protein, increased non-
functional C-terminal splice variants, and increased late INa193-195
(Table 1.1). Interestingly,
mutations in SCN5A have been discovered in patients with HF, where impaired INa leads to
abnormal Ca2+ and Na+ homeostasis and subsequent mechanical dysfunction213. As rapid INa is
essential to normal initiation and propagation of the AP, reduced peak INa in addition to
increased late INa would result in impaired repolarization, prolonged APD, slowed conduction,
and increased susceptibility to arrhythmias.
1.3.2.3 K+ channel remodelling
HF is associated with abnormal repolarization due to reduced K+ currents (Ito, IKs, IK1)
and increased late depolarizing currents (ICa-L, INa). The most consistent ion channel
remodelling in HF is reduced Ito. Transcriptional repression plays a role in regulation of Itof as
the Kv4α subunits Kv4.2 and Kv4.3 in addition to its regulator accessory subunit KChIP2 are
reduced at the mRNA and protein levels in human and experimental models of HF197-199, 214.
Likewise, Kv1.4 mRNA and Itos is reduced in HF197, 200. IKs is also reduced in HF, while levels
of KvLQT1 mRNA and protein have not been reliably different. However, recently, reduced
KCNE1 mRNA as well as a KCNE1 variant have been linked with HF214, 215. Alterations in IKr
function or expression of HERG, or its accessory subunit MiRP1, have not been associated
with HF198, 201. Finally, IK1 is also reduced in HF, however the molecular correlates are
unknown as changes in Kir2.1 mRNA/protein levels have been inconsistent201, 202. This K+ ion
channel remodelling contributes to abnormal repolarization, prolonged APD, and development
of early after depolarizations in HF.

46
1.3.2.4 Gap junction remodelling
The abundance, distribution, and phosphorylation of Cx43 are severely diminished in
HF. Cx43 mRNA and protein expression is reduced, and Cx43 dephosphorylation and
lateralization are increased204-206. Cx43, normally localized at the intercalated disk, is
lateralized and spread uniformly around the cell membrane. Also, the association of Cx43
with zonula occludens-1 is increased in HF and correlates with mislocalization of Cx43216.
Unlike Cx43 remodelling in HF, changes in Cx40 and Cx45 levels that accompany HF have
been inconsistent205, 217, 218. Loss of Cx43 at the intercalated disk would lead to cellular
uncoupling and contribute to conduction slowing and asynchronous contraction.
1.3.3 Electrical remodelling and ET-1
ET-1 induces electrical remodelling by modulating repolarizing K+ current, Ca2+
current, gap-junctional intracellular communication, and Ca2+ handling proteins (Fig. 1.10).
1.3.3.1 Regulation of Ca2+ handling by ET-1
In the heart, ET-1 modulates Ca2+ homeostasis by altering ICa-L, ICa-T, NHE activity,
reverse mode NCX activity, and diastolic Ca2+ release via the IP3R59, 219-222. In human atrial
myocytes, ET-1 stimulates ICa-L via the ETBR when baseline current density is low and
inhibits ICa-L via the ETAR when initial current density in high219. ET-1 induces ICa-T in healthy
and failing adult rat ventricular myocytes220. In neonatal rat ventricular myocytes (NRVM), 24
hrs of 5nM ET-1 exposure induces an increase in NHE activity and subsequent increase in
reverse mode NCX activity59. In a cardiomyopathic rat model, dual ETR blockade normalized
the increased diastolic Ca2+ leak attributed to increased RyR2 and decreased FK-506 binding

Na+
Ca2+
NCX
Ca2+
Figure 1.10. ET-1 induced electrical remodelling in cardiac myocytes. ET-1 modulates the shape and propagation of the cardiac action potential by ion channel / gap junction remodelling. ET-1 disrupts electrical coupling by increasing or decreasing Cx43 and gap junctional intracellular communication. ET-1 triggers an increase in diastolic Ca2+ leak via IP3R on the SR, increases intracellular Ca2+ levels by stimulating NHX activity and thus triggering reverse mode NCX, and reduces ICa-L, Ito, IK , and IK1 and increases ICa-T.
K+Na+
NHX
H+
Ca2+
Ca2+
Ca2+
↓ICa-L, ↑ICa-T↓IK1, IK, Ito↓ ↑ Cx43 / GJIC ↑INHX
↑INCXrev
Cx43
Na+
Ca2+
SR
SERCA
RyR
Ca2+
Ca2+
IP3R
PLB
Ca2+Na+
47

48
proteins221. Lastly, ET-1 induced diastolic Ca2+ leak and spontaneous Ca2+ release events is
dependent on SR Ca2+ release by IP3R2222.
1.3.3.2 Regulation of cardiac repolarization by ET-1
ET-1 alters repolarizing K+ currents in human atrial myocytes, Xenopus oocytes,
neonatal rat ventricular mycoytes, and in a cardiomyopathic hamster model223-226. ET-1
elicites a PKC-dependent reduction in IK1 via phosphorylation of Kir2.2 channels in Xenopus
oocytes expressing the ETAR with the determinants of IK1 (Kir2.1, Kir2.2, Kir2.3) and in
human atrial cardiomyocytes223. Similarly, ET-1 induces a decrease in Ito carried by Kv1.4 or
Kv4.3 by increased PKC/calmodulin kinase II mediated phosphorylation in Xenopus
oocytes227. ET-1 also triggers a ETAR/PKC dependent biphasic response (a transient increase
followed by a decrease) on IKs in Xenopus oocytes225. In a cadiomyopathic hamster model,
electrical remodelling characterized by action potential prolongation and reduced ICa-L, Ito, IK,
and IK1 was ameliorated with ETAR blockade226. In NRVM, 1-3 days of 100nM ET-1
exposure resulted in prolonged APD, increased INCX, and reduced Ito and IK224.
1.3.3.3 Regulation of cardiac conduction by ET-1
ET-1 has been shown to alter Cx43 expression and gap-junctional intracellular
communication in neonatal rat ventricular myocytes, ovarian carcinoma cell lines, cortical
astrocytes, and osteoblatic cells228-232. In NRVM, 24 hrs of 10-1000nM ET-1 treatment caused
an ETAR/ERK dependent increase in phosphorylation and expression of Cx43 accompanied
with increased gap junctional conductance228. However, in another study, NRVM cultured
with 50nM ET-1 for 24 hrs induced a reduction in conduction velocity, with a 40% increase in

49
Cx43 protein (revealed by western blot), despite a 30% reduction in Cx43 expression via
immunofluorescence230. In ovarian carcinoma cells, ET-1 induced reduction in gap-junctional
intracellular communication and Cx43 expression is dependent on ETAR/c-Src229. Culturing
cortical astrocytes with 2-10nM ET-1 for 1-7 days caused an ETBR dependent reduction in
gap-junctional intracellular communication and Cx43 expression231. Finally, osteoblastic cells
cultured with 10nM ET-1 for 2-12 days resulted in reduced mRNA and protein expression of
Cx43 and gap-junctional intracellular communication232.
1.4 RATIONALE, HYPOTHESIS, OBJECTIVES
1.4.1 Rationale
Endothelins are vasoconstrictive peptides involved in the regulation of vascular tone
and pathophyiology of cardiovascular disease. ET-1 can also act as a pro-inflammatory
cytokine and fibrotic factor. In vitro studies have shown ET-1 induces electrical remodelling
of gap junctions, Ca2+ handling proteins, and ion channels. Our mouse model with conditional
cardiac-specific over-expression of human ET-1 will be used to investigate whether ET-1 is
acting primarily as a cytokine mediating inflammation and fibrosis or as a modulator of ion
channel/electrical properties. This model provides the opportunity to assess the role of ET-1 in
electrical remodelling in vivo and its importance to HF initiation and progression, to determine
if ET-1 induced electrical remodelling alone can trigger HF development, and to elucidate
how ET-1 induces cardiac ion channel or gap junction remodelling. Previous attempts to
prevent this model of ET-1 induced cardiomyopathy using selective and non-selective ET
receptor antagonists have not been effective. In this model of ET-1 over-expression, the ability
of ET receptor antagonists to inhibit the ET-1 cascade may be limited in states of high

50
endogenous levels of ET-1, as in HF, due to robust receptor binding kinetics. We propose to
evaluate the use of dual ECE/NEP inhibition to prevent this lethal phenotype.
1.4.2 General hypothesis: Temporal characterization of electrical remodelling will enable
prevention and progression of cardiomyopathy.
1.4.3 Hypotheses
1. Inhibition of ET-1 synthesis using an ECE inhibitor (CGS-26303) will effectively
prevent the ET-1 induced cardiomyopathy.
2. ET-1 induces electrical remodeling by disruption of Connexin-43 and Nav1.5 via
NFκB transcriptional repression.
3. This model will be reversible by the cessation of ET-1 over-expression at the onset of
electrical remodelling.
1.4.4 Objectives
1. To describe the temporal progression of the ET-1 induced electrical remodelling
a. Characterize the electrical defects using surface ECG, octapolar mapping and
programmed stimulation
b. Characterize the progression of electrical remodelling using epicardial mapping
c. Characterize the progression of structural and functional remodelling using
ultrasound biomicroscopy and invasive hemodynamics

51
2. To determine the molecular mechanism(s) leading to ET-1 induced electrical remodelling
a. Identify potential mechanim(s) of ET-1 induce electrical remodelling via western blot
and qRT-PCR for gap junctions and ion channels
b. Confirm molecular mechanism(s) of ET-1 induce electrical remodelling with an in
vitro system
c. Determine if ET-1 induced NFκB activation inhibits Cx43 and Nav1.5 transcription
3. To determine if this cardiomyopathic phenotype can be prevented by inhibiting ET-1 over-
expression
a. Determine if inhibition of ET-1 synthesis using the ECE inhibitor CGS 26303 will
prevent this model
b. Determine if this model can be prevented by inhibition of ET-1 over-expression by re-
introducing DOX at the onset of ET-1 induced electrical remodelling

52
CHAPTER 2. PHARMACOLOGICAL FAILURE OF LONG-TERM DUAL ECE-NEP
INHIBITION WITH CGS-26303 IN AN ET-1 MODEL OF CARDIOMYOPATHY

53
2.1 INTRODUCTION
The ET-1 signaling cascade is hyper-activated in HF; ET-1, BigET-1, ECE-1, and ETA
are elevated in human and animal models of HF64-67, 233. Zinc metalloproteases regulate the
production and degradation of ET-1. ECE is the principal enzyme responsible for the final
rate-limiting step of the synthesis of active ET-1 from its precursor BigET-1, while NEP
contributes to the enzymatic degradation of ET-1 (Fig. 2.1). To investigate the role of ET-1 in
the pathophysiology of cardiovascular disease, we developed a mouse model of temporal and
regional regulation of ET-1. Our lab generated transgenic mice with conditional cardiac-
specific over-expression of human big-endothelin-1 (hET-1) by crossing mice harboring the
tetracycyline transactivator (tTA) under the control of the α-myosin heavy chain promoter,
with a line harboring ET-1 regulated by a tTA-responsive, and thus doxycycline (DOX)-
regulated (DOX-off), transgene for hET-134. Previous reports in our lab have shown that this
model is associated with an inflammatory cardiomyopathy characterized by increased
cytokines, LV dilatation, and contractile dysfunction leading to HF and death 34. Selective
ETA antagonists and combined ETA/ETB antagonists have been developed to treat HF with
inconsistent results79. Previous attempts to prevent this model of ET-1-induced
cardiomyopathy using selective and non-selective ET receptor antagonists have not been
effective34. In this model of ET-1 over-expression, the ability of ET receptor antagonists to
inhibit the ET-1 cascade may be limited by high endogenous ET-1 levels, as occurs in HF, and
robust receptor-binding kinetics. CGS 26303 is a dual ECE/NEP inhibitor that inhibits ECE
with an IC50 of 410 nM. Short-term treatment with CGS 26303 in models of HF and diabetes
has improved LV hemodynamics and contractile function, and reduced cardiac fibrosis and
hypertrophy83, 84, 234. Therefore, we have hypothesized that inhibition of ET-1

prepro ET-1
Big ET-11-38
ET-11-21
ET-11-31
Furin-like enzyme
Chymase
ECE
cleared / inactivated
NEP, ETBR
ETAR ETBR
CGS 26303
Figure 2.1. Schematic of ET-1 synthesis, degradation, and treatment with CGS 26303. The inactive ET-1 precursor preproET-1 is cleaved by furin-like enzyme to generate the 38 amino acid peptide BigET-1. BigET-1 is then either converted to the mature 21 amino acid peptide ET-1 via ECE or is converted to an intermediary 31 amino acid peptide by chymase, and subsequently cleaved to ET-1 by ECE, NEP, or another protease. ET-1 is either cleared by the ETBR or is inactivated by NEP. Preventing the synthesis of mature ET-1 via dual ECE/NEP blockade with CGS 26303 may be a more effective than ET receptor antagonism in conditions with chronically high levels of ET-1.
IC50 (ECE) = 410 nMIC50 (NEP) = 1 nM
54

55
synthesis using CGS-26303, a dual inhibitor of ECE/NEP, as opposed to blocking ET-1 at the
receptor level, will be more effective at treating this model of ET-1-induced cardiomyopathy.
2.2 MATERIALS & METHODS
2.2.1 Experimental animals. Experiments conformed to protocols approved by the
University Health Network Animal Use and Care Committee and were conducted in
accordance with guidelines established by the Canadian Council on Animal Care. Binary
transgenic mice (BT:ET+tTA+) were administered DOX (200 mg/L; Sigma-Aldrich, St. Louis,
MO) in drinking water from conception until 8 wks age. Experiments were conducted in mice
maintained on DOX, withdrawn from DOX to induce gene expression, or withdrawn from
DOX and treated with CGS-26303 for 12 wks. Levels of bigET-1 and ANP were assessed
after 4 wks of treatment. After the treatment period, LV invasive hemodynamics was followed
by assessing inflammatory infiltration, ECE activity, levels of myocardial bigET-1 and plasma
ANP (Fig. 2.2).
2.2.2 Drug administration. Mice were anaesthetized with ketamine (100mg/kg; MTC
Pharmaceuticals, Cambrige, ON) and xylazine (10 mg/kg; Bayer Inc., Etobicoke, ON),
osmotic mini-pumps (model 2004; Alzet, Cupertino, CA) delivering CGS-26303 (5
mg/kg/day; Novartis, Dorval, QC) dissolved in 0.25M NaHCO3 were implanted
subcutaneously in BT mice every 4 wks for the treatment period.

12 wks ET-1 on (-DOX)
12 wks ET-1 on + CGS 26303 (-DOX)
Birth
hemodynamics, histopathology,
ECE activity, ET-1/BigET-1/ANP ELISA’s
8 wks
8 wks ET-1 off (+ DOX)
20 wks12 wks ET-1 off (+DOX)
ON DOX
OFF DOX
CGS 26303
12 wks
ET-1/BigET-1/ANP ELISA’s
Figure 2.2. Schematic of experimental design. Mice were kept on DOX from conception until 8 wks of age, then either maintained on DOX, taken off DOX, or taken off DOX and treated with CGS-26303 for 12 wks. Levels of myocardial ET-1, bigET-1 and plasma ANP were measured by ELISA 4 wks later. After the treatment period, LV hemodynamic measurements were followed by histopathology, ECE activity, and ELISA’s for myocardial bigET-1 and plasma ANP.
56

57
2.2.3 Invasive LV hemodynamics. The mice were anesthetized with Isofluorane (1.5% -
2.5%) and kept on a heating pad to maintain body temperature. The ventral aspect of the neck
was shaved; the mice were placed in a supine position. Using a dissecting microscope (MZ6
Leica, Heerbrugg, Switzerland), the right common carotid artery was exposed via a midline
incision on the ventral side of the neck. The middle segment of the common carotid artery was
dissected and two ligatures with 7-0 silk, and proximal and distally placed around it.
Temporary occlusion of the common carotid artery was achieved with a gentle traction. An
incision hole is made between the two ligatures of the artery, through which a Millar micro-tip
transducer (1.4 F sensor, 2F catheter; Millar Instruments Inc., Houston, TX) was passed
through the common carotid artery and into the LV cavity to measure arterial and LV pressure
tracings. From these tracings HR and ±dP/dT (peak positive and negative rate of LV pressure
change) were calculated. All data were analyzed using computer based acquisition system
(Sonometrics, London, ON).
2.2.4 ET-1 / BigET-1 ELISA. Ventricles were snap frozen, homogenized in 10 times wt/vol
ice cold buffer (10 mM NaCl, 10% glycerol, 1% Triton X-100, 50 mM HEPES, 1mM EDTA,
0.01 PMSF, 4.8μg/mL aprotinin, 4.8μg/mL leupeptin), centrifuged at 20,000g for 20min at
4oC, and filtered through a YM-10 membrane for 3h at 3000g at 4oC (Millipore Canada Ltd,
Etobicoke, ON). The filtrate was then used for precipitation and quantification using a
BigET-1 ELISA kit (ALPCO Diagnostics, Salem, NH). More specifically, the samples,
standards and control sample (human ET-1 / BigET-1) were added to wells pre-coated with
polyclonal sheep anti ET-1 / BigET-1 Ab. A monoclonal anti ET-1 / BigET-1 Ab conjugated
to horse radish peroxidase was then added to the wells and subsequently incubated for 4 hrs at

58
RT in the dark. Next, the plate was incubated for 30 min at RT in the dark with TMB substrate
solution. The reaction was stopped with acidic stopping solution and the absorbance was
measured at 450nm. The colour intensity is proportional to the amount of ET-1 / BigET-1 in
the samples. Sample concentrations were determined from relative standard calibration curves
generated with human ET-1 / BigET-1 samples. Levels of ET-1 peptide are found in
Appendix 1.
2.2.5 ANP ELISA. Blood (~500μl) was collected in pre-cooled tubes coated with EDTA
(VWR), stored on ice until centrifuged at 13,000g for 6min at 4oC. Plasma was separated and
stored at -80oC until assayed for levels of ANP using an ELISA kit (Cayman Chemical; Ann
Arbor, MI) that is based on the competition for binding sites of anti-rat ANP antibodies
between free ANP and rat ANP tracer (linked to acetylcholinesterase). More specifically, the
samples, standards and control samples (rat ANP) were incubated with ANP tracer and rabbit
anti-rat ANP Ab in wells pre-coated with mouse IgG against rabbit overnight at 4oC. The plate
was incubated for 30 min at RT in the dark with Ellman’s Reagent (acetylcholinesterase
substrate), followed by measuring the absorbance at 450nm. The colour intensity is
proportional the amount of tracer complexed in the well and is inversely proportional to the
amount of free ANP in the well. Sample concentrations were determined from relative
standard calibration curves generated with rat ANP samples.
2.2.6 Histopathology. Hearts were perfusion fixed with 4% paraformaldehyde, paraffin-
embedded, sectioned (5-6μm), and stained with hematoxylin-eosin. Three sections per heart

59
were scored (0-4) for cell infiltration by a blinded observer as follows: absence (0), mild (1),
mild-moderate (2), moderate (3), and extensive (4) infiltration or necrosis.
2.2.7 ECE activity. Ventricles were snap frozen, homogenized in ice cold buffer (5%
IGEPAL, 0.1M MES, and 0.2M NaCl, pH 6.0), stored at 4oC for 1 h, and centrifuged at
20,000g for 15min at 4oC. Supernatant (50μg protein) was incubated with 20μM of
fluorogenic ECE substrate (Mca-RPPGFSAFK(Dnp)-OH; R&D systems, Minneapolis, MN)
in 100μL of reaction mixture (0.1mol/L MES, and 0.2 mol/L NaCl, pH 6.0) for 1h at 37 °C.
The rate of the reaction was analyzed by reading excitation at 320nm and emission at 405nm
every 5 min for 1h. An in vitro validation of CGS-26303 was performed by determining ECE
activity in myocardial proteins isolated from a C57BL/6 mouse with an increasing gradient of
CGS-26303 (Fig. 2.3).
2.3 RESULTS
2.3.1 Short term treatment with CGS-26303 inhibits ECE and NEP activity. To determine
if ECE inhibition could prevent the cardiomyopathic phenotype of mice over-expressing ET-1,
BT mice were treated with the dual ECE/NEP inhibitor CGS-26303 for 12 wks. All mice were
maintained on DOX until 8 wks of age. Following this, groups of mice were either maintained
on DOX, withdrawn from DOX, or withdrawn from DOX and treated with CGS-26303 for 12
wks (Fig. 2.2). To confirm the ability of CGS-26303 to inhibit myocardial ECE, CGS-26303
dose-dependently blocked the ex vivo ECE activity of myocardial protein extracts from a
C57BL/6 mouse (Fig. 2.3). To assay if CGS- 26303 was inhibiting ECE/NEP activity in vivo,
myocardial bigET-1 and plasma ANP levels were

Ch
ang
e in
Flu
ore
scen
ce/m
in
-0.5
0.0
0.5
1.0
1.5
2.0
2.5
3.0
Time (m in)
0 10 20 30 40 50
EC
E a
ctiv
ity
-20
0
20
40
60
80
100
Figure 2.3. Ex vivo validation of CGS 26303. ECE activity recorded every 4 min for an hour with an increasing gradient of CGS 26303 (A) and dose-dependent reduction in rate of ECE activity (B) of myocardial proteins isolated from a C57BL/6 mouse.
A
B
0 g0.625 g 1.25 g 2.5 g 5 g 10 g
0 g0.125 g0.625 g2.5 g5 g10 g
60

61
assessed after 4 wks of treatment. As expected, inhibition of ECE/NEP with CGS-26303
treatment caused elevated levels of bigET-1 and ANP (Fig. 2.4A,B), indicating that CGS-
26303 was preventing conversion of bigET-1 to ET-1 and inhibiting ANP degradation.
2.3.2 Long term treatment with CGS-26303 fails to prevent cardiomyopathic phenotype.
At the end of the 12 wks treatment period, invasive hemodynamic analysis was performed to
determine if long-term ECE/NEP inhibition would preserve hemodynamic and contractile
function and/or exhibit any potential survival benefit. Consistent with previous results34, BT
mice withdrawn from DOX developed a significant decline in systolic and diastolic blood
pressures (Fig. 2.5A) and in LV contractile function (Fig. 2.5B), with a significant increase in
inflammatory cell infiltration (Fig. 2.6D). Animals treated with CGS-26303 also manifest the
deterioration in hemodynamic and LV contractile parameters observed in BT mice withdrawn
from DOX (Figs. 2.5A,B); in other words, treatment with CGS-26303 did not result in any
discernable hemodynamic or contractile benefit. Similarly, treatment with CGS-26303 did not
prevent inflammatory cell infiltration (Fig. 2.6D). Finally, no significant survival benefit was
observed in mice treated with CGS-26303 (Fig. 2.7).
To examine if CGS-26303 continued to inhibit endogenous ECE/NEP activity
throughout the entire study period and limit ET-1 production and ANP metabolism, ECE
activity as well as myocardial tissue levels of bigET-1 and plasma levels of ANP were assayed
(Figs. 2.6A,B,C). As expected, DOX withdrawal led to significantly increased levels of
bigET-1 (Fig. 2.6A), and increased levels of ANP and ECE activity (Figs. 2.6B,C). However,
unlike the results observed after 4 wks of treatment, 12 wks of CGS 26303 treatment no
longer prevented the hydrolysis of bigET-1 or the metabolism of ANP (Figs.

AN
P (
pg
/ml)
0
200
400
600
A BB
ig E
T-1
(fm
ol/
ng
)
0
20
40
60
*
*
*
Figure 2.4. Short term treatment with CGS-26303 inhibited ECE and NEP activity. Four weeks of treatment with CGS-26303 led to an accumulation of myocardial bigET-1 and plasma ANP (B). *=P<0.05.
ON DOX (ET-1 off)
OFF DOX (ET-1 on)
CGS 26303 (ET-1 on + treatment)
62

mm
Hg
/s
0
1000
2000
3000
mm
Hg
0
20
40
60
80
100A
B
**
****
**
** **
**
**
**
SBP DBP LVSP
dP/dt-dP/dt+
Figure 2.5. Long term treatment with CGS-26303 fails to preserve hemodynamic and LV contractile integrity in BT mice. Mice over-expressing hET-1 for 12 wks had reduced arterial/LV pressures (A) and LV contractility (B). Treatment with CGS-26303 did not prevent hemodynamic or LV contractile dysfunction. SBP/DBP = arterial systolic/diastolic blood pressure, LVSP = LV systolic blood pressure, dP/dt = peak positive and negative first derivatives of the LV pressure. **=P<0.01
ON DOX (ET-1 off)
OFF DOX (ET-1 on)
CGS 26303 (ET-1 on + treatment)
63

AN
P (
pg
/ml)
0
200
400
600
800
1000
1200B
ig E
T-1
(fm
ol/
ng
)
0
10
20
30
Ch
ang
e in
Flu
ore
scen
ce/m
in
0
1
2
3
4
5
6In
fla
mm
ato
ry S
core
0
1
2
3
4
A B
DC ****
**
**
Figure 2.6. Long term treatment with CGS-26303 fails to inhibit ECE and NEP activity. Myocardial bigET-1 levels (A), plasma ANP levels (B), rate of ECE activity (C), and inflammatory infiltration (D) in BT mice after 12 wks of treament. *=P<0.05, **=P<0.01
ON DOX (ET-1 off)
OFF DOX (ET-1 on)
CGS 26303 (ET-1 on + treatment)
64

Days Post DOX Removal
40 50 60 70 80 90
% S
urvi
val
60
70
80
90
100
on DOX (n=9)
off DOX (n=11)
CGS 26303(n=12)
Figure 2.7. Therapy with CGS-26303 does not improve survival in BT mice. Kaplan-Meier survival curves in BT mice. Mice were maintained on DOX until 8 weeks of age (Day 0), then either continued on DOX, withdrawn from DOX, or withdrawn from DOX and treated with CGS-26303.
65

66
2.6A,B). Surprisingly, CGS-26303 treatment resulted in further increases in endogenous ECE
activity (Fig. 2.6C). Consistent with this, long term treatment with CGS-26303 failed to
restrict ET-1 synthesis, ANP metabolism and ECE activity.
2.4 DISCUSSION
Effectively blunting the ET-1 signaling cascade with ET receptor antagonists has not
been successful. Antagonizing ET-1 signaling via ECE inhibition, thereby preventing the
synthesis of mature ET-1, was thought to represent a better a treatment strategy. The dual
ECE-NEP inhibitor CGS-26303 has proven to be beneficial in many short-term studies;
improving systolic blood pressure and heart to weight ratio in a rat model of diabetes234;
decreasing preload and afterload, increasing cardiac output, and reducing LV hypertrophy,
dilatation, and cardiac fibrosis; and improving systolic blood pressure, LV fractional
shortening, LV perivascular fibrosis, and reducing mRNA levels of collagen type I/ III and
heart to body weight ratio and levels of ET-1 in rat models of HF83, 84. However, whether long
term treatment with CGS-26303 is effective in states of chronically high levels of ET-1, as in
clinical HF, is unknown. In our study, we wanted to determine if treatment with CGS-26303
could prevent our ET-1-induced model of cardiomyopathy.
ECE inhibition failed to prevent our ET-1 induced model of cardiomyopathy. This
may have resulted from indirect effects of ECE/NEP inhibition, or the production of active
ET-1 by CGS-26303-insensitive ECE activity, or failure of CGS-26303 to provide effective
ECE/NEP inhibition after 12 wks of treatment.
Although ECE is the main enzyme involved in ET-1 synthesis, chymases and other
metalloenzymes are able to hydrolyze bigET-1 into active ET-1235. It is possible that other

67
enzymes compensate for the lack of ECE activity, particularly in conditions of high bigET-1
levels; this would result in elevated levels of ET-1 and continued pathophysiological
responses. Whether or not these alternate methods of ET-1 production played a role our model
was beyond the scope of this study.
As CGS-26303 is a more potent inhibitor of NEP (IC50=1nM) than ECE-1
(IC50=410nM), and NEP is involved in an alternate route of ET-1 production and is also
responsible for ET-1 metabolism 235, 236, altering NEP activity could significantly alter the
balance between ET-1 production and degradation. As NEP activity was not inhibited in CGS-
26303 treated mice (Fig. 2.6B), NEP could have played a role in alternative ET-1 synthesis,
and thus limit the effectiveness ECE inhibition by CGS-26303 in this model.
Dose, tissue absorption, and myocardial bigET-1 accumulation may have contributed
to the lack of effectiveness of CGS-26303. The mice received CGS-26303 via mini-pumps at
5mg/kg/day, a dose that prevented ECE and/or NEP activity after 4 wks of treatment in this
study and others237. Scar tissue may have hampered the tissue absorption of the drug, as fresh
mini-pumps were re-implanted every 4 wks into the same subcutaneous area for the duration
of the study. In human and bovine endothelial cells treated with CGS-26303, high levels of
bigET-1 triggered an increase in ECE-1 mRNA, protein and promoter activity238. This study
suggests that chronic treatment with CGS-26303 may lead to reduced responsiveness.
Therefore, it is possible that 12 wks of CGS-26303 treatment and subsequent reduced
therapeutic response in our study was caused by the increase in ECE activity (Fig. 2.6C)
triggered by the accumulation of bigET-1.
In conclusion, we have shown that long-term treatment with CGS-26303 in conditions
with high endogenous levels of bigET-1, as seen in our model of ET-1 induced

68
cardiomyopathy, is unable to effectively inhibit ECE. These data suggest that chymases and/or
other metalloproteases act as alternate methods of ET-1 production, and that the accumulation
of bigET-1 may lead to increased CGS-26303-insensitive ECE activity, circumventing the
efficacy of CGS-26303 (Fig. 2.8). Further studies are necessary to determine if a more
selective ECE inhibitor or a combination of ECE inhibition/ET-1 receptor blockade would be
a more effective treatment strategy.
Acknowledgments: I would like to thank Dr. Abdul Momen (Husain Lab) and Dr. Golam
Kabir (Lewar Centre) for acquiring the invasive LV hemodynamic data, Dr. Almuktafi Sadi
for scoring the cell infiltration data, and Novartis (Summit, NJ) for generously supplying the
CGS-26303.

Figure 2.8. Model depicting short vs. long term treatment with dual ECE-NEP inhibitor. (A) Four weeks of CGS-26303 treatment led to inhibition of ECE and NEP activity, resulting in high levels of BigET-1 and ANP peptides. (B) Twelve weeks of CGS-26303 treatment led to an accumulation of BigET-1 peptide that triggered an increase in expression and activity of ECE, thus ultimately restoring the production of chronically high levels of mature ET-1.
↑ ECE-1 mRNA
ET
A
ET
B
ET-1
↑ ECE activity & ET-1 production
BigET-1
BigET-1 BigET-1
BigET-1
ET-1 ET-1
ET-1 ET-1
ET-1
ET-1ET-1
ET-1ET-1
ET-1ET-1
ECE-1 ECE-1
ECE-1ECE-1 ECE-1
ECE-1
ECE-1ECE-1
ECE-1
ECE-1
ET
A
ET
B
BigET-1
BigET-1
BigET-1
ET-1 ET-1
ECE-1ECE-1 ECE-1 ECE-1 ECE-1
A
B
CGS 26303 CGS 26303
CGS 26303
CGS 26303
ANPBigET-1
BigET-1 BigET-1BigET-1
BigET-1 ANPANP
ANP
ANP
ANP
CGS 26303CGS 26303
ANPANP
BigET-1
ECE-1
69

70
CHAPTER 3. ET-1 INDUCED ELECTRICAL REMODELLING PRECEDES LV
DYSFUNCTION IN ET-1 INDUCED CARDIOMYOPATHY

71
3.1 INTRODUCTION
Mutations in genes encoding ion channels are consistently linked with various rhythm
disorders such as BS and LQT, while mutations in genes encoding sarcomeric and
cytoskeletal proteins are typically associated with contractile dysfunction and HF
development. However, mutations in the Na+ channel SCN5A, the K+ channel β-subunit
KCNE1, and the SERCA2a regulatory protein PLN have also been associated with myocardial
dysfunction and HF96, 213, 215. Thus, it appears abnormal Ca2+ handling and ion channel
function alone can predispose/trigger HF development.
ET-1 is a potent vasoconstrictive peptide that is implicated in the pathogenesis of
several cardiovascular diseases such as HF and atherosclerosis239. ET-1 acts in an autocrine or
paracrine fashion to cause inflammation, fibrosis, cardiac hypertrophy and electrical
remodelling240. ET-1 induces electrical remodelling by modulating repolarizing K+ currents,
Ca2+ currents, gap junctions intracellular communication, and Ca2+ handling proteins59, 220, 223,
228, 230. Our model of conditional cardiac specific over-expression of ET-1 is associated with
an inflammatory cardiomyopathy characterized by increased cytokines, LV dilatation,
contractile dysfunction, progressive widening of QRS, HF and death following DOX
withdrawal34. What is not known is whether ET-1 acts primarily as a cytokine mediating
inflammation and fibrosis or as a modulator of ion channel/electrical properties. We posit that
this model provides a unique opportunity to assess the role of electrical remodelling in the
initiation and progression of HF.
The first objective of this chapter is to describe the temporal progression of the ET-1
induced electrical remodelling by using surface ECG, octapolar mapping, and programmed
stimulation to characterize the electrical defects; using epicardial mapping to characterize the

72
progression of electrical remodelling; and using ultrasound biomicroscopy and invasive
hemodynamics to characterize the progression of structural and functional remodelling. The
second objective of this chapter is to determine if this model can be prevented by inhibition of
ET-1 over-expression by re-introducing DOX at a stage when only electrophysiological
abnormalities are manifest.
3.2 MATERIALS & METHODS
3.2.1 Experimental animal. Experiments conformed to protocols approved by the University
Health Network Animal Use and Care Committee and were in accordance with guidelines of
the Canadian Council on Animal Care. BT and NBT mice were administered DOX (200
mg/L) in drinking water from conception until weaning (3 wks old). Experiments were
conducted in mice withdrawn from DOX for 4 wks (group I), 8 wks (group II), or after 4 wks
of DOX removal with DOX re-administered for an additional 4 wks (group III, Fig. 3.1).
Following acquisition of electrophysiological, hemodynamic, ultrasound biomicroscopy, or
epicardial mapping data, mice were sacrificed by cervical dislocation. Hearts were excised,
rinsed in PBS and snap frozen for RNA/protein extraction for qRT-PCR and western blot
analysis, or perfusion-fixed for histopathology and immunohistochemistry.
3.2.2 Surface ECG and intracardiac electrophysiological evaluation. Mice were
anesthetized with i.p. Ketamine 100mg/kg (MTC Pharmaceuticals, Cambridge, ON) and
Xylazine 10 mg/kg (Bayer Inc., Etobicoke, ON) and kept on a heating pad to maintain body
temperature. Using a dissecting microscope (MZ6 Leica, Heerbrugg, Switzerland), the
external jugular vein was isolated, and two ligatures were placed at the distal area of the

intra-cardiac and epicardial mapping,
hemodynamics and UBM, Histopathology\
epicardial mapping, hemodynamics and UBM,
histopathology
Birth
Weaning (3 wks of age)
0 wks
4 wks
8 wks
Group I
Group II
Group III
4 wks ET-1 on(- DOX)
8 wks ET-1 on(- DOX)
4 wks ET-1 on(- DOX)
4 wks ET-1 off(+ DOX)
8 wks
epicardial mapping, hemodynamics and UBM,
histopathology
ET-1 off(+ DOX)
Figure 3.1. Schematic of experimental design. Mice were kept on DOX from conception until 3 wks of age, then either withdrawn from DOX for 4 wks (Group I), for 8 wks (Group II), or withdrawn from DOX for 4 wks followed by DOX re-administration for an additional 4 wks (Group III). After the treatment periods, terminal experiments (intra-cardiac/epicardial mapping, ultrasound biomicroscopy, or LV invasive hemodynamics) were followed by histopathology.
73

74
isolated vessel. The vessel was tied off at the proximal ligature and tied down to expose the
area between the two ligatures. A small incision was made between the two ligatures of the
external jugular vein. A 2-French octapolar electrode catheter (NuMED Inc., Hopkinton, NY)
was introduced into the hole, and passed into the right atrium, across the tricuspid valve, and
into the right ventricle. The distal ligature was then tied off around the catheter. Surface
electrocardiograms (lead II) were simultaneously recorded along with intracardiac
electrograms from the 8 electrodes spanning the atrium, His bundle, and ventricle (Fig. 3.2A).
Atrial-His (AH), His-ventricular (HV), RR, PR, QRS, and RV-LV intervals (from the onset of
RV activation in intracardiac electrograms to the end of the QRS complex in the surface ECG)
were calculated from these recordings. AV Wenckebach (AVWENK), 2:1 AV block cycle
lengths (AV2:1), and AV effective refractory periods (AVERP) were acquired during right atrial
pacing. AVWENK was obtained by pacing with 12 decremental atrial driving stimuli, beginning
at a cycle length 20 ms less than the RR interval. Decremental pacing was continued until
AV2:1 developed. Next, AVERP was acquired by pacing with 12 atrial driving stimuli, at a
cycle length of 20 ms less than RR interval, and adding a 9th decremental stimulus until
ventricular activation was blocked. VT/ ventricular fibrillation (VF) induction was examined
via right ventricular over-drive pacing and right ventricular pacing with 12 basic stimuli
followed by up to 3 extra decremental test steps. Heart weight to body weight ratio (HW/BW)
was calculated as a measure of hypertrophy.
3.2.3 Epicardial mapping. Mice were anesthetized with Isoflurane (1.5%) and ventilated
with air. Body temperature was maintained by a heating pad. Thoracotomy was performed to
enable the application of a flexible 64 micro-electrode array (4x16) with an inter-electrode

Genotype AVWENK AV2:1 AVERP
(ms)
BT (n=7) 85.93.0 ** 64.83.2 ** 62.63.8 **
NBT (n=10) 68.12.1 51.91.9 45.12.3
Genotype HW / BW(mg/g)
AH HV PR QRS RV-LV
(ms)
BT (n=8) 6.40.5 * 62.13.5 ** 13.71.3 79.13.2 ** 17.62.2 ** 16.71.6 **
NBT (n=7) 4.90.2 45.93.8 11.41.1 62.34.7 9.30.4 11.10.8
A
C
B
BTV
AH
NBT
V
A H
Figure 3.2. Electrophysiological evaluation using intracardiac mapping reveals electrical defects in mice over-expressing ET-1 after 8-10 weeks of transgene induction. (A) Representative His bundle electrograms in BT vs. NBT mice. (B) Baseline conduction intervals, surface electrocardiogram parameters, HW/BW, and RV-LV conduction times in BT vs. NBT mice. (C) AVWENK, AV2:1, and AVERP in BT vs. NBT mice during RA pacing. *=P<0.05, **=P<0.01.
75

76
spacing of 800 µm on the epicardial surface (Fig. 3.3A). The array was positioned around the
heart in 2 orientations (Fig. 3.3B,C). Unipolar electrograms were acquired with custom made
software at 5k samples/s per channel, resulting in a time resolution of 0.2 ms, filtered with a
high- and low-pass filters of 0.5Hz and 1kHz, and amplified with a gain of 500 using
MEA1060 (Multi Channel Systems, Reutlingen, Germany). The sampling frequency was
sufficient to determine local activation time and generate accurate activation maps. Custom
made software (designed by Stéphane Massé; Nanthakumar lab) was used to measure atrial-
to-ventricular activation time (from onset of right atrial activation to onset of ventricular
activation evaluated from one heart beat; Fig. 3.3B); ventricular activation time (from earliest-
to-latest ventricular activation evaluated from one heart beat) and ventricular activation with
repolarization time (from earliest ventricular activation to latest ventricular repolarization
acquired from one heart beat; Fig. 3.3C). Estimates of ventricular repolarization were based
on previous studies that found activation recovery intervals (interval between the minimum
derivative of the QRS and maximum derivative of the T wave in unipolar electrograms) were
related to transmembrane APD241. Matlab (version 2007b) was used to calculate the maximum
negative rate of change of voltage in ventricular electrograms from one heart beat (-dV/dt),
and to produce isochronal activation maps to illustrate the spread of ventricular activation
from earliest-to-latest activation (blue-red) for one heart beat. The moment of local activity
was determined as the time at which the electrogram signal increased significantly above
background noise.
3.2.4 Invasive LV hemodynamics. As found in section 2.2.3.

D BT-d
V/d
t (V
/s)
0
20
40
60
80
100
Ve
ntr
icu
lar
Ac
tiva
tio
n T
ime
(m
s)
0
5
10
15
20
Ear
lies
t A
tria
l-to
-Ven
tric
ula
r A
ctiv
atio
n T
ime
(ms)
0
20
40
60
80
Ven
tric
ula
r A
ctiv
atio
n &
Rep
ola
risa
tio
n T
ime
(ms)
0
10
20
30
40
50
1 2 3 4 5 6 7 8 9 10 11 12 13 14 151
2
3
4
A
1 2 3 4 5 6 7 8 9 10 11 12 13 14 151
2
3
4
NBT
Activation time (ms)
E BTNBT**
LVRV
1 2 3 4 5 6 7 8 9 10 11 12 13 14 151
2
3
4
B C
I II III
LVRVLVRV
Figure 3.3. Temporal progression and prevention of electrical remodeling in mice over-expressing ET-1. Illustration of 4x16 micro-electrode array (A), and placement of array while recording epicardial electrograms (B,C). (D) Ventricular isochronal activation maps in BT vs. NBT mice in group II accompanied by representative epicardial electrograms from micro-electrodes located at 4.5 and 4.14. (E) Earliest atrial-to-ventricular activation time, ventricular activation time, ventricular activation with repolarization time, and -dV/dt measured in sinus rhythm in BT and NBT mice in group I (BT: n=14, NBT: n=24), II (BT: n=21 NBT: n=42), and III (BT: NBT: n=7). *=P<0.05 **=P<0.01.
5 10 15 20 25
**
**
**
**
* **
77

78
3.2.5 Ultrasound biomicroscopy. A Vevo 770 (VisualSonics Inc., Toronto, ON) ultrasound
biomicroscope with a transducer frequency of 30 MHz was used to acquire M-mode and
Doppler recordings in mice anesthetized with 1.5% isoflurane. M-mode was used to evaluate
the change in LV diameter between systole and diastole to determine fractional shortening
(FS%). Doppler was used to assess the velocity of blood flow in the main pulmonary artery
and aorta, and the ratio between early (E: early diastolic filling) and late (A: atrial
contraction) ventricular filling velocity at the mitral valve (LV E/A) and at the tricuspid valve
(RV E/A).
3.2.6 Histopathology. 4µm sections from perfusion-fixed paraffin-embedded hearts at the
level of the papillary muscle were stained with Celestin blue for 5min, counterstained in
hematoxylin, and stained in Picro-Sirius Red for 30 min to colour all collagen fibres red.
Fibrosis was quantified in three endocardial areas of the LV free wall from each section with
Aperio ImageScope using the v9 algorithm (version 10.2.2.2319; Aperio Technologies) to
quantify the area of positively stained pixels. Cardiomyocyte cross-sectional area was
measured at the level of the nucleus using Infinity Analyze version 5.0.2 (Lumenera
Corporation, Ottawa, ON) as a measure of cellular hypertrophy.
3.2.7 Statistical analysis. Data are presented as mean + SEM. ANOVA, and t-tests were
performed using SPSS version 12.0 (Chicago, IL, USA).

79
3.3 RESULTS
3.3.1 Electrical defects in mice with cardiac-specific ET-1 over-expression. All
experimental groups studied and assays undertaken are summarized in Fig. 3.1. To explore the
electrophysiological basis for the QRS prolongation noted in our ET-1 over-expression
model34, intracardiac mapping with an octapolar catheter via transjugular insertion to the RV
was performed in BT and NBT controls 8 wks after withdrawal from DOX at weaning (Fig.
3.2A). Confirming previous findings, PR, QRS, and HW/BW were increased in BT vs. NBT
mice (Fig. 3.2B). Invasive His bundle electrograms revealed BT mice have slowed AH
conduction, without affecting HV intervals (Fig. 3.2B). Prolonged QRS in BT mice without
significant changes in HV conduction were explained by prolonged RV-LV conduction time
in BT vs. NBT littermates (Fig. 3.2B). To investigate the basis of prolonged AH times in BT
mice, we performed right atrial pacing and determined that AVWENK, AV2:1, and AVERP cycle
lengths were prolonged in BT mice (Fig. 3.2C). These data suggest that the extensive AV
nodal and inter-ventricular conduction abnormalities are associated with chronic cardiac ET-1
over-expression.
3.3.2 Electrical remodelling is triggered as early as 4 wks after ET-1 over-expression.
Catheter size limitations in our system restrict intracardiac mapping to mice greater than 20 g
body weight. As such, this technique was not applicable in younger animals. Instead, a
flexible multi-electrode array was wrapped around the heart enabling epicardial mapping and
characterization of the progression of specific electrophysiology parameters in ET-1 over-
expressing mice as young as 4 wks post weaning (Fig. 3.3). Earliest atrial-to-ventricular
activation time, ventricular activation time, ventricular activation and repolarization time,

80
and -dV/dt were studied in sinus rhythm and during pacing. Significant ventricular conduction
delays were observed as early as 4 wks post DOX withdrawal (Group I). Both ventricular
activation time (9.40.3 vs. 8.30.3 ms, P0.01) and ventricular activation with repolarization
time (23.30.8 vs. 17.60.3 ms, P<0.01) were prolonged and -dV/dt was reduced (80.32.5 vs.
89.63.2 V/s, P<0.05) in BT vs. NBT mice in sinus rhythm (Fig. 3.3E) and after pacing for
160 ms (Table 3.1).
Consistent with data obtained from intracardiac catheters, BT mice 8 wks after DOX
withdrawal (Group II) manifest more substantive abnormalities. Earliest atrial-to-ventricular
activation time (76.87.6 vs. 48.22.0 ms, P<0.01), ventricular activation time (15.51.4 vs.
8.40.3 ms, P<0.01), as well as ventricular activation and repolarization time (37.73.5 vs.
19.30.5 ms, P<0.01) were all prolonged, and -dV/dt (25.24.4 vs. 88.22.7 V/s, P<0.01) was
reduced, in BT vs. NBT mice in sinus rhythm (Figs. 3.3D,E) and after pacing for 160 ms
(Table 3.1). Neither VT nor VF was observed in any mice tested. Together, these results
suggested that ET-1 mediates a progressive decline in ventricular conduction.
3.3.3 HF develops by 8 weeks after ET-1 over-expression. Ultrasound biomicroscopy and
LV hemodynamics were used to study heart function in mice 4-8 wks post-DOX withdrawal.
SBP, LVSP, dP/dt+, and dP/dt- were assessed by Millar catheters (Table 3.2), while peak
velocity of blood flow in the main pulmonary artery and aorta, peak RV and LV E/A ratios
and FS% were evaluated by ultrasound biomicroscopy (Table 3.3). No obvious
structural/functional differences were observed in BT mice after 4 wks of ET-1 over-
expression (Group I). By contrast, and consistent with ‘clinical’ evidence of HF, BT mice
exhibited severe structural and functional abnormalities by 8 wks of transgene induction

81
Table 3.1. Temporal progression and prevention of electrical remodelling in mice over-expressing ET-1 during pacing
Parameter
Group I
Group II
Group III
BT (n=7) NBT
(n=18)
BT
(n=9)
NBT
(n=28)
BT
(n=7)
NBT
(n=7)
V Activation (ms) 27.6±3.2** 19.6±1.3 44.2±13.5* 17.6±0.5 18.0±1.0 18.4±0.8
V Activation &
Repolarization (ms) 30.9±1.3** 25.7±0.3 50.6±8.8** 26.6±0.5 26.0±0.9 26.4±1.2
-dV/dt (V/s) 66.2±3.2* 87.9±5.8 28.3±4.7** 76.6±4.7 105.2±9.1 105.3±8.7
*=P<0.05, **=P<0.01, V= ventricular

82
Table 3.2. Progression and prevention of cardiac structural and functional abnormalities as evaluated by invasive hemodynamics
Parameter
Group I
Group II
Group III
BT
(n=5)
NBT
(n=10)
BT
(n=6)
NBT
(n=4)
BT
(n=3)
NBT
(n=4)
HW/BW
(mg/g) 4.78±0.19 4.91±0.10 8.24±0.66* 5.83±0.23 7.47±0.95 6.50±0.33
SBP
(mmHg) 99.6±4.7 91.9±2.1 78.7±1.5** 96.9±3.3 99.7±2.5 107.0±4.7
LVSP
(mmHg) 108.6±8.5 94.9±3.2 66.5±7.1* 94.2±2.0 110.6±1.3 110.7±5.2
+dP/dt (mmHg/s) 2255±170 2572±146 1214±222** 2778±180 2452±128 2548±96
-dP/dt (mmHg/s) 2703±207 2872±145 1578±246** 3198±42 3422±298 3585±183
*=P<0.05, **=P<0.01, HW/BW = heart weight body weight ratio, SBP = arterial systolic blood pressure, LVSP
= LV systolic blood pressure, dP/dt = peak positive and negative first derivatives of the LV pressure

83
Table 3.3. Progression and prevention of cardiac dysfunction as evaluated by ultrasound biomicroscopy
Parameter
Group I
Group II
Group III
BT
(n=6)
NBT
(n=6)
BT
(n=6)
NBT
(n=6)
BT
(n=4)
NBT
(n=4)
Peak Velocity of
Main Pulmonary
Artery (mm/s)
702±14 735±41 404±38** 686 ± 68 815±45 742±36
Peak Velocity
of Aorta (mm/s) 945±39 790±70 655±91** 944±35 980±28 1057±82
RV E/A 0.60±0.03 0.67±0.12 6.54±0.97* 0.59±0.02 0.68±0.05 0.68±0.01
LV E/A 1.68±0.09 1.59±0.15 10.77±2.17** 1.55±0.08 2.02±0.23 1.60±0.13
FS% 28.2±0.7 29.4±1.4 14.4±2.4** 27.0±1.7 26.3±1.1 24.8±2.1
*=P<0.05, **=P<0.01, RV = right ventricle, E/A = ratio of peak early to atrial diastolic inflow velocities, FS =
fractional shortening

84
(Group II, Tables 3.2,3.3). LV systolic dysfunction was exemplified by reduced systolic BP
(78.71.5 vs. 96.12.4 mmHg, P<0.01), peak velocity of aortic flow (65591 vs. 94334
mm/s, P<0.01), FS% (14.42.4 vs. 27.01.7, P<0.01), and dP/dt+ (1214222 vs. 2728137
mmHg/s, P<0.01) (Tables 3.2,3.3). LV diastolic dysfunction was manifest as reduced dP/dt-
(1578246 vs. 319824 mmHg/s, P<0.01), and elevated peak LV E/A ratio (10.82.2 vs.
1.50.1, P<0.01) (Tables 3.2, 3.3). Significant RV systolic (reduced flow in the main
pulmonary artery; 40438 vs. 68668 mm/s, P<0.01) and diastolic dysfunction (elevated peak
RV E/A ratio; 6.51.0 vs. 0.60.0, P<0.01) was also observed (Table 3.3). Consistent with
these results, histopathology of heart sections from mice 4, and 8 wks post ET-1 over-
expression showed that fibrosis (6.4±0.5 vs. 1.1±0.1 %, P<0.01) and hypertrophy (3793±74 vs.
3540±69 µm2, P<0.01) were present only after 8 wks of ET-1 over-expression (Fig. 3.4; BT:
n=223 vs. NBT: n=244 cells).
3.3.4 Inhibiting ET-1 expression at the onset of electrical remodelling prevents
progression to HF. To determine if inhibiting synthesis of ET-1 at the onset of ET-1-induced
electrical remodelling will be effective in preventing the lethal phenotype, DOX was re-
introduced 4 wks after transgene induction and maintained for 4 wks. At the end of this period,
epicardial mapping, ultrasound biomicroscopy, hemodynamics, and histopathology were used
to evaluate the effects of therapy. Re-introducing DOX at the onset of electrical remodelling
completely prevented the development of the electrical, structural and functional decline seen
with age-matched BT mice maintained off DOX (Figs. 3.2-3.4, Tables 3.1-3.3). These data
suggest that in this unique model, electrical remodelling precedes overt manifestations of HF
and that the latter may be prevented by reversal of the former.

% F
ibro
sis
0
2
4
6
BT NBT
I II III
I
II
III
A
B**
50 µm50 µm
Figure 3.4. Myocardial fibrosis appears after 8 wks of ET-1 over-expression. Representative picrosirius red-stained heart sections (A) and % fibrosis (B) from BT and NBT mice in groups I-III (n=4-5 per genotype per group). **=P<0.01
85

86
3.4 DISCUSSION
Cardiac electrical remodelling has traditionally been considered a consequence of
HF242. Here, we present evidence that cardiac-specific over-expression of the vasoactive
agent ET-1 induces a progressive systemic cardiac conduction defect prior to the development
of HF. We have shown that as little as 4 wks of ET-1 excess in the heart can trigger slowed
ventricular conduction, while the overall structure and contractile function of the heart
remains essentially normal. If the ET-1 over-expression is left ‘on’, the scope and severity of
the cardiac electrical dysfunction progress and overt HF develops by 8 wks of ET-1
over-expression. Importantly, turning ‘off’ ET-1 over-expression at the onset of electrical
remodelling was able to reverse the electrical dysfunction and prevent the development of HF.
Electrical remodelling as a trigger for HF development as yet has not been firmly
established. Studies have suggested that left bundle branch block may mediate LV
dysfunction242, however, the molecular mechanism is unknown. Gap junction/ion channel
remodelling leading to mechanical dysfunction in the heart represents a novel concept in the
pathogenesis of HF. As electrical remodelling precedes structural and functional remodelling
(Fig. 3.5), and as these events were reversible by suppressing ET-1 over-expression, our
model supports the view that electrical remodelling can contribute to HF. The alternative
interpretation is that ET-1 induces electrical remodelling first, and subsequently causes
cardiac structural and functional deterioration through an entirely unrelated mechanism. While
ET-1-induced cytokine expression may also play a role in the pathogenesis of HF in our
model34, the fact that the molecular changes described here preceded any evidence of
contractile dysfunction obtained using highly sensitive measures argues against later-stage
inflammation being the only mechanism involved.

Figure 3.5. Model showing effects of 4 vs. 8 wks of ET-1 induction. Electrical remodeling, as seen in Group I mice (A), precedes structural/functional deterioration, as seen in Group II mice (B).
ET
A
ET
B
A
B
ET
A
ET
B
Ventricular DelayReduced –dV/dt
Cell hypertrophyFibrosisInflammationHemodynamic & Contractile declineAV delayVentricular DelayReduced –dV/dt
ET-1ET-1 ET-1ET-1
ET-1ET-1ET-1
ET-1
ET-1
ET-1 ET-1
ET-1ET-1 ET-1ET-1
ET-1ET-1
ET-1ET-1
ET-1
ET-1ET-1
87

88
Recently, mutations in Scn5a have been described leading to dilated cardiomyopathy
with atrial/ventricular arrhythmia and impaired Ca2+ and Na+ homeostasis with mechanical
dysfunction213. Additionally, mice expressing a mutant form of Csx/Nkx2.5 under the -MHC
promoter develop conduction defects with down-regulation of Cx40/Cx43 and HF243.
Although not emphasized in their report, the sequence of events again suggests that
conduction defects develop prior to any structural remodelling.
Acknowledgments: I would like to thank Stéphane Massé (Nanthakumar lab) for providing
technical assistance during the epicardial mapping experiments, the Pathology Department at
the Hospital for Sick Children for performing the picrosirius red staining, as well as Dr. Abdul
Momen (Husain lab) and Dr. Yu-Qing Zhou (TCP Mouse Imaging Centre) for acquiring the
invasive LV hemodynamic and ultrasound biomicroscopy data respectively.

89
CHAPTER 4. REDUCED CONNEXIN-43 AND SODIUM CHANNEL NAV1.5 IS
ASSOCIATED WITH ET-1 INDUCED ELECTRICAL UNCOUPLING

90
4.1 INTRODUCTION
The findings of the previous chapter indicate that our mouse model of ET-1 induced
cardiomyopathy has a progressive ventricular conduction delay that precedes structural or
contractile dysfunction. As ET-1 can modulate Ca2+ currents (ICa-L, ICa-T), Ca2+ handling
proteins (NHE, NCX, IP3R), K+ currents (IK1, Ito, IKs), and gap junctions (Cx43)59, 219-226, 228-232,
we hypothesized that ET-1 induced conduction disturbances in our model will correlate with
remodelled gap junctions and ion channels. Indeed, here we present data revealing that high
levels of myocardial ET-1 leads to reduced LV Cx43, Cx40 and Nav1.5 mRNA and protein,
and Na+ channel conductance. Tbx2, Tbx3, and Nkx2.5 have been shown to mediate
transcriptional repression of Cx43162, 163, while NFκB has been shown to modulate INa and Ito
by repressing SCN5A and KChIP2 expression109, 244. As NFκB has previously been shown to
be activated in our mouse model34, and can act as a transcriptional repressor109, we posited
that NFκB was mediating the ET-1 induced electrical remodelling in our model.
The objectives of this chapter are 1) to determine the potential mechanism(s) of ET-1
induced electrical remodelling via western blot and qRT-PCR for gap junction and ion
channels; 2) to confirm the molecular mechanism(s) of ET-1 induced electrical remodelling
with an in vitro system; 3) to determine if ET-1 induced NFκB activation is responsible for
transcriptional repression of Cx43 and Nav1.5; 4) to determine if this model can be prevented
by inhibition of ET-1 over-expression by re-introducing DOX at the onset of ET-1 induced
electrical remodelling.

91
4.2 MATERIALS & METHODS
4.2.1 Experimental animals. Experiments were conducted in mice withdrawn from DOX for
4 wks (group I), 8 wks (group II), or after 4 wks of DOX removal with DOX re-administered
for an additional 4 wks (group III). Hearts were used for qRT-PCR, western blot, and
immunohistochemistry in groups I, II and III, while Na+ channel conductance and ChIP with
NFκB-p50 Ab were evaluated on group I mice (Fig. 4.1A). In vitro validation of ET-1
induced electrical remodelling was assessed in HL-1 cells and neonatal mouse ventricular
myocytes (Fig. 4.1B,C).
4.2.2 RNA isolation & quantitative real-time RT-PCR analysis. RNA from the LV was
extracted using ice-cold TRIzol reagent (Invitrogen, Burlington, ON), DNased and reverse-
transcribed with Superscript III (Invitrogen). cDNA was subjected to real-time PCR using the
ABI Prism 7900 sequence detection system version 2.3 (Applied Biosystems, Streetsville,
ON) with SYBR green. Mouse-specific primers for GAPDH, IL-6, Nav1.5, Cx40, Cx43, Cx45,
Cav1.2, SERCA2a, Kv1.4, Kv1.5, Kv2.1, Kv4.2, KChIP2, and human primers for ET-1 were
used 34, 245-247 (Table 4.1). Relative standard calibration curves were generated for each primer
set with cDNA produced from one mouse heart. The mRNA level of each gene was
normalized to that of GAPDH. All samples for each primer set were run together in duplicates.
4.2.3 Western blotting. LV tissue was homogenized in ice cold 1% SDS, 5 mM EDTA, 1mM
PMSF, 1X protease inhibitor cocktail, and 1X phosphatase inhibitor (Sigma-Aldrich, Oakville,
ON). 20 µg of protein extract was run on 4-12% SDS-PAGE, transferred to PVDF

24 hrs qRT-PCR for Cx43, Nav1.5, Cav1.2
qRT-PCR and western blot for Cx43, optical mapping
qRT-PCR and western blot for Cx43 and Nav1.5,
Cx43 and p-Cx43 immunohistochemistry,
qRT-PCR and western blot for Cx43 and Nav1.5,Cx43 and p-Cx43 immunohistochemistry,
Na channel conductance and cell capacitance,promoter analysis & ChIP with NFκB Abs
Birth
Weaning (3 wks of age)
0 wks
4 wks
8 wks
Group I
Group II
Group III
4 wks ET-1 on(- DOX)
8 wks ET-1 on(- DOX)
4 wks ET-1 on(- DOX)
4 wks ET-1 off(+ DOX)
8 wks
qRT-PCR and western blot for Cx43 and Nav1.5,
Cx43 and p-Cx43 immunohistochemistry,
ET-1 off(+ DOX)
C
A
Bd0.5 d1.5 d2.5 d3.5
d0 d1 d2
ET-1 exposure
ET-1 exposure
72 hrs
24 hrs
ET-1 exposure
d0
In vivo Ventricular
In vitro Atrial
In vitro Ventricular
92

Figure 4.1. Schematic of experimental designs. A. Mice were kept on DOX from conception until 3 wks of age, then either withdrawn from DOX for 4 wks (Group I), for 8 wks (Group II), or withdrawn from DOX for 4 wks followed by DOX re-administration for an additional 4 wks (Group III). Western blot, immunohistochemistry and qRT-PCR for Cx43 and Nav1.5 were performed on LV tissue samples from mice in all groups. Na channel conductance, cell capacitance, promoter analysis, ChIP with NFκB p50 Ab, and qRT-PCR for NFκB binding sites of various promoter sequences were performed in mice in Group I. HL-1 cells (B) or NMVM (C) cultured with ET-1 (10, 100 nM) underwent qRT-PCR and western blot for Cx43 and/or Nav1.5 , and optical mapping.
93

94
Table 4.1. Real-time PCR Primer Sequences Name Sequence Reference
hET-1 pF 5’-GCTCGTCCCTGATGGATAAA-3’ 34 hET-1 pR 5’-CTGTTGCCTTTGTGGGAAGT-3’ 34 GAPDH pF 5’-GCATGGCCTTCCGTGTTC-3’ 34 GAPDH pR 5’-ATGTCATCATACTTGGCAGGTTTC-3’ 34 Cx40 pF 5’-TCGTCCAGAGCCTCACTCCTC-3’ 245 Cx40 pR 5’-GCCAGAGCGTCTGGATTCTTC-3’ 245 Cx43 pF 5’-TTGACTTCAGCCTCCAAGG-3’ 245 Cx43 pR 5’-AATGAAGAGCACCGACAGC-3’ 245 Cx45 pF 5’-CACTTGGAACACACCCTCTGCTC-3’ 245 Cx45 pR 5’-GGGAGGTGTTCCTCGTGGCT-3’ 245 Cav1.2 pF 5’-GAGTACTGCAGTGAGCCCAGCCTGCTC-3’ Cav1.2 pR 5’-GGCAGAGCGAAGGAAACTCCTCTTTGG-3’ IL-6 pF 5’-GAGGATACCACTCCCAACAGACC-3’ 246 IL-6 pR 5’-AAGTGCATCATCGTTGTTCATACA-3’ 246 Nav1.5 pF 5’-GAAGAAGCTGGGCTCCAAGA-3’ 245 Nav1.5 pR 5’-CATCGAAGGCCTGCTTGGT-3’ 245 SERCA2a pF 5’-TGAGACGCTCAAGTTTGTGG-3’ 247 SERCA2a pR 5’-ATGCAGAGGGCTGGTAGATG-3’ 247 Kv1.4 pF 5’- TATGCCGAAGCCCGAGAG-3’ Kv1.4 pR 5’- CATGTGCTGTTATTTGTGTGGC-3’ Kv1.5 pF 5’- TTATTCTTATGGCTGACGAGTGCAT-3’ Kv1.5 pR 5’- CCCCCCCAAGACTTCATAATTC-3’ Kv2.1 pF 5’- CACACAGCAATAGCGTTCAACTT-3’ Kv2.1 pR 5’- AGGCGTAGACACAGTTCGGC-3’ Kv4.2 pF 5’- GTGTCGGGAAGCCATAGAGGC-3’ Kv4.2 pR 5’- TTACAAGGCAGACACCCTGA-3’ KChIP2 pF 5’- GGCTGTATCACGAAGGAGGAA-3’ KChIP2 pR 5’- CCGTCCTTGTTTCTGTCCATC-3’

95
membrane, blocked in 5% non-fat dry milk in TBS-T (Tris base, NaCl, 0.1% Tween-20, pH
7.6) for 1 h and incubated overnight at 4oC in TBS-T with 5% BSA containing primary rabbit
Abs against Cx43 and p-Cx43 (Ser368; Cell Signaling, Danvers, MA), Cx45 (Invitrogen),
Cx40 (Millipore, Billerica, MA), Nav1.5 (Sigma-Aldrich), and Cav1.2 (Alomone Laboratories,
Jerusalem, Israel), or a primary mouse Ab against SERCA2a (Novus Biologicals, Littleton,
CO, USA). Immunoreactivity was detected with chemiluminescence (Perkin Elmer,
Woodbridge, ON), using a horseradish-peroxidase-conjugated secondary goat anti-rabbit Ab
(Pierce, Pierce, Nepean, ON) in 5% non-fat dry milk in TBS-T for 30 min. Densitometry was
performed using Quantity One (Bio-Rad, Hercules, CA). GAPDH was used as a loading
control to normalize protein samples. All samples from each group were run together.
4.2.4 Immunohistochemistry. 4µm sections from perfusion-fixed paraffin-embedded hearts
at the level of the papillary muscle underwent antigen retrieval by boiling in 10 mM Na citrate
buffer (pH 6.0) for 20 min and blockade of endogenous peroxidase activity with 3% H2O2 for
10 min. Sections were blocked for 1 h in 5% normal goat serum, incubated overnight at 4oC
with primary rabbit anti-Cx43 or p-Cx43 Abs (Cell Signalling), and incubated at RT for 30
min each with a biotinylated goat anti-rabbit secondary Ab (Vector Labs, Burlington, ON),
and an ABC reagent (Vector labs). Staining, visualized with DAB substrate (Vector labs), was
followed by counterstaining with hematoxylin. Ten images from each section were analyzed
with Aperio ImageScope software using the v9 algorithm (version 10.2.2.2319; Aperio
Technologies) to quantify the area of positively stained pixels.

96
4.2.5 HL-1 cell culture. HL-1 cells were cultured at 37oC with 5% CO2 in Claycomb media
(Sigma-Aldrich; supplemented with 10% FBS, 0.1mM Norepinephrine, 2mM L-Glutamine,
and 100U/ml:100μg/ml Penicillin/Streptomycin) on plates coated with fibronectin (12.5μg/ml
in 0.02% gelatin). Cells were cultured for 24 or 72 hrs in media supplemented with 0, 10, or
100 nM ET-1 (Sigma-Aldrich). Microscopy was used to confirm contractile activity and
optical mapping was used to record electrical activity of the monolayer. Protein from cells was
extracted in ice-cold lysis buffer ((mmol/L) 25 Tris pH 7.5, 1 Dithiothreitol, 150 NaCl, 1
PMSF, 1 NaF, 1 Na3VO4, 0.4% NP-40, 0.4% Triton-X-100, 2x complete protease inhibitor
cocktail) and stored at -80oC until used in western blotting. RNA was extracted and treated as
previously described in section 4.2.2.
4.2.6 Optical mapping. HL-1 cells were pre-loaded with 2.5μM di-ANNEPS 4 (Invitrogen)
for 15 min at 37oC, washed and signals recorded in fresh Tyrodes solution. The plate was
placed on a heated stage under a microscope fitted to a high speed camera system (Ultima;
BrainVision, Tokyo, Japan). During the acquisition of optical mapping data, the setup was
maintained in the dark, a green halogen light (530nm) was shined on the monolayer, and the
emitted red fluorescent light (600nm) was recorded from the monolayer through a red filter.
The optical data was digitized with a 14-bit resolution and transformed into pseudo-unipolar
electrograms by averaging the pixel intensity over a 5x5 grid. Pseudo-bipolar electrograms
were then generated by taking the difference between two pseudo-unipolar electrograms. The
spatial resolution of the camera was 6,250 pixels/cm2 (at an optical zoom of 0.6). The mean
conduction velocity (CVx, CVy, CVt) was determined with Matlab (version 2007b) and
individual experiments were normalized to signal rate and to background (without treatment).

97
4.2.7 Isolation of NMVM. NMVM were isolated from 1-2 day old mice 248. Briefly, mice
were decapitated, ventricles quickly excised and minced into 3-4 pieces, washed and
incubated in ice-cold digestion solution [(mmol/L) 137 NaCl, 5.36 KCl, 0.81 MgSO4, 5.55
Dextrose, 0.44 KH2PO4, 0.34 Na2HPO4, 20 HEPES, 0.07 CaCl2, 0.02 Gentimicin, 0.02%
Trypsin, pH 7.5] for 15 hours at 4oC with light shaking. The remaining tissue was serially
digested at 37oC with light shaking in fresh aliquots of digestion buffer supplemented with
150 μg/ml Collagenase II (Worthington Biochemical Corporation, Lakewood, NJ) in 5 min
intervals for 50-60 min. The supernatant was collected and added to FBS, and spun at 4oC for
5 min @ 1000 rpm. To remove fibroblasts, the pellet was resuspended in DMEM/F12
(supplemented with 10% FBS and 100U/ml:100μg/ml Penicillin/Streptomycin), plated in a
10cm dish, and incubated at 37oC for 1 h with 5% CO2. The cell suspension was removed,
counted, plated to 100% confluency and maintained at 37oC with 5% CO2 in DMEM/F12
(supplemented with 10% FBS and 100U/ml:100μg/ml Penicillin/Streptomycin). Contractile
activity was monitored with microscopy. RNA from cells exposed to 10 or 100 nM ET-1
(Sigma-Aldrich) for 24 h were extracted and treated as previously described.
4.2.8 Promoter analysis. Transcription factor binding sites within -1000 to +1000 of the
transcriptional start site of the Cx43 and Nav1.5 (A and B) promoters were analysed using
MatInspector (Genomatix). Clustal analysis was then used to determine if the transcription
factor binding sites were conserved between various species. Primers were designed to target
the NFκB binding sites within the Cx43, Nav1.5 (A and B), NFκBIB, Tbx2, Tbx3 and Nkx2.5
promoters (Table 4.2). Binding to the NFκBIB promoter was used as a positive control.

98
Table 4.2. Real-time PCR Primer Sequences-promoters for ChIP for NFkB binding sites Name Sequence Cx43 (1) pF 5’-ACTGCCCGTGGTCATCTCCTG-3’ Cx43 (1) pR 5’-TGTAGTTTCAATGTGCTGACGAGAAGG-3’ Cx43 (2) pF 5’-AAACGCTTTTACGAGGTATCAGCAC-3’ Cx43 (2) pR 5’-AGGAGAGTTCGGGCTCTCAGAC-3’ Cx43 (3) pF 5’-CCGCTGAGAGTGCAACAGGTAAAAG-3’ Cx43 (3) pR 5’-TGGCGACTACAGAAAAGTGAAGTTAGGC-3’ Nav1.5 (1) pF 5’-CCACCTGTCACATTTCCCGTCTTTAG-3’ Nav1.5 (1) pR 5’-GTGCCCAACTGTGTAGACCTCCATC-3’ Nav1.5 (2) pF 5’-AGCCCCCGCAGCCACTATC-3’ Nav1.5 (2) pR 5’-CGCGGCGATCAGAGAGGAC-3’ Nav1.5 (3) pF 5’-CAGATCTGCGAGTGTGCCTTGTC-3’ Nav1.5 (3) pR 5’-CAGAGCGCATAGACACAGGAGGAG-3’ Tbx2 pF 5’-TGTCAATGCTTTGCACTTGG-3’ Tbx2 pR 5’-CTGGAAAGCGCAGAGGAC-3’ Tbx3 pF 5’-CAGCGGGCAGAAAGATTG-3’ Tbx3 pR 5’-GCTCGCGTAGCTCTGAAGG-3’ Nkx2.5 (1) pF 5’-GGTGGCCGAATACCAAATATC-3’ Nkx2.5 (1) pR 5’-GGGCCTCTCTGGCTTTACTC-3’ Nkx2.5 (2) pF 5’-GGAAGAGGTCTGGGATAGGG-3’ Nkx2.5 (2) pR 5’-CTCGCTCGGTTCGATGTTG-3’ NFkBIB pF 5’-CGAGCGAATGTCCACCTC-3’ NFkBIB pR 5’-CAGTGCTTCCGCCCTATC-3’ Tbx = T-box transcription factor, Nkx2.5 = NK2 transcription factor related, locus 5 (cardiac-specific homeobox), NFκBIB = NFκB inhibitor beta

99
4.2.9 ChIP. Frozen tissue was ground finely with a mortar and pestle and fixed with 1%
formaldehyde in PBS at RT for 10 min on a rotator. Tissue fixation was stopped with 10X
Glycine. Cells were centrifuged at 1,900 rpm for 5 min at 4oC, washed in PBS (supplemented
with 0.5% PMSF) and centrifuged again at 1,900 rpm for 5 min at 4oC. The cells were then
resuspended in 1ml ice-cold lysis buffer (supplemented with 0.5% protease inhibitor cocktail
and 0.5% PMSF), dounced homogenized 30 times (on ice for 30 s between sets of 10),
sonicated for 15 s with a hand-held sonicator to aid in nuclei release, and centrifuged at 5,000
rpm for 10min at 4oC. The nuclear pellet was resuspended in 350μl of shearing buffer
(supplemented with 0.5% protease inhibitor cocktail) and the DNA was sheared by sonicating
in a water bath for 40 min (30 s on, 30 s off) at 4oC. The sheared DNA was centrifuged at
15,000 rpm for 12min at 4oC. Shearing efficiency was determined by running phenol
chloroform extracted DNA on 2% agarose gel. ChIP reactions were performed with NFκB
p50 Ab (Abcam, Cambridge, MA) using the ChIP-IT Express Magnetic Chromatin
Immunoprecipitation Kit (Active Motif, Carsbad, CA). Briefly, immunoprecipitation reactions
consisting of 1µg sheared DNA, 25µl of protein-G magnetic beads, 1µl of protease inhibitor
cocktail, and 2µg of NFκB p50 Ab were incubated on a rotator for 4 hrs at 4oC. Next, the
magnetic beads were washed,the DNA eluted, and cross-links reversed. Then, the samples
were heated to 95oC for 15min and incubated with Proteinase K for 1hr at 37oC. The DNA
from these samples was then purified using a MinElute Kit and used for quantification of
NFκB binding sites within various promoters using qRT-PCR. Sheared DNA was used as
input controls for each primer set for qRT-PCR reactions.

100
4.2.10 Isolation of adult mouse ventricular myocytes. Endo-cardiomyocytes from mice in
Group I were isolated as described previously 249. Briefly, hearts from mice anesthetised with
isoflurane were rapidly removed and retrogradely perfused with Ca2+ free Tyrode’s solution
[(mmol/L) 137 NaCl, 5.4 KCl, 1.0 MgCl2, 0.33 NaH2PO4, 10 D-glucose, 10 HEPES, pH 7.4]
at 37ºC through the aorta for 3-4 min, then perfused with 1.0 mg/mL Collagenase II
(Worthington Biochemical Corporation, Lakewood, NJ) for 10-12 min. A thin layer of
endocardium was removed from the left ventricular fee wall, dissociated and stored in Krebs-
bicarbonate solution at 4ºC [(mmol/L) 120 potassium glutamate, 20 KCl, 20 HEPES, 1.0
MgCl2, 10 D-glucose, 0.5 K-EGTA, and 0.1% bovine serum albumin].
4.2.11 Patch clamp recordings. Isolated myocytes were voltage-clamped by Dr. Jie Liu
(Backx lab) with patch pipettes (1.3-2.3 MΩ) filled with (mmol/L) 1 CaCl2, 135 CsCl, 5 NaCl,
1 MgCl2, 4 MgATP, 10 HEPES, and 10 EGTA (pH 7.2) and superfused with solution
containing (mmol/L) 2.5 NaCl, 130 CsCl, 1 MgCl2, 1 CaCl2, 5 HEPES, and 10 D-glucose (pH
7.35), supplemented with 10µM Nifedipine to block L-type Ca2+ channel and 40 µM NiCl to
block T-type Ca2+ channel. Serial resistance and cell capacitance were compensated by 90%.
The recording protocol consisted of 50ms test steps from –80 mV to +50 mV in increments of
10 mV. Data were analyzed with Clampfit 9. Na+ current I-V curve were plotted and fit with
the following function in which Vm is membrane potential, Vrev is Na current reversal
potential, Gmax is whole cell Na+ channel conductance, Vhalf is Na channel half activation
voltage, k is slope:
Y=(Vm - Vrev) * Gmax * (-1 / (1 + exp((Vm - Vhalf) / k)) + 1)

101
4.2.12 Statistical analysis. Data are presented as mean±SE. Comparisons between BT vs.
NBT in groups I, II, or III and between cells exposed to varying amounts of ET-1(HL-1 or
NMVM) were done by Student’s t-tests using SPSS v12 (SPSS, Chicago, IL).
4.3 RESULTS
4.3.1 ET-1 mediated electrical remodelling correlates with reduced Cx43, p-Cx43, Cx40,
Nav1.5, and Na+ channel conductance. To define the molecular basis for ET-1 induced
electrical remodelling, we examined the mRNA/protein expression levels of genes involved in
action potential generation and propagation through the myocardium (Table 4.3, Fig. 4.2).
Western blot analysis revealed that total Cx43 protein, p-Cx43 to total Cx43 protein ratio, and
Cx40 protein was reduced by 93%, 69%, and 36% respectively in BT vs. NBT mice as early
as 4 wks post ET-1 over-expression (Group I, Figs. 4.2D,E,F,H). qRT-PCR showed that high
levels of hET-1 (transgene) mRNA expression also correlated with significantly reduced
mRNA levels of Cav1.2 (72%), Cx40 (63%), Cx43 (64%) and Nav1.5 (30%) by 4 wks of
transgene induction (Group I, Table 4.3). At 8 wks of ET-1 over-expression, Cav1.2, Cx40,
Cx43, Cx45 and Nav1.5 mRNA expression were reduced by 72%, 69%, 88%, 61%, and 64%
respectively (Group II, Table 4.3), with the p-Cx43 to total Cx43 protein ratio, Cav1.2, Cx40,
and Nav1.5 protein levels being 54%, 69%, 54%, and 71% lower in BT vs. NBT mice (Group
II, Figs. 4.2B,D,E-H). Development of HF also correlated with increased IL-6 mRNA
expression, reduced Cx45 mRNA expression with no appreciable difference at the protein
level, and reduced SERCA2a mRNA and protein expression (Table 4.3, Figs. 4.2A,C,H).

102
Table 4.3. Progression and prevention of molecular remodelling as evaluated by LV mRNA expression levels
Parameter
Group I
Group II
Group III
BT
(n=7)
NBT
(n=8)
BT
(n=6)
NBT
(n=7)
BT
(n=7)
NBT
(n=8)
hET-1 160±36**
(x103) 1.00±0.58
48±13**
(x103) 2.35±1.06 1849±776* 2.91±1.00
IL-6 1.89±0.52 1.00±0.27 12.17±1.79** 2.35±0.47 2.62±0.55 1.25±0.56
SERCA2a 0.32±0.08* 1.00±0.22 0.10±0.02** 0.52±0.12 0.69±0.12 0.94±0.13
Cav1.2 0.28±0.10** 1.00±0.16 0.20±0.01** 0.70±0.05 0.73±0.06 0.78±0.05
Cx40 0.63±0.11 1.00±0.18 0.48±0.08** 1.03±0.12 0.59±0.09 0.75±0.07
Cx45 0.73±0.20 1.00±0.14 0.38±0.09** 0.99±0.16 0.63±0.03 0.73±0.06
Cx43 0.36±0.15* 1.00±0.22 0.13±0.02** 1.01±0.13 0.77±0.14 1.02±0.17
Nav1.5 0.70±0.09** 1.00±0.05 0.36±0.05** 1.00±0.03 0.91±0.09 1.09±0.07
*=P<0.05, **=P<0.01

ph
osp
ho
-Cx4
3/ T
ota
l-C
x43
0
1
2
3
4
SE
RC
A2
a
0.0
0.5
1.0
1.5
2.0
2.5
3.0
Cx4
5
0.0
0.2
0.4
0.6
0.8
1.0
Ca v
1.2
0.0
0.5
1.0
1.5
2.0
2.5
3.0
Cx
43
0
1
2
3
4
Na v
1.5
0.0
0.2
0.4
0.6
0.8
1.0C
A
B BTNBT
E
F
G
D
I II III
H
Figure 4.2. Reduced LV Cx43 and Nav1.5 expression in mice as early as 4 wks after ET-1 induction. Densitometry analysis of SERCA2a (A), Cav1.2 (B), Cx45 (C), Cx40 (D), Cx43 (E), p-Cx43/Total Cx43 (F), and Nav1.5 (G) protein expression, and representative western blots (H) of LV of BT and NBT mice within groups I , II, and III (BT:NBT: n=4-6). *=P<0.05, **=P<0.01.
Cx43
p-Cx43
I II IIINBTBT NBTBT NBTBT
SERCA2aCav1.2
Nav1.5
Cx45
Cx40
I II III
*
*
*
****
*
*
Cx4
0
0.0
0.2
0.4
0.6
0.8
1.0
1.2 **
****
103

104
Immunohistochemistry was performed to explore the nature of disordered Cx43
expression. In agreement with Western blot data, Cx43 and p-Cx43 staining were greatly
reduced in BT mice after only 4 wks of ET-1 over-expression as compared to NBT controls
(Cx43: 3.7±0.3 vs. 9.9±1.1 %, P<0.01; p-Cx43: 2.2±0.3 vs. 7. 8±0.6 %, P<0.01) and further
reduced after 8 wks of ET-1 over-expression (Cx43: 0.2±0.0 vs. 7.9±0.2 %, P<0.01; p-Cx43:
0.2±0.0 vs. 6.3±0.3 %, P<0.01) (Figs. 4.3,4.4). Furthermore, improper localization (i.e.
lateralization) of Cx43 and p-Cx43 was observed in BT mice at the earliest stage (4 wks,
Group I), which became most severe with more prolonged ET-1 over-expression (8 wks,
Group II) (Figs. 4.3A,4.4A).
To determine the functional consequence of reduced Nav1.5 expression, Na+ current
was recorded in LV myocytes isolated from mice 4 wks post DOX withdrawal (Group I). Na+
channel conductance was reduced (0.17±0.01 vs. 0.21±0.01 s, P<0.05) with no difference in
cell capacitance (133.94±8.50 vs. 126.50±7.37 pF, P=NS) in BT (n=18) vs. NBT (n=24) (Fig.
4.5).
To explore the molecular determinants of ET-1 induced delay in repolarization time
seen in Group I mice (Fig. 3.3E), we examined the mRNA expression of several K+ channels
that play a role in action potential repolarization (Kv4.2, Kv2.1, Kv1.4, Kv1.5, KChIP2;
Appendix 2). The K+ channels involved in Ito (Kv4.2, KChIP2) were reduced in BT vs. NBT
mice (Appendix 2).

% o
f C
x43
sta
inin
g
0
2
4
6
8
10
BT NBTI
II
III
A
B**
I II III
**
**
50 µm50 µm
BTNBT
Figure 4.3. Prolonged ET-1 over-expression leads to progressive loss and lateralization of Cx43. Four weeks of ET-1 over-expression led to reduced Cx43 staining, while 8 wks of over-expression caused reduced and lateralized Cx43 staining. Representative Cx43 staining of heart sections (A) and % of Cx43 staining (B) in BT and NBT mice from groups I-III (n=2-4 per genotype per group). **=P<0.01.
105

BT NBT
% p
-Cx4
3 st
ain
ing
0
2
4
6
8
I
II
III
A
B
I II III
**
**
50 µm50 µm
BTNBT
Figure 4.4. Prolonged ET-1 over-expression lead to progressive loss and lateralization of p-Cx43. Diminished cardiac p-Cx43 staining after 4-8 wks of ET-1 over-expression and complete miss-localization of p-Cx43 staining after 8 wks of transgene expression. Representative p-Cx43 staining of hearts sections (A) and % of p-Cx43 staining (B) in BT and NBT mice from groups I-III (n=2-5 per genotype per group) **=P<0.01.
106

Cel
l C
apac
itan
ce (
pF
)
0
20
40
60
80
100
120
140
Cu
rre
nt
(pA
/pF
)
-4
-3
-2
-1
0
A
B
C
BTNBT
*
**
Na
chan
nel
co
nd
uct
anc
e (s
)
0.00
0.05
0.10
0.15
0.20
0.25
Figure 4.5. Na+ channel conductance reduced in LV myocytes isolated from mice 4wks post DOX withdrawal. (CURTESY OF BACKX LAB) Cell capacitance (A), Na+ channel conductance (B), and Na+ current elicited from a 50 ms test step from -80 to -40 mV (C) of LV myocytes isolated from BT (n=18) vs. NBT (n=24) mice in group I. *=P<0.05, **=P<0.01
107

108
4.3.2 In vitro validation of ET-1 induced electrical remodelling. To confirm our in vivo
experiments, HL-1 cells, a mouse atrial cell line, were treated for 24 or 72 hrs with ET-1 (10
or 100 nM) and assessed by western blotting, qRT-PCR, and optical mapping. Application of
exogenous ET-1 (10 or 100 nM) had no effect on Cx43 mRNA, protein, p-Cx43/total Cx43
protein expression, or conduction velocity (Fig. 4.6, 4.7). As our in vivo electrical remodelling
data supports a ventricular phenotype and HL-1 cells are derived from left atrial cells, we next
evaluated whether ET-1 over-expression in the left atria of Group I mice also correlated with
remodeled Cx43 and Nav1.5 (Fig. 4.8). Unlike LV tissue, high levels of hET-1 did not
correspond with reduced levels of Cx43 or Nav1.5 (Fig. 4.8). Therefore, exogenous ET-1
application may not have led to remodeled Cx43/Nav1.5 and associated reductions in
conduction velocity in HL-1 cells as atrial cells may respond differently to chronically high
levels ET-1 as opposed to ventricular cells. Also, indirect factors involved in ET-1 induced
electrical remodelling may be missing from the in vitro milieu.
Next, NMVM were used to confirm the ability of ET-1 to reduce Cx43 levels. Indeed,
24 hrs of exogenous ET-1 (10 and 100 nM) caused reduced Cx43 mRNA expression in
neonatal mouse ventricular myocytes in vitro (Fig. 4.9A). However, 100 nM ET-1 caused
increased levels of Nav1.5 and Cav1.2 mRNA expression (Figs. 4.9B,C). This in vitro
ventricular phenotype may differ to the in vivo ventricular phenotype as the duration of ET-1
treatment was severally limited by the lifespan of healthy contractile NMVM.
4.3.3 ET-1 induced reductions in Cx43 and Nav1.5 may be induced by NFκB. To access
the role of transcription factors in transcriptional repression of Cx43 and Nav1.5, potential

Cx4
3
0.0
0.2
0.4
0.6
0.8
1.0
1.2
1.4
p-C
x43/
To
tal
Cx4
3
0.0
0.5
1.0
1.5
2.0
Cx4
3 m
RN
A
0.0
0.5
1.0
1.5
2.0
2.5
3.0A
B
C
24 hrs 72 hrs
24 hrs 72 hrs
24 hrs 72 hrs
Figure 4.6. ET-1 treatment had no effect on Cx43 mRNA, protein, or p-Cx43/ Total Cx43 protein expression in HL-1 cells. Cx43 mRNA (A), protein (B), and p-Cx43/ Total Cx43 protein (C) levels after 24 or 72 hrs of ET-1 (10 or100 nM) exposure in HL-1 cells.
109

CV
x
0.000
0.002
0.004
0.006
0.008
CV
y
0.000
0.002
0.004
0.006
0.008
CV
t
0.000
0.002
0.004
0.006
0.008
A
B
C
D
24 hrs 72 hrs
24 hrs 72 hrs
24 hrs 72 hrs
Figure 4.7. ET-1 treatment had no effect on conduction velocity in HL-1 cells. A. Representative optical mapping tracing of HL-1 cells cultured without ET-1. CVx (B), Cvy
(C), and CVt (D) treated with ET-1 (10 or 100 nM) for 24 or 72 hrs.
110

Cx
43 m
RN
A
0
2
4
6
8
10
12
14
hE
T-1
mR
NA
0
2
4
6
8
10
12
14
16
18**
BTNBT
Na v
1.5
mR
NA
0
2
4
6
8
A
B
C
Figure 4.8. Four weeks of ET-1 transgene induction had no affect levels of left atrial Cx43 or Nav1.5 mRNA expression. Cx43 (A), Nav1.5 (B), and hET-1 (C) mRNA expression from left atrial tissue from mice in Group I. **=P<0.01
111

Cx4
3 m
RN
A
0.0
0.2
0.4
0.6
0.8
1.0
1.2
Na v
1.5
mR
NA
0.0
0.2
0.4
0.6
0.8
1.0
1.2
Ca
v1.2
mR
NA
0.0
0.2
0.4
0.6
0.8
1.0
1.2
1.4
0 nM10 nM100 nM
**
**
*
A
B
C
Figure 4.9. ET-1 reduces Cx43 levels in NMVM. Effects of 24 h of ET-1 exposure (10, or 100 nM) on expression levels of Cx43 (A) Nav1.5 (B) and Cav1.2 (C) mRNA in NMVMs (n=5 per group). *=P<0.05, **=P<0.01
112

113
transcription factor binding sites within the core promoters of Cx43 and Nav1.5 were
evaluated with Genomatix-MatInspector. The Cx43 promoter contains 4 NFκB, 2 Tbx, and 4
Nkx2.5 binding sites (Appendix 3). The Nav1.5 has two promoter regions, one contains 3
NFκB, 1 Tbx, and 1 Nkx2.5 binding sites, while the other contains 1 NFκB, 5 Tbx, and 4
Nkx2.5 binding sites (Appendix 4). The potential transcription factor binding sites were
located in highly conserved areas (confirmed in 10 species using Clustal). As we previously
showed NFκB is activated in this mouse model and several NFκB binding sites are present in
both the Cx43 and Nav1.5 promoters, we used ChIP with NFκBp50 Ab to quantify the binding
of NFκB to sites within the Cx43 and Nav1.5 promoters using qRT-PCR. As a control, we
examined NFκB binding to NFκBIB. Indeed, NFκB preferentially bound to sites within in the
NFκBIB promoter (Appendix 5). Interestingly, we also found that NFκB preferentially bound
to sites within the Cx43, and Nav1.5 promoters (Figs. 4.10). Also, as we previously found a 16
fold increase in Tbx2 in mice over-expressing ET-1 and found NFκB binding sites in the
promoters of Tbx2, Tbx3, and Nkx2.5, we proceeded to quantify NFκB binding to sites within
these promoters (Appendices 6-8). We found NFκB preferentially bound to a site within the
Tbx2 promoter (Appendix 6), however, NFκB did not preferentially bind to sites within the
Tbx3 or Nkx2.5 promoters (Appendices 7-8). It appears NFκB may directly mediate the ET-1
induced transcriptional repression of Cx43 and Nav1.5, or indirectly via Tbx2.
4.4 DISCUSSION
Our study has shown that as little as 4 wks of ET-1 excess in the heart can trigger
reduced Cx43 and Nav1.5 expression, reduced Na+ channel conductance, and slow

Cx4
3 P
rom
ote
r 1
st N
FB
bin
din
g s
ite
0
20
40
60
80
100
120
Cx4
3 P
rom
ote
r2n
d N
FB
bin
din
g s
ite
0
20
40
60
80
100
120
140
160
Cx
43 P
rom
ote
r 3rd
NFB
bin
din
g s
ite
0
2000
4000
6000
8000AN
a v1.
5 P
rom
ote
r A
1
st N
FB
bin
din
g s
ite
0
10
20
30
40
Na v
1.5
Pro
mo
ter
A
2nd N
FB
bin
din
g s
ite
0
50
100
150
200
250
B
Na v
1.5
Pro
mo
ter
B
NFB
bin
din
g s
ite
0
50
100
150
200
250
BTNBT
Figure 4.10. NFκB p50 preferentially binds to sites within the Cx43 and Nav1.5 promoters in the LV of ET-1 over-expressing mice. qRT-PCR results for the NFκB binding sites within the Cx43 (A) and Nav1.5 (B) promoters from a ChIP using an NFκB p50 Ab on LV tissue from BT vs. NBT mice from Group I.
114

115
ventricular conduction, while the overall structure and contractile function of the heart remain
essentially normal. The cardiac voltage-gated sodium channel Nav1.5 and the dominant gap
junction channel in the working ventricular myocardium Cx43 are the key determinants of
electrical impulse propagation250. Our study suggests that cardiac over-expression of ET-1
leads to impaired conduction via NFκB p50 mediated Cx43 and Nav1.5 remodelling (Fig.
4.11). Although a recent in vitro study found that ET-1 led to increased levels of Cx43 protein
in neonatal rat ventricular myocytes, they, like us, also found reduced intensity and expression
of Cx43 associated with reduced conduction velocity230. In astrocytes, ET-1 causes reduced
Cx43 expression and gap-junctional intercellular communication associated with activated
ETB receptors231. Also, a study recently showed that a loss of Cx43 protein in rabbit LV
myocytes similar to that shown in our model (40-70%) was associated with markedly reduced
cell coupling251.
In addition, studies have shown that TNFα, NHE1, and activated c-Src may be linked
with impaired Cx43 activity. TNFα has been shown to reduce Cx43 promoter activity in a rat
myoblast cell line252, NHE1 was shown to negatively regulate Cx43 expression through a
JNK1/2 dependent pathway in neonatal rat ventricular myocytes253, and activated c-Src has
been shown to reduce gap-junctional intercellular communication via tyrosine
phosphorylation of Cx43254. In human ovarian carcinoma cells, ET-1 mediated gap junction
uncoupling and reduced Cx43 expression is linked with increased tyrosine phosphorylation by
c-Src via activated ETAR229. As ET-1 stimulates TNFα in macrophages255 and activates

ET
A
ET
B
NFκB
↓Cx43
↓Nav1.5 (↓INa)
↓ Cx43-p
Figure 4.11. Model illustrating mechanim(s) of ET-1 induced reductions of Cx43 and Nav1.5 after 4 wks of ET-1 over-expression. ET-1 mediated NFκB activation inhibits transcription of Cx43 and Nav1.5 and increases transcription of Tbx2, ultimately leading to reduced levels of Cx43/Nav1.5 and electrical uncoupling.
ET-1ET-1 ET-1ET-1
ET-1ET-1ET-1
ET-1ET-1
ET-1
ET-1 ET-1
↓ Cx43, Cx40, Nav1.5, Cav1.2, Kv4.2, KChIP2
116

117
the NHE159 and c-Src256 in neonatal rat ventricular myocytes, it is possible that TNFα, NHE1
or c-Src mediate the gap junction remodelling observed in our ET-1 over-expressing mice.
Cx43 is normally located at the intercalated disk in a multi-protein complex with
N-cadherin and Nav1.5 in ventricular myocytes, presumably working in collaboration for
effective electrical coupling between cardiomyocytes. However, in human and animal models
of HF, Cx43 mRNA and protein expression is reduced, and Cx43 dephosphorylation and
lateralization are increased204. Unlike the reduced Cx43 levels seen in human HF, changes in
Cx45 levels that accompany HF have been inconsistent 205, 217. Purkinje fibers isolated from a
canine model of ventricular tachy-pacing induced HF exhibited delayed His-Purkinje
conduction, decreased mRNA and protein expression of Nav1.5 and Cx43, reduced Cx43
phosphorylation, and increased lateralization of Cx43218. Studies have shown the association
of Cx43 with zonula occludens-1 is increased in human HF216 and Cx43 lateralization and
subsequent loss of Cxs from the intercalated disk is linked with increased association of
zonula occludens-1 with activated c-Src257. However, in these studies, changes in Cx43
expression and localization were observed after HF developed. Indeed, we believe the current
study represents the first report of how these changes in Cx43 localization can precede HF,
and how their reversal may prevent progression to HF. However, down-regulation of Cx43
alone may be insufficient to cause HF as mice with cardiac-restricted deletion of Cx43
develop impaired ventricular conduction, spontaneous arrhythmias, and sudden cardiac death,
but not overt HF169. Taken together, these data lead us to propose that electrical remodelling
leading to HF require both reduced expression and/or phosphorylation of Cx43 as well as
impaired Nav1.5 function at the intercalated disk.

118
Na+ channel α and β subunits are localized to t-tubules and the intercalated disk.
Nav1.5 channels located at the intercalated disk are thought to play a key role in propagation
of the action potential between cells, whereas Na+ channels in t-tubules are involved in linking
depolarization of the cell membrane with contraction. Interestingly, only tyrosine
phophorylated β1-subunits are found with Nav1.5 at the intercalated disk258, and normal
functioning Nav1.5 requires dystrophin at the intercalated disk as dystrophin-deficient mice
have severely compromised levels of Nav1.5 expression and increased QRS duration259. HF is
also associated with reduced INa, downregulation of native Nav1.5 mRNA, an upregulation of
2 non-functional C-terminal splice variants, and reduced Nav1.5 protein195. Mutations in
Scn5a gene have been linked with impaired conduction and are associated with Lenègre
disease, the Brugada syndrome, and LQT260. Indeed, SCN5A+/- mice develop impaired
conduction, fibrosis, and diminished and disturbed distribution of Cx43111. A link between
ET-1 over-expression and Nav1.5 downregulation was hypothesized based on our knowledge
of ET-1 acting via NFκB-mediated transcription61, and a report of NFκB p50 binding to and
repressing activity of the SCN5a promoter109. Our study provides additional support that
NFκB p50 mediates ion channel remodelling by transcriptional repression of SCN5a. In
addition, our study suggests that NFκB p50 acts as novel mediator of gap junction remodeling
through transcriptional repression of Cx43.
In conclusion, our study suggests that in the heart, ET-1 leads to impaired ventricular
conduction via NFκB p50 mediated gap junction/ion channel remodelling. The sequence and
reversibility of this cardiomyopathy phenotype suggests that a primary abnormality in
electrophysiology may trigger subsequent ventricular dysfunction and may represent a
therapeutic target in HF. This conceptual paradigm deserves closer examination in human HF,

119
particularly in conditions where early ‘electrical’ interventions may be tested for their ability
to prevent the progression to, or possibly even reverse, the manifestations of ventricular
dysfunction.
Acknowledgments: I would like to thank Stéphane Massé for providing technical assistance
during the optical mapping experiments, Dr. Jie Liu (Backx Lab) for isolating the adult mouse
ventricular myocytes and acquiring the patch-clamp recordings, Talat Afroze (Husain Lab) for
supplying the Cav1.2 primers, and Dr. Kyoung-Han Kim (Backx Lab) for supplying the Kv1.4,
Kv1.5, Kv2.1 Kv4.2 and KChIP2 primers.

120
CHAPTER 5. SUMMARY AND FUTURE DIRECTIONS

121
5.1 SUMMARY AND CONCLUSIONS
In this thesis, we aimed to 1) describe the temporal progression of the ET-1 induced
electrical remodeling, 2) determine the molecular mechanism(s) leading to ET-1 induced
electrical remodeling, and 3) determine if this cardiomyopathic phenotype can be prevented
by inhibiting ET-1 over-expression. We hypothesized that 1) inhibition of ET-1 synthesis
using the ECE inhibitor CGS-26303 will effectively prevent the ET-1 induced
cardiomyopathy, 2) ET-1 induces electrical remodeling by disruption of Connexin-43 and
Nav1.5, via NFκB transcriptional repression, and 3) this model will be reversible by the
cessation of ET-1 over-expression at the onset of electrical remodelling.
Serial evaluation of mice with chronic cardiac over-expression of ET-1 revealed that
electrical remodelling precedes structural and functional remodelling. ET-1 induced electrical
remodelling manifests initially (4 wks post transgene induction) as a ventricular phenotype,
mediating prolonged ventricular activation and reduced ventricular conduction. At the
molecular level, the electrical remodelling is associated with abnormal gap junctions and ion
channels, exemplified by reduced p-Cx43, Cx43 mRNA and protein, Nav1.5 mRNA and
protein, Na+ channel conductance, Kv4.2 and KChIP2 mRNA, as well as NFκB p50
preferentially binding to Cx43 and Nav1.5 promoters. Connecting abnormal clinical
electrophysiological phenotypes with underlying molecular mediators may be beneficial for
identifying targets for individual therapeutic strategies.
Persistent transgene induction (8 wks) eventually triggers biventricular systolic and
diastolic dysfunction, myocardial fibrosis, cardiomyocyte hypertrophy, prolonged ventricular
activation and repolarization, reduced ventricular conduction, and abnormal AV nodal
function. Inhibiting continued ET-1 over-expression at the onset of electrical remodelling

122
reversed the ventricular conduction delay and associated molecular mediators as well as
prevented the development of structural and functional remodelling. The sequence and
reversibility of this phenotype provides further evidence that a primary abnormality in
electrical remodelling may contribute to the pathogenesis of HF. Identifying the molecular
mediators that contribute to HF initiation and progression may facilitate early detection and
prevention of clinical HF.
Dual ECE/NEP inhibition with CGS-26303 (5 mg/kg/day) failed to improve survival,
or preserve hemodynamic and contractile integrity in this model. Our data suggest that high
levels of bigET-1, as seen in HF, may trigger increased ECE activity and/or non-ECE ET-1
synthesis, thus circumventing the efficacy of ECE blockade. Although negative, these results
provide further information of the use ECE/NEP inhibition with CGS 26303 in conditions of
chronic ET-1 expression. Preventing ET-1 production at an early stage may be more effective
than ECE blockade, namely to evade the potential deleterious effects of bigET-1 accumulation
as well as the synthesis of mature ET-1 from non-ECE proteases.
5.2 FUTURE DIRECTIONS
5.2.1 ET-1 and atrial electrical remodelling
In our model, ET-1 initially triggers a ventricular phenotype characterized by impaired
conduction and gap junction/ ion channel remodelling. The ET-1 induced atrial electrical
phenotype characterized by prolonged AH and AV conduction develops post structural and
functional remodelling. The reason for this differential atrial response is unknown, however,
Cx43 remodelling is not apparent in left atrial tissue samples from Group I mice or in HL-1
cells. Similarly, Cx43-/-, Cx43+/-, and cardiac specific Cx43-/- mice develop ventricular

123
conduction delays and arrhythmias with no discernable atrial phenotype. It would be
beneficial to determine whether atrial myocytes lack NFκB p50 activation, and thus deficient
transcriptional repression of gap junctions and ion channels. Ascertaining the mechanism(s)
that underlie the ET-1 induced AV nodal conduction delay by examining the ionic currents
and expression of gap junctions and ion channels in AV nodal cells isolated from Group II
mice would be useful.
5.2.2 ET-1 and K+/Ca2+ channel remodelling
Preliminary investigation revealed 4 weeks of ET-1 over-expression in Group I mice
led to reduced LV Kv4.2 and KChIP2 mRNA (Apendix 2). Additional characterization of
ET-1 induced K+ channel remodelling at the protein level is required to determine if the
reduction at the mRNA level translates into reductions at the protein level. As Kv4.2 and
KChIP2 are the main molecular determinants of Ito, and recent studies have shown that Ito and
INa are functionally and structurally coupled via an interaction between Navβ1 and Kv4α
subunits131, and KChIP2 has been shown to modulate ICa-L through binding to Cav1.2,
evaluating Ito, ICa-L, and INa-L, in ventricular myocytes isolated from Group I mice may be
informative. A recent study suggests that in cardiac disease, NFκB modulates reduced Ito via
down-regulation of KChIP2244. As NFκB is activated in our mouse model and may be
responsible for ET-1 induced repression of Cx43 and Nav1.5 expression, evaluating whether
NFκB plays a role in down-regulating Kv4.2 and KChIP2 expression in Group I mice would
be useful. Furthermore, preliminary promoter analysis with MatInspector showed that both the
Kv4.2 promoter and KChIP2 promoter contains an NFκB binding site.

124
5.2.3 Role of NFκB p50 in ET-1 induced gap junction/ion channel remodelling
NFκB p50 mediates transcriptional repression of Nav1.5109 and contributes to ET-1
induced electrical remodelling in our model by participating in the transcription repression of
Cx43 and Nav1.5. Thus, evaluating whether NFκB p50 inhibition could prevent ET-1 induced
electrical remodelling and/or HF development could provide valuable insight. Inhibiting
NFκB p50 in our model can be accomplished via administration of an NFκB p50 inhibitor
peptide that inhibits the nuclear translocation of NFκB p50261. Conversely, investigating the
effects of increased NFκB p50 in NMVM on ionic currents (Ito, INa) and expression of Cx43,
Nav1.5, Kv4.2 and KChIP2 would be beneficial.
5.2.4 Polymorphisms in ET-1 signaling components
Given our observation that ET-1 plays a role in electrical remodelling and the
pathophysiology of HF, it may be useful to determine if polymorphisms in the key genes
involved in ET-1 signaling (preproET-1, ECE-1, ETA, and ETB) are associated with familial
cardiomyopathy and or inherited arrhythmias. If polymorphisms in these genes are found to
correlate with familial conditions of these sorts, further investigations into the functional
relevance of ET-1 signaling may be indicated. For example, attempts to modify ETA/B receptor
activation with commercially available agents may be considered in these otherwise
untreatable and typically progressive conditions.

125
REFERENCES
1. Jessup M, Abraham WT, Casey DE, Feldman AM, Francis GS, Ganiats TG, et al. 2009 focused update: ACCF/AHA Guidelines for the Diagnosis and Management of Heart Failure in Adults: a report of the American College of Cardiology Foundation/American Heart Association Task Force on Practice Guidelines: developed in collaboration with the International Society for Heart and Lung Transplantation. Circulation 2009;119:1977-2016.
2. Levick SP, Melendez GC, Plante E, McLarty JL, Brower GL, Janicki JS. Cardiac mast cells: the centrepiece in adverse myocardial remodelling. Cardiovasc Res 2011;89:12-19.
3. Stewart JA, Jr., Wei CC, Brower GL, Rynders PE, Hankes GH, Dillon AR, et al. Cardiac mast cell- and chymase-mediated matrix metalloproteinase activity and left ventricular remodeling in mitral regurgitation in the dog. J Mol Cell Cardiol 2003;35:311-319.
4. Deswal A, Petersen NJ, Feldman AM, Young JB, White BG, Mann DL. Cytokines and cytokine receptors in advanced heart failure: an analysis of the cytokine database from the Vesnarinone trial (VEST). Circulation 2001;103:2055-2059.
5. Mann DL. Inflammatory mediators and the failing heart: past, present, and the foreseeable future. Circ Res 2002;91:988-998.
6. Yanagisawa M, Kurihara H, Kimura S, Tomobe Y, Kobayashi M, Mitsui Y, et al. A novel potent vasoconstrictor peptide produced by vascular endothelial cells. Nature 1988;332:411-415.
7. Roubert P, Viossat I, Lonchampt MO, Chapelat M, Schulz J, Plas P, et al. Endothelin receptor regulation by endothelin synthesis in vascular smooth muscle cells: effects of dexamethasone and phosphoramidon. J Vasc Res 1993;30:139-144.
8. Suzuki T, Kumazaki T, Mitsui Y. Endothelin-1 is produced and secreted by neonatal rat cardiac myocytes in vitro. Biochem Biophys Res Commun 1993;191:823-830.
9. Castro MG, Rodriguez-Pascual F, Magan-Marchal N, Reguero JR, Alonso-Montes C, Moris C, et al. Screening of the endothelin1 gene (EDN1) in a cohort of patients with essential left ventricular hypertrophy. Ann Hum Genet 2007;71:601-610.
10. Zhu G, Carlsen K, Carlsen KH, Lenney W, Silverman M, Whyte MK, et al. Polymorphisms in the endothelin-1 (EDN1) are associated with asthma in two populations. Genes Immun 2008;9:23-29.
11. Inoue A, Yanagisawa M, Takuwa Y, Mitsui Y, Kobayashi M, Masaki T. The human preproendothelin-1 gene. Complete nucleotide sequence and regulation of expression. J Biol Chem 1989;264:14954-14959.

126
12. Mawji IA, Robb GB, Tai SC, Marsden PA. Role of the 3'-untranslated region of human endothelin-1 in vascular endothelial cells. Contribution to transcript lability and the cellular heat shock response. J Biol Chem 2004;279:8655-8667.
13. Vallender TW, Lahn BT. Localized methylation in the key regulator gene endothelin-1 is associated with cell type-specific transcriptional silencing. FEBS Lett 2006;580:4560-4566.
14. Stow LR, Gumz ML, Lynch IJ, Greenlee MM, Rudin A, Cain BD, et al. Aldosterone modulates steroid receptor binding to the endothelin-1 gene (edn1). J Biol Chem 2009;284:30087-30096.
15. Yeligar S, Tsukamoto H, Kalra VK. Ethanol-induced expression of ET-1 and ET-BR in liver sinusoidal endothelial cells and human endothelial cells involves hypoxia-inducible factor-1alpha and microrNA-199. J Immunol 2009;183:5232-5243.
16. D'Orleans-Juste P, Plante M, Honore JC, Carrier E, Labonte J. Synthesis and degradation of endothelin-1. Can J Physiol Pharmacol 2003;81:503-510.
17. Honore JC, Plante M, Bkaily G, Rae GA, D'Orleans-Juste P. Pressor and pulmonary responses to ET-1(1-31) in guinea-pigs. Br J Pharmacol 2002;136:819-828.
18. Takuwa Y, Kasuya Y, Takuwa N, Kudo M, Yanagisawa M, Goto K, et al. Endothelin receptor is coupled to phospholipase C via a pertussis toxin-insensitive guanine nucleotide-binding regulatory protein in vascular smooth muscle cells. J Clin Invest 1990;85:653-658.
19. Eguchi S, Hirata Y, Imai T, Marumo F. Endothelin receptor subtypes are coupled to adenylate cyclase via different guanyl nucleotide-binding proteins in vasculature. Endocrinology 1993;132:524-529.
20. Kawanabe Y, Okamoto Y, Hashimoto N, Masaki T. Molecular mechanisms for activation of voltage-independent Ca2+ channels by endothelin-1/endothelin-A receptors. J Cardiovasc Pharmacol 2004;44 Suppl 1:S219-223.
21. Eguchi S, Hirata Y, Marumo F. Endothelin subtype B receptors are coupled to adenylate cyclase via inhibitory G protein in cultured bovine endothelial cells. J Cardiovasc Pharmacol 1993;22 Suppl 8:S161-163.
22. Masaki T, Miwa S, Sawamura T, Ninomiya H, Okamoto Y. Subcellular mechanisms of endothelin action in vascular system. Eur J Pharmacol 1999;375:133-138.
23. Gohla A, Offermanns S, Wilkie TM, Schultz G. Differential involvement of Galpha12 and Galpha13 in receptor-mediated stress fiber formation. J Biol Chem 1999;274:17901-17907.

127
24. Kurihara Y, Kurihara H, Suzuki H, Kodama T, Maemura K, Nagai R, et al. Elevated blood pressure and craniofacial abnormalities in mice deficient in endothelin-1. Nature 1994;368:703-710.
25. Kurihara Y, Kurihara H, Oda H, Maemura K, Nagai R, Ishikawa T, et al. Aortic arch malformations and ventricular septal defect in mice deficient in endothelin-1. J Clin Invest 1995;96:293-300.
26. Kurihara Y, Kurihara H, Maemura K, Kuwaki T, Kumada M, Yazaki Y. Impaired development of the thyroid and thymus in endothelin-1 knockout mice. J Cardiovasc Pharmacol 1995;26 Suppl 3:S13-16.
27. Zhao XS, Pan W, Bekeredjian R, Shohet RV. Endogenous endothelin-1 is required for cardiomyocyte survival in vivo. Circulation 2006;114:830-837.
28. Shohet RV, Kisanuki YY, Zhao XS, Siddiquee Z, Franco F, Yanagisawa M. Mice with cardiomyocyte-specific disruption of the endothelin-1 gene are resistant to hyperthyroid cardiac hypertrophy. Proc Natl Acad Sci U S A 2004;101:2088-2093.
29. Clouthier DE, Hosoda K, Richardson JA, Williams SC, Yanagisawa H, Kuwaki T, et al. Cranial and cardiac neural crest defects in endothelin-A receptor-deficient mice. Development 1998;125:813-824.
30. Hocher B, Thone-Reineke C, Rohmeiss P, Schmager F, Slowinski T, Burst V, et al. Endothelin-1 transgenic mice develop glomerulosclerosis, interstitial fibrosis, and renal cysts but not hypertension. J Clin Invest 1997;99:1380-1389.
31. Hocher B, Schwarz A, Fagan KA, Thone-Reineke C, El-Hag K, Kusserow H, et al. Pulmonary fibrosis and chronic lung inflammation in ET-1 transgenic mice. Am J Respir Cell Mol Biol 2000;23:19-26.
32. Hocher B, Liefeldt L, Thone-Reineke C, Orzechowski HD, Distler A, Bauer C, et al. Characterization of the renal phenotype of transgenic rats expressing the human endothelin-2 gene. Hypertension 1996;28:196-201.
33. Amiri F, Virdis A, Neves MF, Iglarz M, Seidah NG, Touyz RM, et al. Endothelium-restricted overexpression of human endothelin-1 causes vascular remodeling and endothelial dysfunction. Circulation 2004;110:2233-2240.
34. Yang LL, Gros R, Kabir MG, Sadi A, Gotlieb AI, Husain M, et al. Conditional cardiac overexpression of endothelin-1 induces inflammation and dilated cardiomyopathy in mice. Circulation 2004;109:255-261.
35. Baynash AG, Hosoda K, Giaid A, Richardson JA, Emoto N, Hammer RE, et al. Interaction of endothelin-3 with endothelin-B receptor is essential for development of epidermal melanocytes and enteric neurons. Cell 1994;79:1277-1285.

128
36. Bagnall AJ, Kelland NF, Gulliver-Sloan F, Davenport AP, Gray GA, Yanagisawa M, et al. Deletion of endothelial cell endothelin B receptors does not affect blood pressure or sensitivity to salt. Hypertension 2006;48:286-293.
37. Yanagisawa H, Yanagisawa M, Kapur RP, Richardson JA, Williams SC, Clouthier DE, et al. Dual genetic pathways of endothelin-mediated intercellular signaling revealed by targeted disruption of endothelin converting enzyme-1 gene. Development 1998;125:825-836.
38. Yanagisawa H, Hammer RE, Richardson JA, Emoto N, Williams SC, Takeda S, et al. Disruption of ECE-1 and ECE-2 reveals a role for endothelin-converting enzyme-2 in murine cardiac development. J Clin Invest 2000;105:1373-1382.
39. Hafstrom I, Ringertz B, Lundeberg T, Palmblad J. The effect of endothelin, neuropeptide Y, calcitonin gene-related peptide and substance P on neutrophil functions. Acta Physiol Scand 1993;148:341-346.
40. Halim A, Kanayama N, el Maradny E, Maehara K, Terao T. Activated neutrophil by endothelin-1 caused tissue damage in human umbilical cord. Thromb Res 1995;77:321-327.
41. Uchida Y, Ninomiya H, Sakamoto T, Lee JY, Endo T, Nomura A, et al. ET-1 released histamine from guinea pig pulmonary but not peritoneal mast cells. Biochem Biophys Res Commun 1992;189:1196-1201.
42. Zhang JS, Tan YR, Xiang Y, Luo ZQ, Qin XQ. Regulatory peptides modulate adhesion of polymorphonuclear leukocytes to bronchial epithelial cells through regulation of interleukins, ICAM-1 and NF-kappaB/IkappaB. Acta Biochim Biophys Sin (Shanghai) 2006;38:119-128.
43. Browatzki M, Schmidt J, Kubler W, Kranzhofer R. Endothelin-1 induces interleukin-6 release via activation of the transcription factor NF-kappaB in human vascular smooth muscle cells. Basic Res Cardiol 2000;95:98-105.
44. McMillen MA, Huribal M, Cunningham ME, Kumar R, Sumpio BE. Endothelin-1 increases intracellular calcium in human monocytes and causes production of interleukin-6. Crit Care Med 1995;23:34-40.
45. Browatzki M, Pfeiffer CA, Schmidt J, Kranzhofer R. Endothelin-1 induces CD40 but not IL-6 in human monocytes via the proinflammatory transcription factor NF-kappaB. Eur J Med Res 2005;10:197-201.
46. Gonsalves C, Kalra VK. Endothelin-1-induced macrophage inflammatory protein-1beta expression in monocytic cells involves hypoxia-inducible factor-1alpha and AP-1 and is negatively regulated by microRNA-195. J Immunol 2010;185:6253-6264.

129
47. Marini M, Carpi S, Bellini A, Patalano F, Mattoli S. Endothelin-1 induces increased fibronectin expression in human bronchial epithelial cells. Biochem Biophys Res Commun 1996;220:896-899.
48. Rizvi MA, Katwa L, Spadone DP, Myers PR. The effects of endothelin-1 on collagen type I and type III synthesis in cultured porcine coronary artery vascular smooth muscle cells. J Mol Cell Cardiol 1996;28:243-252.
49. Peacock AJ, Dawes KE, Shock A, Gray AJ, Reeves JT, Laurent GJ. Endothelin-1 and endothelin-3 induce chemotaxis and replication of pulmonary artery fibroblasts. Am J Respir Cell Mol Biol 1992;7:492-499.
50. Shi-Wen X, Denton CP, Dashwood MR, Holmes AM, Bou-Gharios G, Pearson JD, et al. Fibroblast matrix gene expression and connective tissue remodeling: role of endothelin-1. J Invest Dermatol 2001;116:417-425.
51. Hafizi S, Wharton J, Chester AH, Yacoub MH. Profibrotic effects of endothelin-1 via the ETA receptor in cultured human cardiac fibroblasts. Cell Physiol Biochem 2004;14:285-292.
52. Freund C, Schmidt-Ullrich R, Baurand A, Dunger S, Schneider W, Loser P, et al. Requirement of nuclear factor-kappaB in angiotensin II- and isoproterenol-induced cardiac hypertrophy in vivo. Circulation 2005;111:2319-2325.
53. Frantz S, Fraccarollo D, Wagner H, Behr TM, Jung P, Angermann CE, et al. Sustained activation of nuclear factor kappa B and activator protein 1 in chronic heart failure. Cardiovasc Res 2003;57:749-756.
54. Hamid T, Guo SZ, Kingery JR, Xiang X, Dawn B, Prabhu SD. Cardiomyocyte NF-kappaB p65 promotes adverse remodelling, apoptosis, and endoplasmic reticulum stress in heart failure. Cardiovasc Res 2011;89:129-138.
55. Kawamura N, Kubota T, Kawano S, Monden Y, Feldman AM, Tsutsui H, et al. Blockade of NF-kappaB improves cardiac function and survival without affecting inflammation in TNF-alpha-induced cardiomyopathy. Cardiovasc Res 2005;66:520-529.
56. Frantz S, Hu K, Bayer B, Gerondakis S, Strotmann J, Adamek A, et al. Absence of NF-kappaB subunit p50 improves heart failure after myocardial infarction. FASEB J 2006;20:1918-1920.
57. Shi-Wen X, Chen Y, Denton CP, Eastwood M, Renzoni EA, Bou-Gharios G, et al. Endothelin-1 promotes myofibroblast induction through the ETA receptor via a rac/phosphoinositide 3-kinase/Akt-dependent pathway and is essential for the enhanced contractile phenotype of fibrotic fibroblasts. Mol Biol Cell 2004;15:2707-2719.

130
58. Jain R, Shaul PW, Borok Z, Willis BC. Endothelin-1 induces alveolar epithelial-mesenchymal transition through endothelin type A receptor-mediated production of TGF-beta1. Am J Respir Cell Mol Biol 2007;37:38-47.
59. Dulce RA, Hurtado C, Ennis IL, Garciarena CD, Alvarez MC, Caldiz C, et al. Endothelin-1 induced hypertrophic effect in neonatal rat cardiomyocytes: involvement of Na+/H+ and Na+/Ca2+ exchangers. J Mol Cell Cardiol 2006;41:807-815.
60. Higazi DR, Fearnley CJ, Drawnel FM, Talasila A, Corps EM, Ritter O, et al. Endothelin-1-stimulated InsP3-induced Ca2+ release is a nexus for hypertrophic signaling in cardiac myocytes. Mol Cell 2009;33:472-482.
61. Hirotani S, Otsu K, Nishida K, Higuchi Y, Morita T, Nakayama H, et al. Involvement of nuclear factor-kappaB and apoptosis signal-regulating kinase 1 in G-protein-coupled receptor agonist-induced cardiomyocyte hypertrophy. Circulation 2002;105:509-515.
62. Liang F, Lu S, Gardner DG. Endothelin-dependent and -independent components of strain-activated brain natriuretic peptide gene transcription require extracellular signal regulated kinase and p38 mitogen-activated protein kinase. Hypertension 2000;35:188-192.
63. Komati H, Maharsy W, Beauregard J, Hayek S, Nemer M. ZFP260 is an inducer of cardiac hypertrophy and a nuclear mediator of endothelin-1 signaling. J Biol Chem 2011;286:1508-1516.
64. Pieske B, Beyermann B, Breu V, Loffler BM, Schlotthauer K, Maier LS, et al. Functional effects of endothelin and regulation of endothelin receptors in isolated human nonfailing and failing myocardium. Circulation 1999;99:1802-1809.
65. Kobayashi T, Miyauchi T, Sakai S, Kobayashi M, Yamaguchi I, Goto K, et al. Expression of endothelin-1, ETA and ETB receptors, and ECE and distribution of endothelin-1 in failing rat heart. Am J Physiol 1999;276:H1197-1206.
66. Wei CM, Lerman A, Rodeheffer RJ, McGregor CG, Brandt RR, Wright S, et al. Endothelin in human congestive heart failure. Circulation 1994;89:1580-1586.
67. Margulies KB, Hildebrand FL, Jr., Lerman A, Perrella MA, Burnett JC, Jr. Increased endothelin in experimental heart failure. Circulation 1990;82:2226-2230.
68. Arai M, Yoguchi A, Iso T, Takahashi T, Imai S, Murata K, et al. Endothelin-1 and its binding sites are upregulated in pressure overload cardiac hypertrophy. Am J Physiol 1995;268:H2084-2091.
69. Zolk O, Quattek J, Sitzler G, Schrader T, Nickenig G, Schnabel P, et al. Expression of endothelin-1, endothelin-converting enzyme, and endothelin receptors in chronic heart failure. Circulation 1999;99:2118-2123.

131
70. Pacher R, Stanek B, Hulsmann M, Koller-Strametz J, Berger R, Schuller M, et al. Prognostic impact of big endothelin-1 plasma concentrations compared with invasive hemodynamic evaluation in severe heart failure. J Am Coll Cardiol 1996;27:633-641.
71. Iwanaga Y, Kihara Y, Inagaki K, Onozawa Y, Yoneda T, Kataoka K, et al. Differential effects of angiotensin II versus endothelin-1 inhibitions in hypertrophic left ventricular myocardium during transition to heart failure. Circulation 2001;104:606-612.
72. Mulder P, Richard V, Derumeaux G, Hogie M, Henry JP, Lallemand F, et al. Role of endogenous endothelin in chronic heart failure: effect of long-term treatment with an endothelin antagonist on survival, hemodynamics, and cardiac remodeling. Circulation 1997;96:1976-1982.
73. Mulder P, Boujedaini H, Richard V, Henry JP, Renet S, Munter K, et al. Long-term survival and hemodynamics after endothelin-a receptor antagonism and angiotensin-converting enzyme inhibition in rats with chronic heart failure: monotherapy versus combination therapy. Circulation 2002;106:1159-1164.
74. Cowburn PJ, Cleland JG, McArthur JD, MacLean MR, McMurray JJ, Dargie HJ. Short-term haemodynamic effects of BQ-123, a selective endothelin ET(A)-receptor antagonist, in chronic heart failure. Lancet 1998;352:201-202.
75. Kiowski W, Sutsch G, Hunziker P, Muller P, Kim J, Oechslin E, et al. Evidence for endothelin-1-mediated vasoconstriction in severe chronic heart failure. Lancet 1995;346:732-736.
76. Spieker LE, Mitrovic V, Noll G, Pacher R, Schulze MR, Muntwyler J, et al. Acute hemodynamic and neurohumoral effects of selective ET(A) receptor blockade in patients with congestive heart failure. ET 003 Investigators. J Am Coll Cardiol 2000;35:1745-1752.
77. Torre-Amione G, Young JB, Colucci WS, Lewis BS, Pratt C, Cotter G, et al. Hemodynamic and clinical effects of tezosentan, an intravenous dual endothelin receptor antagonist, in patients hospitalized for acute decompensated heart failure. J Am Coll Cardiol 2003;42:140-147.
78. Teerlink JR, McMurray JJ, Bourge RC, Cleland JG, Cotter G, Jondeau G, et al. Tezosentan in patients with acute heart failure: design of the Value of Endothelin Receptor Inhibition with Tezosentan in Acute heart failure Study (VERITAS). Am Heart J 2005;150:46-53.
79. Mylona P, Cleland JG. Update of REACH-1 and MERIT-HF clinical trials in heart failure. Cardio.net Editorial Team. Eur J Heart Fail 1999;1:197-200.
80. Kalra PR, Moon JC, Coats AJ. Do results of the ENABLE (Endothelin Antagonist Bosentan for Lowering Cardiac Events in Heart Failure) study spell the end for non-selective endothelin antagonism in heart failure? Int J Cardiol 2002;85:195-197.

132
81. Anand I, McMurray J, Cohn JN, Konstam MA, Notter T, Quitzau K, et al. Long-term effects of darusentan on left-ventricular remodelling and clinical outcomes in the EndothelinA Receptor Antagonist Trial in Heart Failure (EARTH): randomised, double-blind, placebo-controlled trial. Lancet 2004;364:347-354.
82. Mellin V, Jeng AY, Monteil C, Renet S, Henry JP, Thuillez C, et al. Triple ACE-ECE-NEP inhibition in heart failure: a comparison with ACE and dual ECE-NEP inhibition. J Cardiovasc Pharmacol 2005;46:390-397.
83. Mulder P, Barbier S, Monteil C, Jeng AY, Henry JP, Renet S, et al. Sustained improvement of cardiac function and prevention of cardiac remodeling after long-term dual ECE-NEP inhibition in rats with congestive heart failure. J Cardiovasc Pharmacol 2004;43:489-494.
84. Emoto N, Raharjo SB, Isaka D, Masuda S, Adiarto S, Jeng AY, et al. Dual ECE/NEP inhibition on cardiac and neurohumoral function during the transition from hypertrophy to heart failure in rats. Hypertension 2005;45:1145-1152.
85. Tabrizchi R. SLV-306. Solvay. Curr Opin Investig Drugs 2003;4:329-332.
86. Wada A, Tsutamoto T, Ohnishi M, Sawaki M, Fukai D, Maeda Y, et al. Effects of a specific endothelin-converting enzyme inhibitor on cardiac, renal, and neurohumoral functions in congestive heart failure: comparison of effects with those of endothelin A receptor antagonism. Circulation 1999;99:570-577.
87. Fleischer S, Ogunbunmi EM, Dixon MC, Fleer EA. Localization of Ca2+ release channels with ryanodine in junctional terminal cisternae of sarcoplasmic reticulum of fast skeletal muscle. Proc Natl Acad Sci U S A 1985;82:7256-7259.
88. Pessah IN, Waterhouse AL, Casida JE. The calcium-ryanodine receptor complex of skeletal and cardiac muscle. Biochem Biophys Res Commun 1985;128:449-456.
89. Marks AR, Marx SO, Reiken S. Regulation of ryanodine receptors via macromolecular complexes: a novel role for leucine/isoleucine zippers. Trends Cardiovasc Med 2002;12:166-170.
90. Korzick DH. Regulation of cardiac excitation-contraction coupling: a cellular update. Adv Physiol Educ 2003;27:192-200.
91. Priori SG, Napolitano C, Tiso N, Memmi M, Vignati G, Bloise R, et al. Mutations in the cardiac ryanodine receptor gene (hRyR2) underlie catecholaminergic polymorphic ventricular tachycardia. Circulation 2001;103:196-200.
92. Lytton J. Na+/Ca2+ exchangers: three mammalian gene families control Ca2+ transport. Biochem J 2007;406:365-382.
93. Tada M. Calcium cycling proteins of the cardiac sarcoplasmic reticulum. Circ J 2003;67:729-737.

133
94. Frank KF, Bolck B, Erdmann E, Schwinger RH. Sarcoplasmic reticulum Ca2+-ATPase modulates cardiac contraction and relaxation. Cardiovasc Res 2003;57:20-27.
95. Posch MG, Perrot A, Geier C, Boldt LH, Schmidt G, Lehmkuhl HB, et al. Genetic deletion of arginine 14 in phospholamban causes dilated cardiomyopathy with attenuated electrocardiographic R amplitudes. Heart Rhythm 2009;6:480-486.
96. Schmitt JP, Kamisago M, Asahi M, Li GH, Ahmad F, Mende U, et al. Dilated cardiomyopathy and heart failure caused by a mutation in phospholamban. Science 2003;299:1410-1413.
97. Haghighi K, Kolokathis F, Gramolini AO, Waggoner JR, Pater L, Lynch RA, et al. A mutation in the human phospholamban gene, deleting arginine 14, results in lethal, hereditary cardiomyopathy. Proc Natl Acad Sci U S A 2006;103:1388-1393.
98. Qu Y, Boutjdir M. Gene expression of SERCA2a and L- and T-type Ca channels during human heart development. Pediatr Res 2001;50:569-574.
99. Martinez ML, Heredia MP, Delgado C. Expression of T-type Ca(2+) channels in ventricular cells from hypertrophied rat hearts. J Mol Cell Cardiol 1999;31:1617-1625.
100. Thomsen MB, Wang C, Ozgen N, Wang HG, Rosen MR, Pitt GS. Accessory subunit KChIP2 modulates the cardiac L-type calcium current. Circ Res 2009;104:1382-1389.
101. Antzelevitch C, Pollevick GD, Cordeiro JM, Casis O, Sanguinetti MC, Aizawa Y, et al. Loss-of-function mutations in the cardiac calcium channel underlie a new clinical entity characterized by ST-segment elevation, short QT intervals, and sudden cardiac death. Circulation 2007;115:442-449.
102. Cordeiro JM, Marieb M, Pfeiffer R, Calloe K, Burashnikov E, Antzelevitch C. Accelerated inactivation of the L-type calcium current due to a mutation in CACNB2b underlies Brugada syndrome. J Mol Cell Cardiol 2009;46:695-703.
103. Splawski I, Timothy KW, Decher N, Kumar P, Sachse FB, Beggs AH, et al. Severe arrhythmia disorder caused by cardiac L-type calcium channel mutations. Proc Natl Acad Sci U S A 2005;102:8089-8096; discussion 8086-8088.
104. Facer P, Punjabi PP, Abrari A, Kaba RA, Severs NJ, Chambers J, et al. Localisation of SCN10A gene product Na(v)1.8 and novel pain-related ion channels in human heart. Int Heart J 2011;52:146-152.
105. Haufe V, Chamberland C, Dumaine R. The promiscuous nature of the cardiac sodium current. J Mol Cell Cardiol 2007;42:469-477.
106. Maier SK, Westenbroek RE, Schenkman KA, Feigl EO, Scheuer T, Catterall WA. An unexpected role for brain-type sodium channels in coupling of cell surface depolarization to contraction in the heart. Proc Natl Acad Sci U S A 2002;99:4073-4078.

134
107. Maier SK, Westenbroek RE, Yamanushi TT, Dobrzynski H, Boyett MR, Catterall WA, et al. An unexpected requirement for brain-type sodium channels for control of heart rate in the mouse sinoatrial node. Proc Natl Acad Sci U S A 2003;100:3507-3512.
108. Chambers JC, Zhao J, Terracciano CM, Bezzina CR, Zhang W, Kaba R, et al. Genetic variation in SCN10A influences cardiac conduction. Nat Genet 2010;42:149-152.
109. Shang LL, Sanyal S, Pfahnl AE, Jiao Z, Allen J, Liu H, et al. NF-kappaB-dependent transcriptional regulation of the cardiac scn5a sodium channel by angiotensin II. Am J Physiol Cell Physiol 2008;294:C372-379.
110. Papadatos GA, Wallerstein PM, Head CE, Ratcliff R, Brady PA, Benndorf K, et al. Slowed conduction and ventricular tachycardia after targeted disruption of the cardiac sodium channel gene Scn5a. Proc Natl Acad Sci U S A 2002;99:6210-6215.
111. van Veen TA, Stein M, Royer A, Le Quang K, Charpentier F, Colledge WH, et al. Impaired impulse propagation in Scn5a-knockout mice: combined contribution of excitability, connexin expression, and tissue architecture in relation to aging. Circulation 2005;112:1927-1935.
112. Wang Q, Shen J, Splawski I, Atkinson D, Li Z, Robinson JL, et al. SCN5A mutations associated with an inherited cardiac arrhythmia, long QT syndrome. Cell 1995;80:805-811.
113. Chen Q, Kirsch GE, Zhang D, Brugada R, Brugada J, Brugada P, et al. Genetic basis and molecular mechanism for idiopathic ventricular fibrillation. Nature 1998;392:293-296.
114. Tan HL, Bink-Boelkens MT, Bezzina CR, Viswanathan PC, Beaufort-Krol GC, van Tintelen PJ, et al. A sodium-channel mutation causes isolated cardiac conduction disease. Nature 2001;409:1043-1047.
115. Schott JJ, Alshinawi C, Kyndt F, Probst V, Hoorntje TM, Hulsbeek M, et al. Cardiac conduction defects associate with mutations in SCN5A. Nat Genet 1999;23:20-21.
116. Wang DW, Viswanathan PC, Balser JR, George AL, Jr., Benson DW. Clinical, genetic, and biophysical characterization of SCN5A mutations associated with atrioventricular conduction block. Circulation 2002;105:341-346.
117. Wang DW, Desai RR, Crotti L, Arnestad M, Insolia R, Pedrazzini M, et al. Cardiac sodium channel dysfunction in sudden infant death syndrome. Circulation 2007;115:368-376.
118. Benson DW, Wang DW, Dyment M, Knilans TK, Fish FA, Strieper MJ, et al. Congenital sick sinus syndrome caused by recessive mutations in the cardiac sodium channel gene (SCN5A). J Clin Invest 2003;112:1019-1028.

135
119. Darbar D, Kannankeril PJ, Donahue BS, Kucera G, Stubblefield T, Haines JL, et al. Cardiac sodium channel (SCN5A) variants associated with atrial fibrillation. Circulation 2008;117:1927-1935.
120. McNair WP, Ku L, Taylor MR, Fain PR, Dao D, Wolfel E, et al. SCN5A mutation associated with dilated cardiomyopathy, conduction disorder, and arrhythmia. Circulation 2004;110:2163-2167.
121. Vatta M, Ackerman MJ, Ye B, Makielski JC, Ughanze EE, Taylor EW, et al. Mutant caveolin-3 induces persistent late sodium current and is associated with long-QT syndrome. Circulation 2006;114:2104-2112.
122. Cronk LB, Ye B, Kaku T, Tester DJ, Vatta M, Makielski JC, et al. Novel mechanism for sudden infant death syndrome: persistent late sodium current secondary to mutations in caveolin-3. Heart Rhythm 2007;4:161-166.
123. Ueda K, Valdivia C, Medeiros-Domingo A, Tester DJ, Vatta M, Farrugia G, et al. Syntrophin mutation associated with long QT syndrome through activation of the nNOS-SCN5A macromolecular complex. Proc Natl Acad Sci U S A 2008;105:9355-9360.
124. Wu G, Ai T, Kim JJ, Mohapatra B, Xi Y, Li Z, et al. alpha-1-syntrophin mutation and the long-QT syndrome: a disease of sodium channel disruption. Circ Arrhythm Electrophysiol 2008;1:193-201.
125. Watanabe H, Koopmann TT, Le Scouarnec S, Yang T, Ingram CR, Schott JJ, et al. Sodium channel beta1 subunit mutations associated with Brugada syndrome and cardiac conduction disease in humans. J Clin Invest 2008;118:2260-2268.
126. Watanabe H, Darbar D, Kaiser DW, Jiramongkolchai K, Chopra S, Donahue BS, et al. Mutations in sodium channel beta1- and beta2-subunits associated with atrial fibrillation. Circ Arrhythm Electrophysiol 2009;2:268-275.
127. Hu D, Barajas-Martinez H, Burashnikov E, Springer M, Wu Y, Varro A, et al. A mutation in the beta 3 subunit of the cardiac sodium channel associated with Brugada ECG phenotype. Circ Cardiovasc Genet 2009;2:270-278.
128. Medeiros-Domingo A, Kaku T, Tester DJ, Iturralde-Torres P, Itty A, Ye B, et al. SCN4B-encoded sodium channel beta4 subunit in congenital long-QT syndrome. Circulation 2007;116:134-142.
129. Guo W, Jung WE, Marionneau C, Aimond F, Xu H, Yamada KA, et al. Targeted deletion of Kv4.2 eliminates I(to,f) and results in electrical and molecular remodeling, with no evidence of ventricular hypertrophy or myocardial dysfunction. Circ Res 2005;97:1342-1350.

136
130. Kuo HC, Cheng CF, Clark RB, Lin JJ, Lin JL, Hoshijima M, et al. A defect in the Kv channel-interacting protein 2 (KChIP2) gene leads to a complete loss of I(to) and confers susceptibility to ventricular tachycardia. Cell 2001;107:801-813.
131. Deschenes I, Armoundas AA, Jones SP, Tomaselli GF. Post-transcriptional gene silencing of KChIP2 and Navbeta1 in neonatal rat cardiac myocytes reveals a functional association between Na and Ito currents. J Mol Cell Cardiol 2008;45:336-346.
132. Lu Z, Abe J, Taunton J, Lu Y, Shishido T, McClain C, et al. Reactive oxygen species-induced activation of p90 ribosomal S6 kinase prolongs cardiac repolarization through inhibiting outward K+ channel activity. Circ Res 2008;103:269-278.
133. Gomes P, Saito T, Del Corsso C, Alioua A, Eghbali M, Toro L, et al. Identification of a functional interaction between Kv4.3 channels and c-Src tyrosine kinase. Biochim Biophys Acta 2008;1783:1884-1892.
134. Ruiz-Gomez A, Mellstrom B, Tornero D, Morato E, Savignac M, Holguin H, et al. G protein-coupled receptor kinase 2-mediated phosphorylation of downstream regulatory element antagonist modulator regulates membrane trafficking of Kv4.2 potassium channel. J Biol Chem 2007;282:1205-1215.
135. Niwa N, Nerbonne JM. Molecular determinants of cardiac transient outward potassium current (I(to)) expression and regulation. J Mol Cell Cardiol 2010;48:12-25.
136. Delpon E, Cordeiro JM, Nunez L, Thomsen PE, Guerchicoff A, Pollevick GD, et al. Functional effects of KCNE3 mutation and its role in the development of Brugada syndrome. Circ Arrhythm Electrophysiol 2008;1:209-218.
137. Lai LP, Su MJ, Yeh HM, Lin JL, Chiang FT, Hwang JJ, et al. Association of the human minK gene 38G allele with atrial fibrillation: evidence of possible genetic control on the pathogenesis of atrial fibrillation. Am Heart J 2002;144:485-490.
138. Splawski I, Tristani-Firouzi M, Lehmann MH, Sanguinetti MC, Keating MT. Mutations in the hminK gene cause long QT syndrome and suppress IKs function. Nat Genet 1997;17:338-340.
139. Yang Y, Xia M, Jin Q, Bendahhou S, Shi J, Chen Y, et al. Identification of a KCNE2 gain-of-function mutation in patients with familial atrial fibrillation. Am J Hum Genet 2004;75:899-905.
140. Ehrlich JR, Pourrier M, Weerapura M, Ethier N, Marmabachi AM, Hebert TE, et al. KvLQT1 modulates the distribution and biophysical properties of HERG. A novel alpha-subunit interaction between delayed rectifier currents. J Biol Chem 2004;279:1233-1241.

137
141. Chouabe C, Neyroud N, Guicheney P, Lazdunski M, Romey G, Barhanin J. Properties of KvLQT1 K+ channel mutations in Romano-Ward and Jervell and Lange-Nielsen inherited cardiac arrhythmias. EMBO J 1997;16:5472-5479.
142. Peroz D, Dahimene S, Baro I, Loussouarn G, Merot J. LQT1-associated mutations increase KCNQ1 proteasomal degradation independently of Derlin-1. J Biol Chem 2009;284:5250-5256.
143. Sanguinetti MC, Curran ME, Spector PS, Keating MT. Spectrum of HERG K+-channel dysfunction in an inherited cardiac arrhythmia. Proc Natl Acad Sci U S A 1996;93:2208-2212.
144. Chen YH, Xu SJ, Bendahhou S, Wang XL, Wang Y, Xu WY, et al. KCNQ1 gain-of-function mutation in familial atrial fibrillation. Science 2003;299:251-254.
145. Bellocq C, van Ginneken AC, Bezzina CR, Alders M, Escande D, Mannens MM, et al. Mutation in the KCNQ1 gene leading to the short QT-interval syndrome. Circulation 2004;109:2394-2397.
146. Cordeiro JM, Brugada R, Wu YS, Hong K, Dumaine R. Modulation of I(Kr) inactivation by mutation N588K in KCNH2: a link to arrhythmogenesis in short QT syndrome. Cardiovasc Res 2005;67:498-509.
147. Xia M, Jin Q, Bendahhou S, He Y, Larroque MM, Chen Y, et al. A Kir2.1 gain-of-function mutation underlies familial atrial fibrillation. Biochem Biophys Res Commun 2005;332:1012-1019.
148. Priori SG, Pandit SV, Rivolta I, Berenfeld O, Ronchetti E, Dhamoon A, et al. A novel form of short QT syndrome (SQT3) is caused by a mutation in the KCNJ2 gene. Circ Res 2005;96:800-807.
149. Plaster NM, Tawil R, Tristani-Firouzi M, Canun S, Bendahhou S, Tsunoda A, et al. Mutations in Kir2.1 cause the developmental and episodic electrical phenotypes of Andersen's syndrome. Cell 2001;105:511-519.
150. Eckhardt LL, Farley AL, Rodriguez E, Ruwaldt K, Hammill D, Tester DJ, et al. KCNJ2 mutations in arrhythmia patients referred for LQT testing: a mutation T305A with novel effect on rectification properties. Heart Rhythm 2007;4:323-329.
151. Solan JL, Lampe PD. Connexin43 phosphorylation: structural changes and biological effects. Biochem J 2009;419:261-272.
152. Laing JG, Tadros PN, Westphale EM, Beyer EC. Degradation of connexin43 gap junctions involves both the proteasome and the lysosome. Exp Cell Res 1997;236:482-492.

138
153. Lampe PD, TenBroek EM, Burt JM, Kurata WE, Johnson RG, Lau AF. Phosphorylation of connexin43 on serine368 by protein kinase C regulates gap junctional communication. J Cell Biol 2000;149:1503-1512.
154. Lin R, Warn-Cramer BJ, Kurata WE, Lau AF. v-Src phosphorylation of connexin 43 on Tyr247 and Tyr265 disrupts gap junctional communication. J Cell Biol 2001;154:815-827.
155. Cooper CD, Lampe PD. Casein kinase 1 regulates connexin-43 gap junction assembly. J Biol Chem 2002;277:44962-44968.
156. TenBroek EM, Lampe PD, Solan JL, Reynhout JK, Johnson RG. Ser364 of connexin43 and the upregulation of gap junction assembly by cAMP. J Cell Biol 2001;155:1307-1318.
157. Paulson AF, Lampe PD, Meyer RA, TenBroek E, Atkinson MM, Walseth TF, et al. Cyclic AMP and LDL trigger a rapid enhancement in gap junction assembly through a stimulation of connexin trafficking. J Cell Sci 2000;113 ( Pt 17):3037-3049.
158. Cottrell GT, Lin R, Warn-Cramer BJ, Lau AF, Burt JM. Mechanism of v-Src- and mitogen-activated protein kinase-induced reduction of gap junction communication. Am J Physiol Cell Physiol 2003;284:C511-520.
159. Giepmans BN. Gap junctions and connexin-interacting proteins. Cardiovasc Res 2004;62:233-245.
160. Palatinus JA, O'Quinn MP, Barker RJ, Harris BS, Jourdan J, Gourdie RG. ZO-1 determines adherens and gap junction localization at intercalated disks. Am J Physiol Heart Circ Physiol 2011;300:H583-594.
161. Lan Z, Kurata WE, Martyn KD, Jin C, Lau AF. Novel rab GAP-like protein, CIP85, interacts with connexin43 and induces its degradation. Biochemistry 2005;44:2385-2396.
162. Teunissen BE, Jansen AT, van Amersfoorth SC, O'Brien TX, Jongsma HJ, Bierhuizen MF. Analysis of the rat connexin 43 proximal promoter in neonatal cardiomyocytes. Gene 2003;322:123-136.
163. Boogerd KJ, Wong LY, Christoffels VM, Klarenbeek M, Ruijter JM, Moorman AF, et al. Msx1 and Msx2 are functional interacting partners of T-box factors in the regulation of Connexin43. Cardiovasc Res 2008;78:485-493.
164. Reaume AG, de Sousa PA, Kulkarni S, Langille BL, Zhu D, Davies TC, et al. Cardiac malformation in neonatal mice lacking connexin43. Science 1995;267:1831-1834.
165. Vaidya D, Tamaddon HS, Lo CW, Taffet SM, Delmar M, Morley GE, et al. Null mutation of connexin43 causes slow propagation of ventricular activation in the late stages of mouse embryonic development. Circ Res 2001;88:1196-1202.

139
166. Beauchamp P, Choby C, Desplantez T, de Peyer K, Green K, Yamada KA, et al. Electrical propagation in synthetic ventricular myocyte strands from germline connexin43 knockout mice. Circ Res 2004;95:170-178.
167. Thomas SA, Schuessler RB, Berul CI, Beardslee MA, Beyer EC, Mendelsohn ME, et al. Disparate effects of deficient expression of connexin43 on atrial and ventricular conduction: evidence for chamber-specific molecular determinants of conduction. Circulation 1998;97:686-691.
168. Guerrero PA, Schuessler RB, Davis LM, Beyer EC, Johnson CM, Yamada KA, et al. Slow ventricular conduction in mice heterozygous for a connexin43 null mutation. J Clin Invest 1997;99:1991-1998.
169. Gutstein DE, Morley GE, Tamaddon H, Vaidya D, Schneider MD, Chen J, et al. Conduction slowing and sudden arrhythmic death in mice with cardiac-restricted inactivation of connexin43. Circ Res 2001;88:333-339.
170. Eckardt D, Theis M, Degen J, Ott T, van Rijen HV, Kirchhoff S, et al. Functional role of connexin43 gap junction channels in adult mouse heart assessed by inducible gene deletion. J Mol Cell Cardiol 2004;36:101-110.
171. Gutstein DE, Morley GE, Vaidya D, Liu F, Chen FL, Stuhlmann H, et al. Heterogeneous expression of Gap junction channels in the heart leads to conduction defects and ventricular dysfunction. Circulation 2001;104:1194-1199.
172. Kirchhoff S, Nelles E, Hagendorff A, Kruger O, Traub O, Willecke K. Reduced cardiac conduction velocity and predisposition to arrhythmias in connexin40-deficient mice. Curr Biol 1998;8:299-302.
173. Bagwe S, Berenfeld O, Vaidya D, Morley GE, Jalife J. Altered right atrial excitation and propagation in connexin40 knockout mice. Circulation 2005;112:2245-2253.
174. Tamaddon HS, Vaidya D, Simon AM, Paul DL, Jalife J, Morley GE. High-resolution optical mapping of the right bundle branch in connexin40 knockout mice reveals slow conduction in the specialized conduction system. Circ Res 2000;87:929-936.
175. Hagendorff A, Schumacher B, Kirchhoff S, Luderitz B, Willecke K. Conduction disturbances and increased atrial vulnerability in Connexin40-deficient mice analyzed by transesophageal stimulation. Circulation 1999;99:1508-1515.
176. Kreuzberg MM, Schrickel JW, Ghanem A, Kim JS, Degen J, Janssen-Bienhold U, et al. Connexin30.2 containing gap junction channels decelerate impulse propagation through the atrioventricular node. Proc Natl Acad Sci U S A 2006;103:5959-5964.
177. Kruger O, Plum A, Kim JS, Winterhager E, Maxeiner S, Hallas G, et al. Defective vascular development in connexin 45-deficient mice. Development 2000;127:4179-4193.

140
178. Kumai M, Nishii K, Nakamura K, Takeda N, Suzuki M, Shibata Y. Loss of connexin45 causes a cushion defect in early cardiogenesis. Development 2000;127:3501-3512.
179. Nishii K, Kumai M, Egashira K, Miwa T, Hashizume K, Miyano Y, et al. Mice lacking connexin45 conditionally in cardiac myocytes display embryonic lethality similar to that of germline knockout mice without endocardial cushion defect. Cell Commun Adhes 2003;10:365-369.
180. Studer R, Reinecke H, Bilger J, Eschenhagen T, Bohm M, Hasenfuss G, et al. Gene expression of the cardiac Na(+)-Ca2+ exchanger in end-stage human heart failure. Circ Res 1994;75:443-453.
181. O'Rourke B, Kass DA, Tomaselli GF, Kaab S, Tunin R, Marban E. Mechanisms of altered excitation-contraction coupling in canine tachycardia-induced heart failure, I: experimental studies. Circ Res 1999;84:562-570.
182. Shao Q, Ren B, Saini HK, Netticadan T, Takeda N, Dhalla NS. Sarcoplasmic reticulum Ca2+ transport and gene expression in congestive heart failure are modified by imidapril treatment. Am J Physiol Heart Circ Physiol 2005;288:H1674-1682.
183. Mishra S, Gupta RC, Tiwari N, Sharov VG, Sabbah HN. Molecular mechanisms of reduced sarcoplasmic reticulum Ca(2+) uptake in human failing left ventricular myocardium. J Heart Lung Transplant 2002;21:366-373.
184. Netticadan T, Temsah RM, Kawabata K, Dhalla NS. Sarcoplasmic reticulum Ca(2+)/Calmodulin-dependent protein kinase is altered in heart failure. Circ Res 2000;86:596-605.
185. Gupta RC, Mishra S, Mishima T, Goldstein S, Sabbah HN. Reduced sarcoplasmic reticulum Ca(2+)-uptake and expression of phospholamban in left ventricular myocardium of dogs with heart failure. J Mol Cell Cardiol 1999;31:1381-1389.
186. Xiong W, Tian Y, DiSilvestre D, Tomaselli GF. Transmural heterogeneity of Na+-Ca2+ exchange: evidence for differential expression in normal and failing hearts. Circ Res 2005;97:207-209.
187. Flesch M, Schwinger RH, Schiffer F, Frank K, Sudkamp M, Kuhn-Regnier F, et al. Evidence for functional relevance of an enhanced expression of the Na(+)-Ca2+ exchanger in failing human myocardium. Circulation 1996;94:992-1002.
188. Marx SO, Reiken S, Hisamatsu Y, Jayaraman T, Burkhoff D, Rosemblit N, et al. PKA phosphorylation dissociates FKBP12.6 from the calcium release channel (ryanodine receptor): defective regulation in failing hearts. Cell 2000;101:365-376.
189. Go LO, Moschella MC, Watras J, Handa KK, Fyfe BS, Marks AR. Differential regulation of two types of intracellular calcium release channels during end-stage heart failure. J Clin Invest 1995;95:888-894.

141
190. Takahashi T, Allen PD, Lacro RV, Marks AR, Dennis AR, Schoen FJ, et al. Expression of dihydropyridine receptor (Ca2+ channel) and calsequestrin genes in the myocardium of patients with end-stage heart failure. J Clin Invest 1992;90:927-935.
191. Balijepalli RC, Lokuta AJ, Maertz NA, Buck JM, Haworth RA, Valdivia HH, et al. Depletion of T-tubules and specific subcellular changes in sarcolemmal proteins in tachycardia-induced heart failure. Cardiovasc Res 2003;59:67-77.
192. Aimond F, Alvarez JL, Rauzier JM, Lorente P, Vassort G. Ionic basis of ventricular arrhythmias in remodeled rat heart during long-term myocardial infarction. Cardiovasc Res 1999;42:402-415.
193. Zicha S, Maltsev VA, Nattel S, Sabbah HN, Undrovinas AI. Post-transcriptional alterations in the expression of cardiac Na+ channel subunits in chronic heart failure. J Mol Cell Cardiol 2004;37:91-100.
194. Valdivia CR, Chu WW, Pu J, Foell JD, Haworth RA, Wolff MR, et al. Increased late sodium current in myocytes from a canine heart failure model and from failing human heart. J Mol Cell Cardiol 2005;38:475-483.
195. Shang LL, Pfahnl AE, Sanyal S, Jiao Z, Allen J, Banach K, et al. Human heart failure is associated with abnormal C-terminal splicing variants in the cardiac sodium channel. Circ Res 2007;101:1146-1154.
196. Nuss HB, Houser SR. T-type Ca2+ current is expressed in hypertrophied adult feline left ventricular myocytes. Circ Res 1993;73:777-782.
197. Gidh-Jain M, Huang B, Jain P, el-Sherif N. Differential expression of voltage-gated K+ channel genes in left ventricular remodeled myocardium after experimental myocardial infarction. Circ Res 1996;79:669-675.
198. Kaab S, Dixon J, Duc J, Ashen D, Nabauer M, Beuckelmann DJ, et al. Molecular basis of transient outward potassium current downregulation in human heart failure: a decrease in Kv4.3 mRNA correlates with a reduction in current density. Circulation 1998;98:1383-1393.
199. Zicha S, Xiao L, Stafford S, Cha TJ, Han W, Varro A, et al. Transmural expression of transient outward potassium current subunits in normal and failing canine and human hearts. J Physiol 2004;561:735-748.
200. Rose J, Armoundas AA, Tian Y, DiSilvestre D, Burysek M, Halperin V, et al. Molecular correlates of altered expression of potassium currents in failing rabbit myocardium. Am J Physiol Heart Circ Physiol 2005;288:H2077-2087.
201. Li GR, Lau CP, Ducharme A, Tardif JC, Nattel S. Transmural action potential and ionic current remodeling in ventricles of failing canine hearts. Am J Physiol Heart Circ Physiol 2002;283:H1031-1041.

142
202. Li GR, Lau CP, Leung TK, Nattel S. Ionic current abnormalities associated with prolonged action potentials in cardiomyocytes from diseased human right ventricles. Heart Rhythm 2004;1:460-468.
203. Tsuji Y, Zicha S, Qi XY, Kodama I, Nattel S. Potassium channel subunit remodeling in rabbits exposed to long-term bradycardia or tachycardia: discrete arrhythmogenic consequences related to differential delayed-rectifier changes. Circulation 2006;113:345-355.
204. Severs NJ, Bruce AF, Dupont E, Rothery S. Remodelling of gap junctions and connexin expression in diseased myocardium. Cardiovasc Res 2008;80:9-19.
205. Dupont E, Matsushita T, Kaba RA, Vozzi C, Coppen SR, Khan N, et al. Altered connexin expression in human congestive heart failure. J Mol Cell Cardiol 2001;33:359-371.
206. Akar FG, Nass RD, Hahn S, Cingolani E, Shah M, Hesketh GG, et al. Dynamic changes in conduction velocity and gap junction properties during development of pacing-induced heart failure. Am J Physiol Heart Circ Physiol 2007;293:H1223-1230.
207. Muth JN, Bodi I, Lewis W, Varadi G, Schwartz A. A Ca(2+)-dependent transgenic model of cardiac hypertrophy: A role for protein kinase Calpha. Circulation 2001;103:140-147.
208. del Monte F, Williams E, Lebeche D, Schmidt U, Rosenzweig A, Gwathmey JK, et al. Improvement in survival and cardiac metabolism after gene transfer of sarcoplasmic reticulum Ca(2+)-ATPase in a rat model of heart failure. Circulation 2001;104:1424-1429.
209. Miyamoto MI, del Monte F, Schmidt U, DiSalvo TS, Kang ZB, Matsui T, et al. Adenoviral gene transfer of SERCA2a improves left-ventricular function in aortic-banded rats in transition to heart failure. Proc Natl Acad Sci U S A 2000;97:793-798.
210. Iwanaga Y, Hoshijima M, Gu Y, Iwatate M, Dieterle T, Ikeda Y, et al. Chronic phospholamban inhibition prevents progressive cardiac dysfunction and pathological remodeling after infarction in rats. J Clin Invest 2004;113:727-736.
211. Hoshijima M, Ikeda Y, Iwanaga Y, Minamisawa S, Date MO, Gu Y, et al. Chronic suppression of heart-failure progression by a pseudophosphorylated mutant of phospholamban via in vivo cardiac rAAV gene delivery. Nat Med 2002;8:864-871.
212. Jessup M, Greenberg B, Mancini D, Cappola T, Pauly DF, Jaski B, et al. Calcium Upregulation by Percutaneous Administration of Gene Therapy in Cardiac Disease (CUPID): a phase 2 trial of intracoronary gene therapy of sarcoplasmic reticulum Ca2+-ATPase in patients with advanced heart failure. Circulation 2011;124:304-313.

143
213. Nguyen TP, Wang DW, Rhodes TH, George AL, Jr. Divergent biophysical defects caused by mutant sodium channels in dilated cardiomyopathy with arrhythmia. Circ Res 2008;102:364-371.
214. Soltysinska E, Olesen SP, Christ T, Wettwer E, Varro A, Grunnet M, et al. Transmural expression of ion channels and transporters in human nondiseased and end-stage failing hearts. Pflugers Arch 2009;459:11-23.
215. Fatini C, Sticchi E, Marcucci R, Verdiani V, Nozzoli C, Vassallo C, et al. S38G single-nucleotide polymorphism at the KCNE1 locus is associated with heart failure. Heart Rhythm 2010;7:363-367.
216. Bruce AF, Rothery S, Dupont E, Severs NJ. Gap junction remodelling in human heart failure is associated with increased interaction of connexin43 with ZO-1. Cardiovasc Res 2008;77:757-765.
217. Yamada KA, Rogers JG, Sundset R, Steinberg TH, Saffitz JE. Up-regulation of connexin45 in heart failure. J Cardiovasc Electrophysiol 2003;14:1205-1212.
218. Maguy A, Le Bouter S, Comtois P, Chartier D, Villeneuve L, Wakili R, et al. Ion channel subunit expression changes in cardiac Purkinje fibers: a potential role in conduction abnormalities associated with congestive heart failure. Circ Res 2009;104:1113-1122.
219. Boixel C, Dinanian S, Lang-Lazdunski L, Mercadier JJ, Hatem SN. Characterization of effects of endothelin-1 on the L-type Ca2+ current in human atrial myocytes. Am J Physiol Heart Circ Physiol 2001;281:H764-773.
220. Izumi T, Kihara Y, Sarai N, Yoneda T, Iwanaga Y, Inagaki K, et al. Reinduction of T-type calcium channels by endothelin-1 in failing hearts in vivo and in adult rat ventricular myocytes in vitro. Circulation 2003;108:2530-2535.
221. Zhang Y, Huang ZJ, Dai DZ, Feng Y, Na T, Tang XY, et al. Downregulated FKBP12.6 expression and upregulated endothelin signaling contribute to elevated diastolic calcium and arrhythmogenesis in rat cardiomyopathy produced by l-thyroxin. Int J Cardiol 2008;130:463-471.
222. Li X, Zima AV, Sheikh F, Blatter LA, Chen J. Endothelin-1-induced arrhythmogenic Ca2+ signaling is abolished in atrial myocytes of inositol-1,4,5-trisphosphate(IP3)-receptor type 2-deficient mice. Circ Res 2005;96:1274-1281.
223. Kiesecker C, Zitron E, Scherer D, Lueck S, Bloehs R, Scholz EP, et al. Regulation of cardiac inwardly rectifying potassium current IK1 and Kir2.x channels by endothelin-1. J Mol Med 2006;84:46-56.
224. Puglisi JL, Yuan W, Timofeyev V, Myers RE, Chiamvimonvat N, Samarel AM, et al. Phorbol ester and endothelin-1 alter functional expression of Na+/Ca2+ exchange, K+,

144
and Ca2+ currents in cultured neonatal rat myocytes. Am J Physiol Heart Circ Physiol 2011;300:H617-626.
225. Lin C, Nagai M, Ishigaki D, Hayasaka K, Endoh M, Ishii K. Cross-talk between beta(1)-adrenoceptors and ET(A) receptors in modulation of the slow component of delayed rectifier K(+) currents. Naunyn Schmiedebergs Arch Pharmacol 2005;371:133-140.
226. Matsumoto Y, Aihara H, Yamauchi-Kohno R, Reien Y, Ogura T, Yabana H, et al. Long-term endothelin a receptor blockade inhibits electrical remodeling in cardiomyopathic hamsters. Circulation 2002;106:613-619.
227. Hagiwara K, Nunoki K, Ishii K, Abe T, Yanagisawa T. Differential inhibition of transient outward currents of Kv1.4 and Kv4.3 by endothelin. Biochem Biophys Res Commun 2003;310:634-640.
228. Polontchouk L, Ebelt B, Jackels M, Dhein S. Chronic effects of endothelin 1 and angiotensin II on gap junctions and intercellular communication in cardiac cells. Faseb J 2002;16:87-89.
229. Spinella F, Rosano L, Di Castro V, Nicotra MR, Natali PG, Bagnato A. Endothelin-1 decreases gap junctional intercellular communication by inducing phosphorylation of connexin 43 in human ovarian carcinoma cells. J Biol Chem 2003;278:41294-41301.
230. Reisner Y, Meiry G, Zeevi-Levin N, Barac DY, Reiter I, Abassi Z, et al. Impulse conduction and gap junctional remodelling by endothelin-1 in cultured neonatal rat ventricular myocytes. J Cell Mol Med 2009;13:562-573.
231. Rozyczka J, Figiel M, Engele J. Chronic endothelin exposure inhibits connexin43 expression in cultured cortical astroglia. J Neurosci Res 2005;79:303-309.
232. Niger C, Geneau G, Fiorini C, Defamie N, Pointis G, Mesnil M, et al. Endothelin-1 inhibits human osteoblastic cell differentiation: influence of connexin-43 expression level. J Cell Biochem 2008;103:110-122.
233. Morawietz H, Szibor M, Goettsch W, Bartling B, Barton M, Shaw S, et al. Deloading of the left ventricle by ventricular assist device normalizes increased expression of endothelin ET(A) receptors but not endothelin-converting enzyme-1 in patients with end-stage heart failure. Circulation 2000;102:III188-193.
234. Tikkanen I, Tikkanen T, Cao Z, Allen TJ, Davis BJ, Lassila M, et al. Combined inhibition of neutral endopeptidase with angiotensin converting enzyme or endothelin converting enzyme in experimental diabetes. J Hypertens 2002;20:707-714.
235. D'Orleans-Juste P, Houde M, Rae GA, Bkaily G, Carrier E, Simard E. Endothelin-1 (1-31): from chymase-dependent synthesis to cardiovascular pathologies. Vascul Pharmacol 2008;49:51-62.

145
236. Trapani AJ, Beil ME, Cote DT, de Lombaert S, Erion MD, Gerlock TE, et al. Pharmacologic profile of CGS 24128, a potent, long-acting inhibitor of neutral endopeptidase 24.11. J Cardiovasc Pharmacol 1994;23:358-364.
237. Henry PJ, Carr MJ, Goldie RG, Jeng AY. The role of endothelin in mediating virus-induced changes in endothelinB receptor density in mouse airways. Eur Respir J 1999;14:92-97.
238. Raoch V, Martinez-Miguel P, Arribas-Gomez I, Rodriguez-Puyol M, Rodriguez-Puyol D, Lopez-Ongil S. The peptidase inhibitor CGS-26303 increases endothelin converting enzyme-1 expression in endothelial cells through accumulation of big endothelin-1. Br J Pharmacol 2007;152:313-322.
239. Schiffrin EL. Beyond blood pressure: the endothelium and atherosclerosis progression. Am J Hypertens 2002;15:115S-122S.
240. Yang LL, Arab S, Liu P, Stewart DJ, Husain M. The role of endothelin-1 in myocarditis and inflammatory cardiomyopathy: old lessons and new insights. Can J Physiol Pharmacol 2005;83:47-62.
241. Haws CW, Lux RL. Correlation between in vivo transmembrane action potential durations and activation-recovery intervals from electrograms. Effects of interventions that alter repolarization time. Circulation 1990;81:281-288.
242. Liu L, Tockman B, Girouard S, Pastore J, Walcott G, KenKnight B, et al. Left ventricular resynchronization therapy in a canine model of left bundle branch block. Am J Physiol Heart Circ Physiol 2002;282:H2238-2244.
243. Kasahara H, Wakimoto H, Liu M, Maguire CT, Converso KL, Shioi T, et al. Progressive atrioventricular conduction defects and heart failure in mice expressing a mutant Csx/Nkx2.5 homeoprotein. J Clin Invest 2001;108:189-201.
244. Panama BK, Latour-Villamil D, Farman GP, Zhao D, Bolz SS, Kirshenbaum LA, et al. Nuclear Factor kappaB Downregulates the Transient Outward Potassium Current Ito,f Through Control of KChIP2 Expression. Circ Res 2011.
245. Kasi VS, Xiao HD, Shang LL, Iravanian S, Langberg J, Witham EA, et al. Cardiac-restricted angiotensin-converting enzyme overexpression causes conduction defects and connexin dysregulation. Am J Physiol Heart Circ Physiol 2007;293:H182-192.
246. Zaheer A, Sahu SK, Wu Y, Zaheer A, Haas J, Lee K, et al. Diminished cytokine and chemokine expression in the central nervous system of GMF-deficient mice with experimental autoimmune encephalomyelitis. Brain Res 2007;1144:239-247.
247. Gros R, Afroze T, You XM, Kabir G, Van Wert R, Kalair W, et al. Plasma membrane calcium ATPase overexpression in arterial smooth muscle increases vasomotor responsiveness and blood pressure. Circ Res 2003;93:614-621.

146
248. Kim KH, Oudit GY, Backx PH. Erythropoietin protects against doxorubicin-induced cardiomyopathy via a phosphatidylinositol 3-kinase-dependent pathway. J Pharmacol Exp Ther 2008;324:160-169.
249. Liang W, Oudit GY, Patel MM, Shah AM, Woodgett JR, Tsushima RG, et al. Role of Phosphoinositide 3-Kinase alpha, Protein Kinase C, and L-Type Ca2+ Channels in Mediating the Complex Actions of Angiotensin II on Mouse Cardiac Contractility. Hypertension 2010;56:422-429.
250. Kucera JP, Rohr S, Rudy Y. Localization of sodium channels in intercalated disks modulates cardiac conduction. Circ Res 2002;91:1176-1182.
251. Ai X, Zhao W, Pogwizd SM. Connexin43 knockdown or overexpression modulates cell coupling in control and failing rabbit left ventricular myocytes. Cardiovasc Res 2010;85:751-762.
252. Fernandez-Cobo M, Gingalewski C, Drujan D, De Maio A. Downregulation of connexin 43 gene expression in rat heart during inflammation. The role of tumour necrosis factor. Cytokine 1999;11:216-224.
253. Stanbouly S, Kirshenbaum LA, Jones DL, Karmazyn M. Sodium hydrogen exchange 1 (NHE-1) regulates connexin 43 expression in cardiomyocytes via reverse mode sodium calcium exchange and c-Jun NH2-terminal kinase-dependent pathways. J Pharmacol Exp Ther 2008;327:105-113.
254. Toyofuku T, Yabuki M, Otsu K, Kuzuya T, Tada M, Hori M. Functional role of c-Src in gap junctions of the cardiomyopathic heart. Circ Res 1999;85:672-681.
255. Ruetten H, Thiemermann C. Endothelin-1 stimulates the biosynthesis of tumour necrosis factor in macrophages: ET-receptors, signal transduction and inhibition by dexamethasone. J Physiol Pharmacol 1997;48:675-688.
256. Kovacic B, Ilic D, Damsky CH, Gardner DG. c-Src activation plays a role in endothelin-dependent hypertrophy of the cardiac myocyte. J Biol Chem 1998;273:35185-35193.
257. Kieken F, Mutsaers N, Dolmatova E, Virgil K, Wit AL, Kellezi A, et al. Structural and molecular mechanisms of gap junction remodeling in epicardial border zone myocytes following myocardial infarction. Circ Res 2009;104:1103-1112.
258. Malhotra JD, Thyagarajan V, Chen C, Isom LL. Tyrosine-phosphorylated and nonphosphorylated sodium channel beta1 subunits are differentially localized in cardiac myocytes. J Biol Chem 2004;279:40748-40754.
259. Gavillet B, Rougier JS, Domenighetti AA, Behar R, Boixel C, Ruchat P, et al. Cardiac sodium channel Nav1.5 is regulated by a multiprotein complex composed of syntrophins and dystrophin. Circ Res 2006;99:407-414.

147
260. Royer A, van Veen TA, Le Bouter S, Marionneau C, Griol-Charhbili V, Leoni AL, et al. Mouse model of SCN5A-linked hereditary Lenegre's disease: age-related conduction slowing and myocardial fibrosis. Circulation 2005;111:1738-1746.
261. Lin YZ, Yao SY, Veach RA, Torgerson TR, Hawiger J. Inhibition of nuclear translocation of transcription factor NF-kappa B by a synthetic peptide containing a cell membrane-permeable motif and nuclear localization sequence. J Biol Chem 1995;270:14255-14258.

Appendix 1. Myocardial ET- levels after short vs. long term CGS 26303 treatment. Expression of ET-1 peptide after 4 (A) and 12 (B) weeks post ECE inhibitor treatment.
ET
-1 p
epti
de
(fm
ol/n
g)
0
5
10
15
20
ET
-1 p
epti
de
(fm
ol/n
g)
0
2
4
6
8
ON DOX (ET-1 off)
OFF DOX (ET-1 on)
CGS 26303 (ET-1 on + treatment)
A B
148
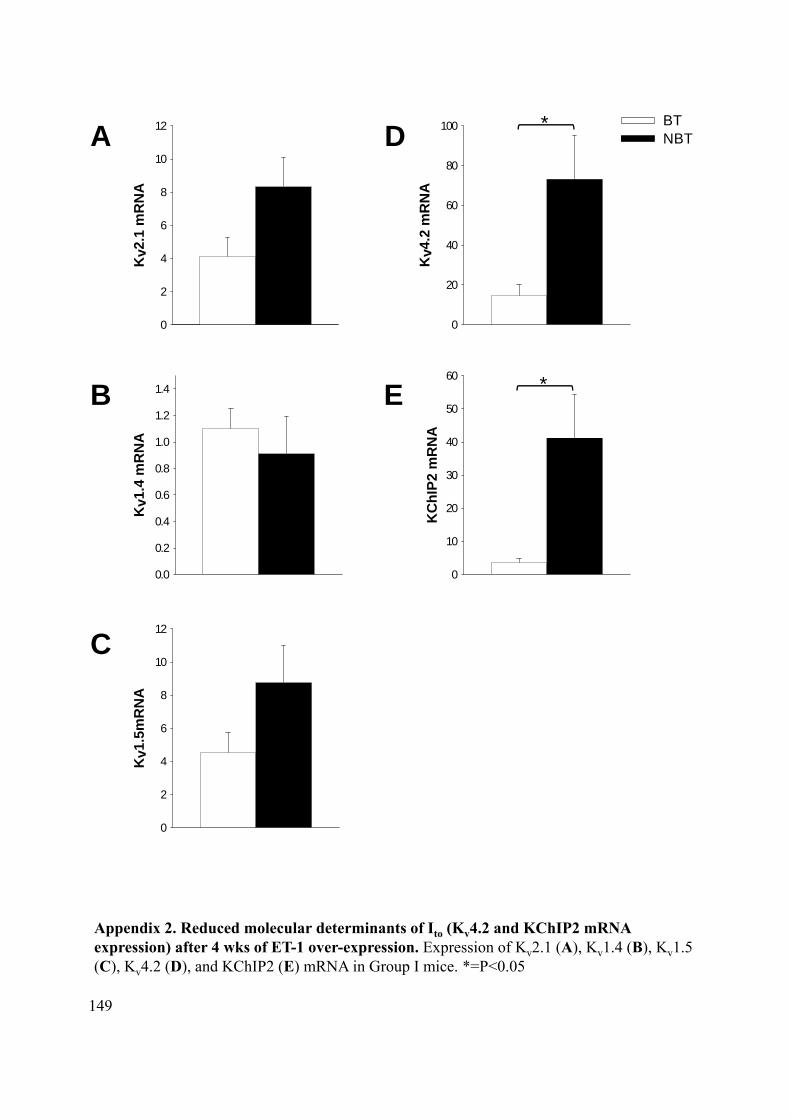
Appendix 2. Reduced molecular determinants of Ito (Kv4.2 and KChIP2 mRNA expression) after 4 wks of ET-1 over-expression. Expression of Kv2.1 (A), Kv1.4 (B), Kv1.5 (C), Kv4.2 (D), and KChIP2 (E) mRNA in Group I mice. *=P<0.05
Kv2
.1 m
RN
A
0
2
4
6
8
10
12
Kv4
.2 m
RN
A
0
20
40
60
80
100K
v1.4
mR
NA
0.0
0.2
0.4
0.6
0.8
1.0
1.2
1.4
Kv1
.5m
RN
A
0
2
4
6
8
10
12
KC
hIP
2 m
RN
A
0
10
20
30
40
50
60
C
A
B
BTNBTD
E *
*
149

GTTCACCTTGTCTCCCCCCCCATTTTTATTTATTGTAATATTATTATTATTATCATTATTATTTAGGAA
ATGTGACCTAAAAGGGACATCTTCTCACTGCCCGTGGTCATCTCCTGAAGGAATGACCCATCCAAC
AGTTTTTTTTTTTAATCTGTGAGGAGTCACAGCCCCGCAGTAGCTGACGTACATCTAGAGCTATTTC
TTACTTTTTTTCCCCTTCTCGTCAGCACATTGAAACTACAACTTTATCTTGACCAGGTTGCTTCTTCTGC
CTGCTCAGCTCCACGCTCGCCAGCCTCCACTCCACCTCCTCCCCGCCTTTTCTTCCTCCCTCCCCTTT
CTCCTAGCCCCTCCTTCCAGTTGAGTCAGTGGCTTGAAACTTTTAAAAGCTCTGTGCTCCAAGTTAA
AAAACGCTTTTACGAGGTATCAGCACTTTTCTTTCATTGGGGGAAAGGCGTGAGGGAAGTACCCAAC
AGCAGCAGACTTTGAAACTTTAAACAGACAGGTCTGAGAGCCCGAACTCTCCTTTTCCTTTGACTTCA
GCCTCCAAGGAGTTCCACCACTTTGGCGTGCCGGCTTCACTTTCATTAAGTGAAAGAGTAAGTTTTG
AAAAAAAGAAAAACCCACTTTACCTAGTAGCGTCTCTTTCTGTAAGGGAAACTCTTCTATGCTTTGT
AAATGCGTTGCGTCTTTGACTTAGGATACTGTAAGTAGAACGACGATCTTCAAAGTGCCTTTGTCAT
TTCCACGTGCTGAGTTTGATTTGGGTTTGTTTAAAAAGTTCCGCTGAGAGTGCAACAGGTAAAAGGCT
CAAACTTTTTCCAACTAGAGTGAAGGAAGGACCAAGTTACTAAACTTAGTTCTTTGTTATGGAGCC
TAACTTCACTTTTCTGTAGTCGCCAATGGAGAAGGTGTTGCGGGGGTGGGGGTGATGGGGGGGCACCT
CAGTCAACTTTGCTTAGCCTGCTTCCTATAGTGCTGGACACTACACGCTTCTTTT
Appendix 3. Cx43 core promoter sequence. Cx43 promoter (-411 to +589 from the transcriptional start site) showing the NFκB binding sites in bold and the accompanying forward and reverse primers.
150

CCGCGCCGCCCGCCGAGCCCGGGAGCCCGAGCGGAGCGGCGGAGCCGAGACGGCGGCGGCGCCCG
TGGGATGTGGGGATCGCGCCCCCGGGGCCGCTGAGCCTGCGCCCGCTGCCCCAAGCCCTACGCCGA
ACCGAGCCCGCGCCGCGCTGCAGCCGCCCACCCCTAGGCGCGGGCCGGGGACTAGCAGGTGAGCG
ATCATCCCCCGTGTCCCGCAGTCCCCCGCTCCCCTCTGTCCGCCGCCGCCGCCCAACTTTCCTCCCCG
ATGGCCCGGAGCCCCCCGGCCGGGTCCCCAGCCCCCGCAGCCACTATCCCCGAGGGGGGAGGGGCG
GGAGAGGCGCAGGTGGCCCGGGCCGGGGTTCCCCTGGTCCTCTCTGATCGCCGCGGGTCTCCTCCC
GTCCCCTCTCCTTCCGGGTTCCCCGTGCGCTGTCGAGGGCCGGGGGATGGGGAGATTCGGCCCCGG
CTCCGGGACCATCGCTGGCGCAGGCTCGCCGAGGGGCTGAGATTGGGACCCGGATTCCCGAGGCC
GGAGACCCCAAGGTCAGAGCAGCGACAAGGCGCGCGGAGAATGTAGCTCTTGAGAGCTGGGGGTG
AAAGGGCTGAGCGGGCACTGACCAGGGCTAGGGAGCAGGCCGAGCGCTTCTTTTCCCTGGGCCCC
GGGGGTGGAGGAACCAGGCACCTGGGCTATGCCCTGGGTTAGACGAGATCTCTGCACCCTAGGAA
GGAATCTGTGCGCTCTATTGGGGGTCAGATCTGCGAGTGTGCCTTGTCTTTGGGAGCTCCCACCACG
CCAAGATCTTGGTGCCCTGTTGTCGCCTTGAAGCGGTCGCATCTCCTCCTGTGTCTATGCGCTCTGAG
AGGGGAGAGGTCTCTGGACATGGAAATTCGGGAGAATTCTCTGCCCTCCTTGCTTTGACCTGTACA
GACTCGCGCACCCGGACAGGAGAGAGAGGAGGTCCCGGACCTCTGCGGTATTTGATCGTCTTTGGA
GGTGCCCTGAG
A
B
Appendix 4. Nav1.5 core promoter sequences. Nav1.5 promoters (-91 to +909, and -343 to +657 respectively from the transcriptional start site) showing the NFκB binding sites in bold and the accompanying forward and reverse primers.
GTCACAGAAGCTTATAGGGGTCACTAATGACATGCCCCATGGAGTGAGCTCAGGGCAAAGCCCTTG
GCTGACAGGAAGAGAGTGTGTCCCTGGCAGACATCTTCAGCAGGTGTCCAGTCTGCCCTCTTGAGC
CGGTGCCCAGCCATTCCCGGTTTTGATGTGGTAATTAGCGGTGCAGCCTCCTGGCTTGGTGTCATAG
TCAGGAGCCCTCTGGCCACATCCTGTTTTGGTGAGCCTGACAGAAATGCATTCTTCTGCCTTGCTGT
CATAGACAGAGTGTGTTCGCACCAGGTTCTGAATCTTTTGAGGCCACCAGGGGCCACATTCTCCCT
GTTTGACGTCACACACATATGCCTGTTGGAAGTCCTCTTCACCAGTGGTCACCGCCTTCCGGTGCTG
GGGGGGAAAGAAAAAACAACCCTTGGGGTTTGCGCTTGTATGTCCCCTTTTATGGGAGAAGAGGG
GACGGTGTGATGAGCCAGAGACCCACCTGTCACATTTCCCGTCTTTAGAATCAGTCTAGCTAGGGACG
GTGCTGCACTCAGGGGATCCCTATGAGATCCTCAGATGGAGGTCTACACAGTTGGGCACACCAGAT
GCCAACATCTGGATGCTAGTTCTTGTGTTCCTGTCCGGCCGGCCGTTGCTGAAAACCCTGGATCCCT
TGGGGGGCAAATGCTGCCTCCAGTTGCTGCCTCTATGCCTCAGGTTTGATTTGCACCTCTTGTGTGA
GGGCATGGGGATTGTGGGGGCACTGGACACCACTCAGGCTGGGAATGTTCCCTGGAGAGGGGGGG
TGGGACCCTGTCCCGTGCAGGGCCAAATCCTGACGTATGCATGCTTCACCTTTTAATTGGAGAAAA
GCCCTTCTGTTTGAGTCTGGGATAAAATGAACGGCATCTCTTCCCATCCCTGCCCTGTGGAGGCCAG
GGAGCCGGTTGTGGTGGAAATGTCTTAAAGGGGGTCAGTTGAAGTGTTTTTACTTTTGTGTGTGGT
GGAC
151

GGACCACTTAGCAACACCCCTCCGAATCCCACTCCCTTCAGGTAGATCCTGTCTCCCGCCCCACCGA
GCCTACCCTCGGGCCCCGGGGCCTGCAGCCCAAAGCCAGGCCCCGGCGCGGCCCGACCCTCCCACC
CTCCCAACAGGGCCCCTAACTCACGGTCCGGCTCGGCCATGGGAACGGGGTGGGGGCCCTCGGGA
CTGTCACCGACTGCTCTGTCCCGTGACGGCAGCGGAAACAAGAAGAGGCCGAGATAGGATAGAGA
ACTACAACTCCCGGCAGGCAATGCGATCCCCATAACCGCTTTCGTACAATGTGCCTCTCATGTTATC
AAACTACAAATCCCAAAAAGCCTGGCGCCGGCGGCCATATTGATAAAGGGCGTCTGGAAAATGTG
GGGAAACTCAGCGAGCGAATGTCCACCTCATTTTTCTGATTGGCTGTGGTGCAGTACAGGGGCGGGG
TGTAGCTATTTAAAACATTTTGATTGGGTATATGAGGGGGCGTGTTGGGGAATTCCCGATAGGGCG
GAAGCACTGGAGCTCATCGCAGAGCCCAGCGACAGGCAGGCGACCACAGGGGGCCACCCGAGGTG
GCTGGGGCCATGGCCGGGGTCGCGTGCTTGGGGAAAACTGCGGATGCCGATGAATGGTGCGACAG
CGGCCTGGGCTCTCTAGGTCCCGACGCAGCGGCTCCCGGAGGACCAGGTCTGGGCGCAGAGCTTGG
CCCAGAGCTGTCGTGGGCGCCCTTAGTCTTTGGCTACGTCACTGAGGATGGGGACACGTGAGTAAA
CCTTAAATTACTAGACTGAGCCCTGGGATCTTACCTGAGTCCCCTTTAGCCTACGATTCCTGTGACT
TCGTATCGTCTCGTTCCTAACTCTTGATTTCCGATGCTGGACTTCCTGGCCCTTAACCCCCAAACACT
AAATTGTGAGACTGGTACGCTTTCACACAGTCAAGTGTCTCACCTTTTTTGAACTCTATAAATTTGA
GCTCCTGA
NF
kBIB
Pro
mo
ter
NFB
bin
din
g s
ite
0.00
0.02
0.04
0.06
0.08
0.10
A
B
Appendix 5. NFκB p50 preferentially binds to sites within the NFκBIB promoter in the LV of ET-1 over-expressing mice. A. NFκBIB promoter sequence (-500 to +500 from the transcriptional start site) showing the NFκB binding sites in bold with the accompanying forward and reverse primers. B. The qRT-PCR results for the NFκB binding site within the NFκBIB promoter from a ChIP using an NFκB p50 Ab on LV tissue from BT vs. NBT mice from Group I.
BTNBT
152

CCGTTCCCGCCCGTGCCGGCCCCATCAGGGTTCTGCCATGGCTCCCCACTCACCGGCTCCGGACACC
TGATTCGGCTCCGGGACCTCGGCCGCCCGAGCCTCCTGATTTGCCCGCCCACCGGCCTCGCTTTCCA
GTGCCCGCAGCCTCCCTCTGAAGTGCATGGACCCGGGGCGGTGACCGGGAGTGGGGTGGGAGGCC
AGGCTGGGTCGCAGGCAAGAGCGGCCGGGCCTCCCGGCGCGGGAGGGAAAGGGCGCCCCCTCTCC
TCTCCGGAACGTGTCAATGCTTTGCACTTGGGGCCGGCATGCGGCTAGGGGGTCCTTCCCCAAGGC
CCCGGGACCCGGGCGCCCCCTGCCTCAGGCCCTTACGGCGGGTCAGATCGGTCCTCTGCGCTTTCCA
GCCCTCGCCCAGGCAGCGGCGGGCGCGGGCGGCGAGGTGGGGGCCAGGCCAGGGGGAGGGGTCTC
GGGGCCCGCTGGCCCGTCATTGGTTAATATTTTATTCTGTTGACATGTTTTCTTACTGCTGAGGCTTC
CGACACCTTCTCCCTGGCCTCCCCTCCCGTCCAGAGCTTGGCCTGAGCTGTCAAAACCCCGCCCCCG
GAGACCCACAATTGGTCCAAAAAGCGTAAAATCAGCAATCAAGGGGGGCCTGGCTCGTTAGCGCA
GGGGATCCGAACAGGGCAGGACATGTGAGATAGTCACAGTTTTCCAGAGATCAGGACAAGATCTA
ACCAGTCGCGCGTGGTCCCCGGCGCCGGAGCCGGCCAGCCCAGCCCAGCCCAGCCCAGCCCAGCCC
CGCGCAGCGCCCCCTCCGCCCCCGCGTCCAGAGCCCTGCGCCCCTTGAGGTGCGCGGGACGGGGAG
CCGGGAGAAGCCGCTGCCGCGCCCGCCGCCCGGGCCGTCCGTCCTCCGCGCGCGCCGCCGCCCGGG
CCGGGGGTCCGAGGCGCGCGCCCCCGGCCCCGGCCCCGGCTCCCAGGAGCCTGGGCCGGATGTCCC
GATGAGAGA
Tb
x2 P
rom
ote
rN
FB
bin
din
g s
ite
0.00
0.02
0.04
0.06
0.08
0.10
0.12
0.14
0.16
A
B
Appendix 6. NFκB p50 preferentially binds to a site within the Tbx2 promoter in the LV of ET-1 over-expressing mice. A. Tbx2 promoter sequence (-500 to +500 from the transcriptional start site) showing the NFκB binding sites in bold and the accompanying forward and reverse primers. B. The qRT-PCR results for the NFκB binding site within the Tbx2 promoter from a ChIP using an NFκB p50 Ab on LV tissue from BT vs. NBT mice from Group I.
BTNBT
153

CTGGCCCGGAGGTGAGGGCAAAGGCTCTCAGGAGACAGGCCCGCAGTTCCAGGCGCTCACTTGTTT
CAGCTGCTTCGTTCTCTCTACACCCACACCCCCATCCGGGTGGAACACTGGTGTCGTCATCCGAATG
GAAGAGGAGCTGGGCTCAAAAGGGTCAGTAAATACATTTTTTAAAAAAAAAAGGAGCGAGCCGAG
CGTAGACTGGGCCAGGCGCGCAGGCCTTGAATGCATTTGCGTGGTTTATTTTTCCGCGTGGGGGCT
CGGTCTCAGTATGTCGGCGCGCGTTCGGGCCCAGAATCCCAAGGGGCATTTGGGTCTCTGCACGTG
GCTGCGGGTGGGGAGCAGGCAGCGGGCAGAAAGATTGGTGGGATCCCCTGCGCTAGTGGCTCGGG
AATCCGAAGCACCCGGGCGCAGGAGCTAGAGGATCTGACTCAGAGGTGGCTGGGCGGTTCCATG
TGGGGCTCCCTTCAGAGCTACGCGAGCCTGGAGCTTTTGTTAAGGCTTATTGGCCGAAAGAGAGGTT
CGGCGGCCAGCTCGGCTAGCCCGGGGCCCGGGAAAGGGAAGAAGCTGCAGATCCGCACAAGAGA
AGCGGCCGCGGGCTTGAACTGTAGCGCTCGGAGCGCGCGAGAGGCGAGCGCCCCTGCCCGGCGCC
TCGGAAGCCGGGCAAGCTGCCTGGCTCCCGCCCTCGCTCCCTCCCCCTTCCTCCCTGGCCCAGCCTC
CTCCCCGGATCCCCCGGCTTGGATGGCTGAGGCCTTTCAGACGTAGGCTGAGCTGAGGAGCCGCGG
CGAGCTCTCAGGCTGCTGCGAACTCTCTTCTGGATCAGCCACCTAGAGGCGACTTTGGTGAGCGCG
CGGCGCCCTGGTGGCTCCCCGCCCTCCCCTCTGATCATGTTGACATAAACGCAGGACAGGCCGTAG
TACCGCGCGGCGCAGCGACGTTCCAGTTTCCGACACCTTCTTTTTATAACTCGGCTCTATTCCCCCA
GCACTCGACCTGT
Tb
x3 P
rom
ote
rN
FB
bin
din
g s
ite
0.00
0.02
0.04
0.06
0.08
0.10
A
B
Appendix 7. NFκB p50 does not preferentially bind to a site within the Tbx3 promoter in the LV of ET-1 over-expressing mice. A. Tbx3 promoter sequence (-764 to +236 from the transcriptional start site) showing the NFκB binding sites in bold and the accompanying forward and reverse primers. B. The qRT-PCR results for the NFκB binding site within the Tbx3 promoter from a ChIP using an NFκB p50 Ab on LV tissue from BT vs. NBT mice from Group I.
BTNBT
154

TCCTTTGTGCCAAAAGAGAGGAGTCTGGGGACCGGGTTGTGATTTTGAGATGCGATGCTGCGGATC
CCAGGAGCTGGGGTTGGCTTTGTTTTTCTAGTACAATCTCCAGGCAGGATCAGGCAGAAATTCCTTT
CTGGGAACTGTTGGGAAATACGGAACGAATAAAGACACTGAGAAGTGAAGAGCAGGCTTCCGAAG
GTGGCCGAATACCAAATATCTCAGAGTAGGGAGAGGGGCTCCTGGCCAGCTCACAGAAGGAGCAGG
GAAGTCCCAGGCCCAGATCCTGTGCTGAGTAAAGCCAGAGAGGCCCAATGGTTTCTGCGAGTTAGA
GGACGTTCCTAGGTTTTCCCTGGGAGCAGAGAGAAGGTCCAGGAACATTGGAGAAGGACTGGAGG
AGGATTAGAACTTATTGCTACCAACCGGAATCCCGCAGGGCTGGTGTAGAGGCCTCTGTCCTCCTC
CGGGTACTGGGAAGTCCAGAAGCCATGCCAACATCAGCTGACCACCCGAGGGACAAACCGGAGTA
TTCCTAATGTAATTCACTCCGAGCATCCTCATGACTTTATTTTCTTTGCAGCCAGAAAAGCTAAAGC
TAAATCCCAGACTGTTAAGAGAACGCTAACTATCCGGGGAAGAGGTCTGGGATAGGGCGCCCGAGA
TTTCTCTTGTTTTGTTTTGTCCAAGCCACTTAGGCATTTCCCAGATGCGCAATTGCGCCAACATCGA
ACCGAGCGAGCCCGTGCACTCGGCGATTAGCTTAAGCGGAGCTGGGTGTCCGGCATAGGACCAGAG
TGATACTCCCTGCCACCCCTACAAGGGCCCTGAGGTCCCCGGCCCGCCGCTCTAACCCGCCACCTCT
CTGCCTCTCTTCCCCTTCAGAGCTGTGCGCGCTGCAGAAGGCAGTGGAGCTGGACAAAGCCGAGAC
GGATGGCGCCGAGAGACCACGCGCACGGCGGCGATCAAGCAACAGCGGTACCTGTCGGCGCCAGA
GCGCGACCAG
Nk
x2.
5 P
rom
ote
r1
st N
FB
bin
din
g s
ite
0.00
0.01
0.02
0.03
0.04
Nk
x2.
5 P
rom
ote
r2
nd
NFB
bin
din
g s
ite
0.00
0.02
0.04
0.06
0.08
0.10
A
B CBTNBT
Appendix 8. NFκB p50 does not preferentially bind to sites within the Nkx2.5 promoterin the LV of ET-1 over-expressing mice. A. Nkx2.5 promoter (-879 to +121 from the transcriptional start site) showing the NFκB binding sites in bold and the accompanying forward and reverse primers. qRT-PCR results for 1st (B), and 2nd (C) NFκB binding sites within the Nkx2.5 promoter from a ChIP using an NFκB p50 Ab on LV tissue from BT vs. NBT mice from Group I.
155