molecular cloning of hyoscyamine gb-hydroxylase, a 2
TRANSCRIPT

THE JOURNAL OF BIOLOGICAL CHEMISTRY (0 1991 by The American Society for Biochemistry and Molecular Biology, Inc.
Val. 266, No. 15, Issue of May 25, pp. 9460-9464,1991 Printed in U. S A .
Molecular Cloning of Hyoscyamine GB-Hydroxylase, a 2-Oxoglutarate-dependent Dioxygenase, from Cultured Roots of Hyoscyamus niger*
(Received for publication, September 13, 1990)
Jun Matsuda, Souichi OkabeS, Takashi Hashimotot, and Yasuyuki Yamada From the Department of Agricultural Chemistry, Faculty of Agriculture, Kyoto Uniuersity, Kyoto 606, Japan
Roots of several solanaceous plants produce anticho- linergic alkaloids, hyoscyamine and scopolamine. Hyoscyamine GB-hydroxylase, a 2-oxoglutarate-de- pendent dioxygenase (EC 1.14.11.11), catalyzes hy- droxylation of hyoscyamine in the biosynthetic path- way leading to scopolamine. We report here on the isolation of cDNA clones encoding the hydroxylase from a cDNA library made from mRNA of the cultured roots of Hyoscyamus niger. The library was screened with three synthetic oligonucleotides that encode amino acid sequences of internal peptide fragments of the purified hydroxylase. Nucleotide sequence analysis of the cloned cDNA revealed an open reading frame that encodes 344 amino acids (Mp = 38,999). All 12 internal peptide fragments determined in the purified enzyme were found in the amino acid sequence deduced from the cDNA. With computer-aided comparison to other proteins we found that the hydroxylase is ho- mologous to two synthases involved in the biosynthesis of B-lactam antibiotics in some microorganisms and the gene products of tomato pTOM13 cDNA and maize A 2 locus which had been proposed to catalyze oxidative reactions in the biosynthesis of ethylene and antho- cyan, respectively. RNA blotting hybridization showed that mRNA of the hydroxylase is abundant in cultured roots and present in plant roots, but absent in leaves, stems, and cultured cells of H. niger.
Hyoscyamine and scopolamine, two major tropane alkaloids found in the family Solanaceae, are important drugs that act mainly on the parasympathetic nervous system. These alka- loids are esters of tropic acid and tropine derivatives: tropine is derived from ornithine and/or arginine, whereas tropic acid is synthesized from phenylalanine (1). Scopolamine, a 6,7- epoxide of hyoscyamine, is formed from hyoscyamine by way of 6P-hydroxyhyoscyamine (Fig. 1). H6H’ that catalyzes the
* The costs of publication of this article were defrayed in part by the payment of page charges. This article must therefore be hereby marked “aduertisement” in accordance with 18 U.S.C. Section 1734 solely to indicate this fact.
The nucleotide sequencefs) reported in this paper has been submitted to the GenBankTM/EMBL Data Bank with accession nunbeds) M62719.
$ Present address: Research and Development Center, Nagase Sangyo Co., Shinmachi 1-1-17, Nishi-Ku, Osaka, 550-90, Japan.
5 To whom correspondence and reprint requests should he ad- dressed.
’ The abbreviations used are: H6H, hyoscyamine GB-hydroxylase; SDS-PAGE, SDS-polyacrlamide gel electrophoresis; TBS, Tris-buff- ered saline; R, a purine nucleoside, A or G; Y, a pyrimidine nucleoside, T or C; N, a nucleoside, A, G, T, or C; DAOC synthase, deacetoxy- cephalosporin C synthase; IPN synthase, isopenicillin N synthase; kb, kilobase(s); bp, base pair(s).
first reaction in the epoxide formation is a 2-oxoglutarate- dependent dioxygenase (EC 1.14.11.11) which requires alka- loid substrate, 2-oxoglutarate, Fez+, molecular oxygen, and ascorbate for catalysis (2, 3). H6H purified from the cultured roots of Hyoscyamus niger is a monomer of 38 kDa, as deter- mined by SDS-PAGE (4).
As originally shown in classical grafting experiments be- tween alkaloid-producing plants and nonproducing plants (5), tropane alkaloids are synthesized mainly in the root and translocated to the aerial part of the plants. Recently, cultured roots were found to be especially active in alkaloid biosyn- thesis (6) and have been widely used for biosynthetic studies of tropane alkaloids. H6H activity, for example, is found at high levels in cultured roots of the Duboisia and Hyoscyamus species, but at very low levels in their cultured shoots and calli (2). Further, immunohistochemical studies using an anti- H6H monoclonal antibody localized H6H protein specifically in the pericycle cells in the root (7). As an initial step to study underlying molecular mechanisms for this tissue- and cell- specific expression of alkaloid biosynthesis, we isolated H6H cDNA from H. niger. Root-specific expression of H6H gene is also demonstrated.
EXPERIMENTAL PROCEDURES
Plant Material-Cultured roots of H. niger were grown a t 25 “C in the dark on a rotary shaker (100 rpm) in Gamborg B5 medium containing 3% (w/v) sucrose and 1 PM indole-3-butyric acid (6). After transfer to the medium without auxin for 1 week, the cultured roots were used for the experiments. Plants were grown in a greenhouse and harvested at the flowering stage. Suspension-cultured cells were grown as described (8).
Protein Sequencing-H6H was purified to homogeneity by a series of chromatographic steps as described (4). Direct amino-terminal sequencing gave negligible yields presumably because of amino-ter- minal blockage. We therefore digested the protein with lysyl endo- peptidase (EC 3.4.21.50; Wako Pure Chemical Inc., Kyoto) and then fractionated internal peptide fragments by reversed-phase high per- formance liquid chromatography using a YMC-PAC A-312 ODS column (6.0 X 105 mm; Yamamura Kagaku, Tokyo), using a linear 5-60% gradient of acetonitrile in 0.1% trifluoroacetic acid. The amino-terminal sequence of 12 peptides was determined by automated Edman degradation with a 470.4 gas phase sequencer (Applied Bio- systems).
Construction and Screening of the cDNA Library-DNA manipu- lations, bacterial growth media, and buffers were as described by Sambrook et al. (9). Poly(A)+ RNA prepared from cultured roots of H. niger was used to synthesize cDNA using avian myeloblastosis virus reverse transcriptase (Life Science Co.) and an oligo(dT) primer having restriction sites of NotI and XhoI. Double-stranded cDNA was blunt-ended, cut by NotI, and then cloned into pBluescript I1 (KS-) (Stratagene) that had been restricted with NotI and EcoRV. Escherichia coli SCS-1 strain was used for transformation and screen- ing.
Three 14-mer oligonucleotide probes used for cDNA screening were designed from amino acid sequences of H6H internal peptide frag- ments. Probe 1, 5’-GGRTTRTCYTGRTT-3’, was constructed by a
9460

Hyoscyamine G@-Hydroxylase cDNA 9461
0 II w ‘ H6H
C02 O’+o
hyoscyamine 6p-hydroxyhyoscyamine scopolamine FIG. 1. Biosynthetic pathway from hyoscyamine to scopolamine. Hyoscyamine 6P-hydroxylase (H6H), a
2-oxoglutarate-dependent dioxygenase, catalyzes the first reaction in this epoxide formation.
complementary nucleotide sequence of Asn-Gln-Asp-Asn-Pro; probe 2, 5’-ATRTCNGCRAAYTC-3’, was constructed from Glu-Phe-Ala- Asp-Ile; and probe 3, 5’-CCRAARTCYTGRCA-3’, from Cys-Gln- Asp-Phe-Gly. Probe degeneracy was 16-fold for probe 1, 32-fold for probe 2, and 16-fold for probe 3. The cDNA library was screened by colony hybridization. Two sets of replicated nitrocellulose filters were prepared and hybridized first with oligonucleotide probe 1. Probes were end-labeled with [cY-~’P]ATP by polynucleotide kinase. cDNA inserts in positive clones obtained in both sets of replicated filters were then analyzed by Southern blotting analysis with probes 1, 2, and 3. From four positive clones that hybridized to all three oligo- nucleotide probes, a clone (designated pBHH1) with the longest cDNA insert was used to transform Escherichia coli MV1184 for nucleotide sequence analysis. The complete nucleotide sequence of the pBHHl cDNA insert was determined on both DNA strands by the dideoxy chain termination method (10). Subsequently, a 20-mer oligonucleotide probe (nucleotides 12 to 31 in Fig. 2B) comprising the 5”untranslated region of the pBHHl cDNA insert was used to screen H. niger cDNA library once more, and five additional clones (pBHH2 to pBHH6) were obtained. The cDNA inserts in the new clones were analyzed on their 5’- and 3’-ends for about 100 bp each.
Construction of Prokaryotic Expression Vector-An NcoI recogni- tion site was introduced at the initiation ATG of the pBHHl insert using an oligonucleotide-directed in vitro mutagenesis system, version 2 (Amersham). The mutated fragment was restricted with NcoI and Sac1 (see Fig. 2 . 4 ) and then introduced to pTV119N (Takara Shuzo Co., Kyoto) at a unique NcoI site. The constructed plasmid was designated pNHH3.
Immunoblot Analysis-After E. coli MV1184 containing pNHH3 or pTV119N was incubated for 2 h at 37 “C in 100 ml of LB medium, 1 ml of 100 mM isopropyl-(3-D-thiogalactoside was added, and incu- bation was continued for 4 h at 37 “C. After centrifugation, the pellet was resuspended in 400 pl of 100 mM phosphate buffer (pH 7.5) containing 5 mM dithiothreitol, lysed by sonication on ice for 15 s (four times), and centrifuged at 15,000 X g for 15 min. The superna- tant was desalted by using a NAP@-5 column (Pharmacia LKB Biotechnology Inc.).
The crude bacterial extracts (30 pg of protein/lane) were separated by SDS-PAGE on a 12.5% gel according to Laemmli (11) and then electrophorectically transferred to polyvinylidene difluoride mem- branes (Immobilon@, Millipore) in 25 mM Tris-HC1 (pH 8.3), 192 mM glycine, and 20% methanol. Nonspecific binding to the membrane was prevented by incubation in 1% bovine serum albumin in 20 mM Tris-HC1 (pH 7.5) and 0.5 M NaCl (TBS) for 1 h a t room temperature. The blocked membranes were incubated with a mouse anti-H6H monoclonal antibody mAb5 (1 pg/ml) (7), which was raised against the H6H purified from cultured roots of H. niger, for 1 h at room temperature with constant shaking. Unbound antibodies were re- moved with two washes of TBS containing 0.05% Tween 20, and the antibodies bound to the membranes were detected using peroxidase- conjugated goat anti-mouse IgG (40 pg/ml; Zymet) and 3,3’-diami- nobenzidine tetrahydrochloride.
Enzyme Assay-Crude bacterial extracts were assayed for H6H activity by measuring, by gas-liquid chromatography, the formation of 6P-hydroxyhyoscyamine from hyoscyamine (3).
RNA Blot Analysis-Total RNA from various tissues of H. niger was denatured and electrophoresed on a formaldehyde, 1.2% agarose gel (15 pg/lane), and then transferred to Immobilon. Filter was hybridized with a 1.47-kb XhoI fragment of pBHHl labeled with 32P by the random hexanucleotide-primed method.
RESULTS AND DISCUSSION
Isolation of cDNA Encoding H. niger H6H”Partial amino acid sequences of 12 internal peptide fragments from purified H. niger H6H were used to design three oligonucleotide probes. About 30,000 independent transformants from a H. niger cDNA library were first screened with probe 1, and about 100 positive clones were obtained. Plasmids from 16 clones that were randomly picked up from the positive clones were restricted with PvuII and analyzed by Southern hybrid- ization using the three probes. Probes 1 and 2 hybridized to 1.2-kb or shorter restriction fragments from all clones, but probe 3 hybridized to 0.5-kb or slightly shorter fragments present only in four clones. Subsequent sequence analysis (see Fig. 2B) revealed that probes 1 and 2 correspond to amino acid sequences close to the carboxyl-terminal of H6H, whereas probe 3 corresponds to a sequence close to the amino-terminal. Thus, it seems likely that a large proportion of the cDNA inserts in our H. niger cDNA library lack 5’-portions of corresponding mRNAs. From the four clones that gave posi- tive fragments with all three probes, a clone with the longest cDNA insert was chosen and designated pBHH1. pBHH1 contained a 1410-bp cDNA insert and, as expected, a 0.5-kb PvuII fragment at the 5‘ end (Fig. 2 A ) .
To demonstrate the function of the polypeptide encoded in pBHH1, we constructed an expression vector (pNHH3) such that an intact protein is translated from the first ATG of the cDNA under the control of the lac2 promoter. The E. coli harboring pNHH3 expressed a 38-kDa protein which was recognized by an H6H-specific antibody on Western blots (Fig. 3) and showed the H6H activity of 1.58 picokatal/mg of protein, whereas the bacteria harboring a control plasmid pTV119N did not show any protein recognized by the anti- body (Fig. 3), nor the detectable H6H activity. These results firmly establish that the cDNA in pBHHl encodes H6H.’
Sequence of H6H cDNA-Five additional cDNA clones were obtained using a 20-bp 5”untranslated region of the cDNA in pBHHl as a probe. These clones had slightly longer 5’-ends and somewhat shorter 3’-ends than the pBHHl cDNA insert. Although only part of these cDNA inserts in each clone was sequenced, no sequence difference was ob- served in the overlapping regions among the six clones, The combined cDNA sequence for H6H is shown in Fig. 2B. A large open reading frame, which begins at the translation
We also constructed a plant expression vector in which the cDNA was placed under the control of the CaMV 35 S promoter with the NOS terminator in pGA482 (Pharmacia) and introduced the gene into tobacco leaf disc cells using Agrobacteriurn-mediated gene trans- fer. The transformed tobacco calli expressed H6H enzyme activity and a 38-kDa protein that was recognized by the anti-H6H antibody on Western blots (T. Hashimoto, D. J. Yun, and Y. Yamada, unpub- lished results).
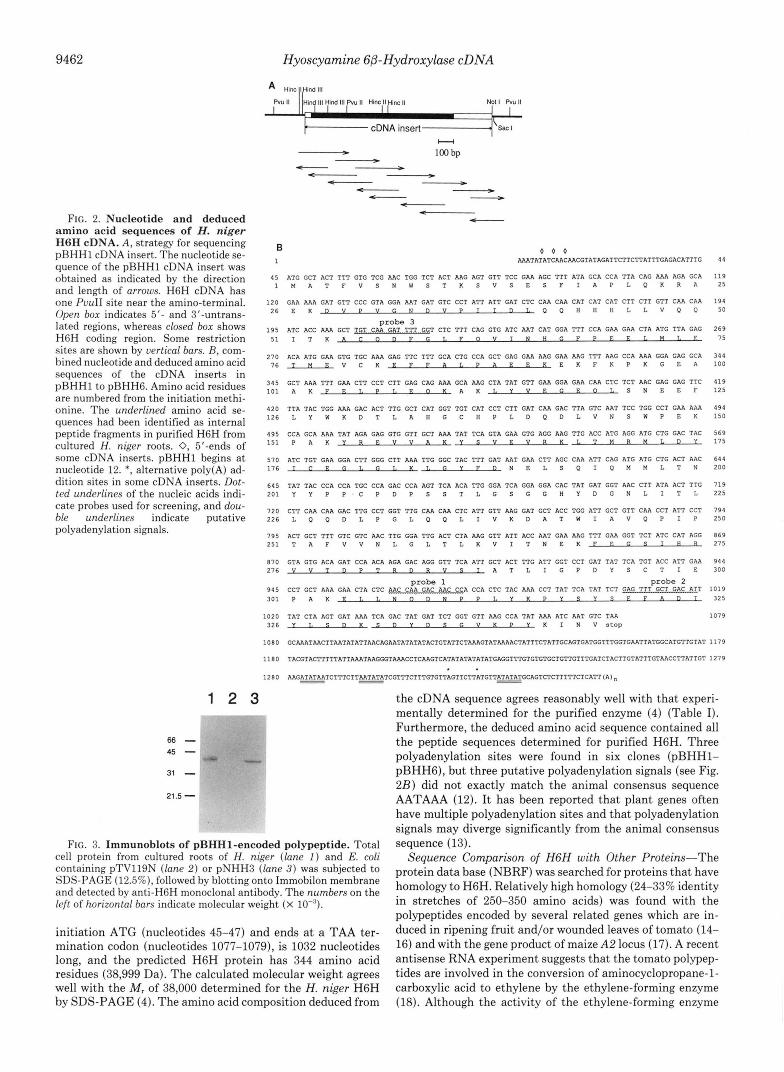
9462 Hyoscyamine GB-Hydroxylase cDNA
No! I pvu II
H - - 100 bp " - - ___) - -
c- - - - - t" FIG. 2. Nucleotide and deduced
amino acid sequences of H. niger H6H cDNA. A, strategy for sequencing pHHH1 cDNA insert. The nucleotide se- quence of the pBHHl cDNA insert was obtained as indicated by the direction and length of arrows. H6H cDNA has one PvuII site near the amino-terminal. Open box indicates 5'- and 3"untrans- lated regions, whereas closed box shows H6H coding region. Some restriction sites are shown by vertical bars. B, com- bined nucleotide and deduced amino acid sequences of the cDNA inserts in pBHHl to pBHH6. Amino acid residues are numbered from the initiation methi- onine. The underlined amino acid se- quences had been identified as internal peptide fragments in purified H6H from cultured H. niger roots. 0, 5'-ends of some cDNA inserts. pBHHl begins a t nucleotide 12. *, alternative poly(A) ad- dition sites in some cDNA .inserts. Dot- ted underlines of the nucleic acids indi- cate probes used for screening, and dou- ble underlines indicate putative polyadenylation signals.
1 2 3 - . .
66- 45 - 31 - 21.5 -
B 1 AAATATATCAACAACGTATAGATTCTTCTTATTTGAGACATTTG
0 0 0
45 ATG GCT ACT TTT GTG TCG AAC TGG TCT ACT AAG AGT GTT TCC GAA AGC TTT ATA GCA CCA TTA CAG AAA AGA GCA I M A T F V S N W S T K S V S E S F I A P L Q K R A
120 GAA AAA GAT GTT CCC GTA GGA AAT GAT GTC CCT ATT ATT GAT CTC CAA CAA CAT CAT CAT CTT CTT GTT CAA CAA ~ ~ E K D V P V G N O V P T I ~ ~ Q ~ H H H L L V O P
probe 3 1 9 5 ATC ACC AAA GCT m C I v I FAT T CTC TTT CAG GTG ATC AAT CAT GGA TTT CCA GAA GAA CTA ATG TTA GAG
5 1 1 T K A C O D F G ~ . F O V ~ N H ~ P P E E 1 ~ M 1 ~ ~
270 ACA ATG GAR GTG TGC AAA GAG TTC TTT GCA CTG CCA GCT GAG GAA AAG GAA AAG TTT AAG CCA AAA GGA GAG GCA 7 6 T M E V C K E F F A L P A E E K E K F K P K G E A
3 4 5 GCT AAA TTT GAA CTT CCT CTT GAG CAG AAA GCA AAG CTA TAT GTT GAA GGA GAA CAA CTC TCT AAC GAG GAG TTC ~ ~ ~ A K F R ~ . P L E ~ K A K ~ . Y ~ E G E ~ ~ . S N E E F
420 TTA TAC TGG AAA GAC ACT TTG GCT CAT GGT TGT CAT CCT CTT GAT CAA GAC TTA GTC AAT TCC TGG CCT GAA AAA 1 2 6 L Y W K D T L A H G C H P L D Q D L V N S W P E K
495 CCA GCA AAA TAT AGA GAG GTG GTT GCT AAA TAT TCA GTA GAA GTG AGG RAG TTG ACC ATG AGG ATG CTG GAC TAC 1 5 1 P A K Y R E V V A K Y S V E V R K l . T M R M L ~ ~
570 ATC TGT GAA GGA CTT GGG CTT ARA TTG GGC TAC TTT GAT AAT GAA CTT AGC CAA ATT CAG ATG ATG CTG ACT ARC 1 7 6 l C E G 1 , ~ l . K l . G V F D N E L S Q I Q M M L T N
645 TAT TAC CCA CCA TGC CCA GAC CCA AGT TCA ACA TTG GGA TCA GGA GGA CAC TAT GAT GGT AAC CTT ATA ACT TTG Z O I Y Y P P C P D P S S T L G S G G H Y D G N L I T L
7 2 0 CTT CAA CAA GAC TTG CCT GOT TTG CAA CAA CTC ATT GTT AAG GAT GCT ACC TGG ATT GCT GTT CAA CCT ATT CCT 2 2 6 L Q Q D L P G L Q Q L l V K D A T W l A V Q P l P
795 ACT GCT TTT GTC GTC AAC TTG GOA TTG ACT CTA AAG GTT ATT ACC AAT GAA AAG TTT GAA GGT TCT ATC CAT AGG 2 5 1 T A F V V N L G L T L K V I T N E K F E G S l H B
8 7 0 GTA GTG ACA GAT CCA ACA AGA GAC AGG GTT TCA ATT GCT ACT TTG ATT GGT CCT GAT TAT TCA TGT ACC ATT GAR 2 7 6 V V T n P T R D R V S l A T L I G P D Y S C T l E
9 4 5 CCT GCT AAA GAR CTA CTC AAC CAA GAC AAC CCA CCA CTC TAC AAA CCT TAT Tcn TAT TCT GAG TTT GCT GAC ATT ~ O ~ P A K P ~ . T . N O ~ N P P ~ . Y K P Y ~ Y S F . F A D ~
probe 1 probe 2
1 0 2 0 TAT CTA AGT GAT AAA TCA GAC TAT GAT TCT GGT GTT RAG CCA TAT AAA ATC ART GTC TAA ~ ~ ~ Y T . B ~ K ~ D Y D ~ ~ V K P Y K I N V ~ ~ ~ ~
1 0 8 0 GCAAATAACTTMTATATTAACAGAATATATATA~~GTATTCTAAAGTTTCATAAAACTATTTCTATTGCAGTGATGGTTTG~GAATTATGGCATGTTGTAT
1 1 8 0 TACGTACTTTTTATTAAATAAG~TAAACCTCAAGTCATATATATATATGAGGTTTGTGTGTGCTGTTGTTTGATCTACTTGTATTT~AACCTTATTGT
1 2 8 0 AAG~AATCTTTCTTAATATATCGTTTCTTffTTGTGTTAGTTCTTATGTT~GCAGT~CTTTTTCTCATT(A) . ~ - -
FIG. 3. Immunoblots of pBHH1-encoded polypeptide. Total cell protein from cultured roots of H. niger (lane 1 ) and E. coli containing pTV119N (lane 2 ) or pNHH3 (lane 3 ) was subjected to SDS-PAGE (12.5%), followed by blotting onto Immobilon membrane and detected by anti-H6H monoclonal antibody. The numbers on the left of horizontal bars indicate molecular weight (x IO-").
initiation ATG (nucleotides 45-47) and ends at a TAA ter- mination codon (nucleotides 1077-1079), is 1032 nucleotides long, and the predicted H6H protein has 344 amino acid residues (38,999 Da). The calculated molecular weight agrees well with the M , of 38,000 determined for the H. niger H6H by SDS-PAGE (4). The amino acid composition deduced from
44
1 1 9 25
194 50
2 6 9 75
344 1 0 0
419 1 2 5
494 1 5 0
569 1 7 5
644 200
719 225
794 250
869 275
3 0 0 944
1 0 1 9 325
1079
1 1 7 9
1 2 7 9
the cDNA sequence agrees reasonably well with that experi- mentally determined for the purified enzyme (4) (Table I). Furthermore, the deduced amino acid sequence contained all the peptide sequences determined for purified H6H. Three polyadenylation sites were found in six clones (pBHH1- pBHHG), but three putative polyadenylation signals (see Fig. 2B) did not exactly match the animal consensus sequence AATAAA (12). It has been reported that plant genes often have multiple polyadenylation sites and that polyadenylation signals may diverge significantly from the animal consensus sequence (13).
Sequence Comparison of H6H with Other Proteins-The protein data base (NBRF) was searched for proteins that have homology to H6H. Relatively high homology (24-33% identity in stretches of 250-350 amino acids) was found with the polypeptides encoded by several related genes which are in- duced in ripening fruit and/or wounded leaves of tomato (14- 16) and with the gene product of maize A2 locus (17). A recent antisense RNA experiment suggests that the tomato polypep- tides are involved in the conversion of aminocyclopropane-l- carboxylic acid to ethylene by the ethylene-forming enzyme (18). Although the activity of the ethylene-forming enzyme

Hyoscyamine G&Hydroxylase cDNA 9463
TABLE I Comparison of the amino acid composition of purified enzyme and
that derived from the H6H cDNA sequence The data for the purified enzyme were obtained from Ref. 4.
Amino acid purified H6H cDNA sequence Analysis of Deduced from
Glycine 23 18 Alanine 19 18 Valine 22 24 Leucine 38 38 Isoleucine 17 18 Serine 18 20 Threonine 20 17 Cysteine 5 6 Methionine 6 7 Aspartic acid + asparagine 33 34 Glutamic acid + glutamine 47 45 Arginine 8 7 Lysine 26 26 Histidine 8 8 Phenylalanine 14 14 Tyrosine 16 16 Typtophan 3 4 Proline 21 24
Total 344 344
can be measured in uiuo by feeding exogenous aminocyclopro- pane-1-carboxylic acid to excised leaves (19), the cofactors and the nature of the enzyme have been unclear. Sequence similarity between H6H and the polypeptides encoded by the ripening and wound-induced tomato genes suggests that the putative ethylene-forming enzyme might be a 2-oxoglutarate- dependent dioxygenase. The tomato-ripening related polypep- tides had also been reported to have amino acid sequence homology with flavone-3-hydroxylase of Antirrhinum majus, another 2-oxoglutarate-dependent dioxygenase involved in anthocyanin biosynthesis (cited in Ref. 18). The A2 locus of Zea mays was recently cloned using transposable elements as gene tags (17). Genetic evidence suggested that the A2 gene product functions in the formation of anthocyan from flavan- 3,4-cis-diol by a dehydration and an oxidation reaction. Al- though the enzyme activity of the gene product is yet to be demonstrated in vitro, the polypeptide encoded by the A2 gene might also be a member of the 2-oxoglutarate-dependent dioxygenases.
Lower but distinct amino acid sequence homology to H6H (20-2594 identity in stretches of 180-270 amino acids) was also found in DAOC synthase and IPN synthase. DAOC synthase is a 2-oxoglutarate-dependent dioxygenase that cat- alyzes late steps in cephalosporin biosynthesis. In Cephalo- sporium acremonium, a single bifunctional enzyme catalyzes both DAOC synthase and diacetylcephalosporin C synthase reactions (20), whereas these reactions are catalyzed by dif- ferent enzymes in Streptomyces claualigerus (21). IPN syn- thase catalyzes the removal of four hydrogen atoms from a tripeptide substrate, a key step common to p-lactam antibi- otics including penicillins and cephalosporins (22). Although IPN synthase also requires Fez+ and ascorbic acid for cataly- sis, it does not utilize 2-oxoglutarate as a co-substrate.
Three regions of relatively high sequence conservation among all or most of the homologous proteins are shown in Fig. 4. An amino acid sequence from His-217 to Asp-229 in H6H is conserved in all of the above-mentioned 2-oxoglutar- ate-dependent dioxygenases (including putative members of this group such as the ripening-related proteins and the A2 gene product), but not in IPN synthase. This sequence might be important for the enzyme function specific to the dioxy- genase (e.g. for binding 2-oxoglutarate co-substrate). In con- trast, two sequences in H6H, from Gly-59 to Gly-67 and from
H6H 51 TOME8 80 pTOM13 4 GTOMA 24 A2 83 DAWS-CA 22 DAOCS-SC 22 FRRCLRI I PNS
78 107 30 50
H6H ..r 217 TOME8 236 pTOM13 157 GTOMA 177
+ D 229 D 248 D 169 D 189
+ H6H 254 27 6 TOME8 272 pTOM13 194
294 216
GTOMA 214 236 A2 DAOCS-CA 224
290 312
DAOCS-SC 223 246
IPNS 252 245 274
FIG. 4. Comparison of amino acid sequence of H6H to other proteins. The deduced amino acid sequences of ripening related proteins (E8, pTOM13, and GTOMA) are from Refs. 14, 15, and 16, respectively, and the maize A2 gene from Ref. 17. The sequences of DAOC synthase are from C. acremonium (DAOCS-CA; Ref. 20) and S. clavaligerus (DAOCS-SC; Ref. 21). IPN synthase genes from seven microorganisms are known (321, and the amino acid sequence of C. acremonium IPN synthase (31) is shown here as a representative sequence. Closed circles below the IPN synthase sequence indicate positions of amino acid residues that are present at the same site in all seven IPN synthase sequences, and open circles indicate positions of similar but not identical residues in all IPN synthase sequences (32). Identical or similar amino acids found at the same site in more than seven sequences shown here are boxed. Arrows indicate con- served histidine residues that may be involved in iron binding (see the text for details). Amino acids are grouped according to Ref. 38.
FIG. 5. Nor the rn blot analysis of H. niger H6H mRNA. Total RNA (15 rg) was isolated from cultured roots (lane I ) , cultured cells (lane 2), roots (lane 3 ) , stem (lane 4 ) , and leaves (lane 5), separated electrophoretically on a 1.2% agarose gel containing formaldehyde, and transferred to nylon membrane. The membrane was hybridized to a "'P-labeled XhoI restriction fragment of H6H cDNA. Horizontal bar indicates a hybridized band a t 1.4 kb.
Val-253 to Val-276, are conserved in all of the above proteins including IPN synthase. This sequence similarity between 2- oxoglutarate-dependent dioxygenases and IPN synthase could support the speculation that both types of enzyme reactions involve similar highly reactive, ferry1 enzyme intermediates (22,23). Accordingly, these homologous sequences might rep- resent part of the iron-binding site. It is interesting to note that no cysteine residues are conserved in the sequences of the above proteins, but 2 histidine residues (His-217 and His- 274 in H6H) are well conserved, which raises a possibility that these residues might be involved in iron binding.
DAOC synthase and IPN synthase share a short homolo-

9464 Hyoscyamine 60-Hydroxylase cDNA
gous amino acid sequence containing the conserved cysteine residue (e.g. Cys-106 in C. acremonium IPN synthase) in all DAOC synthase and IPN synthase genes (20, 24-32). This cysteine, however, is not essential for the IPN synthase re- action because site-directed mutagenesis of Cys-106 to serine residue reduced, but did not completely destroy, IPN synthase activity of C. acremonium (33). Thus, it is not surprising that this cysteine residue is not conserved in other 2-oxoglutarate- dependent dioxygenases.
Another 2-oxoglutarate-dependent dioxygenase whose pri- mary amino acid sequence is known is proline hydroxylase (EC 1.14.11.2), which hydroxylates proline residues in X-Pro- Gly sequences of procollagens. The active enzyme is an asps tetramer, consisting of two different types of inactive mon- omer (34). We could not, however, detect any significant homology between H6H and either subunit, 01 (35, 36) and /3 (37 and references therein), probably because the large cata- lytic site of prolyl-4-hydroxylase may be cooperatively built up by both the 01 and 0 subunits (34).
Tissue-specific Expression of HGH-RNA blot hybridization of total RNA isolated from leaves, stems, roots, cultured roots, and cultured cells of H. niger showed a single transcript of about 1.4 kb only in the root tissues (Fig. 5 ) . Cultured roots contained much more abundant H6H mRNA than plant roots. Based on the number of positive clones obtained (about 100 using probes 1 and 2 ) from the total number of cDNA clones screened (about 30,000), it may be estimated that H6H mRNA consists of roughly 0.3% of the total mRNA in cultured roots. Such strong expression of H6H in cultured roots is explained anatomically by the specific localization of H6H protein in the pericycle, the cell type present only in the young root without secondary growth, such as cultured roots (7).
1. 2.
3.
4.
5.
6.
7.
8.
9.
10.
11.
"
REFERENCES Leete, E. (1990) Planta Med. 56,339-420 Hashimoto, T., and Yamada, Y. (1986) Plant Physiol. 81, 619- 625
Hashimoto, T., and Yamada, Y. (1987) Eur. J . Biochern. 164, 277-285
Yamada, Y., Okabe, S., and Hashimoto, T. (1990) Proc. Jpn . Acad. Ser. B Phys. Biol. Sci. 66, 73-76
Waller, G. R., and Nowacki, E. K. (1978) Alkaloid Biology and Metabolism in Plants, pp. 121-141, Plenum Publishing Corp., New York
Hashimoto, T., Yukimune, Y., and Yamada, Y. (1986) J. Plant Physiol. 124,61-75
Hashimoto, T., Hayashi, A., Amano, Y., Kohno, J., Iwanari, H., Usuda, S., and Yamada, Y. (1991) J. Biol. Chern. 266, 4648- 4653
Hashimoto, T., and Yamada, Y. (1987) Agric. Biol. Chern. 51,
Sambrook, J., Fritsch, E. F., and Maniatis, T. (1989) Molecular Cloning: A Laboratory Method, 2nd Ed., Cold Spring Harbor Laboratory Press, Cold Spring Harbor, NY
Sanger, F., Nicklen, S., and Coulson, A. R. (1977) Proc. Natl. Acad. Sci. U. S. A. 7 4 ~ 5463-5467
Laemmli, U. K. (1970) Nature 227,680-685
2769-2774
12. Proudfoot, N. J., and Brownlee, G. G. (1976) Nature 263, 211- 214
13. Dean, C., Tamaki, S., Dunsmuir, P., Favreau, M., Katayama, C., Dooner, H., and Bedbrook, J. (1986) Nucleic Acids Res. 14, 2229-2240
14. Deikman, J., and Fischer, R. L. (1988) E M B O J . 7,3315-3320 15. Holdsworth, M. J., Bird, C. R., Ray, J., Schuch, W., and Grierson,
D. (1987) Nucleic Acids Res. 15, 731-739 16. Holdsworth, M. J., Schuch, W., and Grierson, D. (1987) Nucleic
Acids Res. 15, 10600 17. Menssen, A., Hohmann, S., Martin, W., Schnable, P. S., Peterson,
P. A,, Saedler, H., and Gierl, A. (1990) E M B O J. 9,3051-3057 18. Hamilton, A. J., Lycett, G. W., and Grierson, D. (1990) Nature
19. Hoffman, N. E., and Yang, S. F. (1982) Plant Physiol. 69, 317- 322
20. Samson, S. M., Dotzlaf, J. E., Slisz, M. L., Becker, G. W., van Frank, R. M., Veal, L. E., Yeh, W.-K., Miller, J. R., Queener, S. W., and Ingolia, T. D. (1987) BiolTechnology 5, 1207-1214
21. Jensen, S. E., Westlake, D. W. S., and Wolfe, S. (1985) J. Antibiot.
22. Baldwin, J . E., and Abraham, E. (1988) Nat. Prod. Rep. 5 , 129- 145
23. Hanauske-Abel, H. M., and Giinzler, V. (1982) J. Theor. Biol. 94,421-455
24. Kovacevic, S., Weigel, B. J., Tobin, M. B., Ingolia, T. D., and Miller, J. R. (1989) J . Bacterid. 171, 754-760
25. Rambn, D., Carramolino, L., Patiiio, C., Sanchez, F., and Peiialva, M. A. (1987) Gene (Amst.) 57, 171-181
26. Leskiw, B. K., Aharonowitz, Y., Mevarech, M., Wolfe, S., Vining, L. C.. Westlake. D. W. S.. and Jensen. S. E. (1988) Gene ( A n s t . )
346,284-287
38,263-265
62, ia7-196 27. Shiffman. D.. Cohen. G.. Aharonowitz. Y.. Palissa. H.. von Doh-
Ten, H.,' Kleinkauf,' H.,' and Mevarech, M. (1990) Nucleic Acids Res. 18,660
28. Shiffman, D., Mevarech, M., Jensen, S. E., Cohen, G., and Ahar- onowitz, Y. (1988) Mol. & Gen. Genet. 214,562-569
29. Weigel, B. J., Burgett, S. G., Chen, V. J., Skatrud, P. L., Frolik, C. A., Queener, S. W., and Ingolia, T. D. (1988) J. Bacteriol.
30. Carr, L. G., Skatrud, P. L., Sheetz M. E., 11, Queener, S. W., and Ingolia, T. D. (1986) Gene (Amst.) 48, 257-266
31. Samson, S. M., Belagaje, R., Blankenship, D. T., Chapman, J. L., Perry, D., Skatrud, P. L., VanFrank, R. M., Abraham, E. P., Baldwin, J. E., Queener, S. W., and Ingolia, T. D. (1985) Nature 318, 191-194
32. Cohen, G., Shiffman, D., Mevarech, M., and Aharonowitz, Y. (1990) Trends Biotechnol. 8, 105-111
33. Samson, S. M., Chapman, J. L., Belagaje, R., Queener, S. W., and Ingolia, T. D. (1987) Proc. Natl. Acad. Sci. U. S. A . 84, 5705- 5709
34. Kivirikko, K. I.. Mvllvla. R.. and Pihlaianiemi, T. (1989) F A S E B
170,3817-3826
J. 3, 1609-1617 .
35. Helaakoski. T.. Vuori. K.. Mvllvla, R.. Kivirikko. K. I., and Pihlajaniemi,' T. (1989) Pro;. kat l . Acad. Sci. U. S. A. 86,
36. Bassuk, J. A,, Kao, W. W.-Y., Herzer, P., Kedersha, N. L., Seyer, J., DeMartino, J. A., Daugherty, B. L., Mark G. E., 111, and Berg, R. A. (1989) Proc. Natl. Acad. Sci. U. S . A. 86, 7382- 7386
37. Parkkonen, T., Kivirikko, K. I., and Pihlajaniemi, T. (1988) Biochern. J . 256, 1005-1011
38. Dayhoff, M. O., Schwartz, R. M., and Orcutt, B. C. (1978) in Atlas of Protein Sequences and Structure (Dayhoff, M. O., ed) Vol. 5, Suppl. 3, pp. 345-352, National Biomedical Research Foundation, Silver Spring, MD
4392-4396