molecular epidemiology and restriction site mapping of adenovirus
TRANSCRIPT

JOURNAL OF CLINICAL MICROBIOLOGY, June 1989, p. 1329-13340095-1137/89/061329-06$02.00/0Copyright ©3 1989, American Society for Microbiology
Molecular Epidemiology and Restriction Site Mapping ofAdenovirus Type 3 Genome Types
THOMAS ADRIAN,1* BARBARA BEST,' JOHN C. HIERHOLZER,2 AND REINHARD WIGAND'
Abteilung fur Virologie, Universitatskliniken, D-6650 Homburg, Saar, Federal Republic of Germany,' and Respiratoryand Enteric Viruses Branch, Division of Viral Diseases, Center for Infectious Diseases, Centers for Disease Control,
Atlanta, Georgia 303332
Received 2 December 1988/Accepted 9 March 1989
One hundred thirty-eight strains of adenovirus type 3 isolated from patients in the United States, WestGermany, and other regions between 1961 and 1984 were analyzed with six restriction endonucleases; 18genome types were found. BgIII was the most discriminative enzyme. Mapping of altered restriction sites wasalso performed for ail six enzymes. The genome types Dl (like the prototype) and D3 prevailed among 45 and47 strains, respectively. Ali genome types could be divided into two groups related to Dl or D3. Several clustersof infections by strains with the same genome type were observed. Only hints of differences were found in thepathogenicities of individual genome types. Dl strains were present in the United States and in Europe; groupD3 prevailed almost exclusively in the United States.
Adenovirus type 3 (Ad3) is one of the more commonhuman adenovirus types, causing mainly diseases of therespiratory tract, pharyngoconjunctival fever, conjunctivi-tis, and generalized illness (7, 16). For Ad7, a type biologi-cally and biochemically related to Ad3, a number of genometypes serologically identical to the prototype but different intheir DNA restriction patterns have been reported by Wadelland co-workers (20, 21, 24). One genome type deviating fromthe prototype was also described for Ad3 (Ad3a [20]),whereas five deviating genome types were found in Scotlandin 1981 and 1982 (14). In a study of the molecular epidemi-ology of adenoviruses, Wadell (20) reported a characteristicdistribution of Ad3 and Ad7 genome types in differentgeographic areas and time periods. From their data, Wadelland co-workers also deduced differences in the pathogenic-ities of certain genome types (21, 23).
Previous results (1, 19-21, 25) suggested that Ad3, 7, and21, members of subgenus B of human adenoviruses, are lessvariable than Adl, 2, 5, and 6 of subgenus C. However,Wadell et al. (20, 21, 23) had used only two restrictionenzymes, namely BamHI and SmaI, for the majority of theAd3 and 7 strains tested. More recently, Li and Wadell (10)analyzed 40 selected Ad7 strains with 12 restriction enzymesand identified 15 genome types; they subsequently identified17 genome types of Ad3 in a group of 59 strains from aroundthe world by using the same set of enzymes (11).
In the study reported here, we examined 138 strains ofAd3 with six endonucleases and identified 18 genome types.Description of their DNA restriction patterns, naming of thegenome types according to our previous proposal (1), andphysical mapping of the genome alterations were the mainobjectives of this study. We also analyzed the distribution ofthe genome types according to time, geographic location,and their association with disease as keys to "molecularepidemiology" (20, 21). A similar comprehensive study ofAd7 will be published elsewhere.
MATERIALS AND METHODSViruses and specimens. Besides the Ad3 prototype strain
GB, Ad3 strains were recovered from 137 patients between
* Corresponding author.
1961 and 1984 (see Fig. 2). A majority of the strains werefrom the United States (28 from Georgia and 66 from otherstates). Twenty-five strains from West Germany comprised16 isolates from Hannover, all isolated in 1981, mostly fromchildren with respiratory diseases, and 8 ocular isolates frompatients with conjunctivitis (1973, 1974, and 1978). Tenisolates from East Asia (see Fig. 2) comprised four fromHong Kong (1967) and six from Taiwan (1982). The remain-ing eight isolates were from sporadic cases in various partsof the world.The viruses were isolated from 80 respiratory, 43 ocular,
21 fecal, and 2 other clinical specimens. In nine cases, morethan one specimen per patient yielded Ad3; the genometypes were always identical within a patient. The viruseswere isolated in Hep2 or HEK cells (Atlanta) or HeLa cells(Homburg) and were propagated in HeLa cell cultures forDNA preparation. Because Ad3 replicates well in cell cul-ture, the isolates were passaged a few times only.
Serological identification. The isolates were typed by neu-tralization or hemagglutination inhibition tests with horse(Atlanta isolates) or rabbit (Homburg isolates) referenceantisera (4). In addition, representative strains of all genometypes were again analyzed by both tests, and all strains wereunequivocally identified as Ad3.DNA restriction analysis and evaluation. For screening
purposes, the DNA of viruses from HeLa cells was isolatedas described elsewhere (22). In some cases the DNA wasisolated from purified virions (6). Portions of the DNA wereanalyzed with the restriction endonucleases BamHI, BglII,BstEII, HindIII, KpnI, and SmaI according to publishedprotocols from Boehringer GmbH, Mannheim, Federal Re-public of Germany. The DNA fragments were stained withethidium bromide, made visible at 303 nm, and photo-graphed on Polaroid 665 film.
If purified DNA fragments were needed, the fragmentswere separated in low-melting-point agarose gels (SeaPlaqueagarose) and cut from the gel, and the DNA was purified byphenol extraction and an additional extraction with phenol-chloroform-isoamyl alcohol (25:24:1).Genome type naming. The naming system was the same as
that for Ad6 genome types (1). The endonucleases werelisted in alphabetical order (see above), and the restriction
1329
Vol. 27, No. 6
1330 ADRIAN ET AL.
TABLE 1. Genome types, representative strains,and enzyme codes of Ad3
Genome Strain Origin Enzyme No. oftype code Place Yr code' isolates
Dl GB Maryland 1953 111111 46D2 17 Louisiana 1961 221122 9D3 119 Illinois 1964 221123 47D4 277 North Carolina 1967 222122 1D5 311 Arizona 1968 231122 7D6 358 Senegal 1969 141111 1D7 725 Oregon 1972 121111 1D8 63 Homburg 1973 111211 1D9 1150 Maine 1976 241123 1D10 1244 Georgia 1977 251123 ilDl 1269 Puerto Rico 1977 231123 2D12 111079 Hannover 1981 161111 1D13 1282 Massachusetts 1981 221124 1D14 118675 Hannover 1981 111113 1D15 1406 Taiwan 1982 223322 2D16 P 1543 Taiwan 1982 233322 4D17 1595 New York 1983 331122 1D18 1657 Missouri 1983 221423 1` Enzyme order: BainHI, Bglll. BsIEII. HindIll, Kpnfl, Snal. DNA
restriction patterns correspond to those shown in Fig. 1.
patterns found were named 1 (prototype) and 2, 3, 4, etc. fordeviating patterns. The order was chosen according to theyear of isolation of the first strain found with the respectivepattern, called the representative strain. Dl, D2, etc. standfor "DNA variant," which is synonymous with "genometype." The representative strains were further named bystrain number, place, and year of isolation (18) (Table 1).The prototype is named Dl and has the enzyme code
111111, while D2 has the code 221122. This means that thegenome type differs from the prototype after BainHI, BgIII,KpnI, and SmaI digestion but not after BstEII and HindIIIdigestion.
Restriction site mapping. Physical maps of the genometypes were constructed by comparing published maps of theprototype (13, 18). The theoretical maps were proven exper-
imentally by Bal3l digestion, hybridization with knownprobes, and double digestion.
Identification of terminal fragments was based on diges-tion with the exonuclease Bal3l (Boehringer) (8). In somecases the kinetics were extended up to 90 min.The linear arrangement of the internal restriction frag-
ments was based on hybridization experiments with 32P-
labeled known DNA fragments (15, 17) or was deduced fromdouble-digest patterns by using a second restriction enzymewith known cleavage sites.
RESULTS
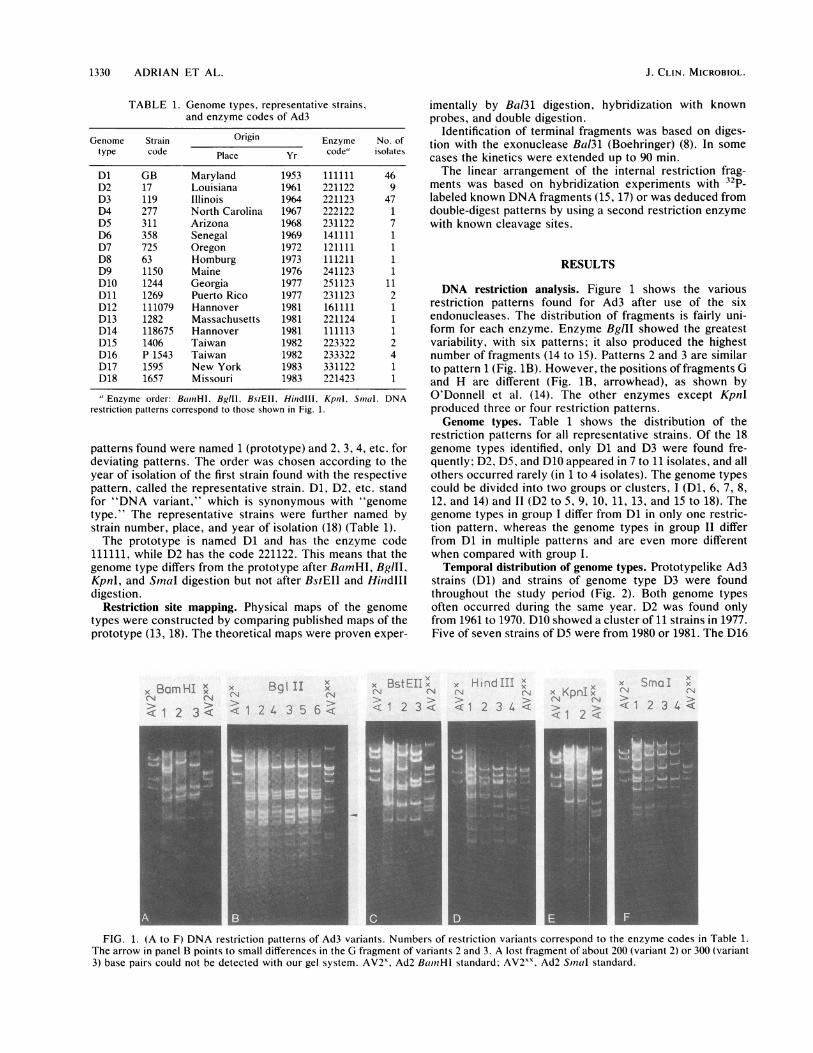
DNA restriction analysis. Figure 1 shows the variousrestriction patterns found for Ad3 after use of the sixendonucleases. The distribution of fragments is fairly uni-form for each enzyme. Enzyme BgIII showed the greatestvariability, with six patterns; it also produced the highestnumber of fragments (14 to 15). Patterns 2 and 3 are similarto pattern 1 (Fig. 1B). However, the positions of fragments Gand H are different (Fig. 1B, arrowhead), as shown byO'Donnell et al. (14). The other enzymes except KpnIproduced three or four restriction patterns.Genome types. Table 1 shows the distribution of the
restriction patterns for all representative strains. Of the 18genome types identified, only Dl and D3 were found fre-quently; D2, D5, and D1O appeared in 7 to 11 isolates, and allothers occurred rarely (in 1 to 4 isolates). The genome typescould be divided into two groups or clusters, I (Dl, 6, 7, 8,12, and 14) and Il (D2 to 5, 9, 10, 11, 13, and 15 to 18). Thegenome types in group I differ from Dl in only one restric-tion pattern, whereas the genome types in group II differfrom Dl in multiple patterns and are even more differentwhen compared with group I.
Temporal distribution of genome types. Prototypelike Ad3strains (Dl) and strains of genome type D3 were foundthroughout the study period (Fig. 2). Both genome typesoften occurred during the same year. D2 was found onlyfrom 1961 to 1970. D10 showed a cluster of 11 strains in 1977.Five of seven strains of D5 were from 1980 or 1981. The D16
x Bg3 II x
< 1 2 4 3 5 6 <
x Bst EI1 x
< 1 2 3 <
x H r c 'II X
< 1 2 3 4'<
x x;I'A s ^ w
<c 1 2 3 4 <`-1: ' «
FIG. 1. (A to F) DNA restriction patterns of Ad3 variants. Numbers of restriction variants correspond to the enzyme codes in Table 1.The arrow in panel B points to small differences in the G fragment of variants 2 and 3. A lost fragment of about 200 (variant 2) or 300 (variant3) base pairs could not be detected with our gel system. AV2x, Ad2 BamifHi standard; AV2XX Ad2 Sinal standard.
Bom HI ,
< 2 3 <
J. CLIN. MICROBIOL.

RESTRICTION MAPPING OF Ad3 GENOME TYPES 1331
53 61 63 65 67 69 71 73 75 77 79 '81 83 85Year
oUSA -FRG EEast Asia other *5 cases *l4 casesFIG. 2. Epidemiology of Ad3 genome types: temporal and geographic distribution. Symbols represent the number of strains and their
distribution (@, five U.S. isolates; M, 14 German isolates). The ordinate (D) indicates the genome type.
strains formed a cluster in 1982, as did five of nine D2 strainsin 1967.
Geographic distribution of genome types. Dl strains werefound in the United States from 1953 to 1981. This genometype predominated in Germany through 1982, and singleisolates came from Peru, Senegal (1972), and Australia(1984). In Germany, only two isolates were not Dl, namelyD12 and D14 from Hannover, which differ from Dl in onerestriction site only. No isolate of D3 and all related genometypes was found among our European isolates, except for asingle isolate of D5 from Spain (1970). Hence the genometypes related to D3 were mostly from the United States.There were several conspicuous clusters of infection by
strains of the same genome type. (i) Of 16 isolates fromHannover (1981), 14 were Dl. (ii) Of eight ocular isolatesfrom Germany, seven were Dl. (iii) Four isolates from HongKong (1967) were D2, all of them associated with upperrespiratory infection. (iv) All five isolates from Ohio (1973)were D3, and all of them were associated with pharyngocon-junctival fever. (v) Of 12 isolates from Georgia (June to July1977), 11 were D10; 7 of them were associated with pharyn-goconjunctival fever. These isolates were from a swimmingpool outbreak (12). (vi) Three isolates from Arizona (March1980) were D5; all were associated with pneumonia. (vii)Four D16 strains were isolated in Taiwan in 1982; strainswith this genome type were not isolated outside Taiwan.
Smaller or larger outbreaks were almost invariably asso-ciated with Ad3 of the same genome type. Patients in thesame outbreak tended to show similar symptoms as well.Genome types and disease. The clinical syndromes ob-
served in patients infected with various genome types arelisted in Table 2. The patients were arranged according to thepredominating syndrome if more than one syndrome wasrecorded. Differences in the pathogenicities or organ tro-pisms of individual genome types may be inferred from anuneven distribution of the clinical syndromes. The rarelyoccurring genome types are summarized in Table 2 for thetwo groups related to Dl or D3.
In view of the small numbers for most genome types andof the limitations in recording clinical syndromes, a formal
statistical analysis was not warranted. Hence only tenden-cies may be noted from the data, besides the geographicdistribution discussed above. Pneumonia and pharyngocon-junctival fever appeared to be less common in Dl than in D3infections, but conjunctivitis was more frequently found inDl than in D3. D10 infections included only one case ofupper respiratory infection and involved predominantlypharyngoconjunctival fever or pneumonia. The four fatalcases were apparently caused by Ad3 alone; no other agentswere identified in the patients. The Dl patient died fromencephalitis; Ad3 was isolated from the brain. The otherthree children died from pneumonia; they were infected withD3, D5, and Dl, which belonged to the group D3-relatedgenome types. The ages of the four children were 3, 1, 6, and1.
Restriction site mapping. The alterations in restrictionfragments for the restriction variants are shown in Fig. 3 foreach enzyme. The maps were adapted from published mapsof the Ad3 prototype 3 (13, 14, 18). These physical mapswere scrutinized by double digestion, Bal3l kinetics, and
TABLE 2. Clinical syndromes in patients infectedwith various genome types of Ad3
No. of patients infected with:Syndrome D8,D4, 9 11,
Dl 12, 14" D2 D3 D5 D10 13, 15-18"
URIb 21 2 5 16 3 1 2Pneumonia 2 1 1 7 3 2 2PCF' 7 1 1 17 1 7 2Conjunctivitisd 10 1 0 3 0 1 3Other 6 0 2 4 0 0 4
Fatal cases 1 0 0 1 1 0 1
' Rare genome types, closely related to Dl or to D2 or 3, respectively(Table 1).
b URI, Upper respiratory infection, including tonsillitis, pharyngitis, orfever.'PCF, Pharyngoconjunctival fever.d Including keratoconjunctivitis.Deaths due to encephalitis (Dl) or pneumonia (D3, D5, and Dl).
18
16
14
12
10.
8
6
4'
2'
il
OeO
0~~~~~
0 6 000 00
0
006 000 000 0 000 00 0 00 0 00 88 0 00 0 00 00 00
0 0 0 0
06 00 O à 0 66,0O 0 0 88Q OnoDoaooo ^ o aa S° °°° ° °o°n8o °
D
18
16
14
12
1i0
4
-2
VOL. 27, 1989
1332 ADRIAN ET AL.
2L t
2,3
i 1 1 i 1 1 1 .f .1 1
4 ô 5Ë tt 1 --.i 1 1 1- 1 1 1 1 1 1 1 1 1
Bam HI
BgI II2
r T
I
3
1-r 1 Bst E Il
, , , HindIII2
1I1 --I Kpn I
-+ Sma I
0 0.1 02 0.3 0.4 0.5 0.6 .7 08 0.9 1
mop units
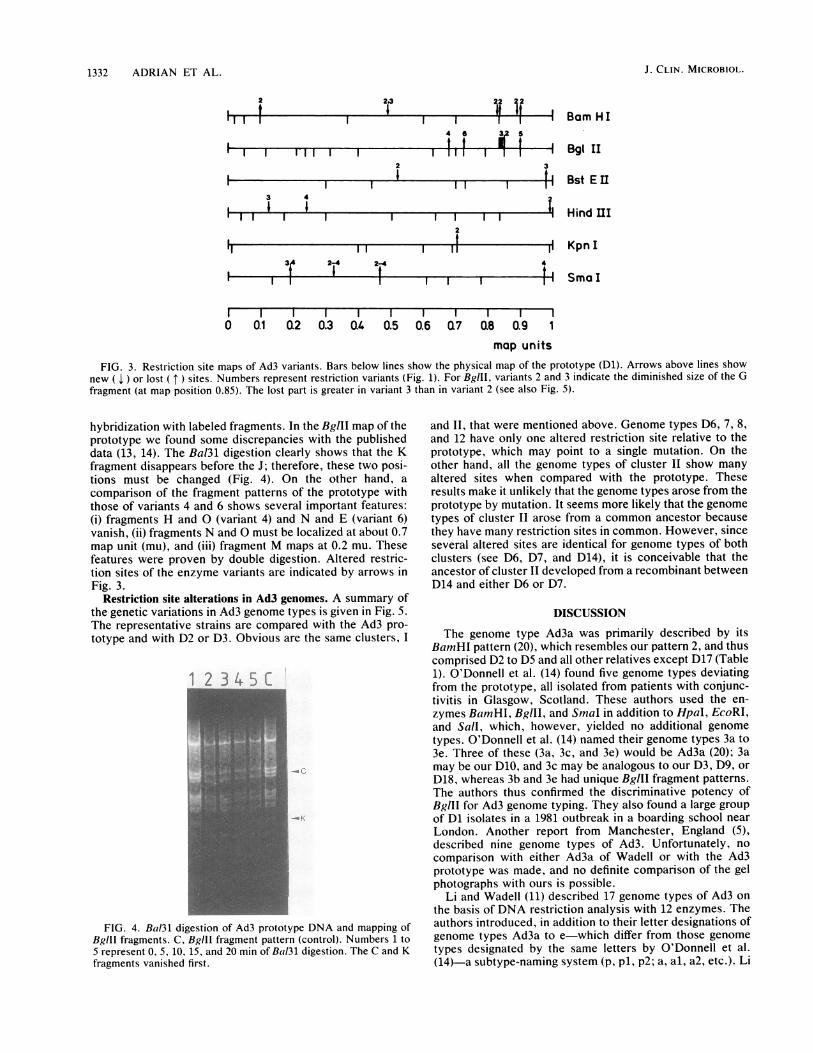
FIG. 3. Restriction site maps of Ad3 variants. Bars below lines show the physical map of the prototype (Dl). Arrows above lines shownew ( 1 ) or lost ( t ) sites. Numbers represent restriction variants (Fig. 1). For BglIl, variants 2 and 3 indicate the diminished size of the Gfragment (at map position 0.85). The lost part is greater in variant 3 than in variant 2 (see also Fig. 5).
hybridization with labeled fragments. In the BglII map of theprototype we found some discrepancies with the publisheddata (13, 14). The Bal3l digestion clearly shows that the Kfragment disappears before the J; therefore, these two posi-tions must be changed (Fig. 4). On the other hand, a
comparison of the fragment patterns of the prototype withthose of variants 4 and 6 shows several important features:(i) fragments H and O (variant 4) and N and E (variant 6)vanish, (ii) fragments N and O must be localized at about 0.7map unit (mu), and (iii) fragment M maps at 0.2 mu. Thesefeatures were proven by double digestion. Altered restric-tion sites of the enzyme variants are indicated by arrows inFig. 3.
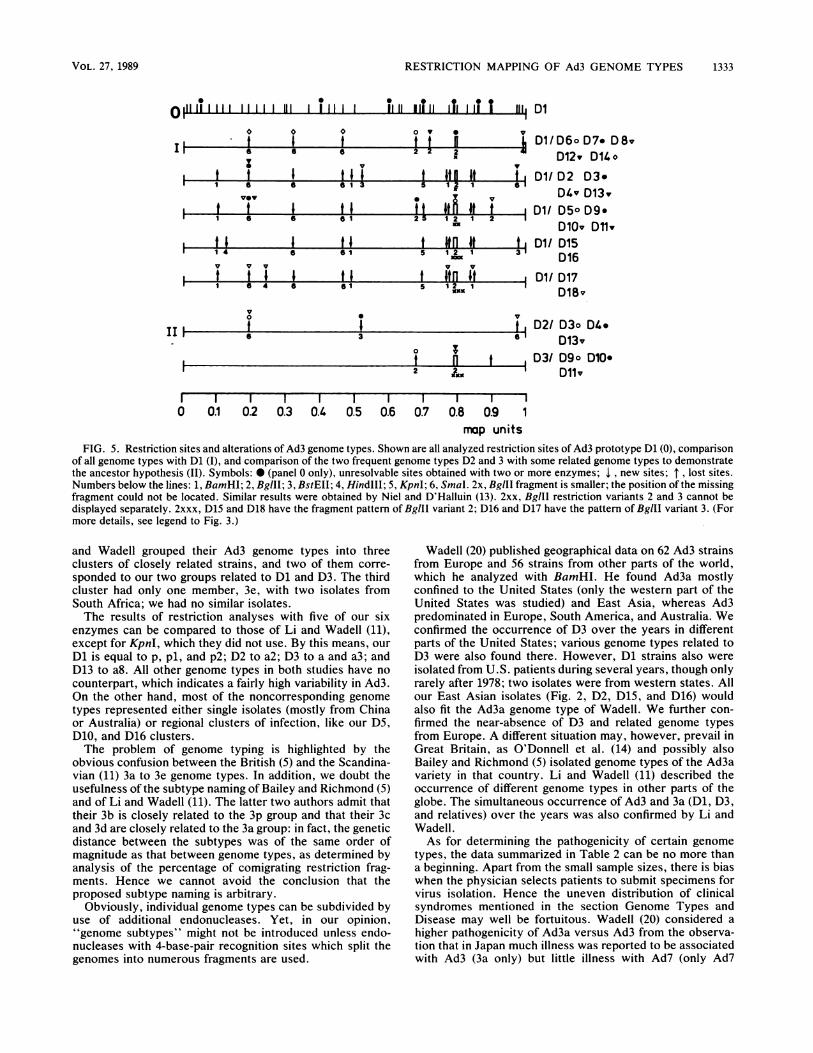
Restriction site alterations in Ad3 genomes. A summary ofthe genetic variations in Ad3 genome types is given in Fig. 5.The representative strains are compared with the Ad3 pro-
totype and with D2 or D3. Obvious are the same clusters, I
1 2 3 4L8
FIG. 4. Bal3l digestion of Ad3 prototype DNA and mapping ofBglll fragments. C, Bglll fragment pattern (control). Numbers 1 to5 represent 0, 5, 10, 15, and 20 min of Bal3l digestion. The C and Kfragments vanished first.
and 11, that were mentioned above. Genome types D6, 7, 8,and 12 have only one altered restriction site relative to theprototype, which may point to a single mutation. On theother hand, all the genome types of cluster Il show manyaltered sites when compared with the prototype. Theseresults make it unlikely that the genome types arose from theprototype by mutation. It seems more likely that the genometypes of cluster Il arose from a common ancestor becausethey have many restriction sites in common. However, sinceseveral altered sites are identical for genome types of bothclusters (see D6, D7, and D14), it is conceivable that theancestor of cluster Il developed from a recombinant betweenD14 and either D6 or D7.
DISCUSSION
The genome type Ad3a was primarily described by itsBamHI pattern (20), which resembles our pattern 2, and thuscomprised D2 to D5 and all other relatives except D17 (Table1). O'Donnell et al. (14) found five genome types deviatingfrom the prototype, all isolated from patients with conjunc-tivitis in Glasgow, Scotland. These authors used the en-
zymes BamHI, BglII, and SmnaI in addition to HpaI, EcoRI,and SalI, which, however, yielded no additional genometypes. O'Donnell et al. (14) named their genome types 3a to3e. Three of these (3a, 3c, and 3e) would be Ad3a (20); 3amay be our D10, and 3c may be analogous to our D3, D9, or
D18, whereas 3b and 3e had unique BglII fragment patterns.The authors thus confirmed the discriminative potency ofBglII for Ad3 genome typing. They also found a large groupof Dl isolates in a 1981 outbreak in a boarding school near
London. Another report from Manchester, England (5),described nine genome types of Ad3. Unfortunately, no
comparison with either Ad3a of Wadell or with the Ad3prototype was made, and no definite comparison of the gelphotographs with ours is possible.
Li and Wadell (11) described 17 genome types of Ad3 on
the basis of DNA restriction analysis with 12 enzymes. Theauthors introduced, in addition to their letter designations ofgenome types Ad3a to e-which differ from those genometypes designated by the same letters by O'Donnell et al.(14)-a subtype-naming system (p. pl, p2; a, al, a2, etc.). Li
3 4I I
1-
h-
l 12-4 24
J. CLIN. MICROBIOL.
3
i 1
2 2 2il it i
RESTRICTION MAPPING OF Ad3 GENOME TYPES 1333
o0,'' "" 1 1 1 1 1 111 ,T Il Tl Il lTIl ,li T î1iîj Dl
t tS S 6y
o v 0
2 2 2-i
|1 6 6 6 ~~ ~ ~~~~13 5 121 6
1 2 2I 1 5 6 1 25 ~1
S146 61 S 1 2 132
v v v v vn
i1 6 4 S 51 S 12 1
D1/ D6o0D7 D 8vD12, D14 o
D1/ D2 D3eD4v D13,v
D1/ D50 D9D10v Dl1,
D1/ D15D16
Dl/ D17D18v
vot
e
ot n2 î
v
t D2/ D3o D4.6- D13v
t 1 D30/ D9o D10-Dlv
0 0.1 0.2 0.3 0.4 0.5 0.6 0.7 0.8 0.9 1map units
FIG. 5. Restriction sites and alterations of Ad3 genome types. Shown are all analyzed restriction sites of Ad3 prototype Dl (0), comparisonof all genome types with Dl (t), and comparison of the two frequent genome types D2 and 3 with some related genome types to demonstratethe ancestor hypothesis (IH). Symbols: * (panel O only), unresolvable sites obtained with two or more enzymes; 1 , new sites; lost sites.Numbers below the lines: 1, BamHI; 2, BglII; 3, BstEII; 4, HindIII; 5, KpnI; 6, SmaI. 2x, BglII fragment is smaller; the position of the missingfragment could not be located. Similar results were obtained by Niel and D'Halluin (13). 2xx, BgIII restriction variants 2 and 3 cannot bedisplayed separately. 2xxx, D15 and D18 have the fragment pattern of BgIlI variant 2; D16 and D17 have the pattern of BglII variant 3. (Formore details, see legend to Fig. 3.)
and Wadell grouped their Ad3 genome types into threeclusters of closely related strains, and two of them corre-sponded to our two groups related to Dl and D3. The thirdcluster had only one member, 3e, with two isolates fromSouth Africa; we had no similar isolates.The results of restriction analyses with five of our six
enzymes can be compared to those of Li and Wadell (11),except for KpnI, which they did not use. By this means, ourDl is equal to p, pl, and p2; D2 to a2; D3 to a and a3; andD13 to a8. All other genome types in both studies have nocounterpart, which indicates a fairly high variability in Ad3.On the other hand, most of the noncorresponding genometypes represented either single isolates (mostly from Chinaor Australia) or regional clusters of infection, like our D5,D10, and D16 clusters.The problem of genome typing is highlighted by the
obvious confusion between the British (5) and the Scandina-vian (11) 3a to 3e genome types. In addition, we doubt theusefulness of the subtype naming of Bailey and Richmond (5)and of Li and Wadell (11). The latter two authors admit thattheir 3b is closely related to the 3p group and that their 3cand 3d are closely related to the 3a group: in fact, the geneticdistance between the subtypes was of the same order ofmagnitude as that between genome types, as determined byanalysis of the percentage of comigrating restriction frag-ments. Hence we cannot avoid the conclusion that theproposed subtype naming is arbitrary.
Obviously, individual genome types can be subdivided byuse of additional endonucleases. Yet, in our opinion,"genome subtypes" might not be introduced unless endo-nucleases with 4-base-pair recognition sites which split thegenomes into numerous fragments are used.
Wadell (20) published geographical data on 62 Ad3 strainsfrom Europe and 56 strains from other parts of the world,which he analyzed with BamHI. He found Ad3a mostlyconfined to the United States (only the western part of theUnited States was studied) and East Asia, whereas Ad3predominated in Europe, South America, and Australia. Weconfirmed the occurrence of D3 over the years in differentparts of the United States; various genome types related toD3 were also found there. However, Dl strains also were
isolated from U.S. patients during several years, though onlyrarely after 1978; two isolates were from western states. Allour East Asian isolates (Fig. 2, D2, D15, and D16) wouldalso fit the Ad3a genome type of Wadell. We further con-firmed the near-absence of D3 and related genome typesfrom Europe. A different situation may, however, prevail inGreat Britain, as O'Donnell et al. (14) and possibly alsoBailey and Richmond (5) isolated genome types of the Ad3avariety in that country. Li and Wadell (11) described theoccurrence of different genome types in other parts of theglobe. The simultaneous occurrence of Ad3 and 3a (Dl, D3,and relatives) over the years was also confirmed by Li andWadell.As for determining the pathogenicity of certain genome
types, the data summarized in Table 2 can be no more thana beginning. Apart from the small sample sizes, there is biaswhen the physician selects patients to submit specimens forvirus isolation. Hence the uneven distribution of clinicalsyndromes mentioned in the section Genome Types andDisease may well be fortuitous. Wadell (20) considered a
higher pathogenicity of Ad3a versus Ad3 from the observa-tion that in Japan much illness was reported to be associatedwith Ad3 (3a only) but little illness with Ad7 (only Ad7
Il
h
VOL. 27, 1989

1334 ADRIAN ET AL.
genome type). An outbreak of eight unusually severe Ad3infections in children was recently described in Israel (9); itwould be of interest to know the genome type(s) involved.Among all adenovirus types studied to date, there is littleconvincing evidence of a relation between genome type andseverity of disease.For further studies of Ad3 genome types, the enzymes
BamHI, BglII, HindIII, and SmaI should be used, possiblysupplemented by PstI (5), while BstEII and KpnI may be asdispensable as EcoRI, HpaI, and SalI are (14). However, Liand Wadell (10) proposed using BamHI, BclI, BgIII, HpaI,SalI, and SmaI, which were sufficient to identify all of their17 genome types. Although we used six restriction enzymesfor all strains, one should bear in mind that with about 60restriction fragments, each indicating 6 base pairs, onlyabout 1% of the genome was scanned and compared. Hencethe extent of variability is incompletely known and willremain so until DNA sequences of parts of the genome oreven of entire genomes are available. However, the degreeof relatedness may be estimated by comigration analysis (2)or by comparing altered restriction sites.We came to the following hypothesis regarding the evolu-
tion of Ad3 genome types. The Dl and D2 or 3 genometypes, circulating in relatively early years, may be consid-ered master strains, from which the genetically relatedgenome types may have developed by stepwise mutations.However, Dl and D2 or 3 are so different that their arisingfrom a coinmon ancestor would have required many muta-tion steps over a long period or several recombinationevents. Since all strains are identical by the serologicaltechniques used, the epitopes for neutralization (hexon) andhemagglutination inhibition (fiber) have not been altered. Itfurther appears that some genome types are more likely tospread and to survive over the years than others that werefound only once. One possible reason for this differentbehavior may be differences in the replication cycle and viralrelease among genome types.
ACKNOWLEDGMENTS
The technical assistance of M. Becker, D. Keller, and G. Schaferis gratefully acknowledged.The work was supported by the Deutsche Forschungsgemein-
schaft (Wi 2/21), Bundesministerium fur Jugend, Familie, Frauenund Gesundheit, Federal Republic of Germany, and Forderervereinder Deutschen Vereinigung zur Bekampfung der Viruskrankheiten,Munich, Federal Republic of Germany.
LITERATURE CITED1. Adrian, T., B. Best, and R. Wigand. 1985. A proposal to name
genome types exemplified by adenovirus type 6. J. Gen. Virol.66:2685-2691.
2. Adrian, T., and W. Heinrich. 1986. COMAP: a comigratinganalysis program for estimating the relationship of adenoviruseson the genome level. Nucleic Acids Res. 14:559-565.
3. Adrian, T., G. Wadell, J. C. Hierbolzer, and R. Wigand. 1986.DNA restriction analysis of adenovirus prototypes 1 to 41.Arch. Virol. 91:277-290.
4. Adrian, T., R. Wigand, and J. C. Hierholzer. 1985. Immunolog-ical and biochemical characterization of adenovirus strains ofsubgenus B. Il. DNA analysis. Arch. Virol. 84:78-89.
5. Bailey, A. S., and S. J. Richmond. 1986. Genetic heterogeneityof recent isolates of adenovirus types 3, 4, and 7. J. Clin.Microbiol. 24:30-35.
6. Doerfler, W. 1969. Nonproductive infection of baby hamsterkidney cells (BHK21) with adenovirus type 12. Virology 38:
587-606.7. Foy, H. M., and J. T. Grayston. 1982. Adenoviruses, p. 67-84.
In A. S. Evans (ed.), Viral infections of humans. Epidemiologyand control, 2nd ed. John Wiley & Sons, Inc., New York.
8. Legerski, R. J., J. L. Hodnett, and H. B. Gray. 1978. Extracel-lular nucleases of pseudomonas Bal3l. III. Use of the double-strand deoxyribonuclease activity as the basis of a convenientmethod for the mapping of fragments of DNA produced bycleavage with restriction enzymes. Nucleic Acids Res. 5:1445-1464.
9. Levy, Y., M. Nitzan, A. Beharab, A. Zeharia, T. Schoenfeld, J.Nutman, L. Rannon, and R. Steinherz. 1986. Adenovirus type 3infection with systematic manifestation in apparently normalchildren. Isr. J. Med. Sci. 22:774-778.
10. Li, Q.-G., and G. Wadell. 1986. Analysis of 15 different genometypes of adenovirus type 7 isolated on five continents. J. Virol.60:331-335.
11. Li, Q.-G., and G. Wadell. 1988. Comparison of 17 differentgenome types of adenovirus type 3 identified among strainsrecovered from six continents. J. Clin. Microbiol. 26:1009-1015.
12. Martone, W. J., J. C. Hierholzer, R. A. Keenlyside, D. W.Fraser, L. J. D'Angelo, and W. G. Winkler. 1980. An outbreakof adenovirus type 3 disease at a private recreation centerswimming pool. Am. J. Epidemiol. 111:229-237.
13. Niel, C., and J. C. D'Halluin. 1984. Restriction maps of humanadenovirus types 2, 3, and 5 for BstEII, MluI, NruI, and Sfi1endonucleases. Gene 31:305-308.
14. O'Donnell, B., E. Bell, S. B. Payne, V. Mautner, and U.Desselberger. 1986. Genome analysis of species 3 adenovirusesisolated during summer outbreaks of conjunctivitis and pharyn-goconjunctival fever in Glasgow and London areas in 1981. J.Med. Virol. 18:213-227.
15. Rigby, P. W. J., M. Dieckman, C. Rhodes, and P. Berg. 1977.Labeling deoxyribonucleic acid to high specific activity in vitroby nick translation with DNA polymerase I. J. Mol. Biol.113:237-251.
16. Snavely, S. R., and C. Liu. 1984. Adenoviruses, p. 779-784. InR. B. Belshe (ed.), Textbook of human virology. PSG Publish-ing Co., Inc., Littleton, Mass.
17. Southern, E. M. 1975. Detection of specific sequences amongDNA fragments separated by gel electrophoresis. J. Mol. Biol.98:503-517.
18. Tibbetts, C. 1977. Physical organization of subgenus B humanadenovirus genomes. J. Virol. 24:564-579.
19. van der Avoort, H. G. A. M., T. Adrian, R. Wigand, A. G.Wermenbol, T. P. L. Zomerdijk, and J. C. de long. 1986.Molecular epidemiology of adenovirus type 21 in The Nether-lands and the Federal Republic of Germany from 1960 to 1985.J. Clin. Microbiol. 24:1084-1088.
20. Wadell, G. 1984. Molecular epidemiology of adenoviruses.Curr. Top. Microbiol. Immunol. 110:191-220.
21. Wadell, G., M. K. Cooney, A. da Costa Linhares, L. de Silva,M. L. Kennett, R. Kono, R. Gui-Fang, K. Lindman, J. P.Nascimento, B. D. Schoub, and C. D. Smith. 1985. Molecularepidemiology of adenoviruses: global distribution of adenovirus7 genome types. J. Clin. Microbiol. 21:403-408.
22. Wadell, G., and J. C. de Jong. 1980. Restriction endonucleasesin identification of a genome type of adenovirus 19 associatedwith keratoconjunctivitis. Infect. Immun. 27:292-296.
23. Wadell, G., J. C. de Jong, and S. Wolontis. 1981. Molecularepidemiology of adenoviruses: alternating appearance of twodifferent genome types of adenovirus 7 during epidemic out-breaks in Europe from 1958 to 1980. Infect. Immun. 34:368-372.
24. Wadell, G., and T. M. Varsanyi. 1978. Demonstration of threedifferent subtypes of adenovirus type 7 by DNA restriction sitemapping. Infect. Immun. 21:238-246.
25. Wigand, R., and T. Adrian. 1986. Classification and epidemiol-ogy of adenoviruses, p. 409-441. In W. Doerfler (ed.), Adeno-virus DNA. The viral genome and its expression. MartinusNijhoff Publishing, Boston.
J. CLIN. MICROBIOL.