monoamine oxidase b (maob)-containing structures in maoa-deficient transgenic mice
TRANSCRIPT

Ž .Brain Research 771 1997 121–132
Research report
Ž .Monoamine oxidase B MAOB -containing structures in MAOA-deficienttransgenic mice
Keiko Ikemoto a,1, Kunio Kitahama a,), Isabelle Seif b, Toshihiro Maeda c, Edward De Maeyer b,Jean-Louis Valatx a
a Departement de Medecine Experimentale, INSERM U52, CNRS ERS 5645, UniÕersite Claude Bernard, 8 aÕenue Rockefeller, 69373 Lyon Cedex 8,´ ´ ´ ´France
b CNRS UMR177, Institut Curie, Orsay, Francec Department of Anatomy, Shiga UniÕersity of Medical Science, Otsu, Japan
Accepted 25 June 1997
Abstract
Ž . Ž .Monoamine oxidase MAO -containing structures were studied for the first time in type A MAO MAOA -deficient transgenic miceŽ .Tg8 derived from C3H strain, using MAO enzyme histochemistry. In this mutant line, MAOA activity was not detected in neurons of
Ž .the locus coeruleus. In contrast, in their dorsal raphe neurons, we noted an intense activity of type B MAO MAOB . Based onŽ .pharmacological MAOA suppression experiments employing a specific inhibitor clorgyline , we confirmed that the localization of
MAOB-positive structures are not different between Tg8 mutant and normal C3H line. Many of MAOB-positive structures which havenot been described previously in the rat, cat and primates were described in this study. In the forebrain, MAOB-containing neurons werediscriminated in the striatum, septal nuclei, major island of Calleja, diagonal band, medial forebrain bundle, ventral pallidum andamygdaloid nucleus. Stained neurons in the thalamus and hypothalamus were much more extensively distributed in the mouse than therat. Pontine laterodorsal tegmental neurons showed MAOB activity. The present data suggest that serotonin, a preferential substrate forMAOA, can be oxidized by MAOB in MAOA-deficient Tg8 mice. q 1997 Elsevier Science B.V.
Keywords: Mouse; Monoamine oxidase; Transgenic animal; Histochemistry; Catecholamine; Serotonin; Aggressive behavior; Species difference
1. Introduction
Ž .The degradative enzyme monoamine oxidase MAO ispresent on the outer membrane of mitochondria and main-tains low cytosolic concentrations of monoamines such as
Ž . Ž . Ž .serotonin 5-HT , noradrenaline NA and dopamine DA .There are two MAO subtypes, MAOA and MAOB. Theyare encoded by separate genes, which are closely linked onthe X chromosome, and share 70% similarity in amino
w xacid sequence 3 . In vitro experiments with brain extractshave shown that MAOA and MAOB have different butoverlapping substrate specificities. DA and tyramine aregood substrates for both types of MAO. 5-HT is a sub-
w xstrate for MAOA better than for MAOB in vitro 20 . In
) Corresponding author.1 Present address: Department of Anatomy, School of Medicine, Fujita
Health University, Toyoake, Aichi 470-11, Japan.
vivo, the substrate specificities of the MAO subtypesmight be regionally modulated, and thus, the relevance ofthe in vitro results is not clear.
The tissue and cellular distributions of MAOA andŽ .MAOB in normal brains primates, cats and rodents have
w xbeen studied by enzyme histochemistry 1,9 , immunohis-w x w xtochemistry 10,14,28,29 , in situ hybridization 18 and
w xradioautography 25 . MAOB is abundant not only inw x w xserotonergic 9,15,19 or histaminergic neurons 16,17 , but
w xalso in astrocytes 10,15,29 . In contrast, the highest levelof MAOA is localized in noradrenergic neurons such as
Ž . w xthe locus coeruleus LC 9,18,25,28,29 , and in fact, itappears to be the sole MAO subtype in these neurons.
At present, to localize MAOB-containing structures,there is no specific antibody to mouse MAOB. In addition,using MAO histochemistry coupled with specific MAOAinhibitors, it has been difficult to totally distinguishMAOB-positive structures from strongly MAOA-positive
0006-8993r97r$17.00 q 1997 Elsevier Science B.V. All rights reserved.Ž .PII S0006-8993 97 00832-9
( )K. Ikemoto et al.rBrain Research 771 1997 121–132122
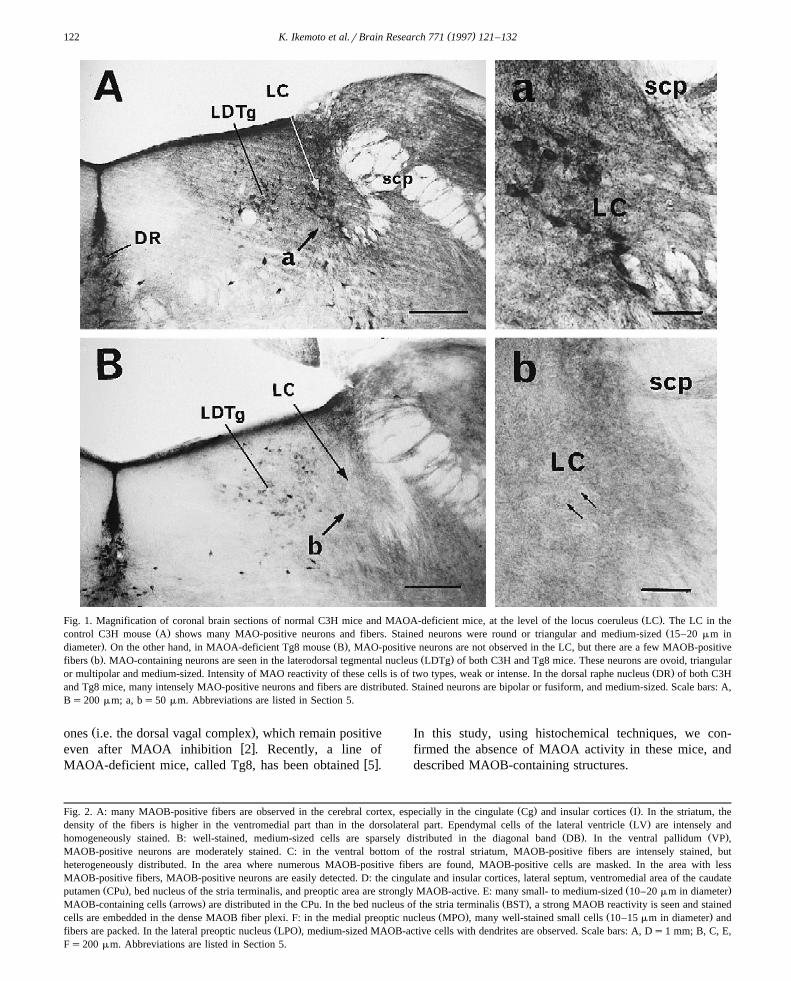
Ž .Fig. 1. Magnification of coronal brain sections of normal C3H mice and MAOA-deficient mice, at the level of the locus coeruleus LC . The LC in theŽ . Žcontrol C3H mouse A shows many MAO-positive neurons and fibers. Stained neurons were round or triangular and medium-sized 15–20 mm in
. Ž .diameter . On the other hand, in MAOA-deficient Tg8 mouse B , MAO-positive neurons are not observed in the LC, but there are a few MAOB-positiveŽ . Ž .fibers b . MAO-containing neurons are seen in the laterodorsal tegmental nucleus LDTg of both C3H and Tg8 mice. These neurons are ovoid, triangular
Ž .or multipolar and medium-sized. Intensity of MAO reactivity of these cells is of two types, weak or intense. In the dorsal raphe nucleus DR of both C3Hand Tg8 mice, many intensely MAO-positive neurons and fibers are distributed. Stained neurons are bipolar or fusiform, and medium-sized. Scale bars: A,Bs200 mm; a, bs50 mm. Abbreviations are listed in Section 5.
Ž .ones i.e. the dorsal vagal complex , which remain positivew xeven after MAOA inhibition 2 . Recently, a line of
w xMAOA-deficient mice, called Tg8, has been obtained 5 .
In this study, using histochemical techniques, we con-firmed the absence of MAOA activity in these mice, anddescribed MAOB-containing structures.
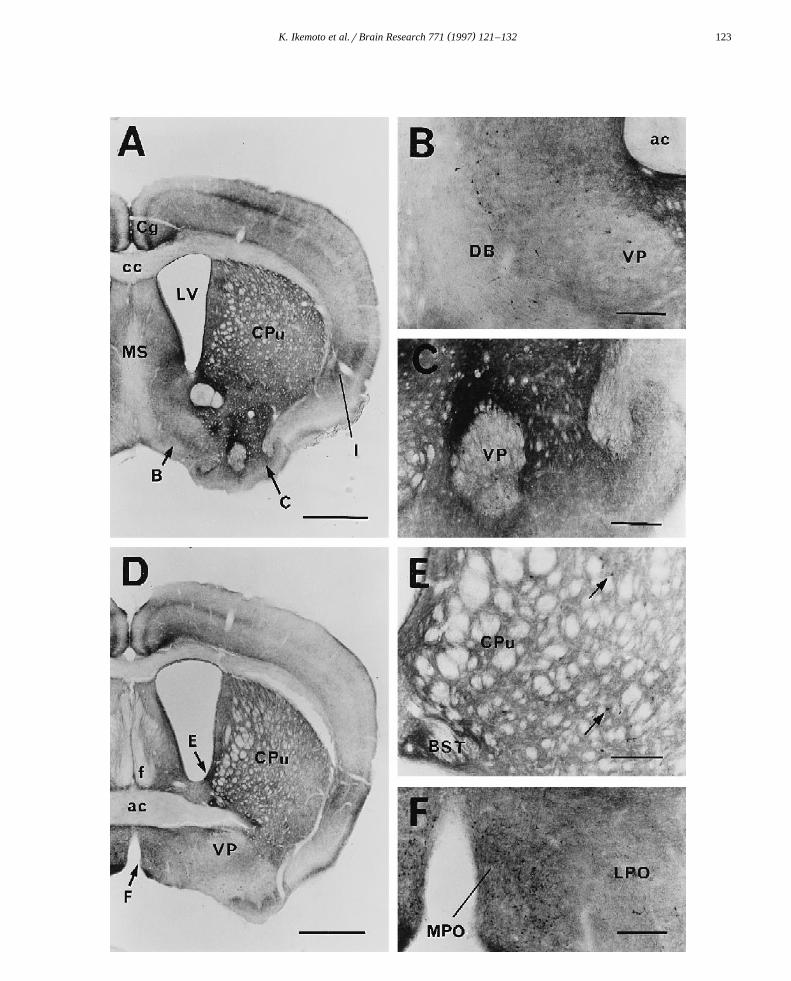
Ž . Ž .Fig. 2. A: many MAOB-positive fibers are observed in the cerebral cortex, especially in the cingulate Cg and insular cortices I . In the striatum, theŽ .density of the fibers is higher in the ventromedial part than in the dorsolateral part. Ependymal cells of the lateral ventricle LV are intensely and
Ž . Ž .homogeneously stained. B: well-stained, medium-sized cells are sparsely distributed in the diagonal band DB . In the ventral pallidum VP ,MAOB-positive neurons are moderately stained. C: in the ventral bottom of the rostral striatum, MAOB-positive fibers are intensely stained, butheterogeneously distributed. In the area where numerous MAOB-positive fibers are found, MAOB-positive cells are masked. In the area with lessMAOB-positive fibers, MAOB-positive neurons are easily detected. D: the cingulate and insular cortices, lateral septum, ventromedial area of the caudate
Ž . Ž .putamen CPu , bed nucleus of the stria terminalis, and preoptic area are strongly MAOB-active. E: many small- to medium-sized 10–20 mm in diameterŽ . Ž .MAOB-containing cells arrows are distributed in the CPu. In the bed nucleus of the stria terminalis BST , a strong MAOB reactivity is seen and stained
Ž . Ž .cells are embedded in the dense MAOB fiber plexi. F: in the medial preoptic nucleus MPO , many well-stained small cells 10–15 mm in diameter andŽ .fibers are packed. In the lateral preoptic nucleus LPO , medium-sized MAOB-active cells with dendrites are observed. Scale bars: A, Ds1 mm; B, C, E,
Fs200 mm. Abbreviations are listed in Section 5.
( )K. Ikemoto et al.rBrain Research 771 1997 121–132 123

( )K. Ikemoto et al.rBrain Research 771 1997 121–132124
2. Materials and methods
Eighteen mice of the normal C3H line as controls and16 mice of the mutant Tg8 line of both sexes, aged from5–12 months, were used. Among them, eight C3H miceand four Tg8 mice were used for pharmacological suppres-sion of both types of MAO. For MAO enzyme histochem-
w xistry, a modified coupled peroxidation method 11,19 wasapplied. Each mouse was anesthetized by an intraperi-toneal injection with sodium pentobarbital, and perfusedthrough the cardiac ventricle with 30 ml of 0.01 M phos-
Žphate-buffered saline 0.01 M phosphate buffer q0.9%.NaCl, pH 7.4 , and subsequently with 100 ml of ice-cold
fixative containing 2% paraformaldehyde and 2% glu-Ž .taraldehyde in 0.1 M phosphate buffer pH 7.4 at a rate of
20 mlrmin. Brains were immediately cut into 50-mmsections with a vibratome and were incubated overnight ina reaction medium under gentle agitation at 48C. Themedium consisted of 7.5 mg tyramine hydrochloride, 2 mg
X Ž .3,3 -diaminobenzidine tetrahydrochloride DAB, Sigma , 5Ž .mg horseradish peroxidase Toyobo, grade III, Japan , 30
mg nickel ammonium sulfate, and 6.5 mg sodium azide, inŽ .10 ml of 0.05 M Tris–HCl buffer pH 7.6 was used. In
case of pharmacological suppression, brain sections wereincubated in 0.05 M Tris–HCl buffer with 10y8 –10y4
mol clorgyline or deprenyl for 30 min just before theincubation in the reaction medium. The former is a specificMAOA inhibitor, and the latter is a specific MAOB in-hibitor. The reaction was terminated by placing the sec-tions in 0.05 M Tris–HCl buffer. No staining was obtainedwhen tyramine was omitted from the medium. The MAO-positive neurons were mapped using a camera lucida. Todefine the anatomical territory, the atlas of Franklin and
w xPaxinos 8 was referred to.
3. Results
3.1. Confirmation of absence of MAOA in Tg8 mice
MAO reactivity was compared between normal C3Hline and transgenic Tg8 line at the level of the LC and
medullary A1 regions, in which noradrenergic neuronsshow a strong MAOA activity in wild-type mice. ManyMAO-containing neurons and fibers were observed in the
Ž .LC Fig. 1A,a and A1 neurons of C3H mice. Stained LCŽneurons were round or triangular and medium-sized 15–20
.mm in diameter . In contrast, we could not detect anyŽMAO activity in the LC and A1 neurons of Tg8 mice Fig.
.1B,b and Fig. 5E,F . After pharmacological MAOA sup-Ž y7 y5 .pression by clorgyline 10 –10 mol , no MAOA activ-
ity was detected in LC and A1 neurons of C3H mice, andthere was no difference in distribution of MAOB-positivestructures between Tg8 and C3H. All of the presumptiveMAOB-containing neurons and glial cells in Tg8 micedisappeared by deprenyl pretreatment at the concentrationmore than 10y5 mol.
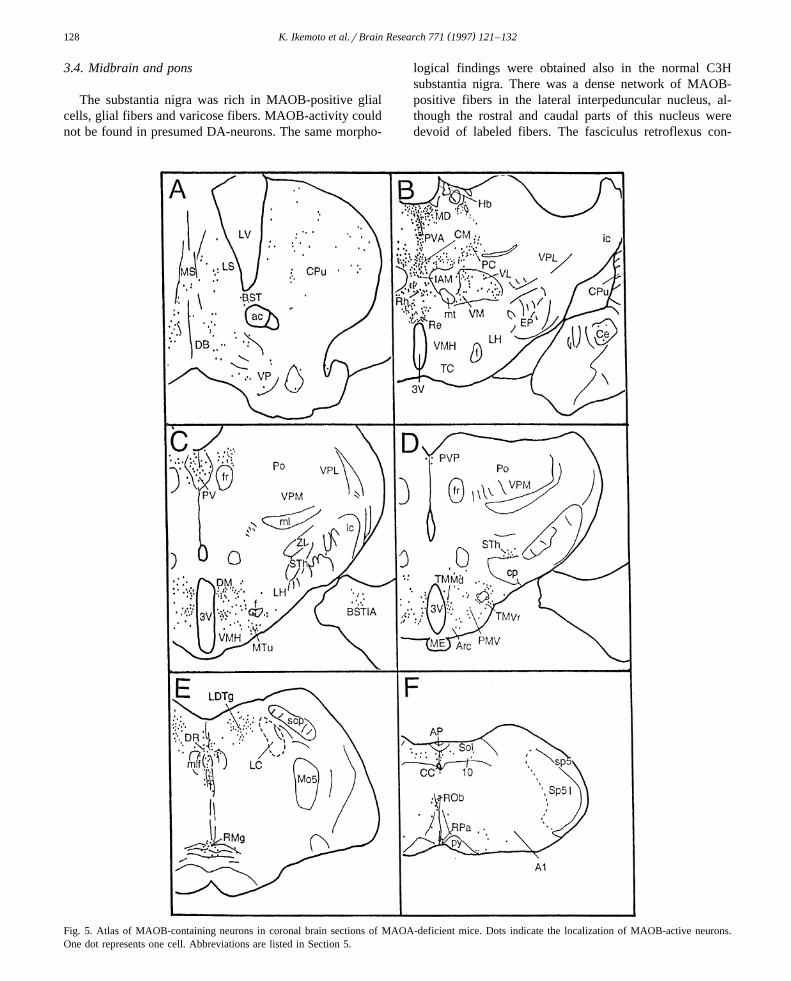
This evidence indicates that histochemically visualizedstructures in Tg8 line were exclusively MAOB-positiveand confirms the absence of MAOA in this mutant. Thismeans that MAO cytochemistry detects exclusivelyMAOB-containing structures in Tg8 line. The localizationof MAO-positive neurons in normal C3H and Tg8 ispresented in Table 1. The overall distribution of MAOB-active neurons in Tg8 is illustrated in a series of semis-chematic coronal sections displayed in rostrocaudal orderin Fig. 5.
3.2. Telencephalon
We observed many MAOB-positive fibers in the cere-Žbral cortex, especially cingulate and insular cortices Fig.
.2A,D and Fig. 3A,C . In the striatum, the fiber density washigh in the ventromedial part and lower in the dorsolateral
Ž . Ž .part Fig. 2A,D . Many small 10–15 mm in diameter - tomedium-sized MAOB-containing neurons were scattered
Ž Ž . .in the caudate putamen Fig. 2E arrows and Fig. 5A . Amoderate to large number of MAOB-containing neuronswere observed in the ventral striatum, bed nucleus of thestria terminalis, medial forebrain bundle, ventral pallidum,major island of Calleja, nucleus accumbens and olfactory
Ž .tubercle Fig. 2A–E and Fig. 5A . The shell part of thenucleus accumbens showed an extremely dense plexus inthe striatum. In a strongly MAOB-positive field of the
Ž .Fig. 3. A: the basolateral amygdaloid nucleus shows strong MAOB-reaction. The hippocampus Hi also contains MAO-B-positive cells in the CA1 regionand a few MAOB-active astrocytes throughout. There are many MAOB-positive cells and fibers in the anterior thalamus. In the anterior hypothalamus,
Ž .many MAOB-containing terminals are homogeneously distributed. B: in the anterior thalamus, the mediodorsal MD and paraventricular thalamic nucleiŽ .PVA show an especially intense MAOB reactivity. Many small MAOB-positive cells are masked by densely packed stained fibers. Numerous
Ž .MAOB-positive cells and fibers are aggregated also in the central medial thalamic nucleus CM . C: in the posterior hypothalamus, there are manyMAOB-positive neurons. The inner wall of the dorsal half of the mammillary recess of the third ventricle shows a strong MAOB activity. The choroid
Ž .plexus of the lateral ventricle demonstrates an intense MAOB activity small arrow . In the posterior thalamus, MAOB-positive cells are hardly observed.Ž .Instead, many MAOB positive terminals are seen. In the fasciculus retroflexus fr , a small amount of MAOB-positive fibers are homogeneously
Ž .distributed, but MAO-B cells are not found. D: MAOB-containing cells are observed in the posterior hypothalamus. Many large )20 mm in diametercells with long dendrite arbors, bipolar, triangular or multipolar in shape, are observed in the mediodorsal and ventrorostral parts of the tuberomammillary
Ž w x . Ž .nucleus TMMd, TMVr 7 , respectively . The dorsomedial nucleus DM contains also MAOB-positive large neurons. Note the presence of severalŽ . Ž .MAOB-positive small neurons 10–15 mm in diameter in the periventricular area medial to TMMd. In the arcuate nucleus Arc , MAOB-positive neurons
are scarcely seen. Scale bars: A, Cs1 mm; for B, Ds200 mm. Abbreviations are listed in Section 5.

( )K. Ikemoto et al.rBrain Research 771 1997 121–132 125
( )K. Ikemoto et al.rBrain Research 771 1997 121–132126
striatum, glial cell bodies were distinguished. In the lateralseptum, small and very weakly stained neurons with very
Žshort processes were embedded in stained fibers Fig. 2A
.and Fig. 5A . Intensely labeled medium-sized neuronsŽwere distributed in the diagonal band Fig. 2B and Fig.
.5A . In the area ventral to the Meynert nucleus, medium-

( )K. Ikemoto et al.rBrain Research 771 1997 121–132 127
Ž .sized neurons spread horizontally not shown . The amyg-daloid nucleus, especially the central subnucleus, includedMAOB-positive neurons, which were embedded in strongly
Ž .stained fibers Fig. 5B . The basolateral subnucleus con-Ž .tained only a dense positive fiber plexus Fig. 3A . In
addition, there were weakly positive medium-sized neu-rons and sparse fibers in the intraamygdaloid division of
Ž .the bed nucleus of the stria terminalis Fig. 5C . In theŽ .hippocampus Fig. 3A , a few medium-sized MAOB-posi-
tive neurons were observed in the CA1 region, andMAOB-active astrocytes were sparsely located throughoutthe hippocampal area. In the ependymal cells of the lateralventricle, an intense MAOB reactivity was obtained in a
Ž .homogeneous manner Fig. 2A,D . We noted a MAOB-ac-tive fiber network in the pineal body. The choroid plexusof the lateral ventricle demonstrated an intense MAOB
Ž .activity indicated by a small arrow in Fig. 3A,C .
3.3. Diencephalon
In the anterior part, the mediodorsal and paraventricularthalamic nuclei showed an especially strong MAOB reac-
Ž .tivity Fig. 3A,B . A large number of small MAOB-posi-tive neurons were masked by densely packed labeled fibersŽ .Fig. 3A,B and Fig. 5B . Numerous MAOB-positive neu-rons and fibers were aggregated also in the central medial,
Žrhomboid, reuniens and ventrolateral thalamic nuclei Fig..3A,B and Fig. 5B . A few stained neurons and fibers were
Ž .seen in the habenular nucleus Fig. 3A,B and Fig. 5Bextending to the posterior portion of the paraventricular
Ž .thalamic nucleus Fig. 5D .Caudally, we noted MAOB-positive neurons in the
Ž .posterior part of the subthalamic nucleus Fig. 5C,D .In the preoptic nucleus, many well-stained small neu-
Ž .rons were observed Fig. 2F . Terminals of MAOB neu-rons were homogeneously seen in the anterior hypothala-mic area. MAOB-containing cells were observed in the
Ž .posterior area of the hypothalamus Fig. 5C,D . ManyŽ .large )20 mm in diameter neurons having long dendrite
arbors which were bipolar, triangular or multipolar inshape, were seen throughout the ventral area of the hypo-thalamus, especially, in the mediodorsal and ventrorostral
w x Žsubnuclei of the tuberomammillary nucleus 7 Fig. 3D.and Fig. 5D . Small weakly stained MAOB neurons are
Ž .clustered in the premammillary nucleus Fig. 5D . Theinner wall of the dorsal half of the mammillary recess was
Ž .strongly MAOB-active Fig. 3C,D .
Table 1The localization and quantity of monoamine oxidase-positive neurons inthe brains of normal C3H mice and mutant Tg8 line
C3H Tg8
Telencephaloncerebral cortex 0 0caudate–putamen qq qqnucleus accumbens qq qqolfactory tubercle q qbed nucleus of stria terminalis q qintraamygdaloid division qq qq
medial forebrain bundle qq qqventral pallidum qq qqmajor island of Calleja qq qqlateral septum qq qqhippocampus CA1 qq qqdentate gyrus qq qq
central amygdaloid nucleus q q
Diencephalonparaventricular thalamic nucleus qqq qqqcentral medial thalamic nucleus qqq qqqmediodorsal thalamic nucleus qqq qqrhomboid thalamic nucleus qq qqreuniens thalamic nucleus qq qqventrolateral thalamic nucleus qq qhabenular nucleus q qsubthalamic nucleus q qtuberomammillary nucleus qqq qqqmedial preoptic nucleus qqq qqqlateral preoptic nucleus qq qdorsomedial hypothalamic nucleus qq qqarcuate hypothalamic nucleus q 0premammillary nucleus qq qq
Midbrain and ponssubstantia nigra 0 0interpeduncular nucleus qq 0fascicular retroflexus 0 0dorsal raphe nucleus qqq qqqraphe centralis qq qqpontis qq qqmagnus qq qq
laterodorsal tegmental nucleus qq qqlocus coeruleus nucleus qqq 0
Medulla oblongataraphe obscurus qq qqpallidus qq qq
nucleus solitary tract qq qqdorsal motor nucleus vagus q qA1 noradrenaline cells qq 0
0, absence of stained neurons; q, a small number of labeled neurons;qq, a moderate number of labeled neurons; qqq, a large number oflabeled neurons.
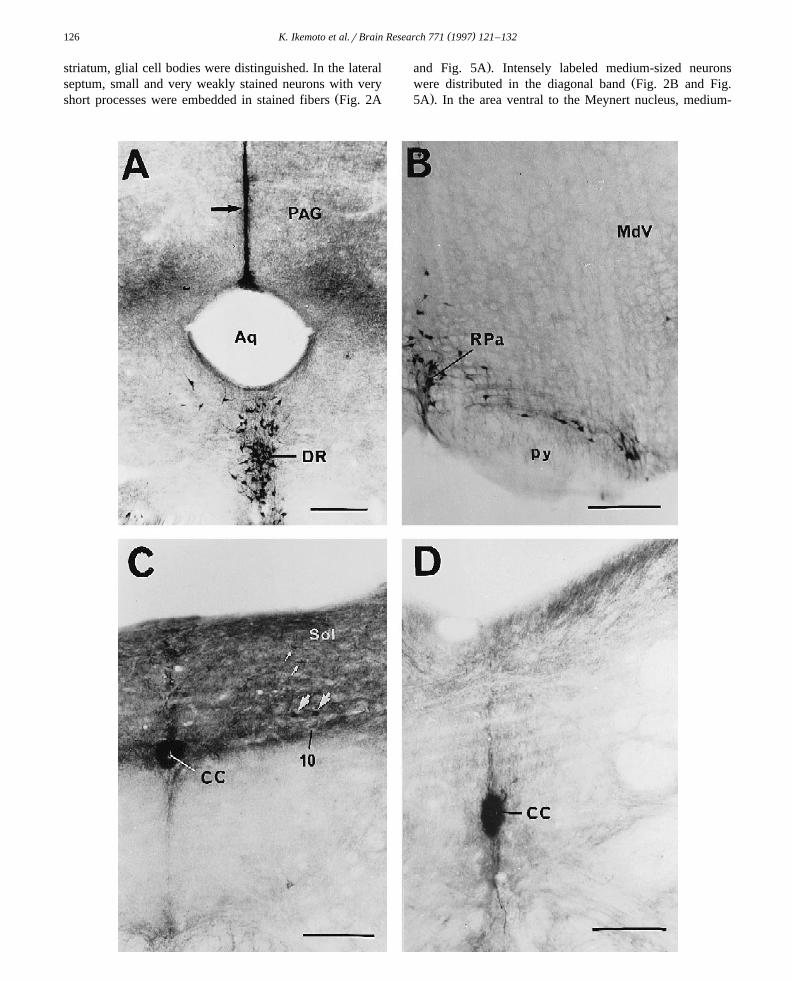
Ž .Fig. 4. A: a vertically extended linear structure with an intense MAOB activity is observed from the dorsal aspect of the aqueduct Aq into the midline ofŽ .the periaqueductal gray PAG . This structure is composed of glial cells. Inner wall of the Aq is also MAOB-positive. MAOB-containing cells in the DR
Ž .are strongly stained. B: the raphe pallidus nucleus RPa in the medulla oblongata contains MAOB-active cells. C: the dorsomedial portion of the medullaoblongata is heavily innervated by MAOB-positive fibers. In this MAOB-positive field, a small number of medium-sized MAOB-positive neurons are seen
Ž . Ž . Ž . Ž .in the nucleus solitary tract Sol small white arrows . A few large cells are embedded in the dorsal motor nucleus vagus 10 large white arrows . IntenseŽ .MAOB activity was observed in the inner wall of the central canal CC . D: inner wall of the CC is intensely MAOB-positive also in the lower medulla
oblongata. Scale bars200 mm. Abbreviations are listed in Section 5.
( )K. Ikemoto et al.rBrain Research 771 1997 121–132128
3.4. Midbrain and pons
The substantia nigra was rich in MAOB-positive glialcells, glial fibers and varicose fibers. MAOB-activity couldnot be found in presumed DA-neurons. The same morpho-
logical findings were obtained also in the normal C3Hsubstantia nigra. There was a dense network of MAOB-positive fibers in the lateral interpeduncular nucleus, al-though the rostral and caudal parts of this nucleus weredevoid of labeled fibers. The fasciculus retroflexus con-
Fig. 5. Atlas of MAOB-containing neurons in coronal brain sections of MAOA-deficient mice. Dots indicate the localization of MAOB-active neurons.One dot represents one cell. Abbreviations are listed in Section 5.

( )K. Ikemoto et al.rBrain Research 771 1997 121–132 129
Ž .tained a small amount of stained fibers Fig. 3C . Anintense MAOB activity was observed in glial cells of thevertically extended linear structure extending from thedorsal aspect of the aqueduct into the midline of the
Ž .periaqueductal gray Fig. 4A . Ependymal cells of theŽinner wall of the aqueduct were also MAOB-active Fig.
.4A .The dorsal raphe nucleus contained many intensely
stained neurons, which are bipolar or fusiform in shape,Ž .and medium-sized Fig. 4A . There were also MAOB-posi-
tive neurons in the raphe centralis, pontis and magnusŽ .nuclei Fig. 5E . We found many MAO-positive neurons
Ž .in the laterodorsal tegmental nucleus LDTg in Tg8 miceŽ .Fig. 1B and Fig. 5E and in normal C3H mice as wellŽ .Fig. 1A , an observation not previously reported. Stainedcells of LDTg were ovoid, triangular or multipolar inshape and medium-sized. The degree of MAO reactivity of
Žthese neurons was of two types, weak or intense Fig..1A,B .
3.5. Medulla oblongata
The raphe obscurus and raphe pallidus nuclei containedŽ .MAOB-active cells Fig. 4B and Fig. 5F . In the rostral
dorsomedial part, the nucleus of the solitary tract wasŽ .heavily innervated by MAOB-positive fibers Fig. 4C , in
which a small number of medium-sized MAOB-positiveŽ .neurons were embedded Fig. 4C and Fig. 5F . A few large
neurons were masked in packed stained fibers present inŽ .the dorsal motor nucleus vagus Fig. 4C and Fig. 5F .
Intense MAOB activity was observed in ependymal cellslying in the wall of the central canal throughout the
Ž .medulla oblongata Fig. 4C,D .
4. Discussion
4.1. Absence of MAOA in Tg8 mouse
In MAOA-deficient transgenic Tg8 line, we confirmedhistochemically the absence of MAOA enzymatic activity,using pharmacological suppression. Absence of MAO-ac-tivity in Tg8 noradrenergic neurons and fibers is its an-other evidence. Therefore, this mutant enables us to studyexclusively MAOB-positive structures, even in the areasuch as the dorsal vagal complex, where both types ofMAO are rich in normal mice. It should be noted that evenafter pharmacological pretreatment, MAOB-positive struc-tures are not always completely distinguishable from verystrong MAOA-positive ones in wild-type strains as re-
w xported in normal rats 2 .
4.2. MAOB neuronal cell distribution in the mouse: speciesdifferences
In the present study, we report the distribution ofMAOB-active structures in normal C3H mice and in its
mutant Tg8 line. Numerous MAOB neurons were found,as in other species, in structures such as the raphe nucleiand tuberomammillary hypothalamic region. Furthermore,in the present, we could note several species differenceswhen compared with the rat, cat and primates.
The major species difference is the presence of MAOBneurons in the mouse basal forebrain, especially in thestriatum, nucleus accumbens, nucleus of the diagonal band,ventral pallidum and preoptic area, as first shown by
w xNakamura et al. 22 . These authors found that MAOBneurons in the striatum correspond to a subpopulation ofcholinergic neurons, that do not contain NADPH-di-
w xaphorase nor GAD 21 . In the mouse brain, we foundweakly stained septal MAOB-positive neurons that Naka-
w xmura et al. 22 failed to demonstrate in normal mice. Thismay be due to the difference in sensitivity of the methodsused.
The present study also demonstrated for the first timethe presence of MAOB-positive neurons in the LDTg anddorsal vagal complex. Such neurons are not detectable inthe rat, cat, or primates. The distribution pattern of LDTg
w xneurons is very similar to that of cholinergic neurons 23 ,and in our preliminary study, some of them are in factcholinergic, and may also contain nitric oxide. LDTg cells
w xare known to project to the thalamus 24 . The functions ofthese MAO-cholinergic neurons are at present unknown.
Some new small species differences in neuronal celldistribution were noted in the thalamic nuclei and hypotha-lamic tuberomammillary region. Thalamic MAOB-activeneurons extend not only in the midline but also in morelaterally located nuclei such as the ventromedial and ven-trolateral thalamic nuclei in the mouse, whereas in the rat,
w xthey are restricted to the midline 1 . The same restrictionhas been reported using in situ hybridization and enzyme
w xradioautography in the rat 18,25 . In the cat, only weaklystained MAO-positive neurons are reported in the paraven-
w xtricular thalamic nucleus 10 .In Tg8 mice, large MAOB neurons were widely dis-
tributed through the entire extent of the ventral posteriorhypothalamic area as reported by Vincent in C57 black
w xmice 27 , whereas in the rat, most of them are restricted toa small region; ventrolaterally situated tuberomammillary
w x w xnucleus 1 . These neurons may contain histamine 16,17w xand many bioactive substances 12 .
Concerning MAOB fiber distribution, we noted denseplexi in the midline of the anterior thalamus, the nucleusaccumbens, shell, and the dorsal vagal complex. Such adensity has not been reported in other species. As we havenot observed such dense 5-HT or histamine plexi, theseplexi may be derived from a large number of small intrin-sic MAOB neurons in these areas. It is noteworthy thatMAOB activity is high in the limbic area, including thecingulate cortex, amygdaloid nucleus and nucleus accum-bens. In this study, a small number of positive neuronswere found in the amygdaloid nucleus and hippocampus.These results suggest that MAOB is more involved in

( )K. Ikemoto et al.rBrain Research 771 1997 121–132130
limbic functions than was believed. In comparison with therat and cat striatum, there were much more MAOB-posi-tive fibers in the mouse. Striatal dopamine should beinactivated by MAOB in this area in Tg8.
The presence of MAOB in glial cells has been reportedw xpreviously 11,19,29 . A vertical plate extending in the
dorsal midline of the periaqueductal gray composed ofMAOB astrocytes has not been described previously in thecat and in the rat. Using Tg8 mice, we confirmed theexistence of MAOB glial cells in the mouse hippocampusand cerebral cortex, which has been mentioned also in the
w xcat 11 . In the Tg8 striatum, we could distinguish MAOBglial cell bodies in a strongly MAOB-positive field. In the
Žrat striatum, there are many MAO-positive astrocytes un-.published data . On the other hand, in the cat, MAOB-
w xpositive glial cells are rare in the striatum 11 . The speciesdifference is evident.
MAOB-activity in mouse ependymal cells along thelateral ventricle is less intense than that reported in the ratw x1,10,11,5,18,25 . Very strong MAOB-active cells lyingalong the surface of the central canal and those in thechoroid plexus are likely to regulate excessive monoaminesin the cerebrospinal fluid.
4.3. Physiological implications
The present study confirmed histochemically the ab-sence of MAOA activity in Tg8 mutant mice. Evidence ofMAOA deficiency in Tg8 mice had been provided by thepresence of extremely high levels of 5-HT in pup’s brainŽ9-fold to 6-fold excess at postnatal day 1 and 12, respec-
.tively and the abnormal accumulation of 5-HT in pup’snoradrenergic neurons of the LC and dopaminergic neu-
w xrons of the substantia nigra and ventral tegmental area 5 .In contrast, in Tg8 adults, 5-HT excess in total brain doesnot exceed twofold, and 5-HT is undetectable in NA andDA neurons. Accordingly, the MAO metabolite of 5-HT,
Ž .5-hydroxyindoleacetic acid 5-HIAA , is present at normallevels in Tg8 adults and at much reduced levels in Tg8
w xpups 5 . In the rodent brain, MAOB activity is low at birthw xand increases during the first month 13,26 . In the present
study, we could not detect MAOA-positive structures inTg8. Based upon biochemical experiments, 5-HT is known
w xto be degraded by MAOA 20 . Therefore, in Tg8, 5-HTmust be degraded by another enzyme, possibly by MAOB.This mechanism should be elucidated in future.
Recently a family has been reported which lacks MAOAcatalytic activity due to a point mutation in the gene
w xencoding MAOA and shows impulsive aggression 4 .MAOA-deficient mutants, Tg8 mice, are also aggressive,and are studied as a good model for abnormal behavior in
w xthis family. In fact, recently Case et al. 5,6 have reportedthe aggressive behavior of Tg8.
Further studies using Tg8 mice will enable us to exam-ine more precisely the ultrastructural organization of
MAOB-containing structures in the central nervous systemwith the aid of electron microscopy.
5. Abbreviations used in Figures
-active enzymatically active3V third ventricle10 dorsal motor nucleus vagusA1 A1 noradrenaline cellsac anterior commissureAP area postremaAq aqueductArc arcuate hypothalamic nucleusBL basolateral amygdaloid nucleusBST bed nucleus of the stria terminalisBSTIA bed nucleus of the stria terminalis, intraamyg-
daloid divisioncc corpus callosumCC central canalCe central amygdaloid nucleusCg cingulate cortexCM central medial thalamic nucleuscp cerebral peduncleCPu caudate putamenDB diagonal bandDM dorsomedial hypothalamic nucleusDR dorsal raphe nucleusEP entopeduncular nucleusf fornixfr fasciculus retroflexusHb habenular nucleusHi hippocampusI insular cortexIAM interanteromedial thalamic nucleusic internal capsuleLC locus coeruleusLDTg laterodorsal tegmental nucleusLH lateral hypothalamic areaLPO lateral preoptic areaLS lateral septal nucleusLV lateral ventricleMD mediodorsal thalamic nucleusMdV medullary reticular nucleus, ventralME median eminenceml medial lemniscusmlf medial longitudinal fasciculusMo5 motor trigeminal nucleusMPO medial preoptic nucleusMS medial septal nucleusmt mammillothalamic tractMTu medial tuberal nucleusPAG periaqueductal grayPC paracentral thalamic nucleusPMV premammillary nucleus, ventralPo posterior thalamic nuclear groupPV paraventricular thalamic nucleusPVA paraventricular thalamic nucleus, anterior

( )K. Ikemoto et al.rBrain Research 771 1997 121–132 131
PVP paraventricular thalamic nucleus, posteriorpy pyramidal tractRe reuniens thalamic nucleusRh rhomboid thalamic nucleusRMg raphe magnus nucleusROb raphe obscurus nucleusRPa raphe pallidus nucleusscp superior cerebellar peduncleSol nucleus solitary tractsp5 spinal trigeminal tractSp5I spinal trigeminal nucleus, interpolarSTh subthalamic nucleusTC tuber cinereum areaTMMd mediodorsal part of the tuberomammillary nu-
cleusTMVr ventrorostral part of the tuberomammillary nu-
cleusVL ventrolateral thalamic nucleusVM ventromedial thalamic nucleusVMH ventromedial hypothalamic nucleusVP ventral pallidumVPL ventral posterolateral thalamic nucleusVPM ventral posteromedial thalamic nucleusZI zona incerta
Acknowledgements
A part of this work was presented at the Society forNeuroscience in Washington in November, 1996. Thisstudy was supported by CNRS ERS 5645, INSERM U52,
Žthe Institut Curie, and CIES Centre International des.Etudiants et Stagiaires .
References
w x1 R. Arai, H. Kimura, T. Maeda, Topographic atlas of monoamineoxidase-containing neurons in the rat brain studied by an improved
Ž .histochemical method, Neuroscience 19 1986 905–925.w x2 R. Arai, H. Kimura, I. Nagatsu, T. Maeda, Preferential localization
of monoamine oxidase type A activity in neurons of the locuscoeruleus and type B activity in neurons of the dorsal raphe nucleusof the rat: a detailed enzyme histochemical study, Brain Res. 745Ž .1997 352–356.
w x3 A.W. Bach, L. Lan, D.L. Johnson, G.W. Abell, M.E. Bembenek,S.W. Kwan, P.H. Seeburg, J.C. Shih, cDNA cloning of human livermonoamine oxidase A and B: Molecular basis of differences in
Ž .enzymatic properties, Proc. Natl. Acad. Sci. USA 85 1988 4934–4938.
w x4 H.G. Brunner, M. Nelen, X.O. Breakefield, H.H. Ropers, B.A. VanOost, Abnormal behavior associated with a point mutation in the
Ž .structural gene for monoamine oxidase A, Science 262 1993578–580.
w x5 O. Cases, I. Seif, J. Grimsby, P. Gaspar, K. Chen, S. Pournin, U.Muller, M. Aguet, C. Babinet, J.C. Shih, E. De Maeyer, Aggressive¨behavior and altered amounts of brain serotonin and norepinephrine
Ž .in mice lacking MAOA, Science 268 1995 1763–1766.w x6 O. Cases, T. Vitalis, I. Seif, E. De Maeyer, C. Sotelo, P. Gaspar,
Lack of barrels in the somatosensory cortex of monoamine oxidaseA-deficient mice: role of a serotonin excess during the critical
Ž .period, Neuron 16 1996 297–307.w x7 H. Ericson, T. Watanabe, C. Kohler, Morphological analysis of the¨
tuberomammillary nucleus in the rat brain: Delineation of subgroupswith antibody against L-histidine decarboxylase as a marker, J.
Ž .Comp. Neurol. 263 1987 1–24.w x8 K.B.J. Franklin, G. Paxinos, The Mouse Brain in Stereotaxic Coor-
dinates, Academic Press, New York, 1997.w x9 K. Kitahama, R. Arai, T. Maeda, M. Jouvet, Demonstration of
monoamine oxidase type B in serotonergic and type A in noradren-ergic neurons in the cat dorsal pontine tegmentum by an improved
Ž .histochemical technique, Neurosci. Lett. 71 1986 19–24.w x10 K. Kitahama, R.M. Denney, T. Maeda, M. Jouvet, Distribution of
type B monoamine oxidase immunoreactivity in the cat brain withŽ .reference to enzyme histochemistry, Neuroscience 44 1991 185–
204.w x11 K. Kitahama, T. Maeda, R.M. Denney, M. Jouvet, Monoamine
oxidase: distribution in the cat brain studied by enzyme- and im-Ž .munohistochemistry: Recent progress, Prog. Neurobiol. 42 1994
53–78.w x12 C. Kohler, H. Ericson, T. Watanabe, J. Polak, S.L. Palay, V. Palay,¨
V. Chan-Palay, Galanin-immunoreactivity in hypothalamic his-tamine neurons: further evidence for multiple chemical messengers
Ž .in the tuberomammillary nucleus, J. Comp. Neurol. 250 198658–64.
w x13 Y. Koide, K. Kobayashi, Developmental changes in the activity andsubstrate specificities of mouse brain monoamine oxidase, Neu-
Ž .rochem. Res. 9 1984 595–606.w x14 C.H. Konradi, E. Svoma, K. Jellinger, P. Riederer, R. Denney, J.
Thibault, Topographic immunocytochemical mapping of monoamineoxidase-A, monoamine oxidase-B and tyrosine hydroxylase in hu-
Ž .man post mortem brain stem, Neuroscience 26 1988 791–802.w x15 P. Levitt, J.E. Pintar, X.O. Breakefield, Immunohistochemical
demonstration of monoamine oxidase B in brain astrocytes andŽ .serotonergic neurons, Proc. Natl. Acad. Sci. USA 79 1982 6385–
6389.w x16 J.S. Lin, P. Fort, K. Kitahama, P. Panula, R.M. Denney, M. Jouvet,
Immunohistochemical evidence for the presence of Type Bmonoamine oxidase in histamine containing neurons in the posterior
Ž .hypothalamus of cats, Neurosci. Lett. 128 1991 61–65.w x17 J.S. Lin, K. Kitahama, P. Fort, P. Panula, R.M. Denney, M. Jouvet,
Histaminergic system in the cat hypothalamus with reference to typeŽ .B monoamine oxidase, J. Comp. Neurol. 330 1993 405–420.
w x18 J.M. Luque, S.W. Kwan, C.W. Abell, M. Da Prada, G. Richards,Cellular expression of mRNAs encoding monoamine oxidases A and
Ž .B in the rat central nervous system, J. Comp. Neurol. 363 1995665–680.
w x19 T. Maeda, H. Imai, R. Arai, H. Tago, T. Nagai, T. Sakumoto, K.Kitahama, B. Onteniente, H. Kimura, An improved peroxidaticoxidation method of MAO histochemistry for neuroanatomical re-search at light and electron microscope levels, Cell Mol. Biol. 33Ž .1987 1–11.
w x20 D.L. Murphy, Substrate-selective monoamine oxidases, Biochem.Ž .Pharmacol. 27 1996 1889–1893.
w x21 S. Nakamura, I. Akiguchi, J. Kimura, A subpopulation of mousestriatal cholinergic neurons show monoamine oxidase activity, Neu-
Ž .rosci. Lett. 161 1993 141–144.w x22 S. Nakamura, I. Akiguchi, J. Kimura, Topographic distributions of
monoamine oxidase-B-containing neurons in the mouse striatum,Ž .Neurosci. Lett. 184 1995 29–31.
w x23 K. Satoh, D.M. Armstrong, H.C. Fibiger, A comparison of thedistribution of central cholinergic neurons as demonstrated byacetylcholinesterase pharmacohistochemistry and choline acetyl-
Ž .transferase immunohistochemistry, Brain Res. Bull. 11 1983 693–720.
w x24 K. Satoh, H.C. Fibiger, Cholinergic neurons of the laterodorsal

( )K. Ikemoto et al.rBrain Research 771 1997 121–132132
tegmental nucleus: efferent and afferent connections, J. Comp. Neu-Ž .rol. 253 1986 277–302.
w x25 J. Saura, R. Kettler, M. Da Prada, J.G. Richards, Quantitativeenzyme radioautography with 3H-Ro 41-1049 and 3H-Ro 19-6327 invitro: Localization and abundance of MAO-A and MAO-B in rat
Ž .CNS, peripheral organs, and human brain, J. Neurosci. 12 19921977–1999.
w x26 D. Tsang, K.P. Ho, H.L. Wen, Ontogenesis of multiple forms ofmonoamine oxidase in rat brain regions and liver, Dev. Neurosci. 8Ž .1986 243–250.
w x27 S.R. Vincent, Histochemical localization of 1-methyl-4-phenyl-1,2,3,6-tetrahydropyridine oxidation in the mouse brain, Neuro-
Ž .science 28 1989 189–199.w x28 K.N. Westlund, R.M. Denney, L.M. Kochersperger, R.M. Rose,
C.W. Abell, Distinct monoamine oxidase A and B populations inŽ .primate brain, Science 230 1985 181–183.
w x29 K.N. Westlund, R.M. Denney, R.M. Rose, C.W. Abell, Localizationof distinct monoamine oxidase A and monoamine oxidase B cell
Ž .populations in human brainstem, Neuroscience 25 1988 439–456.