morphology and anatomy of stems and pedicels - annals of botany
TRANSCRIPT

Annals of Botany 77 : 537–545, 1996
Morphology and Anatomy of Stems and Pedicels of Spring Flush Shoots
Associated with Citrus Fruit Set
YAIR ERNER* and ILAN SHOMER†
Departments of *Citriculture and †Food Science, Agricultural Research Organization, The Volcani Center,
P.O. Box 6, Bet Dagan 50250, Israel
Received: 15 August 1995 Accepted: 10 November 1995
Flowering and vegetative shoots of ‘Shamouti ’ orange [Citrus sinensis (L.) Osbeck] and ‘Marsh’ seedless grapefruit(Citrus paradisi Macf.) were examined for correlation of their morphology and anatomy with fruit set. Fruit set isfavoured on leafy inflorescences whereas abortion is nearly complete on leafless inflorescences. Leafless inflorescencesof ‘Shamouti ’ with one flower were found to have a very thin stem which contained few vascular bundles, whereasthose with three flowers had better-developed vascular systems. The vascular system of leafy inflorescences issignificantly different from that of leafless ones and contains a distinct central xylem cylinder. The vascular area ofleafless inflorescences is only about one-quarter of that of the leafy ones. The vascular system of grapefruit resemblesthat of the ‘Shamouti ’ orange. This study emphasizes the importance of the dimension of the vascular system for fruitset and provides a possible explanation for the better fruit set on both leafy and leafless inflorescences with severalflowers compared with single-flowered inflorescences. #1996 Annals of Botany Company
Key words : Anatomy, citrus, fruit set, leafless inflorescence, leafy inflorescence, pedicel, vascular system, vegetativeshoot.
INTRODUCTION
Fruit set is favoured on leafy inflorescences i.e. shootsbearing leaves and flowers, (LY), whereas abortion ofreproductive organs is nearly total on leafless inflorescencesbearing only flowers, (LS), during the first 2 months afteranthesis (Reece, 1945; Sauer, 1951; Moss, 1970; Moss,Steer and Kriedemann, 1972; Erner, 1989; Davenport,1990). These observations were found to be inconsistentwith regard to fruit set on LS in grapefruit, lemon andkumquat, in which up to 50% of the harvested fruits wereborne on LS (M. Fishler and A. Cohen, pers. comm.).
The preferential set on LY has been explained mainly interms of the supply of photosynthate to the developing fruitfrom the adjacent young leaves, an optimal supply beingneeded to enhance fruit set (Moss, 1970; Moss et al., 1972).
Although fruit set has been positively correlated withcarbohydrate in leaves of the previous season (Ohgaki,Fujita and Ito, 1963; Goldschmidt and Golomb, 1982;Monselise and Goldschmidt, 1982; Goldschmidt et al.,1985; Davenport, 1990), no differences in carbohydratecontent could be found between fruitlets born either on LYor on LS. Both fruitlet types were strong metabolic sinksand competed with other plant parts for carbohydrate(Erner, 1989). Moreover, Fishler, Goldschmidt andMonselise (1983) have shown that the sink power ofgrapefruit may extend great distances in the tree and thatthe fruit can draw its assimilate requirement for developmenteven from a distance of 2 m. Endogenous GA and IAAconcentrations were observed to be similar in these twotypes of fruitlets. On the other hand, ABA concentration
was found to be significantly higher in fruitlets borne on LSthan in those borne on LY up to 7 d after flower opening,and remained higher over the period of measurements(Erner, 1989).
Since ABA has been found to be involved in acceleratingabscission in citrus explants (Sagee, Goren and Riov, 1980)it has been proposed that even non-detectable transientwater stress, especially in single flower LS inflorescences,around midday can cause ABA build-up, which willsubsequently lead to abscission (Erner, 1989; Sagee andErner, 1991). Based on the above, it has been suggested thatfruitlets borne on LY would set fruit which would be morelikely to survive until harvest, owing to better managementof the water status. Leaves have been known to induce andcontrol the development of vascular tissues along the plantaxis by means of a steady polar flow of auxins (Wareing andPhillips, 1978; Sachs, 1981; Aloni, 1987, 1992).
The vascular tissue of the shoot facilitates efficienttransport between the roots to the leaves and}or fruitlet(Jahn, 1978). Thus, we expected that LS would have lessvascular tissue than LY, so that the water transport to theyoung LS-borne fruitlets would be of slower rate than thatto LY-borne fruitlets.
The study described here examined the anatomy of theshoot cross-section at the base and the middle of the pediceland its correlation with the morphology of spring flushshoots [LS, LY, and vegetative shoots (V)]. The nomen-clature of the pedicel mentioned here is the one adopted byDavenport (1990). The possible involvement of the vascularsystem in prevention of fruitlet abscission is demonstratedand discussed.
0305-7364}96}11053709 $25.00}0 # 1996 Annals of Botany Company
Dow
nloaded from https://academ
ic.oup.com/aob/article/78/5/537/2587504 by guest on 01 February 2022

538 Erner and Shomer—Morphology and Anatomy of Stems and Pedicels
MATERIALS AND METHODS
Plant Material
Thirty-year-old ‘Shamouti ’ orange [Citrus sinensis (L.)Osbeck] and ‘Marsh’ seedless grapefruit (Citrus paradisiMacf.), both grafted on sour orange [Citrus aurantium (L.)Osbeck] were used for the experiments. Trees were grownunder optimal citrus-growing conditions on sandy loam soilin the central coastal belt of Israel.
Morphology measurements
Spring-flush shoots were classified into three principaltypes according to Goldschmidt and Monselise (1972) : (a)LS, bearing only one to several flowers, (b) LY, bearingboth leaves and flowers (ranging from one terminal flowerwith one or several leaves to several flowers and leaves) and(c) V, bearing only leaves (normally more than three and upto ten leaves, depending on the species and conditions). In50 samples of each of the various spring-flush shoot types,the diameters were measured when the terminal flowerreached full size just prior to opening, with digital calliperswith 0±01-mm accuracy. Diameter measurements were made5 mm from the shoot base and at mid-pedicel (except for Vshoots, where only the basal measurements could be taken).
Light microscopy
Four types of inflorescence were examined: two leaflessand two leafy, as described in Fig. 33. In addition,vegetative shoots (V, no flowers) were examined. Theinflorescences were sampled when the first petal of the topflower started to open, and the second sampling was taken3 weeks after petal fall. From each of the examinedbranches eight separate tree-samples (1±5 m above groundfrom the south side of the tree, total of 5¬8 for eachvariety) of 2-mm cross-sections were taken for anatomyobservation, more than 5 mm above the shoot base, andfrom the middle of the pedicel.
Sections were fixed in 5% (v}v) glutaraldehyde in 0±1
cacodylate buffer (pH 7) at 4 °C for 2 h. The fixed sectionswere then rinsed several times with the same buffer, washedwith 0±1 phosphate buffer (pH 7), post fixed with 2%(w}v) OsO
%in the phosphate buffer at 4 °C for 2 h,
washed several times with the phosphate buffer and, finally,washed with distilled water. The fixed sections weredehydrated serially with ethanol and embedded in Agar 100Resin (Agar Aid, Cambridge). Cross-sections of 3 µmthickness were prepared by means of a Pyramitome (LKB),stained with solution of toluidine blue and basic fuchsin(0±365 and 0±135 g, respectively, in 60 ml of 30% (v}v)ethyl alcohol, diluted¬10) and examined under immersionoil. Each shoot type was examined in five replicates. Inaddition, the sections were examined in polarized light inorder to verify the identification of the xylem vessels. Majorxylem vessels as well as the total xylem cylinder weremeasured from cross section.
All of the shoot types, from both ‘Shamouti ’ orange and‘Marsh’ grapefruit, were examined microscopically for their
anatomy. Some representative light micrographs are pre-sented here.
RESULTS AND DISCUSSION
The importance of leaves in the inflorescences for fruit set iswell known and has been reported for many varieties andspecies of citrus by a number of investigators (Sauer, 1951;Moss, 1970; Jahn, 1973; Erner and Bravdo, 1983). Thecorrelation found between carbohydrate and fruit-set hasbeen claimed to be the major factor in the enhancement offruit set (Moss, 1970; Goldschmidt and Golomb, 1982).However, Erner (1989), working with LS and LYinflorescences, up to 2 months after anthesis, could notconfirm any significant differences in set which could beattributed to carbohydrate content. Moreover, the results ofcarbohydrate manipulation by means of girdling and GA3also negate the carbohydrate theory (Goldschmidt et al.,1985). Our hypothesis that the conductance of water intothe inflorescence would be higher in LY than in LS led to aneed to clarify the morphology and anatomy of theseinflorescences. Citrus flowers develop mostly on one-year-old branches from the previous spring flush, and can bedivided into several morphological types (Goldschmidt andMonselise, 1972). The relationship between shoot-type in‘Shamouti ’ orange and the cross-sectional area of the stemand pedicel is shown in Fig. 31. The cross-sectional area ofthe stem base (Fig. 31A) of LS is significantly smaller thanthat found in shoots bearing a number of flowers and leaves(LY). LS with one flower formed the majority of shoots on‘Shamouti ’ trees and had a smaller cross-sectional areathan LS shoots bearing more flowers. Moreover, the biggestcross-sectional area of LS stem base (five flowers) was foundto be smaller than that of a vegetative branch (withoutflowers) with four leaves or LY with one flower and threeleaves.
In ‘Shamouti ’ orange the number of flowers on LS wasfound to be strongly positively correlated (r¯ 0±9535) withthe cross-sectional area at the base (Fig. 31A) and negativelycorrelated (r¯®0±9899) with that at the pedicel (Fig. 31B).The increase in the cross-sectional area of LS stem base wasmore than twofold, as the number of flowers increased fromone to five, while the decrease in the area in the pedicel wasabout 17% (Fig. 31). Vegetative spring shoots normally hadfour to 10 leaves ; the number of leaves positively correlated(r¯ 0±9717) with the cross-sectional area at the stem base.
The various combinations of LY with one to manyflowers, or of many leaves with several flowers, emphaticallyillustrate the importance of leaf number for the cross-sectional area at A (Fig. 31). No corresponding differencescould be found in the cross-section at the mid pedicel (B).Most of these various combinations of LY involved fewflowers and few leaves ; only a few combinations of manyflowers with many leaves were found.
Basically similar data were obtained for ‘Marsh’ grape-fruit (Fig. 32). Although the cross-sectional area at the stembase (Fig. 32A) and mid-pedicel (Fig. 32B) in grapefruitwere similar to those in ‘Shamouti ’, in grapefruit, up to50% fruit set has been found on LS (A. Cohen and M.Fishler pers. comm.).
Dow
nloaded from https://academ
ic.oup.com/aob/article/78/5/537/2587504 by guest on 01 February 2022

Erner and Shomer—Morphology and Anatomy of Stems and Pedicels 539
F 1–7. Cross sections of the stems (5 mm from the shoot base) of leafless inflorescences (LS) of ‘Shamouti ’ orange and ‘Marsh’ grapefruit.Figs 1 and 2. ‘Shamouti ’ LS with one flower (¬125 and ¬625, respectively) a typical vascular system with relatively wide cortex. Figs 3 and 4.‘Shamouti ’ LS with three flowers (¬625 and ¬125, respectively) the secondry system and phloem are more developed than that with one flower.Fig. 5. ‘Marsh’ LS with one flower (¬125) seems similar to that of ‘Shamouti ’. Figs 6 and 7. As Fig. 5 but viewed under polarized light (¬125
and ¬625). C, Cambium; Cx, cortex; P, phloem; Pt, pith; V, vessel element ; X, xylem.
Such high set on LS in grapefruit, compared with that in‘Shamouti ’, is due to the greater proportion of LS shootswith more than one flower. The percentages of LS with oneflower and more than one flower are 17 and 36%,respectively for grapefruit, and 59 and 15%, for ‘Shamouti ’,
respectively (Erner, unpubl. res.). Grapefruit LY had manyflowers together with many leaves (up to 10 flowers and sixleaves) while ‘Shamouti ’ had no more than six flowers withfive leaves.
Anatomical studies were made on four main inflorescence
Dow
nloaded from https://academ
ic.oup.com/aob/article/78/5/537/2587504 by guest on 01 February 2022
540 Erner and Shomer—Morphology and Anatomy of Stems and Pedicels
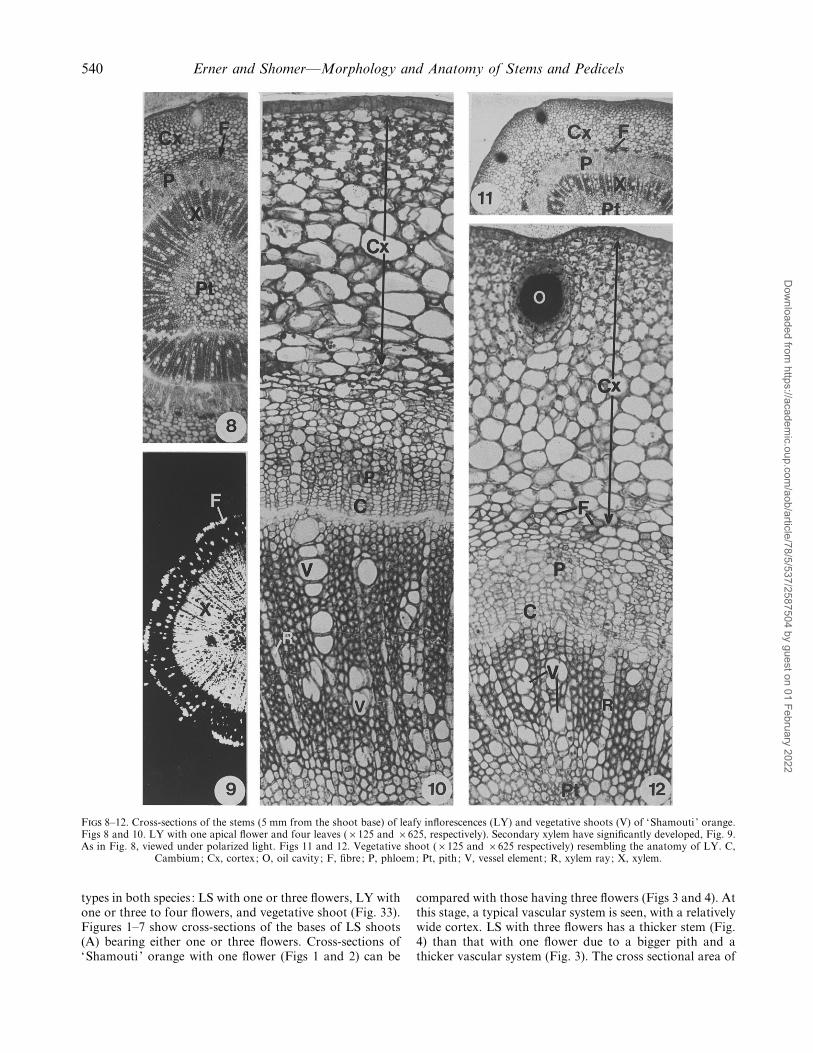
F 8–12. Cross-sections of the stems (5 mm from the shoot base) of leafy inflorescences (LY) and vegetative shoots (V) of ‘Shamouti ’ orange.Figs 8 and 10. LY with one apical flower and four leaves (¬125 and ¬625, respectively). Secondary xylem have significantly developed, Fig. 9.As in Fig. 8, viewed under polarized light. Figs 11 and 12. Vegetative shoot (¬125 and ¬625 respectively) resembling the anatomy of LY. C,
Cambium; Cx, cortex; O, oil cavity ; F, fibre; P, phloem; Pt, pith; V, vessel element ; R, xylem ray; X, xylem.
types in both species : LS with one or three flowers, LY withone or three to four flowers, and vegetative shoot (Fig. 33).Figures 1–7 show cross-sections of the bases of LS shoots(A) bearing either one or three flowers. Cross-sections of‘Shamouti ’ orange with one flower (Figs 1 and 2) can be
compared with those having three flowers (Figs 3 and 4). Atthis stage, a typical vascular system is seen, with a relativelywide cortex. LS with three flowers has a thicker stem (Fig.4) than that with one flower due to a bigger pith and athicker vascular system (Fig. 3). The cross sectional area of
Dow
nloaded from https://academ
ic.oup.com/aob/article/78/5/537/2587504 by guest on 01 February 2022

Erner and Shomer—Morphology and Anatomy of Stems and Pedicels 541
F 13–16. Cross-section of the pedicel from a ‘Shamouti ’ leafless inflorescence (LS) with three flowers (Figs 13 and 15, ¬125 and ¬625,respectively). The vascular system comprises separate bundles, irrespective of the flower number on the rest of the inflorescence. Figs 14 and 16.
As Figs 13 and 15 but viewed under polarized light.
F 17–20. Cross-section of the stem base (A) of ‘Marsh’ grapefruit leafless inflorescences (LS) with one flower (Figs 17–19) and with threeflowers (Fig. 20), 3 weeks after petal fall. The vascular system of LS with one fruitlet is composed of separate bundles, while that with three fruitletsappeared as a vascular cylinder. (Fig. 17, ¬125, Figs 19 and 20, ¬650). Fig. 18. As Fig. 17 but viewed under polarized light. C, Cambium; Cx,
cortex; O, oil cavity ; F, fibre; P, phloem; Pt, pith; VB, vascular bundle ; V, vessel element ; R, xylem ray.
Dow
nloaded from https://academ
ic.oup.com/aob/article/78/5/537/2587504 by guest on 01 February 2022

542 Erner and Shomer—Morphology and Anatomy of Stems and Pedicels
F 21–30. Cross-section of pedicels 3 weeks after petal fall. Figs. 21 and 22. ‘Shamouti ’ leafless inflorescence (LS) with one fruitlet (¬125 and¬650, respectively). The bundles are still separate. Figs 23 and 26. ‘Shamouti ’ LS borne with three flowers (¬125 and ¬650). The vascularsystems are composed of separate bundles, similar to that of LS with one fruitlet. Figs 24 and 25 as Figs 23 and 26 (respectively) but viewed underpolarized light. Figs 27 and 28. ‘Marsh’ grapefruit LS with one fruitlet (¬125 and ¬650). Figs 29 and 30. ‘Marsh’ grapefruit LS borne with threeflowers (¬125 and ¬650). Vessels are larger than those of ‘Shamouti ’ and fibres are well defined. C, Cambium; Cx, cortex; O, oil cavity ; F, fibre;
P, phloem; Pt, pith; V, vessel element ; X, xylem.
the central cylinder of these inflorescence types revealed 0±77and 0±39 mm#, respectively. Moreover, individual xylemvessel measurements of Shamouti showed significantdifferences between LS with one flower and LS with three
flowers, or LY with four leaves and one terminal flower, orvegetative shoot (4±86³0±26; 7±14³0±23; 11±96³0±42;9±96³0±27, respectively). Grapefruit LS with one flower(Fig. 5) exhibits similar anatomy to that of one flowered
Dow
nloaded from https://academ
ic.oup.com/aob/article/78/5/537/2587504 by guest on 01 February 2022

Erner and Shomer—Morphology and Anatomy of Stems and Pedicels 543
A
0
2
4
60
34
56
78
910
8
7
6
5
4
3
2
1
0
Ped
icel
cro
ss s
ecti
onar
ea (
mm
2 )
2
1
010
98
76
54
30
Number of leaves
2
4
6
Num
ber of flowers
B
F. 31. Relationship between inflorescence type of ‘Shamouti ’ orange, and the cross-section of the stem and pedicel, in mm#. A, At 5 mm fromthe stem base; B, At mid pedicel. S.E. does not exceed 20% of each column value.
‘Shamouti ’ LS (Fig. 1), whereas LS with three flowers ofboth varieties were thicker than those with one flower (Fig.31). Xylem vessels were easily identified under polarizedmicroscopy (Figs 6 and 7). When ‘Shamouti LYinflorescence contained four leaves with one terminal flower(Fig. 33, type 3) the secondary xylem was thicker andinduced larger vessel elements and xylem rays (Figs 8–10)than in LS with one flower (Fig. 2) — even more than LSwith three flowers (Fig. 3). The anatomy of the vegetativeshoot (Fig. 33, type 5) was similar to that of the LYinflorescence (Figs 11, 12).
The extent of the development of the vascular system wasfound to depend on the number of leaves, and this wasdependent on the number of leaves and}or flowers.
The vascular system of the pedicel (B in Figs 13–16 and33) in ‘Shamouti ’ LS inflorescence (just before floweropening) was different from that of the inflorescence stembase. Whereas the development of the vascular system in theinflorescence stem base (A) correlates with the number ofleaves, the vascular system of the pedicel (B) in allinflorescence types is built of separate vascular bundles andhas relatively equal anatomical structure.
The shoot base of LS with one flower had a relativelysimple vascular system (Figs 1–2) which, nevertheless, wasmore developed than that of the pedicel (Fig. 15). Threeweeks after anthesis, the differences between stem bases ofLS with one fruitlet and of LS with three fruitlets were verydistinctive: the vascular system of the former (Figs 17–19)
consisted of separate bundles, whereas in that of the latter(Fig. 20), it appeared as a complete ring of secondary xylem,due to cambium activity.
Three weeks after anthesis, the vascular system in thepedicel of LS with one fruitlet still consisted of separatebundles (Fig. 21), although the vascular elements seemedsomewhat larger (Fig. 22) than those of LS with one flower(Fig. 15). No difference was seen between fruitlets borne onLS with three flowers (Figs 23–26) and LS with one flower.The bundles and their vessels could be seen distinctly bypolarized light (Figs 24 and 25). As shown in Figs 27–30,the xylem vessels in the pedicel of ‘Marsh’ grapefruit LS 3weeks after anthesis with one and three flowers (with cross-section areas of 17±23³1±7 and 34±26³3±6 µm#, respectively)were larger than those of the ‘Shamouti ’ orange [(Figs 22and 26) with cross-section areas of 4±86³0±26 and7±14³0±23 µm#].
Calculation of the cross-sectional area of the xylem andpith from the various anatomy samples (Fig. 33) showedthat LS with one flower had almost 50% less vascular areathan LS with three flowers (0±39 and 0±77 mm#, respectively)and almost two thirds less than LY with four leaves and asingle flower (1±07 mm#). Moreover, the cross-sectional areaof the central cylinder was positively correlated (r¯ 0±985)with the diameter of the individual xylem vessels. Thesecalculations, in conjunction with the morphological andanatomical observations, emphasize the significantdifferences between LS with one flower, LS with three
Dow
nloaded from https://academ
ic.oup.com/aob/article/78/5/537/2587504 by guest on 01 February 2022

544 Erner and Shomer—Morphology and Anatomy of Stems and Pedicels
A
01
10
60
3 4 5 6 7 8 9 10
8
7
6
5
4
3
2
1
0
Ped
icel
cro
ss s
ecti
onar
ea (
mm
2 )
2
1
0
109
87
65
43
0 Number of leaves
1
3
10
Num
ber of flowers
B
6
3
6
Num
ber or flowers
F. 32. Relationship between inflorescence type of ‘Marsh’ seedless grapefruit, and the cross-section of the stem and pedicel, in mm#. A, At 5 mmfrom the stem base; B, At mid pedicel. S.E. does not exceed 21% of each column value.
F. 33. Inflorescence types; 1 and 2 represent leafless inflorescence(LS) shoots (no leaves) with one or three flowers, 3 and 4 represent leafyinflorescence (LY) shoots bearing four leaves, with one or severalflowers, and 5 represents a vegetative shoot with four to five leaves. A
and B represent the loci of sampling for anatomy investigation.
flowers and LY. The differences between the major xylemdiameter and the cross-sectional area of LS with pith fromthree flowers and LY with four leaves and one flower arerelatively small and tend to confirm the results which showhigh set of these inflorescences compared with a singleflower of LS inflorescences.
Water uptake and movement measurements in smallfruit-bearing citrus plants showed that the most rapid waterflow takes place in the xylem elements, through which themain part of the transpiration stream flows. The pedicel,
which contains part of the xylem system, showed a higherflow rate than the fruit itself, similar to that in leaf tissue(Mantell, Monselise and Goldschmidt, 1979; Mantell,Goldschmidt and Monselise, 1980). Defoliation of fruit-bearing branches (LY) reduced the entry of water into fruittissues (Mantell et al., 1979, 1980). Moreover, watertransport into fruitlet borne on LS, has been found to besignificantly slower than that in the fruitlet borne on LY(Steven R. Bennet, pers. comm.).
In accordance with the anatomical and the cross-sectionalarea observations in the different inflorescence types, wethink that under limited-water-supply conditions, differentparts of the tree may exhibit differing degrees of stress. LSwith one flower had a significantly smaller vascular systemat the stem base than LS with several flowers andconsiderably less than LY. Therefore, lack of water supplyaround midday would first affect the LS with one flower,leading to an increase in ABA which will later cause fruitletabscission. Since most flowers of ‘Shamouti ’ orangeoccurred on LS with one flower, no fruit set could bedetected on such inflorescences, while there was a highpercentage of set on LS with three flowers. In contrast, mostgrapefruit inflorescences were LS with many flowers or LYwith many flowers and many leaves, so that fruit set could
Dow
nloaded from https://academ
ic.oup.com/aob/article/78/5/537/2587504 by guest on 01 February 2022

Erner and Shomer—Morphology and Anatomy of Stems and Pedicels 545
take place on LS as well as on LY (Erner, unpubl. res.).Moreover, LS ‘Shamouti ’ orange have a maximum of fiveflowers while LS ‘Marsh’ grapefruit contains eight flowers(Figs 31 and 32). The many observations of LY settingbetter than LS (Reece, 1945; Sauer, 1951; Moss, 1970;Moss et al., 1972; Erner, 1989) can be interpreted asresulting from better water supply via the xylem to thesensitive, young fruitlet. Experiments conducted to finddifferences in endogenous gibberelins and IAA could notfind any (Erner, 1989) and exogenous treatments ofgibberellic acid, benzyladenine and 2,4-D could not sub-stitute for the loss of the leaves. Although fruitlet abscissionoccurs during the two months following anthesis, most ofthe fruitlets will abscise during two distinct peaks; 18–22and 32–37 d after anthesis. Optimum temperatures (20–25 °C) will increase significantly fruit set while hot and dryweather will increase fruitlet abscission (Reuther, Batchelorand Webber, 1968). The xylem development in LS with oneflower during the first three weeks after anthesis is very slowand coincides with a high peak of fruitlet abscission.Therefore, those fruitlets borne on LS with three flowers, oron LY, will dominate, simply because of the better supply ofwater and}or substances moving with the transpirationstream (e.g. cytokinins), while insufficient vascular systemdevelopment will cause those borne on LS with one flowerto abscise almost completely.
Since the flow in the xylem is proportional to the fourthpower of the diameter, the resistance of the vascular systemat the base (A) in LS with one flower is higher than that inLS with three flowers or in LY. Flowers and fruitletsadjacent to more developed xylem systems with a betterwater flow, will not suffer from water stress and, therefore,have relatively more chance to set fruits. This seems toaccount for the better set on LY compared with that on LS,in most varieties except for lemon and kumquat.
The interpretation that fruitlets borne on LS have to drawtheir assimilates from older leaves while fruitlets borne onLY can draw theirs from adjacent, young and more efficientleaves (Moss, 1970) seems to be untenable. Erner (1989)could not detect any differences between fruitlets borne onLS or LY in carbohydrate translocation, or any lack ofsource availability. The present work emphasizes theimportance of the xylem for fruit set and provides a possibleexplanation for the better set on LY than on LS when thereis one flower, while LS with several flowers can set as wellas LY.
ACKNOWLEDGEMENTS
We thank Mrs Bracha Artzi and Mrs Rosa Vasiliver fortheir technical assistance. D. M. Joel for reading themanuscript and R. Aloni for reviewing the anatomy figures.This research was partially supported by grants to Y.E.from the Israel-United States (Binational) AgriculturalResearch and Development Fund (BARD), No. IS-1643–89,and by the Citrus Marketing Board of Israel. Contributionfrom the Agricultural Research Organization, TheVolcaniCenter, Bet Dagan, Israel. No. 1177-E, 1993 series.
LITERATURE CITED
Aloni R. 1987. Differentiation of vascular tissues. Annual Re�iew ofPlant Physiology 38 : 179–204.
Aloni R. 1992. The control of vascular differentiation. InternationalJournal of Plant Science 153 : S90–92.
Davenport TL. 1990. Citrus flowering. In: Janick J, ed. Horticulturalre�iew 12 Portland: Timber Press, 349–408.
Erner Y. 1989. Citrus fruit set : carbohydrate, hormone, and leafmineral relationship. In: Wright CJ, ed. Manipulation of fruiting.London: Butterworth & Co., 233–242.
Erner Y, Bravdo B. 1983. The importance of inflorescence leaves in fruitsetting of ‘Shamouti ’ orange. Acta Horticulturae 139 : 107–112.
Fishler M, Goldschmidt EE, Monselise SP. 1983. Leaf area and fruit sizeon girdled grapefruit branches. Journal American Society forHorticultural Science 108 : 218–221.
Goldschmidt EE, Aschkenazi NY, Hrzano AA, Shaffer A, Monselise SP.
1985. A role for carbohydrate levels in the control of flowering incitrus. Scientia Horticulturae 26 : 159–166.
Goldschmidt EE, Golomb A. 1982. The carbohydrate balance ofalternate-bearing citrus trees and the significance of reserves forflowering and fruiting. Journal of the American Society forHorticultural Science 107 : 206–208.
Goldschmidt EE, Monselise SP. 1972. Hormonal control of flowering incitrus and some other woody perennials. In: Carr DJ, ed. Plantgrowth substances 1970. Berlin: Springer-Verlag, 758–765.
Jahn OL. 1973. Inflorescence type and fruiting patterns in ‘Hamlin’and ‘Valencia’ oranges and ‘Marsh’ grapefruit. American Journalof Botany 60 : 663–670.
Jahn OL. 1978. Fruiting and twig diameter in citrus. American Journalof Botany 65 : 1032–1034.
Mantell A, Goldschmidt EE, Monselise SP. 1980. Turnover of tritiatedwater in Calamondin plants. Journal of the American Society forHorticultural Science 105 : 741–744.
Mantell A, Monselise SP, Goldschmidt EE. 1979. Movement of tritiatedwater through young citrus plants. Journal of Experimental Botany30 : 155–164.
Monselise SP, Goldschmidt EE. 1982. Alternate bearing in fruit trees.In: Janick J, ed. Horticultural re�iew 4. New York: Avi PublishingCo, Inc, 128–173.
Moss GI. 1970. Fruit set in sweet orange (Citrus sinensis) : The influenceof inflorescence leaves. Phyton 27 : 141–147.
Moss GI, Steer BT, Kriedemann PE. 1972. The regulatory role ofinflorescence leaves in fruit-setting by sweet orange (Citrussinensis). Physiologia Plantarum 27 : 432–438.
Ohgaki C, Fujita K, Ito H. 1963. Investigations on the cause andcontrol of alternate bearing in Citrus unshiu orange tree. IVNitrogen and carbohydrate contents in the shoots as related toblossoming and fruiting. Journal of the Japanese Society ofHorticultural Science 32 : 157–167.
Reece PC. 1945. Fruit set in the sweet orange in relation to floweringhabit. Journal of the American Society for Horticultural Science 46 :81–86.
Reuther W, Batchelor LD, Webber HJ. 1968. The citrus industry Vol. IIanatomy, physiology, genetics, and reproduction. A CentennialPublication of the University of California, USA.
Sachs, T. 1981. The control of the patterned differentiation of vasculartissues. Ad�ances in Botanical Research 9 : 151–262.
Sagee O, Erner Y. 1991. Gibberellins and abscisic acid content duringflowering and fruit set of ‘Shamouti ’ orange. Scientia Horticulturae48 : 29–39.
Sagee O, Goren R, Riov J. 1980. Abscission of citrus leaf explants,interrelationship of abscisic acid, ethylene, and hydrolytic enzymes.Plant Physiology 66 : 750–753.
Sauer MR. 1951. Growth of orange shoots. Australian Journal ofAgricultural Research 2 : 105–117.
Wareing PF, and Phillips IDJ. 1978. The control and differentiation inplants. Oxford: Pergamon Press.
Dow
nloaded from https://academ
ic.oup.com/aob/article/78/5/537/2587504 by guest on 01 February 2022