multiple factors for organophosphorus resistance in the
TRANSCRIPT

J. Pesticide Sci. 5, 393-402 (1980)
Original Article
Multiple Factors for Organophosphorus Resistance
in the Housefly, Musca domestica L.
Naoki MOTOYAMA, Tatsumi HAYAOKA,* Ken'ichi NOMURA and Walter Carl DAUTERMAN**
Laboratory of Environmental Biology, Faculty of Horticulture, Chiba University, Matsudo, Chiba 271, Japan
**Toxicology Program, Department of Entomology,
North Carolina State University, Raleigh, North Carolina 27650, U. S. A.
(Received January 23, 1980)
Several biochemical factors responsible for organophosphate insecticide resistance in the housefly were investigated. Although no significant difference in substrate specificity for cholinesterase from resistant and susceptible housefly strains was detected, the enzyme from the resistant strains was 5 to 29-fold less sensitive to inhibition depending upon the inhibitor, than the cholinesterase from the susceptible strain. GSH S-transferases and a carboxylesterase which hydrolyzes malathion were also found to be responsible for resistance. The inter-strain difference in GSH S-transferase activity was the result of larger amounts of the same transferase in the resistant strains. On the other hand, the MFO system, cytochrome P-450, phosphorotriester hydrolase, and a-NA carboxylesterases were not involved in resistance. The in vitro degradation of malathion demonstrated that the compound was degraded mainly via carboxylesterase activity which was located in the nuclei, the mito- chondria, and the microsomal fractions, and via GSH S-transferase in the soluble fraction. It was possible to account for the cross resistance and the level of resistance in the housefly strains evaluated by the combination of resistance mechanisms.
INTRODUCTION
Geographical distribution of houseflies re-sistant to insecticides in Japan has been studied by Hayashi et al.1-4 They demon-strated that the resistant populations were widely distributed throughout the entire coun-try. Although flies exhibited cross-resistance to various insecticides, the resistance mecha-nism has not been studied in detail, except those reported by Shono. 5-8 The biochemical and physiological components of insecticide resistance have been reviewed by Oppenoorth and Welling.9 The factors involved in re-
sistance are altered properties at the site of action, increased metabolism, and reduced cuticular penetration. The involvement of carboxylesterases in
malathion resistance has been reported for a number of Japanese strains of the house-fly.13) However, an increase in the level of carboxylester hydrolysis cannot account for cross resistance to the other insecticides which do not contain a carboxyl ester group. The purpose of the present study was to
determine the various factors responsible for organophosphorous resistance in a few Japa-nese strains of the housefly.
MATERIALS AND METHODS
1. Houseflies Houseflies, Musca domestica L., used in the
present study are the insecticide susceptible
* Present address: Toxicology Program, Depart-
ment of Entomology, North Carolina State
University, Raleigh, North Carolina 27650,
U. S. A.

394 日本農薬学会誌 第5巻 第3号 昭和55年8月
Takatsuki strain which was collected in Takatsuki, Osaka, the resistant Chiba-D strain, which was collected in Dejima, Ibaragi, and the resistant Hirokawa strain, originally col-lected in Hirokawa, Wokayama. These flies were kindly supplied by Dr. A. Hayashi of the Public Health Laboratory of Chiba Pre-fecture and maintained without insecticidal
pressure. The response of these housefly strains to certain insecticides has been reported
previously.4
2. Chemicals Insecticides were obtained as a gift from
the following sources: fenitrothion and f enitro-xon from Sumitomo Chemical, Ltd.; diazinon from Nihon Kayaku, Ltd.; malathion, mala-oxon, malathion monoacid, and paraoxon from American Cyanamid; m-tolyl methylcarba-mate (MTMC) from Mitsubishi Chemicals, Ltd. and diazoxon from Ciba-Geigy, Inc. These compounds were either of an analytical grade or used after purification. 0, 0-dimethyl S-(1, 2-dicarbethoxy) [1, 2-14C] phosphorodi-thioate (14C-malathion) was purchased from Amersham/Searle Corporation and had a specific activity of 4.6mCi Jmmol. Water-soluble metabolites of malathion, i. e. des-methyl malathion, malathion diacid, 0, 0-dimethyl phosphorodithioic acid, 0, 0-di-methyl phosphorothioic acid, and 0, 0-di-methyl phosphoric acid were synthesized and are the same as those used in a previous study.14 Reduced glutathione (GSH), 3, 4-dichloronitrobenzene (DCNB), a-naphthyl acetate (a-NA), acetylthiochloline iodide
(AThCh), propionylthiocholine iodide (PThCh), butyrylthiocholine iodide (BThCh) and bovine
plasma albumin (Fraction V) were obtained from Wako Chemical Industries, Ltd. Reduced forms of nicotinamide-adenine dinucleotide
phosphate (NADPH) and j-nitroanisole were obtained from Oriental Yeast Co., Ltd., andKanto Chemical Co., Inc., respectively. Other chemicals used were of the highest grade com-mercially available.
3. Toxicity Evaluations The toxicity of the test compounds was
determined by topical application of 0.5l of acetone solutions containing the insecticides to
individual female flies (4 to 6 days old). Treated flies were held at 25C and the mor-talities were determined 24hr. after treat-ment. LD50 values were calculated by regres-sion analysis.
4. Enzyme Preparation For the cholinesterase assay, fly heads were
homogenized with 0.1M potassium phosphate buffer, pH 8.0 (20 male heads per 10ml) using a Potter-Elvehjem Teflon® homogenizer fitted with a small electric motor (National Baby Motor SC-V535W(R), Matsushita Electric In-dustrial Co, Ltd.) The homogenate was filtered through glass wool, and the filtrate was used as the enzyme source. Subcellular fractions of whole body or abdomen homo-
genates were prepared by differential centrifu-gation using a Hitachi Ultra Centrifuge Model 55PA according to the method described by Hansen and Hodgson.15 Each fraction was resuspended in an appropriate buffer as de-scribed in the text. In order to protect the enzymes from endogenous inhibitors, 1% bovine serum albumin was included in the buffer throughout the preparations.16,17
5. Protein Determination Protein was determined according to the
method of Sedmak and Grossberg18 with bovine plasma albumin (Fraction V) used as a standard.
6. Cholinesterase Assay Cholinesterase activity of fly head homo-
genates was measured according to the method of Ellman et al. The typical incubation mixture contained 1.3ml of 0.1M potassium phosphate buffer, pH 8.0; 0.2ml of the enzyme solution (equivalent to 0.5 male heads), 1.0ml of 0.01M 5, 5'-dithiobis-(2-nitrobenzoic acid) (DTNB) in buffer, and 0.5ml of distilled water containing various concentrations of the different substrates. The reaction mixture was incubated at room temperature (approximately 17 to 19C) and the change in absorbance at 412nm was measured using a Hitachi Spectro-
photometer Model 101. Nonenzymatic hy-drolysis was assayed by replacing the enzyme with the buffer. The in vitro cholinesterase inhibition was determined by adding 5 to 100

Journal of Pesticide Science 5 (3), August 1980 395
l of various concentrations of the inhibitors in acetone simultaneously to the above incuba-tion mixture.
7. GSH S-transferase Assay GSH S-transferase activity was determined spectrophotometrically using DCNB as the substrate.20 The reaction mixture consisted of 1.5ml of the postmicrosomal solution as the enzyme source and 0.1M Tris-HC1 buffer, pH 9.0 in a total volume 3ml. The reaction was run at room temperature (17 to 19C) and a change in absorbance at 344nm was deter-mined using a Hitachi Spectrophotometer Model 101. All reactions were corrected for nonenzymatic conjugation.
8. Mixed function Oxidases (MFO) Assay As an index for MFO activity in housefly
microsomes, 0-demethylation of p-nitroanisole was determined. The assay method was a modification of that described by Hansen and Hodgson.15 The reaction mixture contained 1.5ml of microsomes (equivalent to 15 male abdomens), 0.5ml of 2mM NADPH, 20l of 50mM p-nitroanisole (in ethanol) and 0.1ml Tris-HCL buffer, pH 7.9 in a total volume of 3.0ml. The reaction mixture was incubated at 37C for five minutes, and the change in absorbance at 400nm was recorded using a Hitachi 556 spectrophotometer. All reactions were corrected for nonenzymatic demethyl-ation.
9. Cytochrome P-450 CO-binding Difference Spectrum
CO-binding difference spectra of cytochrome P-450 was measured according to the method of Omura and Sato21 and as cited by Folsom et al.22 To 3ml of a microsomal suspension
(equivalent to 30 male abdomens), a few mgs. of sodium dithionite was added; and then CO was bubbled only into the sample cuvette. The difference spectrum was recorded using a Hitachi 556 spectrophotometer.
10. c-Naphthyl Acetate Hydrolysis Non-specific esterase activity in the sub-
cellular fractions from whole body homo-
genates was determined using a-naphthyl acetate as the substrate according to the meth-
od of van Asperen.23 The reaction mixture consisted of 50al of a subcellular fraction
(equivalent to 0.25male whole bodies) in 0.1M Tris-HC1 buffer, pH 9.0, 1.0ml of 6 x 10-4M aqueous solution of a-naphthyl acetate (meth-anol 0.4%), and 2.0ml of 0.1M sodium phos-
phate buffer pH 7.0. The reaction mixture was incubated at 27C for fifteen min. Non-enzymatic hydrolysis was corrected by sub-stituting the buffer in place of the subcellular fractions.
11. In Vitro Metabolism of Malathion A subcellular fraction (equivalent to five
female abdomens) was incubated with 0.25mM 14C-malathion in 1.0ml of 0.05M potas-sium phosphate buffer, pH 7.0, in the presence or absence of either 4mM GSH or 1.1mM NADPH. The reaction was carried out at 37C and stopped after one hr by the addition and mixing of 1ml of chloroform. After sepa-ration of the phases, an aliquot of the aqueous layer was added to Triton X-100 cocktai124and the radioactivity quantitated with an Aloka Liquid Scintillation Counter Model 650. The nonenzymatic reaction of malathion was corrected by using a heat denatured prepara-tion (1min in a boiling water) in place of the regular incubate.
12. Identification of Metabolites An aliquot of the aqueous layer was ap-
plied to a silica gel G thin layer plate (0.1mm precoated, Eastman) and developed to a height of 15cm with acetonitrile: water (85 15v/v). Non-radioactive standards were co-chromatographed and visualized by spraying with PbCl2-HC1 solution or 2, 6-dibromo-N-chloro p-benzoquinonimine-cyclohexane solu-tion. Silica gel from unsprayed plates was scraped off at 1cm intervals, and the radio-activity was quantitated by liquid scintilla-tion counting.
13. Phosphorotriester Hydrolase Assay Paraoxon was used as the substrate to com-
pare phosphorotriester hydrolase activity in subcellular fractions. The reaction mixture consisted of the subcellular fraction (equi-valent to 10 to 30 male abdomens), 5 to 50lof an ethanolic solution of 50mM paraoxon,

396 日本農薬学会誌 第5巻 第3号 昭和55年8月
and 0.05M potassium phosphate buffer, pH 7.8 in a final volume of 3ml. The reference cuvette contained the reaction mixture with-out the substrate. For control of nonenzymatic hydrolysis, the subcellular fractions were de-leted from both the sample and the reference cuvettes. The change in absorbance at 400nmwas recorded using a Hitachi 556 spectro-
photometer.
RESULTS AND DISCUSSION
1. Cross Resistance to OP Insecticides The LD50 values for the housefly strains
with three organophosphorus (OP) insecti-cides are presented in Table 1. The resistance factor (R/S) for the Hirokawa strain was ex-tremely high for malathion, intermediate for fenitrothion, and relatively low for diazinon. With Chiba-D strain, the resistance factors
for malathion and fenitrothion were approxi-mately in the same range, and that for diazinon was low. The findings would indicate that more than one factor was responsible for re-sistance in the housefly strains under investi-
gation.
2. Cholinesterase Studies The specificity of cholinesterases from the
resistant and susceptible strains of the house-fly was investigated using acetylthiocholine
(AThCh), propionylthiocholine (PThCh), and butyrylthiocholine (BThCh) as substrates. The findings indicated no significant interstrain difference based on the Km and Vmax values. The average Km values obtained under the experimental conditions described were (6.4+1.6) x 10-5M for AThCh, (2.7+0.8) x 10M for PThCh and (7.9+1.3) x 10M for BThCh.
Table 1 Toxicity of some organophosphate insecticides to different housefly strains.
Table 2 Inhibition of flyhead cholinesterase from resistant and susceptible strains of house-
flies by various inhibitors.*
Determined by simultaneous inhibition in the presence of 4 x 10-4M AThCh.

Journal of Pesticide Science 5 (3), August 1980 397
Similarly, the average Vmax values for these substrates were (2.9+0.5) X 10-5M per male head per min, (1.3+0.3) X 10-5M per male head per min and (0.9+0.3) X 10-5M per male head per min, respectively. The in vitro inhibition of cholinesterase with
various concentration of the oxygen analogs of the OP insecticides as well as with MTMC was studied. AThCh was used as the sub-strate at a concentration of 4 X 10-4M. The results are expressed as 150 values and are summarized in Table 2. Cholinesterases from both the Chiba-D and the Hirokawa strains exhibited a marked decrease in sensitivity to inhibition by these compounds. The ratio of I50 values of the resistant and susceptible strains varied with the inhibitor. The largest interstrain difference was observed with feni-troxon. This was followed by malaoxon, dia-zoxon, and MTMC in this order. The results suggest that altered cholinesterases in the
Japanese housefly strains are an important factor in OP resistance. The involvement of insensitive cholinesterase in OP resistance has been reported with mites, ticks, green rice leafhoppers, Nephotetix cincticeps, as well as with a tetrachlorovinphos resistant strain of the housefly.9 The present data would indi-cate that this factor may be a fairly common mechanism for OP resistance in different species and different strains. An attempt was made to inhibit housefly cholinesterases with the n-propyl analog of MTMC, 3-tolyl N-propylcarbamate, since this compound has been reported to inhibit altered cholinesterases to a greater extent than the normal choline-sterases found in the green rice leafhopper.25-27)The potency of this compound to inhibit housefly cholinesterases was extremely poor and is similar to the findings of Takahashi
(personal communication, 1978). Therefore, it was impossible to determine and compare 150 values for the resistant and susceptible strains. Nevertheless, the fact that the ratio of 150 values for the resistant and susceptible strains varied, depending upon the structure of the OP compounds, may suggest the pos-sible existence of a specific inhibitor for in-sensitive cholinesterases of the housefly.
3. GSH S-trans f erase Activity Increased metabolism of OP insecticides by
GSH S-transferases is another important factor for resistance2 and the role of this
group of enzymes in the metabolism of OP insecticides has been recently reviewed.29-32
In the present study, GSH S-transferase activity was determined using DCNB as the substrate, and the results are presented in Fig. 1. It was obvious that there was a marked difference in enzyme activity which, in general,
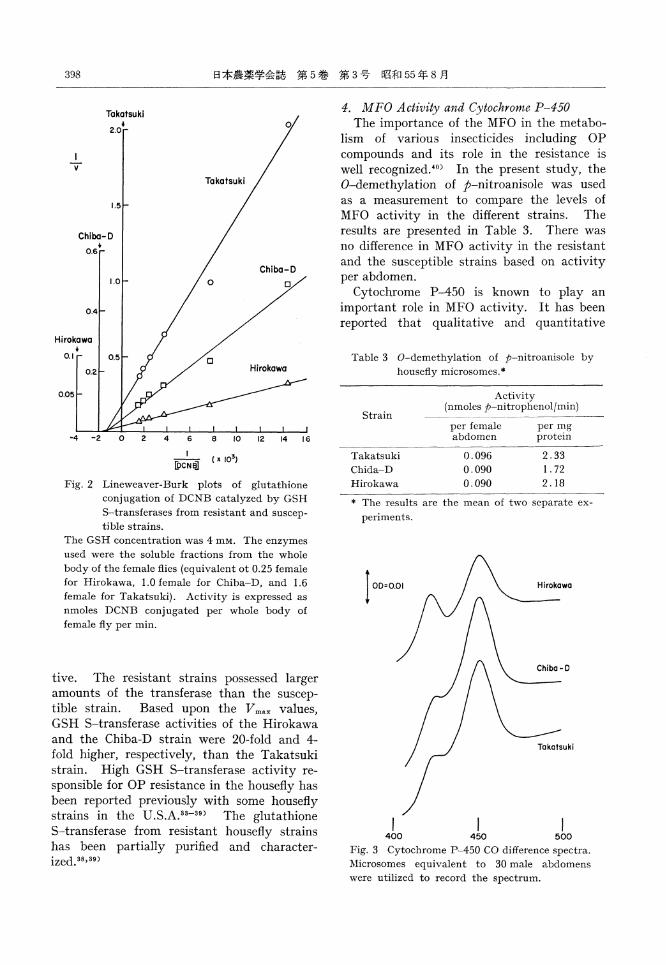
paralleled the resistance levels; namely, the Hirokawa strain had the highest activity fol-lowed by the Chiba-D and then the Takatsuki strain. In order to investigate whether the interstrain difference in GSH S-transf erase activity was quantitative or qualitative, the effect of various concentrations of DCNB on enzyme activity at a constant GSH concent-ration (4mM) was determined. The double reciprocal plot of the data is presented in Fig. 2. The GSH S-transferases from the three strains had similar Km values, but apparently different Vmax values. Therefore, it seems that the interstrain difference was quantita-
Fig. 1 Glutathione S-transferase activity in the soluble fraction from resistant and sus- ceptible housefly strains.
The activity is expressed as nmoles of DCNB conjugated per whole body of female fly.
398 日本農薬学会誌 第5巻 第3号 昭和55年8月
tive. The resistant strains possessed larger amounts of the transf erase than the suscep-
tible strain. Based upon the Vmax values, GSH S-transferase activities of the Hirokawa
and the Chiba-D strain were 20-fold and 4-fold higher, respectively, than the Takatsuki
strain. High GSH S-transferase activity re-sponsible for OP resistance in the housefly has
been reported previously with some housefly strains in the U. S. A. 33-39) The glutathione
S-transferase from resistant housefly strains
has been partially purified and character-ized.38,39
4. MFO Activity and Cytochrome P-450 The importance of the MFO in the metabo-
lism of various insecticides including OP compounds and its role in the resistance is well recognized.40 In the present study, the 0-demethylation of p-nitroanisole was used as a measurement to compare the levels of MFO activity in the different strains. The results are presented in Table 3. There was no difference in MFO activity in the resistant and the susceptible strains based on activity
per abdomen. Cytochrome P-450 is known to play an
important role in MFO activity. It has been reported that qualitative and quantitative
Fig. 2 Lineweaver-Burk plots of glutathione conjugation of DCNB catalyzed by GSH S-transferases from resistant and suscep- tible strains.
The GSH concentration was 4ms. The enzymes used were the soluble fractions from the whole body of the female flies (equivalent of 0.25 female for Hirokawa, 1.0 female for Chiba-D, and 1.6 female for Takatsuki). Activity is expressed as nmoles DCNB conjugated per whole body of female fly per min.
Table 3 0-demethylation of p-nitroanisole by
housefly microsomes.*
* The results are the mean of two separate ex-
periments.
Fig. 3 Cytochrome P-450 CO difference spectra.
Microsomes equivalent to 30 male abdomens
were utilized to record the spectrum.
OD=0.01

Journal of Pesticide Science 5 (3), August 1980 399
differences in cytochrome P-450 have been associated with insecticide resistance,41,42)Therefore, an attempt was made to compare the cytrochrome P-450 CO binding spectrum from the three strains. The data is shown in Fig. 3. Although the amount of cytochrome P-450 in microsomes from the Hirokawa strain appeared to be slightly lower than those from the other two strains, the amount of cytochrome P-420 in the Hirokawa strain was slightly higher than the others. There-fore, it appears that a portion of the P-450 was converted to P-420 during the preparation of the microsomes. No difference in cyto-chrome P-450 in the housefly strains was noted. The results are in agreement with the above observation that the MFO was not as-sociated with the resistance mechanism in the housefly strains evaluated.
5. Hydrolases Phosphorotriester hydrolases (so-called phos-
phatases) have been considered to be an im-portant or sometimes the sole factor for OP resistance in the housefly.28 However, owing to the findings that the same metabolites could be formed via oxidation by MFO, or via conjugation by GSH S-transferases, the importance of the hydrolytic reaction has become more and more debatable.9
In the present study, the enzyme activity was determined using paraoxon as the sub-strate. Under the experimental conditions in which mouse liver microsomes (equivalent to 0.3g of wet liver weight) exhibited significant hydrolytic activity toward paraoxon, forming
0, 0-diethyl phosphoric acid and -nitrophenol, the housefly preparation exhibited little or no activity. Therefore, phosphorotriester hydro-lases are probably not involved in resistance, at least in the housefly strains investigated. A similar observation has also been reported by Shishido and Fukami4 who did not detect any hydrolysis of diazoxon by the fat body of the American cockroach despite the ad-dition of various cofactors, etc. Carboxylesterases which are assayed with
a-naphthyl acetate (NA) as the substrate are often called "nonspecific esterases." Esterase activity toward a-naphthyl acetate has been extensively studied with regard to OP re-sistance in a variety of insect and mite spe-cies. 2S A number of studies have reported either high or low esterase activity which is associated with OP resistance. No consistent relationship between the NA carboxylesterases and the level of resistance has been found. In the present study, a-NA hydrolysis was meas-ured using subcellular fractions from three housefly strains. There was no noticeable inter-strain difference in a-NA carboxyl-esterase activity in any subcellular fraction or in the total activity. In contrast, however, Hayashi and Funaki (personal communication, 1978) found that certain electrophoretic bands active in a-NA hydrolysis were closely related to OP resistance in the housefly. Although the enzymatic identity of these electrophoretic bands was not clarified, a possible role of these enzymes in resistance cannot be ruled out.
6. In Vitro Metabolism of Malathion The in vitro degradation of malathion was
investigated in subcellular fractions from the three housefly strains. The results are sum-marized in Table 5. All the subcellular frac-tions degraded malathion with the highest activity being found in the Hirokawa strain and the lowest in the Takatsuki strain. Micro-somal degradation was not dependent on NADPH, suggesting that the reaction was
probably not mediated by the MFO system. On the other hand, the degradation by the
post-microsomal fractions increased with the addition of GSH, suggesting the involvement of GSH S-transferases in the reaction. The water soluble metabolites of malathion were
Table 4 a-Naphthyl acetate hydrolysis by sub-
cellular fractions from whole houseflies.
* The results are the mean of three separate
experiments.

400 日本農 薬学会誌 第5巻 第3号 昭和55年8月
separated on tlc(Fig. 4). The major metaboliteisolated from each fraction was identified asmalathion monoacid by cochromatography. Malathion degradation-by these fractions wascatalysed by carboxylesterases. Hotivever, themajor water soluble metabolite obtained inthe soluble fraction fortified with GSH wasdesmethyl malathion(Fig. 4, F). The findings
confirm the earlier suggestion that the reactionwas catalyzed by GSH S-transferases. Thetlc also revealed trace amounts of malathiondiacid in certain fractions. It is not clear howthe diacid was formed, but it may be a productof non-enzymatic hydrolysis of malathionmonoacid. The non-enzymatic hydrolysis of malathionthan one biochemical mechanism was re-sponsibie for OP resistance in th. e Japanesehousefly strains studied. Cross resistance andthe level of resistance may be determined bythe availability of these factors. The extremelyhigh level of resistance to malathion in theHirokawa strain can be explained by (1) analtered cholinesterase, (2) high GSH S-trans-ferase activity and (3) high carboxylesteraseactivity. The latter two enzyme systemscatalyze reactions which detoxify malathion. The effect on toxicity of these three factorswould be sufficient to account for more than1,200-fold resistance to malathion. The inter-mediate level of resistance to fenitrothion inthis strain can be explained by the combina-tion of two of these factors, i. e., altered choline-sterase and high GSH S-transferase activity. Although fenitrothion is an excellent sub-strate for O-demethylation by the GSH S-transferases, it is not a substrate for thecarboxylesterase. The low level of diazinonresistance can be explained since only one ofthe three factors, i. e., altered cholinis involved in the resistance of this compound. Diazinon is not a substrate for the carboxyl-esterase and is only a fair substrate for GSHS-transferases. GSH S-transferases are knownto conjugate methoxy compounds better than
Table 5 In vitro degradation of malathion by subcellular fractions from resistant and
susceptible houseflies.*
*The results are the mean of two separate experiments.
Fig. 4 Radiochrom. atograms of water soluble metabolites of 14C-malathion formed by subcellular fractions from the Hirokawa strain.
Upon cochromatography, (a) coincided with malathion dicarboxylic acid, (b) coincided with malathion monocarboxylic acid, and (c) coincid-ed with desmethyl malathion. A: Nuclei, B: Mitochondria, C: Microsomes, D: Microsomes +1.1mM NADPH, E: Soluble fraction, F: Soluble fraction+4mi GSH.

Journal of Pesticide Science 5 (3), August 1980 401
ethoxy compounds and diazinon is an ethoxy compound.32) No evidence was obtained which indicated the involvement of other resistance factors such as MFO system, cytochrome P-450,
phosphorotriester hydrolases, and a-NA carboxylesterases in OP resistance in the
Japanese strains studied. In the present study, the combination of the individual bio-chemical resistance mechanisms may explain cross resistance and the level of resistance.
ACKNOWLEDGEMENTS
This paper is No. 6261 of the Journal Series of the North Carolina Agricultural Research Service, Raleigh, North Caroline. Work supportedd in part by PHS Research Grant ES-00044 from the National Institute of Environmental Health Sciences.
REFERENCES
1) A. Hayashi, M. Hatsukade, M. Hasegawa & K. Hattori: Botyu-Kagaku 36, 41 (1971)
2) A. Hayashi, T. Tanaka & S. Matsuzaki: Botyu- Kagahu 37, 91 (1972)
3) A. Hayashi, Y. Kusui, T. Shinonaga, Y. Ishigaki & R. Kano: Botyu-Kagaku 42, 97 (1977)
4) A. Hayashi, E. Funaki, M. Fuzimagari, R. Kano & K. Nomura: Botyu-Kagaku 42, 198 (1977)
51 T. Shono: Jpn. J. Sanit. Zool. 24, 248 (1974) 6) T. Shono: Botyu-Kag ahu 39, 54 (1974) 7) T. Shono: Botyu-Kagaku 39, 75 (1974) 8) T. Shono: Botyu-Kagahu 39, 80 (1974) 9) F. J. Oppenoorth & L. W. Welling: "Insecticide
Biochemistry and Physiology," ed. by C. F. Wil- kinson, Plenum Press, New York, p. 507, 1976
10) H. Ohkawa, M. Eto, Y. Oh. shima, F. Tanaka & K. Umeda: Botyu-Kagaku 33, 139 (1968)
11) A. Hayashi & M. Hasegawa: Hokkaido Inst. Public Health Bull. 7, 1 (1974)
12) A. Hayashi, R. Kano & S. Ishibashi: Botyu- Kagahu 40, 154 (1975)
13) Y. Niwa, T. Miyata & T. Saito: J. Pesticide Sci. 2, 151 (1977)
14) A. A. Nomeir & W. C. Dauterman: Biochem. Pharmacol. 27, 2975 (1978)
15) L. G. Hansen & E. Hodgson: Biochem. Phayma- col. 20, 1569 (1971)
16) M. Tsukamoto & J. E. Casida: J. Econ. Ento- mol. 60, 619 (1967)
17) N. Motoyama, A. P. Kulkarni, E. Hodgson & W. C. Dauterman: Pestic. Biochem. Physiol. 9, 255 (1978)
18) J. J. Sedmak & S. E. Grossberg: Anal. Biochem. 79, 544 (1977)
19) G. L. Ellman, K. D. Courtney, V. Andres & R. M. Featherstone: Biochem. Pharmacol. 7, 88
(1961) 20) J. Booth, E. Boyland & P. Sims: Biochem. J.
79, 516 (1961) 21) T. Omura & R. Sato: J. Biol. Chem. 239, 2370
(1964) 22) M. D. Folsom, L. G. Hansen, R. M. Philpot, R.
S. H. Yang, W. C. Dauterman & E. Hodgson: Life Sci, 9, 869 (1970)
23) K. van Asperen: J. Insect Physiol. 8, 401 (1962) 24) M. J. Patterson & R. C. Greene: Anal. Chem.
37, 854 (1965) 25) I. Yamamoto, N. Kyomura & Y. Takahashi:
J. Pesticide Sci. 2, 463 (1977) 26) Y. Takahashi, N. Kyomura & I. Yamamoto:
J. Pesticide Sci. 2, 467 (1977) 27) Y. Takahashi, N. Kyomura & I. Yamamoto:
J. Pesticide Sci. 3, 55 (1978) 28) N. Motoyama & W. C. Dauterman: J. Agyic.
Food Chem. 22, 350 (1974) 29) R. S. H. Yang: "Insecticide Biochemistry and
Physiology," ed. by C. F. Wilkinson, Plenum Press, New York, p. 177, 1976
30) T. Shishido & J. Fukami: Bull. Nat. Inst. Agyic. Sci. Sey., No. 32, Tokyo, Japan (1978)
31) J. Fukami: "Pharmacology and Theurapeu- tics," ed. by F. NIatsumura, Pergamon Press,
New York, 1980 32) N. Motoyama & W. C. Dauterman: "Reviews
in Biochemical Toxicology," eds. by E. Hodgson,
J. R. Bend and R. M. Philpot, Vol. 2, Flsevier North-Holland, Inc., New York, 1980
33) N. Motoyama & W. C. Dauterman: Pestic. Biochem. Physiol. 2, 113 (1972)
34) N. Motoyama & W. C. Dauterman: Pestic. Biochem. Physiol. 5, 489 (1975)
35) F. J. Oppenoorth, H. R. Smissaert, W. Welling, L. J. T, van der Pass & K. H. Hitman: Pestic.
Biochem. Physiol. 7, 34 (1977) 36) N. Motoyama & W. C. Dauterman: Pestic.
Biochem. Physiol. 7, 443 (1977) 37) F. J. Oppenoorth, L. J. T. van der Pass & N. W.
Houx: Pestic. Biochem. Physiol. 11, 176 (1979) 38) N. Motoyama & W. C. Dauterman: Insect.
Biochem. 7, 361 (1977) 39) N. Motoyama & W. C. Dauterman: Insect.
Biochem. 8, 337 (1978) 40) T. Nakatsugawa & M. A. Morelli: "Insecticide
Biochemistry and Physiology," ed. by C. F. Wilkinson, Plenum Press, New York, p. 61,
1976 41) E. Hodgson, L. G. Tate, A. P. Kulkarni & F. W.
Plapp, Jr.: J. Agyic. Food Chem. 22, 360 (1974) 42) E. Hodgson & L. G. Tate: "Insecticide Bio-
chemistry and Physiology," ed. by C. F. Wilkin-

402 日本農薬 学会誌 第5巻 第3号 昭和55年8月
son, Plenum Press, New York, p. 115, 1976 43) T. Shishido & J. rukami: Pestic. Biochem.
Physiol. 2, 39 (1972)
要 約
イエ バ エ に お け る 有 機 リン 殺 虫 剤 抵 抗 性 の 生 化
学 的 機 構
本 山直樹, 早岡辰巳
野村健一, W. C. ドウターマ ン
イエバエにおけ る有 機 リン殺 虫剤抵抗 性の機構に関与
す るい くつかの生化 学的要 因について検討 した. コ リン
エステ ラーゼ(ChE)の 基質特異性 に関 しては, 抵 抗性一
感受性系統間に有 意な差 はなか つた. しか し, 抵抗性系
統のChEは 感受性 系統に比較 して, 阻害剤 に対す る感
受性が5~29倍 低 か つた. グル タチオ ンS-ト ランス ファ
ラーゼ と, マ ラソンを加水 分解す るカルボキシルエ ステ
ラーゼ も抵抗性 の機構 に関与 してお り, とくに前者の場
合は, 系統 間にみ られ る活性 の差 は, 酵素の量的な差に
基づ くことが示唆 された. 一方, 薬 物酸化酵素(MFO),
リン酸 トリエステル加水分解酵素, α-ナフチルアセテ ー
ト加水分解酵 素等 の活性, お よびチ トク ローム P-450
の スペ ク トル については, 系統 間 に有意 な 差がなか つ
た. また, 14C-マ ラソンのin vitro におけ る分解代 謝
は, 主 として, 核, ミ トコン ドリア, ミクロゾー ム画分
に存在す るカルボキシルエステラーゼと, 上澄画分に存
在す る グルタチオ ンS-ト ランスフ ァラーゼ によること
がわか った. 本研究 は, 有機 リン殺虫剤抵抗性に関与す
るこのよ うな生化学的諸要因 を分析 し, 組 み合わせ るこ
とによつて, 交差抵抗性 の現象や抵抗性の程度 の違いを
説明す ることが可能であ ることを示唆 した.