multiple penicillin-binding components in bacillus subtilis, bacillus ... · multiple...
TRANSCRIPT

THE JOURNALS OF BIOLOGICAL CHEMISTRY Vol. 247, No. 17, Issue of September 10, pp. 5279-5288, 1972
P&ted in U.S.A.
Multiple Penicillin-binding Components in Bacillus subtilis,
Bacillus cereus, Staphylococcus aureus, and
Escherichia coZi*
(Received for publication, January 24, 1972)
HIDEKAZU SUGINAKA,$ PETER M. BLUMBERG,~ AND JACK L. STROMINGER
From the Biological Laboratories, Harvard University, Cambridge, Massachusetts 02138
SUMMARY
A variety of techniques have been employed to investigate the penicillin-binding components of Bacillus subfilis, Ba- cillus cereus, Staphylococcus aureus, and Escherichia coli. Virtually all the binding sites in these organisms are located in the cell membrane. All the techniques which have been employed suggest that more than one binding component exists in each of these organisms, although S. aureus and E. coli both appear to have only a single major one. A partic- ularly interesting observation was that in Bacillus subtilis one of the two major components which reacts with penicil- lin G does not react readily with cephalothin.
Radioactive penicillin G is irreversibly fixed to the cells of microorganisms which are killed by this antibiotic. Many studies of the nature of the bound penicillin have been carried out over the past 20 years (%14). It has been suggested that the penicillin is bound to a lipoprotein near the surface of the microbial cell. Recent studies from this laboratory have shown that particulate enzyme preparations derived from the cell mem- brane of Bacillus subtilis also bind radioactive penicillin G and that this penicillin G is released on treatment with hydroxyl- amine or with ethanethiol (13). The recovery of penicilloyl hydroxamate and of an a-ethylthiol derivative of penicilloic acid after these treatments suggested that the penicillin was bound as a penicilloyl derivative. Moreover, the inhibition of penicillin binding by thiol reagents such as iodoacetamide sug- gested that the penicilloyl group was substituted in the particu- late enzyme as a thioester. An enzyme present in the particulate enzyme preparation of B. subtilis, D-alanine carboxypeptidase, is “irreversibly” inactivated after the binding of penicillin G, but its activity is restored on treatment with hydroxylamine. Peni-
* This work was supported by research grants from the United States Public Health Service (AM-13230 and AI-09152) and the National Science Foundation (GB-29747X). This publication is number XXVIII in the series, “Biosynthesis of the Peptidoglycan of Bacterial Cell Walls.” Number XXVII is Reference 1.
f Present address, Department of Microbiology, Osaka Uni- versity Dental School, Osaka, Japan.
5 Supported in part by a National Science Foundation Predoc- toral Research Fellowship.
cillin G, therefore, appears to be bound to the D-alanine carboxy- peptidase. These data, however, do not resolve the question of whether there is one or more than one penicillin-binding com- ponent in this preparation and whether the fraction bound to the D-ahiDe earboxypeptidase is a major or a minor (or perhaps the only) penicillin-binding component.
The present experiments were initiated in order to further investigate the nature of the component or components of bac- terial cells which bind radioactive penicillin. Is there one or more than one of these penicillin-binding components?
MATERIALS AND METHODS
[14C]Penicillin G (labeled with [l-*4C]phenylacetic acid) was obtained from Amersham Searle, Arlington Heights, Illinois. It had a specific activity of 26.5 PCi per pmole and yielded 49 cpm per ppmole under the conditions of counting (84% effi- ciency). [3H]Penicillin G (generally labeled in the phenylacetyl moiety) was obtained from the same source. It had a specific activity of 181 PCi per pmole and yielded 110 cpm per ppmole under the conditions of counting (28% efficiency). Samples for counting were in a maximum of 0.5 ml of water. To the sample in a glass scintillation vial, 1 ml of Biosolv, formula BBS-3 (Beckman Instruments), was added followed by 9 ml of scintillation fluid containing 15.2 g of 2,5-diphenyloxazole and 1.14 g of 1,4-bis[2-(4-methyl-5-phenyloxazole)]benzene in 1 gallon of toluene. Counting was carried out in a liquid scin- tillation spectrometer (Packard Instrument Company).
Typical reaction mixtures for measuring binding to whole cells or particulate enzyme preparations are described in the legends to Figs. 1 and 2. After binding, there are a number of alterna- tives for removing unbound penicillin prior to counting the sample. These include washing the particulate material by centrifugation, recovering the particulate material by filtration on Millipore filters and subjecting the incubation mixture to chromatography in isobutyric acid-l N ammonium hydroxide, 5:3, and then counting the material which remains at the origin of the chromatogram. In general, it has been found that treat- ing the incubation mixture with penicillinase prior to any of these procedures reduces the blank and increases the reliability of the measurement. Therefore, all assays were carried out after adding 5 ~1 of penicillinase (800 units per ~1, obtained from Cal- biochem) and incubating at 30” for 10 min. Washing the par- ticulate material after penicillinase treatment was found to be the most reliable method and yielded the lowest blanks. It has
5279
by guest on July 18, 2019http://w
ww
.jbc.org/D
ownloaded from
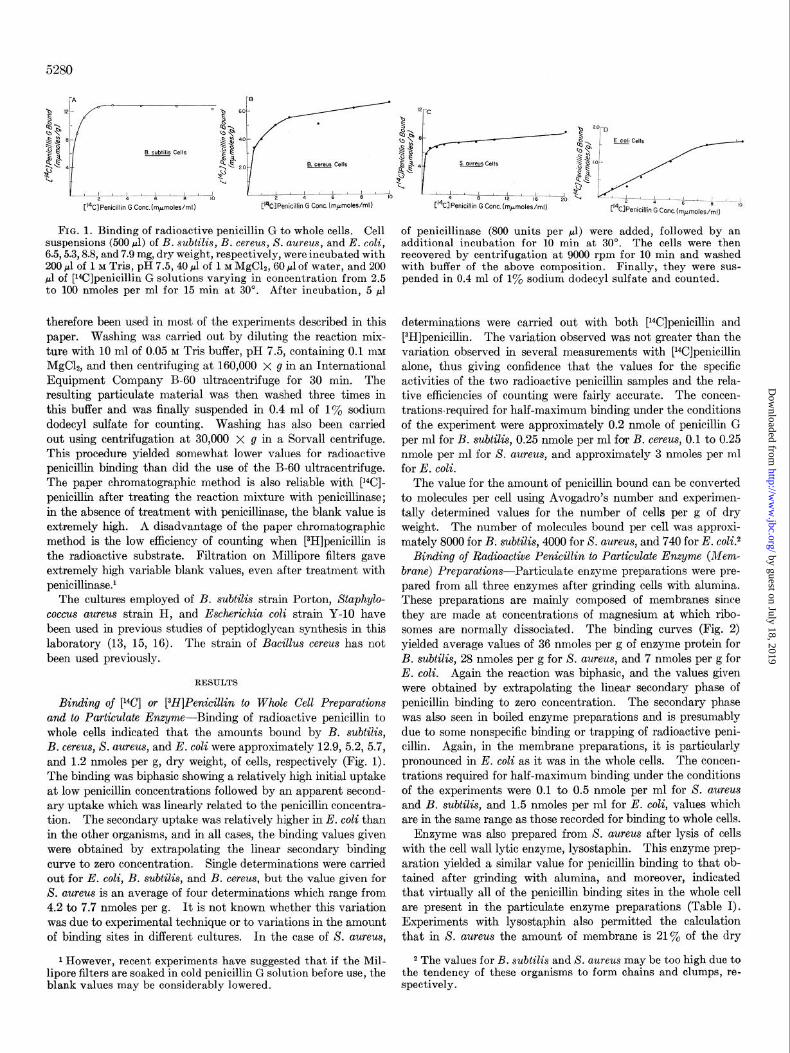
5280
FIG. 1. Binding of radioactive penicillin G to whole cells. Cell suspensions (500 ~1) of B. subtilis, B. cereus, S. aureus, and E. coli, 6.5,5.3,8.8, and 7.9 mg, dry weight, respectively, were incubated with 200 ~1 of 1 M Tris, pH 7.5,40 ~1 of 1 M MgClz, 60 ~1 of water, and 200 ~1 of [l*C]penicillin G solutions varying in concentration from 2.5 to 100 nmoles per ml for 15 min at 30’. After incubation, 5 ~1
therefore been used in most of the experiments described in this paper. Washing was carried out by diluting the reaction mix- ture with 10 ml of 0.05 M Tris buffer, pH 7.5, containing 0.1 InM
MgC12, and then centrifuging at 160,000 x g in an International Equipment Company B-60 ultracentrifuge for 30 min. The resulting particulate material was then washed three times in this buffer and was finally suspended in 0.4 ml of 1% sodium dodecyl sulfate for counting. Washing has also been carried out using centrifugation at 30,000 X g in a Sorvall centrifuge. This procedure yielded somewhat lower values for radioactive penicillin binding than did the use of the B-60 ultracentrifuge. The paper chromatographic method is also reliable with [‘“Cl- penicillin after treating the reaction mixture with penicillinase; in the absence of treatment with penicillinase, the blank value is extremely high. A disadvantage of the paper chromatographic method is the low efficiency of counting when [aH]penicillin is the radioactive substrate. Filtration on Millipore filters gave extremely high variable blank values, even after treatment with penicil1inase.r
The cultures employed of B. subtilis strain Porton, Stctphylo- coccus aureus strain H, and Escherichia coli strain Y-10 have been used in previous studies of peptidoglycan synthesis in this laboratory (13, 15, 16). The strain of Bacillus cereus has not been used previously.
RESULTS
Binding of [“Cl or [3H]Penicillin to Whole Cell Preparations and to Particulate Enzyme-Binding of radioactive penicillin to whole cells indicated that the amounts bound by B. subtilis, B. cereus, X. aureus, and E. coli were approximately 12.9, 5.2, 5.7, and 1.2 nmoles per g, dry weight, of cells, respectively (Fig. 1). The binding was biphasic showing a relatively high initial uptake at low penicillin concentrations followed by an apparent second- ary uptake which was linearly related to the penicillin concentra- tion. The secondary uptake was relatively higher in E. coli than in the other organisms, and in all cases, the binding values given were obtained by extrapolating the linear secondary binding curve to zero concentration. Single determinations were carried out for E. coli, B. subtilis, and B. cereus, but the value given for S. aureus is an average of four determinations which range from 4.2 to 7.7 nmoles per g. It is not known whether this variation was due to experimental technique or to variations in the amount of binding sites in different cultures. In the case of X. aureus,
1 However, recent experiments have suggested that if the Mil- 2 The values for B. subtilis and S. aureus may be too high due to lipore filters are soaked in cold penicillin G solution before use, the the tendency of these organisms to form chains and clumps, re- blank values may be considerably lowered. spectively.
of penicillinase (800 units per ~1) were added, followed by an additional incubation for 10 min at 30”. The cells were then recovered by centrifugation at 9000 rpm for 10 min and washed with buffer of the above composition. Finally, they were sus- pended in 0.4 ml of 1% sodium dodecyl sulfate and counted.
determinations were carried out with both [r4C]penicillin and [3H]penicillin. The variation observed was not greater than the variation observed in several measurements with [14C]penicillin alone, thus giving confidence that the values for the specific activities of the two radioactive penicillin samples and the rela- tive efficiencies of counting were fairly accurate. The concen- trationsrequired for half-maximum binding under the conditions of the experiment were approximately 0.2 nmole of penicillin G per ml for B. subtilis, 0.25 nmole per ml for B. cereus, 0.1 to 0.25 nmole per ml for S. aureus, and approximately 3 nmoles per ml for E. coli.
The value for the amount of penicillin bound can be converted to molecules per cell using Avogadro’s number and experimen- tally determined values for the number of cells per g of dry weight. The number of molecules bound per cell was approxi- mately 8000 for B. subtilis, 4000 for S. aureus, and 740 for E. c01i.~
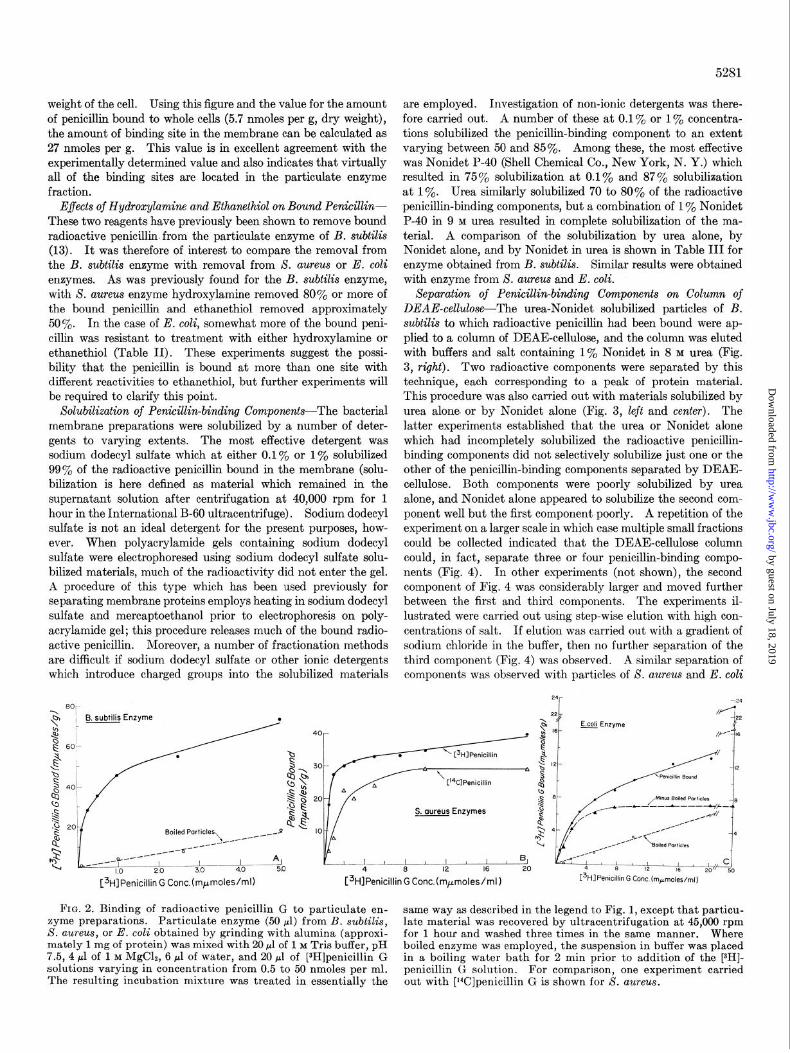
Binding of Radioactive Penicillin to Particulate Enzyme (Mem- brane) Preparations-Particulate enzyme preparations were pre- pared from all three enzymes after grinding cells with alumina. These preparations are mainly composed of membranes since they are made at concentrations of magnesium at which ribo- somes are normally dissociated. The binding curves (Fig. 2) yielded average values of 36 nmoles per g of enzyme protein for B. subtilis, 28 nmoles per g for X. aureus, and 7 nmoles per g for E. coli. Again the reaction was biphasic, and the values given were obtained by extrapolating the linear secondary phase of penicillin binding to zero concentration. The secondary phase was also seen in boiled enzyme preparations and is presumably due to some nonspecific binding or trapping of radioactive peni- cillin. Again, in the membrane preparations, it is ‘particularly pronounced in E. coli as it was in the whole cells. The concen- trations required for half-maximum binding under the conditions of the experiments were 0.1 to 0.5 nmole per ml for S. aureus and B. subtilis, and 1.5 nmoles per ml for E. coli, values which are in the same range as those recorded for binding to whole cells.
Enzyme was also prepared from S. uureus after lysis of cells with the cell wall lytic enzyme, lysostaphin. This enzyme prep- aration yielded a similar value for penicillin binding to that ob- tained after grinding with alumina, and moreover, indicated that virtually all of the penicillin binding sites in the whole cell are present in the particulate enzyme preparations (Table I). Experiments with lysostaphin also permitted the calculation that in X. aureus the amount of membrane is 21% of the dry
by guest on July 18, 2019http://w
ww
.jbc.org/D
ownloaded from
5281
weight of the cell. Using this figure and the value for the amount of penicillin bound to whole cells (5.7 nmoles per g, dry weight), the amount of binding site in the membrane can be calculated as 27 nmoles per g. This value is in excellent agreement with the experimentally determined value and also indicates that virtually all of the binding sites are located in the particulate enzyme fraction.
Efects of Hydroxylamine and Ethanethiol on Bound Penicillin- These two reagents have previously been shown to remove bound radioactive penicillin.from the particulate enzyme of B. subtilis (13). It was therefore of interest to compare the removal from the B. subtilis enzyme with removal from S. aureus or E. coli enzymes. As was previously found for the B. subtilis enzyme, with S. aureus enzyme hydroxylamine removed 80% or more of the bound penicillin and ethanethiol removed approximately 50%. In the case of E. coli, somewhat more of the bound peni- cillin was resistant to treatment with either hydroxylamine or ethanethiol (Table II). These experiments suggest the possi- bility that the penicillin is bound at more than one site with different reactivities to ethanethiol, but further experiments will be required to clarify this point.
Solubilization of Penicillin-binding Components-The bacterial membrane preparations were solubilized by a number of deter- gents to varying extents. The most effective detergent was sodium dodecyl sulfate which at either 0.1 y. or 1 y. solubilized 99% of the radioactive penicillin bound in the membrane (solu- bilization is here defined as material which remained in the supernatant solution after centrifugation at 40,000 rpm for 1 hour in the International B-60 ultracentrifuge). Sodium dodecyl sulfate is not an ideal detergent for the present purposes, how- ever. When polyacrylamide gels containing sodium dodecyl sulfate were electrophoresed using sodium dodecyl sulfate solu- bilized materials, much of the radioactivity did not enter the gel. A procedure of this type which has been used previously for separating membrane proteins employs heating in sodium dodecyl sulfate and mercaptoethanol prior to electrophoresis on poly- acrylamide gel; this procedure releases much of the bound radio- active penicillin. Moreover, a number of fractionation methods are difficult if sodium dodecyl sulfate or other ionic detergents which introduce charged groups into the solubilized materials
SO,-
+ ! B. subtilis Enzyme
[3H]Penicillln G Conc.(mpmoles/ml)
are employed. Investigation of non-ionic detergents was there- fore carried out. A number of these at 0.1 y. or 1 To concentra- tions solubilized the penicillin-binding component to an extent varying between 50 and 85%. Among these, the most effective was Nonidet P-40 (Shell Chemical Co., New York, N. Y.) which resulted in 75% solubilization at 0.1 y. and 87 y. solubilization at 1%. Urea similarly solubilized 70 to 80% of the radioactive penicillin-binding components, but a combination of 1 To Nonidet P-40 in 9 M urea resulted in complete solubilization of the ma- terial. A comparison of the solubilization by urea alone, by Nonidet alone, and by Nonidet in urea is shown in Table III for enzyme obtained from B. subtilis. Similar results were obtained with enzyme from S. aureus and E. coli.
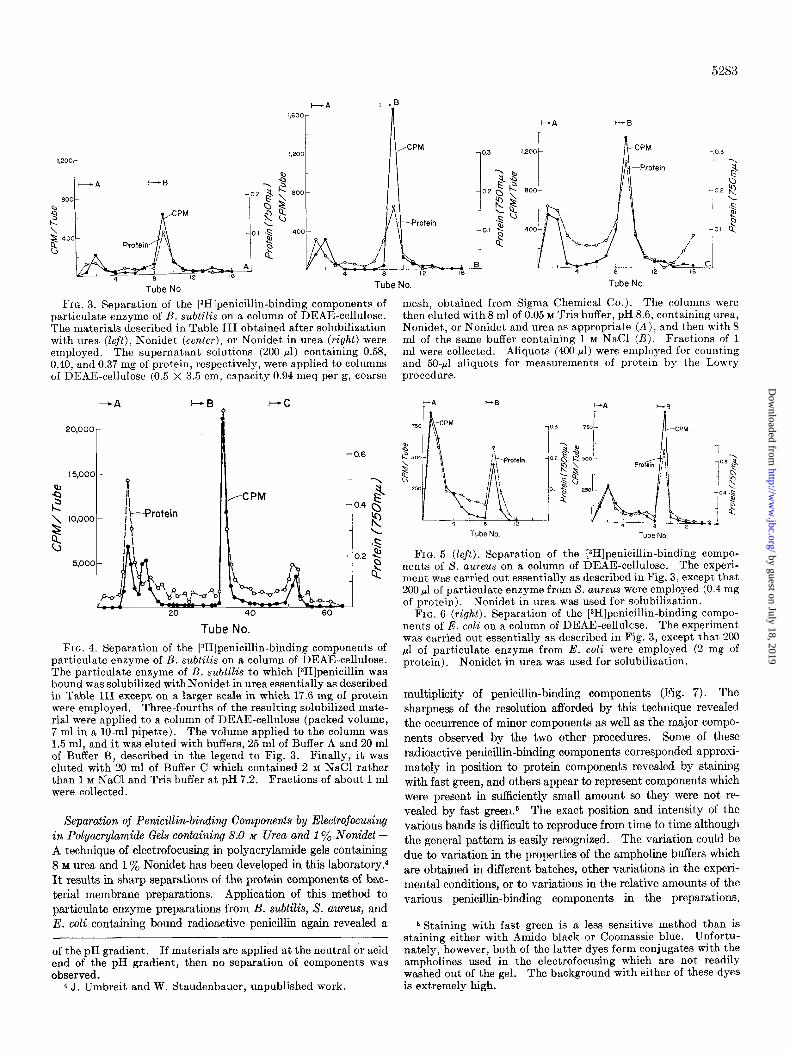
Separation of Penicillin-binding Components on Column of DEAE-cellulose-The urea-Nonidet solubilized particles of B. subtilis to which radioactive penicillin had been bound were ap- plied to a column of DEAE-cellulose, and the column was eluted with buffers and salt containing 1% Nonidet in 8 M urea (Fig. 3, right). Two radioactive components were separated by this technique, each corresponding to a peak of protein material. This procedure was also carried out with materials solubilized by urea alone. or by Nonidet alone (Fig. 3, left and center). The latter experiments established that the urea or Nonidet alone which had incompletely solubilized the radioactive penicillin- binding components did not selectively solubilize just one or the other of the penicillin-binding components separated by DEAE- cellulose. Both components were poorly solubilized by urea alone, and Nonidet alone appeared to solubilize the second com- ponent well but the first component poorly. A repetition of the experiment on a larger scale in which case multiple small fractions could be collected indicated that the DEAE-cellulose column could, in fact, separate three or four penicillin-binding compo- nents (Fig. 4). In other experiments (not shown), the second component of Fig. 4 was considerably larger and moved further between the first and third components. The experiments il- lustrated were carried out using step-wise elution with high con- centrations of salt. If elution was carried out with a gradient of sodium chloride in the buffer, then no further separation of the third component (Fig. 4) was observed. A similar separation of components was observed with particles of S. aureus and E. coli
C3H]Penicillin G Conc.(mpmoles/ml)
4 8 12 [3HlPenicillin G Cont. (m+moles/m~)
FIG. 2. Binding of radioactive penicillin G to particulate en- same way as described in the legend to Fig. 1, except that particu- zyme preparations. Particulate enzyme (50 ~1) from B. subtilis, S. aureus, or E. coli obtained by grinding with alumina (approxi-
late material was recovered by ultracentrifugation at 45,000 rpm for 1 hour and washed three times in the same manner. Where
mately 1 mg of protein) was mixed with 20 ~1 of 1 M Tris buffer, pH boiled enzyme was employed, the suspension in buffer was placed 7.5, 4 ~1 of 1 M MgC12, 6 pl of water, and 20 ~1 of [3H]penicillin G in a boiling water bath for 2 min prior to addition of the [3H]- solutions varying in concentration from 0.5 to 50 nmoles per ml. penicillin G solution. For comparison, one experiment carried The resulting incubation mixture was treated in essentially the out with [rYJpenicillin G is shown for 8. aureus.
by guest on July 18, 2019http://w
ww
.jbc.org/D
ownloaded from

Comparison of binding of [3H]penicillin G to whole cells of S. aureus
TABLE I TABLE II
Reversal of [r4C]penicillin G binding to various particulate enzymes by hydroxylamine or ethanethiol and to particulate enzyme obtained from whole cells
before or after [3H]penicillin G binding
Each reaction tube contained 100 ~1 of cell suspension (9 mg, dry weight), 400 pl of 0.05 M Tris buffer, pH 7.5, containing lo-’ hf MgC12, 200 ~1 of 1 M Tris base, and 40 ~1 of 1 M MgC12, 60 ~1 of water, and 200 ~1 of [3H]penicillin, either 10 nmoles per ml or 37.5 nmoles per ml. After incubation at 30” for 30 min, 1 ml of 0.05 M Tris buffer containing 10-h M MgClz was added, and the mixture was centrifuged at 10,000 rpm for 5 min. The pellet was washed three times. Experiment 1, for a measurement of penicillin binding to whole cells, this pellet was dissolved in 0.4 ml of buffer and then counted. Experiment 2, for measure- ment of the amount of radioactive penicillin-binding component recovered after lysostaphin treatment, the pellet was suspended in 0.4 ml of buffer to which were added 20 ~1 of lysostaphin (10 mg per ml) and 20 ~1 of DNase (1 mg per ml). After incubation at 37” for 30 min, the particulate material was recovered by centrifugation in the International B-60 ultracentrifuge at 50,000 rpm for 1 hour, washed once, resuspended in 0.4 ml of the same buffer and then counted. Experiment 3, for measurement of the amount of penicillin bound to the particulate enzyme obtained, the [zH]penicillin was omitted from the original incubation and replaced by water. The cell pellet which was recovered by centrifugation was resuspended in 1 ml of buffer containing 0.9% NaCl, then 20 pl of lysostaphin were added. After incubation at 37” for 1 hour, the particulate material was recovered by ultracentrifugation at 50,000 rpm for 1 hour. It was then treated with [EH]penicillin in essentially the same way as the treatment of the whole cells, except that 20 ~1 of DNase (5 mg per ml) were included in the incubation mixture. After incubation at 30” for 30 min, the particulate material was recovered by ultracentrifuga- tion and washed two times with buffer, resuspended in 0.4 ml of the buffer and then counted.
Particulate enzyme preparations containing [i”C]penicillin G from B. subtilis, S. aureus, and E. coli (500 ~1) were treated with (a) 500 ~1 of 0.05 M Tris buffer, pH 7.5, containing 1 mM MgC12, (b) 500 ~1 of neutral 2 M NHZOH, or (c) 75 ~1 of ethanethiol in 500 ~1 of buffer. After shaking for 1 hour at 25”, the incubation mixtures were dialyzed against 0.01 M Tris buffer containing 1 mM MgClz overnight. After ultracentrifugation at 50,000 rpm for 30 min, the resulting precipitate was solubilized by treatment with 500 ~1 of 1% Nonidet in 9 M urea. After ultracentrifugation, 25 ~1 of the supernatant solution were counted.
SZUIlple [WPenicillin G bound
cPm % remaining
B. subtilis Control. +NH,OH +CH&HzSH..
S. aureus
8,210 1,910 4,290
23 53
Control. +NHeOH . +CH&H,SH.
E. coli
11,115 1,430 5,760
13 52
Control.................... 4,000 +NH,OH . 2,480 +CH&H&SH. 2,900
62 72
TABLE III
Solubilization of penicillin-binding components of B. subtilis by various methods
Experiment
Amount bound after treatment with following amounts of
[~Hlpenicillin
2.0 nm;es per 7.5 nm$es per
nmoles/g
1. Binding to whole cells. . . 4.58 5.19 2. Binding to whole cells, then re-
The particulate enzyme from B. subtilis to which [3H]penicillin C had been bound (300 ~1) was recovered by centrifugation, and the pellet was then treated with 600 ~1 of 9 M urea, 600 ~1 of 1%; Nonidet, or 600 ~1 of 1% Nonidet in 9 M urea during shaking for 30 min at room temperature. After ultracentrifugation at 45,000 rpm for 30 min, the particulate material was resuspended in 600 ~1 of the same solutions, and the procedure was repeated. The supernatant solutions from the two treatments were combined, and the pellet was resuspended in 0.5 ml of buffer.
covery of particulate enzyme.. 5.50 6.36 3. Recovery of particulate enzyme,
Treatment and fraction Total radioactivity Total protein
l- thenbinding..................... 5.12 6.75
mg
9 M urea
containing bound penicillin (Figs. 5 and 6). In the case of
particles from S. aureus, the relative amounts of the two com- ponents were reversed, the first being considerably larger than
the second. At first, it seemed possible that the separation ob- served was due to an artifact resulting from breakthrough of the
column. However, columns operated at different loads of par-
ticulate material yielded the same results, and as will be shown below, electrofocusing has shown that the column, as operated,
sharply separated neutral and basic proteins in the first peak
from more acidic proteins in the second peak.
Supernatant. Precipitate. .
1% Nonidet
26,800 (78%) 3.00 (83Yo) 7,600 0.64
Supernatant. Precipitate.
1% Nonidet in 9 M urea Supernatant Precipitate. .
25,600 (7399 2.76 (68LTc) 9,600 1.30
38,700 (100%) 75
-.
4.32 (96%) 0.19
Separation of Penicillin-bin&g Compo?zents of B. subtilis by
Electrojocusing on Thin Layer of Sephadex G-75-A technique for separation of proteins by electrofocusing on thin layer plates has been described (17, 18). This technique has been applied to separation of the penicillin-binding components in the particulate
fraction of B. subtilis (see Fig. 15 below). As was seen with
the DEAE-cellulose columns, separation of two major compo- nents was also observed by thin layer electrofocusing. Similar results have been obtained with particles from S. aureus, but the experiments have not yet been carried out with E. colL3
3 One difficulty with the electrofocusing technique should be noted ; that is, some portion of the penicillin-binding component’ is electrophoretically immobile under these conditions. In the experiments shown, the material was applied at the alkaline end
by guest on July 18, 2019http://w
ww
.jbc.org/D
ownloaded from
+-B
-& 4
Tube No. Tube No. Tube No
FIG. 3. Separation of the [3H]penicillin-binding components of mesh, obtained from Sigma Chemical Co.). The columns were particulate enzyme of B. subtilis on a column of DEAE-cellulose. then eluted with 8 ml of 0.05 M Tris buffer, pH 8.6, containing urea, The materials described in Table III obtained after solubilization Nonidet, or Nonidet and urea as appropriate (A), and then with 8 with urea (left), Nonidet (center), or Nonidet in urea (right) were ml of the same buffer containing 1 M NaCl (6). Fractions of 1 employed. The supernatant solutions (200 ~1) containing 0.58, ml were collected. Aliquots (400 ~1) were employed for counting 0.40, and 0.37 mg of protein, respectively, were applied to columns and 50-~1 aliquots for measurements of protein by the Lowry of DEAE-cellulose (0.5 X 3.5 cm, capacity 0.94 meq per g, coarse procedure.
0.6
I5.000
0.4 z Q
10,000 ?
*S 0.2 5
5,000 %
20 40 60
Tube No.
FIG. 4. Separation of the [3H]penicillin-binding components of particulate enzyme of B. subtilis on a column of DEAE-cellulose. The particulate enzyme of B. subtilis to which [3H]penicillin was bound was solubilized with Nonidet in urea essentially as described in Table III except on a larger scale in which 17.6 mg of protein were employed. Three-fourths of the resulting solubilized mate- rial were applied to a column of DEAE-cellulose (packed volume, 7 ml in a IO-ml pipette). The volume applied to the column was 1.5 ml, and it was eluted with buffers, 25 ml of Buffer A and 20 ml of Buffer B, described in the legend to Fig. 3. Finally, it was eluted with 20 ml of Buffer C which contained 2 M NaCl rather than 1 M NaCl and Tris buffer at pH 7.2. Fractions of about 1 ml were collected.
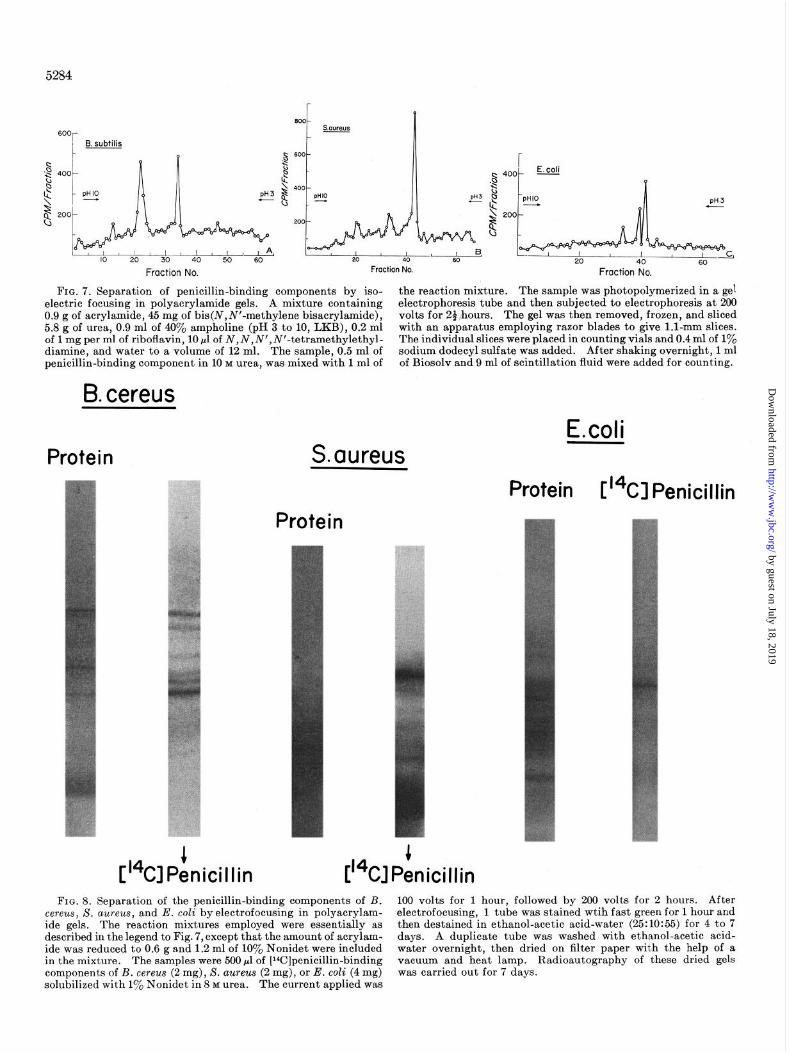
Separation of Penicillin-binding Components by Electrojocusing in Polyacrylamide Gels containing 8.0 M Urea and I yo Nonidet - A technique of electrofocusing in polyacrylamide gels containing 8 M urea and 1 yO Nonidet has been developed in this laboratory.4 It results in sharp separations of the protein components of bac- terial membrane preparations. Application of this method to particulate enzyme preparations from B. subtilis, X. aureus, and E. coli containing bound radioactive penicillin again revealed a
of the pH gradient. If materials are applied at the neutral or acid end of the pH gradient, then no separation of components was observed.
4 J. Umbreit and W. Staudenbauer, unpublished work.
Tube No. Tube No.
FIG. 5 (left). Separation of the [aH]penicillin-binding compo- nents of S. aureus on a column of DEAE-cellulose. The experi- ment was carried out essentially as described in Fig. 3, except that 200 ~1 of particulate enzyme from S. aureus were employed (0.4 mg of protein). Nonidet in urea was used for solubilization.
FIG. 6 (right). Separation of the [aH]penicillin-binding compo- nents of E. coli on a column of DEAE-cellulcse. The experiment was carried out essentially as described in Fig. 3, except that 200 ~1 of particulate enzyme from E. coli were employed (2 mg of protein). Nonidet in urea was used for solubilization.
multiplicity of penicillin-binding components (Fig. 7). The sharpness of the resolution afforded by this technique revealed the occurrence of minor components as well as the major compo- nents observed by the two other procedures. Some of these radioactive penicillin-binding components corresponded approxi- mately in position to protein components revealed by staining with fast green, and others appear to represent components which were present in sufficiently small amount so they were not re- vealed by fast green.6 The exact position and intensity of the various bands is difficult to reproduce from time to time although the general pattern is easily recognized. The variation could be
due to variation in the properties of the ampholine buffers which
are obtained in different batches, other variations in the experi- mental conditions, or to variations in the relative amounts of the various penicillin-binding components in the preparations.
6 Staining with fast green is a less sensitive method than is staining either with Amido black or Coomassie blue. Unfortu- nately, however, both of the latter dyes form conjugates with the ampholines used in the electrofocusing which are not readily washed out of the gel. The background with either of these dyes is extremely high.
by guest on July 18, 2019http://w
ww
.jbc.org/D
ownloaded from
5284
600 B. subtilis
10 20 30 40 50 60
Fraction No. Fraction No Fraction No.
FIG. 7. Separation of penicillin-binding components by iso- the reaction mixture. The sample was photopolymerized in a gel electric focusing in polyacrylamide gels. A mixture containing electrophoresis tube and then subjected to electrophoresis at 200 0.9 g of acrylamide, 45 mg of bis(N,N’-methylene bisacrylamide), volts for 24:hours. The gel was then removed, frozen, and sliced 5.8 g of urea, 0.9 ml of 40% ampholine (pH 3 to 10, LKB), 0.2 ml with an apparatus employing razor blades to give 1.1~mm slices. of 1 mg per ml of riboflavin, 10 pl of N,N,N’,N’-tetramethylethyl- The individual slices were placed in counting vials and 0.4 ml of 1% diamine, and water to a volume of 12 ml. The sample, 0.5 ml of sodium dodecyl sulfate was added. After shaking overnight, 1 ml penicillin-binding component in 10 M urea, was mixed with 1 ml of of Biosolv and 9 ml of scintillation fluid were added for counting.
B. cereus
E.coli Protein S. cwreus
Protein V4Cl Penicillin Protein
FIG. 8. Separation of the penicillin-binding components of B. 100 volts for 1 hour, followed by 200 volts for 2 hours. After cereus, S. aureus, and E. coli byelectrofocusing in polyacrylam- electrofocusing, 1 tube was stained wtih fast green for 1 hour and ide gels. The reaction mixtures employed were essentially as then destained in ethanol-acetic acid-water (25:10:55) for 4 to 7 described in the legend to Fig. 7, except that the amount of acrylam- days. A duplicate tube was washed with ethanol-acetic acid- ide was reduced to 0.6 g and 1.2 ml of 10% Nonidet were included water overnight, then dried on filter paper with the help of a in the mixture. The samples were 500 ~1 of [Wlpenicillin-binding vacuum and heat lamp. Radioautography of these dried gels components of B. cereu~ (2 mg), S. aureus (2 mg), or E. coli (4 mg) was carried out for 7 days. solubilized with 1% Nonidet in 8 M urea. The current applied was
by guest on July 18, 2019http://w
ww
.jbc.org/D
ownloaded from

Variation does not seem to be due to proteolysis occurring in the preparations since treatment of the particulate enzyme of B. subtilis with the protease inhibitor, phenylmethylsulfonyl fluoride, either before or after the binding of the radioactive penicillin, did not result in significant alteration of the number and intensity of various bands. Similarly, no bands were seen in particles
B. subtilis
5285
heated at 100” for 5 mm and then treated with radioactive peni- cillin nor does free radioactive penicillin give any band under the conditions of the experiments; it is presumably either eluted into one of the buffer compartments or eluted from the gel during washing.
The experiments described above were carried out with [3H]-
B. cereus
A
Original
Not Absorbed Absorbed
on DEAE
FIG. 9. Electrofocusing on polyacrylamide gels of membrane proteins of B. subtilis and B. cereus separated on a column of DEAE-cellulose. DEAE-chromatography of penicillin-binding components of B. subtilis and B. cereus was carried out employing 1.5 ml of penicillin-binding component solubilized with 1% Nonidet in 8 M urea. The fractions in each of the two major peaks were pooled, dialyzed against water for 8 hours in the cold room, taken
B
Original
Not Absorbed Absorbed
on DEAE
to dryness by lyophilization, and dissolved in 1.25 ml of 1% Nonidet in 8 M urea. Part of each fraction was employed for electrofocusing as described in the legend to Fig. 8. The control sample was 500 ~1 of the original penicillin-binding components solubilized by Nonidet in urea. After electrofocusing, gels were stained with fast green.
by guest on July 18, 2019http://w
ww
.jbc.org/D
ownloaded from

5286
penicillin-labeled particulate enzyme. When [l%]penicillin-la- beled particulate enzyme was employed, it was possible to make radioautograms of the gels rather than having to slice them. Representative radioautograms for the [‘4C]penicillinbinding components of B. subtilis, B. cereus, S. aureus, and E. coli are shown in Figs. 8 and 16. Two major components were seen in B. subtilis. With B. cereus, five bands were seen, two more in- tense than the other three; the major bands appeared to coincide with prominent bands which stained with fast green. With X. aureus and E. coli, on the other hand, only a single major band was seen in each case, and in both of these cases, this major band did not correspond with a band which was stained by fast green. Close examination of the original gels suggests that there may be a minor fast green staining component in this position (visual- ized in the photograph in the case of E. coli but not seen at all in the photograph in the case of S. aureus). These results then show that under the conditions of these experiments a variety of major and minor bands labeled by radioactive penicillin can be demonstrated in all three organisms. It appears that E. coli and S. aureus contain only a single major penicillin-binding compo- nent. In contrast, B. subtilis contains two prominent compo- nents as well as several minor ones; in B. cereus, two major bands are complemented by three very clear minor ones.
Electrofocusing has also been carried out on the fractions ob- tained from the separation of the membranes of B. subtilis and B. cereus on a column of DEAE-cellulose. Staining with fast green indicates that the column had separated proteins based on charge, that is, neutral and basic proteins were in the fraction which was not absorbed on the column while more acidic proteins were in the fraction which was absorbed to and then eluted from the column (Fig. 9).
Effects of Treatment with Hydroxylamine and Ethunethiol on Penicillin-binding Components-Treatment with hydroxylamine removed 80 to 90% of the bound penicillin. Electrofocusing in polyacrylamide gels of the penicillin-binding components after treatment with hydroxylamine showed no residual radioactive band (Fig. 10). The 10 to 20% of bound penicillin which re- mained after hydroxylamine treatment (Table II) must repre- sent either trapped [14C]penicillin G which is not washed out of
I 20 40 60
Fraction No. FIG. 10. The effect of hydroxylamine on penicillin-binding
components of B. subtilis. The experiment was carried out essentially as described in the legend to Fig. 7, except that 0.98 mg of penicillin-binding component from B. subtilis was employed. The sample for treatment with hydroxylamine was centrifuged to recover particulate material followed by treatment with 1 ml of 1 1~ hydroxylamine at room temperature for 1 hour. After this treatment, the particulate material was again recovered by ultra- centrifugation, washed with Tris buffer, and finally dissolved in 0.5 ml of 10 ivr urea for use.
the preparation or a minor band which is not visualized under the conditions of the electrofocusing experiments. In the case of ethanethiol treatment, 50% of the radioactivity remained in the treated enzyme preparation. Electrofocusing of the penicillin- binding components from B. subtilis treated with ethanethiol re- vealed that one of the major bands and possibly some minor bands remained after this treatment. The question of whether or not the residual radioactivity could be removed by more prolonged or more drastic treatment with ethanethiol requires further in- vestigation (Fig. 11).
Similar results were obtained with penicillin-binding compo- nents of X. aureus. With preparations from E. coli, 60 to 70% of the bound radioactivity remained after treatment with hy- droxylamine or ethanethiol (Table II). Electrofocusing in poly- acrylamide gels revealed that some of the bands had disappeared after these treatments, but most or all of them had been reduced in amount.
Effect of Cephalothin Pretreatment on Penicillin-bin&g Compo- nents of B. subtilti-Treatment of particulate enzyme prepara- tions with unlabeled penicillin G, followed by removal of the un- labeled penicillin G by washing, results in saturation of all or nearly all of the penicillin-binding sites. These sites are then not available during subsequent treatment with radioactive peni- cillin G. This technique has been used to measure the affinity of the penicillin-binding site in S. aureus for various penicillins (19). During the course of other experiments, it was observed using particulate enzyme from B. subtilis that many more of the radioactive penicillin G binding sites were saturated during prior treatment with penicillin G than with cephalothin (Fig. 12A). In the case of S. aureus, however, the number of binding sites
Control
s B 150 + CH, CH2 SH
50 L
I I I I 20 40 60
Fraction No.
FIG. 11. Effect of ethanethiol treatment on [3H]penicillin- binding component from B. subtilis. The experiment was carried out as described in the legend to Fig. 7 employing 1.2 mg of peni- cillin-binding component of B. subtilis. For the ethanethiol treatment, 500 ~1 of [3H]penicillin-binding component were mixed with 1.35 ml of water and 145 ~1 of ethanethiol, followed b.y incu- bation at 25” with shaking for’1 hour. After dilution with-buffer, the particulate material-was recovered by centrifugation and washed twice. It was then susnended in 20 ml of buffer and dialyzed against 4 liters of 0.05*M Tris buffer containing 1 mM MgCI, overnight. It is important to carry out the dialysis because small amounts of residual ethanethiol may interfere with pho- topolymerization of the gel. Finally, the particulate material was recovered by ultracentrifugation and resuspended in 500 ~1 of buffer. For the electrofocusing, 100 ~1 of this material were mixed with 400 pl of 10 M urea.
by guest on July 18, 2019http://w
ww
.jbc.org/D
ownloaded from
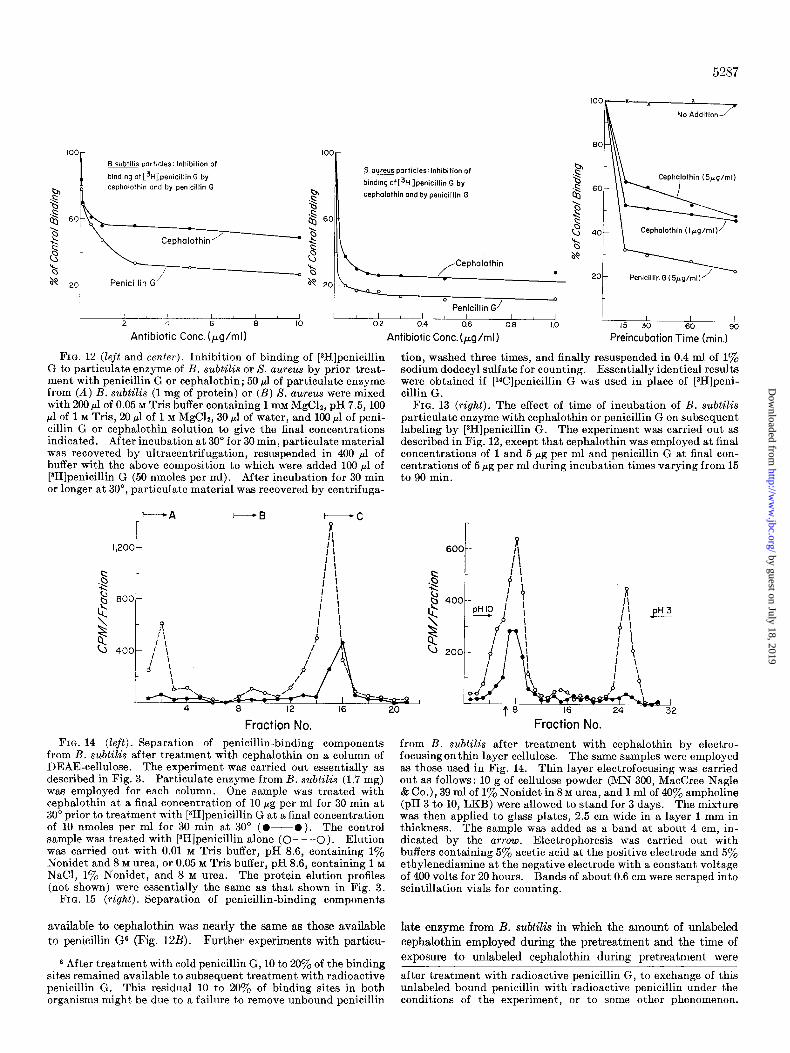
5287
‘ooR---x
x
No Addition
m particles: Inhibition of
binding of [3H]penicillin G by cephalothin and by penicillin G
L-+-l 1 1 I 4 6 8 IO
Antibiotic Cont. (pg/ml)
60 I
Soureus particles:lnhibition of
binding ofC3H]penicillin G by
cephalothin and by penicillin G
Penicillin G/
02 0.4 0.6 0.6 1.0
Antibiotic Conc.(pg/ml)
FIG. 12 (left and center). Inhibition of binding of [3H]penicillin G to particulate enzyme of B. subtilis or S. aureus by prior treat- ment with penicillin G or cephalothin; 50 ~1 of particulate enzyme from (A) B. subtilis (1 mg of protein) or (B) S. aureus were mixed with 200 ~1 of 0.05 M Tris buffer containing 1 mM MgC12, pH 7.5, 100 ~1 of 1 M Tris, 20 ~1 of 1 M MgClz, 30 ~1 of water, and 100 ~1 of peni- cillin G or cephalothin solution to give the final concentrations indicated. After incubation at 30” for 30 min, particulate material was recovered by ultracentrifugation, resuspended in 400 ~1 of buffer with the above composition to which were added 100 ~1 of [3H]penicillin G (50 nmoles per ml). After incubation for 30 min or longer at 30”, particulate material was recovered by centrifuga-
tion, washed three times, and finally resuspended in 0.4 ml of 1% sodium dodecyl sulfate for counting. Essentially identical results were obtained if [‘4C]penicillin G was used in place of [3H]peni- cillin G.
FIG. 13 (right). The effect of time of incubation of B. subtilis particulate enzyme with cephalothin or penicillin G on subsequent labeling by [3H]penicillin G. The experiment was carried out as described in Fig. 12, except that cephalothin was employed at final concentrations of 1 and 5 rg per ml and penicillin G at final con- centrations of 5 rg per ml during incubation times varying from 15 to 90 min.
s l\\L Cephajothin (5pg/ml)
Cephalofhin (Ipg/ml)
Penicillin G (5yg/ml)
I I I I 15 30 60 90
Preincubotion Time (min.)
I I 4 8 12 16 20
Fraction No. Fraction No.
FIG. 14 (left). Separation of penicillin-binding components from B. subtilis after treatment with cephalothin on a column of DEAE-cellulose. The experiment was carried out essentially as described in Fig. 3. Particulate enzyme from B. subtilis (1.7 mg) was employed for each column. One sample was treated with cephalothin at a final concentration of 10 pg per ml for 30 min at 30” prior to treatment, with [3H]penicillin G at a final concentration of 10 nmoles per ml for 30 min at 30” (0-O ). The control sample was treated with [aH]penicillin alone (C- - -0). Elution was carried out with 0.01 M Tris buffer, pH 8.6, containing 1% Nonidet and 8 M urea, or 0.05 M Tris buffer, pH 8.6, containing 1 M NaCl, 1% Nonidet, and 8 M urea. The protein elution profiles (not shown) were essentially the same as that shown in Fig. 3.
FIG. 15 (right). Separation of penicillin-binding components
from B. subtilis after treatment with cephalothin by electro- focusingon thin layer cellulose. The same samples were employed as those used in Fig. 14. Thin layer electrofocusing was carried out as follows: 10 g of cellulose powder (MN 300, MacCree Nagle & Co.), 39 ml of 1% Nonidet in 8 M urea, and 1 ml of 40% ampholine (pH 3 to 10, LKB) were allowed to stand for 3 days. The mixture was then applied to glass plates, 2.5 cm wide in a layer 1 mm in thickness. The sample was added as a band at about 4 cm, in- dicated by the arrow. Electrophoresis was carried out with buffers containing 5% acetic acid at the positive electrode and 5% ethylenediamine at the negative electrode with a constant voltage of 400 volts for 20 hours. Bands of about 0.6 cm were scraped into scintillation vials for counting.
available to cephalothin was nearly the same as those available
to penicillin G6 (Fig. 12B). Further experiments with particu-
6 After treatment with cold penicillin G, 10 to 20% of the binding sites remained available to subsequent treatment with radioactive penicillin G. This residual 10 to 20% of binding sites in both organisms might be due to a failure to remove unbound penicillin
late enzyme from B. subtilis in which the amount of unlabeled
cephalothin employed during the pretreatment and the time of exposure to unlabeled cephalothin during pretreatment were
after treatment with radioactive penicillin G, to exchange of this unlabeled bound penicillin with radioactive penicillin under the conditions of the experiment, or to some other phenomenon.
by guest on July 18, 2019http://w
ww
.jbc.org/D
ownloaded from

5288
Control + Cephalothin
FIG. 16. Separation of penicillin-binding components from B. subtilis after treatment with cephalothin by electrofocusing on polyacrylamide gels. The preparation of the control and ceph- alothin-treated B. subtilis particles containing [aH]penicillin G was carried out essentially as described in Fig. 14. The mixture for photopolymerization was the same as that described in the legend to Fig. 7, except that 1.2 ml of 10% Nonidet were included. One milliliter of this reaction mixture was mixed with 500 ~1 of the B. subtilis particles containing 0.8 mg of protein.
varied (Fig. 13) showed that, indeed, only about 50% of the binding sites available to radioactive penicillin G were irreversi- bly inactivated during pretreatment with unlabeled cephalothin.
Particles from B. subtilis previously treated with unlabeled cephalothin and subsequently treated with radioactive penicillin G were then compared to particles treated only with radioactive penicillin by DEAE-cellulose chromatography, by thin layer electrofocusing, and by electrofocusing in polyacrylamide gels. Using all three techniques, it was observed that one of the major penicillin-binding components of B. subtilis reacted with labeled penicillin G after treatment with unlabeled cephalothin. The other major component was apparently reactive with cepha- lothin since after treatment with this antibiotic, it no longer bound radioactive penicillin G (Figs. 14 to 16).
DISCUSSION
247, 5123-5i31 ’ 2. MAASS, E. A., AND JOHNSON, M. J. (1949) J. Bacterial. 67, 415 3. MAASS. E. A.. AND JOHNSON. M. J. (1949) J. Bacterial. 68, 361 4. COOPER, P. D., AND ROWLE;, D. (i949) ‘Nature 163, 480 5. DANIEL, J. W., JR., AND JOHNSON, M. J. (1954) J. Bacterial. 67,
321 6. EAGLE, H. (1954) J. Exp. Med. 99, 207 7. EAGLE; H. (1954) J. Ezp. Med. 100, 103 8. COOPER, P. D. (1954) J. Gen. Microbial. 10, 236 9. SCHEPARTZ, S. A., AND JOHNSON, M. J. (1956) J. Bacterial. 71,
84 The main thrust of these experiments is that there appear to
be multiple binding components, at least in B. subtilis and B. cereus. While the meaning of this observation is not clear at the present time, a number of possibilities can be considered.
10. MOHBF,RG, J., AND JOHNSON, M. J. (1958) J. Bacterial. 76, 385 11. DUERKSEN, J. D. (1964) Biochim. Biophys. Acta 87, 123 12. ROGERS, H. J. (1967) Biochem. J. 103, 90 13. LAWRENCE. P. J.. AND STROMINGER. J. L. (1970) J. Biol. Chem,
1. Bacterial cells contain multiple penicillin-binding compo- nents. In this case, killing of the bacterial cell might result either from interaction with a given one of the components or else from interaction with any one of several of the penicillin- binding components. Similarly, interaction with some of the components may not result in a lethal effect on the bacterial cell. It is not known which of the binding components in E. coli is the irreversibly inactivated transpeptidase (16). For B. subtilis, results from this laboratory obtained subsequent to much of the work reported here have recently clarified the situation (20). The concentration of cephalothin used in these experiments for
246, 365313659 ’ 14. LAWRENCE, P. J., ROGOLSKY, M., AND HAHN, V. T. (1971) J.
Bacterial. 108, 662 15. HIGASHI. Y.. AND STROMINGER, J. L. (1970) J. Biol. Chem. 246,
3691-3696 16. IZAKI, K., MATSUHASHI, M., AND STROMINGER, J. L. (1968) J.
Biol. Chem. 243, 3180-3192 17. RADOLA, B. J. (1969) Biochim. Biophys. Acta 194, 335-338 18. DELINCBE, H., AND RADOLA, B. J. (1970) Biochim. Biophys.
Acta 200, 404407 19. EDWARDS, J. R., AND PARK, J. T. (1969) J. Bacterial. 99, 459 20. BLUMBERG, P. M., AND STROMINGER, J. L. (1971) Proc. Nat.
Acad. Sci. U. S. A. 68, 2814 21. LEVINE, B. B. (1965) Fed. Proc. 24, 45
prebinding is too low to inactivate the carboxypeptidase. The carboxypeptidase, therefore, must be among the cephalothin-re- sistant penicillin-binding components. Likewise, since the con- centrations of cephalothin used for prebinding are lethal for this organism, the killing site must be among the cephalothin-sensi- tive components (provided that the antibiotic combines irre- versibly with the killing site). In addition, inhibition of the car- boxypeptidase is not lethal for the cell (20) ; one can consequently conclude that at least in this organism one of the penicillin-bind- ing components is dispensable.
Enzymes sensitive to penicillin have not yet been identified in cell-free preparations of S. aureus, but in vivo experiments indi- cate that this organism contains a penicillin-sensitive transpep- tidase. B. cereus has not yet been extensively studied from this point of view.
2. Some of the penicillin-binding components observed (more likely the minor ones) may represent nonenzymatic reaction of penicillin G with various proteins in the bacterial preparation, similar to the nonenzymatic reaction which results in the forma- tion of penicilloyl albumin or penicilloyl polylysine (21). A small amount of radioactive penicillin may still be bound after boiling the particulate enzyme (see Fig. 2), but in any case, boiling pre- vents virtually all of the specific binding of penicillin.
3. The multiple components eluted from the DEAE-column or seen by isoelectrofocusing might be due in part to aggregation or to variations in protein charge (e.g., due to differences in the state of amidation). The technique requires refinement, and further experiments will be needed before one can be certain of the precise number of binding components, especially with regard to the minor components. However, the finding that the prebinding of cephalothin to particulate preparations of B. subtilis can prevent the subsequent binding of penicillin G to only certain of the penicillin-binding components argues very strongly for the ex- istence of at least two classes of penicillin-binding components in this organism.
REFERENCES
1. SANDERMANN. H.. AND STROMINGER. J.L. (1972) J.Biol. Chem.
by guest on July 18, 2019http://w
ww
.jbc.org/D
ownloaded from

Hidekazu Suginaka, Peter M. Blumberg and Jack L. StromingerEscherichia coli, and Staphylococcus aureus
Bacillus subtilis, Bacillus cereus,Multiple Penicillin-binding Components in
1972, 247:5279-5288.J. Biol. Chem.
http://www.jbc.org/content/247/17/5279Access the most updated version of this article at
Alerts:
When a correction for this article is posted•
When this article is cited•
to choose from all of JBC's e-mail alertsClick here
http://www.jbc.org/content/247/17/5279.full.html#ref-list-1
This article cites 0 references, 0 of which can be accessed free at
by guest on July 18, 2019http://w
ww
.jbc.org/D
ownloaded from